Ascertainment Through Family History of Disease Often Decreases the Power of Family-based Association Studies
|
|
|
- Gavin Nelson
- 5 years ago
- Views:
Transcription
1 Behav Genet (2007) 37: DOI 17/s ORIGINAL PAPER Ascertainment Through Family History of Disease Often Decreases the Power of Family-based Association Studies Manuel A. R. Ferreira Æ Pak Sham Æ Mark J. Daly Æ Shaun Purcell Received: 8 September 2006 / Accepted: 15 February 2007 / Published online: 20 March 2007 Ó Springer Science+Business Media, LLC 2007 Abstract Selection of cases with additional affected relatives has been shown to increase the power of the casecontrol association design. We investigated whether this strategy can also improve the power of family-based association studies that use the transmission disequilibrium test (TDT), while accounting for the effects of residual polygenic and environmental factors on disease liability. Ascertainment of parent-offspring trios conditional on the proband having affected first-degree relatives almost always reduced the power of the TDT. For many disease models, this reduction was quite considerable. In contrast, for the same sample size, designs that analyzed more than one affected offspring per family often improved power when compared to the standard parent-offspring trio design. Together, our results suggest that (1) residual polygenic and environmental influences should be considered when estimating the power of the TDT for studies that ascertain families with multiple affected relatives; (2) if trios are selected conditional on having additional affected Edited by David Allison M. A. R. Ferreira (&) M. J. Daly S. Purcell Center for Human Genetic Research, Massachusetts General Hospital, Harvard Medical School, 185 Cambridge St, Boston, MA 02114, USA mferreira@chgr.mgh.harvard.edu P. Sham Institute of Psychiatry, King s College, London, UK P. Sham Genome Research Centre, University of Hong Kong, Pokfulam, Hong Kong M. J. Daly S. Purcell Broad Institute of Harvard & MIT, Cambridge, MA, USA offspring, then it is important to genotype and include in the analysis the additional siblings; (3) the ascertainment strategy should be considered when interpreting results from TDT analyses. Our analytic approach to estimate the asymptotic power of the TDT is implemented online at Keywords Association TDT Power Study design Complex disease Family history Introduction Previous studies have demonstrated that the power of the case-control association design can be greatly improved by selecting familial cases, i.e. affected individuals with additional affected relatives (Risch 2001; Antoniou and Easton 2003; Howson et al. 2005; Li et al. 2006). The explanation for this is that by selecting familial cases, the frequency of the disease allele in the case sample will often increase (Risch 2001). However, this effect is somehow complicated by the number and magnitude of additional loci that contribute to disease risk. As the background heritability increases, the enrichment of the disease allele in familial cases is less pronounced (Risch 2001) and, consequently, the improvement in power as a result of selection is more modest (Li et al. 2006). For very high background heritability, selection may in fact reduce the power of the case-control design (Li et al. 2006). Nonetheless, across a realistic range of disease parameters, selection of familial cases seems to improve power. We were interested in investigating whether ascertainment of familial cases can also improve the power of the most popular test used in family-based association designs, the transmission disequilibrium test (TDT) (Spielman and
2 632 Behav Genet (2007) 37: Ewens 1996). Given the importance of efficient study design, particularly for large-scale association studies, surprisingly few studies have estimated the power of the TDT for different ascertainment strategies. Whittaker and Lewis (1998) showed that for a range of disease models, the power of the TDT improves when trios are selected conditional on having an affected parent. However, the authors implicitly assumed that no other genetic or shared environmental factors contributed to disease risk, which is unrealistic for common, multifactorial diseases. Indeed, Risch (2001) later showed that for diseases with high background heritability, there may be little advantage in ascertaining families with multiple affected offspring as opposed to families with one affected offspring only. In this report, we quantify the power of the TDT for different family designs and disease models. Specifically, we address two separate but related issues: first, can the power of the TDT be improved by ascertaining parentoffspring trios conditional on the proband having affected first-degree relatives? Henceforth we refer to this strategy as ascertainment of trios for family history of disease. Second, for a fixed number of genotypes, which strategy is more powerful, to analyze families with one affected offspring or, alternatively, with multiple affected offspring included in the association analysis ( multiplex design)? The first issue concerns ascertainment only: we compare designs that differ only in the number of affected firstdegree relatives the offspring must have for the trio to be ascertained (design 1 versus designs 2 6, Table 1). In this case, we analyze the same family structure for all designs considered, i.e. two parents and one affected offspring. On the other hand, to address the second issue, we compare family designs that differ in the number of affected offspring used in the TDT (design 1 versus designs 7 and 8, Table 1). Clearly, in this case, the designs differ in terms of both the ascertainment and family structure used for the TDT. Material and methods Below we outline our approach to estimate the asymptotic power of the TDT as a function of the ascertainment scheme, the properties of the disease locus and the residual genetic and environmental influences on disease. Following the standard biometrical model (Falconer 1981), we assume that liability to disease is continuously distributed and determined by a major biallelic locus, with the residual variance partitioned into additive polygenic background, and both shared (i.e. familial) and unique environment components. Individuals that exceed a threshold t in liability are affected. We model the three genotypic means as a (genotype AA), d (Aa) and -a (aa), where a is the additive Table 1 Asymptotic power of the TDT for eight family designs with a fixed total sample size of 1,500 genotyped individuals (a = 0.05) Design a Power of the TDT a Individuals are shown in white if their disease status is unknown and have been genotyped, in black if affected and genotyped, and in grey if affected but not genotyped. The disease prevalence was 5%, the additive test locus (p = 0.2) explained 1% of the variance in liability to disease, the background heritability was 50% and family environmental variance was 10% genetic value of the major locus, d is the dominance deviation, and p and q are the frequency of alleles A and a in the population (q =1 p), respectively. For a given set of a, d and p parameters, the contribution of the major locus to the total variance in liability to disease can be calculated as 2 p q ða þ d ðq pþþ 2 þ ð2 p q dþ 2. To calculate power, we require the expected number of transmitted and untransmitted alleles from heterozygous parents to affected offspring, given ascertainment and the disease model. These quantities are obtained by calculating the probability of each possible family genotypic configuration given the phenotypic ascertainment scheme, PG ð O ; G P ; G M jp O ; P P ; P M ¼ AÞ, where G O represents the set of genotypes for one or more offspring, G P and G M are paternal and maternal genotypes; P represents the
3 Behav Genet (2007) 37: phenotypes (same subscript) and A denotes the phenotypic ascertainment strategy. The standard TDT strategy is to ascertain families with a single affected offspring, disregarding the affection status of other relatives. We also consider designs in which other first-degree relatives (parents or siblings) are also affected. We use Bayes theorem to obtain PG ð O ; G P ; G M jp O ; P P ; P M ¼ AÞ from PG ð O ; G P ; G M Þ and PP ð O ; P P ; P M ¼ AjG O ; G P ; G M Þ. PG ð O ; G P ; G M Þ is the probability of each genotypic configuration in the unselected population; this is a simple function of genotype frequencies assuming Hardy Weinberg equilibrium and Mendelian transmission probabilities. For example, PG ð O ¼ 00 AA 00 ; G P ¼ 00 AA 00 ; G M ¼ 00 Aa 00 Þ ¼ p 2 2pq 0:5 ¼ p 3 q. The probability of the families phenotypic configuration given their genotypic configuration, PP ð O ; P P ; P M ¼ AjG O ; G P ; G M Þ, is given by numerical integration of the liability threshold model multivariate normal distribution with mean vector l (a function of the major locus) and covariance matrix S (a function of the residual polygenic and environmental components). Thus, the measured effect of genotype is specified in a model for the means, whereas the effects of unmeasured polygenic and environmental components are modeled as random effects in the covariance structure. This approach has been well described in a number of publications (e.g. Fulker et al. 1999; Abecasis et al. 2000). Finally, we obtain the expected number of transmitted and untransmitted alleles, b and c, as weighted sums of 0, 1 or 2 transmissions or non-transmissions per offspring per family over PG ð O ; G P ; G M jp O ; P P ; P M ¼ AÞ multiplied by the total number of families (for a fixed family type). The expected TDT test statistic is then computed astdt ¼ ðb cþ2 bþc ; power for a given type-i error rate a corresponds to the area of a non-central v 2 1 with non-centrality parameter = TDT that falls beyond the quantile q(a) from a central v 2 1 (e.g. q = 3.84 for a = 0.05). Note that our analytical approach accounts for the non-independence of parental transmissions to affected offspring and so it is valid for any modes of inheritance and number of affected offspring used in the TDT. The power estimates derived from our approach matched to the second decimal place the empirical power obtained through the analysis of 100,000 dataset replicates (not shown). Results To illustrate the ascertainment strategies and family structures tested, we first considered a single example disease model (Table 1). Both parents were assumed to be genotyped for all designs and the total sample size was fixed at 1,500 individuals. We considered a disease prevalence of 5%, an additive trait locus (p = 0.2) that explained 1% of the variance in liability to the disease (equivalent to a genotype relative risk of 1.9 for the heterozygote), a background heritability of 50% and a family environmental variance of 10%. Under this model, the power of the TDT when analyzing 500 parent-offspring trios ascertained with disregard to the affection status of first-degree relatives of the proband (design 1, which we refer to henceforth as the standard parent-offspring trio design ) was (a = 0.05). However, power decreased steadily as the number of affected first-degree relatives required to ascertain parent-offspring trios increased, reaching for cases with three additional affected relatives (design 6). For this particular disease model, the standard parent-offspring trio design also outperformed the designs that genotyped >1 affected offspring per family (multiplex designs 7 and 8). Next, we investigated whether the detrimental impact of selection on the TDT could be expected across a wider range of disease models. For this purpose, we considered three values for the disease prevalence (1%, 5% and 10%), two values for the variance in liability attributable to the test locus (1% and 2.5%), three genetic models (dominant, additive and recessive), three values for the family environment variance (V C ) (0%, 10% and 20%) and we varied the background heritability (V A ) between 0% and 75%, and the allele frequency p between and For simplicity, we restricted our analysis to five family structures (designs 1 3, 7 and 8). Figure 1 compares the expected TDT test statistic between family design 1 (standard parent-offspring trio design) and two designs that ascertain trios conditional on family history (designs 2 and 3). The color of each cell indicates which design resulted in the largest test statistic, parent-offspring trios (blue) or the trios with one (green) or two (red) additional affected siblings. The ability of family history to improve the power of the TDT was greatly influenced by the family environment variance (V C ), the background heritability (V A ), the allele frequency (p) and the genetic model considered. Disease prevalence and locus effect size had only a minor impact on the results and so data are only presented for a 5% disease prevalence and 1% locus effect size. Ascertainment based on family history reduced power in most scenarios, with the exception of recessive loci, particularly for rare risk alleles and low background heritability and family-specific environmental influences. We then performed the same analysis to compare the multiplex designs (7 and 8) against the standard parentoffspring trio design (design 1), maintaining a constant total sample size. When disease liability was not influenced by the family environment (V C = 0%), analyzing multiplex families was often the most powerful design (Fig. 2). In this case, the value of larger sibships increased as the



 or")

.")

