Task-specific signal transmission from prefrontal cortex in visual selective attention
|
|
|
- Edith Edwards
- 5 years ago
- Views:
Transcription
1 ARTICLES Task-specific signal transmission from prefrontal cortex in visual selective attention Yosuke Morishima 1, Rei Akaishi 1, Yohei Yamada 1, Jiro Okuda 2, Keiichiro Toma 1 & Katsuyuki Sakai 1 29 Nature America, Inc. All rights reserved. Our voluntary behaviors are thought to be controlled by top-down signals from the prefrontal cortex that modulate neural processing in the posterior cortices according to the behavioral goal. However, we have insufficient evidence for the causal effect of the top-down signals. We applied a single-pulse transcranial magnetic stimulation over the human prefrontal cortex and measured the strength of the top-down signals as an increase in the efficiency of neural impulse transmission. The impulse induced by the stimulation transmitted to different posterior visual areas depending on the domain of visual features to which subjects attended. We also found that the amount of impulse transmission was associated with the level of attentional preparation and the performance of visual selective-attention tasks, consistent with the causal role of prefrontal top-down signals. It is generally accepted that top-down signals from the prefrontal cortex are important for cognitive control 1 4.Thesignalsarethoughttobe causal to the task-dependent modulation of neural activity in the posterior cortices and facilitate the processing of task-relevant information 5 7. This idea is supported by the time precedence of prefrontal activity 8,9, correlation of activity between the prefrontal and posterior regions 1 12 and modulation of activity in the posterior regions after prefrontal inactivation The current evidence is, however, insufficient to prove the causal relationship because it does not necessarily indicate the direct influence of the prefrontal signals over the posterior regions. Other studies have shown that microstimulation to the frontal eye field (FEF) in monkeys modulates neural responses in a posterior visual area within 4 ms of the stimulation, consistent with direct transmission of neural impulse from the FEF 18,19. It has also been shown that stimulation of the human FEF modulates the excitability of neurons in the human visual motion sensitive area (hmt+) at a latency of 2 4 ms 2. However, the task-specific nature of the physiological top-down signals remains open because the behavioral task was not manipulated in these studies. Also, the behavioral relevance of the physiological signals can not be tested. Here, we used transcranial magnetic stimulation () as a probe to examine the efficiency of the neural impulse transmission from the prefrontal cortex to posterior regions. The rationale for this technique is that stimulation to a neuronal population in a source region induces a current spread toward the anatomically connected target regions and that the direction and the amount of the current spread are modulated depending on the functional status of the neural network, which is determined by the task at hand. Using a cue-based visual feature attention task, we found that the neural impulse from the FEF induced by was transmitted to different posterior visual areas depending on the attended visual features. RESULTS Behavior We asked normal human subjects to perform a visual-discrimination task for motion direction or face gender. The visual target consisted of a vertical grating moving toward the right or left, superimposed on an image of a male or female face (Fig. 1a). A task-instruction cue preceded the target with a long or short cue-target interval (, stimulus onset asynchrony of 1,5 or 15 ms, respectively). On half of the trials, we applied a single-pulse at 134 ms after the onset of a visual target over the scalp position at electrode FC2, which was located above the junction of the precentral sulcus and superior frontal sulcus in most subjects (Fig. 1b). The mean coordinate of the stimulated region on the cortical surface was (29, 4, 63) based on the MNI (Montreal Neurological Institute) standard brain, which corresponds to the human FEF 21. The response time of the subjects for the visual-discrimination task did not differ significantly depending on the feature of the target to which the subjects attended (F 1,12 ¼.7, P ¼.93; Supplementary Fig. 1 online). The response time was significantly shorter on trials with long compared with trials with short, indicating behavioral preparation effect (F 1,12 ¼ 35.23, P o.1). The interaction between the feature and was not significant (F 1,12 ¼ 2.1, P ¼.16). As for the accuracy of performance, there was a significant main effect of attended feature, with higher accuracy for the face task (F 1,12 ¼ 9.93, P ¼.8). The main effect of and the interaction between the feature and were not significant ( main effect: F 1,12 ¼ 3.64, P ¼.8; interaction effect feature : F 1,12 ¼.49, P ¼.49). Notably, there was no significant main effect of on behavioral performance in terms of response time and accuracy (response time: F 1,12 ¼.11, P ¼.68; accuracy: F 1,12 ¼ 1.19, P ¼.29). This is because we used lowintensity (8% of the active motor threshold, 27 42% of the 1 Department of Cognitive Neuroscience, Graduate School of Medicine, University of Tokyo, Hongo 7-3-1, Bunkyo-ku, Tokyo, Japan, Brain Science Institute, Tamagawa University, Tamagawa-gakuen 6-1-1, Machida-shi, Tokyo, Japan, Correspondence should be addressed to K.S. (ksakai@m.u-tokyo.ac.jp). Received 19 September; accepted 3 November; published online 21 December 28; doi:1.138/nn.2237 NATURE NEUROSCIENCE VOLUME 12 [ NUMBER 1 [ JANUARY 29 85
 Illustration of the timeline of the behavioral task.")

2 ARTICLES a b c Cue Target or 6 (µv), long, short, long 4, short P ms 75 ms 2 (ms) 15 or 1,5 ms 134 ms Nature America, Inc. All rights reserved. Figure 1 Experimental design and -induced ERP. (a) Illustration of the timeline of the behavioral task. Subjects discriminated either the direction of the motion of a vertical grating or the gender of a face on the basis of a cue. The square and circle cues indicate the motion- and face-discrimination tasks, respectively. The stimulus onset asynchrony between the cue and target was either 15 or 1,5 ms. On half of the trials, was given at 134 ms after the onset of a target. (b) Left, scalp position and orientation of the coil. Right, location of the site superimposed on the structural image of the MNI template brain. (c) Mean -induced ERP at occipito-temporal electrode P8. Red and blue lines indicate the motion and face tasks, respectively. The thick and thin lines indicate trials with long and short, respectively. The shaded region corresponds to the 2 4-ms period after the onset of the. No digital filter was applied to the traces. maximum output of the stimulator). We used as a probe to examine the effective connectivity of the neuronal network and not as a tool to manipulate the neural processing in the source region. The effect did not interact significantly with feature and in terms of response time ( feature: F 1,12 ¼ 2.14, P ¼.16; : F 1,12 ¼.83, P ¼.37) and accuracy ( feature: F 1,12 ¼.49, P ¼.49; : F 1,12 ¼.3, P ¼.98). We also confirmed that the did not induce any detectable saccadic eye movements. Task-specific effect on EEG potentials Applying over the right FEF induced changes in the scalp potentials recorded with electroencephalography (EEG). To compare the effect on scalp potentials, we subtracted the event-related potential (ERP) on no trials from that on trials (Fig. 1c). The initial effect, as well as the stimulus-related artifacts localized at the stimulation site, was followed by a long-distance effect over the frontal and posterior regions at 2 4 ms after the (Fig. 2). The peak of the -induced ERP moved to the vertex region at 4 6 ms and then spread again over the frontal and posterior regions, with maximum changes being observed at 1 12 ms. The difference in the ERPs of the and no trials cannot be explained by the baseline shift of the ERP before the, as there was no substantial difference in the ERPs during the and the period of time between the target onset and (Supplementary Fig. 2 online). During an experimental session, the and no trials were given in a pseudo-random order and the subjects were thus unable to anticipate the or the clicking sound that it made. The induced changes in the scalp potential cannot be explained by the latency shift of ERP components in no trials because there were no corresponding ERP components in no trials. Thus, the differential ERPs between the and no trials was considered to be induced purely by the. Notably, the -induced ERPs differed between the face and motion conditions when subjects had sufficient time to prepare for the task (long trials). We observed a significant difference (P o.5) in the -induced ERP at 2 4 ms after over the right posterior parieto-occipital regions (Fig. 2). At this time period, the ERP on electrode P8 was positive in the motion condition and was negative in the face condition (Fig. 1c). At a later time window of 1 16 ms, there was also a significant difference in ERPs (P o.5) on the right posterior and central regions. Notably, there was no significant difference (P 4.5) in the -induced ERP in posterior cortical regions between the two conditions until 8 ms after the on trials with short, in contrast with the feature-specific -induced ERP on trials with long (Fig. 2). More specifically, there was a significant interaction (P o.5) between the task condition and over the right posterior parieto-occipital region at 2 4 ms after the (Fig. 2). This trend was also observed at 2 ms, but disappeared after 4 ms. We considered the possibility that the spread pattern of the induced activation on trials with short reflects anatomical connections with the FEF and that functional connectivity reflects task components that are nonspecific to the attended visual feature. In contrast, this pattern of functional connectivity is modulated by the task components that are specific to the attended visual feature on the long trials. To isolate the attention-related modulation, we subtracted the -induced ERPs on short trials from those on long trials. We focused on the ERPs at 2 4 ms after because the transmission of neural impulse from one cortical region to another has been shown to occur in this time range at a neuronal population level 2, In the motion task, we observed the modulation of the -induced ERP as a positive deflection of the ERP centered over the temporal to occipital regions (Fig. 3 and Supplementary Fig. 3 online). In contrast, we observed the modulation in the face task as a negative deflection of the ERP centered over the right parietal region and a positive deflection centered over the frontal region (z score map in Fig. 3). These differential maps of scalp topography reflect the taskdependent functional modulation in the pattern of cortical impulse transmission from the FEF. Notably, the difference between the motion and face conditions was not just the magnitude or the polarity of scalp potential in a localized region, but was instead the spatial pattern of the scalp potential, which suggests different sources for the task-specific ERP modulations. Task-specific effect on current source densities The idea of different sources for the ERP modulations was confirmed by the task-specific pattern in the cortical distribution of the current source densities that account for the ERP changes at 2 4 ms after the. We first estimated current source density for the -induced ERP on long trials by using low-resolution electromagnetic tomography (LORETA) 25. Among the source regions identified in frontal, parietal and temporal cortices, we found higher current source 86 VOLUME 12 [ NUMBER 1 [ JANUARY 29 NATURE NEUROSCIENCE


