Running title: Cardiosplenic Axis in Heart Failure
|
|
|
- Ashley Junior Hicks
- 6 years ago
- Views:
Transcription

1 Remodeling of the Mononuclear Phagocyte Network Underlies Chronic Inflammation and Disease Progression in Heart Failure: Critical Importance of the Cardiosplenic Axis Mohamed Ameen Ismahil 1, Tariq Hamid 1, Shyam S. Bansal 1, Bindiya Patel 1, Justin R. Kingery 2, and Sumanth D. Prabhu 1 1 Division of Cardiovascular Disease, Department of Medicine, University of Alabama at Birmingham and Birmingham VAMC, Birmingham, AL, and; 2 Department of Medicine, University of Louisville Louisville, KY. Running title: Cardiosplenic Axis in Heart Failure Downloaded from by guest on June 3, 218 Subject codes: [18] Heart failure-basic studies [18] Other myocardial biology [11] Congestive Address correspondence to: Dr. Sumanth D. Prabhu Division of Cardiovascular Disease University of Alabama Birmingham 311 Tinsley Harrison Tower 19 University Blvd Birmingham, AL Tel: /372 Fax: sprabhu@uab.edu This manuscript was sent to Ali J. Marian, Consulting Editor, for review by expert referees, editorial decision, and final disposition. In October 213, the average time from submission to first decision for all original research papers submitted to Circulation Research was days. DOI: /CIRCRESAHA
2 ABSTRACT Rational: The role of mononuclear phagocytes in chronic heart failure (HF) is unknown. Objective: To delineate monocyte, macrophage, and dendritic cell (DC) trafficking in HF and define the contribution of the spleen to cardiac remodeling. Downloaded from by guest on June 3, 218 Methods and Results: We evaluated C7Bl/6 mice with chronic HF, 8 weeks after coronary ligation. As compared with sham-operated controls, HF mice exhibited: 1) increased pro-inflammatory CD11b + F/8 + CD26 macrophages and CD11b + F/8 + Gr-1 hi monocytes in the heart and peripheral blood, respectively, and diminished CD11b + F/8 + Gr-1 hi monocytes in the spleen, 2) significantly augmented CD11c + B22 classical DCs (cdcs) and CD11c +/low B22 + plasmacytoid DCs (pdcs) in both the heart and spleen, and increased cdcs and pdcs in peripheral blood and bone marrow, respectively, 3) augmented CD+ helper and CD8+ cytotoxic T cells in the spleen, and ) profound splenic remodeling with more abundant white pulp follicles, markedly increased size of the marginal zone and germinal centers, and increased expression of alarmins. Splenectomy in mice with established HF reversed pathological cardiac remodeling and inflammation. Splenocytes adoptively transferred from mice with HF, but not from sham-operated mice, homed to the heart and induced long-term LV dilatation, dysfunction, and fibrosis in naïve recipients. Recipient mice also exhibited monocyte activation and splenic remodeling similar to HF mice. Conclusions: Activation of mononuclear phagocytes is central to the progression of cardiac remodeling in HF and heightened antigen processing in the spleen plays a critical role in this process. Splenocytes (presumably splenic monocytes and DCs) promote immune-mediated injurious responses in the failing heart, and retain this memory upon adoptive transfer. Keywords: Heart failure, inflammation, monocytes, dendritic cells, spleen, alarmins, remodeling Nonstandard Abbreviations and Acronyms: HF heart failure DC dendritic cell cdc classical dendritic cell pdc plasmacytoid dendritic cell TNF tumor necrosis factor BM bone marrow DAPI ',6-diamidino-2-phenylindole FITC fluorescein isothiocyanate APC allophycocyanin Cy cyanine dye NC nana crystal PE R-phycoerythrin CD cluster of differentiation Ly6C lymphocyte antigen 6C Gr-1 granulocyte-1 Lin lineage NK1.1 natural killer cell 1.1 B22 mature, resting B cell antigen CBA cytometric bead array DOI: /CIRCRESAHA
3 Downloaded from by guest on June 3, 218 IL Interleukin MCP-1 monocyte chemoattractant protein-1 IFN interferon WGA wheat germ agglutinin SRP subcapsular red pulp Th1 T-helper cell pathway-1 Th2 T-helper cell pathway-2 CCR2 chemokine (C-C motif) receptor 2 DAMP damage associated molecular pattern RP red pulp WP white pulp MZ marginal zone PALS periarteriolar lymphoid sheath CCR2 C-C chemokine receptor type 2 CCL chemokine (C-C motif) ligand CXCR3 chemokine (C-X-C motif) receptor 3 CX3CR1 CX3C chemokine receptor 1 TGF-β transforming growth factor (TGF)-β HMGB1 high mobility group box-1 LPS lipopolysaccharide INTRODUCTION Persistent inflammation is a hallmark of chronic heart failure (HF). 1 In humans, augmented levels of circulating and myocardial pro-inflammatory cytokines correlate with HF disease stage and mortality. 2 Moreover, animal studies support a causal role for inflammatory mediators in many of the pathological responses in the failing heart. 1, 3 These observations formed the basis for the cytokine hypothesis of HF and suggested therapeutic promise for cytokine antagonism. Paradoxically, however, clinical trials of antibody-based neutralization of tumor necrosis factor (TNF), a foundation pro-inflammatory cytokine, failed to show any therapeutic benefit (and was even harmful at high doses). 1, This unexpected clinical response alludes to an overriding and underappreciated complexity of cytokine networks in HF that cannot be effectively targeted in an all-or-none manner. Indeed, to date, despite a wealth of studies establishing pathological inflammatory activation in HF, no large-scale immunomodulatory therapies have been successfully translated to clinical practice. The main translational focus of immune modulation in HF has thus far been directed toward protein mediators such as cytokines. In contrast, much less attention has been given to modulating the underlying immune and inflammatory cell network that serves both as an important source and effector target for cytokines. As HF is a systemic disease triggered by cardiac dysfunction, chronic cytokine elaboration suggests global alterations in inflammatory cell populations both within and beyond the failing heart, such as in lymphoid tissue and bone marrow. Prominent among these cell types are monocyte/macrophage and dendritic cell (DC) populations (mononuclear phagocytes) that are of signal importance in regulating (and generating) immune responses. 6-9 Macrophages and their direct precursors monocytes are central to tissue inflammation and innate immunity, whereas DCs are professional antigenpresenting and -processing cells that are critical for adaptive immune responses and immune memory/tolerance. The functional roles of these cells are complex and heterogeneous, as different subtypes can either promote or suppress a variety of responses including inflammation, cell clearance, wound healing, and autoimmunity. 6-9 DOI: /CIRCRESAHA
4 In acute myocardial infarction, both monocytes and DCs have been shown to contribute importantly to early post-infarction remodeling. 1-1 In contrast, whether and how the myelomonocytic network between lymphoid tissue, bone marrow, and the heart are altered in chronic HF is unknown. Determining this is of utmost importance to understanding inflammatory dysregulation in HF, and for targeting potential therapies directed at specific cell populations. Hence, we undertook a comprehensive analysis of the mononuclear phagocyte network in chronic HF and the contribution of the spleen, the largest secondary lymphoid organ in the body, to this network and to pathological left ventricular (LV) remodeling. Our results demonstrate, to our knowledge for the first time, profound alterations in cardiac and splenic mononuclear cells and splenic tissue niches in chronic HF that: 1) promote the circulation and residence of pro-inflammatory monocytes/macrophages, classical DCs, and plasmacytoid DCs, and 2) regulate both chronic inflammation and the progression of pathological LV remodeling. METHODS Downloaded from by guest on June 3, 218 All studies were performed in compliance with the NIH Guide for the Care and Use of Laboratory Animals (DHHS publication No. [NIH] 8-23, revised 1996). The University of Alabama at Birmingham Institutional Animal Care and Use Committee gave local approval for these studies. A total of 12 mice were used. Mouse models and surgical protocol. Male C7BL/6J mice 1-12 weeks of age (Jackson Laboratories, stock #66) were used. To induce pathological LV remodeling and HF, the mice underwent left thoracotomy and left coronary artery ligation (n=19) as previously described. 3, 1, 16 Sham operated mice (n=12) were used as controls. The mice were followed for 8 weeks after operation and evaluated for the various readouts. Echocardiography. Mouse echocardiography was performed under anesthesia with tribromoethanol (.2 mg/g IP), and light (~1%) isoflurane as needed, using a VisualSonics Vevo 77 High-Resolution System with a RMV77B scanhead. Mice were imaged on a heated, bench-mounted adjustable rail system (Vevo Imaging Station) that allowed steerable and hands-free manipulation of the ultrasound transducer. Isolation of splenocytes. Eight weeks after coronary ligation or sham operation, the spleen was removed aseptically and placed in a 3 mm tissue culture dish with ~ ml of DMEM culture medium (Gibco, Invitrogen). Splenocytes were isolated according to the protocol of Lavelle at al, 17 with slight modifications. Briefly, the spleen was finely minced using a scalpel. Repeated pipetting was used to disperse cells from minced fragments, and single cells were transferred to a fresh tube and kept on ice. For larger tissue pieces, the procedure was repeated several times in the dish with fresh DMEM. The final cell suspension was pipetted through a nylon cell strainer, 1 μm (BD Falcon), into a fresh tube and centrifuged at 1g for min at C. The supernatant was decanted, and the pellet re-suspended in. ml of RBC lysis buffer (ebiosciences) for min at room temperature (RT). The addition of fresh DMEM neutralized the erythrocyte lysis. Pelleted splenocytes were then re-suspended in fresh DMEM (.-1. ml). Isolation of bone marrow (BM) and peripheral blood cells. After mice were sacrificed, femurs and tibiae were dissected from the adherent soft tissue. The tip of each bone was removed with scissors, and the marrow was flushed with DMEM using a 27-gauge needle and aspirated. BM cells were filtered through a 1 μm nylon cell strainer (BD Falcon), sedimented at 1g for min at C, and re-suspended in RBC lysis buffer. After complete erythrocyte lysis, BM cells were re-suspended in fresh 1-2 ml DMEM. DOI: /CIRCRESAHA
5 Peripheral blood (~ μl) was collected in BD Microtainer tubes with EDTA. Erythrocytes were lysed with 2 ml RBC lysis buffer for min on ice. Leukocytes were collected by centrifugation (38g for 1 min at C), and resuspended in ul of ice-cold flow cytometry staining buffer (ebioscience). Downloaded from by guest on June 3, 218 Isolation of mononuclear cells from the heart. Single mononuclear cells were isolated from sham and failing hearts following the method of Austyn et al. 18 Mouse hearts were excised in toto and placed in heparinized saline. After removal of epicardial fatty tissue and the aorta, the heart was finely minced into ~1-2 mm pieces and blood was removed by repeated washing in saline (with stepwise reductions in added heparin). The tissue was digested with collagenase (Worthington, 1mg/ml), trypsin (Gibco, Invitrogen,.1%), and DNAse ( Prime, 1 μg/ml) in 1 ml RPMI media (Gibco) for min at 37 C with occasional shaking. Released cells were separated from solid tissue by filtering through a 1 μm nylon cell strainer (BD Falcon), washed with R1 media (RPMI-16 supplemented with 1% heat-inactivated FCS, 2mM-L-Glutamine, 2 μm-2- Mercaptoethanol, 1% penicillin/streptomycin), and placed on ice. These steps were repeated 2-3 times to digest the remaining tissue. Any residual solid tissue was treated sequentially with EDTA (2 mm) in digestion media for 1 min at 37 o C, collagenase (2 mg/ml) in R1 media for 2 min at 37 o C, and released cells filtered through the strainer. All collected cells were pooled and pelleted at 1g for min at o C. Single cell suspensions were layered on Ficoll gradient solution and centrifuged at 2g for 2 min. To reduce myocyte contamination in the mononuclear cell suspension, 7% of the total volume (excepting cell debris) was collected and washed in R1 media. The presence of nucleated cells was confirmed by DAPI staining during flow cytometry analysis. In a separate aliquot, cell viability (>8%) was confirmed using the trypan blue exclusion method. Flow cytometry. Isolated cell suspensions were incubated for 3 min at RT in a cocktail of fluorophore-labeled mabs (BD Biosciences or as otherwise indicated), as appropriate for the specific study, against: F/8-Pacific Blue; CD8-FITC; CDR/B22-APC; CD11b-6-NC (ebiosciences); Gr-1-APC (ebiosciences); CD11c-PE- Cy7; CD86-PE-Cy; CD26-FITC; Siglec-H-efluor 66 (ebiosciences), CD.1-FITC (Miltenyi Biotech); CD.2-PE. Lineage (Lin)1 antibody cocktail was used for DC identification (to exclude thymocytes: CD9.2-PE, natural killer cells: NK1.1-PE, CD9B-PE, and granulocytes: Gr-1-PE), and Lin2 antibody cocktail was used for macrophage/monocyte subset identification (CD9.2-PE, NK1.1-PE, CD9B-PE, and CDR/B22-PE for B-cells). In separate assessments of splenic T-cells, cell suspensions were incubated with CD3-FITC, CD-6 NC and CD8-6 NC (ebiosciences). Of note, the anti-gr-1 antibody recognizes both Ly6C and Ly6G antigens. 19 Activated macrophages/monocytes were identified as CD11b + F/8 + cells. 2, 21 Pro-inflammatory monocytes were identified as Lin2 CD11b + F/8 + Gr-1 hi and anti-inflammatory monocytes as Lin2 CD11b + F/8 + Gr-1 low cells. 9, 2-22 Pro-inflammatory M1 and anti-inflammatory M2 macrophages in tissue were sub-classified from Lin2 CD11b + F/8 + cells based on the absence or presence of CD26 (mannose receptor) expression. 8 Dendritic cells were identified as Lin1 CD11c + cells. 6, 9, 22 Classical dendritic cells (cdc) were identified as Lin1 CD11c + B22, whereas non-classical plasmacytoid dendritic cells (pdc) were identified as Lin1 CD11c +/low B22 + 6, In some studies, pdcs were identified as Siglec-H + cells. 2 Lymphoid cdcs were further subdivided as CD8 + or CD8, 6, 22 and pdcs as CD86 + or CD86 (classical costimulatory molecule and marker of pdc maturity). 26 Helper and cytotoxic T-cells were characterized as CD3 + CD + and CD3 + CD8 + CD cells, respectively. 27 For the heart, spleen, and BM, identified cells were normalized for total cell population. For peripheral blood, cell numbers were normalized for the total lymphocyte and monocyte gate. Data were acquired on an LSRII flow cytometer (BD Biosciences) and analyzed with FlowJo software, version Immunohistochemistry and confocal microscopy. Formalin-fixed, paraffin-embedded hearts and spleens from sham and HF mice were sectioned at μm DOI: /CIRCRESAHA
6 thickness, deparaffinized and rehydrated; (immuno)histological staining was performed as previously described. 3, 1 Masson s trichrome was used to evaluate tissue fibrosis and general histology (heart and spleen), and Alexa Fluor 88-conjugated wheat-germ agglutinin (WGA; Invitrogen) for myocyte area. Myocardial apoptosis was evaluated using the DeadEnd Fluorometric TUNEL System (Promega) as 1, 28 previously described. Sections were also blocked and then incubated with antibodies against CD11b/ITAM (Epitomics), CD11c (ebiosciences), F/8 (ebiosciences), CD169 (R&D Systems), high mobility group box-1 (HMGB1, IBL International), and CD.2-PE (BD Biosciences), and secondary antibodies conjugated with Alexa 88,, or 67 (Invitrogen). Nuclei were stained with DAPI. Myocyte area and cardiac fibrosis were quantified from -6 high-power fields per section in the remote noninfarcted area of the heart using Image J and Metamorph software, respectively. In the spleen, CD11b positive cells in the subcapsular red pulp were counted in 3- high power fields per section. Data for each group were calculated from 12-1 sections and - mice from each group. The measurements and calculations were conducted in a blinded manner. Confocal microscopy was carried out on an LSM71 microscope (Zeiss) and Z-stack images were acquired according to standard protocols. Downloaded from by guest on June 3, 218 Cytometric bead array (CBA) immunoassay for serum cytokines. Peripheral blood was allowed to clot at RT for 2 h. After centrifugation at 2, rpm for 1 min, serum was collected and stored at 2 C until assays were performed. Serum concentrations of TNF, monocyte chemoattractant protein (MCP)-1, interferon (IFN)-γ, interleukin (IL)-6, IL-1, and IL-12p7 were measured simultaneously using a CBA Mouse Inflammation Kit (BD Biosciences). Briefly, µl of chemokine capture bead mixture was incubated with µl of either recombinant standard or sample and µl of PE-conjugated detection antibody for 2 h at RT. The mixture was washed to remove unbound PE detection reagent before data acquisition on a BD LSRII flow cytometer. Analysis was performed using FCAP Array software. Splenocyte gene expression by quantitative real-time PCR. Mononuclear splenocytes were cultured in serum free DMEM media for 2 h after isolation (2-3 X 1 6 cells/well). The cells were then collected and stored at -8 C in TRIzol reagent (Invitrogen) for subsequent RNA isolation. cdna synthesis and quantitative real-time PCR were performed as described previously. 3, 16, 28 Relative levels of mrna transcripts for TNF, IL-1β, IL-2, IL-, IL-, IL-6, IL-1, IL-13, IFN-γ, inducible nitric oxide synthase (inos), C-C chemokine receptor type 2 (CCR2), CCR, chemokine (C-C motif) ligand (CCL), chemokine (C-X-C motif) receptor 3 (CXCR3), CX3C chemokine receptor 1 (CX3CR1), toll-like receptor (TLR)1, TLR7, TLR9, TLR12, transforming growth factor (TGF)-β, HMGB1, S1A8, S1A9, and galectin-3 were determined using forward and reverse primer pairs detailed in Supplemental Table I, and normalized to 18s rrna expression using the ΔΔC T comparative method. 3 Survival splenectomy and splenocyte adoptive transfer. In a separate sub-group, male (CD.2) C7BL/6J mice (n = 36) underwent coronary ligation or sham operation. Eight weeks later, survival splenectomy (or, in some mice, sham abdominal surgery) was performed using standard surgical techniques. 29 Briefly, mice were anesthetized with tribromoethanol (.2 mg/g IP) and anesthesia was maintained with 1% isoflurane as needed. After sterilizing the surgical area, a ~1 cm incision was made in the left subcostal region, and the peritoneum was opened to exteriorize the spleen. The spleen was retracted away from the pancreatic tail, the splenic bundle was ligated at the hilum using 8. prolene ligature, and then the spleen was removed intact. For sham operations, the splenic bundle and spleen were left intact. The procedure required ~1 1 minutes to complete; totality of splenectomy was ensured by close examination at the time of operation and confirmed during postmortem examination. Splenocytes from sham-operated and HF mice were used for the adoptive transfer studies described below. Splenectomized mice (and sham abdominal surgery controls) were subsequently followed for 8 more weeks with serial echocardiography. DOI: /CIRCRESAHA