 Family environmental")


 or families")

, %")
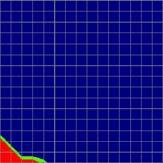
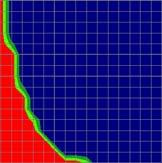
4 634 Behav Genet (2007) 37: Fig. 1 Comparison of the TDT test statistic between the standard parent-offspring trio design and two designs that ascertain trios conditional on family history. The color of each cell indicates which design resulted in the largest test statistic, parent-offspring trios ascertained with disregard to family history (design 1, in blue) or trios ascertained conditional on having one (design 2, in green) or two (design 3, in red) additional affected siblings. The two shades of green identify the design with the second best test statistic, design 1 (dark green) or design 3 (light green). Disease prevalence was 5% and the test locus explained 1% of the variance in disease liability. Total sample size was 1,500 individuals (including parents) for the three designs Allele frequency (p) Family environmental variance (V C ), % V C = 0 V C = 10 V C = 20 A B C D E F G H I Dominant Additive Recessive Background heritability (V A ), % Fig. 2 Comparison of the TDT test statistic between the standard parent-offspring trio design and two multiplex designs. The color of each cell indicates which design resulted in the largest test statistic, parent-offspring trios ascertained with disregard to family history (design 1, in blue) or families with two (design 7, in green) or three (design 8, in red) genotyped affected offspring. The two shades of green identify the design with the second best test statistic, design 1 (dark green) or design 8 (light green). Disease prevalence was 5% and the test locus explained 1% of the variance in disease liability. Total sample size was 1,500 individuals (including parents) for the three designs Allele frequency (p) Family environmental variance (V C ), % V C = 0 V C = 10 V C = 20 A B C D E F G H I Dominant Additive Recessive Background heritability (V A ), %
5 Behav Genet (2007) 37: background heritability and allele frequency decreased. On the other hand, when V C > 0%, larger sibships were beneficial if the background heritability was low to moderate (<50%), particularly when the risk allele was relatively rare. Discussion It is well established that selection of familial cases often improves the power of the case-control association design (Risch 2001; Antoniou and Easton 2003; Howson et al. 2005; Li et al. 2006). Despite a commonly-held belief that such ascertainment also improves the power of familybased tests, this hypothesis has not been thoroughly tested. Our results show that selection of familial cases can improve the power of the TDT for diseases in which additional untested loci and family environmental effects do not strongly influence disease risk. These results are consistent with those reported by Whittaker and Lewis (1998) but are unlikely to be applicable to most common, complex diseases, which are expected to be influenced by many disease loci and, in some cases, by family environmental factors. In this case, our results suggest that enrichment for familial aggregation of disease will often decrease the power of the TDT. For many disease models tested, this reduction in power was quite considerable. One intuitive explanation for these results is as follows. The power of the TDT increases with (a) increased transmission rate of the risk allele to the affected offspring and (b) increased parental heterozygosity. Ascertainment can reduce the transmission rate of the disease allele to affected offspring, especially when residual polygenic and family environmental effects are present. This occurs because ascertainment on family history acts to increase the effective prevalence of the disease (as in this case the probability of developing disease given family history, i.e. the recurrence risk, is substantially greater than the population prevalence) and the power of the TDT is influenced by prevalence. Specifically, the TDT is less powerful for higher prevalences, given the same effects on liability, which is why the undertransmission of the disease allele to unaffecteds contains little or no information for association. On the other hand, selection of familial cases will increase the frequency of the disease variant in founders, which will in turn increase parental heterozygosity, particularly for rarer, more highly penetrant disease alleles (although not when the disease causing variant is the more common allele). However, the increase in parental heterozygosity due to ascertainment is less pronounced with increasing residual polygenic and family environmental effects, again due to the increased effective prevalence. Table 2 Selection conditional on family history influences parental heterozygosity and the transmission rate of the risk allele Allele frequency p Background heritability, % Proportion of heterozygote parents a Proportion of heterozygote parents that transmit the risk allele b Expected TDT test statistic a Defined as bþc 2N, where b and c are the number of heterozygote parents that transmit the risk or non-risk alleles to the affected child, respectively, and N the total number of families b Defined as b bþc, where b and c are defined above Individuals are shown in white if their disease status is unknown and have been genotyped, in black if affected and genotyped, and in grey if affected but not genotyped. The disease prevalence was 5%, the additive test locus explained 1% of the variance in liability to disease and family environmental variance was 0%
6 636 Behav Genet (2007) 37: These effects are demonstrated in Table 2, in which we separate the two determinants of power for the TDT: parental heterozygosity and transmission distortion. We also investigated whether testing multiplex families could be expected to provide a more powerful TDT than testing parent-offspring trios, for a fixed number of genotyped samples. In this case, our results suggest that multiplex families are often beneficial unless the polygenic or the family environmental variances are high. Similar conclusions were reported by Risch (2001). This extra efficiency derives from being able to re-use the same parental genotypes to determine transmissions to more than one affected offspring (for a fixed total sample size). For example, if parental heterozygosity is 0.177, 500 parentoffspring trios would be expected to have ~177 (500 families 2 parents 0.177) informative transmissions, whereas 375 families with two affected offspring would have ~265 (375 families 2 parents offspring). These results may have important implications for the design, analysis and interpretation of association studies of complex diseases. First, residual polygenic and environmental influences should be considered when estimating the power of the TDT for studies that ascertain families with multiple affected relatives. Failure to do so may result in severely biased power analyses. Similarly, if families are ascertained based on having multiple affected offspring but are then broken down into parent-offspring trios to estimate power, this will often lead to biased results. Second, if trios are selected for a family history of disease, then it is important to genotype and include in the analysis the additional affected relatives. Finally, our results imply that the ascertainment strategy should be considered when interpreting results from TDT analyses, since for the same sample size different strategies will lead to different power levels. We have implemented our approach to estimate the asymptotic power of the TDT through a user-friendly module of the Genetic Power Calculator (Purcell et al. 2003) web interface ( The user can specify different ascertainment strategies, family structures and disease models to best describe individual studies. Other accurate methods to calculate the power of the TDT have been described (Knapp 1999; Chen and Deng 2001; Iles 2002) but, to our knowledge, ours is the first to account for the effects of background loci and family