Washington University School of Medicine, St Louis. MO Division of Infectious
|
|
|
- Buddy Horton
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi: /jvi Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 CD8 + T cells use TRAIL to restrict West Nile virus pathogenesis by controlling infection in neurons Bimmi Shrestha 1, Amelia K. Pinto 1, Sharone Green 4, Irene Bosch 5, and Michael. S. Diamond 1,2,3. Departments of Medicine 1, Molecular Microbiology 2, and Pathology & Immunology 3, Washington University School of Medicine, St Louis. MO Division of Infectious Diseases and Immunology 4, University of Massachusetts Medical School, Worcester, MA Massachusetts Institute of Technology 5, Cambridge, MA. Corresponding author: Michael S. Diamond, M.D., PhD, Departments of Medicine, Molecular Microbiology and Pathology & Immunology, Washington University School of Medicine, 660 South Euclid Avenue, Box 8051, St Louis. Missouri Tel: , Fax: , diamond@borcim.wustl.edu Running title: TRAIL controls WNV infection 19 Abstract: 248 words; Figures:
2 ABSTRACT Previous studies in mice have demonstrated that an orchestrated sequence of innate and adaptive immune responses is required to control West Nile virus (WNV) infection in peripheral and central nervous system (CNS) tissues. TNF-related apoptosisinducing ligand (TRAIL, CD253) has been reported to inhibit infection of dengue virus, a closely related flavivirus, in cell culture. To determine the physiological function of TRAIL in the context of flavivirus infection, we compared the pathogenesis of WNV in wild type and TRAIL -/- mice. Mice lacking TRAIL showed increased vulnerability and death after subcutaneous WNV infection. Although no difference in viral burden was detected in peripheral tissues, greater viral infection was detected in the brain and spinal cord at late times after infection, and this was associated with delayed viral clearance in the few surviving TRAIL -/- mice. While priming of adaptive B and T cell responses, and trafficking of immune and antigen-specific cells to the brain was undistinguishable from normal mice, in TRAIL -/- mice, CD8 + T cells showed qualitative defects in their ability to clear WNV infection. Adoptive transfer of WNV-primed wild type but not TRAIL -/- CD8 + T cells to recipient CD8 -/- mice efficiently limited infection in the brain and spinal cord, and analogous results were obtained when wild type or TRAIL -/- CD8 + T cells were added to WNV-infected primary cortical neuron cultures ex vivo. Collectively, our results suggest that TRAIL produced by CD8 + T cells contributes to disease resolution by helping to clear WNV infection from neurons in the central nervous system. 41 2
3 INTRODUCTION West Nile Virus (WNV) is a mosquito-transmitted neurotropic flavivirus that is genetically related to other viruses causing global human disease including dengue virus (DENV), yellow fever virus, and Japanese and tick-borne encephalitis viruses. WNV cycles in nature between birds and Culex mosquitoes but can infect and cause severe neuroinvasive disease in other vertebrate animals including humans (25). While the majority of human WNV infections are asymptomatic, a subset of individuals develops a systemic febrile illness with few progressing to meningitis, encephalitis or an acute flaccid paralysis syndrome (32). Severe and symptomatic WNV infection occurs more frequently in the elderly or immunocompromised and in individuals homozygous for the CCR5Δ32 mutation (11, 18, 19, 24). Since 1999, in the United States, more than 30,000 cases of symptomatic WNV infection have been confirmed, although seroprevalence analysis estimates a much greater number of infections in the population (4). Currently there is no approved therapy or vaccine for WNV in humans. Studies in mice have helped to elucidate how the integrity of the innate and adaptive immune system is required for resistance to WNV infection. Using mice with targeted deletions in individual immune response genes, significant and protective contributions from inflammatory cytokines (e.g., type I and II interferon), chemokines, complement, B cells, and CD4 + and CD8 + T cells have been observed (reviewed in (7, 27)). In particular, receptor-ligand interactions of two members (TNF-α-TNF-α receptor and Fas-Fas ligand) of the tumor necrosis factor (TNF) superfamily of proteins are protective against WNV within the central nervous system. Antigen-specific CD8 + T cells use Fas ligand effector mechanisms to contain WNV infection in Fas-expressing 3
4 neurons in the CNS (34) whereas TNF-α interaction with TNF-R1 protects against WNV infection by regulating migration of protective inflammatory cells into the brain during acute infection (38). TNF-related apoptosis-inducing ligand (CD253 or TRAIL) is a 281 amino acid type II transmembrane protein of the TNF superfamily. The C-terminal extracellular domain is cleaved by cell-associated proteases, resulting in a soluble TRAIL that forms a homotrimer and circulates in the bloodstream. Soluble or cell-associated human TRAIL binds to the death receptors DR4 (TRAIL-R1) and DR5 (TRAIL-R2) on tumor cells, resulting in recruitment of FADD (Fas-associated death domain), activation of caspase-8, and apoptosis of transformed cells. In addition, TRAIL also binds to decoy receptors DcR1 (TRAIL-R3) and DcR2 (TRAIL-R4) that either lack or contain a truncated cytoplasmic domain. TRAIL binding to DcR1 and DcR2 does not promote apoptosis but instead neutralizes TRAIL function (22) or promotes NF-κB activation and transcription of pro-inflammatory genes (13, 20). TRAIL binding to its receptors on non-tumor cells commonly does not trigger apoptosis. Such regulation is controlled by the binding of the endogenous inhibitor of death receptor killing, cellular FLICE-Like inhibitory protein (c- FLIP) to the intracellular domain of TRAIL receptors (23) and by expression of the X- linked inhibitor of apoptosis protein (XIAP) (26). Thus, the physiological role of TRAIL remains less well understood. TRAIL is induced as a consequence of type I and II interferon (IFN) signaling in several cell types (14) and has been reported to have antiinflammatory properties (30), which can mitigate excessive and maladaptive host immune responses in the brain (12). In mice, TRAIL binding to DR5 has been proposed to negatively regulate cellular innate immune responses (9). 4
5 TRAIL also has been suggested to have inhibitory functions against several viruses in vivo. Administration of a blocking but non cell-depleting anti-trail antibody to mice during influenza virus infection delayed virus clearance in the lung (15), and analogously, TRAIL -/- mice showed increased disease severity after influenza infection (2, 3). In addition, type I IFN signaling induced TRAIL expression in NK cells, which facilitated control of encephalomyocarditis virus in vivo (29). Some of the antiviral effect of TRAIL may be mediated by the decoy receptors DcR1 and DcR2, as DR5 -/- mice show lower titers of murine cytomegalovirus in the spleen and higher levels of type I IFN (9). More recently, a role for TRAIL-dependent protection against flaviviruses was suggested (40). DENV infection induced TRAIL expression in blood-derived immune cells, primary myoblasts, and human endothelial cells in a type I IFN-dependent manner, and exogenous recombinant TRAIL inhibited DENV infection in human myeloid cells through an apoptosis-independent mechanism. To determine the physiological relevance of TRAIL in the context of infection by a flavivirus, we compared the virulence of WNV in wild type and TRAIL -/- mice. We found that a deficiency of TRAIL was associated with increased viral burden in the CNS, delayed clearance, and enhanced mortality despite normal priming of adaptive B and T cell immune responses and trafficking to the brain. However, CD8 + T cells showed qualitative defects in their ability to clear WNV infection. Our experiments suggest that CD8 + T cells utilize TRAIL to control WNV infection in neurons
6 MATERIALS AND METHODS Virus. The WNV strain ( ) was isolated in New York in 2000 and passaged once in C6/36 Aedes albopictus cells to generate a stock virus as described previously (10). Mouse experiments. C57BL/6 wild-type inbred mice were commercially obtained (Jackson Laboratories, Bar Harbor, ME). The congenic, backcrossed TRAIL -/- mice were obtained from Dr. Jonathan Weiss (NIH, Bethesda, MD) and re-derived prior to initiation of infection experiments. All mice were genotyped and bred in the animal facilities of the Washington University School of Medicine under pathogen-free conditions, and experiments were performed in accordance with and approval of the Washington university animal studies guidelines. Eight to 10-week-old mice were inoculated with 10 1 and 10 2 plaque-forming units (PFU) by intracranial and subcutaneous route, respectively. Quantitation of tissue viral burden and viremia. To determine the extent of viral spread in vivo, wild type and TRAIL -/- mice were infected with WNV by subcutaneous (via footpad injection) or intracranial inoculation and euthanized at specific time points. Blood was collected by intracardiac puncture, and serum was isolated and stored aliquotted at -80ºC. After extensive tissue perfusion with PBS at 4ºC, organs were harvested, homogenized using a bead beater apparatus, and infectious virus was titered by plaque assay on BHK21-15 cells (8). For measurement of infectious virus in serum, plaque assays were performed on Vero cells (35). Measurement of WNV-specific antibodies. To determine the levels of WNVspecific antibody, IgM and IgG were measured using an enzyme-linked immunosorbent assay (ELISA) with purified WNV E protein as described previously (21). The titer was 6
7 defined as the serum dilution yielding an optical density at 450 nm equivalent to three times above the background of the assay. The titer of neutralizing antibody was determined by using a plaque reduction neutralization assay with BHK21-15 cells (36). Plaques were counted visually, plotted and the plaque reduction neutralization titer for 50% inhibition (PRNT50) was calculated using GraphPad Prism software. Analysis of splenic CD4 + and CD8 + T cells. WNV-specific CD4 + and CD8 + T cells in the spleen were analyzed as described previously (34). Briefly, splenocytes were harvested from wild type and TRAIL -/- mice on day 8 after infection. After lysis of erythrocytes in hypotonic solution, 10 6 cells were stimulated with 0.2 µg/ml of an immunodominant D b -restricted WNV-specific NS4B peptide (SSVWNATTAI) or anti- CD3 antibody (2C11) for 6 hours at 37ºC in the presence of Golgi plug (BD Biosciences). Splenocytes were stained with anti-cd4 or anti-cd8 antibody (BD Biosciences) for 30 minutes at 4ºC. After washing in PBS supplemented with 5% goat serum, cells were fixed with 1% paraformaldehyde in PBS, permeabilized with saponin, and stained with anti-ifn-γ or anti-tnf-α antibody (BD Biosciences) for 30 minutes at 4ºC. After a final series of washes, cells were processed by flow cytometry and the percentage of CD8 + and CD4 + T cells that expressed IFN-γ or TNF-α was determined using CellQuest (BD Biosciences) and FlowJo (Treestar) software. In separate experiments, splenocytes were stained at 4ºC for 30 minutes with FITC-conjugated anti- CD8 antibody (BD Biosciences) and PE-conjugated D b -NS4B peptide (SSVWNATTAI) tetramers (prepared by the NIH tetramer facility, Atlanta GA). Uninfected mice were used as controls in all experiments. The number and percentage of CD4 + CD25 + FoxP regulatory T cells in the spleen of WNV-infected wild type or TRAIL -/- mice was 7