3 ARTICLES, long, long, short, short face, long z score face, short z score (ms) Interaction task 2 z score 3 3 (µv) Figure 2 Scalp maps of the effect on ERP. The top four rows show the time course of the mean voltage scalp maps for the effect on the ERP, which are calculated by subtracting the ERP on no trials from the ERP on trials. The maps are shown separately for each of the long and short and motion and face tasks. The fifth and sixth rows show the scalp distribution of the z scores representing the difference between the motion and face conditions for each of the long and short trials. The positive sign of the z score indicates more positive potentials in the motion compared with the face condition. The bottom row shows the interaction effect between attended feature and. The z score is shown in color only when the P value at each electrode reached significance (P ¼.5). 29 Nature America, Inc. All rights reserved. density in the lateral temporo-occipital region in the motion condition compared with the face condition (Fig. 4a).The change was more pronounced on the right hemisphere, which was the side on which was induced. The maximal difference in the current source density was located at coordinate (53, 67, 8), which corresponds to the hmt+ 26.In contrast, there was an increase in the current source density in the inferior temporal cortex in the face condition compared with the motion condition, which was more pronounced on the right side. The maximal difference was found at (46, 53, 2), which corresponds to the human fusiform face area (FFA) 27. Thus, stimulation of the FEF induced an increase of activity in the hmt+ in the motion condition at 2 4 ms, whereas stimulation induced an increase of activation in the FFA in the face condition at the same latency. For these task-specific posterior visual areas, we calculated the difference in the current source density between long and short trials for each of the motion and face conditions. These differential current source densities can be viewed as reflecting the modulation of -induced activation in the hmt+ and FFA resulting from attentional preparation. We found a significant region task interaction on the differential current source densities (F 1,12 ¼ 7.594, P ¼.17; Fig. 4b), which indicates enhancement of activation in the featurespecific visual areas that are relevant to the task at hand. In fact, the differential activation in the long trials relative to the short trials was significantly larger than zero in task-relevant visual areas (P ¼.3), but was not significantly different from zero in taskirrelevant areas (P ¼.84) (Fig. 4b). The difference between the taskrelevant and task-irrelevant areas was significant (P ¼.17). We assumed that the feature-specific pattern of the -induced ERP reflected the efficiency of the neural impulse transmission from the FEF to posterior visual areas. In trials with long, the subjects had sufficient time to prepare for the task and the top-down signals were being sent to the task-specific posterior regions after. In contrast, the subjects were not prepared in trials with short and the task-specific top-down signals were insufficient to modulate the direction of the impulse induced by. We considered the difference in response time between long and short trials to be the result of the level of attentional preparation, which we have shown to be associated with task-specific direction of the -induced impulse transmission. Association of -induced activation and behavior This idea is further supported by the relationship between the amount of the -induced ERP changes and behavioral response time. In each of the short and long conditions, we examined whether the variability in response time is associated with across-trial variability in the efficiency of impulse transmission from the FEF to posterior visual areas. We split the trials in each condition into fast- and slow-response trials and calculated the difference in the current source densities between them. For the short- condition, we found that the difference in the current source densities in task-relevant regions were significantly larger than zero, indicating higher activation in these regions in fast-response trials than in slow-response trials (P ¼.36; Fig. 4c). Notably, we found that the current source densities in areas irrelevant to the task at hand in these trials with short were also significantly higher in fast-response trials compared with slowresponse trials; the differential current source density was significantly larger than zero (P ¼.29; Fig. 4c). Thus, on trials with short preparation time, the amount of impulse transmission from prefrontal to posterior visual areas was associated with behavioral performance, but the direction of impulse transmission was not task dependent. In contrast, the difference in the induced activation between the fast- and slow-response trials in trials with long was not significantly different from zero in both the task-relevant and task-irrelevant areas (relevant, P ¼.45; irrelevant, P ¼.86; Fig. 4c). In these trials, the level of attentional preparation reached a plateau during the long preparation period and the variability in the response time may have been determined by the bottom-up processing of the target image. EEG potential z score long short long short 1.5 (µv) Figure 3 Mean voltage (top) and z score (bottom) scalp maps representing the difference between long and short trials at 2 4 ms after. For each of the motion (left) and face conditions (right), the ERP on short trials was subtracted from that on long trials. The positive sign of the z scores indicates more positive potential on long compared with short trials NATURE NEUROSCIENCE VOLUME 12 [ NUMBER 1 [ JANUARY 29 87

4 ARTICLES a R b.2 MT+ FFA.2 Task relevant Task irrelevant c.2 Task relevant Task irrelevant MT+ FFA ( 1 3 ) Activity difference (long short ).1..1 Activity difference (long short ).1..1 Activity difference (fast slow response).1. Long Short Nature America, Inc. All rights reserved. Figure 4 -induced current source density in posterior visual areas. (a) Mean -induced current source density at 2 4 ms after on the long trials. The estimated source densities are shown overlaid on the coronal slices of the standard brain at y ¼ 67 (top) and 53 (bottom). The cross-hairs indicate the peak coordinates of the hmt+ (53, 67, 8) (top) and FFA ( ) (bottom) at which there was maximal difference in the current source density between the motion and face conditions. R indicates the right side of the brain. (b) Left, difference in the current source density between long and short trials in the hmt+ and FFA, shown separately for the motion and face conditions. Right, the difference in the current source density between long and short trials is plotted for the task-relevant (hmt+ for motion task and FFA for face task) and task-irrelevant areas (FFA for motion task and hmt+ for face task). Here the data shown in a are log-scaled. Error bars indicate s.e.m. (c) Difference in the current source density between fast- and slow-response trials is plotted for task-relevant and task-irrelevant areas, separately for long and short trials. Here the data shown in a are log-scaled. Error bars indicate s.e.m. The difference in the -induced ERP between the fast- and slowresponse trials cannot be the result of the nonspecific arousal effect. We examined the pupil diameter of the subjects as an index of arousal level and found that it did not differ significantly between the fast- and slowresponse trials. In addition, the global mean field amplitude of the EEG potential induced by the, which has been shown to be associated with arousal level 28, did not differ between the fast- and slow-response trials. Perturbation of the FEF Finally, we confirmed that the FEF is important in feature discrimination by disrupting neural processing in the FEF. We applied double-pulse with higher intensity (65% of the maximal output of the stimulator, which was above the active motor threshold for all subjects) while the subjects performed the same motion- or facediscrimination task. The was given at 34 and 134 ms after the target presentation on either the right FEF or the vertex. There was a significant decrease in the accuracy of performance during trials compared with no trials when was given on the FEF (F 1,5 ¼ 18., P ¼.8), whereas there was no significant effect of when it was given on the vertex control site relative to the no Preparation effect (ms) 1 5 No trials (F 1,5 ¼.6, P ¼.8) (Supplementary Fig. 4 online). These results indicate that the FEF is important in the feature-discrimination task, as reported in previous studies Performance had a tendency to be more impaired in the motion task than in the face task (interaction between and task, F 1,5 ¼ 6., P ¼.57). We also tested the possibility that the perturbation of the right FEF biases the direction of visual motion judgment, but found that the performance did not differ between trials with leftward motion and those with rightward motion; the interaction between and motion direction was not significant (accuracy: F 1,5 ¼.14, P ¼.72; response time: F 1,5 ¼ 1.836, P ¼.23). In addition to a decrease in the accuracy of performance, we found that on the FEF diminished the behavioral advantage of a long preparation time (Fig. 5). The preparation effect, which was calculated by subtracting the response time on long trials from that on short trials, decreased significantly when was given to the FEF (t 5 ¼ 2.64, P ¼.46), but it did not change significantly when the was given to the vertex (t 5 ¼.66, P ¼.95). The effect on preparation was similar for both the motion and face task; the interaction between and task was not significant (F 1,5 ¼.69, P ¼.44). The results suggest that the FEF is important for preparation for the featurediscrimination task and that this occurs during the brief time period within 15 ms of target presentation. This finding complements our data on the task-specific pattern of effective connectivity from the FEF observed at the same time period during the long trials. Higher-intensity on the FEF during this time period disrupts the process of establishing task-specific patterns of effective connectivity and thus diminishes the benefit of attentional preparation in speeding up responses. FEF Vertex Figure 5 Effect of virtual lesion of the FEF on the behavioral preparation effect. In this experiment, was given with higher intensity at 34 and 134 ms after the onset of target visual image to create a virtual lesion. Left, the preparation effect, which was calculated by subtracting the response time on long trials from that on short trials, is plotted separately for trials with on the right FEF and for trials with no. The data for the motion and face conditions are combined. Right, data for a separate session in which the was applied over the vertex are shown. Error bars indicate s.e.m. DISCUSSION We examined whether the efficiency of a -induced impulse transmission reflects the influence of physiological top-down signals from the FEF over posterior visual areas during visual selective attention. A population of neurons in the FEF that have activity specific to the attended visual feature have been shown to be active before the presentation of a target stimulus 33 35, and these neurons are thought to send top-down signals to neurons in the posterior visual areas that are involved in the processing of that feature 36,37. In this situation, it can be thought that there is an increase in the responsiveness of the FEF 88 VOLUME 12 [ NUMBER 1 [ JANUARY 29 NATURE NEUROSCIENCE
