A Listeria-based vaccine that secretes the sand fly salivary protein LJM11 confers long-term protection
|
|
|
- Logan Barrett
- 5 years ago
- Views:
Transcription
1 IAI Accepts, published online ahead of print on 14 April 2014 Infect. Immun. doi: /iai Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 A Listeria-based vaccine that secretes the sand fly salivary protein LJM11 confers long-term protection against vector-transmitted Leishmania major Delbert S. Abi Abdallah a, Alan Pavinski Bitar a, Fabiano Oliveira b, Claudio Meneses b, Justin J. Park a, Susana Mendez a,c *, Shaden Kamhawi b, Jesus G. Valenzuela b, and Hélène Marquis a# Department of Microbiology and Immunology, Cornell University, Ithaca, New York, USA a ; Vector Molecular Biology Section, Laboratory of Malaria and Vector Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Rockville, Maryland, USA b ; Baker Institute for Animal Health, Cornell University, Ithaca, New York, USA c Running title: Listeria-based vaccine against cutaneous leishmaniasis #Address correspondance to: Hélène Marquis, hm72@cornell.edu *Present address : National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, Maryland, USA
2 Abstract Cutaneous leishmaniasis is a sand fly transmitted disease characterized by skin ulcers that carry significant scarring and social stigmatization. Over the past years, there has been cumulative evidence that immunity to specific sand fly salivary proteins confers a significant level of protection against leishmaniasis. In this study, we used an attenuated strain of Listeria monocytogenes as a vaccine expression system for LJM11, a sand fly salivary protein identified as a good vaccine candidate. We observed that mice were best protected against an intradermal needle challenge with Leishmania major and sand fly saliva when vaccinated intravenously. However, this protection was short-term. Importantly, groups of vaccinated mice were protected long-term when challenged with infected sand flies. Protection correlated with smaller lesion size, less scars, and better parasite control between 2 and 6 weeks post-challenge when compared to the control group of mice vaccinated with the parent L. monocytogenes strain not expressing LJM11. Moreover, protection correlated with high numbers of CD4 + IFN-γ + TNF-α +/- IL-10 - and low numbers of CD4 + IFN-γ +/- TNF-α - IL-10 + T cells at two weeks postchallenge. Overall, our data indicate that delivery of LJM11 by Listeria is a promising vaccination strategy against cutaneous leishmaniasis inducing long-term protection against ulcer formation following a natural challenge with infected sand flies. 2
3 36 INTRODUCTION Leishmaniasis is a disease that is endemic in 98 countries of which most are developing countries (1). There are approximately 0.9 to 1.6 million new cases annually and 350 million people at risk of getting infected. The most common form of the disease is cutaneous leishmaniasis, which is characterized by skin ulcers. The lesions commonly self-cure but can leave significant scarring and lead to social stigmatization. The large majority of cases of cutaneous leishmaniasis occur in Afghanistan, Iran, Syria, Algeria, Brazil and Colombia (1). Leishmania spp. are protozoans that are transmitted by phlebotomine sand flies. During feeding, the fly injects saliva into its host. Some of the saliva proteins induce vasodilation and prevent coagulation to facilitate feeding, while others have immunomodulatory effects (2). There is strong evidence that the fly saliva enhances the ability of Leishmania to establish an infection by about a thousand fold (3, 4). Conversely, the development of a delayed-type hypersensitivity (DTH) response to salivary components from uninfected sand flies compromises the ability of Leishmania to establish an infection (5, 6). The immunoprotective value of pre-exposure to the saliva of uninfected sand flies is further emphasized by the observation that the influx of refugees, humanitarian aid workers, tourists, and military personnel in locations with previously low levels of infection leads to dramatic outbreaks of leishmaniasis (7-15). Following these observations, specific salivary proteins have been identified as potent protective immunogens against the establishment of an infection with Leishmania and are being considered as candidates for an anti-leishmania vaccine (16-19). Infected sand flies harbor the flagellated metacyclic promastigote form of Leishmania, which is transmitted into the dermis of a mammalian host during a blood meal. These parasites infect cells, predominantly professional phagocytes such as macrophages, in which they differentiate into nonflagellated amastigotes that divide and proliferate in parasitophorous vacuoles. Interestingly, the initial 3
4 infection does not induce an inflammatory response thus enabling the parasite to expand in numbers (20). However, it has been well documented that resistance to Leishmania correlates with the development of a T H 1 type of immune response driven primarily by interferon gamma (IFN-γ)-producing CD4 + T cells (21). In this work, we used Listeria monocytogenes as a vector to vaccinate against a sand fly salivary protein. L. monocytogenes is an intracellular bacterial pathogen that has the ability of proliferating in the cytosol of infected cells and of spreading from cell to cell without exiting the intracellular milieu (22). Consequently, clearance of an infection is completely cell-mediated (23) and is dependent on the development of a T H 1 immune response that is dominated by IFN-γ-secreting T cells (24). Over the past decade, there has been a strong interest in using L. monocytogenes as a vaccine vector because of its many useful characteristics (25-27). The bacterium can easily be attenuated without affecting its immunogenicity and can secrete large amounts of heterologous proteins in infected cells, leading to the generation of a specific cell-mediated immune response against that heterologous antigen. Therefore, we generated an attenuated Listeria-based vector that expresses a gene coding for the salivary protein LJM11 from the sand fly Lutzomyia longipalpis to assess the efficacy of a Listeria-based vaccine against cutaneous leishmaniasis. We observed a significant decrease in disease burden in mice vaccinated intravenously with Listeria secreting LJM11 and challenged by needle injection with purified parasites and sand fly saliva, or with infected sand flies. Protection was associated with high numbers of CD4 + IFNγ + TNF-α ± IL-10 - and low numbers of CD4 + IFN-γ ± TNF-α - IL-10 + T cells at two weeks post-challenge. MATERIALS AND METHODS Vaccine strain. L. monocytogenes strain DP-L1942, which carries a deletion in the acta gene was used to alleviate the safety concerns regarding a Listeria-based vaccine (28). ActA is a surface protein that mediates actin polymerization, enabling L. monocytogenes to move in the cytosol of host cells and to 4
5 spread from cell to cell without escaping the intracellular milieu (29). The acta deletion mutant is as efficient as the wild-type strain at escaping vacuoles formed upon initial entry into cells and at multiplying in the cytosol of infected cells. However, it is incapable of spreading from cell to cell without escaping from the intracellular milieu, resulting in a 3-log attenuation defect in a mouse model of infection (30). The acta deletion strain confers protective immunity at the same level as the wild-type strain. In an effort to improve vaccine safety and efficacy, the mdrm gene was deleted. This gene codes for a multiple drug resistance transporter that contributes to the activation of Type I interferon in infected cells (31). This innate immune response to a cytosolic infection by L. monocytogenes decreases the ability of the host to control the infection (32, 33). An internal in-frame deletion of the mdrm gene was introduced in strain DP-L1942 by allelic exchange as described (34), generating the control strain HEL The vaccine strain (HEL-1325) was created by introducing in HEL-1207 a construct comprised of the sequences coding for the ActA signal sequence and N-terminal (100 aa) followed by a FLAG tag and the gene coding for LJM11 minus its signal sequence (Fig. 1A). Secretion of LJM11 by intracellular bacteria was verified by pulse-labeling infected J774 mouse macrophage-like cells as described (35). Fitness of the recombinant strains was verified by performing intracellular growth curves in J774 cells as previously described (36). Mice and sand flies. Six to eight week old C57BL/6 mice were used for this study. Lutzomyia longipalpis sand flies were reared at the Laboratory of Malaria and Vector Research (LMVR), NIAID, NIH. Four to seven day old adult flies were dissected to obtain salivary glands, which were stored in PBS at -80 C. Salivary gland homogenates (SGH) were prepared by sonication. Lysates were cleared by centrifugation. When used for animal infections, sand flies were pre-infected with L. major promastigotes as described (6). 5
6 Parasites. Leishmania major WR 2885, a recent isolate originating from Iraq and typed at the Walter Reed Army Institute of Research, was used for this vaccine study (16). Frozen aliquots of L. major promastigotes were thawed, passaged in vitro once or twice and allowed to grow to stationary phase. Parasites were pelleted, washed once and re-suspended in DMEM. A discontinuous Ficoll gradient was prepared with 20% and 10% Ficoll type 400 in MilliQ water, and overlaid with the parasite suspension. After centrifugation at 2000 rpm for 10 min with slow deceleration, the metacyclic promastigotes were recovered above the 10-20% interface. The parasites were washed in DMEM and counts were determined using a hemacytometer. A final suspension was prepared in PBS. Crude Leishmania antigen (CLA). Parasites grown in flasks, as described above, were pelleted by centrifugation, washed three times in PBS and freeze-thawed multiple times to generate a crude lysate. Protein content was measured using a BCA assay and aliquots of CLA in PBS were stored at -80 C. Mouse experiments. Mice were anesthetized with isoflurane and vaccinated intradermally (ID) in the pinna of the ear (10 μl), intramuscularly (IM) in the hamstring (10 μl), subcutaneously at the base of the neck (100 μl), or intravenously in the retro-orbital sinus (100 μl). The vaccination dose was 5 x 10 6 CFU, three times at two-week intervals. Mice were challenged either 3 or 12 weeks after the last boost to assess short and long-term protection, respectively. Anesthetized mice were challenged intradermally in the pinna of the ear with 500 metacyclic promastigotes and SGH (equivalent to half a pair of salivary glands) in a total volume of 10 μl. Sand fly challenges were performed at the NIAID LMVR animal facility as described (6). Ear lesions were measured on a weekly basis starting at day 7 post-challenge. Needle challenges generate single ear lesions that were measured for thickness and diameter. These measurements were transformed into lesion area. Sand fly challenges generate multiple lesions that sometime coalesce. For this reason, we measured the thickness of the biggest ear lesion. 6
