JBC Papers in Press. Published on February 1, 2017 as Manuscript M
|
|
|
- Rosamund Hunt
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on February 1, 2017 as Manuscript M The latest version is at ROS induced by sorafenib overcomes TRAIL resistance Bax/Bak-independent mitochondrial depolarization and reactive oxygen species induction by sorafenib overcome resistance to apoptosis in renal cell carcinoma Bernhard Gillissen 1,2, Anja Richter 1,2, Antje Richter 1, Robert Preissner 5, Klaus Schulze-Osthoff 2,4, Frank Essmann 2,4, and Peter T. Daniel 1,2,3 1 Department of Hematology, Oncology and Tumor Immunology, University Medical Center Charité, Campus Berlin-Buch, Humboldt University Berlin, Germany 2 German Cancer Consortium (DKTK) and German Cancer Research Center (DKFZ), Im Neuenheimer Feld, Heidelberg, Germany 3 Clinical and Molecular Oncology, Max Delbrück Center for Molecular Medicine, Berlin-Buch, Germany 4 Interfaculty Institute for Biochemistry, University of Tübingen, Hoppe-Seyler-Str. 4, Tübingen, Germany 5 Institute of Physiology & Experimental Clinical Research Center, University Medical Center Charité, Berlin, Germany Running title: ROS induced by sorafenib overcomes TRAIL resistance To whom correspondence should be addressed: Dr. Peter Daniel, Clinical and Molecular Oncology, University Medical Center Charité and Max Delbrück Center for Molecular Medicine, Lindenberger Weg 80, Berlin, Germany, phone: ; pdaniel@mdc-berlin.de Keywords: Sorafenib, TRAIL resistance, reactive oxygen species (ROS), apoptosis, B-cell lymphoma 2 (Bcl-2) family, Bax, Bak, cancer ABSTRACT Renal cell carcinoma (RCC) is polyresistant to chemo- and radiotherapy or biologicals including TNF-related apoptosis inducing ligand (TRAIL). Sorafenib, a multikinase inhibitor approved for the treatment of RCC, has been shown to sensitize cancer cells toward TRAIL-induced apoptosis, in particular by downregulation of the Bak-inhibitory Bcl-2 family protein Mcl-1. Here, we demonstrate that sorafenib overcomes TRAIL resistance in RCC by a mechanism that does not rely on Mcl-1 downregulation. Instead, sorafenib induces a rapid dissipation of the mitochondrial membrane potential (ΔΨ m ) that is accompanied by the accumulation of reactive oxygen species (ROS). Loss of ΔΨ m and ROS production induced by sorafenib are independent of caspase activities and do not depend on the presence of the pro-apoptotic Bcl-2 family proteins Bax or Bak indicating that both events are functionally up-stream of the mitochondrial apoptosis signaling cascade. More intriguingly, we find that it is sorafenib-induced ROS accumulation that enables TRAIL to activate caspase-8 in RCC. This leads to apoptosis that involves activation of an amplification loop via the mitochondrial apoptosis pathway. Thus, our mechanistic data indicate that sorafenib bypasses central resistance mechanisms through a direct induction of ΔΨ m breakdown and ROS production. Activation of this pathway might represent a useful strategy to overcome the cell-inherent resistance to cancer therapeutics including TRAIL in multiresistant cancers such as RCC. INTRODUCTION Apoptotic cell death induced by the death ligand TRAIL (tumor necrosis factor related apoptosis inducing ligand) plays an important role in immune surveillance and is a major immune 1 Copyright 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
2 defense mechanism against tumor cells. Consequently, the use of TRAIL, which preferentially kills cancer cells while sparing noncancerous tissue, is a promising concept in anticancer therapy. Recombinant TRAIL and agonistic antibodies against the TRAIL receptors are therefore tested in clinical phase I and II studies (1-3). Binding of TRAIL to the death receptors DR4 (TRAIL-R1) and DR5 (TRAIL-R2) initiates receptor oligomerization and, via the adaptor protein FADD (Fas-associated protein with death domain), recruitment of the initiator caspase-8 to the death domain of the activated receptor (4,5). Formation of this activation platform, the so-called death-inducing signaling complex (DISC), results in autocatalytic activation of caspase-8. In type I cells, active caspase-8 triggers, via direct proteolytic processing of caspase-3, a caspase cascade to induce apoptotic cell death. In type II cells, however, the E3-ligase XIAP (X-linked inhibitor of apoptosis protein) prevents accumulation of active caspase-3 by marking it for proteasomal degradation (6). Therefore, in type II cells efficient caspase-3 activation upon death receptor signaling requires amplification via the mitochondrial apoptosis pathway. Activation of the mitochondrial pathway is achieved by mitochondrial outer membrane permeabilization (MOMP) (7). Upon MOMP, the XIAP inhibitor Smac/DIABLO (second mitochondria-derived activator of caspase/direct IAP-binding protein with low pi) is released from the mitochondrial intermembrane space into the cytosol (8) where it inhibits XIAP and thereby prevents degradation of active caspase-3 (9-11). Another important proapoptotic factor that is released into the cytosol upon MOMP is cytochrome c. Cytochrome c activates the adapter molecule APAF-1, resulting in the formation of the apoptosome, a multiprotein complex in which the initiator caspase-9 is activated (12) for processing of caspase-3 and amplification of the caspase cascade. Upon TRAIL-R ligation, MOMP is induced by caspase-8-mediated cleavage and activation of Bid (BH3-interacting domain death agonist), a proapoptotic protein of the B cell lymphoma 2 (Bcl-2) family (13-15) The proteins of the Bcl-2 family are key regulators of MOMP and show homology in at least one of four Bcl-2 homology (BH1-4) domains. Anti-apoptotic family members (e.g. Bcl-2, Bcl-x L and Mcl-1) are characterized by the presence of all four BH domains. Pro-apoptotic members can be subdivided into the multidomain BH123 homologs (Bax, Bak and Bok), and into the large BH3-only subfamily (e.g. Bid, Bim, Bad, Nbk/Bik, Puma, Noxa) (16). The pro-apoptotic BH123 proteins Bax (Bcl-2 associated x protein) and Bak (Bcl-2 homologous antagonist/killer) drive MOMP and are neutralized by anti-apoptotic family members. BH3-only proteins activate Bax and Bak to induce MOMP indirectly by inhibiting pro-survival Bcl-2 proteins and/or via direct interaction with Bax and Bak (17,18). Deregulation of these apoptosis signaling pathways accounts for resistance to anticancer therapies including the biological TRAIL that often serves as a prototypical targeted reagent to study apoptosis signaling in cancer cells. Strategies to overcome resistance to TRAIL-induced apoptosis comprise combinations with DNAdamaging therapies including the use of