8 measured using the regulatory T cell staining kit (ebioscience). Samples were processed by multi-color flow cytometry on an LSR II flow cytometer (Becton Dickinson) and analyzed with FlowJo software (Treestar). CNS leukocytes. Leukocytes were isolated and quantified from the brains of infected animals as described previously (36). Briefly, eight days after subcutaneous infection with 10 2 PFU of WNV, brains were harvested after extensive perfusion with PBS, dispersed into single-cell suspensions, and digested with 0.05% collagenase D, 0.1 µg/ml trypsin inhibitor TLCK (Nα-p-tosyl-L-lysine chloromethyl ketone), 10 µg/ml DNase I (all from Sigma Chemical), and 10 mm of HEPES ph 7.3 in Hanks balanced salt solution for one hour. Leukocytes were isolated by discontinuous Percoll gradient (70%37%/30%) centrifugation for 30 minutes (850 x g at 4ºC). After washing and counting, cells were stained for CD4, CD8, CD45, and CD11b using directly conjugated antibodies (BD Biosciences) for 30 minutes at 4ºC then fixed with PBS supplemented with 1% paraformaldehye. In some experiments, isolated brain leukocytes were restimulated with D b -restricted NS4B peptide or anti-cd3 antibody for 6 hours and IFNγ- and TNF-α-producing CD8 + T and CD4 + T cells were measured. Alternatively, brain leukocytes were incubated with the NS4B tetramer and anti-cd8 to detect WNV-specific CD8 + T cells. Samples were processed by multi-color flow cytometry on an LSR II flow cytometer and analyzed with FlowJo software. Primary cell isolation and infection. Primary cortical neurons were prepared from 15 days old embryos as described previously (28). Cortical neurons were seeded in 24-well poly-d-lysine- and laminin-coated plates in neurobasal medium containing B27 and L-glutamine (Invitrogen) for 24 hours. The medium was replaced and neurons were 8
9 cultured for three additional days prior to infection. Multistep virus growth analysis was performed with primary cells at a multiplicity of infection (MOI) of Supernatants were harvested at 6, 24, 48 and 72 hours after infection and titrated by plaque assay on BHK21-15 cells, as described previously (8). In some experiments, recombinant TRAIL (Enzo Life Sciences, to 20 μg/ml) was added 24 hours before or one hour after WNV infection. Adoptive transfer of wild type or TRAIL -/- CD8 + T cells. CD8 + T cells from WNV-infected mice were purified and transferred as described previously (37). Splenocytes were harvested from naïve or primed wild type or TRAIL -/- mice at day 7 after WNV infection. CD8 + T cells were purified by positive selection using antibodycoated magnetic beads (Miltenyi Biotec, Auburn, CA) and transferred into CD8 -/- mice one day after WNV infection. In pilot experiments, we confirmed that equivalent numbers and percentages of wild type (4.0% of total lymphocytes) or TRAIL -/- (4.5% of total lymphocytes) primed CD8 + T cells were present in the blood of separate recipient mice two days after adoptive transfer. For virological studies, recipient mice were sacrificed at day 10 after initial infection and tissues were analyzed for viral burden by plaque assay. Addition of CD8 + T cells to WNV-infected neurons. Incubation of purified CD8 + T cells with WNV-infected neurons was performed as described previously (37). Cortical neurons were infected at an MOI of One hour later, unbound virus was removed with four washes of warm medium. Subsequently, purified naïve or WNV-primed CD8 + T cells from wild type or TRAIL -/- mice were added at an effector to target (E:T) ratio of 50:1. Supernatants were harvested at 24 and 48 hours after infection, and WNV 9
10 production was measured by viral plaque assay. In some experiments, recombinant TRAIL (10 μg/ml) or neutralizing anti-trail antibody or isotype control antibody rat IgG2a (BioLegend) was added at the same time as the CD8 + T cells. TRAIL, DR5, FasL and granzyme B staining. Brains were harvested from wild type or TRAIL -/- mice on day 8 after infection. After isolating leukocytes, CD4 + and CD8 + T cells were stained for expression of TRAIL using a biotin labeled anti-mouse CD253 (TRAIL) antibody (ebioscience), for FasL using an anti-mouse CD95L antibody (Biolegend), and for intracellular granzyme B using an anti-mouse granzyme B antibody (Invitrogen) for 30 minutes at 4ºC. Data collection and analysis were performed with a FACSCalibur flow cytometer and CellQuest Software (BD Biosciences). Cortical neurons were isolated as described previously (37), and infected with WNV (MOI of 0.001) for one hour. One day later, cells were fixed with 4% paraformaldehyde in PBS at 4 C for 10 minutes. After several washes in PBS and blocking in 5% normal goat serum, neurons were stained with rabbit anti-mouse DR5 (10 µg/ml anti-trail-r2, Millipore) for one hour at room temperature. After rinsing in PBS supplemented with 0.05% Triton- X-100, cells also were stained with WNV-immune rat serum (1:100 dilution) for one hour at room temperature. Fluorescence was detected after incubation with Alexa 488 (Molecular Probes) and Cy3 (Jackson Laboratories) conjugated secondary antibodies for 30 minutes at room temperature. Cells were visualized after counterstaining with ToPro-3 (Molecular Probes) using a Zeiss 510 Meta LSM confocal microscope Data analysis. All data were analyzed statistically using Prism software (GraphPad4, San Diego, CA). Kaplan-Meier survival curves were analyzed by the log rank test. Differences in viral burdens in mice were analyzed by the Mann-Whitney test. 10
11 Differences in viral infection in primary cell cultures and numbers of T cells were analyzed by an unpaired t test Downloaded from on October 1, 2018 by guest 11
12 RESULTS TRAIL is required to protect mice from lethal WNV infection. Previous in vitro studies suggested that addition of high doses (e.g., 5 to 20 μg/ml) of exogenous recombinant human TRAIL to cell cultures could inhibit infection of DENV, a related flavivirus (40). To assess directly the role of TRAIL in the context of WNV infection in vivo, we compared the survival rates of 8 to 9 week-old wild type and congenic TRAIL -/- mice after subcutaneous infection with 10 2 PFU of a North American WNV isolate (New York, 2000). By day 8 after infection, all wild type and deficient mice showed clinical signs of infection, including reduced activity, weight loss, and hair ruffling. However, survival rates of the TRAIL -/- mice were markedly lower (17% compared to 75%, P < ; Fig 1A). Similarly, we observed a significant decrease in the mean time to death ( versus days, P < 0.04) in TRAIL -/- compared to wild type mice. Thus, an absence of TRAIL results in a more severe phenotype after WNV infection with poorer clinical outcome. A deficiency of TRAIL results in enhanced WNV tissue burden primarily in the central nervous system. To begin to understand how an absence of TRAIL conferred increased susceptibility to WNV infection, we measured the levels of infectious virus in tissues. Wild type and TRAIL -/- mice were infected by subcutaneous route, and the viral loads were analyzed on days 2, 4, 6, 8, and 10 after infection. (a) Peripheral tissues. We observed was no significant difference in the kinetics and magnitude of WNV infection in serum or the spleen between wild type and TRAIL -/- mice throughout the time course (Fig 1B and C, P > 0.3). Moreover, no infectious virus was detected in the liver in either wild type or TRAIL -/- mice (data not shown). In 12
13 comparison, infectious WNV was recovered from the kidney (3 of 10 mice) at day 4 in a subset of TRAIL -/- but not from wild type mice (data not shown). The kidney usually does not support WNV infection in wild type C57BL/6 mice, whereas replication can be observed in congenic mice lacking aspects of cell-intrinsic immunity (e.g., IFNαβR -/-, IRF-7 -/-, or IRF-3 -/- mice (5, 6, 28)). Overall, TRAIL appears to have to a limited role in controlling WNV infection in peripheral organs. (b) CNS tissues. In the brain and spinal cord, similar levels of WNV (10 4 to 10 5 PFU/g) were detected in wild type and TRAIL -/- mice through day 8 after infection. However, by day 10, significantly higher levels were measured in the brain ( versus PFU/g; P = 0.01) and spinal cord ( versus PFU/g; P = 0.01) of TRAIL -/- mice (Fig 1D and 1E). These experiments suggest that TRAIL functions primarily to control WNV in CNS tissues. TRAIL signaling does not directly control WNV replication in the CNS. Based on the viral burden data after subcutaneous infection, we hypothesized that a lack of TRAIL signaling in resident cells of the CNS might contribute to higher WNV titers in the brain through direct inhibitory effects on neuronal infection. To test this, wild type and TRAIL -/- mice were infected with 10 1 PFU of WNV directly into the cerebral cortex via an intracranial route and monitored for local replication and dissemination. Viral burden in the cerebral cortex, subcortical white matter, brain stem, cerebellum, and spinal cord were measured at days 2, 4, and 6 after infection (Fig 2A-E). Somewhat surprisingly, no differences in viral burdens were observed at any time point in the different CNS regions of wild type and TRAIL -/- mice (P > 0.2). Similarly, we observed no difference in relative morbidity between wild type and TRAIL -/- mice after intracranial 13
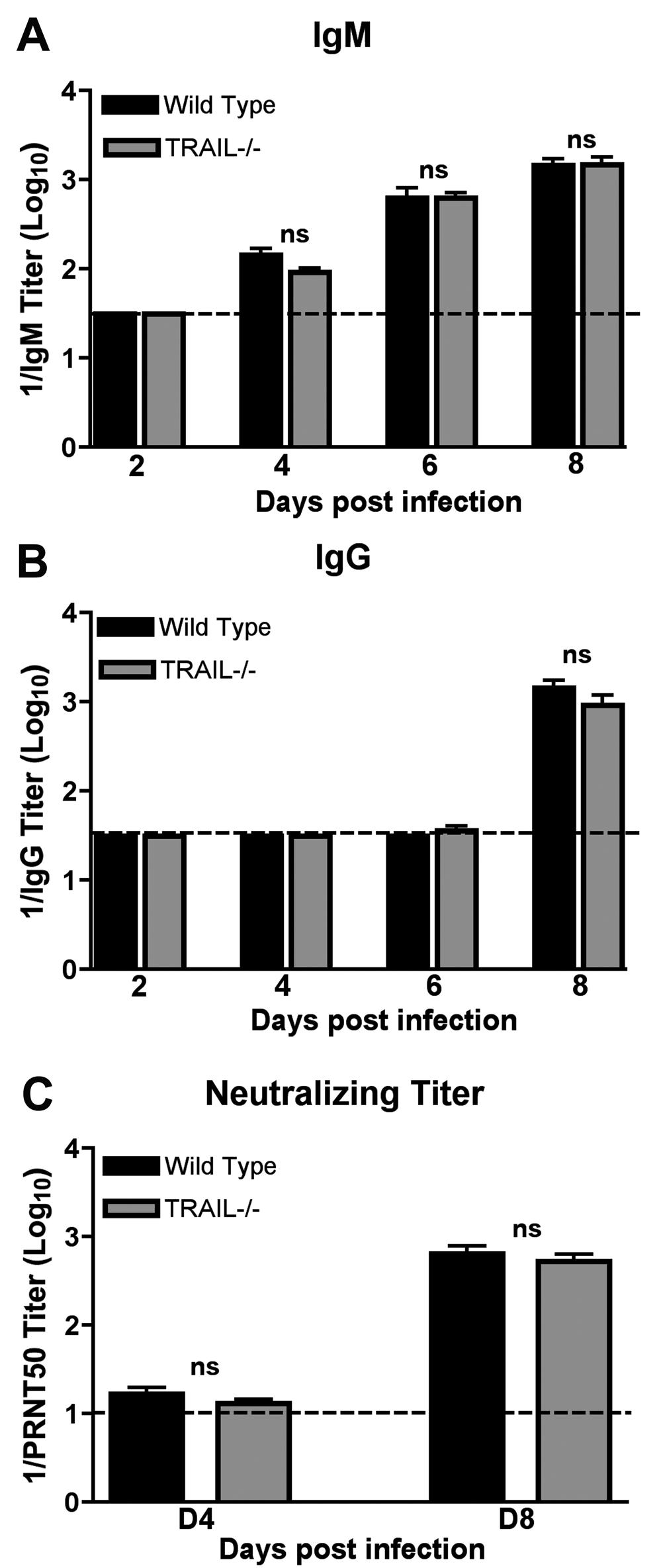
14 WNV infection. These results suggest that TRAIL signaling within the CNS by resident cells does not directly control WNV infection. Adaptive immune responses in TRAIL -/- mice. Since we observed elevated viral burden in CNS tissues after subcutaneous but not intracranial infection, we speculated that TRAIL might modulate adaptive immune functions (B or T cells), which are required for efficient clearance of WNV from the brain (8, 17, 35, 39). To assess this, at different time points after infection, we measured the magnitude of the adaptive immune response to WNV infection. (a) Antibody response. The kinetics and magnitude of anti-wnv IgM and IgG responses were virtually identical in wild type and TRAIL -/- mice (P > 0.9, Fig 3A and B). Similarly, there was no difference in neutralizing titer in wild type and TRAIL -/- mice at days 4 and 8 after infection (P > 0.4, Fig 3C). Thus, TRAIL was not required for the development of WNV specific antibodies, and increased viral burden in the brain and spinal cord was not due to an obvious defect in antibody production. (b) T cell response. To evaluate a possible role of TRAIL in priming of WNVspecific T cells in the periphery, splenocytes from wild type and TRAIL -/- mice were harvested at day 7 after infection and restimulated ex vivo with an immunodominant D b - restricted NS4B peptide or anti-cd3 antibody, and CD4 + and CD8 + T cells were analyzed for the production of intracellular IFN-γ and TNF-α by flow cytometry. We observed no significant differences (P > 0.2) in the percentage of IFN-γ + or TNF-α + CD8 + T cells from wild type and TRAIL -/- mice after incubation with WNV-specific peptide (Fig 4A and B). We did however detect a slight increase in the percentage (7% versus 5%, P < 0.002) but not number (P > 0.2) of NS4B-tetramer positive CD8 + T cells in 14