environment. Acknowledgments MARF was funded by Sidney Sax fellowship from the National Health and Medical Research Council of Australia. SP and PS acknowledge the Medical Research Council grant G and National Eye Institute grant EY We thank three anonymous reviewers for their comments and suggestions on an earlier version of this manuscript. References Abecasis GR, Cardon LR, Cookson WO (2000) A general test of association for quantitative traits in nuclear families. Am J Hum Genet 66: Antoniou AC, Easton DF (2003) Polygenic inheritance of breast cancer: implications for design of association studies. Genet Epidemiol 25: Chen WM, Deng HW (2001) A general and accurate approach for computing the statistical power of the transmission disequilibrium test for complex disease genes. Genet Epidemiol 21:53 67 Falconer DS (1981) Introduction to quantitative genetics. Longman, New York Fulker DW, Cherny SS, Sham PC, Hewitt JK (1999) Combined linkage and association sib-pair analysis for quantitative traits. Am J Hum Genet 64: Howson JM, Barratt BJ, Todd JA, Cordell HJ (2005) Comparison of population- and family-based methods for genetic association analysis in the presence of interacting loci. Genet Epidemiol 29:51 67 Iles MM (2002) On calculating the power of a TDT study comparison of methods. Ann Hum Genet 66: Knapp M (1999) A note on power approximations for the transmission/disequilibrium test. Am J Hum Genet 64: Li M, Boehnke M, Abecasis GR (2006) Efficient study designs for test of genetic association using sibship data and unrelated cases and controls. Am J Hum Genet 78: Purcell S, Cherny SS, Sham PC (2003) Genetic power calculator: design of linkage and association genetic mapping studies of complex traits. Bioinformatics 19: Risch N (2001) Implications of multilocus inheritance for genedisease association studies. Theor Popul Biol 60: Spielman RS, Ewens WJ (1996) The TDT and other family-based tests for linkage disequilibrium and association. Am J Hum Genet 59: Whittaker JC, Lewis CM (1998) The effect of family structure on linkage tests using allelic association. Am J Hum Genet 63:
Introduction to the Genetics of Complex Disease
 Introduction to the Genetics of Complex Disease Jeremiah M. Scharf, MD, PhD Departments of Neurology, Psychiatry and Center for Human Genetic Research Massachusetts General Hospital Breakthroughs in Genome
Introduction to the Genetics of Complex Disease Jeremiah M. Scharf, MD, PhD Departments of Neurology, Psychiatry and Center for Human Genetic Research Massachusetts General Hospital Breakthroughs in Genome
An Introduction to Quantitative Genetics I. Heather A Lawson Advanced Genetics Spring2018
 An Introduction to Quantitative Genetics I Heather A Lawson Advanced Genetics Spring2018 Outline What is Quantitative Genetics? Genotypic Values and Genetic Effects Heritability Linkage Disequilibrium
An Introduction to Quantitative Genetics I Heather A Lawson Advanced Genetics Spring2018 Outline What is Quantitative Genetics? Genotypic Values and Genetic Effects Heritability Linkage Disequilibrium
Dan Koller, Ph.D. Medical and Molecular Genetics
 Design of Genetic Studies Dan Koller, Ph.D. Research Assistant Professor Medical and Molecular Genetics Genetics and Medicine Over the past decade, advances from genetics have permeated medicine Identification
Design of Genetic Studies Dan Koller, Ph.D. Research Assistant Professor Medical and Molecular Genetics Genetics and Medicine Over the past decade, advances from genetics have permeated medicine Identification
Allowing for Missing Parents in Genetic Studies of Case-Parent Triads
 Am. J. Hum. Genet. 64:1186 1193, 1999 Allowing for Missing Parents in Genetic Studies of Case-Parent Triads C. R. Weinberg National Institute of Environmental Health Sciences, Research Triangle Park, NC
Am. J. Hum. Genet. 64:1186 1193, 1999 Allowing for Missing Parents in Genetic Studies of Case-Parent Triads C. R. Weinberg National Institute of Environmental Health Sciences, Research Triangle Park, NC
Complex Multifactorial Genetic Diseases
 Complex Multifactorial Genetic Diseases Nicola J Camp, University of Utah, Utah, USA Aruna Bansal, University of Utah, Utah, USA Secondary article Article Contents. Introduction. Continuous Variation.
Complex Multifactorial Genetic Diseases Nicola J Camp, University of Utah, Utah, USA Aruna Bansal, University of Utah, Utah, USA Secondary article Article Contents. Introduction. Continuous Variation.
BST227 Introduction to Statistical Genetics. Lecture 4: Introduction to linkage and association analysis
 BST227 Introduction to Statistical Genetics Lecture 4: Introduction to linkage and association analysis 1 Housekeeping Homework #1 due today Homework #2 posted (due Monday) Lab at 5:30PM today (FXB G13)
BST227 Introduction to Statistical Genetics Lecture 4: Introduction to linkage and association analysis 1 Housekeeping Homework #1 due today Homework #2 posted (due Monday) Lab at 5:30PM today (FXB G13)
Introduction to linkage and family based designs to study the genetic epidemiology of complex traits. Harold Snieder
 Introduction to linkage and family based designs to study the genetic epidemiology of complex traits Harold Snieder Overview of presentation Designs: population vs. family based Mendelian vs. complex diseases/traits
Introduction to linkage and family based designs to study the genetic epidemiology of complex traits Harold Snieder Overview of presentation Designs: population vs. family based Mendelian vs. complex diseases/traits
Effects of Stratification in the Analysis of Affected-Sib-Pair Data: Benefits and Costs
 Am. J. Hum. Genet. 66:567 575, 2000 Effects of Stratification in the Analysis of Affected-Sib-Pair Data: Benefits and Costs Suzanne M. Leal and Jurg Ott Laboratory of Statistical Genetics, The Rockefeller
Am. J. Hum. Genet. 66:567 575, 2000 Effects of Stratification in the Analysis of Affected-Sib-Pair Data: Benefits and Costs Suzanne M. Leal and Jurg Ott Laboratory of Statistical Genetics, The Rockefeller
Human population sub-structure and genetic association studies
 Human population sub-structure and genetic association studies Stephanie A. Santorico, Ph.D. Department of Mathematical & Statistical Sciences Stephanie.Santorico@ucdenver.edu Global Similarity Map from
Human population sub-structure and genetic association studies Stephanie A. Santorico, Ph.D. Department of Mathematical & Statistical Sciences Stephanie.Santorico@ucdenver.edu Global Similarity Map from
Mendelian & Complex Traits. Quantitative Imaging Genomics. Genetics Terminology 2. Genetics Terminology 1. Human Genome. Genetics Terminology 3
 Mendelian & Complex Traits Quantitative Imaging Genomics David C. Glahn, PhD Olin Neuropsychiatry Research Center & Department of Psychiatry, Yale University July, 010 Mendelian Trait A trait influenced
Mendelian & Complex Traits Quantitative Imaging Genomics David C. Glahn, PhD Olin Neuropsychiatry Research Center & Department of Psychiatry, Yale University July, 010 Mendelian Trait A trait influenced
Transmission Disequilibrium Test in GWAS
 Department of Computer Science Brown University, Providence sorin@cs.brown.edu November 10, 2010 Outline 1 Outline 2 3 4 The transmission/disequilibrium test (TDT) was intro- duced several years ago by
Department of Computer Science Brown University, Providence sorin@cs.brown.edu November 10, 2010 Outline 1 Outline 2 3 4 The transmission/disequilibrium test (TDT) was intro- duced several years ago by
Statistical power and significance testing in large-scale genetic studies
 STUDY DESIGNS Statistical power and significance testing in large-scale genetic studies Pak C. Sham 1 and Shaun M. Purcell 2,3 Abstract Significance testing was developed as an objective method for summarizing
STUDY DESIGNS Statistical power and significance testing in large-scale genetic studies Pak C. Sham 1 and Shaun M. Purcell 2,3 Abstract Significance testing was developed as an objective method for summarizing
Multifactorial Inheritance