chemotherapeutic drugs (19) and irradiation (20), or the inhibition of pro-survival signaling, e.g. the NF-κB (nuclear factor-κb) pathway (21), inhibition of the proteasome (22,23) or inhibition of histone deacetylases (24), all of which have been shown to sensitize tumor cells for TRAIL. In addition, BH3 mimetics, small molecules like ABT-737 or Obatoclax may potentiate TRAILmediated apoptosis through binding to the hydrophobic groove at the surface of anti-apoptotic Bcl-2 proteins thereby blocking their pro-survival function (25,26). Furthermore, the multikinase inhibitor sorafenib sensitizes cancer cells toward TRAIL through alternative mechanisms, e.g. inhibition of STAT3 (27,28), and in particular through downregulation of the Bak inhibitor Mcl-1 (Myeloid cell leukemia 1) (29,30). Downregulation of Mcl-1 enables TRAIL to kill cells via activation of Bak and thus can overcome TRAIL resistance of Bax-deficient cells (31). Sorafenib is approved for the treatment of advanced renal cell carcinomas (RCC) (32-35), a cancer entity that frequently shows resistance not only to conventional radio- and chemotherapy, but also to experimental therapy with TRAIL (22). Here, we show that sorafenib overcomes the TRAIL resistance of various RCC cell lines. Surprisingly, in RCC sorafenib-induced downregulation of Mcl-1 is not causative for the sensitization. Instead, sorafenib induces caspaseand Bax/Bak-independent depolarization of mitochondria accompanied by increased ROS 2
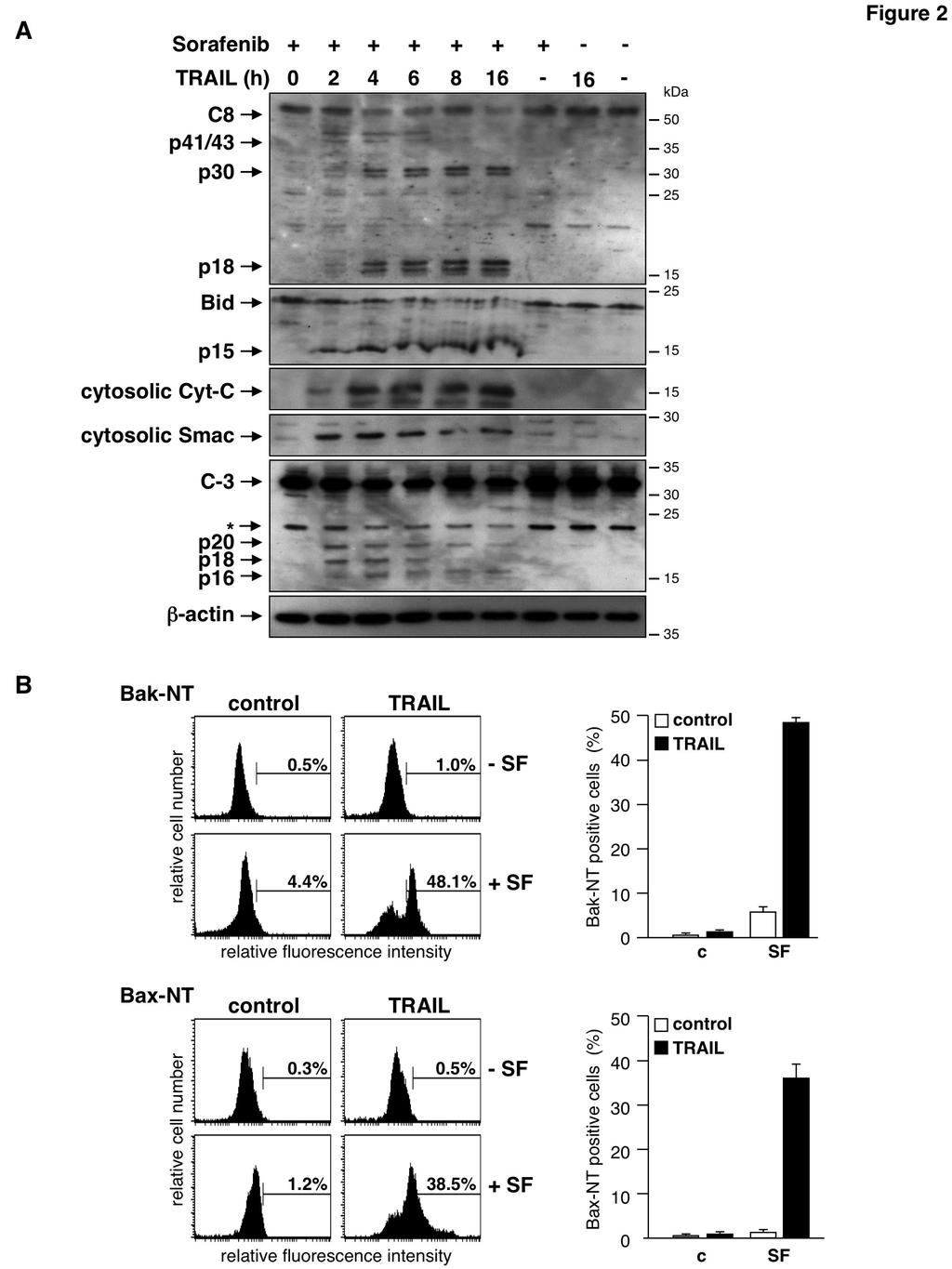
3 accumulation. Accumulation of ROS then overcomes the failure of TRAIL to activate caspase-8 in RCC cells and thereby enables TRAIL to induce apoptosis. RESULTS RCCs frequently display an impressive resistance to anti-cancer therapies including application of the biological TRAIL. We therefore employed TRAIL as a well-defined apoptosis inducer to evaluate strategies to overcome therapy resistance in RCC. To this end, we treated the three RCC cell lines RCC-KP, RCC-26, and RCC-GW, that had revealed high resistance toward TRAIL in dose-response experiments, with TRAIL (50 ng/ml for 24 h), sorafenib (20 µm for 38 h) or preincubated cells for 14 h with sorafenib prior to TRAIL treatment. Induction of apoptosis was analyzed by flow cytometric detection of the relative cellular DNA content and hypodiploid cells were assumed apoptotic. As expected, a TRAIL concentration of 50 ng/ml alone did not induce apoptotic DNA fragmentation in any of the three RCC cell lines (fig. 1). 20 µm sorafenib also did not induce apoptosis in RCC-GW and only marginally induced apoptosis in RCC-KP and RCC-26 cells. Pre-incubation of cells with sorafenib, however, strongly sensitized all cell lines for TRAIL-induced apoptosis. In detail, 45% of RCC-KP, 44% of RCC-26 and 26% of RCC- GW cells showed a hypodiploid, i.e. apoptotic phenotype upon the combined treatment, whereas the treatment with TRAIL or sorafenib as single agents resulted in less than 8% and 15% apoptotic cells, respectively. It has been shown recently that sorafenib sensitizes tumor cells for TRAIL by downregulation of XIAP and in particular by downregulation of Mcl-1. Thus, we analyzed the influence of sorafenib on the expression levels of XIAP and Mcl-1 in RCC lines by immunoblotting. The expression levels of XIAP did not differ in control and sorafenib-treated RCC cell lines (fig. 1B), indicating that XIAP downregulation is not involved in the reversal of apoptosis resistance. However, expression of Mcl-1 was slightly reduced in RCC-KP and RCC-26 cells (but not in RCC-GW cells), suggesting that sorafenib-induced Mcl-1 downregulation might cause TRAIL sensitization. To investigate the relevance of reduced Mcl-1 expression for the sensitization of RCC cell lines, we therefore analyzed TRAILinduced apoptosis in the presence or absence of sirna-mediated knockdown of Mcl-1. In contrast to other cell systems (31) knockdown of Mcl-1 (fig. 1C), surprisingly, did not increase TRAILinduced apoptosis in RCC as compared to controls (fig. 1D). These results indicate that sorafenibmediated downregulation of Mcl-1 is unlikely to cause sensitization of RCC cell lines toward TRAIL. To analyze the mechanism underlying the synergism of sorafenib and TRAIL in RCC, we investigated hallmarks of the mitochondrial apoptosis signaling cascade and death receptor signaling. Western blot analysis showed that the death ligand TRAIL as a single agent failed to induce caspase-8 processing (fig. 2A). Consistently, events located downstream of caspase-8 activation such as Bid cleavage, caspase-3 processing or the release of Smac and cytochrome c into the cytosol, were not detectable upon TRAIL treatment (fig. 2A). Likewise, sorafenib alone did not induce caspase activation or release of pro-apoptotic factors into the cytosol. In contrast, a time course experiment revealed that pre-incubation of cells with sorafenib enabled TRAIL to activate caspase-8, as evidenced by the detection of the caspase-8 subunits p41/43 and p18 after 2 to 4 h of TRAIL treatment. In addition, after 4 h a p30 subunit was detectable, indicating that caspase-8 was also processed via an alternative previously described pathway (36). Activation of caspase-8 was accompanied by processing of Bid to its active form tbid/p15, release of Smac and cytochrome c and processing of pro-caspase-3 to the active subunits (fig. 2A). In accordance with the cleavage of Bid and the release of pro-apoptotic factors into the cytosol, immunofluorescence staining of cells with antibodies specific for active conformers of Bax and Bak, respectively, verified activation of both molecules in response to the combined incubation of cells with TRAIL and sorafenib (fig. 2B). Thus, although TRAIL as a single agent was incapable of efficient caspase-8 activation, pre-treatment of RCC cells with sorafenib enabled TRAIL to activate caspase-8 resulting in Bid cleavage and activation of the mitochondrial apoptosis pathway. In order to delineate the activation of the mitochondrial apoptosis pathway by the combined treatment with sorafenib and TRAIL in more detail, we investigated loss of the mitochondrial membrane potential (ΔΨ m ). To this end, cells were 3
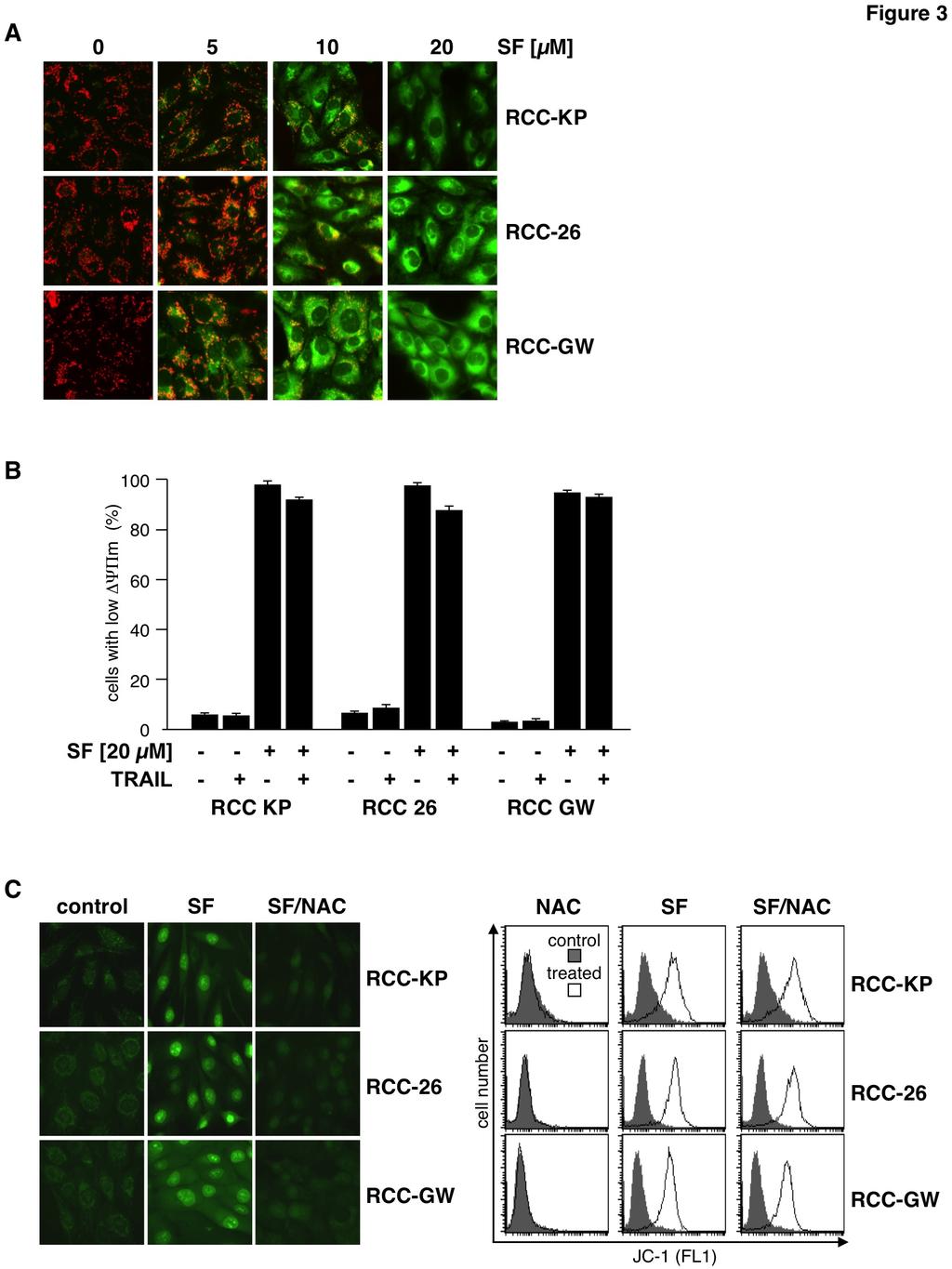