7 Processing of infected ears and flow cytometry experiments. Ear pinna dermal sheets were collected, treated with ethanol, separated and digested in 500 μl of DMEM containing Liberase TL (250 μg/ml, Roche) at 37 C for 1.5 hours. Digested tissues were minced and homogenized. Ear cells were washed with complete tissue culture medium (CTCM: DMEM, 10% FBS, 1 mm Na Pyruvate, 0.1 mm nonessential amino acids, 2 mm L-Glutamine, 30 mm Hepes, and 50 μm -mercaptoethanol). The resulting suspension was filtered, spun down and resuspended in CTCM. Parasite loads in the challenged ears were determined by limiting dilution assays as described (37). Parasite titers were determined from the highest dilution at which growth was visible. Ear cells were mixed with CLA-pre-pulsed bone marrow-derived dendritic cells (DC) to stimulate cytokine production by immune cells, improving the sensitivity of FACS analysis. Briefly, DC were prepared as previously described (38) and incubated with or without CLA (25 μg/ml) for 6 to 7 hours before adding ear cells at a ratio of 1 DC per 5 ear cells. After an overnight incubation, cells were treated for 4 hours with Brefeldin A (10 μg/ml) and PMA (5 ng/ml), collected in FACS buffer (10% normal mouse serum, 0.5% BSA, 0.02%NaN 3 in PBS), and incubated with antibodies against surface and intracellular markers: CD4-APC-H7, CD8a-APC, TNF-PerCP.Cy5.5, IFN-PECy7, and IL10-PE. Cells were collected on a BD Canto flow cytometer and analyzed using FlowJo software. Isotype controls and representative plots of CD4+ T cells stained for IFNγ, TNFα, and IL-10 are shown in figure S1. Integrated median fluorescence intensities (imfi) were calculated for the cells by multiplying the MFI with the frequency of cells producing for the particular cytokine and the data are reported in table S1. Data and statistical analyses. Ratios were obtained by comparing values from each individual mouse from a vaccine group to the calculated mean from the corresponding control group. Statistical analyses were performed using a two-tailed, unpaired, t-test or a One-Way Anova test with Bonferroni post-test analysis. 7
8 Ethics statement. All animal experimental procedures were reviewed and approved by Cornell University Institutional Animal Care and Use Committee, NIAID Animal Care and Use Committee, and US Army Medical Research and Material Command Animal Care and Use Review Office. All animal experiments were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Research Council. RESULTS L. monocytogenes vaccine vector confers the greatest protection against cutaneous leishmaniasis when administered intravenously. We generated an attenuated Listeria vaccine strain (HEL-1325) in which LJM11 was fused to the N-terminus of the Listeria ActA protein (Fig. 1A). This strain secreted large amounts of the fusion protein (55.4 kda) in infected cells (Fig. 1B), and had an intracellular growth rate that was similar to that of the parent strain (Fig. 1C). This attenuated Listeria vaccine strain persisted for several days in tissues of infected mice, inducing a specific T cell immune response against LJM11 (Fig. S2, 1D and S3). Initially, various routes of vaccine delivery were tested to identify parameters conferring the best level of protection in a mouse model of cutaneous leishmaniasis (3). Groups of mice were vaccinated intravenously, intradermally, subcutaneously, or intramuscularly, and challenged intradermally in the ear pinna with purified promastigotes and sand fly SGH. An age-matched unvaccinated group of mice was included in the challenge. Mice vaccinated intravenously, intradermally and intramuscularly had diminished lesion development when compared to unvaccinated mice (Fig. 2A). These differences were statistically significant at all time points for the intravenous group (p values between and <0.0001), between 4 and 11 weeks post-challenge for the intradermal group (p values between and ), and at 8 and 10 weeks post-challenge for the intramuscular group (p values of and 8
9 ). Mice vaccinated by the subcutaneous route developed lesions that were similar in size to the unvaccinated group The data were also analyzed by calculating the area under the curve for each individual mouse as a measure of disease burden over the duration of the entire study (Fig. 2B). The group vaccinated intravenously suffered a disease burden that was about three folds less than the unvaccinated and subcutaneously vaccinated groups, and about two folds less than the intradermally and intramuscularly vaccinated groups. The overall protection was highly significant for the group vaccinated intravenously (p <0.001) when compared to the group vaccinated subcutaneously and to the control group. Protection was also significant for the group vaccinated intradermally (p <0.05) when compared to the control group. The experiment described above did not include a vaccine vector control for any of the four routes of vaccination tested. To test the possibility that protection conferred by intravenous vaccination was due to a non-specific immune response to the vaccine vector, we performed an experiment in which we vaccinated mice intravenously with the vaccine vector (HEL-1207) or with PBS. Mice were challenged as described three weeks post-vaccination with L. major and sand fly saliva, and lesion areas were recorded over a period of 11 weeks. The results show no difference between these two groups (Fig. 2C), suggesting that the L. monocytogenes vaccine vector does not confer protection against cutaneous leishmaniasis. These data taken together indicate that while most routes of vaccination confer a certain level of protection against challenge, the intravenous route of vaccination confers the greatest protection Vaccinated mice are protected short-term against a needle challenge. In this next set of experiments, mice were vaccinated intravenously with either L. monocytogenes expressing LJM11 (HEL-1325=Vaccine group) or the isogenic L. monocytogenes strain not expressing LJM11 (HEL-1207=Control group) to 9
10 assess the specificity of the protective immune response. Short-term immune protection was assessed by challenging the two groups of mice intradermally three weeks after the last boost either with purified parasites mixed with sand fly SGH (needle challenge) or infected Lutzomyia longipalpis sand flies (sand fly challenge). Ear lesion sizes were recorded weekly. Because a needle challenge generates a single ear lesion, we measured lesion thickness and diameter, and transformed these measurements into lesion area. Contrarily, a sand fly challenge generates multiple lesions that sometime coalesce. For this reason, we measured the thickness of the biggest ear lesion. In addition to measuring lesion sizes, a subset of mice from each group was sacrificed at 2, 6, and 11 weeks post-challenge to determine parasite counts and characterize the immune response in the infected ears. Results from the short-term needle (STN) challenge groups indicated that mice in the vaccine group developed smaller lesions than those in the control group between 3 and 10 weeks post-challenge (p values between and <0.0001) (Fig. 3A), concurrently exhibiting a significantly smaller disease burden (p <0.0001) (Fig. 3B). Ear lesions were monitored carefully for the development of scars or ulcers and those that developed either over the duration of the experiment were scored. Five percent of mice in the vaccine group and 55% of mice in the control group developed scars or ulcers (p=0.010) (Fig. 3C). Ear parasite numbers were about the same in both groups of mice at 2 and 11 weeks post-challenge (Fig. 3D). However, between 2 and 6 weeks post-challenge, ear parasite numbers increased by three logs in the control group as opposed to one log in the vaccine group. The difference in ear parasite numbers between vaccine and control groups at 6 weeks post-challenge was statistically significant (p=0.0011). Overall, results from the STN challenge indicated that the vaccine is effective in decreasing overall disease burden, preventing the formation of scars, and decreasing parasite replication between 2 and 6 weeks post-challenge. 10
11 Results from the short-term sand fly (STSF) challenge groups indicated that mice developed similar size lesions in vaccine and control groups (Fig. 3E) with no significant difference in disease burden (Fig. 3F). Scars developed in 65 and 40 percent of the mice in control and vaccine groups respectively (Fig. 3G), but this difference was not statistically significant. Ear parasite numbers were approximately 1.5 logs higher in the vaccine group compared to the control group at 2 weeks post-challenge (Fig. 3H). However, between 2 and 6 weeks post-challenge, ear parasite numbers increased by nearly three logs in the control group as opposed to one log in the vaccine group. By 11 weeks post-challenge, ear parasite numbers were similar in both groups of mice. Overall, results from the STSF challenge indicated that the vaccine did not confer protection, although the vaccine group controlled ear parasite numbers much more efficiently than the control group between 2 and 6 weeks post-challenge. Vaccinated mice are protected long-term against a sand fly challenge. Long-term protection assays were performed by challenging mice 12 weeks post-vaccination. Results from the long-term needle (LTN) challenge indicated that the vaccine group (HEL-1325) of mice had significantly smaller ear lesions than the control group (HEL-1207) at 6 and 7 weeks post-challenge (p values of and 0.004, respectively) (Fig. 4A). However, the overall burden of disease was not significantly different between the two groups (Fig. 4B). Scars developed in 62 and 45 percent of the mice in control and vaccine groups respectively (Fig. 4C). Ear parasite numbers were similar between the two groups at all time points (Fig. 4D). Overall, results from the long-term needle challenge (LTN) indicated that the vaccine did not confer long-term protection in mice. Results from the long-term sand fly (LTSF) challenge indicated that the vaccine group (HEL-1325) developed significantly smaller ear lesions between 4 and 8 weeks post-challenge compared to the control group (HEL-1207) (p values between and ) (Fig. 4E), concurrently exhibiting a significantly smaller disease burden (p=0.019) (Fig. 4F). Scars developed in 52 and 15 percent of the mice 11