15 TRAIL -/- mice (Fig 4C and D). In comparison, no difference in the percentage or absolute numbers of CD4 + T cells producing IFN-γ and TNF-α was observed between wild type mice and TRAIL -/- mice (P > 0.4) (Fig 4E and F). Finally, we also did not observe a difference (P > 0.1) in the percentage or number of CD4 + CD25 + FoxP3 + regulatory T cells in WNV-infected wild type or TRAIL -/- mice (Fig 4G, and data not shown). Overall, these experiments demonstrate that an absence of TRAIL does not affect induction of adaptive B and T cell responses after WNV infection. Trafficking of leukocytes to the brain is not impaired in TRAIL -/- mice. Although peripheral T cell responses were unchanged in TRAIL -/- mice after WNV infection, we assessed whether the enhanced viral replication in the CNS of TRAIL -/- mice might be related to blunted activation of resident microglia or migration of protective macrophages and CD8 + T cells. To assess this, leukocytes were isolated from brains of wild type and TRAIL -/- mice at day 8 after infection by gradient centrifugation and analyzed by flow cytometry. Notably, we failed to observe a difference in percentages or numbers of activated CD11b + CD45 lo microglia or macrophages (CD11b + CD45 hi ) (P > 0.4; Fig 5A and B). We did find, however, a higher percentage and number (P < 0.03, Fig 5C and D) of total CD8 + T cells in the brains of TRAIL -/- mice. This correlated with increased numbers of CD8 + T cells from TRAIL -/- mice expressing IFN-γ (P < 0.04, Fig 5E) after WNV-specific peptide restimulation ex vivo, or that stained directly with NS4B tetramers (P = 0.05, Fig 5F). In comparison, while similar percentages and numbers (P > 0.4) of CD4 + T cells and CD19 + B220 + B cells were observed, we measured greater numbers of IFN-γ + CD4 + T cells after ex vivo restimulation from TRAIL -/- compared to wild type mice (P < 0.04, Fig 5G). Overall, these data show no clear deficiency in 15
16 recruitment or activation of leukocytes in the brains of WNV-infected TRAIL -/- mice; in general, higher levels of T cells were observed likely secondary to the increased viral burden. Adoptive transfer of wild type and TRAIL -/- CD8 + T cells. In addition to Fas-Fas ligand or perforin/granzyme cytolysis mechanisms, CD8 + T cells and NK cells can secrete TRAIL to lyse target cells (1, 3, 41). Since an absence of CD8 + T cells or their specific effector functions resulted in a failure to control WNV replication in the brain and spinal cord (34, 35, 37, 39), we hypothesized that a deficiency of TRAIL in CD8 + T cells could explain the enhanced viral replication in the CNS of TRAIL -/- mice. To evaluate this, we performed adoptive transfer studies. Primed CD8 + T cells were purified from the spleens of WNV-infected wild type or TRAIL -/- mice on day 7 by positive selection using antibody-coated magnetic beads. We adoptively transferred 10 x 10 6 CD8 + T cells into congenic CD8 -/- mice one day after WNV infection. Nine days later, we assessed the viral burden in spleen and CNS tissues of recipient CD8 -/- mice. Adoptive transfer of 10 x 10 6 naïve wild type CD8 + T cells failed to control infection, as high levels of WNV were apparent in the spleen and CNS of recipient CD8 -/- mice (Fig 6A). Transfer of 10 x 10 6 WNV-primed wild type CD8 + T cells, however, efficiently controlled infection, as no virus was detected in the target tissues. In comparison, while adoptive transfer of primed TRAIL -/- CD8 + T cells cleared virus from the spleen, they failed to control WNV in the CNS, as recipient mice (4 of 4) sustained infection (~10 4 to 10 5 PFU/g). The viral burden in the CNS of CD8 -/- mice receiving TRAIL -/- CD8 + T cells nonetheless was lower than those receiving naïve wild type cells ( PFU/g versus PFU/g, P = 0.01), confirming TRAIL-independent clearance mechanisms by CD8 + T 16
17 345 cells To corroborate the role of TRAIL in CD8 + T cell-mediated clearance of WNV from the CNS, a lower number of (3 x 10 6 ) of primed CD8 + T cells was adoptively transferred (Fig 6B). Nine days after transfer, WNV was not detected (< PFU/g) in the spleen of recipient CD8 -/- mice that had received primed wild type cells; however, infectious WNV was recovered from mice receiving primed TRAIL -/- CD8 + T cells (~ PFU/g). Higher levels of WNV also were recovered from the CNS of CD8 -/- mice receiving 3 x 10 6 primed CD8 + T cells from TRAIL -/- compared to wild type mice ( to PFU/g, P < 0.02). Consistent with these findings, adoptive transfer of 3 x 10 6 WNVprimed but not naïve CD8 + T cells into recipient TRAIL -/- mice resulted in an improved survival rate (71% versus 20%, P = 0.05). Overall, the adoptive transfer experiments demonstrate that a deficiency of TRAIL in CD8 + T cells negatively impacts control of WNV in mice. Delayed WNV clearance in the CNS of TRAIL -/- mice. Given our adoptive transfer study results, we hypothesized that TRAIL was required in the CNS for efficient clearance by CD8 + T cells. As prior studies showed that a deficiency of CD8 + T cell effector molecules (e.g., perforin or Fas-ligand) caused WNV persistence in the CNS for several weeks (34, 37), we evaluated the impact of a TRAIL deficiency on the kinetics of viral clearance in the brain and spinal cord. Infectious virus was measured in the few surviving wild type and TRAIL -/- mice at days 15, 21, and 28 after inoculation (Fig 6C and D). Low levels (10 2 to 10 3 PFU/g) of infectious WNV were detected on day 15 in 12.5% (1 of 8) and 25% (2 of 8) of brains and spinal cords, respectively of wild type mice. No infectious virus was recovered from the CNS at 21 days or after in wild type 17
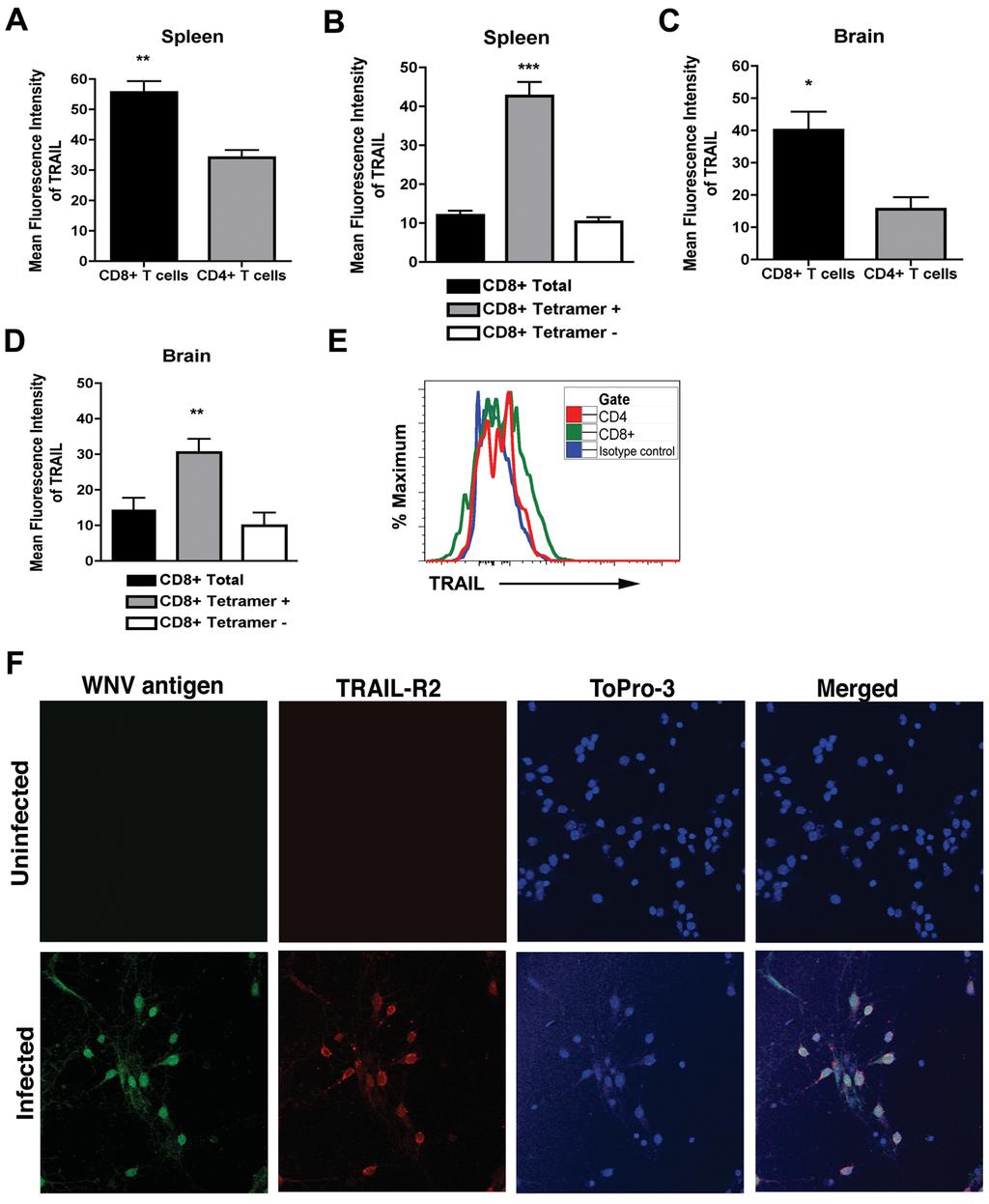
18 mice. In comparison, higher levels of WNV (10 2 to 10 6 PFU/g) were detected in the CNS (67%; 6 of 9, P < 0.005) of TRAIL -/- mice on day 15. Moreover, on day 21, 10 (1 of 10) to 20% (2 of 10) of TRAIL -/- mice had detectable yet low levels (10 2 to 10 3 PFU/g) of infectious virus in the brain and spinal cord, respectively. By day 28, no infectious virus was recovered from the CNS of any TRAIL -/- mice. Thus, in the surviving mice, an absence of TRAIL resulted in somewhat delayed clearance of infections WNV from the brain and spinal cord. TRAIL -/- CD8 + T cells are impaired in their ability to clear WNV from infected neurons. To demonstrate directly that CD8 + T cells require TRAIL to control WNV infection of neurons, we used a viral, clearance assay with primary cortical neurons (37). One hour after infection, bulk WNV-primed wild type or TRAIL -/- splenic CD8 + T cells were added to neurons at an effector to target ratio of 50:1. At 24 and 48 hours, the level of infectious virus in the supernatants of neuronal cultures was titrated by plaque assay. As seen previously (34), we observed reduced (2 to ~1,300-fold, P < 0.03) viral yield in supernatants from neurons incubated with wild type primed compared to naïve or no CD8 + T cells at 24 and 48 hours, respectively (Fig 6E and F). In comparison, WNVprimed TRAIL -/- CD8 + T cells less efficiently restricted infection in neurons, with no difference at 24 hours (P > 0.7) and a smaller (23-fold, P < 0.02) reduction at 48 hours. These experiments suggest that WNV primed CD8 + T cells control neuronal infection, in part through TRAIL-dependent mechanisms. As additional confirmation, we assessed surface expression of TRAIL on WNVprimed CD8 + T cells in the spleen and the brain. Wild type mice were infected with 10 2 PFU by subcutaneous inoculation, and at day 8 splenocytes and brain leukocytes were 18
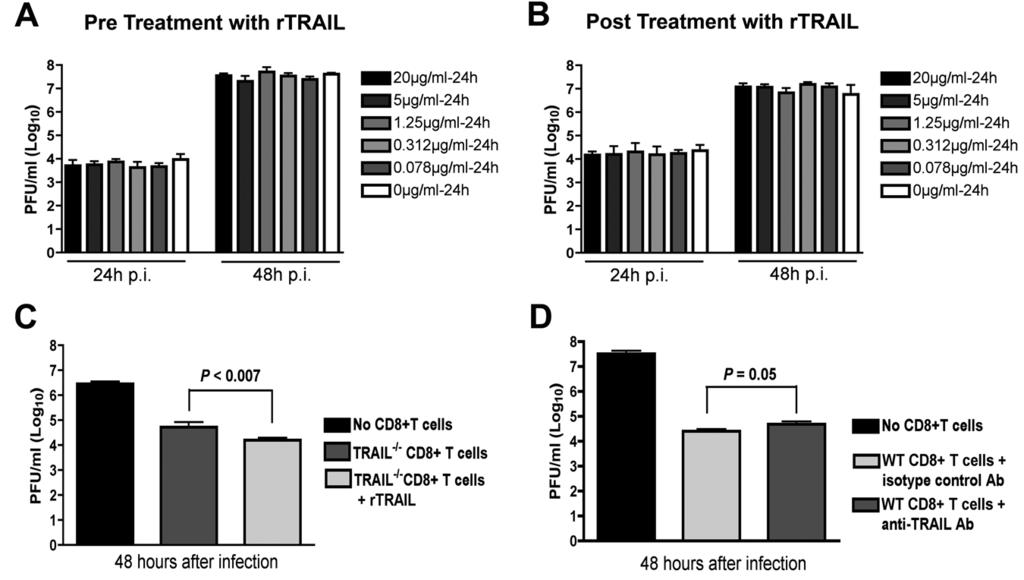
19 harvested and stained with anti-trail antibody. In the spleen, TRAIL was expressed at higher levels on CD8 + T cells compared to CD4 + T cells (mean fluorescence intensity of 56 ± 4 for CD8 + T cells and 34 ± 2 for CD4 + T cells, P < 0.006) (Fig 7A). Among total CD8 + T cells, higher levels (P < ) of TRAIL were measured on NS4B tetramer positive CD8 + T cells (Fig 7B). Analogous to data from the spleen, higher levels of TRAIL were detected on CD8 + T cells isolated from the brain compared to CD4 + T cells (mean fluorescence intensity of 40 ± 6 for CD8 + T cells and 16 ± 4 for CD4 + T cells, P < 0.02) (Fig 7C and E), and greater surface expression of TRAIL was measured on NS4B tetramer positive compared to tetramer negative CD8 + T cells (Fig 7D). These data confirm that WNV-specific CD8 + T cells express TRAIL in the spleen and brain isolated leukocytes after WNV infection. Given that CD8 + T cells in the brain expressed TRAIL, we assessed whether WNV-infected neurons expressed its cognate ligand, DR5. In the absence of infection, cortical neurons showed no apparent DR5 expression (Fig 7F, upper panels). However, within 24 hours, WNV infection resulted in efficient surface expression of DR5 (Fig 7F, lower panels). Thus, WNV infection rapidly induces DR5 expression on neurons, which could sensitize them to the inhibitory effects of TRAIL. CD8 + T cells require TRAIL to optimally control WNV infection in neurons. Our CD8 + T cell adoptive transfer and co-incubation experiments with neurons strongly suggested an inhibitory effect of TRAIL on viral replication in neurons. However, they did not provide direct insight into mechanism. We speculated that TRAIL might have a direct antiviral effect, as was seen previously with the related flavivirus, DENV (40). To address this, we added increasing concentrations of recombinant TRAIL (0.078 to 20 μg/ml) one day prior to WNV infection (MOI of 0.001) of primary cortical neurons in the 19
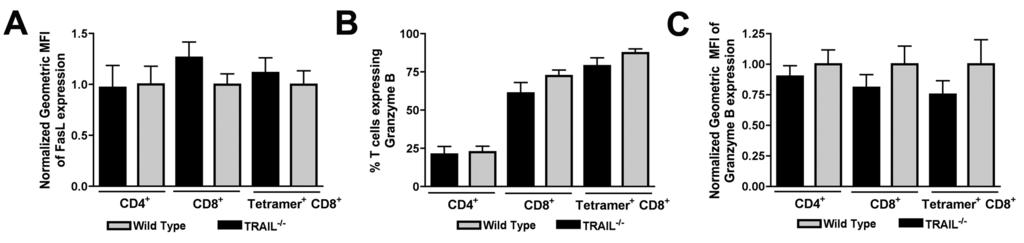