4 incubated with the cationic dye JC-1 that exhibits specific mitochondrial localization and red fluorescence in vital cells and cytosolic localization and green fluorescence upon loss of ΔΨ m. In control cells fluorescence microscopy showed a red dotted staining pattern, indicating accumulation of JC-1 in the mitochondria. In contrast, upon sorafenib treatment cells showed a dose-dependent increase of diffuse (cytosolic) green fluorescence (fig. 3A). FACS analysis of JC-1 green fluorescence intensity revealed loss of ΔΨ m in almost all cells upon treatment with sorafenib alone and, naturally, also upon combined treatment with sorafenib and TRAIL (fig. 3B). Although sorafenib by itself was sufficient to induce loss of ΔΨ m it failed, as a single agent, to induce apoptosis in RCC cells (fig. 2A). Mitochondrial dysfunction, such as loss of ΔΨ m, is commonly associated with production of reactive oxygen species (ROS). To analyze whether sorafenib-induced depolarization of the mitochondria is accompanied by increased intracellular ROS levels, cells were incubated with the cell-permeable dye CellRox Green that exhibits green fluorescence and binding to DNA only upon oxidation. Fluorescence microscopy of control cells showed a weak dotted green fluorescence pattern in all three cell lines, indicating basal ROS production at the mitochondria (fig. 3C left). Upon treatment with sorafenib, the staining pattern changed to a bright nuclear fluorescence demonstrating oxidation of CellRox Green and binding of oxidized CellRox Green to nuclear DNA. Co-incubation of sorafenib treated cells with the antioxidative agent N-acetylcysteine (NAC) efficiently prevented bright nuclear fluorescence indicating inhibition of ROS accumulation (fig. 3C left). However, although NAC efficiently prevented sorafenib-induced ROS accumulation, it failed to prevent sorafenib-induced depolarization of mitochondria, as shown by JC-1 staining and FACS analysis of sorafenib- and NAC-treated cells (fig. 3C right). Hence, sorafenib-induced mitochondrial depolarization is independent and occurs up-stream of ROS accumulation. To further study the role of caspases in the mechanism underlying sorafenib-mediated sensitization of RCC cells toward TRAIL-induced cell death, RCC cells were pre-treated with sorafenib and subsequently exposed to TRAIL in the presence of the caspase-8 inhibitor z-letd- FMK, the caspase-3/7 inhibitor z-devd-fmk or the pan-caspase inhibitor Q-VD-OPh. After 6 h of incubation, cells were stained with annexin V- FITC/PI. Flow cytometric analysis revealed that all inhibitors potently reduced the proportion of apoptotic cells, with Q-VD-OPh showing the strongest effect (fig. 4A) almost completely inhibiting sorafenib/trail-induced apoptosis. Interestingly, FACS analysis of identically treated cells stained with JC-1 showed that Q-VD-OPh had no effect on sorafenib-induced mitochondrial depolarization (fig. 4B). Also, like mitochondrial depolarization, sorafenib-induced ROS accumulation was not influenced by Q-VD-OPh and thus is independent of caspase activity, as shown by the bright CellRox Green staining induced by sorafenib in Q-VD-OPh-pretreated RCC cells (fig. 4C). Sorafenib-induced depolarization of mitochondria and ROS accumulation thus are independent and functionally up-stream of caspase activation. TRAIL alone did not induce caspase-8 activation in RCC-26 cells but efficiently triggered processing of caspase-8 in cells pre-incubated with sorafenib (fig. 2A). Therefore, we investigated whether sorafenib-induced accumulation of ROS is instrumental for caspase-8 activation upon TRAIL treatment. To this end, RCC cells were treated with sorafenib and TRAIL and active caspase-8 was labeled with the fluorescent irreversible caspase-8 inhibitor FAM-LETD-FMK. Subsequent flow cytometric analysis revealed that the presence of NAC strongly reduced the number of cells positive for active caspase-8 labeling upon sorafenib/trail treatment. For example, only 4% of RCC-26 control cells were positive for active caspase-8 whereas about 38% of sorafenib/trailtreated cells were fluorescent for active caspase-8 (fig. 5A). Inhibition of ROS accumulation by coincubation with NAC strongly reduced the fraction of RCC-26 cells positive for active caspase-8 from 38% to 15% (fig. 5A). NAC-mediated inhibition of sorafenib/trail-induced caspase-8 processing was confirmed by immunoblotting. The processing of caspase-8 to its subunits in RCC-26 cells exposed to sorafenib and TRAIL was impaired by the ROS scavenger NAC (fig. 5B), indicating that ROS production by sorafenib is crucial for TRAIL to activate caspase-8. We next asked whether the sensitizing effect of sorafenib toward TRAIL-induced apoptosis is mediated by excessive ROS production. Therefore, RCC cells were pre-incubated for 1 h with NAC or 4
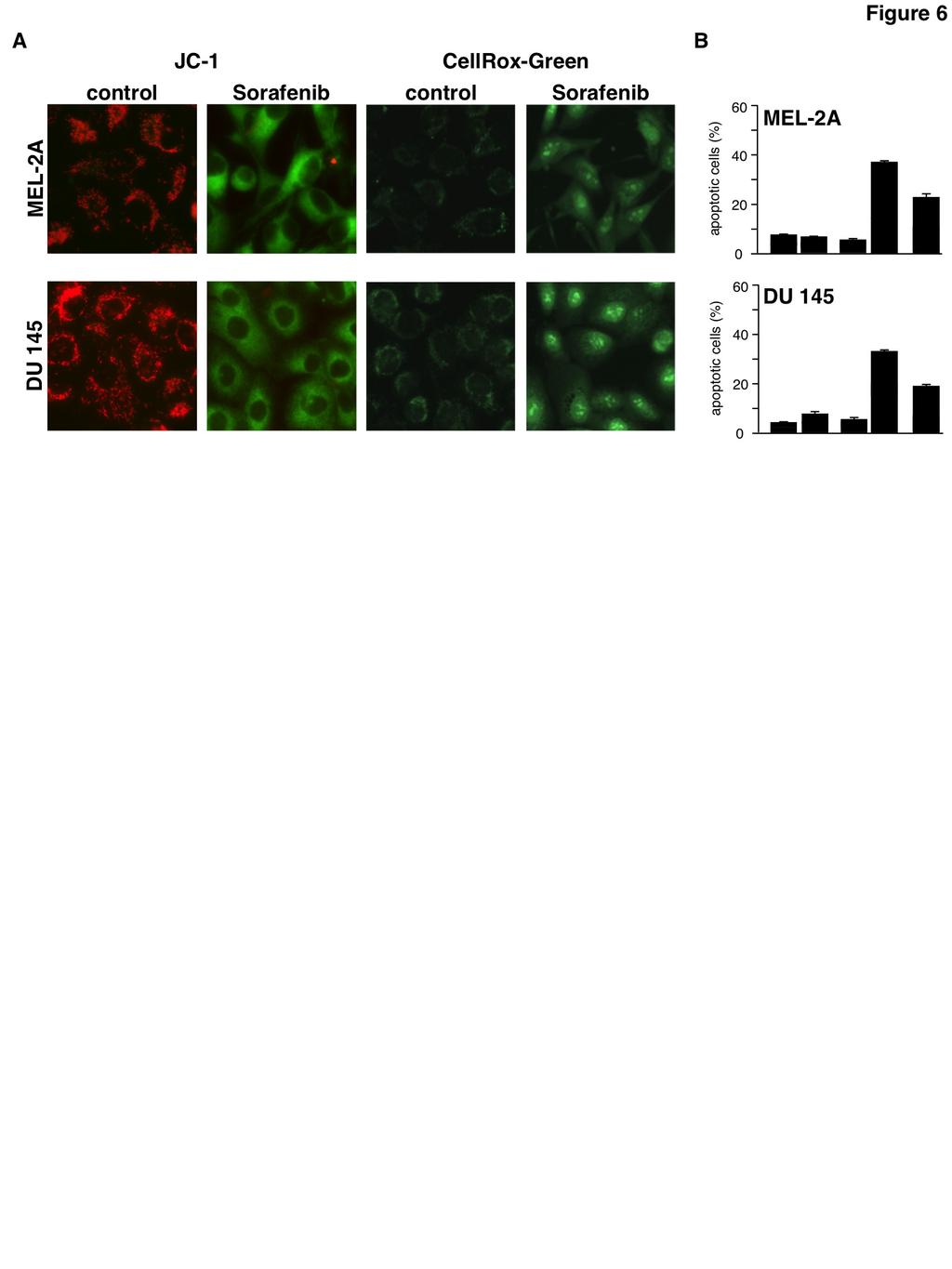
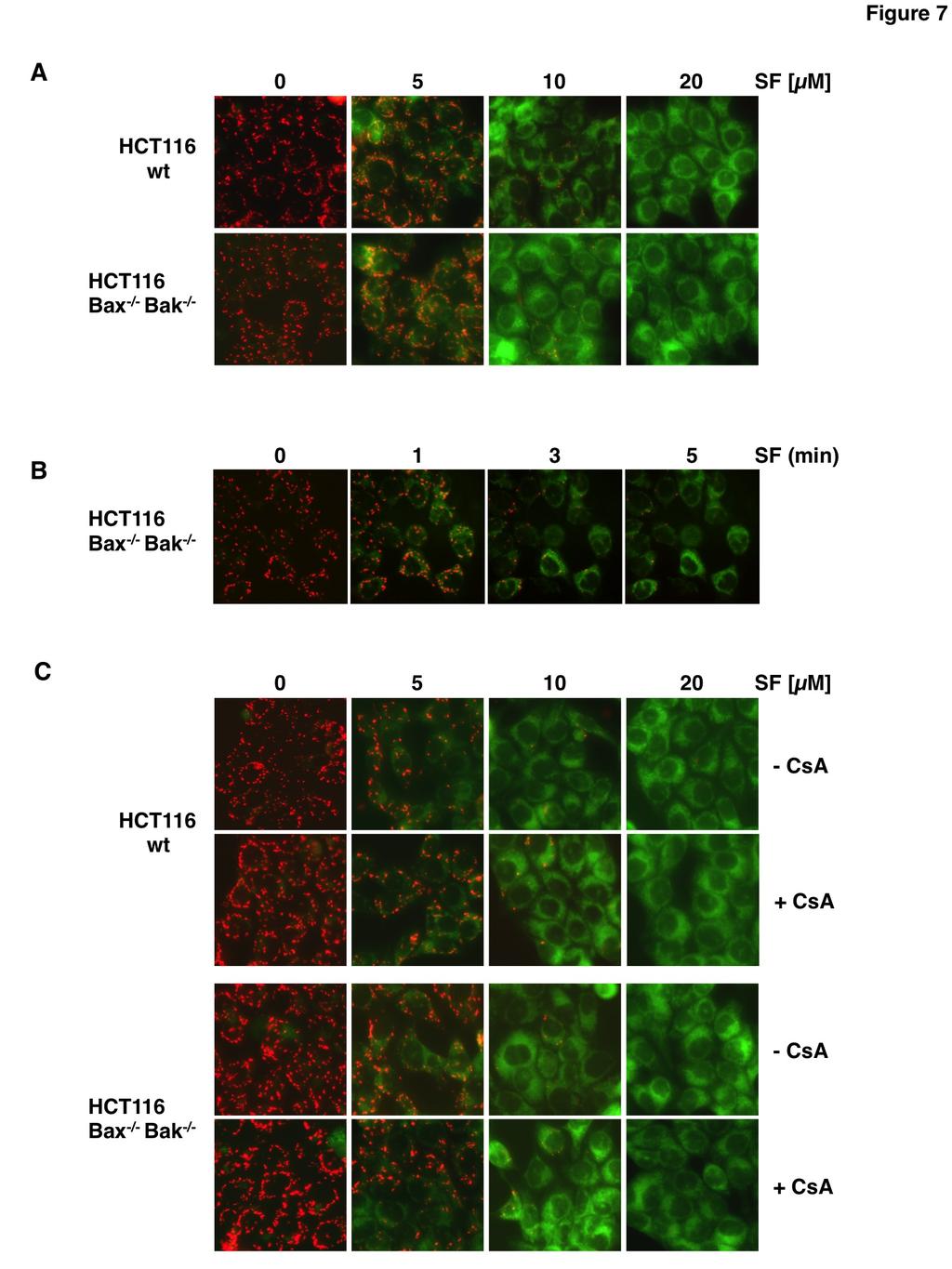
5 left untreated prior to addition of sorafenib for 14 h. Then TRAIL was added and additional 6 h later induction of apoptosis was assessed by flow cytometric analysis of annexin V-FITC/PI stained cells. In cells pre-incubated with NAC, the proportion of apoptotic cells (early apoptotic cells: annexin V-FITC+/PI- and late apoptotic cells: annexin V-FITC+/PI+) was reduced by approximately 50% in each cell line, i.e. from 65% to 30% in RCC-KP cells, from 60% to 28% in RCC-26 cells and from 36% to 19% in RCC-GW cells (fig. 5C). These data demonstrate that induction of apoptosis by the combination of sorafenib and TRAIL is strongly reduced upon blocking of ROS accumulation. Since inhibition of ROS accumulation by NAC abrogates sorafenibmediated sensitization of RCC cells toward TRAIL-induced apoptosis we conclude that ROS accumulation is crucial for the TRAIL-sensitizing effect of sorafenib. In order to test whether sorafenib-induced ROS production is a general mechanism that sensitizes cells to TRAIL-induced apoptosis, we incubated Mel-2a melanoma and DU145 prostate carcinoma cells with sorafenib and TRAIL. In both cell lines, sorafenib induced depolarization of the mitochondria and ROS formation (fig. 6A). Moreover, sorafenib sensitized Mel-2a and DU145 cells toward TRAIL-induced apoptosis, which was efficiently inhibited by the ROS scavenger NAC (fig. 6B). During canonical activation of the intrinsic apoptosis pathway, the BH123 proteins Bax and Bak mediate the dissipation of the mitochondrial membrane potential. To elucidate whether Bax and Bak are also involved in the depolarization of mitochondria induced by sorafenib, we used HCT116-wt colon cancer cells and the isogenic knockout cell line HCT116-Bax -/- /Bak -/- (37). Fluorescence microscopy of JC-1-stained cells showed that sorafenib induced depolarization in both cell lines in a concentration-dependent manner (fig. 7A), which became detectable already 1 to 3 min after the addition of sorafenib (fig. 7B). These results demonstrate that sorafenib-induced depolarization of mitochondria is a very early event and, more intriguingly, is not mediated by Bax or Bak. Besides Bax and Bak, the opening of the mitochondrial permeability