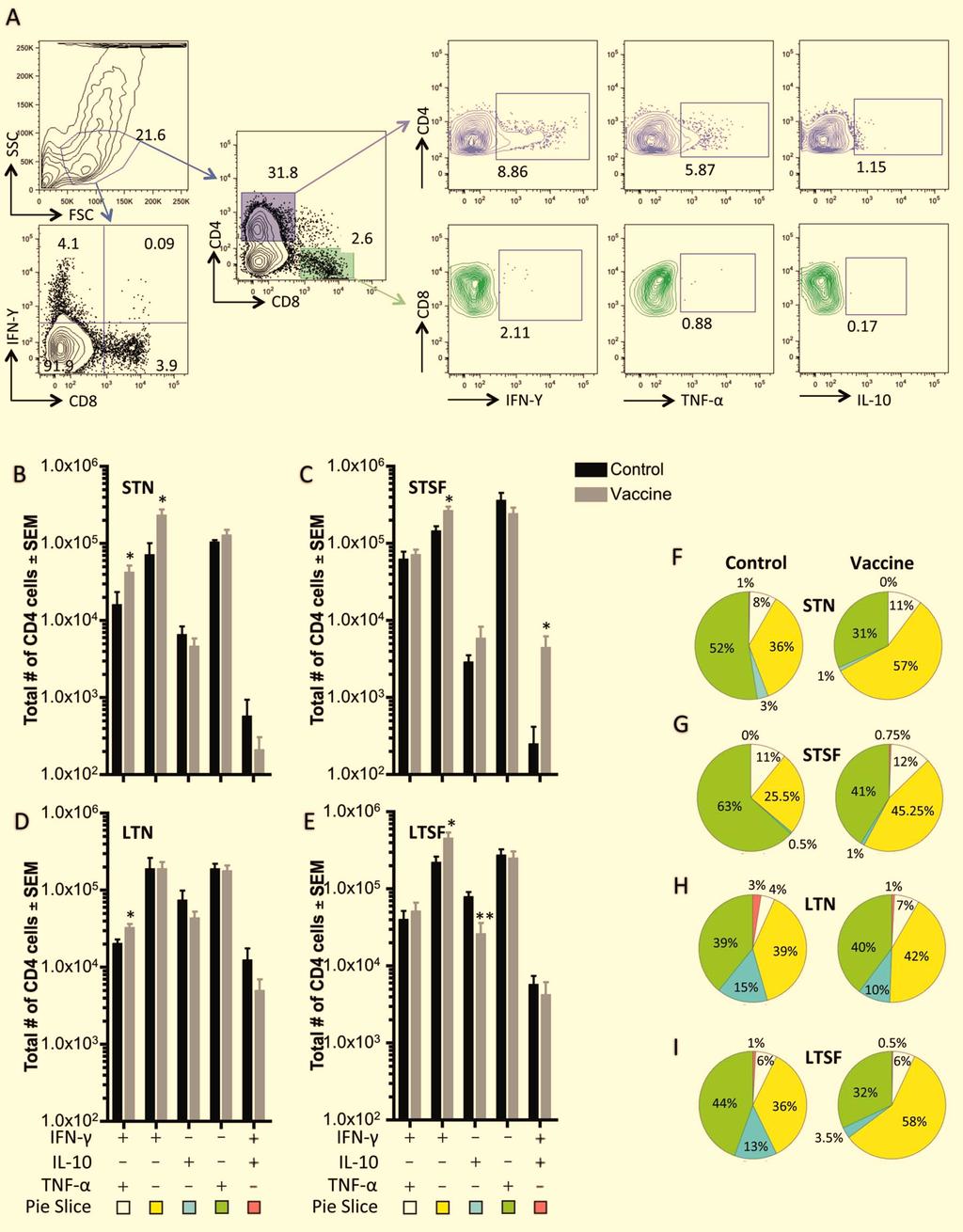
12 in control and vaccine groups respectively, and this difference was statistically significant (p=0.017) (Fig. 4G). Similarly to the STSF challenge, ear parasite numbers started 1.5 logs higher in the vaccine group compared to the control group (Fig. 4H). However, these numbers decreased by a log in the vaccine group, whereas they increased by 1.5 logs in the control group between 2 and 6 weeks post-challenge. Parasite numbers did not change in either groups between 6 and 11 weeks post-challenge. Overall, results from the LTSF challenge indicated that although parasite numbers were not significantly different between the vaccine and control groups by the end of the experiment, the vaccine is effective in decreasing overall disease burden, and preventing the formation of scars. A combination of high CD4 + IFN-γ + TNF-α ± IL-10 - and low CD4 + IFN-γ ± TNF-α - IL-10 + T cell populations at two weeks post-challenge is predictive of protection. Analysis of CD4 + and CD8 + specific T cells recruited at the site of challenge indicated that more than 97% on average of the specific immune response to Leishmania was mediated by CD4 + T cells (Fig. 5A). At two weeks post-challenge, there were significantly higher numbers of CD4 + IFN-γ + TNF-α ± IL-10 - T cells in every vaccine group compared to the respective control groups (Fig. 5B-E). The share CD4 + IFN-γ + TNF-α ± IL-10 - T cells occupied was 68% and 64% in the protected groups (STN and LTSF vaccine groups), and between 36% and 57% in the nonprotected groups (LTN and STSF vaccine groups and all control groups) (Fig. 5F-I). There were no significant differences between control and vaccine groups in the number of cells producing TNF-α for any of the experimental challenges, although the share CD4 + IFN-γ ± TNF-α + IL-10 - T cells occupied was lower, 42% and 38%, in the protected groups (STN and LTSF), and between 43% and 74% in the nonprotected groups (LTN and STSF vaccine groups and all control groups) (Fig. 5F-I). The data also indicates a significantly smaller number of CD4 + IFN-γ - TNF-α - IL-10 + T cells in the protected LTSF vaccine group (Fig. 5E) and a significantly larger number of CD4 + IFN-γ ± TNF-α - IL-10 + T cells in the non-protected STSF vaccine group compared to their respective control groups (Fig. 5C). Although the share IL-10 (with or 12
13 without IFN-γ) occupied in the protected groups was not always lower than in the non-protected groups as a whole, the protected vaccine groups (STN and LTSF) had fold less CD4 + IFN-γ ± TNF-α - IL-10 + T cells than their respective control groups, whereas this difference was 0.3 and 1.6 fold between the nonprotected vaccine groups (LTN and STSF) and their respective control groups (Fig. 5F-I). The balance between CD4 + IFN-γ + IL-10 - and CD4 + IFN-γ ± IL-10 + T cells has been shown to be an indicator of protection in the mouse model of cutaneous leishmaniasis (39-41). Therefore, to assess this balance we calculated the vaccine/control ratios for CD4 + T cells expressing these cytokines ± TNFα at 2 weeks post-challenge and for disease burden. The ratios eliminate the inherent cytokine staining variability between samples processed on different days and lesion size variability for challenges performed with different preparations of metacyclic promastigotes or infected sand flies. For example, control and vaccine groups from a STN challenge were challenged together and processed together, but on a different day than control and vaccine groups from a STSF challenge. The protected STN and LTSF challenge groups had nearly a six-fold difference in the ratios of CD4 + IFN-γ + TNF-α ± IL-10 - (ratios of 2.3) and CD4 + IFN-γ ± TNF-α - IL-10 + (ratios of 0.4) respectively, with low disease burden ratios of 0.2 and 0.4 respectively (Fig. 6). The unprotected LTN challenge group had ratios of 1.3 and 0.7 for CD4 + IFN-γ + TNFα ± IL-10 - and CD4 + IFN-γ ± TNF-α - IL-10 + respectively, with a disease burden ratio of 0.6. The other unprotected STSF challenge group had ratios of 1.9 and 3.5 for CD4 + IFN-γ + TNF-α ± IL-10 - and CD4 + IFN-γ ± TNF-α - IL-10 + respectively, with a disease burden ratio of 0.8. Overall, the results reveal a pattern in which a combination of high CD4 + IFN-γ + TNF-α ± IL-10 - and low CD4 + IFN-γ ± TNF-α - IL-10 + T cell numbers at 2 weeks post-challenge is predictive of lower disease burden over the course of the experiment. Remarkably, no significant differences or patterns were observed in the anti-leishmania immune response of protected and non-protected mice at 6 and 11 weeks post-challenge (data not shown). DISCUSSION 13
14 In this study, we used L. monocytogenes as a vaccine delivery vector for LJM11, a sand fly salivary protein that has been shown to be a potent vaccine antigen (16, 42). Our results indicate that delivery of LJM11 by Listeria is a promising vaccination strategy against cutaneous leishmaniasis inducing long-term protection against the formation of ulcers and scars and resulting in an overall decrease in disease burden following a natural challenge with infected sand flies. In addition, these studies reinforce the value of LJM11 as a protective immunogen and validate the use of L. monocytogenes as a delivery vaccine vector. Given that the formulation for a vaccine can have great effects on its efficacy, we set out to determine whether Listeria could be used as an effective vaccine delivery model. Among the advantages of using this system for vaccination are that L. monocytogenes can be attenuated without affecting its ability to induce a strong cell-mediated T H 1 immune response (30), and the antigen of interest is secreted directly in the cytosol of host cells bypassing the need for DNA or protein purification. Our Listeria vaccine vector can express an abundant amount of LJM11 in infected cells and vaccinated mice developed a robust immune response to this protein. Cutaneous leishmaniasis is by definition an infection of the skin, an organ that is populated by specialized sets of dendritic cells that contribute to homing receptor imprinting on T cells (43-45). Accordingly, mice vaccinated intradermally were protected against disease, whereas those vaccinated intramuscularly or subcutaneously were not, presumably because intradermal delivery of the antigen lead to the imprinting of a larger number of T cells that readily migrated to the skin upon challenge. However, the best level of protection was observed in mice vaccinated intravenously. Upon intravenous injection, bacteria are delivered systemically to a large number of secondary lymphatic tissues within minutes of injection. Seemingly, this would lead to production of a larger quantity of antigen, exposition of the antigen to different subsets of dendritic cells, and a more robust induction of T cell proliferation at central and peripheral sites. However, an intravenous vaccine would not be practical, as it would 14
15 require specialized technical personnel to administer, creating an increased level of complexity in areas where cutaneous leishmaniasis is endemic. A future challenge will be to find ways to improve protection conferred by an intradermal vaccine. An initial approach could be to combine the vaccine with an inhibitor of IL-10, as this cytokine activates regulatory T cells, compromising clearance of the infection (40). Two different types of challenges were used to determine the short and long-term protection effects of the vaccine: a needle challenge in which mice were injected with purified metacyclic promastigotes mixed with sand fly saliva, and a natural challenge in which mice were exposed to infected sand flies. The needle challenge is more practical, less time consuming, more economical, less traumatic for the mice, and more precise than a natural challenge. For these reasons, most experiments were performed using the needle challenge. Ultimately, when optimal vaccine parameters were determined, we aimed at testing vaccine efficacy against a natural challenge using infected sand flies. During a natural challenge, sand flies feed multiple times on the mouse ear, causing more inflammation than a needle challenge. In addition, the challenge dose cannot be controlled during a natural challenge, adding a variable that is absent from the needle challenge. Nevertheless, challenging mice with infected sand flies is imperative to the assessment of vaccine efficacy, as this type of challenge reproduces more closely what happens when an individual becomes infected. Conferring protection long after the last boost and preferably for the life of the host is of primary concern when developing a vaccine. Vaccinated mice were protected short-term against a needle challenge, but not long-term. On the contrary, vaccinated mice were protected long-term against a sand fly challenge, but not short-term. There are obvious differences in these two types of challenges as described above. Needle injections are administered quickly and at a single site, while sand flies bite repeatedly creating more trauma and inflammation at the site of infection. Furthermore, sand flies secrete a mucin-like gel that has been reported to exacerbate infection (46). It would be reasonable to 15