20 absence of CD8 + T cells. Supernatants were collected from neurons at 24 and 48 hours after infection and virus burden was analyzed by plaque assay. Notably, we observed no reduction in WNV yield with any dose of TRAIL when added prior to infection (Fig 8A). Even when experiments were repeated with addition of recombinant TRAIL immediately after infection, no direct antiviral effect of TRAIL was observed (Fig 8B). Given these results, we suspected that the antiviral effect of TRAIL on WNV-infected neurons needed to occur in the context of CD8 + T cell-mediated clearance. Indeed, the addition of recombinant TRAIL to primed TRAIL -/- CD8 + T cells reduced WNV-infection in cortical neurons (Fig 8C; no TRAIL versus with TRAIL, 2.5-fold; P < 0.007). Reciprocally, addition of a neutralizing anti-trail antibody to wild type primed CD8 + T cells also resulted in increased WNV infection of neurons (Fig 8D: isotype control versus anti- TRAIL antibody, 3.2-fold; P = 0.05). Thus, TRAIL provides an antiviral signal to neurons but only when CD8 + T cells were present. Because of the modest effects of exogenous TRAIL and anti-trail antibodies on CD8 + T cell-mediated clearance of WNV infection in neurons, we speculated that TRAIL -/- CD8 + T cells might have additional defects in their effector functions. As prior studies had already established protective effects of cytolytic granules (perforin/granzyme) and cell-death (Fas ligand) pathways in T cell protection against WNV (34, 37), we assessed their relative expression in wild type and TRAIL -/- CD8 + T cells in the brain. Relative FasL expression was unchanged on the surface of TRAIL -/- CD4 + and CD8 + T cells in the brain (P > 0.2) (Fig 9A). This result was also observed on WNV-specific NS4B-tetramer-positive CD8 + T cells. Similarly, in the brain, granzyme B was expressed at similar levels (P > 0.2) in CD4 + and CD8 + T cells and NS4B tetramer- 20
21 positive CD8 + T cells (Fig 9B and C). Thus, the functional defect of TRAIL -/- CD8 + T cells in the context of clearance of WNV infection in the brain was not due to impaired expression of proteins associated with alternate cytolytic mechanisms Downloaded from on October 1, 2018 by guest 21
22 DISCUSSION TRAIL has been implicated in the control of viral infections by several different mechanisms including cytotoxic and direct antiviral effects. We initiated this study because primary human cell culture experiments showed that TRAIL was induced after DENV infection, and treatment of human myeloid cells with exogenous, recombinant TRAIL inhibited DENV infection through an apoptosis-independent mechanism (40). To address the physiological function of TRAIL in vivo in the context of infection by a flavivirus, we assessed WNV pathogenesis in wild type and TRAIL -/- mice. Mice lacking TRAIL showed relatively normal levels of WNV infection in peripheral tissues but sustained elevated viral titers in the brain and spinal cord that was associated with delayed clearance and increased mortality compared to wild type mice. No difference in WNV infection was observed in the brain and spinal cord after direct intracranial inoculation. While priming of adaptive B and T cell responses in the periphery, and trafficking of immune and antigen-specific cells to the brain was normal in TRAIL -/- mice, CD8 + T cells showed qualitative defects in their ability to clear WNV infection. Adoptive transfer studies suggested that the increased susceptibility associated with a TRAIL deficiency was due to defects in CD8 + T cell clearance mechanisms. Indeed, primed TRAIL -/- CD8 + T cells less efficiently controlled WNV infection in primary neuronal cultures ex vivo. Our experiments suggest that CD8 + T cells utilize TRAIL to control WNV pathogenesis and infection of neurons in the CNS. This conclusion is based on several experimental observations: (a) increased viral burden in the brain and spinal cord at day 10 but not day 6 or 8 after infection in TRAIL -/- mice; (b) delayed clearance of WNV 22
23 infection in the few surviving TRAIL -/- mice; (c) less efficient clearance of infection in CD8 -/- mice after adoptive transfer of WNV-primed TRAIL -/- compared to wild type CD8 + T cells; and (d) less efficient control of WNV infection in neurons ex vivo by primed TRAIL -/- compared to wild type CD8 + T cells. This phenotype was not due to TRAILdependent effects on CD8 + T cell priming or trafficking, as equivalent percentages and numbers of polyfunctional antigen-specific CD8 + T cells were detected in WNV-infected TRAIL -/- and wild type mice. Our results are consistent with recent studies with influenza virus, which showed that a deficiency of TRAIL enhanced morbidity and viral replication through qualitatively decreased influenza-specific CD8 + T cell responses in the lung (3). Prior studies have shown that CD8 + T cells also utilize perforin and Fas ligand to clear WNV from infected neurons (34, 37). Analogous to our current studies with TRAIL - /- mice, perforin and Fas-ligand-deficient (gld) mice had elevated viral titers in the brain and spinal cord resulting in increased mortality compared to wild type mice, and in the few mice that survived infection persistent WNV infection was detected for several weeks. However, there appears to be a functional hierarchy, as a deficiency of perforin or Fas ligand resulted in less efficient clearance of WNV infection in the spleen, whereas this phenotype was absent in TRAIL -/- mice. Thus, while CD8 + T cells utilize multiple effector molecules to clear WNV from neurons in the CNS, the viral burden and survival phenotypes in TRAIL -/- mice were subordinate compared to those observed previously in perforin -/- and gld mice. Prior cell culture studies with DENV and HIV suggested that an antiviral effect of exogenous treatment of TRAIL occurs in the absence of apoptosis (33, 40). Although an analogous effect of recombinant TRAIL on WNV infection in cultured primary neurons 23
24 was not observed, an inhibitory effect was seen in the context of a protective CD8 + T cell response. The lack of direct antiviral effect of exogenous TRAIL against WNV was consistent with some of our in vivo results: (a) we observed no difference in viral burden in peripheral tissues after subcutaneous infection; (b) we failed to detect differences in viral replication in multiple CNS regions after direct intracranial inoculation of wild type and TRAIL -/- mice. However, TRAIL did show an antiviral effect in the context of CD8 + T cell mediated clearance of WNV infection in neurons. This observation was most apparent in reconstitution experiments showing that the addition of exogenous TRAIL to TRAIL -/- CD8 + T cells enhanced the antiviral effect against WNV in neurons. What is the precise mechanism by which TRAIL facilitates optimal CD8 + T cell-mediated control of WNV infection in neurons? While studies with transformed cells in culture have shown that exogenous TRAIL sensitized respiratory syncytial virus- or human cytomegalovirusinfected cells to apoptosis (16, 31), an absence of TRAIL did not specifically affect cell death as judged by TUNEL staining of neurons in the brain or in culture (B. Shrestha and M. Diamond, unpublished data). Although further signaling analysis is warranted, our data is more consistent with a model in which TRAIL produced by CD8 + T cells provides a direct antiviral signal to neurons. Why did recombinant TRAIL alone fail to inhibit WNV infection directly in culture? We speculate that TRAIL derived by WNV-specific CD8 + T cells is necessary but not sufficient to confer the inhibitory effect, and requires additional signals through other cytolytic pathways (e.g. Fas ligand-fas and perforin/granzyme) to achieve an optimal inhibitory effect. Alternatively, activated WNV-specific effector CD8 + T cells secrete other cytokines (e.g., IFN-γ or TNF-α), which modulate expression of TRAIL ligands (e.g., DR4 and DR5) or signaling 24
25 components in neurons to levels that are sufficient for functional consequences. WNV infection alone can induce DR5 expression on neuronal targets in culture and in vivo. 513 In summary, our study demonstrates that CD8 + T cells require TRAIL to optimally restrict WNV infection in the brain and spinal cord. Thus, CD8 + T cells utilize at least three separate effector mechanisms (TRAIL, Fas-ligand, and perforin/granzyme) to control WNV infection and injury of neurons. For highly neurovirulent viruses (e.g., New York strain of WNV), clearance mechanisms by effector CD8 + T cells appear to limit dissemination and disease and outweigh the possible pathological effects of immune targeted neuronal injury. A more complete understanding of the effector mechanisms responsible for clearance of WNV infection may inform novel vaccine strategies that optimize CD8 + T cell-mediated immunity against highly virulent viruses that target neurons in the CNS. 25
26 FIGURE LEGENDS Figure 1. Survival and viral titer analysis of wild type and TRAIL -/- mice infected with WNV. A. Kaplan-Meier survival curves. Wild type (n = 40) and TRAIL -/- (n = 29) mice were infected via subcutaneous route with 10 2 PFU of WNV and monitored for mortality for 28 days. Survival differences were judged by the log rank test and were statistically significant (P < ). B to E. Viremia and WNV tissue burden in wild type and TRAIL -/- mice. Infectious WNV levels in (B) serum, (C) spleen, (D) brain and (E) spinal cord of wild type and TRAIL -/- mice were measured by viral plaque assay from samples harvested at the indicated time points. Data are expressed as log 10 PFU per gram of tissue or ml of serum, and reflect results for 8 to 13 mice per time point between days 2 and 10. For viral burden experiments, the horizontal bars represent the mean titers, dotted line represents the limit of sensitivity of viral detection, and asterisks indicate statistically significant differences (P < 0.05) between wild type and TRAIL -/- mice as determined by the Mann-Whitney test. Figure 2. WNV burden in the CNS after intracranial infection. Wild type and TRAIL -/- mice were infected with 10 1 PFU of WNV via an intracranial route. Different regions of brain (A, cerebellum; B, brain stem; C, cerebral cortex; D, subcortex; and E, spinal cord) were harvested at the indicated time points. Tissue homogenates were analyzed for viral burden by plaque assay. Data are shown as PFU per gram of tissue for 5 to 8 mice per time point. The dotted line represents the limit of detection. None of the differences achieved statistical significance. Figure 3. WNV-specific antibody responses in wild type and TRAIL -/- mice. Wild type and TRAIL -/- mice were infected with WNV via a subcutaneous route and serum was collected at the indicated time points. The development of WNV-specific (A) 26
27 IgM or (B) IgG was determined by ELISA using purified WNV E protein. Data are the average of 5 mice at day 2 and 10 mice at days 4 through 8. (C) Neutralizing antibody response. Neutralizing titers were determined by a PRNT assay. All samples were serially diluted in duplicate and data are expressed as the reciprocal PRNT50, the antibody titer that reduced plaque number by 50%. Data are the average of 10 mice per time point. None of the differences achieved statistical significance (as indicated by n.s.). Figure 4. T cell responses in the spleen of wild type and TRAIL -/- mice after WNV infection. A-D. CD8 + T cells. WNV-infected splenocytes from wild type or TRAIL -/- mice were harvested at day 7 and stimulated ex vivo for six hours with (A, B) an immunodominant D b -restricted NS4B peptide. Cells were co-stained for CD8 and intracellular IFN-γ or TNF-α and processed by flow cytometry. Data is presented as the (A) percentage or (B) total number of positive cells. The (C) percentage and (D) total numbers of antigen specific CD8 + T cells in uninfected or WNV-infected wild type and TRAIL -/- mice as determined by binding to a D b -restricted NS4B tetramer. Asterisks indicate statistically significant (P < 0.05) differences between wild type and TRAIL -/- mice. E-F. CD4 + T cells. WNV-infected splenocytes from wild type or TRAIL -/- mice were harvested at day 7 and stimulated ex vivo for six hours with a stimulatory anti-cd3 antibody. Cells were co-stained for CD4 and intracellular IFN-γ or TNF-α and processed by flow cytometry. Data is presented as the (E) percentage or (F) total number of positive cells. Data from panels A-F represents from the 6 to 10 mice from three independent experiments. In panels A, C, and E, the percentage of IFN-γ, TNF-α, or NS4B tetramer positive cells represents the fraction of total gated CD8 + or CD4 + T cells. G. Regulatory T cells. Splenocytes from WNV-infected wild type (naïve or infected) and TRAIL -/- mice 27