transition pore (MPTP) induces MOMP and causes loss of ΔΨ m. However, cyclosporin A (CsA), which acts as a potent inhibitor of MPTP opening, did not prevent sorafenib-induced mitochondrial depolarization in HCT116-wt and HCT116-Bax -/- /Bak -/- cells (fig. 7C), demonstrating that sorafenib-induced depolarization is independent of both Bax/Bak and the MPTP. The rapid, Bax/Bak- and MPTP-independent mitochondrial depolarization induced by sorafenib resembles the mechanism induced by classical uncouplers of the mitochondrial respiratory chain like carbonyl cyanide m-chlorophenylhydrazone (CCCP). We therefore next asked whether CCCP sensitizes RCC-26 cells to TRAIL-induced apoptosis in a ROS-dependent mechanism. As supposed, flow cytometric analysis of JC-1-stained cells showed that CCCP treatment induced dissipation of ΔΨ m (fig. 8A), which was accompanied by ROS accumulation as shown by CellRox staining (fig. 8B). Furthermore, incubation with CCCP also increased TRAILinduced apoptosis evidenced by annexin V- FITC/PI staining (fig. 8C). Both effects induced by CCCP, increased ROS accumulation and increased apoptosis induction by TRAIL, were considerably reduced by pre-incubation with NAC. However, in the presence of NAC still a small portion of cells was positive for annexin V-FITC/PI, indicating that CCCP in combination with TRAIL might also, albeit to a lesser extent, induce necrosis-like cell death. Comparable to sorafenib, CCCP-induced mitochondrial depolarization was detectable 1 to 3 min upon treatment and independent of Bax and Bak (fig.7d). As mitochondrial depolarization is independent of Bax or Bak, we next asked whether in RCC cells also activation of caspase-8 and induction of apoptosis by the combined treatment with TRAIL and sorafenib is independent of Bax and Bak. To this end, expression of Bax and Bak in RCC-26 cells was downregulated by transfection with appropriate sirnas and cells were subsequently incubated with sorafenib and TRAIL. Despite efficient knockdown of Bax and Bak expression in RCC-26 cells (fig. 9A), caspase-8 activation upon combined sorafenib/trail treatment was not reduced (fig. 9B), indicating that caspase-8 activation is Bax and Bak-independent. However, despite unaltered caspase-8 activation, apoptosis induction by sorafenib/trail was decreased but not completely inhibited by knockdown of Bax and Bak (fig. 9C). In cells transfected with control sirna, 60% of the cells were positive for annexin V-FITC staining upon sorafenib/trail treatment. 5
6 This was reduced to 40% upon transfection of the cells with Bax and Bak sirna. Transfection of cells with Bak sirna did not completely abolish Bak expression. Thus, activation of residual Bak, mediated by caspase-8 cleaved and activated Bid, might account for the remaining induction of apoptosis. To completely block the mitochondrial amplification loop and to inhibit activation of residual Bak by tbid, we downregulated Bid expression additionally to Bax and Bak (fig. 10A). Similar to the Bax/Bak knockdown, simultaneous knockdown of Bid, Bax and Bak reduced but did not completely block sorafenib/trail-induced apoptosis (fig. 10B). This indicates that Bax/Bakregulated events of mitochondrial apoptosis signaling only partially contribute to apoptosis mediated by the combination of sorafenib/trail. Overall, induction of apoptosis upon sorafenib/trail treatment appears to be rather mediated by a type I pathway, which might further boost downstream activation of the mitochondrial pathway. DISCUSSION Resistance to classical chemotherapy or targeted treatment modalities including TRAIL is the major obstacle in oncology. RCC is one of the most chemoresistant tumor types and only the introduction of multi-targeted kinase inhibitors has provided some, rather limited therapeutic progress. Here, we provide evidence for an interesting new mechanism how sorafenib may be utilized to overcome resistance to apoptosis induction. As a model agent, we employed the death ligand TRAIL that induces apoptosis through a Bax/Bakcontrolled (type II) mitochondrial pathway in most solid tumors. The multi-kinase inhibitor sorafenib is approved for the treatment of advanced hepatocellular and renal cell carcinoma and shows promising antitumor activity as a single agent. However, primary and acquired drug resistance decreases the rate and duration of tumor response and impedes survival benefits (38,39). Nevertheless, the multiple targets of sorafenib make it an attractive choice for combination therapy with TRAIL and, consequently, sorafenib was shown to sensitize various cancer cells to TRAIL-induced apoptosis (30,40,41). Here, we present a new mechanism by which sorafenib overcomes TRAIL resistance of RCC cells. Increased ROS levels, generated upon sorafenibinduced mitochondrial depolarization, enable TRAIL to activate caspase-8 and to induce cell death. The anticancer effect of sorafenib is assumed to rely on its ability to inhibit cancer cell proliferation and to block angiogenesis through inhibition of Raf, PDGFR, and VEGFR (38). In addition, sorafenib induces apoptosis and sensitizes tumor cells to chemo- and radiotherapy by modulating STAT3 (42), Akt (43), NF-κB (29,44), and apoptosis signaling pathways. Particularly sorafenib-mediated downregulation of Mcl-1 seems to be crucial to sensitize cancer cells for TRAIL-induced cell death (29,45). Downregulation of Mcl-1, a specific Bak antagonist, lowers the apoptotic threshold and enables TRAIL to induce apoptosis via a Bakdependent mitochondrial pathway. Hence, even TRAIL resistance caused by Bax deficiency is overcome by sorafenib-induced Mcl-1 downregulation (31). However, RCC cells treated with sorafenib do not show consistently decreased levels of Mcl-1, indicating that in these cells downregulation of Mcl-1 is unlikely to be involved in overcoming TRAIL resistance. Furthermore, the specific sirna-mediated knockdown of Mcl-1 did not affect TRAIL resistance in RCC cells. In the present study, we further demonstrate that sorafenib sensitizes RCC cells for apoptosis induced by TRAIL through a mechanism that depends on sorafenib-induced ROS accumulation. This conclusion is supported by the prevention of sorafenib-induced ROS accumulation by NAC, which results in the inhibition of sorafenib/trailinduced apoptosis. Importantly, NAC does not inhibit mitochondrial depolarization indicating that depolarization per se is insufficient to enhance TRAIL-induced apoptosis and rather generation of ROS downstream of mitochondrial depolarization provokes sensitization toward TRAIL. It has been shown that in FADD-deficient acute lymphoblastic leukemia cells ROS induction is a critical regulator of necroptotic cell death induced by a Smac mimetic in combination with TNFα. Consistent with the definition of necroptosis, this cell death is caspase-independent (46,47). In contrast, ROS mediated cell death induced by sorafenib/trail is blocked by the caspase inhibitor Q-VD-OPh, indicating induction of apoptotic cell death. ROS production has been previously also reported during sorafenib-mediated cytotoxicity in hepatocarcinoma and EBV-transformed B cells 6