16 speculate that the extra inflammatory reaction caused by sand fly bites combined with a strong recall response in the short-term challenge might have hastened the development of a regulatory T cell response, abrogating the protective immune response to infection (39). However, vaccination with the purified protein was shown to confer short and long-term protection against a sand fly challenge (16). Perhaps, the structure of LJM11 is modified when made as a chimeric protein by Listeria, potentially affecting the presentation of important amino acids. These results emphasize the need to use a challenge method that is as close as possible to a natural challenge for truly testing vaccine efficacy. At 11 weeks post-challenge, parasite numbers were equivalent in all the groups, despite the fact that the overall disease burden and the number of mice with scars were significantly lower in the protected groups (STN and LTSF). It was not uncommon to find mice with equal parasite numbers but very different pathology at 11 weeks post-challenge (Fig. S4). Nonetheless, parasite numbers were significantly lower at 6 weeks post-challenge in the protected vaccinated groups compared to their respective control groups, and did not significantly increase between 6 and 11 weeks. These results suggest that early control of parasite replication is most important to prevent lesion development and that parasite clearance is not essential to prevent the pathology. Vaccinated mice challenged with infected sand flies had higher parasite numbers than control mice at two weeks post-challenge. Control and vaccinated groups were challenged simultaneously and analysis of flies post-infection indicated no differences between the percent of fed flies between mice. Therefore, it appears that the recall response to LJM11 was favorable to initial parasite survival and/or growth following a sand fly challenge. The immune response in the hours and days following challenge ought to be characterized to address these differences. Crucial to the control of an infection with Leishmania is the development of a T H 1 immune response, characterized mainly by the production of IFN-γ by CD4 + T cells (16, 47). Although CD8 + T cells can also serve as a source of IFN-γ, their cytolytic effect exacerbates immunopathology and mediates the 16
17 development of metastatic lesions (48). However, we focused our analysis on CD4 + T cells because they consistently accounted for more than 97% of the total numbers of T cells specific to L. major at 2 weeks post-infection. Induction of multifunctional CD4 + T cells producing IFN-γ with or without TNF-α, and few CD4+ T cells producing IL-10 with or without IFN-γ is also very important to disease control (40, 49). Accordingly, our results indicate that protected vaccinated groups generated high CD4 + IFN-γ + TNF-α ± IL and low CD4 + IFN-γ ± TNF-α - IL-10 + T cell numbers. This immune profile at 2 weeks post-challenge was predictive of the outcome disease burden for these 11-week long experiments. We conclude that the Listeria-based vaccine secreting the sand fly salivary protein LJM11 during intracellular infection generates an immune response that contributes to curbing disease burden and reducing the number of scars and ulcers in mice challenged with infected sand flies three months postvaccination. Protection correlates with a high ratio of CD4 + IFN-γ + TNF-α ± IL-10 - to CD4 + IFN-γ ± TNF-α - IL at 2 weeks post challenge and lower parasite numbers at 6 weeks post-challenge. ACKNOWLEDGEMENTS The study was funded by the Department of Defense Peer Reviewed Medical Research Program of the Office of the Congressionally Directed Medical Research Programs (W81XWH ), and in part by the Intramural Research Program of the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health. We are thankful to Dr. Regis Gomes for advice during the duration of this project. REFERENCES 1. Alvar J, Velez ID, Bern C, Herrero M, Desjeux P, Cano J, Jannin J, den Boer M Leishmaniasis worldwide and global estimates of its incidence. PLoS One 7:e
18 Gomes R, Oliveira F The immune response to sand fly salivary proteins and its influence on Leishmania immunity. Front. Immunol. 3: Belkaid Y, Kamhawi S, Modi G, Valenzuela J, Noben-Trauth N, Rowton E, Ribeiro J, Sacks DL Development of a natural model of cutaneous leishmaniasis: powerful effects of vector saliva and saliva preexposure on the long-term outcome of Leishmania major infection in the mouse ear dermis. J. Exp. Med. 188: Titus RG, Ribeiro JM Salivary gland lysates from the sand fly Lutzomyia longipalpis enhance Leishmania infectivity. Science 239: Andrade BB, de Oliveira CI, Brodskyn CI, Barral A, Barral-Netto M Role of sand fly saliva in human and experimental leishmaniasis: current insights. Scand. J. Immunol. 66: Kamhawi S, Belkaid Y, Modi G, Rowton E, Sacks D Protection against cutaneous leishmaniasis resulting from bites of uninfected sand flies. Science 290: Anderson JM, Samake S, Jaramillo-Gutierrez G, Sissoko I, Coulibaly CA, Traore B, Soucko C, Guindo B, Diarra D, Fay MP, Lawyer PG, Doumbia S, Valenzuela JG, Kamhawi S Seasonality and prevalence of Leishmania major infection in Phlebotomus duboscqi Neveu- Lemaire from two neighboring villages in central Mali. PLoS Negl. Trop. Dis. 5:e Aronson SM The Bagdad boil deploys to the United States. Med. Health R I 90: Bhutto AM, Soomro RA, Nonaka S, Hashiguchi Y Detection of new endemic areas of cutaneous leishmaniasis in Pakistan: a 6-year study. Int. J. Dermatol. 42: Jacobson RL Leishmaniasis in an era of conflict in the Middle East. Vector Borne Zoonotic Dis. 11: Pavli A, Maltezou HC Leishmaniasis, an emerging infection in travelers. Int. J. Infect. Dis. 14:e
19 Postigo JA Leishmaniasis in the World Health Organization eastern mediterranean region. Int. J. Antimicrob. Agents 36 Suppl 1:S Reyburn H, Rowland M, Mohsen M, Khan B, Davies C The prolonged epidemic of anthroponotic cutaneous leishmaniasis in Kabul, Afghanistan: 'bringing down the neighbourhood'. Trans. R. Soc. Trop. Med. Hyg. 97: van Thiel PP, Leenstra T, de Vries HJ, van der Sluis A, van Gool T, Krull AC, van Vugt M, de Vries PJ, Zeegelaar JE, Bart A, van der Meide WF, Schallig HD, Faber WR, Kager PA Cutaneous leishmaniasis (Leishmania major infection) in Dutch troops deployed in northern Afghanistan: epidemiology, clinical aspects, and treatment. Am. J. Trop. Med. Hyg. 83: Yaghoobi-Ershadi MR, Jafari R, Hanafi-Bojd AA A new epidemic focus of zoonotic cutaneous leishmaniasis in central Iran. Ann. Saudi Med. 24: Gomes R, Oliveira F, Teixeira C, Meneses C, Gilmore DC, Elnaiem DE, Kamhawi S, Valenzuela JG Immunity to sand fly salivary protein LJM11 modulates host response to vectortransmitted Leishmania conferring ulcer-free protection. J. Invest. Dermatol. 132: Gomes R, Teixeira C, Teixeira MJ, Oliveira F, Menezes MJ, Silva C, de Oliveira CI, Miranda JC, Elnaiem DE, Kamhawi S, Valenzuela JG, Brodskyn CI Immunity to a salivary protein of a sand fly vector protects against the fatal outcome of visceral leishmaniasis in a hamster model. Proc. Natl. Acad. Sci. U S A 105: Oliveira F, Lawyer PG, Kamhawi S, Valenzuela JG Immunity to distinct sand fly salivary proteins primes the anti-leishmania immune response towards protection or exacerbation of disease. PLoS Negl. Trop. Dis. 2:e Tavares NM, Silva RA, Costa DJ, Pitombo MA, Fukutani KF, Miranda JC, Valenzuela JG, Barral A, de Oliveira CI, Barral-Netto M, Brodskyn C Lutzomyia longipalpis Saliva or Salivary 19
20 Protein LJM19 Protects against Leishmania braziliensis and the Saliva of Its Vector, Lutzomyia intermedia. PLoS Negl. Trop. Dis. 5:e Belkaid Y, Mendez S, Lira R, Kadambi N, Milon G, Sacks D A natural model of Leishmania major infection reveals a prolonged "silent" phase of parasite amplification in the skin before the onset of lesion formation and immunity. J. Immunol. 165: Sacks D, Noben-Trauth N The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2: Tilney LG, Portnoy DA Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J. Cell Biol. 109: Mackaness GB Cellular resistance to infection. J. Exp. Med. 116: Pamer EG Immune responses to Listeria monocytogenes. Nat. Rev. Immunol. 4: Brockstedt DG, Dubensky TW Promises and challenges for the development of Listeria monocytogenes-based immunotherapies. Expert Rev. Vaccines 7: Gravekamp C, Paterson Y Harnessing Listeria monocytogenes to target tumors. Cancer Biol. Ther. 9: Le DT, Dubenksy TW, Jr., Brockstedt DG Clinical development of Listeria monocytogenesbased immunotherapies. Semin. Oncol. 39: Brundage RA, Smith GA, Camilli A, Theriot JA, Portnoy DA Expression and phosphorylation of the Listeria monocytogenes ActA protein in mammalian cells. Proc. Natl. Acad. Sci. USA 90: Kocks C, Gouin E, Tabouret M, Berche P, Ohayon H, Cossart P L. monocytogenes-induced actin assembly requires the acta gene product, a surface protein. Cell 68:
21 Brockstedt DG, Giedlin MA, Leong ML, Bahjat KS, Gao Y, Luckett W, Liu W, Cook DN, Portnoy DA, Dubensky TW, Jr Listeria-based cancer vaccines that segregate immunogenicity from toxicity. Proc. Natl. Acad. Sci. U S A 101: Crimmins GT, Herskovits AA, Rehder K, Sivick KE, Lauer P, Dubensky TW, Jr., Portnoy DA Listeria monocytogenes multidrug resistance transporters activate a cytosolic surveillance pathway of innate immunity. Proc. Natl. Acad. Sci. U S A 105: Auerbuch V, Brockstedt DG, Meyer-Morse N, O'Riordan M, Portnoy DA Mice lacking the type I interferon receptor are resistant to Listeria monocytogenes. J. Exp. Med. 200: Rayamajhi M, Humann J, Penheiter K, Andreasen K, Lenz LL Induction of IFN-alphabeta enables Listeria monocytogenes to suppress macrophage activation by IFN-gamma. J. Exp. Med. 207: Camilli A, Tilney LG, Portnoy DA Dual roles of plca in Listeria monocytogenes pathogenesis. Mol. Microbiol. 8: Snyder A, Marquis H Restricted translocation across the cell wall regulates secretion of the broad-range phospholipase C of Listeria monocytogenes. J. Bacteriol. 185: Marquis H Tissue culture cell assays used to analyze Listeria monocytogenes, p. 9B.4.1-9B In Downey T (ed.), Current Protocols In Microbiology. John Wiley & Sons, Inc., Hoboken, NJ. 37. Mendez S, Gurunathan S, Kamhawi S, Belkaid Y, Moga MA, Skeiky YA, Campos-Neto A, Reed S, Seder RA, Sacks D The potency and durability of DNA- and protein-based vaccines against Leishmania major evaluated using low-dose, intradermal challenge. J. Immunol. 166: Abi Abdallah DS, Egan CE, Butcher BA, Denkers EY Mouse neutrophils are professional antigen-presenting cells programmed to instruct Th1 and Th17 T-cell differentiation. Int. Immunol. 23:
22 Anderson CF, Oukka M, Kuchroo VJ, Sacks D CD4(+)CD25(-)Foxp3(-) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J. Exp. Med. 204: Darrah PA, Hegde ST, Patel DT, Lindsay RW, Chen L, Roederer M, Seder RA IL-10 production differentially influences the magnitude, quality, and protective capacity of Th1 responses depending on the vaccine platform. J. Exp. Med. 207: Stober CB, Lange UG, Roberts MT, Alcami A, Blackwell JM IL-10 from regulatory T cells determines vaccine efficacy in murine Leishmania major infection. J. Immunol. 175: Xu X, Oliveira F, Chang BW, Collin N, Gomes R, Teixeira C, Reynoso D, My Pham V, Elnaiem DE, Kamhawi S, Ribeiro JM, Valenzuela JG, Andersen JF Structure and function of a "yellow" protein from saliva of the sand fly Lutzomyia longipalpis that confers protective immunity against Leishmania major infection. J. Biol. Chem. 286: Kupper TS Old and new: recent innovations in vaccine biology and skin T cells. J. Invest. Dermatol. 132: Larregina AT, Falo LD, Jr Changing paradigms in cutaneous immunology: adapting with dendritic cells. J. Invest. Dermatol. 124: Sigmundsdottir H, Butcher EC Environmental cues, dendritic cells and the programming of tissue-selective lymphocyte trafficking. Nat. Immunol. 9: Rogers ME The role of Leishmania proteophosphoglycans in sand fly transmission and infection of the mammalian host. Front. Microbiol. 3: McSorley S, Proudfoot L, O'Donnell CA, Liew FY Immunology of murine leishmaniasis. Clin Dermatol 14:
23 Novais FO, Carvalho LP, Graff JW, Beiting DP, Ruthel G, Roos DS, Betts MR, Goldschmidt MH, Wilson ME, de Oliveira CI, Scott P Cytotoxic T cells mediate pathology and metastasis in cutaneous leishmaniasis. PLoS Pathog. 9:e Darrah PA, Patel DT, De Luca PM, Lindsay RW, Davey DF, Flynn BJ, Hoff ST, Andersen P, Reed SG, Morris SL, Roederer M, Seder RA Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat. Med. 13: Downloaded from on September 11, 2018 by guest 23
24 506 FIGURE LEGENDS FIG 1 Listeria vaccine vector. A: Schematic representation of the L. monocytogenes vaccine construct. The various sequence parts code for: acta P: acta promoter; ActA SS: ActA signal sequence; 100 amino acids of the ActA N-terminus; FLAG: Flag tag; and LJM11: Lutzomyia longipalpis salivary gland protein LJM11. B: Infected J774 cells were pulse labeled with 35 S-met in the presence of host protein synthesis inhibitors. Host cells were lysed and secreted bacterial proteins were resolved by SDS-PAGE and detected by autoradiography. Molecular mass markers in kda are indicated on the far left; the location of ActA/LJM11 chimera and a degradation product (*) are indicated on the far right; HEL-1207 (L. monocytogenes acta mdrm), and HEL-1325 (HEL-1207 acta/ljm11). C: Intracellular growth kinetics of L. monocytogenes in J774 cells. D: Immune response to LJM11 in splenocytes from vaccinated mice. Mice were vaccinated intravenously as described in Methods with the control vector (HEL-1207) or the vaccine vector (HEL-1325). Three weeks later, spleens were collected and splenocytes were incubated overnight with dendritic cells that had been pulsed with or without LJM11. Levels of secreted IFN-γ and IL-10 were measured by ELISA (limit of detection was 15 and 70 pg/ml for IFN-γ and IL-10, respectively). Data were analyzed using the One way Anova test with Bonferroni post-test analysis. ** p<0.01. n=3 per mouse group. FIG 2 L. monocytogenes vaccine vector confers the greatest protection against cutaneous leishmaniasis when administered intravenously. Lesion areas of mice vaccinated using different routes. Mice were primed and boosted twice at two-week intervals with the vaccine vector (HEL-1325) by the intradermal (ID, dark blue x), intramuscular (IM, magenta diamond), subcutaneous (SC, light blue circle) or intravenous (IV, red cross) route, while a group of age-matched mice was left unvaccinated (CTRL, black square). Three weeks after the last boost, mice were challenged with L. major plus half a pair of sand fly salivary glands. A: Ear lesion areas (diameter x thickness) were measured weekly and plotted. B: Disease burden of mice vaccinated and challenged. Area under the curve was measured for each individual 24
25 mouse and plotted. Experiment was repeated twice with n=6 per mouse group. C: Mice were primed and boosted intravenously twice at two-week intervals with the vaccine vector control (HEL-1207) or PBS, and challenged three weeks later as described above. Ear lesion areas are reported. Data were analyzed using the One-way Anova statistical test with Bonferroni post-test analysis. *** p<0.001, * p<0.05. FIG 3 Short-term needle and sand fly challenges. Mice were vaccinated intravenously three times at two-week intervals, and challenged intradermally three weeks after the last boost. Control group (HEL- 1207); Vaccine group (HEL-1325). A-D: Mice were challenged by needle injection with L. major plus half a pair of salivary gland homogenate. E-H: Mice were challenged with infected sand flies. A and E: Lesion area measurements for 10 weeks post-challenge. B and F: Disease burden calculated by measuring the area under the curve for each individual mouse. C and G: Percent of mice that developed scars over the duration of 10 weeks. D and H: Parasite numbers per ear at 2, 6 and 11 weeks post challenge. Experiments were repeated twice and data shown are pooled from both experiments; n of 30 mice per group (10 processed at each time-point). Data were analyzed using the T-test comparing vaccine and control groups. *** p<0.001, **p<0.01, * p<0.05. The control group in 3A is the same as the vector control group in 2C. FIG 4 Long-term needle and sand fly challenges. Mice were vaccinated intravenously three times at twoweek intervals, and challenged intradermally three months after the last boost. Control group (HEL- 1207); Vaccine group (HEL-1325). A-D: Mice were challenged by needle injection with L. major plus half a pair of salivary gland homogenate. E-H: Mice were challenged with infected sand flies. A and E: Lesion area measurements for 10 weeks post-challenge. B and F: Disease burden calculated by measuring the area under the curve for each individual mouse. C and G: Percent of mice that developed scars over the duration of 10 weeks. D and H: Parasite numbers per ear at 2, 6 and 11 weeks post challenge. Experiments were repeated twice and data shown are pooled from both experiments; n of 30 mice per 25
26 group (10 processed at each time-point). Data were analyzed using the T-test comparing vaccine and control groups. *** p<0.001, **p<0.01, * p< FIG 5 Recall response to vaccine at 2 weeks post-challenge. Cells collected from ears were incubated overnight with dendritic cells pre-pulsed with CLA and then treated for 4 hours with brefeldin A and PMA. Cells were stained for surface markers and intracellular cytokines, and analyzed by flow cytometry. Results were analyzed by gating for CD4 surface expression and intracellular cytokine expression (IFN-γ and TNF-α, IFN-γ, IL-10, or TNF-α). The total number of Leishmania-specific cytokine producing CD4 + T cells in both control (HEL-1207; black bars) and vaccine (HEL-1325; grey bars) groups of mice is shown for (A) short-term needle challenge (STN), (B) long-term needle challenge (LTN), (C) short-term sand fly challenge (STSF), and (D) long-term sand fly challenge (LTSF). E-H: Data were converted into pie charts showing the contribution of each cytokine combination to the total number of cytokine producing CD4 + T cells. Experiments were repeated twice and data shown are pooled from both experiments. N-10 (n=5 mice per group per experimental repeat). Data were analyzed using the T-test comparing vaccine and control groups. **p<0.01, * p<0.05. FIG 6 Cytokine production at 2 weeks post-challenge predicts disease burden. Total numbers of CD4 + IFN-γ + TNF-α ± IL-10 - and CD4 + IFN-γ ± TNF-α - IL-10 + T cells were calculated for control (HEL-1207) and vaccine (HEL-1325) groups of mice and plotted as Vaccine/Control ratios. Ratios for CD4 + IFN-γ + TNF-α ± IL-10 - are shown in blue (triangles), CD4 + IFN-γ ± TNF-α - IL-10 + ratios are shown in red (squares) and disease burden ratios are shown in grey (circles). Cytokine ratios were calculated from data at 2 weeks post-challenge while disease burden ratios are for the duration of the entire study. Cytokine ratios are plotted on the left y-axis while the disease burden ratios are plotted on the right y-axis. STN: short-term needle challenge, LTN: long-term needle challenge, STSF: short-term sand fly challenge, LTSF: long-term sand fly challenge. Experiments were repeated twice and data shown is pooled from both experiments. n=10 (n=5 mice per group per repeat). 26
27
28
29
30
31
INFLAMMATORY CELL INFILTRATION AND HIGH ANTIBODY PRODUCTION IN BALB/c MICE CAUSED BY NATURAL EXPOSURE TO LUTZOMYIA LONGIPALPIS BITES
 Am. J. Trop. Med. Hyg., 72(1), 2005, pp. 94 98 Copyright 2005 by The American Society of Tropical Medicine and Hygiene INFLAMMATORY CELL INFILTRATION AND HIGH ANTIBODY PRODUCTION IN BALB/c MICE CAUSED
Am. J. Trop. Med. Hyg., 72(1), 2005, pp. 94 98 Copyright 2005 by The American Society of Tropical Medicine and Hygiene INFLAMMATORY CELL INFILTRATION AND HIGH ANTIBODY PRODUCTION IN BALB/c MICE CAUSED
Analysis of Enhancing Effect of Sand Fly Saliva on Leishmania Infection in Mice
 INFCTION AND IMMUNITY, May 1991, p. 1592-1598 19-9567/91/51592-7$2./ Copyright X) 1991, American Society for Microbiology Vol. 59, No. 5 Analysis of nhancing ffect of Sand Fly Saliva on Leishmania Infection
INFCTION AND IMMUNITY, May 1991, p. 1592-1598 19-9567/91/51592-7$2./ Copyright X) 1991, American Society for Microbiology Vol. 59, No. 5 Analysis of nhancing ffect of Sand Fly Saliva on Leishmania Infection
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Animal Models of Leishmaniasis Relevant to Vaccine Development. David Sacks Laboratory of Parasitic Diseases NIAID, NIH
 Animal Models of Leishmaniasis Relevant to Vaccine Development David Sacks Laboratory of Parasitic Diseases NIAID, NIH Leishmaniasis: A spectrum of diseases associated with a diversity of parasite species,
Animal Models of Leishmaniasis Relevant to Vaccine Development David Sacks Laboratory of Parasitic Diseases NIAID, NIH Leishmaniasis: A spectrum of diseases associated with a diversity of parasite species,
Carregaro et al. BMC Microbiology 2013, 13:102