7 For adoptive transfer studies, splenocytes were prepared as described previously. 3 Briefly, single cell suspensions were prepared from spleens isolated from sham-operated (n=8) and HF mice (n=7). The erythrocytes were lysed with RBC lysis buffer (ebioscience, CA). Single cell suspensions were then layered on Ficoll-Paque (GE Healthcare) gradient and centrifuged at 2g for 2 min to separate mononuclear cells and exclude granulocytes and residual erythrocytes. Mononuclear cells were collected, washed three times with sterile PBS, and resuspended in sterile PBS. Cells were then pooled from either sham or HF mice and resuspended in ml PBS. Mononuclear cells were injected (~17 X 1 6 cells in 1 μl PBS per animal via tail vein) into naïve CD.1 C7BL/6J mice (7 mice receiving sham splenocytes and 7 mice receiving HF splenocytes). Downloaded from by guest on June 3, 218 As an additional control for adoptive transfer, we also isolated splenocytes from CD.2 C7BL/6J mice (male, 6-8 weeks) treated with either lipopolysaccharide (LPS) to induce systemic inflammation or PBS control. LPS (. mg/kg) or PBS (.2 ml) was injected i.p. daily for two consecutive days (n = per group). Spleens were aseptically harvested 2 h after the last i.p. dose, and the mononuclear cell population was isolated and injected via tail vein into naïve CD.1 C7BL/6 recipient mice (n = 7 per group receiving either LPS-splenocytes or PBS-splenocytes) as above. Statistical analysis. Continuous data are summarized as mean ± standard deviation (SD). Statistical comparisons were performed using the unpaired student s t-test when comparing two groups. For comparisons of more than two groups, two-way ANOVA was used with Bonferroni post-test to adjust for multiple comparisons. A p value of <. was considered statistically significant. RESULTS Profound structural remodeling in the heart 8 weeks after coronary ligation. Four weeks after coronary ligation, mice exhibit significant pathological cardiac remodeling and hemodynamic signs of HF. 3, 1 Here, we evaluated mice at 8 w to ensure the establishment of chronic ischemic HF. Typical M-mode echocardiograms and ventricular sections from sham-operated and HF mice (Supplemental [S] Figure IA), and histological sections of remote (non-infarcted) myocardium stained with Masson s trichrome and WGA (Figure SIB) demonstrated substantial myocardial scarring, LV dilatation, LV systolic dysfunction, and increased fibrosis and myocyte size in HF mice. Group data (Figure SIC) confirmed significantly increased LV end-diastolic and end-systolic volume (EDV and ESV), reduced LV ejection fraction (EF), and increased (~2-fold) remote zone fibrosis and myocyte area. Normalized heart and lung weights were also increased. These findings indicated marked LV remodeling and pulmonary edema in HF mice. Activated M1 polarized macrophages infiltrate the failing heart. Activated macrophages were defined as cells with dual expression of CD11b (Mac-1) and F/8 surface markers. 2, 21 As seen in Figure 1A, as compared with sham, failing hearts exhibited a significant increase in both overall mononuclear cells and infiltrating activated macrophages (19.2 ± 1.9 vs. 1.9 ± 2.9%; p=.33). Augmented tissue macrophages were further confirmed by Mac-1 immunostaining (Figure 1B), which revealed a ~-fold increase in macrophages in failing hearts over sham (p <.1). Lin2 negativity (see Methods) and CD26 expression were used to sub-classify macrophages as either M1 (pro-inflammatory, Lin2 CD11b + F/8 + CD26 ) or M2 (anti-inflammatory, Lin2 CD11b + F/8 + CD26 + ) by flow cytometry. As seen in Figure 1C, both sham and failing hearts exhibited a greater abundance of tissue M2 macrophages in relation to M1 macrophages, consistent with DOI: /CIRCRESAHA
8 prior studies. 31 M1 macrophages significantly increased in the failing heart over sham (6.2 ±.6 vs. 3.8 ±.7%, p=.19), whereas there was no change in M2 macrophages (p=.28), thus favoring a proinflammatory milieu. In support of this, HF mice (compared with sham) exhibited significant (or near significant) elevations of circulating pro-inflammatory IFN-γ, IL-6, and MCP-1 in serum (Figure SII). No differences among groups were observed for serum levels of IL12p7, TNFα, and IL-1. Classical and plasmacytoid DCs increase in the failing heart. Downloaded from by guest on June 3, 218 CD11c + DCs are antigen-presenting cells (APCs) categorized as classical and non-classical DCs. 22 cdcs include migratory DCs (present in most organs, including the heart) that activate immune responses after detection of danger signals, and lymphoid tissue-resident DCs that do not migrate but collect and process antigens in one lymphoid organ throughout their lifespan. 22 Non-classical DCs include pdcs that are present in an immature form at steady-state, but mature with dendritic form and costimulatory molecule expression upon inflammatory activation. As seen in Figure 2A, CD11c 6, 22, 26 immunostaining revealed distinctly increased tissue DC infiltration in the interstitium of the failing heart (border and remote zone) as compared with sham. DC subsets in the heart were quantified by flow cytometry using Lin1 negativity (see Methods), with cdcs identified as Lin1 CD11c + B22 and pdcs as Lin1 CD11c +/low B , 2 pdcs were further classified as CD86 low and CD86 hi, to indicate immature and mature pdcs, respectively. 26 Both DC populations increased in the failing heart, cdcs by ~1. fold (p=.23) and pdcs by ~.3 fold (p=.12) over sham (Figure 2B). Moreover, pdcs over a range of maturation stages were present in the failing heart. As seen in Figure 2C, the number of mature CD86 hi and immature CD86 low pdcs in sham hearts was very low. However, both pdc populations were markedly increased in HF hearts as compared with sham (~9-fold higher CD86 hi pdcs, p=.9; ~22-fold higher CD86 low pdcs, p <.1). Splenic remodeling is prominent in chronic HF. Masson s trichrome stains of the spleen revealed striking alterations of the splenic architecture in HF. As seen under low power in Figure 3A, the HF spleen exhibited greater numbers of white pulp follicles (primarily composed of lymphocytes), with greater variability and irregularities of follicular size and shape, and greater numbers of large germinal centers as compared with spleens from sham-operated mice. Higher power views revealed a striking increase in size of the marginal zone surrounding the white pulp, a site that plays an important role in antigen screening and processing. 32 The changes in tissue architecture suggested immune activation and heightened antigen processing in the spleen. 32 Additionally, there was reduced mononuclear cell abundance in the red pulp, especially the subcapsular red pulp (SRP), in HF spleens. To quantify monocytes/macrophages and DCs in the spleen, flow cytometry and immunostaining were performed. As seen in Figure 3B, CD11b/Mac-1 and F/8 immunostains revealed ~3.-fold fewer activated monocytes in the spleens of HF mice as compared with sham, primarily in the red pulp and SRP. This was confirmed by flow cytometry of the splenic mononuclear cell population (Figure 3C), which revealed a significant (p =.11) ~2-fold reduction in CD11b + F/8 + cells in HF spleens. These changes were primarily related to pro-inflammatory (CD11b + F/8 + Gr-1 hi ) monocytes without significant change in anti-inflammatory (CD11b + F/8 + Gr-1 low ) monocytes. In contrast to activated monocytes, lymphoid tissue-resident DCs populations were markedly increased in the spleens of HF mice. As seen in Figures A-B, both CD11c + B22 cdc and CD11c +/low B22 + pdc populations were augmented in HF spleens. Increased splenic pdcs in HF were also observed using Siglec-H as a pdc marker (Figure SIII). In lymphoid organs, CD8 + and CD8 cdcs differ in their efficiency for antigen cross-presentation to T-cells. 6, 22, 33 As seen in Figure C, there was a significant ~1.8-fold increase in both CD8 + and CD8 cdcs in spleens from HF mice as compared with sham, together with significantly increased numbers of DOI: /CIRCRESAHA
9 CD + and CD8 + T-cells (Figure SIV). Importantly, while both CD8 + and CD8 cdcs prime T-cell responses, CD8 + DCs have a greater propensity to stimulate CD8 + cytotoxic T-cells 6, 33 and Th-1 CD + helper T-cells 3 that would promote a pro-inflammatory milieu. In addition to increased splenic DC abundance in HF, CD11c immunostaining revealed a striking spatial redistribution of DCs (Figure D). In sham-operated mice, splenic CD11c + DCs were primarily seen in the marginal zone. In contrast, in HF spleens, DCs were localized within the white pulp and often in germinal centers, suggesting a critical role for DCs in the heightened antigen processing occurring in HF. Augmented circulating cdcs and pro-inflammatory monocytes in HF. Downloaded from by guest on June 3, 218 HF mice exhibited a marked ~2-fold increase in circulating CD11b + F/8 + activated monocytes (p=.1) as compared with sham (Figure A). These changes were again related to pro-inflammatory monocytes, analogous to the polarity of tissue macrophages in the failing heart (Figure 1), and consistent with the systemic elaboration of pro-inflammatory mediators such as MCP-1 (Figure SII). Notably, circulating anti-inflammatory monocytes decreased substantially in HF mice. cdcs and pdcs were also detected in the circulation of sham and HF mice (Figure B). Circulating CD8 + and CD8 cdcs were both increased ~2-3-fold in HF mice as compared with sham, while circulating pdcs were similar between the groups. In contrast, bone marrow from HF mice exhibited significantly (~1.-fold) increased pdcs, but no change in cdcs or activated monocytes when compared with sham bone marrow (Figure SV). The spleen regulates the progression of pathological LV remodeling in chronic HF. The above data indicated global pro-inflammatory alterations of the APC network in HF. As the largest lymphoid organ in the body, the spleen would be expected to be the critical lynchpin for their pathogenesis and, by analogy, for the progression of cardiac remodeling. To examine this as a proof-ofprinciple, two approaches were used. First, splenectomy (or sham splenectomy) was performed in mice with HF, 8 w post-coronary ligation, and the effects on LV remodeling were evaluated 8 w later (Figure 6A). Splenectomy in sham-operated mice did not induce any changes in LV size or function. Figure 6B depicts representative M-mode echocardiograms from one mouse at baseline, 8 w post-ligation (HF), and 16 w post-ligation/8 w post-splenectomy (HF/splenectomy). LV enlargement and dysfunction significantly improved post-splenectomy. Group data (Figure 6C) confirmed that splenectomy in HF, over a period of 8 weeks, induced significant (p <.) reductions in EDV and ESV and improvement (22% relative change, 8% absolute change) in LVEF, indicating reversal of pathological remodeling, whereas HF mice with sham abdominal surgery at 8 w continued to show progressive LV remodeling and functional decline. Moreover, cardiac CD11b and CD11c immunostaining revealed a profound ~3-fold reduction in tissue macrophages and DCs in the failing heart after splenectomy (Figure 6D). Also, as illustrated in Figure SVI, splenectomy did not reduce serum levels of TNF, MCP-1, IL-6, IL12p7, and IFN-γ in HF mice (IL-6 levels were actually mildly increased after splenectomy). Taken together, this suggests that the improved LV remodeling following splenectomy was not simply related to changes in circulating cytokines, but rather to loss of splenic mobilization and subsequent diminished tissue infiltration of inflammatory cells. Second, we performed adoptive transfer of mononuclear splenocytes isolated from either shamoperated or HF (CD.2) mice into naïve (CD.1) syngeneic C7Bl/6 mice and serially evaluated LV remodeling over an 8-week period (Figure 7A). Flow cytometry revealed persistence of donor splenocytes in the peripheral blood of recipients at a level of ~1% by one week, and <1-2% by 2 weeks post-transfer (Figure SVII). Mice that received splenocytes from sham-operated mice exhibited no appreciable changes in LV size or ejection fraction over an 8-week period after cell transfer (Figure 7B-7C). However, mice that received splenocytes from HF mice had significant (p <.) LV dilatation (~3% and ~6% increase in EDV and ESV, respectively) and LVEF decline evident by 8 weeks. Analogous to HF mice with splenectomy, serum levels of TNF, MCP-1, IL-6, IL12p7, and IFN-γ were similar between mice DOI: /CIRCRESAHA
10 receiving HF-splenocytes and sham-splenocytes (Figure SVIII), suggesting that the remodeling responses were not secondary to high systemic cytokines. Moreover, to exclude a non-specific response, we performed parallel experiments using splenocytes from mice given i.p. LPS (or PBS control) for 2 days. While this LPS regimen significantly augmented circulating pro-inflammatory CD11b + F/8 + Gr-1 + cells as compared with PBS-treated mice (Figure SIXA), adoptive transfer of LPS-splenocytes (or PBSsplenocytes) had no discernible effects on LV size or systolic function (Figure SIXB, Figure 7D), suggesting that the HF-splenocyte response was specific to the HF disease state itself rather than nonspecific inflammation. HF splenocytes express inflammatory mediators and alarmins and home to the heart to induce pathological injury. Downloaded from by guest on June 3, 218 Gravimetric analysis (Figure 7E) revealed increased heart and lung weights in HF-splenocyte recipient mice, as compared with sham-splenocyte recipients, indicative of cardiac hypertrophy and lung edema. Peripheral blood flow cytometry 8 w post-transfer also revealed augmented activated CD.1 monocytes, indicating reproduction of chronic inflammation in recipients (Figure 7F). Moreover, as compared with sham-splenocyte recipient mice, the spleens of HF-splenocyte mice were hypertrophied and exhibited histological changes indicative of heightened antigen processing similar to that observed in mice with HF (Figure 7G). Histological analysis of HF-splenocyte recipient hearts revealed augmented interstitial fibrosis as compared with sham-splenocyte recipient hearts (Figure 8A); fibrotic foci often coincided with retained CD.2 + donor cells. These hearts also exhibited a ~1-fold increase in cardiomyocyte apoptosis (Figure 8B), suggesting HF-splenocyte-induced cell loss as a potential trigger for myocardial fibrosis. To confirm cardiac homing of HF splenocytes, we performed immunostaining for CD169, a marker for splenic metallophilic marginal zone macrophages. 3 HF-splenocyte recipient hearts, but not sham-splenocyte recipient hearts, harbored multiple foci of CD.2 + CD169 + cells in the perivascular and interstitial regions (Figure 8C). We also observed CD169 + macrophages in the border zone of failing hearts 8 w after coronary ligation, but not in sham-operated hearts (data not shown). Hence, mononuclear splenocytes activated in HF home to the heart to induce tissue injury and remodeling, and retain memory sufficient to induce similar inflammation and long-term pathological remodeling in otherwise naïve hearts. We examined inflammatory gene expression in splenocytes and levels of splenic alarmins as potential mechanisms for immune cell-mediated injury. As illustrated in Figure SX, as compared to sham, splenocytes from HF mice exhibited significantly increased expression of a number of pro-inflammatory cytokines/mediators (TNF, IL-2, IL-, IL-6, inos, IFN-γ), and chemokine receptors/ligands (CCR2, CX3CR1, CCL, CXCR3) and pattern recognition receptors (PRRs: TLR-7, TLR-9, TLR-12) of signal importance in the activation of DCs. 36 In contrast, of the anti-inflammatory mediators examined (IL-, IL- 1, IL-13, TGF-β), only IL-1 was upregulated. Hence, HF-splenocytes are potently activated and serve as sources of inflammatory mediators that can initiate and/or amplify tissue injury. PRR upregulation, cell homing to the heart, and the capacity to reproduce systemic inflammation and cardiac injury and remodeling upon HF-splenocyte adoptive transfer suggests prior priming of the transferred immune cells by cardiac antigens. Potential candidates for the latter are damage-associated molecular patterns (DAMPs) or alarmins, TLR ligands released upon cardiac injury that can activate innate immune cells both locally and globally. 37, 38 As shown in Figure SXIA, immunostaining of HF spleens revealed robustly increased levels of both intracellular and extracellular HMGB1, a prototypical alarmin, in the white pulp, marginal zone, and SRP, suggesting ongoing HMGB1 antigen presentation to T-cells. HF splenocytes themselves further exhibited significant upregulation of several alarmins, including HMGB1, S1A8, S1A9, and galectin-3 (Figure SXIB) that can amplify the inflammatory response. 37 Moreover, adoptively transferred HF splenocytes were also intermittently observed to robustly express HMGB1 in recipient hearts (Figure SXIC). Taken in totality, the adoptive transfer studies provide DOI: /CIRCRESAHA
