EVOLUTION OF THE THERIAN MAMMALS IN THE LATE CRETACEOUS OF ASIA PART I. DELTATHERIDIIDAE
|
|
|
- Britney Wilkins
- 6 years ago
- Views:
Transcription
1 ZOFIA KIELAN-JAWOROWSKA EVOLUTION OF THE THERIAN MAMMALS IN THE LATE CRETACEOUS OF ASIA PART I. DELTATHERIDIIDAE (plates XXVIII-XXXV) Abstract. - A Late Cretaceous mammalian family of metatherian-eutherian grade from Mongolia, the Deltatheridiidae, is described.it contains two monotypic genera: Deltatheridium and Deltatheroides. A mandible of De/tatheroides cretacicus GREGORY & SIMPSON from the Djadokhta Formation is described for the first time. Two subspecies of De/tatheridium pretritubercu/are GREGORY & SIMPSON are recognized: the nominal subspecies from the Djadokhta Formation and Deltatheridium pretritubercu/are tardum subsp. n. from the younger Baron Goyot Formation and Khermeen Tsav formation. De/tatheroides cretacicus, De/tatheridium pretritubercu/are pretritubercu/are and Deltatheridium pretritubercu/are tardum form a morphological sequence, De/tatheroides being the most primitive. The snout of De/tatheridium shows resemblance to primitive marsupials, however, the points of resemblance are primitive features. Dental formula of the Deltatheridiidae is. The shearing surfaces of the molars of De/tatheridium are discussed and it ISshown that bothsurfaces and 2 (CROMPTON,1971) were present, with surface 2 slightly larger. The Deltatheridiidae are very different from the known eutherian mammals of the same age of Asia. They probably derive from the Ea rly Cretaceous Aegialodontidae. The forms closely related to the DeItatheridiidae appear to be the Pappotheridiidae from the Alb ian of Texas. Potamote/ses from the Early Campanian ofalberta, classified as a therian incertae sedis, is not related to the DeItatheridiidae. The Late Cretaceous metatherian family Stagodontidae shows some resemblance to the DeItatheridiidae but it is possible that the similarities between the two families are due to the con vergence. The DeItatheridiidae cannot be placed with confidence in either Eutheria or Metatheria (in spite of their metatherian dental formula) ; if they prove to be rnetatherians, they will constitute a specialised-aberrant group. In this paper they are simply classified as a primitive group of therian mammals. The de- - scendants of the DeItatheridiidae are not known. Contents Introduction Acknowledgements Description of the material. Theria of metatherian-eutherian grade Family Deltatheridiidae GREGORY & SIMPSON, 1926 Genus Deltatheridium GREGORY & SIMPSON, 1926 De/tatheridiumpretritubercu/are tardum subsp,n. Shearing surfaces in De/tatheridium..... Genus Deltatheroides GREGORY & SIMPSON, Deltatheroides cretacicus GREGORY & SIMPSON, Discussion Evolution in the DeItatheridiidae Comparison of the DeItather idiidae with other therians of rnetatherian-eutherian grad e. Comparison of the DeItatheridiidae with the marsupials Comparison of the DeItatheridiidae with eutherian mammals of the Late Cretaceous of Asia. References Page
2 104. ZOFIA KIELAN-JAWOROWSKA INTRODUCTION GREGORY and SIMPSON (1926) erected the family Deltatheridiidae, to which they assigned Deltatheridium, Deltatheroides and tentatively also Hyotheridium, all from the Late Cretaceous Djadokhta Formation of Mongolia. The material of all these genera, on which the original description of GREGORY and SIMPSON was based, is housed in the American Museum of Natural History and is scanty and poorly preserved. The poor state of preservation of the teeth in Deltatheridium and Deltatheroides led GREGORY and SIMPSON to the statement that in the De1tatheridiidae the paracone and metacone are (1926, p. 6): "barely if at all separated or very closely 'approximated"; this has subsequently been shown (BUTLER & KIELAN-JAwo ROWSKA, 1973) not to be the case. The postcanine upper teeth were recognised by GREGORY and SIMPSON in Deltatheridium as three premolars and three molars, while in Deltatheroides as four premolars and three molars. A comparison of the original specimens with the drawings of GREGORY and SIMPSON (Text-figs la, B) indicates that the only obvious difference between the two genera is the presence in Deltatheroides of an additional vestigial upper molar, which is evidently absent in Deltatheridium. B c Fig. 1 Comparison of the occlusal views of the upper teeth of A, Deltatheroides ; Band C. Deltatheridium. A and B are redrawn from GREGORY & SIMPSON (1926), C is a reconstruction based on the material of Deltatheridium pretrituberculare tardum from the Barun Goyot Formation. The lower dentition of Deltatheroides was not known and is described for the first time in this paper. In Deltatheroides there are seven lower po stcanine teeth. The postcanine lower dentition of Deltatheridium was recognised by GREGORY and SIMPSON (1926) as three premolars and three molars, however, McKENNA et al. (1972) have shown that in the type specimen of Deltatheridium pretrituberculare there are also seven lower postcanine teeth. GREGORY and SIMPSON had confused the fourth molar with an enlarged talonid of the third molar. The Deltatheridiidae have been discussed subsequently by vario us authors, the most important being the paper by VAN VALEN (1966), who erected the new order of mammals
3 DELTATHERIDIIDAE 105 the Deltatheridia. VAN VALEN included in the Deltatheridia the Palaeoryctidae, Didymoconidae, Oxyaenidae and Hyaenodontidae and concluded that the family Deltatheridiidae as defined by GREGORY and SIMPSON (1926) was a junior synonym of the Palaeoryctidae. He placed Deltatheridium and Hyotheridium in a subfamily Deltatheridiinae and Deltatheroides in the Didelphodontinae and placed both these subfamilies in the Palaeoryctidae. McKENNA et al. (1972) and SZALAY and McKENNA (1971) revised the genus Deltatheridium. They included in the Deltatheridiidae Deltatheridium and two Mongolian Paleocene genera: Sarcodon (= Opisthopsalis) and Hyracolestes and removed from the Deltatheridiidae Deltatheroides and Hyotheridium. The dental formula of the Deltatheridiidae has been re-? cognised by SZALAY and McKENNA (1971) as 2142' SZALAY and McKENNA (1971) stated in the diagnosis of the Deltatheridiidae that the last premolar is somewhat molariform, while in fact the fourth postcanine teeth both in the upper and lower jaws are in Deltatheridium completely molarized and do not differ from subsequent molars. BUTLER and KIELAN-JAWOROWSKA (1973) have shown that the dental formula of the Deltatheridiidae (to which they assigned only two Late Cretaceous Asian genera Deltatheriduim.? basi. 1 and Deltatheroides) IS? ' and claimed that It agrees better WIth asic marsupia formula than with the eutherian formula. In spite of the marsupial dental formula we did not classify the Deltatheridiidae as the marsupials, but as Theria of metatherian-eutherian grade. The same approach is followed in the present paper. The Cretaceous therian mammals were known from Asia until recently only from the Djadokhta Formation (GREGORY & SIMPSON, 1926; KrnLAN-JAwOROWSKA, 1969). The age ofthe Djadokhta Formation has been estimated on the basis ofmultituberculates as?coniacian or Santonian (KrnLAN-JAWOROWSKA, 1970), but on the basis of further studies of multituberculates I now think that it is more probable that the Djadokhta Formation is of Santonian age. In 1970 and 1971 in Asia new Late Cretaceous mammalian fauna was discovered by the Polish-Mongolian Palaeontological Expeditions (KIELAN-JAWOROWSKA& BARSBOLD, 1972) in the Barun Goyot Formation (previously called also Lower Nemegt Beds), the age of which has been estimated as? Middle Campanian (KIELAN-JAWOROWSKA, 1974) and in its stratigraphic equivalent informally called Khermeen Tsav formation at the localities of Khermeen Tsaw I and Khermeen Tsav Il (see KIELAN-JAWOROWSKA, 1975). In 1969 the Soviet-Mongolian Palaeontological Expedition discovered in the Gobi Desert in the locality of Khovboor a new mammalian fauna of Early Cretaceous (Aptian or Albian) age (TROFIMOV, 1972). In addition to the representatives.of the Triconodonta, Symmetrodonta and Multituberculata, this fauna contains true eutherian mammals, which are the presumable ancestors of two Djadokhta Formation genera: Kennalestes and Zalambdalestes. The representatives of the Deltatheridiidae have not been reported from the Early Cretaceous of Mongolia (TROFIMOV, 1972, BELIAJEVA et al., 1974). During my visit to the Palaeontological Institute of the USSR Academy of Sciences in Moscow in June of 1974, thanks to the courtesy of Dr. B. A. TROFIMOV I was allowed to examine the collection of Early Cretaceous mammals from Khovboor, housed in this Institute. In this collection I have seen a mandible with two molars (PIN ), which in my opinion is similar to Aegialodon from the Lower Wealden of Great Britain (K. A. KERMACK et al., 1965). This specimen will be described by Dr. B. A. TROFIMOV. Also Dr. D. DASHZEVEG kindly informed me that in the collection from Khovboor, assembled by him in 1964 and housed at the Laboratory of Stratigraphy and Palaeontology, Geological In stitute of the Academy of Sciences of the
4 106 ZOFIA KIELAN-JAWOROWSKA Mongolian People's Republic in Ulan Bator, there is a single lower molar? M 2 similar to Aegialodon. It seems possible that the Aegialodontidae are the ancestors of the Deltatheridiidae (see p. 118). The terminology used in the present paper for describing the elements of the tooth structure is that of VAN VALEN (1966) and SZALAY (1969), while the shearing surfaces are described according to terminology introduced by CROMPTON (1971). SZALAY (1969) de-. signated the cingulum extending from the paraconule antero-labially the paracingulum, while he named the ridge on the anterior edge of the paracingulum the preparaconule crista. The paracingulum may extend labially to the parastyle, or it may fade out before reaching the para style, as is the case in Deltatheridium. In the majority of primitive mammals on the parastylar region there is a groove which extends from the parastyle upwards and inwards, along which the protoconid moves. I shall refer to this as the groove for the protoconid (Text-fig. 2 A). If the paracingulum is long and meets the parastyle, the groove for the protoconid crosses the paracingulum. In more mature specimens this groove is extensively worn (Text-fig. 2 B). as prdg pplc \-J 4 pa pacg pac m~ 1a A 8 Fig. 2 Comparison of the oblique anterior views of M2 of A, De/tatheridium pretrituberculare tardum (ZPAL MgM-Ij91) and B, M" of? Ge/astops sp. (MCZ 19734) to illustrate the structure of the groove for the protoconid. Not to scale. For abbreviations see p, 107 pas sf The following abbreviations are introduced here to indicate the directions of described elements: long. -longitudinal, parallel to the plane of symmetry of the animal and tr. transverse, perpendicular and at right angles to the plane of symmetry. Abbreviations used for institutions: AMNH - American Museum of Natural History (New York). MCZ - Museum of Comparative Zoology, Harvard University (Cambridge, USA). PIN - Palaeontological Institute, USSR Academy of Sciences (Moscow). SMP-SMU - Shuler Museum of Paleontology, Southern Methodist University (Dallas). UA - University of Alberta (Edmonton). ZPAL - Palaeozoological Institute of the Polish Academy of Sciences (Warsaw).
5 DELTATHERIDIIDAE 107 Abbreviations used in figures: "c" cusp "c" (CROMPTON, 1971) end entoconid hyd hypoconid hyld hypoconulid me metacone med metaconid mtl metaconule pa paracone pac paracrista pacg paracingulum pad pal pas pplc pr prd prdg st 1a, I b-5 paraconid paraconule parastyle preparaconule crista protocone protoconid groove for the protoconid stylocone matching shearing surfaces ACKNOWLEDGEMENTS The present paper has been written in part at the Palaeozoological Institute of the Polish Academy of Sciences in Warsaw and completed during my stay in the Museum of Comparative Zoology, as a visiting Alexander Agassiz professor at Harvard University in Cambridge, USA, in I would like to express my deep gratitude to Professor A. W. CROMPTON, Director of the Museum of Comparative Zoology at Harvard, who during my stay in Cambridge has.always been willing to both assist me and to discuss questions connected with my work. He kindly read the manuscript of the present paper, and made valuable suggestions. Harvard University provided me with good working facilities during my stay; some of the drawings published in this paper (Text-figs 2; 6, 8) and some of the photographs (PIs XXIX, XXX (partly), XXXI, XXXlI and XXXV (partly) were made by Mrs. P. CHAUDHURI and Mrs. A. COLE MAN, respectively, both from the staff of the Museum of Comparative Zoology. The scanning electron microscope photographs were taken by Mr. G. R. PIERCE of Harvard University. The remaining drawings were made by Mrs. K. BUDZYNSKA, and the photographs by Mr. W. SKAR ZYNSKI both from the Palaeozoological Institute in Warsaw. I wish also to thank Professor W. A. CLEMENS (University of California, Berkeley), Professor R. C. Fox (University of Alberta, Edmonton), Professor M. C. McKENNA (American Museum of Natural History, New York), Professor R. H. SLAUGHTER (Southern Methodist University, Dallas), Dr. B. A.- TROFIMOV (palaeontological Institute, USSR Academy of Sciences, Moscow) and Dr. W. D. TURNBULL (Field Museum of Natural History, Chicago) for providing me with the opportunity to study the specimens in their charge. I am also grateful to Professor R. C. Fox and Professor M. C. McKENNA for reading a part of the manuscript of this paper and their useful comments. DESCRIPTION OF THE MATERIAL THERIA OF METATHERIAN-EUTHERIAN GRADE Family DELTATHERIDIIDAE GREGORY & SIMPSON, 1926 Diagnosis. - Medium sized Cretaceous mammals, length of the skull varying around 4 cm. Snout shortened, nasals strongly expanded posteriorly, contacting the lacrimals. Lacrimals with large facial wing and large lacrimal foramen, strongly resembling that of primitive marsupials, especially Didelphis. Jugal deep and long. No palatine fenestra. Dental formula ' Canines very large, single rooted. P' single rooted, p2 and p3 double rooted with basal cusps. Premolars not molarized. Stylar shelf of molars very large, with oblique
6 108 ZOFIA KIELAN-JAWOROWSKA grooves extending from paracrista and metacrista to the middle of the ectoflexus. Parastyle and stylocone large, metastylar area large with one style only. Paracone and metacone placed in the centre of the tooth width (tr.), paracone larger than the metacone. Conules large, convex, ' but hardly discernible in worn specimens. Paracingulum present but not very wide, disappearing at a level above the tip of the paracone, metacingulum absent. Groove for protoconid not very wide. Precingulum and postcingulum absent. Protocone small, low and pointed in unworn specimens. Lower molars : protoconid tall, metaconid smaller than paraconid, talonid narrow (tr.) and lies behind the lingual part of the trigonid, weak crest extending from apex of metaconid to cristid obliqua. Included genera: Deltatheridium GREGORY & SIMPSON, 1926, and Deltatheroides GREGORY & SIMPSON, Stratigraphic and geographic range. - Late Cretaceous of Mongolia. Genus DELTATIIERIDIUM GREGORY & SIMPSON, 1926 Type species: Deltatheridium pretrituberculare GREGORY & SIMPSON, 1926, the only species known, within which the two subspecies are recognised in this paper: Deltatheridiumpretrituberculare pretrituberculare GREGORY & SIMPSON, 1926, and Deltatheridium pretrituberculare tardum subsp, n.? Generic and specific diagnosis, - Dental formula Narrow spaces between C_pl and Pl ~P2. Conules large, rounded, on M2 labial to the paraconule a small accessory conule is present on the paracingulum. Deep notch in the metacrista, just labial to the metacone base, smaller one on the paracrista. PI very small, with two roots arranged obliquely with respect to the length of the jaw. Carnassial notch present on the paraconid protoconid blade (paracristid) on all the molars. M 4 with double cusped trigonid (metaconid absent) and vestigial talonid 1. Stratigraphic and geographic range. - Known only from the Djadokhta Formation (locality of Bayn Dzak), from the Baron Goyot Formation (locality of Nemegt), and from the Khermeen Tsav formation, stratigraphic equivalent of the Baron Goyot Formation (locality of Khermeen Tsav II), all in the Gobi Desert,' Mongolia. Deltatheridium pretrituberculare tardum subsp. n. (Pis XXVIII-XXXIV, XXXV, Fig. 2; Text-fig. 1B, C, 2A, 3, 5, 6, 7B, 8 left) Holotype: ZPAL MgM-I/91, strongly compressed and damaged anterior part of the skull of a young individual, associat ed with incomplete right and left mandibles. Left C-M, right MI-M', left P,-M., right I-M. and roots of M. Type horizon and locality : Late Cretaceous (? Middle Campanian), Kherrneen Tsav formation, stratigraphic equivalent of the Barun Goyot Formation, Khermeen Tsav 11, Gobi Desert, Mongolia. Derivation of the name : Lat. tardum-iate, occurring later than the nominal subspecies.? iff. I.. h. Diagnosis. - Dental formula DJ ers from the nomina subspecies In avmg a somewhat shorter skull and only one lower incisor, whereas there are two in D. pretrituber- \ culare pretrituberculare. 1 It appears from the investigation on the development of cheek teeth in some modern marsupials (ARCHER, 1972 that P2 are lost during the ontogenetic development and the premolars retained are PI, P3 and P4. Because there is no data as to which teeth were lost in the ontogeny of the Deltatheridiidae, and as the homology of their teeth with those of the marsupials is not clear, I regard it reasonable to designate the premolars in the Deltatheridiidae in the traditional way, i.e., PI, P2 and P3.
