Molecular mechanism of cell-autonomous circadian gene. expression of Period2, a crucial regulator of the mammalian
|
|
|
- Virgil Russell
- 5 years ago
- Views:
Transcription
1 Molecular mechanism of cell-autonomous circadian gene expression of Period2, a crucial regulator of the mammalian circadian clock Makoto Akashi*, Tomoko Ichise*, Takayoshi Mamine and Toru Takumi* *Osaka Bioscience Institute, Suita, Osaka , Japan Life Science Laboratory, Material Laboratories, Sony Corporation, Shinagawa, Tokyo , Japan Corresponding author: Toru Takumi ( takumi@obi.or.jp) Osaka Bioscience Institute (OBI), Furuedai, Suita, Osaka , Japan Tel: Fax: A running title: Molecular mechanism of Per2 oscillation Key words: circadian rhythm, cell-autonomous oscillation, transcription, period2, E-box
2 Abstract Although circadian transcription of Period2 (Per2) is fundamental for the generation of circadian rhythm, the molecular mechanism remains unclear. Here we report that cell-autonomous circadian transcription of Per2 is driven by two transcriptional elements, one for rhythm generation and the other for phase control. The former contains the E-box-like sequence (CACGTT) that is sufficient and indispensable to drive oscillation, and indeed circadian transcription factors site-specifically bind to it. Furthermore, the nature of this atypical E-box is different from that of the classical circadian E-box. The current feedback loop model is based mainly on Period1. Our results provide not only compelling evidence in support of this model but also an explanation for a general basic mechanism to produce various patterns in the phase and amplitude of cell-autonomous circadian gene expression. 2
3 INTRODUCTION In nearly all organisms, behavioral and physiological processes display approximately 24 hr rhythms that are controlled by circadian pacemakers (Pittendrigh, 1993). The circadian organization of physiology and behavior in mammals is governed by the suprachiasmatic nuclei (SCN), a defined pair of cell clusters in the anteroventral hypothalamus (Ralph et al., 1990). Circadian clocks can count time only approximately and must be adjusted every day by the photoperiod in order to be in harmony with the outside world (Menaker, 2003). Circadian oscillators also exist in most peripheral cells and even in cultured cells (Balsalobre et al., 1998; Yamazaki et al., 2000). It is thought that the phase of these peripheral timekeepers is reset by signals regulated by the SCN pacemaker (Akashi and Nishida, 2000; Schibler and Sassone-Corsi, 2002). The molecular makeup of circadian clocks has been the subject of intense genetic and biochemical investigation in various organisms, including cyanobacteria, Neurospora, higher plants, Drosophila, and mammals (Dunlap, 1999; Kondo and Ishiura, 2000; Allada et al., 2001; Williams and Sehgal, 2001; Young and Kay, 2001; Reppert and Weaver, 2002). Over the last several years, orthologs of most Drosophila circadian clock genes have been cloned from mammals (Albrecht and Eichele, 2003; Lowrey and Takahashi, 2004). Although mper2 was literally cloned as a secondary mammalian 3
4 period gene (Albrecht et al., 1997; Takumi et al., 1998), gene-knockout analysis revealed that an mper2 mutant displays a loss of circadian rhythmicity, revealing a prominent role for mper2 in the mammalian clock (Zheng et al., 1999). Additionally familial advanced sleep phase syndrome has been attributed to a missense mutation in hper2 (Toh et al., 2001). These studies demonstrate that a robust circadian fluctuation in Per2 transcription is an essential event for the generation of circadian rhythm. Circadian oscillators appear to have been highly conserved throughout evolution and to involve transcription translation negative feedback loops for the regulation of clock genes (Dunlap, 1999; Young and Kay, 2001). In mammals, in vitro studies have shown that the expression of Per1 (Period1) is driven by the CLOCK/BMAL1 transcription complex through an E-box enhancer and that PER proteins, together with CRY (Cryptochrome) proteins, serve to regulate the CLOCK/BMAL1 transcription complex negatively (Gekakis et al., 1998; Kume et al., 1999; Hida et al., 2000). This model has been thought to be applicable to other Per and Cry genes. However, several problems remain to be solved in the current model. First, there is a wide variation in the phase and amplitude of circadian accumulation of Per1, Per2, Per3, Cry1, and Cry2 mrna levels in tissues and cultured cells (Yamamoto et al., 2004). The current feedback loop model in mammals cannot explain the mechanism to 4
5 generate this variation. It is importantly to note that the phase and amplitude of circadian transcription cannot be verified by using an overexpression-based transient reporter assay. Second, although Per1, which has been analyzed in detail, serves as a foundation for the mammalian model, it has been reported recently that Per1 is not absolutely required for the generation and maintenance of circadian rhythms (Cermakian et al., 2001; Zheng et al., 2001). This issue remains controversial, therefore, since Bae et al. found that the loss of Per1 can result in arrhythmic mice (Bae et al., 2001). Thus, it is unclear whether the current model really reflects the core clock mechanism. Gene knockout studies of clock genes support the current model; for example, genetic deficiency of Bmal1 results in the down-regulation of Per expression (Vitaterna et al., 1999; Bunger et al., 2000; Bae et al., 2001). However, these studies cannot prove whether the regulation is direct or indirect. Overexpression-based transient reporter assay has shown that transcription of the Per2 gene, a key component for rhythm generation, is also upregulated by coexpression of circadian transcription factors (BMAL1, CLOCK and NPAS2), as shown for the Per1 gene (Travnickova-Bendova et al., 2002; Kaasik and Lee, 2004). Yet, while Per1 contains 5 classical circadian E-boxes (Hida et al., 2000; Yamamoto et al., 2004), no evolutionarily conserved E-box (the classical circadian E-box) has been identified in the upstream sequence of Per2. 5
6 Therefore it remains unknown whether these circadian transcription factors directly activate the Per2 gene in a site-specific manner. Herein we report the detailed analysis of the mechanism of a robust cell-autonomous circadian fluctuation in Per2 transcription, not by transient reporter assays but by monitoring transcriptional fluctuation of the luciferase reporter gene over several days. An approximately 20-bp region located near the transcription start site (TSS) was indispensable to drive cell-autonomous rhythmic transcription of Per2, whereas another region, located upstream from it, was shown to be responsible for phase control of cell-autonomous circadian transcription. An E-box-like sequence exists in the core region (the former region), and indeed circadian transcription factors activated Per2 transcription through site-specific binding to this element. The fact that this identified small region, indispensable for cell-autonomous rhythmic transcription of Per2, contains a functional E-box-like sequence is of great significance. Our results thus validate the current model by demonstrating that this model is actually applicable to cell-autonomous circadian fluctuation of Per2, a core component for the generation of circadian rhythm. We also show that this atypical E-box has properties different from those of the classical circadian E-box, which result in a pattern of circadian transcription different from that of Per1. Our results provide not only compelling evidence in support of the current 6
7 feedback loop model in mammals but also an explanation for a general mechanism that generates wide variation in the expression patterns of circadian genes. 7
8 MATERIALS AND METHODS Plasmid construction A bacterial artificial chromosome (BAC) clone containing the complete genomic sequence of the mouse Per2 (mper2) gene was purchased from the BACPAC Resource Center (BPRC) at Children's Hospital Oakland Research Institute. The mper2 promoter region was isolated and cloned in the pgl3-basic vector (Promega). The mper2 region spans from to +110 (+1 is the putative TSS). Cell culture, transfection and reporter assay NIH3T3 cells were cultured and transfected as described previously (Akashi et al., 2002). For real-time PCR analyses, cells were immediately frozen in liquid nitrogen and stored at 80 o C until processed for RNA. Cell lysates were used in the Dual Luciferase assay System (Promega) as described previously (Akashi and Takumi, 2005) Real-time monitoring of luciferase activity in living cells NIH3T3 cells were cultured, transfected with mper2-luc, and incubated for 24 hours. Then the medium was exchanged for serum-rich medium (DMEM, supplemented with 50% serum). Two hours later this medium was replaced with normal culture medium. In the presence of 0.1 mm luciferin, light emission was measured and integrated for 1 min at intervals of 15 min with a photomulti-plier tube (Hamamatsu Photonics) as described 8
9 previously (Akashi and Takumi, 2005). Data analysis Phase and period measurements were calculated as in previous studies (Abe et al., 2002; Yamazaki et al., 2002; Yoo et al., 2004). Data sets were detrended by subtracting the 24 hr running average from the raw data. The maximum differences between the smoothed curves for each cycle (the peak and the trough) were used to calculate the amplitude of each cycle. Animals Mice were housed under a strict 12:12 hour light/dark condition. Tissues were immediately frozen in liquid nitrogen and stored at 80 o C until processed for RNA. All protocols of experiments using animals in this study were approved by the OBI (Osaka Bioscience Institute) Animal Research Committee. Pull-down experiment Mouse liver extracts were prepared at 4-hr intervals by homogenizing the tissue in ice-cold incubation buffer (Akashi et al., 2002), and then the extracts were incubated with an 80-bp double-stranded biotinylated oligonucleotide that had been immobilized on streptavidin-sepharose beads (Amersham). After having been washed with the incubation buffer, the resulting bound protein was subjected to immunoblot analysis. The 9
10 designed sequences (response elements underlined) were the following: wild-type (mper2, -105~+15): 5 CTCAGGTTCCGCCCCGCCAGTATGCAAATGAGGTGGCACTCCGACCAATGG CGCGCGCAGGGGCGGGCTCAGCGCGCGCGGTCACGTTTTCCACTATGTGACA GCGGAGGGCGACGCGGC3 mutant (mper2, -105~+15): 5 CTCAGGTTCCGCCCCGCCAGTATGCAAATGAGGTGGCACTCCGACCAATGG CGCGCGCAGGGGCGGGCTCAGCGCGCGCGGTCTTTCCACTATGTGACAGCG GAGGGCGACGCGGC3 wild-type (mper1, 3E-box): 5 GAAAGCTTTAGCCACGTGACAGTGAGGGGCACCCCTTAACGACACGTGGG CCCTCAATTGAGCACCCAAGTCCACGTGCAGGGATGTGTGGGGGCAGGGCC TGGCATTATGCAACCCGCCTCCCAGCCTC3 mutant (mper1, 3E-box): 5 GAAAGCTTTAGCCGACAGTGAGGGGCACCCCTTAACGACGGGCCCTCAAT TGAGCACCCAAGTCCGCAGGGATGTGTGGGGGCAGGGCCTGGCATTAACCC GCCTCCCAGCCTC3 10
11 Real-time quantitative RT-PCR Quantification of relative RNA levels by the SYBER Green real-time PCR technology was done as described previously (Yamamoto et al., 2004). Briefly, DNase-treated total RNA (2.5 μg) was reverse-transcribed by using an oligo(dt) primer and Superscript reverse transcriptase (Invitrogen). The cdna equivalent to 20 ng total RNA was PCR-amplified in an ABI PRISM 7900 HT sequence detection system (Applied Biosystems). Forward primers and reverse primers were as follow: Gapdh forward: 5 -CATCCACTGGTGCTGCCAAGGCTGT-3 ; Gapdh reverse: 5 -ACAACCTGGTCCTCAGTGTAGCCCA-3 mper1 forward: 5 -CAGGCTAACCAGGAATATTACCAGC-3 ; mper1 reverse: 5 -CACAGCCACAGAGAAGGTGTCCTGG-3 mper2 forward: 5 -GGCTTCACCATGCCTGTTGT-3 ; mper2 reverse: 5 -GGAGTTATTTCGGAGGCAAGTGT-3 The relative levels of each RNA were normalized to the corresponding Gapdh RNA levels. Relative RNA levels were then expressed as percentage of the maximal value obtained for each experiment. 11
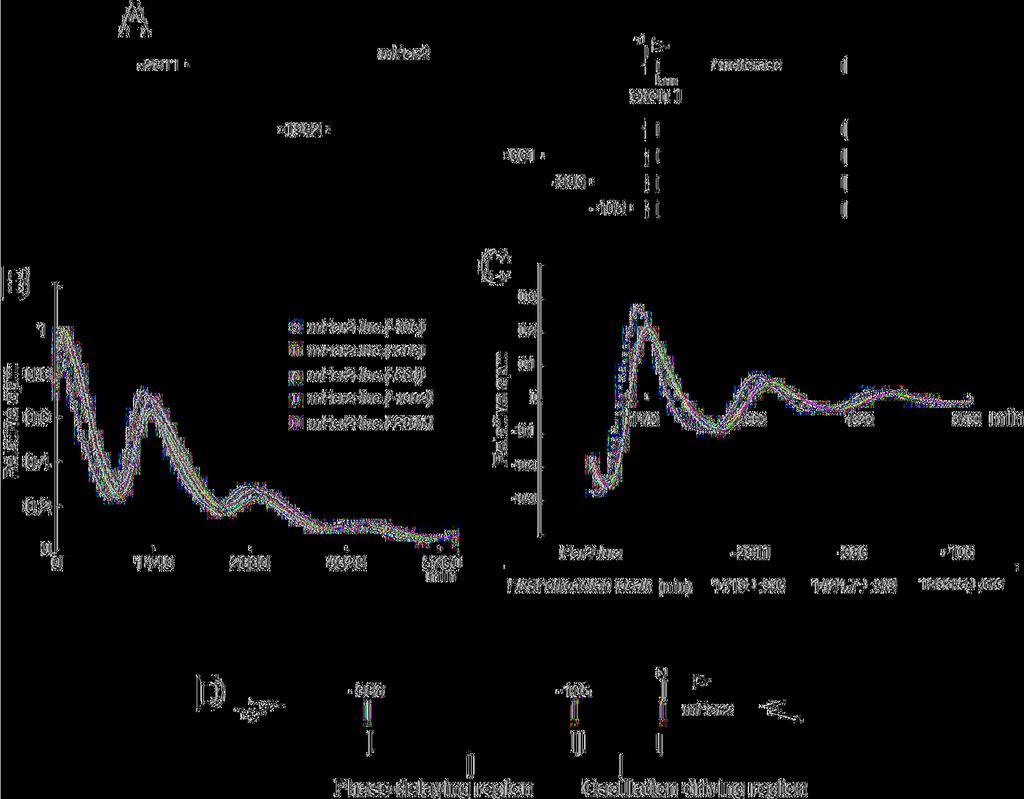
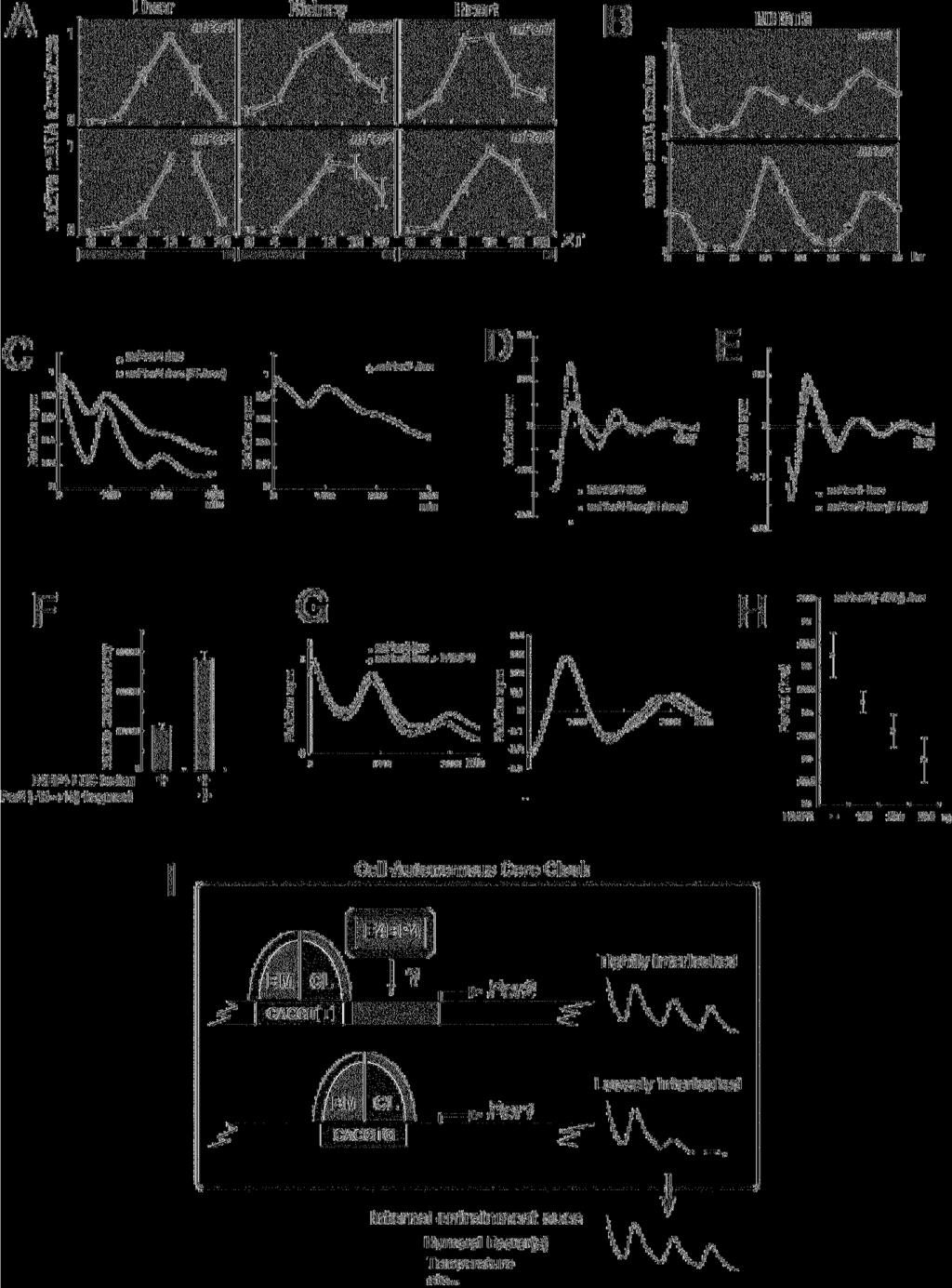
