The Role of E2F Transcription Factors in Oncogene-Mediated. Mammary Tumorigenesis
|
|
|
- Hubert Russell
- 5 years ago
- Views:
Transcription
1 The Role of E2F Transcription Factors in Oncogene-Mediated Mammary Tumorigenesis A Senior Honors Thesis Presented in partial fulfillment of the requirements for graduation with distinction in Molecular Genetics in the College of Biological Sciences of The Ohio State University By Melissa House The Ohio State University June 2007
2 Abstract The Myc and Neu oncogenes are commonly amplified in breast cancer, and in order to determine their roles, one must study the pathways they regulate. It has been found that there may be a cross-regulatory relationship between Myc and Neu through the action of E2F transcription factors. Establishing and understanding any potential link between the Myc and Neu oncogenes would enhance our understanding of gene regulatory pathways in breast cancer, and possibly have clinical implications. Based largely on in-vitro studies, Myc had been previously suggested to act through a select number of E2F family members. Similarly, studies have also linked Neu to the action of the E2F pathway. In order to study the relevance of E2F transcription factors in these two pathways, I examined mammary tumor formation initiated by either the Myc and Neu oncogenes in mice deficient for the E2F1, E2F2, or E2F3 genes. Because there exists a high degree of functional redundancy, combinations of the E2Fs were also deleted. Development of glands deficient in the E2Fs as well as tumor start dates were examined in order to determine the contributions made by the specific E2Fs. It was found that E2F3 did not play a significant role in Myc-mediated tumorigenesis, but it did play a critical role in Neu-mediated tumorigenesis. E2Fs 1 and 2 had varying roles of importance between the two pathways, and even when mice were deficient for multiple E2Fs, tumorigenesis was able to occur. The data implies that the Rb-E2F pathway is only mildly involved in Mycmediated mammary tumorigenesis, but that this pathway is more critical for Neu mediated tumorigenesis. This suggests that Myc most likely acts through several additional regulatory pathways, and Neu is dependent on the Cyclin D1-Rb E2F pathway. 1
3 Introduction Control mechanisms of the cell cycle affect many aspects of development. Whether it is determining when a cell should proliferate, commit to apoptosis, or remain quiescent, many of the decisions depend on a balance of regulators of the cell cycle. The cell cycle is considered to involve an ordered series of events that occur during each cell division, and that allows cells to proliferate appropriately. It includes the G1 phase, or initial growth phase of the cellular machinery, the S phase, or DNA replication phase, the G2, or second growth phase, and M phase, or mitotic phase. During these different stages of the cycle, several checkpoints exist to ensure the cell division proceeds under the appropriate set of signals and results in two genetically identical daughter cells. During G1, a cell can also leave to a state called G0, where it is no longer growing, but instead is in a quiescent state. Cell external signals and cell internal information in combination determine whether a cell enters the division cycle. External signals usually only affect the decision until the cell is committed, or at the Restriction point (van den Heuval, 2005). Progression thereafter is controlled only by internal cell cycle machinery. Rb and E2F transcription factors are the central components of this restriction control mechanism. It is imperative to know the details of these checkpoints and inducers of cell growth because deregulation of the cell cycle can lead to damaged, mutated, and eventually cancerous cells that may continue to proliferate un-checked. Many genes have been found to be involved in cell division regulation, including Rb. The Rb gene product can act as a transcriptional repressor of genes involved in S phase progression by associating with E2F transcription factors. It has primarily been found to associate with E2F activators in order to prevent them promoting 2
4 transcription, or can be recruited along with other associated chromatin remodeling complexes and repressive E2Fs to inhibit transcription (Stevaux & Dyson, 2002). When Rb is deleted, the E2F activators are de-regulated, leading to aberrant cellular proliferation and eventually to tumor formation. Rb itself is regulated by different upstream regulators, such as Cyclin D1, which phosphorylates and inactivates Rb from being able to associate with E2Fs (Nevins 2001). Only the underphosphorylated form of Rb is functional in E2F binding. The Cyclin D1-Rb-E2F pathway is under intense study because the Cyclin D1 gene is amplified in 20% of human breast cancers (Dickson 1995), and the Cyclin D1 protein is over-expressed in over 50% of human mammary carcinomas (Bartkova et al. 1994). There have been eight E2F transcription factors found thus far. Evidence exists to indicate that each E2F factors may have specific functions. E2F factors appear to be key components in a cell cycle checkpoint that determines whether a cell will arrest in G1 or enter S phase. A direct correlation has been made between the ability of Rb to bind E2F and to arrest cells in the G1 phase of the cell cycle. E2Fs 1, 2, and 3 are thought to be the transcriptional activators, so it implies that when Rb is phosphorylated, it leaves these three transcription factors free to activate expression of important transcriptional machinery and avoid G1 arrest (Wu et al. 2001). These three E2Fs have a very closely related structure, and can associate exclusively with Rb, while other E2Fs can associate with the other two Rb-related gene products, the p107 and p130 pocket proteins (Johnson & Schneider-Broussard, 1998). E2Fs 1-3 are found almost exclusively in the nucleus because they have a nuclear localization signal on their amino termini. E2F1, E2F2, and E2F3 have been shown to induce serum starved REF52 cells to enter S phase (DeGregori 3
5 et al., 1997). E2Fs 4 and 5 cannot induce S phase under these conditions, and this is attributed to the fact that they lack nuclear localization signals. E2F1 expression also has the ability to overcome G1 phase block induced by Rb (Zhu et al.1993), TGF-beta (Schwarz et al. 1995), and gamma irradiation (DeGregori et al. 1995), and overexpression of E2F1 has enabled cells to overcome inhibitory activities of several tumor suppressor proteins. Further evidence that E2F plays a critical role in G1-S phase transition comes from the fact that a DP1 dominant negative mutant, which forms E2F heterodimers inactive in DNA binding, causes cells to arrest in G1 (Wu et al. 1996). On the contrary, over-expression of E2F1 has also been shown to induce cells to undergo apoptosis via causing accumulation of the tumor suppressor p53 (Qin et al. 1994). This data alone goes to show that E2F transcription factors also have their own unique functions outside of possible redundancy. Because the Cyclin D1-Rb-E2F pathway plays a critical role in the control of the cell cycle and under normal conditions, it is important to investigate the effects it may have when it is deregulated during cancer progression. Cyclin D1 is one of the most commonly overexpressed oncogenes in breast cancer, in as many as 50% of the cases. Without Cyclin D1 the mammary gland cannot develop normally, and with overexpression of Cyclin D1 benign mouse mammary epithelial cells can transform to become malignant (Wang et al. 2007). Another commonly over-expressed gene in breast cancer is the c-myc proto-oncogene. Loss of the gene results in embryonic death, thus it is also imperative to cell growth. Evidence has linked c-myc and Neu to the regulation of Cyclin D1, where it has been shown that over-expression of c-myc and Neu causes an up-regulation of Cyclin D1. In turn, Cyclin D1 function is linked to the control of E2F. 4
6 Cyclin D1-cdk complexes can phosphorylate the Rb tumor suppressor, and in its hyperphosphorylated state, Rb can no longer bind to E2F factors. The aim of this project is to evaluate the possible link between Myc-Cyclin D1 and Neu-Cyclin D1 pathways to the Cyclin D1-Rb-E2F pathway (Figure 1), and to examine how exactly these oncogenes mediate tumorigenesis. In order to understand the possible links in tumorigenesis, I have evaluated Myc-mediated and Neu-mediated mammary tumorigenesis in mice that are deficient in various E2Fs. Because E2Fs may also have an important role during normal proliferation, I have also evaluated the role of E2Fs during mammary development. By examining these two aspects-tumorigenesis and normal development-the role of the E2Fs in the control of proliferation could be compared under normal or cancer conditions. We used two mouse models of breast cancer to evaluate the role of E2Fs in tumorigenesis for several reasons. First, many of the events in the step-wise progression to human cancer can be recapitulated in mice. Second, the mice genome can be efficiently and specifically altered by experimentation. Third, the Myc and Neu oncogenes are highly relevant because they are amplified in a high proportion of human breast cancers. Because over-expression of specific genes in the entire mouse may have undesirable consequences on mouse physiology, we used a system to localize the expression of genes or deletion of genes to the mammary gland epithelium. To accomplish this, we used the mouse mammary tumor virus (MMTV-rtTa) promoter to drive the expression of the tetracycline activator. The MMTV promoter can be used to target expression of genes to the mammary epithelium because the tetracycline activator is only transcribed in the mammary tissue. In the presence of doxycycline, the 5