7 (48,49). The increased ROS production might be a consequence of activation of a NAD(P)H oxidase (48) or sorafenib-induced inactivation of mitochondrial complex I (NADH dehydrogenase) (50). In neuroblastoma cells, downregulation of complex I, induced by sorafenib, results in a caspase- and Bcl-2 independent loss of mitochondrial transmembrane potential (ΔΨ m ) that is accompanied by increased ROS levels and induction of cell death (50). In contrast to neuroblastoma cells, however, in which the decrease of ΔΨ m became evident after 12 h of treatment, in RCC cells depolarization was already detectable 1 to 3 min after addition of sorafenib. This almost instant depolarization makes a mechanism involving downregulation of the NADH dehydrogenase most unlikely. Interestingly, it has been shown that sorafenib, in contrast to the kinase inhibitors imatinib, dasatinib, and sunitinib, can directly impair mitochondrial function. Sorafenib can act as an inhibitor of the oxidative phosphorylation complexes and/or as an uncoupler of oxidative phosphorylation (51), and both mechanisms might explain the rapid sorafenib-induced mitochondrial depolarization. Although the exact mechanism of ROS induction is elusive, uncoupling of oxidative phosphorylation by sorafenib is compatible with our finding that sorafenib-mediated depolarization is independent of Bax, Bak and opening of the mitochondrial permeability transition pore. In line with this hypothesis, uncoupling of oxidative phosphorylation is accompanied by generation of ROS and results in sensitization toward TRAIL, as shown for the classical uncoupler CCCP. CCCP sensitizes human colon carcinoma cells to TRAILinduced apoptosis by enhancing caspase-8 activation via generation of ROS (52). ROS in turn modulate apoptosis signaling by assisting in the activation of initiator caspases (53). Likewise our results indicate that ROS act as up-stream signaling molecules that facilitate caspase-8 activation after receptor ligation, ultimately resulting in induction of apoptosis in RCC. ROS accumulation, in contrast to cell death, is not inhibited by caspase inhibitors. ROS accumulation is therefore caspase-independent and represents an up-stream event in sorafenib/trail-induced apoptosis. Furthermore, sorafenib/trailtriggered caspase-8 activation is inhibited by NAC, indicating that ROS accumulation plays a causal role in facilitating caspase-8 activation by TRAIL. Once caspase-8 is activated, however, additional sorafenib-induced mechanisms are likely to amplify TRAIL-induced caspase-8 activation and apoptosis in RCC cells. Such an amplification of caspase-8 activation via the mitochondrial pathway upon depolarization and ROS accumulation has been shown for TRAILresistant human colon carcinoma cell lines. Depolarization and ROS accumulation induced by CCCP sensitizes these cells for TRAIL-induced caspase-8 activation and apoptosis but complete processing of caspase-8 in response to TRAIL required a functional mitochondrial amplification loop (52). In RCC cells the initial step in overcoming TRAIL resistance seems to be the increased TRAIL-induced caspase-8 activation facilitated by sorafenib-mediated ROS accumulation, which enables the death receptors to kill cells via a type I, largely Bax/Bak-independent apoptosis pathway. However, a participation of the mitochondrial cell death pathway in RCC cell lines upon sorafenib/trail treatment is obvious, because of the reduced induction of apoptosis upon Bax and Bak knockdown. Thus, it is likely that enhancement of the mitochondrial amplification loop e.g. by a downregulation of Mcl-1 further boosts sorafenib/trail-induced apoptosis. Both, the death ligand TRAIL and sorafenib have been proposed as promising antitumor agents although their therapeutic use is restricted due to primary and acquired resistance of tumors. However, we show here that the combined use of TRAIL and sorafenib can overcome resistance of various RCC cell lines. Based on the presented data, we suggest a new pathway that depends on sorafenib-induced ROS accumulation, which in turn mediates increased caspase-8 activation (fig. 11). This mechanism explains the sensitization of RCC cells to TRAIL-induced cell death by sorafenib. Importantly, we show that sorafenibinduced depolarization and ROS accumulation are not confined to RCC and might be more generally involved in sorafenib/trail-induced apoptosis. Thus, the combined use of TRAIL and sorafenib might be a useful strategy to broaden the therapeutic efficacy of both agents. Interestingly, while our finding of sorafenib-induced ROS production in mitochondria is so far restricted to in-vitro settings, elevated serum levels of advanced oxidation protein products have been recently associated with an improved overall survival of sorafenib-treated patients (48). 7
8 Experimental procedures Cell culture RCC lines were cultured as described (54). In brief, cells were grown in RPMI 1640 supplemented with 10% fetal calf serum (FCS), 100 U/ml penicillin and 0.1 mg/ml streptomycin (Life Technologies GmbH, Darmstadt, Germany). HCT116 wild-type cells and the isogenic double knockout subline HCT116-Bax -/- /Bak -/- were kindly provided by Dr. R.J. Youle, National Institutes of Health, Bethesda, MD, USA (37). Cells were grown in DMEM supplemented with 10% FCS, 100,000 U/l penicillin and 0.1 g/l streptomycin at 37 C with 5% CO 2 in a humidified atmosphere. Media and culture reagents were from Invitrogen (Karlsruhe, Germany). Antibodies and reagents Anti-Smac mab (#2954), anti-caspase-8 (1C12) mab (#9746), anti-bid (#2002), and anti-xiap mab (3B6) (#2045) were purchased from Cell Signaling Technology, Inc. (Boston, USA). Anti- Bak mab (clone TC102) was from Calbiochem (Merck KGaA, Darmstadt, Germany), anti-bax mab (clone YTH-2D2) was purchased from Trevigen (Gaithersburg, USA) and anti-mcl-1 (H- 260) from Santa Cruz Biotechnology (Santa Cruz, USA). Anti-Caspase-3 Ab (#AF605) was from R&D Systems GmbH (Wiesbaden-Nordenstadt, Germany), and anti-actin mab (AC-74) from Sigma-Aldrich (Taufkirchen, Germany). Secondary anti-rabbit, anti-goat, and anti-mouse horseradish peroxidase (HRP)-conjugated antibodies were from Promega (Mannheim, Germany) or