on October 4, 2018 by guest
|
|
|
- Joel Mathews
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 22 December 2010 J. Virol. doi: /jvi Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. 1 The IFITM proteins inhibit HIV-1 infection Jennifer Lu 1,2, Qinghua Pan 1, Liwei Rong 1, Shan-Lu Liu 3 and Chen Liang 1,2,3 McGill AIDS Centre, Lady Davis Institute-Jewish General Hospital 1, Montreal, Quebec, Canada H3T 1E2. Department of Medicine 2, Department of Microbiology and Immunology 3, McGill University, Montreal, Quebec, Canada H3A 2B4. Word count: Abstract, 126; Text, Address correspondence to: Chen Liang, Ph.D. chen.liang@mcgill.ca Tel: Fax: Jennifer Lu and Qinghua Pan contributed equally to this work. Downloaded from on October 4, 2018 by guest 1
2 ABSTRACT Type I interferon protects cells from virus infection through the induction of a group of genes collectively named interferon stimulated genes (ISGs). In this study, we have utilized shrna to deplete ISGs in SupT1 cells in order to identify ISGs that suppress the production of human immunodeficiency virus type 1 (HIV-1). Among the ISG candidates thus identified are interferon induced transmembrane (IFITM) proteins, including IFITM1, IFITM2 and IFITM3, that potently inhibit HIV-1 replication at least partially through interfering with virus entry. Further mutagenesis analysis shows that the intracellular region, rather than the N- and C-terminal extracellular domains, is essential for the antiviral activity of IFITM1. Altogether, these data suggest that the IFITM proteins serve as important components of the innate immune system to restrict HIV-1 infection. Downloaded from on October 4, 2018 by guest 2
3 INTRODUCTION Mammalian cells produce type I interferon in response to virus infection (reviewed in references (8, 25)). Viral proteins and viral nucleic acids are detected by pathogen recognition receptors (PRRs) as pathogen associated molecular patterns (PAMPs). These PRRs are either membrane-associated, such as toll-like receptor 3 (TLR3) and TLR7 on endosomes, or cytosolic such as retinoic acid-inducible gene I (RIG-I), melanoma differentiation-associated gene 5 (MDA5), nucleotide-binding oligomerization domain containing 2 (NOD2), and absent in melanoma 2 (AIM2). These recognition events activate signaling transduction pathways that trigger production of type I interferon and other cytokines. Type I interferon then augments the expression of hundreds of genes named interferon stimulated genes (ISGs) that subvert virus replication by a variety of mechanisms (reviewed in references (50, 53)). Human immunodeficiency virus type 1 (HIV-1) infection also triggers type I interferon production (17). Plasmacytoid dendritic cells, the major interferon producers, release large quantities of interferon partially through recognition of HIV-1 RNA by TLR7 (23). Although the clinical benefit of engaging interferon in treating HIV-1 infection is uncertain due to the controversial results of clinical trials (16, 31, 51, 60), HIV-1 replication in cultured cells is clearly inhibited by interferon (4, 19, 22, 24, 48, 49, 68). This inhibition must have exerted a selection pressure on HIV-1 in vivo in light of the multiple measures that the virus has evolved to evade interferon restriction. For example, HIV-1 infection causes a gradual decrease in the number of plasmacytoid dendritic cells as well as a reduced capacity of these cells to produce interferon (13, 35, 62). In addition, 3
4 HIV-1 infection causes depletion of interferon regulatory factor 3 (IRF-3) that plays a key role in toll-like receptor and RIG-I mediated innate immune signaling (12, 45). Equally important, HIV-1 encodes proteins that antagonize individual ISGs. Examples include Vpu (viral protein u) that counteracts BST-2 (bone marrow stromal cell antigen 2, also known as tetherin/cd317/hm1.24) that otherwise blocks virus release from the cell surface (44, 64), and Vif (viral infectivity factor) that antagonizes APOBEC3G (apolipoprotein B mrna editing enzyme 3G), a cytidine deaminase that causes hypermutation of HIV-1 cdna (57). Interferon impedes multiple steps of HIV-1 replication at both the early and late phases (1, 11, 18-22, 24, 28-30, 40, 49, 58, 59, 67, 68). These inhibition activities are the functions of distinct ISGs. For example, PKR (protein kinase R) inhibits HIV-1 production by suppressing protein translation (43). RNase L degrades HIV-1 RNA, thus restricts HIV-1 replication (36). TRIM22 (tripartite motif protein 22) disrupts Gag assembly (3). BST-2 blocks the release of HIV-1 particles from the cell surface (44, 64). ISG15 impedes HIV-1 virus production by causing ISGylation of viral Gag protein and cellular factors such as CHMP5 (chromatin modifying protein 5) (46, 47). APOBEC3G restricts HIV-1 replication by causing hypermutation of viral cdna during reverse transcription (37, 57). TRIM5alpha from old world monkeys targets viral core following HIV-1 entry and destroys viral reverse transcription complex (37, 63). Although APOBEC3G and TRIM5alpha are constitutively expressed, their levels are elevated by interferon, which potentiates their antiviral function (10, 54). Recent studies by Brass et al identified the interferon-induced transmembrane (IFITM) proteins as potent inhibitors 4
5 of influenza A H1N1 virus, West Nile virus and dengue virus in their genome-wide RNAi screen (6). In this study, we further show that IFITM proteins also inhibit HIV-1 replication MATERIALS AND METHODS Plasmid DNA, viruses and antibodies. The cdna clones of IFITM1, IFITM2 and IFITM3 were kindly provided by Ju-Tao Guo (26). The cdna sequences of these ifitm genes were amplified by PCR using primer pairs IFITM1-S/IFITM1-A, IFITM2-S/IFITM2-A and IFITM3-S/IFITM3-A (primer sequences are provided in supplementary Table S1). The PCR products were digested with restriction enzymes BamHI and EcoRI, and inserted into the pretrox-tight-pur retroviral vector (Clontech) to create DNA constructs Tet-IFITM1, Tet-IFITM2 and Tet- IFITM3. A Flag tag was attached to the N-terminus of each IFITM. IFITM1 mutants were generated by PCR using primers that are listed in supplementary Table S1. The infectious HIV-1 proviral DNA clone BH10 was obtained from the NIH AIDS Research and Reference Reagent Program. The NLEY1-IRES and the NLEY1-ES-IRES DNA constructs were kindly provided by David Levy (33), the pcmv-blam-vpr plasmid DNA by Warner Greene (9). HIV-1 stocks were generated by transfecting 293T cells with proviral DNA clones BH10, NLEY1-IRES or NLEY1-ES-IRES. It is noted that both the BH10 and NLEY1-IRES viruses are infectious, whereas the NLEY1-ES-IRES virus lacks HIV-1 envelope protein and needs to be pseudotyped for entry. When necessary, the vesicular stomatitis virus (VSV) glycoprotein (G) was used to pseudotype HIV-1 5
6 97 98 particles. Amounts of viruses in the stock were determined by measuring the activity of viral reverse transcriptase or the level of viral CA(p24) antigen Anti-Flag and anti-tubulin antibodies were purchased from Sigma, anti-hiv-1 p24 antibody from ID Lab Inc., anti-ifitm1, anti-ifitm2 and anti-ifitm3 antibodies from Proteintech Group, PE-conjugated anti-human CD4 antibody from BD Biosciences. IFN α2b and G418 were purchased from Invitrogen, puromycin and doxycycline from Sigma. Microarray SupT1 cells were cultured in RPMI1640 media supplemented with 10% FBS, 1% penicillin/streptomycin, and 1% glutamine. Following treatment with IFN α2b (1000 U/ml) for 8 or 16 hours, total cellular RNA was extracted using the Trizol reagent (Invitrogen). RNA samples were submitted to McGill University and Génome Québec Innovation Centre for microarray analysis using the Illumina expression BeadChip. Screening for anti-hiv-1 ISGs The MISSION shrna transduction particles were purchased from Sigma to knock down ISGs in SupT1 cells. Each ISG was targeted on average by 3 to 5 shrna clones. A control shrna was also purchased from Sigma that has the scrambled RNA target sequence. HIV-1 stock was prepared by transfecting 293T cells with a subtype B HIV-1 DNA named BH10 together with a plasmid expressing the glycoprotein (G) of VSV. VSV G protein was used to enhance infection efficiency of wild type HIV-1 particles, 6
7 and the VSV G-containing wild type HIV-1 particles were only used in screening experiments. The level of viruses was titrated by infecting TZM-bl indicator cells (65) First, stable SupT1 cell lines were created to express each individual shrna. This was achieved by infecting 1x10 6 SupT1 with 50 µl of the MISSION shrna transduction particles supplemented with polybrene (5 µg/ml) in a 24-well plate. After 48 hours, puromycin (2 µg/ml) was added to select for stably transduced cells. Cells (10 6 ) were then treated with IFN α2b (500 U/ml) for 16 hours before infection with HIV-1. After the inoculums were removed, cells were kept in culture for a further 24-hour period before the levels of infectious HIV-1 in the supernatants were measured by infecting the TZM-bl cells. An ISG is considered as a candidate if more than two of its shrna clones alleviated IFN α2b-posed restriction by above 2 fold. The candidate ISGs were then tested in a second round of HIV-1 infection for validation. Creating doxycycline-inducible IFITM cell lines Retroviral particles were prepared by transfecting 1 µg of each of the Tet-IFITM1, Tet- IFITM2 and Tet-IFITM3 plasmid DNA into the packaging cells GP2-293 (Clontech) together with 0.1 µg of the VSV G DNA. These viruses were used to infect SupT1 cells together with viruses expressing the rtta activator (Clontech). Stably transduced cell lines were selected in the RPMI1640 medium supplemented with puromycin (2 µg/ml), G418 (1 mg/ml) and tetracycline-free serum (Clontech). To check the expression of IFITM proteins, cell lines were exposed to doxycycline (500 ng /ml) for 16 hours before the cell lysates were collected and examined by Western blots with an anti-flag antibody 7
