The Role of Adult Hippocampal Neurogenesis in Reducing Interference
|
|
|
- Claud Burke
- 5 years ago
- Views:
Transcription
1 Behavioral Neuroscience 212 American Psychological Association 212, Vol. 126, No. 3, /12/$12. DOI: 1.137/a28252 The Role of Adult Hippocampal Neurogenesis in Reducing Interference Paul Luu University of Toronto Lulu Gao University of Toronto Jan Martin Wojtowicz University of Toronto Orriana C. Sill Cornell University Suzanna Becker McMaster University David M. Smith Cornell University Adult neurogenesis in the hippocampal dentate gyrus plays an important role in learning and memory. However, the precise contribution of the new neurons to hippocampal function remains controversial. Emerging evidence suggests that neurogenesis is important for pattern separation and for mitigating interference when similar items must be learned at different times. In the present study, we directly test this prediction using a recently developed olfactory memory task that has those specific features. In this task, rats learn two highly interfering lists of odor pairs, one after the other, in either the same or in different contexts. Consistent with our hypothesis, focal cranial irradiation, resulting in selective reduction of neurogenesis within the dentate gyrus, significantly impaired the ability to overcome interference during learning of the second list. The ability to learn a single odor list was unimpaired. We also show that irradiation had no effect on learning in a hippocampal-dependent spatial alternation task. Although both tasks involved learning interfering responses, the time course for learning the interfering items differed. Learning the interfering odor lists took place sequentially, over the course of several sessions, whereas learning the interfering spatial locations took place concurrently, within each session. Thus, the gradual addition of new neurons may have provided a pattern separation mechanism for the olfactory task but not for the maze task. These findings demonstrate a role for neurogenesis in resolving interference and they are consistent with models suggesting a critical role for neurogenesis in pattern separation. Keywords: hippocampus, memory interference, adult neurogenesis, learning and memory, dentate gyrus Adult neurogenesis has emerged as an integral and crucial process within the hippocampus. Due to their strategic location in the dentate gyrus, new neurons are thought to play a key role in learning and memory, as well as in some related hippocampal functions, such as the regulation of drug addiction, emotions, and stress (Becker & Wojtowicz, 7; Deng, Aimone, & Gage, Paul Luu, Lulu Gao, and Jan Martin Wojtowicz, Department of Physiology, University of Toronto; Orriana C. Sill and David M. Smith, Department of Psychology, Cornell University; Suzanna Becker, Department of Psychology, Neuroscience and Behaviour, McMaster University. Paul Luu and Orriana C. Sill contributed equally to this project. This work was supported by National Institutes of Health grant MH8389 to David M. Smith; a Canadian Institutes of Health Research operating grant to Jan Martin Wojtowicz and Natural Sciences and Engineering Research Council discovery grant and Discovery Accelerator Supplement to Suzanna Becker. Technical assistance by Ms. Yao Fang Tan is greatly appreciated. Dr. Howard Dobson and Ms. Kim Stewart provided expert help with rat irradiation procedure. Correspondence concerning this article should be addressed to David M. Smith, Department of Psychology, 252 Uris Hall, Cornell University, Ithaca, NY dms248@cornell.edu 21; Jacobs, van Praag, & Gage, ; Noonan, Bulin, Fuller, & Eisch, 21; Schoenfeld & Gould, 211). Computational studies predict a role for new neurons in pattern separation and interference reduction on the basis of unique properties of these cells (Becker, 5). One such property is the ability of new neurons to undergo a complete turnover while they grow and become transformed from one developmental stage to another during the course of several days to weeks (Deng et al., 21). Behavioral tasks that included pattern separation have already been exploited in experiments (Clelland et al., 9; Creer, Romberg, Saksida, van Praag, & Bussey, 21). These studies have shown that mice with reduced neurogenesis are impaired on such tasks. Paradoxically, Saxe et al. (7) observed improvement in a working memory task involving pattern separation. The discrepancy may be accounted for by the reliance upon different memory systems in these tasks, with the former (Clelland et al., 9; Creer et al., 21) being hippocampal dependent, whereas the latter task (Saxe et al., 7) may rely more on extrahippocampal working memory circuits in the prefrontal cortex. Nonetheless, all these tasks have in common that they require the animal to represent and separate events occurring almost simultaneously, or interleaved within a single experimental test session. 381
2 382 LUU, SILL, GAO, BECKER, WOJTOWICZ, AND SMITH In contrast, Aimone, Wiles, and Gage (6), and Becker and Wojtowicz (Becker, 5; Becker & Wojtowicz, 7) proposed another form of pattern separation, for events separated by days, specifically related to turnover of new cells. Our model predicts that hippocampal neurogenesis should be critical when subjects must form two distinct memories for highly interfering items, as long as the two learning experiences occur at different times, so that distinct populations of new neurons are available for the encoding of each item. In the present study, we directly test this prediction using a recently developed task that has those specific features (Butterly, Petroccione, & Smith, 212). In this task, rats learn two highly interfering lists of odor pairs, one after the other in different contexts. For comparison, we also examined the role of neurogenesis in another hippocampal-dependent task that we have used previously (blocked spatial alternation, Smith & Mizori, 6b) and that also involved learning interfering responses, but within the same session. Thus, the present study examined the role of adult neurogenesis in pattern separation and memory interference within and across experimental sessions. In addition, recognizing a dynamic, reciprocal relationship between learning and neurogenesis, we deployed a battery of tests to estimate the nber of young neurons and their rate of proliferation and survival in relation to behavioral performance. Method Animals and Time Line of the Study Fifty-four male Long-Evans rats were obtained from Charles River (Quebec) in five batches, at approximately every two months. The animals were 3 months old on arrival and were kept in the animal facility at Guelph University (Ontario, Canada) for 1 week prior to any procedures. Rats in the irradiated group were anesthetized and underwent procedures for cranial irradiation at the Guelph Veterinary Clinic adjacent to the animal facility, as described previously (Winocur, Wojtowicz, Sekeres, Snyder, & Wang, 6). rats were anesthetized, but were not exposed to irradiation. Ten rats (six controls and four irradiated) served as untrained cage controls, and were kept in small animal cages at the University of Toronto for the duration of the experiment. They received injections of bromodeoxyuridine (BrdU) at 4 weeks prior to the perfusion, in synchrony with the trained rats. The remaining 44 irradiated and nonirradiated animals were transported by air to Cornell University (Ithaca, NY) for all behavioral tests and kept there until the experiments were completed. Behavioral training began 5 weeks after the irradiation. The rats (n 44) were first trained on the olfactory discrimination task, which lasted approximately three weeks. After completing the olfactory discrimination task, 1 of the rats (five controls, five irradiated) were given injections of BrdU at mg/kg ip and perfused 1 week later in order to determine whether olfactory training affects the subsequent production of new neurons. One week after completing the olfactory task, 32 of the rats (eight rats in each of four groups, described below) began training on the plus-maze task, which took up to 15 days to complete. One week prior to the plus-maze task, the rats were injected with a single dose of BrdU at mg/kg ip in order to determine whether plus-maze training affects the survival of new neurons. The timing of the BrdU injection was planned specifically to detect changes in neuronal survival of 1 2-week-old neurons during the plus-maze task as described previously for other hippocampal-dependent tasks (Sisti, Glass, & Shors, 7; Tronel et al., 21). Because the olfactory discrimination task occurred prior to the BrdU injection, we did not expect the neuronal survival of the BrdU-positive cells to be affected by the olfactory training. The animals health was monitored throughout the duration of the study. The weights of the control and irradiated rats were monitored and did not differ [control , mean standard error of the mean (SEM); irradiated , at the outset of training, t (24).31, p.41]. The animals were perfused exactly 25 days after BrdU injection, regardless of how quickly they reached the criterion in the plus-maze task. Perfusion, Sectioning, and Sampling Each animal was deeply anesthetized with isoflurane and then intracardially perfused with 3 ml.1 M phosphate buffered saline (PBS, ph 7.4) followed by ml cold 4% paraformaldehyde (PFA, ph 7.4, 4 C) in PBS. The brain was carefully removed from the skull and placed in 4% PFA at 4 C for 24 hours. Later, the brain was stored in.1% sodi azide in PBS until sectioning. The right hippocampus from each animal was carefully isolated and coronally sectioned at 3- m thickness using a vibratome. Twelve sections were sampled evenly across the whole length of the hippocampus and stained for several markers of neurogenesis as described below. In a subset of animals (five controls, five irradiated), the left hemisphere was sectioned coronally at 4 m. Two sections from several regions of the brain were sampled representing the olfactory bulb (OB), the rostral migratory stream (RMS), and the subventricular zone (SVZ). Stereotaxic coordinates were 6.7 mm, 4.2 mm, 3.2 mm,.7 mm, and.92 mm (Paxinos & Watson, 1998). Calci-Binding Protein/BrdU Immunohistochemistry Double labeling of BrdU-positive cells with calci-binding protein (CaBP) was used to identify newly born dentate granule cells. The hippocampal sections were incubated with 1 N hydrochloric acid for 3 min at 45 C followed by three 5-min washes. The sections were incubated with anti-brdu primary antibody (rat, 1: in.3% Triton-X PBS, Serotec) for 24 hr at 4 C, followed by three 5-min washes. The sections were incubated with the secondary antibody (rabbit anti-rat IgG Alexa Fluor 488, 1: in.3% Triton-X PBS, Molecular Probes) for 2 hr at room temperature, followed by three 5-min washes. Then, the sections were incubated with anti-calbindin primary antibody (rabbit, 1: in.3% Triton-X PBS, Chemicon) for 72 hr at 4 C, followed by three 5-min washes. The sections were incubated with the secondary antibody (goat anti-rabbit IgG Alexa Fluor 568, 1: in.3% Triton-X PBS, Molecular Probes) for 2 hr at room temperature. Finally, the sections were washed three times and mounted on slides with mounting medi (Fluoromount, Sigma). Doublecortin Immunohistochemistry Doublecortin (DCX) labeling was used to identify recently born neurons. Free-floating sections were incubated with a primary goat
3 HIPPOCAMPAL NEUROGENESIS AND INTERFERENCE 383 anti-dcx antibody (1:, Santa Cruz Biotechnology, 24 hr at 4 C), followed by Alexa488 donkey anti-goat secondary antibody (1:; Invitrogen; 2 hr at room temperature). All antibodies were diluted in PBS containing.3% Triton X-1. Ki67 Immunohistochemistry Ki67 labeling was used to observe cell proliferation at the time of perfusion. Sections were incubated with anti-ki67 primary antibody (rabbit, 1: in.3% Triton-X PBS, Vector Laboratories) for 18 hr at room temperature, followed by three 5-min washes. Then, the sections were incubated with secondary antibody (goat anti-rabbit IgG Alexa Fluor 568, 1: in.3% Triton-X PBS, Molecular Probes) for 2 hr at room temperature, followed by three 5-min washes. Finally, the sections were washed three times and mounted on slides with mounting medi (Fluoromount, Sigma). Cell Counting In hippocampal sections, the single-labeled cells were counted under a fluorescent microscope (4 ) in the subgranular zone, excluding the upper and lower edges of the sections. The doublelabeled cells (CaBP and BrdU) were counted using a confocal microscope (Leica). The total nber of cells (per dentate gyrus) was obtained by multiplying the average nber of cells per section by the total nber of sections, in each animal. In coronal sections used for estimates of cell nber in the SVZ and RMS, cells expressing BrdU were counted within the region covered by DCX-positive cells. In the OB, BrdU-positive cells were counted within four square visual fields (5 5 m) located within the area covered by NeuN-positive cells. All counts were done on a fluorescent microscope. The cells nbers are given as densities (per mm 2 of area examined). General Behavioral Methods and Rationale We trained rats on two different behavioral tasks that we have used previously and that have been shown to be hippocampal dependent in our laboratory (Butterly et al., 212; Smith & Mizori, 6a). In Experiment 1, rats learned two lists of interfering odor pairs. They learned the first list over the course of several daily training sessions, followed by training on the second list during subsequent training days. In Experiment 2, the rats were trained to remember and approach two different reward locations on a plus maze. Rewards were placed on the east arm for the first half of each training session and on the