7 tetracycline activator it encodes can bind to a tetracycline responsive promoter (tet-on), thereby activating specific gene expression (Figure 2). In this system, the Myc transgene contains the human Myc cdna under the control of the tetracycline-responsive minimal promoter (Teto-Myc). The tetracycline activator then binds to the tet-on promoter only in the presence of doxycycline, and fires off transcription of c-myc only in the mammary epithelium. As indicated in the figure, without doxycycline, the activator cannot bind to the tet-on promoter. At the time of the experiment, a Teto-Neu transgene was unavailable, therefore the Neu mice were bred to have MMTV-c-Neu. This allows for the Neu gene to be under the direct control of the MMTV promoter, and consequently also upregulated only in the mammary gland. Because the mice were able to survive with no real defects from the deficiency of the transcription factors, E2Fs 1 and 2 were able to be conventionally deleted in the entire mouse. The alleles for each E2Fs 1 and 2 were mutant/mutant in every cell. On the other hand, loss of E2F3 causes embryonic lethality, so it was necessary for it to be conditionally deleted only in the mammary gland. To circumvent this problem, the mice were bred to have E2F3 floxed alleles. These floxed alleles are flanked by loxp sites on either end of exon 3 of the E2F3 gene. Cre recombinase can cut at these loxp sites, excising the exon and rendering the product inactive. For that reason, the Teto-Cre transgene was created to be under the control of the MMTV-rtTa encoded tetracycline activator and doxycycline also, encoding Cre recombinase only in the mammary gland. As a result, Cre mediated ablation of E2F3 will occur exclusively in the mammary gland. Because of the possibility of functional redundancy, the E2Fs were also deleted in combination to investigate potential phenotypes. 6
8 Materials and Methods Generation of the Transgenic Mice Because it was necessary to have the oncogenes and cre recombinase only active in the mammary gland, tissue specificity was created by incorporating the mouse mammary tumor virus (MMTV-rtTa). The mice for the tumor were always positive for MMTVrtTa and Teto-Myc or MMTV-c-Neu, and depending on what transcription factor was to be studied, Teto-Cre could have been positive or negative. The development mice were always positive for MMTV-rtTa, and depending on what was to be studied were positive or negative for the oncogenes and Teto-Cre. E2F3 was always floxed for both alleles, and because E2F1 and E2F2 were conventional knock outs, the alleles were mutant/mutant. Induction of Myc and Cre In order for c-myc and Cre recombinase to be induced in the mammary gland, the tetracycline activator encoded by the MMTV gene needed doxycycline in order to bind to the tet-on promoters. The Neu line of mice did not need doxycycline for oncogene expression, but they did need it for Teto-Cre expression. The doxycycline was given to the mice through their water supply. I initiated doxycycline into their diet at 30 days of age in a concentration of 2mg/ml in distilled water, and they received it until time of harvest. Mouse Husbandry I checked for tumor initiation twice a week after the mice were on doxy for three months. The primary tumor location and the date were noted, in order to distinguish it from other tumors that may have developed. Some mice were reserved for analyzing the mammary 7
9 gland prior to tumor initiation, or developmental studies. In order to maintain the mouse lines, it was necessary to keep some mice aside from the study and set them up with males for breeding. General Tissue Collection The mice were harvested at 45 days after tumor initiation, and the tumors were measured for their size in millimeters. Tissue was collected from all tumors that had formed, and placed them in three tubes immediately on dry ice for future DNA, RNA, and protein isolation. I also re-tailed the mice to isolate DNA for re-genotyping. These tubes were then placed at -80 C. Any tumor tissue that still remained was placed in Neutral Buffered Formalin. The heart, lungs, kidney, liver, and spleen were examined for metastasis, and then placed in Neutral Buffered Formalin as well. When mice were harvested for developmental studies, it was 30 days after doxycycline induction. Two glands were removed, position 4 was used for a whole mount stain and position 9 was placed in Neutral Buffered Formalin. Tissue Preparation for the Analysis of Mammary Gland Development The gland in position 4 used for a Carmine whole mount stain was placed in a 3:1 solution of 95% Ethanol and Glacial Acetic Acid (Carnoy s Fixative) overnight at 4 C, followed by a series of consecutive hydrations of 70%, 50%, and 30% Ethanol, followed by 2 washes of distilled water. They were then stained overnight with Carmine stain at 4 C, and dehydrated with consecutive solutions of 70%, 90%, 95% and two washes of 100% ethanol. The slides were then placed in Xylene until they were mounted with Biomedia Permount and glass slide covers. The other gland that was placed in Formalin 8
10 was processed and embedded into paraffin sections. The 5um sections were stained with Hematoxylin and Eosin (H&E). Tissue Preparation of Tumor Mice The frozen tumor tissue could be used for DNA, RNA, or protein isolation. I isolated DNA from the tumor by incubating the sample with Lysis Buffer and Protinease K at 55 C overnight. 6M saturated NaCl was added, and sample was left on ice for 10 minutes. After centrifuging, the supernatant was added to 700ul isopropanol and centrifuged again. The supernatant was then removed, and the pellet was resuspended in 200ul of TE buffer. I also isolated protein from the tissue by adding Protein Extraction Buffer (Hepes, KCl, 0.5M EDTA, 0.5M EGTA, Glycerol, and NP40) with the Protinease inhibitors 50mM NaF, 1 MM NaVO4, 1mM PMSF, 1ug/ml aprotinin, pepstain, and leupeptin to the sample. It was then homogenized, and the volume adjusted based on a sample weight at 120mg/ml ratio. The sample was kept on ice for 10 minutes, and centrifuged at 4 C. The supernatant was transferred to a new tube, and then was separated into 4-6 aliquots and kept at -80 C. The rest of the tumor was kept in Formalin until it was processed and embedded into paraffin sections. The 5um sections were also stained with Hematoxylin and Eosin(H&E). Data Compilation Microsoft Excel was used in order to create Kaplan-Meyer curves of the tumor mice population. The graphs showed the percentage of the population that remained tumor free in order to see if the loss of the transcription factors had any affect on the rate of tumorigenesis. Excel was also used to create the bar graphs of average tumor size and number of tumors per mouse. 9
11 Southern Blot Analysis Southern blot analysis was completed to determine if E2F3 was deleted efficiently in the tumor. I digested the DNA with the restriction enzyme EcoRV and used an E2F3 radioactively labeled DNA probe. Southern analysis was completed on all tumors that were considered positive for Teto-Cre, and also on several cre negative tumors as a control. After I accumulated all the data, in order to quantify the percentage of deletion I determined the volume of both bands using the Typhoon Phosphoimager Screening ImageQuant Program, and divided the knock out band s volume by the total volume of both bands. Western Blot Analysis Protein expression of the transcription factors in the tumors was analyzed by using the Western Blot technique. Using the aforementioned protein samples I isolated, I diluted the protein with PBS and bromophenol blue dye to a 30ug concentration. I heat shocked them in boiling water and ran them on 10% separating and 6% stacking gels. The separated protein was then transferred on to a membrane. Using the E2F1, 2, or 3 primary antibodies in 5% milk in TBST, followed by their respective secondary antibodies, I probed the membranes and then observed the results on X-Ray film. β-galactosidase staining The mammary gland in position 4 was also used for β-galactosidase in mice with the LacZ reporter gene. It was harvested and preserved in a fixative (4% paraformaldehyde, 0.1M phosphate buffer ph 7.3, and 25% glutaraldehyde) for 2-6 hours at 4 C, and then 10
12 stained (0.8x PBS, 5mM potassium ferricyanide, 5mM potassium ferrocyanide, 0.8mg/ml X-gal, 0.1% deoxycholate, 0.2% NP-40, and 2mM magnesium chloride) overnight at room temperature. The gland was washed in PBS overnight, then dehydrated as the Carmine slides were and placed in Xylene until mounting. Statistics T-tests were performed to determine p values on each of the Kaplan-Meyer curves by Julie Stephens, MS, and Soledad Fernandez, PhD, for The Center for Biostatistics at The Ohio State University. Results When analyzing the data from this study, it is imperative that we are positive of the genotypes of the mice. I isolated DNA from the tail taken at harvest, and used PCR techniques to verify the alleles for each gene being studied: MMTV-c-Neu, Teto-Myc, MMTV-rtTa, Teto-Cre, and the applicable E2Fs to the mouse. The E2F3 PCR was particularly important, because it was necessary for both alleles to be floxed in order for the conditional system to be effective (Figure 3). If there was a mistake in genotyping for any of the transgenes, the observed result would be false. Upon re-genotyping, mice that were not re-confirmed with the original genotype were re-genotyped again to see if the mouse should be changed to a different group. A small portion of those results were uninterpretable, and those particular mice were censored. To compare the roles of the E2Fs in the Myc and Neu pathways, I examined the mammary development of Myc-overexpressing and Neu-overexpressing glands deficient in the different transcription factors. I also studied mammary glands with deficiencies in 11