8 (Sigma). Doxycycline-inducible IFITM MT-2 and 293 cell lines were created following the same protocol HIV-1 virion fusion assay The fusion assay was performed as described (9). HIV-1 particles containing the β- lactamase-vpr (BlaM-Vpr) chimera proteins were produced by transfecting 293T cells with 3 µg of the BH10 DNA and 1 µg of pcmv-blam-vpr DNA using Lipofectamine 2000 (Invitrogen). Virus particles thus generated bear HIV-1 envelope proteins gp120/gp41. After 48 hours, the culture supernatants were first clarified by passing through a 0.22 µm filter, and then concentrated by ultracentrifugation at 100,000xg for 1 hour at 4 o C. Pelleted viruses were suspended in complete DMEM media, aliquoted, and stored at -80 C. SupT1 cell lines were exposed to doxycycline (500 ng/ml) for 16 hours before spinoculation with HIV-1 particles containing BlaM-Vpr. After a 2-hour incubation at 37 C, cells were washed with CO 2 -independent medium (Invitrogen) and loaded with CCF2/AM substrate (Invitrogen) by incubating cells with 100 µl of loading solution for 1 hour at room temperature in the dark. The loading solution was prepared by mixing 2 µl of CCF2/AM (1 nm) with 8 µl of 0.1% acetic acid containing 100 mg/ml Pluronic- F127surfactant (solution B provided by Invitrogen with the CCF2/AM loading kit) and 1 ml of CO 2 -independent medium. Finally, cells were washed with 200 µl of development media and the BlaM reaction was developed in 200 µl development media for 16 hours at room temperature in the dark. The development media was prepared by mixing 10 µl of 8
9 probenecid (250 mm) with 1 ml CO 2 -independent medium and 100 µl tetracycline-free FBS. After washing once with cold 1xphosphate buffered saline (containing 2% FBS), cells were fixed with 1% paraformaldehyde. The levels of CCF2/AM and its cleaved products were measured by flow cytometry. sirna knockdown of IFITM proteins in TZM-bl cells Short interfering RNA (sirna) oligos were purchased from Ambion to target IFITM1 (sirna ID #: s16192 (GGUCCACCGUGAUCAACAUtt)), IFITM2 (sirna ID #: s20771 (CCACGUACUCUAUCUUCCAtt), s (GCCCUUGACCUGUAUUCC Att)), and IFITM3 (sirna ID #: s (CCCACGUACUCCAACUUCCtt)). TZM-bl cells were seeded in 24-well plates one day before sirna transfection was performed with Lipofectamine 2000 (Invitrogen). Two sirna mixtures were used to simultaneously deplete IFITM1, IFITM2 and IFITM3, these mixtures are (s16192/s20771/s195035) and (s16192/s230492/s195035). An amount of 10 nm of each sirna was used in transfection. After two sequential sirna transfections, cells were exposed to the wild type HIV-1 named BH10 for 2 hours before viruses were washed off. Luciferase activity in TZM-bl cells was measured 40 hours later. Levels of IFITM proteins were assessed by Western blotting using anti-ifitm1, anti-ifitm2, and anti-ifitm3 antibodies, respectively. Virus infection assays IFITM SupT1 cells (10 6 cells for each line) were treated with doxycycline (500 ng/ml) for 16 hours before being infected with HIV-1 named NLEY1-IRES equivalent to 100 ng 9
10 CA(p24). The NLEY1-IRES virus bears HIV-1 envelope protein, is infectious and expresses the yellow fluorescent protein (YFP) (33). The inoculated viruses were washed off after an overnight incubation. After 24 hours, cells were washed with cold 1x phosphate buffered saline and fixed with 1% paraformaldehyde (in 1x phosphate buffered saline). YFP-positive cells were scored by flow cytometry. Amounts of viruses in the supernatants were determined by infecting the TZM-bl indicator cells. For spreading infection, 2x10 6 IFITM SupT1 cells were induced with doxycycline (500 ng/ml) for 16 hours before being infected with wild type HIV-1 (BH10) equivalent to 20 ng CA(p24). Viral replication was monitored by measuring levels of viral reverse transcriptase activity in supernatants over different time intervals. Measuring HIV-1 reverse transcription products. SupT1 cells (2x10 6 ) were exposed to the wild type BH10 viruses (equivalent to 100 ng CA(p24)) for 1 hour at 37 o C before the virus inoculums were washed off. As a control, nevirapine (1 µm) was used to block reverse transcription. Cells were collected 16 hours after infection, total DNA was extracted with the DNeasy tissue kit (Qiagen). Real-time PCR was performed with equal DNA amounts of different samples using the following primer pairs: 5 - TTAGACCAGATCTGAGCCTGGGAG-3 /GGGTCTGAGGGATCTCTAGT TACC-3 to amplify the early viral DNA and 5 -TGTGTGCCCGTCTGTTGTG-3 /5 - GAGTCCTGCGTCGAGA-3 to amplify the late viral DNA (34). The reactions were performed with the LightCycler FastStart DNA Master SYBR Green 1 system (Roche) in accordance with manufacturer s instructions. The PCR conditions are 95 o C for 10 10
11 seconds, 62 o C for 5 seconds and 72 o C for 7 seconds. In order to measure the viral DNA that is integrated into cellular chromosomal DNA, the first round of PCR was performed with primers 5 GCCTCCCAAAGTGCTGGGATTACAG-3 /5 -GTTCCTGCTATGTC ACTTCC-3 that bind to Alu and HIV-1 Gag DNA sequences. The reactions were performed at 94 o C for one minute to denature DNA templates followed by 12 cycles at 94 o C for 30 seconds, 50 o C for 30 seconds and 72 o C for 3.3 minutes. The amplified DNA products were quantified in real-time PCR as described above with primers 5 - TTAAGCCTCAATAAAGCTTGCC-3 /5 -GTTCGGGCGCCACTGCTAGA-3 that amplify HIV-1 LTR region (73). Immunofluorescence microscopy. The 293 cells were seeded into slide chambers prior to transfection with vector DNA expressing Flag tagged IFITM1, IFITM2 and IFITM3. Forty hours after transfection, cells were rinsed with serum-free DMEM and incubated with Alexa fluor 555-conjugated transferrin (5 µg/ml in serum-free DMEM, Invitrogen) for 10 minutes before fixation in 4% paraformaldehyde (in 1x phosphate buffered saline) for 10 minutes at room temperature. After permeabilization with 0.1% Triton X-100 for 10 minutes at room temperature and incubation in a blocking solution (2% FBS) for 2 hours, cells were stained with the rabbit anti-flag primary antibody (1:500 dilution, 2 hours at room temperature) and the Alexa fluor 488-conjugated secondary anti-rabbit antibody (1:2000 dilution, 40 minutes at room temperature). Images were taken using the Zeiss PASCAL laser scanning confocal microscope. 11
12 RESULTS Knockdown of IFITM1 increases HIV-1 production in the presence of IFN α2b. With an aim to identify ISGs that have anti-hiv-1 activity, we first tested the effect of interferon α2b (IFN α2b ) on HIV-1 production in several human cell lines as well as in cord blood mononuclear cells (CBMCs) in order to find a cell line that is suitable for the screening assay. The results showed a greater than 100 fold decrease of HIV-1 production in both SupT1 and CBMCs following IFN α2b treatment as opposed to a less pronounced HIV-1 reduction in Jurkat, U937 and 293T cells (supplementary Fig. S1). We thus proceeded with SupT1 cells and performed microarray to determine the genes that were up-regulated by IFN α2b. A total of 94 genes showed a more than 2 fold increase in their mrna expression (supplementary Table S2). In order to determine which of these ISGs inhibit HIV-1 production, we used the MISSION shrna products from Sigma to knock down each individual ISG. Sixty-seven out of these 94 ISGs were covered by shrna clones that were available at Sigma (supplementary Table S2). We first created stably transduced shrna SupT1 cell lines, including a scrambled shrna cell line as the control, then treated these cells with IFN α2b for 16 hours before infection with wild type HIV-1 (BH10). Production of infectious HIV-1 particles was monitored by infecting the TZM-bl indicator cells (65). An ISG was selected as an anti-hiv-1 candidate if at least two of its shrna clones increased virus production by above 2 fold than that from the control cell line in the presence of IFN α2b. Six such ISGs were identified, they are IFITM1, IFI44, PARP14, RTP4, EPSTI1 and TDRD7 (Fig. 1A-C). Further experiments showed that IFITM1 was substantially downregulated by each of the five shrna clones and that four out of these five shrna increased HIV-1 production by above two fold 12
13 (Fig. 1D). It is notable that interferon-mediated inhibition of HIV-1 production was not fully alleviated by knocking down any of the 67 ISGs. It is therefore possible that the ISG with the strongest inhibition activity was not covered in the 67 ISGs tested in this study, or that multiple ISGs were needed to strongly suppress HIV-1 infection. IFITM proteins diminish HIV-1 replication in SupT1 cells. In order to assess the anti- HIV-1 activity of an individual ISG more specifically, we utilized the Tet-On expression system and generated SupT1 stable cell lines that were able to express a single ISG in response to tetracycline (or doxycycline). With this approach, we found that doxycyclineinduced IFITM1 strongly inhibited HIV-1 replication in SupT1 cells (Fig. 2A, B and C), which is consistent with the increased HIV-1 production seen with shrna knockdown of IFITM1 (Fig. 1D). IFITM1 has three homologs in the human genome, these include IFITM2, IFITM3 and IFITM5 (38, 42). Unlike IFITM1, 2 and 3, IFITM5 is solely expressed in osteoblasts and is involved in bone mineralization (42), thus is not further investigated in this study. When the effect of IFITM2 and IFITM3 on HIV-1 replication was examined using the Tet-On system, we found that IFITM2 and IFITM3 also markedly suppressed HIV-1 infection albeit to a lesser extent than that of IFITM1 (Fig. 2C). Further experiments showed that the induced expression of IFITM1, IFITM2 or IFITM3 did not affect cell proliferation, cell cycle or cell surface expression of CD4 (supplementary Fig. S2), supporting a likely direct inhibition of HIV-1 replication by IFITM1, IFITM2 and IFITM3. 13