11 direct evidence for activated splenocytes as disease mediators in HF, as part of a pathophysiologically relevant cardio-splenic axis that is both necessary and sufficient for the progression of LV remodeling. DISCUSSION Downloaded from by guest on June 3, 218 There are several novel findings of this study. First, there are profound alterations of the mononuclear phagocyte network in established ischemic HF that encompass the failing heart, spleen, peripheral blood, and bone marrow and form the foundation for chronic inflammation in this disease. These were comprised of: 1) increased pro-inflammatory macrophages and monocytes in the failing heart and peripheral blood, together with a reduction in the activated monocyte reservoir in the spleen, 2) augmented cdcs and pdcs in both the failing heart and spleen, and increased cdcs and pdcs in the blood and bone marrow, respectively, and 3) increased levels of both CD8 + and CD8 cdc subtypes in the spleen and blood. Augmented circulating levels of IFN-γ, MCP-1, and IL-6 accompanied these immune cell profiles. Second, there is profound splenic remodeling in HF indicative of ongoing, heightened antigen processing. This occurred in concert with robust expression of alarmins and proinflammatory mediators, and augmented splenic CD + and CD8 + T-cell populations, consistent with heightened T cell activation and differentiation secondary to antigen-mhc presentation by DCs. Third, the spleen has an obligatory role in the progression of cardiac remodeling and inflammation in chronic HF, as splenectomy reversed remodeling and attenuated tissue macrophage and DC infiltration. Fourth, HFderived mononuclear splenocytes, not otherwise activated ex vivo, home to the heart and induce long-term cardiac remodeling and dysfunction, systemic inflammation, and splenic remodeling akin to HF in previously normal mice. These data indicate that mononuclear splenocytes in chronic HF: 1) are highly activated, potentially in response to cardiac-derived alarmins, and traffic to the heart to induce immune cell-mediated injury, and that 2) this capacity is retained upon adoptive transfer into naïve animals. Taken together, we conclude that activation of mononuclear phagocytes is indispensable for the progression of pathological remodeling and that the splenic microenvironment in HF plays a critical role in this process. Altered inflammatory cell network in HF. Recent studies have established a critical role for monocytes/macrophages 11-1 and DCs 1 in tissue healing early after myocardial infarction. Nahrendorf, Swirski, and co-workers 12-1 have demonstrated that monocyte recruitment to the heart following acute infarction is dynamic and high, and is dependent on extramedullary splenic hematopoiesis and mobilization of splenic monocytes (with accompanying loss of the splenic monocyte reservoir). This recruitment followed two phases an initial infiltration of proinflammatory (Ly-6C hi ) monocytes (days 1- post- infarction) that promote tissue digestion, followed by reparative (Ly-6C low ) monocytes that resolve inflammation and promote tissue healing. Wang et al 39 have shown an analogous pro-inflammatory state in the bone marrow (increased CD11b +, Gr-1 + and/or Ly6C high cells) early after infarction that diminishes the reparative efficacy of bone marrow-derived progenitors. This impaired efficacy resolved after the first week, consistent with an acute, time-limited process. Extending these prior studies, our data reveal that chronic ischemic HF is characterized by Gr-1 hi monocytes in the blood and CD26 macrophages in the failing heart well after the formation of healed infarct scar. Hence, infiltrating monocytes/macrophages in the chronically failing heart reestablish a proinflammatory and injurious phenotype that promotes ongoing remote (and border) zone tissue 7, 8, 21 remodeling. Importantly, the splenic pro-inflammatory monocyte reservoir remains depleted in chronic HF, especially in the SRP, similar to that observed early after infarction. 1 Unlike in the spleen, and contrary to the early post-infarct period, 39 we observed no changes in the activated monocyte population in the bone marrow in HF. Therefore, our results suggest that in chronic HF, there is persistent splenic DOI: /CIRCRESAHA
12 mobilization of pro-inflammatory monocytes to the heart resulting in tissue infiltration by M1 macrophages that promote ongoing tissue remodeling. Persistently elevated serum levels of MCP-1 (CCR2), a C-C chemokine that plays a critical role in the trafficking and tissue recruitment of proinflammatory monocytes after myocardial infarction, 7, 11, provide further support for this scenario. These data are also consistent with human studies demonstrating increased numbers of activated monocytes in the circulation of patients with advanced HF. 1 Downloaded from by guest on June 3, 218 DCs are specialized for the processing and presentation of antigens to T cells, upon upregulation of MHC and co-stimulatory molecules (e.g., CD86). Depending on the DC functional subset, activation state, and maturation stage 2 a variety of immune responses can be engendered, including helper or cytotoxic T-cell activation as well as T-cell immune tolerance. 6, 22, 26, 33, 3 DCs infiltrate into the heart early after infarction, peaking at 7 days, and are immunoprotective, as selective DC ablation immediately after infarction worsened remodeling, enhanced inflammatory cytokines, and increased cardiac infiltration of pro-inflammatory monocytes. 1 Our results demonstrate that in chronic HF, well after the infarct healing period, there is a marked increase in cdc and pdc populations in the spleen and in the failing heart itself, together with increased circulating cdcs and augmented bone marrow pdcs. Moreover, contrary to early post-infarction, this increase in DCs is accompanied by the mobilization and infiltration of pro-inflammatory monocytes and macrophages to the failing heart, suggesting a chronic and inappropriately activated deleterious immune response. Notably, while cdcs are potent activators of both CD8 and CD T-cells that can promote tissue damage, 6 mature pdcs are poorly immunostimulatory and favor the formation of regulatory T cells and self-tolerance, 26, which can mitigate the same. In our study, the failing heart exhibited augmented levels of both immature and mature pdcs (Figure 2), presumably reflecting varying capacities for the induction of self-tolerance in response to tissue injury. Hence, the balance between cdcs and pdcs, as well as the relative abundance of maturing pdcs, may be of considerable importance in the maintenance, or alternatively resolution, of chronic inflammation in the failing heart. Increased CD + and CD8 + T-cells in the spleen accompanied the augmented levels of DCs, suggesting ongoing processing and presentation of antigens by DCs to T-cells, thereby leading to T-cell activation and differentiation, and subsequent generation of effector and/or memory immune responses. While the specific antigens responsible for triggering immune cell activation are not definitively answered by our study, we found marked upregulation of chemokine and TLR pattern-recognition receptors known to be differentially expressed on DCs 36 in HF splenocytes, and augmented extra- and intra-cellular levels of the HMGB1 in HF spleens (Figures SX-SXI). These findings suggest that the ongoing release, collection, and processing of DAMPs, presumably derived from injured myocardium, contribute importantly to sustained activation of immune cells in chronic HF. Among the cdc subsets, CD8 + DCs are considered more robust stimulators of CD8 + cytotoxic T-lymphocytes 6, 33 and Th-1 CD + helper T-cell responses; 3 they have, however, also been implicated in tolerance induction. 6, 3 Dissecting the specific antigen- and cell-mediated determinants of immune activation versus tolerance in HF would be critical for guiding potential therapeutic immunomodulation. In this regard, Eriksson et al 6 previously demonstrated that bone marrow-derived DCs self-loaded with α-myosin heavy chain peptide, and activated with lipopolysaccharide and a CD stimulatory antibody, induced CD + T-cell-mediated myocarditis and subsequent HF. We propose that chronic remodeling in HF following myocardial infarction is also driven, at least in part, by sustained DC activation in response to cardiac antigens. Indeed, HF is generally considered a Th1 activation state this would necessarily implicate DCs, regulators of CD + T helper-cell polarization, as critical mediators of the inflammatory response. A newly uncovered robust cardiosplenic axis in chronic HF. Global alterations of the mononuclear phagocyte network suggest significant underlying changes in the spleen, as two important functions of the spleen are to filter blood and remove and/or initiate DOI: /CIRCRESAHA
13 immune responses to circulating antigens. 32 Our results demonstrate striking changes in splenic architecture (more follicles, large germinal centers and prominent marginal zones) and cell populations (augmented CD8 + /CD8 cdcs and pdcc, shift in the spatial distribution of DCs to the white pulp germinal centers, augmented CD + and CD8 + T-cells) that signify amplified antigen processing in HF. Also, the spleen exhibited depletion of SRP monocytes, connoting a reduced ability of the spleen in HF to remove (by phagocytosis) senescent erythrocytes and circulating particulates. 32 Hence, profound splenic remodeling occurs in chronic HF that supports immune system activation on one hand and diminished circulating antigenic clearance on the other. Downloaded from by guest on June 3, 218 To evaluate the importance of these splenic abnormalities in HF, we selected two approaches to establish both the necessity and the sufficiency of the spleen for pathological cardiac remodeling. To establish necessity, we performed splenectomy in mice with chronic HF and evaluated LV structure and function 8 weeks later. Splenectomy both stabilized and actually reversed cardiac remodeling, with longterm improvement in LVEF and reduction in chamber size, and suppression of CD11b + and CD11c + cell infiltration in the failing heart. Importantly, splenectomy did not significantly diminish circulating proinflammatory cytokines, suggesting primary effects referable to the cardiac recruitment of splenic immune cell populations rather than global levels of inflammatory mediators. Hence, the spleen exacerbates disease progression and cardiac inflammation in chronic HF. These results are counter to the beneficial role of the spleen early after infarction. Splenectomy at the time of infarction or 3 d later worsened scar formation and subsequent LV remodeling, 12 an effect thought to be due to wound healing responses engendered by the splenic monocyte reservoir. In contrast to this early effect, our data establish that in chronic HF, the spleen, with sustained depletion of SRP monocytes and increased antigenic processing, assumes a much more pro-inflammatory role and promotes mobilization of immune cell populations to the heart that exacerbate detrimental LV remodeling. To establish sufficiency of the spleen, we performed adoptive transfer of mononuclear splenocytes into otherwise normal naïve mice. Importantly, our study design did not include additional exogenous mitogenic stimulation of splenocytes as has been reported previously in a study in rats. 7 Transfer of splenocytes from mice with HF, but not from sham-operated mice, recapitulated multiple facets of chronic pathological remodeling in naïve mice. Notably, there were no discernible early effects on LV function, indicating a low likelihood of an acute myocarditis after transfer. While donor splenocytes disappeared from the recipient mouse circulation within 2 weeks, pathological changes were manifested in recipient hearts much later, at 8 weeks, and included LV dilatation, hypertrophy (by gravimetric analysis), systolic dysfunction, and myocardial apoptosis and fibrosis. Moreover, we found evidence of donor splenocytes homing to recipient hearts, localized primarily at sites of interstitial fibrosis and in perivascular regions. Intriguingly, systemic changes were also evident including splenic hypertrophy with architectural changes similar to those seen in HF, and increased levels of circulating activated monocytes consistent with an inflammatory state. This effect was specific to splenocytes from HF, as there were no substantial changes in systemic cytokine levels, and as adoptive transfer of splenocytes from mice treated with LPS had no discernible cardiac effects. Moreover, the high level of expression of cytokine/chemokine mediators, TLRs, and alarmins in HF splenocytes indicates that these cells were potently activated, and suggested that the recapitulation of systemic inflammation and tissue injury/remodeling in naïve hearts occurred as a result of prior priming of splenic immune cells to cardiac antigens in HF. Hence, taken together, the results suggest that the spleen is a fundamental mediator of chronic inflammation and remodeling in HF, that activated splenocytes (potentially in response to cardiac alarmins) home to the heart to induce tissue injury, and that HF splenocytes retain memory sufficient for recreating the injurious immune responses in otherwise normal animals. These data suggest that remodeling progression in chronic HF is an immune cell-mediated phenomenon, and that targeting specific mononuclear cell populations within the spleen and heart (e.g., monocyte/macrophages and DCs), or the specific antigens responsible for their activation, DOI: /CIRCRESAHA
14 may comprise a more feasible approach to therapeutic immunomodulation in this disease. The relative importance of each specific cell type in this disease process, and the identity and importance of specific antigens and alarmins, will require further study. In summary, we have demonstrated profound remodeling of the mononuclear phagocyte network and the spleen in HF that forms the basis for chronic inflammation. These changes, which are consistent with ongoing, enhanced antigen processing in the spleen and sustained inflammatory/immune cellmediated injury in the failing heart, are of pathophysiological importance for the progression of HF, independent of other factors. Hence, we propose the novel paradigm that mononuclear phagocytes are obligatory disease mediators in HF, and that progression of pathological cardiac remodeling is dependent, at least in part, on autoimmune injury in the heart induced by these cell populations. Downloaded from by guest on June 3, 218 SOURCES OF FUNDING This work was supported by a VA Merit Award (S.D.P.), NIH grants HL-7882 and HL-991 (S.D.P.), and an AHA SDG award 836N (T.H.). ACKNOWLEDGMENT The authors gratefully acknowledge James F. George, PhD and his laboratory for their assistance in performing the splenocyte adoptive transfer studies. DISCLOSURES Conflicts of interest: none. REFERENCES 1. Mann DL. Inflammatory mediators and the failing heart: Past, present, and the foreseeable future. Circ Res. 22;91: Bozkurt B, Mann DL, Deswal A. Biomarkers of inflammation in heart failure. Heart Fail Rev. 21;1: Hamid T, Gu Y, Ortines RV, Bhattacharya C, Wang G, Xuan YT, Prabhu SD. Divergent tumor necrosis factor receptor-related remodeling responses in heart failure: Role of nuclear factor-κb and inflammatory activation. Circulation. 29;119: Braunwald E. Biomarkers in heart failure. N Engl J Med. 28;38: Prabhu SD. Cytokine-induced modulation of cardiac function. Circ Res. 2;9: Kushwah R, Hu J. Complexity of dendritic cell subsets and their function in the host immune system. Immunology. 211;133: Ingersoll MA, Platt AM, Potteaux S, Randolph GJ. Monocyte trafficking in acute and chronic inflammation. Trends Immunol. 211;32: Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat Immunol. 21;11: Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K. Development of monocytes, macrophages, and dendritic cells. Science. 21;327: Anzai A, Anzai T, Nagai S, Maekawa Y, Naito K, Kaneko H, Sugano Y, Takahashi T, Abe H, Mochizuki S, Sano M, Yoshikawa T, Okada Y, Koyasu S, Ogawa S, Fukuda K. Regulatory role of dendritic cells in postinfarction healing and left ventricular remodeling. Circulation. 212;12: Frangogiannis NG. Regulation of the inflammatory response in cardiac repair. Circ Res. 212;11: DOI: /CIRCRESAHA