5 ARTICLES 29 Nature America, Inc. All rights reserved. neurons, projecting to task-relevant visual areas, to the. Also, there is an increase in the responsiveness of neurons in the posterior visual areas to the inputs from the FEF induced by the. Both the pre- and postsynaptic mechanisms reflect the physiological top-down signals from the FEF during visual selective attention and we quantified their effects as a change in the -induced activation in the posterior visual areas. We propose that the -induced ERP in posterior areas reflects mainly orthodromic impulse transmission along the axons of FEF neurons, as scalp EEGs reflect summation of postsynaptic potentials in a local region: EEGs can not detect the firing of posterior visual neurons triggered by antidromic impulse. There still remains the possibility that antidromic firings of neurons may have resulted in organized oscillation in the gamma frequency range in the posterior visual areas 38,39. FEF neurons have been shown to increase firing in response to a visual stimulus at a specific spatial location or with a specific visual feature 37. However, the influences the activity of neurons in a region of several centimeters and activates millions of FEF neurons with different preferences for spatial location and visual feature. It remains unknown whether the neural impulse from the FEF at 134 ms after the visual target presentation codes specific information about the target. Here we found that the impulses from the FEF code information about the task that is to be performed, which is the domain of the visual feature to be attended. The important feature of the top-down control signal is its taskspecific nature. We have shown that stimulation of the prefrontal cortex induces activation in posterior visual areas in a manner that is specific to the task at hand. To date, studies have found task-dependent modulation of functional connectivity between the prefrontal and posterior cortices, but these findings tell us nothing about the direction of the influence (Supplementary Data and Supplementary Fig. 5 online) 11,12,4. Some studies have used mathematical modeling and have estimated the strength of task-specific influence from the prefrontal cortex over the posterior regions, but the estimated values are known to be strongly biased by the arbitrary choice of an anatomical model 4. Others have used multiple pulses of high-intensity to disrupt neural processing in the frontal or parietal region while subjects performed a visual-attention task 16,17,41. They found changes in EEG potentials later than 1 ms after the, and it is therefore hard to establish the direct influence of the site over the regions in which the changes in EEG potentials were observed. Also, the effects that were observed in those studies may reflect compensatory mechanisms after the perturbation by. Our study goes further than previous studies, showing transmission ofsignalsfromaspecificsourceregion,thefef,tospecifictarget regions, hmt+and FFA. Notably, the target regions were identified in an exploratory manner, rather than on the basis of pre-determined regions of interest. In addition, we observed effects at 2 4 ms after, suggesting that there was a direct cortico-cortical impulse transmission from the FEF to the posterior regions 2,22 24, although it remains possible that the effects were instead the result of oligosynaptic impulse transmission. Thus, the functional modulation of the neural circuit that weobservedwasspecificintermsofthesourceandtargetandwasalso specific in terms of the temporal relationship between the cause and effect. -induced current has been shown to spread along anatomical connections from the stimulation site 22 and this can be abolished during sleep 42. Here, we found a substantial change in the direction of the current spread during voluntary control of attention. The task-specific patterns in the -induced ERP cannot be accounted for by the difference in the magnitude of regional activation between the face and motion conditions, not only in the FEF, but also in all other cortical regions. On no trials, there was no significant difference in EEG potential between the two task conditions during the time window of ms after the target onset, which corresponds to the time window of 2 4 ms after on trials (P 4.5; Supplementary Fig. 6 online). Thus, the -EEG technique has uncovered changes in effective connectivity, which did not accompany changes in regional activation. Furthermore, our study found evidence supporting the behavioral importance of the top-down signals. The task-specific modulation of the direction of the impulse transmission from the FEF to posterior visual areas was associated with a behavioral preparation effect, which is determined by the. In addition, the amount of effective impulse transmission from the FEF was associated with across-trial variability in behavioral response on trials with short preparation, which may reflect the level of attentional preparation. However, the regional selectivity of the FEF projection was not observed on trials with short because of insufficient task preparation. In contrast, on trials with long, the effective connectivity from the FEF was directed specifically to the task-relevant visual areas. On these trials, the preparation for the target may have been saturated and the strength of effective connectivity from the FEF did not differ between fast- and slow-response trials. This suggests a mechanism of visual selective attention whereby nonspecific activation of visual neurons is followed by selective activation of task-relevant neurons 43,44. The tight association with behavior supports our argument that the efficiency in the transmission of the -induced impulse reflects physiological top-down signals that control our voluntary behavior. This idea is also supported by the finding that the perturbation of the FEF with higher-intensity resulted in an increase in errors in the featurediscrimination task and a decrease in the behavioral advantage of long preparation time, which suggests a failure to establish task-specific patterns of effective connectivity. We have not just shown an association between a single prefrontal region and behavior, but rather have found a tight functional link between a prefrontal region and posterior regions, as well as an association between the prefrontal efferent signals and behavior. Our results also demonstrate the feasibility of our technique for characterizing, at a specific time point of cognitive processing, the functional status of a neural network, which can be dissociated from the pattern of regional activation. METHODS Subjects. Fifteen normal subjects (two female, aged 19 26) participated in the -EEG experiments. We excluded two subjects from the analysis as a result of a low correct rate in the motion task (o6%). In addition, six normal subjects (two female, aged 19 32) participated in the -inactivation experiments. All subjects gave written informed consent before participating in this study. The study was approved by the ethics committee of the Graduate School of Medicine at the University of Tokyo. Cued discrimination task. Subjects were presented with a task instruction cue that was followed by a target stimulus on the black background of a LCD monitor. The target stimulus was a short, 75-ms movie (refresh rate of 6 Hz) with a visual angle of 6 degrees. Each frame of the movie was a composite bitmap image of a vertical sinusoidal grating and a grayscale face image (Softopia Japan Foundation; Fig. 1a). A black and white random-noise pattern was also added to the image to equate the behavioral response time between the motion- and face-discrimination tasks. The sinusoidal grating was shifted to the left or right at a speed of 121 s 1. A visual target was preceded by a white square or disc, which instructed the motion- and face-discrimination task, respectively. The duration of the cue presentation was 117 ms and the stimulus onset asynchrony between the cue and target was 15 ms for half of the trials and 1,5 ms for the other half. Subjects responded to the target with a button NATURE NEUROSCIENCE VOLUME 12 [ NUMBER 1 [ JANUARY 29 89
6 ARTICLES 29 Nature America, Inc. All rights reserved. press using either the index or middle finger of the right hand. Thus, the task was designed in a two-by-two fashion, (motion and face task) (long and short ). In one experimental session, 2 trials were given for each condition in a pseudo-random order with an intertrial interval of 2 3 s. Each subject was tested in 12 sessions on 2 separate days. EEG recording and. Foreachsubject,wefirstdeterminedthethreshold intensity of the necessary to evoke motor response. Subjects wore an EEG cap (Nexstim) and were seated on a chair with their hand resting on the desk and their right index finger extended and uplifted. Using a figure eight shaped flat coil (Magstim 2), we delivered a single-pulse to the scalp position corresponding to the left primary motor cortex (adjusted by moving the coil center in a step of 1 cm) while an electromyographic (EMG) recording was made from the right first dorsal interosseous muscle. The coil was placed tangentially over the scalp, such that the evoked current passed in a posterior to anterior direction in the brain. We defined the active motor threshold as the lowest intensity that evoked more than five small EMG responses (around 5 mv) in a series of ten stimulations. We then asked the subjects to perform the motion- or face-discrimination task while we recorded EEG from 6 scalp electrodes. The amplifier of the EEG was specially designed to allow pulses to be delivered over the electrode caps (eximia EEG, Nexstim). This was achieved by closing the circuit for 2 ms on trigger pulse from the magnetic stimulator 22.Wewereableto record artifact-free EEG signals 1 ms after the. The EEG signals were referenced to the left ear, filtered at the frequency of.1 5 Hz and sampled at 1,45 Hz. To reduce the artifacts resulting from the click sound of the pulse, the subjects wore earplugs. On half of the trials for each condition, a single-pulse (8% of active motor threshold) was delivered to the right FEF at 134 ms after the onset of the visual target. We chose this time to maximize the difference in the influence of the FEF stimulation between the motion and face tasks. This is based on the finding that the FEF neurons show differential responses to target and distractor at about 1 ms after the presentation of the image in a visual selective-attention task in monkeys 33,34. It has also been shown that the ERP that is observed at 17 ms after presentation of a face image, called N17, is enhanced when subjects attended to the face 45, suggesting that the top-down signal acts on posterior visual areas at this time period. With a cortico-cortical transmission time of 2 4 ms, we decided that the attentional modulation of the effect would be maximized when the was given over the FEF at B13 ms after the presentation of a target image. The order of the and no trials was randomized in each experimental session. The coil was placed tangentially over the FC2 electrode, with the handle of the coil pointing backwards and laterally approximately 45 degrees to the interhemispheric line. After the EEG recording session, the position of the coil was co-registered with the structural magnetic resonance imaging of the brain for each subject using the Brainsight frameless stereotaxy system (Brainsight, Magstim) for 6 of the 15 subjects. In the normalized structural magnetic resonance imaging, the center of the coil was clustered in a spherical region with a diameter of 12 mm across subjects and