14 IFITM2 and IFITM3 impede HIV-1 entry. We next asked at what step(s) HIV-1 replication was affected by IFITM proteins. To answer this question, we used in the following one-cycle infection assays an infectious HIV-1 reporter virus named NLEY1- IRES that expresses yellow fluorescence protein (YFP) (33). Cells were first treated with doxycycline to induce IFITM expression, then infected with the NLEY1-IRES virus for 40 hours. The number of YFP-positive cells was scored by flow cytometry (Fig. 3A), and the amounts of infectious viruses in supernatants were determined by infecting the TZMbl indicator cells (Fig. 3B). The results showed that IFITM2 and IFITM3 reduced the number of infected cells by 40-50%, and decreased the amounts of infectious viruses by 4 to 5 fold; interestingly, IFITM1 exerted a less pronounced inhibition effect (Fig. 3A, B). Similar observations were made in MT-2 cells (supplementary Fig. S3). When SupT1 cells were infected by the NLEY1-IRES virus prior to the addition of doxycycline, neither the number of infected cells nor the amounts of infectious HIV-1 particles was evidently affected by any of the induced IFITM proteins (Fig. 3C). These results indicate that IFITM proteins likely target an early step in HIV-1 replication. In support of this notion, HIV-1 production was not affected by ectopic expression of IFITM proteins in 293 cells that were transfected with HIV-1 DNA clone BH10 (data not shown). We also measured the effect of IFITM proteins on infection by HIV-1 named NLEY1-ES-IRES that was pseudotyped with VSV G protein. The results revealed that IFITM3 reduced virus infection by approximately 55% whereas IFITM1 and IFITM2 led to 30% reduction (Fig. 3D). The more dramatic impact of IFITM3 on VSV G protein-mediated infection is consistent with a recent report showing that IFITM3 inhibits VSV infection by targeting distinct steps of virus life cycle (66). 14
15 We then performed HIV-1 virion fusion assay to test whether IFITM proteins affect virus entry. The fusion virus particles were prepared by transfecting 293T cells with an infectious HIV-1 proviral DNA clone BH10 together with the BlaM-Vpr DNA. The HIV- 1 entry inhibitor T20 (1 µg/ml) effectively blocked the entry of BH10/BlaM-Vpr viruses, but did not affect the entry mediated by VSV G protein (Fig. 4A). When the BH10/BlaM- Vpr viruses were used to infect the IFITM cell lines, IFITM2 and IFITM3 diminished HIV-1 entry by 2 to 3 fold whereas IFITM1 did not exert any effect in this regard (Fig. 3B, C). Since IFITM1 differs from IFITM2 and IFITM3 mainly by the lack of an amino acid stretch in the N-terminal region, we speculated that the elongated N terminal region may allow IFITM2 and IFITM3 to affect virus entry. To test this, we created three IFITM3 mutants named (1-6), (1-11) and (1-16) that lacked various lengths of the N terminal sequences (Fig. 4D). The results of HIV-1 virion fusion assays showed that the (1-6) mutant and the wild type IFITM3 diminished virus entry by about 40% as opposed to a 20% reduction caused by the (1-11) and the (1-16) mutants (Fig. 4E), suggesting a role of the 7-FFSPV-11 peptide in restricting virus entry. Taken together, these data suggest that IFITM2 and IFITM3 interfere with HIV-1 entry, which represents one mechanism underlying their anti-hiv-1 activity. IFITM2 and IFITM3 associate with endocytosis. Virus entry involves binding to the receptor molecules on cell surface and subsequent endocytosis of virus particles (39). Recent studies suggest that HIV-1 entry also involves the endocytosis process (41). Different viruses escape the endosome pathway at different stages. Some viruses such as 15
16 influenza virus travel to the late endosomes to accomplish membrane fusion. We envision that in order to elicit a direct effect on virus entry, IFITM proteins may need to associate with the endocytosis pathway. To test this, we fed cells with Alexa fluor 555-conjugated transferrin and determined whether IFITM proteins co-localize with the endocytosed transferrin. Although IFITM1 barely exhibited any co-localization with transferrin, a strong association was observed between the endocytosed transferrin with IFITM2 or IFITM3 (Fig. 5). This result suggests that IFITM2 and IFITM3, but not IFITM1, associate with the endocytosis pathway and may exert an effect on the fate of the endocytosed cargos such as virus particles. IFITM1 suppresses HIV-1 Gag production. IFITM1 does not affect HIV-1 entry, yet efficiently inhibits virus replication in SupT1 cells. In order to gain insights into this restriction activity of IFITM1, we first measured the effect of IFITM1 on HIV-1 reverse transcription by real-time PCR. Viral DNA was not detected by PCR in SupT1 cells that were infected by HIV-1 in the presence of a reverse transcriptase inhibitor named nevirapine (1 µm) (data not shown), which validates that the amplified DNA products from HIV-1 infected cells in the absence of nevirapine treatment are reverse transcribed viral DNA. Consistent with the 40% to 50% decrease of virus entry caused by IFITM2 and IFITM3, both the early and the late HIV-1 cdna products diminished by a similar magnitude (Fig. 6A). No effect of IFITM1 was observed on the production of HIV-1 cdna (Fig. 6A). We next performed Alu-PCR to assess the level of HIV-1 DNA that was integrated into cellular chromosomal DNA. The results did not reveal any effect of IFITM1 on HIV-1 DNA integration (Fig. 6A). Interestingly, when HIV-1 Gag protein 16
17 and its processing products including p40 and p24 were examined in Western blots, a significant reduction was detected in SupT1 cells that expressed either of the three IFITM proteins (Fig. 6B), suggesting that IFITM1 as well as IFITM2 and IFITM3 adversely affect viral Gag expression, which must have led to diminished virus production. The C-terminal sequence of IFITM1 regulates its antiviral activity. It is interesting to note that IFITM1 does not affect HIV-1 entry but inhibits HIV-1 replication in SupT1 cells more severely than IFITM2 and IFITM3. In order to understand the antiviral activity of IFITM1, we performed mutagenesis studies to determine which domains of IFITM1 are critical for inhibiting HIV-1 replication. IFITM1 can be divided into five domains interspersed by its two transmembrane domains (Fig. 7A). We first deleted the C-terminal sequence of IFITM1 and generated three mutants named ( ), ( ) and ( ) (Fig. 7A, B). The results of spreading infection assays showed that these three mutants inhibited HIV-1 replication in SupT1 cells even more effectively than did the wild type IFITM1 (Fig. 7C). Consistent with this, these mutants reduced the number of infected cells and the amounts of infectious virus particles to a higher degree than the wild type IFITM1 (Fig. 7D). In order to exclude the possibility that the increased antiviral activity of these IFITM1 mutants is a result of their artificially high expression, we compared the doxycycline-induced expression of one mutant ( ) to that of the IFN α2b-induced endogenous IFITM1. Comparable levels between ( ) and endogenous IFITM1 were shown by the results of Western blots (Fig. 7E). These data suggest that the C-terminal sequence of IFITM1 is not only dispensable for the antiviral 17
18 activity of IFITM1, but that its presence may rather impair the antiviral function of this protein We next assessed the role of the N-terminal sequence of IFITM1 in inhibiting HIV-1 replication. Two mutants were accordingly created by deleting either 21 or 29 amino acids from the N-terminus in the context of the ( ) deletion that not only inhibited HIV-1 replication more strongly than wild type IFITM1 but also severely diminished the number of infected cells in the one-cycle infection assay. Despites its low expression level, the (1-21)/ ( ) mutant inhibited HIV-1 replication as efficiently as the wild type IFITM1 and the ( ) mutant (Fig. 8). HIV-1 replication was not affected by the (1-29)/ ( ) mutant due to the loss of its expression (Fig. 8). These results suggest that at least the first 21 amino acids of the N- terminal region are not required for IFITM1 to inhibit HIV-1 replication. The intracellular region of IFITM1 was also subjected to mutagenesis analysis by changing every four amino acids to alanines. Eight mutants were thus created (Fig. 8A). Results of Western blots showed that these mutants were not equally well expressed (Fig. 8B). Nevertheless, the VTG and QY mutants suppressed HIV-1 replication as effectively as the wild type IFITM1 and the ( ) mutant (Fig. 8C, D). MVGK and ST partially repressed HIV-1 replication, whereas the SVKS, RDRK, KC and LNI mutants did not exert an evident inhibitory effect (Fig. 8C, D). However, given the low expression levels of these latter six mutants, the data are insufficient to conclude a direct involvement of the mutated amino acids in inhibiting HIV-1 infection. Nonetheless, the 18
19 results do suggest that the amino acids altered in the VTG and QY mutants do not have a role in the IFITM1 anti-hiv-1 function Knockdown of IFITM proteins increases susceptibility of TZM-bl cells to HIV-1 infection. IFITM proteins were recently shown to inhibit infection of influenza A virus, West Nile virus and Dengue virus (6, 27). However, an inhibition of HIV-1 infection was not measured in the HeLa-CD4+ cells when IFITM3 was knocked down with sirna oligos (6). Consistent with this, we also failed to observe an increase in HIV-1 infection when depleting each of the three IFITM proteins in the TZM-bl cells (data not shown). However, we subsequently noted that TZM-bl, which is a HeLa-derived HIV-1 indicator cell line (65), expresses considerable amounts of IFITM2 and IFITM3 proteins (Fig. 9A). Since both IFITM2 and IFITM3 inhibited HIV-1 entry, we decided to deplete all three IFITM proteins by transfecting the TZM-bl cells with sirna oligo mixtures that target all three ifitm genes (Fig. 9B). Indeed, this treatment led to a 2 to 3 fold increase in HIV-1 infection (Fig. 9C), which suggests that the endogenous IFITM proteins play a role in restricting HIV-1 infection. DISCUSSION There exist four human ifitm genes including ifitm1, ifitm2, ifitm3 and ifitm5 (38, 42). IFITM5 is expressed in osteoblasts and plays a role in bone mineralization (42). IFITM1, IFITM2 and IFITM3 are ubiquitously expressed with IFITM1 and IFITM3 responding to interferon stimulation due to the presence of a functional ISRE (interferon stimulation response element) in their promoters (38). IFITM proteins have been reported to play a 19