15 Downloaded from by guest on June 3, Leuschner F, Rauch PJ, Ueno T, Gorbatov R, Marinelli B, Lee WW, Dutta P, Wei Y, Robbins C, Iwamoto Y, Sena B, Chudnovskiy A, Panizzi P, Keliher E, Higgins JM, Libby P, Moskowitz MA, Pittet MJ, Swirski FK, Weissleder R, Nahrendorf M. Rapid monocyte kinetics in acute myocardial infarction are sustained by extramedullary monocytopoiesis. J Exp Med. 212;29: Nahrendorf M, Pittet MJ, Swirski FK. Monocytes: Protagonists of infarct inflammation and repair after myocardial infarction. Circulation. 21;121: Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science. 29;32: Wang G, Hamid T, Keith RJ, Zhou G, Partridge CR, Xiang X, Kingery JR, Lewis RK, Li Q, Rokosh DG, Ford R, Spinale FG, Riggs DW, Srivastava S, Bhatnagar A, Bolli R, Prabhu SD. Cardioprotective and antiapoptotic effects of heme oxygenase-1 in the failing heart. Circulation. 21;121: Hamid T, Guo SZ, Kingery JR, Xiang X, Dawn B, Prabhu SD. Cardiomyocyte NF-κB p6 promotes adverse remodelling, apoptosis, and endoplasmic reticulum stress in heart failure. Cardiovasc Res. 211;89: Lavelle GC, Sturman L, Hadlow WJ. Isolation from mouse spleen of cell populations with high specific infectivity for scrapie virus. Infect Immun. 1972;: Austyn JM, Hankins DF, Larsen CP, Morris PJ, Rao AS, Roake JA. Isolation and characterization of dendritic cells from mouse heart and kidney. J Immunol. 199;12: Fleming TJ, Fleming ML, Malek TR. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C mab to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J Immunol. 1993;11: Robbins CS, Swirski FK. The multiple roles of monocyte subsets in steady state and inflammation. Cell Mol Life Sci. 21;67: Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2;: Shortman K, Naik SH. Steady-state and inflammatory dendritic-cell development. Nat Rev Immunol. 27;7: Hadeiba H, Lahl K, Edalati A, Oderup C, Habtezion A, Pachynski R, Nguyen L, Ghodsi A, Adler S, Butcher EC. Plasmacytoid dendritic cells transport peripheral antigens to the thymus to promote central tolerance. Immunity. 212;36:38-2. Hochrein H, O'Keeffe M, Wagner H. Human and mouse plasmacytoid dendritic cells. Human Immunol. 22;63: Blasius AL, Colonna M. Sampling and signaling in plasmacytoid dendritic cells: The potential roles of Siglec-H. Trends Immunol. 26;27: Ito T, Yang M, Wang YH, Lande R, Gregorio J, Perng OA, Qin XF, Liu YJ, Gilliet M. Plasmacytoid dendritic cells prime IL-1-producing T-regulatory cells by inducible costimulator ligand. J Exp Med. 27;2: Kitchen SG, Jones NR, LaForge S, Whitmire JK, Vu BA, Galic Z, Brooks DG, Brown SJ, Kitchen CM, Zack JA. CD on CD8+ T cells directly enhances effector function and is a target for HIV infection. Proc Natl Acad Sci USA. 2;11: Ismahil MA, Hamid T, Haberzettl P, Gu Y, Chandrasekar B, Srivastava S, Bhatnagar A, Prabhu SD. Chronic oral exposure to the aldehyde pollutant acrolein induces dilated cardiomyopathy. Am J Physiol Heart Circ Physiol. 211;31:H Wang HX, Yi SQ, Li J, Terayama H, Naito M, Hirai S, Qu N, Yi N, Itoh M. Effects of splenectomy on spontaneously chronic pancreatitis in aly/aly mice. Clin Dev Immunol. 21;21:6189 DOI: /CIRCRESAHA
16 Downloaded from by guest on June 3, Kodama M, Matsumoto Y, Fujiwara M. In vivo lymphocyte-mediated myocardial injuries demonstrated by adoptive transfer of experimental autoimmune myocarditis. Circulation. 1992;8: Pinto AR, Paolicelli R, Salimova E, Gospocic J, Slonimsky E, Bilbao-Cortes D, Godwin JW, Rosenthal NA. An abundant tissue macrophage population in the adult murine heart with a distinct alternatively-activated macrophage profile. PloS One. 212;7:e Cesta MF. Normal structure, function, and histology of the spleen. Toxicol Pathol. 26;3: Joffre OP, Segura E, Savina A, Amigorena S. Cross-presentation by dendritic cells. Nat Rev Immunol. 212;12: Maldonado-Lopez R, De Smedt T, Michel P, Godfroid J, Pajak B, Heirman C, Thielemans K, Leo O, Urbain J, Moser M. CD8alpha+ and CD8alpha- subclasses of dendritic cells direct the development of distinct T-helper cells in vivo. J Exp Med. 1999;189: den Haan JM, Kraal G. Innate immune functions of macrophage subpopulations in the spleen. J Innate Immun. 212;: Kaisho T. Pathogen sensors and chemokine receptors in dendritic cell subsets. Vaccine. 212;3: Chan JK, Roth J, Oppenheim JJ, Tracey KJ, Vogl T, Feldmann M, Horwood N, Nanchahal J. Alarmins: Awaiting a clinical response. J Clin Invest. 212;122: Timmers L, Pasterkamp G, de Hoog VC, Arslan F, Appelman Y, de Kleijn DP. The innate immune response in reperfused myocardium. Cardiovasc Res. 212;9: Wang X, Takagawa J, Lam VC, Haddad DJ, Tobler DL, Mok PY, Zhang Y, Clifford BT, Pinnamaneni K, Saini SA, Su R, Bartel MJ, Sievers RE, Carbone L, Kogan S, Yeghiazarians Y, Hermiston M, Springer ML. Donor myocardial infarction impairs the therapeutic potential of bone marrow cells by an interleukin-1-mediated inflammatory response. Sci Transl Med. 211;3:1ra19. Dewald O, Zymek P, Winkelmann K, Koerting A, Ren G, Abou-Khamis T, Michael LH, Rollins BJ, Entman ML, Frangogiannis NG. CCL2/monocyte chemoattractant protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ Res. 2;96: Conraads VM, Bosmans JM, Schuerwegh AJ, Goovaerts I, De Clerck LS, Stevens WJ, Bridts CH, Vrints CJ. Intracellular monocyte cytokine production and CD1 expression are up-regulated in severe vs. mild chronic heart failure. J Heart Lung Transplant. 2;2: Mahnke K, Schmitt E, Bonifaz L, Enk AH, Jonuleit H. Immature, but not inactive: The tolerogenic function of immature dendritic cells. Immunol Cell Biol. 22;8: Shortman K, Heath WR. The CD8+ dendritic cell subset. Immunol Rev. 21;23: Matta BM, Castellaneta A, Thomson AW. Tolerogenic plasmacytoid DC. Eur J Immunol. 21;: Brownlie RJ, Zamoyska R. T-cell receptor signalling networks: Branched, diversified and bounded. Nat Rev Immunol. 213;13: Eriksson U, Ricci R, Hunziker L, Kurrer MO, Oudit GY, Watts TH, Sonderegger I, Bachmaier K, Kopf M, Penninger JM. Dendritic cell-induced autoimmune heart failure requires cooperation between adaptive and innate immunity. Nat Med. 23;9: Maisel A, Cesario D, Baird S, Rehman J, Haghighi P, Carter S. Experimental autoimmune myocarditis produced by adoptive transfer of splenocytes after myocardial infarction. Circ Res. 1998;82:8-63 DOI: /CIRCRESAHA
17 FIGURE LEGENDS Figure 1. Activated macrophages infiltrate the failing heart. A, Cardiac mononuclear cells were isolated upon tissue digestion and gradient centrifugation purification. Top panels, Representative forward- and side-scatter profiles and live cell gates from a sham and HF heart. Bottom panels, Identification of Lin2 (defined in text) CD11b + F8 + cells, expressed as a percentage of total live cells, and corresponding group data from sham and HF hearts. B, Representative Mac-1 immunostains in sham and HF hearts and corresponding quantitation of Mac-1 + cells (arrows). C, Flow cytometric quantitation of classically-activated M1 (Lin2 CD11b + F/8 + CD26 ) and alternatively-activated M2 (Lin2 CD11b + F/8 + CD26 + ) macrophages in sham and HF hearts. Mϕ, macrophage; n = -/group. Downloaded from by guest on June 3, 218 Figure 2. Infiltrating dendritic cells (DCs) are increased in the failing heart. A, Representative confocal images of CD11c immunostained sections from sham and HF hearts (remote zone) and corresponding group data. The arrows indicate infiltrating CD11c + DCs; n = /group. B, Representative scatter plots for Lin1 (defined in text) cardiac mononuclear cells further separated into CD11c + B22 classical DCs and CD11c +/low B22 + plasmacytoid DCs, expressed as a percentage of total live cells, together with quantitative group data for cdc and pdc populations in sham and HF hearts. N = 8/group. C, Flow cytometric quantitation of CD86 hi and CD86 low pdcs in sham and HF hearts. N = -/group. Figure 3. Splenic remodeling in heart failure. A, Representative Masson s trichrome stains of spleens from sham-operated and HF mice. The low power views (Top) depict increased white pulp (WP) follicles with greater number of large germinal centers (clear areas in the WP) in the spleen from the HF mouse. The higher power views (Middle) illustrate the increased size of the marginal zone (MZ) surrounding the WP in the HF spleen. The bottom panels highlight the subcapsular red pulp (SRP), which exhibits fewer mononuclear cells in HF as compared with sham. PALS, periarteriolar lymphoid sheath. B, Representative confocal images of CD11b + SRP monocytes (Top) and F/8 + monocytes (Bottom) in the RP of sham and HF spleens, together with quantitative group data for CD11b + cells in the SRP. C, Representative live cell gates and scatter plots from sham and HF spleens identifying CD11b + F8 + cells, further subdivided by low or high Gr-1 expression (percentage of total live cells), and corresponding group data. N = -6/group. Figure. Remodeling of the splenic DC population in HF. A, Representative scatter plots for splenic mononuclear cells and live cell gates, with subsequent identification of Lin1 CD11c + B22 classical DCs (cdcs) and CD11c + B22 + plasmacytoid DCs (pdcs), and further subdivision of cdcs as CD8 + or CD8 cells. B and C, Quantitative group data for cdcs and pdcs, and CD8 + and CD8 cdcs, expressed as a percentage of total live cell population in sham and HF spleens. N = -6/group. D, Representative confocal images of CD11c + DCs in sham and HF spleens demonstrating prominent spatial redistribution of DCs to the white pulp germinal centers in the HF spleen. Figure. Altered circulating mononuclear phagocytes in HF. A, Lin2 CD11b + F/8 + monocytes were identified from the monocyte-lymphocyte gate of peripheral blood leukocytes, and further subdivided as high or low Gr-1 expressing cells (pro- and anti-inflammatory monocytes, respectively). Quantitative group data for circulating monocyte subsets in sham and HF mice are shown; n = /group. B, Schema for flow cytometric identification of circulating Lin1 CD11c + B22 cdcs and CD11c + B22 + pdcs, and CD8 + and CD8 cdcs, expressed as a percentage of the monocyte-lymphocyte gate of peripheral blood cells determined from side and forward scatter (SSC, FSC) plots as shown, together with quantitative group data for the same in sham and HF mice. In parallel studies, peripheral blood pdcs were alternatively identified as Siglec-H + cells and expressed as a percentage of the monocyte-lymphocyte gate. N = -12/group. DOI: /CIRCRESAHA
18 Figure 6. The spleen influences pathological cardiac remodeling in HF. A, Schema for splenectomy studies in mice with chronic HF. Splenectomy (or sham abdominal surgery) was performed in HF mice 8 weeks after coronary ligation (or sham operation), and remodeling was assessed by echocardiography over an additional 8 weeks. B, M-mode echocardiograms from one mouse at baseline, 8 weeks postligation (HF), and 16 weeks post-ligation/8 weeks post-splenectomy (HF/splenectomy). C, Quantitative group echocardiographic data for LV end-diastolic and end-systolic volume (EDV and ESV) and LV ejection fraction (EF) in sham operated and ligated HF mice 8 weeks after splenectomy (n = 7 sham mice, n = 11 HF mice) or sham abdominal surgery (n = HF mice). D. Representative confocal images of CD11b (for macrophages, arrows) and CD11c (for DCs, arrows) immunostained heart sections from HF mice (16 weeks post-ligation) with or without splenectomy at 8 weeks and corresponding quantitation (n = 3-/group). Downloaded from by guest on June 3, 218 Figure 7. HF-derived splenocytes induce pathological cardiac remodeling upon adoptive transfer. A, Schema for splenocyte adoptive transfer experiments. Splenocytes were isolated from CD.2 sham and HF mice 8 weeks after coronary ligation or sham operation and transferred to syngeneic CD.1 mice. Recipient mice were then followed for an 8-week period. B, M-mode echocardiograms from recipient mice 8 weeks after receiving splenocytes from sham or HF mice. C, Serial group echocardiographic data for EDV, ESV, and EF over the 8-week follow-up period after cell transfer (n = 7/group). D, Similar echocardiographic data in parallel groups of mice following adoptive transfer of splenocytes derived from donors treated with either lipopolysaccharide (LPS) or PBS control (n = 7/group). E, Representative hearts from mice receiving HF- or sham-splenocytes, and corresponding heart and lung gravimetric group data. F, Flow cytometry scatter plots and quantitation of circulating CD11b + F/8 + monocytes in recipient mice 8 weeks after cell transfer. G, Examples of spleens harvested from mice receiving HF- or sham-splenocytes, corresponding spleen gravimetry, and representative trichrome-stained histological sections from the same. N = -6/group for E-G. Figure 8. Example trichrome and CD.2 immunostains (A) and TUNEL stains (B) of hearts from HF- or sham-splenocyte recipient mice with corresponding quantitation of myocardial fibrosis and apoptosis. CD.2 + donor cells were frequently observed in interstitial and perivascular regions in HFsplenocyte recipient hearts (A). C, Representative confocal images of CD.2 and CD169 immunostained sections from sham- and HF-splenocyte recipient hearts and corresponding group data for CD.2 + CD169 + cells (arrows). N = 3-/group for A-C. DOI: /CIRCRESAHA
19 Novelty and Significance What Is Known? Sustained local and systemic inflammatory activation are hallmarks of chronic heart failure (HF). Inflammatory cells such as mononuclear phagocytes, i.e., monocytes/macrophages and dendritic cells (DCs) are both important sources and effector targets for pro-inflammatory cytokines. Although monocytes/macrophages and DCs are known to contribute importantly to cardiac remodeling early after myocardial infarction, their role in chronic HF is unknown. What New Information Does This Article Contribute? Downloaded from by guest on June 3, 218 Anomalies of the global mononuclear phagocyte network underlie the persistent inflammation in chronic HF, and promote the cardiac residence of pro-inflammatory monocytes/macrophages, classical DCs (cdcs), and plasmacytoid DCs (pdcs). In HF, the spleen exhibits heightened antigen processing, together with reductions in the splenic monocyte reservoir but augmentation of cdcs and pdcs, and increased expression of alarmins and pro-inflammatory mediators. The cardiosplenic plays an obligatory role in the pathogenesis and progression of cardiac remodeling in HF, as splenectomy reversed LV remodeling and inflammation, whereas adoptive transfer of HF-derived mononuclear splenocytes into naïve mice reproduced long-term cardiac remodeling. Whether inflammatory cells can be manipulated for therapeutic benefit in HF is unknown. In this study we show profound alterations in the mononuclear phagocyte network in HF that include: 1) increased proinflammatory macrophages and monocytes in the failing heart and circulation, and 2) augmented cdcs and pdcs in the heart and spleen. We also demonstrate marked splenic remodeling in HF indicative of heightened antigen processing, in concert with robust splenocyte expression of alarmins and proinflammatory mediators, and augmented splenic CD + and CD8 + T-cells. Finally, we show that the spleen is indispensable for remodeling progression and inflammation in HF, as: 1) splenectomy reversed remodeling and attenuated cardiac macrophage and DC infiltration, and 2) mononuclear splenocytes from HF mice homed to the heart and induced long-term cardiac remodeling, apoptosis, fibrosis, and dysfunction, and systemic inflammation upon adoptive transfer into naïve mice. Hence, we propose the novel paradigm that adverse LV remodeling in chronic ischemic HF is in part immune cell-mediated, potentially in response to cardiac-derived alarmins, and that targeting specific monocyte/macrophage and DC populations within the spleen and heart, or the antigens responsible for their activation, may comprise a more feasible approach to therapeutic immunomodulation in this disease. DOI: /CIRCRESAHA











