JBC Papers in Press. Published on June 13, 2013 as Manuscript M
|
|
|
- Spencer Nash
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on June 13, 2013 as Manuscript M The latest version is at Novel signaling pathway promotes contextual discrimination Acquisition of Contextual Discrimination Involves the Appearance of a Ras-GRF1/p38 Map Kinase- Mediated Signaling Pathway that Promotes LTP Shan-Xue Jin 1, Junko Arai 1, Xuejun Tian 1, Rajendra Kumar-Singh 2, Larry A. Feig 1,3, From the Departments of Biochemistry 1, and Ophthalmology 2, Tufts University School of Medicine, Boston, MA, *Running title: Novel LTP pathway contributes to contextual discrimination To whom correspondence should be addressed: Larry A. Feig, Department of Biochemistry, Tufts University School of Medicine, 136 Harrison Ave. Boston MA, Tel: , larry.feig@tufts.edu Keywords: Ras-GRF1, contextual discrimination, p38 Map kinase, synaptic plasticity, calciumpermeable AMPA receptors Background: The ability to distinguish similar experiences is a critical aspect of memory. Results: Contextual discrimination involves LTP promoted by calcium-permeable AMPA-type glutamate receptors, Ras-GRF1 and p38 Map kinase. Conclusion: A newly discovered, LTP-supporting, signaling pathway contributes to a key component of memory formation. Significance: Defects in this signaling pathway may contribute to cognitive loss associated with neurological disorders. SUMMARY Ras-GRF1 is a guanine nucleotide exchange factor with the ability to activate Ras and Rac GTPases in response to elevated calcium levels. We previously showed that beginning at 1- month of age, Ras-GRF1 mediates NMDA-type glutamate receptor (NMDAR)-induction of LTD in the CA1 region of the hippocampus of mice. Here we show that beginning at 2- months of age, when mice first acquire the ability to discriminate between closely related contexts, Ras-GRF1 begins to contribute to the induction of LTP in the CA1 hippocampus by mediating the action of calcium-permeable, AMPA-type glutamate receptors (CP- AMPARs). Surprisingly, LTP-induction by CP-AMPARs through Ras-GRF1 occurs via activation of p38 Map kinase rather than Erk Map kinase, which has more frequently been linked to LTP. Moreover, contextual discrimination is blocked by knockdown of Ras-GRF1 expression specifically in the CA1 hippocampus, infusion of a p38 Map kinase inhibitor into the CA1 hippocampus, or the injection of an inhibitor of CP-AMPARs. These findings implicate the CA1 hippocampus in the developmentally dependent capacity to distinguish closely related contexts through the appearance of a novel LTP-supporting signaling pathway. It is well known that the hippocampus plays a critical role in processing contextual information relating to episodic memory. Moreover, the contribution of the hippocampus to learning and memory develops late among brain regions, appearing first ~21 days after birth in mammals (1). Among this category of functions, the hippocampus is particularly important for discriminating similar contexts, which requires processing of not only spatial but also temporal and relational information (2). In fact, clearly distinct roles for the dentate gyrus, CA3 and CA1 regions of the hippocampus in this form of learning have been identified (3). However, the biochemical underpinnings of these functions remain poorly understood. Likely candidates include signaling components that regulate synaptic plasticity in each of these hippocampus segments. 1 Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc.
2 One class of signaling molecules that have been implicated in synaptic plasticity are the Ras-GRF1 and Ras-GRF2 guanine nucleotide exchange factors (4). These proteins have the capacity to activate Ras and Rac GTPases through their CDC25 and DH domains, respectively. Both proteins can be activated by calcium-dependent binding of calmodulin to their IQ motifs. Beginning at ~1 month of age Ras-GRF1 mediates NMDA receptor (NMDAR)-induced LTD, whereas Ras-GRF2 mediates NMDAR-induced LTP in the CA3/CA1 synapse of the hippocampus (5). At this site, Ras-GRF2 mediates LTP induced specifically by NMDARs that contain GluN2A subunits (6). Consistent with their defective LTP, Ras-GRF2 knockout mice display defective contextual fear conditioning memory (Feig, unpublished observations). In contrast, Ras-GRF1 mice display more subtle fear memory defects in that contextual fear memory is weakly defective (7,8), but contextual discrimination is completely lost (7). How Ras-GRF1 contributes to contextual discrimination is not known. Map kinases play integral roles in neuronal functions related to synaptic plasticity associated with learning and memory(9). Erk Map kinase in synapses of mature animals can be activated by a variety of upstream receptors, including the Trk family of tyrosine kinase receptors (10) and both NMDA type (11) and calcium-permeable AMPA type glutamate receptors (11,12). Its activation is a critical step in promoting many forms of LTP, including protein synthesis-independent early-ltp induced by theta-burst stimulation that is thought to contribute to short-term memory (13). It is also critical for protein-dependent late-ltp and longterm memory (14). In contrast, p38 Map kinase has been most often associated with LTD, although conflicting results have been obtained (5,15-17). However, p38 Map kinase activity has been associated with theta-burst induced early- LTP that is associated with enhanced learning after exposure to an enriched environment (18). In this paper, we demonstrate that p38 Map kinase activation is also involved in an agedependent LTP mechanism, mediated by calciumpermeable AMPA receptors and Ras-GRF1, that contributes to the acquisition at ~2-months of age of the ability of mice to distinguish closely related contexts. EXPERIMENTAL PROCEDURES Mice. ras-grf1(-/-) mice in a C57Bl6 background were described previously (5,19). All procedures involving mice were in accordance with the animal welfare guidelines of Tufts University. Extracellular field experiments. Mutant and control mice (1-6 month-old) were anesthetized with halothane and decapitated. The transverse acute hippocampal slices (350 µm) were cut in ice-cold oxygenated sucrose-enhanced artificial cerebrospinal fluid (ACSF) containing 206 mm sucrose, 2 mm KCl, 2 mm MgSO 4, 1.25 mm NaH 2 PO 4, 1 mm CaCl 2, 1 mm MgCl 2, 26 mm NaHCO 3, 10 mm D-glucose, ph 7.4. saturated with 95% O 2 and 5% CO 2 (ph7.4), in which they were allowed to recover for at least 90 min before recording. Recordings were performed in the same solution at room temperature in a chamber submerged in ACSF. To record field EPSPs (fepsps) in the CA1, CA3 and DG regions of the hippocampus respectively, standard procedures were used. 1) For CA1, Schaffer collaterals were stimulated with a unipolar stimulating electrode (World Precision Instruments) placed in the lateral CA1 subfield. A borosilicate glass recording electrode filled with ACSF was positioned in stratum radiatum of CA1. 2) For CA3, the mossy fiber CA3 pyramidal neuron responses were induced by the stimulation of mossy fiber axons with a unipolar stimulating electrode. A borosilicate glass recording electrode filled with ACSF were placed at least 500 µm from the stimulating electrodes along the trajectory of the mossy fiber pathway. D-APV (50 µm) was added in ACSF to prevent contamination with the NMDA receptor-dependent pathway converging on CA3 neurons. 3) For DG, stimulating and recording electrodes were placed in the medial aspect of the dentate molecular layer. 10 µm bicuculline methiodide was added in ACSF to prevent GABA A -mediated inhibition. Test stimuli were applied at low frequency (0.05 Hz) at a stimulus intensity that elicited an fepsp amplitude that was 50% of maximum, and the test responses were recorded for 10 min before the experiment was begun to ensure stability of the response. To induce LTP, we used three protocols in this study: (1) High frequency stimulation (HFS)- induced LTP in the CA1 and CA3: two 2
3 consecutive trains (1 s) of stimuli at 100 Hz separated by 20 s were applied to the CA1 and CA3 areas respectively. (2) theta-burst stimulation (TBS) in CA1: 15 bursts of four pulses at 100 Hz delivered at an interburst interval of 200 ms were applied to the CA1 area; (3) four bursts of 50 pulses at 100 Hz, 30 s between bursts were applied to the dentate gyrus. Traces were obtained by pclamp 9.2 and analyzed using the Clampfit 9.2. Data analysis was as follows: The fepsp magnitude was measured using the initial fepsp slope and three consecutive slopes (1 min) were averaged and normalized to the mean value recorded 10 min before conditioning stimulus. Data were pooled across animals of the same treatment and genotype and are presented as mean ±SEM. Values expressed here represent 50 min time points after conditioning stimulus was initiated. The following statistical analysis was carried out: the same time window samples of the control (cont) and treatment were compared using paired, two-tailed Student s t test. An effect was considered significant if p < Hippocampal brain slices preparation for immunofluorescence. The brain was removed quickly and submerged in ice-cold oxygenated sucrose-replaced ACSF cutting solution [containing the following (in mm): 240 sucrose, 2.5 KCI, 1.25 NaH 2 PO 4, 0.5 CaCl 2, 25 NaHCO 3, 7 MgCl 2, ph 7.4]. After dissection, slices (350 µm, transverse) were incubated in ACSF that contained the following (in mm): 124 NaCl, 2.8 KCl, 1.5 MgSO 4, 1.25 NaH 2 PO 4, 2.5 CaCl 2, 26 NaHCO 3, 10 D-glucose saturated with 95% O 2 and 5% CO 2 (ph7.4), in which they were allowed to recover for at least 2 hr before an experiment. Then a single slice was transferred to the recording chamber and submerged beneath a continuously perfusing the same ACSF. A unipolar stimulating electrode were placed in the stratum radiatum, and HFS or TBS stimulation was delivered with an intensity of 140 µa. Slices were fixed in 4% paraformaldehyde 10 min after HFS stimulation for overnight, then replaced with sucrose 24 h, and re-cut in cryostat at 18 µm. the sections were blocked with 2% Donkey serum in 0.3% Triton X-100 for 1 hr at room temperature (RT) and incubated over two nights at 4 C with anti-p-p38 antibody (anti-rabbit, 1:400; Cell Signaling). The sections were then incubated for 1 hr at RT with Cy3-conjugated secondary antibody (1:300; Jackson ImmunoResearch, West Grove, PA). The stained sections were examined with a Nikon (Tokyo, Japan) fluorescence microscope, and images were captured with a CCD spot camera, and subsequently analyzed with ImageJ software (National Institute of Health). For cell