7 DELTATHERIDIIDAE 109 T able 1 Dimensions of Deltatheroides cretacicus and two subspecies of Deltatheridium pretrituberculare in mm. Species Deltat heroides cretacicus Deltatheridium pretri tuberculare pretrituberculare Deltatheridium pretrituberculare tardum Format ion Djadokhta Formation K herrneen Tsav formation Barun Goyot Formation Locality Bayn Dzak Khermeen Tsav 11 Nernegt Mu s. cat. No. ZPAL AMNH ZPAL ZPAL MgM-I / MgM-I/91 MgM-I/102 MgM-I/60 MgM-I/136 Width across upper canines? Width acro ss posterior margins ofinfrao rbital foramen 10.8 Length ofthe palatal part of maxilla (suture with premaxilla-transverse suture) 9.5 Width of the palate - acro ss M' (between the root s)? pl_p3 ant-post, PC_M" ant.-post. (along median cusps) pc ant.-post tr J.O M' ant.-post tr, Depth of the lower jaw beneath M. (labial) 5.5? Pi-M i ant.-post. 12.0?1 l.o 8.3 Mi-M 3 ant-post. 7.5 P, ant.-post tr M, ant.-post. ext. 3.3? tr, P 3 ant.-post. ext tr
8 110 ZOFIA KIELAN-JAWOROWSKA Material. - In addition to the type specimen, there are four specimens: ZPAL MgM-Ij60, Barun Goyot Formation, Nemegt, Red Monadnocks; strongly damaged rostral part of the skull, with left C, P' and pz and right P3_M3. ZPAL MgM-I/102, Khermeen Tsav formation, Khermeen Tsav II; rostral part of the skull with C-M3 preserved on both sides. ZPAL MgM-I/1l3, Barun Goyot Formation, Nemegt, Eastern Sayr; badly damaged fragment of left mandible with Mz-M4 ZPAL MgM-I/136, Barun Goyot Formation, Nemegt, Eastern Sayr; fragment of right maxilla with MI_Ms, associated with fragment of the left mandible with roots of M 1, Mz-Ms. Geographic and stratigraphic range. - Known only from the type horizon, from the locality Khermeen Tsav II and from the Barun Goyot Formation, from the locality of Nemegt, Gobi Desert, Mongolia. Dimensions. See Table I. Description. - The skull (Text-fig. 3). On the basis of ZPAL MgM-I/102, which is not distorted, one can state that the snout, which is narrow in the anterior part, widens strongly in the region of P3. The anterior margin of the orbit is placed opposite the P3_MI embrasure. The dorsal border of the maxilla in lateral view is strongly vaulted, being the highest above PZ. The infraorbital foramen in MgM-I/I02 is 1.3 mm long, situated above the anterior part of P", surrounded posteriorly by the thickened rim of the maxilla; it is higher than figured by GREGORY and SIMPSON (1926, Fig. 8) in D. pretrituberculare pretrituberculare. The infraorbital Fig. 3 Deltatheridium pretriturberculare tardum: A, (ZPAL MgM-I/I02) reconstruction of the anterior part of the skull in lateral view and B, reconstruction of the upper dentition ofthe same species in crown view, based on ZPAL MgM-I/I02 and /91.
9 DELTATHERIDIIDAE 111 canal is about 2.7 mm long. The nasals are moderately wide anteriorly, become narrower opposite p2 and then expand strongly laterally to reach the lacrimals. The naso-frontal suture in studied specimens is not preserved. The lacrimal has a comparatively large exposure on the lateral wall of the face, being somewhat pointed at the anterodorsal corner, between the nasal and maxilla. The lacrimal foramen in MgM-Ij102 is 1.1 mm long and is situated in the emargination of the bone, the anterior part of which, i.e., the part in front of the foramen lies more external than the posterior part. The posterior border of the lacrimal within the orbit is not visible, but its ventral border extends posteriorly along the upper and inner parts of the zygomatic arch, as far back as the posterior part of :P4. The jugal extends anteriorly, to the level of the anterior margin of the lacrimal, the distance between its anterior margin and the infraorbital foramen in this specimen is 0.9 mm. In lateral view, above MI and M2 the jugal occupies the upper half of the depth of the zygomatic arch. The zygomatic arch appears to be deeper than reconstructed by GREGORY and SIMPSON (1926, Fig. 8), for D. pretrituberculare pretrituberculare. Palatal region. Palatine fenestra are absent. The palatal surfaces of maxillae are moderately concave. The transverse part of the palatal bone is roughly rectangular, the transverse palatine suture is situated opposite MI_M2 embrasure. The anterior palatine foramen is placed in the middle of the length (tr.) on each side of the transverse suture. In front of it a faint palatine groove, which narrows and shallows anteriorly, extends approximately to a position opposite to P3. Posterior palatine foramina are visible on both sides of MgM-Ij102, smaller than the anterior ones and situated just behind them. Mandible. The mandibles which are preserved in ZPAL MgM-Ij91 (young individual) and in MgM-Ij136 (fragment), vary strongly in size, but otherwise are similar to those in D. pretrituberculare pretrituberculare. The mental foramina cannot be seen. Dentition. Upper teeth. The examination of the type, specimen of D. pretrituberculare pretrituberculare (AMNH 21706) confirmed VAN V ALEN'S view that there are four alveoli for the incisors preserved on the left side of the specimen, while on the right one only three are discernible, but the first one may not be preserved due to the damage of this part of the.. premaxilla (Text-fig. 4). The number of incisors in D. pretrituberculare tardum cannot be ascertained; as they are not preserved in the studied specimens from ZPAL collections, except for one fragmentary root in front of the canine on the left side of ZPAL MgM-IjI02. The canine is large and curved and in ZPAL MgM-I/91 it is 4.8 mm long and in MgM-Ij60 is 5.3 mm long. P' is small and single-rooted, directed obliquely anteriorly, with a small posterior basal cusp. There is a narrow gap 0.4 mm long in MgM-Ij102 between C and P', and 0.55 mm long between P' and P2. p2 and p3 are double-rooted, transversely compressed, P2 lower than P3. p2 has a small anterior cingulum and comparatively large posterior cusp. P" is similar in shape to p2, but larger and higher. This is the highest of the postcanine teeth. The anterior cingulum is more strongly developed than in p2, the posterior basal cusp is larger, surrounded by a distinct ridge. The posterior ridge of the main cusp meets the lingual side of the basal cusp. MI is roughly triangular, slightly longer than wide, asymmetrical because of the stronger development of the metastylar region. In labial view MI is lower than M2. The ectoflexus is very deep and the stylar shelf very wide, due to the lingual (central) position of the paracone and metacone. The metastylar region is strongly elongated posterolaterally with a single cusp (metastyle), situated on its postero-lateral corner. The parastylar region is in occlusal view smaller than the metastylar region and three styles: para style, stylocone and poorly developed, unnamed style, designated here as style BI (BENSLEY, 1906), adhering tightly to the stylocone are present. The latter cannot be designated as a mesostyle (style C) as it is situated in the
10 112 ZOFIA KIELAN-JAWOROWSKA parastylar region and not in the middle ofthe ectoflexus as the mesostyle usually is. The stylocone is situated more lingually than the two remaining styles of the parastylar region. Along the middle of the ectoflexus small and indistinct crenulations, visible only on unworn specimens are present. The paracrista and metacrista are developed as very strong ridges. On the metacrista. labially to the base of the metacone there is a deep notch. A similar, but shallower CJl 3 3 Fig. 4 Deltatheridium pretrituberculare pretrituberculare: (AMNH 21706). Anterior part of the snout in palatal view. showing four alveoli for the incisors on the left side. notch is present on the paracrista labially to the base of the protocone. Due to the powerful development of the metastylar region, the metacrista is much longer than the paracrista. The stylar area in occlusal view consists of three surfaces: anterior, central and posterior. The central is separated from the anterior and posterior by oblique grooves, which extend from the notches on the paracrista and metacrista to the deepest part of the ectoflexus. The shearing surface above the metacrista is vertical and is not visible in occlusal view. The shearing surface above the paracrista is directed obliquely a nd can be seen in occlusal view. The paracone and metacone are situated in the middle of the tooth width (tr.), the paracone lies slightly more lingually than the metacone and they have closely approximated bases, but their tips are widely separated. The paracone is higher than the metacone. The conules are comparatively large, swollen cusps, well discernible only in young specimens. The paracingulum extends from the paraconule along the anterior wall of the tooth, it is distinct and fades out at (or slightly beyond) a level above the tip of the paracone. The preparaconule crista is absent. The groove for the protoconid is well developed, widest (long.) in its middle part, disappearing opposite the paracone. A metacingulum is not present. The protocone is small and situated much higher than the paracone and metacone. The precingulum and postcingulum are absent. The lingual part of the tooth is very narrow (long.) in respect to the wide (long.) labial part. The upper anterior and po sterior margins of the crown, i.e., the parts in contact with the maxilla, are strongly concave in the middle. In labial view: the parastylar region is large and has a form of a three-lobed petal; the ventral surface of the metastylar region labial to the metacone is strongly concave; the metacone is visible and the paracone is partly obscured by the descending parastylar region.
11 DELTATHERIDIIDAE 113 M~ is very similar to MI but is larger and more symmetrical, the parastylar and metastylar regions are almost equally developed. The paracrista in M2 in much longer than in MI. In ZPAL MgM-Ij91 on the right M2, labially to the paraconule, on the paracingulum an extra conule, similar to that in Pappotherium is present. On the left M2 of the same specimen the paracingulum in this region is damaged, while in all other specimens the teeth are worn and the conules are hardly discernible. The incurvature on the crown-maxilla boundary between the roots, along both embrasures, is shallower than in M', the paracingulum is poorly developed, but more prominent than in MI, the groove for the protoconid is wider than in.m'. Otherwise all the details of the structure of M2 are the same as in MI ~ M" is strongly asymmetrical, with a very large parastylar region elongated into a narrow process and the metastylar region hardly developed. The parastylar region is bordered labially by a prominent ridge, and the para style and stylocone are hardly discernible. The paracone and metacone are situated in the middle of the tooth width, the paracone being much higher than the metacone. The paracrista is prominent, with a distinct notch, the paracingulum faint and weaker than in M2. The paraconule is well developed and the metaconule not discernible. The groove for the protoconid is as wide as in M2. Due to the lack of metastylar region, the metacrista is poorly developed, but the remaining details of the tooth structure are the same as in M2. The embrasures between MI_M2and M2_Ma are very deep and the maxilla forms a highly domed roof. Lower teeth (Text-fig. 5). In the right mandible of ZPAL MgM-Ij91, in front of the broken canine, the root of a single incisor is present. This incisor was semi-procumbent and oval in outline. The anterior margin of the mandible is preserved, and because there is apparently no space for more than one incisor, it appears that a single lower incisor is characteristic of D. pretrituberculare tardum. In other specimens the lower incisors are not preserved. This is the only specimen in which the canine is preserved and its tip is damaged. It is a large tooth, oval in outline and directed vertically and slightly forwards. PI was present in the right mandible but was damaged and lost during preparation. It was a very small, cone-like, double-rooted tooth, with roots almost perpendicular to the longitudinal axis of the mandible so that the lingual root lies further anteriorly than the labial. This is a result of the narrow space between C and P2. A similar condition is present in Deltatheroides cretacicus (ZPAL MgM-Jj29). P z is cone-like, double-rooted (as are all that follow), is much taller than PI> has a prominent main cusp and a comparatively large posterior basal cusp. The basal cusp (in lateral view) projects posteriorly beyond the base of the posterior root. Along the posterior edge of the main cusp there is a crest, which reaches the middle of the posterior cusp. P, is higher than P2, has a large anterior cusp and a small incipient talonid. The anterior cusp is tall and piercing and wide at the base. On the posterior edge of the anterior cusp there is a crest, which reaches the incipient talonid on the labial side and continues along the length of the talonid as the cristid obliqua, but the latter is directed, in a longitudinal rather than an oblique direction. Labially to the cristid obliqua the talonid slopes downwards, while lingually it is slightly basined. This basined part, placed on the lingual side of the tooth, corresponds to the talonids of M1-Ma, which in these teeth lie behind the labial parts of trigonids. Cusps on the incipient. talonid cannot be recognised. M1 is somewhat smaller than M2 and Ma. The trigonid is larger (long. and tr.) than the talonid, which is distinctly narrower than the trigonid' and placed rather lingually. The protoconid is the highest and largest of the trigonid cusps, the metaconid the smallest. The paraconid and metaconid are well separated by a wide furrow. On the anterior edge of the 8 - Palaeoritologia Polonica No. ~3
12 114 ZOFIA KIELAN-JAWOROWSKA paraconid there is a basal cingulum, directed obliquely downwards. The paraconid and protoconid anterior surfaces form a cutting blade, with a distinct carnassial notch in the centre of the paracristid. On the labial side, the base of the protoconid slightly overhangs the bases of the roots. Posterior wall of the protoconid and metaconid are not vertical but slightly oblique. Along the posterior wall of the metaconid a faint crest (metaconid crest) extends 5mm Fig. 5 Deltatheridium pretrituberculare tardus: reconstruction of the mandible and dentition in occlusal, outer and internal. views; based on ZPAL MgM-I/91 and MgM-I/136. from the tip of the metaconid downwards to the cristid obliqua. This crest separates shearing surfaces 1 and 5. The surface of the talonid is in all specimens strongly worn but the three talonid cusps entoconid, hypoconulid and hypoconid can be recognised. A hypoconulid is situated at the very end of the talonid. The hypoconid and entoconid are arranged symmetrically on both sides of the hypoconuiid and fairly distant from it. The floor of the talonid basin falls away rapidly on the lingual side and an entocristid is not present. Consequently
13 DELTATHERIDIIDAE 115 a well developed talonid basin is not present. The hypoflexid for the reception of the paracone is prominent and shearing surfaces 1 and 3 are well developed. M 2 is the largest of all the cheek teeth, M 4 very strongly reduced. M 2 is similarly shaped as M 1, but differs from it in having more robust protoconid, which projects more strongly labially over the base of the tooth. The metaconid is more strongly separated from the paraconid than in M 1, the prefossid being somewhat larger. The relative size of the talonid is the same as in M 1 The talonid projects beyond the posterior root of the tooth. M, differs from M 2 in being slightly smaller and in having the protoconid more robust in relation to the other cusps. M 4 is very small and shorter (both tr. and especially long.) than M, and M 2 The trigonid consists of two cusps ' (protoconid and paraconid), and the metaconid is absent. The protoconid is higher than the paraconid and does not overhang the root labially as in Ma. The talonid is rudimentary and cannot be seen in the labial view, because it is hidden behind the coronoid process. It is narrow, situated lingually and lacks clearly defined cusps. Variability. - The specimens assigned here to D. pretrituberculare tardum differ considerably in size. ZPAL lyigm-ij136 is about 1 5 times larger than MgM-Ij91 in respect to the depthof the lower jaws. MgM-Ij91 belongs to a young individual in which M, is just erupting. The differences between these specimens concern not only the size of the bones but also of the lower teeth, which in the larger jaw are on the average about 1.3 times larger than in the smaller specimen. The differences between the upper teeth are less striking, the upper teeth in the specimens MgM-Ij136 andj60 are on the average 1.2 times larger than in MgM-Ij91, while the specimen MgM-Ijl02 is intermediate between them. As no differences except the size are observed between the studied specimens and all the details of their tooth structure are identical, I regard all the specimens discussed here as conspecific. SHEARING SURFACES IN DELTATHERIDIUM Deltatheridium (Text-fig. 6) has strong shearing along both the anterior and posterior walls of the molars. The anterior walls of the upper molars are oriented obliquely with regard to the median suture. On the anterior wall of the upper molars one can recognise the shearing surface la (CROMPTON, 1971). The surface Ibis hardly recognisable because of the poor development of the paracingulum. The paracrista, forming the leading edge of the shearing surface 1a, is a very strong ridge with a distinct but not very deep carnassial notch. The main shearing along the anterior wall of the upper molars was on the surface 1a. On the posterior wall of the trigonid, there is a faint ridge extending from the tip of the metaconid to the cristid obliqua; this separates shearing surfaces 1 and 5. Shearing surface 1 is not divided into two parts as in Aegialodon. The posterior wallof the upper molars labial to the metacone is vertical and it forms the strong shearing surface 2, which shears against the paraconid-protoconid blade (paracristid) on the lower molar. On the leading edges of shearing surfaces 2 on both the upper and lower molars there are very deep carnassial notches. The presence of these notches, characteristic for carnassial teeth (see CROMPTON, 1974, Fig. 9) indicates that although in Deltatheridium the shearing took place both along the anterior and posterior walls of the molars, shearing surface 2 was slightly larger as in marsupials. This conclusion is supported by study of the type of wear on the paracrista and metacrista in Deltatheridium. The shearing surfaces 3 and 4 on either side of the hypoconid are rather small, 3 somewhat larger than the 4. Matching surfaces I are present on the lingual slopes of the paracone and metacone. The protocone is comparatively small and pointed. The talonid is small and lacks 8"


14 116 ZOFIA KIELAN-JAWOROWSKA a lingual wall, consequently shearing surface 6 is not present. The preprotocrista and postprotocrista do not play an important part in shearing in Deltatheridium, whereas the paracrista and metacrista are long and form the leading edges of the main shearing surfaces. A OCCLUSAL B CROWN st 1mm pad c D pr 5 POST M2 2 ANT M2 Fig. 6 Diagrammatic drawings of the upper and lower molars of Deltatheridium pretrituberculate tardum in four different views to illustrate the shearing surfaces: A, occlusal views of M' and M.; B, crown views of M', M' and M.; C, anterior view ofm' and posterior of M.; D, posterior view of M' and anterior view of Ma- The bold arrows on Figs C and D indicate the direction of movement of the lower teeth against the upper. For abbreviations see p. 107.