west arm for the second half, so the two reward locations were learned concurrently. Experiment 1: Olfactory Discrimination Task Subjects were 44 adult male Long-Evans rats. Prior to training, the rats were placed on a restricted feeding regimen (8 85% of free-feeding weight). The rats were trained to dig in cups of odorized bedding material to retrieve buried food rewards (45 mg sucrose pellets, Bioserve, Inc., Frenchtown, NJ). All of the rats were first trained on one list of odor pairs. They were then given training on a second list of odor pairs either in the same context or a different context, yielding a 2 2 design with irradiation condition (control or irradiated) and context condition (same or different) as factors. One rat was excluded due to experimenter error in the training procedure, resulting in 11 rats in each group, except the control different condition which had 1 rats. The two contexts differed along the following dimensions: color of the chamber (white or black), color of surrounding area (either an open experimental room with black painted walls or a 6 8 feet area enclosed by white plastic blinds), substrate in the chamber (uncovered Plexiglas floor or a black rubber mat), the 65-dB continuous background-masking noise (white noise or pink noise) and the ambient odor left by wiping out the chamber with baby wipes (unscented or scented, Rite Aid, Inc.) prior to each training session. The rats were trained in Plexiglas chambers (45 cm wide 6 cm long 4 cm deep) equipped with a removable divider, which separated the odor presentation area from an area where the rats waited during the intertrial interval (ITI). Odor cues were presented in ceramic cups (8.25 cm in diameter, 4.5 cm deep). The digging cups fit into circular cutouts cemented to the floor of the chamber to discourage the rats from moving the cups or tipping them over. Twenty-four pure odorants served as cues (for details see Butterly et al., 212). Briefly, the amount of each odorant was calculated so that it produced an equivalent vapor phase-partial pressure when mixed with 5 ml of mineral oil (Clelland et al., 9). Ten ml of each odorant solution was then mixed with 2 L of corncob bedding material and stored in covered containers. Prior to training, the rats were acclimated to each of the two contexts for two 1-min sessions in order to control for possible effects of novelty on neurogenesis. The rats were then shaped to dig in cups of bedding to retrieve buried rewards. After the rats had learned to reliably retrieve the rewards from the bottom of the cups, they began training on the first of two lists of odor pairs. Each list contained eight odor pairs (16 individual odors). The two odors comprising each pair were always presented together, in separate cups. Within each odor pair, one odor always contained a buried reward and the other did not. The predictive value of the odors (rewarded or nonrewarded) was counterbalanced across subjects and their presentation locations for each trial (left or right side of the chamber) were randomized. The daily training sessions consisted of 64 trials (eight trials with each odor pair, presented in an unpredictable sequence). At the start of each trial, the experimenter placed the two cups containing the odorized bedding in the assigned locations (left or right) and removed the divider so that the rat could approach the cups. The rat was allowed to dig until he retrieved the reward. A digging response was recorded if the rat displaced any of the bedding, except when stepping into the cup without investigating. After consing the reward, the rat was returned to the waiting area and the divider was replaced. During an ITI of approximately 1 s, the experimenter prepared the cups for the next trial. The rats were given daily training sessions on List 1 until they reached a behavioral criterion of 9% correct choices on two consecutive sessions. After reaching the criterion, the rats were given five training sessions on a second list of eight odor pairs. The rats were trained in either the same context where they learned the first list or in a different context. The training sessions for List 2 were carried out
4 384 LUU, SILL, GAO, BECKER, WOJTOWICZ, AND SMITH in the same manner as the List 1 training sessions, except that the second list contained eight new odor pairs. In order to induce high levels of interference between the two lists, each of the new odor pairs for List 2 consisted of a novel odor and an odor which had previously been presented in List 1. Of the eight odors taken from List 1, half had been rewarded previously and half had not. For example, if the first two odor pairs on List 1 were A /B and C /D, the first two odor pairs on List 2 would be X /A and D /Y. This ensured that the rats could not adopt a strategy of simply approaching the novel odor (or avoiding the familiar odor) within each new odor pair. Previous studies indicated that the rats could not smell the buried rewards (Butterly et al., 212). Nevertheless, a subset of the rats (n 18) were tested to ensure that they were not able to directly detect the pellets. After the completion of training, the rats were given a session consisting of 24 trials (three trials with each of the eight rewarded odors from List 2). On each trial, the rats were presented with two cups containing the same odor. However, only one of the cups was baited. If the rats could directly detect the pellets, they would be expected to perform better than chance (5%). The rats chose the baited cup 49.53% of the time, which did not differ from chance, t (17).265, p.8. The long-term effects of the olfactory task on neurogenesis were measured by estimating the nber of proliferating, Ki-67-positive progenitors and immature, DCX-positive neurons. Experiment 2: Plus-Maze Task The subjects were 32 adult male Long-Evans rats which had previously been trained in the olfactory discrimination task for Experiment 1. The rats were trained to approach the east arm of a plus maze for reward during the first half of each training session and to approach the west arm during the second half. In this experiment, we sought to determine whether suppression of neurogenesis would impair learning in this task and whether training on this task would increase the survival of new neurons, as has been reported in other hippocampal-dependent tasks (Epp, Haack, & Galea, 211; Sisti et al., 7; Tronel et al., 21). To this end, all of the rats were given a single injection of BrdU ( mg per kg ip) 7 days prior to beginning training. In order to control for exercise, handling, and exposure to the rewards, the rats were divided into two groups. One group received regular training sessions and the other served as a yoked control group in which each rat was given the same nber of sessions as a trained rat, but they were given control sessions that did not permit learning about predictable reward locations (described below). If learning to discriminate the go-east and go-west trials induced neurogenesis that was greater than that seen in the yoked controls, then the neurogenesis could not be attributed to factors other than learning. The rats were trained on a plus maze (12 cm across) that occupied a circular area enclosed by curtains (3 m in diameter). Distinctive objects were attached to the curtain to serve as distal visual cues. Prior to training, they were given several sessions in which they were acclimated to the plus maze and trained to retrieve chocolate milk rewards (.2 ml Nestle s Quik) from cups at the ends of the maze arms. The rats in the trained condition were then given daily training trials consisting of two blocks of 15 trials each. During the first block of every training session, the reward was always placed at the end of the east arm. During the second block, the reward was always placed at the end of the west arm. Trials began when the rat was placed on the maze facing outward at the end of an arm and ended when the rat arrived at the reward. During an ITI of approximately 2 s, the rats were placed on a platform adjacent to the maze. The position of the ITI platform was constant throughout training. The start positions for each trial were randomly designated from among the three nonreward arms. Training continued with the same two reward locations presented each day until the rats attained a behavioral criterion of at least 75% correct choices on two consecutive sessions. Training was discontinued if the rat did not achieve this criterion in 15 days. Each rat in the yoked control condition was given the same nber of sessions as his trained counterpart. The training trials for the yoked control rats were identical to those described above, except that instead of placing the rewards in predictable locations (i.e., the east and west arms, as above), the rewards were placed on randomly designated arms. rats choose the rewarded arm on 64.3% of the trials, on average, over all training sessions. The training procedures for the yoked controls were designed to approximate this as closely as possible. On each trial, two randomly designated arms were baited (although the rat was only allowed to run until he found one of the rewards). With two of the three nonstart arms baited, the rats pick a rewarded arm 66.6% of the time by chance. Additionally, analysis of the total nber of arm entries confirmed that trained rats and yoked controls ran similar distances on the maze, F(1, 28).71, p.41. Thus, the yoked control rats were given the same nber of training trials and rewards, but they could not learn to remember and approach two different predictable reward locations. Results Effects of Irradiation on Neurogenesis Neurogenesis was selectively reduced in the dentate gyrus (DG) but not in the olfactory tract at 9 weeks and 14 weeks after irradiation (end of the study). A comparison of cell densities at 9 weeks (n 5 per group) and at 14 weeks (n 16 per group) showed a significant effect of irradiation, DCX, F(1, 41) , p.1, but no main effect of time, F(1, 41) 3.82, p.58. The average reduction of neurogenesis after 14 weeks, as measured by the nber of DCX new neurons, was 85%, t (3) 9.29, p.1. In contrast, there was no effect of irradiation on neurogenesis in the olfactory tract. The density of BrdU cells counted in five regions of the olfactory system was not affected by the irradiation procedures [analysis of variance (ANOVA) with irradiation condition and location showed no effect of irradiation, F(1, 11).331, p.556, and no interactions of the irradiation and location factors, Figure 1]. Experiment 1: Olfactory Discrimination Task Behavior. As expected, suppression of neurogenesis did not impair learning of the first list of odors. and irradiated rats did not differ in the nber of training sessions needed to reach the criterion [irradiated mean (M) 4.41 sessions, control M 4.14 sessions, t (41) 1.6, p.3] and there were no differences between control and irradiated rats in terms of their performance
5 HIPPOCAMPAL NEUROGENESIS AND INTERFERENCE 385 on the final training session of List 1, F(1, 39) 2.5, p.45. Importantly, these results indicate that the irradiated rats did not have a general impairment in olfactory sensory processing or olfactory learning. However, suppression of neurogenesis did cause a significant impairment in performance on the second list. The percent correct on each day of training were submitted to a repeated-measures ANOVA with training session (5 days of training on List 2) as the within-subject factor and irradiation condition (control or irradiated) and context condition (same or different) as between subjects factors (see Figure 2). This analysis revealed a significant main effect of training session, F(1, 39) 12.76, p.1, a significant main effect of irradiation, F(1, 39) 12.76, p.1, with controls performing significantly better than irradiated subjects, and a main effect of context, F(1, 39) 4.9, p.5, with subjects in the different-context condition performing significantly better than subjects in the same-context condition. However, there was no significant interaction of the context and irradiation conditions, F(1, 39) 1.52, p.23. This result suggests that the suppression of neurogenesis impaired performance regardless of the context condition, in contrast to our previous findings that muscimol lesions of the dorsal hippocampus selectively impaired performance in the different-context condition but not in the samecontext condition (Butterly et al., 212). Whereas the muscimol lesions had no impact on performance in the same-context condition, the suppression of neurogenesis may have had more widespread effect on performance, including impairment in the samecontext condition. This may have occurred because neurogenesis was suppressed throughout the hippocampus, whereas the muscimol lesions were specific to the dorsal hippocampus. We revisit this issue in the general Discussion. Assessment of Interference in each condition. Our hypothesis was that new neurons play a beneficial role in learning because they provide a means of overcoming interference for items learned at different times. The effects of interference can be assessed by comparing performance on the two lists. When there is little opportunity for interference (e.g., when learning noninterfering material), performance on the second list is facilitated by prior experience on the first list (Butterly et al., 212). However, if proactive interference occurs, performance on the second list will not be facilitated and can even be impaired by prior learning on the first list. Since interference is typically most pronounced during the initial stages of learning, we compared performance during the first three sessions of List 2 to performance during the same sessions of List 1 (see Figure 3). A similar pattern of results was