12 Results and Discussion Cell-autonomous circadian transcription of Per2 is regulated by two or more transcriptional regulatory elements The core clock is a cell-autonomous system. Therefore, we should examine whether transcriptional regulatory elements function under a cell-autonomous condition. Our approach enables us to exclude internal environmental cues such as blood-borne factors and body temperature. To identify the transcriptional regulatory elements for cell-autonomous circadian transcription of Per2, we monitored transcriptional fluctuation of Per2 in real-time by using a deletion series of Per2 promoter-reporter constructs (Fig. 1A). Note that exogenously transfected transgenes have no effect on endogenous cell-autonomous circadian oscillation and, therefore, that the deletion or mutation of constructs does not affect the endogenous pacemaker. NIH3T3 cells were transfected with the Per2-luc construct and then stimulated with a high concentration of serum. After the serum shock, in the presence of luciferin, light emission was measured and integrated for 1 min at intervals of 15 min. By using this in vitro luminescence reporter system, we can monitor cell-autonomous oscillators. Clearly, Per2-controlled fluctuations of luminescence resulted in a series of readily appreciable peaks and troughs, as exemplified in Figure 1. Almost the same phases and amplitudes were observed in the 12
13 cells transfected with the constructs from 2811 to -386, whereas a phase advance (approximately 2 hours earlier) was detected only when using the Per2 (-105) construct (Fig. 1B). To better calculate phase differences, data sets were detrended, and the time of the first peak was used as a phase marker (Fig. 1C). Again the Per2 (-105) construct showed a 2-h phase advance compared to the other constructs, as expected in Fig. 1B. Thus, these data suggest that a phase delaying element is located between -386 and -106 bp upstream from the TSS(s) and that a rhythm-generating element exists between -105 and +1 bp upstream from the TSS(s) (Fig. 1D). These results demonstrate that two or more transcriptional regulatory elements, i.e., a phase delaying element and a rhythm-generating element located upstream from the TSS(s), are required for cell-autonomous circadian gene expression of Per2. Consistent with this cell-autonomous phenomenon, Yoo et al. has very recently reported that a 210-bp fragment upstream from the TSS(s) drives Per2 circadian oscillation in vivo (Yoo et al., 2005). An approximately 20-bp region located near the TSS(s) was indispensable to drive cell-autonomous rhythmic transcription of Per2 To identify transcription factors required to drive cell-autonomous rhythmic transcription of Per2, we exactly narrowed the rhythm-generating region by monitoring in real-time 13
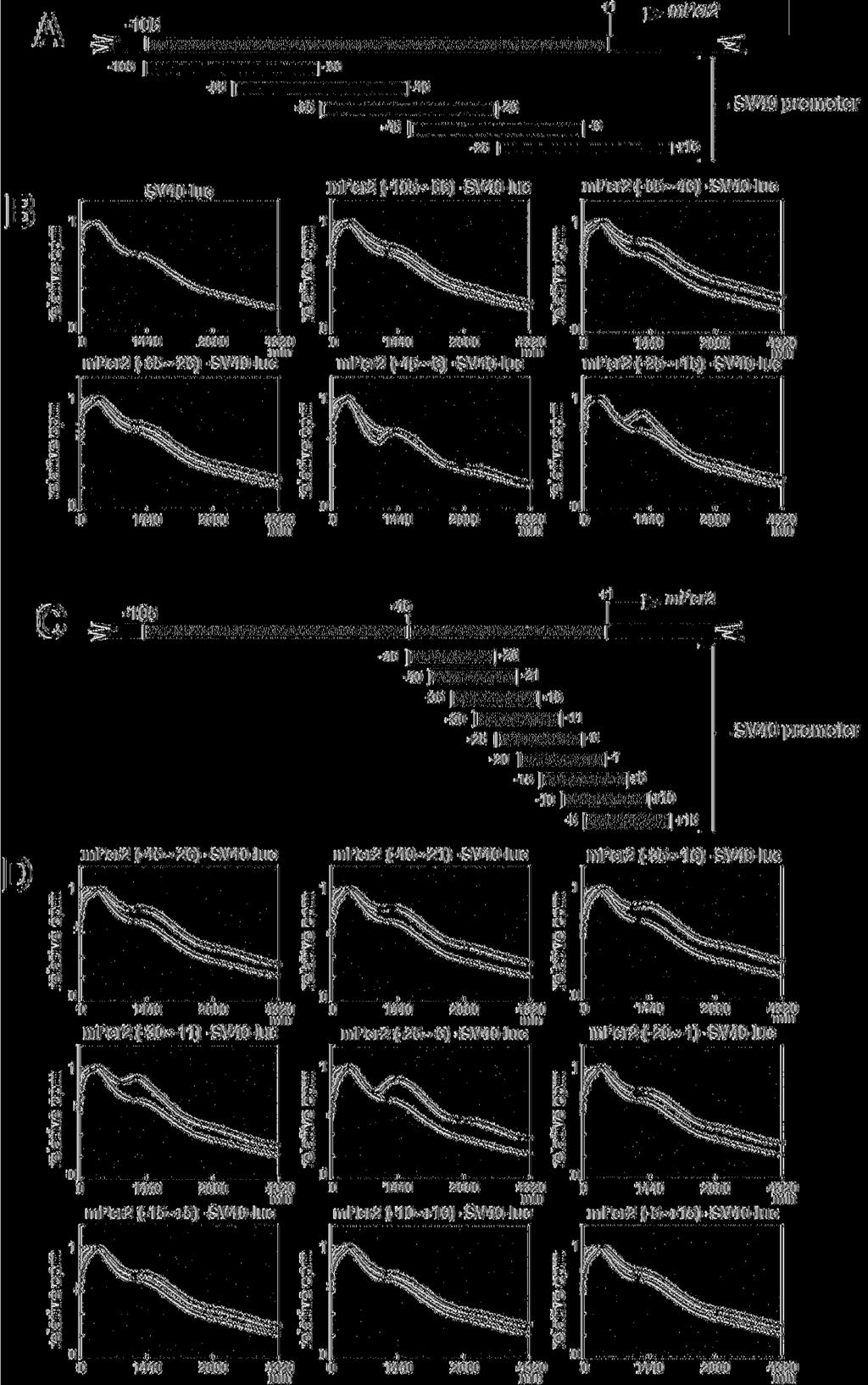
14 Per2-controlled fluctuations of luminescence in cells transfected with another set of constructs, in which a fragment from Per2 (-105) was inserted into the upstream of the SV40 promoter (Fig. 2A and 2C). When 40-bp fragments overlapping by 20 bp were used (Fig. 2A), the Per2 (-45~-6)-SV40-luc construct and the Per2 (-25~+15)-SV40-luc construct clearly showed Per2-controlled fluctuations of luminescence as compared with the other constructs (Fig. 2B). The detrended data suggests that the SV40 promoter activity exhibits not a drastic but significant circadian fluctuation (Supplemental Fig. 1A), indicating that this promoter contains some circadian enhancer elements. The detrended bioluminescence datasets make clear that only the Per2 (-45~-6)-SV40-luc and Per2 (-25~+15)-SV40-luc construct exhibit a higher amplitude of oscillation than the SV40 promoter. Next, to compare the magnitude of fluctuation in these two constructs, the amplitude in each cycle was calculated by subtracting the value of the trough from that of the peak (Supplemental Fig. 1B). The higher amplitude of oscillation was maintained over the cycles in Per2 (-45~-6)-SV40-luc. The Per2 (-45~-6)-SV40-luc/Per2 (-25~+15)-SV40-luc amplitude ratio was obtained by dividing the Per2 (-45~-6)-SV40-luc wave amplitude by the Per2 (-25~+15)-SV40-luc wave amplitude in each cycle (Supplemental Fig. 1C), demonstrating that the amplitude of Per2 (-45~-6)-SV40-luc gradually grew to be 2-fold higher than that of Per2 14
15 (-25~+15)-SV40-luc, as cycle number increases. Taken together, the region ranging from -45 to +15 was shown to possess the ability to drive circadian oscillation of transcription. Next, to further define this region, we used 20-bp fragments overlapping by 15 bp (Fig. 2C). The results show that the Per2 (-25~-6) fragment possessed the most potent ability to oscillate SV40 promoter activity (Fig. 2D). The detrended data highlighted the Per2 (-25~-6) and Per2 (-30~-11) fragments ability to drive oscillation (Supplemental Fig. 1D). The Per2 (-25~-6)-SV40-luc maintained a higher amplitude in each cycle than that of Per2 (-30~-11)-SV40-luc (Supplemental Fig. 1E). Additionally, the Per2 (-25~-6)-SV40-luc/Per2 (-30~-11)-SV40-luc amplitude ratio in each cycle demonstrated that the amplitude of Per2 (-25~-6)-SV40-luc was also considered, being 2-fold higher over cycles than that of Per2 (-30~-11)-SV40-luc (Supplemental Fig. 1F). The merge of Per2 (-25~-6)-SV40-luc (Supplemental Fig. 1D) and Per2 (-45~-6)-SV40-luc (Supplemental Fig. 1A) illustrated almost the same pattern of oscillation, confirming that the Per2 (-25~-6) region is necessary and sufficient to drive Per2 (-45~-6)-SV40-luc oscillation (Supplemental Fig. 1G). This region (-25~-6), therefore, contains the core element for rhythmic transcription of Per2. We also constructed mutants lacking 10 bp in the 105~+6 region, and monitored transcriptional fluctuation of Per2 in real-time (Fig. 3A). The amplitude of 15
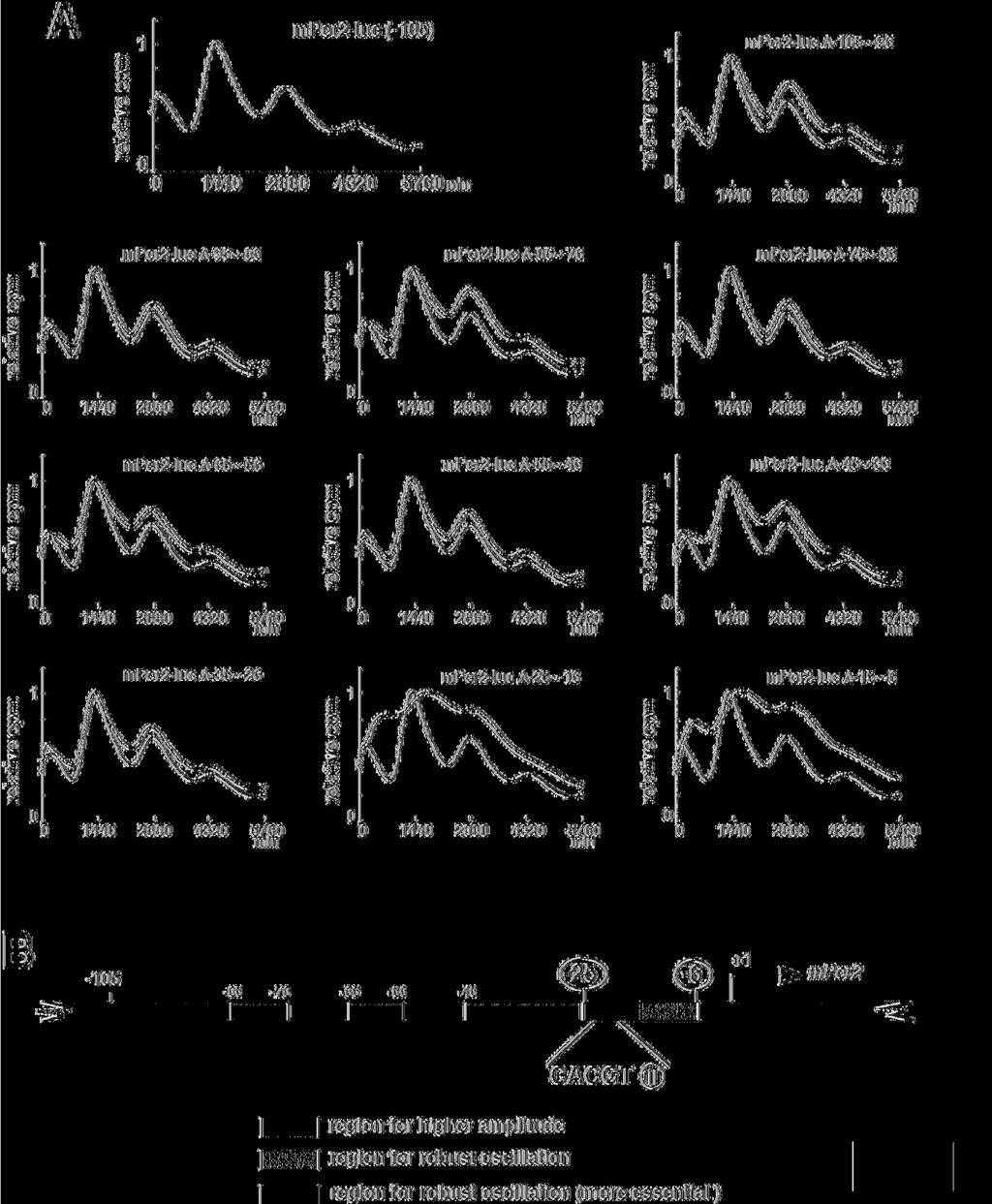
16 circadian transcription of Per2 was strongly diminished in mper2-luc Δ (-25~-16) and mper2-luc Δ (-15~-6). Furthermore, both of these two constructs exhibited a phase delay, illustrating that both 10-bp regions contribute to maintain the original phase. Other regions were required for amplification, whereas these 2 regions were essential for driving rhythmic transcription of Per2. After removal of baseline changes (Supplemental Fig. 2A), the period was obtained from regression analysis of a circadian marker (trough) in Supplemental Figure 2B. As compared to wild-type Per2-luc, mper2-luc Δ (-95~-86), mper2-luc Δ (-85~-76), mper2-luc Δ (-75~-66), mper2-luc Δ (-65~-56), mper2-luc Δ (-55~-46) and mper2-luc Δ (-45~-36) showed a 20 to 40-min shorter period. Notably, the -75~-46 region seemed to largely contribute to the determination of period length. Basically, exogenously transfected transgenes have no effect on endogenous cell-autonomous circadian oscillation. Therefore, these slight changes in period lengths may be attributable to desynchronization of reporter expression from the endogenous oscillator and elicitation of transcriptional noise, because these mutations resulted in loss of robust transcriptional oscillation. The amplitude of mper2-luc Δ (-85~-76), mper2-luc Δ (65~-56), mper2-luc Δ (-45~-36) or mper2-luc Δ (-35~-26) was significantly smaller over cycles, compared to wild-type Per2-luc (Supplemental Fig. 2C), and deletion of -15~-6 or -25~-16 caused a remarkable inhibition of circadian fluctuation, as shown in 16
17 Figure 3A. The others showed only a slight decrease of the initial amplitude. In order to compare the damping pattern in the constructs, the amplitude in the initial cycle was set to 100 (Supplemental Fig. 2D). While mper2-luc Δ (-25~-16) showed a slightly irregular damping pattern, the others had almost the same damping pattern as that of wild-type Per2-luc. Taken together, the -85~-76, 65~-56, -45~-36 and -35~-26 regions work to enhance the amplitude of transcriptional oscillation, whereas the -15~-6 and -25~-16 regions are most essential to drive circadian oscillation. There are no regions that obviously affect the period length and damping rate. To rule out the possibility that these mutations disrupt the core promoter activity, we examined the basal transcriptional activity of these constructs by conducting reporter experiments. These mutations did not interfere with the core promoter activity, because these constructs did not show severe attenuation of the transcriptional activity found for the full-length construct (Supplemental Fig. 3). These results are consistent with those in Figure 2B and 2D. Taken together, our data indicate that the -25 to -6 region upstream from the TSS is indispensable for robust cell-autonomous circadian gene expression of Per2. Sequence inspection revealed an E-box-like sequence (CACGTT, instead of the known clock E-box sequence CACGTG) in this region (Fig. 3B). Endogenous circadian transcription factors bind site-specifically to the Per2 17
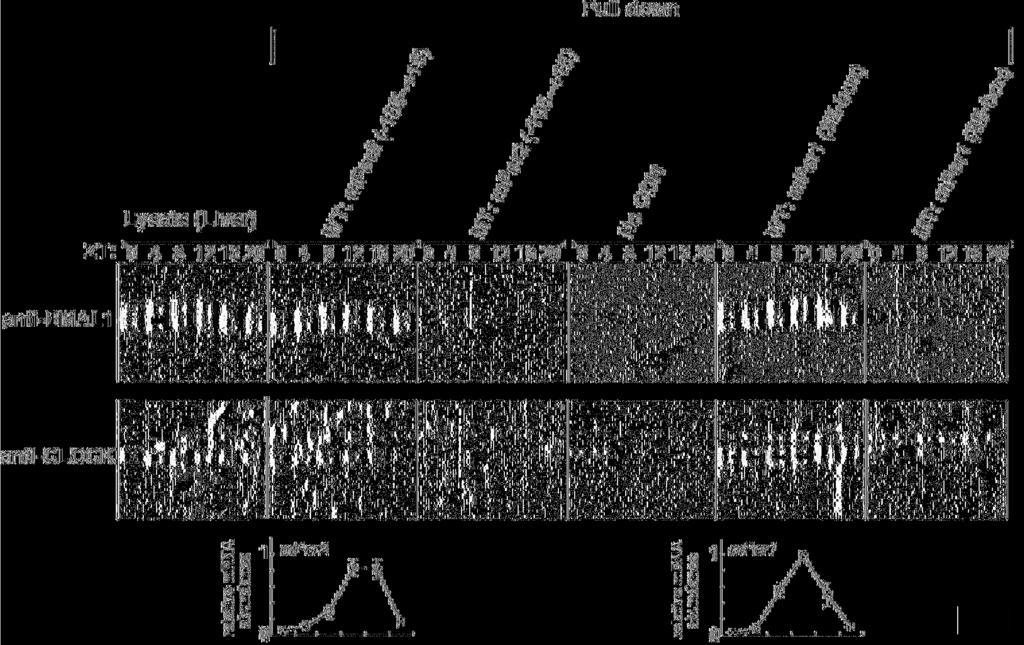
18 E-box-like sequence To examine whether circadian transcription factors indeed bind to the Per2 E-box-like sequence, we investigated the binding of endogenous BMAL1 and CLOCK to Per2 (-105~+15) double-stranded DNA fragments immobilized on streptavidin beads (Fig. 4). After having been entrained to LD (12-h light/12-h dark) cycles, BALB/c mice were transferred to DD (constant dark). Liver lysates were prepared at 4-hr intervals and immunoblotted with anti-bmal1 or anti-clock antibody. The shifted bands correspond to phosphorylated BMAL1, as reported recently (Lee et al., 2001; Kondratov et al., 2003; Tamaru et al., 2003). The phosphorylation levels peaked at ZT (Zeitgeber Time) 8 ~ ZT12. By pull-down experiments using the Per2 promoter fragment immobilized to beads, protein precipitation of BMAL1 and phosphorylated BMAL1 was observed. The peak of phosphorylated BMAL1 bound to the Per2 promoter correlated with that of Per2 mrna expression in the liver (lower panel). This correlation indicates that phosphorylated BMAL1 activated transcription of the Per2 gene, consistent with the report that formation of the CLOCK/BMAL1 complex is followed by their co-dependent phosphorylation (Kondratov et al., 2003). When we used the Per2 promoter fragment with a mutated E-box-like sequence, precipitated BMAL1 protein and phosphorylated BMAL1 were almost completely undetectable. Thus, BMAL1 and phosphorylated 18
19 BMAL1 were confirmed to specifically recognize and bind to the Per2 E-box-like sequence. We also confirmed that the BMAL1-CLOCK-mediated transcription of the Per2 gene was dependent on this E-box-like sequence by performing traditional luciferase assays (data not shown). As a positive control, when a DNA fragment containing 3 different Per1 E-boxes was used for pull-down assays, a similar pattern of BMAL1 binding was detected. Mutation of these Per1 E-boxes completely inhibited BMAL1 binding. As observed in the Per2 pull-down assays, the peak of phosphorylated BMAL1 bound to the Per1 E-boxes correlated with that of Per1 mrna expression (lower panel). Our results indicate that the Per2 E-box-like sequence, as well as the classical circadian E-box, binds to endogenous circadian transcription factors and that transcriptional activation of Per2 and Per1 correlates with phosphorylation of BMAL1. The Per2 E-box-like sequence has functional characteristics different from those of the classical circadian E-box Both Per2 and Per1 mrna expression showed circadian oscillations with high amplitude in peripheral tissues (Fig. 5A), whereas the amplitude of Per1 mrna rhythms was significantly lower than that of Per2 mrna rhythms in serum-stimulated NIH3T3 cells (Fig. 5B). This observation was reproducible when Rat-1 fibroblasts were used (data not shown; Balsalobre et al., 1998). A very recent report demonstrated that in vitro 19