counting in stratum pyramidal region of stimulated regions ( µm from the location of the stimulating electrode), the three most representative and/or most complete sections from each slice were chosen and labeled cells in a 20 X optic field on each section was counted. Viral vectors- GRF1 mirna adeno-associated virus: An AAV8-shRNA vector pfb-u6-r2- CMVmCherry expressing mirna against GRF1 (seq- TGCTGTTGACAGTGAGCGCGCTGATCAATA TCTACAGAAATAGTGAAGCCACAGATGTA TTTCTGTAGATATTGATCAGCTTGCCTACT GCCTCGAA) or a similar control random sequence, along with mcherry or GFP under the control of the U6 and CMV promoters, respectively were generated by Virovek ( and were produced at working concentrations of ~1.6 X10 13 /ml. Adenovirus expressing Ras-GRF1 from the synapsin promoter was generated as described (20,21). Briefly, the synapsin promoter and polya regions were digested out of a plasmid provided by Dr. Colin Sumners (University of Florida), then ligated into pshuttle (20). Then Ras-GRF1 cdna was inserted in the resulting plasmid. pshsyngrf1 was digested with PmeI, then used to co-transform E.coli strain BJ5183 along with padeasy-i. Colonies were screened for the presence of GRF1 gene and the padeasy-i backbone by colony PCR. The positive clones were tested for the correct orientation by restriction enzyme digestion. The plasmid was then digested with PacI and transfected into 293 cells. Then the culture supernatant containing viral particles was harvested. Cleared supernatant was used for subsequent serial infection of 293 cells. Resultant viral samples were subjected to adenovirus purification using Adenopure kit (Puresyn Inc.). The virus was concentrated by filtration, yielding a concentration greater than 1x1012 vp/ml. Stereotaxic surgery. 2 month-old ras-grf1 knockout mice or WT mice were anesthetized with an i.p. injection of ketamine (100 mg/kg)/ xylazine 3
4 (10 mg/kg). Once anesthetized, each mouse was placed in a stereotaxic frame (myneurolab, St. Louis, MO). A surgical incision was made along the midline of the head to expose the skull. Two holes were made in the skull overlying the dorsal hippocampus. Coordinates for CA1 injection or cannula implantation into the mice: 2.0 mm posterior to Bregma, ±1.9 mm lateral from the midline, 1.9 mm or 0.9 mm (Cannula implantation) beneath the surface of the skull). Injections were performed with a 10 µl Hamilton syringe fitted with a custom made blunt ended 30- gauge needle (Hamilton). Each injection consisted of 1 µl of Adenovirus expressing GRF1 or Adeno- Associated virus (AAV-8) expressing GRF1 shrna with mcherry infused at a rate of 0.06 µl/min. An infusion pump controlling the plunger on the Hamilton syringe precisely regulated the rate of injection. The needle was then left in place for 8 min prior to withdrawal from the brain. Electrophysiological recordings or behavioral procedures began 7-11 days after stereotaxic injection of the viral vectors. In some experiments, a stainless steel double guide cannulas (21 gauge) were implanted bilaterally into the hemisphere. Mice were given at least 1 week to recover after cannula implantation. The 28-gauge injection cannula was 1 mm longer than the guide. For intrahippocampal infusion, Birb0796 was prepared by dissolving in DMSO and was diluted in sterile saline to a concentration of mg/ml in 10% DMSO. Vehicle-treated animals were infused with 90% saline and 10% DMSO. 1 µl /side of either drug or vehicle were performed in freely moving animals at a rate of 0.15 µl /min using a syringe infusion pump. Behavior assays. Mice were tested in contextual discrimination as previously described (7). In this task the mice were trained to discriminate between two chambers, one in which they were shocked and another in which they were not. On day 1 the mice were pre-exposed to the two contexts for 10 min. On day 2 the mice were shocked in one context (paired): after 148 seconds a 2 second shock (0.75 ma) was delivered and the mice remained for another 30 seconds in the context. Additionally, the mice were exposed for 3 min to the other context (non-paired) during which time no shocks were delivered. On day 3 the mice were shocked again in the paired context and exposed to the non-paired context. On day 4 the amount of freezing was measured for 3 min in each of the two contexts. In some experiments Adeno-associated virus expressing, GFP, control random shrna or shrna against GRF1 were injected stereotaxically into the CA1 hippocampus 7-10 days before beginning the experiment. In other experiments, the CP-AMPA receptor selective blocker IEM-1460 (Tocris Bioscience, Ellisville, MI) was dissolved in filtered saline and administered via intraperitoneal injections. In another group of experiments, animals received an IP injection of either saline or IEM-1460 each day of the experiments and 30 minutes later were trained in context A on day 1, day 2, day 3 respectively. In another group of experiments, animals received an intrahippocampal infusion of either vehicle, SB (0.75 µg/side) or Birb0796 (1.056 µg/side) each day of the experiments and 30 minutes later were trained in context A on day 1, day 2, day 3 respectively. animals received an intrahippocampal infusion of either vehicle, SB or Birb0796 each day of the experiments and 30 minutes later were trained in context A on day 1, day 2, day 3 respectively. Contextual fear conditioning was assessed by measuring the amount of freezing displayed by mice after they were placed back into the context where a single shock had been delivered 24 hrs previously. Mice were pretreated with shrna against GRF1, BIRB0796 or IEM as described above. The results were tested with a one-way analysis of variance (ANOVA). Histology. After the behavioral experiments, mice were killed by ketamine/xylazine overdose and intracardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (ph 7.4). Brains were sliced coronally (40 µm). The section was mounted on gelatin-subbed slides and stained with crystal violet. The location of the site of infusion was evaluated and recorded for each animal. RESULTS Ras-GRF1 begins to contribute to HFS-induced LTP at 2-months of age in the CA1, but not CA3 or dentate gyrus regions of the hippocampus. Using Ras-GRF knockout mice, we previously showed that Ras-GRF1 is predominantly involved in mediating NMDAR activation of LTD, while Ras- GRF2 is predominantly involved in mediating NMDAR activation of LTP at the CA3/CA1 4
5 synapse beginning at ~pn day 30 (early adolescence) in mice (5). However, because most behavior assays are performed with mice that are at least approaching late adolescence at 2- months of age, synaptic plasticity was retested in older Ras-GRF1 knockout mice. To our surprise, 2- month old Ras-GRF1 knockout mice (as well as 6- month old mice (data not shown)) displayed defective LTP-induced by high frequency stimulation (HFS) (two trains of 100 Hz tetanus) (Fig. 1A)[WT, ±4.73% (n=11 slices from six mice) vs ±2.54% (n=12 slices from eight grf1 (-/-) mice; p<0.01]. Interestingly, LTP generated by thetaburst stimulation (TBS) (15 bursts of four pulses at 100 Hz delivered at an interburst interval of 200 ms) was normal in 2 month old Ras-GRF1 knockout mice (Fig. 1B) [WT, ±2.77% (n=8 slices from five mice) vs ±1.69% (n=9 slices from five grf1(-/-) mice; p>0.05] as we showed previously for 1-month old mice (5). 2- month old Ras-GRF2 knockout mice show the opposite results, normal HFS-LTP but defective TBS-LTP (data not shown) Confirming that this defect was not due to the indirect effects of the loss of the Ras-GRF1 in neighboring brain regions or during development, defective HFS LTP in Ras-GRF1 knockout mice was restored to wild-type levels by re-expression of Ras-GRF1 in principle neurons of the CA1 hippocampus after injection of adenovirusencoding Ras-GRF1 into the CA1 hippocampus of 2 month-old Ras-GRF1 knockout mice (Fig. 1C) [Ras-GRF1, ±2.31% (n=10 slices from 5 mice); GFP, ±3.16% (n=13 slices from nine); p<0.01]. Moreover, Ras-GRF1 expression was suppressed acutely by stereotaxic injection of adeno-associated virus expressing shrna against GRF1 and the red cherry fluorophore into the CA1 region of the dorsal hippocampus. The shrna sequence is specific because it blocks the expression of co-transfected Ras-GRF1, but not Ras-GRF2, in HEK 293 cells (Fig. 2A). As expected, HFS-induced LTP was suppressed compared to infection with a control random shrna-encoding virus (Fig. 2B upper graph) (AAV-control, ±1.75%; n=8; AAVshRNA-GRF1, HFS, ±2.32%; n=9; p<0.01). The effect of virus infection was not as severe as that observed in the GRF1 knockout mice presumably because not all pyramidal cells were infected and GRF1 loss was not complete. Importantly, theta-burst induced LTP was not suppressed by GRF1 knockdown (Fig. 2B, lower graph)(aav-grf1shrna, TBS, ±1.03%; n=7, p>.05). Finally, infection of a similar virus expressing shrna against GRF2 had the expected opposite effect, suppression of theta-burst LTP, but not HFS-induced LTP (data not shown). The dentate gyrus and CA3 regions of the hippocampus have been implicated in distinct aspects of hippocampus-mediated learning and memory including pattern separation required for distinguishing closely related contexts (3). Thus, the role of Ras-GRF1 in LTP in these regions was also tested. In contrast to the CA1, HFS-induced LTP was normal in both of these regions of the hippocampus from 2-month old Ras-GRF1 knockout mice ([LTP, DG, WT, ±1.22% (n=9 slices from six mice) vs ±0.64% (n=10 slices from six grf1(-/-) mice; p>0.05; CA3, WT, ±1.51% (n=8 slices from five mice) vs ±1.63% (n=9 slices from five grf1(-/-) mice; p>0.05). Overall, these findings show that Ras-GRF1 plays an age-, stimulation-, and synapse-dependent role in promoting LTP induction in the hippocampus. Ras-GRF1 induces LTP in the CA1 of 2-month old mice through activation of p38 MAP kinase. One of the known properties of Ras-GRF proteins is their ability to activate the Ras/Erk Map kinase pathway, which can promote LTP. However, we showed previously that Ras-GRF1 is a weak inducer of NMDA-mediated LTP in the CA1 compared to Ras-GRF2 in 1-month old mice, at least in part, because it is less effective than Ras- GRF2 in activating Erk Map kinase (5). Nevertheless, it was possible that Ras-GRF1 acquires the ability to promote LTP through Erk Map kinase by 2-months of age. To test this possibility, hippocampal slices from 2-month old wild-type mice were pre-exposed to U0126, an inhibitor of Erk activation or buffer only, for 30 minutes before LTP induction. Fig. 3A shows that as demonstrated previously (13,22), HFS-induced LTP was not blocked by Erk inhibition [Control, ±4.73% (n=11); U0126, ±2.79% (n=9); P>0.05] even though HFS-induced LTP was associated with an increase in Erk activity in CA1 neurons that was blocked by pretreatment with U0126 (data not shown). In contrast, TBSinduced LTP in 2-month old mice, which is 5