 Carregaro et al. BMC Microbiology 213, 13:12 RESEARCH ARTICLE Open Access Dual effect of Lutzomyia longipalpis saliva on Leishmania braziliensis infection is mediated by distinct saliva-induced cellular
Carregaro et al. BMC Microbiology 213, 13:12 RESEARCH ARTICLE Open Access Dual effect of Lutzomyia longipalpis saliva on Leishmania braziliensis infection is mediated by distinct saliva-induced cellular
Harnessing (and Controlling) L. monocytogenes to Treat. and Prevent Human Disease. Thomas W. Dubensky, Jr., Ph.D. Cerus Corporation Concord, CA
 L. monocytogenes to Treat. and Prevent Human Disease. Thomas W. Dubensky, Jr., Ph.D. Cerus Corporation Concord, CA") Harnessing (and Controlling) L. monocytogenes to Treat and Prevent Human Disease Thomas W. Dubensky, Jr., Ph.D. Cerus Corporation Concord, CA Presentation Overview Rationale for L. monocytogenes based
Harnessing (and Controlling) L. monocytogenes to Treat and Prevent Human Disease Thomas W. Dubensky, Jr., Ph.D. Cerus Corporation Concord, CA Presentation Overview Rationale for L. monocytogenes based
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Shigella Pathogenesis and Vaccine Development
 Shigella Pathogenesis and Vaccine Development Ryan Ranallo, Ph.D. Department of Enteric Infections Division of Communicable Diseases and Immunology Walter Reed Army Institute of Research Causes of Travelers
Shigella Pathogenesis and Vaccine Development Ryan Ranallo, Ph.D. Department of Enteric Infections Division of Communicable Diseases and Immunology Walter Reed Army Institute of Research Causes of Travelers
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Enhanced Leishmania braziliensis Infection Following Pre-Exposure to Sandfly Saliva
 Enhanced Leishmania braziliensis Infection Following Pre-Exposure to Sandfly Saliva Tatiana R. de Moura 1, Fabiano Oliveira 1,2, Fernanda O. Novais 1, José Carlos Miranda 1, Jorge Clarêncio 1, Ivonise
Enhanced Leishmania braziliensis Infection Following Pre-Exposure to Sandfly Saliva Tatiana R. de Moura 1, Fabiano Oliveira 1,2, Fernanda O. Novais 1, José Carlos Miranda 1, Jorge Clarêncio 1, Ivonise
Increased IL-12 induced STAT-4 signaling in CD8 T cells. from aged mice
 Increased IL-2 induced STAT-4 signaling in CD8 T cells from aged mice Erin Rottinghaus * Abstract: Aging is associated with poor immune function leading to increased susceptibility to infectious diseases
Increased IL-2 induced STAT-4 signaling in CD8 T cells from aged mice Erin Rottinghaus * Abstract: Aging is associated with poor immune function leading to increased susceptibility to infectious diseases
L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity
 Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Series Editors Samuel J. Black, University of Massachusetts, Amherst, MA, US.A. J. Richard Seed, University of North Carolina, Chapel Hill, NC, US.A.
 LEISHMANIA World Class Parasites VOLUME 4 Volumes in the World Class Parasites book series are written for researchers, students and scholars who enjoy reading about excellent research on problems of global
LEISHMANIA World Class Parasites VOLUME 4 Volumes in the World Class Parasites book series are written for researchers, students and scholars who enjoy reading about excellent research on problems of global
Delayed-Type Hypersensitivity to Sand Fly Saliva in Humans from a Leishmaniasis-Endemic Area of Mali Is T H 1-Mediated and Persists to Midlife
 ORIGINAL ARTICLE Delayed-Type Hypersensitivity to Sand Fly Saliva in Humans from a Leishmaniasis-Endemic Area of Mali Is T H -Mediated and Persists to Midlife Fabiano Oliveira, Bourama Traoré 2, Regis
ORIGINAL ARTICLE Delayed-Type Hypersensitivity to Sand Fly Saliva in Humans from a Leishmaniasis-Endemic Area of Mali Is T H -Mediated and Persists to Midlife Fabiano Oliveira, Bourama Traoré 2, Regis
Electron micrograph of phosphotungstanic acid-stained exosomes derived from murine
 1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Naive, memory and regulatory T lymphocytes populations analysis
 Naive, memory and regulatory T lymphocytes populations analysis Jaen Olivier, PhD ojaen@beckmancoulter.com Cellular Analysis application specialist Beckman Coulter France Introduction Flow cytometric analysis
Naive, memory and regulatory T lymphocytes populations analysis Jaen Olivier, PhD ojaen@beckmancoulter.com Cellular Analysis application specialist Beckman Coulter France Introduction Flow cytometric analysis
Rapid antigen-specific T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
Vaccine Delivery and TLR Ligands Influence the Quality of T cell Responses in NHP. Robert A. Seder, M.D. Vaccine Research Center NIAID, NIH
 Vaccine Delivery and TLR Ligands Influence the Quality of T cell Responses in NHP Robert A. Seder, M.D. Vaccine Research Center NIAID, NIH Vaccines Against HIV, Malaria and Tuberculosis Will Require Neutralizing
Vaccine Delivery and TLR Ligands Influence the Quality of T cell Responses in NHP Robert A. Seder, M.D. Vaccine Research Center NIAID, NIH Vaccines Against HIV, Malaria and Tuberculosis Will Require Neutralizing
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Supplementary Information. Tissue-wide immunity against Leishmania. through collective production of nitric oxide
 Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Animal models for infectious diseases caused by parasites: Leishmaniasis
 Drug Discovery Today: Disease Models Vol. 1, No. 1 2004 DRUG DISCOVERY TODAY DISEASE MODELS Editors-in-Chief Jan Tornell AstraZeneca, Sweden Denis Noble University of Oxford, UK Infectious diseases Animal
Drug Discovery Today: Disease Models Vol. 1, No. 1 2004 DRUG DISCOVERY TODAY DISEASE MODELS Editors-in-Chief Jan Tornell AstraZeneca, Sweden Denis Noble University of Oxford, UK Infectious diseases Animal
Post Kala-azar Dermal Leishmaniasis (PKDL) from the field to the cellular and the subcellular levels
 from the field to the cellular and the subcellular levels") Post Kala-azar Dermal Leishmaniasis (PKDL) from the field to the cellular and the subcellular levels A M EL Hassan Institute of Endemic Diseases University of Khartoum Introduction PKDL is a VL related
Post Kala-azar Dermal Leishmaniasis (PKDL) from the field to the cellular and the subcellular levels A M EL Hassan Institute of Endemic Diseases University of Khartoum Introduction PKDL is a VL related
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Received 11 September 2003/Returned for modification 17 November 2003/Accepted 8 December 2003
 INFECTION AND IMMUNITY, Mar. 2004, p. 1298 1305 Vol. 72, No. 3 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.3.1298 1305.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Lutzomyia
INFECTION AND IMMUNITY, Mar. 2004, p. 1298 1305 Vol. 72, No. 3 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.3.1298 1305.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Lutzomyia
NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity
 NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity Peiwen Kuo Scientist, Research Biology Nektar Therapeutics August 31 st, 2018 Emerging
NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity Peiwen Kuo Scientist, Research Biology Nektar Therapeutics August 31 st, 2018 Emerging
Engineering an in vitro human immune system for rapid vaccine evaluation
 Engineering an in vitro human immune system for rapid vaccine evaluation William Warren, Ph.D. VaxDesign Corporation Orlando, FL 32826 wwarren@vaxdesign.com www.vaxdesign.com 1 MIMIC Technology Overview
Engineering an in vitro human immune system for rapid vaccine evaluation William Warren, Ph.D. VaxDesign Corporation Orlando, FL 32826 wwarren@vaxdesign.com www.vaxdesign.com 1 MIMIC Technology Overview
CUTANEOUS LEISHMANIASIS
 CUTANEOUS LEISHMANIASIS Why are you neglecting me? A WHO initiative to control Cutaneous Leishmaniasis in selected Old World areas This document has been produced as the result of a WHO Informal Consultative
CUTANEOUS LEISHMANIASIS Why are you neglecting me? A WHO initiative to control Cutaneous Leishmaniasis in selected Old World areas This document has been produced as the result of a WHO Informal Consultative
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplemental Information. Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection. Cell Host & Microbe, Volume 15
 Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
La risposta immune all infezione da virus ebola. Chiara Agrati, PhD
 La risposta immune all infezione da virus ebola Chiara Agrati, PhD Pathogenetic mechanisms This virus infection is able to: - disable the immune system, preventing an effective protective immune response
La risposta immune all infezione da virus ebola Chiara Agrati, PhD Pathogenetic mechanisms This virus infection is able to: - disable the immune system, preventing an effective protective immune response
Role of Listeriolysin O in Cell-to-Cell Spread of Listeria monocytogenes
 INFECTION AND IMMUNITY, Feb. 2000, p. 999 1003 Vol. 68, No. 2 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Role of Listeriolysin O in Cell-to-Cell Spread
INFECTION AND IMMUNITY, Feb. 2000, p. 999 1003 Vol. 68, No. 2 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Role of Listeriolysin O in Cell-to-Cell Spread
Original Article Comparison of fine needle aspiration with biopsy in the diagnosis of cutaneous leishmaniasis Ikramullah Khan, Rifat Yasmin
 Original Article Comparison of fine needle aspiration with biopsy in the diagnosis of cutaneous leishmaniasis Ikramullah Khan, Rifat Yasmin Department of Dermatology, Pakistan Institute of Medical Sciences,
Original Article Comparison of fine needle aspiration with biopsy in the diagnosis of cutaneous leishmaniasis Ikramullah Khan, Rifat Yasmin Department of Dermatology, Pakistan Institute of Medical Sciences,
Immunologically Induced and Elicited Local
 INFECTION AND IMMUNITY, Dec. 1970, p. 757-761 Copyright 1970 American Society for Microbiology Vol. 2, No. 6 Printed in U.S.A. Immunologically Induced and Elicited Local Resistance to Staphylococcus aureus
INFECTION AND IMMUNITY, Dec. 1970, p. 757-761 Copyright 1970 American Society for Microbiology Vol. 2, No. 6 Printed in U.S.A. Immunologically Induced and Elicited Local Resistance to Staphylococcus aureus
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies
 NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
SUMMARY. Cutaneous leishmaniasis with only skin involvement: single to multiple skin ulcers, satellite lesions and nodular lymphangitis.