the mean coordinate of the brain surface just below the coil center was (29, 4, 63) in the MNI space (Fig. 1b). This position is considered to be the FEF in the human brain 21. In a separate session, we also applied on the primary somatosensory cortex (3 cm posterior to hand motor region) in three subjects as control experiments and found that the -induced ERPs did not differ between the motion and face tasks. Recording eye movement. The pupillary position of the left eye was recorded at 6 Hz with a ViewPoint eye tracker (Arrington Research) and PC-6 software. The eye tracker was adjusted before starting each session. We confirmed that there were no saccadic eye movements between 15 ms before and 3 ms after the presentation of a target. The pupil diameter of the subjects was also recorded at the same sampling rate. The pupil diameter showed a gradual decrease after the presentation of a target, peaking at roughly 5 ms after the presentation. The difference in arousal level is known to affect the pupil size after this light reflex 46. Thus, the changes in the pupil diameter relative to the pre-target period were calculated and averaged in a time window of 5 8 ms after the target onset and compared between fast- and slowresponse trials to see whether the difference in arousal level accounted for the variability in behavioral response speed. We found that the pupil size did not differ between the fast- and slow-response trials. EEG data analysis. After rejecting trials with blinking and EMG artifacts, we first averaged, for each condition, the ERPs that were time-locked to the onset of the for trials and to the corresponding time point for no trials. We then used EEGLAB 47, implemented on MatLab (Mathworks), for rereference of the EEG data to an average reference. EEGLAB was also used for statistical analysis and data visualization. After pre-processing, we examined the effect on EEG potentials for each condition by subtracting the ERPs on no trials from those on trials. The baseline of the ERP was corrected with reference to that obtained during the 2-ms period before the onset of. Although we used -compatible EEGs, -related artifacts were still observed for a short period after (o1 ms). We also observed artifacts around 2 21 ms after the, when the stimulator started to recharge its energystorage capacitor after each single-pulse of the. Click sounds associated with the -induced auditory evoked potentials around 8 ms after the 48. However, we primarily focused on the EEG data collected during the 2 4-msperiodaftertheandconsiderthisperiodtobefreeoftheseartifacts. In addition, as the main aim of our study was to compare the -induced ERP changes across conditions, -induced artifacts do not affect the analysis. We compared -induced ERPs between face and motion conditions for each of the long and short trials and examined the task-specific modulation of the pattern of neural impulse transmission. We then subtracted, for each of the face and motion conditions, the -induced ERPs on short trials from those on long trials and examined the components of the ERPs that were modulated by the preparation of specific attention tasks. To obtain the time course of the overall amount of electrical activity induced by, we calculated the global mean field amplitude by the formula sffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi P N i¼1 ðv i VÞ 2 ; N where N is the number of channels, V i is the voltage measured with channel i, and V is the mean of the measured voltages. This value has been shown to be associated with the arousal level of the subjects 28 Estimation of current source density. To estimate the cortical distribution of the current source density that accounts for the -induced ERP, we used LORETA 25. LORETA computes the three-dimensional distribution of neuronal generators of the observed electrical activity in a three-shell spherical head model. The head model includes scalp, skull and brain compartments. The brain compartment was restricted to the cortical gray matter of the standard brain of the MNI, which was resampled to 2,394 cubic voxels at a 7-mm resolution. To determine EEG electrode positions in the brain images, we used previously reported coordinates 49. In that study, the positions of scalp EEG electrodes were determined on the basis of the cartesian coordinates in the bestfitting sphere, and the position data were merged with the magnetic resonance imaging of the template brain. It was reported that the test-retest variability across subject was, on average, 4.8 mm. The LORETA functional images represent the electrical activity at each voxel as the squared magnitude (that is, power) of the computed current density. We obtained current source density at each time point for the grand-averaged -induced ERPs across subjects in each condition. We log-scaled and averaged the current source density during the 2 4-ms period after the. We then compared the current source density between the motion and face conditions on trials with long and identified the peak coordinate that showed the maximal difference in the current source density between the two conditions in the middle temporal and fusiform gyrus. Next, we calculated the current source densities for each subject from these peak coordinates on the long and short trials. The difference in the current source densities between the long and short trials was calculated for each region, task and subject, and we applied two-way ANOVA with repeated measures. We also compared the current source densities between task-relevant and task-irrelevant regions using two-tailed paired t tests. Finally, we calculated the difference in the current source densities between the fast- and slow-response trials for each of the long and short trial conditions in the motion and face tasks. We 9 VOLUME 12 [ NUMBER 1 [ JANUARY 29 NATURE NEUROSCIENCE
7 ARTICLES 29 Nature America, Inc. All rights reserved. used one-sample t test to examine whether the difference in the current source density between the fast- and slow-response trials was significantly larger than zero. -perturbation experiments. We used the same motion- and face-discrimination task for our -perturbation experiments as we used in the main -EEG experiments. In one experimental session, 12 trials for each of the four conditions (short- and long- conditions for each of the face and motion tasks) were given in a pseudo-random order with an intertrial interval of 2 3 s. Subjects performed the task for four sessions. On half of the trials, we delivered double-pulse using a figure eight shaped flat coil (Magstim Rapid) to the scalp position corresponding to the right FEF at 34 and 134 ms after the onset of a visual target. In this experiment, the was used to perturb the neural processing in the FEF. The effect is thought to be mediated by silencing task-related neurons as well as by stimulating neurons that are not related to the task. The order of the and no trials was randomized in an experimental session. The stimulus intensity was set at 65% of maximum output of the stimulator. The same subjects performed the task for another four sessions and a double-pulse of the same intensity was delivered to the vertex in half of the trials. The coil was placed tangentially over the vertex with the handle of the coil pointing backwards and aligned to the interhemispheric line. The did not elicit any saccadic eye movements for both the FEF- and vertex- sessions. For statistical analysis of behavioral data, we carried out three-way ANOVAs (motion or face task, or no and long or short ) separately for the FEF- and vertex- sessions. Note: Supplementary information is available on the Nature Neuroscience website. ACKNOWLEDGMENTS We thank M. Okamoto and N. Yamamoto for assistance with EEG recording. This work was supported by a Grant-in-Aid for Scientific Research (A) and a Grant-in-Aid for Young Scientists (S) from the Japan Society for the Promotion of Science, and a Grant-in-Aid from the Center of Excellence Program, Center for Brain Medical Science, from the Ministry of Education, Culture, Sports, Science and Technology, Japan. AUTHOR CONTRIBUTIONS Y.M. designed the task, conducted the experiments and analyzed the data. R.A., Y.Y., J.O. and K.T. contributed to the experiments and analysis. K.S. conceptualized the original idea for the study. Y.M. and K.S. wrote the paper. Published online at Reprints and permissions information is available online at reprintsandpermissions/ 1. Desimone, R. & Duncan, J. Neural mechanisms of selective visual attention. Annu. Rev. Neurosci. 18, (1995). 2. Kastner, S. & Ungerleider, L.G. Mechanisms of visual attention in the human cortex. Annu. Rev. Neurosci. 23, (2). 3. Miller, E.K. & Cohen, J.D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 24, (21). 4. Corbetta, M. & Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 3, (22). 5. Moran, J. & Desimone, R. Selective attention gates visual processing in the extrastriate cortex. Science 229, (1985). 6. Reynolds, J.H. & Chelazzi, L. Attentional modulation of visual processing. Annu. Rev. Neurosci. 27, (24). 7. Maunsell, J.H. & Treue, S. Feature-based attention in visual cortex. Trends Neurosci. 29, (26). 8. Brass, M., Ullsperger, M., Knoesche, T.R., von Cramon, D.Y. & Phillips, N.A. Who comes first? The role of the prefrontal and parietal cortex in cognitive control. J. Cogn. Neurosci. 17, (25). 9. Grent- t-jong, T. & Woldorff, M.G. Timing and sequence of brain activity in top-down control of visual-spatial attention. PLoS Biol. 5, e12 (27). 1. Buchel, C. & Friston, K.J. Modulation of connectivity in visual pathways by attention: cortical interactions evaluated with structural equation modeling and fmri. Cereb. Cortex 7, (1997). 11. Sakai, K. & Passingham, R.E. Prefrontal interactions reflect future task operations. Nat. Neurosci. 6, (23). 12. Sakai, K. & Passingham, R.E. Prefrontal set activity predicts rule-specific neural processing during subsequent cognitive performance. J. Neurosci. 26, (26). 13. Fuster, J.M., Bauer, R.H. & Jervey, J.P. Functional interactions between inferotemporal and prefrontal cortex in a cognitive task. Brain Res. 33, (1985). 14. Tomita, H., Ohbayashi, M., Nakahara, K., Hasegawa, I. & Miyashita, Y. Top-down signal from prefrontal cortex in executive control of memory retrieval. Nature 41, (1999). 15. Barcelo, F., Suwazono, S. & Knight, R.T. Prefrontal modulation of visual processing in humans. Nat. Neurosci. 3, (2). 16. Taylor, P.C., Nobre, A.C. & Rushworth, M.F. FEF affects visual cortical activity. Cereb. Cortex 17, (27). 17. Taylor, P.C., Nobre, A.C. & Rushworth, M.F. Subsecond changes in top down control exerted by human medial frontal cortex during conflict and action selection: a combined transcranial magnetic stimulation electroencephalography study. J. Neurosci. 27, (27). 18. Moore, T. & Armstrong, K.M. Selective gating of visual signals by microstimulation of frontal cortex. Nature 421, (23). 19. Armstrong, K.M. & Moore, T. Rapid enhancement of visual cortical response discriminability by microstimulation of the frontal eye field. Proc. Natl. Acad. Sci. USA 14, (27). 2. Silvanto, J., Lavie, N. & Walsh, V. Stimulation of the human frontal eye fields modulates sensitivity of extrastriate visual cortex. J. Neurophysiol. 96, (26). 21. Grosbras, M.H., Laird, A.R. & Paus, T. Cortical regions involved in eye movements, shifts of attention and gaze perception. Hum. Brain Mapp. 25, (25). 22. Ilmoniemi, R.J. et al. Neuronal responses to magnetic stimulation reveal cortical reactivity and connectivity. Neuroreport 8, (1997). 23. Pascual-Leone, A. & Walsh, V. Fast backprojections from the motion to the primary visual area necessary for visual awareness. Science 292, (21). 