Protease-mediated entry via endosome of human coronavirus 229E ACCEPTED. Miyuki Kawase, Kazuya Shirato, Shutoku Matsuyama and Fumihiro Taguchi*
|
|
|
- Howard Davis
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 29 October 2008 J. Virol. doi: /jvi Copyright 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Protease-mediated entry via endosome of human coronavirus 229E By Miyuki Kawase, Kazuya Shirato, Shutoku Matsuyama and Fumihiro Taguchi* Laboratory of Respiratory Viral Infections, Department of Virology III, National Institute of Infectious Diseases, Musashi-Murayama, Tokyo, Japan Running head: Cell entry mechanism of human coronavirus 229E Corresponding author: Fumihiro Taguchi Laboratory of Viral Respiratory Infections Department of Virology III National Institute of Infectious Diseases, Musashi-Murayama Tokyo, Japan e. mail: phone: (ext. 3533) 5
2 Abstract Human coronavirus 229E, classified as a group I coronavirus, utilizes human aminopeptidase N (APN) as a receptor; however, its entry mechanism has not yet been fully elucidated. We found that HeLa cells infected with 229E via APN formed syncitia when treated with trypsin or other proteases, but not in a low ph environment, a finding consistent with syncytia formation by SARS coronavirus (SARS-CoV). In addition, trypsin induced cleavage of the 229E S protein. By using infectious viruses and pseudotype viruses bearing the 229E S protein, we found that its infection was profoundly blocked by lysosomotropic agents, as well as by protease inhibitors that also prevented infection with SARS-CoV, but not that caused by murine coronavirus MHV-JHMV, which enters cells directly from the cell surface. We found that Cathepsin L (CPL) inhibitors blocked 229E infection the most remarkably among a variety of protease inhibitors tested. Furthermore, 229E infection was inhibited in CPL knockdown cells by sirna, when compared with a normal counterpart producing CPL. However, its inhibition was not so remarkable as that found with SARS-CoV infection, which seems to indicate that while CPL is involved in the fusogenic activation of 229E S protein in endosomal infection, not yet identified proteases could also play a part in that activity. We also found 229E virion S protein to be cleaved by CPL. Furthermore, as with SARS-CoV, 229E entered cells directly from the cell surface, when cell-attached viruses were treated with trypsin. These findings suggest that 229E takes an endosomal pathway for cell entry, and proteases like CPL are involved in this mode of entry. 6
3 Introduction Human coronavirus (HCoV) 229E, a causative agent of the human common cold (44), is classified as a group I coronavirus and is an enveloped virus having RNA consisting of a single, positive-stranded genome of about 30 kilobases (21). Human aminopeptidase N (APN), a surface metalloprotease found on the apical membranes of a variety of cells, serves as a receptor for HCoV 229E (17, 49) Spike (S) protein of HCoV 229E, categorized as a class I fusion protein, is responsible for the binding to APN and entry into cells (3, 6). Although the S protein of group II coronavirus mouse hepatitis virus (MHV) is cleaved by a host cell derived protease into two subunits, N terminal S1 and C terminal S2 during biogenesis in the exocytic pathway (35), 229E S is not cleaved in cells and S protein on the virion is an uncleaved form (6). However, the 229E S protein consists of two regions corresponding to S1 and S2 of the cleaved subunits of MHV. The former is responsible for receptor binding and the latter for entry. The receptor-binding site of 229E S encompasses the internal region of S1, consisting of amino acids 417 to 547 (3, 41), and differs from the location of the MHV receptor-binding site, which is in the N terminal 330 amino acids of the S1 subunit (19, 36). S1 is variable among different coronaviruses; even in a given coronavirus group, S1 is highly divergent (6). However, the S2 or S2 corresponding regions of coronaviruses share common structural and biological features (5, 6, 9). These observations suggest a common entry mechanism utilized by various coronaviruses. Enveloped viruses enter cells through the fusion of their envelope with the 7
4 plasma membrane or endosomal membrane. Human immunodeficiency virus (HIV) enters cells via a plasma membrane (7, 12, 45). The binding to the receptor/co-receptor induces conformational changes together with the fusion activation of gp160 of HIV, which, in turn, facilitates fusion of the viral envelope and plasma membrane (7, 12). This mechanism is utilized by a number of retroviruses, paramyxoviruses and coronaviruses (12, 45). prototypically utilizes an endosomal pathway for entry. In contrast, the influenza virus Its hemaglutinin (HA) protein is not activated by binding to its receptor, but, rather, is activated for fusion in the acidic environment of the endosome (low ph-dependent entry). A similar mode of entry is used by vesicular stomatitits virus (VSV) and others as well (45). A third mode of entry was reported as a mechanism for Ebola virus that enters cells via an endosomal compartment, and acidic condition in the endosomes is critical for entry (8, 39, 47). However, it is not the acidic conditions, but, rather, proteases that trigger conformational changes and fusion activation of GP protein (protease-dependent entry) (8). SARS-CoV has been also reported to enter cells in a protease-dependent fashion as well (33, 34). Cells infected with a number of non-fusogenic coronaviruses, such as SARS-CoV or MHV-2, form syncytia after trypsin treatment (26, 30, 34, 50). Syncytia formation was also observed in the presence of trypsin in cells infected with porcine epidemic diarrhea virus or bovine coronavirus (20, 40). The 229E-infected cells also formed syncytia in the presence of some trypin-like proteases and the infection was inhibited by cystein and the trypsin-like protease inhibitor leupeptin (2). These 8
5 features of 229E are shared by SARS-CoV (26, 34), which suggests the possibility that 229E enters into cells in a fashion similar to that of SARS-CoV. In the present study, we have tested this possibility by using HeLa cells permissive to 229E infection. Our results suggest that 229E enters cells via endosomes in which proteases active at a low ph environment, most likely CPL and other proteases, are involved in the fusogenic activation of 229E S protein, leading us to conclude that the 229E cell entry mechanism is similar to that of SARS-CoV. Cells: Materials and methods HeLa cells maintained in our institute, expressing human APN and permissive to 229E infection, were transformed to express the receptor for SARS-CoV, human angiotensin-converting enzyme 2 (ACE2) (23), as well as the MHV receptor, carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) (10, 11, 46). HeLa cells expressing ACE2 were established by transfecting the expression plasmid for ACE2 and through selection in a medium containing G418, as described previously (51). The cell line that showed high sensitivity to SARS-CoV (designated HeLa-A cells) was further transfected with an expression plasmid containing the CEACAM1 gene (24, 31) and selected in a medium containing Blastcydine. Cell line sensitive 229E, SARS-CoV and MHV, expressing ACE2 and CEACAM1 (HeLa-AC), was selected. Those cells were grown and maintained in Dulbecco s modified minimal essential medium (DMEM, Nissui, Tokyo, Japan) containing 5% and 1% fetal bovine serum 9
6 (FBS, Sigma, St Louis, MO) as growth medium (GM) and maintenance medium (MM), respectively. All of the HeLa, HeLa-A and HeLa-AC cell lines we isolated were proved to be sensitive to vesicular stomatitis virus (VSV) infection as well. Virus and virus infection: HCoV 229E was kindly provided by Dr. R. Nomura (28), and initially grown in L132 cells, a subline of HeLa cells. The viruses were then grown in HeLa cells. For the propagation of 229E as a seed virus, viruses were inoculated onto HeLa cells with a multiplicity of infection (MOI) of 0.5~1 and adsorbed at 34 C for 1 h. Then, cells were cultured with MM at 34 o C for 2 days. Culture supernatants, as well as infected cells, were used after ultra-sonication as described previously (27). Viruses divided into a small volume were stored at 80 o C until use. MHV-2 and MHV-JHMV sp-4 also used in this experiment were prepared as described elsewhere (38, 48). Pseudotype VSV that expresses green fluorescent protein (GFP) and harbors SARS-CoV S protein or VSV G protein was prepared as described previously (15). The production of a VSV pseudotype bearing 229E S protein is described below. Plaque assay Plaque assay was done using HeLa cells prepared in 24-well plates with a modified plaque assay for MHV as described previously (37). Confluent HeLa cells in 24-well plates (Falcon, Lincoln Park, NJ) were inoculated with 50 µl of 10-fold serially diluted virus samples and incubated at 34 o C for min. Then, cells were overlaid 10
7 with 0.5 ml of MM containing 1% carboxylmethyl cellulose (CMC, Wako, Tokyo, Japan) at 34 o C for 2 days. Cells were then washed once with phosphate buffered saline, ph 7.2 (PBS) and treated with 200 µg/ml of trypsin at room temperature (RT) for 5 min. After removal of trypsin, cells were further incubated at 34 o C for 2-3 h in MM, until syncytia were visible. Alternatively, infected cells were cultured with DMEM+10 % tryptose phosphate broth (TPB, Difco, Detroit, MI) + 1 % CMC containing 5 µg/ml of trypsin for 24 h. Cells were then fixed with formalin and stained with crystal violet. Plaques of syncytia were counted under light microscopy. Blockade of 229E infection by anti-apn antibodies. HeLa cells cultured in GM in a 96-well plate (Smilon, Tokyo, Japan) were treated with serially diluted anti-apn antibody WM15 (Biolegend, San Diego, CA) at 4 o C for 45 min. Then, cells were infected with ca. 150 PFU of 229E, incubated at 4 o C for 45 min and further incubated at 34 o C in GM for 20 to 24 h in the presence of anti-apn antibody. After fixing with cold ethanol/acetone (1:1) for 5 min, infected cells were examined for 229E antigen with anti-229e antibodies raised using rabbit and FITC-labeled, anti-rabbit serum. Antigen-positive cells were counted under fluorescence microscopy. The inhibition of infection by the antibody was calculated in comparison with the antigen-positive cell number obtained without treatment of APN antiserum. Generation of VSV pseudotyped with 229E S protein. 11