13 the transcription factors, but that did not express the Myc or Neu transgenes. Mice were harvested 30 days after the addition of doxy to the water supply (see Materials and Methods). Mammary glands were harvested and stained with carmine stain. After staining, I evaluated the development by looking at whole mount pictures (Figures 4 and 5). In this analysis, the glands appeared to have normal duct architecture, with typical budding formation at the end of the ducts. Comparison of the control to the Teto-Cre positive, E2F3 f/f showed minimal change in the development of the gland, and the comparison of the Teto-Myc positive control to the Teto-Myc, Teto-Cre, E2F3 f/f gland did not show a very large change. The Neu positive control compared to the MMTV-c- Neu, Teto-Cre, E2F3 f/f gland did show some loss of budding and growth. The majority of E2F2 deficient glands showed an acceleration of bud formation when compared to the wild type control. This shows E2F2 as a potential tumor suppressor, contrary to past studies (Wu et al. 2001). Because growth was accelerated without E2F2 present, it implies E2F2 may be a repressor of cell proliferation. E2F1 m/m mice show a slight delay in growth of the mammary tissue as compared to the wild type control. When the Myc and Neu proto-oncogenes are overexpressed in combination with E2F1 m/m the majority of the glands exhibit rescued growth to a normal rate. E2F2 m/m and E2F3 f/f mice showed varying evidence of both loss of growth and acceleration of growth for both Myc and Neu positive, Teto-Cre positive mice. E2F1 m/m and E2F3 f/f mice showed slower growth than the control in both Myc and Neu mice, suggesting these two both contribute to cellular proliferation. Overall, the development data provides preliminary information on the roles the E2F factors might play, but alone it does not provide conclusive evidence of their functions. 12
14 β-galactosidase staining was used to ensure efficient deletion of E2F3 in the mammary gland by viewing expression of cre recombinase. The lacz reporter gene was utilized to test for Cre expression. Where Cre is expressed, lacz is expressed also, and I stained for lacz expression. Glands that were positive for Teto-Cre stained for complete expression across the entire mammary gland, which can be seen in the whole mount view as well as a 2.5x magnification (Figure 6). The X-gal stain is uniform across the entire gland, which implies that Cre is expressed efficiently and E2F3 should be deleted efficiently as well. Those negative for cre stained for no expression. Southern blot analysis is in progress to confirm this finding. While the development data provides clues as to the roles of these particular E2Fs in the Myc and Her-2 pathways, the focal point of the proposed analysis was to compare tumor initiation dates among Myc and Neu induced mammary adenocarcinomas. It was then necessary to harvest the mouse in order to obtain molecular analysis data of the tumors, (Figure 7) and compare the phenotypes and number of tumors among the different genotypes. Evaluation of average tumor size among the different genotypes showed no distinct difference among Myc mice, but the Neu mice show smaller tumors when the transcription factors are lost (Figure 8). Average primary tumor size showed the same trend, with the Myc mice not showing a definitive trend and the Neu mice developing smaller tumors when the E2Fs were affected (Figure 9). Comparison of the average number of tumors per mouse shows that all genotypes were around the same range for the Myc mice, except for the loss E2F1 and 3 in combination (Figure 10). This category did only contain 5 mice for the Myc line, but it showed a smaller number of 13
15 tumors per mouse than the other genotypes. The Neu genotypes did not significantly differ in their number of tumors per mouse. Careful analysis of tumor start dates and genotypes of the mice resulted in several important findings. Upon comparison of the percentage of tumor free mice in the different genotype populations, it shows that E2F3 deletion alone does not have a very significant effect in the Myc pathway (p=0.971) (statistically significant at 0.01). The Kaplan-Meyer curve of E2F3 looks almost identical to that of Myc induced tumors alone, with a slight acceleration initially, but then it crosses back over to a delay. Neu tumors on the other hand, do show a significant delay when E2F3 is deficient in the mouse (p<0.0001) (Figure 11). E2F2 deficient mice suffered an acceleration of tumorigenesis as compared to the Myc positive control induced tumors (p=0.018). This further emphasizes the preliminary data seen from the development study that E2F2 may play a tumor suppressor role in the Myc pathway. It appears that the loss of E2F2 and E2F3 in combination even further accelerate tumorigenesis in Myc, but this had an overall p value of and was not considered significant. This is most likely because the sample size in this genotype was only 11 mice. When examining the Neu tumor data, it shows that E2F2 delays tumorigenesis in this case, contrasting the Myc pathway (p=0.003, statistically significant at ) (Figure 12). When 2 and 3 were lost in combination, tumor development was delayed with a similar rate as E2F2 loss alone (p=0.004). After evaluation of the loss of E2F1 in Myc induced tumors compared to the Myc control, it appears that E2F1 causes a delay in tumorigenesis. Initially, the rate is about the same as Myc control tumors, but the tumors then start to occur later. There are only 14 mice in this category, so it is not seen as significant with a p value of.632. Loss of E2F1 and 14
16 E2F3 combined seemed to have a major effect on delaying Myc-mediated tumorigenesis, with a noticeable curve of a much slower rate than the Myc induced tumors alone. This is also not considered significant, because there are only 5 mice in the category (p=0.074). Loss of E2F1 in the Neu tumors also caused a delay, but was found insignificant with a p value of Loss of both E2Fs 1 and 3 caused a much larger delay than deletion of E2F1 alone, with a p value of (Figure 13). This suggests that E2Fs 1 and 3 in this case may have functional redundancy. Completion of Southern Blot analysis allowed me to examine whether the Cre recombinase deletion was efficient in the model, and was also used to investigate whether E2F3 was required for tumor formation (Figure 14). If the tumor tissue all contained E2F3 wild type alleles, then it would imply that there was a selection process and malignant cells all needed to contain E2F3. But if the tissue had E2F3 deletion or some level of it, as in the Myc case, it follows that E2F3 is not necessarily required for cells to become over-prolific. It also shows that Cre recombinase was effective. When comparing E2F3 floxed/floxed mice to the other genotypes, their tumors were indeed E2F3 deficient, therefore the phenotype that followed can be attributed to the loss of E2F3. In the 29 Myc positive, E2F3 floxed/floxed mice, 16 had a deletion percentage of 75% of higher, 11 were between 50%-75% deletion, 1 was in between 25%-50% deletion, and only 1 had 0% deletion. Of the 14 Myc positive, E2F2 mutant/mutant, and E2F3 floxed/floxed mice, 5 tumors had 75% or greater deletion, 8 were between 50%- 75%, and 1 was between 25% and 50%. In the Neu case, the Southern results were much different, with the majority of the tumors being non deleted for E2F3. Only 13%, or 4 out of 30, had a deletion of greater than 90% for E2F3. The rest of the tumors were 15
17 essentially 0% deletion. This follows that E2F3 was indeed required for tumorigenesis to occur in the neu-mediated mice, calling for a much higher importance of this transcription factor than in the Myc-mediated pathway. In results not shown, normal E2F3 protein expression analyzed by Western blot is a faint band for a normal mouse mammary gland. As shown here, E2F3 protein expression for Myc and Neu is significantly higher (Figure 15a). The thick band on the top is indicative of strong E2F3 expression, linking the both oncogenes to increased E2F expression. This result shows that E2Fs are indeed a target of the two oncogenes, especially Myc, even though they don t play a significant role in tumorigenesis rates. Due to antibody problems, the Myc-E2F2 western was the only one to work. It was found that while the control mammary gland shows a faint band for E2F2 protein expression, the Myc positive tumors showed absolutely no E2F2 expression (Figure 15b). This result complies with the findings that E2F2 acts as a tumor suppressor in the Myc pathway. The data shows that when tumors are able to occur, it is in the absence of E2F2 protein and its tumor suppressor function. Discussion It can be seen that loss of E2Fs 1-3 still allow tumorigenesis to occur, and loss of E2F2 even accelerates tumor formation in the Myc mouse mammary model. Even when E2Fs 1 and 3 are lost in combination, tumor formation still occurs. So while these transcription factors seem to play somewhat of a role in affecting the rate of tumorigenesis, they do not seem to be crucial for growth to occur in the Myc model. It appears that by delaying the tumor progression, E2Fs can act as activators of cellular 16
18 proliferation. Because the loss of 1 and 3 in combination was much stronger of a phenotype than the loss of the two individually, it seems these two have functional redundancy, and can make up for the other s role when it is absent. E2F1 deficiency by itself does have the stronger phenotype than E2F3 loss alone, so it suggests that it may play a larger role in regulating cell growth in the Myc pathway. But, the small population of E2F1 deficient Myc tumor mice does not allow for a strong conclusion to be made. The loss of these activators does not seem to show a strong phenotype in the development studies, whether singularly or in combination, so this data does not provide sufficient evidence to back this claim either. While no solid assertion can be made regarding the exact roles of E2Fs 1 and 3 and how they interact with one another in the Myc pathway, it does remain interesting how much tumorigenesis in the E2F1/3 deficient mice was delayed. E2F2 increases the rate of tumor occurrence in the mouse population, and even shows an altered development of the mammary gland in 60 day old virgin mice, with an obvious increase in branching and bud formation. In unpublished data, our lab has found E2F2 may cause apoptosis in Myc-overexpressing cells. Therefore, without E2F2, apoptosis is prevented, and tumor formation is able to occur at a faster rate in the Mycoverexpressing mice. While it was not statistically significant that the loss of E2F2 and 3 in combination delayed tumorigenesis even further, it does still suggest the synergistic effects of the E2F transcription factors. When E2F2 is deficient, E2F3 may play a reverse role than it does with E2F1 in order to make up for what the growth regulation was lacking without 2 present. There is evidence of E2F1 activating cellular 17