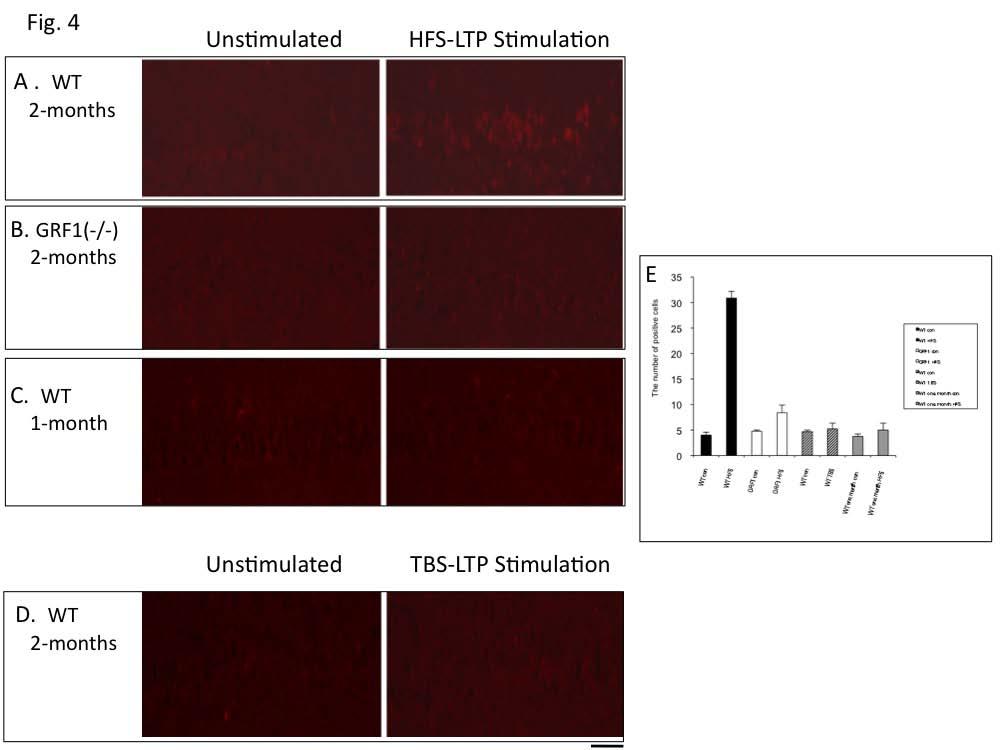
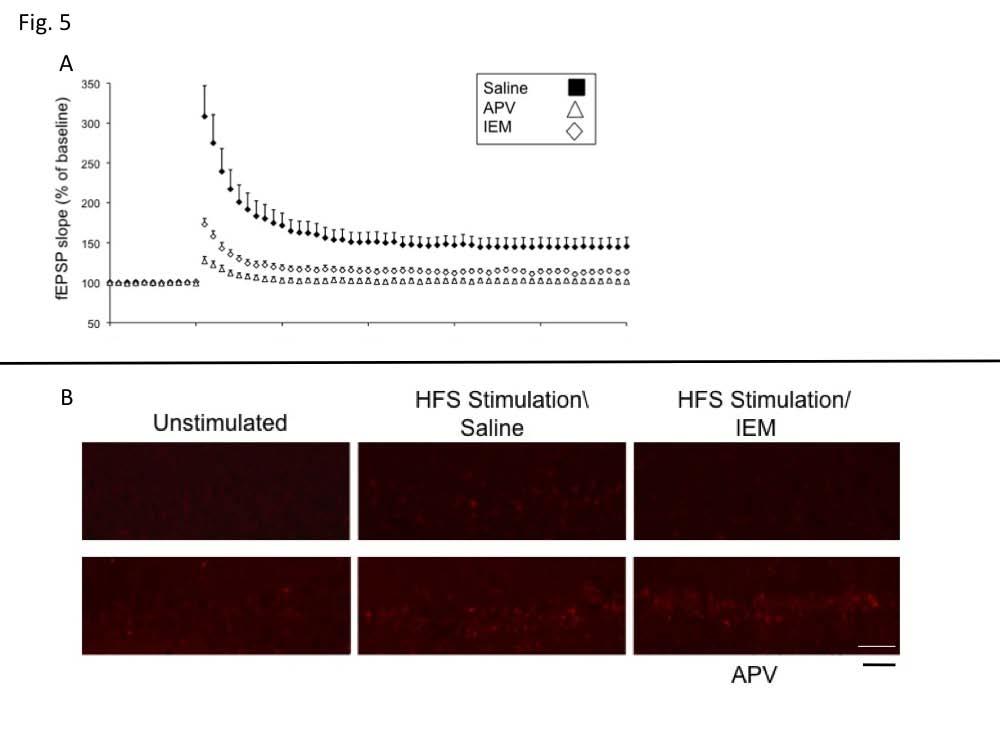
6 mediated by Ras-GRF2/Erk signaling (see (5)), was suppressed by treatment with U0126 (Fig. 3B) [Control, ±2.77% (n=8); U0126, ±1.50% ( n=7); P<0.01]. Ras-GRF1 can also mediate p38 activation by NMDA receptors potentially through its Racactivating DH domain or through interaction with JIP2, a p38 scaffold that binds to the N-terminal region of Ras-GRF1 (5,23). Thus, we pretreated hippocampus slices from 2 month-old mice with the p38 inhibitor, SB (10 um) and found that HFS-induced, LTP was suppressed (Fig. 3C) (Control, ±4.73% (n=11); SB203580, ±1.45%; n=8; p<0.01). Inhibition of p38 with SB20380 was quite specific, since as expected it did not block TBS LTP in 2-month old mice nor HFS LTP in 1-month old animals. Neither of these forms of LTP is affected in Ras- GRF1 knockout mice (data not shown). Nevertheless, since the SB inhibitor is not totally specific for p38, we tested another p38 inhibitor, Birb0796. It also suppressed HFS LTP (Fig. 3C.) (Birb0796, ±1.47%; n=8;p<0.01) but not TBS LTP (Fig. 3D) in 2-month old mice. Specific inhibition by both of these molecules is considered strong proof that p38 MAP kinase is involved (24). To gain additional support for the idea that this form of LTP is mediated by p38 Map kinase, HFS- LTP was induced in hippocampal slices from 2- month old mice and p38 activation was measured by immunofluorescence using antibodies specific for its active form. HFS activated p38 in principal cells of the CA1 hippocampus of control mice (Fig. 4A) but not in hippocampal slices from Ras-GRF1 knockout mice (Fig. 4B). Moreover, LTP induced by HFS treatment did not activate p38 in 1-month old mice when GRF1 does not yet have the capacity to promote LTP (Fig. 4C). Finally, theta-burst LTP, which is not blocked by p38 inhibitors in 2 month-old mice did not activate p38 (Fig. 4D) (for quantification of data see Fig. 4E)). Together these data strongly support the idea that HFS-induce LTP that begins at 2-month of age depends upon Ras-GRF1 mediated activation of p38 Map kinase. CP-AMPARs mediate HFS-induced LTP in the CA1 but not CA3 or dentate gyrus. Both NMDA and CP-AMPAR types of glutamate receptors are known to bind to Ras-GRF1(12,25), and both permeate calcium, an activator of Ras-GRF proteins. Thus, to determine which receptor type mediates HFS induction of LTP by Ras-GRF1 in the CA1 hippocampus of 2-month mice, hippocampal slices were pretreated with inhibitors of either NMDARs (APV) or CP-AMPARs (IEM). Either inhibitor blocked HFS-LTP (Fig. 5A) (Control, ±4.73% (n=11); IEM, ±1.61% (n=8); p<0.01; (Control, ±4.73% (n=11); APV, ±0.71% (n=7); p<0.01]), but only the CP-AMPAR inhibitor blocked HFS-induced p38 activation (Fig. 5B) These findings imply that CP- AMPAR/GRF1/p38 signaling adds an additional component to LTP induction that complements that involving NMDARs. Moreover, the CP-AMPAR inhibitor did not block LTP induction in the dentate gyrus or CA3 regions of the hippocampus from 2-month old mice, or the CA1 region from 1-month old mice, situations where GRF1 and p38 do not contribute to LTP [DG, Control, ±1.22% (n=9) vs IEM, ±1.09% (n=8); p>0.05; CA3, Control, ±1.51% (n=8) vs IEM, ±1.64% (n=8); p>0.05 and CA1 (one month old), Control, ±3.32% (n=8) vs ±2.49% (n=9); p>0.05.] Also, the p38 inhibitor has already been shown not to block LTP in the dentate gyrus (26). Together, the results described above define a novel, CP-AMPAR/Ras-GRF1/p38 Map kinase signaling cascade that promotes LTP specifically in the CA1, only after mice reach ~2-months of age. Contextual Discrimination is mediated by Ras- GRF1 in the CA1 hippocampus, p38 MAP kinase and CP-AMPARs. We previously showed that 2- month old Ras-GRF1 knockout mice fail to display contextual fear discrimination but have little if any defect in contextual fear conditioning (7). Thus, when 2-month old Ras-GRF1 knockout mice were exposed to foot shock in a specific environment and then placed back in that environment 24 hours later, they recognized the context and became immobile (froze) expecting a shock with a frequency that was similar to control mice treated similarly. However, Ras-GRF1 knockout mice failed to detect similar contexts. In particular, mice were trained to discriminate two closely related contexts, only one in which they received foot shocks. Freezing was then assessed in the non-shock environment two days later. Fig. 6A shows that wild-type mice recognized that the 6
7 related environment was different from the one in which they received a foot shock and froze less than when placed in the shock context. Consistent with our previous results, Ras-GRF1 knockout mice froze equally in the non-shock related context as they did in the shock-context (Fig. 6A) [WT (n=7), froze 31.35±5.90% of the testing time in the "paired" context versus 18.25±6.06% in the "non-paired" context, F(1,12)=5.39, p<0.05; GRF1KO (n=8), froze 27.09±5.93% of the testing time in the "paired" context versus 25±6.03% in the "non-paired" context, F(1,14)=0.06, p=0.81]. Because these mice lacked Ras-GRF1 in all tissues, we tested its role in the CA1 hippocampus by assaying the consequence of knocking down Ras-GRF1 expression specifically in the CA1 as described in Fig. 2. In particular, adenoassociated viruses expressing shrna against Ras- GRF1 or control viruses were injected bilaterally into the CA1 hippocampus of 2 month-old mice. Seven to nine days later, mice were compared in contextual discrimination assays. Fig. 6B shows that control, but not GRF1 shrna, virus-treated mice displayed contextual discrimination [AAVcontrol (n=8), froze 55.13±5.56% of the testing time in the "paired" context versus 40.08±4.37% in the "non-paired" context, F(1,14)=4.53, p<0.05; AAV-GRF1 shrna (n=8), froze 57.54±6.25% of the testing time in the "paired" context versus 56.56±3.49% in the "non-paired" context, F(1,14)=0.02, p=0.89]. To test the role of p38 Map kinase in contextual discrimination, the p38 MAP kinase inhibitors, SB or Birb0796, or vehicle was infused through a cannula placed above the CA1 region 30 minutes before mice were habituated to the two related contexts. Mice were then used in contextual discrimination assays. While vehicleinfused mice retained the ability to distinguish related contexts, BIRB0796 or SB infused mice did not (Fig. 6C and 6D) Vehicle (n=11), froze 53.01±2.86% of the testing time in the "paired" context versus ±4.81% in the "nonpaired" context, F(1,20)=8.24, p<0.01; Birb (n=9), froze 59.03±5.79% of the testing time in the "paired" context versus 62.37±5.77% in the "nonpaired" context, F(1,16)=0.17, p=0.69]; SB (n=9), froze /- 9.89% in paired versus /- 6.45% in unpaired contex, F(1,16)=0.52, p=0.48. Importantly, like Ras-GRF1 shrna knockdown mice, both sets of p38 inhibitor infused mice displayed normal contextual fear conditioning (data not shown). Finally, to test the role of CP-AMPARs in contextual discrimination, the CP-AMAPR inhibitor, IEM, or saline was introduced via intraperitaneal injection 30 minutes before mice were habituated to the related contexts. Like mice with suppressed GRF1 expression and inhibited p38map kinase in the CA1, these mice also did not recognize a difference in the non-shock context and froze just as often when placed in the non-shock context as when they were placed in the shock environment (Fig. 6E) [saline(n=10), froze 47.48±4.10% of the testing time in the "paired" context versus 31.19±5.36% in the "non-paired" context, F(1,20)=5.83, p<0.05; IEM (n=6), froze 42.52±12.98% of the testing time in the "paired" context versus 44.48±10.05% in the "non-paired" context, F(1,10)=0.01, p=0.91] but did display normal contextual fear conditioning (data not sh0wn). Because we did not detect a contribution of CP- AMPARs, GRF1, or p38 MAP kinase in HFSinduced LTP in the CA1 hippocampus of 1-month old mice, we tested them for the ability to distinguish closely related contexts. Fig. 6F shows that, in contrast to 2-months old mice, 1-month old mice do not yet have the capacity to distinguish related contexts. Overall, these findings implicate the appearance of a CP-AMPAR/Ras-GRF1/p38 MAP kinase LTP pathway in the acquisition of the ability to distinguish closely related contexts during early adolescent development in mice. DISCUSSION This study uses electrophysiology, genetics, immunohistochemistry and behavioral analysis to reveal that the postnatal emergence of the ability of mice to distinguish closely related contexts at ~ 2 months of age involves the appearance of a newly discovered signaling pathway involving CP-AMPARs, Ras-GRF1 and p38 Map kinase that contributes to LTP in the CA3/CA1 synapse of the hippocampus. One of the most striking findings of this work is the detection of a role for p38 Map kinase, rather than Erk Map kinase in promoting LTP, since p38 has mainly been implicated in promoting LTD (5,15-17) or inhibiting LTP (27,28). For example, LTD induced through metabotropic glutamate receptors in the CA1 (15) 7