8 To generate the VSV pseudotyped with 229E S protein, we first of all isolated the cdna to 229E S protein, with 19 amino-acid truncation from the C-terminal. The S protein with 19 amino-acid deletion was reported to be efficiently incorporated into VSV pseudotype virus (15). The gene encoding this protein was amplified by a reverse-transcribed polymerase chain reaction (RT-PCR) with a specific primer set (sense: anti-sense 5 -CCGCTCGAGgccgccaccATGTTTGTTTTGCTTGTTGCATATGCC-3, primer: 5 -CGGGGTACCtcaTCTAATAGAAGATGCAAAACAACTAAAG-3 ), and then cloned into the XhoI-KpnI site of the mammalian expression vector, ptarget (Promega, Madison, WI). The sequence of the constructed plasmid was confirmed using the BigDye Terminator Cycle Sequencing Kit ver 3.1 and the ABI PRISM 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA), and the plasmid was designated as ptarget 229ES-del19. The 293T/17 cells obtained from the American Tissue Culture Collection (ATCC) (catalog no. CRL-11268) were transfected with the above expression plasmid using TransIT-293 transfection reagent (Mirus Bio, Madison, WI) and incubated at 34 o C for 40 h. Those cells were then infected with VSV G*-G, in which the VSV G gene is replaced by a GFP gene and harbors VSV G protein, kindly provided by Dr. M. A. Whitt, GTx, Inc, TN, USA, and incubated at 34 o C for 1 h. After 4 washes with PBS, those cells were further incubated at 34 o C for 24 h. The culture fluid was collected, centrifuged at 1,000 rpm for 5 min at 4ºC, divided into a small volume and stored at -80ºC until use. As controls, VSV pseudotype viruses bearing SARS-CoV S and VSV G proteins were generated in the same way as reported 12
9 previously (15). The infectivity of those pseudotype viruses was determined by the number of GFP-positive cells and was shown as infectious units (IU). Proteases: The proteases used in this study are trypsin (Sigma, T-8802); thermolysin (Sigma, P1512); chymotrypsin (Sigma, C-3142); dispase (Roche, , Branchburg, NJ); papain (Worthington Biochemicals, 53J6521, Freehold, NJ); proteinase K (Wako, Tokyo, Japan); collagenase (Sigma, C-5183) and elastase (Sigma, E-0258). These proteases were dissolved in PBS and used at the indicated concentration in PBS. Cells infected with 229E cultured in MM were washed once with PBS and treated with various concentrations of proteases at RT for 5 min. Then, proteases were removed and cells were further incubated in MM for 2 to 3 h until the syncytia became detectable under light microscopy. Treatment of virions with proteases: To see whether trypsin induced cleavage of 229E S protein, we used concentrated virions. 229E in culture fluid consisting of DMEM+ 10 % TPB was clarified by centrifugation at 5,000 rpm for 20 min, and the supernatants were concentrated by ca. 10-fold by ultrafiltration (Amicon Ultra, Ultracel-100K, Milipore, MA). These virions were treated with trypsin at different concentrations at RT for 5 min and then analyzed by Western blot after SDS-polyacrylamide gel electrophoresis (PAGE), as described previously (24). HeLa cells infected with 229E at an MOI of 1 were treated with 13
10 trypsin at 1 day postinfection at RT for 5 min and the proteins were analyzed by Western blot after SDS-PAGE. Virions were also treated with 36 µg/ml of CPL (Calbiochem, #219402) or 36 µg/ml of CPB (Calbiochem, #219362) in a buffer containing 130 mm NaCl, 40 mm HEPES, ph 5.0 and 40 mm MES, ph5.0 at 37 o C for 30 min. Then, the resultant virions were analyzed by Western blot with anti-s2 antibody. Treatment of cells with lysosomotropic agents and protease inhibitors: To see the effect of bafilomycin (Baf in short, Sigma) or ammonium chloride (NH 4 Cl) (Wako) that blocks the acidification of the endosome, we used infectious viruses or pseudotype VSV bearing 229E S, SARS-CoV S or VSV G protein. HeLa-AC cells in 24-well plates were treated with Baf (1 µm) or NH 4 Cl (50 mm) in MM for 1 h at 37 o C and were infected with 5x10 5 pfu of 229E, MHV-JHM or MHV-2. After incubation for 1 h at 34 o C (229E) or 37 o C (MHVs) in the presence of those agents, cells were washed once with PBS and fed in MM in the presence of agents for 3 h. Then, the medium was changed to MM without agents. At 24 and 10 h after infection with 229E or MHVs, virus titers in cells were examined by a plaque assay. Unless otherwise stated, the adsorption and incubation of infected cells were carried out at 34 o C and 37 o C for 229E and MHVs, respectively. In some experiments, HeLa cells in 24-well plates were treated with 500 nm Baf in MM from 1 h before to 4 h after 229E infection or from 4 to 24 h after infection of 5x10 5 PFU of 229E and virus titers were examined at 24 h. In the analysis using VSV pseudotype, HeLa-A cells prepared in 96-well plates 14
11 were treated with different concentrations of Baf for 60 min. After removing Baf, we infected the cells with pseudotype viruses of approximately 500 IU in 50 µl of medium containing Baf and incubated at 37 o C for 1 h for adsorption. After washing cells with PBS, we incubated them for a further 24 h in the presence of Baf. GFP-positive cells were photographed under Keyence fluorescence microscopy (Keyence Corporation, Osaka, Japan) and counted by using Image Measurement & Analyzing software VH-H1A5 ver 2.6 (Keyence). The inhibitory effect of the Baf was calculated by dividing GFP-positive cell numbers in treated cells by the number obtained from untreated cells. NH 4 Cl was also used in a manner similar to that employed with Baf. Cells were also treated with a variety of protease inhibitors to determine the involvement of protease in the entry process of 229E by using VSV pseudotypes bearing either 229E S protein, SARS-CoV S protein, or VSV G protein. The protease inhibitors used were as follows: cystein protease and trypsin-like protease inhibitor Leupeptin (Roche); CPL inhibitor EST (Calbiochem); MDL28107 (Sigma); CPB inhibitor CA-074 (Sigma); cathepsin K (CPK) inhibitor II (Z-L-NHNHCONHNH-LF-Boc, Calbiochem); cathepsin S (CPS) inhibitor (Z-FL-COCHO, Calbiochem) and CPS L inhibitor II (Z-FY-CHO, Calbiochem). Cells were treated with these protease inhibitors in a manner similar to the treatment with Baf and their effect on the infection of pseudotypes with 229E S, SARS-CoV S and VSV G proteins was determined by the quantification of GFP-positive cells as described above. Knock down of CPL by sirna. 15
12 Small interfering RNA (sirna) oligonucleotides targeting CPL (L ) and control non-targeting sirna (D ) were purchased from Darmacon (Chicago, IL). HeLa-A cells were treated with those sirnas according to the manufacturer s recommendation. Briefly, 50% subconfluent cells prepared one day before use in 96-well plates were incubated with GM containing 100 nm of sirna for 3 to 4 days. Then, those cells were infected with pseudotype VSV-bearing 229E S, VSV-G or SARS-CoV S protein. The GFP-positive cells were counted 24 h later under Keyence fluorescence microscopy and the percentage of inhibition was obtained by comparing the GFP-positive cells treated with control non-targeting sirna. Potentiation of 229E infection from cell surface: 229E was facilitated to infect from the cell surface, as described previously for the cell-surface infection of SARS-CoV (26, 42). HeLa-A cells were treated with 500 nm Baf for 60 min at 37 o C to block the endosomal pathway and then cells were kept on ice. Those cells were infected with pseudotype VSV bearing the 229E S protein on ice for 30 min and then treated with 100 µg/ml trypsin at RT for 5 min. Cells were further incubated at 37 o C for 24 h in the presence of 500 nm Baf. Virus infection was estimated by the expression of GFP as described previously (15). VSV pseudotypes bearing SARS-CoV S protein or VSV G protein were also used as controls. Western blot Samples for Western blot analysis were lysed in a lysis buffer and subjected to 16
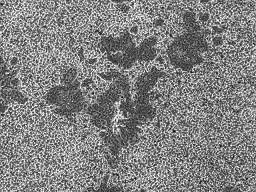
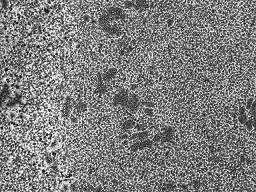
13 SDS-PAGE, as described previously (25). The proteins in the gel were transferred onto a PDF membrane (25), and S protein was detected with anti-s2 antibodies raised with a synthetic peptide derived from the C terminal region of the S protein in rabbits, kindly provided by Dr. R. Nomura. Western blot analysis of CPL was done with anti-cat-l (Athens Res. & Tech., Athens). Then, the bands were visualized using Supersignal West Dura (Pierce, Rockford, IL, USA) with LAS-1000PLUS (Fujifilm, Tokyo, Japan). Results Syncytium formation of 229E-infected HeLa cells after treatment with proteases. Permissive cell lines to human coronavirus 229E are restricted. cells have been generally used for infectious assay of 229E (28, 41). L132 or MCR-5 L132 is a sub-clone derived from HeLa cells, although these two cell lines are morphologically distinct. We have examined whether the HeLa cells maintained in our institute were permissive to 229E infection or not. When HeLa cells were infected with 229E and cultured for 2 days, there appeared to be no cytopathic changes (Fig. 1A). However, 229E-inoculated cells were found to be antigen-positive, when examined by indirect immunofluorescence using anti-229e antibodies (Fig. 1A), which indicated that the HeLa cells we used are permissive to 229E infection. Since cells infected with other coronaviruses without inducing syncytia formation, such as SARS-CoV and MHV-2, developed syncytia following trypsin treatment (26, 34, 50), we examined the effect of 17