seen when all five sessions of List 2 were analyzed. rats that learned the two lists in different contexts performed significantly better on the second list than on the first, paired-samples t test: t (9) 3.96, p.5. That is, when contextual information was available to disambiguate the two conflicting lists, control rats did not experience interference and performance was facilitated on the second list. In contrast, control rats that learned the two lists in the same context showed no such facilitation [i.e., no significant change in performance from List 1 to List 2, t (1) 1.7, p.12]. rats were not able to use contextual information to disambiguate the two lists and performance was not facilitated on List 2, t (1).61, p.56. It was interesting that irradiated rats in the same-context condition showed an even greater evidence of interference, as performance was significantly worse on List 2 than List 1, t (1) 2.33, p.5. Measures of neurogenesis for the rats in the olfactory discrimination experiment were subjected to an ANOVA with irradiation condition and training condition (trained or untrained cage controls) as factors (see Figure 4). Rats in the same-context and different-context training groups did not differ in terms of neurogenesis, e.g., DCX, F(1, 37).72, p.4, so these groups were combined for these analyses. Although there was a main effect of irradiation, DCX-labeled cells, F(1, 39) 65.22, p.1, there was no effect of training condition DCX, F(1, 37).6, p.82, nor was there an interaction of the irradiation and training condition factors, F(1, 37) 2.846, p.1. Analysis of the Ki67- positive cells revealed similar results. Thus, the olfactory learning did not have long-term effects on levels of neuronal production in terms of proliferation or neuronal differentiation. Experiment 2: Plus-Maze Task Behavior. In contrast to the odor task, irradiated rats showed no evidence of impairment on the plus-maze task. and irradiated rats did not differ in terms of the nber of sessions needed to reach the behavioral criterion, control M 7.5 sessions, irradiated M 9. sessions, t (16).614, p.55, Figure 5a. Two control rats and three irradiated rats failed to reach the criterion within the 15 sessions that were given. and irradiated rats also exhibited similar levels of performance throughout training. To assess this, the percentage of trials with a correct response were submitted to a repeated-measures ANOVA with group (control and irradiated) as a between-subjects factor and training stage as a within-subjects factor (three stages, including the first, middle, and last training sessions). The rats took a variable nber of training sessions to reach the criterion, so the middle training session was simply the session that was half way between the first and last session. For those rats that received an even nber of sessions, the average of the two middle-most sessions was used. For example, for a rat that required 12 sessions to reach the criterion, the average performance on Sessions 6 and 7 was used as the middle session. This analysis showed no difference between groups, F(1, 14).16, p.69, and no interaction of the group and training stage variables, F(1, 28).75, p.48, Figure 5b. The latency to reach the reward and various measures of inflexible behavioral responding (e.g., right or left turn biases) were also assessed and no group differences were found. Effects of maze training on neurogenesis and survival. Neurogenesis was compared among all animals participating in the plus maze during weeks 9 14 of the study. There was a clear effect of irradiation, but none of the markers (BrdU, DCX, Ki67) revealed any differences between trained rats, yoked controls, and cage controls. This lack of differences held for both nonirradiated and irradiated animals (see Figure 6). Measures of neurogenesis (BrdU, DCX, Ki67) for the rats trained in the plus-maze task were analyzed by ANOVA with irradiation condition and training condition (three groups: trained, yoked controls, and untrained caged controls) as factors. One irradiated and three nonirradiated animals were excluded from immunohistochemical analysis due to poor fixation of the tissue. There was a significant main effect of irradiation on the survival of BrdU cells born 1 week prior to the beginning of learning, F(1,





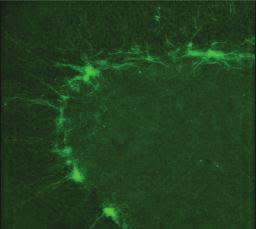









6 386 LUU, SILL, GAO, BECKER, WOJTOWICZ, AND SMITH A OB RMS SVZ IRR CON Radiation B DG DCX CaBP BrdU CaBP/BrdU CON IRR C D E * # DCX+ cells/dg * # BrdU/CaBP+ cells/dg IRR BrdU + cells/mm Figure 1 (opposite).
7 HIPPOCAMPAL NEUROGENESIS AND INTERFERENCE 387 A Correct (%) B Correct (%) // Last Day 1 Day 2 Day 3 Day 4 Day 5 Different Same Different -Different -Same -Different -Same Same Figure 2. Olfactory discrimination performance. In Panel A, the average percent correct choices are shown for control (open symbols) and irradiated rats (filled symbols) and for the different-context (solid lines) and samecontext conditions (dashed lines). Performance data are shown for the final session of List 1 training (Last) and the five training sessions of List 2. In Panel B, the same data are shown averaged across all 5 days of training on List 2. 4) 4.354, p.1, but there was no effect of training condition, F(1, 4).311, p.51, nor was there an interaction of irradiation condition and training condition, F(1, 4).162, p.162. Similar results were seen in the rate of maturation, as indicated by CaBP/BrdU labeling, main effect of irradiation: F(1, 37) , p.1, all others ns, and in DCX-labeled cells, main effect of irradiation: F(1, 39) , p.1, all others ns. Discussion Suppression of hippocampal neurogenesis significantly impaired performance on the olfactory discrimination task. Although performance on the first list was entirely unaffected by the loss of neurogenesis, the rats performed significantly worse than controls when they were confronted with a second list of interfering items and irradiated rats experienced significantly more interference than controls. These results therefore support accounts which suggest that hippocampal neurogenesis plays a critical role in mitigating interference (Aimone et al., 6; Becker & Wojtowicz, 7). It was interesting that irradiation produced no impairment in the plus-maze task. Although the olfactory-discrimination task and the maze task differ in a nber of ways, both tasks are impaired by temporary inactivation of the hippocampus (Butterly et al., 212; Smith & Mizori, 6a) and both tasks induce interference. One potentially important difference between the two tasks is the different time courses for learning the interfering items. In the odor-discrimination task, the rats learned the two lists of interfering items sequentially, over the course of several days. In contrast, the competing responses of the maze task were trained concurrently with both responses rewarded within each training session. Thus, the plus maze places a high demand on spatial working memory, requiring the animal to remember the current reward location and to ignore the other location. The lack of an impairment in the irradiated animals on this task is consistent with the finding that rodents with reduced neurogenesis actually outperform controls on a working memory version of the eightarm radial maze (Saxe et al., 7). On the other hand, the impairment seen in the sequentially learned olfactory task supports the hypothesis that the gradual addition of new neurons is an important mechanism for differentially encoding potentially interfering memories more widely separated in time (Becker & Wojtowicz, 7). Specifically, we proposed the cohorts of newly born neurons to be selectively sensitive to the incoming perforant path synaptic inputs and that they transmit the signals encoding common experiences to CA3. Following a relatively brief sensitive period of reduced firing thresholds and heightened plasticity, the cohort would progress to a further state of maturation wherein neurons are less responsive to afferent stimulation, permitting the next wave of young adult neurons to be preferentially recruited. The synaptic mechanisms responsible for the sensitive period include reduced GABA-ergic inhibition and enhanced NMDAdependent plasticity (Becker & Wojtowicz, 7; Deng et al., 21; Snyder, Kee, & Wojtowicz, 1). This putative mech- Figure 1 (opposite). Two neurogenic regions in the rat brain. Images in A show staining for NeuN (green) and BrdU (red) in the olfactory bulb (OB), and DCX (green) and BrdU (red) in the rostral migratory stream (RMS) and the subventricular zone (SVZ). Images in B show staining for DCX, CaBP, and BrdU as indicated in the dentate gyrus (DG). Irradiation was applied selectively to the rear of the brain in order to reduce neurogenesis in the DG, but not in the olfactory system. There was too little CaBP/BrdU to image in irradiated rats. Irradiation caused an 85% reduction in neurogenesis in the dentate gyrus (plots C, D), but had no effect on any of the olfactory regions (plot E).
8 388 LUU, SILL, GAO, BECKER, WOJTOWICZ, AND SMITH Performance Change Different Same Different Same Figure 3. Neurogenesis and interference. Change in performance from List 1 to List 2, computed as the average percent correct during the first three sessions of List 2 minus the average percent correct during the first three sessions of List 1, is shown for each of the experimental groups. Facilitation is indicated by better performance on List 2 than on List 1 (positive values) while interference is indicated by worse performance on List 2 (negative values). anism causes similar events spaced across several days to be encoded by distinct populations of young dentate gyrus neurons. The young neurons in turn contribute to distinct memory traces being formed in downstream regions. In contrast, the maze task may not benefit from neurogenesis because concurrently learning the competing responses does not allow distinct neural populations to differentially encode them. The odor task was specifically designed to induce interference through the use of overlapping odors on the two lists. Nevertheless, interference is also an important aspect of the maze task. It involves serial reversals of the go-east and go-west rules, which produce strong interference, and the choice point presents the rat with an array of cues that have been associated with both reward locations. This likely leads to intrusions of the memory for the incorrect reward location and most errors consisted of entries into the incorrect reward arm, rather than random entries into arms that were never rewarded (data not shown). Thus, both tasks involve interference. It was important, however, that the type of interference differs in the two tasks. In the odor task, similar patterns (overlapping odor pairs) must be mapped to different responses. If the rat can encode the odor pairs learned in the two lists as separate events, using new neurons to generate distinctive memory traces for the overlapping inputs, the task of learning the correct response to an overlapping odor pair becomes greatly simplified as the overlap has been reduced. On the other hand, for the spatial reversal learning task, a single spatial location (the choice point) must be associated with multiple competing responses. The interference cannot be resolved by separating similar inputs, but requires learning the reward value of alternative responses to a given input. These observations suggest that the presence of interference, by itself, is not sufficient to engage neurogenesis dependent pattern separation processes. Instead, neurogenesis may be specifically beneficial for resolving the interference arising from overlapping inputs, particularly when the memories are acquired over the course of a sufficiently long time frame. Of course, the olfactory task and the maze task differed in other ways. For example, the two tasks differ in terms of modality (visuospatial vs. olfactory). However, the impairment seen in the olfactory task was not likely due to general olfactory processing deficits, since irradiation did not cause damage to the rostral migratory stream leading to the olfactory bulb, and the irradiated rats were entirely unimpaired in learning the first list of eight odor pairs. The spatial component of the maze task is probably not an important factor in the differential effects of the irradiation on the two tasks since neurogenesis has been shown to be important for some spatial tasks (Clelland et al., 9). The observation that olfactory tasks are not consistently impaired, and spatial tasks are not consistently spared, suggests that modality is not the critical factor. Moreover, the fact that the two tasks are hippocampal dependent indicates that they both engage general hippocampal mechanisms, despite the modality differences. The two tasks of the present study also differed in terms of contextual manipulations. The olfactory discrimination task involved an explicit manipulation of the environmental context, whereas the maze task did not. However, our results suggested that irradiation impaired performance, regardless of the contextual manipulation. This result stands in contrast to our previous finding that temporary muscimol lesions of the dorsal hippocampus selec- # Cells # Cells CONTROL CONTROL DCX/DG Ki67/DG Untrained Trained IRRADIATED Untrained Trained IRRADIATED Figure 4. Olfactory discrimination training and neurogenesis. Estimates of neurogenesis in rats trained in the olfactory discrimination task and untrained control rats are shown for irradiated and control subjects. The nber of DCX cells per dentate gyrus is shown in A and the nber of Ki67 cells is shown in B. The trained group includes rats from the different-context and same-context training conditions since these groups did not differ.