 S e s s i o n 6 Medical Genetics Multifactorial Inheritance and Population Genetics J a v a d J a m s h i d i F a s a U n i v e r s i t y o f M e d i c a l S c i e n c e s, Novemb e r 2 0 1 7 Multifactorial
S e s s i o n 6 Medical Genetics Multifactorial Inheritance and Population Genetics J a v a d J a m s h i d i F a s a U n i v e r s i t y o f M e d i c a l S c i e n c e s, Novemb e r 2 0 1 7 Multifactorial
Chapter 2. Linkage Analysis. JenniferH.BarrettandM.DawnTeare. Abstract. 1. Introduction
 Chapter 2 Linkage Analysis JenniferH.BarrettandM.DawnTeare Abstract Linkage analysis is used to map genetic loci using observations on relatives. It can be applied to both major gene disorders (parametric
Chapter 2 Linkage Analysis JenniferH.BarrettandM.DawnTeare Abstract Linkage analysis is used to map genetic loci using observations on relatives. It can be applied to both major gene disorders (parametric
Lecture 1 Mendelian Inheritance
 Genes Mendelian Inheritance Lecture 1 Mendelian Inheritance Jurg Ott Gregor Mendel, monk in a monastery in Brünn (now Brno in Czech Republic): Breeding experiments with the garden pea: Flower color and
Genes Mendelian Inheritance Lecture 1 Mendelian Inheritance Jurg Ott Gregor Mendel, monk in a monastery in Brünn (now Brno in Czech Republic): Breeding experiments with the garden pea: Flower color and
Nature Genetics: doi: /ng Supplementary Figure 1
 Supplementary Figure 1 Illustrative example of ptdt using height The expected value of a child s polygenic risk score (PRS) for a trait is the average of maternal and paternal PRS values. For example,
Supplementary Figure 1 Illustrative example of ptdt using height The expected value of a child s polygenic risk score (PRS) for a trait is the average of maternal and paternal PRS values. For example,
Comparison of Linkage-Disequilibrium Methods for Localization of Genes Influencing Quantitative Traits in Humans
 Am. J. Hum. Genet. 64:1194 105, 1999 Comparison of Linkage-Disequilibrium Methods for Localization of Genes Influencing Quantitative Traits in Humans Grier P. Page and Christopher I. Amos Department of
Am. J. Hum. Genet. 64:1194 105, 1999 Comparison of Linkage-Disequilibrium Methods for Localization of Genes Influencing Quantitative Traits in Humans Grier P. Page and Christopher I. Amos Department of
Nonparametric Linkage Analysis. Nonparametric Linkage Analysis
 Limitations of Parametric Linkage Analysis We previously discued parametric linkage analysis Genetic model for the disease must be specified: allele frequency parameters and penetrance parameters Lod scores
Limitations of Parametric Linkage Analysis We previously discued parametric linkage analysis Genetic model for the disease must be specified: allele frequency parameters and penetrance parameters Lod scores
STATISTICAL GENETICS 98 Transmission Disequilibrium, Family Controls, and Great Expectations
 Am. J. Hum. Genet. 63:935 941, 1998 STATISTICAL GENETICS 98 Transmission Disequilibrium, Family Controls, and Great Expectations Daniel J. Schaid Departments of Health Sciences Research and Medical Genetics,
Am. J. Hum. Genet. 63:935 941, 1998 STATISTICAL GENETICS 98 Transmission Disequilibrium, Family Controls, and Great Expectations Daniel J. Schaid Departments of Health Sciences Research and Medical Genetics,
Ch. 23 The Evolution of Populations
 Ch. 23 The Evolution of Populations 1 Essential question: Do populations evolve? 2 Mutation and Sexual reproduction produce genetic variation that makes evolution possible What is the smallest unit of
Ch. 23 The Evolution of Populations 1 Essential question: Do populations evolve? 2 Mutation and Sexual reproduction produce genetic variation that makes evolution possible What is the smallest unit of
Transmission Disequilibrium Methods for Family-Based Studies Daniel J. Schaid Technical Report #72 July, 2004
 Transmission Disequilibrium Methods for Family-Based Studies Daniel J. Schaid Technical Report #72 July, 2004 Correspondence to: Daniel J. Schaid, Ph.D., Harwick 775, Division of Biostatistics Mayo Clinic/Foundation,
Transmission Disequilibrium Methods for Family-Based Studies Daniel J. Schaid Technical Report #72 July, 2004 Correspondence to: Daniel J. Schaid, Ph.D., Harwick 775, Division of Biostatistics Mayo Clinic/Foundation,
Pedigree Construction Notes
 Name Date Pedigree Construction Notes GO TO à Mendelian Inheritance (http://www.uic.edu/classes/bms/bms655/lesson3.html) When human geneticists first began to publish family studies, they used a variety
Name Date Pedigree Construction Notes GO TO à Mendelian Inheritance (http://www.uic.edu/classes/bms/bms655/lesson3.html) When human geneticists first began to publish family studies, they used a variety
Mendelian Inheritance. Jurg Ott Columbia and Rockefeller Universities New York
 Mendelian Inheritance Jurg Ott Columbia and Rockefeller Universities New York Genes Mendelian Inheritance Gregor Mendel, monk in a monastery in Brünn (now Brno in Czech Republic): Breeding experiments
Mendelian Inheritance Jurg Ott Columbia and Rockefeller Universities New York Genes Mendelian Inheritance Gregor Mendel, monk in a monastery in Brünn (now Brno in Czech Republic): Breeding experiments
MULTIFACTORIAL DISEASES. MG L-10 July 7 th 2014
 MULTIFACTORIAL DISEASES MG L-10 July 7 th 2014 Genetic Diseases Unifactorial Chromosomal Multifactorial AD Numerical AR Structural X-linked Microdeletions Mitochondrial Spectrum of Alterations in DNA Sequence
MULTIFACTORIAL DISEASES MG L-10 July 7 th 2014 Genetic Diseases Unifactorial Chromosomal Multifactorial AD Numerical AR Structural X-linked Microdeletions Mitochondrial Spectrum of Alterations in DNA Sequence
Effect of Genetic Heterogeneity and Assortative Mating on Linkage Analysis: A Simulation Study
 Am. J. Hum. Genet. 61:1169 1178, 1997 Effect of Genetic Heterogeneity and Assortative Mating on Linkage Analysis: A Simulation Study Catherine T. Falk The Lindsley F. Kimball Research Institute of The
Am. J. Hum. Genet. 61:1169 1178, 1997 Effect of Genetic Heterogeneity and Assortative Mating on Linkage Analysis: A Simulation Study Catherine T. Falk The Lindsley F. Kimball Research Institute of The
Statistical Genetics : Gene Mappin g through Linkag e and Associatio n
 Statistical Genetics : Gene Mappin g through Linkag e and Associatio n Benjamin M Neale Manuel AR Ferreira Sarah E Medlan d Danielle Posthuma About the editors List of contributors Preface Acknowledgements
Statistical Genetics : Gene Mappin g through Linkag e and Associatio n Benjamin M Neale Manuel AR Ferreira Sarah E Medlan d Danielle Posthuma About the editors List of contributors Preface Acknowledgements
DEFINITIONS: POPULATION: a localized group of individuals belonging to the same species
 DEFINITIONS: POPULATION: a localized group of individuals belonging to the same species SPECIES: a group of populations whose individuals have the potential to interbreed and produce fertile offspring
DEFINITIONS: POPULATION: a localized group of individuals belonging to the same species SPECIES: a group of populations whose individuals have the potential to interbreed and produce fertile offspring
Alzheimer Disease and Complex Segregation Analysis p.1/29
 Alzheimer Disease and Complex Segregation Analysis Amanda Halladay Dalhousie University Alzheimer Disease and Complex Segregation Analysis p.1/29 Outline Background Information on Alzheimer Disease Alzheimer
Alzheimer Disease and Complex Segregation Analysis Amanda Halladay Dalhousie University Alzheimer Disease and Complex Segregation Analysis p.1/29 Outline Background Information on Alzheimer Disease Alzheimer
Inheritance Of Alzheimer s Disease Investigated By Complex Segregation Analysis
 Inheritance Of Alzheimer s Disease Investigated By Complex Segregation Analysis Manzano S, MD, PhD 1, Baron M, MD, PhD 2, Gomez-Tortosa E, MD, PhD 3, Barquero MS, MD, PhD 1, Jimenez-Escrig A, MD, PhD 4.
Inheritance Of Alzheimer s Disease Investigated By Complex Segregation Analysis Manzano S, MD, PhD 1, Baron M, MD, PhD 2, Gomez-Tortosa E, MD, PhD 3, Barquero MS, MD, PhD 1, Jimenez-Escrig A, MD, PhD 4.