19 proliferation while also causing apoptosis via p53, and the data in this study suggests that E2F2 and 3 may also be able to have opposite effects in regulating cell proliferation. When evaluating Neu tumors, the analysis is much different. Because E2F3 caused a significant delay in Neu-mediated tumors, it shows a much more significant role. The fact that essentially no tumors developed without the presence of E2F3 allows for the assertion that it is crucial for tumorigenesis in this pathway. One could safely say that E2F3 acts as a growth activator in the Neu pathway. E2F2 may cause controversy, because in this pathway it seems to act as a growth activator. This experiment proposes dual roles for E2F2, as a growth activator in Neu and as a growth repressor in Myc. E2F1 also seems to act as a growth activator, because tumorigenesis was also delayed when it was lost in this case. The synergistic effects of the activators lost in combination do not cause for a much larger delay in tumorigenesis as would be thought in this pathway, because all the E2Fs seem to show a very important role. Analysis of all three lost would be interesting, but is hard to accomplish because of the severe growth defects these mice have in multiple tissues, which could complicate the interpretation of the results. The evidence from this experiment does suggest that the transcription factors play a role in the Myc-mediated pathway to tumorigenesis, but it appears Myc also acts through other growth factors in order to cause cells to proliferate rapidly. It is known that c-myc causes up-regulation of Cyclin D1, but it is possible that c-myc acts through factors other than the Cyclin D1-Rb-E2F pathway in order to cause transcription of genes needed for cell growth. When Cyclin D1 was deficient in Ras and c-neu mice, mammary tumorigenesis was avoided all together. On the contrary, when Cyclin D1 was 18
20 deficient in Wnt-1 and c-myc mice, tumorigenesis did indeed occur, albeit tumorigenesis was slightly delayed in these settings (Yu, Geng, & Sicinski 2001). This evidence alone shows that the c-myc pathway is able to overcome the barrier the loss of the Cyclin D1- Rb-E2F pathway can create, and can help explain why there was not a large effect in the rate of tumorigenesis when these transcription factors were disrupted in the c-myc mice. C-Myc is able to act through other factors to cause exponential growth, and investigation of these other targets of the c-myc oncogene is essential for disrupting Myc-mediated tumorigenesis. This past Cyclin D1 study can also help to explain why there was a much larger effect on the tumorigenesis of Her-2 mice when E2F3 was lost. Cyclin D1 was imperative for mammary tumors to form in the Neu-mediated pathway, so this indicates that Her-2 acts only through the Cyclin D1-Rb-E2F pathway. Because E2F occurs later in the pathway than Cyclin D1, it is possible that other factors are able to contribute against the loss of E2F3 and still allow tumorigenesis to occur. Because majority of the tumors do still have E2F3 present in the cells, this shows how essential E2F3 is in Neumediated tumorigenesis to cause those cells to proliferate. Examining the data between the Neu and c-myc oncogenes, it has been shown that the importance of E2F function is signaling-specific, but could it also be tissue specific? In Cyclin D1 deficient mice with Ras and Neu oncogenes, skin tumors as well as salivary tumors were still able to form, and mouse embryo fibroblasts grew viciously in soft agar and caused tumors in nude mice. This shows that not only is the role of Cyclin D1 and therefore E2F function dependent on the pathway, but it is also dependent on the tissue in which it is present. I would predict that loss of E2Fs 1 and 3 in c-myc 19
21 mice in any tissue would still allow for tumorigenesis, and that loss of E2F2 might still increase the rapidity of tumor formation but no more than the rate in mammary epithelial cells. I would also predict that loss of E2Fs 1, 2, and 3 would allow for tumorigenesis in other tissues in the Neu model as well. This study has provided further background information regarding oncogenic pathways and breast cancers. Breast cancer is highly hereditary and dependent on genetic mutation more than environmental factors, so knowledge regarding these pathways is imperative. About 50% of human breast cancers contain amplification and/or overexpression of the c-neu gene, and the data this experiment provides could potentially help to shut down those tumor causing pathways. Because E2F3 provides a crucial role in Neu-mediated growth, it can be a valid target for therapeutics involving Neu mammary tumors. This data also further emphasizes the past study of the importance of Cyclin D1 in the Neu-mediated pathway, and because of evidence of tissue specificity, anti-therapy to this pathway could be invaluable to Neu-mediated breast cancers. The c-myc aspect of the study has led us to search for more targets of the Myc pathway, in order to discover how potent the oncogene really is and to narrow down more growth targets that could be involved in several types of tissues and cancers. C-Myc is commonly overexpressed in breast cancer, so narrowing down the growth factors it signals through will lead to therapeutics for Myc-mediated breast cancers as well. Signaling pathways in mammalian development are highly complex, as this study helps to emphasize, and the more information known about these pathways, the more likely we are to combat the most common malignancy among women. 20
22 References 1. Bartkova, J. et al: Cyclin D1 protein expression and function in human breast cancer. Int. J. Cancer 57, (1994) 2. DeGregori, J., G. Leone, K. Ohtani, A. Miron & J. R. Nevins: E2F-1 accumulation bypasses a G1 arrest resulting from the inhibition of G1 cyclin-dependent kinase activity. Genes Dev 9, (1995) 3. DeGregori, J., G. Leone, A. Miron, L. Jakoi & J. R. Nevins: Distinct roles for E2F proteins in cell growth control and apoptosis. Proc Natl Acad Sci USA 94, (1997) 4. Dickson, C. et al: Amplification of chromosome band 11q13 and a role for cyclin D2 in human breast cancer. Cancer Lett. 90, (1995) 5. Johnson, D.G., R. Schneider-Broussard: Role of E2F in cell cycle control and cancer. Frontiers in Bioscience 3, (1998) 6. Lukas, J., B. O. Petersen, K. Holm, J. Bartek & K. Helin: Deregulated expression of E2F family members induces S-phase entry and overcomes p16ink4a-mediated growth suppression. Mol Cell Biol 16, (1996) 7. Nevins, J.R: The Rb/E2F pathway and cancer. Human Molecular Genetics 7, (2001) 8. Qin, X.-Q., Livingston, W. G. Kaelin Jr., & P. D. Adams: Deregulated transcription factor E2F-1 expression leads to S-phase entry and p53-mediated apoptosis. Proc Natl Acad Sci USA 91, (1994) 21
23 9. Schwarz, J. K., C. H. Bassing, I. Kovesdi, M. B. Datto, M. Blazing, S. George, X-F. Wang, & J. R. Nevins: Expression of the E2F1 transcription factor overcomes type b transforming growth factor-mediated growth suppression. Proc Natl Acad Sci USA 92, (1995) 10. Stevaux, O. & N.J. Dyson: A revised picture of the E2F transcriptional network and Rb function. Cell Biol 14, (2002) 11. van den Heuvel, S. Cell cycle regulation (September 21, 2005), WormBook ed. The C. elegans Research Community, WormBook, doi/ /wormbook , Wang, Y., A. Thakur, Y. Sun, J. Wu, H. Biliran, A. Bollig, & D. Joshua Liao: Sunergistic effect of cyclin D1 and c-myc leads to more aggressive and invasive mammary tumors in severe combined immunodeficient mice. Cancer Res 67 (8), (2007) 13. Wu, C. L., M. Classon, N. Dyson & E. Harlow: Expression of dominant-negative mutant DP-1 blocks cell cycle progression in G1. Mol Cell Biol 16, (1996) 14. Wu, L., C. Timmers, B. Maiti, H. Saavedra, L. Sang, G. Chong, F. Nuckolls, P. Giangrande, F. Wright, S. Field, M. Greenberg, S. Orkin, J. Nevins, M. Robinson, & G. Leone: The E2F1-3 transcription factors are essential for cellular proliferation. Nature 414, (2001) 15. Yu, Q., Y. Geng, P. Sicinski: Specific Protection against breast cancers by cyclin D1 ablation. Nature 411, (2001) 22
24 16. Zhu, L., S. van den Heuvel, K. Helin, A. Fattaey, M. Ewen, D. Livingston, N. Dyson & E. Harlow: Inhibition of cell proliferation by p107, a relative of the retinoblastoma protein. Genes Dev 7, (1993) 23
25 a. Neu Wnt Ras Myc Cyclin D1/D2 Cdk4? E2F1/E2F2/E2F3 Mammary Adenocarcinoma b. loxp Exon 3 loxp E2F3 floxed allele +Cre E2F3 mutant allele Fig. 1. a) Proposed pathway for Neu and c-myc, acting through Cyclin D1 and the E2F transcription factors to cause over-proliferation b) An E2F3 f/f allele. Exon 3 of the E2F3 gene was flanked by lox-p sites, where Cre recombinase was then able excise the exon and recombine the DNA, creating a mutant allele.
26 Fig. 2. A tet-on expression system was used to conditionally delete E2F3 and overexpress Myc in mouse mammary glands. The system is made specific for mammary glands by the mouse mammary tumor virus (MMTV) gene. Doxycycline is used to activate the transcriptional activator rtta so that it can bind to the tet-on promoter and activate transcription of the target gene.
27 M +/+ f/f / +/+ +/f +/ C 100 bp- 200 bp- 300 bp- E2F3 / E2F3f/f E2F3+/+ 400 bp- Fig. 3. Polymerase Chain Reaction for the E2F3 gene. The M lane represents a DNA marker, and the following lanes represent the respective genotypes. The final lane is the control lane. The top band is the E2F3 mutant band, the middle represents the E2F3 band flanked by loxp sites ( floxed ), and the lowest band represents the E2F3 wild type band.





























28 control E2F3 f/f E2F1 -/- E2F1 -/- E2F3 f/f E2F2 -/- E2F2 -/- E2F3 f/f Myc+ Myc E2F3 f/f Myc E2F2 -/- Myc E2F2 -/- E2F3 f/f Myc E2F1 -/- Myc E2F1 -/- E2F3 f/f Neu+ Neu E2F3 f/f Neu E2F2 -/- Neu E2F2 -/- E2F3 f/f Neu E2F1 -/- Neu E2F1 -/- E2F3 f/f Fig. 4. Carmine mammary gland whole mounts of 60 day old virgin mice, 30 days after doxycycline induction, in order to examine the development of the mammary tissue and ducts lacking the individual transcription factors. All mice were positive for MMTV-rtTa. E2F3 f/f mice were positive for Teto-Cre. The labels coordinate to the specific oncogene that was overexpressed and E2F factor genotype.