9 HIPPOCAMPAL NEUROGENESIS AND INTERFERENCE 389 (%) Sessions ACorrect tively impaired performance in the different-context condition but not in the same-context condition (Butterly et al., 212). In that study, the muscimol lesions had no discernible effect on performance in the same-context condition, suggesting a highly specific deficit in the ability to use contextual information to resolve interference. The present results suggest that hippocampal neurogenesis may play a more general role in resolving interference regardless of whether there is an environmental contextual component. The present results also raise the possibility that hippocampal neurogenesis may play an important role in a different kind of context. Ongoing neurogenesis may provide an internal context that gradually varies over time, allowing overlapping events to be separated into distinct memory traces when they are well separated in time, even in the absence of differentiating environmental contexts. This is consistent with the idea that ongoing neural processes form an ever-changing temporal context and that individual events are embedded within this temporal context, in a manner that allows for distinct representations for similar events that occur at different times (Manns, Howard, & Eichenba, 7; Polyn & Kahana, 8). As mentioned above, the recruitment of new neurons may serve to tag each memory trace with its own unique temporal context, thereby reducing interference (Aimone et al., 6; Becker, 5; Becker, Macqueen, & Wojtowicz, 9). In the present study, irradiated rats exhibited proactive interference from previously learned odor pairs. Interestingly, Winocur and colleagues (212) demonstrated retroactive interference in irradiated animals. Specifically, after animals learned distinct stimulus response associations based on black versus white visual cues, only those subsequently given a confusing interfering event a gray cue that had no predictive value for reward later 2 B First Middle Last Figure 5. Plus-maze performance. The nber of sessions needed to reach the behavioral criterion in the plus-maze task are shown in A. Plot B illustrates the percentage of trials with a correct response for control (open) and irradiated rats (filled). Data are shown for the first training session, the session midway through training, and the final training session. Figure 6. Plus-maze training and neurogenesis. Effects of spatial learning on neurogenesis are shown for rats trained in the plus-maze task, for yoked controls (see Method for details) and for untrained, cage controls. The survival of BrdU cells born 1 week before the start of training was not affected by learning (trained group), nor was it affected by handling or exposure to the training apparatus (yoked controls), as compared with the untrained controls (Plot A). Similarly, the rate of maturation as indicated by CaBP/BrdU labeling was the same in trained rats, yoked controls, and untrained controls (Plot B). The density of DCX cells was also the same in trained rats, yoked controls, and untrained controls (Plot C).
10 39 LUU, SILL, GAO, BECKER, WOJTOWICZ, AND SMITH showed impaired performance on the original black white discrimination. Their findings were interpreted as supporting the notion that intact animals used new hippocampal neurons to form separate contextualized memories for the original and interfering events, whereas irradiated animals may have relied upon a striatal stimulus response strategy. However, they did not exclude the possibility that the irradiated animals showed stimulus generalization after exposure to the gray cue and were no longer able to maintain the distinction between the original black and white cues. Such generalization could have accounted for their results. Our present experiments deal specifically with this possibility by showing that neurogenesis was critical for mitigating interference between overlapping associations learned at different times, and was not due to a basic deficit in olfactory stimulus discrimination. As mentioned above, learning in the plus-maze task was not impaired by irradiation. In addition, our experiments failed to show any reciprocal effect of learning on neurogenesis. There were no differences in the nbers of new neurons between rats trained in the olfactory task and untrained controls. However, since the measurements were performed at the end of the study, after the olfactory and plus-maze tasks were completed, any effect of olfactory training may have been obscured by the subsequent maze training. Nevertheless, olfactory training did not produce an effect on neurogenesis large enough that it could be detected even after maze training. However, recent studies (Dupret et al., 7; Epp, Spritzer, & Galea, 7) suggest that the effects of training on neuronal survival may be subtle, with training-induced neuronal survival being restricted to specific phases of learning. One-weekold cells may survive at a higher rate in trained rats, whereas other cells born during later phases of training may show reduced survival, resulting in no net effect. In smary, the results of this study confirm that new neurons are involved in hippocampal-dependent processes that resolve memory interference. At the mechanistic level, interference between successive learning episodes may be related to the mechanism of pattern separation, as proposed by theoretical models (Aimone, Wiles, & Gage, 9; Becker & Wojtowicz, 7). Further experimentation exploring these ideas seems warranted. References Aimone, J. B., Wiles, J., & Gage, F. H. (6). Potential role for adult neurogenesis in the encoding of time in new memories. Nature Neuroscience, 9, doi:1.138/nn177 Aimone, J. B., Wiles, J., & Gage, F. H. (9). Computational influence of adult neurogenesis on memory encoding. Neuron, 61, doi: 1.116/j.neuron Becker, S., Macqueen, G., & Wojtowicz, J. M. (9). Computational modeling and empirical studies of hippocampal neurogenesis-dependent memory: Effects of interference, stress, and depression. Brain Research, 1299, doi:1.116/j.brainres Becker, S., & Wojtowicz, J. M. (7). A model of hippocampal neurogenesis in memory and mood disorders. Trends in Cognitive Science, 11, doi:1.116/j.tics Becker, S. (5). A computational principle for hippocampal learning and neurogenesis. Hippocampus, 15, doi:1.12/hipo.95 Butterly, D. A., Petroccione, M. A., & Smith, D. M. (212). Hippocampal context processing is critical for interference free recall of odor memories in rats. Hippocampus, 22, doi: 1.12/hipo.2953,n/an/a Clelland, C. D., Choi, M., Romberg, C., Clemenson, G. D., Jr., Fragniere, A., Tyers, P.,... Bussey, T. J. (9). A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science, 32, doi:1.1126/science Creer, D. J., Romberg, C., Saksida, L. M., van Praag, H., & Bussey, T. J. (21). Running enhances spatial pattern separation in mice. Proceedings of the National Academy of Sciences of the United States of America, 17, doi:1.173/pnas Deng, W., Aimone, J. B., & Gage, F. H. (21). New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nature Reviews Neuroscience, 1, doi:1.138/ nrn2822 Dupret, D., Fabre, A., Dobrossy, M. D., Panatier, A., Rodriguez, J. J., Lamarque, S.,... Abrous, D. N. (7). Spatial learning depends on both the addition and removal of new hippocampal neurons. PLoS Biology, 5, e214. doi:1.1371/journal.pbio.5214 Epp, J. R., Haack, A. K., & Galea, L. A. (211). Activation and survival of immature neurons in the dentate gyrus with spatial memory is dependent on time of exposure to spatial learning and age of cells at examination. Neurobiology of Learning and Memory, 95, doi:1.116/ j.nlm Epp, J. R., Spritzer, M. D., & Galea, L. A. (7). Hippocampus-dependent learning promotes survival of new neurons in the dentate gyrus at a specific time during cell maturation. Neuroscience, 149, doi: 1.116/j.neuroscience Jacobs, B. L., van Praag, H., & Gage, F. H. (). Adult brain neurogenesis and psychiatry: A novel theory of depression. Molecular Psychiatry, 5, doi:1.138/sj.mp.4712 Manns, J. R., Howard, M. W., & Eichenba, H. (7). Gradual changes in hippocampal activity support remembering the order of events. Neuron, 56, doi:1.116/j.neuron Noonan, M. A., Bulin, S. E., Fuller, D. C., & Eisch, A. J. (21). Reduction of adult hippocampal neurogenesis confers vulnerability in an animal model of cocaine addiction. The Journal of Neuroscience, 3, doi:1.1523/jneurosci Paxinos, G., & Watson, C. (1998). The rat brain in stereotaxic coordinates. (Fourth ed.). New York, NY: Academic Press. Polyn, S. M., & Kahana, M. J. (8). Memory search and the neural representation of context. Trends in Cognitive Sciences, 12, doi:1.116/j.tics Saxe, M. D., Malleret, G., Vronskaya, S., Mendez, I., Garcia, A. D., Sofroniew, M. V.,... Hen, R. (7). Paradoxical influence of hippocampal neurogenesis on working memory. Proceedings of the National Academy of Sciences of the United States of America, 14, doi:1.173/pnas Schoenfeld, T. J., & Gould, E. (211). Stress, stress hormones, and adult neurogenesis. Experimental Neurology, 233, Sisti, H. M., Glass, A. L., & Shors, T. J. (7). Neurogenesis and the spacing effect: Learning over time enhances memory and the survival of new neurons. Learning & Memory, 14, doi:1.111/ lm Smith, D. M., & Mizori, S. J. Y. (6a). Hippocampal place cells, context, and episodic memory. Hippocampus, 16, doi: 1.12/hipo.228 Smith, D. M., & Mizori, S. J. Y. (6b). Learning-related development of context-specific neuronal responses to places and events: The hippocampal role in context processing. The Journal of Neuroscience, 26, doi:1.1523/jneurosci Snyder, J. S., Kee, N., & Wojtowicz, J. M. (1). Effects of adult neurogenesis on synaptic plasticity in the rat dentate gyrus. Journal of Neurophysiology, 85, Tronel, S., Fabre, A., Charrier, V., Oliet, S. H., Gage, F. H., & Abrous,
A Comparison of the Effects of Temporary Hippocampal Lesions on Single and Dual Context Versions of the Olfactory Sequence Memory Task
 Behavioral Neuroscience 2012 American Psychological Association 2012, Vol. 126, No. 4, 588 592 0735-7044/12/$12.00 DOI: 10.1037/a0028824 BRIEF COMMUNICATION A Comparison of the Effects of Temporary Hippocampal
Behavioral Neuroscience 2012 American Psychological Association 2012, Vol. 126, No. 4, 588 592 0735-7044/12/$12.00 DOI: 10.1037/a0028824 BRIEF COMMUNICATION A Comparison of the Effects of Temporary Hippocampal
Terminology. Terminology. Terminology. Terminology. Terminology. Bromodeoxyuridine
 Kateřina Náměstková, Zuzana Šimonová, Eva Syková Behavioural Brain Research Bromodeoxyuridine : Doublecortin : DCX Glial Fibrillary Acidic Protein : GFAP Trace eye blink conditioning 1 Volume 163 : pp.