15 DELTATHERIDIIDAE 117 G enus DELTATHEROIDES GREGORY & SIMPSON, 1926 Type species: Deltatheroides eretaeieus GREGORY & SIMPSON, 1926, the only known species of the genus.? 1 34 Generic and specific diagnosis. - Dental formula? Resembles strongly Deltatheridium in size, proportions and structure of the dentition. Differs from Deltatheridium in presence of vestigial M4 (absent in Deltatheridium) and in the structure of M4, which has a completely developed trigonid, with metaconid present (absent in Deltatheridium) and normally developed talonid (Text-fig. 7). Stratigraphic and geographic range. - Known only from the Late Cretaceous (?Santonian) Djadokhta Formation of Bayn Dzak, Gobi Desert, Mongolia. Deltatheroides cretacicus GREGORY & SIMPSON, 1926 (pi. XXXV, Fig. 1; Text-figs 1A and 7A) 1926 Deltatheroides eretacieus sp.n.; GREGORY & SIMPSON, PI. 11, Figs le, 9, 10. Material. - AMNH partial face with badly damaged dentition, described and figured by GREGORY and SIMPSON, ZPAL MgM-I/29 incomplete left mandible with roots of canine and of PI and somewhat damaged P 2-M4. Both specimens are from the Djadokhta Formation of Bayn Dzak. Dimensions. See Table 1. Description and discussion. - The upper dentition in the AMNH specimen is strongly damaged and does not merit description. No differences from the upper dentition of Deltatheridium pretrituberculare (both subspecies) are observed, except for the presence in Deltatheroides cretacicus of vestigial M4. Fig. 7 Comparison of the internal views of M 4 of: A, Deltatheroides eretacieus and E, Deltatheridiumpretritubereulare tardum, not to scale. 8 The mandible ZPAL MgM-I/29 found in Bayn Dzak is assigned to Deltatheroides cretacicus for the following reasons : the upper molars of Deltatheridium and Deltatheroides are almost identical and therefore one would expect that the similar lower molars would be present in both genera; the lower molars in ZPAL MgM-I/29 are almost identical to those of Deltatheridium; in Deltatheridium M4 is absent, M4 has a reduced trigonid, without a metaconid and a strongly reduced talonid; in MgM-I/29 M4 has a three-cusped trigonid and a metaconid which sheared against M'{present in Deltatheroides);the mandible of ZPAL MgM-I/29 agrees in size with AMNH
16 118 ZOFIA KIELAN-JAWOROWSKA DISCUSSION EVOLUTION IN THE DELTATHERIDIIDAE Comparing the teeth structure of Deltatheroides cretacicus and two subspecies of Deltatheridium pretrituberculare one can demonstrate that the three forms are very closely related. Deltatheroides cretacicus and Deltatheridium pretrituberculare pretrituberculare are contemporaries, but Deltatheroides cretacicus has a structure from which Deltatheridium pretrituberculare pretrituberculare could be derived. Deltatheridium pretrituberculare tardum was found in younger formation than the two preceeding forms. In the evolution in this group shortening of the skull took place and this was correlated with the loss of anterior and posterior teeth. The most primitive member of the family is Deltatheroides cretacicus, which has four upper and lower molars, M 4 with completely developed trigonid, while M4 is somewhat reduced. The next in the structural sequence is Deltatheridium pretrituberculare pretrituberculare which has two lower incisors (unknown in Deltatheroides), and which has lost M4, while in M 4 the talonid is more reduced than in Deltatheroides. The last form in the sequence is Deltatheridium pretrituberculare tardum in which there is only one lower incisor, and M 4 is extremely reduced. The trigonid consists of only two cusps (protoconid and paraconid), the metaconid being absent, and the talonid is extremely small and lacks the typical cusps. The details of the structure of M 4of Deltatheridium pretrituberculare pretrituberculare are unknown because in AMNH specimen this tooth is very badly damaged. However, in Deltatheridium pretrituberculare pretrituberculare M4 is absent and one can presume the loss of the metaconid in M 4 in this subspecies, because M 4 sheared only against the metacrista of M3. COMPARISON OF THE DELTATHERIDIIDAE WITH OTHER THERIA OF METATHERIAN-EUTHERIAN GRADE The most primitive known therian mammal is Aegialodon represented by a single lower molar from the Lower Wealden of Great Britain (K. A. KERMACK et al., 1965). The forms similar the Aegialodon are now known from the Early Cretaceous of Asia, although they have not been described as yet (see p. 105). Also a lower molar from the Albian of Texas, identified as Trinity molar type 6, SMP-SMU (SLAUGHTER, 1971) is approximately at the same level of organization as Aegialodon, although has ' a slightly longer talonid. All these forms could possibly be placed in the same family the Aegialodontidae. The Aegialodontidae are similar to the Deltatheridiidae in that the paraconid and metaconid are of the same height or the paraconid is slightly larger and both are considerably shorter than the protoconid; the talonid is narrow (bucco-lingually), is lingually placed and lacks a medial wall. However, in the Aegialodontidae the molars are smaller than in the Deltatheridiidae and have a smaller talonid basin, which indicates that in the upper molars the protocone was smaller than in the Deltatheridiidae. A primitive feature of the Aegialodontidae is the presence of both an anterior buccal cingulum and an anterior lingual cusp or ridge. The lingual cusp or ridge is lost in the Deltatheridiidae, although it is retained in some Cretaceous mammals (e.g. in some marsupials). It seems possible that the Aegialodontidae are the ancestors of the Deltatheridiidae. PATTERSON (1956) described several isolated lower fragmentary upper molars from the
17 DELTATHERIDIIDAE 119 Trinity sandstone (Albian) of Texas, which he classified as Theria of metatherian-eutherian grade. Slaughter (1965) described from the same beds but a different locality last two upper molars which he designated Pappotherium pattersoni, and for which he created the family Pappotheriidae within the subclass Theria. In the same paper he described several lower molars from the same beds, without naming them. The Trinity therians have been subsequently described by SLAUGHTER (1968a, 1968b, 1968c, 1971), CROMPTON (1971) and TURNBULL (1971). The upper molars of Pappotherium strongly resemble those of Deltatheridium (Text-fig. 8), although Pappotherium is much smaller than Deltatheridium. The parastylar region has the same form in both genera, the parastyle and the styloco ne in both cases are prominent, the difference is that in Deltatheridium an extra cusp, closely adhering to the posterior surface of the stylocone is present; this gives the parastylar region in labial view the appearance of a three-lobed petal, while in Pappotherium this petal has only two lobes. The groove for the protoconid in Pappotherium is wider than in Deltatheridium, especially in its most labial part. In both genera the paracingulum does not meet the groove. The ectoflexus is strongly concave in both genera and there are indistinct crenulations along its margin, the mesostyle (stylar, cusp C) is not present. The metastylar region is large and strongly elongated postero-labially in Deltatheridium and supports a single cusp (metastyle) whereas in Pappotherium it is less elongated and supports two cusps. In both genera on the stylar shelf one can recognise three surfaces, separated by grooves extending from the lingual corners of the paracrista and metacrista towards the middle part of the ectoflexus, On the metacrista in Pappotherium a small cusp "c" (CROMPTON, 1971) is present, while in Deltatheridium there is a carnassial notch on the metacrista. The paracone and the metacone are similarly shaped, in both genera the paracone being larger and placed more lingually and in both the paracone and the metacone are partially concave on their labial faces. Conules are present in both genera; they may be slightly larger in Deltatheridium but in both cases they are not winged (i.e., lack a postparaconule crista). In one M2 of Deltatheridium pretrituberculare tardum an extra conule is present on the paracingulum, labially to the paraconule, as in Pappotherium. The groove for the protoconid in Pappotherium is wider than in Deltatheridium, especially in its labial part. Also, the paracingulum is wider in Pappotherium than in Deltatheridium, but in both genera it does not meet the groove for the protoconid. The protocone is small and pointed and at a horizontal level considerably higher than the paracone in both genera. The lingual side of the tooth is relatively narrower (long.) in Deltatheridium than in Pappotherium. The lower teeth of Pappotherium are not known. CROMPTON (1971) has chosen a lower molar SMP-SMU from the Trinity sandstone as a possible lower molar of Pappotherium, while SLAUGHTER (1971) has tentatively assigned a left lower jaw fragment SMP-SMU to Pappotherium. The mandible fragment is according to SLAUGHTER an eutherian because a replacing tooth for the penultimate premolar is preserved. The lower molars chosen by CROMP TON and SLAUGHTER as possibly belonging to Pappotherium differ considerably from each other and I think that it is impossible to demonstrate unequivocally that either of them matches the upper molars of Pappotherium. The study of the structure of the upper molars of Pappotherium and Deltatheridium shows that in both genera both shearing surfaces I and 2 are used. However, in Pappotherium the main shearing was on surface I, whereas in Deltatheridium it was on surface 2. In this respect Deltatheridium is somewhat more similar to marsupials than Pappotherium. If the relative sizes of the shearing surfaces in Deltatheridium and Pappotherium described above are correct, one would expect that the lower molars of Pappotherium may differ from those of Deltatheridium, and the former may have a paraconid smaller than the
18 120 ZOFIA KIELAN-JAWOROWSKA A B ~ 5 la c D B1 2 E 5 pr d 9 pas pr \.~, Fig. 8 Comparison of M2 of Deltatheridium pretrituberculare tardum (left) (ZPAL MgM-I j91) and Pappotheriumpattersoni (right) (SMP-SMU )in A, crown, B, anterior, C, posterior, D, external and E, internal views. For abbre viation see p. 107.
19 DELTATHERIDIIDAE 121 metaconid. In spite of considerable similarity between the upper molars of Pappotherium and Deltatheridium, they probably differ slightly in the shearing mechanism and I therefore think it reasonable to leave them for the time being in separate families. The structure of the upper molars in these two genera suggests-that they possibly share a common ancestor. Another genus resembling Deltatheridium in certain respects and classified as a therian incertae sedis is Potamotelses from the Early Campanian Upper Milk River Formation of Alberta (Fox, 1972). Potamotelses upper molar has a large stylar region, the paracone is somewhat larger than the metacone and it is placed slightly more lingually, the protocone is small and situated at a level far above that ofthe paracone. The stylar region is proportionally shorter (tr.) than in Deltatheridium, especially the metastylar region. In both genera there is only one cusp - metastyle on the metastylar region. The parastylar region is very differently shaped. The parastylar region anterior to the stylocone is wide (tr.), whereas it is narrow in Potamotelses (see Fox, 1972, Fig. la). In Deltatheridium the paracingulum extends from the paraconule along the anterior wall of the tooth to a point above the tip of the paracone, in Potamotelses only a possible remnant of the paracingulum can be recognised above the paracone tip and conules are lacking. The ectoflexus is less strongly incurved in Potamotelses than in Deltatheridium, and the crenulations along its margin are larger in Potamotelses than in Deltatheridium. The metacrista is much shorter in Potamotelses than in Deltatheridium and Fox claims that a cusp "c" (CROMPTON, 1971)is present in Potamotelses, while in Deltatheridium a carnassial notch is present in the metacrista. The protocone is very differently shaped in the two genera, it is narrow (tr.) and pointed in Deltatheridium, while in Potamotelses it is not pointed and the protofossa is wider (tr.) and less basined. In both Potamotelses and Deltatheridium precingula and postcingula are absent. The lower molars of Potamotelses resemble those of Deltatheridium in having the paracone larger than the metacone and distinctly separated. However, the trigonid is taller and narrower in Deltatheridium than in Potamotelses. The posterior wall of trigonid has a different structure in the two genera. In Potamotelses there is a distinct groove in the external surface of the metaconid; this clearly separates the two shearing surfaces: on the posterior wall of the protoconid (surface 1) and metaconid (surface 5). In Deltatheridium this groove is absent and only a slight ridge separates the two shearing surfaces, as is.the case in most Cretaceous mammals with tribosphenic dentition. In Potamotelses the marked shearing surface 1 is confined to the protoconid and surface 5 to the metaconid, whereas in Deltatheridium the shearing surface 1 is present on both the protoconid and part of the metaconid, while the shearing surface 5 is small and extends only along the lingual part of the posterior wall of the metaconid. In other words, the typical "trapshear mechanism" (CROMPTON, 1974), well developed in Deltatheridium, is not present in Potamotelses. The carnassial notches present in Deltatheridium on the protocrista and metacrista, as well as on the paracristid, are absent from Potamotelses. The talonid in Potamotelses has a structure very different from that of Deltatheridium, it is wide (tr.) with three or four well developed talonid cusps, while in Deltatheridium the talonid is very narrow and trigonid cusps very poorly developed. The above comparison of the structure of the molars of Deltatheridium and Potamotelses, as well as the analysis of the shearing surfaces in Deltatheridium published in this paper, show that the two genera differ markedly. Deltatheridium has well developed shearing surfaces along both anterior and posterior walls of the upper molars, shearing surface 2 being larger than the shearing surface 1, whereas in Potamotelses (based on the upper tooth) the shearing surfaces are not as well developed. In Deltatheridium the entocristid is very low, the protocone tip is exposed lingually when the teeth are in centric occlusion (Text-fig. 6A), and the shearing surface 6 is not present, whereas in Potamotelses, in which the talonid basin