20 role in a number of cell functions such as oncogenesis (7, 15), cell adhesion (14) and immune cell signaling (5). High levels of IFITM proteins have also been observed in certain types of tumors such as colorectal tumors and astrocytoma (2, 56). Nonetheless, these proteins do not seem to play a vital role in embryogenesis, since mice that are knocked out for all ifitm genes appear to develop normally (32). As interferon stimulated genes, the antiviral function of IFITM proteins has not been well appreciated until the recent discovery of their strong inhibition activity against influenza A virus, West Nile virus, Dengue virus and vesicular stomatitis virus (6, 27, 66). Studies also show that palmitoylation of the cysteine residues are required for the antiviral function of IFITM3 (72). Our current study demonstrates that IFITM proteins also inhibit HIV-1 infection, thus expanding the spectrum of viruses under the control of this small ISG family. One mechanism behind the restriction activity of IFITM proteins involves impeding virus entry. Studies by Brass and colleagues have shown that MLV particles that were pseudotyped with envelope proteins from influenza A virus, West Nile virus, and Dengue virus, but not from alphaviruses, were sensitive to restriction by IFITM proteins (6). One implication of this observation is that IFITM proteins inhibit the entry step that is mediated by the envelope proteins of certain viruses. This function of IFITM is now supported by the results of an assay that directly measures HIV-1 entry (Fig. 4). Brass and colleagues depleted the endogenous IFITM3 protein in a CD4+ HeLa cell line and did not observe an effect on infection by an HIV-1 IIIb strain (6). This is most likely because HeLa cells express all three IFITM proteins and each IFITM protein is a potent 20
21 inhibitor of HIV-1 infection. Indeed, when we performed similar experiments in TZM-bl cells that are HeLa-derived CD4-expressing cells, an increase in HIV-1 infection was observed only when all three IFITM proteins were simultaneously depleted by sirna oligos (Fig. 9). IFITM proteins are membrane-associated, a feature that is considered necessary for directly interfering with virus entry (6). The IFITM protein family is characterized by two transmembrane domains and a highly conserved intracellular region. The N- and C- termini project away from the plasma membrane toward the extracellular environment (6, 61, 66). Interestingly, this topological presentation inversely mirrors that of BST-2 that inhibits the release of a variety of enveloped viruses from the cell surface (reviewed in reference (55)). Although the molecular mechanism behind the IFITM restriction action remains to be elucidated, it seems unlikely that they directly target viral envelope proteins or viral receptors given the different viruses that IFITM proteins inhibit. Since virus entry often proceeds through endocytosis (reviewed in reference (39)), it is possible that IFITM proteins are involved in endocytosis such as to detect and eliminate virus invaders before infection is established. In support of this speculation, it has been shown that IFITM1 is associated with the CD81/antibody complexes when they are endocytosed by cells (61). Our results further demonstrate an association of IFITM2 and IFITM3 with endocytosed transferrin (Fig. 5). Among the viruses tested thus far, arenaviruses are resistant to restriction by IFITM proteins (6). This suggests that arenavirus envelope proteins may have a mechanism to 21
22 evade IFITM restriction. Entry of arenaviruses, such as Lassa virus (LASV), lymphocytic choriomeningitivirus (LCMV) and machupo virus (MACH), begins with endocytosis at the plasma membrane, and completes at the late endosome with low-ph-triggered membrane fusion (reviewed in reference (52)). It is noted that LASV and LCMV exploit alpha-dystroglycan (alpha-dg) as the receptor, and enter cells via an endocytosis process that is independent of clatherin, dynamin and caveolin; whereas MACH uses transferrin receptor 1 to enter cells via clatherin-mediated endocytosis. In addition, the glycoproteins (GPs) of arenaviruses contain a stable signal peptide (SSP) of unusual length that becomes an integral component of the mature GP complex (including GP1, GP2 and SSP) (69-71). This SSP component may have a role in communicating the cytoplasmic domain (CTD) with the fusion-active ectodomain, which represents a unique feature of arenavirus GP-mediated entry. Further studies should determine which of these features or other unknown properties of arenavirus GPs allow these viruses to be resistant to IFITM proteins. Targeting virus entry may not be the only mechanism by which IFITM proteins inhibit HIV-1 replication. For example, the 2 3-fold decrease in entry as a result of IFITM2 or IFITM3 expression does not seem to fully account for the over 5-fold decrease in HIV-1 production as being measured in the one-cycle virus infection assay (Fig. 3B). It is possible that the results of the virion fusion assay may have underestimated the impact of IFITM proteins on virus entry in the context of a natural HIV-1 infection during which cells are exposed to a much lower dose of viruses compared with a large quantity of concentrated HIV-1 particles that were used for the virion fusion assay. It is also notable 22
23 that IFITM1 does not affect HIV-1 entry, yet strongly inhibits HIV-1 replication. This latter observation adds to the argument that IFITM proteins may affect more than one step of HIV-1 replication The effect of interferon on HIV-1 entry was recently assessed by Goujon and Malim in a number of human immune cell lines (20). Their results revealed a moderate inhibitory effect in CD4+ T cells, which is, to some extent, consistent with the 2 to 3 fold reduction in HIV-1 entry seen with IFITM2 or IFITM3 in our study (Fig. 4). Given the constitutive expression of IFITM proteins in different cell lines (Fig. 9A), the exact impact of IFITM proteins on HIV-1 entry awaits further experimentation using IFITM knockout cells. In summary, our results demonstrate that IFITM proteins restrict HIV-1 infection. Given the evidence that other viruses including influenza A virus, West Nile virus and Dengue virus are inhibited by IFITM proteins, this small ISG family emerges as an integral component of the interferon-mediated innate immune system that helps to defend against important human viral pathogens. ACKNOWLEDGEMENTS We thank Warner Greene, David Levy and Ju-Tao Guo for providing valuable reagents, and Vicky Cheng for critically reading the manuscript. This work was supported by funding from the Canadian Institutes of Health Research and the Canadian Foundation for AIDS Research. 23
24 FIGURE LEGENDS Figure 1. Effect of ISG knockdown on HIV-1 production in the presence of IFN α2b. Each ISG was targeted with 3 to 5 shrna clones. The shrna transduced SupT1 cells were treated with IFN α2b for 16 hours before being infected with a subtype B HIV-1 named BH10. Production of infectious HIV-1 particles was determined by infecting the TZM-bl indicator cells. Fold of change for each shrna was calculated by dividing the value of infectious HIV-1 made from the shrna SupT1 cells by that from the control shrna SupT1 cells. The results are summarized in three graphs (A-C). The candidate genes are highlighted by arrows. (D). Western blots of SupT1 cells that were stably transduced with either the control shrna vector or the shifitm1 clones. Cells were treated with IFN α2b (500 U/ml) for 20 hours prior to Western blotting analysis. Fold change of HIV-1 production for each shifitm1 clone is shown. Results shown are the average of two independent infection experiments. Figure 2. IFITM proteins inhibit HIV-1 replication in SupT1 cells. (A) Illustration of the domain structures of IFITM proteins. The transmembrane domains are highlighted in boxes. A Flag tag was attached to the N-termini of IFITM proteins. The conserved amino acid residues are highlighted in red letters. (B) Doxycycline-induced expression of IFITM proteins in SupT1 cells. Stably transduced SupT1 cells were exposed to different amounts of doxycycline (0, 50 ng/ml, 500 ng/ml) for 16 hours before cell lysates were harvested and subjected to Western blots using anti-flag antibodies to detect the expression of IFITM proteins. Control represents a cell line that was stably transduced with the empty retroviral vector. (C) HIV-1 replication in IFITM-expressing SupT1 cells. Cells were 24
25 treated with doxycycline (500 ng/ml) for 16 hours before they were exposed to wild type HIV-1. Virus production over different time intervals was monitored by measuring viral reverse transcriptase activity in the culture supernatants. Results shown represent four independent infection experiments. Figure 3. IFITM proteins inhibit an early step of HIV-1 replication. (A) IFITM2 and IFITM3 diminished the number of HIV-1 infected cells. Following doxycycline treatment (0, 500 ng/ml) for 16 hours, SupT1 cells were infected with the NLEY1-IRES virus that expresses YFP. Forty hours after infection, the number of YFP-positive cells was scored by flow cytometry. Results of three independent experiments are summarized in the bar graph shown in (B). (B) IFITM proteins suppressed production of infectious HIV-1 particles. The amounts of infectious virus particles were determined by infecting the TZM-bl indicator cells. Fold of change is calculated by dividing the values from doxycycline treated cells (+Dox) with the values from non-treated cells (-Dox). Results shown are the average of three independent infection experiments. (C) SupT1 cells were infected with the NLEY1-IRES virus for 2 hours before exposure to doxycycline (500 ng/ml) treatment. Effects of IFITM proteins on the number of infected cells and the amounts of infectious HIV-1 are summarized in the bar graph. (D) The IFITM SupT1 cell lines were first treated with Dox (500 ng/ml) for 16 hours, then infected with the NLEY1- ES-IRES viruses that were pseudotyped with VSV G protein. The number of YFPpositive cells was scored 40 hours after infection. Results of three independent infections were summarized in the bar graph. 25