Kateřina Náměstková, Zuzana Šimonová, Eva Syková Behavioural Brain Research Bromodeoxyuridine : Doublecortin : DCX Glial Fibrillary Acidic Protein : GFAP Trace eye blink conditioning 1 Volume 163 : pp.
Acetylcholine again! - thought to be involved in learning and memory - thought to be involved dementia (Alzheimer's disease)
") Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function Michael E. Hasselmo * and Bradley P. Wyble Acetylcholine again! - thought to
Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function Michael E. Hasselmo * and Bradley P. Wyble Acetylcholine again! - thought to
Supplementary Fig. 1: TBR2+ cells in different brain regions.
 Hip SVZ OB Cere Hypo Supplementary Fig. 1: TBR2 + cells in different brain regions. Three weeks after the last tamoxifen injection, TBR2 immunostaining images reveal a large reduction of TBR2 + cells in
Hip SVZ OB Cere Hypo Supplementary Fig. 1: TBR2 + cells in different brain regions. Three weeks after the last tamoxifen injection, TBR2 immunostaining images reveal a large reduction of TBR2 + cells in
Limbic system outline
 Limbic system outline 1 Introduction 4 The amygdala and emotion -history - theories of emotion - definition - fear and fear conditioning 2 Review of anatomy 5 The hippocampus - amygdaloid complex - septal
Limbic system outline 1 Introduction 4 The amygdala and emotion -history - theories of emotion - definition - fear and fear conditioning 2 Review of anatomy 5 The hippocampus - amygdaloid complex - septal
Supplementary Methods
 1 Supplementary Methods Social Preference Test Subjects Seventy-four Long-Evans, male rats served as subjects (S-rats) in the foodpreference test, with 40 assigned to the CXT-Same (CXT-S) Condition and
1 Supplementary Methods Social Preference Test Subjects Seventy-four Long-Evans, male rats served as subjects (S-rats) in the foodpreference test, with 40 assigned to the CXT-Same (CXT-S) Condition and
Development of a Touch-Screen-Based Paradigm for Assessing Working Memory in the Mouse
 http://dx.doi.org/10.5607/en.2015.24.1.84 Exp Neurobiol. 2015 Mar;24(1):84-89. pissn 1226-2560 eissn 2093-8144 Original Article Development of a Touch-Screen-Based Paradigm for Assessing Working Memory
http://dx.doi.org/10.5607/en.2015.24.1.84 Exp Neurobiol. 2015 Mar;24(1):84-89. pissn 1226-2560 eissn 2093-8144 Original Article Development of a Touch-Screen-Based Paradigm for Assessing Working Memory
Neural stem cells and the neurobiology of ageing. Chen Siyun 1, Dawe G.S. 2
 ABSTRACT Neural stem cells and the neurobiology of ageing Chen Siyun 1, Dawe G.S. 2 Department of Physics, Faculty of Science, National University of Singapore 10 Kent Ridge Road, Singapore 117546 The
ABSTRACT Neural stem cells and the neurobiology of ageing Chen Siyun 1, Dawe G.S. 2 Department of Physics, Faculty of Science, National University of Singapore 10 Kent Ridge Road, Singapore 117546 The
The Effects of Chemotherapy on Cognitive Behavior and Neurogenesis in an Animal Model of Pre- and Post- Menopausal Females
 The Effects of Chemotherapy on Cognitive Behavior and Neurogenesis in an Animal Model of Pre- and Post- Menopausal Females Samantha Pavlock (Medical Student) Pradeep Bhide and Deirdre McCarthy (Faculty
The Effects of Chemotherapy on Cognitive Behavior and Neurogenesis in an Animal Model of Pre- and Post- Menopausal Females Samantha Pavlock (Medical Student) Pradeep Bhide and Deirdre McCarthy (Faculty
Behavioural Brain Research
 Behavioural Brain Research 227 (2012) 464 469 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Adult hippocampal neurogenesis and
Behavioural Brain Research 227 (2012) 464 469 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Adult hippocampal neurogenesis and
Memory: Computation, Genetics, Physiology, and Behavior. James L. McClelland Stanford University
 Memory: Computation, Genetics, Physiology, and Behavior James L. McClelland Stanford University A Playwright s Take on Memory What interests me a great deal is the mistiness of the past Harold Pinter,
Memory: Computation, Genetics, Physiology, and Behavior James L. McClelland Stanford University A Playwright s Take on Memory What interests me a great deal is the mistiness of the past Harold Pinter,
Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare. Venkat R. Lakshminarayanan and Laurie R. Santos
 Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare Venkat R. Lakshminarayanan and Laurie R. Santos Supplemental Experimental Procedures Subjects Seven adult capuchin monkeys were tested.
Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare Venkat R. Lakshminarayanan and Laurie R. Santos Supplemental Experimental Procedures Subjects Seven adult capuchin monkeys were tested.
EFFECTS OF NITRIC OXIDE SYNTHASE INHIBITOR N G -NITRO-L-ARGININE METHYL ESTER ON SPATIAL AND CUED LEANING
 Pergamon Neuroscience Vol. 83, No. 3, pp. 837 841, 1998 Copyright 1998 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved PII: S0306-4522(97)00457-0 0306 4522/98 $19.00+0.00
Pergamon Neuroscience Vol. 83, No. 3, pp. 837 841, 1998 Copyright 1998 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved PII: S0306-4522(97)00457-0 0306 4522/98 $19.00+0.00
Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons
 Animal Learning & Behavior 1999, 27 (2), 206-210 Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons BRIGETTE R. DORRANCE and THOMAS R. ZENTALL University
Animal Learning & Behavior 1999, 27 (2), 206-210 Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons BRIGETTE R. DORRANCE and THOMAS R. ZENTALL University
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/330/6009/1408/dc1 Supporting Online Material for Paradoxical False Memory for Objects After Brain Damage Stephanie M. McTighe, Rosemary A. Cowell, Boyer D. Winters,
www.sciencemag.org/cgi/content/full/330/6009/1408/dc1 Supporting Online Material for Paradoxical False Memory for Objects After Brain Damage Stephanie M. McTighe, Rosemary A. Cowell, Boyer D. Winters,
Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context
 Animal Learning & Behavior 1998, 26 (3), 257 263 Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context BRIGETTE R.
Animal Learning & Behavior 1998, 26 (3), 257 263 Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context BRIGETTE R.
UCLA UCLA Previously Published Works
 UCLA UCLA Previously Published Works Title The role of postnatal neurogenesis in supporting remote memory and spatial metric processing Permalink https://escholarship.org/uc/item/87668203 Authors Kesner,
UCLA UCLA Previously Published Works Title The role of postnatal neurogenesis in supporting remote memory and spatial metric processing Permalink https://escholarship.org/uc/item/87668203 Authors Kesner,
The Timing of Differentiation of Adult Hippocampal Neurons Is Crucial for Spatial Memory
 The Timing of Differentiation of Adult Hippocampal Neurons Is Crucial for Spatial Memory PLoS BIOLOGY Stefano Farioli-Vecchioli 1[, Daniele Saraulli 2,3[, Marco Costanzi 2,3[, Simone Pacioni 4[, Irene
The Timing of Differentiation of Adult Hippocampal Neurons Is Crucial for Spatial Memory PLoS BIOLOGY Stefano Farioli-Vecchioli 1[, Daniele Saraulli 2,3[, Marco Costanzi 2,3[, Simone Pacioni 4[, Irene
Geometrical and Spatial Cues
 The Huron University College Journal of Learning and Motivation Volume 45 Issue 1 Article 20 2007 Geometrical and Spatial Cues Josée Viau Follow this and additional works at: http://ir.lib.uwo.ca/hucjlm
The Huron University College Journal of Learning and Motivation Volume 45 Issue 1 Article 20 2007 Geometrical and Spatial Cues Josée Viau Follow this and additional works at: http://ir.lib.uwo.ca/hucjlm
Modeling of Hippocampal Behavior
 Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
Assessing mouse behaviors: Modeling pediatric traumatic brain injury
 Assessing mouse behaviors: Modeling pediatric traumatic brain injury Bridgette Semple Ph.D. Postdoctoral Fellow, Noble Laboratory Department of Neurological Surgery, UCSF Pediatric Traumatic Brain Injury
Assessing mouse behaviors: Modeling pediatric traumatic brain injury Bridgette Semple Ph.D. Postdoctoral Fellow, Noble Laboratory Department of Neurological Surgery, UCSF Pediatric Traumatic Brain Injury
Nature Neuroscience: doi: /nn Supplementary Figure 1. Behavioral training.
 Supplementary Figure 1 Behavioral training. a, Mazes used for behavioral training. Asterisks indicate reward location. Only some example mazes are shown (for example, right choice and not left choice maze
Supplementary Figure 1 Behavioral training. a, Mazes used for behavioral training. Asterisks indicate reward location. Only some example mazes are shown (for example, right choice and not left choice maze
Cultural Differences in Cognitive Processing Style: Evidence from Eye Movements During Scene Processing
 Cultural Differences in Cognitive Processing Style: Evidence from Eye Movements During Scene Processing Zihui Lu (zihui.lu@utoronto.ca) Meredyth Daneman (daneman@psych.utoronto.ca) Eyal M. Reingold (reingold@psych.utoronto.ca)
Cultural Differences in Cognitive Processing Style: Evidence from Eye Movements During Scene Processing Zihui Lu (zihui.lu@utoronto.ca) Meredyth Daneman (daneman@psych.utoronto.ca) Eyal M. Reingold (reingold@psych.utoronto.ca)
Reduction of Adult Hippocampal Neurogenesis Confers Vulnerability in an Animal Model of Cocaine Addiction
 304 The Journal of Neuroscience, January 6, 2010 30(1):304 315 Behavioral/Systems/Cognitive Reduction of Adult Hippocampal Neurogenesis Confers Vulnerability in an Animal Model of Cocaine Addiction Michele
304 The Journal of Neuroscience, January 6, 2010 30(1):304 315 Behavioral/Systems/Cognitive Reduction of Adult Hippocampal Neurogenesis Confers Vulnerability in an Animal Model of Cocaine Addiction Michele
BIOMED 509. Executive Control UNM SOM. Primate Research Inst. Kyoto,Japan. Cambridge University. JL Brigman
 BIOMED 509 Executive Control Cambridge University Primate Research Inst. Kyoto,Japan UNM SOM JL Brigman 4-7-17 Symptoms and Assays of Cognitive Disorders Symptoms of Cognitive Disorders Learning & Memory
BIOMED 509 Executive Control Cambridge University Primate Research Inst. Kyoto,Japan UNM SOM JL Brigman 4-7-17 Symptoms and Assays of Cognitive Disorders Symptoms of Cognitive Disorders Learning & Memory
Transfer of memory retrieval cues attenuates the context specificity of latent inhibition
 Scholarly Commons Psychology Faculty Publications 2015 Transfer of memory retrieval cues attenuates the context specificity of latent inhibition James F. Briggs Timothy A. Toth Brian P. Olson Jacob G.