20 cultured fibroblasts harbor self-sustained and cell-autonomous circadian clocks similar to those operative in SCN neurons (Nagoshi et al., 2004). Therefore, the high amplitude of Per1 mrna oscillation in peripheral tissues largely depends on the extracellular environment such as blood-borne factors and body temperature, which changes cyclically around the clock, rather than on the cell autonomous core clock. In support of this idea, in mper1::luc transgenic animals, peripheral organs fail to express persistent circadian rhythms in reporter gene activity (Yamazaki et al., 2000). In contrast, in mper2-luciferase knockin mice, peripheral tissues in explant cultures show robust and self-sustained circadian rhythms (Yoo et al., 2004). On the other hand, gene knockout studies have indicated that mper1-deficient mice display a persistent circadian rhythm (Bae et al., 2001; Cermakian et al., 2001; Zheng et al., 2001), whereas mice deficient for mper2 have no circadian rhythms in locomotor activity (Zheng et al., 1999; Bae et al., 2001; Zheng et al., 2001). Consistent with these behavioral phenotypes, disruption of mper2 results in reduced levels of clock gene expression in the SCN; and in contrast, mice homozygous for the targeted mper1 allele have unaltered SCN gene expression rhythms (Bae et al., 2001). Taken together, these reports also indicate that the Per1 gene might not be tightly incorporated into the cell-autonomous core clock mechanism. However, this issue remains controversial, because the loss of Per1 can result in 20
21 arrhythmic mice, in prolonged constant dark conditions (Bae et al., 2001). Also, as for the above in vivo experiments, the difference between transgenics and knockins should be recognised; the latter are in their native context and have significantly more potential cis-acting sequences surrounding them. If there are important elements that are distant from the TSS, they might be lost in the Per1 transgenics. Per1 is highly sensitive to various extracellular stimuli in vitro (Akashi and Nishida, 2000; Balsalobre et al., 2000); and also in vivo, circadian changes in the extracellular environment can readily induce expression of the Per1 gene. Thus, the dramatic circadian changes in Per1 mrna accumulation in peripheral tissues (Fig. 5A) may be attributed to this mechanism. Increased expression of the Per1 gene in response to changes in the extracellular environment may be functional in the entrainment of peripheral oscillators. In fact, in peripheral tissues of mper1-deficient mice, the phase of clock gene expression is not only delayed, but the peak of expression is broadened (Cermakian et al., 2001) A mechanism by which the Per2 E-box-like sequence generates a higher amplitude of circadian gene expression than does the classical circadian E-box As shown in Figure 3A, even when the region including the E-box-like sequence was deleted, the promoter activity still fluctuated in a circadian manner. These data suggest 21
22 that the neighboring -15 to -6 region also contains another element that regulates circadian transcription of Per2 by cooperating with the E-box-like sequence. This -15 to -6 region contains a consensus E4BP4 binding site (8/10 bp match), suggesting that DBP and E4BP4 may cooperate with BMAL1-CLOCK/PER-CRY to drive robust circadian gene expression of Per2. In Figure 5C, we substituted the Per2 E-box-like sequence (CACGTT) with the classical circadian E-box (CACGTG) by site-directed mutagenesis and monitored in real-time its transcriptional fluctuation. This substitution did not markedly affect either basal promoter activity or BMAL1-CLOCK-induced transactivation (data not shown). Interestingly, this 1-bp substitution resulted in small amplitude of circadian gene expression of Per2 (Fig. 5C, left panel), as observed in Per1-luc (Fig. 5C, right panel). The detrended bioluminescence datasets made it clear that compared to Per2-luc, Per2-luc (E-box) showed a small amplitude, and additionally that its period time-dependently became longer (Fig. 5D). Interestingly, Per2-luc (E-box) had a very similar pattern of amplitude, period and damping rate to those of Per1-luc (Fig. 5E). Thus, our data suggest that the Per2 E-box-like sequence generates high amplitude of circadian gene expression through cooperation between these 2 distinct elements (the 25 to 16 region and the 15 to 6 region) and that the 1-bp difference (CACGTT) is indispensable for this cooperation. 22
23 To examine whether E4BP4 controls transcriptional oscillation of Per2, we studied the binding of E4BP4 to Per2 (-45~+15). To confirm that the E4BP4-LUCIFERASE fusion protein binds to the Per2 fragment, we pulled down the fusion protein by using biotinylated double-strand Per2 (-45~+15) containing the putative E4BP4 consensus (Fig. 5F). In the presence of this Per2 fragment, the luciferase activity in the precipitate was enhanced, demonstrating the E4BP4 binding to this fragment. Next, in order to examine the role of E4BP4 protein in cell-autonomous transcriptional oscillation of Per2, we monitored the Per2 (-105)-luc activity in real time. As shown in Figure 5G, coexpression of E4BP4 resulted in a gradual reduction of the basal transcriptional activity (Left) and a shortened period length of circadian transcription (Right, detrended data). The expression of E4BP4 shortened the period in a dose-dependent manner (Fig. 5H). These results indicate that E4BP4 may be a transcriptional repressor in Per2 transcription and control the period length of Per2 oscillation. As shown in Fig. 3B and 5C, Per2 Δ (-15~-6)-luc and Per2 (E-box)-luc did not show deep troughs in circadian transcription, as wild type Per2-luc did, and therefore, we speculate that E4BP4 may be required for the trough formation in circadian transcription of Per2. 23
24 Conclusions The current feedback loop model has been based mainly on Per1. A publication that appeared after this paper was submitted demonstrated that a 210-bp region including the E-box-like sequence is sufficient for Per2 oscillation in vivo (Yoo et al., 2005). Consistent with this report, our data indicate that the -25 to -6 region upstream from the TSS, including the same E-box-like sequence, is indispensable for cell-autonomous circadian gene expression of Per2. Not only the E-box described by Yoo et al. and Ueda et al. (Ueda et al., 2005; Yoo et al., 2005) but also the 10-bp nucleotides next to the E-box are necessary for this oscillatory regulation. Furthermore, our biochemical data show that endogenous BMAL1 and CLOCK site-specifically bind to the E-box in a phosphorylation- and time-dependent manner (Fig. 4), which is novel observation. Importantly, our experiments were mainly performed on the cell-based real-time assay, because the core clock is a cell-autonomous system. In order to investigate if transcriptional oscillation is driven in the core clock system, we need to examine whether transcriptional regulatory elements function under a cell-autonomous condition. Our approach enables us to exclude internal environmental cues such as blood-borne factors and body temperature. Thus, these two reports verified that the current feedback loop model reflects a core clock mechanism. 24
25 The current molecular model for the mammalian circadian clock has been established mainly on the basis of data obtained by conventional reporter assays on the Per1 promoter, implying that several issues remain to be clarified. First, it is difficult to interpret whether the experimental system based on ectopic overexpression reflects physiological phenomena. The E-box consensus sequence, which consists of only 6 bp nucleotides, could appear at some frequency in the promoter region of many genes, leading to the possibility that overexpressed BMAL1 and CLOCK nonspecifically bind to E-box and E-box-like sequences and activate transcription (Munoz et al., 2002). Second, these transient assays are not suitable to monitor temporal changes, and therefore the obtained data do not reflect mechanisms of circadian rhythm generation. Mutant mice also could not clarify the role of BMAL1 and CLOCK in Per1 oscillation, due to the possibility of indirect effects. Thus, to date there is no compelling evidence that BMAL1 and CLOCK regulate circadian transcription of Per1, and that E-boxes are indispensable for circadian fluctuation of Per1 transcription. Third, the BMAL1: CLOCK regulation might be specific for Per1 transcription, because detailed analyses have not been performed on the involvement of BMAL1 and CLOCK in circadian transcription of other clock genes. So far at least we could not exclude the possibility that other as yet unknown components have pivotal roles in circadian transcription of other 25
26 clock genes. In this report, by the real-time monitoring of bioluminescence in cultured cells, we demonstrated that the BMAL1 and CLOCK binding region is included in only the 20-bp region essential for circadian transcription of Per2, an indispensable gene for the mammalian circadian clock. Our monitoring system was performed under the condition that the endogenous cell-autonomous circadian pacemaker was indeed operating, without the use of overexpression. Our results are summarized in Figure 5I. Next to the Per2 E-box-like sequence, there is another region that generates more robust cell-autonomous oscillation in transcription of Per2. A combination of the E-box-like sequence and this cooperating element increases the amplitude of cell-autonomous rhythmic transcription of Per2, and consequently the amplitude of Per2 mrna rhythms is significantly higher than that of Per1 mrna rhythms. The 1-bp difference (CACGTT) from the classical circadian E-box is indispensable for this combination. This discovery suggests that the atypical E-box (CACGTT) might be more functional for circadian gene expression rather than the classical E-box (CACGTG) that has been believed to be as the BMAL1:CLOCK binding consensus. However, we still need to note that the reduction in clock-gene amplitude may not necessarily indicate that the pacemaker amplitude is reduced. Future studies will define the precise relationship between the amplitude of clock gene oscillation and that 26
27 of pacemaker function. The cell-autonomous core clock generates overt circadian oscillations in Per2 transcription, whereas the high amplitude of Per1 oscillation in vivo largely depends on the extracellular environment, which changes cyclically around the clock, rather than on the core clock. This increased expression of the Per1 gene in response to circadian changes of the extracellular environment may be functional in the entrainment of peripheral oscillators. Among genes that are expressed in a circadian manner, there are clock genes that are central to the timing mechanism and output genes that directly or indirectly mediate physiology under circadian control. The phases and amplitudes in the circadian expression of these genes are different from each other (Panda et al., 2002; Storch et al., 2002). Nevertheless, it is thought that a few enhancer elements, such as E-box, RORE, and DBPE, generate a broad range of phases and amplitudes in circadian gene expression (Yamamoto et al., 2004). Our results provide an explanation for why a small number of elements generate various patterns of circadian gene expression. We showed that two or more transcriptional regulatory elements, such as a phase delaying element and a rhythm-generating element, are required for robust circadian gene expression of Per2, illustrating that even when the same rhythm-generating element regulates several different genes, combinations with other enhancer elements can advance or delay the 27
28 phase of these circadian gene expressions. The copy number and various combinations of elements would create unlimited patterns in phases. On the other hand, we showed that the Per2 E-box-like element might generate high amplitude of circadian gene expression by cooperating with a neighboring element. This result demonstrates that synergistic cooperation of several elements may generate more dynamic oscillation in circadian gene expression. In fact, we found that 2 ROR response elements synergistically function in circadian transcription of Bmal1 (Akashi and Takumi, 2005), illustrating that cooperation of elements can enhance the amplitude. The molecular mechanism by which circadian expression of clock and clock-related genes shows a variety of phases and amplitudes will be more clearly revealed by further detailed analyses of the regulatory mechanism for circadian transcription of each known clock gene. 28