24. Matsumoto, R. et al. Functional connectivity in human cortical motor system: a corticocortical evoked potential study. Brain 13, (27). 25. Pascual-Marqui, R.D. et al. Low-resolution brain electromagnetic tomography (LORETA) functional imaging in acute, neuroleptic-naive, first-episode, productive schizophrenia. Psychiatry Res. 9, (1999). 26. Tootell, R.B. et al. Functional analysis of human MT and related visual cortical areas using magnetic resonance imaging. J. Neurosci. 15, (1995). 27. Kanwisher, N., McDermott, J. & Chun, M.M. The fusiform face area: a module in human extrastriate cortex specialized for face perception. J. Neurosci. 17, (1997). 28. Kahkonen, S. et al. Ethanol modulates cortical activity: direct evidence with combined and EEG. Neuroimage 14, (21). 29. Grosbras, M.H. & Paus, T. Transcranial magnetic stimulation of the human frontal eye field: effects on visual perception and attention. J. Cogn. Neurosci. 14, (22). 3. Grosbras, M.H. & Paus, T. Transcranial magnetic stimulation of the human frontal eye field facilitates visual awareness. Eur. J. Neurosci. 18, (23). 31. O Shea, J., Muggleton, N.G., Cowey, A. & Walsh, V. Timing of target discrimination in human frontal eye fields. J. Cogn. Neurosci. 16, (24). 32. O Shea, J., Muggleton, N.G., Cowey, A. & Walsh, V. Human frontal eye fields and spatial priming of pop-out. J. Cogn. Neurosci. 19, (27). 33. Bichot, N.P. & Schall, J.D. Effects of similarity and history on neural mechanisms of visual selection. Nat. Neurosci. 2, (1999). 34. Thompson, K.G. & Bichot, N.P. A visual salience map in the primate frontal eye field. Prog. Brain Res. 147, (25). 35. Peng, X., Sereno, M.E., Silva, A.K., Lehky, S.R. & Sereno, A.B. Shape selectivity in primate frontal eye field. J. Neurophysiol. 1, (28). 36. Stanton, G.B., Bruce, C.J. & Goldberg, M.E. Topography of projections to posterior cortical areas from the macaque frontal eye fields. J. Comp. Neurol. 353, (1995). 37. Egner, T. et al. Neural integration of top-down spatial and feature-based information in visual search. J. Neurosci. 28, (28). 38. Lakatos, P., Chen, C.M., O Connell, M.N., Mills, A. & Schroeder, C.E. Neuronal oscillations and multisensory interaction in primary auditory cortex. Neuron 53, (27). 39. Lakatos, P., Karmos, G., Mehta, A.D., Ulbert, I. & Schroeder, C.E. Entrainment of neuronal oscillations as a mechanism of attentional selection. Science 32, (28). 4. Horwitz, B. et al. Investigating the neural basis for functional and effective connectivity. Application to fmri. Phil. Trans. R. Soc. Lond. B 36, (25). 41. Fuggetta, G., Pavone, E.F., Walsh, V., Kiss, M. & Eimer, M. Cortico-cortical interactions in spatial attention: a combined ERP/ study. J. Neurophysiol. 95, (26). 42. Massimini, M. et al. Breakdown of cortical effective connectivity during sleep. Science 39, (25). 43. Khayat, P.S., Spekreijse, H. & Roelfsema, P.R. Attention lights up new object representations before the old ones fade away. J. Neurosci. 26, (26). 44. Roelfsema, P.R., Tolboom, M. & Khayat, P.S. Different processing phases for features, figures and selective attention in the primary visual cortex. Neuron 56, (27). 45. Bentin, S., Allison, T., Puce, A., Perez, E. & McCarthy, G. Electrophysiological studies of face perception in humans. J. Cogn. Neurosci. 8, (1996). 46. Bradley, M.M., Miccoli, L., Escrig, M.A. & Lang, P.J. The pupil as a measure of emotional arousal and autonomic activation. Psychophysiology 45, (28). 47. Delorme, A. & Makeig, S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 134, 9 21 (24). 48. Nikouline, V., Ruohonen, J. & Ilmoniemi, R.J. The role of the coil click in assessed with simultaneous EEG. Clin. Neurophysiol. 11, (1999). 49. Towle, V.L. et al. The spatial location of EEG electrodes: locating the best-fitting sphere relative to cortical anatomy. Electroencephalogr. Clin. Neurophysiol. 86, 1 6 (1993). NATURE NEUROSCIENCE VOLUME 12 [ NUMBER 1 [ JANUARY 29 91
8 Task-specific signal transmission from prefrontal cortex in visual selective attention Yosuke Morishima, Rei Akaishi, Yohei Yamada, Jiro Okuda, Keiichiro Toma, Katsuyuki Sakai Response time (ms) Long Short Response time (ms) Long Short no no Correct rate (%) Correct rate (%) no 5 no Supplementary Fig. 1 Behavioral results. Mean response time and correct rate are plotted for the long and short trials in the motion and face conditions. Error bars indicate s.e.m. Nature Neuroscience: doi:1.138/nn.2237
9 Aligned at Cue onset Aligned at onset 1 (µv) onset (µv) ) 6,, no,, no Long (ms) Target onset (ms) (µv) onset (µv) 6 1 P8 Short Target onset 5 1 (ms) (ms) 1 12 Supplementary Fig. 2 Single-condition ERP waveforms from the electrode P8 on the scalp separately shown for the long (upper panels) and short trials (bottom panels). On the left are shown the waveforms for the whole task epoch, aligned at the onset of the task cue. On the right are shown the waveforms for the 2 ms epoch from the onset, aligned at the onset. Nature Neuroscience: doi:1.138/nn.2237





 1.")












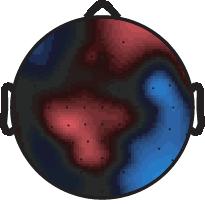


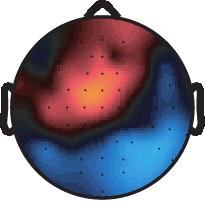











10 2~ ~2 2~4 EEG long short EEG long short Z score long short Z score long short 4~6 6~8 8~1 1~12 12~14 14~16 16~18 18~2 (ms) 1.5 (µv) Supplementary Fig. 3 Mean voltage and z-score scalp maps for the differential -induced ERPs between long and short trials as in Fig. 3, but are now shown for the time period from 2 to 2 ms after the. Top two rows show the difference in the mean voltage of -induced ERPs for the contrast of the long minus short trials. Bottom two rows show the z-scores for the contrast. The positive sign of the z-scores indicates more positive potential on the long compared to short trials. Nature Neuroscience: doi:1.138/nn.2237
Stimulation of the Frontal Eye Field Reveals Persistent Effective Connectivity after Controlled Behavior
 The Journal of Neuroscience, March 24, 2010 30(12):4295 4305 4295 Behavioral/Systems/Cognitive Stimulation of the Frontal Eye Field Reveals Persistent Effective Connectivity after Controlled Behavior Rei
The Journal of Neuroscience, March 24, 2010 30(12):4295 4305 4295 Behavioral/Systems/Cognitive Stimulation of the Frontal Eye Field Reveals Persistent Effective Connectivity after Controlled Behavior Rei
Attention: Neural Mechanisms and Attentional Control Networks Attention 2
 Attention: Neural Mechanisms and Attentional Control Networks Attention 2 Hillyard(1973) Dichotic Listening Task N1 component enhanced for attended stimuli Supports early selection Effects of Voluntary
Attention: Neural Mechanisms and Attentional Control Networks Attention 2 Hillyard(1973) Dichotic Listening Task N1 component enhanced for attended stimuli Supports early selection Effects of Voluntary
Supplemental Material
 Supplemental Material Recording technique Multi-unit activity (MUA) was recorded from electrodes that were chronically implanted (Teflon-coated platinum-iridium wires) in the primary visual cortex representing
Supplemental Material Recording technique Multi-unit activity (MUA) was recorded from electrodes that were chronically implanted (Teflon-coated platinum-iridium wires) in the primary visual cortex representing
Effect of Transcranial Magnetic Stimulation(TMS) on Visual Search Task
 on Visual Search Task") INTERNATIONAL JOURNAL OF APPLIED BIOMEDICAL ENGINEERING VOL.2, NO.2 2009 1 Effect of Transcranial Magnetic Stimulation(TMS) on Visual Search Task K. Iramina, Guest member ABSTRACT Transcranial magnetic
INTERNATIONAL JOURNAL OF APPLIED BIOMEDICAL ENGINEERING VOL.2, NO.2 2009 1 Effect of Transcranial Magnetic Stimulation(TMS) on Visual Search Task K. Iramina, Guest member ABSTRACT Transcranial magnetic
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Control of visuo-spatial attention. Emiliano Macaluso
 Control of visuo-spatial attention Emiliano Macaluso CB demo Attention Limited processing resources Overwhelming sensory input cannot be fully processed => SELECTIVE PROCESSING Selection via spatial orienting
Control of visuo-spatial attention Emiliano Macaluso CB demo Attention Limited processing resources Overwhelming sensory input cannot be fully processed => SELECTIVE PROCESSING Selection via spatial orienting
Introduction to TMS Transcranial Magnetic Stimulation
 Introduction to TMS Transcranial Magnetic Stimulation Lisa Koski, PhD, Clin Psy TMS Neurorehabilitation Lab Royal Victoria Hospital 2009-12-14 BIC Seminar, MNI Overview History, basic principles, instrumentation
Introduction to TMS Transcranial Magnetic Stimulation Lisa Koski, PhD, Clin Psy TMS Neurorehabilitation Lab Royal Victoria Hospital 2009-12-14 BIC Seminar, MNI Overview History, basic principles, instrumentation
Selective Attention. Modes of Control. Domains of Selection
 The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing
 Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
Extrastriate Visual Areas February 27, 2003 A. Roe
 Extrastriate Visual Areas February 27, 2003 A. Roe How many extrastriate areas are there? LOTS!!! Macaque monkey flattened cortex Why? How do we know this? Topography Functional properties Connections
Extrastriate Visual Areas February 27, 2003 A. Roe How many extrastriate areas are there? LOTS!!! Macaque monkey flattened cortex Why? How do we know this? Topography Functional properties Connections
FINAL PROGRESS REPORT
 (1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
(1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
Mental representation of number in different numerical forms
 Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Twelve right-handed subjects between the ages of 22 and 30 were recruited from the
 Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Transcranial direct current stimulation modulates shifts in global/local attention
 University of New Mexico UNM Digital Repository Psychology ETDs Electronic Theses and Dissertations 2-9-2010 Transcranial direct current stimulation modulates shifts in global/local attention David B.
University of New Mexico UNM Digital Repository Psychology ETDs Electronic Theses and Dissertations 2-9-2010 Transcranial direct current stimulation modulates shifts in global/local attention David B.
The Contribution of TMS-EEG Coregistration in the Exploration of the Human Connectome
 IRCCS San Giovanni di Dio Fatebenefratelli The Contribution of TMS-EEG Coregistration in the Exploration of the Human Connectome Marta Bortoletto Carlo Miniussi Cognitive Neuroscience Section, The Saint
IRCCS San Giovanni di Dio Fatebenefratelli The Contribution of TMS-EEG Coregistration in the Exploration of the Human Connectome Marta Bortoletto Carlo Miniussi Cognitive Neuroscience Section, The Saint
Report. Spatial Attention Can Be Allocated Rapidly and in Parallel to New Visual Objects. Martin Eimer 1, * and Anna Grubert 1 1
 Current Biology 24, 193 198, January 20, 2014 ª2014 The Authors http://dx.doi.org/10.1016/j.cub.2013.12.001 Spatial Attention Can Be Allocated Rapidly and in Parallel to New Visual Objects Report Martin
Current Biology 24, 193 198, January 20, 2014 ª2014 The Authors http://dx.doi.org/10.1016/j.cub.2013.12.001 Spatial Attention Can Be Allocated Rapidly and in Parallel to New Visual Objects Report Martin
Transcranial Magnetic Stimulation
 Transcranial Magnetic Stimulation Session 4 Virtual Lesion Approach I Alexandra Reichenbach MPI for Biological Cybernetics Tübingen, Germany Today s Schedule Virtual Lesion Approach : Study Design Rationale
Transcranial Magnetic Stimulation Session 4 Virtual Lesion Approach I Alexandra Reichenbach MPI for Biological Cybernetics Tübingen, Germany Today s Schedule Virtual Lesion Approach : Study Design Rationale
Sum of Neurally Distinct Stimulus- and Task-Related Components.
 SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
The neurolinguistic toolbox Jonathan R. Brennan. Introduction to Neurolinguistics, LSA2017 1
 The neurolinguistic toolbox Jonathan R. Brennan Introduction to Neurolinguistics, LSA2017 1 Psycholinguistics / Neurolinguistics Happy Hour!!! Tuesdays 7/11, 7/18, 7/25 5:30-6:30 PM @ the Boone Center
The neurolinguistic toolbox Jonathan R. Brennan Introduction to Neurolinguistics, LSA2017 1 Psycholinguistics / Neurolinguistics Happy Hour!!! Tuesdays 7/11, 7/18, 7/25 5:30-6:30 PM @ the Boone Center
Selective bias in temporal bisection task by number exposition
 Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
Manuscript under review for Psychological Science. Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory
 Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
Biomedical Research 2013; 24 (3): ISSN X
: ISSN X") Biomedical Research 2013; 24 (3): 359-364 ISSN 0970-938X http://www.biomedres.info Investigating relative strengths and positions of electrical activity in the left and right hemispheres of the human brain
Biomedical Research 2013; 24 (3): 359-364 ISSN 0970-938X http://www.biomedres.info Investigating relative strengths and positions of electrical activity in the left and right hemispheres of the human brain
Beyond Blind Averaging: Analyzing Event-Related Brain Dynamics. Scott Makeig. sccn.ucsd.edu
 Beyond Blind Averaging: Analyzing Event-Related Brain Dynamics Scott Makeig Institute for Neural Computation University of California San Diego La Jolla CA sccn.ucsd.edu Talk given at the EEG/MEG course
Beyond Blind Averaging: Analyzing Event-Related Brain Dynamics Scott Makeig Institute for Neural Computation University of California San Diego La Jolla CA sccn.ucsd.edu Talk given at the EEG/MEG course
Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials
 Chinese Science Bulletin 2007 Science in China Press Springer-Verlag Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials SONG Yan 1, PENG
Chinese Science Bulletin 2007 Science in China Press Springer-Verlag Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials SONG Yan 1, PENG
Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry
 International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
TMS of the right angular gyrus modulates priming of pop-out in visual search: combined TMS-ERP evidence
 J Neurophysiol 106: 3001 3009, 2011. First published August 31, 2011; doi:10.1152/jn.00121.2011. TMS of the right angular gyrus modulates priming of pop-out in visual search: combined TMS-ERP evidence
J Neurophysiol 106: 3001 3009, 2011. First published August 31, 2011; doi:10.1152/jn.00121.2011. TMS of the right angular gyrus modulates priming of pop-out in visual search: combined TMS-ERP evidence
Behavioral/Systems/Cognitive. June Hung, 1,2 Jon Driver, 2 and Vincent Walsh 2 1
 15904 The Journal of Neuroscience, November 2, 2011 31(44):15904 15913 Behavioral/Systems/Cognitive Visual Selection and the Human Frontal Eye Fields: Effects of Frontal Transcranial Magnetic Stimulation
15904 The Journal of Neuroscience, November 2, 2011 31(44):15904 15913 Behavioral/Systems/Cognitive Visual Selection and the Human Frontal Eye Fields: Effects of Frontal Transcranial Magnetic Stimulation
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED. Dennis L. Molfese University of Nebraska - Lincoln
 DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
Supporting Information
 Supporting Information Lingnau et al. 10.1073/pnas.0902262106 Fig. S1. Material presented during motor act observation (A) and execution (B). Each row shows one of the 8 different motor acts. Columns in
Supporting Information Lingnau et al. 10.1073/pnas.0902262106 Fig. S1. Material presented during motor act observation (A) and execution (B). Each row shows one of the 8 different motor acts. Columns in
From Single-trial EEG to Brain Area Dynamics
 From Single-trial EEG to Brain Area Dynamics a Delorme A., a Makeig, S., b Fabre-Thorpe, M., a Sejnowski, T. a The Salk Institute for Biological Studies, 10010 N. Torey Pines Road, La Jolla, CA92109, USA
From Single-trial EEG to Brain Area Dynamics a Delorme A., a Makeig, S., b Fabre-Thorpe, M., a Sejnowski, T. a The Salk Institute for Biological Studies, 10010 N. Torey Pines Road, La Jolla, CA92109, USA
Nature Methods: doi: /nmeth Supplementary Figure 1. Activity in turtle dorsal cortex is sparse.
 Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab
 TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab This report details my summer research project for the REU Theoretical and Computational Neuroscience program as
TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab This report details my summer research project for the REU Theoretical and Computational Neuroscience program as
Electrophysiological Indices of Target and Distractor Processing in Visual Search
 Electrophysiological Indices of Target and Distractor Processing in Visual Search Clayton Hickey 1,2, Vincent Di Lollo 2, and John J. McDonald 2 Abstract & Attentional selection of a target presented among
Electrophysiological Indices of Target and Distractor Processing in Visual Search Clayton Hickey 1,2, Vincent Di Lollo 2, and John J. McDonald 2 Abstract & Attentional selection of a target presented among
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B
 Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Neural Correlates of Human Cognitive Function:
 Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
The Central Nervous System
 The Central Nervous System Cellular Basis. Neural Communication. Major Structures. Principles & Methods. Principles of Neural Organization Big Question #1: Representation. How is the external world coded
The Central Nervous System Cellular Basis. Neural Communication. Major Structures. Principles & Methods. Principles of Neural Organization Big Question #1: Representation. How is the external world coded
Early posterior ERP components do not reflect the control of attentional shifts toward expected peripheral events
 Psychophysiology, 40 (2003), 827 831. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research BRIEF REPT Early posterior ERP components do not reflect the
Psychophysiology, 40 (2003), 827 831. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research BRIEF REPT Early posterior ERP components do not reflect the
CS/NEUR125 Brains, Minds, and Machines. Due: Friday, April 14
 CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
Visual activity in the human frontal eye eld
 Cognitive Neuroscience NeuroReport NeuroReport 10, 925±930 (1999) ALTHOUGH visual information processing in the monkey frontal eye eld (FEF) has been well demonstrated, the contribution of its human homologue
Cognitive Neuroscience NeuroReport NeuroReport 10, 925±930 (1999) ALTHOUGH visual information processing in the monkey frontal eye eld (FEF) has been well demonstrated, the contribution of its human homologue
Event-Related fmri and the Hemodynamic Response
 Human Brain Mapping 6:373 377(1998) Event-Related fmri and the Hemodynamic Response Randy L. Buckner 1,2,3 * 1 Departments of Psychology, Anatomy and Neurobiology, and Radiology, Washington University,
Human Brain Mapping 6:373 377(1998) Event-Related fmri and the Hemodynamic Response Randy L. Buckner 1,2,3 * 1 Departments of Psychology, Anatomy and Neurobiology, and Radiology, Washington University,
Outline of Talk. Introduction to EEG and Event Related Potentials. Key points. My path to EEG
 Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature14066 Supplementary discussion Gradual accumulation of evidence for or against different choices has been implicated in many types of decision-making, including value-based decisions
doi:10.1038/nature14066 Supplementary discussion Gradual accumulation of evidence for or against different choices has been implicated in many types of decision-making, including value-based decisions
L'application de la TMS dans l'étude des fonctions cognitives
 L'application de la TMS dans l'étude des fonctions cognitives Chotiga Pattamadilok Laboratoire Parole et Langage Aix-en-Provence RMN- Oct. 2016 Magnetic stimulation Silvanus P. Thompson: on inserting the
L'application de la TMS dans l'étude des fonctions cognitives Chotiga Pattamadilok Laboratoire Parole et Langage Aix-en-Provence RMN- Oct. 2016 Magnetic stimulation Silvanus P. Thompson: on inserting the
Selective Attention to Face Identity and Color Studied With fmri
 Human Brain Mapping 5:293 297(1997) Selective Attention to Face Identity and Color Studied With fmri Vincent P. Clark, 1 * Raja Parasuraman, 2 Katrina Keil, 1 Rachel Kulansky, 1 Sean Fannon, 2 Jose Ma.
Human Brain Mapping 5:293 297(1997) Selective Attention to Face Identity and Color Studied With fmri Vincent P. Clark, 1 * Raja Parasuraman, 2 Katrina Keil, 1 Rachel Kulansky, 1 Sean Fannon, 2 Jose Ma.