Southern Biotechnology Associates (Birmingham, USA). RNase A was from Carl Roth GmbH (Karlsruhe, Germany), recombinant human TRAIL from R&D Systems GmbH, sorafenib (BAY ) from Enzo Life Sciences GmbH (Lörrach Germany), and NAC from Sigma- Aldrich. Pan-caspase inhibitor Q-VD-OPh, caspase-3 inhibitor z-devd-fmk, caspase-8 inhibitor z-letd-fmk was purchased from Merck KGaA. Small interfering RNA SmartPool On-Target plus Mcl-1, Bid, Bax, Bak and control sirnas were purchased from Dharmacon (Lafayette, USA). Transfection of cells was carried out by use of DharmaFECT Transfection Reagent 1 according to the manufacturer s instructions. Downregulation of respective proteins was confirmed by immunoblotting 24 h after transfection. Immunoblotting After trypsination, cells were washed twice with ice-cold PBS and lysed in 10 mm Tris-HCl ph 7.5, 137 mm NaCl, 1% Triton X-100, 2 mm EDTA, 1 mm pepstatin, 1 mm leupeptin, and 100 µm phenylmethyl sulfonylfluoride (PMSF). Protein concentration was determined using the Thermo Scientific Pierce BCA Protein Assay Kit (Life Technologies GmbH). Equal amounts of protein were separated by SDS-PAGE, electroblotted onto nitrocellulose membrane and visualized as described (55). For analysis of cytochrome c and SMAC release, cytosolic extracts were prepared according to a method described previously (55). Briefly, after induction of apoptosis, cells were harvested, washed in PBS, equilibrated in hypotonic buffer (20 mm HEPES ph 7.4, 10 mm KCl, 2 mm MgCl 2, 1 mm EDTA) supplemented with 100 µm PMSF and 0.75 mg/ml digitonin (Sigma-Aldrich) and incubated on ice for 3 min. Debris was pelleted by centrifugation at 10,000 x g at 4 C for 5 min and the supernatant was subjected to Western blot analysis. Mitochondrial outer membrane permeabilization Cells were harvested by trypsination and collected by centrifugation at 300 x g at 4 C for 5 min. Mitochondrial outer membrane permeabilization was assessed by staining the cells with JC-1 (5,5,6,6 -tetrachloro-1,1,3,3 tetraethyl-benzimidazolylcarbocyanin iodide; Life Technologies). JC-1 is a cationic dye that exhibits membrane potential-dependent accumulation and formation of red fluorescent J-aggregates in mitochondria with a high membrane potential. At low membrane potential (upon MOMP) JC-1 shows a cytosolic green fluorescence. Mitochondrial permeability transition was analyzed by flow cytometry and cells with increased green fluorescence were assumed as cells with reduced mitochondrial membrane potential (ΔΨ m ). Detection of apoptotic cell death by flow cytometry DNA fragmentation was analyzed as described (56). Briefly, cells were harvested and collected by 8
9 centrifugation at 300 x g for 5 min, washed with PBS at 4 C, and fixed in PBS/2% (vol/vol) formaldehyde on ice for 30 min. After fixation, cells were incubated with ethanol/pbs (2:1, vol/vol) for 15 min on ice, pelleted, and resuspended in PBS containing 40 µg/ml RNase A. After incubation for 30 min at 37 C, cells were pelleted and finally resuspended in PBS containing 50 µg/ml propidium iodide. Relative cellular DNA content was quantified using a FACScan (Becton Dickinson, Heidelberg, Germany) and the fraction of hypodiploid cells was calculated using CELLQuest software. Data is given in % hypodiploid (subg1), which reflects the number of apoptotic cells. Alternatively, cell death was determined by staining cells with annexin V- fluorescein isothiocyanate (FITC) and propidium iodide (PI) (57). Briefly, cells were washed twice with cold PBS and resuspended in 10 mm Hepes (ph7.4), 140 mm NaCl, 2.5 mm CaCl 2 at 1x10 6 cells/ml. To 100 µl of the solution (1 x 10 5 cells), 5 µl of annexin V-FITC (BD Biosciences, Heidelberg, Germany) and 10 µl PI (20 mg/ml, Sigma-Aldrich) were added. Analysis was performed using a FACScan and CELLQuest software (Becton Dickinson). Detection of Bax/Bak conformational change Activation of Bax and Bak, i.e. their conformational change and exposure of an N- terminal epitope, was detected by the use of primary antibodies specific for the Bax or Bak N- terminal domains (Bax-NT from Upstate Biotechnology, and Bak-NT from Merck) and fluorescently labeled secondary antibodies. After fluorescent staining cells were subjected to flow cytometry (58). In brief 1 x 10 5 cells were harvested, washed in PBS and fixed for 30 min with 0.5% paraformaldehyde in PBS. After washing, cells were resuspended in PBS supplemented with 1% FCS and 0.1% saponin and incubated with the Bax-NT or Bak-NT specific antibody. Thereafter, cells were washed in PBS and incubated with a secondary FITC-labeled goatanti-rabbit or goat-anti-mouse IgG (H + L) antibodies (Jackson ImmunoResearch Laboratories, INC., Newmarket, UK) at a final concentration of 1 µg/ml for 30 min at 4 C in the dark. After washing and resuspension cells were immediately analyzed by flow cytometry. Caspase-8 activation Cells with active caspase-8 were detected using the FAM-FLICA TM Caspase-8 Assay Kit (Immunochemistry Technology, LLC, Bloomington, USA) according to the manufacturer's instructions. This kit employs a carboxyfluorescein-labeled fluoromethyl ketone peptide inhibitor of caspase-8 (FAM-LETD-FMK), which is cell-permeable and a non-cytotoxic fluorochrome. Detection of ROS Reactive oxygen species (ROS) in cells were analyzed by use of the fluorogenic CellROX Green reagent (Invitrogen, Life Technologies), according to the manufacturer s instructions. In brief, 1x10 5 cells were plated in a 12-well plate and incubated with the indicated drugs. Upon treatment CellROX reagent was added to a final concentration of 5 µm and cells were incubated for 60 min at 37 C. Microscopy images were taken using an Axiovert 200 fluorescence microscope (Carl Zeiss, Inc.) equipped with a Hamamatsu ORCA-ER digital camera using the Openlab software (Improvision). Acknowledgments: This work was kindly supported by the Stiftung Urologische Forschung Berlin. Conflict of interest: The authors declare no conflict of interest Author Contributions B. G. performed experiments, analyzed the experiments and wrote the manuscript A. R. and A. R. performed and analyzed experiments. R. P. analyzed data and reviewed the manuscript. F. E. performed experiments, analyzed the experiments and wrote the manuscript. K. S. O. analyzed data and wrote the manuscript. P. D. supervised the work, analyzed the experiments and wrote the manuscript. All authors reviewed the results and approved the final version of the manuscript. 9