 SUMMARY Leishmaniasis is a disease affecting predominantly people in the developing countries; 350 million people worldwide are at risk and yearly more than 2 million new cases occur. Leishmaniasis is
SUMMARY Leishmaniasis is a disease affecting predominantly people in the developing countries; 350 million people worldwide are at risk and yearly more than 2 million new cases occur. Leishmaniasis is
Leishmaniasis, Kala Azar(The Black Fever)
") Leishmaniasis, Kala Azar(The Black Fever) By Lawrence Hall Etiologic agent Protist obligate intracellular parasite, Transmission Vectors Phylum: Euglenozoa (genus Leishmania) Over 21 species that infect
Leishmaniasis, Kala Azar(The Black Fever) By Lawrence Hall Etiologic agent Protist obligate intracellular parasite, Transmission Vectors Phylum: Euglenozoa (genus Leishmania) Over 21 species that infect
Detailed step-by-step operating procedures for NK cell and CTL degranulation assays
 Supplemental methods Detailed step-by-step operating procedures for NK cell and CTL degranulation assays Materials PBMC isolated from patients, relatives and healthy donors as control K562 cells (ATCC,
Supplemental methods Detailed step-by-step operating procedures for NK cell and CTL degranulation assays Materials PBMC isolated from patients, relatives and healthy donors as control K562 cells (ATCC,
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Product Datasheet. Ly-6G6C Antibody (NIMP-R14) NB Unit Size: 0.05 mg. Store at 4C. Do not freeze. Publications: 23
 NB Unit Size: 0.05 mg. Store at 4C. Do not freeze. Publications: 23") Product Datasheet Ly-6G6C Antibody (NIMP-R14) NB600-1387 Unit Size: 0.05 mg Store at 4C. Do not freeze. Publications: 23 Protocols, Publications, Related Products, Reviews, Research Tools and Images at:
Product Datasheet Ly-6G6C Antibody (NIMP-R14) NB600-1387 Unit Size: 0.05 mg Store at 4C. Do not freeze. Publications: 23 Protocols, Publications, Related Products, Reviews, Research Tools and Images at:
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland Approved for public release; distribution unlimited
 AD (Leave blank) Award Number: W81XWH-07-1-0345 TITLE: Second-Generation Therapeutic DNA Lymphoma Vaccines PRINCIPAL INVESTIGATOR: Larry W. Kwak, M.D., Ph.D. CONTRACTING ORGANIZATION: University of Texas
AD (Leave blank) Award Number: W81XWH-07-1-0345 TITLE: Second-Generation Therapeutic DNA Lymphoma Vaccines PRINCIPAL INVESTIGATOR: Larry W. Kwak, M.D., Ph.D. CONTRACTING ORGANIZATION: University of Texas
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection
 Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells
 ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
Third line of Defense
 Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
% of live splenocytes. STAT5 deletion. (open shapes) % ROSA + % floxed
 % ROSA + % floxed") Supp. Figure 1. a 14 1 1 8 6 spleen cells (x1 6 ) 16 % of live splenocytes 5 4 3 1 % of live splenocytes 8 6 4 b 1 1 c % of CD11c + splenocytes (closed shapes) 8 6 4 8 6 4 % ROSA + (open shapes) % floxed
Supp. Figure 1. a 14 1 1 8 6 spleen cells (x1 6 ) 16 % of live splenocytes 5 4 3 1 % of live splenocytes 8 6 4 b 1 1 c % of CD11c + splenocytes (closed shapes) 8 6 4 8 6 4 % ROSA + (open shapes) % floxed
Supplementary Figures
 Supplementary Figures Supplementary Fig. 1. Surface thiol groups and reduction of activated T cells. (a) Activated CD8 + T-cells have high expression levels of free thiol groups on cell surface proteins.
Supplementary Figures Supplementary Fig. 1. Surface thiol groups and reduction of activated T cells. (a) Activated CD8 + T-cells have high expression levels of free thiol groups on cell surface proteins.
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific
 SUPPLEMENTARY FIGURE LEGEND Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific CD4 + T cells correlates with intracellular CTLA-4 levels. (a) Comparative CTLA-4 levels
SUPPLEMENTARY FIGURE LEGEND Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific CD4 + T cells correlates with intracellular CTLA-4 levels. (a) Comparative CTLA-4 levels
3. Lymphocyte proliferation (fig. 15.4): Clones of responder cells and memory cells are derived from B cells and T cells.
: Clones of responder cells and memory cells are derived from B cells and T cells.") Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
The humoral immune responses to IBV proteins.
 The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
Study Guide 23, 24 & 47
 Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis
Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis
Memory NK cells during mousepox infection. Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science
 Memory NK cells during mousepox infection Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science Infectious Diseases are a Major Cause of Death Worldwide May 14 th 1796 Prevalence
Memory NK cells during mousepox infection Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science Infectious Diseases are a Major Cause of Death Worldwide May 14 th 1796 Prevalence
ASH 2011 aktualijos: MSC TPŠL gydyme. Mindaugas Stoškus VULSK HOTC MRMS
 ASH 2011 aktualijos: MSC TPŠL gydyme Mindaugas Stoškus VULSK HOTC MRMS #3042. Yukiyasu Ozawa et al. Mesenchymal Stem Cells As a Treatment for Steroid-Resistant Acute Graft Versus Host Disease (agvhd);
ASH 2011 aktualijos: MSC TPŠL gydyme Mindaugas Stoškus VULSK HOTC MRMS #3042. Yukiyasu Ozawa et al. Mesenchymal Stem Cells As a Treatment for Steroid-Resistant Acute Graft Versus Host Disease (agvhd);
Perspective in novel TB vaccine development Mohamed Ridha BARBOUCHE M.D., Ph.D. Department of Immunology Institut Pasteur de Tunis
 Perspective in novel TB vaccine development Mohamed Ridha BARBOUCHE M.D., Ph.D. Department of Immunology Institut Pasteur de Tunis Existing TB Vaccine is not effective for global TB epidemic control BCG
Perspective in novel TB vaccine development Mohamed Ridha BARBOUCHE M.D., Ph.D. Department of Immunology Institut Pasteur de Tunis Existing TB Vaccine is not effective for global TB epidemic control BCG
IOM Immunization Safety Review 11/12/2001. Immunological Competition and the Infant Immune Response to Vaccines
 IOM Immunization Safety Review 11/12/2001 Immunological Competition and the Infant Immune Response to Vaccines Richard Insel University of Rochester Goals Neonatal and Infant Immune System Broad Effects
IOM Immunization Safety Review 11/12/2001 Immunological Competition and the Infant Immune Response to Vaccines Richard Insel University of Rochester Goals Neonatal and Infant Immune System Broad Effects
Production of Exosomes in a Hollow Fiber Bioreactor
 Production of Exosomes in a Hollow Fiber Bioreactor John J S Cadwell, President and CEO, FiberCell Systems Inc INTRODUCTION Exosomes are small lipid membrane vesicles (80-120 nm) of endocytic origin generated
Production of Exosomes in a Hollow Fiber Bioreactor John J S Cadwell, President and CEO, FiberCell Systems Inc INTRODUCTION Exosomes are small lipid membrane vesicles (80-120 nm) of endocytic origin generated
Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder I Nakaya, Kousik
 SUPPLEMENTARY FIGURES 1-19 T H 2 response to cysteine-proteases requires dendritic cell-basophil cooperation via ROS mediated signaling Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder
SUPPLEMENTARY FIGURES 1-19 T H 2 response to cysteine-proteases requires dendritic cell-basophil cooperation via ROS mediated signaling Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder
/01/$ DOI: /IAI Received 21 July 2000/Returned for modification 26 September 2000/Accepted 13 October 2000
 INFECTION AND IMMUNITY, Jan. 2001, p. 194 203 Vol. 69, No. 1 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.1.194 203.2001 Susceptibility to Secondary Francisella tularensis Live Vaccine Strain Infection in
INFECTION AND IMMUNITY, Jan. 2001, p. 194 203 Vol. 69, No. 1 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.1.194 203.2001 Susceptibility to Secondary Francisella tularensis Live Vaccine Strain Infection in
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES
 HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
Rapamycin reduced inhibition of V 1 T cells towards V 4 T cells through down-regulating IL-4
 Rapamycin reduced inhibition of V 1 T cells towards V 4 T cells through down-regulating IL-4 Wanjun Huang Biomedical Translational Research Institute, Jinan University, Guangzhou 510632, China. Abstract
Rapamycin reduced inhibition of V 1 T cells towards V 4 T cells through down-regulating IL-4 Wanjun Huang Biomedical Translational Research Institute, Jinan University, Guangzhou 510632, China. Abstract
Supplemental Information. T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism
 Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
The IL-7 Receptor A Key Factor in HIV Pathogenesis
 The IL-7 Receptor A Key Factor in HIV Pathogenesis Paul MacPherson PhD, MD, FRCPC Associate Professor Division of Infectious Diseases Ottawa Hospital, General Campus Ottawa Hospital Research Institute
The IL-7 Receptor A Key Factor in HIV Pathogenesis Paul MacPherson PhD, MD, FRCPC Associate Professor Division of Infectious Diseases Ottawa Hospital, General Campus Ottawa Hospital Research Institute
Memory CD4 T Cells Enhance Primary CD8 T-Cell Responses
 INFECTION AND IMMUNITY, July 2007, p. 3556 3560 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00086-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Memory CD4 T Cells
INFECTION AND IMMUNITY, July 2007, p. 3556 3560 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00086-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Memory CD4 T Cells
Interferon γ regulates idiopathic pneumonia syndrome, a. Th17 + CD4 + T-cell-mediated GvH disease
 Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Nanomedicine & Drug Delivery Systems Group Research Institute for Medicines and Pharmaceutical Sciences (imed.ul)