29 ACKNOWLEDGMENTS We thank Setsuko Tsuboi, Yukari Shima and Daniel Trcka for their expert technical assistance, as well as Takuro Yamamoto, Yasukazu Nakahata, Atsuko Takano, Mayumi Yoshida, and Pabel Delgado for help, reagents, and discussions. We also thank Eisuke Nishida and Keiji Tanaka for general support. We are grateful to Teruya Tamaru for providing the anti-bmal1 antibody, and to Shin Yamazaki for helpful suggestion on data analysis. This work was supported in part by research grants from MEXT. The support of fellowships from the Japan Society for the Promotion of Science (M.A.) is also acknowledged. 29
30 References Abe, M., Herzog, E.D., Yamazaki, S., Straume, M., Tei, H., Sakaki, Y., Menaker, M., and Block, G.D. (2002). Circadian rhythms in isolated brain regions. J Neurosci 22, Akashi, M., and Nishida, E. (2000). Involvement of the MAP kinase cascade in resetting of the mammalian circadian clock. Genes Dev 14, Akashi, M., and Takumi, T. (2005). The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat Struct Mol Biol 12, Akashi, M., Tsuchiya, Y., Yoshino, T., and Nishida, E. (2002). Control of intracellular dynamics of mammalian period proteins by casein kinase I epsilon (CKIepsilon) and CKIdelta in cultured cells. Mol Cell Biol 22, Albrecht, U., and Eichele, G. (2003). The mammalian circadian clock. Curr Opin Genet Dev 13, Albrecht, U., Sun, Z.S., Eichele, G., and Lee, C.C. (1997). A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell 91, Allada, R., Emery, P., Takahashi, J.S., and Rosbash, M. (2001). Stopping time: the genetics of fly and mouse circadian clocks. Annu Rev Neurosci 24,
31 Bae, K., Jin, X., Maywood, E.S., Hastings, M.H., Reppert, S.M., and Weaver, D.R. (2001). Differential functions of mper1, mper2, and mper3 in the SCN circadian clock. Neuron 30, Balsalobre, A., Damiola, F., and Schibler, U. (1998). A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 93, Balsalobre, A., Marcacci, L., and Schibler, U. (2000). Multiple signaling pathways elicit circadian gene expression in cultured Rat-1 fibroblasts. Curr Biol 10, Bunger, M.K., Wilsbacher, L.D., Moran, S.M., Clendenin, C., Radcliffe, L.A., Hogenesch, J.B., Simon, M.C., Takahashi, J.S., and Bradfield, C.A. (2000). Mop3 is an essential component of the master circadian pacemaker in mammals. Cell 103, Cermakian, N., Monaco, L., Pando, M.P., Dierich, A., and Sassone-Corsi, P. (2001). Altered behavioral rhythms and clock gene expression in mice with a targeted mutation in the Period1 gene. Embo J 20, Dunlap, J.C. (1999). Molecular bases for circadian clocks. Cell 96, Gekakis, N., Staknis, D., Nguyen, H.B., Davis, F.C., Wilsbacher, L.D., King, D.P., Takahashi, J.S., and Weitz, C.J. (1998). Role of the CLOCK protein in the mammalian circadian mechanism. Science 280,
32 Hida, A., Koike, N., Hirose, M., Hattori, M., Sakaki, Y., and Tei, H. (2000). The human and mouse Period1 genes: five well-conserved E-boxes additively contribute to the enhancement of mper1 transcription. Genomics 65, Kaasik, K., and Lee, C.C. (2004). Reciprocal regulation of haem biosynthesis and the circadian clock in mammals. Nature 430, Kondo, T., and Ishiura, M. (2000). The circadian clock of cyanobacteria. Bioessays 22, Kondratov, R.V., Chernov, M.V., Kondratova, A.A., Gorbacheva, V.Y., Gudkov, A.V., and Antoch, M.P. (2003). BMAL1-dependent circadian oscillation of nuclear CLOCK: posttranslational events induced by dimerization of transcriptional activators of the mammalian clock system. Genes Dev 17, Kume, K., Zylka, M.J., Sriram, S., Shearman, L.P., Weaver, D.R., Jin, X., Maywood, E.S., Hastings, M.H., and Reppert, S.M. (1999). mcry1 and mcry2 are essential components of the negative limb of the circadian clock feedback loop. Cell 98, Lee, C., Etchegaray, J.P., Cagampang, F.R., Loudon, A.S., and Reppert, S.M. (2001). Posttranslational mechanisms regulate the mammalian circadian clock. Cell 107, Lowrey, P.L., and Takahashi, J.S. (2004). Mammalian circadian biology: elucidating 32
33 genome-wide levels of temporal organization. Annu Rev Genomics Hum Genet 5, Menaker, M. (2003). Circadian rhythms. Circadian photoreception. Science 299, Munoz, E., Brewer, M., and Baler, R. (2002). Circadian Transcription. Thinking outside the E-Box. J Biol Chem 277, Nagoshi, E., Saini, C., Bauer, C., Laroche, T., Naef, F., and Schibler, U. (2004). Circadian gene expression in individual fibroblasts: cell-autonomous and self-sustained oscillators pass time to daughter cells. Cell 119, Panda, S., Antoch, M.P., Miller, B.H., Su, A.I., Schook, A.B., Straume, M., Schultz, P.G., Kay, S.A., Takahashi, J.S., and Hogenesch, J.B. (2002). Coordinated transcription of key pathways in the mouse by the circadian clock. Cell 109, Pittendrigh, C.S. (1993). Temporal organization: reflections of a Darwinian clock-watcher. Annu Rev Physiol 55, Ralph, M.R., Foster, R.G., Davis, F.C., and Menaker, M. (1990). Transplanted suprachiasmatic nucleus determines circadian period. Science 247, Reppert, S.M., and Weaver, D.R. (2002). Coordination of circadian timing in mammals. Nature 418,
34 Schibler, U., and Sassone-Corsi, P. (2002). A web of circadian pacemakers. Cell 111, Storch, K.F., Lipan, O., Leykin, I., Viswanathan, N., Davis, F.C., Wong, W.H., and Weitz, C.J. (2002). Extensive and divergent circadian gene expression in liver and heart. Nature 417, Takumi, T., Matsubara, C., Shigeyoshi, Y., Taguchi, K., Yagita, K., Maebayashi, Y., Sakakida, Y., Okumura, K., Takashima, N., and Okamura, H. (1998). A new mammalian period gene predominantly expressed in the suprachiasmatic nucleus. Genes Cells 3, Tamaru, T., Isojima, Y., van der Horst, G.T., Takei, K., Nagai, K., and Takamatsu, K. (2003). Nucleocytoplasmic shuttling and phosphorylation of BMAL1 are regulated by circadian clock in cultured fibroblasts. Genes Cells 8, Toh, K.L., Jones, C.R., He, Y., Eide, E.J., Hinz, W.A., Virshup, D.M., Ptacek, L.J., and Fu, Y.H. (2001). An hper2 phosphorylation site mutation in familial advanced sleep phase syndrome. Science 291, Travnickova-Bendova, Z., Cermakian, N., Reppert, S.M., and Sassone-Corsi, P. (2002). Bimodal regulation of mperiod promoters by CREB-dependent signaling and CLOCK/BMAL1 activity. Proc Natl Acad Sci U S A 99,
35 Ueda, H.R., Hayashi, S., Chen, W., Sano, M., Machida, M., Shigeyoshi, Y., Iino, M., and Hashimoto, S. (2005). System-level identification of transcriptional circuits underlying mammalian circadian clocks. Nat Genet 37, Vitaterna, M.H., Selby, C.P., Todo, T., Niwa, H., Thompson, C., Fruechte, E.M., Hitomi, K., Thresher, R.J., Ishikawa, T., Miyazaki, J., Takahashi, J.S., and Sancar, A. (1999). Differential regulation of mammalian period genes and circadian rhythmicity by cryptochromes 1 and 2. Proc Natl Acad Sci U S A 96, Williams, J.A., and Sehgal, A. (2001). Molecular components of the circadian system in Drosophila. Annu Rev Physiol 63, Yamamoto, T., Nakahata, Y., Soma, H., Akashi, M., Mamine, T., and Takumi, T. (2004). Transcriptional oscillation of canonical clock genes in mouse peripheral tissues. BMC Mol Biol 5, 18. Yamazaki, S., Numano, R., Abe, M., Hida, A., Takahashi, R., Ueda, M., Block, G.D., Sakaki, Y., Menaker, M., and Tei, H. (2000). Resetting central and peripheral circadian oscillators in transgenic rats. Science 288, Yamazaki, S., Straume, M., Tei, H., Sakaki, Y., Menaker, M., and Block, G.D. (2002). Effects of aging on central and peripheral mammalian clocks. Proc Natl Acad Sci U S A 99,
36 Yoo, S.H., Ko, C.H., Lowrey, P.L., Buhr, E.D., Song, E.J., Chang, S., Yoo, O.J., Yamazaki, S., Lee, C., and Takahashi, J.S. (2005). A noncanonical E-box enhancer drives mouse Period2 circadian oscillations in vivo. Proc Natl Acad Sci U S A 102, Yoo, S.H., Yamazaki, S., Lowrey, P.L., Shimomura, K., Ko, C.H., Buhr, E.D., Siepka, S.M., Hong, H.K., Oh, W.J., Yoo, O.J., Menaker, M., and Takahashi, J.S. (2004). PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci U S A 101, Young, M.W., and Kay, S.A. (2001). Time zones: a comparative genetics of circadian clocks. Nat Rev Genet 2, Zheng, B., Albrecht, U., Kaasik, K., Sage, M., Lu, W., Vaishnav, S., Li, Q., Sun, Z.S., Eichele, G., Bradley, A., and Lee, C.C. (2001). Nonredundant roles of the mper1 and mper2 genes in the mammalian circadian clock. Cell 105, Zheng, B., Larkin, D.W., Albrecht, U., Sun, Z.S., Sage, M., Eichele, G., Lee, C.C., and Bradley, A. (1999). The mper2 gene encodes a functional component of the mammalian circadian clock. Nature 400, Figure Legends Figure 1. A phase-delaying element and a rhythm-generating element are required 36
37 for robust cell-autonomous circadian gene expression of Per2 (A) Schematic representation of deletion mutants of the mper2 promoter. +1 corresponds to the transcription start site. (B) Transcriptional oscillation of mper2 was monitored by using the cell culture-based luminescent reporter assay. NIH3T3 cells were transfected with the mper2-luc construct and then stimulated with a high concentration of serum. After the serum shock, in the presence of luciferin, light emission was measured and integrated for 1 min at intervals of 15 min (vertical scale: relative counts per minute, horizontal scale: 1440 minutes = 1 day). Peak values of the curves were set to 1. A representative result of three independent experiments is shown. (C) The signals obtained in (B) were detrended. The time of the first peak was calculated as a phase marker (mean ± SEM; n = 4). (D) Schematic representation of the results obtained from the deletion analysis. Figure 2. Identification of the region responsible for driving cell-autonomous circadian oscillation of Per2 transcription (A) Schematic representation of a set of constructs in which a 40-bp fragment from Per2 (-105~+15) was inserted into the upstream of the SV40 promoter attached to the luciferase gene. These fragments overlap by 20 bp. (B) Transcriptional oscillation of mper2-sv40-luc was monitored. NIH3T3 cells were transfected and then stimulated 37
38 with a high concentration of serum. After the serum shock, light emission was measured and integrated for 1 min at intervals of 15 min (vertical scale: relative counts per minute, horizontal scale: 1440 minutes = 1 day). Peak values of the curves were set to 1. A representative result of three independent experiments is shown. For accurate comparison, thin lines show the curve for SV40-luc. (C) Schematic representation of another set of constructs, in which a 20-bp fragment from Per2 (-45~+15) was inserted into the upstream of the SV40 promoter attached to the luciferase gene. These fragments overlap by 15 bp. (D) Transcriptional oscillation of mper2-sv40-luc was monitored. Peak values of the curves were set to 1 (horizontal scale: 1440 minutes = 1 day). A representative result of three independent experiments is shown. For accurate comparison, thin lines show the curve for SV40-luc. Figure 3. Mapping of the rhythm-generating element by using deletion mutants of Per2 (A) Transcriptional oscillation of a series of mper2-luc 10-bp deletion constructs was monitored in real time. NIH3T3 cells were transfected and then stimulated with a high concentration of serum. Peak values of the curves were set to 1 (vertical scale: relative counts per minute, horizontal scale: 1440 minutes = 1 day). A representative result of three independent experiments is shown. For accurate comparison, thin lines show the 38