20 122 ZOFIA KIELAN-JAWOROWSKA is very large, the protocone tip is not visible lingually when the teeth are in centric occlusion because of the high lingual wall of the talonid, and the shearing surface 6 is very well developed. The above comparison shows that, in spite of the apparent similarity between the molars of Potamotelses and Deltatheridium, they differ basically in many points and the analysis of their occlusion shows that they functioned in quite different ways. The conclusion is that Deltatheridium and Potamotelses are not closely related and belong to different groups of mammals, Deltatheridium representing a primitive type of molar occlusion, Potamotelses a strongly advanced one. Fox (1972) compared Potamotelses with Aegialodon (K. A. KERMACK et al., 1965) and claimed a great similarity between the lower molars of these genera. However, on the posterior wall of the trigonid surface in Aegialodon (K. A. KERMACK et al. I, c. Fig. 9) there are three shearing surfaces, whereas only two in Potamotelses. I do not recognise the floor of the groove, separating the posterior surfaces of the protoconid and metaconid in Potamotelses, as being homologous to the shearing surface "d" described by K. A. KERMACK et al. (1965) for Aegialodon and present on the posterior surface of the metaconid. It appears from the analysis of the type of shearing surfaces in Aegialodon and a reconstruction of the upper molars in this genus done by CROMPTON (1971) that the separate middle shearing surface (surface "d" ofk. A. KER MACK et al., 1965) was probably caused by the protoconule, which was not aligned with the paracrista, and that the paracingulum extending from the protoconule labially was rather well developed. As the shearing surface I b (lead by paracingulum) on the upper molar was probably arranged at a different angle than the shearing surface la (lead by paracrista), the matching shearing surfaces on the trigonid have to be arranged also at different angles to each other. Thus it is very probable that in Aegialodon, which is regarded by Fox (1972) as close to the ancestors of Potamotelses, the paracingulum and paraconule were probably developed. COMPARISON OF THE DELTATHERlDIIDAE WITH THE MARSUPIALS The fragments of skull of Deltatheridium, which are preserved, show striking similarities to those of primitive marsupials, such as, e.g., Didelphis. The similarities are in the structure of the nasals, which are expanded posteriorly and contact the lacrimals; in details of the structure of the lacrimal, which in both groups has a very large facial wing and the lacrimal foramen situated in the same position; and in the structure of the zygomatic arch, which in both groups is deep, with the jugal extending far anteriorly to a level opposite the anterior margin of the lacrimal and posteriorly as far as the glenoid fossa. The characters which the Deltatheridiidae and primitive marsupials have in common are primitive features and some of them (e.g. expanded nasals and long jugal) also occur in the Late Cretaceous eutherian mammals from Asia (KIELAN-JAWOROWSKA, 1969 and 1975). However, the Deltatheridiidae are in the structure of the snout closer to the primitive marsupials than the Cretaceous eutherian mammals from Asia. Deltatheridium differs from most marsupials in lacking palatal vacuities; however, the modern marsupial genus Philander from Panama has no palatal vacuities and the structure of the palate is similar to that of Deltatheridium, The Deltatheridiidae have a metatherian dental formula, but it is possible that the primitive therians had four molars, the last one being lost in eutherians. If this is correct, then the metatherian dental formula in the Deltatheridijdae, with a tendency towards the loss of the last molar, would not necessarily imply that they are marsupials. The structure of the molars of the Deltatheridiidae is different from that of most of the American Cretaceous marsupials (CLEMENS, 1966;
21 DELTATHERIDIIDAE 123 SLAUGHTER, 1968b; LILLEGRAVEN, 1969; SAHNI, 1972; SIGE, 1973). Also, the early Cretaceous forms such as Holoclemensia differ from the Deltatheridiidae in-pos sessing a mesostyle (absent in Deltatheridiidae) and in having a parastylar region which is much larger than the metastylar region. The lower molars have not been found in occlusion with the uppers of Holoclemensia; however, lower mol ars that have been assigned to Holoclemensia by SLAUGHTER (1971) differ from those of the D eltatheridiidae in having the paraconid smaller than the metaconid and the entoconid twinned with hypoconulid. TuRNBULL (1971) assigned to Holoclemensia two different types of lower molars, but these are clearly not from congeneric forms. In one of these types, represented, e.g., by the specimen PM 1005 in the Field Museum collection (TuRNBuLL, 1971, Fig. 5 c), the paraconid is very small, smaller than the metaconid, whereas in the other type represented, e.g., by the specimen PM 965 from the same collection (TuRNBULL, I. c. Fig. 4, middle row, second figure from the top) the paraconid is larger than the metaconid, as is the case in the Deltatheridiidae. It follows, therefore, that the identification of isolated lower teeth as belonging to the genus Holoclemensia is doubtful. Of the Cretaceous marsupials only the Stagodontidae are in some respects similar to the Deltatheridiidae; for example, in both the paraconid is larger than the metaconid. However, in the Stagodontidae the entoconid and hypoconulid are twinned, as in all primitive marsupials, which is not the case in the Deltatheridiidae. The talonid is narrow (tr.) in the Deltatheridiidae, but comparatively wide in the Stagodontidae. The upper molars in both groups show some resemblance in that they lack a stylar cusp C, but the general pattern of the upper molars is different; the stylar shelf in the Stagodontidae is not as wide (tr.) as in the Deltatheridiidae; the conules in the Stagodontidae are winged (CLEMENS, 1966; Fox, 1971) and the paracingulum in the Stagodontidae is more extensive than in the Deltatheridiidae; the paracone in the Stagodontidae is smaller than the metacone, while the reverse is the case for the Deltatheridiidae. COMPARISON OF THE DELTATHERIDIIDAE WITH EUTHERIAN MAMMALS OF THE LATE CRETACEOUS OF ASIA The known therian fauna from the Late Cretaceous of Asia contains seven monotypic genera (see Table 2). Four of these genera: Kennalestes, A sioryctes, Zalambdalestes and Barunlestes are regarded as true eutherians and they are assigned to the three families:?leptictidae, Palaeoryctidae and Zalambdalestidae (KIELAN-JAWOROWSKA, 1969 and 1975). Hyotheridium from the Djadokhta Formation is known from a single specimen (AMNH 21702) which is so badly damaged that only a few features can be recognised. For this reason Hyotheridium cannot be assigned to a specific family. However, it should be stressed that the strong, single rooted canines and the short snout of Hyotheridium are much more reminiscent of the DeItatheridiidae than of the true eutherian mammals from the same formation. Because.of the paucity of the material I shall not discuss Hyotheridium further. All the eutherian genera listed above from the Late Cretaceous of Asia are highly specialised in different direction s (and assigned to three different famili es), but despite of thi s have many features in common, which suggest that they have derived from a common ancestor; these features differentiate them from the Deltatheridiidae. Deltatheridiidae are characterised by a short snout (which suggests a carnivorous speciali sation), nasal s which are expanded posteriorly to contact the lacrim als, lacrimal large with an extensive facial wing, large lacrimal foramen and zygomatic arches strong and deep, with jugal reaching far backwards. The -
22 124 ZOFIA KIELAN-JAWOROWSKA T able 2 The Late Cretaceous therian fauna of Asia InfracIass Family I I Djadokhta Formation (1 Santonian) Species Barun Goyot Formation and Khermeen Tsav formation (1 Middle Campanian) 'l Leptictidae Kennalestes gobiensis Eutheria Palaeoryctid ae Asioryctes nemegetensis Zalambdalestidae Zalambdalestes lechei Barunlestes butleri Deltatheroides cretacicus Theria of DeItatheridiidae Deltatheridium pretrituberculare Deltatheridium pretrituberculare tardum metatherian- pretrituberculare eutherian grade Uncertain family Hyotheridium dobsoni eutherian mammals listed above are in contradistinction characterised by: slender, strongly elongated snouts (especially so in Zalambdalestes), posteriorly expanded nasals, which probably contact the lacrimals, comparatively large lacrimals, although they are relatively smaller than in the Deltatheridiidae and have smaller lacrimal foramina. In these eutherian mammals the zygomatic arches are very slender, the jugal is less deep than in the Deltatheridiidae, but reaches far backwards. Both the Deltatheridiidae and the eutherian mammals have more than three upper incisors. There are probably four in the Deltatheridiidae, probably four in Kennalestes, five in Asioryctes and possibly three in the Zalambdalestidae. The number of lower incisors is reduced in the Deltatheridiidae to two or one, while in Kennalestes there are probably three or more lower incisors, four in A sioryctes and three (the first one unusually enlarged) in the Zalambdalestidae. The maxillary dentition is quite different in the Deltatheridiidae and in the Asiatic Cretaceous eutherian mammals. In the Deltatheridiidae there is a strong, single-rooted canine, followed by three non-molarized premolars and three to four molars, whereas in the here discussed eutherian mammals the canine is strong but double-rooted and situated some distance to the rear of the premaxillo-maxillary suture. The only exception is Barunlestes, which was probably derived from Zalambdalestes and which has a single-rooted upper canine. It should be noted that in Kennalestes the deciduous upper canine is also single-rooted and it is only the permanent one which is double-rooted. The double-rooted canine in these eutherian mammals is separated from a small P' by a narrow space, both P' and p2 are double-rooted and do not come into occlusion with the lower premolars. The strongest tooth in the whole series is P", with a high, piercing paracone. In the Zalambdalestidae P" has a strongly developed protocone, while in Kennalestes and A sioryctes there is an incipient protocone. P' is semimolariform in all the listed eutherian genera. In the Zalambdalestidae it is strongly elongated
23 DELTATHERlDIIDAE 125 transversely, the protocone is well developed and only an incipient metacone is present as a cuspule on the metacrista. In Kennalestes and in Asioryctes p4 shows a more advanced degree of rnolarization, the protocone is developed as in the molars and an incipient metacone, which is smaller than that of the molars, is present. As far as the lower dentition is concerned, the lower incisors, although differently developed in various genera discussed here, are in all cases semi-procumbent; the lower canine is in Kennalestes and Asioryctes double-rooted, while it is single-rooted in the Zalambdalestidae, the two first premolars are in all the genera comparatively short, third and fourth lower premolars are tall piercing teeth, P 4 is semimolariform in the Zalambdalestidae, but not so in Kennalestes or in Asioryctes. The above comparison shows that all the Late Cretaceous Asian eutherian mammals are similar to one another in the general appearance and organisation of the dentition when seen in lateral view (Kielan-Jaworowska, 1975, figs 1 and 2) and differ from the Deltatheridiidae. It is difficult to draw any conclusion as to the homology of the teeth in the Asiatic eutherian mammals and the Deltatheridiidae. As stated above, in the Deltatheridiidae the upper canine is strong and single-rooted, while in the Late Cretaceous Asiatic eutherian mammals it is with one exception double-rooted, situated some distance behind the premaxillomaxillary suture. Single-rooted, strong canines are a feature of primitive (Triassic and Jurassic) mammals and ofthe cynodonts (SIMPSON, 1928; PARRINGTON, 1971; MILLS, 1971; CROMPTON, 1974). The possibility exists that the double-rooted tooth, designated as an upper canine in the Asiatic Cretaceous eutherians, is not homologous with the single-rooted canine of the Deltatheridiidae and of the Triassic mammals and cynodonts, but represents the first premolar, the primitive canine presumably having been lost. This would possibly explain why there is always a space between the premaxillo-maxillary suture and the double-rooted canine of the Asiatic eutherians. In Asioryctes, which has five upper incisors (KIELAN-JAwOROWSKA, 1975), the last incisor is situated just within the premaxillo-maxillary suture and it is possible that this tooth is a reduced canine rather than an incisor. If this hypothesis is correct, there are five upper premolars and three molars in the eutherian genera being discussed, the first premolar acting as a canine and showing a tendency in evolution to the replaced by the reduced, singlerooted tooth. Whether we accept the first maxillary tooth in the Late Cretaceous eutherian mammals as the true canine, homologous to that of the Deltatheridiidae, or as the first premolar, the general organization of the dentition of the Deltatheridiidae and the Asiatic eutherian mammals is quite different. The differences in the skull and teeth structure between the Deltatheridiidae and the Late Cretaceous Asiatic eutherians show that these groups are not very closely related and, if they have derived from a common ancestor, the divergence took place long ago before the Late Cretaceous, presumably at the beginning of the Early Cretaceous. It follows that the Deltatheridiidae are in the general organization of the dentition and in the coronal structure of the molars different from undoubted eutherian and metatherian mammals. They probably represent a third, separate evolutionary line, (what has been also suggested by LILLEGRAVEN, 1974) and are therefore classified as Theria of metatherianeutherian grade. Among the Asian eutherian mammals I do not know any forms which could be with certainty recognised as possible descendants of the Deltatheridiidae. However, certain North American Paleocene Palaeoryctidae, such as Didelphodus have upper molars strikingly similar to those of Deltatheridium, but this is not true for all North American Palaeoryctidae. However, this must not be taken to imply that some of the North American Palaeoryctidae are