8 (29) and dentate gyrus of the hippocampus functions through p38. Evidence suggests p38 also mediates NMDA receptor-induced LTD, although the degree of inhibition of this form of LTD by p38 inhibitors has ranged from complete (16), to partial (5) to none (17). Some of this discrepancy may arise the fact that these studies used varied methods to detect LTD, different rodents and/or animals of different ages animals. One example where p38 has been implicated in LTP induction mediated by NMDA receptors is via theta-burst stimulation of the CA1 after exposure of adolescent mice to an enriched environment (18). In that case, a camp/p38 Map kinase signaling pathway emerges after enrichment that compliments the existing Erk Map kinase pathway regulated by Ras-GRF2. Apparently, p38 Map kinase can have multiple functions in cells, a concept consistent with the fact that kinases exist in distinct signaling complexes containing different scaffold proteins that can target them to specific subsets of their many substrates in cells (30). Of the ~100 p38 putative substrates identified (31) most are involved in gene transcription or translation regulation, which occurs at a time scale too slow to contribute to HFS-LTP. However some tyrosine kinase receptors, cytoplasmic kinases and proteins involved in endocytosis also have been proposed to be substrates, and thus could potentially influence synaptic plasticity. Which p38 substrates are involved here remains to be determined. This form of LTP appears to be a postsynaptic process, since no change in paired pulse facilitation was observed, indicating a lack of presynaptic involvement (data not shown). Another interesting aspect of this CP- AMPAR/Ras-GRF1/p38 Map kinase pathway is that it contributes to HFS, but not TBS, -induced early LTP. HFS usually involves one train of 100 stimuli at 100 Hz, while theta burst stimulation is typically four 100 Hz pulses with 200 ms interburst intervals, a protocol designed to match endogenous theta waves (32). Although both of these types of tetanization produce similar magnitudes of early LTP when applied to the Schaffer collateral innervating synapses in the CA1 hippocampus, differences in their mechanisms have been noted. For example, although HFS is often described as a more intense form of stimulation, theta-burst actually leads to more calcium influx (33). In addition, theta burst stimulation leads to significant inhibitory postsynaptic potentials soon after tetanus, whereas HFS stimulation produces a distinct NMDAmediated suppression of these inhibitory responses (33). Which, if any of these differences, contribute to the specificity in evoking the CP-AMPAR/Ras- GRF1/p38 pathway at 2 months of age remains to be determined. Finally, while theta-burst LTP is considered a more physiological stimulus, our finding that the appearance of contextual discrimination at ~2-months of age correlates with the appearance of HFS-LTP mediated by GRF1 and p38, rather than theta-burst LTP mediated by Erk Map kinase that has been present since birth, suggests that HFS-LTP contributes to physiological events in the hippocampus. Furthermore, our finding that a new signaling cascade contributes to HFS-induced LTP only after 2 months of age during late adolescence highlights how the biochemical underpinnings of LTP change with age along with the changing role of synaptic plasticity. For example, until ~1 month of age the hippocampus does not contribute to learning and memory, so that synaptic plasticity at this time is likely involved in promoting the final maturation of this brain region (1,34). Soon after birth, camp is a predominant inducer of LTP in the CA1 hippocampus, and then ~ 2 weeks of age CaMKII activity takes over (35). At 1 month of age, when contextual fear learning is first enabled, Ras-GRF2 begins to contribute to LTP in the CA1 along with CamKII (5). The present study shows that at 2-months of age more refined learning required for discriminating closely related contexts becomes possible due, at least in part, to the appearance of an additional signaling pathway that contributes to LTP, involving CP-AMPAR, GRF1 and p38 Map kinase. These findings also highlight how GRF1 plays distinct roles in different regions of the brain. Previous studies showed that GRF1 contributes to TBS-induced LTP in the basal amygdala (36) but not the CA1 hippocampus (5). Here we show that GRF1 contributes to HFS-induced LTP in the CA1, but not the CA3 or dentate gyrus of the hippocampus. Moreover, GRF1 mediates dopamine signaling through Erk activation in the striatum (37), whereas it is a poor mediator of NMDA receptor activation of Erk in the CA1 (5). In addition, GRF1 affects visual recognition 8
9 memory and both LTP and LTD in the perirhinal cortex via regulation of Erk (38). Finally, GRF1 mediates CP-AMPAR activation of Erk in the cortex at 1 month of age (12), but p38 activation starting at 2 months of age in the CA1, but not CA3 or dentate gyrus, regions of the hippocampus. The biochemical underpinnings of these differences have not yet been revealed. However, it most likely involves discretionary use of its known interactions with a plethora of scaffold proteins that target the protein to distinct effector proteins in cells (23,39,40). Calcium-permeable AMPA type glutamate receptors, CP-AMPARs, consist of homodimers of GluA1 subunits that are permeable to calcium, making them likely mediators of synaptic plasticity (41). Their role in synaptic plasticity in inhibitory interneurons has been well established, however their role in CA1 pyramidal neurons has been controversial. Plant et al first demonstrated that the early phase of LTP in these neurons involves signaling through CP-AMPARs at synapses (42), but this was quickly refuted (43). However, additional studies have reaffirmed that this class of receptors represent a significant fraction (~8-10%) of the AMPA receptor population in these cells (44). They appear to contribute specifically to the early phase of LTP in an age-dependent manner (45). Increased CP- AMPAR content in mice increases the magnitude of HFS-LTP in the CA1, and their inhibition blocks NMDA-independent learning, where animals have a prior experience on a similar behavioral task (46). Here, we expand these finding by showing that CP-AMPARs begin to function through Ras-GRF1 to regulate p38 at about ~2 months of age in mice to contribute to contextual discrimination. The present study also reveals that the CP- AMPARs, Ras-GRF1, and p38 contribution to HFS-LTP is limited to the CA1 region, as inhibition of any of these proteins in the CA3 or dentate gyrus had no effect on this form of synaptic plasticity. Most studies have pinpointed specific roles for the dentate gyrus and CA3 regions in pattern separation, an important component of contextual discrimination (47). In particular, the DG and CA3 have been proposed to play contrasting roles in memory processing, with the dentate gyrus functioning as a pattern separator splitting the signal input from the entorhinael cortex into distinct parts and encoding stimuli as novel. The CA3 contributes to pattern completion that matches incoming stimuli from EC with previous familiar experiences. The CA1 generates the output of the hippocampus and therefore the end-point of its processing properties. The CA1 has been proposed to be a mismatch detector (48), since it receives both direct input about current events from EC and stored information from CA3. However, evidence suggests that this region also adds a temporal context to events (49), another key aspect of pattern separation involved in contextual discrimination studied here, where contexts are experienced temporally and retrieval of stored information during testing is required. Which of these aspects of contextual discrimination involve synaptic plasticity provided by the CP-AMPAR/Ras-GRF1/p38/LTP pathway is presently under investigation. 9
10 REFERENCES 1. Dumas, T. C. (2005) Hippocampus 15, Yassa, M. A., Lacy, J. W., Stark, S. M., Albert, M. S., Gallagher, M., and Stark, C. E. (2011) Hippocampus 21, Langston, R. F., Stevenson, C. H., Wilson, C. L., Saunders, I., and Wood, E. R. (2010) Behav Brain Res 215, Feig, L. A. (2011) Genes Cancer 2, Li, S., Tian, X., Hartley, D. M., and Feig, L. A. (2006) J Neurosci 26, Jin, S. X., and Feig, L. A. (2010) PLoS One 5, e Giese, K. P., Friedman, E., Telliez, J. B., Fedorov, N. B., Wines, M., Feig, L. A., and Silva, A. J. (2001) Neuropharmacology 41, d'isa, R., Clapcote, S. J., Voikar, V., Wolfer, D. P., Giese, K. P., Brambilla, R., and Fasano, S. (2011) Front Behav Neurosci 5, Sweatt, J. D. (2001) J Neurochem 76, Klesse, L. J., and Parada, L. F. (1999) Microsc Res Tech 45, Xia, Z., Dudek, H., Miranti, C. K., and Greenberg, M. E. (1996) J Neurosci 16, Tian, X., and Feig, L. A. (2006) J Biol Chem 13. Selcher, J. C., Weeber, E. J., Christian, J., Nekrasova, T., Landreth, G. E., and Sweatt, J. D. (2003) Learn Mem 10, Adams, J. P., Roberson, E. D., English, J. D., Selcher, J. C., and Sweatt, J. D. (2000) Acta Neurobiol Exp (Wars) 60, Bolshakov, V. Y., Carboni, L., Cobb, M. H., Siegelbaum, S. A., and Belardetti, F. (2000) Nat Neurosci 3, Zhu, J. J., Qin, Y., Zhao, M., Van Aelst, L., and Malinow, R. (2002) Cell 110, Peineau, S., Nicolas, C. S., Bortolotto, Z. A., Bhat, R. V., Ryves, W. J., Harwood, A. J., Dournaud, P., Fitzjohn, S. M., and Collingridge, G. L. (2009) Mol Brain 2, Li, S., Tian, X., Hartley, D. M., and Feig, L. A. (2006) Curr Biol 16, Tian, X., Gotoh, T., Tsuji, K., Lo, E. H., Huang, S., and Feig, L. A. (2004) Embo J 23, He, T. C., Zhou, S., da Costa, L. T., Yu, J., Kinzler, K. W., and Vogelstein, B. (1998) Proc Natl Acad Sci U S A 95, Cashman, S. M., Sadowski, S. L., Morris, D. J., Frederick, J., and Kumar-Singh, R. (2002) Mol Ther 6, Kelleher, R. J., 3rd, Govindarajan, A., Jung, H. Y., Kang, H., and Tonegawa, S. (2004) Cell 116, Buchsbaum, R. J., Connolly, B. A., and Feig, L. A. (2002) Mol Cell Biol 22, Bain, J., Plater, L., Elliott, M., Shpiro, N., Hastie, C. J., McLauchlan, H., Klevernic, I., Arthur, J. S., Alessi, D. R., and Cohen, P. (2007) Biochem J 408, Krapivinsky, G., Krapivinsky, L., Manasian, Y., Ivanov, A., Tyzio, R., Pellegrino, C., Ben-Ari, Y., Clapham, D. E., and Medina, I. (2003) Neuron 40, Saleshando, G., and O'Connor, J. J. (2000) Neurosci Lett 288, Izumi, Y., Tokuda, K., and Zorumski, C. F. (2008) Hippocampus 18, Butler, M. P., O'Connor, J. J., and Moynagh, P. N. (2004) Neuroscience 124, Rush, A. M., Wu, J., Rowan, M. J., and Anwyl, R. (2002) J Neurosci 22, Brown, M. D., and Sacks, D. B. (2009) Cell Signal 21, Trempolec, N., Dave-Coll, N., and Nebreda, A. R. (2013) Cell 152, e Albensi, B. C., Oliver, D. R., Toupin, J., and Odero, G. (2007) Exp Neurol 204,
11 33. Perez, Y., Chapman, C. A., Woodhall, G., Robitaille, R., and Lacaille, J. C. (1999) Neuroscience 90, Rudy, J. W., and Morledge, P. (1994) Behav Neurosci 108, Malenka, R. C., and Bear, M. F. (2004) Neuron 44, Brambilla, R., Gnesutta, N., Minichiello, L., White, G., Roylance, A. J., Herron, C. E., Ramsey, M., Wolfer, D. P., Cestari, V., Rossi-Arnaud, C., Grant, S. G., Chapman, P. F., Lipp, H. P., Sturani, E., and Klein, R. (1997) Nature 390, Fasano, S., D'Antoni, A., Orban, P. C., Valjent, E., Putignano, E., Vara, H., Pizzorusso, T., Giustetto, M., Yoon, B., Soloway, P., Maldonado, R., Caboche, J., and Brambilla, R. (2009) Biol Psychiatry 66, Silingardi, D., Angelucci, A., De Pasquale, R., Borsotti, M., Squitieri, G., Brambilla, R., Putignano, E., Pizzorusso, T., and Berardi, N. (2011) Front Behav Neurosci 5, Buchsbaum, R. J., Connolly, B. A., and Feig, L. A. (2003) J Biol Chem 278, Connolly, B. A., Rice, J., Feig, L. A., and Buchsbaum, R. J. (2005) Mol Cell Biol 25, Man, H. Y. (2011) Curr Opin Neurobiol 21, Plant, K., Pelkey, K. A., Bortolotto, Z. A., Morita, D., Terashima, A., McBain, C. J., Collingridge, G. L., and Isaac, J. T. (2006) Nat Neurosci 9, Adesnik, H., and Nicoll, R. A. (2007) J Neurosci 27, Rozov, A., Sprengel, R., and Seeburg, P. H. (2012) Front Mol Neurosci 5, Lu, Y., Allen, M., Halt, A. R., Weisenhaus, M., Dallapiazza, R. F., Hall, D. D., Usachev, Y. M., McKnight, G. S., and Hell, J. W. (2007) EMBO J 26, Wiltgen, B. J., Royle, G. A., Gray, E. E., Abdipranoto, A., Thangthaeng, N., Jacobs, N., Saab, F., Tonegawa, S., Heinemann, S. F., O'Dell, T. J., Fanselow, M. S., and Vissel, B. (2010) PLoS One Yassa, M. A., and Stark, C. E. (2011) Trends Neurosci 34, Duncan, K., Ketz, N., Inati, S. J., and Davachi, L. (2012) Hippocampus 22, Kesner, R. P., and Hunsaker, M. R. (2010) Behav Brain Res 215,