39 curve for mper2-luc (-105). (B) Schematic representation of the results obtained from the 10-bp deletion analysis, taken together with those in Figure 2. Sequence inspection reveals an E-box-like sequence (CACGTT) in the region essential for circadian transcription of Per2. Figure 4. Temporal patterns of the site-specific binding of endogenous circadian transcription factors to the Per2 E-box-like sequence (Upper and middle panels) Mouse liver extracts were harvested at 4-h intervals, and then subjected to immunoblot analysis with anti-bmal1 antibody (upper panels) or anti-clock antibody (middle panels). Mouse liver extracts were incubated with a double-stranded biotinylated oligonucleotide including the consensus-predicted Per2 E-box-like sequence (CACGTT) or 3 different Per1 E-boxes (wild-type: WT, mutant: MT), which was immobilized on streptavidin-sepharose beads. The negative control samples were treated with the beads without an oligonucleotide (No ODN). The resulting precipitates were subjected to immunoblot analysis with anti-bmal1 antibody (upper panels) or anti-clock antibody (middle panels). (Lower panels) Temporal expression patterns of the mper2 and mper1 genes in mouse liver were assayed by real-time quantitative RT-PCR. Each value represents the average of 3 independent RT-PCR experiments. The relative levels were normalized to the corresponding Gapdh RNA 39
40 levels. Peak values of the mper2 and mper1 curves were set to 1. Figure 5. A potential mechanism by which the cell-autonomous core clock generates more overt circadian oscillations in Per2 transcription than in Per1 transcription (A and B) Temporal expression patterns of the mper1 and mper2 genes, in mouse peripheral tissues (A) and in serum-stimulated NIH3T3 cells (B) were assayed by real-time quantitative RT-PCR. Mice were kept in a 12-h light: 12-h dark cycle (LD, lights on 8 a.m.; lights off 8 p.m.) for 2 weeks to establish entrainment. Three animals were sacrificed at the times given on the abscissae of the diagrams. Each value represents the average of 3 independent RT-PCR experiments. The relative levels of each RNA were normalized to the corresponding Gapdh RNA levels. Peak values of the mper1 and mper2 curves were set to 1. (C) Transcriptional oscillation of mper2-luc, mper2-luc (E-box), and mper1-luc was monitored in real time. NIH3T3 cells were transfected and then stimulated with a high concentration of serum. Peak values of the curves were set to 1 (vertical scale: relative counts per minute, horizontal scale: 1440 minutes = 1 day). A representative result of three independent experiments is shown. (D and E) The signals obtained in (C) were detrended. (F) COS7 cells were transfected with the E4BP4-luciferase fusion expression vector. Cell extracts were incubated with the 40
41 double-stranded biotinylated Per2 (-45~+15) oligonucleotide, including the consensus-predicted E4BP4 response elements, which was immobilized on streptavidin-sepharose beads. The negative control samples were treated with streptavidin-sepharose beads without an oligonucleotide. The resulting precipitates were subjected to luciferase assays. Data represent the mean ± SEM of triplicate samples. (G) Transcriptional oscillation of Per2 (-105)-luc was monitored, in the presence or absence of E4BP4 (Left). The signals obtained were detrended (Right). (H) With increasing dose of the E4BP4 expression plasmid, transcriptional oscillation of Per2 (-105)-luc was monitored. The periods were obtained from analysis of a circadian marker. Data represent the mean ± SEM of triplicate samples. (I) Schematic model representing BMAL1:CLOCK-mediated control of cell-autonomous Per1 and Per2 oscillation. A combination of the E-box-like sequence and a cooperating element increases the amplitude of rhythmic transcription of Per2, and consequently the amplitude of Per2 mrna rhythms is significantly higher than that of Per1 mrna rhythms. The 1-bp difference (CACGTT) from the classical circadian E-box may be indispensable for this combination. The cell-autonomous core clock generates overt circadian oscillations in Per2 transcription, whereas the high amplitude of Per1 oscillation in vivo largely depends on the extracellular environment, which changes cyclically around the clock, 41
42 rather than on the core clock. Thus, the Per1 gene might not be tightly incorporated into the cell-autonomous core clock mechanism. 42
43
44
45
46
47
Advance in circadian rhythm genetics in mammals
 16 2 2004 4 Chinese Bulletin of Life Sciences Vol. 16, No. 2 Apr., 2004 1004-0374 (2004) 02-0104-05 1 100101 2 434025 9 24, Q41 A Advance in circadian rhythm genetics in mammals XU Zu-Yuan 1,2 (1 Beijing
16 2 2004 4 Chinese Bulletin of Life Sciences Vol. 16, No. 2 Apr., 2004 1004-0374 (2004) 02-0104-05 1 100101 2 434025 9 24, Q41 A Advance in circadian rhythm genetics in mammals XU Zu-Yuan 1,2 (1 Beijing
CIRCADIAN SIGNALING NETWORKS
 Transcription Regulation And Gene Expression in Eukaryotes Cycle G2 (lecture 13709) FS 2014 P. Matthias and RG Clerc Roger G. Clerc 07.05.2014 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «Slave clocks»
Transcription Regulation And Gene Expression in Eukaryotes Cycle G2 (lecture 13709) FS 2014 P. Matthias and RG Clerc Roger G. Clerc 07.05.2014 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «Slave clocks»
Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 # )
") Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 #13709-01) CIRCADIAN SIGNALING NETWORKS RG. Clerc May 19. 2010 www.fmi.ch/training/teaching Circadian rythms : most physiological processes
Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 #13709-01) CIRCADIAN SIGNALING NETWORKS RG. Clerc May 19. 2010 www.fmi.ch/training/teaching Circadian rythms : most physiological processes
Report. Circadian Clock Gene Bmal1 Is Not Essential; Functional Replacement with its Paralog, Bmal2
 Current Biology 20, 316 321, February 23, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2009.12.034 Circadian Clock Gene Bmal1 Is Not Essential; Functional Replacement with its Paralog,
Current Biology 20, 316 321, February 23, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2009.12.034 Circadian Clock Gene Bmal1 Is Not Essential; Functional Replacement with its Paralog,
Multiple circadian transcriptional elements cooperatively regulate cell-autonomous transcriptional oscillation of Period3, a mammalian clock gene
 JBC Papers in Press. Published on August 15, 2017 as Manuscript M117.806836 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m117.806836 Multiple circadian transcriptional elements cooperatively
JBC Papers in Press. Published on August 15, 2017 as Manuscript M117.806836 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m117.806836 Multiple circadian transcriptional elements cooperatively
Molecular Signals of Mammalian Circadian Clock
 Kobe J. Med. Sci., Vol. 50, No. 4, pp. 101-109, 2004 Molecular Signals of Mammalian Circadian Clock JING ZHANG, XIN DONG, YOSHITO FUJIMOTO, and HITOSHI OKAMURA Division of Molecular Brain Science, Department
Kobe J. Med. Sci., Vol. 50, No. 4, pp. 101-109, 2004 Molecular Signals of Mammalian Circadian Clock JING ZHANG, XIN DONG, YOSHITO FUJIMOTO, and HITOSHI OKAMURA Division of Molecular Brain Science, Department
Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2
 Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2 P. Matthias and RG Clerc Pharmazentrum Hörsaal 2 16h15-18h00 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «slave
Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2 P. Matthias and RG Clerc Pharmazentrum Hörsaal 2 16h15-18h00 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «slave
Clicker Question. The Need to Decompose. Mechanism and Reduction: Decomposing Circadian Clocks
 Mechanism and Reduction: Decomposing Circadian Clocks Clicker Question On the Deductive-Nomological (DN) model of reduction, which of the following does not figure in providing the explanation (i.e., is
Mechanism and Reduction: Decomposing Circadian Clocks Clicker Question On the Deductive-Nomological (DN) model of reduction, which of the following does not figure in providing the explanation (i.e., is
Supplementary Information for
 Supplementary Information for Involvement of urinary bladder Connexin43 and the circadian clock in the coordination of diurnal micturition rhythm Hiromitsu Negoro, 1,2 Akihiro Kanematsu, 1,3 Masao Doi,
Supplementary Information for Involvement of urinary bladder Connexin43 and the circadian clock in the coordination of diurnal micturition rhythm Hiromitsu Negoro, 1,2 Akihiro Kanematsu, 1,3 Masao Doi,
Nature Genetics: doi: /ng.3731
 Supplementary Figure 1 Circadian profiles of Adarb1 transcript and ADARB1 protein in mouse tissues. (a) Overlap of rhythmic transcripts identified in the previous transcriptome analyses. The mouse liver
Supplementary Figure 1 Circadian profiles of Adarb1 transcript and ADARB1 protein in mouse tissues. (a) Overlap of rhythmic transcripts identified in the previous transcriptome analyses. The mouse liver
SUPPLEMENTARY INFORMATION
 Supplementary Table 2 Mouse circadian s and observed circadian and physiological phenotypes. Gene Circadian phenotype Ref. Associated physiological abnormality Ref. Bmal1/Mop3 (Arntl) Loss-of-circadian
Supplementary Table 2 Mouse circadian s and observed circadian and physiological phenotypes. Gene Circadian phenotype Ref. Associated physiological abnormality Ref. Bmal1/Mop3 (Arntl) Loss-of-circadian
T. WU 1, Y. NI 1, F. ZHUGE 1, Z. FU 1. Introduction
 Physiol. Res. 59: 581-590, 2010 Resetting Process of Peripheral Circadian Gene Expression after the Combined Reversal of Feeding Schedule and Light/Dark Cycle Via a 24-h Light Period Transition in Rats
Physiol. Res. 59: 581-590, 2010 Resetting Process of Peripheral Circadian Gene Expression after the Combined Reversal of Feeding Schedule and Light/Dark Cycle Via a 24-h Light Period Transition in Rats
The Success of Decomposition
 11/21/11 Mechanism and Levels of Organization: Recomposing and Situating Circadian Clocks The Success of Decomposition Moving beyond per, researchers in the 1990s and early 2000s identified many clock
11/21/11 Mechanism and Levels of Organization: Recomposing and Situating Circadian Clocks The Success of Decomposition Moving beyond per, researchers in the 1990s and early 2000s identified many clock
The Human PER1 Gene is Inducible by Interleukin-6
 Journal of Molecular Neuroscience Copyright 2002 Humana Press Inc. All rights of any nature whatsoever reserved. ISSN0895-8696/02/18:105 110/$11.25 The Human PER1 Gene is Inducible by Interleukin-6 Dirk
Journal of Molecular Neuroscience Copyright 2002 Humana Press Inc. All rights of any nature whatsoever reserved. ISSN0895-8696/02/18:105 110/$11.25 The Human PER1 Gene is Inducible by Interleukin-6 Dirk
Involvement of the MAP kinase cascade in resetting of the mammalian circadian clock
 RESEARCH COMMUNICATION Involvement of the MAP kinase cascade in resetting of the mammalian circadian clock Makoto Akashi 1 and Eisuke Nishida 1,2 1 Department of Biophysics, Graduate School of Science
RESEARCH COMMUNICATION Involvement of the MAP kinase cascade in resetting of the mammalian circadian clock Makoto Akashi 1 and Eisuke Nishida 1,2 1 Department of Biophysics, Graduate School of Science
Phase Shifts of Circadian Transcripts in Rat Suprachiasmatic Nucleus
 The Second International Symposium on Optimization and Systems Biology (OSB 08) Lijiang, China, October 31 November 3, 2008 Copyright 2008 ORSC & APORC, pp. 109 114 Phase Shifts of Circadian Transcripts
The Second International Symposium on Optimization and Systems Biology (OSB 08) Lijiang, China, October 31 November 3, 2008 Copyright 2008 ORSC & APORC, pp. 109 114 Phase Shifts of Circadian Transcripts
Strong Resetting of the Mammalian Clock by Constant Light Followed by Constant Darkness
 The Journal of Neuroscience, November 12, 2008 28(46):11839 11847 11839 Behavioral/Systems/Cognitive Strong Resetting of the Mammalian Clock by Constant Light Followed by Constant Darkness Rongmin Chen,
The Journal of Neuroscience, November 12, 2008 28(46):11839 11847 11839 Behavioral/Systems/Cognitive Strong Resetting of the Mammalian Clock by Constant Light Followed by Constant Darkness Rongmin Chen,
Oscillation and Light Induction of timeless mrna in the Mammalian Circadian Clock
 The Journal of Neuroscience, 1999, Vol. 19 RC15 1of6 Oscillation and Light Induction of timeless mrna in the Mammalian Circadian Clock Shelley A. Tischkau, 1 Jeffrey A. Barnes, 1 Fang-Ju Lin, 2 Edith M.