20 Downloaded from by guest on June 3, 218 A B 2 p=.332 SSC-A Resident MΦ CD11b+F8+ cells ( % cell population) FSC-A F8 1μm C CD11b 2 p=.1 Resident classically activated Mϕϕ CD11b+F8+ CD26- cells (% cell population) Resident alternatively activated Mϕϕ CD11b+F8+ CD26+ cells (%cell population) C p= p= Mac-1 positive cells /high power field () () Figure 1


 B22/CDR 2μm DAPI")

 plasmacytoid DC")
 6 3 2 8 8 1.")



21 Downloaded from by guest on June 3, 218 A -Remote zone B Lin - (CD9.2/NK1.1/CD9B/Ly6G) B22/CDR 2μm DAPI CD11c DAPI CD11c 2μm HF-Border zone Magnified area CD11c 8 7 p= p= cells n) plasmacytoid DC CD11c +/low CDR/B22+cells (% cell population) classical DC CD11c+CDR/B22- (% cell populatio DAPI CD11c 2μm p=.12 1μm C p= p=. + cells/hpf CD86 hi pdc (% cell population) CD86 low pdc (% cell population) 2 1 CD11c Figure 2






22 Downloaded from by guest on June 3, 218 A B Sham HF DAPI CD11b DAPI CD11b MZ 1mm 1mm RP MZ WP RP WP Sham 2μm HF 2μm.2mm.1mm.2mm.1mm WP Sham PI F8 PI F8 WP RP RP MZ CD11b+ cells/hp PF μm WP 2 μm HF p=.68 Figure 3A-3B
 Lin - CD11b+F8+ Sham")




")


23 C FSC-A Lin - (CD9.2/NK1.1/CD9B) Lin - CD11b+F8+ Sham Sham Gr-1 low Gr-1 hi HF HF Gr-1 low Gr-1 hi SSC-A Lin-1 F CD11b CD11b Gr-1 p= p= p=.329 CD11b+F8+ cells (total % population) F8 Pro-inflammatory monocytes CD11b+F8+Gr-1hi cells (total % population) Anti-inflammatory monocytes CD11b+F8+Gr-1 low cells (total % population) Figure 3C Downloaded from by guest on June 3, 218













24 A B p=.23 Sham p=.23 classical DC CD11c+CDR/B B22- cells (total % population) plasmacytoid DC CD11c +/low CDR/B22+ cells (total % population) HF C 1.2 p= B22/CDR SSC-A p=.83 CD8- cdcs D11c+CDR/B22-CD8- cells (total % population) CD8+ cdc CD11c+CDR/B22 C 2-CD8+ cells (total % popu ulation) FSC-A CD8 D MZ CD11c CD11c CD11b DAPI RP MZ.6..2 CD11c. CD11c CD11b DAPI WP PALS 6 Magnified area C MZ PALS WP WP MZ MZ RP Sham 2μm HF RP MZ 2μm μm Figure Downloaded from by guest on June 3, 218
25 A 1 p=.1 p= p=.1 yteation) Activated Monocytes CD11b+F8+ cells Proinflammatory monocytes Lin-CD11b+F8+Gr-1 hi cells (% Lymphocyte- Monocyte population) Anti-inflammatory monocytes Lin-CD11b+F8+Gr-1 low cells (% Lymphocyte- Monocyte population) 1 2 Figure A Downloaded from by guest on June 3, 218 (% Lymphocy Monocyte popul

 CD11c CD11c p=.1 1.2 1..8.6. 1.6 1.")
 plasmacytoid DC")
 (% 7 6 3 2")

26 B FSC-A Lin - (CD9.2/NK1.1/CD9B/Ly6G) cdc Sham HF p= SSC-A B22/CDR CD8 Siglec-H cells Lymphocyte, Monocyte population) CD11c CD11c p= p=. CD8- cdcs CD11c+ +B22-CD8- cells (% Lymphocyte- Monocyte population) plasmacytoid DC CD11c +/low B22+ cells (% Lymphocyte- Monocyte population) (% p= Figure B Downloaded from by guest on June 3, classical DC CD11c+B22- cells (% Lymphocyte- population) Monocyte
 16 Wk Echo C ED DV (μl) 3 2 1-1 Spleen removed p=.316 Spleen removed p=.136 p=.")

27 Downloaded from by guest on June 3, 218 A B Baseline HF HF/Splenectomy CD.2 MI HF 8 Wk Spx (or Sham Spx) 16 Wk Echo C ED DV (μl) Spleen removed p=.316 Spleen removed p=.136 p=.72 8wk 16wk 8wk 16wk 8wk 16wk Spleen HF ES SV (μl) 3 p=.18 Spleen removed Spleen removed p=.26 p=.8 1 8wk 16wk 8wk 16wk 8wk 16wk n HF EF % Spleen removed p=.261 Spleen removed p=.3737 p=.127 8wk 16wk 8wk 16wk 8wk 16wk n HF Figure 6A-6C











28 Downloaded from by guest on June 3, 218 HF with spleen HF without spleen CD11b DAPI CD11b DAPI 8 p= CD11b fluorescenc ce intensity μm μm 2μm 2μm 3 Spleen + - HF CD11c DAPI CD11c DAPI p= cells/hpf 2 CD11c+ 2μm 2μm Spleen + - HF 3 Figure 6D

 8 p=.1 6 2 ES V (μl) 3 2 1 2 8 Wk Echo p=.")

 2 ESV (μl) 1 1 EF % 8 7 6 PBS-Splenocytes LPS-Splenocytes n")
29 Downloaded from by guest on June 3, 218 A CD.2 Spleen Mononuclear cells CD Wk FACS B Sham-Splenocytes HF-Splenocytes MI Tissue Harvest HF (or Sham) 8 Wk Naïve C EDV V (μl) 8 p= ES V (μl) Wk Echo p=.3 2mm EF E % ms p= ms Sham-Splenocytes HF-Splenocytes n = 7 per group D Splenocyte IV Injection 6 Weeks Splenocyte IV Injection Weeks Splenocyte IV Injection Weeks EDV (μl) 2 ESV (μl) 1 1 EF % PBS-Splenocytes LPS-Splenocytes n = 7 per group Weeks Splenocyte IV Injection Weeks Splenocyte IV Injection Weeks Splenocyte IV Injection Figure 7A-7D
30 E Sham-Splenocyte HF-Splenocyte p= p= Lung wt/tibia length (mg/cm) Heart wt/tibia length (mg/cm) F G Lin - (CD9.2 NK1.1 CD9B B22) Sham splenocyte transfer MZ 1.8 cmm 1.8 cmm p=.286 Splenocyte Injection IV p=.2 MZ mm.mm HF splenocyte transfer MZ CD11b CD11bb +F8+ (Percent monocyte /lymphocytee population) Spleen wt/tibia length (mg/cm) F8 MZ.mm Figure 7E-7G Downloaded from by guest on June 3, 218


31 Downloaded from by guest on June 3, 218 A Sham splenocyte transfer HF splenocyte transfer.2 mm.2 mm B CD.2 DAPI CD.2 DAPI p=.1 1 TnI TUNEL DAPI 2μm 2 μm TnI TUNEL DAPI 1 μm 2μm 2 μm ence CD D.2 fluoresc intensity UNEL positive nuclei /HPF T 1 Splenocyte Injection IV p=.3 3 Splenocyte Injection IV 3 Figure 8A-B








32 DAPI CD.2 DAPI CD169 DAPI DAPI CD.2CD169 3 p=.17 Sham-splenocyte CD.2+CD169 fluorescence intensity C 2μm DAPI CD169 DAPI DAPI CD.2 CD169 DAPI CD.2 CD.2 CD Splenocyte Injection IV μm 2μm Figure 8C Downloaded from by guest on June 3, 218 HF-splenocyte