24 126 ZOFIA KIELAN-JAWOROWSKA the descendants of Asiatic Deltatheridiidae especially in view of the fact theat the Palaeoryctydiae have a eutherian dental formula. It appears that the forms at present classified as the Palaeoryctidae are not a homogeneous group, and should be revised. Polska Akademia Nauk Zaklad Paleoozologii, Warszawa, AI. Zwirki i Wigury 93 January, 1974 ADDENDUM When this paper was submitted to publication, a paper by Fox (1974) appeared, in which he describes a single upper molariform tooth from the upper part of the Edmonton Formation, a single lower molar and a lower molar trigonid from the Lance Formation and a lower molar talonid from the Oldman Formation, which show strong similarities to the Asian Deltatheridiidae. On this basis Fox claims that the Deltatheridiidae occured inthe Late Cretaceous of North America. Although I agree that the similarities between these teeth and those of the Deltatheridiidae are striking, I think that the occurrence of the Deltatheridiidae in North America cannot be regarded as proven, as we do not know the dental formulae of the animals to which these teeth belong, and the similarities in the structure of isolated teeth may be due to convergence. REFERENCES ARCHER, M The development of the cheek teeth in Antechinus flavipes (Marsupialia, Dasyuridae). - Australian Mammalogy, 1, 68. BELIAJEVA, E. I. - see EEnHEBA, E. H. BENSLEY, A. B The hornologies of the stylar cusps in the upper molars of the Didelphidae. - Univ. Toronto Stud. Bioi. Ser., 5, , Toronto. BUTLER, P. M The evolution ofcarnassial dentition in the Mammalia. - Proc. Zoo I. Soc., 116, 2, , London. - & KIELAN-JAWOROWSKA, Z Is Deltatheridium a marsupial? - Nature, 245, , London. CLEMENS, W. A. Jr Fossil mammals of the type Lance Formation Wyoming. Part II. Marsupialia, - Univ. Calif. Pubs. Geol. sa., 62, 1-122, Berkeley. - & MILLS, J. R. E Review of Peramus tenuirostris Owen (Eupantotheria, Mammalia). - Bull. Brit. Mus. {Nat. Hist.} Geol., 20, 3, , London. CROMPTON, A. W The origin of the tribosphenic molar. - In : Early Mammals, D. M. Kermack & K. A. Kermack (eds.). Suppl, No. I, Zool. J. Linn. Soc., 50, 65-87, London The dentitions and relationships of the Southern African Triassic mammals Erythrotherium parringtoni and Megazostrodon rudnerae. - Bull. Brit. Mus. {Nat. Hist.} Geol., 24, 7, , London. Fox, R. C Marsupial mammals from the early Campanian Milk River Formation, Alberta. In : Early Mammals, D. M. Kerrnack & K. A. Kermack (eds.). Suppl. No. I, Zool. J. Linn. Soc., 50, , London A primitive therian mammal from the Upper Cretaceous ofalberta. - Canad. J. Earth Scis., 9,11, Deltatheroides - like mammals from the Upper Cretaceous of North America. - Nature, 249, 392, London. GREGORY, W. K. & SIMPSON, G. G Cretaceous mammal skulls from Mongolia. - Amer. Mus. Novit., 225, 1-20, New York. KERMACK, D. M., KERMACK, K. A. & MussETT, F The Welsh pantothere Kuehneotherium praecursoris. - J. Linn. Soc. {Zool.}, 47, 312, , London. KERMACK, K. A., LEES, P. M. & MussETT, F Aegialodon dawsoni, a new trituberculosectorial tooth from the Lower Wealden, - Proc. Roy. Soc., B., 162, , London. KIELAN-JAWORowSKA, Z Preliminary data on the upper Cretaceous eutherian mammals from Bayn Dzak, Gobi Desert. - Results Pol. Mong. Pal. Exp. I, Pal. Pol., 19, , Pis XXII-XXVII, Warszawa Multituberculate succession in the late Cretaceous of the Gobi Desert (Mongolia). - Results..., V. Ibidem, 30, 23 44, Pis V-XXI,
25 DELTATHERIDIlDAE Preliminary description of two new eutherian genera from the Late Cretaceous of Mongolia. - Re sults..., VI. Ibidem, 33, & BARSBOLD, R Narrative of the Poli sh-mongolian Palaeontological Expeditions Re sults..., IV. Ibidem, 27, KREBS, B Evolution of the mandible and lower dentition in dryolestids (Pantotheria, Mammalia). -In: Early Mammals, D. M. Kermack & K. A. Kermack (eds.), Suppl. No. 1, Zool. J. Linn. Soc., 50, , London. LILLEGRAVEN, J. A Latest Cretaceous mammals ofthe upper part ofthe Edmonton Formation ofalberta, Canada, and review of marsupial-placental dichotomy in mammalian evolution. - Univ. Kans. Paleont. Contr., 50, (Vertebrata 12) 1-122, Lawrence Biogeographical con siderations of the marsupial-placental dichotomy. - Ann. Rev. Ecol. Syst., 5, McKENNA, M. c., MELLElT, J. S. & SZALAY, F. S Relationships of the Cretaceous mammal Deltatheridium. J. Pal., 45, 3, MILLS, J. R. E The dentitions of Peramus and Amphitherium. - Proc. Linn. Soc. Lond., 175, 2, ,2 Pis, London The dentition of Morganucodon. -In: Early Mammals, D. M. Kermack & K. A. Kermack (eds.), Suppl. No. 1, Zool. J. Linn. Soc., 50, 29-63, London. PARRINGTON, F. R On the Upper Triassic mammals. - Phil. Trans. Roy. Soc. Land. B., 261,838, , London. PAlTERSON, B Early Cretaceous mammals and the evolution of mammalian molar teeth. - Fieldiana: Geology, 13, 1, 1-105, Chicago. SAHNI, A The vertebrate fauna of the Judith River Formation, Montana. - Bull. Amer. Mus. Not. Hist., 147, 6, , New York. SIGE, B La faunule de marnrniferes du Cretace superieur de Laguna Umayo (Andes peruviennes), - Bull. Mus. Not. Hist. Nat., 3, ser, 99, , PIs I-Il, Paris. ' SIMPSON, G. G A catalogue of the Mesozoi c Mammalia in the Geological Department of the British Museum. Brit. Mus. (Nat. H istory), 1-215, London. SLAUGHTER, B. H A therian from the Lower Cretaceous (Albian) of Texas. - Postilla, 93, 1-18, New Haven Earliest known eutherian mammals and the evolution of premolar occlusion. - Tex, J. Sci., 20, 1, 3-12, Houston. 1968b. Earliest known marsupials. -Science, 62, ; Washington. 1968c. Holoclemensia instead of Clemensia. -Ibidem, 162, Mid-Cretaceous (Albian) therians of the Butler Farm local fauna, Texas. - In : Early Mammals, D. M. Kermack & K. A. Kerrnack (eds.), Suppl. No. 1, Zool. J. Linn. Soc., 50, , London. SZALAY, F. S Mixodectidae, Microsyopidae, and the insectivore-primate transition. - Bull. Amer. Mus. Not. Hist., 140, 4, , Pis 17-57, New York. SZALAY, F. S. & McKENNA, M. C Beginning of the age of mammals in Asia: the late Paleocene Gashato Fauna, Mongolia. -Ibidem, 144, 4, TURNBULL, W. D The Trinity therians: their bearing on evolution of marsupials and other therians. - In: Dental Morphology and Evolution, A. A. Dahlberg (ed.), Univ. Chicago Press, , Chicago. TRoFIMov, B. A. see TpO<I>HMOB, E. A. VAN VALEN, L Deltatheridia, a new order of mammals. - Bull. Amer. Mus. Not. Hist., 132, I, 1-126, PIs 8, New York. TPO<I>liMOB, E. A Mnexomrraromae Ha Hjf)J{Hero Mena MOHronHH. - Ceccaa nocbrll\ehhar loo-netiho eo ~HH p0)f{~ehhr arcanexaxa A. A. EOPI1CRHa, TIaneOHT. HHCT. AH CCCP, 65-67, Mocxsa. EEnREBA, E. 11., TpO<I>HMOB, E. A., PElUETOB, B OCHoBHble sransr saomouaa MneHOn,llTalOll\llX B Iloa~ HeM Meaoaoe-IlaneoreHe nerrrparrsnoii AaIlH. - <l>ayha H 6HOCTpaTllrpa!pIIR MeaoaOR H HaHHoaOR MOHronmr. - COBM. COB.-MoHL TIaneOHM. 3Hcn. Tpyobl, I, 19-45, Mocrcaa,
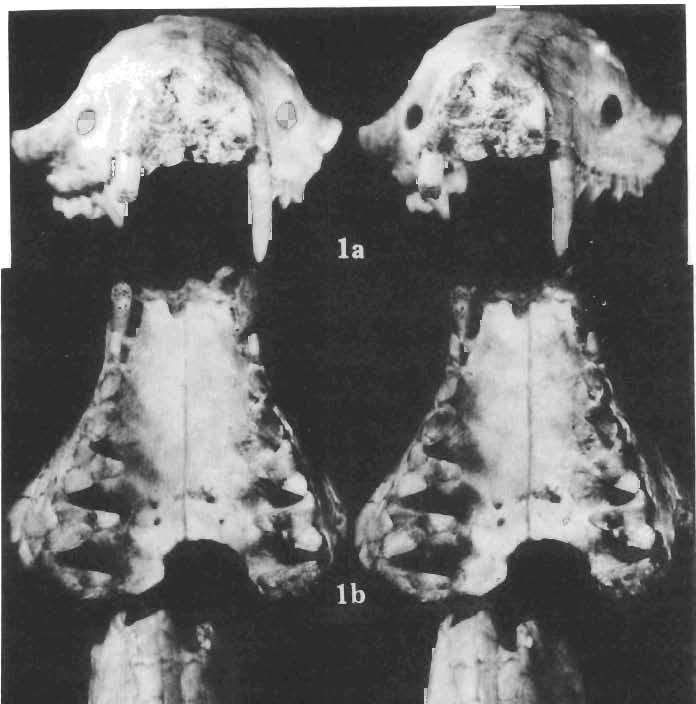
26 EXPLANATIONS OF PLATES PLATE XXVIII Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upp er Cretaceous, Kherrneen Tsav formation, Khermeen Tsav 11, Gobi Desert, Mongolia Fig. I a. Part of the rostrum with right and left C-M", right canine broken off, ZPAL MgM-IjI02, stereo-photograph in anterior view. F ig. I b. Stereo-photograph of the same in occlusal view. Fig. 1c. Stereo-photograph of the same in dorsal view. Fig. I d. Stereo-photograph of the same in right lateral view. Fig. I e. The same in left lateral view. All x 4 Photo : W. Skariynsk! PLATE XXIX Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Barun Goyot Formation, Nemegt, Red monadnocks, Nemegt Basin, Gobi Desert, Mongolia Fig. I a. Stereo-photograph of the incomplete rostral part of the skull, with strongl y damaged cranial roof with left C, pt and P' and right PO-M", in right lateral view, ZPAL MgM-Ij60, x 4. Fig. I b. Stereo-photograph of the same in occlusal view, x 4. Photo: W. Skarzyns k i Upper Cretaceous, Kherrneen Tsav formation, Kh ermeen Tsav 11, Gobi Desert, Mongolia Fig. 2a. Holotype, (young individual), (see also Plates XXX-XXXIV). Stereo -photograph of the left maxilla with broken C, pt_m", ZPAL MgM-Ij91, x 6. Fig. 2b. Stereo-photograph of the same in occlusal view, X 6. Photo : A. Coleman
27 DELTATHERIDIlDAE 129 PLATE XXX Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Khermeen Tsav formation, Khermeen Tsav Il, Gobi Desert, Mongolia Fig. 1a. Badly damaged skull, consisting of fragments of the cranial roof and both maxillae, associated with incomplete right and left mandible, before the final preparation in left lateral view, holotype, (see also Plates XXIX and XXXI-XXXIV) ZPAL MgM-I j91, x 6. Fig. 1b. Stereo-photograph of the left mandible, of the same specimen with P2-M., in occlusal view, X 6. Fig. 1c. Stereo -photograph of the right mandible, belonging to the same specimen, with root of single incisor, root of C, roots of P, and with P2-M2 and roots of M. in occlusal view, X 6. Upper Cretaceous, Barun Goyot Formation, Nernegt, Eastern Sayr, Nemegt Basin, Gobi Desert, Mongolia Fig. 2a. Stereo-photograph of the fragment of the right maxilla with M'-M' in outer view. Left mandible of the same specimen is figured on PI. XXXV, ZPAL MgM-Ij136, X 4. Fig. 2b. Stereo-photograph of the same in occlusal view, X 4. Fig. 2c. Stereo-photograph of the same in inner view, X 4. Photo: W. Skarzynskl and A. Coleman PLATE XXXI Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Khermeen Tsav formation, Khermeen Tsav 11, Gobi Desert, Mongolia Fig. 1a. Stereo-photograph of the right maxilla with damaged P' and P2_M2 in outer view; holotype (see also Plates XXIX, XXX, XXXII-XXXIV), ZPAL MgM-Ij9I. Fig. 1b. Stereo-photograph of the same in inner view, showing part ofthe damaged cranial roof. Fig. 1c. Stereo-photograph of the left mandible of the same specimen with P2-M. in outer view. Fig. 1d. Stereo-photograph of the same in inner view. All X Palaeontologia Polonica No. 33 Photo: A. Coleman
28 130 ZOFIA KIELAN-JAWOROWSKA PLATE xxxn Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Kherrneen Tsav formation, Khermeen Tsav I, Gobi Desert, Mongolia Fig. 1a. Stereo-photograph of the left maxilla with C-M3 in inner view; holotype (see also Plates XXIX-XXXI, XXXIII and XXXIV), ZPAL MgM-I/91. Fig. 1b. Stereo-photograph of the right maxilla of the same specimen in occlusal view. Fig. 1c. Stereo-photograph of the right mandible of the same specimen with root of a single incisor, broken of canine, broken off Ph P2-M2, roots of M,; in inner view. Fig. 1d. Stereo-photograph of the same in outer view. All X 4 Photo: A. Coleman PLATE XXXIII Deltatheridium pretrituberculare tardum subsp. n. Upper Cretaceous, Khermeen Tsav formation, Khermeen Tsav I, Gobi Desert, Mongolia Page 108 Scanning electron microscope stereo-photographs Fig. la. Left MS in occlusal view; holotype (see also Plates XXIX-XXXII and XXXIV), ZPAL MgM-I/91. Fig. 1b. The same in inner view. Fig. 1c. Left maxilla of the same with MI_M2 in occlusal view. Fig. 1d. The same in inner view. Fig. 1e. The same in anterior view. All X 11.5 Photo:.H. G. Pierce PLATE XXXIV Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Khermeen Tsav formation, Khermeen Tsav I, Gobi Desert, Mongolia Scanning electron microscope stereo-photographs Fig. 1a. Right M2-M3 in oblique anterior view; holotype (see also Plates XXIX-XXXTIT), ZPAL MgM-T/9I. Fig. lb. The same in posterior view. Fig. 1c. M3 of the same in occlusal view. Fig. Id. M2 of the same in occlusal view. Fig. le. Left maxilla of the same specimen with Ml-M" in inner view. All X 11.5 Photo: R.' G. Pierce
29 DELTATHERIDIIDAE 131 PLATE XXXV Deltatheroides cretacicus GREGORY & SIMPSON. Page 117 Upper Cretaceou s, Djadokhta Formation, Bayn Dzak, Ruins, Gobi Desert, Mongolia Fig. 1a. Stereo-photograph of the damaged left mandible with fragmentary root of C, roots of Pi' and Pa-M. in outer view, ZPAL MgM-I/29, x 4. Fig. 1b. Stereo-photograph of the same in occlusal view, x 4. Fig. 1c. Stereo-photograph of the same in inner view, X 4. Photo: A. Coleman Deltatheridium pretrituberculare tardum subsp. n. Page 108 Upper Cretaceous, Barun Goyot Formation, Nemegt, Eastern Sayr, Nemegt Basin, Gobi Desert, Mongolia Fig. 2a. Stereo-photograph of the fragment of the left mandible with roots of p., Ml-Ma and fragment of root of Ma in outer view, x 4. Fragment of the right maxilla found in association with this specimen is figured on Plate XXX, ZPAL MgM-I/136. Fig. 2b. Stereo-photograph of the same in occlusal view, x 4. Fig. 2c. Stereo-photograph of the same in inner view, x 4. Photo: W. Skarlyliskl




30 Palaeontologia Polonica, No. 33, 1975 PI. XXVIII Z. KIELAN-JAWOROWSKA: THE DELTATHERIDIIDAE








31 Palaeontologia Polonica, No. 33, 1975 PI. XXIX Z. KIELAN-JAWOROWSKA: THE DELTATHERIDIIDAE










32 Palaeontologia Polonica, No. 33, 1975 PI. XXX N ll:i,... V,.~'.'" "'.~ t '. "' '.,''7",-, -,'_.» ' ';...,.,.. '.:.i.";': ;"...~....'-. -'. ~. ~ " i.i Z. KIELAN-]AWOROWSKA : THE D ELTATHERlDIIDAE
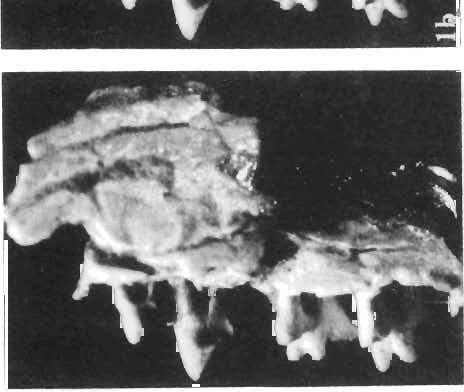
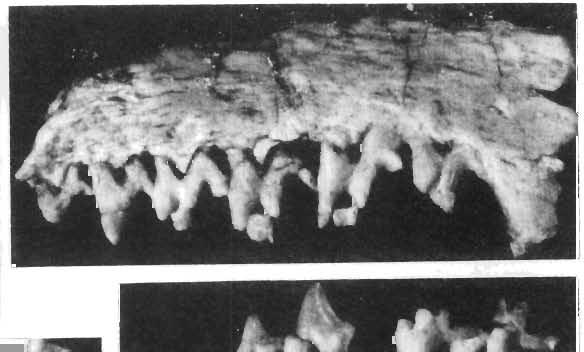




33 Palaeontologia Polonica, No. 33, 1975 PI. XXXI Z. KIELAN-JAWOROWSKA: THE DELTATHERIDIlDAE



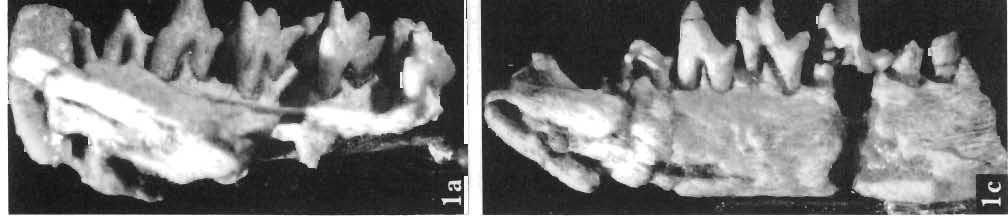

34 Palaeontologia Polonica, No. 33, 1975 PI. XXXlI Z. KlELAN-JAWOROWSKA: THE DELTATHERIDIIDAE






35 Palaeontologia Polonica, No. 33, 1975 PI. XXXIlI Z. KIELAN-]AWOROWSKA: THE DELTATHERlDlIDAB






36 Palaeontologia Polonica, No. 33, 1975 PI. XXXIV z. KIELAN-JAWOROWSKA : THE DELTATHERlDIIDAE






37 Palaeontologia Polonica. No. 33, 1975 PI. XXXV Z. KIELAN -JAWOROWSKA: THE DELTATHERIDIIDAE
P3. (Figs. 3A-F, J-K, 4O-Q, 9C) P3 of P. gaoi bears three roots, the mesial and lingual
 P3 of P. gaoi bears three roots, the mesial and lingual") Boyer, Scott, and Fox, 2011: Supplementary File 2 Description of dentition of Pronothodectes gaoi P3. (Figs. 3A-F, J-K, 4O-Q, 9C) P3 of P. gaoi bears three roots, the mesial and lingual of which are subequal
Boyer, Scott, and Fox, 2011: Supplementary File 2 Description of dentition of Pronothodectes gaoi P3. (Figs. 3A-F, J-K, 4O-Q, 9C) P3 of P. gaoi bears three roots, the mesial and lingual of which are subequal
Fig. 12 Trigonias?osborni Lucas, SMNH PI637.2, left maxillary fragment with pi to MI; occlusal view, x I. DESCRIPTION
 Fig. 12 Trigonias?osborni Lucas, SMNH PI637.2, left maxillary fragment with pi to MI; occlusal view, x I. DESCRIPTION SMNH P1637.1 has the teeth somewhat cracked but otherwise well preserved. p 2 is very
Fig. 12 Trigonias?osborni Lucas, SMNH PI637.2, left maxillary fragment with pi to MI; occlusal view, x I. DESCRIPTION SMNH P1637.1 has the teeth somewhat cracked but otherwise well preserved. p 2 is very
PRELIMINARY DATA ON THE UPPER CRETACEOUS EUTHERIAN MAMMALS FROM BAYN DZAK, GOBI DESERT
 ZOFIA KIELAN-JAWOROWSKA PRELIMINARY DATA ON THE UPPER CRETACEOUS EUTHERIAN MAMMALS FROM BAYN DZAK, GOBI DESERT (WST~PNE DANE 0 G6RNO-KREDOWYCH SSAKACH LOZYSKOWYCH Z BAJN DZAK NA PUSTYNI GOB!) (Plates XXII-XXVII)
ZOFIA KIELAN-JAWOROWSKA PRELIMINARY DATA ON THE UPPER CRETACEOUS EUTHERIAN MAMMALS FROM BAYN DZAK, GOBI DESERT (WST~PNE DANE 0 G6RNO-KREDOWYCH SSAKACH LOZYSKOWYCH Z BAJN DZAK NA PUSTYNI GOB!) (Plates XXII-XXVII)
PAL A EON T 0 LOG I C A POLONICA PERCY M. BUTLER PLACENTALS
 ACT A Vol. 22 PAL A EON T 0 LOG I C A POLONICA ------------- 1977 Nr.3 PERCY M. BUTLER EVOLUTIONARY RADIATION OF THE CHEEK TEETH OF CRETACEOUS PLACENTALS Abstract. - Included is a comparative study of
ACT A Vol. 22 PAL A EON T 0 LOG I C A POLONICA ------------- 1977 Nr.3 PERCY M. BUTLER EVOLUTIONARY RADIATION OF THE CHEEK TEETH OF CRETACEOUS PLACENTALS Abstract. - Included is a comparative study of
Characters.-M1 more like that in Procynodictis than like that of Uintacyon. Prodaphcenus (?) robustus in known features of jaw and lower dentition,
 robustus in known features of jaw and lower dentition,") VOL. 20, 1934 GEOLOGY: C. STOCK NEW CREODONTA FROM THE SESPE UPPER EOCENE, CALIFORNIA By CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
VOL. 20, 1934 GEOLOGY: C. STOCK NEW CREODONTA FROM THE SESPE UPPER EOCENE, CALIFORNIA By CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
Key points for starting off
 Key points for starting off First off, the five questions to ask yourself about a loose tooth before identifying it are: 1. 2. 3. 4. 5. Category (incisor, canine, premolar or molar)? Permanent or deciduous?