Scholarly Commons Psychology Faculty Publications 2015 Transfer of memory retrieval cues attenuates the context specificity of latent inhibition James F. Briggs Timothy A. Toth Brian P. Olson Jacob G.
Why do we have a hippocampus? Short-term memory and consolidation
 Why do we have a hippocampus? Short-term memory and consolidation So far we have talked about the hippocampus and: -coding of spatial locations in rats -declarative (explicit) memory -experimental evidence
Why do we have a hippocampus? Short-term memory and consolidation So far we have talked about the hippocampus and: -coding of spatial locations in rats -declarative (explicit) memory -experimental evidence
Interference with spatial working memory: An eye movement is more than a shift of attention
 Psychonomic Bulletin & Review 2004, 11 (3), 488-494 Interference with spatial working memory: An eye movement is more than a shift of attention BONNIE M. LAWRENCE Washington University School of Medicine,
Psychonomic Bulletin & Review 2004, 11 (3), 488-494 Interference with spatial working memory: An eye movement is more than a shift of attention BONNIE M. LAWRENCE Washington University School of Medicine,
Hippocampal mechanisms of memory and cognition. Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT
 Hippocampal mechanisms of memory and cognition Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT 1 Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission.
Hippocampal mechanisms of memory and cognition Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT 1 Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission.
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
A model of the interaction between mood and memory
 INSTITUTE OF PHYSICS PUBLISHING NETWORK: COMPUTATION IN NEURAL SYSTEMS Network: Comput. Neural Syst. 12 (2001) 89 109 www.iop.org/journals/ne PII: S0954-898X(01)22487-7 A model of the interaction between
INSTITUTE OF PHYSICS PUBLISHING NETWORK: COMPUTATION IN NEURAL SYSTEMS Network: Comput. Neural Syst. 12 (2001) 89 109 www.iop.org/journals/ne PII: S0954-898X(01)22487-7 A model of the interaction between
Attentional Blink Paradigm
 Attentional Blink Paradigm ATTENTIONAL BLINK 83 ms stimulus onset asychrony between all stimuli B T D A 3 N P Z F R K M R N Lag 3 Target 1 Target 2 After detection of a target in a rapid stream of visual
Attentional Blink Paradigm ATTENTIONAL BLINK 83 ms stimulus onset asychrony between all stimuli B T D A 3 N P Z F R K M R N Lag 3 Target 1 Target 2 After detection of a target in a rapid stream of visual
A Study of Hippocampal Structure-function Relations. Along the Septo-temporal Axis
 1 A Study of Hippocampal Structure-function Relations Along the Septo-temporal Axis Leonard E. Jarrard 1, Lisa P. Luu 1, & Terry L. Davidson 2 1Department of Psychology and Neuroscience Program, Washington
1 A Study of Hippocampal Structure-function Relations Along the Septo-temporal Axis Leonard E. Jarrard 1, Lisa P. Luu 1, & Terry L. Davidson 2 1Department of Psychology and Neuroscience Program, Washington
The effects of subthreshold synchrony on the perception of simultaneity. Ludwig-Maximilians-Universität Leopoldstr 13 D München/Munich, Germany
 The effects of subthreshold synchrony on the perception of simultaneity 1,2 Mark A. Elliott, 2 Zhuanghua Shi & 2,3 Fatma Sürer 1 Department of Psychology National University of Ireland Galway, Ireland.
The effects of subthreshold synchrony on the perception of simultaneity 1,2 Mark A. Elliott, 2 Zhuanghua Shi & 2,3 Fatma Sürer 1 Department of Psychology National University of Ireland Galway, Ireland.
Adolescent Prozac Exposure Enhances Sensitivity to Cocaine in Adulthood INTRODUCTION
 INTRODUCTION Epidemiologic reports indicate that mood disorders in children and adolescents are quite common, with up to 70% of depressed children and adolescents experiencing a recurrence within 5 years
INTRODUCTION Epidemiologic reports indicate that mood disorders in children and adolescents are quite common, with up to 70% of depressed children and adolescents experiencing a recurrence within 5 years
Theta sequences are essential for internally generated hippocampal firing fields.
 Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Are Both Embryonic Migratory Pathways Preserved in the Adult Brain Cerebral Cortex?
 Prague Medical Report / Vol. 107 (2006) No. 1, p. 71 80 71) Are Both Embryonic Migratory Pathways Preserved in the Adult Brain Cerebral Cortex? Šimonová Z., Dutt J. Department of Neuroscience of the Institute
Prague Medical Report / Vol. 107 (2006) No. 1, p. 71 80 71) Are Both Embryonic Migratory Pathways Preserved in the Adult Brain Cerebral Cortex? Šimonová Z., Dutt J. Department of Neuroscience of the Institute
Exercising Our Minds: Effects of Exercise on Brain Structure & Function
 Public Lecture Exercising Our Minds: Effects of Exercise on Brain Structure & Function Dr. Brian R. Christie Division of Medical Sciences, University of Victoria Cellular and Physiological Sciences, UBC
Public Lecture Exercising Our Minds: Effects of Exercise on Brain Structure & Function Dr. Brian R. Christie Division of Medical Sciences, University of Victoria Cellular and Physiological Sciences, UBC
Adult Neurogenesis in the Hippocampus From a Perspective of Discrimination and Generalization: A Hypothesis
 Physiol. Res. 66: 441-448, 2017 REVIEW Adult Neurogenesis in the Hippocampus From a Perspective of Discrimination and Generalization: A Hypothesis A. PISTIKOVA 1*, H. BROZKA 1*, A. STUCHLIK 1 * These authors
Physiol. Res. 66: 441-448, 2017 REVIEW Adult Neurogenesis in the Hippocampus From a Perspective of Discrimination and Generalization: A Hypothesis A. PISTIKOVA 1*, H. BROZKA 1*, A. STUCHLIK 1 * These authors
<student name> Undergraduate Research Grant Proposal
 Undergraduate Research Grant Proposal A. Project Description Objective of research: The objective of this study is to determine if hippocampal dopamine D1 receptors facilitate novel object
Undergraduate Research Grant Proposal A. Project Description Objective of research: The objective of this study is to determine if hippocampal dopamine D1 receptors facilitate novel object
Memory Systems Interaction in the Pigeon: Working and Reference Memory
 Journal of Experimental Psychology: Animal Learning and Cognition 2015 American Psychological Association 2015, Vol. 41, No. 2, 152 162 2329-8456/15/$12.00 http://dx.doi.org/10.1037/xan0000053 Memory Systems
Journal of Experimental Psychology: Animal Learning and Cognition 2015 American Psychological Association 2015, Vol. 41, No. 2, 152 162 2329-8456/15/$12.00 http://dx.doi.org/10.1037/xan0000053 Memory Systems
Chapter 6: Hippocampal Function In Cognition. From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D.
 Chapter 6: Hippocampal Function In Cognition From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Grid Cell The Hippocampus Serves a Role in Multimodal Information Processing and Memory
Chapter 6: Hippocampal Function In Cognition From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Grid Cell The Hippocampus Serves a Role in Multimodal Information Processing and Memory
Modeling the Role of Working Memory and Episodic Memory in Behavioral Tasks
 Modeling the Role of Working Memory and Episodic Memory in Behavioral Tasks Eric A. Zilli 1,2 * and Michael E. Hasselmo 1,2 HIPPOCAMPUS 00:000 000 (2007) ABSTRACT: The mechanisms of goal-directed behavior
Modeling the Role of Working Memory and Episodic Memory in Behavioral Tasks Eric A. Zilli 1,2 * and Michael E. Hasselmo 1,2 HIPPOCAMPUS 00:000 000 (2007) ABSTRACT: The mechanisms of goal-directed behavior
Object Substitution Masking: When does Mask Preview work?
 Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
A systems neuroscience approach to memory
 A systems neuroscience approach to memory Critical brain structures for declarative memory Relational memory vs. item memory Recollection vs. familiarity Recall vs. recognition What about PDs? R-K paradigm
A systems neuroscience approach to memory Critical brain structures for declarative memory Relational memory vs. item memory Recollection vs. familiarity Recall vs. recognition What about PDs? R-K paradigm
Psychology Neuroscience and Behaviour, McMaster University, Canada. Provisional
 Neurogenesis paradoxically decreases both pattern separation and memory interference Rory Finnegan 1, Suzanna Becker 2* 1 Psychology Neuroscience and Behaviour, McMaster University, Canada, 2 Department
Neurogenesis paradoxically decreases both pattern separation and memory interference Rory Finnegan 1, Suzanna Becker 2* 1 Psychology Neuroscience and Behaviour, McMaster University, Canada, 2 Department
Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions
 Learning & Behavior 2009, 37 (4), 299-304 doi:10.3758/lb.37.4.299 Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions REBECCA RAYBURN-REEVES
Learning & Behavior 2009, 37 (4), 299-304 doi:10.3758/lb.37.4.299 Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions REBECCA RAYBURN-REEVES
Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements
 Y. Isomura et al. 1 Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements Yoshikazu Isomura, Rie Harukuni, Takashi Takekawa, Hidenori Aizawa & Tomoki Fukai
Y. Isomura et al. 1 Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements Yoshikazu Isomura, Rie Harukuni, Takashi Takekawa, Hidenori Aizawa & Tomoki Fukai
Brain and Cognitive Sciences 9.96 Experimental Methods of Tetrode Array Neurophysiology IAP 2001
 Brain and Cognitive Sciences 9.96 Experimental Methods of Tetrode Array Neurophysiology IAP 2001 An Investigation into the Mechanisms of Memory through Hippocampal Microstimulation In rodents, the hippocampus
Brain and Cognitive Sciences 9.96 Experimental Methods of Tetrode Array Neurophysiology IAP 2001 An Investigation into the Mechanisms of Memory through Hippocampal Microstimulation In rodents, the hippocampus
TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab
 TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab This report details my summer research project for the REU Theoretical and Computational Neuroscience program as
TMS Disruption of Time Encoding in Human Primary Visual Cortex Molly Bryan Beauchamp Lab This report details my summer research project for the REU Theoretical and Computational Neuroscience program as
Session Goals. Principles of Brain Plasticity
 Presenter: Bryan Kolb Canadian Centre for Behavioural Neuroscience University of Lethbridge Date: January 12, 2011 The FASD Learning Series is part of the Alberta government s commitment to programs and
Presenter: Bryan Kolb Canadian Centre for Behavioural Neuroscience University of Lethbridge Date: January 12, 2011 The FASD Learning Series is part of the Alberta government s commitment to programs and
Encoding of Elements and Relations of Object Arrangements by Young Children
 Encoding of Elements and Relations of Object Arrangements by Young Children Leslee J. Martin (martin.1103@osu.edu) Department of Psychology & Center for Cognitive Science Ohio State University 216 Lazenby
Encoding of Elements and Relations of Object Arrangements by Young Children Leslee J. Martin (martin.1103@osu.edu) Department of Psychology & Center for Cognitive Science Ohio State University 216 Lazenby
Invariant Effects of Working Memory Load in the Face of Competition
 Invariant Effects of Working Memory Load in the Face of Competition Ewald Neumann (ewald.neumann@canterbury.ac.nz) Department of Psychology, University of Canterbury Christchurch, New Zealand Stephen J.