14 trypsin on 229E-infected cells. As shown in Fig. 1A and B, cells infected with 229E formed syncytia with 229E antigens after treatment with trypsin. These results showed that our HeLa cells were susceptible for 229E infection, and infected cells developed syncytia when treated with trypsin. However, cells infected with 229E treated with a buffer of ph5.0 did not exhibit syncytia formation (Fig. 1B). These features of 229E are similar to those of SARS-CoV and MHV-2. We have also examined the effects of a variety of proteases on fusion formation of 229E-infected HeLa cells. Infected cells were treated with proteases at the highest concentration that does not give overt effects in non-infected HeLa cells. As shown in Fig. 1C, a variety of proteases, such as proteinase K and thermolysin, induced large syncytia as efficiently as trypsin did, while elastase, dispase, chymotrypsin and papain induced syncytia slightly or apparently smaller than the syncytia induced by trypsin treatment. Collagenase treatment failed to induce syncytia. The findings of syncytia formation by those proteases and failure of collagenase to induce syncytia on 229E-infected cells were quite similar to those relative to syncytia formation by SARS-CoV, as described previously (26). Cleavage of the S protein by trypsin. We have examined, by using 229E virions, whether syncytia formation is attributed to S protein cleavage, as reported for SARS-CoV and MHV (26, 30). 229E virions concentrated by ultrafiltration, as described in Materials and Methods, were treated with varied concentrations of trypsin at RT for 5 min, and the S protein was 18
15 analyzed by Western blot using anti-229e S antibodies. 229E-infected or mock-infected cells were also treated with trypsin and analyzed by Western blot. As shown in Fig. 2, the virions, without trypsin treatment, contained a protein of ca. 200 kda, a finding which suggested that this protein corresponds to the uncleaved 229E S. Likewise, in 229E-infected cells untreated with trypsin, only a 200-kDa band was detected. After treatment with trypsin, two other bands, one of 80 to 85 kda and the other of ca. 150 kda, became visible in the virions as well as in the infected cells. The smaller band was always found after treatment with trypsin, although a band of ca. 150 kda was not seen in some cases, especially when virions were treated with low concentrations of trypsin (data not shown). Since the antibody used for this analysis is against the C terminal region of the S, the 80- to 85-kDa protein seems to correspond to the S2 subunit of MHV, while a 150-kDa protein would result from the cleavage at the site further upstream from the cleavage site producing the 80- to 85-kDa protein. A 150- kda protein would not be involved in the fusion formation, since this band was not always found in cell lysates after treatment by trypsin, even when cell- to-cell fusion was induced. There may be a site in the S protein highly sensitive to trypsin treatment, which would result in the production of a150-kda fragment. APN expressed on HeLa cells and utilized as a receptor for 229E. Since it was revealed that trypsin treatment induced syncytia formation in 229Einfected cells, we have established a plaque assay as described in Materials and Methods. As shown in Fig. 3A, plaques consisting of fused cells were produced 1 to 2 19
16 days after infection in the presence of trypsin. The assay showed the one-hit kinetics, as shown in Fig. 3B, indicating that a plaque is produced by an infectious virus, and this plaque assay is highly reliable for infectivity testing. Since 229E is known to utilize human APN as a receptor for infection (17, 49), we have examined whether the infection of HeLa cells is mediated by the APN. The infection of 229E was blocked by anti-apn antibody in a concentration-dependent manner (Fig. 3C) and not blocked by control antibody (data not shown), indicating that our HeLa cells are infected via APN, and no alternative receptor is used for the infection. Effects of lysosomotropic agents on infection of 229E. As 229E produced syncytia formation in the presence of trypsin that induces the cleavage of its S protein, being similar to SARS-CoV and MHV-2 that take an endosomal pathway for infection, we deduced that 229E enters into cells in a fashion similar to those coronaviruses. To test this possibility, we examined the effects on 229E infection of the lysosomotropic agents Baf or NH 4 Cl, known to prevent the acidification of the endosome and block infection via an endosomal pathway. First of all, we examined whether Baf treatment in an early phase of infection prevents infection or not. HeLa cells were continuously treated with Baf from 1 h before 229E infection to 4 h after infection, and the cells were cultured in the absence of Baf thereafter. Alternatively, cells were treated with Baf from 4 h to 24 h of infection. Virus titers in the culture at 24 h infection were monitored. As shown in Fig. 4A, 229E infection was heavily (by ca fold) inhibited when cells were treated at -1 to 4 h of infection, 20
17 while it was not so greatly influenced when cells were treated from 4 h after infection, which suggested to us that Baf treatment impaired an early phase of 229E infection, but not the phase later than 4 h. This finding could indicate that 229E takes an endosomal pathway and the acidic conditions of endosomes are critical for its cell entry. We also examined the effects of Baf and NH 4 Cl on the infection by 229E of HeLa-AC cells under the control of MHV-2 that requires acidic endosomal conditions for cell entry, and also of JHMV that enters cells directly from the cell surface. As shown in Fig. 4B, the infection of MHV-2 was extensively suppressed by treatment with Baf and NH 4 Cl, yet JHMV infection was hardly influenced, as expected due to their entry mechanisms shown by previous reports (16). 229E infection in those cells was suppressed more efficiently than in MHV-2 by both of those agents, a finding leading us to suggest that 229E takes an endosomal pathway and the acidic conditions of the endosomes are necessary for its infection. Furthermore, we have confirmed these observations by using pseudotype VSV harboring the 229E S protein. Pseudotype VSV carrying SARS-CoV S protein or VSV glycoprotein were used as control viruses that take an endosomal pathway. HeLa-A cells treated with either Baf or NH 4 Cl were infected with those pseudotypes, and infection was monitored by GFP expression at 24 h of infection. As shown in Fig. 4C, the infection of all of three pseudotypes was suppressed in an agent-concentration-dependent fashion. This result strengthened the above observation that 229E enters into cells via the endosomes of the acidic environment, in a manner similar to the infection of SARS-CoV and VSV. 21
18 Effects of protease inhibitors on infection of 229E. The above data led us to the possibility that 229E takes an endosomal pathway and the acidic condition of the endosomes is necessary for its infection, although we could not clarify whether this virus requires proteases active in the endosome, as revealed with SARS-CoV, or a low ph environment is sufficient for its infection as shown by VSV; infection of the former but not the latter is blocked by protease inhibitors. To address this issue, cells treated with a variety of protease inhibitors were infected with pseudotypes bearing 229E S, SARS-S or VSV-G, and their infection was estimated using GFP expression. As shown in Fig. 5, infection of the 229E pseudotype and that of SARS-CoV was prevented in a concentration-dependent manner by the treatment with leupeptin (cystein protease and trypsin-like protease inhibitor), MDL and EST (both CPL inhibitors), as well as CPL inhbitor II, which implied that CPL is utilized by 229E for its infection. Infection of the pseudotype with 229E S protein was partially blocked by treatment with CPK inhibitor, but the inhibition was less prominent than that of the pseudotype with SARS-CoV S protein. Those pseudotypes were not drastically affected with the treatment of CA-074 (CPB inhibitor). In contrast, the infection of the VSV-G pseudotype was affected by none of those protease inhibitors. CPS inhibitor displayed an inhibitory effect in all of those pseudotype infections, even for one with VSV G protein that requires no proteases thus far reported, leading us to the suggestion that CPS is not involved in the infection of 229E and the inhibitory effects may be due to the side effects of CPS inhibitor. These results collectively suggested that 229E enters the cell in a protease-dependent fashion similar to that of SARS-CoV, and CPL is 22
19 the primary candidate for activation of 229E entry into cells Effect of CPL knock-down by sirna. To determine the role of CPL on 229E infection more directly, we knocked-down the CPL expression by sirna as described in Materials and Methods. Cells treated with sirna for 3-4 days were examined for CPL expression. As shown in Fig. 6A, CPL expression in cells treated with CPL-siRNA, but not with control RNA, was reduced as examined by Western blot. These cells were infected with a pseudotype bearing 229E S, SARS-CoV S or VSV-G and their infection was monitored by GFP expression. As shown in Fig. 6B, infection of SARS-CoV pseudotype was blocked by ca. 80 %, while VSV-G pseudotype was not at all blocked, a finding similar to the observations obtained from the protease inhibitor experiments. The infection of 229E pseudotype was also blocked by the treatment with sirna, yet it was not so drastically blocked as the infection with SARS-CoV pseudotype. We consistently observed a reduction in infection of about 20-45%, which suggested to us that although CPL is involved in the infection initiated by 229E, some other proteases could also participate in the infection process. Cleavage of virion S protein by CPL. Since protease inhibitors and sirna analysis suggested that CPL is the most probable protease to induce the cleavage and fusion activation of 229E S protein, we have treated 229E virion S protein with CPL and analyzed the resultant S protein by 23
20 Western blot. As shown in Fig. 6C, 229E S protein was cleaved, resulting in the appearance of a fragment of ca. 100 kda, while the cleavage was not found following the treatment with CPB. This suggested that 229E S protein is cleaved and activated for fusion by CPL. We have also treated virions with trypsin and compared the cleavage products of CPL and trypsin. As shown in Fig. 6C, the cleavage product of CPL, ca. 100 kda, was slightly larger than that of trypsin of ca. 80 kda. We also treated 229E-infected cells with CPL to see whether CPL induced cell- to-cell fusion, as revealed by Bosch et al for SARS-CoV-infected cells (4), but it failed to induce the cell-to-cell fusion (data not shown), although treatment with trypsin did induce the fusion, as seen in Fig. 1. Potentiation of 229E infection from the cell surface. The results described above indicate that 229E takes an endosomal pathway for cell entry, and the CPL and some other proteases that are active at low ph environment are critical for the fusogenic activation of the S protein. We then examined whether 229E is facilitated to enter cells from the cell surface, when receptor-bound 229E is treated with trypsin that activates S protein fusion activity. This phenomenon was observed in SARS-CoV and MHV-2 infections (26, 30, 33). HeLa-A cells treated with Baf, inhibitor of endosomal infection, were adsorbed with pseudotype VSV bearing either 229E S, SARS-CoV S or VSV-G at 4 o C for 30 min and then treated with trypsin. Infection of those pseudotypes was monitored by the expression of GFP. As shown in Fig. 7, the infection of pseudoypes with 229E S, as well as with SARS-CoV S, was 24