26 Figure 4. Effect of IFITM proteins on HIV-1 entry. (A) T20 blocks the infection of SupT1 cells by the BH10/BlaM-Vpr viruses. SupT1 cells were infected with the BH10/BlaM-Vpr or the NLEY1-ES-IRES/VSV G/BlaM-Vpr viruses in the absence or presence of entry inhibitor T20 (1 µg/ml) for 2 hours before cells were harvested to detect BlaM activity. Number of cells with successful HIV-1 entry was scored in window Q2. (B, C) The IFITM SupT1 cell lines were treated with Dox (500 ng/ml) for 16 hours before infected with the BH10/BlaM-Vpr viruses. Results of three independent HIV- 1/BlaM-Vpr fusion experiments are summarized in the bar graph. (D) Illustration of IFITM3 mutations. The N-terminal sequences of IFITM3 were deleted to create mutants (1-6), (1-11) and (1-16). Their expression in the Dox-inducible SupT1 cell lines was examined in Western blots. (E) The (1-11) and (1-16) mutants are less effective in inhibiting HIV-1 entry than the wild type IFITM3. Results of three independent HIV-1 fusion experiments are shown in the bar graph. Figure 5. Association of IFITM2 and IFITM3 with endocytosis. The 293 cells were transiently transfected with vector DNA that expressed Flag-tagged IFITM1, IFITM2 or IFITM3. Forty hours after transfection, cells were fed with Alexa fluor 555-conjugated transferrin (red) at 37 o C for 10 minutes before fixed with 4% paraformadehyde (in 1x phosphate buffered saline). IFITM proteins (green) were detected by immunostaining with the anti-flag antibody. Figure 6. Effect of IFITM proteins on HIV-1 cdna synthesis and viral Gag expression. (A) The IFITM-inducible SupT1 cells were infected with the HIV-1(BH10) viruses. 26
27 Real-time PCR was performed using primers that amplify either the early or the late viral cdna products. Levels of integrated viral DNA were determined by Alu-PCR. Results shown are the average of three independent infections. (B) Western blots of HIV-1 Gag protein expression in IFITM-inducible SupT1 cells that were infected with the HIV- 1(BH10) viruses. Results shown represent three independent infection experiments. Figure 7. The anti-hiv-1 activity of IFITM1 does not depend on its C-terminal sequence. (A) Illustration of IFITM1 deletion mutants. Sequences from IFITM1 C-terminus were deleted. (B) Doxycycline-induced expression of IFITM1 mutants in SupT1 cells. (C) HIV-1 replication in SupT1 cells that express IFITM1 mutants. SupT1 cells were cultured in media with or without doxycycline (500 ng/ml). Results shown represent three independent spreading infection experiments. (D) Infection of SupT1 cells with the NLEY1-IRES virus. For details refer to the legend in Figure 3B. (E) Comparison of doxycycline-induced expression of IFITM1 and ( ) to IFN α2b-induced endogenous IFITM1. Cell lines were exposed to doxycycline (500 ng/ml), IFN α2b (500 U/ml) or both for 16 hours before levels of IFITM1 and ( ) were assessed in Western blots using anti-ifitm1 antibody. Tubulin was probed as the internal control. Figure 8. Roles of the N-terminal sequence and the loop sequence of IFITM1 in suppressing HIV-1 infection. (A) Illustration of IFITM1 mutants. All mutants lack the C- terminal sequence from amino acids 108 to 125. (B) Expression of the IFITM1 mutants in stably transduced SupT1 cell lines upon treatment with doxycycline (500 ng/ml). (C) Replication of HIV-1 in SupT1 cell lines that express IFITM1 mutants. Results shown 27
28 represent three independent spreading infection experiments. (D) Effects of IFITM1 mutants on HIV-1 infection in the one-cycle infection assays. For details refer to the legend in Figure 3B Figure 9. Knockdown of IFITM1, IFITM2 and IFITM3 increases HIV-1 infection of the TZM-bl cells. (A) Expression of endogenous IFITM1, IFITM2 and IFITM3 in different cell lines. Western blots were probed with anti-ifitm1, -IFITM2 or -IFITM3 antibodies. (B) Western blots to assess the knockdown of IFITM1, IFITM2 and IFITM3 in TZM-bl cells that were transfected with mixtures of sirna oligos. (C) Following transfection with sirna oligos, TZM-bl cells were exposed to HIV-1 infection. Levels of luciferase activity in the TZM-bl cell lysates were measured 40 hours after infection. The values (relative luciferase units, RLU) represent the susceptibility of TZM-bl cells to HIV-1 infection under different sirna treatment. Results shown are the average of three independent experiments. 28
29 614 Reference List Agy, M. B., R. L. Acker, C. H. Sherbert, and M. G. Katze Interferon treatment inhibits virus replication in HIV-1- and SIV-infected CD4+ T-cell lines by distinct mechanisms: evidence for decreased stability and aberrant processing of HIV-1 proteins. Virology 214: Andreu, P., S. Colnot, C. Godard, P. Laurent-Puig, D. Lamarque, A. Kahn, C. Perret, and B. Romagnolo Identification of the IFITM family as a new molecular marker in human colorectal tumors. Cancer Res 66: Barr, S. D., J. R. Smiley, and F. D. Bushman The interferon response inhibits HIV particle production by induction of TRIM22. PLoS Pathog 4:e Bednarik, D. P., J. D. Mosca, N. B. Raj, and P. M. Pitha Inhibition of human immunodeficiency virus (HIV) replication by HIV-trans-activated alpha 2-interferon. Proc Natl Acad Sci U S A 86: Bradbury, L. E., G. S. Kansas, S. Levy, R. L. Evans, and T. F. Tedder The CD19/CD21 signal transducing complex of human B lymphocytes includes the target of antiproliferative antibody-1 and Leu-13 molecules. J Immunol 149: Brass, A. L., I. C. Huang, Y. Benita, S. P. John, M. N. Krishnan, E. M. Feeley, B. J. Ryan, J. L. Weyer, L. van der Weyden, E. Fikrig, D. J. Adams, R. J. Xavier, M. Farzan, and S. J. Elledge The IFITM proteins mediate cellular resistance to influenza A H1N1 virus, West Nile virus, and dengue virus. Cell 139: Brem, R., K. Oraszlan-Szovik, S. Foser, B. Bohrmann, and U. Certa Inhibition of proliferation by 1-8U in interferon-alpha-responsive and nonresponsive cell lines. Cell Mol Life Sci 60: Brennan, K., and A. G. Bowie. Activation of host pattern recognition receptors by viruses. Curr Opin Microbiol. 9. Cavrois, M., J. Neidleman, M. Bigos, and W. C. Greene Fluorescence resonance energy transfer-based HIV-1 virion fusion assay. Methods Mol Biol 263: Chen, K., J. Huang, C. Zhang, S. Huang, G. Nunnari, F. X. Wang, X. Tong, L. Gao, K. Nikisher, and H. Zhang Alpha interferon potently enhances the anti-human immunodeficiency virus type 1 activity of APOBEC3G in resting primary CD4 T cells. J Virol 80: Dianzani, F., C. Castilletti, M. Gentile, H. R. Gelderblom, F. Frezza, and M. R. Capobianchi Effects of IFN alpha on late stages of HIV-1 replication cycle. Biochimie 80: Doehle, B. P., F. Hladik, J. P. McNevin, M. J. McElrath, and M. Gale, Jr Human immunodeficiency virus type 1 mediates global disruption of innate antiviral signaling and immune defenses within infected cells. J Virol 83:
30 Donaghy, H., A. Pozniak, B. Gazzard, N. Qazi, J. Gilmour, F. Gotch, and S. Patterson Loss of blood CD11c(+) myeloid and CD11c(-) plasmacytoid dendritic cells in patients with HIV-1 infection correlates with HIV-1 RNA virus load. Blood 98: Evans, S. S., R. P. Collea, J. A. Leasure, and D. B. Lee IFN-alpha induces homotypic adhesion and Leu-13 expression in human B lymphoid cells. J Immunol 150: Evans, S. S., D. B. Lee, T. Han, T. B. Tomasi, and R. L. Evans Monoclonal antibody to the interferon-inducible protein Leu-13 triggers aggregation and inhibits proliferation of leukemic B cells. Blood 76: Fernandez-Cruz, E., J. M. Lang, J. Frissen, V. Furner, M. Chateauvert, C. A. Boucher, P. Dowd, and J. Stevens Zidovudine plus interferon-alpha versus zidovudine alone in HIV-infected symptomatic or asymptomatic persons with CD4+ cell counts > 150 x 10(6)/L: results of the Zidon trial. Zidon Study Group. AIDS 9: Francis, M. L., and M. S. Meltzer Induction of IFN-alpha by HIV-1 in monocyte-enriched PBMC requires gp120-cd4 interaction but not virus replication. J Immunol 151: Gendelman, H. E., L. Baca, J. A. Turpin, D. C. Kalter, B. D. Hansen, J. M. Orenstein, R. M. Friedman, and M. S. Meltzer Restriction of HIV replication in infected T cells and monocytes by interferon-alpha. AIDS Res Hum Retroviruses 6: Gendelman, H. E., L. M. Baca, J. Turpin, D. C. Kalter, B. Hansen, J. M. Orenstein, C. W. Dieffenbach, R. M. Friedman, and M. S. Meltzer Regulation of HIV replication in infected monocytes by IFN-alpha. Mechanisms for viral restriction. J Immunol 145: Goujon, C., and M. H. Malim. Characterization of the interferon-{alpha} induced post-entry block to HIV-1 infection in primary human macrophages and T cells. J Virol. 21. Hansen, B. D., P. L. Nara, R. K. Maheshwari, G. S. Sidhu, J. G. Bernbaum, D. Hoekzema, M. S. Meltzer, and H. E. Gendelman Loss of infectivity by progeny virus from alpha interferon-treated human immunodeficiency virus type 1-infected T cells is associated with defective assembly of envelope gp120. J Virol 66: Hartshorn, K. L., D. Neumeyer, M. W. Vogt, R. T. Schooley, and M. S. Hirsch Activity of interferons alpha, beta, and gamma against human immunodeficiency virus replication in vitro. AIDS Res Hum Retroviruses 3: Heil, F., H. Hemmi, H. Hochrein, F. Ampenberger, C. Kirschning, S. Akira, G. Lipford, H. Wagner, and S. Bauer Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 303: Ho, D. D., K. L. Hartshorn, T. R. Rota, C. A. Andrews, J. C. Kaplan, R. T. Schooley, and M. S. Hirsch Recombinant human interferon alfa-a suppresses HTLV-III replication in vitro. Lancet 1:
31 Ishii, K. J., S. Koyama, A. Nakagawa, C. Coban, and S. Akira Host innate immune receptors and beyond: making sense of microbial infections. Cell Host Microbe 3: Jiang, D., H. Guo, C. Xu, J. Chang, B. Gu, L. Wang, T. M. Block, and J. T. Guo Identification of three interferon-inducible cellular enzymes that inhibit the replication of hepatitis C virus. J Virol 82: Jiang, D., J. M. Weidner, M. Qing, X. B. Pan, H. Guo, C. Xu, X. Zhang, A. Birk, J. Chang, P. Y. Shi, T. M. Block, and J. T. Guo. Identification of Five Interferon-induced Cellular Proteins That Inhibit West Nile Virus and Dengue Virus Infection. J Virol. 28. Kornbluth, R. S., P. S. Oh, J. R. Munis, P. H. Cleveland, and D. D. Richman Interferons and bacterial lipopolysaccharide protect macrophages from productive infection by human immunodeficiency virus in vitro. J Exp Med 169: Kornbluth, R. S., P. S. Oh, J. R. Munis, P. H. Cleveland, and D. D. Richman The role of interferons in the control of HIV replication in macrophages. Clin Immunol Immunopathol 54: Korth, M. J., M. D. Taylor, and M. G. Katze Interferon inhibits the replication of HIV-1, SIV, and SHIV chimeric viruses by distinct mechanisms. Virology 247: Lane, H. C., V. Davey, J. A. Kovacs, J. Feinberg, J. A. Metcalf, B. Herpin, R. Walker, L. Deyton, R. T. Davey, Jr., J. Falloon, and et al Interferonalpha in patients with asymptomatic human immunodeficiency virus (HIV) infection. A randomized, placebo-controlled trial. Ann Intern Med 112: Lange, U. C., D. J. Adams, C. Lee, S. Barton, R. Schneider, A. Bradley, and M. A. Surani Normal germ line establishment in mice carrying a deletion of the Ifitm/Fragilis gene family cluster. Mol Cell Biol 28: Levy, D. N., G. M. Aldrovandi, O. Kutsch, and G. M. Shaw Dynamics of HIV-1 recombination in its natural target cells. Proc Natl Acad Sci U S A 101: Li, X. Y., F. Guo, L. Zhang, L. Kleiman, and S. Cen APOBEC3G inhibits DNA strand transfer during HIV-1 reverse transcription. J Biol Chem 282: Lichtner, M., R. Rossi, F. Mengoni, S. Vignoli, B. Colacchia, A. P. Massetti, I. Kamga, A. Hosmalin, V. Vullo, and C. M. Mastroianni Circulating dendritic cells and interferon-alpha production in patients with tuberculosis: correlation with clinical outcome and treatment response. Clin Exp Immunol 143: Maitra, R. K., and R. H. Silverman Regulation of human immunodeficiency virus replication by 2',5'-oligoadenylate-dependent RNase L. J Virol 72: Malim, M. H., and M. Emerman HIV-1 accessory proteins--ensuring viral survival in a hostile environment. Cell Host Microbe 3: Martensen, P. M., and J. Justesen Small ISGs coming forward. J Interferon Cytokine Res 24:
32 Mercer, J., M. Schelhaas, and A. Helenius. Virus entry by endocytosis. Annu Rev Biochem 79: Michaelis, B., and J. A. Levy HIV replication can be blocked by recombinant human interferon beta. AIDS 3: Miyauchi, K., Y. Kim, O. Latinovic, V. Morozov, and G. B. Melikyan HIV enters cells via endocytosis and dynamin-dependent fusion with endosomes. Cell 137: Moffatt, P., M. H. Gaumond, P. Salois, K. Sellin, M. C. Bessette, E. Godin, P. T. de Oliveira, G. J. Atkins, A. Nanci, and G. Thomas Bril: a novel bone-specific modulator of mineralization. J Bone Miner Res 23: Nagai, K., A. H. Wong, S. Li, W. N. Tam, A. R. Cuddihy, N. Sonenberg, M. B. Mathews, J. Hiscott, M. A. Wainberg, and A. E. Koromilas Induction of CD4 expression and human immunodeficiency virus type 1 replication by mutants of the interferon-inducible protein kinase PKR. J Virol 71: Neil, S. J., T. Zang, and P. D. Bieniasz Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature 451: Okumura, A., T. Alce, B. Lubyova, H. Ezelle, K. Strebel, and P. M. Pitha HIV-1 accessory proteins VPR and Vif modulate antiviral response by targeting IRF-3 for degradation. Virology 373: Okumura, A., G. Lu, I. Pitha-Rowe, and P. M. Pitha Innate antiviral response targets HIV-1 release by the induction of ubiquitin-like protein ISG15. Proc Natl Acad Sci U S A 103: Pincetic, A., Z. Kuang, E. J. Seo, and J. Leis. The interferon-induced gene ISG15 blocks retrovirus release from cells late in the budding process. J Virol 84: Pitha, P. M Multiple effects of interferon on the replication of human immunodeficiency virus type 1. Antiviral Res 24: Poli, G., J. M. Orenstein, A. Kinter, T. M. Folks, and A. S. Fauci Interferon-alpha but not AZT suppresses HIV expression in chronically infected cell lines. Science 244: Randall, R. E., and S. Goodbourn Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures. J Gen Virol 89: Rivero, J., M. Limonta, A. Aguilera, M. Fraga, and P. Lopez Saura Use of recombinant interferon-alpha in human immunodeficiency virus (HIV)-infected individuals. Biotherapy 8: Rojek, J. M., and S. Kunz Cell entry by human pathogenic arenaviruses. Cell Microbiol 10: Sadler, A. J., and B. R. Williams Interferon-inducible antiviral effectors. Nat Rev Immunol 8: Sakuma, R., A. A. Mael, and Y. Ikeda Alpha interferon enhances TRIM5alpha-mediated antiviral activities in human and rhesus monkey cells. J Virol 81: Sauter, D., A. Specht, and F. Kirchhoff. Tetherin: holding on and letting go. Cell 141:
33 Seyfried, N. T., L. C. Huysentruyt, J. A. Atwood, 3rd, Q. Xia, T. N. Seyfried, and R. Orlando Up-regulation of NG2 proteoglycan and interferoninduced transmembrane proteins 1 and 3 in mouse astrocytoma: a membrane proteomics approach. Cancer Lett 263: Sheehy, A. M., N. C. Gaddis, J. D. Choi, and M. H. Malim Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 418: Shirazi, Y., and P. M. Pitha Alpha interferon inhibits early stages of the human immunodeficiency virus type 1 replication cycle. J Virol 66: Shirazi, Y., and P. M. Pitha Interferon alpha-mediated inhibition of human immunodeficiency virus type 1 provirus synthesis in T-cells. Virology 193: Skillman, D. R., J. L. Malone, C. F. Decker, K. F. Wagner, R. L. Mapou, M. J. Liao, D. Testa, and M. S. Meltzer Phase I trial of interferon alfa-n3 in early-stage human immunodeficiency virus type 1 disease: evidence for drug safety, tolerance, and antiviral activity. J Infect Dis 173: Smith, R. A., J. Young, J. J. Weis, and J. H. Weis Expression of the mouse fragilis gene products in immune cells and association with receptor signaling complexes. Genes Immun 7: Soumelis, V., I. Scott, F. Gheyas, D. Bouhour, G. Cozon, L. Cotte, L. Huang, J. A. Levy, and Y. J. Liu Depletion of circulating natural type 1 interferon-producing cells in HIV-infected AIDS patients. Blood 98: Stremlau, M., C. M. Owens, M. J. Perron, M. Kiessling, P. Autissier, and J. Sodroski The cytoplasmic body component TRIM5alpha restricts HIV- 1 infection in Old World monkeys. Nature 427: Van Damme, N., D. Goff, C. Katsura, R. L. Jorgenson, R. Mitchell, M. C. Johnson, E. B. Stephens, and J. Guatelli The interferon-induced protein BST-2 restricts HIV-1 release and is downregulated from the cell surface by the viral Vpu protein. Cell Host Microbe 3: Wei, X., J. M. Decker, H. Liu, Z. Zhang, R. B. Arani, J. M. Kilby, M. S. Saag, X. Wu, G. M. Shaw, and J. C. Kappes Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob Agents Chemother 46: Weidner, J. M., D. Jiang, X. B. Pan, J. Chang, T. M. Block, and J. T. Guo. Interferon-induced cell membrane proteins, IFITM3 and tetherin, inhibit vesicular stomatitis virus infection via distinct mechanisms. J Virol. 67. Yamada, O., N. Hattori, T. Kurimura, M. Kita, and T. Kishida Inhibition of growth of HIV by human natural interferon in vitro. AIDS Res Hum Retroviruses 4: Yamamoto, J. K., F. Barre-Sinoussi, V. Bolton, N. C. Pedersen, and M. B. Gardner Human alpha- and beta-interferon but not gamma- suppress the in vitro replication of LAV, HTLV-III, and ARV-2. J Interferon Res 6:
34 York, J., and J. H. Nunberg A novel zinc-binding domain is essential for formation of the functional Junin virus envelope glycoprotein complex. J Virol 81: York, J., and J. H. Nunberg Distinct requirements for signal peptidase processing and function in the stable signal peptide subunit of the Junin virus envelope glycoprotein. Virology 359: York, J., and J. H. Nunberg Role of the stable signal peptide of Junin arenavirus envelope glycoprotein in ph-dependent membrane fusion. J Virol 80: Yount, J. S., B. Moltedo, Y. Y. Yang, G. Charron, T. M. Moran, C. B. Lopez, and H. C. Hang. Palmitoylome profiling reveals S-palmitoylation-dependent antiviral activity of IFITM3. Nat Chem Biol 6: Yu, J. J., T. L. Wu, M. K. Liszewski, J. Dai, W. J. Swiggard, C. Baytop, I. Frank, B. L. Levine, W. Yang, T. Theodosopoulos, and U. O'Doherty A more precise HIV integration assay designed to detect small differences finds lower levels of integrated DNA in HAART treated patients. Virology 379: Downloaded from on October 4, 2018 by guest 34
35 Fold change B Fold change A Figure 1 IRF7 IRF9 DDX58 DDX60 MDA5 PARP9 PARP10 PARP12 PARP14 SP100 SP110 MYD88 NM1 ISG20 ADAR PLSCR1 RSAD2 HLA-C BST-2 GBP1 GBP2 RTP Fold change of virus production α Tubulin α IFITM1 shcon shifitm1 D OAS1 OAS2 OAS3 OASL STAT1 STAT2 TRIM21 TRIM22 ISG15 HERC5 HERC6 USP18 UBE2L6 MX1 MX2 IFIT1 IFIT2 IFIT3 IFIT5 IFITM1 IFI6 IFI27 IFI35 IFI44 PKR PSMB9 GZMB NT5C3 TAP1 RASGRP3 LAP3 LGP2 CDA DNAPTP6 EHD4 KIAA1618 PRIC285 RARRES3 XAF1 EPSTI1 SAMD9 TDRD7 MT2A LY6E DTX3L Fold change C

36 A IFITM3 IFITM2 IFITM1 IFITM3 IFITM2 IFITM1 MNHTVQTFFSPVNSGQPPNYEMLKEEHEVAVLGAPHNPAPPTSTVIHIRSETSVPDHVVW 60 MNHIVQT-FSPVNSGQPPNYEMLKEEQEVAMLGGPHNPAPPTSTVIHIRSETSVPDHVVW MHKEEHEVAVLGPPPSTILPRSTVINIHSETSVPDHVVW 39 SLFNTLFMNPCCLGFIAFAYSVKSRDRKMVGDVTGAQAYASTAKCLNIWALILGILMTIL 120 SLFNTLFMNTCCLGFIAFAYSVKSRDRKMVGDVTGAQAYASTAKCLNIWALILGIFMTIL 119 SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDVTGAQAYASTAKCLNIWALILGILMTIG 99 B C Viral RT activity (x105 CPM) IFITM3 LIVIPVL IFQAYG IFITM2 LVIIPVL VVQAQR IFITM1 FILLLVFGSVTVYHIMLQIIQEKRGY 125 control IFITM1 IFITM2 IFITM control -Dox +Dox -Dox +Dox IFITM Doxycycline α Flag α Tubulin -Dox +Dox Days after infection IFITM IFITM3 14 -Dox 12 +Dox Figure 2

 1.2 1.0 0.8 0.6 0.4 0.2 0 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0 19.")



37 A D un-infected cells un-infected cells - Dox + Dox control 17.90% 16.60% - Dox + Dox IFITM3 IFITM2 IFITM1 B Fold change (+Dox/-Dox) C Fold change (+Dox/-Dox) % 18.34% 16.73% 9.70% 21.09% 10.42% YFP change of infected cell numbers change of infectious HIV-1 particles control IFITM1 IFITM2 IFITM3 change of infected cell numbers change of infectious HIV-1 particles control IFITM1 IFITM2 IFITM3 Fold change (+Dox/-Dox) IFITM3 IFITM2 IFITM1 control % 37.21% 39.05% 25.57% 39.36% 24.74% 37.11% 16.29% control IFITM1 IFITM2 IFITM3 Figure 3
 3.1% 1.")