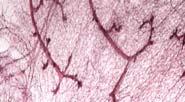





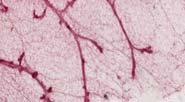
















29 control E2F3 f/f E2F2 -/- E2F2 -/- E2F3 f/f E2F1 -/- E2F1 -/- E2F3 f/f Myc+ Myc E2F3 f/f Myc E2F2 -/- Myc E2F2 -/ - E2F3 f/f Myc E2F1 -/- Myc E2F1 -/ - E2F3 f/ Neu+ Neu E2F3 f/f Neu E2F2 -/- Neu E2F2 -/- E2F3 f/f Neu E2F1 -/- Neu E2F1 -/- E2F3 f/f Fig. 5. Carmine stained mammary glands of 60 day old virgin mice, 30 days after doxycycline induction, 2.5x magnification to view the duct and budding formation. All mice were positive for MMTV-rtTa, and those indicated with E2F3 f/f are positive for Teto-Cre. The labels coordinate to the respective oncogene expression and E2F factor alleles.















30 a. b. c. d. Fig. 6. Examination of Cre expression via Xgal staining for the FSP-cre/Rosaloxp reporter gene system a) MMTV-rtTa, Teto-Cre positive, Rosaloxp positive whole mount showing complete Cre expression and implying efficient E2F3 deletion b) 2.5x magnification of Xgal stained whole mount c) MMTV-rtTa positive, Teto-Cre negative, Rosaloxp positive whole mount control, showing no blue staining, and therefore no E2F3 deletion d) 2.5x magnification of Xgal stained control










31 a. b. c. Fig. 7. Harvest of the tumor mouse a) The outside of the mouse, tumor at mammary gland number 7. b) The outside of the mouse, showing the relative tumor size c) The inside of the mouse, tumors at mammary glands 6 and 8.
32 a. Av e rage Tumor Size Area (mm^2) Myc+ Myc+ E2F2-/- Myc+ E2F1-/- Myc+ E2F3f/f Myc+, E2F2-/-, E2F3f/f Myc+ E2F1-/- E2F3 f/f b Average Tumor Size Area (mm^2) Neu+ Neu+ E2F2-/- Neu+ E2F1-/- Neu+ E2F3f/f Neu+ E2F2-/- E2F3f/f Neu+ E2F1-/- E2F3f/f Fig. 8. a) Bar graph exemplifying the average tumor size of Teto-Myc positive mice, with a calculated standard deviation for each genotype. Each mouse was MMTV-rtTa positive, and those with E2F3 f/f alleles were Teto-Cre positive b) Bar graph showing average tumor size of MMTV-c-Neu positive mice, with a calculated standard deviation for each genotype. Each mouse was MMTV-rtTa positive, and those with E2F3 f/f alleles were Teto-Cre positive.
33 a. 800 Average Primary Tumor Size Area (mm^2) Myc+ Myc+ E2F3f/f Myc+ E2F2-/- Myc+ E2F2-/- E2F3f/f Myc+ E2F1-/- E2F3 f/f Myc+ E2F1/3- b Average Primary Tumor Size Area (mm^2) Neu+ Neu+ E2F3f/f Neu+ E2F2-/- Neu+ E2F2-/- E2F3f/f Neu+ E2F1-/- Neu+ E2F1-/- E2F3f/f Fig. 9. a) Bar graph exemplifying the primary tumor size of Teto-Myc positive mice, with a calculated standard deviation for each genotype. Each mouse was MMTV-rtTa positive, and those with E2F3 floxed alleles were Teto-Cre positive b) Bar graph exemplifying the primary tumor size of MMTV-c-Neu mice, also with a calculated standard deviation for each genotype. Each mouse was MMTV-rtTa positive, and those with E2F3 floxed alleles were Teto-Cre positive.
34 a. 3.5 Average Number of Tumors per Mouse 3 Number of Tumors Myc+ Myc+ E2F3f/f Myc+ E2F2-/- Myc+ E2F2-/- E2F3f/f Myc+ E2F1-/- Myc+ E2F1-/- E2F3f/f b. Average Number of Tumors per Mouse Number of Tumors Neu+ Neu+ E2F3f/f Neu+ E2F2-/- Neu+ E2F2-/- E2F3f/f Neu+ E2F1-/- Neu+ E2F1-/- E2F3f/f Fig. 10. a) Bar graph of the average number of tumors per Teto-Myc positive mouse, with a calculated standard deviation for each genotype. Every mouse was MMTV-rtTa positive, and those with E2F3f/f alleles were Teto-Cre positive. b) Bar graph of the average number of tumors per MMTV-c-Neu mouse, with a calculated standard deviation for each genotype. Each mouse was MMTV-rtTa positive, and those with E2F3f/f alleles were Teto-Cre positive.
35 a. Myc v. Myc-E2F3 120 Percentage of tumor free mice Days from Birth Myc n=33 T50=243 Myc, E2F3f/f n=22 T50=215 b. Neu v. Neu-E2F3 120 Percentage of tumor free mice Days from birth Neu n=53 T50=206 Neu E2F3f/f n=34 T50=284 Fig. 11. a) Kaplan-Meyer curve of the tumor free population of the Teto-Myc and MMTV-rtTa positive mice (blue), n=33, versus Teto-Myc, MMTV-rtTa, and Teto-Cre positive, E2F3f/f mice (pink), n=22. The p-value was for this pairwise comparison. b) Kaplan-Meyer curve of the tumor free population of MMTV-c-Neu and MMTV-rtTa positive mice (blue), n=53, versus MMTV-c-Neu, MMTV-rtTa, and Teto-Cre positive, E2F3f/f mice (pink), n=34. The p-value was less than for this comparison.
36 a. Myc v Myc-E2F2 v MycE2F2/3 120 Percentage of Tumor Free Mice Days from birth Myc n=33 T50=243 Myc, E2F2-/- n=26 T50=213 Myc, E2F2-/- E2F3f/f n=11 T50=203 b. Neu v Neu-E2F2 v Neu-E2F2/3 120 Percentage of Tumor Free Mice Days from birth Neu n=53 T50=206 Neu E2F2-/- n=18 T50=274 Neu E2F2-/- E2F3f/f n=26 T50=281 Fig. 12. a) Kaplan-Meyer pairwise comparison of the percentage of tumor free mice. Teto- Myc, MMTV-rtTa positive mice (blue), n=33, versus Teto-Myc, MMTV-rtTa positive, E2F2 mutant mice (yellow), n=26, versus Teto-Myc, MMTV-rtTa, Teto-Cre positive, E2F2 mutant, E2F3 f/f mice (teal), n=11. The p value of Myc v. Myc-E2F2 is 0.018, and the p value of Myc v. Myc E2F2/3 is b) Kaplan-Meyer pairwise comparison of MMTV-c-Neu, MMTV-rtTa positive mice (blue), n=53, to MMTV-c-Neu, MMTV-rtTa positive, E2F2 mutant mice (yellow), n=18, to MMTV-c-Neu, MMTV-rtta, Teto-Cre positive, E2F2 mutant, E2F3 f/f mice (teal), n=26. The p value of Neu v Neu-E2F2 is 0.003, and the p-value of Neu v Neu E2F2/3 is
37 a. Myc v Myc-E2F1 v Myc-E2F1/3 120 Percentage of tumor free mice Days from birth Myc n=33 T50=243 Myc, E2F1-/- n=14 T50=256 Myc, E2F1-/- E2F3f/f n=5 T50=285.5 b. Neu v. Neu-E2F1 v. Neu-E2F1/3 120 Percentage of tumor free mice Days from birth Neu n=53 T50=206 Neu, E2F1-/- n=19 T50=230.5 Neu, E2F1-/- E2F3f/f n=13 T50=294.5 Fig. 13. a) A Kaplan-Meyer pairwise comparison of Teto-Myc, MMTV-rtTa positive mice, n=33, versus Teto-Myc, MMTV-rtTa, E2F1 mutant mice, n=14, and Teto-Myc, MMTV-rtTa, Teto-Cre positive, E2F1 mutant, E2F3f/f mice, n=5. The p value of Myc versus Myc-E2F1 was 0.632, and the p-value of Myc versus Myc-E2F1 & 3 was b) A Kaplan-Meyer pairwise comparison of MMTV-c-Neu, MMTV-rtTa positive mice, n=53, versus MMTV-c-Neu, MMTV-rtTa positive, E2F1 mutant mice, n=19, versus MMTV-c-Neu, MMTV-rtTa, Teto-Cre positive, E2F1 mutant, E2F3f/f mice, n=13. The p value of Neu versus Neu-E2F1 was 0.036, and the p value of Neu versus Neu-E2F1 & 3 was
 E2F3 southern blot phosphoimage of Teto-Myc, MMTV-rtTa, and Teto-Cre positive, E2F3 floxed mice. The top band is E2F3 floxed/floxed band, the bottom band is E2F3 mutant band.")
 E2F3 southern blot phosphoimage of MMTV-c-Neu, MMTV-rtTa, and Teto-Cre positive, E2F3 floxed mice.")