The Genetic Epidemiology of Cancer: Interpreting Family and Twin Studies and Their Implications for Molecular Genetic Approaches
 Vol. 10, 733 741, July 2001 Cancer Epidemiology, Biomarkers & Prevention 733 Review The Genetic Epidemiology of Cancer: Interpreting Family and Twin Studies and Their Implications for Molecular Genetic
Vol. 10, 733 741, July 2001 Cancer Epidemiology, Biomarkers & Prevention 733 Review The Genetic Epidemiology of Cancer: Interpreting Family and Twin Studies and Their Implications for Molecular Genetic
Introduction to Quantitative Genetics
 Introduction to Quantitative Genetics 1 / 17 Historical Background Quantitative genetics is the study of continuous or quantitative traits and their underlying mechanisms. The main principals of quantitative
Introduction to Quantitative Genetics 1 / 17 Historical Background Quantitative genetics is the study of continuous or quantitative traits and their underlying mechanisms. The main principals of quantitative
Tutorial on Genome-Wide Association Studies
 Tutorial on Genome-Wide Association Studies Assistant Professor Institute for Computational Biology Department of Epidemiology and Biostatistics Case Western Reserve University Acknowledgements Dana Crawford
Tutorial on Genome-Wide Association Studies Assistant Professor Institute for Computational Biology Department of Epidemiology and Biostatistics Case Western Reserve University Acknowledgements Dana Crawford
Pedigree Analysis Why do Pedigrees? Goals of Pedigree Analysis Basic Symbols More Symbols Y-Linked Inheritance
 Pedigree Analysis Why do Pedigrees? Punnett squares and chi-square tests work well for organisms that have large numbers of offspring and controlled mating, but humans are quite different: Small families.
Pedigree Analysis Why do Pedigrees? Punnett squares and chi-square tests work well for organisms that have large numbers of offspring and controlled mating, but humans are quite different: Small families.
Single Gene (Monogenic) Disorders. Mendelian Inheritance: Definitions. Mendelian Inheritance: Definitions
 Disorders. Mendelian Inheritance: Definitions. Mendelian Inheritance: Definitions") Single Gene (Monogenic) Disorders Mendelian Inheritance: Definitions A genetic locus is a specific position or location on a chromosome. Frequently, locus is used to refer to a specific gene. Alleles are
Single Gene (Monogenic) Disorders Mendelian Inheritance: Definitions A genetic locus is a specific position or location on a chromosome. Frequently, locus is used to refer to a specific gene. Alleles are
Non-Mendelian inheritance
 Non-Mendelian inheritance Focus on Human Disorders Peter K. Rogan, Ph.D. Laboratory of Human Molecular Genetics Children s Mercy Hospital Schools of Medicine & Computer Science and Engineering University
Non-Mendelian inheritance Focus on Human Disorders Peter K. Rogan, Ph.D. Laboratory of Human Molecular Genetics Children s Mercy Hospital Schools of Medicine & Computer Science and Engineering University
Summary. Introduction. Atypical and Duplicated Samples. Atypical Samples. Noah A. Rosenberg
 doi: 10.1111/j.1469-1809.2006.00285.x Standardized Subsets of the HGDP-CEPH Human Genome Diversity Cell Line Panel, Accounting for Atypical and Duplicated Samples and Pairs of Close Relatives Noah A. Rosenberg
doi: 10.1111/j.1469-1809.2006.00285.x Standardized Subsets of the HGDP-CEPH Human Genome Diversity Cell Line Panel, Accounting for Atypical and Duplicated Samples and Pairs of Close Relatives Noah A. Rosenberg
Genome-wide association studies (case/control and family-based) Heather J. Cordell, Institute of Genetic Medicine Newcastle University, UK
 Heather J. Cordell, Institute of Genetic Medicine Newcastle University, UK") Genome-wide association studies (case/control and family-based) Heather J. Cordell, Institute of Genetic Medicine Newcastle University, UK GWAS For the last 8 years, genome-wide association studies (GWAS)
Genome-wide association studies (case/control and family-based) Heather J. Cordell, Institute of Genetic Medicine Newcastle University, UK GWAS For the last 8 years, genome-wide association studies (GWAS)
Using Imputed Genotypes for Relative Risk Estimation in Case-Parent Studies
 American Journal of Epidemiology Published by Oxford University Press on behalf of the Johns Hopkins Bloomberg School of Public Health 011. Vol. 173, No. 5 DOI: 10.1093/aje/kwq363 Advance Access publication:
American Journal of Epidemiology Published by Oxford University Press on behalf of the Johns Hopkins Bloomberg School of Public Health 011. Vol. 173, No. 5 DOI: 10.1093/aje/kwq363 Advance Access publication:
Imaging Genetics: Heritability, Linkage & Association
 Imaging Genetics: Heritability, Linkage & Association David C. Glahn, PhD Olin Neuropsychiatry Research Center & Department of Psychiatry, Yale University July 17, 2011 Memory Activation & APOE ε4 Risk
Imaging Genetics: Heritability, Linkage & Association David C. Glahn, PhD Olin Neuropsychiatry Research Center & Department of Psychiatry, Yale University July 17, 2011 Memory Activation & APOE ε4 Risk
Statistical Tests for X Chromosome Association Study. with Simulations. Jian Wang July 10, 2012
 Statistical Tests for X Chromosome Association Study with Simulations Jian Wang July 10, 2012 Statistical Tests Zheng G, et al. 2007. Testing association for markers on the X chromosome. Genetic Epidemiology
Statistical Tests for X Chromosome Association Study with Simulations Jian Wang July 10, 2012 Statistical Tests Zheng G, et al. 2007. Testing association for markers on the X chromosome. Genetic Epidemiology
Decomposition of the Genotypic Value
 Decomposition of the Genotypic Value 1 / 17 Partitioning of Phenotypic Values We introduced the general model of Y = G + E in the first lecture, where Y is the phenotypic value, G is the genotypic value,
Decomposition of the Genotypic Value 1 / 17 Partitioning of Phenotypic Values We introduced the general model of Y = G + E in the first lecture, where Y is the phenotypic value, G is the genotypic value,
Lab Activity 36. Principles of Heredity. Portland Community College BI 233
 Lab Activity 36 Principles of Heredity Portland Community College BI 233 Terminology of Chromosomes Homologous chromosomes: A pair, of which you get one from mom, and one from dad. Example: the pair of
Lab Activity 36 Principles of Heredity Portland Community College BI 233 Terminology of Chromosomes Homologous chromosomes: A pair, of which you get one from mom, and one from dad. Example: the pair of
Non-parametric methods for linkage analysis
 BIOSTT516 Statistical Methods in Genetic Epidemiology utumn 005 Non-parametric methods for linkage analysis To this point, we have discussed model-based linkage analyses. These require one to specify a
BIOSTT516 Statistical Methods in Genetic Epidemiology utumn 005 Non-parametric methods for linkage analysis To this point, we have discussed model-based linkage analyses. These require one to specify a
(b) What is the allele frequency of the b allele in the new merged population on the island?
 What is the allele frequency of the b allele in the new merged population on the island?") 2005 7.03 Problem Set 6 KEY Due before 5 PM on WEDNESDAY, November 23, 2005. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. Two populations (Population One
2005 7.03 Problem Set 6 KEY Due before 5 PM on WEDNESDAY, November 23, 2005. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. Two populations (Population One
Genome-wide Association Analysis Applied to Asthma-Susceptibility Gene. McCaw, Z., Wu, W., Hsiao, S., McKhann, A., Tracy, S.
 Genome-wide Association Analysis Applied to Asthma-Susceptibility Gene McCaw, Z., Wu, W., Hsiao, S., McKhann, A., Tracy, S. December 17, 2014 1 Introduction Asthma is a chronic respiratory disease affecting
Genome-wide Association Analysis Applied to Asthma-Susceptibility Gene McCaw, Z., Wu, W., Hsiao, S., McKhann, A., Tracy, S. December 17, 2014 1 Introduction Asthma is a chronic respiratory disease affecting
Evidence for linkage of nonsyndromic cleft lip with or without cleft palate to a region on chromosome 2
 (2003) 11, 835 839 & 2003 Nature Publishing Group All rights reserved 1018-4813/03 $25.00 www.nature.com/ejhg ARTICLE Evidence for linkage of nonsyndromic cleft lip with or without cleft palate to a region
(2003) 11, 835 839 & 2003 Nature Publishing Group All rights reserved 1018-4813/03 $25.00 www.nature.com/ejhg ARTICLE Evidence for linkage of nonsyndromic cleft lip with or without cleft palate to a region
Systems of Mating: Systems of Mating:
 8/29/2 Systems of Mating: the rules by which pairs of gametes are chosen from the local gene pool to be united in a zygote with respect to a particular locus or genetic system. Systems of Mating: A deme
8/29/2 Systems of Mating: the rules by which pairs of gametes are chosen from the local gene pool to be united in a zygote with respect to a particular locus or genetic system. Systems of Mating: A deme
For more information about how to cite these materials visit
 Author(s): Kerby Shedden, Ph.D., 2010 License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution Share Alike 3.0 License: http://creativecommons.org/licenses/by-sa/3.0/
Author(s): Kerby Shedden, Ph.D., 2010 License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution Share Alike 3.0 License: http://creativecommons.org/licenses/by-sa/3.0/
Lecture 13: May 24, 2004
 Lecture 13: May 24, 2004 CH14: Mendel and the gene idea *particulate inheritance parents pass on discrete heritable units *gene- unit of inheritance which occupies a specific chromosomal location (locus)
Lecture 13: May 24, 2004 CH14: Mendel and the gene idea *particulate inheritance parents pass on discrete heritable units *gene- unit of inheritance which occupies a specific chromosomal location (locus)
Parental Effects. Genomic Imprinting
 Parental Effects A parental effect refers to a situation where, conditional on the individual s own genotype, the phenotype of an individual depends upon the mother s or father s phenotype or genotype.