28 were harvested at day 7 and stained with antibodies to CD4, CD25 and FoxP3. Data is expressed as the percentage of CD4 + T cells that stained positive for CD25 and FoxP3. Data were pooled from 6 mice from 2 independent experiments. H. Flow cytometry profiles. Representative flow cytometry profiles of intracellular IFN-γ staining (upper panels) and NS4b tetramer staining (bottom panels) of splenic CD8 + T cells from wild type or TRAIL -/- mice at day 7 after WNV infection. Figure 5. Accumulation of leukocytes in the CNS of WNV-infected wild type and TRAIL -/- mice. Wild type and TRAIL -/- mice were infected with 10 2 PFU of WNV by subcutaneous route. Eight days later, brains were harvested and leukocytes were isolated by Percoll gradient centrifugation. A-B. The percentage (A) and total number (B) of microglia (CD11b + CD45 lo ) and macrophages (CD11b + CD45 hi ) are shown. C-D. The percentage (C) and total number (D) of CD8 + T cells, CD4 + T cells, and CD19 + B220 + B cells are shown. E-F. The number of WNV-specific CD8 + T cells in the brain as judged by (E) NS4B peptide restimulation and intracellular IFN-γ staining or (F) NS4B tetramer staining. G. The number of activated CD4 + T cells in the brain as judged after incubation with a stimulatory anti-cd3 antibody and intracellular IFN-γ staining. Data represent the average of two independent experiments with 6 mice per group. Asterisks indicate statistically significant (P < 0.05) differences between wild type and TRAIL -/- mice. H. Representative flow cytometry profiles of CD3 + CD8 + T cells in the brain (left) from TRAIL -/- mice eight days after WNV infection. Surface NS4b tetramer staining (middle) or intracellular IFN-γ staining after NS4B peptide restimulation ex vivo is shown (right) on brain CD8 + T cells. 28
29 Figure 6. Role of TRAIL in CD8 + T cell mediated control of WNV infection in the brain. A-B. Viral yield in different tissues after adoptive transfer of WNV-primed CD8 + T cells. Donor CD8 + T cells (A, 10 x 10 6 ; B, 3 x 10 6 ) were purified from naïve or WNV-primed wild type and TRAIL -/- mice and transferred into recipient CD8 -/- mice one day after infection. The spleen, brain, and spinal cord of recipient CD8 -/- mice were harvested on day 10 after initial infection and WNV was titrated by plaque assay. Data represents results from 3 to 4 mice in two independent experiments. The dotted line represents the limit of sensitivity of the assay. C-D. WNV persistence in CNS tissues. Wild type and TRAIL -/- mice were infected with WNV and (C) brain and (D) spinal cord were harvested at days 15, 21, and 28 after infection, and viral yield was titrated by plaque assay. Results are from 4 to 9 mice per time point. E-F. Control of WNV infection in primary neurons ex vivo after addition of primed CD8 + T cells. Cortical neurons were infected with WNV at an MOI of After one hour, purified naïve or WNV-primed CD8 + T cells from wild type or TRAIL -/- mice were added at an E:T ratio of 50:1. After (E) 24 or (F) 48 hours, supernatants were harvested and WNV production was measured by plaque assay. Asterisks indicate statistically significant differences in this Figure (*, P < 0.05, ** P < and *** P< ), and n.s. indicates no significant difference. Figure 7. CD8 + T cells express TRAIL. Splenocytes and brain leukocytes from WNV-infected wild type mice were harvested at day 7 and stained for surface expression of TRAIL on CD4 and CD8 + T cells. Data were analyzed by flow cytometry and are expressed as the mean fluorescence intensity only of the positive cells. A. TRAIL expression on total CD4 + and CD8 + T cells in the spleen. B. TRAIL expression on NS4B tetramer positive and tetramer negative CD8 + T cells in the spleen. C. TRAIL expression 29
30 on total CD4 + and CD8 + T cells in the brain. D. TRAIL expression on NS4B tetramer positive and tetramer negative CD8 + T cells in the brain. Data are pooled from four mice, and asterisks indicate differences that are statistically significant. E. Representative flow cytometry profiles after staining with anti-trail antibody on the surface of CD4 + and CD8 + T cells from the brains of mice infected with WNV. F. DR5 expression on neurons. Neurons were uninfected or infected with WNV (MOI of 0.001) for 24 hours. Cells were harvested and stained for surface DR5 and intracellular WNV antigen, counter-stained with ToPro3, and analyzed by confocal microscopy. Representative data from multiple images are shown. Figure 8. Mechanism by which TRAIL inhibits WNV infection in neurons. A. Recombinant TRAIL does not have a direct antiviral effect on cortical neurons. Primary cortical neurons were treated with increasing doses (0.078 to 20 μg/ml) of recombinant TRAIL for 24 hours prior to infection with WNV (MOI of 0.001). One hour later free virus and TRAIL were removed by washing. Supernatants were harvested 24 and 48 hours after infection and titrated by plaque assay. Results are representative of three independent experiments performed in triplicate. B. Post-exposure addition of recombinant TRAIL does not confer an antiviral effect against WNV infection of cortical neurons. Neurons were infected with WNV (MOI of 0.001). One hour later, free virus was removed by extensive washing. Subsequently, increasing doses (0.078 to 20 μg/ml) of recombinant TRAIL were added and supernatants were harvested 24 hours later and titrated by plaque assay. Results are representative of three independent experiments performed in triplicate. None of the doses of recombinant TRAIL conferred a statistically significant antiviral effect. C. Addition of recombinant TRAIL enhances the antiviral 30
31 effect of primed TRAIL -/- CD8 + T cells. Cortical neurons were infected with WNV at an MOI of After one hour, purified WNV-primed CD8 + T cells from TRAIL -/- mice were added at an E:T ratio of 50:1. At the same time, recombinant TRAIL (10 μg/ml) was added to some of the cell cultures. Supernatants were harvested 48 hours later, and WNV production was measured by plaque assay. D. Addition of neutralizing anti-trail antibody decreases the antiviral effect of primed wild type CD8 + T cells. Cortical neurons were infected with WNV at an MOI of After one hour, purified WNV-primed CD8 + T cells from wild type mice were added at an E:T ratio of 50:1. At the same time, a neutralizing anti-trail or isotype control antibody (10 μg/ml) was added to cells. Supernatants were harvested 48 hours later, and WNV production was measured by plaque assay. Data in panels C and D was pooled from six replicate samples generated from independent experiments. Figure 9. FasL and granzyme B expression on CD8 + T cells from the brains of WNV-infected wild type and TRAIL -/- mice. Brain leukocytes from WNV-infected wild type mice were harvested at day 8 and stained for surface expression of FasL and intracellular expression of granzyme B. Data were analyzed by flow cytometry, and all results are pooled from two independent experiments. A. FasL expression on total CD4 + and CD8 + T cells and on NS4B tetramer positive CD8 + T cells in the brain. Note, 100% of brain T cells expressed FasL on their surface. Data is expressed as the normalized mean fluorescence intensity of the FasL positive cells. B-C. Granzyme B expression on total CD4 + and CD8 + T cells and on NS4B tetramer positive CD8 + T cells in the brain. Data is expressed as the percentage of granzyme B positive cells (B) and the normalized 31
32 mean fluorescence intensity of the granzyme B positive cells (C). Data are pooled from nine mice, and none of the differences were statistically significant Downloaded from on October 1, 2018 by guest 32
33 REFERENCES 1. Beraza, N., Y. Malato, L. E. Sander, M. Al-Masaoudi, J. Freimuth, D. Riethmacher, G. J. Gores, T. Roskams, C. Liedtke, and C. Trautwein Hepatocyte-specific NEMO deletion promotes NK/NKT cell- and TRAILdependent liver damage. J Exp Med 206: Brincks, E. L., P. Gurung, R. A. Langlois, E. A. Hemann, K. L. Legge, and T. S. Griffith The magnitude of the T cell response to a clinically significant dose of influenza virus is regulated by TRAIL. J Immunol 187: Brincks, E. L., A. Katewa, T. A. Kucaba, T. S. Griffith, and K. L. Legge CD8 T cells utilize TRAIL to control influenza virus infection. J Immunol 181: Busch, M. P., D. J. Wright, B. Custer, L. H. Tobler, S. L. Stramer, S. H. Kleinman, H. E. Prince, C. Bianco, G. Foster, L. R. Petersen, G. Nemo, and S. A. Glynn West Nile virus infections projected from blood donor screening data, United States, Emerg Infect Dis 12: Daffis, S., M. A. Samuel, B. C. Keller, M. Gale, Jr., and M. S. Diamond Cell-specific IRF-3 responses protect against West Nile virus infection by interferon-dependent and independent mechanisms. PLoS Pathog 3:e Daffis, S., M. A. Samuel, M. S. Suthar, B. C. Keller, M. Gale, Jr., and M. S. Diamond Interferon regulatory factor IRF-7 induces the antiviral alpha interferon response and protects against lethal West Nile virus infection. J Virol 82: Diamond, M. S., E. Mehlhop, T. Oliphant, and M. A. Samuel The host immunologic response to West Nile encephalitis virus. Front Biosci 14: Diamond, M. S., B. Shrestha, A. Marri, D. Mahan, and M. Engle B cells and antibody play critical roles in the immediate defense of disseminated infection by West Nile encephalitis virus. J Virol 77: Diehl, G. E., H. H. Yue, K. Hsieh, A. A. Kuang, M. Ho, L. A. Morici, L. L. Lenz, D. Cado, L. W. Riley, and A. Winoto TRAIL-R as a negative regulator of innate immune cell responses. Immunity 21:
34 Ebel, G. D., J. Carricaburu, D. Young, K. A. Bernard, and L. D. Kramer Genetic and phenotypic variation of West Nile virus in New York, Am J Trop Med Hyg 71: Glass, W. G., D. H. McDermott, J. K. Lim, S. Lekhong, S. F. Yu, W. A. Frank, J. Pape, R. C. Cheshier, and P. M. Murphy CCR5 deficiency increases risk of symptomatic West Nile virus infection. J Exp Med 203: Hoffmann, O., J. Priller, T. Prozorovski, U. Schulze-Topphoff, N. Baeva, J. D. Lunemann, O. Aktas, C. Mahrhofer, S. Stricker, F. Zipp, and J. R. Weber TRAIL limits excessive host immune responses in bacterial meningitis. J Clin Invest 117: Hu, W. H., H. Johnson, and H. B. Shu Tumor necrosis factor-related apoptosis-inducing ligand receptors signal NF-kappaB and JNK activation and apoptosis through distinct pathways. J Biol Chem 274: Indraccolo, S., U. Pfeffer, S. Minuzzo, G. Esposito, V. Roni, S. Mandruzzato, N. Ferrari, L. Anfosso, R. Dell'Eva, D. M. Noonan, L. Chieco-Bianchi, A. Albini, and A. Amadori Identification of genes selectively regulated by IFNs in endothelial cells. J Immunol 178: Ishikawa, E., M. Nakazawa, M. Yoshinari, and M. Minami Role of tumor necrosis factor-related apoptosis-inducing ligand in immune response to influenza virus infection in mice. J Virol 79: Kotelkin, A., E. A. Prikhod'ko, J. I. Cohen, P. L. Collins, and A. Bukreyev Respiratory syncytial virus infection sensitizes cells to apoptosis mediated by tumor necrosis factor-related apoptosis-inducing ligand. J Virol 77: Lanteri, M. C., K. M. O'Brien, W. E. Purtha, M. J. Cameron, J. M. Lund, R. E. Owen, J. W. Heitman, B. Custer, D. F. Hirschkorn, L. H. Tobler, N. Kiely, H. E. Prince, L. C. Ndhlovu, D. F. Nixon, H. T. Kamel, D. J. Kelvin, M. P. Busch, A. Y. Rudensky, M. S. Diamond, and P. J. Norris Tregs control the development of symptomatic West Nile virus infection in humans and mice. J Clin Invest 119: Lim, J. K., C. Y. Louie, C. Glaser, C. Jean, B. Johnson, H. Johnson, D. H. McDermott, and P. M. Murphy Genetic deficiency of chemokine 34
35 receptor CCR5 is a strong risk factor for symptomatic West Nile virus infection: a meta-analysis of 4 cohorts in the US epidemic. J Infect Dis 197: Lim, J. K., D. H. McDermott, A. Lisco, G. A. Foster, D. Krysztof, D. Follmann, S. L. Stramer, and P. M. Murphy CCR5 deficiency is a risk factor for early clinical manifestations of West Nile virus infection but not for viral transmission. J Infect Dis 201: Lin, Y., A. Devin, A. Cook, M. M. Keane, M. Kelliher, S. Lipkowitz, and Z. G. Liu The death domain kinase RIP is essential for TRAIL (Apo2L)- induced activation of IkappaB kinase and c-jun N-terminal kinase. Mol Cell Biol 20: Mehlhop, E., and M. S. Diamond Protective immune responses against West Nile virus are primed by distinct complement activation pathways. J Exp Med 203: Merino, D., N. Lalaoui, A. Morizot, P. Schneider, E. Solary, and O. Micheau Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2. Mol Cell Biol 26: Micheau, O Cellular FLICE-inhibitory protein: an attractive therapeutic target? Expert Opin Ther Targets 7: Murray, K., S. Baraniuk, M. Resnick, R. Arafat, C. Kilborn, K. Cain, R. Shallenberger, T. L. York, D. Martinez, J. S. Hellums, D. Hellums, M. Malkoff, N. Elgawley, W. McNeely, S. A. Khuwaja, and R. B. Tesh Risk factors for encephalitis and death from West Nile virus infection. Epidemiol Infect 134: Petersen, L. R., A. A. Marfin, and D. J. Gubler West Nile virus. Jama 290: Rigaud, S., M. C. Fondaneche, N. Lambert, B. Pasquier, V. Mateo, P. Soulas, L. Galicier, F. Le Deist, F. Rieux-Laucat, P. Revy, A. Fischer, G. de Saint Basile, and S. Latour XIAP deficiency in humans causes an X-linked lymphoproliferative syndrome. Nature 444:
36 Samuel, M. A., and M. S. Diamond Pathogenesis of West Nile virus infection: A balance between virulence, innate and adaptive immunity, and viral evasion. J Virol 80: Samuel, M. A., and M. S. Diamond Type I IFN protects against lethal West Nile Virus infection by restricting cellular tropism and enhancing neuronal survival. J Virol 79: Sato, K., S. Hida, H. Takayanagi, T. Yokochi, N. Kayagaki, K. Takeda, H. Yagita, K. Okumura, N. Tanaka, T. Taniguchi, and K. Ogasawara Antiviral response by natural killer cells through TRAIL gene induction by IFNalpha/beta. Eur J Immunol 31: Secchiero, P., F. Corallini, M. G. di Iasio, A. Gonelli, E. Barbarotto, and G. Zauli TRAIL counteracts the proadhesive activity of inflammatory cytokines in endothelial cells by down-modulating CCL8 and CXCL10 chemokine expression and release. Blood 105: Sedger, L. M., D. M. Shows, R. A. Blanton, J. J. Peschon, R. G. Goodwin, D. Cosman, and S. R. Wiley IFN-gamma mediates a novel antiviral activity through dynamic modulation of TRAIL and TRAIL receptor expression. J Immunol 163: Sejvar, J. J., A. V. Bode, A. A. Marfin, G. L. Campbell, J. Pape, B. J. Biggerstaff, and L. R. Petersen West Nile Virus-associated flaccid paralysis outcome. Emerg Infect Dis 12: Shepard, B. D., D. De Forni, D. R. McNamara, A. Foli, S. A. Rizza, R. S. Abraham, K. Knutson, P. J. Wettstein, F. Lori, and A. D. Badley Beneficial effect of TRAIL on HIV burden, without detectable immune consequences. PLoS One 3:e Shrestha, B., and M. S. Diamond Fas Ligand interactions contribute to CD8+ T cell-mediated control of West Nile virus infection in the central nervous system. J Virol 81: Shrestha, B., and M. S. Diamond The role of CD8+ T cells in the control of West Nile virus infection. J Virol 78:
37 Shrestha, B., T. Ng, H. J. Chu, M. Noll, and M. S. Diamond The relative contribution of antibody and CD8+ T cells to vaccine immunity against West Nile encephalitis virus. Vaccine 26: Shrestha, B., M. A. Samuel, and M. S. Diamond CD8+ T cells require perforin to clear West Nile virus from infected neurons. J Virol 80: Shrestha, B., B. Zhang, W. E. Purtha, R. S. Klein, and M. S. Diamond Tumor necrosis factor alpha protects against lethal West Nile virus infection by promoting trafficking of mononuclear leukocytes into the central nervous system. J Virol 82: Wang, Y., M. Lobigs, E. Lee, and A. Mullbacher CD8+ T cells mediate recovery and immunopathology in West Nile virus encephalitis. J Virol 77: Warke, R. V., K. J. Martin, K. Giaya, S. K. Shaw, A. L. Rothman, and I. Bosch TRAIL is a novel antiviral protein against dengue virus. J Virol 82: Zamai, L., M. Ahmad, I. M. Bennett, L. Azzoni, E. S. Alnemri, and B. Perussia Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp Med 188:
38
39
40
41
42
43
44
45
46
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Interferon regulatory factor-1 (IRF-1) shapes both innate and CD8+ T cell immune responses against West Nile virus infection
 shapes both innate and CD8+ T cell immune responses against West Nile virus infection") Washington University School of Medicine Digital Commons@Becker Open Access Publications 2011 Interferon regulatory factor-1 (IRF-1) shapes both innate and CD8+ T cell immune responses against West Nile
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2011 Interferon regulatory factor-1 (IRF-1) shapes both innate and CD8+ T cell immune responses against West Nile
A temporal role Of Type I interferon signaling in CD8+ T Cell maturation during acute West Nile virus infection
 Washington University School of Medicine Digital Commons@Becker Open Access Publications 2011 A temporal role Of Type I interferon signaling in CD8+ T Cell maturation during acute West Nile virus infection
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2011 A temporal role Of Type I interferon signaling in CD8+ T Cell maturation during acute West Nile virus infection
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
The innate immune adaptor molecule MyD88 restricts West Nile replication and
 JVI Accepts, published online ahead of print on 29 September 2010 J. Virol. doi:10.1128/jvi.01026-10 Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
JVI Accepts, published online ahead of print on 29 September 2010 J. Virol. doi:10.1128/jvi.01026-10 Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
NOTES. The Naturally Attenuated Kunjin Strain of West Nile Virus Shows Enhanced Sensitivity to the Host Type I Interferon Response
 JOURNAL OF VIROLOGY, June 2011, p. 5664 5668 Vol. 85, No. 11 0022-538X/11/$12.00 doi:10.1128/jvi.00232-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. NOTES The Naturally Attenuated
JOURNAL OF VIROLOGY, June 2011, p. 5664 5668 Vol. 85, No. 11 0022-538X/11/$12.00 doi:10.1128/jvi.00232-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. NOTES The Naturally Attenuated
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
NK cell flow cytometric assay In vivo DC viability and migration assay
 NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary Materials for
 immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
Follow this and additional works at: Part of the Medicine and Health Sciences Commons
 Washington University School of Medicine Digital Commons@Becker Open Access Publications 2009 Induction of IFN-beta and the innate antiviral response in myeloid cells occurs through an IPS-1-dependent
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2009 Induction of IFN-beta and the innate antiviral response in myeloid cells occurs through an IPS-1-dependent
Absence of mouse 2B4 promotes NK cell mediated killing of activated CD8 + T cells, leading to prolonged viral persistence and altered pathogenesis
 Research article Absence of mouse 2B4 promotes NK cell mediated killing of activated CD8 + T cells, leading to prolonged viral persistence and altered pathogenesis Stephen N. Waggoner, 1 Ruth T. Taniguchi,
Research article Absence of mouse 2B4 promotes NK cell mediated killing of activated CD8 + T cells, leading to prolonged viral persistence and altered pathogenesis Stephen N. Waggoner, 1 Ruth T. Taniguchi,
Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific
 SUPPLEMENTARY FIGURE LEGEND Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific CD4 + T cells correlates with intracellular CTLA-4 levels. (a) Comparative CTLA-4 levels
SUPPLEMENTARY FIGURE LEGEND Supplementary Figure 1. Enhanced detection of CTLA-4 on the surface of HIV-specific CD4 + T cells correlates with intracellular CTLA-4 levels. (a) Comparative CTLA-4 levels
D CD8 T cell number (x10 6 )
") IFNγ Supplemental Figure 1. CD T cell number (x1 6 ) 18 15 1 9 6 3 CD CD T cells CD6L C CD5 CD T cells CD6L D CD8 T cell number (x1 6 ) 1 8 6 E CD CD8 T cells CD6L F Log(1)CFU/g Feces 1 8 6 p
IFNγ Supplemental Figure 1. CD T cell number (x1 6 ) 18 15 1 9 6 3 CD CD T cells CD6L C CD5 CD T cells CD6L D CD8 T cell number (x1 6 ) 1 8 6 E CD CD8 T cells CD6L F Log(1)CFU/g Feces 1 8 6 p
Human Immunodeficiency Virus Type-1 Myeloid Derived Suppressor Cells Inhibit Cytomegalovirus Inflammation through Interleukin-27 and B7-H4
 Human Immunodeficiency Virus Type-1 Myeloid Derived Suppressor Cells Inhibit Cytomegalovirus Inflammation through Interleukin-27 and B7-H4 Ankita Garg, Rodney Trout and Stephen A. Spector,,* Department
Human Immunodeficiency Virus Type-1 Myeloid Derived Suppressor Cells Inhibit Cytomegalovirus Inflammation through Interleukin-27 and B7-H4 Ankita Garg, Rodney Trout and Stephen A. Spector,,* Department
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
Detailed step-by-step operating procedures for NK cell and CTL degranulation assays
 Supplemental methods Detailed step-by-step operating procedures for NK cell and CTL degranulation assays Materials PBMC isolated from patients, relatives and healthy donors as control K562 cells (ATCC,
Supplemental methods Detailed step-by-step operating procedures for NK cell and CTL degranulation assays Materials PBMC isolated from patients, relatives and healthy donors as control K562 cells (ATCC,
Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).
 Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).") Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
IMMUNOLOGICAL MEMORY. CD4 T Follicular Helper Cells. Memory CD8 T Cell Differentiation
 IMMUNOLOGICAL MEMORY CD4 T Follicular Helper Cells Memory CD8 T Cell Differentiation CD4 T Cell Differentiation Bcl-6 T-bet GATA-3 ROR t Foxp3 CD4 T follicular helper (Tfh) cells FUNCTION Provide essential
IMMUNOLOGICAL MEMORY CD4 T Follicular Helper Cells Memory CD8 T Cell Differentiation CD4 T Cell Differentiation Bcl-6 T-bet GATA-3 ROR t Foxp3 CD4 T follicular helper (Tfh) cells FUNCTION Provide essential
IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia
 Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
Supplemental Information. T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism
 Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to
 conjugated to") Class II tetramer staining Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to PE were combined with dominant HIV epitopes (DRB1*0101-DRFYKTLRAEQASQEV, DRB1*0301- PEKEVLVWKFDSRLAFHH,
Class II tetramer staining Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to PE were combined with dominant HIV epitopes (DRB1*0101-DRFYKTLRAEQASQEV, DRB1*0301- PEKEVLVWKFDSRLAFHH,
Critical Role for Alpha/Beta and Gamma Interferons in Persistence of Lymphocytic Choriomeningitis Virus by Clonal Exhaustion of Cytotoxic T Cells
 JOURNAL OF VIROLOGY, Sept. 2001, p. 8407 8423 Vol. 75, No. 18 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.18.8407 8423.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Critical
JOURNAL OF VIROLOGY, Sept. 2001, p. 8407 8423 Vol. 75, No. 18 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.18.8407 8423.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Critical
Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection
 Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
Supplemental Materials for. Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to. FTY720 during neuroinflammation
 Supplemental Materials for Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to FTY7 during neuroinflammation This file includes: Supplemental Table 1. EAE clinical parameters of
Supplemental Materials for Effects of sphingosine-1-phosphate receptor 1 phosphorylation in response to FTY7 during neuroinflammation This file includes: Supplemental Table 1. EAE clinical parameters of
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured
 Naïve CD4 + T cells were cultured") Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Micro 204. Cytotoxic T Lymphocytes (CTL) Lewis Lanier
 Lewis Lanier") Micro 204 Cytotoxic T Lymphocytes (CTL) Lewis Lanier Lewis.Lanier@ucsf.edu Lymphocyte-mediated Cytotoxicity CD8 + αβ-tcr + T cells CD4 + αβ-tcr + T cells γδ-tcr + T cells Natural Killer cells CD8 + αβ-tcr
Micro 204 Cytotoxic T Lymphocytes (CTL) Lewis Lanier Lewis.Lanier@ucsf.edu Lymphocyte-mediated Cytotoxicity CD8 + αβ-tcr + T cells CD4 + αβ-tcr + T cells γδ-tcr + T cells Natural Killer cells CD8 + αβ-tcr
Supporting Information
 Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature775 4 O.D. (595-655) 3 1 -ζ no antibody isotype ctrl Plated Soluble 1F6 397 7H11 Supplementary Figure 1 Soluble and plated anti- Abs induce -! signalling. B3Z cells stably expressing!
doi: 1.138/nature775 4 O.D. (595-655) 3 1 -ζ no antibody isotype ctrl Plated Soluble 1F6 397 7H11 Supplementary Figure 1 Soluble and plated anti- Abs induce -! signalling. B3Z cells stably expressing!
Interferon γ regulates idiopathic pneumonia syndrome, a. Th17 + CD4 + T-cell-mediated GvH disease
 Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Mukesh Kumar 1,2, Kelsey Roe 1,2, Maile O Connell 1,2 and Vivek R. Nerurkar 1,2* JOURNAL OF NEUROINFLAMMATION
 Kumar et al. Journal of Neuroinflammation (215) 12:178 DOI 1.1186/s12974-15-4-y JOURNAL OF NEUROINFLAMMATION RESEARCH Open Access Induction of virus-specific effector immune cell response limits virus
Kumar et al. Journal of Neuroinflammation (215) 12:178 DOI 1.1186/s12974-15-4-y JOURNAL OF NEUROINFLAMMATION RESEARCH Open Access Induction of virus-specific effector immune cell response limits virus
Resolution of a chronic viral infection after interleukin-10 receptor blockade
 ARTICLE Resolution of a chronic viral infection after interleukin-10 receptor blockade Mette Ejrnaes, 1 Christophe M. Filippi, 1 Marianne M. Martinic, 1 Eleanor M. Ling, 1 Lisa M. Togher, 1 Shane Crotty,
ARTICLE Resolution of a chronic viral infection after interleukin-10 receptor blockade Mette Ejrnaes, 1 Christophe M. Filippi, 1 Marianne M. Martinic, 1 Eleanor M. Ling, 1 Lisa M. Togher, 1 Shane Crotty,
Chapter 10 (pages ): Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Shenghua Zhou, Rong Ou, Lei Huang, and Demetrius Moskophidis*
 JOURNAL OF VIROLOGY, Jan. 2002, p. 829 840 Vol. 76, No. 2 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.2.829 840.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Critical Role
JOURNAL OF VIROLOGY, Jan. 2002, p. 829 840 Vol. 76, No. 2 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.2.829 840.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Critical Role
Supplementary Figure 1. Efficient DC depletion in CD11c.DOG transgenic mice
 Supplementary Figure 1. Efficient DC depletion in CD11c.DOG transgenic mice (a) CD11c.DOG transgenic mice (tg) were treated with 8 ng/g body weight (b.w.) diphtheria toxin (DT) i.p. on day -1 and every
Supplementary Figure 1. Efficient DC depletion in CD11c.DOG transgenic mice (a) CD11c.DOG transgenic mice (tg) were treated with 8 ng/g body weight (b.w.) diphtheria toxin (DT) i.p. on day -1 and every
SUPPLEMENTARY INFORMATION
 Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
Effector Mechanisms of Cell-Mediated Immunity
 Effector Mechanisms of Cell-Mediated Immunity Dr. Julia Rempel Section of Hepatology 789-3825 jdrempel@cc.umanitoba.ca 804D JBRC Topics: I. Types of Cell-Mediated Immunity II. Migration of Effector T Lymphocytes
Effector Mechanisms of Cell-Mediated Immunity Dr. Julia Rempel Section of Hepatology 789-3825 jdrempel@cc.umanitoba.ca 804D JBRC Topics: I. Types of Cell-Mediated Immunity II. Migration of Effector T Lymphocytes
Production of Interferon Alpha by Dengue Virus-infected Human Monocytes
 J. gen. Virol. (1988), 69, 445-449. Printed in Great Britain 445 Key words: IFN-ct/dengue virus/monocytes Production of Interferon Alpha by Dengue Virus-infected Human Monocytes By ICHIRO KURANE AND FRANCIS
J. gen. Virol. (1988), 69, 445-449. Printed in Great Britain 445 Key words: IFN-ct/dengue virus/monocytes Production of Interferon Alpha by Dengue Virus-infected Human Monocytes By ICHIRO KURANE AND FRANCIS
Lineage relationship and protective immunity of memory CD8 T cell subsets
 23 Nature Publishing Group http://www.nature.com/natureimmunology Lineage relationship and protective immunity of memory CD8 T cell subsets E. John Wherry 1 *,Volker Teichgräber 1 *,Todd C. Becker 1,David
23 Nature Publishing Group http://www.nature.com/natureimmunology Lineage relationship and protective immunity of memory CD8 T cell subsets E. John Wherry 1 *,Volker Teichgräber 1 *,Todd C. Becker 1,David
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
Direct ex vivo characterization of human antigen-specific CD154 + CD4 + T cells Rapid antigen-reactive T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central
General Overview of Immunology. Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center
 General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
Supplemental Information. CD4 + CD25 + Foxp3 + Regulatory T Cells Promote. Th17 Cells In Vitro and Enhance Host Resistance
 Immunity, Volume 34 Supplemental Information D4 + D25 + + Regulatory T ells Promote Th17 ells In Vitro and Enhance Host Resistance in Mouse andida albicans Th17 ell Infection Model Pushpa Pandiyan, Heather
Immunity, Volume 34 Supplemental Information D4 + D25 + + Regulatory T ells Promote Th17 ells In Vitro and Enhance Host Resistance in Mouse andida albicans Th17 ell Infection Model Pushpa Pandiyan, Heather
Supplementary Information. Tissue-wide immunity against Leishmania. through collective production of nitric oxide
 Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity
 Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Increased IL-12 induced STAT-4 signaling in CD8 T cells. from aged mice
 Increased IL-2 induced STAT-4 signaling in CD8 T cells from aged mice Erin Rottinghaus * Abstract: Aging is associated with poor immune function leading to increased susceptibility to infectious diseases
Increased IL-2 induced STAT-4 signaling in CD8 T cells from aged mice Erin Rottinghaus * Abstract: Aging is associated with poor immune function leading to increased susceptibility to infectious diseases
CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads (Miltenyi Biotec) for depletion of CD25 +
 and labeled with anti-pe-microbeads (Miltenyi Biotec) for depletion of CD25 +") Supplements Supplemental Materials and Methods Depletion of CD25 + T-cells from PBMC. Fresh or HD precultured PBMC were stained with the conjugate CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads
Supplements Supplemental Materials and Methods Depletion of CD25 + T-cells from PBMC. Fresh or HD precultured PBMC were stained with the conjugate CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Cover Page. The handle holds various files of this Leiden University dissertation.