Invariant Effects of Working Memory Load in the Face of Competition Ewald Neumann (ewald.neumann@canterbury.ac.nz) Department of Psychology, University of Canterbury Christchurch, New Zealand Stephen J.
Learning to Use Episodic Memory
 Learning to Use Episodic Memory Nicholas A. Gorski (ngorski@umich.edu) John E. Laird (laird@umich.edu) Computer Science & Engineering, University of Michigan 2260 Hayward St., Ann Arbor, MI 48109 USA Abstract
Learning to Use Episodic Memory Nicholas A. Gorski (ngorski@umich.edu) John E. Laird (laird@umich.edu) Computer Science & Engineering, University of Michigan 2260 Hayward St., Ann Arbor, MI 48109 USA Abstract
Visual working memory as the substrate for mental rotation
 Psychonomic Bulletin & Review 2007, 14 (1), 154-158 Visual working memory as the substrate for mental rotation JOO-SEOK HYUN AND STEVEN J. LUCK University of Iowa, Iowa City, Iowa In mental rotation, a
Psychonomic Bulletin & Review 2007, 14 (1), 154-158 Visual working memory as the substrate for mental rotation JOO-SEOK HYUN AND STEVEN J. LUCK University of Iowa, Iowa City, Iowa In mental rotation, a
October 2, Memory II. 8 The Human Amnesic Syndrome. 9 Recent/Remote Distinction. 11 Frontal/Executive Contributions to Memory
 1 Memory II October 2, 2008 2 3 4 5 6 7 8 The Human Amnesic Syndrome Impaired new learning (anterograde amnesia), exacerbated by increasing retention delay Impaired recollection of events learned prior
1 Memory II October 2, 2008 2 3 4 5 6 7 8 The Human Amnesic Syndrome Impaired new learning (anterograde amnesia), exacerbated by increasing retention delay Impaired recollection of events learned prior
The Journal of Physiology Neuroscience
 J Physiol 594.7 (216) pp 1855 1873 1855 The Journal of Physiology Neuroscience Physical exercise increases adult hippocampal neurogenesis in male rats provided it is aerobic and sustained Miriam S. Nokia
J Physiol 594.7 (216) pp 1855 1873 1855 The Journal of Physiology Neuroscience Physical exercise increases adult hippocampal neurogenesis in male rats provided it is aerobic and sustained Miriam S. Nokia
The hippocampus and contextual memory retrieval in Pavlovian conditioning
 Behavioural Brain Research 110 (2000) 97 108 www.elsevier.com/locate/bbr The hippocampus and contextual memory retrieval in Pavlovian conditioning Stephen Maren *, William Holt Department of Psychology
Behavioural Brain Research 110 (2000) 97 108 www.elsevier.com/locate/bbr The hippocampus and contextual memory retrieval in Pavlovian conditioning Stephen Maren *, William Holt Department of Psychology
Human Cognitive Developmental Neuroscience. Jan 27
 Human Cognitive Developmental Neuroscience Jan 27 Wiki Definition Developmental cognitive neuroscience is an interdisciplinary scientific field that is situated at the boundaries of Neuroscience Psychology
Human Cognitive Developmental Neuroscience Jan 27 Wiki Definition Developmental cognitive neuroscience is an interdisciplinary scientific field that is situated at the boundaries of Neuroscience Psychology
Brain Research Bulletin 67 (2005) Toronto, Ont., Canada M6A 2E1 b Department of Psychology, University of New Mexico, Albuquerque,
 Toronto, Ont., Canada M6A 2E1 b Department of Psychology, University of New Mexico, Albuquerque,") Brain Research Bulletin 67 (2005) 62 76 Differential contributions of hippocampus, amygdala and perirhinal cortex to recognition of novel objects, contextual stimuli and stimulus relationships Sandra N.
Brain Research Bulletin 67 (2005) 62 76 Differential contributions of hippocampus, amygdala and perirhinal cortex to recognition of novel objects, contextual stimuli and stimulus relationships Sandra N.
Supplemental Information. A Visual-Cue-Dependent Memory Circuit. for Place Navigation
 Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
Memory retention the synaptic stability versus plasticity dilemma
 Memory retention the synaptic stability versus plasticity dilemma Paper: Abraham, Wickliffe C., and Anthony Robins. "Memory retention the synaptic stability versus plasticity dilemma." Trends in neurosciences
Memory retention the synaptic stability versus plasticity dilemma Paper: Abraham, Wickliffe C., and Anthony Robins. "Memory retention the synaptic stability versus plasticity dilemma." Trends in neurosciences
Time Experiencing by Robotic Agents
 Time Experiencing by Robotic Agents Michail Maniadakis 1 and Marc Wittmann 2 and Panos Trahanias 1 1- Foundation for Research and Technology - Hellas, ICS, Greece 2- Institute for Frontier Areas of Psychology
Time Experiencing by Robotic Agents Michail Maniadakis 1 and Marc Wittmann 2 and Panos Trahanias 1 1- Foundation for Research and Technology - Hellas, ICS, Greece 2- Institute for Frontier Areas of Psychology
Memory, Attention, and Decision-Making
 Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
S100B+ Cell Density day Spine Formation (%)
") b * CP * HC CP Th DAPI S100B ** 50 40 30 * 20 10 0-10 -20 * * ** 9 6 3 0 0-50 50-100 100-150 150-200 Distance from Prism Face (um) 1-day Spine Formation (%) 1-day Spine Elimination (%) 12 Distance from
b * CP * HC CP Th DAPI S100B ** 50 40 30 * 20 10 0-10 -20 * * ** 9 6 3 0 0-50 50-100 100-150 150-200 Distance from Prism Face (um) 1-day Spine Formation (%) 1-day Spine Elimination (%) 12 Distance from
NIH Public Access Author Manuscript Nat Neurosci. Author manuscript; available in PMC 2006 September 5.
 NIH Public Access Author Manuscript Published in final edited form as: Nat Neurosci. 2006 August ; 9(8): 1004 1006. Maternal presence serves as a switch between learning fear and attraction in infancy
NIH Public Access Author Manuscript Published in final edited form as: Nat Neurosci. 2006 August ; 9(8): 1004 1006. Maternal presence serves as a switch between learning fear and attraction in infancy
Interplay of Hippocampus and Prefrontal Cortex in Memory
 Current Biology 23, R764 R773, September 9, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.05.041 Interplay of Hippocampus and Prefrontal Cortex in Memory Review Alison
Current Biology 23, R764 R773, September 9, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.05.041 Interplay of Hippocampus and Prefrontal Cortex in Memory Review Alison
CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems
 CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems Compare Chap 31 of Purves et al., 5e Chap 24 of Bear et al., 3e Larry Wittie Computer Science, StonyBrook University http://www.cs.sunysb.edu/~cse511
CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems Compare Chap 31 of Purves et al., 5e Chap 24 of Bear et al., 3e Larry Wittie Computer Science, StonyBrook University http://www.cs.sunysb.edu/~cse511
The Meaning of the Mask Matters
 PSYCHOLOGICAL SCIENCE Research Report The Meaning of the Mask Matters Evidence of Conceptual Interference in the Attentional Blink Paul E. Dux and Veronika Coltheart Macquarie Centre for Cognitive Science,
PSYCHOLOGICAL SCIENCE Research Report The Meaning of the Mask Matters Evidence of Conceptual Interference in the Attentional Blink Paul E. Dux and Veronika Coltheart Macquarie Centre for Cognitive Science,
Value Transfer in a Simultaneous Discrimination Appears to Result From Within-Event Pavlovian Conditioning
 Journal of Experimental Psychology: Animal Behavior Processes 1996, Vol. 22. No. 1, 68-75 Copyright 1996 by the American Psychological Association. Inc. 0097-7403/96/53.00 Value Transfer in a Simultaneous
Journal of Experimental Psychology: Animal Behavior Processes 1996, Vol. 22. No. 1, 68-75 Copyright 1996 by the American Psychological Association. Inc. 0097-7403/96/53.00 Value Transfer in a Simultaneous
A Memory Model for Decision Processes in Pigeons
 From M. L. Commons, R.J. Herrnstein, & A.R. Wagner (Eds.). 1983. Quantitative Analyses of Behavior: Discrimination Processes. Cambridge, MA: Ballinger (Vol. IV, Chapter 1, pages 3-19). A Memory Model for
From M. L. Commons, R.J. Herrnstein, & A.R. Wagner (Eds.). 1983. Quantitative Analyses of Behavior: Discrimination Processes. Cambridge, MA: Ballinger (Vol. IV, Chapter 1, pages 3-19). A Memory Model for
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05772 SUPPLEMENTARY INFORMATION Supplemental figure 1. Enrichment facilitates learning. a. Images showing a home cage and a cage used for environmental enrichment (EE). For EE up to
doi: 10.1038/nature05772 SUPPLEMENTARY INFORMATION Supplemental figure 1. Enrichment facilitates learning. a. Images showing a home cage and a cage used for environmental enrichment (EE). For EE up to
Templates for Rejection: Configuring Attention to Ignore Task-Irrelevant Features
 Journal of Experimental Psychology: Human Perception and Performance 2012, Vol. 38, No. 3, 580 584 2012 American Psychological Association 0096-1523/12/$12.00 DOI: 10.1037/a0027885 OBSERVATION Templates
Journal of Experimental Psychology: Human Perception and Performance 2012, Vol. 38, No. 3, 580 584 2012 American Psychological Association 0096-1523/12/$12.00 DOI: 10.1037/a0027885 OBSERVATION Templates
How to measure rodent behavior and perform a neurological screen.