Parental Effects A parental effect refers to a situation where, conditional on the individual s own genotype, the phenotype of an individual depends upon the mother s or father s phenotype or genotype.
Review and Evaluation of Methods Correcting for Population Stratification with a Focus on Underlying Statistical Principles
 Original Paper DOI: 10.1159/000119107 Published online: March 31, 2008 Review and Evaluation of Methods Correcting for Population Stratification with a Focus on Underlying Statistical Principles Hemant
Original Paper DOI: 10.1159/000119107 Published online: March 31, 2008 Review and Evaluation of Methods Correcting for Population Stratification with a Focus on Underlying Statistical Principles Hemant
Analysis of single gene effects 1. Quantitative analysis of single gene effects. Gregory Carey, Barbara J. Bowers, Jeanne M.
 Analysis of single gene effects 1 Quantitative analysis of single gene effects Gregory Carey, Barbara J. Bowers, Jeanne M. Wehner From the Department of Psychology (GC, JMW) and Institute for Behavioral
Analysis of single gene effects 1 Quantitative analysis of single gene effects Gregory Carey, Barbara J. Bowers, Jeanne M. Wehner From the Department of Psychology (GC, JMW) and Institute for Behavioral
GENETICS - NOTES-
 GENETICS - NOTES- Warm Up Exercise Using your previous knowledge of genetics, determine what maternal genotype would most likely yield offspring with such characteristics. Use the genotype that you came
GENETICS - NOTES- Warm Up Exercise Using your previous knowledge of genetics, determine what maternal genotype would most likely yield offspring with such characteristics. Use the genotype that you came
UNIT 1-History of life on earth! Big picture biodiversity-major lineages, Prokaryotes, Eukaryotes-Evolution of Meiosis
 Where are we in this course??? UNIT 1-History of life on earth! Big picture biodiversity-major lineages, Prokaryotes, Eukaryotes-Evolution of Meiosis Today we will start with UNIT 2 A. Mendel and the Gene
Where are we in this course??? UNIT 1-History of life on earth! Big picture biodiversity-major lineages, Prokaryotes, Eukaryotes-Evolution of Meiosis Today we will start with UNIT 2 A. Mendel and the Gene
Publications (* denote senior corresponding author)
") Publications (* denote senior corresponding author) 1. Sha Q, Zhang K, * Zhang SL (2016) A nonparametric regression approach to control for population stratification in rare variant association studies.
Publications (* denote senior corresponding author) 1. Sha Q, Zhang K, * Zhang SL (2016) A nonparametric regression approach to control for population stratification in rare variant association studies.
Biostatistics Faculty Publications
 University of Kentucky UKnowledge Biostatistics Faculty Publications Biostatistics 7-24-2009 On Quality Control Measures in Genome-Wide Association Studies: A Test to Assess the Genotyping Quality of Individual
University of Kentucky UKnowledge Biostatistics Faculty Publications Biostatistics 7-24-2009 On Quality Control Measures in Genome-Wide Association Studies: A Test to Assess the Genotyping Quality of Individual
IS IT GENETIC? How do genes, environment and chance interact to specify a complex trait such as intelligence?
 1 IS IT GENETIC? How do genes, environment and chance interact to specify a complex trait such as intelligence? Single-gene (monogenic) traits Phenotypic variation is typically discrete (often comparing
1 IS IT GENETIC? How do genes, environment and chance interact to specify a complex trait such as intelligence? Single-gene (monogenic) traits Phenotypic variation is typically discrete (often comparing
Power Calculation for Testing If Disease is Associated with Marker in a Case-Control Study Using the GeneticsDesign Package
 Power Calculation for Testing If Disease is Associated with Marker in a Case-Control Study Using the GeneticsDesign Package Weiliang Qiu email: weiliang.qiu@gmail.com Ross Lazarus email: ross.lazarus@channing.harvard.edu
Power Calculation for Testing If Disease is Associated with Marker in a Case-Control Study Using the GeneticsDesign Package Weiliang Qiu email: weiliang.qiu@gmail.com Ross Lazarus email: ross.lazarus@channing.harvard.edu
Statistical Evaluation of Sibling Relationship
 The Korean Communications in Statistics Vol. 14 No. 3, 2007, pp. 541 549 Statistical Evaluation of Sibling Relationship Jae Won Lee 1), Hye-Seung Lee 2), Hyo Jung Lee 3) and Juck-Joon Hwang 4) Abstract
The Korean Communications in Statistics Vol. 14 No. 3, 2007, pp. 541 549 Statistical Evaluation of Sibling Relationship Jae Won Lee 1), Hye-Seung Lee 2), Hyo Jung Lee 3) and Juck-Joon Hwang 4) Abstract
Evolution II.2 Answers.
 Evolution II.2 Answers. 1. (4 pts) Contrast the predictions of blending inheritance for F1 and F2 generations with those observed under Mendelian inheritance. Blending inheritance predicts both F1 and
Evolution II.2 Answers. 1. (4 pts) Contrast the predictions of blending inheritance for F1 and F2 generations with those observed under Mendelian inheritance. Blending inheritance predicts both F1 and
PopGen4: Assortative mating
 opgen4: Assortative mating Introduction Although random mating is the most important system of mating in many natural populations, non-random mating can also be an important mating system in some populations.
opgen4: Assortative mating Introduction Although random mating is the most important system of mating in many natural populations, non-random mating can also be an important mating system in some populations.
Mendelian Genetics. Activity. Part I: Introduction. Instructions
 Activity Part I: Introduction Some of your traits are inherited and cannot be changed, while others can be influenced by the environment around you. There has been ongoing research in the causes of cancer.
Activity Part I: Introduction Some of your traits are inherited and cannot be changed, while others can be influenced by the environment around you. There has been ongoing research in the causes of cancer.
Lecture 6 Practice of Linkage Analysis
 Lecture 6 Practice of Linkage Analysis Jurg Ott http://lab.rockefeller.edu/ott/ http://www.jurgott.org/pekingu/ The LINKAGE Programs http://www.jurgott.org/linkage/linkagepc.html Input: pedfile Fam ID
Lecture 6 Practice of Linkage Analysis Jurg Ott http://lab.rockefeller.edu/ott/ http://www.jurgott.org/pekingu/ The LINKAGE Programs http://www.jurgott.org/linkage/linkagepc.html Input: pedfile Fam ID
Chapter 4 PEDIGREE ANALYSIS IN HUMAN GENETICS
 Chapter 4 PEDIGREE ANALYSIS IN HUMAN GENETICS Chapter Summary In order to study the transmission of human genetic traits to the next generation, a different method of operation had to be adopted. Instead
Chapter 4 PEDIGREE ANALYSIS IN HUMAN GENETICS Chapter Summary In order to study the transmission of human genetic traits to the next generation, a different method of operation had to be adopted. Instead
Genetics Review. Alleles. The Punnett Square. Genotype and Phenotype. Codominance. Incomplete Dominance
 Genetics Review Alleles These two different versions of gene A create a condition known as heterozygous. Only the dominant allele (A) will be expressed. When both chromosomes have identical copies of the
Genetics Review Alleles These two different versions of gene A create a condition known as heterozygous. Only the dominant allele (A) will be expressed. When both chromosomes have identical copies of the
CS2220 Introduction to Computational Biology
 CS2220 Introduction to Computational Biology WEEK 8: GENOME-WIDE ASSOCIATION STUDIES (GWAS) 1 Dr. Mengling FENG Institute for Infocomm Research Massachusetts Institute of Technology mfeng@mit.edu PLANS
CS2220 Introduction to Computational Biology WEEK 8: GENOME-WIDE ASSOCIATION STUDIES (GWAS) 1 Dr. Mengling FENG Institute for Infocomm Research Massachusetts Institute of Technology mfeng@mit.edu PLANS
Quantitative Genetic Methods to Dissect Heterogeneity in Complex Traits
 Virginia Commonwealth University VCU Scholars Compass Theses and Dissertations Graduate School 2012 Quantitative Genetic Methods to Dissect Heterogeneity in Complex Traits T. Bernard Bigdeli Virginia Commonwealth
Virginia Commonwealth University VCU Scholars Compass Theses and Dissertations Graduate School 2012 Quantitative Genetic Methods to Dissect Heterogeneity in Complex Traits T. Bernard Bigdeli Virginia Commonwealth
C. Incorrect! Resemblance is not described by heritability. Heritability is a concept that is easily confused, so memorize the definition.