12 FOOTNOTES * This work was supported by NIH grants RO1 MH to LAF and by the Tufts Center for Neuroscience Research grant P30 NS To whom crrespondence may be addressed: Department of Biochemistry, Tufts University School of Medicine, 136 Harrison Ave, Boston, MA, 02111, USA, Tel.:(617) ; larry.feig@tufts.edu 4 The abbreviations used are: CP-AMPARs, calcium permeable 2-amino-3-(3-hydroxy-5-methyl-isoxazol- 4-yl)propanoic acid receptors; Ras-GRF1, Ras guanine nucleotide releasing factor 1; Ras-GRF2, Ras guanine nucleotide releasing factor 2; NMDA, N-methyl-D-aspartate; LTP, long-term potentiation; LTD, long-term depression; ACF, artificial cerebrospinal fluid ; fepsp, field excitatory postsynaptic potential; TBS, theta-burst; HFS, high frequency stimuliation; BIRB, BIRB0796. FIGURE LEGENDS Figure 1. GRF1 knockout mice display defective HFS- but not TBS-induced LTP in CA1 hippocampus of 2 month-old mice. Hippocampus slices from two month-old wild type (filled square) or GRF1 knockout mice (open square) were stimulated with high frequency (HFS) (A) or theta-burst (TBS) stimulation (B). The recorded fepsp slope using field recordings is expressed as a percentage of baseline +/- SEM (HFS) (WT; n=11 slices from six mice; grf1(-/-); n=12 slices from eight grf1 (-/-) mice) (TBS) WT; n=8 slices from five mice; grf1(-/-) n=9 slices from five mice. Insets, Representative fepsps recorded. (C) Control GFP expressing adenovirus (filled diamonds) or adenovirus expressing GRF1 (open diamonds) were injected into the CA1 hippocampus days later HFS LTP was measured in isolated hippocampal slices. Data represent the mean +/- SEM (Ras-GRF1; n=10 slices from 5 mice); (GFP; n=13 slices from nine mice). Inset, CA1 hippocampus was stained with anti-grf1 antibodies. (calibration: horizontal, 10ms, vertical, 1 mv, scale bar, 50um. Figure 2. shrna knockdown of GRF1 in the CA1 hippocampus of 2-month old mice suppresses HFS, but not TBS induced LTP. (A) A mammalian expression vector expressing GRF1 or GRF2 was cotransfected with an empty control vector or a vector expressing shrna against GRF2 or GRF1 into HEK 293 cells. 3 days later, lysates of cells were probed with antibodies that recognize GRF1 and GRF2. (B) Either control scrambled shrna- or GRF1 shrna-expressing adeno-associated viruses co-expressing red cherry were stereotaxically injected into the CA1 hippocampus days later, brain slices were isolated. HFS (top graph) or TBS (lower graph)-induced LTP in the CA1 were then recorded (control AAV-random shrna (closed figures) n=7; shrna-grf1 AAV (open figures), n=9. Data represent the mean +/- SEM. Inset, a hippocampal slice from an infected mouse detecting red-cherry fluorescence. (scale bar, 250 µm) Figure 3. HFS induced LTP, but not TBS induced LTP, in the CA1 hippocampus of 2-month old mice is dependent upon p38. (A and B) Hippocampal slices were preincubated with either vehicle (filled squares), or MAP kinase inhibitor U0126 (20 µm) (open squares). (C) Hippocampal were preincubated with either vehicle (closed square), p38 MAP kinase inhibitors SB (10 µm) (open square) or Birb0796 (10 µm) (open diamond) and tested for HFS-LTP (A and C) or TBS- LTP (B and D) n>6. Data represent the mean +/- SEM. Figure 4. HFS, but not TBS-induced LTP, in 2-month old mice activates p38 MAP kinase activity in principal neurons of the CA1 hippocampus. Hippocampal slices from 2 month-old (A, B and D) or 1 month-old (C) WT or GRF1 knockout mice were stimulated with HFS-LTP induction (A-C) or TBS-LTP induction. The tissues were fixed 15 minutes later and stained with anti-p-p38 MAP kinase antibody. (E) 12
13 Stained cells/visual field were quantified in each experiment date represent the average of 3 fields +/- SEM. (scale bar, 50 µm) Figure 5. HFS-induced LTP is blocked by inhibitors of either NMDARs or CP-AMPARs receptors, but only inhibitors of CP-AMPARs block HFS-induced p38 activation. HFS LTP was induced in brain slices from WT mice in the presence of saline, NMDA receptor inhibitor, APV, (100 µm) or the CP-AMPA receptor inhibitor IEM-1460 (30 µm). fepsp slopes were then measured in some slices (A) (data represent the mean +/- SEM of > 6 slices) and others were stained with p-p38 MAP kinase antibodies (B) Data is representative of 3 fields. (scale bar, 50 µm) Figure 6. Contextual discrimination is blocked by expression of GRF1 shrna in the CA1 hippocampus, p38 inhibitor infusion into the CA1 hippocampus or injection of a CP-AMPAR inhibitor. Mice were exposed to a contextual discrimination paradigm as described (7). The amount of freezing was measured when mice were placed in the cage where they were shocked (paired- filled bar) and in the related cage in which they were not (non-paired- open bar). A) WT mice were compared with grf1(-/-) mice B) Adenoassociated virus expressing GFP (or random shrna-similar results) or an shrna against GRF1 were stereotaxically injected into the dorsal CA1 hippocampus of mice 8-10 days before beginning the experiment. C) Saline or the p38 MAP kinase inhibitor Birb0796 (1 µl of mg/ml) were infused through a cannula into the hippocampus of mice 30 minutes before each exposure to fear conditioning equipment. D) Saline or the p38 MAP kinase inhibitor SB (1 µl of 0.75 mg/ml) were infused through a cannula into the hippocampus of mice 30 minutes before each exposure to fear conditioning equipment. E) Saline or the CP-AMPAR inhibitor, IEM-1460 (8mg/kg), was injected ip into mice 30 minutes before each exposure to the fear conditioning equipment. F) WT mice at 1-month of age were used. Data represent the mean +/- SEM of at least 9 animals each. 13
14
15
16
17
18
19
2 The abbreviations used are: NMDAR, NMDA receptor; CP-AMPAR, calciumpermeable
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 288, NO. 30, pp. 21703 21713, July 26, 2013 2013 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. Acquisition of Contextual
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 288, NO. 30, pp. 21703 21713, July 26, 2013 2013 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. Acquisition of Contextual
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Synaptic plasticityhippocampus. Neur 8790 Topics in Neuroscience: Neuroplasticity. Outline. Synaptic plasticity hypothesis
 Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/317/5841/183/dc1 Supporting Online Material for Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses Gertrudis Perea and Alfonso Araque* *To whom
www.sciencemag.org/cgi/content/full/317/5841/183/dc1 Supporting Online Material for Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses Gertrudis Perea and Alfonso Araque* *To whom
BIPN140 Lecture 12: Synaptic Plasticity (II)
") BIPN140 Lecture 12: Synaptic Plasticity (II) 1. Early v.s. Late LTP 2. Long-Term Depression 3. Molecular Mechanisms of Long-Term Depression: NMDA-R dependent 4. Molecular Mechanisms of Long-Term Depression:
BIPN140 Lecture 12: Synaptic Plasticity (II) 1. Early v.s. Late LTP 2. Long-Term Depression 3. Molecular Mechanisms of Long-Term Depression: NMDA-R dependent 4. Molecular Mechanisms of Long-Term Depression:
Bidirectional modifications in synaptic efficacy, exemplified
 Capture of a protein synthesis-dependent component of long-term depression Beth S. Kauderer* and Eric R. Kandel* Howard Hughes Medical Institute and *Center for Neurobiology and Behavior, College of Physicians
Capture of a protein synthesis-dependent component of long-term depression Beth S. Kauderer* and Eric R. Kandel* Howard Hughes Medical Institute and *Center for Neurobiology and Behavior, College of Physicians
Cellular Neurobiology BIPN140
 Cellular Neurobiology BIPN140 1st Midterm Exam Ready for Pickup By the elevator on the 3 rd Floor of Pacific Hall (waiver) Exam Depot Window at the north entrance to Pacific Hall (no waiver) Mon-Fri, 10:00
Cellular Neurobiology BIPN140 1st Midterm Exam Ready for Pickup By the elevator on the 3 rd Floor of Pacific Hall (waiver) Exam Depot Window at the north entrance to Pacific Hall (no waiver) Mon-Fri, 10:00
Synaptic Plasticity and the NMDA Receptor
 Synaptic Plasticity and the NMDA Receptor Lecture 4.2 David S. Touretzky November, 2015 Long Term Synaptic Plasticity Long Term Potentiation (LTP) Reversal of LTP Long Term Depression (LTD) Reversal of
Synaptic Plasticity and the NMDA Receptor Lecture 4.2 David S. Touretzky November, 2015 Long Term Synaptic Plasticity Long Term Potentiation (LTP) Reversal of LTP Long Term Depression (LTD) Reversal of
Supplemental information Acid-sensing ion channel 1a contributes to hippocampal LTP inducibility through multiple mechanisms
 Supplemental information Acid-sensing ion channel 1a contributes to hippocampal LTP inducibility through multiple mechanisms Ming-Gang Liu, Hu-Song Li, Wei-Guang Li, Yan-Jiao Wu, Shi-Ning Deng, Chen Huang,
Supplemental information Acid-sensing ion channel 1a contributes to hippocampal LTP inducibility through multiple mechanisms Ming-Gang Liu, Hu-Song Li, Wei-Guang Li, Yan-Jiao Wu, Shi-Ning Deng, Chen Huang,
mtorc2 controls actin polymerization required for consolidation of long-term memory
 CORRECTION NOTICE Nat. Neurosci.; doi:1.138/nn.3351 mtorc2 controls actin polymerization required for consolidation of long-term memory Wei Huang, Ping Jun Zhu, Shixing Zhang, Hongyi Zhou, Loredana Stoica,
CORRECTION NOTICE Nat. Neurosci.; doi:1.138/nn.3351 mtorc2 controls actin polymerization required for consolidation of long-term memory Wei Huang, Ping Jun Zhu, Shixing Zhang, Hongyi Zhou, Loredana Stoica,
Ube3a is required for experience-dependent maturation of the neocortex
 Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/312/5779/1533/dc1 Supporting Online Material for Long-Term Potentiation of Neuron-Glia Synapses Mediated by Ca 2+ - Permeable AMPA Receptors Woo-Ping Ge, Xiu-Juan Yang,
www.sciencemag.org/cgi/content/full/312/5779/1533/dc1 Supporting Online Material for Long-Term Potentiation of Neuron-Glia Synapses Mediated by Ca 2+ - Permeable AMPA Receptors Woo-Ping Ge, Xiu-Juan Yang,
Synaptic plasticity. Activity-dependent changes in synaptic strength. Changes in innervation patterns. New synapses or deterioration of synapses.