33 Downloaded from by guest on June 3, 218 Remodeling of the Mononuclear Phagocyte Network Underlies Chronic Inflammation and Disease Progression in Heart Failure: Critical Importance of the Cardiosplenic Axis Mohamed A Ismahil, Tariq Hamid, Shyam S Bansal, Bindiya Patel, Justin R Kingery and Sumanth D Prabhu Circ Res. published online November 1, 213; Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 7231 Copyright 213 American Heart Association, Inc. All rights reserved. Print ISSN: Online ISSN: The online version of this article, along with updated information and services, is located on the World Wide Web at: Data Supplement (unedited) at: Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: Subscriptions: Information about subscribing to Circulation Research is online at:
34 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement EXPANDED METHODS All studies were performed in compliance with the NIH Guide for the Care and Use of Laboratory Animals (DHHS publication No. [NIH] 8-23, revised 1996). The University of Alabama at Birmingham Institutional Animal Care and Use Committee gave local approval for these studies. The total of 12 mice were used. Mouse models and surgical protocol. Male C7BL/6J mice 1-12 weeks of age (Jackson Laboratories, stock #66) were used. To induce pathological LV remodeling and HF, the mice underwent left thoracotomy and left coronary artery ligation (n=19) as previously described. 1-3 Sham operated mice (n=12) were used as controls. Anesthesia was induced in mice with tribromoethanol (.2 mg/g IP). Mice were then intubated and ventilated with a MiniVent Mouse Ventilator (Type 8, Harvard Apparatus) at 12-1 breaths/minute, with anesthesia maintained using 1-2% isoflurane and body temperature kept at 37 C using heat lamps and heating pads. Using sterile technique, mice were subjected to a thoracotomy in the th intercostal space. With the aid of a dissecting microscope, the proximal left coronary artery was visualized and permanently occluded with 8.-prolene suture, 1 mm distal to the left atrial appendage border. Successful occlusion was confirmed by the production of pallor and dyskinesia in the distal myocardium. In sham animals, the suture was passed but not tied. The chest was then closed in layers using. silk and the mice were weaned off isoflurane anesthesia. Upon recovery of spontaneous respiration, the intubation tube was removed and the mice were recovered in a temperature-controlled area supplemented with 1% oxygen. The mice were then followed for 8 weeks after operation and evaluated for the various readouts. Echocardiography. Mouse echocardiography was performed under anesthesia with tribromoethanol (.2 mg/g IP), and light (~1%) isoflurane as needed, using a VisualSonics Vevo 77 High-Resolution System with a RMV77B scanhead. Mice were imaged on a heated, bench-mounted adjustable rail system (Vevo Imaging Station) that allowed steerable and hands-free manipulation of the ultrasound transducer. Two echocardiographers performed the 1
35 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement studies and were blinded as to the specific experimental group. Left ventricular (LV) enddiastolic and end-systolic volumes (EDV and ESV) were planimetered from the long-axis images using the modified Simpson s method. LV systolic function was indexed by single plane LV ejection fraction (EF = [EDV-ESV]/EDV). Isolation of splenocytes. Eight weeks after coronary ligation or sham operation, the spleen was removed aseptically and placed in a 3 mm tissue culture dish with ~ ml of DMEM culture medium (Gibco, Invitrogen). Splenocytes were isolated according to the protocol of Lavelle at al, with slight modifications. Briefly, the spleen was finely minced using a scalpel. Repeated pipetting was used to disperse cells from minced fragments, and single cells were transferred to a fresh tube and kept on ice. For larger tissue pieces, the procedure was repeated several times in the dish with fresh DMEM. The final cell suspension was pipetted through a nylon cell strainer, 1 μm (BD Falcon), into a fresh tube and centrifuged at 1g for min at C. The supernatant was decanted, and the pellet re-suspended in. ml of RBC lysis buffer (ebiosciences) for min at room temperature (RT). The addition of fresh DMEM neutralized the erythrocyte lysis. Pelleted splenocytes were then re-suspended in fresh DMEM (.-1. ml). Isolation of bone marrow (BM) and peripheral blood cells. After mice were sacrificed, femurs and tibiae were dissected from the adherent soft tissue. The tip of each bone was removed with scissors, and the marrow was flushed with DMEM using a 27-gauge needle and aspirated. BM cells were filtered through a 1 μm nylon cell strainer (BD Falcon), sedimented at 1g for min at C, and re-suspended in RBC lysis buffer. After complete erythrocyte lysis, BM cells were re-suspended in fresh 1-2 ml DMEM. Peripheral blood (~ μl) was collected in BD Microtainer tubes with EDTA. Erythrocytes were lysed with 2 ml RBC lysis buffer for min on ice. Leukocytes were collected by centrifugation (38g for 1 min at C), and resuspended in ul of ice-cold flow cytometry staining buffer (ebioscience). 2
36 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement Isolation of mononuclear cells from the heart. Single mononuclear cells were isolated from sham and failing hearts following the method of Austyn et al. Mouse hearts were excised in toto and placed in heparinized saline. After removal of epicardial fatty tissue and the aorta, the heart was finely minced into ~1-2 mm pieces and blood was removed by repeated washing in saline (with stepwise reductions in added heparin). The tissue was digested with collagenase (Worthington, 1mg/ml), trypsin (Gibco, Invitrogen,.1%), and DNAse ( Prime, 1 μg/ml) in 1 ml RPMI media (Gibco) for min at 37 C with occasional shaking. Released cells were separated from solid tissue by filtering through a 1 μm nylon cell strainer (BD Falcon), washed with R1 media (RPMI-16 supplemented with 1% heat-inactivated FCS, 2mM-L- Glutamine, 2 μm-2-mercaptoethanol, 1% penicillin/streptomycin), and placed on ice. These steps were repeated 2-3 times to digest the remaining tissue. Any residual solid tissue was treated sequentially with EDTA (2 mm) in digestion media for 1 min at 37 o C, collagenase (2 mg/ml) in R1 media for 2 min at 37 o C, and released cells filtered through the strainer. All collected cells were pooled and pelleted at 1g for min at o C. Single cell suspensions were layered on Ficoll gradient solution and centrifuged at 2g for 2 min. To reduce myocyte contamination in the mononuclear cell suspension, 7% of the total volume (excepting cell debris) was collected and washed in R1 media. The presence of nucleated cells was confirmed by DAPI staining during flow cytometry analysis. In a separate aliquot, cell viability (>8%) was confirmed using the trypan blue exclusion method. Flow Cytometry. Isolated cell suspensions were incubated for 3 min at RT in a cocktail of fluorophore-labeled mabs (BD Biosciences or as otherwise indicated), as appropriate for the specific study, against: F/8-Pacific Blue; CD8-FITC; CDR/B22-APC; CD11b-6-NC (ebiosciences); Gr-1-APC (ebiosciences); CD11c-PE-Cy7; CD86-PE-Cy; CD26-FITC; Siglec- H-efluor 66 (ebiosciences), CD.1-FITC (Miltenyi Biotech); CD.2-PE. Lineage (Lin)1 antibody cocktail was used for DC identification (to exclude thymocytes: CD9.2-PE, natural killer cells: NK1.1-PE, CD9B-PE, and granulocytes: Gr-1-PE), and Lin2 antibody cocktail was 3
37 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement used for macrophage/monocyte subset identification (CD9.2-PE, NK1.1-PE, CD9B-PE, and CDR/B22-PE for B-cells). In separate assessments of splenic T-cells, cell suspensions were incubated with CD3-FITC, CD-6 NC and CD8-6 NC (ebiosciences). Of note, the anti-gr-1 antibody recognizes both Ly6C and Ly6G antigens. 6 Activated macrophages/monocytes were identified as CD11b + F/8 + cells. 7, 8 Proinflammatory monocytes were identified as Lin2 CD11b + F/8 + Gr-1 hi and anti-inflammatory monocytes as Lin2 CD11b + F/8 + Gr-1 low cells. 7-1 Pro-inflammatory M1 and anti-inflammatory M2 macrophages in tissue were sub-classified from Lin2 CD11b + F/8 + cells based on the absence or presence of CD26 (mannose receptor) expression. 11 Dendritic cells were identified as Lin1 CD11c + cells. 9, 1, 12 Classical dendritic cells (cdc) were identified as Lin1 CD11c + B22, whereas non-classical plasmacytoid dendritic cells (pdc) were identified as Lin1 CD11c +/low B , 12-1 In some studies, pdcs were identified as Siglec-H + cells. 1 Lymphoid cdcs were further subdivided as CD8 + or CD8, 1, 12 and pdcs as CD86 + or CD86 (classical costimulatory molecule and marker of pdc maturity). 16 Helper and cytotoxic T-cells were characterized as CD3 + CD + and CD3 + CD8 + CD cells, respectively. 17 For the heart, spleen, and BM, identified cells were normalized for total cell population. For peripheral blood, cell numbers were normalized for the total lymphocyte and monocyte gate. Data were acquired on an LSRII flow cytometer (BD Biosciences) and analyzed with FlowJo software, version Immunohistochemistry and confocal microscopy. Formalin-fixed, paraffin-embedded hearts and spleens from sham and HF mice were sectioned at μm thickness, deparaffinized and rehydrated; (immuno)histological staining was performed as previously described. 1, 3 Masson s trichrome was used to evaluate tissue fibrosis and general histology (heart and spleen), and Alexa Fluor 88-conjugated wheat-germ agglutinin (WGA; Invitrogen) for myocyte area. Myocardial apoptosis was evaluated using the DeadEnd Fluorometric TUNEL System (Promega) as previously described. 3, 18 This assay kit catalytically incorporates fluorescein-12-
38 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement dutp at the 3 -ends of fragmented DNA in apoptotic cells using recombinant terminal deoxynucleotidyl transferase (rtdt). Deparaffinized and rehydrated tissue sections were treated with Proteinase K (2 μg/ml) for 1 min at 37 C and then fixed with % methanol-free formaldehyde solution in PBS. All subsequent steps were performed following the manufacturer's instructions. Cardiomyocytes were identified by staining with anti-troponin I antibody (Santa Cruz Biotechnology) followed by Alexa Fluor -conjugated secondary antibody (Invitrogen) using the protocol below, and sections were counterstained with,6- diamidino-2-phenylindole (DAPI, Invitrogen) to label nuclei blue. TUNEL-positive nuclei (cyan staining) were visualized directly by confocal microscopy (Zeiss LSM71) with nuclear staining confirmed using z-axis sections. Images were taken with a 63x objective lens at five different locations in each tissue section. DNase (1 U/ml)-treated sections were used as positive controls. Sections without rtdt treatment were used as negative controls. For immunostaining, deparaffinized and rehydrated sections were incubated for 2 min with 1 mmol/l citric acid (ph 6.) and then treated with enzymatic antigen retrieval to recover antigenicity. Nonspecific binding was blocked with % normal goat serum and.% saponin (Sigma) in PBS (ph 7.) for 3 min. Sections were blocked and then incubated with the appropriate primary antibody in PBS with 1% BSA and.% saponin for 1 h at 37 C against: CD11b/ITAM (Epitomics), CD11c (ebiosciences), F/8 (ebiosciences), CD169 (R&D Systems), high mobility group box-1 (HMGB1, IBL International), and CD.2-PE (BD Biosciences). Tissue sections were then incubated for 3 min at room temperature with respective secondary secondary antibodies conjugated with Alexa 88,, or 67 (Invitrogen), and counterstained with DAPI to label nuclei. Images were made with a 63x objective lens at five different locations in each tissue section. Mean fluorescence intensity was evaluated using imagej 1.7V software for at least five images/heart. Myocyte area and cardiac fibrosis were quantified from -6 high-power fields per section in the remote non-infarcted area of the heart using Image J and Metamorph software,
39 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement respectively. In the spleen, CD11b positive cells in the subcapsular red pulp were counted in 3- high power fields per section. Data for each group were calculated from 12-1 sections and - mice from each group. The measurements and calculations were conducted in a blinded manner. Confocal microscopy was carried out on an LSM71 microscope (Zeiss) and Z-stack images were acquired according to standard protocols. Cytometric bead array (CBA) immunoassay for serum cytokines. Peripheral blood was allowed to clot at RT for 2 h. After centrifugation at 2, rpm for 1 min, serum was collected and stored at 2 C until assays were performed. Serum concentrations of TNF, monocyte chemoattractant protein (MCP)-1, interferon (IFN)-γ, interleukin (IL)-6, IL-1, and IL- 12p7 were measured simultaneously using a CBA Mouse Inflammation Kit (BD Biosciences). Briefly, µl of chemokine capture bead mixture was incubated with µl of either recombinant standard or sample and µl of PE-conjugated detection antibody for 2 h at RT. The mixture was washed to remove unbound PE detection reagent before data acquisition on a BD LSRII flow cytometer. Analysis was performed using FCAP Array software. Splenocyte gene expression by quantitative real-time PCR. Mononuclear splenocytes were cultured in serum free DMEM media for 2 h after isolation (2-3 X 1 6 cells/well). The cells were then collected and stored at -8 C in TRIzol reagent (Invitrogen) for subsequent RNA isolation. Total RNA levels were quantified and sample purity assessed using the ratio of absorbance at 26 and 28 nm by NanoDrop 1 Spectrophotometer (Thermo Scientific). Subsequent cdna synthesis and quantitative real-time PCR using SYBR Green (Life Technologies) were performed as described previously. 1, 2, 18 Briefly, total RNA ( ng) was reversed transcribed using High Efficiency cdna Synthesis Kit (Life Technologies). Relative levels of mrna transcripts for TNF, IL-1β, IL-2, IL-, IL-, IL-6, IL-1, IL-13, IFN-γ, inducible nitric oxide synthase (inos), C-C chemokine receptor type 2 (CCR2), CCR, chemokine (C-C motif) ligand (CCL), chemokine (C-X-C motif) receptor 3 (CXCR3), CX3C chemokine receptor 1 (CX3CR1), toll-like receptor (TLR)1, TLR7, TLR9, TLR12, transforming growth factor 6
40 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement (TGF)-β, HMGB1, S1A8, S1A9, and galectin-3 were determined using forward and reverse primer pairs detailed in Supplemental Table I, and normalized to 18s rrna expression using the C T comparative method. 1 Survival splenectomy and splenocyte adoptive transfer. In a separate sub-group, male (CD.2) C7BL/6J mice (n = 36) underwent coronary ligation or sham operation. Eight weeks later, survival splenectomy (or, in some mice, sham abdominal surgery) was performed using standard surgical techniques. 19 Briefly, mice were anesthetized with tribromoethanol (.2 mg/g IP) and anesthesia was maintained with 1% isoflurane as needed. After sterilizing the surgical area, a ~1 cm incision was made in the left subcostal region, and the peritoneum was opened to exteriorize the spleen. The spleen was retracted away from the pancreatic tail, the splenic bundle was ligated at the hilum using 8. prolene ligature, and then the spleen was removed intact. For sham operations, the splenic bundle and spleen were left intact. The procedure required ~1 1 minutes to complete; totality of splenectomy was ensured by close examination at the time of operation and confirmed during postmortem examination. Splenocytes from sham-operated and HF mice were used for the adoptive transfer studies described below. Splenectomized mice (and sham abdominal surgery controls) were subsequently followed for 8 more weeks with serial echocardiography. For adoptive transfer studies, splenocytes were prepared as described previously. 2 Briefly, single cell suspensions were prepared from spleens isolated from sham-operated (n=8) and HF mice (n=7). The erythrocytes were lysed with RBC lysis buffer (ebioscience, CA). Single cell suspensions were then layered on Ficoll-Paque (GE Healthcare) gradient and centrifuged at 2g for 2 min to separate mononuclear cells and exclude granulocytes and residual erythrocytes. Mononuclear cells were collected, washed three times with sterile PBS, and resuspended in sterile PBS. Cells were then pooled from either sham or HF mice and resuspended in ml PBS. Mononuclear cells were injected (~17 X 1 6 cells in 1 μl PBS per animal via tail vein) into naïve CD.1 C7BL/6J mice (7 mice receiving sham splenocytes and 7
41 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement 7 mice receiving HF splenocytes). As an additional control for adoptive transfer, we also isolated splenocytes from CD.2 C7BL/6J mice (male, 6-8 weeks) treated with either lipopolysaccharide (LPS) to induce systemic inflammation or PBS control. LPS (. mg/kg) or PBS (.2 ml) was injected i.p. daily for two consecutive days (n = per group). Spleens were aseptically harvested 2 h after the last i.p. dose, and the mononuclear cell population was isolated and injected via tail vein into naïve CD.1 C7BL/6 recipient mice (n = 7 per group receiving either LPS-splenocytes or PBS-splenocytes) as above. Statistical Analysis. Continuous data are summarized as mean ± standard deviation (SD). Statistical comparisons were performed using the unpaired student s t-test when comparing two groups. For comparisons of more than two groups, two-way ANOVA was used with Bonferroni post-test to adjust for multiple comparisons. A p value of <. was considered statistically significant. REFERENCES 1. Hamid T, Gu Y, Ortines RV, Bhattacharya C, Wang G, Xuan YT, Prabhu SD. Divergent tumor necrosis factor receptor-related remodeling responses in heart failure: Role of nuclear factor-κb and inflammatory activation. Circulation. 29;119: Hamid T, Guo SZ, Kingery JR, Xiang X, Dawn B, Prabhu SD. Cardiomyocyte NF-κB p6 promotes adverse remodelling, apoptosis, and endoplasmic reticulum stress in heart failure. Cardiovasc Res. 211;89: Wang G, Hamid T, Keith RJ, Zhou G, Partridge CR, Xiang X, Kingery JR, Lewis RK, Li Q, Rokosh DG, Ford R, Spinale FG, Riggs DW, Srivastava S, Bhatnagar A, Bolli R, Prabhu SD. Cardioprotective and antiapoptotic effects of heme oxygenase-1 in the failing heart. Circulation. 21;121: Lavelle GC, Sturman L, Hadlow WJ. Isolation from mouse spleen of cell populations with high specific infectivity for scrapie virus. Infect Immun. 1972;: Austyn JM, Hankins DF, Larsen CP, Morris PJ, Rao AS, Roake JA. Isolation and characterization of dendritic cells from mouse heart and kidney. J Immunol. 199;12: Fleming TJ, Fleming ML, Malek TR. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C mab to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J Immunol. 1993;11:
42 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement 7. Robbins CS, Swirski FK. The multiple roles of monocyte subsets in steady state and inflammation. Cell Mol Life Sci. 21;67: Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2;: Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K. Development of monocytes, macrophages, and dendritic cells. Science. 21;327: Shortman K, Naik SH. Steady-state and inflammatory dendritic-cell development. Nature reviews. Immunology. 27;7: Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat Immunol. 21;11: Kushwah R, Hu J. Complexity of dendritic cell subsets and their function in the host immune system. Immunology. 211;133: Hadeiba H, Lahl K, Edalati A, Oderup C, Habtezion A, Pachynski R, Nguyen L, Ghodsi A, Adler S, Butcher EC. Plasmacytoid dendritic cells transport peripheral antigens to the thymus to promote central tolerance. Immunity. 212;36:38-1. Hochrein H, O'Keeffe M, Wagner H. Human and mouse plasmacytoid dendritic cells. Human Immunol. 22;63: Blasius AL, Colonna M. Sampling and signaling in plasmacytoid dendritic cells: The potential roles of Siglec-H. Trends Immunol. 26;27: Ito T, Yang M, Wang YH, Lande R, Gregorio J, Perng OA, Qin XF, Liu YJ, Gilliet M. Plasmacytoid dendritic cells prime IL-1-producing T-regulatory cells by inducible costimulator ligand. J Exp Med. 27;2: Kitchen SG, Jones NR, LaForge S, Whitmire JK, Vu BA, Galic Z, Brooks DG, Brown SJ, Kitchen CM, Zack JA. CD on CD8+ T cells directly enhances effector function and is a target for HIV infection. Proc Natl Acad Sci USA. 2;11: Ismahil MA, Hamid T, Haberzettl P, Gu Y, Chandrasekar B, Srivastava S, Bhatnagar A, Prabhu SD. Chronic oral exposure to the aldehyde pollutant acrolein induces dilated cardiomyopathy. Am J Physiol Heart Circ Physiol. 211;31:H Wang HX, Yi SQ, Li J, Terayama H, Naito M, Hirai S, Qu N, Yi N, Itoh M. Effects of splenectomy on spontaneously chronic pancreatitis in aly/aly mice. Clin Dev Immunol. 21;21: Kodama M, Matsumoto Y, Fujiwara M. In vivo lymphocyte-mediated myocardial injuries demonstrated by adoptive transfer of experimental autoimmune myocarditis. Circulation. 1992;8:
43 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement Supplemental Table I. Forward and Reverse Primers for Real-Time PCR Gene Name Gene ID Accession # Sequence TNF-α NM_13693 F: CAGCCGATGGGTTGTACCTT R: GGCAGCCTTGTCCCTTGA IL-1β NM_8361 F: AGTTGACGGACCCCAAAAGA R: GGACAGCCCAGGTCAAAGG IL NM_8366 F: AAACTAAAGGGCTCTGACAACACA R: CACCACAGTTGCTGACTCATCA IL NM_21283 F: GGAGATGGATGTGCCAAACG R: CGAGCTCACTCTCTGTGGTGTT IL NM_18 F: TGCACTTGAGTGTTCTGACTCTCA R: TGTGCTCATGGGAATCTCCAT IL NM_31168 F: CCACGGCCTTCCCTACTTC R: TTGGGAGTGGTATCCTCTGTGA IL NM_18 F: GATGCCCCAGGCAGAGAA R: CACCCAGGGAATTCAAATGC IL NM_83 F: TTGAGGAGCTGAGCAACATCAC R: CCATGCTGCCGTTGCA IFN-γ 1978 NM_8337 F: TTGGCTTTGCAGCTCTTCCT R: TGACTGTGCCGTGGCAGTA inos NM_1927 F: AGACCTCAACAGAGCCCTCA R: GCAGCCTCTTGTCTTTGACC CCR NM_991 F: CTGCTCAACTTGGCCATCTCT R: GTGAGCCCAGAATGGTAATGTG CCR 1277 NM_9917 F: GTCAAACGCTTTTGCAAACG R: TGAGCTTGCACGATCAGGATT CCL 23 NM_1363 F: TCCAATCTTGCAGTCGTGTTTG R: TCTGGGTTGGCACACACTTG CXCR NM_991 F: TTGCCCTCCCAGATTTCATC R: TGGCATTGAGGCGCTGAT CX3CR1 131 NM_9987 F: TCGGTCTGGTGGGAAATCTG R: GGCTTCCGGCTGTTGGT TLR NM_1276 F: GGCTTTGCAGGAACTCAATGTA R: CCCCGCACCCAGGAA TLR NM_ F: CAGTGAACTCTGGCCGTTGA R: CAAGCCGGTTGTTGGAGAA TLR NM_31178 F: CCTTCGTGGTGTTCGATAAGG R: CACCCGCAGCTCGTTATACA TLR NM_2823 F: CTGCCACTGTGCCAATGC 1
44 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement R: ATATGTACGTTTTAGTGGACCGCTTA TGF-β 2183 NM_1177 F: GCAGTGGCTGAACCAAGGA R: AGCAGTGAGCGCTGAATCG HMGB NM_139 F: CCCCAATGCACCCAAGAG R: GGCGGTACTCAGAACAGAACAAG Galectin NM_1193 F: CACAATCATGGGCACAGTGAA R: TTCCCTCTCCTGAAATCTAGAACAA S1A8 221 NM_136 F: GGCCTTGAGCAACCTCATTG R: AGAGGGCATGGTGATTTCCTT S1A9 222 NM_911 F: TCATGGAGGACCTGGACACA R: CAGCATCATACACTCCTCAAAGCT 11
45 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure I. Long-term pathological cardiac remodeling after coronary ligation. A, Representative M-mode echocardiograms (Left) and Masson s trichrome-stained short-axis histological sections (Right) from sham-operated mice and mice with permanent coronary ligation and heart failure (HF), 8 weeks after operation. Histological images were taken with a 2X objective; scale bar 1 mm. B, Higher power histological images of trichrome (left) and Alexa Fluor 88-conjugated wheat-germ agglutinin (WGA) stains of sham and HF heart sections. C, quantitative group data for gravimetric, histological, and echocardiographic analyses from sham and HF mice. EDV and ESV, end-diastolic and end-systolic volume; EF, ejection fraction. Fibrosis and myocyte area were determined in myocardium remote from the infarction. N = - 7/group. Supplemental Figure II. Serum cytokine levels in sham and HF mice as determined by cytometric bead array immunoassay. IL, interleukin; TNF, tumor necrosis factor; IFN, interferon, MCP, monocyte chemotactic protein. N = 3-/group. Supplemental Figure III. Representative flow cytometry scatter plots for SSC versus Siglec-H fluorescence in mononuclear splenocytes from sham and HF mice, expressed as percentage of the total mononuclear cell population, and corresponding group data. An unstained control sample (Left) is also shown for comparison. N = -/group. Supplemental Figure IV. Representative live cell gates and scatter plots from sham and HF spleens identifying CD3 + CD + and CD3 + CD8 + CD cells as a percentage of total live mononuclear cell population, and corresponding group data. N = -/group. Supplemental Figure V. Quantitative group data for activated monocytes (Lin2 CD11b + F/8 + cells), classical DCs (Lin1 CD11c + B22 cells), and plasmacytoid DCs (Lin1 CD11c +/low B22 + cells) in bone marrow from sham and HF mice. N = -6/group. Supplemental Figure VI. Serum cytokine levels as determined by cytometric bead array immunoassay in mice with HF (16 weeks after coronary ligation) that underwent either 12
46 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement splenectomy (HF without spleen) or sham abdominal surgery (HF with spleen) 8 weeks after ligation. All values normalized to HF with spleen group. TNF, tumor necrosis factor; MCP, monocyte chemotactic protein; IL, interleukin; IFN, interferon. N = -6/group. Supplemental Figure VII. Representative peripheral blood leukocyte flow cytometry scatter plots for recipient CD.1 + versus donor CD.2 + cells (monocyte-lymphocyte gate) two weeks following splenocyte adoptive transfer (AT) demonstrating low levels of residual circulating CD.2 + donor cells (1.6% in this example). An unstained blood sample is shown for comparison. Supplemental Figure VIII. Serum cytokine levels as determined by cytometric bead array immunoassay in mice that received either sham-splenocytes or HF-splenocytes, measured 8 weeks after adoptive transfer. All values normalized to sham-splenocyte group. TNF, tumor necrosis factor; MCP, monocyte chemotactic protein; IL, interleukin; IFN, interferon. N = 3-/group. Supplemental Figure IX. A, Lin2 CD11b + F/8 + circulating monocytes were identified from the monocyte-lymphocyte gate of peripheral blood leukocytes from mice treated with either lipopolysaccharide (LPS) or PBS control for two days. CD11b + F/8 + cells were then subdivided as Gr-1 low or high. Representative scatter plots from PBS and LPS treated mice (and unstained sample control) are shown in the top panels and quantitative group data for circulating monocyte subsets in the bottom panels. N = /group. B, Representative M-mode echocardiograms from recipient mice given mononuclear splenocytes from either PBS- or LPStreated mice, at baseline (prior to transfer) and 8 weeks after adoptive cell transfer. Supplemental Figure X. Normalized gene expression of select pro- and antiinflammatory mediators and TLR pattern-recognition receptors (PRRs) in mononuclear splenocytes isolated from sham-operated and HF mice. *p <. vs. sham; n = -6/group. Supplemental Figure XI. A, Representative splenic immunostains for the alarmin HMGB1 from sham-operated and HF mice and corresponding quantitation for HMGB1 13
47 Ismahil et al. CIRCRES/213/3172/R2 Online Supplement fluorescence intensity. WP, white pulp, RP, red pulp, MZ, marginal zone. B, Normalized gene expression of the alarmins HMGB1, S1A8, S1A9, and galectin-3 in mononuclear splenocytes isolated from sham and HF mice. n = -/group for A and B. C, Representative confocal images of HMGB1 (green), CD.2 (red), and DAPI (blue, for nuclei) (immuno)staining of hearts from CD.1 mice 8 weeks after adoptive transfer of CD.2 sham- or HF-splenocytes. The yellow arrow indicates a CD.2 + HMGB1 + cell cluster in the HF-splenocyte recipient heart. The green arrows indicate other areas of HMGB1 expression in recipient myocardium. 1