 Genetics - Problem Drill 09: Genetics and Environment No. 1 of 10 1. Heritability is. (A) The fraction of a population's phenotype, which is strictly due to genetic factors. (B) The fraction of a population's
Genetics - Problem Drill 09: Genetics and Environment No. 1 of 10 1. Heritability is. (A) The fraction of a population's phenotype, which is strictly due to genetic factors. (B) The fraction of a population's
Your DNA extractions! 10 kb
 Your DNA extractions! 10 kb Quantitative characters: polygenes and environment Most ecologically important quantitative traits (QTs) vary. Distributions are often unimodal and approximately normal. Offspring
Your DNA extractions! 10 kb Quantitative characters: polygenes and environment Most ecologically important quantitative traits (QTs) vary. Distributions are often unimodal and approximately normal. Offspring
During the hyperinsulinemic-euglycemic clamp [1], a priming dose of human insulin (Novolin,
![During the hyperinsulinemic-euglycemic clamp [1], a priming dose of human insulin (Novolin,](/thumbs/72/66792834.jpg "During the hyperinsulinemic-euglycemic clamp [1], a priming dose of human insulin (Novolin,") ESM Methods Hyperinsulinemic-euglycemic clamp procedure During the hyperinsulinemic-euglycemic clamp [1], a priming dose of human insulin (Novolin, Clayton, NC) was followed by a constant rate (60 mu m
ESM Methods Hyperinsulinemic-euglycemic clamp procedure During the hyperinsulinemic-euglycemic clamp [1], a priming dose of human insulin (Novolin, Clayton, NC) was followed by a constant rate (60 mu m
Some basics about mating schemes
 Some basics about mating schemes Ab F. Groen and Liesbeth Van der Waaij Animal Breeding and Genetics Group, Wageningen Institute of Animal Sciences, Wageningen University, P.O. Box 338, 6700 AH, Wageningen,
Some basics about mating schemes Ab F. Groen and Liesbeth Van der Waaij Animal Breeding and Genetics Group, Wageningen Institute of Animal Sciences, Wageningen University, P.O. Box 338, 6700 AH, Wageningen,
STATISTICAL ANALYSIS FOR GENETIC EPIDEMIOLOGY (S.A.G.E.) INTRODUCTION
 INTRODUCTION") STATISTICAL ANALYSIS FOR GENETIC EPIDEMIOLOGY (S.A.G.E.) INTRODUCTION Release 3.1 December 1997 - ii - INTRODUCTION Table of Contents 1 Changes Since Last Release... 1 2 Purpose... 3 3 Using the Programs...
STATISTICAL ANALYSIS FOR GENETIC EPIDEMIOLOGY (S.A.G.E.) INTRODUCTION Release 3.1 December 1997 - ii - INTRODUCTION Table of Contents 1 Changes Since Last Release... 1 2 Purpose... 3 3 Using the Programs...
A Comparison of Sample Size and Power in Case-Only Association Studies of Gene-Environment Interaction
 American Journal of Epidemiology ª The Author 2010. Published by Oxford University Press on behalf of the Johns Hopkins Bloomberg School of Public Health. This is an Open Access article distributed under
American Journal of Epidemiology ª The Author 2010. Published by Oxford University Press on behalf of the Johns Hopkins Bloomberg School of Public Health. This is an Open Access article distributed under
INTRODUCTION TO GENETIC EPIDEMIOLOGY (1012GENEP1) Prof. Dr. Dr. K. Van Steen
 Prof. Dr. Dr. K. Van Steen") INTRODUCTION TO GENETIC EPIDEMIOLOGY (1012GENEP1) Prof. Dr. Dr. K. Van Steen DIFFERENT FACES OF GENETIC EPIDEMIOLOGY 1 Basic epidemiology 1.a Aims of epidemiology 1.b Designs in epidemiology 1.c An overview
INTRODUCTION TO GENETIC EPIDEMIOLOGY (1012GENEP1) Prof. Dr. Dr. K. Van Steen DIFFERENT FACES OF GENETIC EPIDEMIOLOGY 1 Basic epidemiology 1.a Aims of epidemiology 1.b Designs in epidemiology 1.c An overview
Complex Traits Activity INSTRUCTION MANUAL. ANT 2110 Introduction to Physical Anthropology Professor Julie J. Lesnik
 Complex Traits Activity INSTRUCTION MANUAL ANT 2110 Introduction to Physical Anthropology Professor Julie J. Lesnik Introduction Human variation is complex. The simplest form of variation in a population
Complex Traits Activity INSTRUCTION MANUAL ANT 2110 Introduction to Physical Anthropology Professor Julie J. Lesnik Introduction Human variation is complex. The simplest form of variation in a population
Quantitative genetics: traits controlled by alleles at many loci
 Quantitative genetics: traits controlled by alleles at many loci Human phenotypic adaptations and diseases commonly involve the effects of many genes, each will small effect Quantitative genetics allows
Quantitative genetics: traits controlled by alleles at many loci Human phenotypic adaptations and diseases commonly involve the effects of many genes, each will small effect Quantitative genetics allows
GENETIC LINKAGE ANALYSIS
 Atlas of Genetics and Cytogenetics in Oncology and Haematology GENETIC LINKAGE ANALYSIS * I- Recombination fraction II- Definition of the "lod score" of a family III- Test for linkage IV- Estimation of
Atlas of Genetics and Cytogenetics in Oncology and Haematology GENETIC LINKAGE ANALYSIS * I- Recombination fraction II- Definition of the "lod score" of a family III- Test for linkage IV- Estimation of
Patterns of Parental Transmission and Familial Aggregation Models in Bipolar Affective Disorder
 American Journal of Medical Genetics (Neuropsychiatric Genetics) 81:397 404 (1998) Patterns of Parental Transmission and Familial Aggregation Models in Bipolar Affective Disorder Maria Grigoroiu-Serbanescu,
American Journal of Medical Genetics (Neuropsychiatric Genetics) 81:397 404 (1998) Patterns of Parental Transmission and Familial Aggregation Models in Bipolar Affective Disorder Maria Grigoroiu-Serbanescu,
Genetics of common disorders with complex inheritance Bettina Blaumeiser MD PhD
 Genetics of common disorders with complex inheritance Bettina Blaumeiser MD PhD Medical Genetics University Hospital & University of Antwerp Programme Day 6: Genetics of common disorders with complex inheritance
Genetics of common disorders with complex inheritance Bettina Blaumeiser MD PhD Medical Genetics University Hospital & University of Antwerp Programme Day 6: Genetics of common disorders with complex inheritance
IB BIO I Genetics Test Madden
 Name Date Multiple Choice 1. What does the genotype X H X h indicate? A. A co-dominant female B. A heterozygous male C. A heterozygous female D. A co-dominant male 2. A pure breeding tall plant with smooth
Name Date Multiple Choice 1. What does the genotype X H X h indicate? A. A co-dominant female B. A heterozygous male C. A heterozygous female D. A co-dominant male 2. A pure breeding tall plant with smooth
Research Article Power Estimation for Gene-Longevity Association Analysis Using Concordant Twins
 Genetics Research International, Article ID 154204, 8 pages http://dx.doi.org/10.1155/2014/154204 Research Article Power Estimation for Gene-Longevity Association Analysis Using Concordant Twins Qihua
Genetics Research International, Article ID 154204, 8 pages http://dx.doi.org/10.1155/2014/154204 Research Article Power Estimation for Gene-Longevity Association Analysis Using Concordant Twins Qihua
Mapping quantitative trait loci in humans: achievements and limitations
 Review series Mapping quantitative trait loci in humans: achievements and limitations Partha P. Majumder and Saurabh Ghosh Human Genetics Unit, Indian Statistical Institute, Kolkata, India Recent advances
Review series Mapping quantitative trait loci in humans: achievements and limitations Partha P. Majumder and Saurabh Ghosh Human Genetics Unit, Indian Statistical Institute, Kolkata, India Recent advances
The Association Design and a Continuous Phenotype
 PSYC 5102: Association Design & Continuous Phenotypes (4/4/07) 1 The Association Design and a Continuous Phenotype The purpose of this note is to demonstrate how to perform a population-based association