") Faculty of Pharmacy of the University of Lisbon Nanomedicine & Drug Delivery Systems Group Research Institute for Medicines and Pharmaceutical Sciences (imed.ul) Leishmania in southern Europe. (Duiardin,
Faculty of Pharmacy of the University of Lisbon Nanomedicine & Drug Delivery Systems Group Research Institute for Medicines and Pharmaceutical Sciences (imed.ul) Leishmania in southern Europe. (Duiardin,
Fluid movement in capillaries. Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system
 Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
International Journal of Integrative Biology A journal for biology beyond borders ISSN
 Research International Journal of Integrative Biology A journal for biology beyond borders ISSN 0973-8363 Infectivity of Leishmania infantum treated with amphotericin B plus Phlebotomus salivary gland
Research International Journal of Integrative Biology A journal for biology beyond borders ISSN 0973-8363 Infectivity of Leishmania infantum treated with amphotericin B plus Phlebotomus salivary gland
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the
 Diagrammatic representation of the") Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
D CD8 T cell number (x10 6 )
") IFNγ Supplemental Figure 1. CD T cell number (x1 6 ) 18 15 1 9 6 3 CD CD T cells CD6L C CD5 CD T cells CD6L D CD8 T cell number (x1 6 ) 1 8 6 E CD CD8 T cells CD6L F Log(1)CFU/g Feces 1 8 6 p
IFNγ Supplemental Figure 1. CD T cell number (x1 6 ) 18 15 1 9 6 3 CD CD T cells CD6L C CD5 CD T cells CD6L D CD8 T cell number (x1 6 ) 1 8 6 E CD CD8 T cells CD6L F Log(1)CFU/g Feces 1 8 6 p
Malaria parasite vaccine development Strategies & Targets
 Malaria parasite vaccine development Strategies & Targets Tulane University Ahmed Aly Most malaria disease deaths are among children and pregnant women A child or a pregnant woman dies of malaria nearly
Malaria parasite vaccine development Strategies & Targets Tulane University Ahmed Aly Most malaria disease deaths are among children and pregnant women A child or a pregnant woman dies of malaria nearly
Writing Effective Grant Proposals
 WritingEffectiveGrantProposals SUPPLEMENTALHANDOUT EXERCISES (toaccompanypowerpointslidepresentation) PamelaDerish ScientificPublicationsManager DepartmentofSurgery,UCSF tel415.885 7686 Pamela.Derish@ucsfmedctr.org
WritingEffectiveGrantProposals SUPPLEMENTALHANDOUT EXERCISES (toaccompanypowerpointslidepresentation) PamelaDerish ScientificPublicationsManager DepartmentofSurgery,UCSF tel415.885 7686 Pamela.Derish@ucsfmedctr.org
Autoimmune Diseases. Betsy Kirchner CNP The Cleveland Clinic
 Autoimmune Diseases Betsy Kirchner CNP The Cleveland Clinic Disclosures (financial) No relevant disclosures Learning Objectives Explain the pathophysiology of autoimmune disease Discuss safe administration
Autoimmune Diseases Betsy Kirchner CNP The Cleveland Clinic Disclosures (financial) No relevant disclosures Learning Objectives Explain the pathophysiology of autoimmune disease Discuss safe administration
A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System
 Cell Host & Microbe, Volume 7 Supplemental Information A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System Igor E. Brodsky, Noah
Cell Host & Microbe, Volume 7 Supplemental Information A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System Igor E. Brodsky, Noah
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
Cover Page. The handle holds various files of this Leiden University dissertation
 Cover Page The handle http://hdl.handle.net/1887/35908 holds various files of this Leiden University dissertation Author: Soema, Peter Title: Formulation of influenza T cell peptides : in search of a universal
Cover Page The handle http://hdl.handle.net/1887/35908 holds various files of this Leiden University dissertation Author: Soema, Peter Title: Formulation of influenza T cell peptides : in search of a universal
Foot and Mouth Disease Vaccine Research and Development in India
 Foot and Mouth Disease Vaccine Research and Development in India R.Venkataramanan Indian Veterinary Research Institute, Hebbal, Bangalore 560 024 Foot and Mouth Disease in India Present Status Large Susceptible
Foot and Mouth Disease Vaccine Research and Development in India R.Venkataramanan Indian Veterinary Research Institute, Hebbal, Bangalore 560 024 Foot and Mouth Disease in India Present Status Large Susceptible
DTA3/COFUND Programme Research Project Proforma
 Page 1 General Information Project Code Partner University Faculty/School/Department/Research Centres First supervisor Please provide name and weblink Second supervisor Please provide name and weblink
Page 1 General Information Project Code Partner University Faculty/School/Department/Research Centres First supervisor Please provide name and weblink Second supervisor Please provide name and weblink
ASSESSMENT OF LYMPHOCYTE SUBGROUPS IN CHRONIC BRUCELLOSIS BEFORE AND AFTER IMMUNOTHERAPY
 ASSESSMENT OF LYMPHOCYTE SUBGROUPS IN CHRONIC BRUCELLOSIS BEFORE AND AFTER IMMUNOTHERAPY D. Tahamzadeh 1, A. Massoud *1, G. Samar 2 and B. Nikbin 1 1) Department of Immunology, School of Medicine, Tehran
ASSESSMENT OF LYMPHOCYTE SUBGROUPS IN CHRONIC BRUCELLOSIS BEFORE AND AFTER IMMUNOTHERAPY D. Tahamzadeh 1, A. Massoud *1, G. Samar 2 and B. Nikbin 1 1) Department of Immunology, School of Medicine, Tehran
Supplementary Material
 Supplementary Material Supplementary Figure 1. NOS2 -/- mice develop an analogous Ghon complex after infection in the ear dermis and show dissemination of Mtb to the lung. (A) WT and NOS2 -/- mice were
Supplementary Material Supplementary Figure 1. NOS2 -/- mice develop an analogous Ghon complex after infection in the ear dermis and show dissemination of Mtb to the lung. (A) WT and NOS2 -/- mice were
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
World Health Organization Department of Communicable Disease Surveillance and Response
 WHO Report on Global Surveillance of Epidemic-prone Infectious Diseases World Health Organization Department of Communicable Disease Surveillance and Response This document has been downloaded from the
WHO Report on Global Surveillance of Epidemic-prone Infectious Diseases World Health Organization Department of Communicable Disease Surveillance and Response This document has been downloaded from the
Immunity and Infection. Chapter 17
 Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
Polio vaccines and polio immunization in the pre-eradication era: WHO position paper. Published in WER 4 June, 2010
 Polio vaccines and polio immunization in the pre-eradication era: WHO position paper Published in WER 4 June, 2010 Epidemiology & Background Poliomyelitis (polio) is an acute communicable disease of humans
Polio vaccines and polio immunization in the pre-eradication era: WHO position paper Published in WER 4 June, 2010 Epidemiology & Background Poliomyelitis (polio) is an acute communicable disease of humans
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1: Chemokine receptor expression profiles of CCR6 + and CCR6 - CD4 + IL-17A +/ex and Treg cells. Quantitative PCR analysis of chemokine receptor transcript abundance
SUPPLEMENTARY FIGURES Supplementary Figure 1: Chemokine receptor expression profiles of CCR6 + and CCR6 - CD4 + IL-17A +/ex and Treg cells. Quantitative PCR analysis of chemokine receptor transcript abundance
Chapter 7 Conclusions
 VII-1 Chapter 7 Conclusions VII-2 The development of cell-based therapies ranging from well-established practices such as bone marrow transplant to next-generation strategies such as adoptive T-cell therapy
VII-1 Chapter 7 Conclusions VII-2 The development of cell-based therapies ranging from well-established practices such as bone marrow transplant to next-generation strategies such as adoptive T-cell therapy
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes
 Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes Undergraduate Honors Thesis The Ohio State University, College of Pharmacy Division of Medicinal Chemistry and Pharmacognosy
Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes Undergraduate Honors Thesis The Ohio State University, College of Pharmacy Division of Medicinal Chemistry and Pharmacognosy
Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection
 Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection Victoria L. Smith, Yong Cheng, Barry R. Bryant and Jeffrey S. Schorey Supplementary Figure 1: Unprocessed
Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection Victoria L. Smith, Yong Cheng, Barry R. Bryant and Jeffrey S. Schorey Supplementary Figure 1: Unprocessed
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES
 CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
Approved for Public Release; Distribution Unlimited
 AD Award Number: W81XWH-04-1-0186 TITLE: A Novel Therapeutic System for the Treatment of Occult Prostate Cancer PRINCIPAL INVESTIGATOR: Shongyun Dong, M.D., Ph.D. CONTRACTING ORGANIZATION: University of
AD Award Number: W81XWH-04-1-0186 TITLE: A Novel Therapeutic System for the Treatment of Occult Prostate Cancer PRINCIPAL INVESTIGATOR: Shongyun Dong, M.D., Ph.D. CONTRACTING ORGANIZATION: University of
Page # Lecture 8: Immune Dysfunction - Immunopathology. Four Types of Hypersensitivity. Friend of Foe? Autoimmune disease Immunodeficiency
 Lecture 8: Immune Dysfunction - Immunopathology Autoimmune disease Immunodeficiency Allergy and Asthma Graft rejection and Lupus Friend of Foe? Four Types of Hypersensitivity Allergic Responses - Type
Lecture 8: Immune Dysfunction - Immunopathology Autoimmune disease Immunodeficiency Allergy and Asthma Graft rejection and Lupus Friend of Foe? Four Types of Hypersensitivity Allergic Responses - Type
Chapter 22: The Lymphatic System and Immunity
 Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due