 IFITM3 (1-6) (1-11) (1-16) + - + - + - + - Dox 1.2 1.0 0.8 0.")
38 A C Cleaved CCF2 (Blue) B Cleaved CCF2 (Blue) control IFITM1 IFITM2 IFITM3 -CCF2 control +DOX 5.7% 5.4% IFITM1 +DOX 4.4% 4.4% IFITM2 +DOX 5.1% 1.8% IFITM3 +DOX +CCF2 +T20 9.6% 1.2% +T % 18.2% Uncleaved CCF2 (Green) 3.1% 1.6% BH10 BH10+ BlaM-Vpr NLEY1-ES-IRES +BlaM-Vpr BH10+BlaM-Vpr Fold change (+Dox/-Dox) D control IFITM1 IFITM2 IFITM MNHTVQFFSPVNSGQP E Fold change (+Dox/-Dox) IFITM3 (1-6) (1-11) (1-16) Dox α Flag α Tubulin control IFITM3 (1-6) (1-11) (1-16) Uncleaved CCF2 (Green) Figure 4




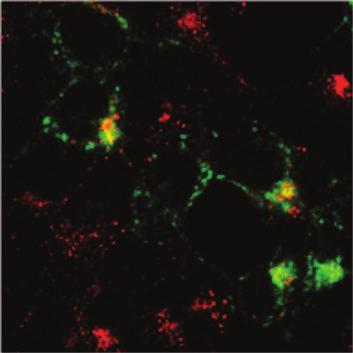


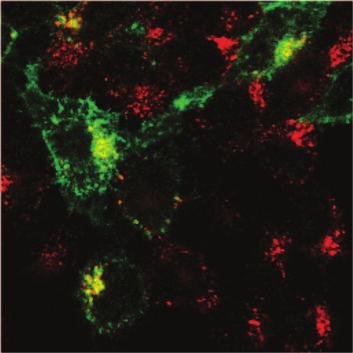
39 IFITM1 IFITM2 IFITM3 α Flag Transferrin Merge Figure 5



40 A Fold change (+Dox/-Dox) B control IFITM1 IFITM2 IFITM3 Dox Early cdna Late cdna Integrated cdna control IFITM1 IFITM2 IFITM3 Figure 6 pr55 Gag p40 p24 IFITM Tubulin
 2 4 6 8 10 12 14 0 2 4 6 8 1 3 5 7 0 -Dox +Dox -Dox +Dox (112-125) (108-125) 2 4 6 8 10 12 14 16 18 20 22 24 26 2 4 6 8 10 12 14")
41 A IFITM B control IFITM1 ( ) ( ) ( ) ( ) ( ) ( ) α Flag α tubulin C Viral RT activity (x105 CPM) Dox +Dox -Dox +Dox -Dox +Dox control IFITM1 ( ) Dox +Dox -Dox +Dox ( ) ( ) Days after infection D E Fold change (+Dox/-Dox) control control IFITM1 ( ) IFITM1 ( ) change of infected cell numbers change of infectious HIV-1 particles ( ) IFN α2b + + Dox (ng/ml) ( ) α IFITM1 α Tubulin α IFITM1 α Tubulin α IFITM1 α Tubulin Figure 7
 SVKS RDRK MVGD VTG QY ST KC LNI 2.5 2.0 1.5 1.0 0.")
42 A IFITM1 (1-21)( ) (1-29)( ) ( ) C 250 control 200 IFITM NO DOX DOX NO DOX DOX B D Fold change (+Dox/-Dox) SVKS RDRK MVGD VTG QY ST KC LNI ( ) control SLFNTLFLNWCCLGFIAFAYAAAARDRKMVGDVTGAQAYASTAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSAAAAMVGDVTGAQAYASTAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKAAAAVTGAQAYASTAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDAAAAQAYASTAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDVTGAAAAASTAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDVTGAQAYAAAAKCLNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDVTGAQAYASTAAALNIWALILGILMTIG SLFNTLFLNWCCLGFIAFAYSVKSRDRKMVGDVTGAQAYASTAKCAAAWALILGILMTIG (1-21)( ) IFITM1 (1-29)( ) ( ) ( ) SVKS (1-21) RDRK (1-29) MVGD SVKS VTG QY RDRK ST MVGD KC VTG LNI QY + ( ) α Flag α Tubulin change of infected cell number change of infectious HIV-1 particles ST KC LNI RT activity (10 4 CPM) ( ) NO DOX DOX (1-29)( ) NO DOX DOX RDRK NO DOX DOX VTG NO DOX DOX ST NO DOX DOX LNI NO DOX DOX (1-21)( ) NO DOX DOX SVKS NO DOX DOX MVGD NO DOX DOX QY NO DOX DOX KC NO DOX DOX Days after infection Figure 8
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
SUPPLEMENTARY INFORMATION
 sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
Intrinsic cellular defenses against virus infection
 Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Pre-made Reporter Lentivirus for JAK-STAT Signaling Pathway
 Pre-made Reporter for JAK-STAT Signaling Pathway Cat# Product Name Amounts LVP937-P or: LVP937-P-PBS ISRE-GFP (Puro) LVP938-P or: LVP938-P-PBS ISRE-RFP (Puro) LVP939-P or: LVP939-P-PBS ISRE-Luc (Puro)
Pre-made Reporter for JAK-STAT Signaling Pathway Cat# Product Name Amounts LVP937-P or: LVP937-P-PBS ISRE-GFP (Puro) LVP938-P or: LVP938-P-PBS ISRE-RFP (Puro) LVP939-P or: LVP939-P-PBS ISRE-Luc (Puro)
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Innate Immunity & Inflammation
 Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Inhibition of trna 3 Lys -Primed Reverse Transcription by Human APOBEC3G during Human Immunodeficiency Virus Type 1 Replication
 JOURNAL OF VIROLOGY, Dec. 2006, p. 11710 11722 Vol. 80, No. 23 0022-538X/06/$08.00 0 doi:10.1128/jvi.01038-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Inhibition of trna
JOURNAL OF VIROLOGY, Dec. 2006, p. 11710 11722 Vol. 80, No. 23 0022-538X/06/$08.00 0 doi:10.1128/jvi.01038-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Inhibition of trna
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Pre-made Lentiviral Particles for Fluorescent Proteins
 Pre-made Lentiviral Particles for Fluorescent Proteins Catalog# Product Name Amounts Fluorescent proteins expressed under sucmv promoter: LVP001 LVP001-PBS LVP002 LVP002-PBS LVP011 LVP011-PBS LVP012 LVP012-PBS
Pre-made Lentiviral Particles for Fluorescent Proteins Catalog# Product Name Amounts Fluorescent proteins expressed under sucmv promoter: LVP001 LVP001-PBS LVP002 LVP002-PBS LVP011 LVP011-PBS LVP012 LVP012-PBS
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Human Immunodeficiency Virus
 Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T
 Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
FIG S1 Examination of eif4b expression after virus infection. (A) A549 cells
 A549 cells") Supplementary Figure Legends FIG S1 Examination of expression after virus infection. () 549 cells were infected with herpes simplex virus (HSV) (MOI = 1), and harvested at the indicated times, followed
Supplementary Figure Legends FIG S1 Examination of expression after virus infection. () 549 cells were infected with herpes simplex virus (HSV) (MOI = 1), and harvested at the indicated times, followed
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
VIRAL TITER COUNTS. The best methods of measuring infectious lentiviral titer
 VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v)
") SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
Pre-made Reporter Lentivirus for MAPK/ERK Signal Pathway
 Pre-made Reporter for MAPK/ERK Signal Pathway Cat# Product Name Amounts LVP957-P or: LVP957-P-PBS SRE-GFP (Puro) LVP958-P or: LVP958-P-PBS SRE-RFP (Puro) LVP959-P or: LVP959-P-PBS SRE-Luc (Puro) LVP960-P
Pre-made Reporter for MAPK/ERK Signal Pathway Cat# Product Name Amounts LVP957-P or: LVP957-P-PBS SRE-GFP (Puro) LVP958-P or: LVP958-P-PBS SRE-RFP (Puro) LVP959-P or: LVP959-P-PBS SRE-Luc (Puro) LVP960-P
VIROLOGY. Engineering Viral Genomes: Retrovirus Vectors
 VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
Pre-made Reporter Lentivirus for NF-κB Signal Pathway
 Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Reviewers' comments: Reviewer #1 (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 (Remarks to the Author): In this manuscript, Song et al. identified FBXW7 as a new positive regulator for RIG-Itriggered type I IFN signaling pathway. The authors observed
Reviewers' comments: Reviewer #1 (Remarks to the Author): In this manuscript, Song et al. identified FBXW7 as a new positive regulator for RIG-Itriggered type I IFN signaling pathway. The authors observed
Supplemental Information
 Cell Host & Microbe, Volume 14 Supplemental Information HIV-1 Induces the Formation of Stable Microtubules to Enhance Early Infection Yosef Sabo, Derek Walsh, Denis S. Barry, Sedef Tinaztepe, Kenia de
Cell Host & Microbe, Volume 14 Supplemental Information HIV-1 Induces the Formation of Stable Microtubules to Enhance Early Infection Yosef Sabo, Derek Walsh, Denis S. Barry, Sedef Tinaztepe, Kenia de
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document
 Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or
 Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
Regulation of cell signaling cascades by influenza A virus
 The Second International Symposium on Optimization and Systems Biology (OSB 08) Lijiang, China, October 31 November 3, 2008 Copyright 2008 ORSC & APORC, pp. 389 394 Regulation of cell signaling cascades
The Second International Symposium on Optimization and Systems Biology (OSB 08) Lijiang, China, October 31 November 3, 2008 Copyright 2008 ORSC & APORC, pp. 389 394 Regulation of cell signaling cascades
QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24)
") Product Manual QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24) Catalog Number VPK-107 VPK-107-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction
Product Manual QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24) Catalog Number VPK-107 VPK-107-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction
Jumpstart your research with ViraPower Lentiviral Expression Systems
 ViraPower Lentiviral Expression Systems Jumpstart your research with ViraPower Lentiviral Expression Systems With ViraPower Lentiviral Systems you can: Efficiently transduce both dividing and non-dividing
ViraPower Lentiviral Expression Systems Jumpstart your research with ViraPower Lentiviral Expression Systems With ViraPower Lentiviral Systems you can: Efficiently transduce both dividing and non-dividing
Patricia Fitzgerald-Bocarsly
 FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
Supporting Information
 Supporting Information Palmisano et al. 10.1073/pnas.1202174109 Fig. S1. Expression of different transgenes, driven by either viral or human promoters, is up-regulated by amino acid starvation. (A) Quantification
Supporting Information Palmisano et al. 10.1073/pnas.1202174109 Fig. S1. Expression of different transgenes, driven by either viral or human promoters, is up-regulated by amino acid starvation. (A) Quantification
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05732 SUPPLEMENTARY INFORMATION Supplemental Data Supplement Figure Legends Figure S1. RIG-I 2CARD undergo robust ubiquitination a, (top) At 48 h posttransfection with a GST, GST-RIG-I-2CARD
doi: 10.1038/nature05732 SUPPLEMENTARY INFORMATION Supplemental Data Supplement Figure Legends Figure S1. RIG-I 2CARD undergo robust ubiquitination a, (top) At 48 h posttransfection with a GST, GST-RIG-I-2CARD
MedChem 401~ Retroviridae. Retroviridae
 MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Fayth K. Yoshimura, Ph.D. September 7, of 7 RETROVIRUSES. 2. HTLV-II causes hairy T-cell leukemia
 1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
RAISON D ETRE OF THE IMMUNE SYSTEM:
 RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Data Sheet. Notch Pathway Reporter Kit Catalog # 60509
 Data Sheet Notch Pathway Reporter Kit Catalog # 60509 6042 Cornerstone Court W, Ste B Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. NOTCH signaling
Data Sheet Notch Pathway Reporter Kit Catalog # 60509 6042 Cornerstone Court W, Ste B Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. NOTCH signaling
genome edited transient transfection, CMV promoter
 Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
SUPPLEMENTARY INFORMATION. Divergent TLR7/9 signaling and type I interferon production distinguish
 SUPPLEMENTARY INFOATION Divergent TLR7/9 signaling and type I interferon production distinguish pathogenic and non-pathogenic AIDS-virus infections Judith N. Mandl, Ashley P. Barry, Thomas H. Vanderford,
SUPPLEMENTARY INFOATION Divergent TLR7/9 signaling and type I interferon production distinguish pathogenic and non-pathogenic AIDS-virus infections Judith N. Mandl, Ashley P. Barry, Thomas H. Vanderford,
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
How HIV Causes Disease Prof. Bruce D. Walker
 How HIV Causes Disease Howard Hughes Medical Institute Massachusetts General Hospital Harvard Medical School 1 The global AIDS crisis 60 million infections 20 million deaths 2 3 The screen versions of
How HIV Causes Disease Howard Hughes Medical Institute Massachusetts General Hospital Harvard Medical School 1 The global AIDS crisis 60 million infections 20 million deaths 2 3 The screen versions of
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set
 p24 / Capsid Protein p24 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent Protein.