Key points for starting off First off, the five questions to ask yourself about a loose tooth before identifying it are: 1. 2. 3. 4. 5. Category (incisor, canine, premolar or molar)? Permanent or deciduous?
PERISSODACTYLA OF THE SESPE EOCENE, CALIFORNIA
 260 PALEONTOLOGY: C. STOCK PROC. N. A. S. PERISSODACTYLA OF THE SESPE EOCENE, CALIFORNIA BY CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
260 PALEONTOLOGY: C. STOCK PROC. N. A. S. PERISSODACTYLA OF THE SESPE EOCENE, CALIFORNIA BY CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
NOVITATES AMEIRIICAN MUSEUM NOTES ON AUSTRALIAN MARSUPIALS RARE OR LITTLE-KNOWN IN THE UNITED STATES BY G. H. H. TATE
 AMEIRIICAN MUSEUM NOVITATES PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CITY OF NEW YORK JULY 17, 1951 NUMBER 1528 NOTES ON AUSTRALIAN MARSUPIALS RARE OR LITTLE-KNOWN IN THE UNITED STATES BY G.
AMEIRIICAN MUSEUM NOVITATES PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CITY OF NEW YORK JULY 17, 1951 NUMBER 1528 NOTES ON AUSTRALIAN MARSUPIALS RARE OR LITTLE-KNOWN IN THE UNITED STATES BY G.
Arrangement of the artificial teeth:
 Lecture Prosthodontic Dr. Osama Arrangement of the artificial teeth: It s the placement of the teeth on a denture with definite objective in mind or it s the setting of teeth on temporary bases. Rules
Lecture Prosthodontic Dr. Osama Arrangement of the artificial teeth: It s the placement of the teeth on a denture with definite objective in mind or it s the setting of teeth on temporary bases. Rules
1. What is the highest and sharpest cusp on the lower first deciduous molar? 2. Which of the following is NOT the correct location of an embrasure?
 1 1. What is the highest and sharpest cusp on the lower first deciduous molar? a. mesiobuccal b. distobuccal c. distolingual d.mesiolingual 2. Which of the following is NOT the correct location of an embrasure?
1 1. What is the highest and sharpest cusp on the lower first deciduous molar? a. mesiobuccal b. distobuccal c. distolingual d.mesiolingual 2. Which of the following is NOT the correct location of an embrasure?
Morphology of an Anatomic Crown. By: Assistant Professor Dr. Baydaa Ali Al - Rawi
 Morphology of an Anatomic Crown By: Assistant Professor Dr. Baydaa Ali Al - Rawi October 4, 2009 Elevated landmarks Depressed landmarks A) Elevated landmarks : 1. Dental lobe : is one of the primary centers
Morphology of an Anatomic Crown By: Assistant Professor Dr. Baydaa Ali Al - Rawi October 4, 2009 Elevated landmarks Depressed landmarks A) Elevated landmarks : 1. Dental lobe : is one of the primary centers
Primary Teeth Chapter 18. Dental Anatomy 2016
 Primary Teeth Chapter 18 Dental Anatomy 2016 Primary Teeth - Introduction Synonyms deciduous teeth, baby teeth, temporary teeth, milk teeth. There are 20 primary teeth, designated as A thru T in the Universal
Primary Teeth Chapter 18 Dental Anatomy 2016 Primary Teeth - Introduction Synonyms deciduous teeth, baby teeth, temporary teeth, milk teeth. There are 20 primary teeth, designated as A thru T in the Universal
Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences,
 A new symmetrodont mammal (Trechnotheria: Zhangheotheriidae) from the Early Cretaceous of China and trechnotherian character evolution Shundong Bi 1,2 *, Xiaoting Zheng 3,4, Jin Meng 5,2, Xiaoli Wang 3,4,
A new symmetrodont mammal (Trechnotheria: Zhangheotheriidae) from the Early Cretaceous of China and trechnotherian character evolution Shundong Bi 1,2 *, Xiaoting Zheng 3,4, Jin Meng 5,2, Xiaoli Wang 3,4,
Identification of Mammal Skulls
 Identification of Mammal Skulls Mammalian teeth are heterodont. That is, they are different as you move from front to rear in the tooth row. This contrasts with the homodont condition of most toothed vertebrates.
Identification of Mammal Skulls Mammalian teeth are heterodont. That is, they are different as you move from front to rear in the tooth row. This contrasts with the homodont condition of most toothed vertebrates.
Fundamental & Preventive Curvatures of Teeth and Tooth Development. Lecture Three Chapter 15 Continued; Chapter 6 (parts) Dr. Margaret L.
 Dr. Margaret L.") Fundamental & Preventive Curvatures of Teeth and Tooth Development Lecture Three Chapter 15 Continued; Chapter 6 (parts) Dr. Margaret L. Dennis Proximal contact areas Contact areas are on the mesial and
Fundamental & Preventive Curvatures of Teeth and Tooth Development Lecture Three Chapter 15 Continued; Chapter 6 (parts) Dr. Margaret L. Dennis Proximal contact areas Contact areas are on the mesial and
THE SNOUT OF PAULOCNUS PETRIFACTUS (MAMMALIA, EDENTATA)
") THE SNOUT OF PAULOCNUS PETRIFACTUS (MAMMALIA, EDENTATA) by D. A. HOOIJER Rijksmuseum van Natuurlijke Historie, Leiden A specimen of the ground sloth discovered by Mr. P. Stuiver in Curasao, Paulocnus petrifactus
THE SNOUT OF PAULOCNUS PETRIFACTUS (MAMMALIA, EDENTATA) by D. A. HOOIJER Rijksmuseum van Natuurlijke Historie, Leiden A specimen of the ground sloth discovered by Mr. P. Stuiver in Curasao, Paulocnus petrifactus
PERMANENT MANDIBULAR INCISORS
 PERMANENT MANDIBULAR INCISORS (Central and Lateral) DR.AHMED AL-JOBORY LEC. 5 PERMANENT MANDIBULAR INCISORS ARE 4 IN NUMBER : 2 CENTRAL (RIGHT &LEFT) AND 2 LATERAL INCISORS (RIGHT &LEFT). CHARACTERISTIC
PERMANENT MANDIBULAR INCISORS (Central and Lateral) DR.AHMED AL-JOBORY LEC. 5 PERMANENT MANDIBULAR INCISORS ARE 4 IN NUMBER : 2 CENTRAL (RIGHT &LEFT) AND 2 LATERAL INCISORS (RIGHT &LEFT). CHARACTERISTIC
AMERICAN MUSEUM NOVITATES
 AMERICAN MUSEUM NOVITATES Number 511 Published by TNe AmzuacAwsMumoIJNATU2AL HISTorCir Dec. 15, 1931 56.9,31 (119:729.1) GENERA AND NEW SPECIES OF GROUND SLOTHS FROM THE PLEISTOCENE OF CUBA BY W. D. MATTHEW1
AMERICAN MUSEUM NOVITATES Number 511 Published by TNe AmzuacAwsMumoIJNATU2AL HISTorCir Dec. 15, 1931 56.9,31 (119:729.1) GENERA AND NEW SPECIES OF GROUND SLOTHS FROM THE PLEISTOCENE OF CUBA BY W. D. MATTHEW1
Bones Ethmoid bone Inferior nasal concha Lacrimal bone Maxilla Nasal bone Palatine bone Vomer Zygomatic bone Mandible
 splanchnocranium - Consists of part of skull that is derived from branchial arches - The facial bones are the bones of the anterior and lower human skull Bones Ethmoid bone Inferior nasal concha Lacrimal
splanchnocranium - Consists of part of skull that is derived from branchial arches - The facial bones are the bones of the anterior and lower human skull Bones Ethmoid bone Inferior nasal concha Lacrimal
European Veterinary Dental College
 European Veterinary Dental College EVDC Training Support Document Preparation of Radiograph Sets (Cat and Dog) Document version : evdc-tsd-radiograph_positioning_(dog_and_cat)-20120121.docx page 1 of 13
European Veterinary Dental College EVDC Training Support Document Preparation of Radiograph Sets (Cat and Dog) Document version : evdc-tsd-radiograph_positioning_(dog_and_cat)-20120121.docx page 1 of 13
Infratemporal fossa: Tikrit University college of Dentistry Dr.Ban I.S. head & neck Anatomy 2 nd y.
 Infratemporal fossa: This is a space lying beneath the base of the skull between the lateral wall of the pharynx and the ramus of the mandible. It is also referred to as the parapharyngeal or lateral pharyngeal
Infratemporal fossa: This is a space lying beneath the base of the skull between the lateral wall of the pharynx and the ramus of the mandible. It is also referred to as the parapharyngeal or lateral pharyngeal
Central Incisor DR.Ahmed Al-Jobory B.D.S.,M.Sc. Conservative Department
 Dental Anatomy Lecture 3 Central Incisor DR.Ahmed Al-Jobory B.D.S.,M.Sc. Conservative Department The permanent maxillary Incisors Maxillary incisor are four in number. The maxillary central incisor is
Dental Anatomy Lecture 3 Central Incisor DR.Ahmed Al-Jobory B.D.S.,M.Sc. Conservative Department The permanent maxillary Incisors Maxillary incisor are four in number. The maxillary central incisor is
Insectivora from the Upper Aragonian and the Lower Vallesian of the Daroca- Villafeliche area in the Calatayud- Teruel Basin (Spain)
") De Jong, Upper Aragonian and Lower Vallesian Insectívora 253 Insectivora from the Upper Aragonian and the Lower Vallesian of the Daroca- Villafeliche area in the Calatayud- Teruel Basin (Spain) F. de Jong
De Jong, Upper Aragonian and Lower Vallesian Insectívora 253 Insectivora from the Upper Aragonian and the Lower Vallesian of the Daroca- Villafeliche area in the Calatayud- Teruel Basin (Spain) F. de Jong
New South Wales, 2052, Australia Version of record first published: 08 Jan 2013.
 This article was downloaded by: [58.165.130.223] On: 06 February 2013, At: 03:54 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
This article was downloaded by: [58.165.130.223] On: 06 February 2013, At: 03:54 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
Upper arch. 1Prosthodontics. Dr.Bassam Ali Al-Turaihi. Basic anatomy & & landmark of denture & mouth
 1Prosthodontics Lecture 2 Dr.Bassam Ali Al-Turaihi Basic anatomy & & landmark of denture & mouth Upper arch Palatine process of maxilla: it form the anterior three quarter of the hard palate. Horizontal
1Prosthodontics Lecture 2 Dr.Bassam Ali Al-Turaihi Basic anatomy & & landmark of denture & mouth Upper arch Palatine process of maxilla: it form the anterior three quarter of the hard palate. Horizontal
DOUBLE TEETH IN THE SPERM WHALE (PHYSETER MACROCEPHALUS L.)
") DOUBLE TEETH IN THE SPERM WHALE (PHYSETER MACROCEPHALUS L.) by H. BOSCHMA Recently a fairly large number of teeth of the sperm whale were acquired for the collections of the Rijksmuseum van Natuurlijke
DOUBLE TEETH IN THE SPERM WHALE (PHYSETER MACROCEPHALUS L.) by H. BOSCHMA Recently a fairly large number of teeth of the sperm whale were acquired for the collections of the Rijksmuseum van Natuurlijke
MAMMALS FROM THE BLUE ASH LOCAL FAUNA (LATE OLIGOCENE), SOUTH DAKOTA. RODENTIA, PART 2: FAMILIES FLORENTIAMYIDAE AND GEOMYIDAE
, SOUTH DAKOTA. RODENTIA, PART 2: FAMILIES FLORENTIAMYIDAE AND GEOMYIDAE") Paludicola 7(1):14-25 November 2008 by the Rochester Institute of Vertebrate Paleontology MAMMALS FROM THE BLUE ASH LOCAL FAUNA (LATE OLIGOCENE), SOUTH DAKOTA. RODENTIA, PART 2: FAMILIES FLORENTIAMYIDAE
Paludicola 7(1):14-25 November 2008 by the Rochester Institute of Vertebrate Paleontology MAMMALS FROM THE BLUE ASH LOCAL FAUNA (LATE OLIGOCENE), SOUTH DAKOTA. RODENTIA, PART 2: FAMILIES FLORENTIAMYIDAE
Lecture 2 Maxillary central incisor
 Lecture 2 Maxillary central incisor Generally The deciduous tooth appears in the mouth at 3 18 months of age, with 6 months being the average and is replaced by the permanent tooth around 7 8 years of
Lecture 2 Maxillary central incisor Generally The deciduous tooth appears in the mouth at 3 18 months of age, with 6 months being the average and is replaced by the permanent tooth around 7 8 years of
Arrangement of posterior artificial teeth Standardized parameters Curve of Wilson Curve of Spee
 . Arrangement of posterior artificial teeth Posterior teeth are set up in tight centric occlusion. The mandibular teeth are set in the wax occlusion rim over the residual ridge in their ideal buccolingual
. Arrangement of posterior artificial teeth Posterior teeth are set up in tight centric occlusion. The mandibular teeth are set in the wax occlusion rim over the residual ridge in their ideal buccolingual
Figure (2-6): Labial frenum and labial notch.
: Labial frenum and labial notch.") The anatomy of the edentulous ridge in the maxilla and mandible is very important for the design of a complete denture. The consistency of the mucosa and architecture of the underlying bone is different
The anatomy of the edentulous ridge in the maxilla and mandible is very important for the design of a complete denture. The consistency of the mucosa and architecture of the underlying bone is different
3. The Jaw and Related Structures
 Overview and objectives of this dissection 3. The Jaw and Related Structures The goal of this dissection is to observe the muscles of jaw raising. You will also have the opportunity to observe several
Overview and objectives of this dissection 3. The Jaw and Related Structures The goal of this dissection is to observe the muscles of jaw raising. You will also have the opportunity to observe several
Techniques of local anesthesia in the mandible
 Techniques of local anesthesia in the mandible The technique of choice for anesthesia of the mandible is the block injection and this is attributed to the absence of the advantages which are present in
Techniques of local anesthesia in the mandible The technique of choice for anesthesia of the mandible is the block injection and this is attributed to the absence of the advantages which are present in
Medical NBDE-II. Dental Board Exams Part I.
 Medical NBDE-II Dental Board Exams Part I http://killexams.com/exam-detail/nbde-ii Question: 149 Anatomically, the term "clinical root" can be defined as which of the following: A. The space in the tooth
Medical NBDE-II Dental Board Exams Part I http://killexams.com/exam-detail/nbde-ii Question: 149 Anatomically, the term "clinical root" can be defined as which of the following: A. The space in the tooth
THE RIGHT INTERPRETATION OF THE CHEEKTEETH TUBERCLES OP TITANOMYS.