38 a. wt H 89.5% 100% 95.8% 79.2% 69.9% 86.3% 93.5% E2F3 f/f E2F3 deleted b. 0% 0% 0% 95% 0% 0% 0% 0% 0% 98% E2F3 f/f E2F3 deleted Fig. 14. a) E2F3 southern blot phosphoimage of Teto-Myc, MMTV-rtTa, and Teto-Cre positive, E2F3 floxed mice. The top band is E2F3 floxed/floxed band, the bottom band is E2F3 mutant band. Lane wt is a Teto-Cre negative control, Lane H is a heterozygous floxed/wild type for the E2F3 alleles that is Teto-Cre positive. The following lanes are indicated by their respective deletion percentages. b) E2F3 southern blot phosphoimage of MMTV-c-Neu, MMTV-rtTa, and Teto-Cre positive, E2F3 floxed mice. The top band is the E2F3 floxed band, and the bottom band is the E2F3 mutant. The lanes are indicated by their respective deletion percentages.



39 a. b. Neu tumors Myc tumors T0* T1 T2 T3 T4 T5 T6 T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 E2F3a c. C T1 T 2 T3 T4 T5 T6 T7 T8 T9 T10 T11 T12 M E2F2 Fig. 15. a) Western blot for E2F3 protein expression of MMTV-c-Neu positive tumors. E2F3a band is the top band of the lane. Lane T0* is an E2F3 deleted control. b) Western blot for E2F3 protein expression of Teto-Myc positive tumors. The top band is indicative of E2F3a expression. c) Western blot for E2F2 protein expression of Teto-Myc positive tumors. Lane C is an E2F2 control sample. The middle band is indicative of E2F2 protein expression.
The Role of E2Fs in Mouse Mammary Gland Development. A Senior Honors Thesis. April Sandy. The Ohio State University June 2006
 The Role of E2Fs in Mouse Mammary Gland Development A Senior Honors Thesis Presented in Partial Fulfillment of the Requirements for graduation with distinction in Microbiology in the College of Biological
The Role of E2Fs in Mouse Mammary Gland Development A Senior Honors Thesis Presented in Partial Fulfillment of the Requirements for graduation with distinction in Microbiology in the College of Biological
Supplementary Information Titles Journal: Nature Medicine
 Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Generating Mouse Models of Pancreatic Cancer
 Generating Mouse Models of Pancreatic Cancer Aom Isbell http://www2.massgeneral.org/cancerresourceroom/types/gi/index.asp Spring/Summer 1, 2012 Alexandros Tzatsos, MD PhD Bardeesy Lab: Goals and Objectives
Generating Mouse Models of Pancreatic Cancer Aom Isbell http://www2.massgeneral.org/cancerresourceroom/types/gi/index.asp Spring/Summer 1, 2012 Alexandros Tzatsos, MD PhD Bardeesy Lab: Goals and Objectives
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Multistep nature of cancer development. Cancer genes
 Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
(Stratagene, La Jolla, CA) (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment
 (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment") SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
Early Embryonic Development
 Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
BIOL2005 WORKSHEET 2008
 BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
Neoplasia 18 lecture 6. Dr Heyam Awad MD, FRCPath
 Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Problem Set 8 Key 1 of 8
 7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
TITLE: A Mouse Model to Investigate the Role of DBC2 in Breast Cancer
 AD Award Number: W81XWH-04-1-0325 TITLE: A Mouse Model to Investigate the Role of DBC2 in Breast Cancer PRINCIPAL INVESTIGATOR: Valerie Boka CONTRACTING ORGANIZATION: University of Texas Health Science
AD Award Number: W81XWH-04-1-0325 TITLE: A Mouse Model to Investigate the Role of DBC2 in Breast Cancer PRINCIPAL INVESTIGATOR: Valerie Boka CONTRACTING ORGANIZATION: University of Texas Health Science
A class of genes that normally suppress cell proliferation. p53 and Rb..ect. suppressor gene products can release cells. hyperproliferation.
 Tumor Suppressor Genes A class of genes that normally suppress cell proliferation. p53 and Rb..ect Mutations that inactivate the tumor suppressor gene products can release cells from growth suppression
Tumor Suppressor Genes A class of genes that normally suppress cell proliferation. p53 and Rb..ect Mutations that inactivate the tumor suppressor gene products can release cells from growth suppression
Problem Set 5 KEY
 2006 7.012 Problem Set 5 KEY ** Due before 5 PM on THURSDAY, November 9, 2006. ** Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. You are studying the development
2006 7.012 Problem Set 5 KEY ** Due before 5 PM on THURSDAY, November 9, 2006. ** Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. You are studying the development
ERK1/2/MAPK pathway-dependent regulation of the telomeric factor TRF2
 ERK1/2/MAPK pathway-dependent regulation of the telomeric factor TRF2 SUPPLEMENTARY FIGURES AND TABLE Supplementary Figure S1: Conservation of the D domain throughout evolution. Alignment of TRF2 sequences
ERK1/2/MAPK pathway-dependent regulation of the telomeric factor TRF2 SUPPLEMENTARY FIGURES AND TABLE Supplementary Figure S1: Conservation of the D domain throughout evolution. Alignment of TRF2 sequences
Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy
 Kavya Puchhalapalli CALS Honors Project Report Spring 2017 Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy Abstract Malignant brain tumors including medulloblastomas and primitive neuroectodermal
Kavya Puchhalapalli CALS Honors Project Report Spring 2017 Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy Abstract Malignant brain tumors including medulloblastomas and primitive neuroectodermal
Hunk is required for HER2/neu-induced mammary tumorigenesis
 Research article Hunk is required for HER2/neu-induced mammary tumorigenesis Elizabeth S. Yeh, 1 Thomas W. Yang, 1 Jason J. Jung, 1 Heather P. Gardner, 1 Robert D. Cardiff, 2 and Lewis A. Chodosh 1 1 Department
Research article Hunk is required for HER2/neu-induced mammary tumorigenesis Elizabeth S. Yeh, 1 Thomas W. Yang, 1 Jason J. Jung, 1 Heather P. Gardner, 1 Robert D. Cardiff, 2 and Lewis A. Chodosh 1 1 Department
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway
 Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Introduction to Genetics
 Introduction to Genetics Table of contents Chromosome DNA Protein synthesis Mutation Genetic disorder Relationship between genes and cancer Genetic testing Technical concern 2 All living organisms consist
Introduction to Genetics Table of contents Chromosome DNA Protein synthesis Mutation Genetic disorder Relationship between genes and cancer Genetic testing Technical concern 2 All living organisms consist
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Prof. R. V. Skibbens
 Prof. R. V. Skibbens December 2, 2011 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality The Cell Cycle clock goes in only one direction S-phase cells
Prof. R. V. Skibbens December 2, 2011 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality The Cell Cycle clock goes in only one direction S-phase cells
TARGETS OF CYCLIN D1-CDK
 TARGETS OF CYCLIN D1-CDK FIRST TARGET OF THE COMPLEX CYCLIN D-KINASI: prb, IS THE PRODUCT OF THE GENE CONFERRING SUSCEPTIBILITY TO RETINOBLASTOMA - ABSENT OR MUTATED IN SEVERAL HUMAN CANCERS - TRANSCRIPTIONL
TARGETS OF CYCLIN D1-CDK FIRST TARGET OF THE COMPLEX CYCLIN D-KINASI: prb, IS THE PRODUCT OF THE GENE CONFERRING SUSCEPTIBILITY TO RETINOBLASTOMA - ABSENT OR MUTATED IN SEVERAL HUMAN CANCERS - TRANSCRIPTIONL
Introduction. Cancer Biology. Tumor-suppressor genes. Proto-oncogenes. DNA stability genes. Mechanisms of carcinogenesis.
 Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer
 AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Karyotype analysis reveals transloction of chromosome 22 to 9 in CML chronic myelogenous leukemia has fusion protein Bcr-Abl
 Chapt. 18 Cancer Molecular Biology of Cancer Student Learning Outcomes: Describe cancer diseases in which cells no longer respond Describe how cancers come from genomic mutations (inherited or somatic)
Chapt. 18 Cancer Molecular Biology of Cancer Student Learning Outcomes: Describe cancer diseases in which cells no longer respond Describe how cancers come from genomic mutations (inherited or somatic)
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Cell Cycle and Cancer
 142 8. Cell Cycle and Cancer NOTES CELL CYCLE G 0 state o Resting cells may re-enter the cell cycle Nondividing cells (skeletal and cardiac muscle, neurons) o Have left the cell cycle and cannot undergo
142 8. Cell Cycle and Cancer NOTES CELL CYCLE G 0 state o Resting cells may re-enter the cell cycle Nondividing cells (skeletal and cardiac muscle, neurons) o Have left the cell cycle and cannot undergo
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
number Done by Corrected by Doctor Maha Shomaf
 number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
Supporting Information
 Supporting Information Franco et al. 10.1073/pnas.1015557108 SI Materials and Methods Drug Administration. PD352901 was dissolved in 0.5% (wt/vol) hydroxyl-propyl-methylcellulose, 0.2% (vol/vol) Tween
Supporting Information Franco et al. 10.1073/pnas.1015557108 SI Materials and Methods Drug Administration. PD352901 was dissolved in 0.5% (wt/vol) hydroxyl-propyl-methylcellulose, 0.2% (vol/vol) Tween
Study of the H-Ras-Rb Axis in Oncogene Induced Senescence. Honors Research Thesis. Presented in partial fulfillment of the requirements for graduation
 Study of the H-Ras-Rb Axis in Oncogene Induced Senescence Honors Research Thesis Presented in partial fulfillment of the requirements for graduation with honors research distinction in Biology in the undergraduate
Study of the H-Ras-Rb Axis in Oncogene Induced Senescence Honors Research Thesis Presented in partial fulfillment of the requirements for graduation with honors research distinction in Biology in the undergraduate
Cancer Genetics. What is Cancer? Cancer Classification. Medical Genetics. Uncontrolled growth of cells. Not all tumors are cancerous
 Session8 Medical Genetics Cancer Genetics J avad Jamshidi F a s a U n i v e r s i t y o f M e d i c a l S c i e n c e s, N o v e m b e r 2 0 1 7 What is Cancer? Uncontrolled growth of cells Not all tumors
Session8 Medical Genetics Cancer Genetics J avad Jamshidi F a s a U n i v e r s i t y o f M e d i c a l S c i e n c e s, N o v e m b e r 2 0 1 7 What is Cancer? Uncontrolled growth of cells Not all tumors
Molecular biology :- Cancer genetics lecture 11
 Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
Lecture 10. G1/S Regulation and Cell Cycle Checkpoints. G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint
 Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Probe. Hind III Q,!?R'!! /0!!!!D1"?R'! vector. Homologous recombination
 Supple-Zhang Page 1 Wild-type locus Targeting construct Targeted allele Exon Exon3 Exon Probe P1 P P3 FRT FRT loxp loxp neo vector amh I Homologous recombination neo P1 P P3 FLPe recombination Q,!?R'!!