The Tools: Imaging the Living Brain
 The Tools: Imaging the Living Brain I believe the study of neuroimaging has supported the localization of mental operations within the human brain. -Michael I. Posner, 2003 Neuroimaging methods Since Descarte
The Tools: Imaging the Living Brain I believe the study of neuroimaging has supported the localization of mental operations within the human brain. -Michael I. Posner, 2003 Neuroimaging methods Since Descarte
Supplementary Note Psychophysics:
 Supplementary Note More detailed description of MM s subjective experiences can be found on Mike May s Perceptions Home Page, http://www.senderogroup.com/perception.htm Psychophysics: The spatial CSF was
Supplementary Note More detailed description of MM s subjective experiences can be found on Mike May s Perceptions Home Page, http://www.senderogroup.com/perception.htm Psychophysics: The spatial CSF was
Event-Related Potentials Recorded during Human-Computer Interaction
 Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
ABSTRACT 1. INTRODUCTION 2. ARTIFACT REJECTION ON RAW DATA
 AUTOMATIC ARTIFACT REJECTION FOR EEG DATA USING HIGH-ORDER STATISTICS AND INDEPENDENT COMPONENT ANALYSIS A. Delorme, S. Makeig, T. Sejnowski CNL, Salk Institute 11 N. Torrey Pines Road La Jolla, CA 917,
AUTOMATIC ARTIFACT REJECTION FOR EEG DATA USING HIGH-ORDER STATISTICS AND INDEPENDENT COMPONENT ANALYSIS A. Delorme, S. Makeig, T. Sejnowski CNL, Salk Institute 11 N. Torrey Pines Road La Jolla, CA 917,
Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory
 Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
Attentional Control 1. Identifying the neural systems of top-down attentional control: A meta-analytic approach
 Attentional Control 1 Identifying the neural systems of top-down attentional control: A meta-analytic approach Barry Giesbrecht & George R. Mangun Center for Mind & Brain University of California, Davis
Attentional Control 1 Identifying the neural systems of top-down attentional control: A meta-analytic approach Barry Giesbrecht & George R. Mangun Center for Mind & Brain University of California, Davis
Preparatory attention in visual cortex
 Ann. N.Y. Acad. Sci. ISSN 0077-8923 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Issue: The Year in Cognitive Neuroscience REVIEW ARTICLE Preparatory attention in visual cortex Elisa Battistoni, 1 Timo Stein,
Ann. N.Y. Acad. Sci. ISSN 0077-8923 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Issue: The Year in Cognitive Neuroscience REVIEW ARTICLE Preparatory attention in visual cortex Elisa Battistoni, 1 Timo Stein,
The Role of Working Memory in Visual Selective Attention
 Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Oscillations: From Neuron to MEG
 Oscillations: From Neuron to MEG Educational Symposium, MEG UK 2014, Nottingham, Jan 8th 2014 Krish Singh CUBRIC, School of Psychology Cardiff University What are we trying to achieve? Bridge the gap from
Oscillations: From Neuron to MEG Educational Symposium, MEG UK 2014, Nottingham, Jan 8th 2014 Krish Singh CUBRIC, School of Psychology Cardiff University What are we trying to achieve? Bridge the gap from
The Journal of Neuroscience For Peer Review Only
 Page 1 of 31 Section: Behavioral System Cognitive Senior Editor: John Maunsell Number of figures and tables: 3 Figures and 4 Supplementary items Number of pages: 25 Title: Different Effects of Voluntary
Page 1 of 31 Section: Behavioral System Cognitive Senior Editor: John Maunsell Number of figures and tables: 3 Figures and 4 Supplementary items Number of pages: 25 Title: Different Effects of Voluntary
Decoding a Perceptual Decision Process across Cortex
 Article Decoding a Perceptual Decision Process across Cortex Adrián Hernández, 1 Verónica Nácher, 1 Rogelio Luna, 1 Antonio Zainos, 1 Luis Lemus, 1 Manuel Alvarez, 1 Yuriria Vázquez, 1 Liliana Camarillo,
Article Decoding a Perceptual Decision Process across Cortex Adrián Hernández, 1 Verónica Nácher, 1 Rogelio Luna, 1 Antonio Zainos, 1 Luis Lemus, 1 Manuel Alvarez, 1 Yuriria Vázquez, 1 Liliana Camarillo,
Nature Medicine: doi: /nm.4084
 Supplementary Figure 1: Sample IEDs. (a) Sample hippocampal IEDs from different kindled rats (scale bar = 200 µv, 100 ms). (b) Sample temporal lobe IEDs from different subjects with epilepsy (scale bar
Supplementary Figure 1: Sample IEDs. (a) Sample hippocampal IEDs from different kindled rats (scale bar = 200 µv, 100 ms). (b) Sample temporal lobe IEDs from different subjects with epilepsy (scale bar
Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling
 Dipole modeling") Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Mental Representation of Number in Different Numerical Forms
 Current Biology, Vol. 13, 2045 2050, December 2, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/j.cub.2003.11.023 Mental Representation of Number in Different Numerical Forms Anna Plodowski,
Current Biology, Vol. 13, 2045 2050, December 2, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/j.cub.2003.11.023 Mental Representation of Number in Different Numerical Forms Anna Plodowski,
fmri (functional MRI)
") Lesion fmri (functional MRI) Electroencephalogram (EEG) Brainstem CT (computed tomography) Scan Medulla PET (positron emission tomography) Scan Reticular Formation MRI (magnetic resonance imaging) Thalamus
Lesion fmri (functional MRI) Electroencephalogram (EEG) Brainstem CT (computed tomography) Scan Medulla PET (positron emission tomography) Scan Reticular Formation MRI (magnetic resonance imaging) Thalamus
The neural systems that mediate human perceptual decision making
 The neural systems that mediate human perceptual decision making Hauke R. Heekeren*, Sean Marrett and Leslie G. Ungerleider Abstract Perceptual decision making is the act of choosing one option or course
The neural systems that mediate human perceptual decision making Hauke R. Heekeren*, Sean Marrett and Leslie G. Ungerleider Abstract Perceptual decision making is the act of choosing one option or course
ERP Measures of Multiple Attention Deficits Following Prefrontal Damage
 INO056 10/18/04 6:34 PM Page 339 CHAPTER 56 ERP Measures of Multiple Attention Deficits Following Prefrontal Damage Leon Y. Deouell and Robert T. Knight ABSTRACT Maintaining a goal-directed behavior requires
INO056 10/18/04 6:34 PM Page 339 CHAPTER 56 ERP Measures of Multiple Attention Deficits Following Prefrontal Damage Leon Y. Deouell and Robert T. Knight ABSTRACT Maintaining a goal-directed behavior requires
Active suppression after involuntary capture of attention
 Psychon Bull Rev (2013) 20:296 301 DOI 10.3758/s13423-012-0353-4 BRIEF REPORT Active suppression after involuntary capture of attention Risa Sawaki & Steven J. Luck Published online: 20 December 2012 #
Psychon Bull Rev (2013) 20:296 301 DOI 10.3758/s13423-012-0353-4 BRIEF REPORT Active suppression after involuntary capture of attention Risa Sawaki & Steven J. Luck Published online: 20 December 2012 #
Timing and Sequence of Brain Activity in Top-Down Control of Visual-Spatial Attention
 Timing and Sequence of Brain Activity in Top-Down Control of Visual-Spatial Attention Tineke Grent- t-jong 1,2, Marty G. Woldorff 1,3* PLoS BIOLOGY 1 Center for Cognitive Neuroscience, Duke University,
Timing and Sequence of Brain Activity in Top-Down Control of Visual-Spatial Attention Tineke Grent- t-jong 1,2, Marty G. Woldorff 1,3* PLoS BIOLOGY 1 Center for Cognitive Neuroscience, Duke University,
Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures
 Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings
 Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings J. Besle, P. Lakatos, C.A. Schevon, R.R. Goodman, G.M. McKhann, A. Mehta, R.G. Emerson, C.E.
Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings J. Besle, P. Lakatos, C.A. Schevon, R.R. Goodman, G.M. McKhann, A. Mehta, R.G. Emerson, C.E.
Retinotopy & Phase Mapping
 Retinotopy & Phase Mapping Fani Deligianni B. A. Wandell, et al. Visual Field Maps in Human Cortex, Neuron, 56(2):366-383, 2007 Retinotopy Visual Cortex organised in visual field maps: Nearby neurons have
Retinotopy & Phase Mapping Fani Deligianni B. A. Wandell, et al. Visual Field Maps in Human Cortex, Neuron, 56(2):366-383, 2007 Retinotopy Visual Cortex organised in visual field maps: Nearby neurons have
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 11: Attention & Decision making Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis
Introduction to Computational Neuroscience Lecture 11: Attention & Decision making Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis
 or osteoarthritis") Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Introduction to Physiological Psychology
 Introduction to Physiological Psychology Review Kim Sweeney ksweeney@cogsci.ucsd.edu www.cogsci.ucsd.edu/~ksweeney/psy260.html Today n Discuss Final Paper Proposal (due 3/10) n General Review 1 The article
Introduction to Physiological Psychology Review Kim Sweeney ksweeney@cogsci.ucsd.edu www.cogsci.ucsd.edu/~ksweeney/psy260.html Today n Discuss Final Paper Proposal (due 3/10) n General Review 1 The article
How does the ventral pathway contribute to spatial attention and the planning of eye movements?
 R. P. Würtz and M. Lappe (Eds.) Dynamic Perception, Infix Verlag, St. Augustin, 83-88, 2002. How does the ventral pathway contribute to spatial attention and the planning of eye movements? Fred H. Hamker
R. P. Würtz and M. Lappe (Eds.) Dynamic Perception, Infix Verlag, St. Augustin, 83-88, 2002. How does the ventral pathway contribute to spatial attention and the planning of eye movements? Fred H. Hamker
STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM
 STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM STRUCTURAL ORGANIZATION OF THE BRAIN The central nervous system (CNS), consisting of the brain and spinal cord, receives input from sensory neurons and directs
STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM STRUCTURAL ORGANIZATION OF THE BRAIN The central nervous system (CNS), consisting of the brain and spinal cord, receives input from sensory neurons and directs
Myers Psychology for AP*
 Myers Psychology for AP* David G. Myers PowerPoint Presentation Slides by Kent Korek Germantown High School Worth Publishers, 2010 *AP is a trademark registered and/or owned by the College Board, which
Myers Psychology for AP* David G. Myers PowerPoint Presentation Slides by Kent Korek Germantown High School Worth Publishers, 2010 *AP is a trademark registered and/or owned by the College Board, which
Combining microstimulation and fmri in an awake behaving monkey
 NEAAPM Peter Neurath Symposium Combining microstimulation and fmri in an awake behaving monkey Presented by: Leeland B. Ekstrom April 2006 Advisors: Wim Vanduffel & Bruce Rosen MGH / MIT / HMS Martinos
NEAAPM Peter Neurath Symposium Combining microstimulation and fmri in an awake behaving monkey Presented by: Leeland B. Ekstrom April 2006 Advisors: Wim Vanduffel & Bruce Rosen MGH / MIT / HMS Martinos
Practicalities of EEG Measurement and Experiment Design YATING LIU
 Practicalities of EEG Measurement and Experiment Design YATING LIU 2014.02.04 Content Designing Experiments: Discuss & Pilot Event Markers Intra- and Intertrial Timing How Many Trials You Will Need How
Practicalities of EEG Measurement and Experiment Design YATING LIU 2014.02.04 Content Designing Experiments: Discuss & Pilot Event Markers Intra- and Intertrial Timing How Many Trials You Will Need How
Introduction to Electrophysiology
 Introduction to Electrophysiology Dr. Kwangyeol Baek Martinos Center for Biomedical Imaging Massachusetts General Hospital Harvard Medical School 2018-05-31s Contents Principles in Electrophysiology Techniques
Introduction to Electrophysiology Dr. Kwangyeol Baek Martinos Center for Biomedical Imaging Massachusetts General Hospital Harvard Medical School 2018-05-31s Contents Principles in Electrophysiology Techniques
Cognitive Neuroscience Attention
 Cognitive Neuroscience Attention There are many aspects to attention. It can be controlled. It can be focused on a particular sensory modality or item. It can be divided. It can set a perceptual system.