The Journal of Neuroscience, 1999, Vol. 19 RC15 1of6 Oscillation and Light Induction of timeless mrna in the Mammalian Circadian Clock Shelley A. Tischkau, 1 Jeffrey A. Barnes, 1 Fang-Ju Lin, 2 Edith M.
Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms
 Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms Hanspeter Herzel Institute for Theoretical Biology (ITB) Charité and Humboldt University Berlin Molecular Chronobiology SCN-neuron nucleus
Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms Hanspeter Herzel Institute for Theoretical Biology (ITB) Charité and Humboldt University Berlin Molecular Chronobiology SCN-neuron nucleus
Circadian Rhythms in Physiology and Behavior. The Persistence of Memory, Salvador Dali, 1931
 Circadian Rhythms in Physiology and Behavior The Persistence of Memory, Salvador Dali, 1931 Homeostasis and Rhythms? Homeostasis (Bernard, 1878): All the vital mechanisms, however varied they may be, have
Circadian Rhythms in Physiology and Behavior The Persistence of Memory, Salvador Dali, 1931 Homeostasis and Rhythms? Homeostasis (Bernard, 1878): All the vital mechanisms, however varied they may be, have
Supplementary Figure 1. Expression of the inducible tper2 is proportional to Dox/Tet concentration in Rosa-DTG/Per2 Per2-luc/wt MEFs.
 Supplementary Figure 1. Expression of the inducible tper2 is proportional to Dox/Tet concentration in Rosa-DTG/Per2 Per2-luc/wt MEFs. (a) Dose-responsive expression of tper2 by Dox. Note that there are
Supplementary Figure 1. Expression of the inducible tper2 is proportional to Dox/Tet concentration in Rosa-DTG/Per2 Per2-luc/wt MEFs. (a) Dose-responsive expression of tper2 by Dox. Note that there are
Cellular DBP and E4BP4 proteins are critical for determining the period length of the circadian oscillator
 FEBS Letters 55 () 7 journal homepage: www.febsletters.org Cellular and proteins are critical for determining the period length of the circadian oscillator Daisuke Yamajuku,, Yasutaka Shibata, Masashi
FEBS Letters 55 () 7 journal homepage: www.febsletters.org Cellular and proteins are critical for determining the period length of the circadian oscillator Daisuke Yamajuku,, Yasutaka Shibata, Masashi
Stochastic simulations
 Stochastic simulations Application to circadian clocks Didier Gonze Circadian rhythms Circadian rhythms allow living organisms to live in phase with the alternance of day and night... Circadian rhythms
Stochastic simulations Application to circadian clocks Didier Gonze Circadian rhythms Circadian rhythms allow living organisms to live in phase with the alternance of day and night... Circadian rhythms
The dominant circadian pacemaker in the mammalian brain is
 The methamphetamine-sensitive circadian oscillator does not employ canonical clock genes Jennifer A. Mohawk, Matthew L. Baer, and Michael Menaker 1 Department of Biology, University of Virginia, Charlottesville
The methamphetamine-sensitive circadian oscillator does not employ canonical clock genes Jennifer A. Mohawk, Matthew L. Baer, and Michael Menaker 1 Department of Biology, University of Virginia, Charlottesville
Light and Glutamate-Induced Degradation of the Circadian Oscillating Protein BMAL1 during the Mammalian Clock Resetting
 The Journal of Neuroscience, October 15, 2000, 20(20):7525 7530 Light and Glutamate-Induced Degradation of the Circadian Oscillating Protein BMAL1 during the Mammalian Clock Resetting Teruya Tamaru, 1
The Journal of Neuroscience, October 15, 2000, 20(20):7525 7530 Light and Glutamate-Induced Degradation of the Circadian Oscillating Protein BMAL1 during the Mammalian Clock Resetting Teruya Tamaru, 1
In mammals, a circadian pacemaker located in the suprachiasmatic
 PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues Seung-Hee Yoo*, Shin Yamazaki, Phillip L. Lowrey*, Kazuhiro Shimomura*,
PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues Seung-Hee Yoo*, Shin Yamazaki, Phillip L. Lowrey*, Kazuhiro Shimomura*,
PROMOTER ANALYSIS OF MAMMALIAN CLOCK CONTROLLED GENES
 65 PROMOTER ANALYSIS OF MAMMALIAN CLOCK CONTROLLED GENES KATARZYNA BOŻEK1 SZYMON M. KIE LBASA 2 k.bozek@biologie.hu-berlin.de kielbasa@molgen.mpg.de ACHIM KRAMER 3 HANSPETER HERZEL 1 achim.kramer@charite.de
65 PROMOTER ANALYSIS OF MAMMALIAN CLOCK CONTROLLED GENES KATARZYNA BOŻEK1 SZYMON M. KIE LBASA 2 k.bozek@biologie.hu-berlin.de kielbasa@molgen.mpg.de ACHIM KRAMER 3 HANSPETER HERZEL 1 achim.kramer@charite.de
Intercellular Coupling Confers Robustness against Mutations in the SCN Circadian Clock Network
 Intercellular Coupling Confers Robustness against Mutations in the SCN Circadian Clock Network Andrew C. Liu, 1,2,11 David K. Welsh, 1,3,4,11 Caroline H. Ko, 6,7 Hien G. Tran, 1,2 Eric E. Zhang, 1,2 Aaron
Intercellular Coupling Confers Robustness against Mutations in the SCN Circadian Clock Network Andrew C. Liu, 1,2,11 David K. Welsh, 1,3,4,11 Caroline H. Ko, 6,7 Hien G. Tran, 1,2 Eric E. Zhang, 1,2 Aaron
Functional central rhythmicity and light entrainment, but not liver and muscle rhythmicity, are Clock independent
 Am J Physiol Regul Integr Comp Physiol 291: R1172 R1180, 2006. First published May 18, 2006; doi:10.1152/ajpregu.00223.2006. Functional central rhythmicity and light entrainment, but not liver and muscle
Am J Physiol Regul Integr Comp Physiol 291: R1172 R1180, 2006. First published May 18, 2006; doi:10.1152/ajpregu.00223.2006. Functional central rhythmicity and light entrainment, but not liver and muscle
Biological Clocks. Lu Chen, Ph.D. MCB, UC Berkeley. Why Does Melatonin Now Outsell Vitamin C??
 Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 Why Does Melatonin Now Outsell Vitamin C?? Wake / sleep complaints are extremely prevalent. Much melatonin is consumed in an attempt to overcome the
Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 Why Does Melatonin Now Outsell Vitamin C?? Wake / sleep complaints are extremely prevalent. Much melatonin is consumed in an attempt to overcome the
BioNSi: A Discrete Biological Network Simulator Tool
 BioNSi: A Discrete Biological Network Simulator Tool Amir Rubinstein 1, Noga Bracha 1, Liat Rudner 1, Noga Zucker 1, Hadas E. Sloin 2, and Benny Chor 1 1 Blavatnik School of Computer Science, Tel Aviv
BioNSi: A Discrete Biological Network Simulator Tool Amir Rubinstein 1, Noga Bracha 1, Liat Rudner 1, Noga Zucker 1, Hadas E. Sloin 2, and Benny Chor 1 1 Blavatnik School of Computer Science, Tel Aviv
Stochastic simulations
 Circadian rhythms Stochastic simulations Circadian rhythms allow living organisms to live in phase with the alternance of day and night... Application to circadian clocks Didier Gonze Circadian rhythms
Circadian rhythms Stochastic simulations Circadian rhythms allow living organisms to live in phase with the alternance of day and night... Application to circadian clocks Didier Gonze Circadian rhythms
Circadian Rhythms in Isolated Brain Regions
 The Journal of Neuroscience, January 1, 2002, 22(1):350 356 Circadian Rhythms in Isolated Brain Regions Michikazu Abe, 1 * Erik D. Herzog, 1 * Shin Yamazaki, 1 Marty Straume, 1 Hajime Tei, 2 Yoshiyuki
The Journal of Neuroscience, January 1, 2002, 22(1):350 356 Circadian Rhythms in Isolated Brain Regions Michikazu Abe, 1 * Erik D. Herzog, 1 * Shin Yamazaki, 1 Marty Straume, 1 Hajime Tei, 2 Yoshiyuki
Molecular components of the mammalian circadian clock
 doi:10.1093/hmg/ddl207 R271 R277 Molecular components of the mammalian circadian clock Caroline H. Ko 1,3 and Joseph S. Takahashi 1,2, * 1 Department of Neurobiology and Physiology, 2 Howard Hughes Medical
doi:10.1093/hmg/ddl207 R271 R277 Molecular components of the mammalian circadian clock Caroline H. Ko 1,3 and Joseph S. Takahashi 1,2, * 1 Department of Neurobiology and Physiology, 2 Howard Hughes Medical
All mammalian cells investigated to date seem to possess
 Clock Genes in the Heart Characterization and Attenuation With Hypertrophy Martin E. Young, Peter Razeghi, Heinrich Taegtmeyer Abstract We investigated whether the heart, like other mammalian organs, possesses
Clock Genes in the Heart Characterization and Attenuation With Hypertrophy Martin E. Young, Peter Razeghi, Heinrich Taegtmeyer Abstract We investigated whether the heart, like other mammalian organs, possesses
Sleep-Wake Cycle I Brain Rhythms. Reading: BCP Chapter 19
 Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Peripheral Clocks: Keeping Up with the Master Clock
 Peripheral Clocks: Keeping Up with the Master Clock E. KOWALSKA AND S.A. BROWN University of Zurich, Institute for Pharmacology and Toxicology, 8057 Zurich, Switzerland Circadian clocks influence most
Peripheral Clocks: Keeping Up with the Master Clock E. KOWALSKA AND S.A. BROWN University of Zurich, Institute for Pharmacology and Toxicology, 8057 Zurich, Switzerland Circadian clocks influence most
Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v)
") SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
Molecular and Cellular Endocrinology
 Molecular and Cellular Endocrinology 349 (2012) 38 44 Contents lists available at ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce Review The role of clock
Molecular and Cellular Endocrinology 349 (2012) 38 44 Contents lists available at ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce Review The role of clock
Phase-resetting Responses in Clock Null Mutant Mice
 Phase-resetting Responses in Clock Null Mutant Mice Vinhfield X. Ta Program in Biological Sciences Northwestern University Evanston, Illinois May 7, 2007 Joseph S. Takahashi Senior Thesis Advisor Jason
Phase-resetting Responses in Clock Null Mutant Mice Vinhfield X. Ta Program in Biological Sciences Northwestern University Evanston, Illinois May 7, 2007 Joseph S. Takahashi Senior Thesis Advisor Jason
Eicosapentaenoic Acid Reverses the Oleic Acid-induced Reduction in Per1 mrna Expression in Cultured Rat Hepatocytes
 8 Ivyspring International Publisher Research Paper Journal of Biomedicine 2018; 3: 8-12. doi: 10.7150/jbm.23267 Eicosapentaenoic Acid Reverses the Oleic Acid-induced Reduction in Per1 mrna Expression in
8 Ivyspring International Publisher Research Paper Journal of Biomedicine 2018; 3: 8-12. doi: 10.7150/jbm.23267 Eicosapentaenoic Acid Reverses the Oleic Acid-induced Reduction in Per1 mrna Expression in
mcry1 and mcry2 Are Essential Components of the Negative Limb of the Circadian Clock Feedback Loop
 Cell, Vol. 98, 193 205, July 23, 1999, Copyright 1999 by Cell Press mcry1 and mcry2 Are Essential Components of the Negative Limb of the Circadian Clock Feedback Loop Kazuhiko Kume,* # Mark J. Zylka,*
Cell, Vol. 98, 193 205, July 23, 1999, Copyright 1999 by Cell Press mcry1 and mcry2 Are Essential Components of the Negative Limb of the Circadian Clock Feedback Loop Kazuhiko Kume,* # Mark J. Zylka,*
Entrainment of the circadian clock in humans: mechanism and implications for sleep disorders.