 Synaptic plasticity Activity-dependent changes in synaptic strength. Changes in innervation patterns. New synapses or deterioration of synapses. Repair/changes in the nervous system after damage. MRC Centre
Synaptic plasticity Activity-dependent changes in synaptic strength. Changes in innervation patterns. New synapses or deterioration of synapses. Repair/changes in the nervous system after damage. MRC Centre
Supplementary Methods
 1 Supplementary Methods Social Preference Test Subjects Seventy-four Long-Evans, male rats served as subjects (S-rats) in the foodpreference test, with 40 assigned to the CXT-Same (CXT-S) Condition and
1 Supplementary Methods Social Preference Test Subjects Seventy-four Long-Evans, male rats served as subjects (S-rats) in the foodpreference test, with 40 assigned to the CXT-Same (CXT-S) Condition and
SUPPLEMENTARY INFORMATION. Supplementary Figure 1
 SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
Cellular mechanisms of information transfer: neuronal and synaptic plasticity
 Cellular mechanisms of information transfer: neuronal and synaptic plasticity Ivan Pavlov (UCL Institute of Neurology, UK) Anton Chizhov (Ioffe Physical Technical Institute) Pavel Zykin (St.-Petersburg
Cellular mechanisms of information transfer: neuronal and synaptic plasticity Ivan Pavlov (UCL Institute of Neurology, UK) Anton Chizhov (Ioffe Physical Technical Institute) Pavel Zykin (St.-Petersburg
Cellular Neurobiology / BIPN 140
 SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
The molecular analysis of long-term plasticity in the mammalian
 Genetic evidence for a protein-kinase-a-mediated presynaptic component in NMDA-receptor-dependent forms of long-term synaptic potentiation Yan-You Huang*, Stanislav S. Zakharenko*, Susanne Schoch, Pascal
Genetic evidence for a protein-kinase-a-mediated presynaptic component in NMDA-receptor-dependent forms of long-term synaptic potentiation Yan-You Huang*, Stanislav S. Zakharenko*, Susanne Schoch, Pascal
Zhu et al, page 1. Supplementary Figures
 Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05772 SUPPLEMENTARY INFORMATION Supplemental figure 1. Enrichment facilitates learning. a. Images showing a home cage and a cage used for environmental enrichment (EE). For EE up to
doi: 10.1038/nature05772 SUPPLEMENTARY INFORMATION Supplemental figure 1. Enrichment facilitates learning. a. Images showing a home cage and a cage used for environmental enrichment (EE). For EE up to
Part 11: Mechanisms of Learning
 Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Supporting Information
 ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
A form of long-lasting, learning-related synaptic plasticity in the hippocampus induced by heterosynaptic low-frequency pairing
 A form of long-lasting, learning-related synaptic plasticity in the hippocampus induced by heterosynaptic low-frequency pairing Yan-You Huang, Christopher Pittenger*, and Eric R. Kandel Center for Neurobiology
A form of long-lasting, learning-related synaptic plasticity in the hippocampus induced by heterosynaptic low-frequency pairing Yan-You Huang, Christopher Pittenger*, and Eric R. Kandel Center for Neurobiology
Memory Systems II How Stored: Engram and LTP. Reading: BCP Chapter 25
 Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Supplementary Figure 1. Basic properties of compound EPSPs at
 Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
1.0. FSL NMDAR-fEPSP 0.8. amplitude (mv) Intensity (µa) 2.0 SD FSL Time (ms)
 Intensity (µa) 2.0 SD FSL Time (ms)") a 2.5 1. AMPAR-fEPSP slope (mv/ms) 2. 1. NMDAR-fEPSP amplitude (mv).8.6.4.5.2. 2 4 6 8. 1 2 3 4 5 Intensity (µa) Intensity (µa) b 2. PPF Ratio (fepsp2/fepsp1) 1..5. 5 1 2 5 Time (ms) Supplementary Figure
a 2.5 1. AMPAR-fEPSP slope (mv/ms) 2. 1. NMDAR-fEPSP amplitude (mv).8.6.4.5.2. 2 4 6 8. 1 2 3 4 5 Intensity (µa) Intensity (µa) b 2. PPF Ratio (fepsp2/fepsp1) 1..5. 5 1 2 5 Time (ms) Supplementary Figure
Supplementary Figure S1. Effect of stress during withdrawal on expression of sensitization to repeated cocaine exposure in WT and D2R / mice.
 Supplementary Figure S1. Effect of stress during withdrawal on expression of sensitization to repeated cocaine exposure in WT and D2R / mice. The time course of locomotor activity for WT (a, b) or D2R
Supplementary Figure S1. Effect of stress during withdrawal on expression of sensitization to repeated cocaine exposure in WT and D2R / mice. The time course of locomotor activity for WT (a, b) or D2R
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Behavioural effects of ketamine in non-stressed and stressed mice. Naive C57BL/6 adult male mice (n=10/group) were given a single dose of saline vehicle or ketamine (3.0 mg/kg,
Supplementary Figure 1. Behavioural effects of ketamine in non-stressed and stressed mice. Naive C57BL/6 adult male mice (n=10/group) were given a single dose of saline vehicle or ketamine (3.0 mg/kg,
Nature Neuroscience: doi: /nn Supplementary Figure 1. Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons
 Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
CASE 49. What type of memory is available for conscious retrieval? Which part of the brain stores semantic (factual) memories?
 memories?") CASE 49 A 43-year-old woman is brought to her primary care physician by her family because of concerns about her forgetfulness. The patient has a history of Down syndrome but no other medical problems.
CASE 49 A 43-year-old woman is brought to her primary care physician by her family because of concerns about her forgetfulness. The patient has a history of Down syndrome but no other medical problems.
Linking Neuronal Ensembles by Associative Synaptic Plasticity
 Linking Neuronal Ensembles by Associative Synaptic Plasticity Qi Yuan 1,2, Jeffry S. Isaacson 2, Massimo Scanziani 1,2,3 * 1 Department of Neurobiology, Center for Neural Circuits and Behavior, University
Linking Neuronal Ensembles by Associative Synaptic Plasticity Qi Yuan 1,2, Jeffry S. Isaacson 2, Massimo Scanziani 1,2,3 * 1 Department of Neurobiology, Center for Neural Circuits and Behavior, University
Supplementary Methods. the ventrolateral orbitofrontal cortex (VLO) and basolateral amygdala (BLA). AAV8-CaMKII-HAhM
 and basolateral amygdala (BLA). AAV8-CaMKII-HAhM") Supplementary Materials, Zimmermann et al. Supplementary Methods Surgery. AAV5-CaMKII-HA-hM 4 D(Gi)-IRES-mCitrine or AAV5-CaMKII-GFP was infused into the ventrolateral orbitofrontal cortex (VLO) and basolateral
Supplementary Materials, Zimmermann et al. Supplementary Methods Surgery. AAV5-CaMKII-HA-hM 4 D(Gi)-IRES-mCitrine or AAV5-CaMKII-GFP was infused into the ventrolateral orbitofrontal cortex (VLO) and basolateral
9.01 Introduction to Neuroscience Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Declarative memory conscious,
MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Declarative memory conscious,
Supplementary Information
 Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
Is action potential threshold lowest in the axon?
 Supplementary information to: Is action potential threshold lowest in the axon? Maarten H. P. Kole & Greg J. Stuart Supplementary Fig. 1 Analysis of action potential (AP) threshold criteria. (a) Example
Supplementary information to: Is action potential threshold lowest in the axon? Maarten H. P. Kole & Greg J. Stuart Supplementary Fig. 1 Analysis of action potential (AP) threshold criteria. (a) Example
Suppl. Information Supplementary Figure 1. Strategy/latency analysis of individual mice during maze learning. a,
 Goal-oriented searching mediated by ventral hippocampus early in trial-and-error learning Ruediger, S, Spirig, D., Donato, F., Caroni, P. Suppl. Information Supplementary Figure 1. Strategy/latency analysis
Goal-oriented searching mediated by ventral hippocampus early in trial-and-error learning Ruediger, S, Spirig, D., Donato, F., Caroni, P. Suppl. Information Supplementary Figure 1. Strategy/latency analysis
Supplemental Information. A Visual-Cue-Dependent Memory Circuit. for Place Navigation
 Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
The Neurobiology of Learning and Memory
 The Neurobiology of Learning and Memory JERRY W. RUDY University of Colorado, Boulder Sinauer Associates, Inc. Publishers Sunderland, Massachusetts 01375 Table of Contents CHAPTER 1 Introduction: Fundamental
The Neurobiology of Learning and Memory JERRY W. RUDY University of Colorado, Boulder Sinauer Associates, Inc. Publishers Sunderland, Massachusetts 01375 Table of Contents CHAPTER 1 Introduction: Fundamental
Synaptic Plasticity and Memory
 Synaptic Plasticity and Memory Properties and synaptic mechanisms underlying the induction of long-term potentiation (LTP) The role of calcium/calmodulin-dependent kinase II (CamKII) in the induction,
Synaptic Plasticity and Memory Properties and synaptic mechanisms underlying the induction of long-term potentiation (LTP) The role of calcium/calmodulin-dependent kinase II (CamKII) in the induction,
Neurons of the Bed Nucleus of the Stria Terminalis (BNST)
") Neurons of the Bed Nucleus of the Stria Terminalis (BNST) Electrophysiological Properties and Their Response to Serotonin DONALD G. RAINNIE a Harvard Medical School and Department of Psychiatry, Brockton
Neurons of the Bed Nucleus of the Stria Terminalis (BNST) Electrophysiological Properties and Their Response to Serotonin DONALD G. RAINNIE a Harvard Medical School and Department of Psychiatry, Brockton
When cells are already maximally potentiated LTP is occluded.
 When cells are already maximally potentiated LTP is occluded. Stein, V et al., (2003) J Neurosci, 23:5503-6606. Also found in Rat Barrel Cortex Ehrlich & Malinow (2004) J. Neurosci. 24:916-927 Over-expression
When cells are already maximally potentiated LTP is occluded. Stein, V et al., (2003) J Neurosci, 23:5503-6606. Also found in Rat Barrel Cortex Ehrlich & Malinow (2004) J. Neurosci. 24:916-927 Over-expression
GABA B Receptor-Mediated Presynaptic Inhibition Has History-Dependent Effects on Synaptic Transmission during Physiologically Relevant Spike Trains
 The Journal of Neuroscience, June 15, 2003 23(12):4809 4814 4809 Brief Communication GABA B Receptor-Mediated Presynaptic Inhibition Has History-Dependent Effects on Synaptic Transmission during Physiologically
The Journal of Neuroscience, June 15, 2003 23(12):4809 4814 4809 Brief Communication GABA B Receptor-Mediated Presynaptic Inhibition Has History-Dependent Effects on Synaptic Transmission during Physiologically
Supplementary Information
 Hyperpolarization-activated cation channels inhibit EPSPs by interactions with M-type K + channels Meena S. George, L.F. Abbott, Steven A. Siegelbaum Supplementary Information Part 1: Supplementary Figures
Hyperpolarization-activated cation channels inhibit EPSPs by interactions with M-type K + channels Meena S. George, L.F. Abbott, Steven A. Siegelbaum Supplementary Information Part 1: Supplementary Figures
Neuroscience 201A (2016) - Problems in Synaptic Physiology
 - Problems in Synaptic Physiology") Question 1: The record below in A shows an EPSC recorded from a cerebellar granule cell following stimulation (at the gap in the record) of a mossy fiber input. These responses are, then, evoked by stimulation.