 Cover Page The handle http://hdl.handle.net/1887/23854 holds various files of this Leiden University dissertation. Author: Marel, Sander van der Title: Gene and cell therapy based treatment strategies
Cover Page The handle http://hdl.handle.net/1887/23854 holds various files of this Leiden University dissertation. Author: Marel, Sander van der Title: Gene and cell therapy based treatment strategies
Rapid antigen-specific T cell enrichment (Rapid ARTE)
") Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
Direct ex vivo characterization of human antigen-specific CD154+CD4+ T cell Rapid antigen-specific T cell enrichment (Rapid ARTE) Introduction Workflow Antigen (ag)-specific T cells play a central role
Control of Virus-Specific CD8 T-Cell Exhaustion and Immune-Mediated Pathology by E3 Ubiquitin Ligase Cbl-b during Chronic Viral Infection
 JOURNAL OF VIROLOGY, Apr. 2008, p. 3353 3368 Vol. 82, No. 7 0022-538X/08/$08.00 0 doi:10.1128/jvi.01350-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Control of Virus-Specific
JOURNAL OF VIROLOGY, Apr. 2008, p. 3353 3368 Vol. 82, No. 7 0022-538X/08/$08.00 0 doi:10.1128/jvi.01350-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Control of Virus-Specific
Dual Targeting Nanoparticle Stimulates the Immune
 Dual Targeting Nanoparticle Stimulates the Immune System to Inhibit Tumor Growth Alyssa K. Kosmides, John-William Sidhom, Andrew Fraser, Catherine A. Bessell, Jonathan P. Schneck * Supplemental Figure
Dual Targeting Nanoparticle Stimulates the Immune System to Inhibit Tumor Growth Alyssa K. Kosmides, John-William Sidhom, Andrew Fraser, Catherine A. Bessell, Jonathan P. Schneck * Supplemental Figure
Viral Persistence Alters CD8 T-Cell Immunodominance and Tissue Distribution and Results in Distinct Stages of Functional Impairment
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4911 4927 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4911 4927.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Viral Persistence
JOURNAL OF VIROLOGY, Apr. 2003, p. 4911 4927 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4911 4927.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Viral Persistence
Bead Based Assays for Cytokine Detection
 Bead Based Assays for Cytokine Detection September 27, 2014 6 th EFIS-EJI South East European Immunology School SEEIS 2014 Timisoara, Romania The Cells of the Immune System The Immune Reaction (Th2) (Th1)
Bead Based Assays for Cytokine Detection September 27, 2014 6 th EFIS-EJI South East European Immunology School SEEIS 2014 Timisoara, Romania The Cells of the Immune System The Immune Reaction (Th2) (Th1)
Immunology - Lecture 2 Adaptive Immune System 1
 Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
for six pairs of mice. (b) Representative FACS analysis of absolute number of T cells (CD4 + and
 Representative FACS analysis of absolute number of T cells (CD4 + and") SUPPLEMENTARY DATA Supplementary Figure 1: Peripheral lymphoid organs of SMAR1 -/- mice have an effector memory phenotype. (a) Lymphocytes collected from MLNs and Peyer s patches (PPs) of WT and SMAR1
SUPPLEMENTARY DATA Supplementary Figure 1: Peripheral lymphoid organs of SMAR1 -/- mice have an effector memory phenotype. (a) Lymphocytes collected from MLNs and Peyer s patches (PPs) of WT and SMAR1
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Materials for
 www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
Cell-mediated Immunity
 Cellular & Molecular Immunology Cell-mediated Immunity Nicholas M. Ponzio, Ph.D. Department of Pathology & Laboratory Medicine April 6, 2009 Today s Presentation: Overview Cellular Interactions In Humoral
Cellular & Molecular Immunology Cell-mediated Immunity Nicholas M. Ponzio, Ph.D. Department of Pathology & Laboratory Medicine April 6, 2009 Today s Presentation: Overview Cellular Interactions In Humoral
Immunity to Viruses. Patricia Fitzgerald-Bocarsly September 25, 2008
 Immunity to Viruses Patricia Fitzgerald-Bocarsly September 25, 2008 The Immune System Deals with a Huge Range of Pathogens Roitt, 2003 Immune Responses to Viruses Viruses are dependent on the host cell
Immunity to Viruses Patricia Fitzgerald-Bocarsly September 25, 2008 The Immune System Deals with a Huge Range of Pathogens Roitt, 2003 Immune Responses to Viruses Viruses are dependent on the host cell
Supplemental Information. Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection. Cell Host & Microbe, Volume 15
 Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
Canberra, Australia). CD11c-DTR-OVA-GFP (B6.CD11c-OVA), B6.luc + and. Cancer Research Center, Germany). B6 or BALB/c.FoxP3-DTR-GFP mice were
. CD11c-DTR-OVA-GFP (B6.CD11c-OVA), B6.luc + and. Cancer Research Center, Germany). B6 or BALB/c.FoxP3-DTR-GFP mice were") Supplemental Materials and Methods Mice Female C57BL/6 (B6, I-E null, H-2 b ), BALB/c (H-2 d ) + ), FVB/N (H-2 q, I-E null, CD45.1 + ), and B6D2F1 (H-2 b/d ) mice were purchased from the Animal Resources
Supplemental Materials and Methods Mice Female C57BL/6 (B6, I-E null, H-2 b ), BALB/c (H-2 d ) + ), FVB/N (H-2 q, I-E null, CD45.1 + ), and B6D2F1 (H-2 b/d ) mice were purchased from the Animal Resources
The Role of CD4 T Cells in the Pathogenesis of Murine AIDS
 JOURNAL OF VIROLOGY, June 2006, p. 5777 5789 Vol. 80, No. 12 0022-538X/06/$08.00 0 doi:10.1128/jvi.02711-05 Copyright 2006, American Society for Microbiology. All Rights Reserved. The Role of CD4 T Cells
JOURNAL OF VIROLOGY, June 2006, p. 5777 5789 Vol. 80, No. 12 0022-538X/06/$08.00 0 doi:10.1128/jvi.02711-05 Copyright 2006, American Society for Microbiology. All Rights Reserved. The Role of CD4 T Cells
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human
 with 5% human") Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
Supplementary Figure 1.
 Supplementary Figure 1. Female Pro-ins2 -/- mice at 5-6 weeks of age were either inoculated i.p. with a single dose of CVB4 (1x10 5 PFU/mouse) or PBS and treated with αgalcer or control vehicle. On day
Supplementary Figure 1. Female Pro-ins2 -/- mice at 5-6 weeks of age were either inoculated i.p. with a single dose of CVB4 (1x10 5 PFU/mouse) or PBS and treated with αgalcer or control vehicle. On day
Pearson r = P (one-tailed) = n = 9
 = n = 9") 8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells
 The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells Jeremy Herskowitz, Emory University Meagan A. Jacoby, Emory University Samuel Speck, Emory University
The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells Jeremy Herskowitz, Emory University Meagan A. Jacoby, Emory University Samuel Speck, Emory University
Supporting Information
 Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Cell Mediated Immunity CELL MEDIATED IMMUNITY. Basic Elements of Cell Mediated Immunity (CMI) Antibody-dependent cell-mediated cytotoxicity (ADCC)
 Antibody-dependent cell-mediated cytotoxicity (ADCC)") Chapter 16 CELL MEDIATED IMMUNITY Cell Mediated Immunity Also known as Cellular Immunity or CMI The effector phase T cells Specificity for immune recognition reactions TH provide cytokines CTLs do the
Chapter 16 CELL MEDIATED IMMUNITY Cell Mediated Immunity Also known as Cellular Immunity or CMI The effector phase T cells Specificity for immune recognition reactions TH provide cytokines CTLs do the
and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the
 Diagrammatic representation of the") Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Persistence of West Nile Virus in the Central Nervous System and Periphery of Mice
 Persistence of West Nile Virus in the Central Nervous System and Periphery of Mice Kim K. Appler 1, Ashley N. Brown 1,2 a, Barbara S. Stewart 1,2, Melissa J. Behr 1,2 b, Valerie L. Demarest 1, Susan J.
Persistence of West Nile Virus in the Central Nervous System and Periphery of Mice Kim K. Appler 1, Ashley N. Brown 1,2 a, Barbara S. Stewart 1,2, Melissa J. Behr 1,2 b, Valerie L. Demarest 1, Susan J.
Out-of-sequence signal 3 as a mechanism for virusinduced immune suppression of CD8 T cell responses
 University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 9-25-2014 Out-of-sequence signal 3 as a mechanism for virusinduced immune suppression
University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 9-25-2014 Out-of-sequence signal 3 as a mechanism for virusinduced immune suppression
Supplementalgfigureg1gSchematicgdiagramgofgtumor1modellingg
 SChinjectionh F:LuchLCLsh IVhinjectionh T:cellsh Monitorhforhtumorh growthhandhxeno: reactivehgvhd GVLgexperimentg kcbgvsgpbgt1cellse Xeno1reactiveg experimentg kcbgvsgpbgt1cellse IVhinjectionh 5xh,N^6
SChinjectionh F:LuchLCLsh IVhinjectionh T:cellsh Monitorhforhtumorh growthhandhxeno: reactivehgvhd GVLgexperimentg kcbgvsgpbgt1cellse Xeno1reactiveg experimentg kcbgvsgpbgt1cellse IVhinjectionh 5xh,N^6
Dengue vaccine induced CD8 + T cell immunity confers protection in the context of enhancing, interfering maternal antibodies
 Downloaded from http:// on February 14, 2018. Dengue vaccine induced CD8 + T cell immunity confers protection in the context of enhancing, interfering maternal antibodies Jian Hang Lam, 1,2 Yen Leong Chua,
Downloaded from http:// on February 14, 2018. Dengue vaccine induced CD8 + T cell immunity confers protection in the context of enhancing, interfering maternal antibodies Jian Hang Lam, 1,2 Yen Leong Chua,
NKG2D Receptor Signaling Enhances Cytolytic Activity by Virus-Specific CD8 T Cells: Evidence for a Protective Role in Virus-Induced Encephalitis
 JOURNAL OF VIROLOGY, Mar. 2008, p. 3031 3044 Vol. 82, No. 6 0022-538X/08/$08.00 0 doi:10.1128/jvi.02033-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. NKG2D Receptor Signaling
JOURNAL OF VIROLOGY, Mar. 2008, p. 3031 3044 Vol. 82, No. 6 0022-538X/08/$08.00 0 doi:10.1128/jvi.02033-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. NKG2D Receptor Signaling
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors Presented by Jurg Rohrer, PhD, BD Biosciences 23-10780-00 Outline Introduction Cytokines Transcription
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors Presented by Jurg Rohrer, PhD, BD Biosciences 23-10780-00 Outline Introduction Cytokines Transcription
Acid sphingomyelinase is a critical regulator of cytotoxic granule secretion by
 Supplementary Data Acid sphingomyelinase is a critical regulator of cytotoxic granule secretion by primary T lymphocytes Jasmin Herz, Julian Pardo, Hamid Kashkar, Michael Schramm, Elza Kuzmenkina, Erik
Supplementary Data Acid sphingomyelinase is a critical regulator of cytotoxic granule secretion by primary T lymphocytes Jasmin Herz, Julian Pardo, Hamid Kashkar, Michael Schramm, Elza Kuzmenkina, Erik
CD14 + S100A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer
 CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
Optimizing Intracellular Flow Cytometry
 Optimizing Intracellular Flow Cytometry Detection of Cytokines, Transcription Factors, and Phosphoprotein by Flow Cytometry Presented by Erika O Donnell, PhD, BD Biosciences 23-14876-00 Outline Basic principles
Optimizing Intracellular Flow Cytometry Detection of Cytokines, Transcription Factors, and Phosphoprotein by Flow Cytometry Presented by Erika O Donnell, PhD, BD Biosciences 23-14876-00 Outline Basic principles
Cell-specific IRF-3 responses protect against West Nile virus infection by interferon-dependent and - independent mechanisms
 Washington University School of Medicine Digital Commons@Becker Open Access Publications 2007 Cell-specific IRF-3 responses protect against West Nile virus infection by interferon-dependent and - independent
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2007 Cell-specific IRF-3 responses protect against West Nile virus infection by interferon-dependent and - independent
The role of IL-6 in the expression of PD-1 and PDL-1 on CNS cells following infection with Theiler s murine encephalomyelitis virus
 JVI Accepts, published online ahead of print on 21 August 2013 J. Virol. doi:10.1128/jvi.01967-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 The role of IL-6 in the expression
JVI Accepts, published online ahead of print on 21 August 2013 J. Virol. doi:10.1128/jvi.01967-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 The role of IL-6 in the expression
W/T Itgam -/- F4/80 CD115. F4/80 hi CD115 + F4/80 + CD115 +
 F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
Natural Killer Cells as Novel Helpers in Anti-Herpes Simplex Virus Immune Response
 JOURNAL OF VIROLOGY, Nov. 2008, p. 10820 10831 Vol. 82, No. 21 0022-538X/08/$08.00 0 doi:10.1128/jvi.00365-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Natural Killer Cells
JOURNAL OF VIROLOGY, Nov. 2008, p. 10820 10831 Vol. 82, No. 21 0022-538X/08/$08.00 0 doi:10.1128/jvi.00365-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Natural Killer Cells
staining and flow cytometry
 Detection of influenza virus-specific T cell responses by intracellular by cytokine intracellular staining cytokine staining and flow cytometry Detection of influenza virus-specific T cell responses and
Detection of influenza virus-specific T cell responses by intracellular by cytokine intracellular staining cytokine staining and flow cytometry Detection of influenza virus-specific T cell responses and
RAISON D ETRE OF THE IMMUNE SYSTEM:
 RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
Nature Immunology: doi: /ni Supplementary Figure 1. Production of cytokines and chemokines after vaginal HSV-2 infection.
 Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Supplementary Information:
 Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
SUPPORTING INFORMATIONS
 SUPPORTING INFORMATIONS Mice MT/ret RetCD3ε KO α-cd25 treated MT/ret Age 1 month 3 mnths 6 months 1 month 3 months 6 months 1 month 3 months 6 months 2/87 Survival 87/87 incidence of 17/87 1 ary tumor
SUPPORTING INFORMATIONS Mice MT/ret RetCD3ε KO α-cd25 treated MT/ret Age 1 month 3 mnths 6 months 1 month 3 months 6 months 1 month 3 months 6 months 2/87 Survival 87/87 incidence of 17/87 1 ary tumor
West Nile Virus. By Frank Riusech
 West Nile Virus By Frank Riusech Disease Etiology: West Nile virus(wnv), genus, flavivirus is positive- stranded RNA arbovirus (arthropod- borne), belonging to the Flaviviridae family. Included in this
West Nile Virus By Frank Riusech Disease Etiology: West Nile virus(wnv), genus, flavivirus is positive- stranded RNA arbovirus (arthropod- borne), belonging to the Flaviviridae family. Included in this
T Cell Activation, Costimulation and Regulation
 1 T Cell Activation, Costimulation and Regulation Abul K. Abbas, MD University of California San Francisco 2 Lecture outline T cell antigen recognition and activation Costimulation, the B7:CD28 family
1 T Cell Activation, Costimulation and Regulation Abul K. Abbas, MD University of California San Francisco 2 Lecture outline T cell antigen recognition and activation Costimulation, the B7:CD28 family