 Impulse: The Premier Journal for Undergraduate Publications in the Neurosciences Entrainment of the circadian clock in humans: mechanism and implications for sleep disorders. David Metcalfe Warwick Medical
Impulse: The Premier Journal for Undergraduate Publications in the Neurosciences Entrainment of the circadian clock in humans: mechanism and implications for sleep disorders. David Metcalfe Warwick Medical
Biological Clocks. Lu Chen, Ph.D. MCB, UC Berkeley. What is biological clock?
 Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 What is biological clock? All eukaryotes and some prokaryotes display changes in gene activity, biochemistry, physiology, and behavior that wax and wane
Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 What is biological clock? All eukaryotes and some prokaryotes display changes in gene activity, biochemistry, physiology, and behavior that wax and wane
Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells.
 SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells. A) Cirbp mrna expression levels in various mouse tissues collected around the clock
SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells. A) Cirbp mrna expression levels in various mouse tissues collected around the clock
Neurons and Hormones 1. How do animals perform the right behaviors at the right time? In the right context?
 Neurons and Hormones 1 How do animals perform the right behaviors at the right time? In the right context? Active at night only What if conflicting signals? Magnetic cues are always present But migrate
Neurons and Hormones 1 How do animals perform the right behaviors at the right time? In the right context? Active at night only What if conflicting signals? Magnetic cues are always present But migrate
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
The Nobel Assembly at Karolinska Institutet has today decided to award. the 2017 Nobel Prize in Physiology or Medicine. jointly to
 The Nobel Assembly at Karolinska Institutet has today decided to award the 2017 Nobel Prize in Physiology or Medicine jointly to Jeffrey C. Hall, Michael Rosbash and Michael W. Young for their discoveries
The Nobel Assembly at Karolinska Institutet has today decided to award the 2017 Nobel Prize in Physiology or Medicine jointly to Jeffrey C. Hall, Michael Rosbash and Michael W. Young for their discoveries
Time after time: inputs to and outputs from the mammalian circadian oscillators
 632 Review Time after time: inputs to and outputs from the mammalian circadian oscillators David Morse and Paolo Sassone-Corsi Oscillating levels of clock gene transcripts in the suprachiasmatic nucleus
632 Review Time after time: inputs to and outputs from the mammalian circadian oscillators David Morse and Paolo Sassone-Corsi Oscillating levels of clock gene transcripts in the suprachiasmatic nucleus
An Abrupt Shift in the Day/Night Cycle Causes Desynchrony in the Mammalian Circadian Center
 The Journal of Neuroscience, July 9, 2003 23(14):6141 6151 6141 Behavioral/Systems/Cognitive An Abrupt Shift in the Day/Night Cycle Causes Desynchrony in the Mammalian Circadian Center Mamoru Nagano, 1
The Journal of Neuroscience, July 9, 2003 23(14):6141 6151 6141 Behavioral/Systems/Cognitive An Abrupt Shift in the Day/Night Cycle Causes Desynchrony in the Mammalian Circadian Center Mamoru Nagano, 1
Scientific Background Discoveries of Molecular Mechanisms Controlling the Circadian Rhythm
 Scientific Background Discoveries of Molecular Mechanisms Controlling the Circadian Rhythm The 2017 Nobel Prize in Physiology or Medicine is awarded to Jeffrey C. Hall, Michael Rosbash and Michael W. Young
Scientific Background Discoveries of Molecular Mechanisms Controlling the Circadian Rhythm The 2017 Nobel Prize in Physiology or Medicine is awarded to Jeffrey C. Hall, Michael Rosbash and Michael W. Young
Review. The Network of Time: Understanding the Molecular Circadian System. Till Roenneberg and Martha Merrow
 Current Biology, Vol. 13, R198 R207, March 4, 2003, 2003 Elsevier Science Ltd. All rights reserved. PII S0960-9822(03)00124-6 The Network of Time: Understanding the Molecular Circadian System Review Till
Current Biology, Vol. 13, R198 R207, March 4, 2003, 2003 Elsevier Science Ltd. All rights reserved. PII S0960-9822(03)00124-6 The Network of Time: Understanding the Molecular Circadian System Review Till
Phenotypic Rescue of a Peripheral Clock Genetic Defect via SCN Hierarchical Dominance
 Cell, Vol. 110, 107 117, July 12, 2002, Copyright 2002 by Cell Press Phenotypic Rescue of a Peripheral Clock Genetic Defect via SCN Hierarchical Dominance Matthew P. Pando, 1,3 David Morse, 1,4 Nicolas
Cell, Vol. 110, 107 117, July 12, 2002, Copyright 2002 by Cell Press Phenotypic Rescue of a Peripheral Clock Genetic Defect via SCN Hierarchical Dominance Matthew P. Pando, 1,3 David Morse, 1,4 Nicolas
2 The abbreviations used are: per, period; tim, timeless; Cry, cryptochrome;
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 290, NO. 7, pp. 4367 4382, February 13, 2015 2015 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. The Zebrafish Period2
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 290, NO. 7, pp. 4367 4382, February 13, 2015 2015 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. The Zebrafish Period2
Mammalian Circadian Clock: The Roles of Transcriptional Repression and Delay
 Mammalian Circadian Clock: The Roles of Transcriptional Repression and Delay Yoichi Minami, Koji L. Ode, and Hiroki R. Ueda Abstract The circadian clock is an endogenous oscillator with a 24-h period.
Mammalian Circadian Clock: The Roles of Transcriptional Repression and Delay Yoichi Minami, Koji L. Ode, and Hiroki R. Ueda Abstract The circadian clock is an endogenous oscillator with a 24-h period.
Molecular Clocks in Mouse Skin
 See related commentary on pg ORIGINAL ARTICLE Molecular Clocks in Mouse Skin Miki Tanioka,,, Hiroyuki Yamada,, Masao Doi,, Hideki Bando, Yoshiaki Yamaguchi, Chikako Nishigori and Hitoshi Okamura, Clock
See related commentary on pg ORIGINAL ARTICLE Molecular Clocks in Mouse Skin Miki Tanioka,,, Hiroyuki Yamada,, Masao Doi,, Hideki Bando, Yoshiaki Yamaguchi, Chikako Nishigori and Hitoshi Okamura, Clock
Circadian Gene Expression in the Suprachiasmatic Nucleus
 Circadian Gene Expression in the Suprachiasmatic Nucleus 901 Circadian Gene Expression in the Suprachiasmatic Nucleus M U Gillette and S-H Tyan, University of Illinois at Urbana-Champaign, Urbana, IL,
Circadian Gene Expression in the Suprachiasmatic Nucleus 901 Circadian Gene Expression in the Suprachiasmatic Nucleus M U Gillette and S-H Tyan, University of Illinois at Urbana-Champaign, Urbana, IL,
From the reveller to the lark
 In Focus: Chronobiology From the reveller to the lark The internal clock changes with age Prof. Dr. Anne Eckert, Neurobiological Laboratory for Brain Aging and Mental Health, Psychiatric University Clinics
In Focus: Chronobiology From the reveller to the lark The internal clock changes with age Prof. Dr. Anne Eckert, Neurobiological Laboratory for Brain Aging and Mental Health, Psychiatric University Clinics
CHAPTER12. Synthesis
 CHAPTER12 Synthesis 149 Chapter 12 The tau mutation and non-circadian rhythms Biological rhythms cover a wide range of frequencies, from milliseconds to years. In this thesis we have shown that an allele
CHAPTER12 Synthesis 149 Chapter 12 The tau mutation and non-circadian rhythms Biological rhythms cover a wide range of frequencies, from milliseconds to years. In this thesis we have shown that an allele
Supplementary information
 Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone
 Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone Steven A. Thau MD Chief, Pulmonary, Sleep Department. Phelps Hospital, Northwell Health Internal Clock Examples
Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone Steven A. Thau MD Chief, Pulmonary, Sleep Department. Phelps Hospital, Northwell Health Internal Clock Examples
The Circadian Clock: Regulating the Rhythm of Life
 The Circadian Clock: Regulating the Rhythm of Life Christopher G. Gomez 1 1BIOC 462aH, University of Arizona Abstract The human body follows a daily cycle of sleep, of hunger which is independent of the
The Circadian Clock: Regulating the Rhythm of Life Christopher G. Gomez 1 1BIOC 462aH, University of Arizona Abstract The human body follows a daily cycle of sleep, of hunger which is independent of the
Parathyroid Hormone, But Not Melatonin, Resets The Bone Circadian Clock
 Parathyroid Hormone, But Not Melatonin, Resets The Bone Circadian Clock Naoki Okubo 1,2,3, Yoichi Minami 1,3, Hiroyoshi Fujiwara 2, Tatsuya Kunimoto 1,2,3, Toshihiro Hosokawa 1,2,3, Ryo Oda 2, Toshikazu
Parathyroid Hormone, But Not Melatonin, Resets The Bone Circadian Clock Naoki Okubo 1,2,3, Yoichi Minami 1,3, Hiroyoshi Fujiwara 2, Tatsuya Kunimoto 1,2,3, Toshihiro Hosokawa 1,2,3, Ryo Oda 2, Toshikazu
Altered Entrainment and Feedback Loop Function Effected by a Mutant Period Protein
 The Journal of Neuroscience, February 1, 2000, 20(3):958 968 Altered Entrainment and Feedback Loop Function Effected by a Mutant Period Protein Peter Schotland, Melissa Hunter-Ensor, Todd Lawrence, and
The Journal of Neuroscience, February 1, 2000, 20(3):958 968 Altered Entrainment and Feedback Loop Function Effected by a Mutant Period Protein Peter Schotland, Melissa Hunter-Ensor, Todd Lawrence, and
Daily injection of insulin attenuated impairment of liver circadian clock oscillation in the streptozotocin-treated diabetic mouse
 FEBS Letters 572 (2004) 206 210 FEBS 28689 Daily injection of insulin attenuated impairment of liver circadian clock oscillation in the streptozotocin-treated diabetic mouse Koji Kuriyama, Kei Sasahara,
FEBS Letters 572 (2004) 206 210 FEBS 28689 Daily injection of insulin attenuated impairment of liver circadian clock oscillation in the streptozotocin-treated diabetic mouse Koji Kuriyama, Kei Sasahara,
Circadian Molecular Clocks Tick along Ontogenesis
 Physiol. Res. 57 (Suppl. 3): S139-S148, 2008 MINIREVIEW Circadian Molecular Clocks Tick along Ontogenesis A. SUMOVÁ, Z. BENDOVÁ, M. SLÁDEK, R. EL-HENNAMY, K. MATĚJŮ, L. POLIDAROVÁ, S. SOSNIYENKO, H. ILLNEROVÁ
Physiol. Res. 57 (Suppl. 3): S139-S148, 2008 MINIREVIEW Circadian Molecular Clocks Tick along Ontogenesis A. SUMOVÁ, Z. BENDOVÁ, M. SLÁDEK, R. EL-HENNAMY, K. MATĚJŮ, L. POLIDAROVÁ, S. SOSNIYENKO, H. ILLNEROVÁ
Circular RNAs (circrnas) act a stable mirna sponges
 act a stable mirna sponges") Circular RNAs (circrnas) act a stable mirna sponges cernas compete for mirnas Ancestal mrna (+3 UTR) Pseudogene RNA (+3 UTR homolgy region) The model holds true for all RNAs that share a mirna binding
Circular RNAs (circrnas) act a stable mirna sponges cernas compete for mirnas Ancestal mrna (+3 UTR) Pseudogene RNA (+3 UTR homolgy region) The model holds true for all RNAs that share a mirna binding
The Suprachiasmatic Nucleus Entrains, But Does Not Sustain, Circadian Rhythmicity in the Olfactory Bulb
 The Journal of Neuroscience, January 21, 2004 24(3):615 619 615 Brief Communication The Suprachiasmatic Nucleus Entrains, But Does Not Sustain, Circadian Rhythmicity in the Olfactory Bulb Daniel Granados-Fuentes,
The Journal of Neuroscience, January 21, 2004 24(3):615 619 615 Brief Communication The Suprachiasmatic Nucleus Entrains, But Does Not Sustain, Circadian Rhythmicity in the Olfactory Bulb Daniel Granados-Fuentes,
The tim SL Mutant Affects a Restricted Portion of the Drosophila melanogaster Circadian Cycle
 JO RutURNAL ila et al. OF / tim BIO SL LOGICAL MUTANT RHYTHMS / October 1998 The tim SL Mutant Affects a Restricted Portion of the Drosophila melanogaster Circadian Cycle Joan E. Rutila, * Olga Maltseva,
JO RutURNAL ila et al. OF / tim BIO SL LOGICAL MUTANT RHYTHMS / October 1998 The tim SL Mutant Affects a Restricted Portion of the Drosophila melanogaster Circadian Cycle Joan E. Rutila, * Olga Maltseva,
Cell Type-Specific Functions of Period Genes Revealed by Novel Adipocyte and Hepatocyte Circadian Clock Models
 Cell Type-Specific Functions of Period Genes Revealed by Novel Adipocyte and Hepatocyte Circadian Clock Models Chidambaram Ramanathan 1, Haiyan Xu 1, Sanjoy K. Khan 1, Yang Shen 1, Paula J. Gitis 2,3,
Cell Type-Specific Functions of Period Genes Revealed by Novel Adipocyte and Hepatocyte Circadian Clock Models Chidambaram Ramanathan 1, Haiyan Xu 1, Sanjoy K. Khan 1, Yang Shen 1, Paula J. Gitis 2,3,
Key words: antisense oligonucleotide; circadian rhythm; firing rhythm; mper1; phase shift; suprachiasmatic nucleus
 The Journal of Neuroscience, February 1, 1999, 19(3):1115 1121 Inhibition of Light- or Glutamate-Induced mper1 Expression Represses the Phase Shifts into the Mouse Circadian Locomotor and Suprachiasmatic
The Journal of Neuroscience, February 1, 1999, 19(3):1115 1121 Inhibition of Light- or Glutamate-Induced mper1 Expression Represses the Phase Shifts into the Mouse Circadian Locomotor and Suprachiasmatic