10 REFERENCES 1. Falschlehner, C., Ganten, T. M., Koschny, R., Schaefer, U., and Walczak, H. (2009) TRAIL and other TRAIL receptor agonists as novel cancer therapeutics. Adv Exp Med Biol 647, Gonzalvez, F., and Ashkenazi, A. (2010) New insights into apoptosis signaling by Apo2L/TRAIL. Oncogene 29, Yang, A., Wilson, N. S., and Ashkenazi, A. (2010) Proapoptotic DR4 and DR5 signaling in cancer cells: toward clinical translation. Curr Opin Cell Biol 22, Chaudhary, P. M., Eby, M., Jasmin, A., Bookwalter, A., Murray, J., and Hood, L. (1997) Death receptor 5, a new member of the TNFR family, and DR4 induce FADD-dependent apoptosis and activate the NF-kappaB pathway. Immunity 7, Kischkel, F. C., Lawrence, D. A., Chuntharapai, A., Schow, P., Kim, K. J., and Ashkenazi, A. (2000) Apo2L/TRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity 12, Suzuki, Y., Nakabayashi, Y., and Takahashi, R. (2001) Ubiquitin-protein ligase activity of X-linked inhibitor of apoptosis protein promotes proteasomal degradation of caspase-3 and enhances its antiapoptotic effect in Fas-induced cell death. Proc Natl Acad Sci U S A 98, Green, D. R., and Kroemer, G. (2004) The pathophysiology of mitochondrial cell death. Science 305, Du, C., Fang, M., Li, Y., Li, L., and Wang, X. (2000) Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 102, Scaffidi, C., Fulda, S., Srinivasan, A., Friesen, C., Li, F., Tomaselli, K. J., Debatin, K. M., Krammer, P. H., and Peter, M. E. (1998) Two CD95 (APO-1/Fas) signaling pathways. Embo J 17, Jost, P. J., Grabow, S., Gray, D., McKenzie, M. D., Nachbur, U., Huang, D. C., Bouillet, P., Thomas, H. E., Borner, C., Silke, J., Strasser, A., and Kaufmann, T. (2009) XIAP discriminates between type I and type II FAS-induced apoptosis. Nature 460, Gillissen, B., Richter, A., Overkamp, T., Essmann, F., Hemmati, P. G., Preissner, R., Belka, C., and Daniel, P. T. (2013) Targeted therapy of the XIAP/proteasome pathway overcomes TRAIL-resistance in carcinoma by switching apoptosis signaling to a Bax/Bak-independent 'type I' mode. Cell Death Dis 4, e Zou, H., Li, Y., Liu, X., and Wang, X. (1999) An APAF-1.cytochrome c multimeric complex is a functional apoptosome that activates procaspase-9. The Journal of biological chemistry 274, Yamada, H., Tada-Oikawa, S., Uchida, A., and Kawanishi, S. (1999) TRAIL causes cleavage of bid by caspase-8 and loss of mitochondrial membrane potential resulting in apoptosis in BJAB cells. Biochem Biophys Res Commun 265, Luo, X., Budihardjo, I., Zou, H., Slaughter, C., and Wang, X. (1998) Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 94, Slee, E. A., Keogh, S. A., and Martin, S. J. (2000) Cleavage of BID during cytotoxic drug and UV radiation-induced apoptosis occurs downstream of the point of Bcl-2 action and is catalysed by caspase-3: a potential feedback loop for amplification of apoptosis-associated mitochondrial cytochrome c release. Cell Death Differ 7, Daniel, P. T., Schulze-Osthoff, K., Belka, C., and Guner, D. (2003) Guardians of cell death: the Bcl-2 family proteins. Essays Biochem 39, van Delft, M. F., and Huang, D. C. (2006) How the Bcl-2 family of proteins interact to regulate apoptosis. Cell Res 16, Bender, T., and Martinou, J. C. (2013) Where killers meet--permeabilization of the outer mitochondrial membrane during apoptosis. Cold Spring Harb Perspect Biol 5, a Schmelz, K., Wieder, T., Tamm, I., Muller, A., Essmann, F., Geilen, C. C., Schulze-Osthoff, K., Dorken, B., and Daniel, P. T. (2004) Tumor necrosis factor alpha sensitizes malignant cells to 10
11 chemotherapeutic drugs via the mitochondrial apoptosis pathway independently of caspase-8 and NFkappaB. Oncogene 23, Marini, P., Schmid, A., Jendrossek, V., Faltin, H., Daniel, P. T., Budach, W., and Belka, C. (2005) Irradiation specifically sensitises solid tumour cell lines to TRAIL mediated apoptosis. BMC Cancer 5, Keane, M. M., Rubinstein, Y., Cuello, M., Ettenberg, S. A., Banerjee, P., Nau, M. M., and Lipkowitz, S. (2000) Inhibition of NF-kappaB activity enhances TRAIL mediated apoptosis in breast cancer cell lines. Breast Cancer Res Treat 64, Brooks, A. D., Jacobsen, K. M., Li, W., Shanker, A., and Sayers, T. J. (2010) Bortezomib sensitizes human renal cell carcinomas to TRAIL apoptosis through increased activation of caspase-8 in the death-inducing signaling complex. Mol Cancer Res 8, Chen, K. F., Yeh, P. Y., Hsu, C., Hsu, C. H., Lu, Y. S., Hsieh, H. P., Chen, P. J., and Cheng, A. L. (2009) Bortezomib overcomes tumor necrosis factor-related apoptosis-inducing ligand resistance in hepatocellular carcinoma cells in part through the inhibition of the phosphatidylinositol 3-kinase/Akt pathway. J Biol Chem 284, Inoue, S., Mai, A., Dyer, M. J., and Cohen, G. M. (2006) Inhibition of histone deacetylase class I but not class II is critical for the sensitization of leukemic cells to tumor necrosis factor-related apoptosisinducing ligand-induced apoptosis. Cancer Res 66, Huang, S., and Sinicrope, F. A. (2008) BH3 mimetic ABT-737 potentiates TRAIL-mediated apoptotic signaling by unsequestering Bim and Bak in human pancreatic cancer cells. Cancer Res 68, Mott, J. L., Bronk, S. F., Mesa, R. A., Kaufmann, S. H., and Gores, G. J. (2008) BH3-only protein mimetic obatoclax sensitizes cholangiocarcinoma cells to Apo2L/TRAIL-induced apoptosis. Mol Cancer Ther 7, Huang, S., and Sinicrope, F. A. (2010) Sorafenib inhibits STAT3 activation to enhance TRAILmediated apoptosis in human pancreatic cancer cells. Mol Cancer Ther 9, Chen, K. F., Tai, W. T., Liu, T. H., Huang, H. P., Lin, Y. C., Shiau, C. W., Li, P. K., Chen, P. J., and Cheng, A. L. (2010) Sorafenib Overcomes TRAIL Resistance of Hepatocellular Carcinoma Cells through the Inhibition of STAT3. Clin Cancer Res 16, Ricci, M. S., Kim, S. H., Ogi, K., Plastaras, J. P., Ling, J., Wang, W., Jin, Z., Liu, Y. Y., Dicker, D. T., Chiao, P. J., Flaherty, K. T., Smith, C. D., and El-Deiry, W. S. (2007) Reduction of TRAIL-induced Mcl-1 and ciap2 by c-myc or sorafenib sensitizes resistant human cancer cells to TRAIL-induced death. Cancer Cell 12, Abdulghani, J., Allen, J. E., Dicker, D. T., Liu, Y. Y., Goldenberg, D., Smith, C. D., Humphreys, R., and El-Deiry, W. S. (2013) Sorafenib Sensitizes Solid Tumors to Apo2L/TRAIL and Apo2L/TRAIL Receptor Agonist Antibodies by the Jak2-Stat3-Mcl1 Axis. PLoS One 8, e Gillissen, B., Wendt, J., Richter, A., Muer, A., Overkamp, T., Gebhardt, N., Preissner, R., Belka, C., Dorken, B., and Daniel, P. T. (2010) Endogenous Bak inhibitors Mcl-1 and Bcl-xL: differential impact on TRAIL resistance in Bax-deficient carcinoma. J Cell Biol 188, Wilhelm, S., Carter, C., Lynch, M., Lowinger, T., Dumas, J., Smith, R. A., Schwartz, B., Simantov, R., and Kelley, S. (2006) Discovery and development of sorafenib: a multikinase inhibitor for treating cancer. Nat Rev Drug Discov 5, Iyer, R., Fetterly, G., Lugade, A., and Thanavala, Y. (2010) Sorafenib: a clinical and pharmacologic review. Expert Opin Pharmacother 11, Kane, R. C., Farrell, A. T., Saber, H., Tang, S., Williams, G., Jee, J. M., Liang, C., Booth, B., Chidambaram, N., Morse, D., Sridhara, R., Garvey, P., Justice, R., and Pazdur, R. (2006) Sorafenib for the treatment of advanced renal cell carcinoma. Clinical cancer research : an official journal of the American Association for Cancer Research 12, Bracarda, S., Caserta, C., Sordini, L., Rossi, M., Hamzay, A., and Crino, L. (2007) Protein kinase inhibitors in the treatment of renal cell carcinoma: sorafenib. Ann Oncol 18 Suppl 6, vi