 2μm Sham HF 3")


")
 Sham 1ms")
 Trichrome stain")
 B Sham")
48 A Sham HF HF Sham 2 mm Sham 1 2μm 1 p= ESV ( L) EDV ( L) 2μm Sham HF p= p mm.2mm HF p=.1 p=.39 p=.21 HF 1. EF %.2mm p=.22 1 Myocyte area ( m2) C WGA stain Myocardial fibrosis (%) Sham 1ms Lung wt/tibia length (m g/m m ) Trichrome stain 1ms Heart Wt/tibia length (mg/mm) B Sham HF Sham HF Supplemental Figure I
49 A IL-6 pg/ml p=.7 3 Wt HF MCP-1 pg/ml p=. 3 Wt HF IFN- pg/ml p=.13 Wt HF IL12p7 pg/ml p=.28 3 Wt HF IL1 pg/ml p=. Wt HF TNF pg/ml p= Wt HF Supplemental Figure II
50 Unstained SSC-A Siglec-H Siglec-H population (total percent population) p=.37 Supplemental Figure III

51 Sham CD3+CD+ 17. CD3+CD8+CD CD3+CD p=.27 SSC-A FSC-A HF CD3 CD3+CD CD CD3 CD3+CD8+CD- 9.9 CD8 CD3+CD8+CD p=.36 Supplemental Figure IV
52 3 2 1 p= p= p= Supplemental Figure V activated monocytes CD11b+F8+ cells (total % population) classical DC CD11c+B22- cells (total % population) plasmacytoid DC CD11c +/low B22+ cells (total % population) 6
53 Supplemental Figure VI
54 Unstained 2 Wks after AT CD.1 CD.2 Supplemental Figure VII
55 Supplemental Figure VIII
56 Unstained PBS LPS F8 CD11b 1 1 p=.287 CD11b + F8 + cells/ (% Lymphocyte, Monocyte population) ) PBS LPS Challenge for 8hrs p=.9 CD11b + F8 + Gr-1 hi cells/ (% Lymphocyte, Monocyte population ) PBS LPS Challenge for 8hrs p=.1932 CD11b + F8 + Gr-1 low cells/ (% Lymphocyte, Monocyte population ) PBS LPS Challenge for 8hrs Supplemental Figure IXA




57 Baseline PBS-Splenocytes 8 Wks PBS-Splenocytes 2mm 1ms LPS-Splenocytes 1ms LPS-Splenocytes 2mm 1ms 1ms Supplemental Figure IXB
58 2. NS 2. 3 * * * * * IL-2/18s IL-6/18s inos/18s TNF /18s IFN- /18s. Pro inflammatory cytokines IL-1 /18s NS * * * * * NS CCR2/18s CX3CR1/18s CCL/18s CXCR3/18s IL-/18s NS () () Pro inflammatory chemokines CCR/18s * IL-1/18s Anti inflammatory cytokines IL-13/18s IL-/18s NS TLR1/18s TGF /18s * * * 2 1 TLR12/18s TLR9/18s TLR7/18s NS Supplemental Figure X Innate immunity/prr

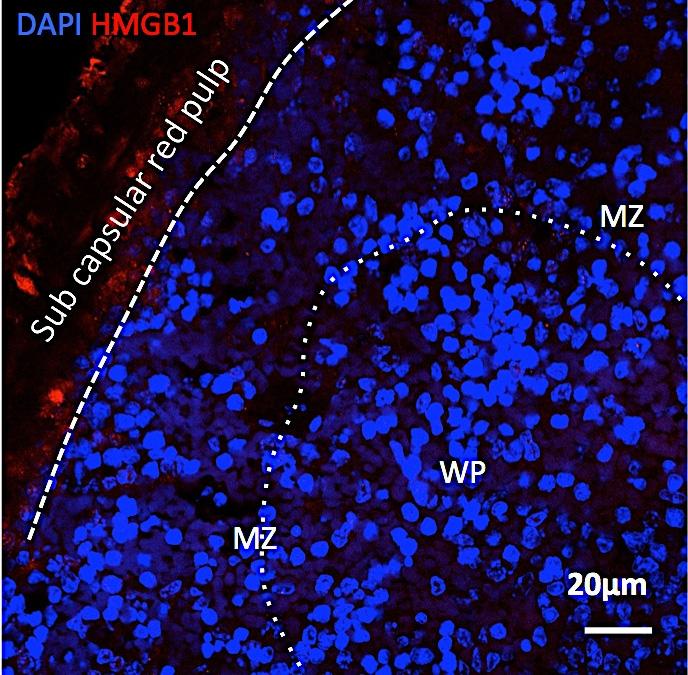
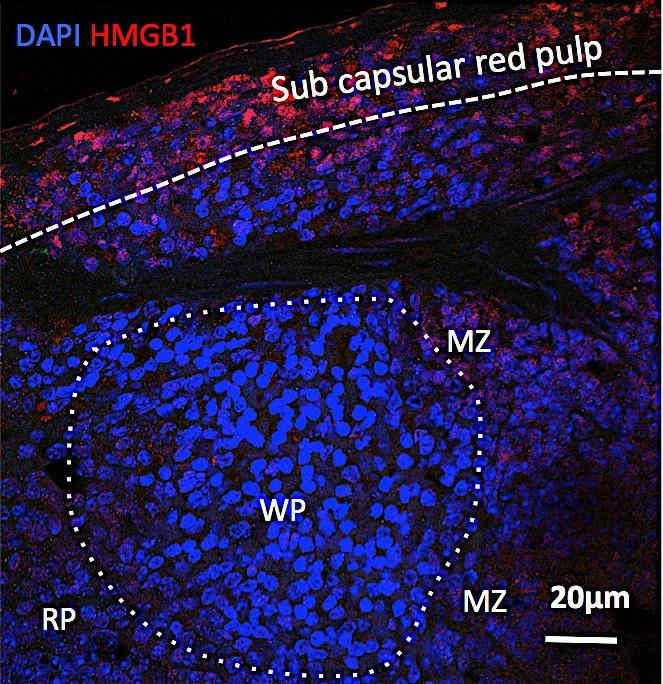

59 HF HMGB1 fluorescence intensity Sham Sham p=.396 HF Supplemental Figure XIA
60 3 p = p =.19 HMGB1/18s 2 1 S1A8/18s p =.3 p =.6 S1A9/18s Galectin-3/18s Supplemental Figure XIB