21 blocked by the Baf (Baf +, Try -) treatment, but the infection was facilitated when cell-adsorbed viruses were treated with trypsin (Baf +, Try +), while that with VSV G failed to infect even with trypsin treatment (Baf +, Try +). These results indicate that viruses with 229E S, as well as SARS-CoV S proteins, enter cells from the cell surface in the presence of trypsin, while that with VSV-G fails to do so. These results suggested that 229E enters cells directly from cell surface in the presence of protease that activates fusion potential of 229E S protein. Discussion In the present study, we have employed a few different approaches to analyze the 229E cell entry mechanism. We used infectious 229E together with the control viruses MHV-2 and JHMV; the former takes an endosomal pathway in a protease-dependent fashion, and the latter enters cells directly from a plasma membrane (16, 30). We also took advantage of pseudotype VSV bearing 229E S together with those bearing SARS-CoV S protein and VSV G protein. SARS-CoV enters in a protease-dependent fashion and VSV in a ph-dependent fashion, although those two viruses enter from an endosomal compartment of acidic environment. To compare the cell entry feature of 229E with those viruses, we established a cell line that expresses the receptors for SARS-CoV and MHV using HeLa cells originally permissive to 229E and VSV. By using these viruses, pseudotypes and cells, we showed that 229E enters cells by endocytosis, and proteases active at low ph condition are critical for its 25
22 infection. CPL is a possible protease that enables the fusogenic activation of the 229E S protein; however, some other not-yet-identified proteases active at low ph environment and also sensitive to cystein-protease inhibitor could be involved in this activation. Thus, 229E is the third coronavirus (first in the group 1 coronavirus) shown to enter cells in a protease-dependent fashion. Uncleavability of 229E S protein is shared by group 2 coronaviruses, SARS-CoV and MHV-2, as well as most of group 1 members, namely transmissible gastroenteritis virus, feline infectious peritonitis virus and the like (6). It is of interest to see whether those group 1 coronaviruses also utilize a protease-dependent entry like that of 229E The entry pathway of 229E has not been adequately studied, although the interaction of its receptor and S protein has been well investigated (3, 41). Nomura et al. reported that 229E enters cells in a caveolin-mediated fashion (28). They showed that caveolin-1 and 229E receptor APN co-localized on the lipid raft on the cell membrane and that inoculated 229E co-localized with caveolin-1. In addition to the binding of 229E to ANP molecules associated with caveolin-1 molecule, they further showed that knockdown of caveolin-1 by sirna reduced the infection of 229E, albeit the reduction was moderate (28). In contrast, our data shown in this study revealed that the blockade of endosomal pathway, e.g., the treatment of Baf and NH 4 Cl, reduced the infection quite efficiently, by 2 orders of magnitude. Generally, viruses that are endocytosed via caveolae are targeted to the endoplasmic reticulum or Golgi, and not to the endosome (32). However, it could happen that 229E incorporated into caveolae is trafficked to the endosome, although it is not a general pathway of caveolin-mediated infection or 26
23 caveolin-mediated endocytosis. A detailed study of the 229E entry pathway could delineate a new mechanism of viral invasion into the cell. The fusion activity of the coronavirus S proteins is thought to be independent of the cleavage site of the S protein. SARS-CoV S protein is fusogenically activated when cleaved at amino acid position 667 by trypsin and also at 678 by CPL (4, 22). We recently observed that SARS-CoV S protein cleaved at position 798, a position ca. 120 amino acids downstream of the cleavage sites of trypsin and CPL, is also fusogenic (43). As shown in this study, 229E S protein can be activated for the fusion of, at least, envelope-cell membranes, even if the protein is cleaved in a different position in the S protein. Judging from the size of cleavage products, ca. 100 kda and 80 kda by CPL and trypsin treatment, respectively, CPL cleavage site would be around the putative S1-S2 junction region (amino acid 576 from an initiator of S), which was assumed by the sequence alignment of various coronavirus S proteins (14), while that of trypsin is located further downstream. Since the fusion peptide on coronavirus S protein is not located at the N terminus of the membrane-anchored subunit, being different from the envelope proteins of HIV or influenza virus, multiple forms of the S2 subunit could be functional in virus-cell fusion, if the fusion peptide is located at a position accessible to the target membrane. Analysis of the precise cleavage site and fusion peptide of the 229E S protein will delineate the mechanism of how S2 interacts with the target membranes. It should be noted that three different modes of cell entry are utilized by various coronaviruses; one from the plasma membrane and the others from an endosomal 27
24 membrane in a low ph-dependent or a protease-dependent fashion. The MHV-JHMV strain is thought to enter cells from the cell surface, since its infection results in an extensive cell- to-cell fusion in a wide range of phs and is not influenced by the treatment of cells with lysosomotropic agents (16). In contrast, SARS-CoV enters cells via an endosomal compartment under a low-ph environment (26, 34). SARS-CoV S protein is activated by a protease (protease-dependent entry), such as CPS L, in the endosome (4, 18, 33). Endosomal conditions of acidity are critical for protease activation that induces cleavage of the SARS-CoV S protein. Previous and present studies on MHV-2 (30) and 229E, respectively, showed that those utilize a mechanism for cell entry similar to that of SARS-CoV. On the other hand, a mutant MHV isolated from persistently infected cells was reported to utilize an endosomal pathway similar to that of influenza s low ph-dependent entry (16). Quite recently avian infectious bronchitis virus (IBV) and MHV-A59 also have been reported to take a similar entry mode; moreover, the S protein of those viruses is fusogenically activated under a low ph environment, and no protease is required for activation (9, 13), although the entry mechanism of MHV-A59 is still controversial, since entry from cell surface and other mechanism of fusion activation have been reported (29, 30, 52). These observations indicate that coronavirus S protein is an appropriate tool to study the viral entry mechanism from molecular aspects. Three different coronaviruses, SARS-CoV, MHV-2 and 229E, have been revealed to enter cells directly from the cell surface, when the virion S proteins adsorbed to the cell surface receptor are cleaved with proteases, such as trypsin that 28
25 activate the fusion activity of those S protein. In agreement with those findings, cleaved S protein of SARS-CoV was recently shown to mediate the infection from cell surface (43). In SARS-CoV infection, the cell surface entry seemed to be more efficient than entry via the endosomes (26), which could account for extraordinarily high replication in the lungs where elastase, a protease to enable SARS-CoV entry from the cell surface (26), is secreted during the development of mild pneumonia. This efficient infection in the lungs could be a trigger for SARS, which was reproduced recently in mice infected with SARS-CoV and respiratory bacteria (1). Although 229E fails to induce a serious respiratory disease in humans, selective infection in the upper respiratory tract could be attributed to the specific proteases that could enable 229E infection from the cell surface. Since 229E receptor APN is expressed in a wide variety of cells in the body (17), and 229E infection is limited in the upper respiratory tract, a host factor other than receptor that determines the tissue-specificity of 229E must exist. Protease that enhances the infection, as observed in SARS-CoV infection (1, 26), may also participate in the pathogenesis of 229E infection. To delineate the pathogenesis of 229E, an analysis of such proteases would be an important direction. Acknowledgements We thank Naoko Matsubara for her technical assistance and Dr. Shigeru Morikawa for his valuable suggestions to use HeLa cells for 229E infection. We also thank Dr. Ryuji Nomura for 229E and anti-229e S serum and Dr. M. A. Whitt for VSV pseudotype system. This work was financially supported by grant-in-aids from the 29
26 Ministry of Education, Culture, Sports, Science and Technology of Japan (grant No and No ). References 1. Ami, Y., N. Nagata, K. Shirato, R. Watanabe, N. Iwata, K. Nakagaki, S. Fukushi, M. Saijo, S. Morikawa, and F. Taguchi Co-infection of respiratory bacterium with severe acute respiratory syndrome coronavirus induces an exacerbated pneumonia in mice. Microbiol Immunol 52: Appleyard, G., and M. Tisdale Inhibition of the growth of human coronavirus 229E by leupeptin. J Gen Virol 66 ( Pt 2): Bonavia, A., B. D. Zelus, D. E. Wentworth, P. J. Talbot, and K. V. Holmes Identification of a receptor-binding domain of the spike glycoprotein of human coronavirus HCoV-229E. J Virol 77: Bosch, B. J., W. Bartelink, and P. J. Rottier Cathepsin L functionally cleaves the severe acute respiratory syndrome coronavirus class I fusion protein upstream of rather than adjacent to the fusion peptide. J Virol 82: Bosch, B. J., B. E. Martina, R. Van Der Zee, J. Lepault, B. J. Haijema, C. Versluis, A. J. Heck, R. De Groot, A. D. Osterhaus, and P. J. Rottier Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc Natl Acad Sci U S A 101: Cavanagh, D The coronavirus surface glycoprotein, p In S. G. 30