Question 1: The record below in A shows an EPSC recorded from a cerebellar granule cell following stimulation (at the gap in the record) of a mossy fiber input. These responses are, then, evoked by stimulation.
Ionotropic glutamate receptors (iglurs)
") Ionotropic glutamate receptors (iglurs) GluA1 GluA2 GluA3 GluA4 GluN1 GluN2A GluN2B GluN2C GluN2D GluN3A GluN3B GluK1 GluK2 GluK3 GluK4 GluK5 The general architecture of receptor subunits Unique properties
Ionotropic glutamate receptors (iglurs) GluA1 GluA2 GluA3 GluA4 GluN1 GluN2A GluN2B GluN2C GluN2D GluN3A GluN3B GluK1 GluK2 GluK3 GluK4 GluK5 The general architecture of receptor subunits Unique properties
Differential Effect of TEA on Long-Term Synaptic Modification in Hippocampal CA1 and Dentate Gyrus in vitro
 Neurobiology of Learning and Memory 76, 375 387 (2001) doi:10.1006/nlme.2001.4032, available online at http://www.idealibrary.com on Differential Effect of TEA on Long-Term Synaptic Modification in Hippocampal
Neurobiology of Learning and Memory 76, 375 387 (2001) doi:10.1006/nlme.2001.4032, available online at http://www.idealibrary.com on Differential Effect of TEA on Long-Term Synaptic Modification in Hippocampal
Neuronal Plasticity, Learning and Memory. David Keays Institute of Molecular Pathology
 Neuronal Plasticity, Learning and Memory David Keays Institute of Molecular Pathology http://keayslab.org Structure 1. What is learning and memory? 2. Anatomical basis 3. Cellular basis 4. Molecular
Neuronal Plasticity, Learning and Memory David Keays Institute of Molecular Pathology http://keayslab.org Structure 1. What is learning and memory? 2. Anatomical basis 3. Cellular basis 4. Molecular
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
Roles of NMDA NR2B Subtype Receptor in Prefrontal Long-Term Potentiation and Contextual Fear Memory
 Neuron, Vol. 47, 859 872, September 15, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.08.014 Roles of NMDA NR2B Subtype Receptor in Prefrontal Long-Term Potentiation and Contextual Fear
Neuron, Vol. 47, 859 872, September 15, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.08.014 Roles of NMDA NR2B Subtype Receptor in Prefrontal Long-Term Potentiation and Contextual Fear
Bidirectional NMDA receptor plasticity controls CA3 output and heterosynaptic metaplasticity
 Bidirectional NMDA receptor plasticity controls CA output and heterosynaptic metaplasticity David L. Hunt, Nagore Puente, Pedro Grandes, Pablo E. Castillo a NMDAR EPSC (pa) - - -8-6 -4 - st 5 nd 5 b NMDAR
Bidirectional NMDA receptor plasticity controls CA output and heterosynaptic metaplasticity David L. Hunt, Nagore Puente, Pedro Grandes, Pablo E. Castillo a NMDAR EPSC (pa) - - -8-6 -4 - st 5 nd 5 b NMDAR
DOMINIQUE DEBANNE*, BEAT H. GÄHWILER, AND SCOTT M. THOMPSON MATERIALS AND METHODS
 Proc. Natl. Acad. Sci. USA Vol. 93, pp. 11225 11230, October 1996 Neurobiology Cooperative interactions in the induction of long-term potentiation and depression of synaptic excitation between hippocampal
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 11225 11230, October 1996 Neurobiology Cooperative interactions in the induction of long-term potentiation and depression of synaptic excitation between hippocampal
Supplementary Figure 1: Kv7 currents in neonatal CA1 neurons measured with the classic M- current voltage-clamp protocol.
 Supplementary Figures 1-11 Supplementary Figure 1: Kv7 currents in neonatal CA1 neurons measured with the classic M- current voltage-clamp protocol. (a), Voltage-clamp recordings from CA1 pyramidal neurons
Supplementary Figures 1-11 Supplementary Figure 1: Kv7 currents in neonatal CA1 neurons measured with the classic M- current voltage-clamp protocol. (a), Voltage-clamp recordings from CA1 pyramidal neurons
Requirements for LTP Induction by Pairing in Hippocampal CA1 Pyramidal Cells
 Requirements for LTP Induction by Pairing in Hippocampal CA1 Pyramidal Cells HUAN-XIN CHEN, NIKOLAI OTMAKHOV, AND JOHN LISMAN Volen Center for Complex Systems, Biology Department, Brandeis University,
Requirements for LTP Induction by Pairing in Hippocampal CA1 Pyramidal Cells HUAN-XIN CHEN, NIKOLAI OTMAKHOV, AND JOHN LISMAN Volen Center for Complex Systems, Biology Department, Brandeis University,
How Nicotinic Signaling Shapes Neural Networks
 How Nicotinic Signaling Shapes Neural Networks Darwin K. Berg Division of Biological Sciences University of California, San Diego Nicotinic Cholinergic Signaling Uses the transmitter ACh to activate cation-selective
How Nicotinic Signaling Shapes Neural Networks Darwin K. Berg Division of Biological Sciences University of California, San Diego Nicotinic Cholinergic Signaling Uses the transmitter ACh to activate cation-selective
previously shown (10), however, this manipulation by itself does not reliably result in the development of a large
, however, this manipulation by itself does not reliably result in the development of a large") Proc. Nati. Acad. Sci. USA Vol. 85, pp. 9346-9350, December 1988 Neurobiology Long-term potentiation differentially affects two components of synaptic responses in hippocampus (plasticity/n-methyl-d-aspartate/d-2-amino-5-phosphonovglerate/facilitation)
Proc. Nati. Acad. Sci. USA Vol. 85, pp. 9346-9350, December 1988 Neurobiology Long-term potentiation differentially affects two components of synaptic responses in hippocampus (plasticity/n-methyl-d-aspartate/d-2-amino-5-phosphonovglerate/facilitation)
<student name> Undergraduate Research Grant Proposal
 Undergraduate Research Grant Proposal A. Project Description Objective of research: The objective of this study is to determine if hippocampal dopamine D1 receptors facilitate novel object
Undergraduate Research Grant Proposal A. Project Description Objective of research: The objective of this study is to determine if hippocampal dopamine D1 receptors facilitate novel object
Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex
 Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Fear conditioning induces associative long-term potentiation in the amygdala
 11 December 1997 Nature 390, 604-607 (1997) Macmillan Publishers Ltd. Fear conditioning induces associative long-term potentiation in the amygdala MICHAEL T. ROGAN, URSULA V. STÄUBLI & JOSEPH E. LEDOUX
11 December 1997 Nature 390, 604-607 (1997) Macmillan Publishers Ltd. Fear conditioning induces associative long-term potentiation in the amygdala MICHAEL T. ROGAN, URSULA V. STÄUBLI & JOSEPH E. LEDOUX
TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki
 TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki Rich Turner (turner@gatsby.ucl.ac.uk) Gatsby Unit, 22/04/2005 Rich T. Introduction Interneuron def = GABAergic non-principal cell Usually
TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki Rich Turner (turner@gatsby.ucl.ac.uk) Gatsby Unit, 22/04/2005 Rich T. Introduction Interneuron def = GABAergic non-principal cell Usually
DNA and Histone Methylation in Learning and Memory
 EXPERIMENTAL BIOLOGY ANNUAL MEETING April 2010 DNA and Histone Methylation in Learning and Memory J. David Sweatt Dept of Neurobiology McKnight Brain Institute UAB School of Medicine The Molecular Basis
EXPERIMENTAL BIOLOGY ANNUAL MEETING April 2010 DNA and Histone Methylation in Learning and Memory J. David Sweatt Dept of Neurobiology McKnight Brain Institute UAB School of Medicine The Molecular Basis
Supplementary Materials for VAMP4 directs synaptic vesicles to a pool that selectively maintains asynchronous neurotransmission
 Supplementary Materials for VAMP4 directs synaptic vesicles to a pool that selectively maintains asynchronous neurotransmission Jesica Raingo, Mikhail Khvotchev, Pei Liu, Frederic Darios, Ying C. Li, Denise
Supplementary Materials for VAMP4 directs synaptic vesicles to a pool that selectively maintains asynchronous neurotransmission Jesica Raingo, Mikhail Khvotchev, Pei Liu, Frederic Darios, Ying C. Li, Denise
Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression
 The Journal of Neuroscience, November 1, 1997, 17(21):8613 8620 Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression Sukwoo
The Journal of Neuroscience, November 1, 1997, 17(21):8613 8620 Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression Sukwoo
Glycine-induced long-term potentiation is associated with structural and functional modifications of
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 9451 9456, August 1997 Neurobiology Glycine-induced long-term potentiation is associated with structural and functional modifications of -amino-3-hydroxyl-5- methyl-4-isoxazolepropionic
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 9451 9456, August 1997 Neurobiology Glycine-induced long-term potentiation is associated with structural and functional modifications of -amino-3-hydroxyl-5- methyl-4-isoxazolepropionic
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
CONTEXT. LTP (long term potentiation) definition. LTP as a interesting mechanism for learning and memory
 definition. LTP as a interesting mechanism for learning and memory") CONTEXT LTP (long term potentiation) definition LTP as a interesting mechanism for learning and memory LTP is due primarily to a pre or post- synaptic modification? (Increased Glut release or increased
CONTEXT LTP (long term potentiation) definition LTP as a interesting mechanism for learning and memory LTP is due primarily to a pre or post- synaptic modification? (Increased Glut release or increased
Basal Ganglia Anatomy, Physiology, and Function. NS201c
 Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Phosphatidylinositol 3-Kinase Regulates the Induction of Long-Term Potentiation through Extracellular Signal- Related Kinase-Independent Mechanisms
 The Journal of Neuroscience, May 1, 2003 23(9):3679 3688 3679 Phosphatidylinositol 3-Kinase Regulates the Induction of Long-Term Potentiation through Extracellular Signal- Related Kinase-Independent Mechanisms
The Journal of Neuroscience, May 1, 2003 23(9):3679 3688 3679 Phosphatidylinositol 3-Kinase Regulates the Induction of Long-Term Potentiation through Extracellular Signal- Related Kinase-Independent Mechanisms
Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA
 Contribution of Calcium Channel α 2 δ Binding Sites to the Pharmacology of Gabapentin and Pregabalin Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA Disclosure Information Charlie Taylor, PhD
Contribution of Calcium Channel α 2 δ Binding Sites to the Pharmacology of Gabapentin and Pregabalin Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA Disclosure Information Charlie Taylor, PhD
Synaptic Transmission: Ionic and Metabotropic
 Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Nature Neuroscience: doi: /nn Supplementary Figure 1. Trial structure for go/no-go behavior
 Supplementary Figure 1 Trial structure for go/no-go behavior a, Overall timeline of experiments. Day 1: A1 mapping, injection of AAV1-SYN-GCAMP6s, cranial window and headpost implantation. Water restriction
Supplementary Figure 1 Trial structure for go/no-go behavior a, Overall timeline of experiments. Day 1: A1 mapping, injection of AAV1-SYN-GCAMP6s, cranial window and headpost implantation. Water restriction
Synaptic plasticity and hippocampal memory
 Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
COGNITIVE SCIENCE 107A. Hippocampus. Jaime A. Pineda, Ph.D.