Supple-Zhang Page 1 Wild-type locus Targeting construct Targeted allele Exon Exon3 Exon Probe P1 P P3 FRT FRT loxp loxp neo vector amh I Homologous recombination neo P1 P P3 FLPe recombination Q,!?R'!!
7.012 Problem Set 6 Solutions
 Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
The Biology and Genetics of Cells and Organisms The Biology of Cancer
 The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney
 2013 Course University of Sydney") BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
Cancer and Gene Alterations - 1
 Cancer and Gene Alterations - 1 Cancer and Gene Alteration As we know, cancer is a disease of unregulated cell growth. Although we looked at some of the features of cancer when we discussed mitosis checkpoints,
Cancer and Gene Alterations - 1 Cancer and Gene Alteration As we know, cancer is a disease of unregulated cell growth. Although we looked at some of the features of cancer when we discussed mitosis checkpoints,
Regulation of cell cycle. Dr. SARRAY Sameh, Ph.D
 Regulation of cell cycle Dr. SARRAY Sameh, Ph.D Control of cell cycle: Checkpoints Are the cell cycle controls mechanisms in eukaryotic cells. These checkpoints verify whether the processes at each phase
Regulation of cell cycle Dr. SARRAY Sameh, Ph.D Control of cell cycle: Checkpoints Are the cell cycle controls mechanisms in eukaryotic cells. These checkpoints verify whether the processes at each phase
Genome 371, Autumn 2018 Quiz Section 9: Genetics of Cancer Worksheet
 Genome 371, Autumn 2018 Quiz Section 9: Genetics of Cancer Worksheet All cancer is due to genetic mutations. However, in cancer that clusters in families (familial cancer) at least one of these mutations
Genome 371, Autumn 2018 Quiz Section 9: Genetics of Cancer Worksheet All cancer is due to genetic mutations. However, in cancer that clusters in families (familial cancer) at least one of these mutations
Chapter 12. Regulation of Cell Division. AP Biology
 Chapter 12. Regulation of Cell Division Coordination of cell division! Multicellular organism " need to coordinate across different parts of organism! timing of cell division! rates of cell division "
Chapter 12. Regulation of Cell Division Coordination of cell division! Multicellular organism " need to coordinate across different parts of organism! timing of cell division! rates of cell division "
Effec<ve Use of PI3K and MEK Inhibitors to Treat Mutant K Ras G12D and PIK3CA H1047R Murine Lung Cancers
 Effec
Effec
Lecture 8 Neoplasia II. Dr. Nabila Hamdi MD, PhD
 Lecture 8 Neoplasia II Dr. Nabila Hamdi MD, PhD ILOs Understand the definition of neoplasia. List the classification of neoplasia. Describe the general characters of benign tumors. Understand the nomenclature
Lecture 8 Neoplasia II Dr. Nabila Hamdi MD, PhD ILOs Understand the definition of neoplasia. List the classification of neoplasia. Describe the general characters of benign tumors. Understand the nomenclature
CELL CYCLE REGULATION AND CANCER. Cellular Reproduction II
 CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
Reviewers' comments: Reviewer #1 (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 (Remarks to the Author): In this study the authors analysed 18 deep penetrating nevi for oncogenic genomic changes (single nucleotide variations, insertions/deletions,
Reviewers' comments: Reviewer #1 (Remarks to the Author): In this study the authors analysed 18 deep penetrating nevi for oncogenic genomic changes (single nucleotide variations, insertions/deletions,
Introduction to Cancer Biology
 Introduction to Cancer Biology Robin Hesketh Multiple choice questions (choose the one correct answer from the five choices) Which ONE of the following is a tumour suppressor? a. AKT b. APC c. BCL2 d.
Introduction to Cancer Biology Robin Hesketh Multiple choice questions (choose the one correct answer from the five choices) Which ONE of the following is a tumour suppressor? a. AKT b. APC c. BCL2 d.
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2607 Figure S1 Elf5 loss promotes EMT in mammary epithelium while Elf5 overexpression inhibits TGFβ induced EMT. (a, c) Different confocal slices through the Z stack image. (b, d) 3D rendering
DOI: 10.1038/ncb2607 Figure S1 Elf5 loss promotes EMT in mammary epithelium while Elf5 overexpression inhibits TGFβ induced EMT. (a, c) Different confocal slices through the Z stack image. (b, d) 3D rendering
Early cell death (FGF) B No RunX transcription factor produced Yes No differentiation
 B No RunX transcription factor produced Yes No differentiation") Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Chapt 15: Molecular Genetics of Cell Cycle and Cancer
 Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
Tumor suppressor genes D R. S H O S S E I N I - A S L
 Tumor suppressor genes 1 D R. S H O S S E I N I - A S L What is a Tumor Suppressor Gene? 2 A tumor suppressor gene is a type of cancer gene that is created by loss-of function mutations. In contrast to
Tumor suppressor genes 1 D R. S H O S S E I N I - A S L What is a Tumor Suppressor Gene? 2 A tumor suppressor gene is a type of cancer gene that is created by loss-of function mutations. In contrast to
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Prof. R. V. Skibbens
 Prof. R. V. Skibbens September 8, 2017 BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell growth and division What are the goals? I Cell Cycle what is this? response to
Prof. R. V. Skibbens September 8, 2017 BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell growth and division What are the goals? I Cell Cycle what is this? response to
Genetics and Cancer Ch 20
 Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
Determination Differentiation. determinated precursor specialized cell
 Biology of Cancer -Developmental Biology: Determination and Differentiation -Cell Cycle Regulation -Tumor genes: Proto-Oncogenes, Tumor supressor genes -Tumor-Progression -Example for Tumor-Progression:
Biology of Cancer -Developmental Biology: Determination and Differentiation -Cell Cycle Regulation -Tumor genes: Proto-Oncogenes, Tumor supressor genes -Tumor-Progression -Example for Tumor-Progression:
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Functional Limitations
 Regulation of the Cell Cycle Chapter 12 Pg. 228 245 Functional Limitations Various factors determine whether and when a cell divides. Two functional limitations for cell size limit growth or influence
Regulation of the Cell Cycle Chapter 12 Pg. 228 245 Functional Limitations Various factors determine whether and when a cell divides. Two functional limitations for cell size limit growth or influence
Regulation of Cell Division. AP Biology
 Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS
 CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS Summary of the regulation of cyclin/cdk complexes during celll cycle Cell cycle phase Cyclin-cdk complex inhibitor activation Substrate(s) G1 Cyclin D/cdk 4,6
CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS Summary of the regulation of cyclin/cdk complexes during celll cycle Cell cycle phase Cyclin-cdk complex inhibitor activation Substrate(s) G1 Cyclin D/cdk 4,6
BIO360 Fall 2013 Quiz 1
 BIO360 Fall 2013 Quiz 1 1. Examine the diagram below. There are two homologous copies of chromosome one and the allele of YFG carried on the light gray chromosome has undergone a loss-of-function mutation.
BIO360 Fall 2013 Quiz 1 1. Examine the diagram below. There are two homologous copies of chromosome one and the allele of YFG carried on the light gray chromosome has undergone a loss-of-function mutation.
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 AD Award Number: W81XWH-05-1-0262 TITLE: Role of CDK4 in Breast Development and Cancer PRINCIPAL INVESTIGATOR: Haritha Reddy CONTRACTING ORGANIZATION: Temple University Philadelphia PA 19122-6024 REPORT
AD Award Number: W81XWH-05-1-0262 TITLE: Role of CDK4 in Breast Development and Cancer PRINCIPAL INVESTIGATOR: Haritha Reddy CONTRACTING ORGANIZATION: Temple University Philadelphia PA 19122-6024 REPORT
1. Basic principles 2. 6 hallmark features 3. Abnormal cell proliferation: mechanisms 4. Carcinogens: examples. Major Principles:
 Carcinogenesis 1. Basic principles 2. 6 hallmark features 3. Abnormal cell proliferation: mechanisms 4. Carcinogens: examples Carcinogenesis Major Principles: 1. Nonlethal genetic damage is central to
Carcinogenesis 1. Basic principles 2. 6 hallmark features 3. Abnormal cell proliferation: mechanisms 4. Carcinogens: examples Carcinogenesis Major Principles: 1. Nonlethal genetic damage is central to
Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the
 Schematics of the Sellg allele (top), the targeting vector, the") Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the targeted allele in ES cells, and the mutant allele in
Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the targeted allele in ES cells, and the mutant allele in
A protocol for enhancement of the AAV-mediated expression of transgenes
 A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
A Genetic Program for Embryonic Development
 Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Activation of cellular proto-oncogenes to oncogenes. How was active Ras identified?