27 Siddell (ed.), In the "Coronaviridae". Plenum press, New York. 7. Chan, D. C., and P. S. Kim HIV entry and its inhibition. Cell 93: Chandran, K., N. J. Sullivan, U. Felbor, S. P. Whelan, and J. M. Cunningham Endosomal proteolysis of the Ebola virus glycoprotein is necessary for infection. Science 308: Chu, V. C., L. J. McElroy, V. Chu, B. E. Bauman, and G. R. Whittaker The avian coronavirus infectious bronchitis virus undergoes direct low-ph-dependent fusion activation during entry into host cells. J Virol 80: Dveksler, G. S., M. N. Pensiero, C. B. Cardellichio, R. K. Williams, G. S. Jiang, K. V. Holmes, and C. W. Dieffenbach Cloning of the mouse hepatitis virus (MHV) receptor: expression in human and hamster cell lines confers susceptibility to MHV. J Virol 65: Dveksler, G. S., M. N. Pensiero, C. W. Dieffenbach, C. B. Cardellichio, A. A. Basile, P. E. Elia, and K. V. Holmes Mouse hepatitis virus strain A59 and blocking antireceptor monoclonal antibody bind to the N-terminal domain of cellular receptor. Proc Natl Acad Sci U S A 90: Eckert, D. M., and P. S. Kim Mechanisms of viral membrane fusion and its inhibition. Annu Rev Biochem 70: Eifart, P., K. Ludwig, C. Bottcher, C. A. de Haan, P. J. Rottier, T. Korte, and A. Herrmann Role of endocytosis and low ph in murine hepatitis virus strain A59 cell entry. J Virol 81:
28 14. Follis, K. E., J. York, and J. H. Nunberg Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell-cell fusion but does not affect virion entry. Virology 350: Fukushi, S., T. Mizutani, M. Saijo, S. Matsuyama, N. Miyajima, F. Taguchi, S. Itamura, I. Kurane, and S. Morikawa Vesicular stomatitis virus pseudotyped with severe acute respiratory syndrome coronavirus spike protein. J Gen Virol 86: Gallagher, T. M., C. Escarmis, and M. J. Buchmeier Alteration of the ph dependence of coronavirus-induced cell fusion: effect of mutations in the spike glycoprotein. J Virol 65: Holmes, K. V. a. C., S. R Coronavirus receptors, p In S. G. Siddell (ed.), In the "Coronaviridae". Plenum press, New York. 18. Huang, I. C., B. J. Bosch, F. Li, W. Li, K. H. Lee, S. Ghiran, N. Vasilieva, T. S. Dermody, S. C. Harrison, P. R. Dormitzer, M. Farzan, P. J. Rottier, and H. Choe SARS coronavirus, but not human coronavirus NL63, utilizes cathepsin L to infect ACE2-expressing cells. J Biol Chem 281: Kubo, H., S. Takase-Yoden, and F. Taguchi Neutralization and fusion inhibition activities of monoclonal antibodies specific for the S1 subunit of the spike protein of neurovirulent murine coronavirus JHMV c1-2 variant. J Gen Virol 74 ( Pt 7): Kusanagi, K., H. Kuwahara, T. Katoh, T. Nunoya, Y. Ishikawa, T. Samejima, and M. Tajima Isolation and serial propagation of porcine epidemic diarrhea 32
29 virus in cell cultures and partial characterization of the isolate. J Vet Med Sci 54: Lai, M. M., and D. Cavanagh The molecular biology of coronaviruses. Adv Virus Res 48: Li, F., M. Berardi, W. Li, M. Farzan, P. R. Dormitzer, and S. C. Harrison Conformational states of the severe acute respiratory syndrome coronavirus spike protein ectodomain. J Virol 80: J 23. Li, W., M. J. Moore, N. Vasilieva, J. Sui, S. K. Wong, M. A. Berne, M. Somasundaran, J. L. Sullivan, K. Luzuriaga, T. C. Greenough, H. Choe, and M. Farzan Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426: Matsuyama, S., and F. Taguchi Impaired entry of soluble receptor-resistant mutants of mouse hepatitis virus into cells expressing MHVR2 receptor. Virology 273: Matsuyama, S., and F. Taguchi Receptor-induced conformational changes of murine coronavirus spike protein. J Virol 76: Matsuyama, S., M. Ujike, S. Morikawa, M. Tashiro, and F. Taguchi Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection. Proc Natl Acad Sci U S A 102: Miura, H. S., K. Nakagaki, and F. Taguchi N-terminal domain of the murine coronavirus receptor CEACAM1 is responsible for fusogenic activation and conformational changes of the spike protein. J Virol 78:
30 28. Nomura, R., A. Kiyota, E. Suzaki, K. Kataoka, Y. Ohe, K. Miyamoto, T. Senda, and T. Fujimoto Human coronavirus 229E binds to CD13 in rafts and enters the cell through caveolae. J Virol 78: Pu, Y., and X. Zhang Mouse hepatitis virus type 2 enters cells through a clathrin-mediated endocytic pathway independent of Eps15. J. Virol. 82: Qiu, Z., S. T. Hingley, G. Simmons, C. Yu, J. Das Sarma, P. Bates, and S. R. Weiss Endosomal proteolysis by cathepsins is necessary for murine coronavirus mouse hepatitis virus type 2 spike-mediated entry. J Virol 80: Saeki, K., N. Ohtsuka, and F. Taguchi Identification of spike protein residues of murine coronavirus responsible for receptor-binding activity by use of soluble receptor-resistant mutants. J Virol 71: Sieczkarski, S. B., and G. R. Whittaker Viral entry. Curr Top Microbiol Immunol 285: Simmons, G., D. N. Gosalia, A. J. Rennekamp, J. D. Reeves, S. L. Diamond, and P. Bates Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc Natl Acad Sci U S A 102: Simmons, G., J. D. Reeves, A. J. Rennekamp, S. M. Amberg, A. J. Piefer, and P. Bates Characterization of severe acute respiratory syndrome-associated coronavirus (SARS-CoV) spike glycoprotein-mediated viral entry. Proc Natl Acad Sci U S A 101: Sturman, L. S., C. S. Ricard, and K. V. Holmes Proteolytic cleavage of 34
31 the E2 glycoprotein of murine coronavirus: activation of cell-fusing activity of virions by trypsin and separation of two different 90K cleavage fragments. J Virol 56: Suzuki, H., and F. Taguchi Analysis of the receptor-binding site of murine coronavirus spike protein. J Virol 70: Taguchi, F., and S. Matsuyama Soluble receptor potentiates receptor-independent infection by murine coronavirus. J Virol 76: Taguchi, F., S. G. Siddell, H. Wege, and V. ter Meulen Characterization of a variant virus selected in rat brains after infection by coronavirus mouse hepatitis virus JHM. J Virol 54: Takada, A., C. Robison, H. Goto, A. Sanchez, K. G. Murti, M. A. Whitt, and Y. Kawaoka A system for functional analysis of Ebola virus glycoprotein. Proc Natl Acad Sci U S A 94: Toth, T. E Trypsin-enhanced replication of neonatal calf diarrhea coronavirus in bovine embryonic lung cells. Am J Vet Res 43: Tusell, S. M., S. A. Schittone, and K. V. Holmes Mutational analysis of aminopeptidase N, a receptor for several group 1 coronaviruses, identifies key determinants of viral host range. J Virol 81: Ujike, M., H. Nishikawa, A. Otaka, N. Yamamoto, M. Matsuoka, E. Kodama, N. Fujii, and F. Taguchi Heptad repeat-derived peptides block protease-mediated direct entry from the cell surface of severe acute respiratory syndrome coronavirus but not entry via the endosomal pathway. J Virol 35
32 82: Watanabe, R., S. Matsuyama, K. Shirato, M. Maejima, S. Fukushi, S. Morikawa and F. Taguchi. (2008) Entry from cell surface of SARS coronavirus with cleaved S protein as revealed by pseudotype virus bearing cleaved S protein. J. Virol. In press. 44. Wege, H., S. Siddell, and V. ter Meulen The biology and pathogenesis of coronaviruses. Curr Top Microbiol Immunol 99: White, J. M Viral and cellular membrane fusion proteins. Annu Rev Physiol 52: Williams, R. K., G. S. Jiang, and K. V. Holmes Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc Natl Acad Sci U S A 88: Wool-Lewis, R. J., and P. Bates Characterization of Ebola virus entry by using pseudotyped viruses: identification of receptor-deficient cell lines. J Virol 72: Yamada, Y. K., K. Takimoto, M. Yabe, and F. Taguchi Acquired fusion activity of a murine coronavirus MHV-2 variant with mutations in the proteolytic cleavage site and the signal sequence of the S protein. Virology 227: Yeager, C. L., R. A. Ashmun, R. K. Williams, C. B. Cardellichio, L. H. Shapiro, A. T. Look, and K. V. Holmes Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 357:
33 50. Yoshikura, H., and S. Tejima Role of protease in mouse hepatitis virus-induced cell fusion. Studies with a cold-sensitive mutant isolated from a persistent infection. Virology 113: Zamoto, A., F. Taguchi, S. Fukushi, S. Morikawa, and Y. K. Yamada Identification of ferret ACE2 and its receptor function for SARS-coronavirus. Adv Exp Med Biol 581: Zelus, B. D., J. H. Schickli, D. M. Blau, S. R. Weiss and K. V. Holmes Conformational changes in the spike glycoprotein of murine coronavirus are induced at 37 degree C either by soluble murine CEACAM1 receptors or by ph 8. J. Virol. 77: Figure legends Fig. 1: Syncytia formation of 229E-infected HeLa cells induced by trypsin and other protease treatment. (A) HeLa cells infected with 229E were cultured for 2 days and then treated or left untreated with trypsin (100 µg/ml) at RT for 5 min. Cells were cultured for an additional 2 h and observed for syncytia formation and viral antigen via phase contrast (PC) and fluorescent (FA) microscopy, respectively. (B) Cells infected (Infect) with 229E or mock infected (Mock) were cultured for 2 days and treated with 100 µg/ml of trypsin (Try) or DMEM adjusted to ph 5.0 (ph 5.0) at RT for 5 min. After 2 h culture, cells were fixed with formaldehyde and stained with crystal violet. Cells without any treatment 37
34 were shown as controls (None). (C) Syncytia formation of cells infected with 229E was induced by treatment with various proteases. HeLa cells infected with 229E and cultured for 2 days were treated with a variety of proteases at RT for 5 min and cultured for an additional 2 h. Cells were fixed and stained as described above. Following concentrations were employed for treatment: Protease (-) (no protease), proteinase K (8µg/ml), Elastase (2mg/ml), Thermolysin (200µg/ml), Dispase (1unit/ml), Papain (0.19unit/ml), Chymotrypsin (2mg/ml) and Collagenase (1mg/ml). Fig. 2: Western blotting analysis of the 229E S protein after treatment with trypsin. 229E virions concentrated as described in Materials and Methods were treated with trypsin (1: 0 µg/ml, 2: 50 µg/ml, 3: 100 µg/ml) at RT for 5 min. Also, 229E-infected and mock-infected HeLa cells were treated with trypsin (1: 0 µg/ml, 2: 250 µg/ml, 3: 500 µg/ml). The resultant samples were analyzed by Western blotting using anti-229e S antibodies raised with a synthetic peptide corresponding to the C terminus region of the S protein. Uncleaved S protein and cleaved S2 protein are indicated by black and white arrow heads, respectively. Fig. 3: Characterization of HeLa cells permissive to 229E infection. (A): HeLa cells infected with 229E and cultured for 1 day in the presence or absence of trypsin (5 ng/ml) in DMEM containing 10 % TPB were fixed and stained as described 38