 COGNITIVE SCIENCE 107A Hippocampus Jaime A. Pineda, Ph.D. Common (Distributed) Model of Memory Processes Time Course of Memory Processes Long Term Memory DECLARATIVE NON-DECLARATIVE Semantic Episodic Skills
COGNITIVE SCIENCE 107A Hippocampus Jaime A. Pineda, Ph.D. Common (Distributed) Model of Memory Processes Time Course of Memory Processes Long Term Memory DECLARATIVE NON-DECLARATIVE Semantic Episodic Skills
Modeling of Hippocampal Behavior
 Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
The clathrin adaptor Numb regulates intestinal cholesterol. absorption through dynamic interaction with NPC1L1
 The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
Fig. 4. The activity of Pkc -transduced neurons is required for enhanced learning. After gene transfer, rats were tested on [] vs. +.
![Fig. 4. The activity of Pkc -transduced neurons is required for enhanced learning. After gene transfer, rats were tested on [] vs. +.](/thumbs/90/102196023.jpg "Fig. 4. The activity of Pkc -transduced neurons is required for enhanced learning. After gene transfer, rats were tested on [] vs. +.") Research Interests Advanced cognitive learning is encoded in distributed circuits that span multiple forebrain areas. Further, synaptic plasticity and neural network theories hypothesize that essential
Research Interests Advanced cognitive learning is encoded in distributed circuits that span multiple forebrain areas. Further, synaptic plasticity and neural network theories hypothesize that essential
Short- and long-lasting consequences of in vivo nicotine treatment
 Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
Key words: fear conditioning; APV; ifenprodil; verapamil; LTP; amygdala
 The Journal of Neuroscience, June 15, 2002, 22(12):5239 5249 NMDA Receptors and L-Type Voltage-Gated Calcium Channels Contribute to Long-Term Potentiation and Different Components of Fear Memory Formation
The Journal of Neuroscience, June 15, 2002, 22(12):5239 5249 NMDA Receptors and L-Type Voltage-Gated Calcium Channels Contribute to Long-Term Potentiation and Different Components of Fear Memory Formation
Direct current stimulation modulates LTP and LTD: activity-dependence and dendritic effects
 City University of New York (CUNY) CUNY Academic Works Master's Theses City College of New York 2016 Direct current stimulation modulates LTP and LTD: activity-dependence and dendritic effects Gregory
City University of New York (CUNY) CUNY Academic Works Master's Theses City College of New York 2016 Direct current stimulation modulates LTP and LTD: activity-dependence and dendritic effects Gregory
Requirement of Translation But Not Transcription for the Maintenance of Long-Term Depression in the CA1 Region of Freely Moving Rats
 The Journal of Neuroscience, November 15, 2000, 20(22):8572 8576 Requirement of Translation But Not Transcription for the Maintenance of Long-Term Depression in the CA1 Region of Freely Moving Rats Denise
The Journal of Neuroscience, November 15, 2000, 20(22):8572 8576 Requirement of Translation But Not Transcription for the Maintenance of Long-Term Depression in the CA1 Region of Freely Moving Rats Denise
Ca 2+ Permeable AMPA Receptor Induced Long-Term Potentiation Requires PI3/MAP Kinases but Not Ca/CaM- Dependent Kinase II
 Ca 2+ Permeable AMPA Receptor Induced Long-Term Potentiation Requires PI3/MAP Kinases but Not Ca/CaM- Dependent Kinase II Suhail Asrar 1,2, Zikai Zhou 1,2, Wei Ren 3, Zhengping Jia 1,2 1 Neurosciences
Ca 2+ Permeable AMPA Receptor Induced Long-Term Potentiation Requires PI3/MAP Kinases but Not Ca/CaM- Dependent Kinase II Suhail Asrar 1,2, Zikai Zhou 1,2, Wei Ren 3, Zhengping Jia 1,2 1 Neurosciences
Sample Lab Report 1 from 1. Measuring and Manipulating Passive Membrane Properties
 Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Plasticity of Cerebral Cortex in Development
 Plasticity of Cerebral Cortex in Development Jessica R. Newton and Mriganka Sur Department of Brain & Cognitive Sciences Picower Center for Learning & Memory Massachusetts Institute of Technology Cambridge,
Plasticity of Cerebral Cortex in Development Jessica R. Newton and Mriganka Sur Department of Brain & Cognitive Sciences Picower Center for Learning & Memory Massachusetts Institute of Technology Cambridge,
Differential Mechanisms of Transmission at Three Types of Mossy Fiber Synapse
 The Journal of Neuroscience, November 15, 2000, 20(22):8279 8289 Differential Mechanisms of Transmission at Three Types of Mossy Fiber Synapse Katalin Toth, Gregory Suares, J. Josh Lawrence, Emily Philips-Tansey,
The Journal of Neuroscience, November 15, 2000, 20(22):8279 8289 Differential Mechanisms of Transmission at Three Types of Mossy Fiber Synapse Katalin Toth, Gregory Suares, J. Josh Lawrence, Emily Philips-Tansey,
Synaptic plasticity. Mark van Rossum. Institute for Adaptive and Neural Computation University of Edinburgh
 Synaptic plasticity Mark van Rossum Institute for Adaptive and Neural Computation University of Edinburgh 1 Human memory systems 2 Psychologists have split up memory in: Declarative memory * Episodic memory
Synaptic plasticity Mark van Rossum Institute for Adaptive and Neural Computation University of Edinburgh 1 Human memory systems 2 Psychologists have split up memory in: Declarative memory * Episodic memory
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature6416 Supplementary Notes Spine Ca 2+ signals produced by glutamate uncaging We imaged uncaging-evoked [Ca 2+ ] transients in neurons loaded with a green Ca 2+ - sensitive indicator (G;
doi: 1.138/nature6416 Supplementary Notes Spine Ca 2+ signals produced by glutamate uncaging We imaged uncaging-evoked [Ca 2+ ] transients in neurons loaded with a green Ca 2+ - sensitive indicator (G;
Nature Neuroscience: doi: /nn Supplementary Figure 1. Large-scale calcium imaging in vivo.
 Supplementary Figure 1 Large-scale calcium imaging in vivo. (a) Schematic illustration of the in vivo camera imaging set-up for large-scale calcium imaging. (b) High-magnification two-photon image from
Supplementary Figure 1 Large-scale calcium imaging in vivo. (a) Schematic illustration of the in vivo camera imaging set-up for large-scale calcium imaging. (b) High-magnification two-photon image from
Lecture: Introduction to nervous system development
 Lecture: Introduction to nervous system development Prof. Ilan Davis, Department of Biochemistry. Wellcome Senior Research Fellow Senior Research Fellow, Jesus College ilan.davis@bioch.ox.ac.uk http://www.ilandavis.com
Lecture: Introduction to nervous system development Prof. Ilan Davis, Department of Biochemistry. Wellcome Senior Research Fellow Senior Research Fellow, Jesus College ilan.davis@bioch.ox.ac.uk http://www.ilandavis.com
Terminology. Terminology. Terminology. Terminology. Terminology. Bromodeoxyuridine
 Kateřina Náměstková, Zuzana Šimonová, Eva Syková Behavioural Brain Research Bromodeoxyuridine : Doublecortin : DCX Glial Fibrillary Acidic Protein : GFAP Trace eye blink conditioning 1 Volume 163 : pp.
Kateřina Náměstková, Zuzana Šimonová, Eva Syková Behavioural Brain Research Bromodeoxyuridine : Doublecortin : DCX Glial Fibrillary Acidic Protein : GFAP Trace eye blink conditioning 1 Volume 163 : pp.
Chapter 9: Biochemical Mechanisms for Information Storage at the Cellular Level. From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D.
 Chapter 9: Biochemical Mechanisms for Information Storage at the Cellular Level From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Chapter 9: Dendritic Spine Figure 1 Summary: Three Primary
Chapter 9: Biochemical Mechanisms for Information Storage at the Cellular Level From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Chapter 9: Dendritic Spine Figure 1 Summary: Three Primary
Electrophysiological Effects of Three Groups of Glutamate Metabotropic Receptors in Rat Piriform Cortex
 Cellular and Molecular Neurobiology, Vol. 26, Nos. 4 6, July/August 2006 ( C 2006) DOI: 10.1007/s10571-006-9102-4 Electrophysiological Effects of Three Groups of Glutamate Metabotropic Receptors in Rat
Cellular and Molecular Neurobiology, Vol. 26, Nos. 4 6, July/August 2006 ( C 2006) DOI: 10.1007/s10571-006-9102-4 Electrophysiological Effects of Three Groups of Glutamate Metabotropic Receptors in Rat
Lisa M. Giocomo & Michael E. Hasselmo
 Mol Neurobiol (2007) 36:184 200 DOI 10.1007/s12035-007-0032-z Neuromodulation by Glutamate and Acetylcholine can Change Circuit Dynamics by Regulating the Relative Influence of Afferent Input and Excitatory
Mol Neurobiol (2007) 36:184 200 DOI 10.1007/s12035-007-0032-z Neuromodulation by Glutamate and Acetylcholine can Change Circuit Dynamics by Regulating the Relative Influence of Afferent Input and Excitatory
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11775 Supplementary Discussion Based on our data, we propose that LTP requires a large extrasynaptic pool of surface receptors regardless of their subunit composition.
SUPPLEMENTARY INFORMATION doi:10.1038/nature11775 Supplementary Discussion Based on our data, we propose that LTP requires a large extrasynaptic pool of surface receptors regardless of their subunit composition.
Astrocyte signaling controls spike timing-dependent depression at neocortical synapses
 Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06994 A phosphatase cascade by which rewarding stimuli control nucleosomal response A. Stipanovich*, E. Valjent*, M. Matamales*, A. Nishi, J.H. Ahn, M. Maroteaux, J. Bertran-Gonzalez,
doi: 10.1038/nature06994 A phosphatase cascade by which rewarding stimuli control nucleosomal response A. Stipanovich*, E. Valjent*, M. Matamales*, A. Nishi, J.H. Ahn, M. Maroteaux, J. Bertran-Gonzalez,
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/328/5983/1288/dc1 Supporting Online Material for Induction of Fear Extinction with Hippocampal-Infralimbic BDNF Jamie Peters, Laura M. Dieppa-Perea, Loyda M. Melendez,
www.sciencemag.org/cgi/content/full/328/5983/1288/dc1 Supporting Online Material for Induction of Fear Extinction with Hippocampal-Infralimbic BDNF Jamie Peters, Laura M. Dieppa-Perea, Loyda M. Melendez,
Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus
 Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus Authors: Jeffrey Lopez-Rojas a1, Martin Heine b1 and Michael R. Kreutz ac1 a Research Group Neuroplasticity, b Research
Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus Authors: Jeffrey Lopez-Rojas a1, Martin Heine b1 and Michael R. Kreutz ac1 a Research Group Neuroplasticity, b Research
Cellular and molecular mechanisms of memory storage Synaptic plasticity and Alzheimer s disease Reward learning and addiction
 Brian Wiltgen, Ph.D. Curriculum Vitae University of Virginia Department of Psychology Charlottesville, VA 22904 Telephone: (434) 924-1458 E-mail: bw4fh@virginia.edu RESEARCH INTERESTS Cellular and molecular
Brian Wiltgen, Ph.D. Curriculum Vitae University of Virginia Department of Psychology Charlottesville, VA 22904 Telephone: (434) 924-1458 E-mail: bw4fh@virginia.edu RESEARCH INTERESTS Cellular and molecular