 prfp-vector RFP Exon1 Intron RFP Exon2 prfp-mir-124 mir-93/124 RFP Exon1 Intron RFP Exon2 Untransfected prfp-vector prfp-mir-93 prfp-mir-124 Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent
prfp-vector RFP Exon1 Intron RFP Exon2 prfp-mir-124 mir-93/124 RFP Exon1 Intron RFP Exon2 Untransfected prfp-vector prfp-mir-93 prfp-mir-124 Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent
Vif Proteins of Human and Simian Immunodeficiency Viruses Require Cellular CBFβ to Degrade APOBEC3 Restriction Factors
 JVI Accepts, published online ahead of print on 28 December 2011 J. Virol. doi:10.1128/jvi.06950-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13
JVI Accepts, published online ahead of print on 28 December 2011 J. Virol. doi:10.1128/jvi.06950-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13
Constitutive Reporter Lentiviral Vectors Expressing Fluorescent Proteins
 Constitutive Reporter Lentiviral Vectors Expressing Fluorescent Proteins www.vectalys.com/products/ Constitutive Reporter Lentiviral Vectors Catalog Number referring to this User Manual: 0008VCT; 0009VCT;
Constitutive Reporter Lentiviral Vectors Expressing Fluorescent Proteins www.vectalys.com/products/ Constitutive Reporter Lentiviral Vectors Catalog Number referring to this User Manual: 0008VCT; 0009VCT;
Doctoral Degree Program in Marine Biotechnology, College of Marine Sciences, Doctoral Degree Program in Marine Biotechnology, Academia Sinica, Taipei,
 Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
Reoviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP
, RBP, CBSP") Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Supplementary information
 Supplementary information Exosomes mediate the cell-to-cell transmission of interferon alpha-induced antiviral activity Jianhua Li, Kuancheng Liu, Yang Liu, Yan Xu, Fei Zhang, Huijuan Yang, Jiangxia Liu,
Supplementary information Exosomes mediate the cell-to-cell transmission of interferon alpha-induced antiviral activity Jianhua Li, Kuancheng Liu, Yang Liu, Yan Xu, Fei Zhang, Huijuan Yang, Jiangxia Liu,
Lecture 11. Immunology and disease: parasite antigenic diversity
 Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Overview: Chapter 19 Viruses: A Borrowed Life
 Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Viral Genetics. BIT 220 Chapter 16
 Viral Genetics BIT 220 Chapter 16 Details of the Virus Classified According to a. DNA or RNA b. Enveloped or Non-Enveloped c. Single-stranded or double-stranded Viruses contain only a few genes Reverse
Viral Genetics BIT 220 Chapter 16 Details of the Virus Classified According to a. DNA or RNA b. Enveloped or Non-Enveloped c. Single-stranded or double-stranded Viruses contain only a few genes Reverse
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
CRISPRaTest Functional dcas9-activator Assay Kit v1 Last update: 2018/07/04 Cellecta, Inc.
 CRISPRaTest Functional dcas9-activator Assay Kit v1 Last update: 2018/07/04 Cellecta, Inc. Copyright (c) 2018 Cellecta, Inc. All Rights Reserved. Table of Contents 1. CRISPRaTest Functional dcas9-activator
CRISPRaTest Functional dcas9-activator Assay Kit v1 Last update: 2018/07/04 Cellecta, Inc. Copyright (c) 2018 Cellecta, Inc. All Rights Reserved. Table of Contents 1. CRISPRaTest Functional dcas9-activator
Supplementary Figure 1
 Supplementary Figure 1 Constitutive EGFR signaling does not activate canonical EGFR signals (a) U251EGFRInd cells with or without tetracycline exposure (24h, 1µg/ml) were treated with EGF for 15 minutes
Supplementary Figure 1 Constitutive EGFR signaling does not activate canonical EGFR signals (a) U251EGFRInd cells with or without tetracycline exposure (24h, 1µg/ml) were treated with EGF for 15 minutes
Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).
 Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).") Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Phenomena first observed in petunia
 Vectors for RNAi Phenomena first observed in petunia Attempted to overexpress chalone synthase (anthrocyanin pigment gene) in petunia. (trying to darken flower color) Caused the loss of pigment. Bill Douherty
Vectors for RNAi Phenomena first observed in petunia Attempted to overexpress chalone synthase (anthrocyanin pigment gene) in petunia. (trying to darken flower color) Caused the loss of pigment. Bill Douherty
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Viruses. Picture from:
 Viruses Understand the structure of bacteriophages & human immunodeficiency virus (HIV) Appreciate that viruses replicate in host cells (thereby destroying them) Picture from: http://eands.caltech.edu/articles/lxvii1/viruses.html
Viruses Understand the structure of bacteriophages & human immunodeficiency virus (HIV) Appreciate that viruses replicate in host cells (thereby destroying them) Picture from: http://eands.caltech.edu/articles/lxvii1/viruses.html
HIV INFECTION: An Overview
 HIV INFECTION: An Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL MBBS II SEMINAR VJ
HIV INFECTION: An Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL MBBS II SEMINAR VJ
QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA)
") New and Improved Product Manual QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA) Catalog Numbers VPK-108-HIV-p24 96 tests VPK-108-HIV-p24-5 5 x 96 tests FOR RESEARCH USE ONLY Not for use in diagnostic
New and Improved Product Manual QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA) Catalog Numbers VPK-108-HIV-p24 96 tests VPK-108-HIV-p24-5 5 x 96 tests FOR RESEARCH USE ONLY Not for use in diagnostic
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis
 Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis") Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Antiviral Drugs Lecture 5
 Antiviral Drugs Lecture 5 Antimicrobial Chemotherapy (MLAB 366) 1 Dr. Mohamed A. El-Sakhawy 2 Introduction Viruses are microscopic organisms that can infect all living cells. They are parasitic and multiply
Antiviral Drugs Lecture 5 Antimicrobial Chemotherapy (MLAB 366) 1 Dr. Mohamed A. El-Sakhawy 2 Introduction Viruses are microscopic organisms that can infect all living cells. They are parasitic and multiply
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/366/ra25/dc1 Supplementary Materials for Viral entry route determines how human plasmacytoid dendritic cells produce type I interferons Daniela Bruni, Maxime
www.sciencesignaling.org/cgi/content/full/8/366/ra25/dc1 Supplementary Materials for Viral entry route determines how human plasmacytoid dendritic cells produce type I interferons Daniela Bruni, Maxime
SUPPLEMENTARY FIGURES AND TABLE
 SUPPLEMENTARY FIGURES AND TABLE Supplementary Figure S1: Characterization of IRE1α mutants. A. U87-LUC cells were transduced with the lentiviral vector containing the GFP sequence (U87-LUC Tet-ON GFP).
SUPPLEMENTARY FIGURES AND TABLE Supplementary Figure S1: Characterization of IRE1α mutants. A. U87-LUC cells were transduced with the lentiviral vector containing the GFP sequence (U87-LUC Tet-ON GFP).
Retroviruses. ---The name retrovirus comes from the enzyme, reverse transcriptase.
 Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set
 gp120 / Glycoprotein 120 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
DATA SHEET. Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter calf thymus DNA.
 Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
Ready-to-use Lentiviral Particles for intracelular labeling
 Ready-to-use Lentiviral Particles for intracelular labeling (LocLight TM Living cell imaging lentivirus for sub-cellular localization) LocLight TM cell organelle labeling lentivirus are provided as 200ul/per
Ready-to-use Lentiviral Particles for intracelular labeling (LocLight TM Living cell imaging lentivirus for sub-cellular localization) LocLight TM cell organelle labeling lentivirus are provided as 200ul/per