 Dominant Acting Oncogenes Eugene E. Marcantonio, M.D. Ph.D. Oncogenes are altered forms of normal cellular genes called proto-oncogenes that are involved in pathways regulating cell growth, differentiation,
Dominant Acting Oncogenes Eugene E. Marcantonio, M.D. Ph.D. Oncogenes are altered forms of normal cellular genes called proto-oncogenes that are involved in pathways regulating cell growth, differentiation,
HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
MIT OpenCourseWare http://ocw.mit.edu HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:
 into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:") Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Cancer. October is National Breast Cancer Awareness Month
 Cancer October is National Breast Cancer Awareness Month Objectives 1: Gene regulation Explain how cells in all the different parts of your body develop such different characteristics and functions. Contrast
Cancer October is National Breast Cancer Awareness Month Objectives 1: Gene regulation Explain how cells in all the different parts of your body develop such different characteristics and functions. Contrast
Breeding scheme, transgenes, histological analysis and site distribution of SB-mutagenized osteosarcoma.
 Supplementary Figure 1 Breeding scheme, transgenes, histological analysis and site distribution of SB-mutagenized osteosarcoma. (a) Breeding scheme. R26-LSL-SB11 homozygous mice were bred to Trp53 LSL-R270H/+
Supplementary Figure 1 Breeding scheme, transgenes, histological analysis and site distribution of SB-mutagenized osteosarcoma. (a) Breeding scheme. R26-LSL-SB11 homozygous mice were bred to Trp53 LSL-R270H/+
Regarding techniques of proteomics, there is:
 Molecular الحلقة biology 14 واألخيرة To put you back in the context; the discussion was about Trancriptomics (the study of transcription). The following topic will be PROTEOMICS, which is the study of
Molecular الحلقة biology 14 واألخيرة To put you back in the context; the discussion was about Trancriptomics (the study of transcription). The following topic will be PROTEOMICS, which is the study of
Control of Cell Cycle. Unit 2 Part f III
 Control of Cell Cycle Unit 2 Part f III How often do cells divide and why? The timing and rate of cell division in different parts of the plant or animals are crucial to normal growth, development and
Control of Cell Cycle Unit 2 Part f III How often do cells divide and why? The timing and rate of cell division in different parts of the plant or animals are crucial to normal growth, development and
SAMPLE LITERATURE Please refer to included weblink for correct version. Morphology of Cancer Cells. Edvo-Kit #990. Experiment Objective:
 Edvo-Kit #990 Morphology of Cancer Cells Experiment Objective: 990 In this lab, students will explore the morphology of normal and cancerous cells by staining pre-fi xed cells with methylene blue and eosin.
Edvo-Kit #990 Morphology of Cancer Cells Experiment Objective: 990 In this lab, students will explore the morphology of normal and cancerous cells by staining pre-fi xed cells with methylene blue and eosin.
MOLECULAR BASIS OF ONCOGENESIS
 MOLECULAR BASIS OF ONCOGENESIS MUDr. Jiří Vachtenheim, CSc. 1 Cell processes which result also in cell cycle effects. Differentiation. Differentiated cells are usually in the G0 phase of the cell cycle.
MOLECULAR BASIS OF ONCOGENESIS MUDr. Jiří Vachtenheim, CSc. 1 Cell processes which result also in cell cycle effects. Differentiation. Differentiated cells are usually in the G0 phase of the cell cycle.
Genome of Hepatitis B Virus. VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department
 Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
Prof. R. V. Skibbens. Cell Cycle, Cell Division and Cancer (Part 2)
") Prof. R. V. Skibbens November 22, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality - clocks go in only one direction G1 doesn t have replication-inducing
Prof. R. V. Skibbens November 22, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality - clocks go in only one direction G1 doesn t have replication-inducing
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Supporting Information Table of Contents
 Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
AP VP DLP H&E. p-akt DLP
 A B AP VP DLP H&E AP AP VP DLP p-akt wild-type prostate PTEN-null prostate Supplementary Fig. 1. Targeted deletion of PTEN in prostate epithelium resulted in HG-PIN in all three lobes. (A) The anatomy
A B AP VP DLP H&E AP AP VP DLP p-akt wild-type prostate PTEN-null prostate Supplementary Fig. 1. Targeted deletion of PTEN in prostate epithelium resulted in HG-PIN in all three lobes. (A) The anatomy
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary Figure 1. Generation of a conditional allele of the Kindlin-2 gene. (A) A restriction map of the relevant genomic region of Kindlin-2 (top), the targeting construct
SUPPLEMENTARY INFORMATION Supplementary Figure 1. Generation of a conditional allele of the Kindlin-2 gene. (A) A restriction map of the relevant genomic region of Kindlin-2 (top), the targeting construct
CANCER. Inherited Cancer Syndromes. Affects 25% of US population. Kills 19% of US population (2nd largest killer after heart disease)
") CANCER Affects 25% of US population Kills 19% of US population (2nd largest killer after heart disease) NOT one disease but 200-300 different defects Etiologic Factors In Cancer: Relative contributions
CANCER Affects 25% of US population Kills 19% of US population (2nd largest killer after heart disease) NOT one disease but 200-300 different defects Etiologic Factors In Cancer: Relative contributions
CELL BIOLOGY - CLUTCH CH CANCER.
 !! www.clutchprep.com CONCEPT: OVERVIEW OF CANCER Cancer is a disease which is primarily caused from misregulated cell division, which form There are two types of tumors - Benign tumors remain confined
!! www.clutchprep.com CONCEPT: OVERVIEW OF CANCER Cancer is a disease which is primarily caused from misregulated cell division, which form There are two types of tumors - Benign tumors remain confined
Mitochondrial DNA Isolation Kit
 Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
CHANTALE T. GUY, M5C. A Thesis. for the Degree. Doctor of Philosophy. McMaster University. (c) Copyright by Chantale T.
 Copyright by Chantale T.") ROLE AND MECHANISM OF ACTION OF TYROSINE KINASES IN MAMMARY TUMORIGENESIS By CHANTALE T. GUY, M5C. A Thesis Submitted to the School of Graduate Studies in Partial Fulfilment of the Requirements for the
ROLE AND MECHANISM OF ACTION OF TYROSINE KINASES IN MAMMARY TUMORIGENESIS By CHANTALE T. GUY, M5C. A Thesis Submitted to the School of Graduate Studies in Partial Fulfilment of the Requirements for the
Src-INACTIVE / Src-INACTIVE
 Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Protein Dephosphorylation Methods
 Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Regulation of Cell Division (Ch. 12)
") Regulation of Cell Division (Ch. 12) Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development &
Regulation of Cell Division (Ch. 12) Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development &
Biochemistry of Cancer and Tumor Markers
 Biochemistry of Cancer and Tumor Markers The term cancer applies to a group of diseases in which cells grow abnormally and form a malignant tumor. It is a long term multistage genetic process. The first
Biochemistry of Cancer and Tumor Markers The term cancer applies to a group of diseases in which cells grow abnormally and form a malignant tumor. It is a long term multistage genetic process. The first
Williams Lab Recipes ANTIBIOTICS
 Williams Lab Recipes ANTIBIOTICS 1000x Ampicillin (sodium salt) 100mg/ml recipe 1. Measure out 1 g of Ampicillin tri hydrate 2. Add Milli-Q H2O to 10 ml 3. Add ~.1 g of NaOH pellets (half pellet or more
Williams Lab Recipes ANTIBIOTICS 1000x Ampicillin (sodium salt) 100mg/ml recipe 1. Measure out 1 g of Ampicillin tri hydrate 2. Add Milli-Q H2O to 10 ml 3. Add ~.1 g of NaOH pellets (half pellet or more
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Author's response to reviews
 Author's response to reviews Title: Wnt1 is epistatic to Id2 in modeling mammary gland development and in causing mammary tumors Authors: Susan Marino (smarino@cmgm.stanford.edu) Claire Romelfanger (clairefrog@yahoo.com)
Author's response to reviews Title: Wnt1 is epistatic to Id2 in modeling mammary gland development and in causing mammary tumors Authors: Susan Marino (smarino@cmgm.stanford.edu) Claire Romelfanger (clairefrog@yahoo.com)
Regulation of Cell Division
 Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Regulation of the Cell Cycle
 Regulation of the Cell Cycle 21 I. OVERVIEW Quiescent differentiated cell / can be induced to re-enter the active cell cycle. urvival Cell division Apoptosis 1 Daughter cells Apoptic cell enescent cell
Regulation of the Cell Cycle 21 I. OVERVIEW Quiescent differentiated cell / can be induced to re-enter the active cell cycle. urvival Cell division Apoptosis 1 Daughter cells Apoptic cell enescent cell
mirna Dr. S Hosseini-Asl
 mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region