 An Organ Systems Approach to Experimental Targeting of the Metabolic Syndrome How to measure rodent behavior and perform a neurological screen. Fiona Harrison, PhD Department of Medicine Vanderbilt University
An Organ Systems Approach to Experimental Targeting of the Metabolic Syndrome How to measure rodent behavior and perform a neurological screen. Fiona Harrison, PhD Department of Medicine Vanderbilt University
A model of parallel time estimation
 A model of parallel time estimation Hedderik van Rijn 1 and Niels Taatgen 1,2 1 Department of Artificial Intelligence, University of Groningen Grote Kruisstraat 2/1, 9712 TS Groningen 2 Department of Psychology,
A model of parallel time estimation Hedderik van Rijn 1 and Niels Taatgen 1,2 1 Department of Artificial Intelligence, University of Groningen Grote Kruisstraat 2/1, 9712 TS Groningen 2 Department of Psychology,
Suppl. Information Supplementary Figure 1. Strategy/latency analysis of individual mice during maze learning. a,
 Goal-oriented searching mediated by ventral hippocampus early in trial-and-error learning Ruediger, S, Spirig, D., Donato, F., Caroni, P. Suppl. Information Supplementary Figure 1. Strategy/latency analysis
Goal-oriented searching mediated by ventral hippocampus early in trial-and-error learning Ruediger, S, Spirig, D., Donato, F., Caroni, P. Suppl. Information Supplementary Figure 1. Strategy/latency analysis
Behavioural Brain Research
 Behavioural Brain Research 215 (2010) 197 208 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review The role of the human hippocampus
Behavioural Brain Research 215 (2010) 197 208 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review The role of the human hippocampus
Gene co-expression networks in the mouse, monkey, and human brain July 16, Jeremy Miller Scientist I
 Gene co-expression networks in the mouse, monkey, and human brain July 16, 2013 Jeremy Miller Scientist I jeremym@alleninstitute.org Outline 1. Brief introduction to previous WGCNA studies in brain 2.
Gene co-expression networks in the mouse, monkey, and human brain July 16, 2013 Jeremy Miller Scientist I jeremym@alleninstitute.org Outline 1. Brief introduction to previous WGCNA studies in brain 2.
CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA. in which stimulus control developed was studied; of subjects differing in the probability value
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 551-559 NUMBER 4 (JULY) PROBABILITY OF REINFORCEMENT AND THE DEVELOPMENT OF STIMULUS CONTROL' CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA Pigeons
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 551-559 NUMBER 4 (JULY) PROBABILITY OF REINFORCEMENT AND THE DEVELOPMENT OF STIMULUS CONTROL' CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA Pigeons
The Neurophysiology of Memory
 The Neurophysiology of Memory WENDY A. SUZUKI a,c AND HOWARD EICHENBAUM b a Center for Neural Science, New York University, 4 Washington Place, Room 809, New York, New York 10003, USA b Laboratory of Cognitive
The Neurophysiology of Memory WENDY A. SUZUKI a,c AND HOWARD EICHENBAUM b a Center for Neural Science, New York University, 4 Washington Place, Room 809, New York, New York 10003, USA b Laboratory of Cognitive
Remembering the Past to Imagine the Future: A Cognitive Neuroscience Perspective
 MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
Auditory Scene Analysis
 1 Auditory Scene Analysis Albert S. Bregman Department of Psychology McGill University 1205 Docteur Penfield Avenue Montreal, QC Canada H3A 1B1 E-mail: bregman@hebb.psych.mcgill.ca To appear in N.J. Smelzer
1 Auditory Scene Analysis Albert S. Bregman Department of Psychology McGill University 1205 Docteur Penfield Avenue Montreal, QC Canada H3A 1B1 E-mail: bregman@hebb.psych.mcgill.ca To appear in N.J. Smelzer
Prss56, a novel marker of adult neurogenesis in the mouse brain. - Supplemental Figures 1 to 5- Brain Structure and Function
 Prss56, a novel marker of adult neurogenesis in the mouse brain - Supplemental Figures 1 to 5- Brain Structure and Function Alexandre Jourdon 1,2, Aurélie Gresset 1, Nathalie Spassky 1, Patrick Charnay
Prss56, a novel marker of adult neurogenesis in the mouse brain - Supplemental Figures 1 to 5- Brain Structure and Function Alexandre Jourdon 1,2, Aurélie Gresset 1, Nathalie Spassky 1, Patrick Charnay
Mnemonic representations of transient stimuli and temporal sequences in the rodent hippocampus in vitro
 Supplementary Material Mnemonic representations of transient stimuli and temporal sequences in the rodent hippocampus in vitro Robert. Hyde and en W. Strowbridge Mossy ell 1 Mossy ell Mossy ell 3 Stimulus
Supplementary Material Mnemonic representations of transient stimuli and temporal sequences in the rodent hippocampus in vitro Robert. Hyde and en W. Strowbridge Mossy ell 1 Mossy ell Mossy ell 3 Stimulus
Synaptic plasticity and hippocampal memory
 Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE ORDER TERRY W.
 RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE ORDER TERRY W.") JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2000, 73, 319 331 NUMBER 3(MAY) STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2000, 73, 319 331 NUMBER 3(MAY) STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE
The spacing and lag effect in free recall
 The spacing and lag effect in free recall Michael J. Kahana, Bradley R. Wellington & Marc W. Howard Center for Complex Systems and Department of Psychology Brandeis University Send correspondence to: Michael
The spacing and lag effect in free recall Michael J. Kahana, Bradley R. Wellington & Marc W. Howard Center for Complex Systems and Department of Psychology Brandeis University Send correspondence to: Michael
A Dynamic Field Theory of Visual Recognition in Infant Looking Tasks
 A Dynamic Field Theory of Visual Recognition in Infant Looking Tasks Sammy Perone (sammy-perone@uiowa.edu) and John P. Spencer (john-spencer@uiowa.edu) Department of Psychology, 11 Seashore Hall East Iowa
A Dynamic Field Theory of Visual Recognition in Infant Looking Tasks Sammy Perone (sammy-perone@uiowa.edu) and John P. Spencer (john-spencer@uiowa.edu) Department of Psychology, 11 Seashore Hall East Iowa
Untangling the Influences of Voluntary Running, Environmental Complexity, Social Housing and Stress on Adult Hippocampal Neurogenesis
 Untangling the Influences of Voluntary Running, Environmental Complexity, Social Housing and Stress on Adult Hippocampal Neurogenesis Catherine-Alexandra Grégoire, David Bonenfant, Adalie Le Nguyen, Anne
Untangling the Influences of Voluntary Running, Environmental Complexity, Social Housing and Stress on Adult Hippocampal Neurogenesis Catherine-Alexandra Grégoire, David Bonenfant, Adalie Le Nguyen, Anne
Behavioural Brain Research
 Behavioural Brain Research 227 (2012) 450 458 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Use it or lose it: How neurogenesis
Behavioural Brain Research 227 (2012) 450 458 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Use it or lose it: How neurogenesis
INFLUENCE OF RETROACTIVE INTERFERENCE ON THE CONTEXT SHIFT EFFECT
 INFLUENCE OF RETROACTIVE INTERFERENCE ON THE CONTEXT SHIFT EFFECT A thesis submitted to Kent State University in partial fulfillment of the requirements for the degree of Master of Arts By Erin Marie Fleming
INFLUENCE OF RETROACTIVE INTERFERENCE ON THE CONTEXT SHIFT EFFECT A thesis submitted to Kent State University in partial fulfillment of the requirements for the degree of Master of Arts By Erin Marie Fleming
Supporting Information: Cognitive capacities for cooking in chimpanzees Felix Warneken & Alexandra G. Rosati
 Supporting Information: Cognitive capacities for cooking in chimpanzees Felix Warneken & Alexandra G. Rosati Subject Information Name Sex Age Testing Year 1 Testing Year 2 1 2 3 4 5a 5b 5c 6a 6b 7 8 9
Supporting Information: Cognitive capacities for cooking in chimpanzees Felix Warneken & Alexandra G. Rosati Subject Information Name Sex Age Testing Year 1 Testing Year 2 1 2 3 4 5a 5b 5c 6a 6b 7 8 9
Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) 108 (b-c) (d) (e) (f) (g)
 108 (b-c) (d) (e) (f) (g)") Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) In four mice, cre-dependent expression of the hyperpolarizing opsin Arch in pyramidal cells
Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) In four mice, cre-dependent expression of the hyperpolarizing opsin Arch in pyramidal cells
Running head: EFFECTS OF ALCOHOL ON OBJECT RECOGNITION. Impairing Effects of Alcohol on Object Recognition. A Senior Honors Thesis
 Running head: EFFECTS OF ALCOHOL ON OBJECT RECOGNITION Alcohol on Object Recognition 1 Impairing Effects of Alcohol on Object Recognition A Senior Honors Thesis Presented in Partial Fulfillment of the
Running head: EFFECTS OF ALCOHOL ON OBJECT RECOGNITION Alcohol on Object Recognition 1 Impairing Effects of Alcohol on Object Recognition A Senior Honors Thesis Presented in Partial Fulfillment of the
Cerebral Cortex. Edmund T. Rolls. Principles of Operation. Presubiculum. Subiculum F S D. Neocortex. PHG & Perirhinal. CA1 Fornix CA3 S D
 Cerebral Cortex Principles of Operation Edmund T. Rolls F S D Neocortex S D PHG & Perirhinal 2 3 5 pp Ento rhinal DG Subiculum Presubiculum mf CA3 CA1 Fornix Appendix 4 Simulation software for neuronal
Cerebral Cortex Principles of Operation Edmund T. Rolls F S D Neocortex S D PHG & Perirhinal 2 3 5 pp Ento rhinal DG Subiculum Presubiculum mf CA3 CA1 Fornix Appendix 4 Simulation software for neuronal
ASHI 712. The Neuroscience of Human Memory. Dr. Olave E. Krigolson LECTURE 2: Short Term Memory and Sleep and Memory
 ASHI 712 The Neuroscience of Human Memory Dr. Olave E. Krigolson krigolson@uvic.ca LECTURE 2: Short Term Memory and Sleep and Memory Working / Short Term Memory Sunglasses Chair Dress Earrings Boots Bed
ASHI 712 The Neuroscience of Human Memory Dr. Olave E. Krigolson krigolson@uvic.ca LECTURE 2: Short Term Memory and Sleep and Memory Working / Short Term Memory Sunglasses Chair Dress Earrings Boots Bed
Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory
 Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory Anna Smith Outline 1. 2. 3. 4. 5. Background in Memory Models Models of Consolidation The Hippocampus Competitive Trace Theory
Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory Anna Smith Outline 1. 2. 3. 4. 5. Background in Memory Models Models of Consolidation The Hippocampus Competitive Trace Theory
Experience-dependent recovery of vision following chronic deprivation amblyopia. Hai-Yan He, Baisali Ray, Katie Dennis and Elizabeth M.
 Experience-dependent recovery of vision following chronic deprivation amblyopia Hai-Yan He, Baisali Ray, Katie Dennis and Elizabeth M. Quinlan a 3. 2.5 2. 1.5.5 Deprived eye Non-deprived VCtx * * b 8.
Experience-dependent recovery of vision following chronic deprivation amblyopia Hai-Yan He, Baisali Ray, Katie Dennis and Elizabeth M. Quinlan a 3. 2.5 2. 1.5.5 Deprived eye Non-deprived VCtx * * b 8.
This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail.
 This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail. Author(s): Nokia, Miriam; Lensu, Sanna; Ahtiainen, Juha; Johansson,
This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail. Author(s): Nokia, Miriam; Lensu, Sanna; Ahtiainen, Juha; Johansson,