Circadian Gene Expression in Individual Fibroblasts: Cell-Autonomous and Self-Sustained Oscillators Pass Time to Daughter Cells
 Cell, Vol. 119, 693 705, November 24, 2004, Copyright 2004 by Cell Press Circadian Gene Expression in Individual Fibroblasts: Cell-Autonomous and Self-Sustained Oscillators Pass Time to Daughter Cells
Cell, Vol. 119, 693 705, November 24, 2004, Copyright 2004 by Cell Press Circadian Gene Expression in Individual Fibroblasts: Cell-Autonomous and Self-Sustained Oscillators Pass Time to Daughter Cells
MOLECULAR BASES OF CIRCADIAN RHYTHMS
 Annu. Rev. Cell Dev. Biol. 2001. 17:215 53 Copyright c 2001 by Annual Reviews. All rights reserved MOLECULAR BASES OF CIRCADIAN RHYTHMS Stacey L. Harmer, Satchidananda Panda, and Steve A. Kay Department
Annu. Rev. Cell Dev. Biol. 2001. 17:215 53 Copyright c 2001 by Annual Reviews. All rights reserved MOLECULAR BASES OF CIRCADIAN RHYTHMS Stacey L. Harmer, Satchidananda Panda, and Steve A. Kay Department
The Ticking CLOCK of HSV-2 Pathology Rebecca J. Bayliss 1 and Vincent Piguet 1,2,3
 The Ticking CLOCK of HSV-2 Pathology Rebecca J. Bayliss 1 and Vincent Piguet 1,2,3 1 Division of Infection and Immunity, Cardiff University School of Medicine, Cardiff, UK; 2 Division of Dermatology, Women
The Ticking CLOCK of HSV-2 Pathology Rebecca J. Bayliss 1 and Vincent Piguet 1,2,3 1 Division of Infection and Immunity, Cardiff University School of Medicine, Cardiff, UK; 2 Division of Dermatology, Women
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
N.T.Kumar*,Thiago R,Sandro B,Vinicius R,Lisane V,Vanderlei P Versor Inovação,Santo André SP,Brazil. id* :
 General Theoretical Aspects of Cell Mechanics from Circadian Oscillator Point of View Based on Higher Order Logic(HOL) A Short Communication to Develop Informatics Framework Involving Bio-Chemical Concepts
General Theoretical Aspects of Cell Mechanics from Circadian Oscillator Point of View Based on Higher Order Logic(HOL) A Short Communication to Develop Informatics Framework Involving Bio-Chemical Concepts
Nonredundant Roles of the mper1 and mper2 Genes in the Mammalian Circadian Clock
 Cell, Vol. 105, 683 694, June 1, 2001, Copyright 2001 by Cell Press Nonredundant Roles of the mper1 and mper2 Genes in the Mammalian Circadian Clock Binhai Zheng, 1,5 Urs Albrecht, 2,7 Krista Kaasik, 1
Cell, Vol. 105, 683 694, June 1, 2001, Copyright 2001 by Cell Press Nonredundant Roles of the mper1 and mper2 Genes in the Mammalian Circadian Clock Binhai Zheng, 1,5 Urs Albrecht, 2,7 Krista Kaasik, 1
Representing Time in Scientific Diagrams
 Representing Time in Scientific Diagrams William Bechtel (bechtel@ucsd.edu) Daniel Burnston (dburnston@ucsd.edu) Benjamin Sheredos (bsheredos@ucsd.edu) Department of Philosophy and Center for Circadian
Representing Time in Scientific Diagrams William Bechtel (bechtel@ucsd.edu) Daniel Burnston (dburnston@ucsd.edu) Benjamin Sheredos (bsheredos@ucsd.edu) Department of Philosophy and Center for Circadian
V3: Circadian rhythms program for today
 V3: Circadian rhythms program for today (1) Case study: circadian effects on drug response (2) Circadian rhythms are closely connected to metabolism (3) Circadian clock genes generate epigenetic effects
V3: Circadian rhythms program for today (1) Case study: circadian effects on drug response (2) Circadian rhythms are closely connected to metabolism (3) Circadian clock genes generate epigenetic effects
Modeling the circadian clock: from molecular mechanism to physiological disorders
 Modeling the circadian clock: from molecular mechanism to physiological disorders Jean-Christophe Leloup* and Albert Goldbeter Summary Based on genetic and biochemical advances on the molecular mechanism
Modeling the circadian clock: from molecular mechanism to physiological disorders Jean-Christophe Leloup* and Albert Goldbeter Summary Based on genetic and biochemical advances on the molecular mechanism
The Drosophila melanogaster life cycle
 The Drosophila melanogaster life cycle Eclosion The first phenotype described in Drosophila as an endogenous rhythm (1954): Number of eclosed flies Hamblen et al., Gene3cs 1998 rhythm with a periodicity
The Drosophila melanogaster life cycle Eclosion The first phenotype described in Drosophila as an endogenous rhythm (1954): Number of eclosed flies Hamblen et al., Gene3cs 1998 rhythm with a periodicity
Smooth-Muscle BMAL1 Participates in Blood Pressure Circadian Rhythm Regulation
 University of Kentucky UKnowledge Physiology Faculty Publications Physiology 1-2015 Smooth-Muscle BMAL1 Participates in Blood Pressure Circadian Rhythm Regulation Zhongwen Xie University of Kentucky, zxie1@uky.edu
University of Kentucky UKnowledge Physiology Faculty Publications Physiology 1-2015 Smooth-Muscle BMAL1 Participates in Blood Pressure Circadian Rhythm Regulation Zhongwen Xie University of Kentucky, zxie1@uky.edu
SERIES ARTICLE Circadian Rhythms
 Circadian Rhythms 3. Circadian Timing Systems: How are they Organized? Koustubh M Vaze and Vijay Kumar Sharma (left) Koustubh Vaze is a Research Associate in the Evolutionary and Organismal Biology Unit,
Circadian Rhythms 3. Circadian Timing Systems: How are they Organized? Koustubh M Vaze and Vijay Kumar Sharma (left) Koustubh Vaze is a Research Associate in the Evolutionary and Organismal Biology Unit,
SUPPLEMENTAL DATA. Lumen area ( m 2 )
") Elastin Lumen area ( m 2 ) Media to lumen ratio (x1) H.E. Medium thickness ( m) Medium area ( m 2 ) SUPPLEMENTAL DATA A (Bmal1 flox/flox ) (SM-Bmal1 -/- ) B 1 8 8 6 6 4 4 2 2 1µm 5 8 4 6 3 2 4 1 2 Supplemental
Elastin Lumen area ( m 2 ) Media to lumen ratio (x1) H.E. Medium thickness ( m) Medium area ( m 2 ) SUPPLEMENTAL DATA A (Bmal1 flox/flox ) (SM-Bmal1 -/- ) B 1 8 8 6 6 4 4 2 2 1µm 5 8 4 6 3 2 4 1 2 Supplemental
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
Food-entrained circadian rhythms are sustained in arrhythmic Clk/Clk mutant mice
 Am J Physiol Regul Integr Comp Physiol 285: R57 R67, 2003. First published March 20, 2003; 10.1152/ajpregu.00023.2003. Food-entrained circadian rhythms are sustained in arrhythmic Clk/Clk mutant mice SiNae
Am J Physiol Regul Integr Comp Physiol 285: R57 R67, 2003. First published March 20, 2003; 10.1152/ajpregu.00023.2003. Food-entrained circadian rhythms are sustained in arrhythmic Clk/Clk mutant mice SiNae
Biological rhythms. Types of biological rhythms
 Biological rhythms Types of biological rhythms 2/33 what do we call rhythm in a living organism? physiological events occurring at approximately regular times internally controlled rhythms: breathing,
Biological rhythms Types of biological rhythms 2/33 what do we call rhythm in a living organism? physiological events occurring at approximately regular times internally controlled rhythms: breathing,
Diagrams as Vehicles for Scientific Reasoning. Adele Abrahamsen Center for Research in Language University of California, San Diego
 Diagrams as Vehicles for Scientific Reasoning William Bechtel Department of Philosophy, Center for Chronobiology, and Interdisciplinary Program in Cognitive Science University of California, San Diego
Diagrams as Vehicles for Scientific Reasoning William Bechtel Department of Philosophy, Center for Chronobiology, and Interdisciplinary Program in Cognitive Science University of California, San Diego
Molecular Architecture of the Circadian Clock in Mammals
 Molecular Architecture of the Circadian Clock in Mammals Joseph S. Takahashi Abstract The circadian clock mechanism in animals involves an autoregulatory transcriptional feedback loop in which CLOCK and
Molecular Architecture of the Circadian Clock in Mammals Joseph S. Takahashi Abstract The circadian clock mechanism in animals involves an autoregulatory transcriptional feedback loop in which CLOCK and
Entrainment of the mouse circadian clock by sub-acute physical and psychological
 Supplementary Information Entrainment of the mouse circadian clock by sub-acute physical and psychological Yu Tahara 1, Takuya Shiraishi 1, Yosuke Kikuchi 1, Atsushi Haraguchi 1, Daisuke Kuriki 1, Hiroyuki
Supplementary Information Entrainment of the mouse circadian clock by sub-acute physical and psychological Yu Tahara 1, Takuya Shiraishi 1, Yosuke Kikuchi 1, Atsushi Haraguchi 1, Daisuke Kuriki 1, Hiroyuki
Methods in Enzymology. Volume 393. Circadian Rhythms. Michael W. Young
 Methods in Enzymology Volume 393 Circadian Rhythms Michael W. Young CONTRIBUTORS TO VOLUME 393 PREFACE VOLUME IN SERIES x i XVI I xix Section I. Genetic Approaches to Circadian Clocks 1. Analysis of Circadian
Methods in Enzymology Volume 393 Circadian Rhythms Michael W. Young CONTRIBUTORS TO VOLUME 393 PREFACE VOLUME IN SERIES x i XVI I xix Section I. Genetic Approaches to Circadian Clocks 1. Analysis of Circadian
Systems Biology of Mammalian Circadian Clocks
 Annu. Rev. Physiol. 2010. 72:579 603 First published online as a Review in Advance on November 13, 2009 The Annual Review of Physiology is online at physiol.annualreviews.org This article s doi: 10.1146/annurev-physiol-073109-130051
Annu. Rev. Physiol. 2010. 72:579 603 First published online as a Review in Advance on November 13, 2009 The Annual Review of Physiology is online at physiol.annualreviews.org This article s doi: 10.1146/annurev-physiol-073109-130051
Deletion of the Mammalian Circadian Clock Gene BMAL1/Mop3 Alters Baseline Sleep Architecture and the Response to Sleep Deprivation
 CIRCADIAN RHYTHMS Deletion of the Mammalian Circadian Clock Gene BMAL1/Mop3 Alters Baseline Sleep Architecture and the Response to Sleep Deprivation Aaron Laposky, PhD 1 ; Amy Easton, PhD 1 ; Christine
CIRCADIAN RHYTHMS Deletion of the Mammalian Circadian Clock Gene BMAL1/Mop3 Alters Baseline Sleep Architecture and the Response to Sleep Deprivation Aaron Laposky, PhD 1 ; Amy Easton, PhD 1 ; Christine
Drosophila Photoreceptors Contain an Autonomous Circadian Oscillator That Can Function without period mrna Cycling
 The Journal of Neuroscience, January 15, 1998, 18(2):741 750 Drosophila Photoreceptors Contain an Autonomous Circadian Oscillator That Can Function without period mrna Cycling Yuzhong Cheng 1 and Paul
The Journal of Neuroscience, January 15, 1998, 18(2):741 750 Drosophila Photoreceptors Contain an Autonomous Circadian Oscillator That Can Function without period mrna Cycling Yuzhong Cheng 1 and Paul
There is growing recognition of the critical contribution
 Glucocorticoid Signaling Synchronizes the Liver Circadian Transcriptome Akhilesh B. Reddy, 1 Elizabeth S. Maywood, 1 Natasha A. Karp, 2 Verdun M. King, 3 Yusuke Inoue, 4 Frank J. Gonzalez, 4 Kathryn S.
Glucocorticoid Signaling Synchronizes the Liver Circadian Transcriptome Akhilesh B. Reddy, 1 Elizabeth S. Maywood, 1 Natasha A. Karp, 2 Verdun M. King, 3 Yusuke Inoue, 4 Frank J. Gonzalez, 4 Kathryn S.
In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described
 Supplemental Methods In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described previously 1 2 using 32P-labeled 211 bp fragment from 3 HS1. Footprinting reaction mixes contained
Supplemental Methods In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described previously 1 2 using 32P-labeled 211 bp fragment from 3 HS1. Footprinting reaction mixes contained
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION
 Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
Revisiting existing drugs finds molecules that control body clocks ~ Discovery of an anti-aging supplement that reduces jet lag in mice ~
 Revisiting existing drugs finds molecules that control body clocks ~ Discovery of an anti-aging supplement that reduces jet lag in mice ~ April 17, 2018 Using drug repurposing, a team of researchers led
Revisiting existing drugs finds molecules that control body clocks ~ Discovery of an anti-aging supplement that reduces jet lag in mice ~ April 17, 2018 Using drug repurposing, a team of researchers led
RESEARCH ARTICLE Significant dissociation of expression patterns of the basic helix loop helix transcription factors Dec1 and Dec2 in rat kidney
 1257 The Journal of Experimental Biology 214, 1257-1263 2011. Published by The Company of Biologists Ltd doi:10.1242/jeb.052100 RESEARCH ARTICLE Significant dissociation of expression patterns of the basic
1257 The Journal of Experimental Biology 214, 1257-1263 2011. Published by The Company of Biologists Ltd doi:10.1242/jeb.052100 RESEARCH ARTICLE Significant dissociation of expression patterns of the basic
University of Groningen. Life before the Clock Roenneberg, Till; Merrow, Martha. Published in: Journal of Biological Rhythms
 University of Groningen Life before the Clock Roenneberg, Till; Merrow, Martha Published in: Journal of Biological Rhythms DOI: 10.1177/0748730402238231 IMPORTANT NOTE: You are advised to consult the publisher's
University of Groningen Life before the Clock Roenneberg, Till; Merrow, Martha Published in: Journal of Biological Rhythms DOI: 10.1177/0748730402238231 IMPORTANT NOTE: You are advised to consult the publisher's