 XXIII. ANNALES MUSEI NATIONALIS HUNGARICI. 96. THE RIGHT INTERPRETATION OF THE CHEEKTEETH TUBERCLES OP TITANOMYS. By Dr. J. ÉHIK. (With 5 figures.) The lophodont character of the upper molars of Titanomys
XXIII. ANNALES MUSEI NATIONALIS HUNGARICI. 96. THE RIGHT INTERPRETATION OF THE CHEEKTEETH TUBERCLES OP TITANOMYS. By Dr. J. ÉHIK. (With 5 figures.) The lophodont character of the upper molars of Titanomys
The Premolars. Chapter 17 Permanent Posterior Teeth (p )
") The Premolars Chapter 17 Permanent Posterior Teeth (p. 230-244) General Information Function: u Hold and grind food u Work with molars in mastication. u Even without molars one may be able to chew well
The Premolars Chapter 17 Permanent Posterior Teeth (p. 230-244) General Information Function: u Hold and grind food u Work with molars in mastication. u Even without molars one may be able to chew well
Dental Anatomy High Yield Notes. **Atleast 35 questions comes from these areas of old lectures**
 Dental Anatomy High Yield Notes **Atleast 35 questions comes from these areas of old lectures** This review notes compiled and prepared by my sister for her own study, as a last day review session for
Dental Anatomy High Yield Notes **Atleast 35 questions comes from these areas of old lectures** This review notes compiled and prepared by my sister for her own study, as a last day review session for
Dr.Sepideh Falah-kooshki
 Dr.Sepideh Falah-kooshki MAXILLA Premaxillary/median palatal suture (radiolucent). Incisive fossa and foramen (radiolucent). Nasal passages (radiolucent). Nasal septum (radiopaque). Anterior nasal spine
Dr.Sepideh Falah-kooshki MAXILLA Premaxillary/median palatal suture (radiolucent). Incisive fossa and foramen (radiolucent). Nasal passages (radiolucent). Nasal septum (radiopaque). Anterior nasal spine
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to The Journal of Geology.
 Description of a New Species of Petalodus (P. Securiger) from the Carboniferous of Illinois Author(s): O. P. Hay Reviewed work(s): Source: The Journal of Geology, Vol. 3, No. 5 (Jul. - Aug., 1895), pp.
Description of a New Species of Petalodus (P. Securiger) from the Carboniferous of Illinois Author(s): O. P. Hay Reviewed work(s): Source: The Journal of Geology, Vol. 3, No. 5 (Jul. - Aug., 1895), pp.
The os coxae or hip bone consists of three flat bones, ilium, ischium and pubis, which fuse together to form the acetabulum.
 The os coxae The os coxae or hip bone consists of three flat bones, ilium, ischium and pubis, which fuse together to form the acetabulum. The ilium extends from the acetabulum upwards forming the lateral
The os coxae The os coxae or hip bone consists of three flat bones, ilium, ischium and pubis, which fuse together to form the acetabulum. The ilium extends from the acetabulum upwards forming the lateral
Anatomic Relations Summary. Done by: Sohayyla Yasin Dababseh
 Anatomic Relations Summary Done by: Sohayyla Yasin Dababseh Anatomic Relations Lecture 1 Part-1 - The medial wall of the nose is the septum. - The vestibule lies directly inside the nostrils (Nares). -
Anatomic Relations Summary Done by: Sohayyla Yasin Dababseh Anatomic Relations Lecture 1 Part-1 - The medial wall of the nose is the septum. - The vestibule lies directly inside the nostrils (Nares). -
Concepts of occlusion Balanced occlusion. Monoplane occlusion. Lingualized occlusion. Figure (10-1)
") Any contact between teeth of opposing dental arches; usually, referring to contact between the occlusal surface. The static relationship between the incising or masticatory surfaces of the maxillary or
Any contact between teeth of opposing dental arches; usually, referring to contact between the occlusal surface. The static relationship between the incising or masticatory surfaces of the maxillary or
6610 NE 181st Street, Suite #1, Kenmore, WA
 660 NE 8st Street, Suite #, Kenmore, WA 9808 www.northshoredentalacademy.com.08.900 READ CHAPTER The Professional Dental Assistant (p.-9) No Key Terms Recall Questions:,,,, and 6 CLASS SYLLABUS DAY READ
660 NE 8st Street, Suite #, Kenmore, WA 9808 www.northshoredentalacademy.com.08.900 READ CHAPTER The Professional Dental Assistant (p.-9) No Key Terms Recall Questions:,,,, and 6 CLASS SYLLABUS DAY READ
Lecture. Permanent maxillary premolars
 Lecture Permanent maxillary premolars Permanent premolars The maxillary premolars are four in number: two in the right and two in the left. They are posterior to the canines and anterior to the molars.
Lecture Permanent maxillary premolars Permanent premolars The maxillary premolars are four in number: two in the right and two in the left. They are posterior to the canines and anterior to the molars.
Phylogenetics Lab: Character Descriptions
 Phylogenetics Lab: Character Descriptions 1) Osseous Auditory Canal. 0= absent, 1= present. Does the organism have a bony auditory canal? This will look like a hole or opening in the skull behind the jaw
Phylogenetics Lab: Character Descriptions 1) Osseous Auditory Canal. 0= absent, 1= present. Does the organism have a bony auditory canal? This will look like a hole or opening in the skull behind the jaw
 1 of 9 7/11/2012 11:29 AM For many years, producers, veterinarians, and exhibitors (at cattle shows) have used cattle dentition to make general age determinations. Dentition will vary from herd-to-herd
1 of 9 7/11/2012 11:29 AM For many years, producers, veterinarians, and exhibitors (at cattle shows) have used cattle dentition to make general age determinations. Dentition will vary from herd-to-herd
TOOTHED CETACEAN FROM
 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 76, NUMBER 7 DESCRIPTION OF AN APPARENTLY NEW TOOTHED CETACEAN FROM, SOUTH CAROLINA (With Two Plates) BY REMINGTON KELLOGG Bureau of Biological Survey, U. S.
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 76, NUMBER 7 DESCRIPTION OF AN APPARENTLY NEW TOOTHED CETACEAN FROM, SOUTH CAROLINA (With Two Plates) BY REMINGTON KELLOGG Bureau of Biological Survey, U. S.
Bones of the skull & face
 Bones of the skull & face Cranium= brain case or helmet Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. The cranium is composed of eight bones : frontal Occipital
Bones of the skull & face Cranium= brain case or helmet Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. The cranium is composed of eight bones : frontal Occipital
DEVELOPING ANALOGUE/SUBTITUTE FOR THE MANDIBULAR DENTURE BEARING AREA. Dr Muhammad Rizwan Memon FCPS Assistant Professor
 DEVELOPING ANALOGUE/SUBTITUTE FOR THE MANDIBULAR DENTURE BEARING AREA Dr Muhammad Rizwan Memon FCPS Assistant Professor Crest of Residual Ridge Buccal Shelf Shape of supporting structure Mylohyoid Ridge
DEVELOPING ANALOGUE/SUBTITUTE FOR THE MANDIBULAR DENTURE BEARING AREA Dr Muhammad Rizwan Memon FCPS Assistant Professor Crest of Residual Ridge Buccal Shelf Shape of supporting structure Mylohyoid Ridge
TRAUMA TO THE FACE AND MOUTH
 Dr.Yahya A. Ali 3/10/2012 F.I.C.M.S TRAUMA TO THE FACE AND MOUTH Bailey & Love s 25 th edition Injuries to the orofacial region are common, but the majority are relatively minor in nature. A few are major
Dr.Yahya A. Ali 3/10/2012 F.I.C.M.S TRAUMA TO THE FACE AND MOUTH Bailey & Love s 25 th edition Injuries to the orofacial region are common, but the majority are relatively minor in nature. A few are major
Dental Morphology and Vocabulary
 Dental Morphology and Vocabulary Palate Palate Palate 1 2 Hard Palate Rugae Hard Palate Palate Palate Soft Palate Palate Palate Soft Palate 4 Palate Hard Palate Soft Palate Maxillary Arch (Maxilla) (Uppers)
Dental Morphology and Vocabulary Palate Palate Palate 1 2 Hard Palate Rugae Hard Palate Palate Palate Soft Palate Palate Palate Soft Palate 4 Palate Hard Palate Soft Palate Maxillary Arch (Maxilla) (Uppers)
Skull-2. Norma Basalis Interna Norma Basalis Externa. Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology
 Skull-2 Norma Basalis Interna Norma Basalis Externa Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology Norma basalis interna Base of the skull- superior view The interior of the base of the
Skull-2 Norma Basalis Interna Norma Basalis Externa Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology Norma basalis interna Base of the skull- superior view The interior of the base of the
Variation in arch shape and dynamics of shape change from infancy to early childhood
 University of Iowa Iowa Research Online Theses and Dissertations Spring 2017 Variation in arch shape and dynamics of shape change from infancy to early childhood Gisela Lilian Borget University of Iowa
University of Iowa Iowa Research Online Theses and Dissertations Spring 2017 Variation in arch shape and dynamics of shape change from infancy to early childhood Gisela Lilian Borget University of Iowa
Periodontal Disease. Radiology of Periodontal Disease. Periodontal Disease. The Role of Radiology in Assessment of Periodontal Disease
 Radiology of Periodontal Disease Steven R. Singer, DDS srs2@columbia.edu 212.305.5674 Periodontal Disease! Includes several disorders of the periodontium! Gingivitis! Marginal Periodontitis! Localized
Radiology of Periodontal Disease Steven R. Singer, DDS srs2@columbia.edu 212.305.5674 Periodontal Disease! Includes several disorders of the periodontium! Gingivitis! Marginal Periodontitis! Localized
You know you would like to stop swearing at the computer after each shot. Troubleshooting oral radiography
 You know you would like to stop swearing at the computer after each shot Troubleshooting oral radiography Goals of oral radiology Achieve diagnostic images of the teeth and surrounding bone. Images should
You know you would like to stop swearing at the computer after each shot Troubleshooting oral radiography Goals of oral radiology Achieve diagnostic images of the teeth and surrounding bone. Images should
Adam J. Freeman, D.D.S., 22 Imperial Avenue Westport, Connecticut
 Adam J. Freeman, D.D.S., 22 Imperial Avenue Westport, Connecticut 06880 203 227-3709 Forensic Dental Report January 20, 2009 Introduction The information in this report details the examination and analysis
Adam J. Freeman, D.D.S., 22 Imperial Avenue Westport, Connecticut 06880 203 227-3709 Forensic Dental Report January 20, 2009 Introduction The information in this report details the examination and analysis
Prosthetic Options in Implant Dentistry. Hakimeh Siadat, DDS, MSc Associate Professor
 Prosthetic Options in Dentistry Hakimeh Siadat, DDS, MSc Associate Professor Dental Research Center, Department of Prosthodontics & Dental s Faculty of Dentistry, Tehran University of Medical Sciences
Prosthetic Options in Dentistry Hakimeh Siadat, DDS, MSc Associate Professor Dental Research Center, Department of Prosthodontics & Dental s Faculty of Dentistry, Tehran University of Medical Sciences
Case Report: Long-Term Outcome of Class II Division 1 Malocclusion Treated with Rapid Palatal Expansion and Cervical Traction
 Case Report Case Report: Long-Term Outcome of Class II Division 1 Malocclusion Treated with Rapid Palatal Expansion and Cervical Traction Roberto M. A. Lima, DDS a ; Anna Leticia Lima, DDS b Abstract:
Case Report Case Report: Long-Term Outcome of Class II Division 1 Malocclusion Treated with Rapid Palatal Expansion and Cervical Traction Roberto M. A. Lima, DDS a ; Anna Leticia Lima, DDS b Abstract:
Remember from the first year embryology Trilaminar disc has 3 layers: ectoderm, mesoderm, and endoderm
 Development of face Remember from the first year embryology Trilaminar disc has 3 layers: ectoderm, mesoderm, and endoderm The ectoderm forms the neural groove, then tube The neural tube lies in the mesoderm
Development of face Remember from the first year embryology Trilaminar disc has 3 layers: ectoderm, mesoderm, and endoderm The ectoderm forms the neural groove, then tube The neural tube lies in the mesoderm
PH-04A: Clinical Photography Production Checklist With A Small Camera
 PH-04A: Clinical Photography Production Checklist With A Small Camera Operator Name Total 0-49, Passing 39 Your Score Patient Name Date of Series Instructions: Evaluate your Series of photographs first.
PH-04A: Clinical Photography Production Checklist With A Small Camera Operator Name Total 0-49, Passing 39 Your Score Patient Name Date of Series Instructions: Evaluate your Series of photographs first.
Attachment G. Orthodontic Criteria Index Form Comprehensive D8080. ABBREVIATIONS CRITERIA for Permanent Dentition YES NO
 First Review IL HFS Dental Program Models Second Review Ortho cad Attachment G Orthodontic Criteria Index Form Comprehensive D8080 Ceph Film X-Rays Photos Narrative Patient Name: DOB: ABBREVIATIONS CRITERIA
First Review IL HFS Dental Program Models Second Review Ortho cad Attachment G Orthodontic Criteria Index Form Comprehensive D8080 Ceph Film X-Rays Photos Narrative Patient Name: DOB: ABBREVIATIONS CRITERIA
GOMPHOTHERIUM ELEGANS, FROM THE PLEISTO-
 DESCRIPTION OF A NEW SPECIES OF MASTODON, GOMPHOTHERIUM ELEGANS, FROM THE PLEISTO- CENE OF KANSAS. By Oliver P. Hay, Research As.iu.iiitc of the Carnegie In-stituti'in of Wn-:]iingto)i. The specimen here
DESCRIPTION OF A NEW SPECIES OF MASTODON, GOMPHOTHERIUM ELEGANS, FROM THE PLEISTO- CENE OF KANSAS. By Oliver P. Hay, Research As.iu.iiitc of the Carnegie In-stituti'in of Wn-:]iingto)i. The specimen here
Partners. 6. Occipital crest for attachment of 6. No occipital crest, neck muscles not
 Name Period Partners Primate and Human Evolution- A Skull Comparison Introduction Skulls are one of the most descriptive parts of an individual s skeleton. Skulls alone can give clues as to the age, sex,
Name Period Partners Primate and Human Evolution- A Skull Comparison Introduction Skulls are one of the most descriptive parts of an individual s skeleton. Skulls alone can give clues as to the age, sex,
Tikrit University collage of dentistry Dr.Ban I.S. head & neck anatomy 2 nd y. Lec [5] / Temporal fossa :
![Tikrit University collage of dentistry Dr.Ban I.S. head & neck anatomy 2 nd y. Lec [5] / Temporal fossa :](/thumbs/88/115294566.jpg "Tikrit University collage of dentistry Dr.Ban I.S. head & neck anatomy 2 nd y. Lec [5] / Temporal fossa :") Lec [5] / Temporal fossa : Borders of the Temporal Fossa: Superior: Superior temporal line. Inferior: gap between zygomatic arch and infratemporal crest of sphenoid bone. Anterior: Frontal process of the
Lec [5] / Temporal fossa : Borders of the Temporal Fossa: Superior: Superior temporal line. Inferior: gap between zygomatic arch and infratemporal crest of sphenoid bone. Anterior: Frontal process of the
GENERAL SCOPE AND USES OF PHYSICAL/BIOLOGICAL ANTHROPOLOGY. Paper No. & Title: B.A./B.Sc. (Honours) 2 dn semester. (Practical)
 2 dn semester. (Practical)") GENERAL SCOPE AND USES OF PHYSICAL/BIOLOGICAL ANTHROPOLOGY Course name: Physical Anthropology Paper No. & Title: B.A./B.Sc. (Honours) 2 dn semester (Practical) Topic No. & Title: 5/12 (Part-I) Drawing
GENERAL SCOPE AND USES OF PHYSICAL/BIOLOGICAL ANTHROPOLOGY Course name: Physical Anthropology Paper No. & Title: B.A./B.Sc. (Honours) 2 dn semester (Practical) Topic No. & Title: 5/12 (Part-I) Drawing
ANATOMY OF JUNGLE CAT SKULL (FELIS CHAUS, SCHREBER, 1777)
") CASE REPORT ZOOS' PRINT JOURNAL (): 0-0 ANATOMY OF JUNGLE CAT SKULL (FELIS CHAUS, SCHREBER, ) P.O. Nameer, P.O. Naseer, M.O. Ipe and P.A. Ommer Assistant Professor, Department of Wildlife Sciences, College
CASE REPORT ZOOS' PRINT JOURNAL (): 0-0 ANATOMY OF JUNGLE CAT SKULL (FELIS CHAUS, SCHREBER, ) P.O. Nameer, P.O. Naseer, M.O. Ipe and P.A. Ommer Assistant Professor, Department of Wildlife Sciences, College
POSTILLA PEABODY MUSEUM YALE UNIVERSITY NUMBER NOV OCCLUSION IN DOCODON (MAM MALIA, DOCODONTA) FARISH A. JENKINS, JR.