35 above. The syncytia formed, indicated by arrows, were observed under microscopy as plaques. (B): To evaluate the plaque assay developed using HeLa cells and trypsin, 229E of ca. 120 PFU/50 µl were diluted by two-fold steps and inoculated into 3 wells of HeLa cells prepared in 24-well plates. The relationship of the virus dilution and plaque number was depicted. (C): HeLa cells prepared in 96-well plates were treated with anti-human APN sera at 4 o C for 45 min, and then allowed to be adsorbed with ca. 150 PFU of 229E at 4 o C for 45 min in the presence of antiserum. Next, those cells were cultured for 1 day in the presence of the antiserum. The infected cells were counted under fluorescence microscopy after cells had been stained with antibody against 229E and secondary FITC-labeled anti-rabbit serum. Cells in which infection was blocked by APN antibody were compared to those without APN antibody- treatment. Fig. 4: Effect of lysosomotropic agents on 229E infection. (A): HeLa cells in 24-well plates were treated with 100 nm of Baf from 1 h before infection to 4 h postinfection (early) or from 4-24 h post 229E infection (late). Virus titers were determined in cells at 24 h post infection.. HeLa cells were infected with an MOI of 1. (B): HeLa-AC cells prepared in 24-well plates were treated with 1000 µm of Baf or 50 mm NH 4 Cl for 1 h at 37 o C. Then, cells were infected with 1x10 5 PFU of virus (229E, MHV-JHM or MHV-2) in DMEM containing each agent and incubated at 34 o C (229E) or 37 o C (MHV) for 1 h. After washing once, cells were cultured with DMEM containing each agent for 3 h and then medium 39
36 was changed to DMEM without agent. Virus titers were examined by plaque assay in cells at 24 h (229E) or 10 h (MHV) after infection. (C): HeLa-A cells prepared in 96-well plates were treated with different concentrations of Baf or NH 4 Cl for 1 h, infected with ca. 500 IU of pseudotype VSV bearing 229E S (solid line with circle), SARS-CoV S (broken line with square) or VSV G protein (dotted line with triangle) and cultured for 24 h in the presence of each agent. Then, GFP-positive cells were photographed by Keyence fluorescence microscopy and counted. The percentage of infection was was calculated as follows: (GFP-positive cell number in the presence of agent)/(gfp-positive cell number in the absence of agent) x100. Fig. 5: Effects of protease inhibitors on pseudotype virus infection. HeLa-A cells pretreated with different concentrations of protease inhibitors for 1 h were infected with ca.500 IU of pseudotype VSV bearing 229E S (solid line with circle); SARS-CoV S (broken line with square) or VSV G protein (dotted line with triangle) and cultured at 37 o C for 24 h in the presence of protease inhibitors. The percentage of infection was determinedas described in Fig. 4C. Fig. 6: Effect of CPL knockdown by sirna on pseudotype infection (A and B) and digestion of 229E S by CPL (C). (A): HeLa-A cells were treated with sirna for CPL (1) and control nonsense RNA (2) or were left untreated (3) and cultured for 4 days. Amounts of CPL expressed in those cells were examined 40
37 by Western blotting. Arrowhead indicates the position of CPL. (B): HeLa-A cells transfected with CPL sirna and cultured for 4 days (black column) were infected with ca. 500 IU pseudotype VSV with 229E S, SARS-CoV S or VSV G protein. Their infection rates (GFP-positive cell number) were compared to those in cells transfected with control RNA (grey column). (C): 229E virions concentrated as described in Materials and Methods were treated with 36 µg/ ml of CPL (2), CPB (3) or mock-treated (1) at 37 o C for 30 min in a buffer of ph 5.0, and S protein was analyzed by Western blotting using anti-229e S antibodies as described in Fig. 2 legend. As a control, virions were also treated (5) or mock-treated (4) with 50 µg/ml of trypsin at 37 o C for 30 min in a buffer of ph. 70 and analyzed by Western blot. Fig. 7: Infection with 229E S bearing pseudotype VSV from cell surface. HeLa-A cells prepared in 96-well plates were treated with 100 nm Baf for 1 h at 37 o C and then allowed to adsorb VSV bearing 229E S, SARS-CoV S or VSV G protein on ice for 30 min. The plate was left at RT for 10 min, and adsorbed virus was treated with 100 µg/ml trypsin for 5 min. Then, those cells were cultured for 24 h at 37 o C and GFP-positive cell numbers were calculated as described above (Baf +, Try +). GFP-positive cell numbers were also determined in the groups of cells treated with: Baf alone (Baf +, Try -); trypsin alone (Baf -, Try +); or maintained as untreated control cells (Baf -, Try -). The number of GFP-positive cells in each group was shown as a percentage in 41
38 comparison with that of GFP-positive cells without any treatment. 42
 B")












39 A FA PC Try (+) B Try Infect C A C Protease (-) Dispase Proteinace K Papain ph 5.0 D E T P E C Mock Try (-) ownloaded from on September 24, 2018 by guest Fig. 1 Elastase Thermolysin Chymotrypsin Collagenase None


40 Fig Cells Virion Infected Uninfected wnloaded from on September 24, 2018 by guest
 Plaque number (PFU) Try (-) 120 D E T P 120 100")

41 A B C A 60 E C 0 Try (+) Virus dilution 20 Block of infection (%) Plaque number (PFU) Try (-) 120 D E T P ownloaded from on September 24, 2018 by guest Fig. 3 0 C No Ab Antibody dilution (5 x 2x)
42 Fig. 4 A Virus titer (log 10 PFU/ml) Cr early late Virus titer (log 10 PFU/ml) Cr 229E Baf NH 4 Cl B Cr MHV-JHM Baf NH 4 Cl Cr MHV-2 Baf NH 4 Cl Infection (%) Infection (%) wnloaded from on September 24, 2018 by guest C SARS Bafilomycin (nm) 229E VSV NH 4 Cl (mm)
43 Fig. 5 Infection (%) Infection (%) Leupeptin 229E CPK inhibitor VSV SARS CA CPS inhibitor EST CPL inhibitor II MDL wnloaded from on September 24, 2018 by guest µm µm
 260 160 110 80 60 1 2 3 wnloaded from http://jvi.asm.")
44 Fig. 6 A Mw (kda) Infection (%) B E SARS- CoV VSV C Mw (kda) wnloaded from on September 24, 2018 by guest 4 5
Protease-Mediated Entry via the Endosome of Human Coronavirus 229E
 JOURNAL OF VIROLOGY, Jan. 2009, p. 712 721 Vol. 83, No. 2 0022-538X/09/$08.00 0 doi:10.1128/jvi.01933-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Protease-Mediated Entry
JOURNAL OF VIROLOGY, Jan. 2009, p. 712 721 Vol. 83, No. 2 0022-538X/09/$08.00 0 doi:10.1128/jvi.01933-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Protease-Mediated Entry
Received 8 July 2008/Accepted 3 September 2008
 JOURNAL OF VIROLOGY, Dec. 2008, p. 11985 11991 Vol. 82, No. 23 0022-538X/08/$08.00 0 doi:10.1128/jvi.01412-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Entry from the Cell
JOURNAL OF VIROLOGY, Dec. 2008, p. 11985 11991 Vol. 82, No. 23 0022-538X/08/$08.00 0 doi:10.1128/jvi.01412-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Entry from the Cell
Title Simultaneous treatment of human bronchial epithelial cells with serine and cysteine protease inhibitors prevents SARS coronavirus entry
 JVI Accepts, published online ahead of print on 11 April 212 J. Virol. doi:1.1128/jvi.94-12 Copyright 212, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16
JVI Accepts, published online ahead of print on 11 April 212 J. Virol. doi:1.1128/jvi.94-12 Copyright 212, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16
Severe acute respiratory syndrome (SARS) is caused by a
 is caused by a") Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection Shutoku Matsuyama*, Makoto Ujike*, Shigeru Morikawa, Masato Tashiro*, and Fumihiro Taguchi* Division of Respiratory
Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection Shutoku Matsuyama*, Makoto Ujike*, Shigeru Morikawa, Masato Tashiro*, and Fumihiro Taguchi* Division of Respiratory
Coronaviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Endosomal Proteolysis by Cathepsins Is Necessary for Murine Coronavirus Mouse Hepatitis Virus Type 2 Spike-Mediated Entry
 JOURNAL OF VIROLOGY, June 2006, p. 5768 5776 Vol. 80, No. 12 0022-538X/06/$08.00 0 doi:10.1128/jvi.00442-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Endosomal Proteolysis
JOURNAL OF VIROLOGY, June 2006, p. 5768 5776 Vol. 80, No. 12 0022-538X/06/$08.00 0 doi:10.1128/jvi.00442-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Endosomal Proteolysis
The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding
 Korean J. Microbiol. Biotechnol. Vol. 39, No. 2, 140 145 (2011) The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding Lee, Dong-Kyu 1, Se-Yeoun
Korean J. Microbiol. Biotechnol. Vol. 39, No. 2, 140 145 (2011) The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding Lee, Dong-Kyu 1, Se-Yeoun
Department of Virology III, National Institute of Infectious Diseases, Tokyo ; and 2
 Jpn. J. Infect. Dis., 64, 499-505, 2011 Original Article Receptor-Independent Infection by Mutant Viruses Newly Isolated from the Neuropathogenic Mouse Hepatitis Virus srr7 Detected through a Combination
Jpn. J. Infect. Dis., 64, 499-505, 2011 Original Article Receptor-Independent Infection by Mutant Viruses Newly Isolated from the Neuropathogenic Mouse Hepatitis Virus srr7 Detected through a Combination
Role of Endocytosis and Low ph in Murine Hepatitis Virus Strain A59 Cell Entry
 JOURNAL OF VIROLOGY, Oct. 2007, p. 10758 10768 Vol. 81, No. 19 0022-538X/07/$08.00 0 doi:10.1128/jvi.00725-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Role of Endocytosis
JOURNAL OF VIROLOGY, Oct. 2007, p. 10758 10768 Vol. 81, No. 19 0022-538X/07/$08.00 0 doi:10.1128/jvi.00725-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Role of Endocytosis
Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
 J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
Virus Entry/Uncoating
 Virus Entry/Uncoating Delivery of genome to inside of a cell Genome must be available for first step of replication The Problem--barriers to infection Virion Barriers: Non-enveloped viruses capsid Enveloped
Virus Entry/Uncoating Delivery of genome to inside of a cell Genome must be available for first step of replication The Problem--barriers to infection Virion Barriers: Non-enveloped viruses capsid Enveloped
Coronaviruses cause acute, mild upper respiratory infection (common cold).
.") Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES. Held September 13-18, 1992, in Chantilly France
 PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES Held September 13-18, 1992, in Chantilly France INDEX Adenovirus expression, ofprcv S protein, 469 oftgev S gene fragments, 455 A59 virus,
PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES Held September 13-18, 1992, in Chantilly France INDEX Adenovirus expression, ofprcv S protein, 469 oftgev S gene fragments, 455 A59 virus,
Endosomal Proteolysis of the Ebola Virus Glycoprotein Is Necessary for Infection
 Endosomal Proteolysis of the Ebola Virus Glycoprotein Is Necessary for Infection Kartik Chandran, 1 Nancy J. Sullivan, 2 Ute Felbor, 3 Sean P. Whelan, 4 James M. Cunningham 1,4 * 1 Department of Medicine,
Endosomal Proteolysis of the Ebola Virus Glycoprotein Is Necessary for Infection Kartik Chandran, 1 Nancy J. Sullivan, 2 Ute Felbor, 3 Sean P. Whelan, 4 James M. Cunningham 1,4 * 1 Department of Medicine,
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Severe Acute Respiratory Syndrome (SARS) Coronavirus
 Coronavirus") Severe Acute Respiratory Syndrome (SARS) Coronavirus Coronaviruses Coronaviruses are single stranded enveloped RNA viruses that have a helical geometry. Coronaviruses are the largest of RNA viruses with
Severe Acute Respiratory Syndrome (SARS) Coronavirus Coronaviruses Coronaviruses are single stranded enveloped RNA viruses that have a helical geometry. Coronaviruses are the largest of RNA viruses with
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Conformational Changes in the Spike Glycoprotein of Murine Coronavirus Are Induced at 37 C either by Soluble Murine CEACAM1 Receptors or by ph 8
 JOURNAL OF VIROLOGY, Jan. 2003, p. 830 840 Vol. 77, No. 2 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.2.830 840.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Conformational
JOURNAL OF VIROLOGY, Jan. 2003, p. 830 840 Vol. 77, No. 2 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.2.830 840.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Conformational
The Avian Coronavirus Infectious Bronchitis Virus Undergoes Direct Low-pH-Dependent Fusion Activation during Entry into Host Cells
 JOURNAL OF VIROLOGY, Apr. 2006, p. 3180 3188 Vol. 80, No. 7 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.7.3180 3188.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. The Avian Coronavirus
JOURNAL OF VIROLOGY, Apr. 2006, p. 3180 3188 Vol. 80, No. 7 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.7.3180 3188.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. The Avian Coronavirus
The isolation of a novel human betacoronavirus was first accomplished
 Middle East Respiratory Syndrome Coronavirus Infection Mediated by the Transmembrane Serine Protease TMPRSS2 Kazuya Shirato, Miyuki Kawase, Shutoku Matsuyama Department of Virology III, National Institute
Middle East Respiratory Syndrome Coronavirus Infection Mediated by the Transmembrane Serine Protease TMPRSS2 Kazuya Shirato, Miyuki Kawase, Shutoku Matsuyama Department of Virology III, National Institute
Supporting Information
 Supporting Information Millet and Whittaker 10.1073/pnas.1407087111 SI Materials and Methods Plasmids and Viruses. Influenza virus strain A/WSN/1933(H1N1) HA-encoding and neuraminidase-encoding (NA-encoding)
Supporting Information Millet and Whittaker 10.1073/pnas.1407087111 SI Materials and Methods Plasmids and Viruses. Influenza virus strain A/WSN/1933(H1N1) HA-encoding and neuraminidase-encoding (NA-encoding)
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Part I. Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents.
 Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
JBC Papers in Press. Published on October 11, 2016 as Manuscript M
 JBC Papers in Press. Published on October 11, 2016 as Manuscript M116.740746 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m116.740746 Molecular mechanism for PEDV entry Cell entry of
JBC Papers in Press. Published on October 11, 2016 as Manuscript M116.740746 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m116.740746 Molecular mechanism for PEDV entry Cell entry of
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
11/15/2011. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Viral Genomes. Structural Features and Characteristics
 Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches
 Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches Development of novel nanoparticle-base vaccines for infectious bronchitis PI, Mazhar I. Khan; CoPI, Peter
Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches Development of novel nanoparticle-base vaccines for infectious bronchitis PI, Mazhar I. Khan; CoPI, Peter
SOME PROPERTIES OF ECHO AND COXSACKIE VIRUSES IN TISSUE CULTURE AND VARIATIONS BY HEAT
 THE KURUME MEDICAL JOURNAL Vol. 9, No. 1, 1962 SOME PROPERTIES OF ECHO AND COXSACKIE VIRUSES IN TISSUE CULTURE AND VARIATIONS BY HEAT SHIGERU YAMAMATO AND MASAHISA SHINGU Department of Microbiology, Kurume
THE KURUME MEDICAL JOURNAL Vol. 9, No. 1, 1962 SOME PROPERTIES OF ECHO AND COXSACKIE VIRUSES IN TISSUE CULTURE AND VARIATIONS BY HEAT SHIGERU YAMAMATO AND MASAHISA SHINGU Department of Microbiology, Kurume
MCB130 Midterm. GSI s Name:
 1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
 VIROLOGICA SINICA 2016, 31 (1): 49 56 DOI: 10.1007/s12250-015-3690-4 RESEARCH ARTICLE Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
VIROLOGICA SINICA 2016, 31 (1): 49 56 DOI: 10.1007/s12250-015-3690-4 RESEARCH ARTICLE Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
Received 3 September 2002/Accepted 15 January 2003
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Distinct Mechanisms of Entry by Envelope Glycoproteins of Marburg and Ebola (Zaire) Viruses
 Viruses") JOURNAL OF VIROLOGY, May 2000, p. 4933 4937 Vol. 74, No. 10 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Distinct Mechanisms of Entry by Envelope Glycoproteins
JOURNAL OF VIROLOGY, May 2000, p. 4933 4937 Vol. 74, No. 10 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Distinct Mechanisms of Entry by Envelope Glycoproteins
Pre-made Lentiviral Particles for Fluorescent Proteins
 Pre-made Lentiviral Particles for Fluorescent Proteins Catalog# Product Name Amounts Fluorescent proteins expressed under sucmv promoter: LVP001 LVP001-PBS LVP002 LVP002-PBS LVP011 LVP011-PBS LVP012 LVP012-PBS
Pre-made Lentiviral Particles for Fluorescent Proteins Catalog# Product Name Amounts Fluorescent proteins expressed under sucmv promoter: LVP001 LVP001-PBS LVP002 LVP002-PBS LVP011 LVP011-PBS LVP012 LVP012-PBS
PERSISTENT INFECTIONS WITH HUMAN PARAINFLUENZAVIRUS TYPE 3 IN TWO CELL LINES
 71 PERSISTENT INFECTIONS WITH HUMAN PARAINFLUENZAVIRUS TYPE 3 IN TWO CELL LINES Harold G. Jensen, Alan J. Parkinson, and L. Vernon Scott* Department of Microbiology & Immunology, University of Oklahoma
71 PERSISTENT INFECTIONS WITH HUMAN PARAINFLUENZAVIRUS TYPE 3 IN TWO CELL LINES Harold G. Jensen, Alan J. Parkinson, and L. Vernon Scott* Department of Microbiology & Immunology, University of Oklahoma
xcelligence Real-Time Cell Analyzers
 xcelligence Real-Time Cell Analyzers Application Note No. 9 A New Way to Monitor Virus-Mediated Cytopathogenicity Introduction One of the most important procedures in virology is the measurement of viral
xcelligence Real-Time Cell Analyzers Application Note No. 9 A New Way to Monitor Virus-Mediated Cytopathogenicity Introduction One of the most important procedures in virology is the measurement of viral
VIROLOGY. Engineering Viral Genomes: Retrovirus Vectors
 VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
The Infectious Cycle. Lecture 2 Biology W3310/4310 Virology Spring You know my methods, Watson --SIR ARTHUR CONAN DOYLE
 The Infectious Cycle Lecture 2 Biology W3310/4310 Virology Spring 2016 You know my methods, Watson --SIR ARTHUR CONAN DOYLE The Infectious Cycle Virologists divide the infectious cycle into steps to facilitate
The Infectious Cycle Lecture 2 Biology W3310/4310 Virology Spring 2016 You know my methods, Watson --SIR ARTHUR CONAN DOYLE The Infectious Cycle Virologists divide the infectious cycle into steps to facilitate
Virus Entry. Steps in virus entry. Penetration through cellular membranes. Intracellular transport John Wiley & Sons, Inc. All rights reserved.
 Virus Entry Steps in virus entry Penetration through cellular membranes Intracellular transport Steps in virus entry How do virions get into cells? Viruses of bacteria, archaea, algae and plants use different
Virus Entry Steps in virus entry Penetration through cellular membranes Intracellular transport Steps in virus entry How do virions get into cells? Viruses of bacteria, archaea, algae and plants use different
Ali Alabbadi. Bann. Bann. Dr. Belal
 31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
Temperature-Sensitive Mutants Isolated from Hamster and
 JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
Phosphate buffered saline (PBS) for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay
 for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay") Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Attachment and internalization of feline infectious peritonitis virus in feline blood monocytes and Crandell feline kidney cells
 Journal of General Virology (2007), 88, 2527 2532 DOI 10.1099/vir.0.82991-0 Short Communication Attachment and internalization of feline infectious peritonitis virus in feline blood monocytes and Crandell
Journal of General Virology (2007), 88, 2527 2532 DOI 10.1099/vir.0.82991-0 Short Communication Attachment and internalization of feline infectious peritonitis virus in feline blood monocytes and Crandell
Gastroenteritis and viral infections
 Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12005 a S Ψ ΨΨ ΨΨ ΨΨ Ψ Ψ Ψ Ψ Ψ ΨΨ Ψ Ψ ΨΨΨΨΨΨ S1 (a.a. 1 751) S2 (a.a. 752 1353) S1 Fc S1 (a.a. 1-747) Fc b HCoV-EMC-S1-Fc SARS-CoV-S1-Fc HCoV-EMC-S1-Fc SARS-CoV-S1-Fc kda - 170 - - 130
doi:10.1038/nature12005 a S Ψ ΨΨ ΨΨ ΨΨ Ψ Ψ Ψ Ψ Ψ ΨΨ Ψ Ψ ΨΨΨΨΨΨ S1 (a.a. 1 751) S2 (a.a. 752 1353) S1 Fc S1 (a.a. 1-747) Fc b HCoV-EMC-S1-Fc SARS-CoV-S1-Fc HCoV-EMC-S1-Fc SARS-CoV-S1-Fc kda - 170 - - 130
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
SUPPLEMENTARY INFORMATION
 sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
In Vitro Cultivation of Human Rotavirus in MA 104 Cells
 Acute Diarrhea: Its Nutritional Consequences in Children, edited by J. A. Bellanti. Nestle, Vevey/Raven Press, New York 1983. ETIOLOGIC AGENTS OF ACUTE DIARRHEA In Vitro Cultivation of Human Rotavirus
Acute Diarrhea: Its Nutritional Consequences in Children, edited by J. A. Bellanti. Nestle, Vevey/Raven Press, New York 1983. ETIOLOGIC AGENTS OF ACUTE DIARRHEA In Vitro Cultivation of Human Rotavirus
DATA SHEET. Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter calf thymus DNA.
 Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
Sequence analysis of the S gene of recombinant MHV-2/A59 coronaviruses reveals three candidate mutations associated with demyelination and hepatitis
 Journal of NeuroVirology, 7: 432± 436, 2001 c 2001 Taylor & Francis ISSN 1355± 0284/01 $12.00+.00 Sequence analysis of the S gene of recombinant MHV-2/A59 coronaviruses reveals three candidate mutations
Journal of NeuroVirology, 7: 432± 436, 2001 c 2001 Taylor & Francis ISSN 1355± 0284/01 $12.00+.00 Sequence analysis of the S gene of recombinant MHV-2/A59 coronaviruses reveals three candidate mutations
Sarah A. Kopecky and Douglas S. Lyles*
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
Supplemental Information Dose Response Parameters for Gain of Function Pathogens
 Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents. Acellular Agents
 viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
Ralf Wagner Paul-Ehrlich-Institut
 www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
EVALUATION OF THE EFFECTIVENESS OF A 7% ACCELERATED HYDROGEN PEROXIDE-BASED FORMULATION AGAINST CANINE PARVOVIRUS
 Final report submitted to Virox Technologies, Inc. EVALUATION OF THE EFFECTIVENESS OF A 7% ACCELERATED HYDROGEN PEROXIDE-BASED FORMULATION AGAINST CANINE PARVOVIRUS Syed A. Sattar, M.Sc., Dip. Bact., M.S.,
Final report submitted to Virox Technologies, Inc. EVALUATION OF THE EFFECTIVENESS OF A 7% ACCELERATED HYDROGEN PEROXIDE-BASED FORMULATION AGAINST CANINE PARVOVIRUS Syed A. Sattar, M.Sc., Dip. Bact., M.S.,
(;[rowth Charaeteristies of Influenza Virus Type C in Avian Hosts
 Archives of Virology 58, 349--353 (1978) Archives of Virology by Springer-Verlag 1978 (;[rowth Charaeteristies of Influena Virus Type C in Avian Hosts Brief Report By M ~R A~N D. AUSTIn, A. S. MONTO, and
Archives of Virology 58, 349--353 (1978) Archives of Virology by Springer-Verlag 1978 (;[rowth Charaeteristies of Influena Virus Type C in Avian Hosts Brief Report By M ~R A~N D. AUSTIn, A. S. MONTO, and
Chapter 5. Virus isolation and identification of measles and rubella in cell culture
 Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
Department of Molecular Medicine, University of Padua, Via A. Gabelli 63, Padua, Italy
 Short communication Amiodarone affects Ebola virus binding and entry into target cells Cristiano Salata 1, Denis Munegato 1, Francesco Martelli 1, Cristina Parolin 1, Arianna Calistri 1, Aldo Baritussio
Short communication Amiodarone affects Ebola virus binding and entry into target cells Cristiano Salata 1, Denis Munegato 1, Francesco Martelli 1, Cristina Parolin 1, Arianna Calistri 1, Aldo Baritussio
Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests
 Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests A. Wise, Matti Kiupel,, C. Isenhour, R. Maes Coronaviruses
Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests A. Wise, Matti Kiupel,, C. Isenhour, R. Maes Coronaviruses
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
287 Current Topics in Microbiology and Immunology
 287 Current Topics in Microbiology and Immunology Editors R.W. Compans, Atlanta/Georgia M.D. Cooper, Birmingham/Alabama T. Honjo, Kyoto H. Koprowski, Philadelphia/Pennsylvania F. Melchers, Basel M.B.A.
287 Current Topics in Microbiology and Immunology Editors R.W. Compans, Atlanta/Georgia M.D. Cooper, Birmingham/Alabama T. Honjo, Kyoto H. Koprowski, Philadelphia/Pennsylvania F. Melchers, Basel M.B.A.
Cleavage Inhibition of the Murine Coronavirus Spike Protein by a Furin-Like Enzyme Affects Cell-Cell but Not Virus-Cell Fusion
 JOURNAL OF VIROLOGY, June 2004, p. 6048 6054 Vol. 78, No. 11 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.11.6048 6054.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Cleavage
JOURNAL OF VIROLOGY, June 2004, p. 6048 6054 Vol. 78, No. 11 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.11.6048 6054.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Cleavage
Broad Spectrum Protease Inhibitor Cocktail
 Broad Spectrum Protease Inhibitor Cocktail Catalog number: AR1182 Boster s Broad Spectrum Protease Inhibitor Cocktail is a complex of various protease inhibitors, which has been tested for inhibiting proteases
Broad Spectrum Protease Inhibitor Cocktail Catalog number: AR1182 Boster s Broad Spectrum Protease Inhibitor Cocktail is a complex of various protease inhibitors, which has been tested for inhibiting proteases
AP Biology. Viral diseases Polio. Chapter 18. Smallpox. Influenza: 1918 epidemic. Emerging viruses. A sense of size
 Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24)
") Product Manual QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24) Catalog Number VPK-107 VPK-107-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction
Product Manual QuickTiter Lentivirus Titer Kit (Lentivirus-Associated HIV p24) Catalog Number VPK-107 VPK-107-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction
Plasmid-Driven Formation of Influenza Virus-Like Particles
 JOURNAL OF VIROLOGY, Jan. 2000, p. 547 551 Vol. 74, No. 1 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Plasmid-Driven Formation of Influenza Virus-Like
JOURNAL OF VIROLOGY, Jan. 2000, p. 547 551 Vol. 74, No. 1 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Plasmid-Driven Formation of Influenza Virus-Like
The clathrin adaptor Numb regulates intestinal cholesterol. absorption through dynamic interaction with NPC1L1
 The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
Chapter 13 Viruses, Viroids, and Prions. Biology 1009 Microbiology Johnson-Summer 2003
 Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
VIRAL TITER COUNTS. The best methods of measuring infectious lentiviral titer
 VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma
 Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
The chemical fate of biological pollutants in treatment processes
 The chemical fate of biological pollutants in treatment processes http://pubs.rsc.org/en/content/articlehtml/2011/sm/c1sm060 92a http://www.epa.gov/nerlcwww/polio.htm Source: Centers for Disease Control
The chemical fate of biological pollutants in treatment processes http://pubs.rsc.org/en/content/articlehtml/2011/sm/c1sm060 92a http://www.epa.gov/nerlcwww/polio.htm Source: Centers for Disease Control
19 Viruses BIOLOGY. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Structural Features and Characteristics
 9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS
 Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3
 Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
Conformational States of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein Ectodomain
 JOURNAL OF VIROLOGY, July 2006, p. 6794 6800 Vol. 80, No. 14 0022-538X/06/$08.00 0 doi:10.1128/jvi.02744-05 Copyright 2006, American Society for Microbiology. All Rights Reserved. Conformational States
JOURNAL OF VIROLOGY, July 2006, p. 6794 6800 Vol. 80, No. 14 0022-538X/06/$08.00 0 doi:10.1128/jvi.02744-05 Copyright 2006, American Society for Microbiology. All Rights Reserved. Conformational States
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or
 Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Host Restriction of Friend Leukemia Virus. Role of the Viral Outer Coat (mice/fv-1 locus/vesicular stomatitis virus)
") Proc. Nat. Acad. Sci. USA Vol. 70, No. 9, pp. 2549-2553, September 1973 Host Restriction of Friend Leukemia Virus. Role of the Viral Outer Coat (mice/fv-1 locus/vesicular stomatitis virus) THEODORE G.
Proc. Nat. Acad. Sci. USA Vol. 70, No. 9, pp. 2549-2553, September 1973 Host Restriction of Friend Leukemia Virus. Role of the Viral Outer Coat (mice/fv-1 locus/vesicular stomatitis virus) THEODORE G.
Test Report. Efficacy of A New JM Nanocomposite Material in Inhibiting Respiratory Syncytial Virus Cellular Infection
 Test Report Efficacy of A New JM Nanocomposite Material in Inhibiting Respiratory Syncytial Virus Cellular Infection Test Reagent New JM Nanocomposite Material Project Commissioner JM Material Technology,
Test Report Efficacy of A New JM Nanocomposite Material in Inhibiting Respiratory Syncytial Virus Cellular Infection Test Reagent New JM Nanocomposite Material Project Commissioner JM Material Technology,
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
The Murine Coronavirus Mouse Hepatitis Virus Strain A59 from Persistently Infected Murine Cells Exhibits an Extended Host Range
 JOURNAL OF VIROLOGY, Dec. 1997, p. 9499 9507 Vol. 71, No. 12 0022-538X/97/$04.00 0 Copyright 1997, American Society for Microbiology The Murine Coronavirus Mouse Hepatitis Virus Strain A59 from Persistently
JOURNAL OF VIROLOGY, Dec. 1997, p. 9499 9507 Vol. 71, No. 12 0022-538X/97/$04.00 0 Copyright 1997, American Society for Microbiology The Murine Coronavirus Mouse Hepatitis Virus Strain A59 from Persistently
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Viruse associated gastrointestinal infection
 Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
Dr. Ahmed K. Ali Attachment and entry of viruses into cells
 Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Influenza B Hemagglutinin / HA ELISA Pair Set
 Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Riham. Bann. Bann. Belal
 30 Riham Bann Bann Belal Concepts to be covered in this lecture: 1. The Infectious Cycle 2. Virus cultivation 3. Viruses cytopathic effects 4. Viruses Quantification, Plaque assay, End-point dilution assay
30 Riham Bann Bann Belal Concepts to be covered in this lecture: 1. The Infectious Cycle 2. Virus cultivation 3. Viruses cytopathic effects 4. Viruses Quantification, Plaque assay, End-point dilution assay
QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA)
") New and Improved Product Manual QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA) Catalog Numbers VPK-108-HIV-p24 96 tests VPK-108-HIV-p24-5 5 x 96 tests FOR RESEARCH USE ONLY Not for use in diagnostic
New and Improved Product Manual QuickTiter Lentivirus Quantitation Kit (HIV p24 ELISA) Catalog Numbers VPK-108-HIV-p24 96 tests VPK-108-HIV-p24-5 5 x 96 tests FOR RESEARCH USE ONLY Not for use in diagnostic
Superinfection with Vaccinia Virus
 JOURNAL OF VIROLOGY, Aug. 1975, p. 322-329 Copyright 1975 American Society for Microbiology Vol. 16, No. 2 Printed in U.S.A. Abortive Infection of a Rabbit Cornea Cell Line by Vesicular Stomatitis Virus:
JOURNAL OF VIROLOGY, Aug. 1975, p. 322-329 Copyright 1975 American Society for Microbiology Vol. 16, No. 2 Printed in U.S.A. Abortive Infection of a Rabbit Cornea Cell Line by Vesicular Stomatitis Virus:
Size nm m m
 1 Viral size and organization Size 20-250nm 0.000000002m-0.000000025m Virion structure Capsid Core Acellular obligate intracellular parasites Lack organelles, metabolic activities, and reproduction Replicated
1 Viral size and organization Size 20-250nm 0.000000002m-0.000000025m Virion structure Capsid Core Acellular obligate intracellular parasites Lack organelles, metabolic activities, and reproduction Replicated