PSYC 5102: Association Design & Continuous Phenotypes (4/4/07) 1 The Association Design and a Continuous Phenotype The purpose of this note is to demonstrate how to perform a population-based association
A test of quantitative genetic theory using Drosophila effects of inbreeding and rate of inbreeding on heritabilities and variance components #
 Theatre Presentation in the Commision on Animal Genetics G2.7, EAAP 2005 Uppsala A test of quantitative genetic theory using Drosophila effects of inbreeding and rate of inbreeding on heritabilities and
Theatre Presentation in the Commision on Animal Genetics G2.7, EAAP 2005 Uppsala A test of quantitative genetic theory using Drosophila effects of inbreeding and rate of inbreeding on heritabilities and
Mx Scripts Library: Structural Equation Modeling Scripts for Twin and Family Data
 Behavior Genetics, Vol. 35, No. 4, July 2005 (Ó 2005) DOI: 10.1007/s10519-005-2791-5 Mx Scripts Library: Structural Equation Modeling Scripts for Twin and Family Data D Posthuma 1 and D. I. Boomsma 1 Received
Behavior Genetics, Vol. 35, No. 4, July 2005 (Ó 2005) DOI: 10.1007/s10519-005-2791-5 Mx Scripts Library: Structural Equation Modeling Scripts for Twin and Family Data D Posthuma 1 and D. I. Boomsma 1 Received
INTRODUCTION TO GENETIC EPIDEMIOLOGY (EPID0754) Prof. Dr. Dr. K. Van Steen
 Prof. Dr. Dr. K. Van Steen") INTRODUCTION TO GENETIC EPIDEMIOLOGY (EPID0754) Prof. Dr. Dr. K. Van Steen DIFFERENT FACES OF GENETIC EPIDEMIOLOGY 1 Basic epidemiology 1.a Aims of epidemiology 1.b Designs in epidemiology 1.c An overview
INTRODUCTION TO GENETIC EPIDEMIOLOGY (EPID0754) Prof. Dr. Dr. K. Van Steen DIFFERENT FACES OF GENETIC EPIDEMIOLOGY 1 Basic epidemiology 1.a Aims of epidemiology 1.b Designs in epidemiology 1.c An overview
Role of Genomics in Selection of Beef Cattle for Healthfulness Characteristics
 Role of Genomics in Selection of Beef Cattle for Healthfulness Characteristics Dorian Garrick dorian@iastate.edu Iowa State University & National Beef Cattle Evaluation Consortium Selection and Prediction
Role of Genomics in Selection of Beef Cattle for Healthfulness Characteristics Dorian Garrick dorian@iastate.edu Iowa State University & National Beef Cattle Evaluation Consortium Selection and Prediction
Pre-AP Biology Unit 7 Genetics Review Outline
 Unit 7 Genetics Review Outline Pre-AP Biology 2017-2018 LT 1 - I can explain the relationships among alleles, genes, chromosomes, genotypes, and phenotypes. This target covers application of the vocabulary
Unit 7 Genetics Review Outline Pre-AP Biology 2017-2018 LT 1 - I can explain the relationships among alleles, genes, chromosomes, genotypes, and phenotypes. This target covers application of the vocabulary
Lecture 17: Human Genetics. I. Types of Genetic Disorders. A. Single gene disorders
 Lecture 17: Human Genetics I. Types of Genetic Disorders A. Single gene disorders B. Multifactorial traits 1. Mutant alleles at several loci acting in concert C. Chromosomal abnormalities 1. Physical changes
Lecture 17: Human Genetics I. Types of Genetic Disorders A. Single gene disorders B. Multifactorial traits 1. Mutant alleles at several loci acting in concert C. Chromosomal abnormalities 1. Physical changes
PRINCIPLE OF INHERITANCE AND
 29 CHAPTER 5 PRINCIPLE OF INHERITANCE AND VARIATION MULTIPLE-CHOICE QUESTIONS 1. All genes located on the same chromosome: a. Form different groups depending upon their relative distance b. Form one linkage
29 CHAPTER 5 PRINCIPLE OF INHERITANCE AND VARIATION MULTIPLE-CHOICE QUESTIONS 1. All genes located on the same chromosome: a. Form different groups depending upon their relative distance b. Form one linkage
IN A GENOMEWIDE scan in the Irish Affected Sib
 ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH Vol. 30, No. 12 December 2006 A Joint Genomewide Linkage Analysis of Symptoms of Alcohol Dependence and Conduct Disorder Kenneth S. Kendler, Po-Hsiu Kuo,
ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH Vol. 30, No. 12 December 2006 A Joint Genomewide Linkage Analysis of Symptoms of Alcohol Dependence and Conduct Disorder Kenneth S. Kendler, Po-Hsiu Kuo,
Regression-based Linkage Analysis Methods
 Chapter 1 Regression-based Linkage Analysis Methods Tao Wang and Robert C. Elston Department of Epidemiology and Biostatistics Case Western Reserve University, Wolstein Research Building 2103 Cornell Road,
Chapter 1 Regression-based Linkage Analysis Methods Tao Wang and Robert C. Elston Department of Epidemiology and Biostatistics Case Western Reserve University, Wolstein Research Building 2103 Cornell Road,
Multifactorial Inheritance. Prof. Dr. Nedime Serakinci
 Multifactorial Inheritance Prof. Dr. Nedime Serakinci GENETICS I. Importance of genetics. Genetic terminology. I. Mendelian Genetics, Mendel s Laws (Law of Segregation, Law of Independent Assortment).
Multifactorial Inheritance Prof. Dr. Nedime Serakinci GENETICS I. Importance of genetics. Genetic terminology. I. Mendelian Genetics, Mendel s Laws (Law of Segregation, Law of Independent Assortment).
Supplementary Figures
 Supplementary Figures Supplementary Fig 1. Comparison of sub-samples on the first two principal components of genetic variation. TheBritishsampleisplottedwithredpoints.The sub-samples of the diverse sample
Supplementary Figures Supplementary Fig 1. Comparison of sub-samples on the first two principal components of genetic variation. TheBritishsampleisplottedwithredpoints.The sub-samples of the diverse sample
The plant of the day Pinus longaeva Pinus aristata
 The plant of the day Pinus longaeva Pinus aristata Today s Topics Non-random mating Genetic drift Population structure Big Questions What are the causes and evolutionary consequences of non-random mating?
The plant of the day Pinus longaeva Pinus aristata Today s Topics Non-random mating Genetic drift Population structure Big Questions What are the causes and evolutionary consequences of non-random mating?
Independent genome-wide scans identify a chromosome 18 quantitative-trait locus influencing dyslexia theoret read
 Independent genome-wide scans identify a chromosome 18 quantitative-trait locus influencing dyslexia Simon E. Fisher 1 *, Clyde Francks 1 *, Angela J. Marlow 1, I. Laurence MacPhie 1, Dianne F. Newbury
Independent genome-wide scans identify a chromosome 18 quantitative-trait locus influencing dyslexia Simon E. Fisher 1 *, Clyde Francks 1 *, Angela J. Marlow 1, I. Laurence MacPhie 1, Dianne F. Newbury
Chapter 7: Pedigree Analysis B I O L O G Y
 Name Date Period Chapter 7: Pedigree Analysis B I O L O G Y Introduction: A pedigree is a diagram of family relationships that uses symbols to represent people and lines to represent genetic relationships.
Name Date Period Chapter 7: Pedigree Analysis B I O L O G Y Introduction: A pedigree is a diagram of family relationships that uses symbols to represent people and lines to represent genetic relationships.
LTA Analysis of HapMap Genotype Data
 LTA Analysis of HapMap Genotype Data Introduction. This supplement to Global variation in copy number in the human genome, by Redon et al., describes the details of the LTA analysis used to screen HapMap
LTA Analysis of HapMap Genotype Data Introduction. This supplement to Global variation in copy number in the human genome, by Redon et al., describes the details of the LTA analysis used to screen HapMap
Activity 15.2 Solving Problems When the Genetics Are Unknown
 f. Blue-eyed, color-blind females 1 2 0 0 g. What is the probability that any of the males will be color-blind? 1 2 (Note: This question asks only about the males, not about all of the offspring. If we
f. Blue-eyed, color-blind females 1 2 0 0 g. What is the probability that any of the males will be color-blind? 1 2 (Note: This question asks only about the males, not about all of the offspring. If we