 FARISH A. JENKINS, JR.") POSTILLA PEABODY MUSEUM YALE UNIVERSITY NUMBER 139. 20 NOV. 1969 OCCLUSION IN DOCODON (MAM MALIA, DOCODONTA) FARISH A. JENKINS, JR. POSTILLA Published by the Peabody Museum of Natural History, Yale University
POSTILLA PEABODY MUSEUM YALE UNIVERSITY NUMBER 139. 20 NOV. 1969 OCCLUSION IN DOCODON (MAM MALIA, DOCODONTA) FARISH A. JENKINS, JR. POSTILLA Published by the Peabody Museum of Natural History, Yale University
Cephalometric Analysis
 Cephalometric Analysis of Maxillary and Mandibular Growth and Dento-Alveolar Change Part III In two previous articles in the PCSO Bulletin s Faculty Files, we discussed the benefits and limitations of
Cephalometric Analysis of Maxillary and Mandibular Growth and Dento-Alveolar Change Part III In two previous articles in the PCSO Bulletin s Faculty Files, we discussed the benefits and limitations of
Figure S1: Distal Humerus
 Figure S: Distal Humerus Criteria The distal part of the medial epicondyle forms The distal part of the medial epicondyle forms an a right angle. oblique angle. Viewed from the lateral aspect, the pit
Figure S: Distal Humerus Criteria The distal part of the medial epicondyle forms The distal part of the medial epicondyle forms an a right angle. oblique angle. Viewed from the lateral aspect, the pit
Human Male Asian Skeleton, Robust
 Human Male Asian Skeleton, Robust Product Number: Specimen Evaluated: Skeletal Inventory: SC-287 Original Specimen Near-complete human skeleton with 28 teeth. Osteological Observations: This is a clean,
Human Male Asian Skeleton, Robust Product Number: Specimen Evaluated: Skeletal Inventory: SC-287 Original Specimen Near-complete human skeleton with 28 teeth. Osteological Observations: This is a clean,
The orbit-1. Dr. Heba Kalbouneh Assistant Professor of Anatomy and Histology
 The orbit-1 Dr. Heba Kalbouneh Assistant Professor of Anatomy and Histology Orbital plate of frontal bone Orbital plate of ethmoid bone Lesser wing of sphenoid Greater wing of sphenoid Lacrimal bone Orbital
The orbit-1 Dr. Heba Kalbouneh Assistant Professor of Anatomy and Histology Orbital plate of frontal bone Orbital plate of ethmoid bone Lesser wing of sphenoid Greater wing of sphenoid Lacrimal bone Orbital
Lips and labial mucosa
 Lips and labial mucosa External portion of the lips: the vermilion border and the skin Vermilion border : the exposed red portion of the lip, covered by mucous membrane, no mucous glands Boundary: the
Lips and labial mucosa External portion of the lips: the vermilion border and the skin Vermilion border : the exposed red portion of the lip, covered by mucous membrane, no mucous glands Boundary: the
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Stirton, R. A., and L. F. Marcus, 1966. Generic and specific diagnoses in the gigantic macropodid genus Procoptodon. Records of the Australian Museum 26(16): 349
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Stirton, R. A., and L. F. Marcus, 1966. Generic and specific diagnoses in the gigantic macropodid genus Procoptodon. Records of the Australian Museum 26(16): 349
Subfamily Amphicyonina.
 Article XX1.-NEW C A N I D B FROM T H E MIOCENE OF COLORADO. Cynarctus, new genus. Subfamily Amphicyonina. Family Ca~zida, Dentition 3.1.4.3. Carnassials reduced and molars enlarged, talonids bicuspid
Article XX1.-NEW C A N I D B FROM T H E MIOCENE OF COLORADO. Cynarctus, new genus. Subfamily Amphicyonina. Family Ca~zida, Dentition 3.1.4.3. Carnassials reduced and molars enlarged, talonids bicuspid
With judicious treatment planning, the clinical
 CLINICIAN S CORNER Selecting custom torque prescriptions for the straight-wire appliance Earl Johnson San Francisco, Calif Selecting custom torque prescriptions based on the treatment needs of each patient
CLINICIAN S CORNER Selecting custom torque prescriptions for the straight-wire appliance Earl Johnson San Francisco, Calif Selecting custom torque prescriptions based on the treatment needs of each patient
Morphological Scoring of Dental Casts Using the Arizona State University Dental Anthropology System
 University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2004 Morphological Scoring of
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2004 Morphological Scoring of
Oligocene squalodont (Cetacea: Mammalia) from the Ashiya Group, Japan
 from the Ashiya Group, Japan") Bull. Kitakyushu Mus. Nat. Hist., 8: 75-80. December 27, 1988 Oligocene squalodont (Cetacea: Mammalia) from the Ashiya Group, Japan Yoshihiko Okazaki Kitakyushu Museum of Natural History, Kitakyushu, 805,
Bull. Kitakyushu Mus. Nat. Hist., 8: 75-80. December 27, 1988 Oligocene squalodont (Cetacea: Mammalia) from the Ashiya Group, Japan Yoshihiko Okazaki Kitakyushu Museum of Natural History, Kitakyushu, 805,
Indirect retainers. 1 i
 8 1 i Indirect retainers Factors Influencing Effectiveness Indirect Retainers Auxiliary Functions Indirect Retainers Forms Indirect Retainers Auxiliary occlusal rest Canine extensions fiom occlusal rests
8 1 i Indirect retainers Factors Influencing Effectiveness Indirect Retainers Auxiliary Functions Indirect Retainers Forms Indirect Retainers Auxiliary occlusal rest Canine extensions fiom occlusal rests
Anatomy and Physiology. Bones, Sutures, Teeth, Processes and Foramina of the Human Skull
 Anatomy and Physiology Chapter 6 DRO Bones, Sutures, Teeth, Processes and Foramina of the Human Skull Name: Period: Bones of the Human Skull Bones of the Cranium: Frontal bone: forms the forehead and the
Anatomy and Physiology Chapter 6 DRO Bones, Sutures, Teeth, Processes and Foramina of the Human Skull Name: Period: Bones of the Human Skull Bones of the Cranium: Frontal bone: forms the forehead and the
DENT Advanced Topics in Removable Prosthodontics, Winter 2008
 University of Michigan Deep Blue deepblue.lib.umich.edu 2008-01 DENT 718 - Advanced Topics in Removable Prosthodontics, Winter 2008 Shotwell, Jeffrey Shotwell, J. (2008, April 23) Advanced Topics in Removable
University of Michigan Deep Blue deepblue.lib.umich.edu 2008-01 DENT 718 - Advanced Topics in Removable Prosthodontics, Winter 2008 Shotwell, Jeffrey Shotwell, J. (2008, April 23) Advanced Topics in Removable
Any of the vertebra in the cervical (neck) region of the spinal column. The cervical vertebra are the smallest vertebra in the spine, reflective of th
 region of the spinal column. The cervical vertebra are the smallest vertebra in the spine, reflective of th") Any of the vertebra in the cervical (neck) region of the spinal column. The cervical vertebra are the smallest vertebra in the spine, reflective of the fact that they support the least load. In humans,
Any of the vertebra in the cervical (neck) region of the spinal column. The cervical vertebra are the smallest vertebra in the spine, reflective of the fact that they support the least load. In humans,
Examination of Wear Striation Direction
 A Preliminary Scanning Electron Microscope Examination of Wear Striation Direction on Primate Teeth ALAN S. RYAN Department of Anthropology, The University ofmichigan, Ann Arbor, Michigan 48109 Wear striations
A Preliminary Scanning Electron Microscope Examination of Wear Striation Direction on Primate Teeth ALAN S. RYAN Department of Anthropology, The University ofmichigan, Ann Arbor, Michigan 48109 Wear striations
Dr.Noor Hashem Mohammad Lecture (5)
") Dr.Noor Hashem Mohammad Lecture (5) 2016-2017 If the mandible is discarded, the anterior part of this aspect of the skull is seen to be formed by the hard palate. The palatal processes of the maxillae
Dr.Noor Hashem Mohammad Lecture (5) 2016-2017 If the mandible is discarded, the anterior part of this aspect of the skull is seen to be formed by the hard palate. The palatal processes of the maxillae
Dentalelle Tutoring - Faulty Radiographs
 Dentalelle Tutoring - Faulty Radiographs Errors in improperly exposing or processing dental films can produce undesirable dental radiographs of nondiagnostic quality. These are known as faulty radiographs.
Dentalelle Tutoring - Faulty Radiographs Errors in improperly exposing or processing dental films can produce undesirable dental radiographs of nondiagnostic quality. These are known as faulty radiographs.
ORTHODONTICS Treatment of malocclusion Assist.Lec.Kasem A.Abeas University of Babylon Faculty of Dentistry 5 th stage
 Lec: Treatment of class I malocclusion Class I occlusion can be defined by Angles, classification as the mesiobuccal cusp of the upper 1 st permanent molar occlude with the developmental groove of the
Lec: Treatment of class I malocclusion Class I occlusion can be defined by Angles, classification as the mesiobuccal cusp of the upper 1 st permanent molar occlude with the developmental groove of the
Temporal fossa Infratemporal fossa Pterygopalatine fossa Terminal branches of external carotid artery Pterygoid venous plexus
 Outline of content Temporal fossa Infratemporal fossa Pterygopalatine fossa Terminal branches of external carotid artery Pterygoid venous plexus Boundary Content Communication Mandibular division of trigeminal
Outline of content Temporal fossa Infratemporal fossa Pterygopalatine fossa Terminal branches of external carotid artery Pterygoid venous plexus Boundary Content Communication Mandibular division of trigeminal
THE CANADA AND ALBERTA BSE SURVEILLANCE PROGRAM (CABSESP) GUIDELINES FOR AGE VERIFICATION IN CATTLE 1
 GUIDELINES FOR AGE VERIFICATION IN CATTLE 1") AGRICULTURE AND RURAL DEVELOPMENT Food Safety and Animal Health Division, Animal Health Branch 9 th Floor, O.S. Longman Building 6909-116 Street Edmonton, Alberta T6H 4P2 Telephone 780-644-2148 Fax 780-422-5734
AGRICULTURE AND RURAL DEVELOPMENT Food Safety and Animal Health Division, Animal Health Branch 9 th Floor, O.S. Longman Building 6909-116 Street Edmonton, Alberta T6H 4P2 Telephone 780-644-2148 Fax 780-422-5734
IMPACTED CANINES. Unfortunately, this important tooth is the second most common tooth to be impacted after third molars
 IMPACTED CANINES After we talked about impacted third molars, today we ll discuss about maxillary impacted canines in upper dental arch, how to manage these cases as a dental surgeon. You will study about
IMPACTED CANINES After we talked about impacted third molars, today we ll discuss about maxillary impacted canines in upper dental arch, how to manage these cases as a dental surgeon. You will study about
Postnatal Growth. The study of growth in growing children is for two reasons : -For health and nutrition assessment
 Growth of The Soft Tissues Postnatal Growth Postnatal growth is defined as the first 20 years of growth after birth krogman 1972 The study of growth in growing children is for two reasons : -For health
Growth of The Soft Tissues Postnatal Growth Postnatal growth is defined as the first 20 years of growth after birth krogman 1972 The study of growth in growing children is for two reasons : -For health
Chapter XII: Temporal Expanding Processes, Including Those in the Sylvian Fissure and the Insula
 Acta Radiologica ISSN: 0001-6926 (Print) (Online) Journal homepage: https://www.tandfonline.com/loi/iaro20 Chapter XII: Temporal Expanding Processes, Including Those in the Sylvian Fissure and the Insula
Acta Radiologica ISSN: 0001-6926 (Print) (Online) Journal homepage: https://www.tandfonline.com/loi/iaro20 Chapter XII: Temporal Expanding Processes, Including Those in the Sylvian Fissure and the Insula
Hedgehogs (Erinaceidae, Lipotyphla) from the Miocene of Pakistan, with description of a new species of Galerix
 from the Miocene of Pakistan, with description of a new species of Galerix") Hedgehogs (Erinaceidae, Lipotyphla) from the Miocene of Pakistan, with description of a new species of Galerix The Harvard community has made this article openly available. Please share how this access
Hedgehogs (Erinaceidae, Lipotyphla) from the Miocene of Pakistan, with description of a new species of Galerix The Harvard community has made this article openly available. Please share how this access
evolution and development of primate teeth
 evolution and development of primate teeth diversity of mammalian teeth upper left molars buccal mesial distal lingual Jernvall & Salazar-Ciudad 07 trends in dental evolution many similar single-cusped
evolution and development of primate teeth diversity of mammalian teeth upper left molars buccal mesial distal lingual Jernvall & Salazar-Ciudad 07 trends in dental evolution many similar single-cusped
Muscles of mastication [part 1]
![Muscles of mastication [part 1]](/thumbs/76/73586850.jpg "Muscles of mastication [part 1]") Muscles of mastication [part 1] In this lecture well have the muscles of mastication, neuromuscular function, and its relationship to the occlusion morphology. The fourth determinant of occlusion is the
Muscles of mastication [part 1] In this lecture well have the muscles of mastication, neuromuscular function, and its relationship to the occlusion morphology. The fourth determinant of occlusion is the
Permanent 2 nd Maxillary Molars
 Permanent 2 nd Maxillary Molars In comparison to the first max molar First molars appears in the oral cavity at the age of 6 years old.. While 2 nd molar 3 rd molar Max. 2 nd molar have long roots (sometimes
Permanent 2 nd Maxillary Molars In comparison to the first max molar First molars appears in the oral cavity at the age of 6 years old.. While 2 nd molar 3 rd molar Max. 2 nd molar have long roots (sometimes
Intraoral radiographic techniques Introduction There are three main types of intraoral radiographs: Periapical radiograph Bitewing radiograph
 Intraoral radiographic techniques Introduction There are three main types of intraoral radiographs: Periapical radiograph Bitewing radiograph Occlusal radiograph The anatomic area of interest and type
Intraoral radiographic techniques Introduction There are three main types of intraoral radiographs: Periapical radiograph Bitewing radiograph Occlusal radiograph The anatomic area of interest and type
Advanced Probing Techniques
 Module 21 Advanced Probing Techniques MODULE OVERVIEW The clinical periodontal assessment is one of the most important functions performed by dental hygienists. This module begins with a review of the
Module 21 Advanced Probing Techniques MODULE OVERVIEW The clinical periodontal assessment is one of the most important functions performed by dental hygienists. This module begins with a review of the
What is the primary job of a forensic anthropologist? What are the three main things that can determined from a skeleton?
 What is the primary job of a forensic anthropologist? What are the three main things that can determined from a skeleton? What three areas of a skeleton can be used to determine sex? Physical anthropologists
What is the primary job of a forensic anthropologist? What are the three main things that can determined from a skeleton? What three areas of a skeleton can be used to determine sex? Physical anthropologists
Amalgam restoration of posterior proximal cavities with deep and concave gingival outlines
 Amalgam restoration of posterior proximal cavities with deep and concave gingival outlines M. Darveniza, MDSc, FRACDS* Abstract A technique for making a matrix and wedges for amalgam restoration of deep
Amalgam restoration of posterior proximal cavities with deep and concave gingival outlines M. Darveniza, MDSc, FRACDS* Abstract A technique for making a matrix and wedges for amalgam restoration of deep
Dr. Sami Zaqout, IUG Medical School
 The skull The skull is composed of several separate bones united at immobile joints called sutures. Exceptions? Frontal bone Occipital bone Vault Cranium Sphenoid bone Zygomatic bones Base Ethmoid bone
The skull The skull is composed of several separate bones united at immobile joints called sutures. Exceptions? Frontal bone Occipital bone Vault Cranium Sphenoid bone Zygomatic bones Base Ethmoid bone
#45 Ortho-Tain, Inc PREVENTIVE ERUPTION GUIDANCE -- PREVENTIVE OCCLUSAL DEVELOPMENT
 #45 Ortho-Tain, Inc. 1-800-541-6612 PREVENTIVE ERUPTION GUIDANCE -- PREVENTIVE OCCLUSAL DEVELOPMENT Analysis and Diagnosis of Occlusion: The ideal child of 5 y ears of age that probably has the best chance
#45 Ortho-Tain, Inc. 1-800-541-6612 PREVENTIVE ERUPTION GUIDANCE -- PREVENTIVE OCCLUSAL DEVELOPMENT Analysis and Diagnosis of Occlusion: The ideal child of 5 y ears of age that probably has the best chance
Chapter 2. Material and methods
 Chapter 2 Material and methods Material and methods Summary This chapter describes the subjects and methods being used in this study. Between 1986 and 1997 9 expeditions were undertaken in remote areas
Chapter 2 Material and methods Material and methods Summary This chapter describes the subjects and methods being used in this study. Between 1986 and 1997 9 expeditions were undertaken in remote areas