Cognitive Neuroscience Attention There are many aspects to attention. It can be controlled. It can be focused on a particular sensory modality or item. It can be divided. It can set a perceptual system.
The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction
 The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction FORUM ON NEUROSCIENCE AND NERVOUS SYSTEM DISORDERS Board on Health Sciences Policy Glutamate-related
The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction FORUM ON NEUROSCIENCE AND NERVOUS SYSTEM DISORDERS Board on Health Sciences Policy Glutamate-related
Reward prediction error signals associated with a modified time estimation task
 Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
Time Course of Attention Reveals Different Mechanisms for Spatial and Feature-Based Attention in Area V4
 Neuron, Vol. 47, 637 643, September 1, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.07.020 Time Course of Attention Reveals Different Mechanisms for Spatial and Feature-Based Attention
Neuron, Vol. 47, 637 643, September 1, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.07.020 Time Course of Attention Reveals Different Mechanisms for Spatial and Feature-Based Attention
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
Using EEG to Explore How rtms Produces Its Effects on Behavior
 Brain Topogr (21) 22:281 293 DOI 1.17/s1548-9-118-1 ORIGINAL PAPER Using EEG to Explore How rtms Produces Its Effects on Behavior Jeffrey S. Johnson Massihullah Hamidi Bradley R. Postle Received: 16 June
Brain Topogr (21) 22:281 293 DOI 1.17/s1548-9-118-1 ORIGINAL PAPER Using EEG to Explore How rtms Produces Its Effects on Behavior Jeffrey S. Johnson Massihullah Hamidi Bradley R. Postle Received: 16 June
Neuroscience Tutorial
 Neuroscience Tutorial Brain Organization : cortex, basal ganglia, limbic lobe : thalamus, hypothal., pituitary gland : medulla oblongata, midbrain, pons, cerebellum Cortical Organization Cortical Organization
Neuroscience Tutorial Brain Organization : cortex, basal ganglia, limbic lobe : thalamus, hypothal., pituitary gland : medulla oblongata, midbrain, pons, cerebellum Cortical Organization Cortical Organization
European Journal of Neuroscience. Functional specificity of premotor-motor cortical interactions during action selection
 Functional specificity of premotor-motor cortical interactions during action selection Journal: Manuscript ID: Manuscript Type: Date Submitted by the Author: EJN-2007-06-12587.R1 Research Report n/a Complete
Functional specificity of premotor-motor cortical interactions during action selection Journal: Manuscript ID: Manuscript Type: Date Submitted by the Author: EJN-2007-06-12587.R1 Research Report n/a Complete
Dynamic Changes in Phase-Amplitude Coupling Facilitate Spatial Attention Control in Fronto-Parietal Cortex
 Dynamic Changes in Phase-Amplitude Coupling Facilitate Spatial Attention Control in Fronto-Parietal Cortex Sara M. Szczepanski 1,2 *, Nathan E. Crone 3, Rachel A. Kuperman 4, Kurtis I. Auguste 5,6, Josef
Dynamic Changes in Phase-Amplitude Coupling Facilitate Spatial Attention Control in Fronto-Parietal Cortex Sara M. Szczepanski 1,2 *, Nathan E. Crone 3, Rachel A. Kuperman 4, Kurtis I. Auguste 5,6, Josef
The good, the bad, and the ugly of top-down executive control. Taraz Geola Lee. A dissertation submitted in partial satisfaction of the
 The good, the bad, and the ugly of top-down executive control By Taraz Geola Lee A dissertation submitted in partial satisfaction of the Requirements for the degree of Doctor of Philosophy in Psychology
The good, the bad, and the ugly of top-down executive control By Taraz Geola Lee A dissertation submitted in partial satisfaction of the Requirements for the degree of Doctor of Philosophy in Psychology
Supplemental Material
 1 Supplemental Material Golomb, J.D, and Kanwisher, N. (2012). Higher-level visual cortex represents retinotopic, not spatiotopic, object location. Cerebral Cortex. Contents: - Supplemental Figures S1-S3
1 Supplemental Material Golomb, J.D, and Kanwisher, N. (2012). Higher-level visual cortex represents retinotopic, not spatiotopic, object location. Cerebral Cortex. Contents: - Supplemental Figures S1-S3
CROSSMODAL PLASTICITY IN SPECIFIC AUDITORY CORTICES UNDERLIES VISUAL COMPENSATIONS IN THE DEAF "
 Supplementary Online Materials To complement: CROSSMODAL PLASTICITY IN SPECIFIC AUDITORY CORTICES UNDERLIES VISUAL COMPENSATIONS IN THE DEAF " Stephen G. Lomber, M. Alex Meredith, and Andrej Kral 1 Supplementary
Supplementary Online Materials To complement: CROSSMODAL PLASTICITY IN SPECIFIC AUDITORY CORTICES UNDERLIES VISUAL COMPENSATIONS IN THE DEAF " Stephen G. Lomber, M. Alex Meredith, and Andrej Kral 1 Supplementary
ERP Studies of Selective Attention to Nonspatial Features
 CHAPTER 82 ERP Studies of Selective Attention to Nonspatial Features Alice Mado Proverbio and Alberto Zani ABSTRACT This paper concentrates on electrophysiological data concerning selective attention to
CHAPTER 82 ERP Studies of Selective Attention to Nonspatial Features Alice Mado Proverbio and Alberto Zani ABSTRACT This paper concentrates on electrophysiological data concerning selective attention to
Journal of Experimental Psychology: Human Perception and Performance
 Journal of Experimental Psychology: Human Perception and Performance Item and Category-Based Attentional Control During Search for Real-World Objects: Can You Find the Pants Among the Pans? Rebecca Nako,
Journal of Experimental Psychology: Human Perception and Performance Item and Category-Based Attentional Control During Search for Real-World Objects: Can You Find the Pants Among the Pans? Rebecca Nako,
Brain and Cognition. Cognitive Neuroscience. If the brain were simple enough to understand, we would be too stupid to understand it
 Brain and Cognition Cognitive Neuroscience If the brain were simple enough to understand, we would be too stupid to understand it 1 The Chemical Synapse 2 Chemical Neurotransmission At rest, the synapse
Brain and Cognition Cognitive Neuroscience If the brain were simple enough to understand, we would be too stupid to understand it 1 The Chemical Synapse 2 Chemical Neurotransmission At rest, the synapse
Submitted report on Sufi recordings at AAPB 2013 in Portland. Not for general distribution. Thomas F. Collura, Ph.D. July, 2013
 Submitted report on Sufi recordings at AAPB 2013 in Portland Not for general distribution. Thomas F. Collura, Ph.D. July, 2013 Summary of EEG findings The intent of the EEG monitoring was to see which
Submitted report on Sufi recordings at AAPB 2013 in Portland Not for general distribution. Thomas F. Collura, Ph.D. July, 2013 Summary of EEG findings The intent of the EEG monitoring was to see which
Organization of the nervous system. The withdrawal reflex. The central nervous system. Structure of a neuron. Overview
 Overview The nervous system- central and peripheral The brain: The source of mind and self Neurons Neuron Communication Chemical messengers Inside the brain Parts of the brain Split Brain Patients Organization
Overview The nervous system- central and peripheral The brain: The source of mind and self Neurons Neuron Communication Chemical messengers Inside the brain Parts of the brain Split Brain Patients Organization
Combining tdcs and fmri. OHMB Teaching Course, Hamburg June 8, Andrea Antal
 Andrea Antal Department of Clinical Neurophysiology Georg-August University Goettingen Combining tdcs and fmri OHMB Teaching Course, Hamburg June 8, 2014 Classical Biomarkers for measuring human neuroplasticity
Andrea Antal Department of Clinical Neurophysiology Georg-August University Goettingen Combining tdcs and fmri OHMB Teaching Course, Hamburg June 8, 2014 Classical Biomarkers for measuring human neuroplasticity
Behavioural Brain Research
 Behavioural Brain Research 191 (2008) 141 147 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Combining TMS and EEG to study cognitive
Behavioural Brain Research 191 (2008) 141 147 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Combining TMS and EEG to study cognitive
Seeing through the tongue: cross-modal plasticity in the congenitally blind
 International Congress Series 1270 (2004) 79 84 Seeing through the tongue: cross-modal plasticity in the congenitally blind Ron Kupers a, *, Maurice Ptito b www.ics-elsevier.com a Center for Functionally
International Congress Series 1270 (2004) 79 84 Seeing through the tongue: cross-modal plasticity in the congenitally blind Ron Kupers a, *, Maurice Ptito b www.ics-elsevier.com a Center for Functionally
Report. Stimulation of Category-Selective Brain Areas Modulates ERP to Their Preferred Categories
 Current Biology 21, 1894 1899, November 22, 2011 ª2011 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2011.09.030 Stimulation of Category-Selective Brain Areas Modulates ERP to Their Preferred Categories
Current Biology 21, 1894 1899, November 22, 2011 ª2011 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2011.09.030 Stimulation of Category-Selective Brain Areas Modulates ERP to Their Preferred Categories
The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition
 The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition Chair: Jason Samaha, University of Wisconsin-Madison Co-Chair: Ali Mazaheri, University of Birmingham
The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition Chair: Jason Samaha, University of Wisconsin-Madison Co-Chair: Ali Mazaheri, University of Birmingham