61 F DAPI HMGB1 CD.2 DAPI HMGB1 CD.2 DAPI CD.2 DAPI HMGB1 Sham splenocyte 2 μm HF splenocyte 2 μm Supplemental Figure XIC
Integrative Physiology. Remodeling of the Mononuclear Phagocyte Network Underlies Chronic Inflammation and Disease Progression in Heart Failure
 Integrative Physiology Remodeling of the Mononuclear Phagocyte Network Underlies Chronic Inflammation and Disease Progression in Heart Failure Critical Importance of the Cardiosplenic Axis Mohamed Ameen
Integrative Physiology Remodeling of the Mononuclear Phagocyte Network Underlies Chronic Inflammation and Disease Progression in Heart Failure Critical Importance of the Cardiosplenic Axis Mohamed Ameen
Supporting Information
 Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplemental Table I.
 Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Primary Adult Naïve CD4+ CD45RA+ Cells. Prepared by: David Randolph at University of Alabama, Birmingham
 Primary Adult Naïve CD4+ CD45RA+ Cells Prepared by: David Randolph (drdrdr@uab.edu) at University of Alabama, Birmingham Goal: To obtain large numbers of highly pure primary CD4+ CD45RO- CD25- cells from
Primary Adult Naïve CD4+ CD45RA+ Cells Prepared by: David Randolph (drdrdr@uab.edu) at University of Alabama, Birmingham Goal: To obtain large numbers of highly pure primary CD4+ CD45RO- CD25- cells from
Hematopoiesis. Hematopoiesis. Hematopoiesis
 Chapter. Cells and Organs of the Immune System Hematopoiesis Hematopoiesis- formation and development of WBC and RBC bone marrow. Hematopoietic stem cell- give rise to any blood cells (constant number,
Chapter. Cells and Organs of the Immune System Hematopoiesis Hematopoiesis- formation and development of WBC and RBC bone marrow. Hematopoietic stem cell- give rise to any blood cells (constant number,
SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplemental Table 1. Primer sequences for transcript analysis
 Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Rapid antigen-specific T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
SUPPLEMENTARY INFORMATION. CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative. breast cancer patients
 SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
Eosinophils are required. for the maintenance of plasma cells in the bone marrow
 Eosinophils are required for the maintenance of plasma cells in the bone marrow Van Trung Chu, Anja Fröhlich, Gudrun Steinhauser, Tobias Scheel, Toralf Roch, Simon Fillatreau, James J. Lee, Max Löhning
Eosinophils are required for the maintenance of plasma cells in the bone marrow Van Trung Chu, Anja Fröhlich, Gudrun Steinhauser, Tobias Scheel, Toralf Roch, Simon Fillatreau, James J. Lee, Max Löhning
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
Application Information Bulletin: Human NK Cells Phenotypic characterizing of human Natural Killer (NK) cell populations in peripheral blood
 cell populations in peripheral blood") Application Information Bulletin: Human NK Cells Phenotypic characterizing of human Natural Killer (NK) cell populations in peripheral blood Christopher A Fraker, Ph.D., University of Miami - Miami, Florida
Application Information Bulletin: Human NK Cells Phenotypic characterizing of human Natural Killer (NK) cell populations in peripheral blood Christopher A Fraker, Ph.D., University of Miami - Miami, Florida
NK cell flow cytometric assay In vivo DC viability and migration assay
 NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Supplemental Materials for. Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to. FTY720 during neuroinflammation
 Supplemental Materials for Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to FTY7 during neuroinflammation This file includes: Supplemental Table 1. EAE clinical parameters of
Supplemental Materials for Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to FTY7 during neuroinflammation This file includes: Supplemental Table 1. EAE clinical parameters of
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
T H 1, T H 2 and T H 17 polarization of naïve CD4 + mouse T cells
 A complete workflow for cell preparation, isolation, polarization and analysis T H 1, T H 2 and T H 17 polarization of naïve CD4 + mouse T cells Introduction Workflow CD4 + T helper (T H) cells play a
A complete workflow for cell preparation, isolation, polarization and analysis T H 1, T H 2 and T H 17 polarization of naïve CD4 + mouse T cells Introduction Workflow CD4 + T helper (T H) cells play a
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Targeting tumour associated macrophages in anti-cancer therapies. Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018
 Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10188 Supplementary Figure 1. Embryonic epicardial genes are down-regulated from midgestation stages and barely detectable post-natally. Real time qrt-pcr revealed a significant down-regulation
doi:10.1038/nature10188 Supplementary Figure 1. Embryonic epicardial genes are down-regulated from midgestation stages and barely detectable post-natally. Real time qrt-pcr revealed a significant down-regulation
Serafino et al. Thymosin α1 activates complement receptor-mediated phagocytosis in human monocyte-derived macrophages. SUPPLEMENTARY FIGURES
 Supplementary Fig. S1. Evaluation of the purity and maturation of macrophage cultures tested by flow cytometry. The lymphocytic/monocytic cellular fraction was isolated from buffy coats of healthy donors
Supplementary Fig. S1. Evaluation of the purity and maturation of macrophage cultures tested by flow cytometry. The lymphocytic/monocytic cellular fraction was isolated from buffy coats of healthy donors
The encephalitogenicity of TH17 cells is dependent on IL-1- and IL-23- induced production of the cytokine GM-CSF
 CORRECTION NOTICE Nat.Immunol. 12, 568 575 (2011) The encephalitogenicity of TH17 cells is dependent on IL-1- and IL-23- induced production of the cytokine GM-CSF Mohamed El-Behi, Bogoljub Ciric, Hong
CORRECTION NOTICE Nat.Immunol. 12, 568 575 (2011) The encephalitogenicity of TH17 cells is dependent on IL-1- and IL-23- induced production of the cytokine GM-CSF Mohamed El-Behi, Bogoljub Ciric, Hong
Supporting Information
 Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Innate Immunity II. Integration. Lindsay Nicholson Advanced Immunology L2
 Innate Immunity II Integration Lindsay Nicholson Advanced Immunology L2 l.nicholson@bristol.ac.uk Lecture 1 Defining Innate Immunity Recognition and effector mechanisms (I) Lecture 2 Recognition and effector
Innate Immunity II Integration Lindsay Nicholson Advanced Immunology L2 l.nicholson@bristol.ac.uk Lecture 1 Defining Innate Immunity Recognition and effector mechanisms (I) Lecture 2 Recognition and effector
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
General Overview of Immunology. Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center
 General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
In vitro human regulatory T cell expansion
 - 1 - Human CD4 + CD25 + regulatory T cell isolation, Workflow in vitro expansion and analysis In vitro human regulatory T cell expansion Introduction Regulatory T (Treg) cells are a subpopulation of T
- 1 - Human CD4 + CD25 + regulatory T cell isolation, Workflow in vitro expansion and analysis In vitro human regulatory T cell expansion Introduction Regulatory T (Treg) cells are a subpopulation of T
ALETA : MODE OF ACTION
 ALETA : MODE OF ACTION Lakshmibai Vasanthakumari Bindhu. PhD ABSTRACT Previous research has shown that beta-glucan has immunomodulatory effects. Paramylon is the beta-glucan found in Aleta and forms granules.
ALETA : MODE OF ACTION Lakshmibai Vasanthakumari Bindhu. PhD ABSTRACT Previous research has shown that beta-glucan has immunomodulatory effects. Paramylon is the beta-glucan found in Aleta and forms granules.
In vitro human regulatory T cell expansion
 - 1 - Human CD4 + CD25 + CD127 dim/- regulatory T cell Workflow isolation, in vitro expansion and analysis In vitro human regulatory T cell expansion Introduction Regulatory T (Treg) cells are a subpopulation
- 1 - Human CD4 + CD25 + CD127 dim/- regulatory T cell Workflow isolation, in vitro expansion and analysis In vitro human regulatory T cell expansion Introduction Regulatory T (Treg) cells are a subpopulation
HARVARD MEDICAL SCHOOL
 The Role of the Various Monocyte Types in Atherothrombotic Disease Protective vs. Detrimental Effects and Therapeutic Implications Matthias Nahrendorf MGH Center for Systems Biology http://csb.mgh.harvard.edu/investigator/matthias_nahrendorf
The Role of the Various Monocyte Types in Atherothrombotic Disease Protective vs. Detrimental Effects and Therapeutic Implications Matthias Nahrendorf MGH Center for Systems Biology http://csb.mgh.harvard.edu/investigator/matthias_nahrendorf
Cover Page. The handle holds various files of this Leiden University dissertation.
 Cover Page The handle http://hdl.handle.net/1887/23854 holds various files of this Leiden University dissertation. Author: Marel, Sander van der Title: Gene and cell therapy based treatment strategies
Cover Page The handle http://hdl.handle.net/1887/23854 holds various files of this Leiden University dissertation. Author: Marel, Sander van der Title: Gene and cell therapy based treatment strategies
Supplementary Information. Tissue-wide immunity against Leishmania. through collective production of nitric oxide
 Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1: Chemokine receptor expression profiles of CCR6 + and CCR6 - CD4 + IL-17A +/ex and Treg cells. Quantitative PCR analysis of chemokine receptor transcript abundance
SUPPLEMENTARY FIGURES Supplementary Figure 1: Chemokine receptor expression profiles of CCR6 + and CCR6 - CD4 + IL-17A +/ex and Treg cells. Quantitative PCR analysis of chemokine receptor transcript abundance
BD Flow Cytometry Reagents Multicolor Panels Designed for Optimal Resolution with the BD LSRFortessa X-20 Cell Analyzer
 Multicolor Panels Designed for Optimal Resolution with the BD LSRFortessa X-2 Cell Analyzer Proper multicolor panel design takes into account fluorochrome brightness, antigen density, co-expression, and
Multicolor Panels Designed for Optimal Resolution with the BD LSRFortessa X-2 Cell Analyzer Proper multicolor panel design takes into account fluorochrome brightness, antigen density, co-expression, and
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5
 sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
Supplemental Information. T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism
 Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
MACROPHAGE "MONOCYTES" SURFACE RECEPTORS
 LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH
 FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Immunology for the Rheumatologist
 Immunology for the Rheumatologist Rheumatologists frequently deal with the immune system gone awry, rarely studying normal immunology. This program is an overview and discussion of the function of the
Immunology for the Rheumatologist Rheumatologists frequently deal with the immune system gone awry, rarely studying normal immunology. This program is an overview and discussion of the function of the
Nature Immunology: doi: /ni Supplementary Figure 1. Examples of staining for each antibody used for the mass cytometry analysis.
 Supplementary Figure 1 Examples of staining for each antibody used for the mass cytometry analysis. To illustrate the functionality of each antibody probe, representative plots illustrating the expected
Supplementary Figure 1 Examples of staining for each antibody used for the mass cytometry analysis. To illustrate the functionality of each antibody probe, representative plots illustrating the expected
Part III Innate and Adaptive Immune Cells: General Introduction
 Innate and Adaptive Immune Cells: General Introduction Iván López-Expósito As an organ specialized in food digestion and nutrient absorption, the intestinal mucosa presents a huge surface area (almost
Innate and Adaptive Immune Cells: General Introduction Iván López-Expósito As an organ specialized in food digestion and nutrient absorption, the intestinal mucosa presents a huge surface area (almost
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
M.Sc. III Semester Biotechnology End Semester Examination, 2013 Model Answer LBTM: 302 Advanced Immunology
 Code : AS-2246 M.Sc. III Semester Biotechnology End Semester Examination, 2013 Model Answer LBTM: 302 Advanced Immunology A. Select one correct option for each of the following questions:- 2X10=10 1. (b)
Code : AS-2246 M.Sc. III Semester Biotechnology End Semester Examination, 2013 Model Answer LBTM: 302 Advanced Immunology A. Select one correct option for each of the following questions:- 2X10=10 1. (b)
Additional file 6. Summary of experiments characterizing development of adaptive immune response
 Additional file 6. Summary of experiments characterizing development of adaptive immune response Tests performed to evaluate the development of adaptive immunity in patients are summarized in Table 1.
Additional file 6. Summary of experiments characterizing development of adaptive immune response Tests performed to evaluate the development of adaptive immunity in patients are summarized in Table 1.
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured
 Naïve CD4 + T cells were cultured") Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Evaluation of directed and random motility in microslides Assessment of leukocyte adhesion in flow chambers
 Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
MagniSort Mouse CD4 Naive T cell Enrichment Kit Catalog Number: RUO: For Research Use Only. Not for use in diagnostic procedures.
 Page 1 of 2 MagniSort Mouse CD4 Naive T cell Enrichment Kit RUO: For Research Use Only. Not for use in diagnostic procedures. Mouse splenocytes were unsorted (left) or sorted with the MagniSort Mouse CD4
Page 1 of 2 MagniSort Mouse CD4 Naive T cell Enrichment Kit RUO: For Research Use Only. Not for use in diagnostic procedures. Mouse splenocytes were unsorted (left) or sorted with the MagniSort Mouse CD4
Interferon γ regulates idiopathic pneumonia syndrome, a. Th17 + CD4 + T-cell-mediated GvH disease
 Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Effector T Cells and
 1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
Antigen Presentation and T Lymphocyte Activation. Abul K. Abbas UCSF. FOCiS
 1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
Pair-fed % inkt cells 0.5. EtOH 0.0
 MATERIALS AND METHODS Histopathological analysis Liver tissue was collected 9 h post-gavage, and the tissue samples were fixed in 1% formalin and paraffin-embedded following a standard procedure. The embedded
MATERIALS AND METHODS Histopathological analysis Liver tissue was collected 9 h post-gavage, and the tissue samples were fixed in 1% formalin and paraffin-embedded following a standard procedure. The embedded
TCR, MHC and coreceptors
 Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
COMPONENT NAME COMPONENT # QUANTITY STORAGE SHELF LIFE FORMAT. Store at 2-8 C. Do not freeze. Store at 2-8 C. Do not freeze.
 This document is available at www.stemcell.com/pis EasySep Mouse Monocyte Isolation Kit Catalog #19861 For processing 1 x 10^9 cells Description Isolate untouched and highly purified monocytes from mouse
This document is available at www.stemcell.com/pis EasySep Mouse Monocyte Isolation Kit Catalog #19861 For processing 1 x 10^9 cells Description Isolate untouched and highly purified monocytes from mouse
Nature Immunology: doi: /ni Supplementary Figure 1. Production of cytokines and chemokines after vaginal HSV-2 infection.
 Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
Therapeutic effect of baicalin on experimental autoimmune encephalomyelitis. is mediated by SOCS3 regulatory pathway
 Therapeutic effect of baicalin on experimental autoimmune encephalomyelitis is mediated by SOCS3 regulatory pathway Yuan Zhang 1,2, Xing Li 1,2, Bogoljub Ciric 1, Cun-gen Ma 3, Bruno Gran 4, Abdolmohamad
Therapeutic effect of baicalin on experimental autoimmune encephalomyelitis is mediated by SOCS3 regulatory pathway Yuan Zhang 1,2, Xing Li 1,2, Bogoljub Ciric 1, Cun-gen Ma 3, Bruno Gran 4, Abdolmohamad
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
B6/COLODR/SPL/11C/83/LAP/#2.006 B6/COLODR/SPL/11C/86/LAP/#2.016 CD11C B6/COLODR/SPL/11C/80/LAP/#2.011 CD11C
 CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
The development of T cells in the thymus
 T cells rearrange their receptors in the thymus whereas B cells do so in the bone marrow. The development of T cells in the thymus The lobular/cellular organization of the thymus Immature cells are called
T cells rearrange their receptors in the thymus whereas B cells do so in the bone marrow. The development of T cells in the thymus The lobular/cellular organization of the thymus Immature cells are called
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Cell-mediated Immunity
 Cellular & Molecular Immunology Cell-mediated Immunity Nicholas M. Ponzio, Ph.D. Department of Pathology & Laboratory Medicine April 6, 2009 Today s Presentation: Overview Cellular Interactions In Humoral
Cellular & Molecular Immunology Cell-mediated Immunity Nicholas M. Ponzio, Ph.D. Department of Pathology & Laboratory Medicine April 6, 2009 Today s Presentation: Overview Cellular Interactions In Humoral
In vitro human regulatory T cell suppression assay
 Human CD4 + CD25 + regulatory T cell isolation, in vitro suppression assay and analysis In vitro human regulatory T cell suppression assay Introduction Regulatory T (Treg) cells are a subpopulation of
Human CD4 + CD25 + regulatory T cell isolation, in vitro suppression assay and analysis In vitro human regulatory T cell suppression assay Introduction Regulatory T (Treg) cells are a subpopulation of
Immune response to infection
 Immune response to infection Dr. Sandra Nitsche (Sandra.Nitsche@rub.de ) 20.06.2018 1 Course of acute infection Typical acute infection that is cleared by an adaptive immune reaction 1. invasion of pathogen
Immune response to infection Dr. Sandra Nitsche (Sandra.Nitsche@rub.de ) 20.06.2018 1 Course of acute infection Typical acute infection that is cleared by an adaptive immune reaction 1. invasion of pathogen
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Darwinian selection and Newtonian physics wrapped up in systems biology
 Darwinian selection and Newtonian physics wrapped up in systems biology Concept published in 1957* by Macfarland Burnet (1960 Nobel Laureate for the theory of induced immune tolerance, leading to solid
Darwinian selection and Newtonian physics wrapped up in systems biology Concept published in 1957* by Macfarland Burnet (1960 Nobel Laureate for the theory of induced immune tolerance, leading to solid
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Technical Resources. BD Immunocytometry Systems. FastImmune Intracellular Cytokine Staining Procedures
 FastImmune Intracellular Cytokine Staining Procedures BD has developed protocols for the detection of intracellular cytokines in activated lymphocytes and in activated monocytes. The procedures have been
FastImmune Intracellular Cytokine Staining Procedures BD has developed protocols for the detection of intracellular cytokines in activated lymphocytes and in activated monocytes. The procedures have been
Frontiers of Science Foundation Scholar. Toby Bothwell. Freshman, University of Arkansas. final research manuscripts
 Frontiers of Science Foundation Scholar Toby Bothwell Oklahoma city, OKlahoma Freshman, University of Arkansas final research manuscripts The Effect of Estradiol and Estrogen Receptors α and β on Bone
Frontiers of Science Foundation Scholar Toby Bothwell Oklahoma city, OKlahoma Freshman, University of Arkansas final research manuscripts The Effect of Estradiol and Estrogen Receptors α and β on Bone
5/1/13. The proportion of thymus that produces T cells decreases with age. The cellular organization of the thymus
 T cell precursors migrate from the bone marrow via the blood to the thymus to mature 1 2 The cellular organization of the thymus The proportion of thymus that produces T cells decreases with age 3 4 1
T cell precursors migrate from the bone marrow via the blood to the thymus to mature 1 2 The cellular organization of the thymus The proportion of thymus that produces T cells decreases with age 3 4 1
Pearson r = P (one-tailed) = n = 9
 = n = 9") 8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained
 Representative pictures of H&E-stained") 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
Fluorochrome Panel 1 Panel 2 Panel 3 Panel 4 Panel 5 CTLA-4 CTLA-4 CD15 CD3 FITC. Bio) PD-1 (MIH4, BD) ICOS (C398.4A, Biolegend) PD-L1 (MIH1, BD)
 PD-1 (MIH4, BD) ICOS (C398.4A, Biolegend) PD-L1 (MIH1, BD)") Additional file : Table S. Antibodies used for panel stain to identify peripheral immune cell subsets. Panel : PD- signaling; Panel : CD + T cells, CD + T cells, B cells; Panel : Tregs; Panel :, -T, cdc,
Additional file : Table S. Antibodies used for panel stain to identify peripheral immune cell subsets. Panel : PD- signaling; Panel : CD + T cells, CD + T cells, B cells; Panel : Tregs; Panel :, -T, cdc,
Sipper BK Experimental Animal Co. (Shanghai, China) and bred in a specific. pathogen-free environment. The animal study protocol was approved by the
 and bred in a specific. pathogen-free environment. The animal study protocol was approved by the") Supplementary information, Data S1 Materials and Methods Mice, Ad vectors and reagents Female C57BL/6 mice, 8-10 weeks of age, were purchased from Joint Ventures Sipper BK Experimental Animal Co. (Shanghai,
Supplementary information, Data S1 Materials and Methods Mice, Ad vectors and reagents Female C57BL/6 mice, 8-10 weeks of age, were purchased from Joint Ventures Sipper BK Experimental Animal Co. (Shanghai,
Manipulating the Tumor Environment
 Manipulating the Tumor Environment Vincenzo Bronte Verona University Hospital vincenzo.bronte@univr.it Escape from immune control can be viewed as one of the «Hallmarks of Cancer» D. Hanahan and R. A.
Manipulating the Tumor Environment Vincenzo Bronte Verona University Hospital vincenzo.bronte@univr.it Escape from immune control can be viewed as one of the «Hallmarks of Cancer» D. Hanahan and R. A.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
Pretargeting and Bioorthogonal Click Chemistry-Mediated Endogenous Stem Cell Homing for Heart Repair
 Pretargeting and Bioorthogonal Click Chemistry-Mediated Endogenous Stem Cell Homing for Heart Repair Mouse Model of Myocardial Infarction (MI) All animal work was compliant with the Institutional Animal
Pretargeting and Bioorthogonal Click Chemistry-Mediated Endogenous Stem Cell Homing for Heart Repair Mouse Model of Myocardial Infarction (MI) All animal work was compliant with the Institutional Animal
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity
 Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Supplementary Materials for
 www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
Regulation of anti-tumor immunity through migration of immune cell subsets within the tumor microenvironment Thomas F. Gajewski, M.D., Ph.D.
 Regulation of anti-tumor immunity through migration of immune cell subsets within the tumor microenvironment Thomas F. Gajewski, M.D., Ph.D. Professor, Departments of Pathology and Medicine Program Leader,
Regulation of anti-tumor immunity through migration of immune cell subsets within the tumor microenvironment Thomas F. Gajewski, M.D., Ph.D. Professor, Departments of Pathology and Medicine Program Leader,
Alternatively Activated Macrophages Determine the Repair of the Infarcted
 Alternatively Activated Macrophages Determine the Repair of the Infarcted Adult Murine Heart (Shiraishi et al.) List of Supplemental Materials Supplemental Methods Supplemental Figure 1. Cardiac CD206
Alternatively Activated Macrophages Determine the Repair of the Infarcted Adult Murine Heart (Shiraishi et al.) List of Supplemental Materials Supplemental Methods Supplemental Figure 1. Cardiac CD206
Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells
 ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
Supplemental Information. Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection. Cell Host & Microbe, Volume 15
 Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
well for 2 h at rt. Each dot represents an individual mouse and bar is the mean ±
 Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY
 ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through