12 36. Hoffmann, J. C., Pappa, A., Krammer, P. H., and Lavrik, I. N. (2009) A new C-terminal cleavage product of procaspase-8, p30, defines an alternative pathway of procaspase-8 activation. Mol Cell Biol 29, Wang, C., and Youle, R. J. (2012) Predominant requirement of Bax for apoptosis in HCT116 cells is determined by Mcl-1's inhibitory effect on Bak. Oncogene 31, Ibrahim, N., Yu, Y., Walsh, W. R., and Yang, J. L. (2012) Molecular targeted therapies for cancer: sorafenib mono-therapy and its combination with other therapies (review). Oncol Rep 27, Berasain, C. (2013) Hepatocellular carcinoma and sorafenib: too many resistance mechanisms? Gut 62, Chen, K. F., Chen, H. L., Shiau, C. W., Liu, C. Y., Chu, P. Y., Tai, W. T., Ichikawa, K., Chen, P. J., and Cheng, A. L. (2013) Sorafenib and its derivative SC-49 sensitize hepatocellular carcinoma cells to CS-1008, a humanized anti-tnfrsf10b (DR5) antibody. Br J Pharmacol 168, Nojiri, K., Sugimoto, K., Shiraki, K., Tameda, M., Inagaki, Y., Ogura, S., Kasai, C., Kusagawa, S., Yoneda, M., Yamamoto, N., Takei, Y., Nobori, T., and Ito, M. (2013) Sorafenib and TRAIL have synergistic effect on hepatocellular carcinoma. Int J Oncol 42, Yang, F., Brown, C., Buettner, R., Hedvat, M., Starr, R., Scuto, A., Schroeder, A., Jensen, M., and Jove, R. (2010) Sorafenib induces growth arrest and apoptosis of human glioblastoma cells through the dephosphorylation of signal transducers and activators of transcription 3. Mol Cancer Ther 9, Oh, S. J., Erb, H. H., Hobisch, A., Santer, F. R., and Culig, Z. (2012) Sorafenib decreases proliferation and induces apoptosis of prostate cancer cells by inhibition of the androgen receptor and Akt signaling pathways. Endocr Relat Cancer 19, Kuo, Y. C., Lin, W. C., Chiang, I. T., Chang, Y. F., Chen, C. W., Su, S. H., Chen, C. L., and Hwang, J. J. (2012) Sorafenib sensitizes human colorectal carcinoma to radiation via suppression of NFkappaB expression in vitro and in vivo. Biomed Pharmacother 66, Meng, X. W., Lee, S. H., Dai, H., Loegering, D., Yu, C., Flatten, K., Schneider, P., Dai, N. T., Kumar, S. K., Smith, B. D., Karp, J. E., Adjei, A. A., and Kaufmann, S. H. (2007) Mcl-1 as a buffer for proapoptotic Bcl-2 family members during TRAIL-induced apoptosis: a mechanistic basis for sorafenib (Bay )-induced TRAIL sensitization. The Journal of biological chemistry 282, Laukens, B., Jennewein, C., Schenk, B., Vanlangenakker, N., Schier, A., Cristofanon, S., Zobel, K., Deshayes, K., Vucic, D., Jeremias, I., Bertrand, M. J., Vandenabeele, P., and Fulda, S. (2011) Smac mimetic bypasses apoptosis resistance in FADD- or caspase-8-deficient cells by priming for tumor necrosis factor alpha-induced necroptosis. Neoplasia 13, Schenk, B., and Fulda, S. (2015) Reactive oxygen species regulate Smac mimetic/tnfalpha-induced necroptotic signaling and cell death. Oncogene 34, Coriat, R., Nicco, C., Chereau, C., Mir, O., Alexandre, J., Ropert, S., Weill, B., Chaussade, S., Goldwasser, F., and Batteux, F. (2012) Sorafenib-induced hepatocellular carcinoma cell death depends on reactive oxygen species production in vitro and in vivo. Mol Cancer Ther 11, Park, G. B., Choi, Y., Kim, Y. S., Lee, H. K., Kim, D., and Hur, D. Y. (2014) ROS-mediated JNK/p38-MAPK activation regulates Bax translocation in Sorafenib-induced apoptosis of EBVtransformed B cells. Int J Oncol 44, Bull, V. H., Rajalingam, K., and Thiede, B. (2012) Sorafenib-induced mitochondrial complex I inactivation and cell death in human neuroblastoma cells. J Proteome Res 11, Will, Y., Dykens, J. A., Nadanaciva, S., Hirakawa, B., Jamieson, J., Marroquin, L. D., Hynes, J., Patyna, S., and Jessen, B. A. (2008) Effect of the multitargeted tyrosine kinase inhibitors imatinib, dasatinib, sunitinib, and sorafenib on mitochondrial function in isolated rat heart mitochondria and H9c2 cells. Toxicol Sci 106, Izeradjene, K., Douglas, L., Tillman, D. M., Delaney, A. B., and Houghton, J. A. (2005) Reactive oxygen species regulate caspase activation in tumor necrosis factor-related apoptosis-inducing ligandresistant human colon carcinoma cell lines. Cancer research 65,
13 53. Perez-Cruz, I., Carcamo, J. M., and Golde, D. W. (2007) Caspase-8 dependent TRAIL-induced apoptosis in cancer cell lines is inhibited by vitamin C and catalase. Apoptosis 12, Sturm, I., Stephan, C., Gillissen, B., Siebert, R., Janz, M., Radetzki, S., Jung, K., Loening, S., Dorken, B., and Daniel, P. T. (2006) Loss of the tissue-specific proapoptotic BH3-only protein Nbk/Bik is a unifying feature of renal cell carcinoma. Cell Death Differ 13, von Haefen, C., Wieder, T., Essmann, F., Schulze-Osthoff, K., Dorken, B., and Daniel, P. T. (2003) Paclitaxel-induced apoptosis in BJAB cells proceeds via a death receptor-independent, caspases-3/-8- driven mitochondrial amplification loop. Oncogene 22, Gillissen, B., Essmann, F., Hemmati, P. G., Richter, A., Richter, A., Oztop, I., Chinnadurai, G., Dorken, B., and Daniel, P. T. (2007) Mcl-1 determines the Bax dependency of Nbk/Bik-induced apoptosis. J Cell Biol 179, Gillissen, B., Essmann, F., Graupner, V., Starck, L., Radetzki, S., Dorken, B., Schulze-Osthoff, K., and Daniel, P. T. (2003) Induction of cell death by the BH3-only Bcl-2 homolog Nbk/Bik is mediated by an entirely Bax-dependent mitochondrial pathway. The EMBO journal 22, Hemmati, P. G., Muer, A., Gillissen, B., Overkamp, T., Milojkovic, A., Wendt, J., Dorken, B., and Daniel, P. T. (2010) Systematic genetic dissection of p14arf-mediated mitochondrial cell death signaling reveals a key role for p21cdkn1 and the BH3-only protein Puma/bbc3. J Mol Med (Berl) 88, FOOTNOTES The abbreviations used are: Bad, Bcl-2-associated agonist of cell death; Bak, Bcl-2 homologous antagonist/killer; Bax, Bcl-2 associated x protein; Bcl-2, B cell lymphoma 2; Bcl-xL, long splice variant of Bcl-x; BH3, Bcl-2 homology region 3; Bid, BH3-interacting domain death agonist; Bim, Bcl-2 interacting mediator of cell death; Bok, Bcl-2 related ovarian killer; cflip, cellular FLICE-like inhibitory protein; DISC, death-inducing signaling complex; FADD, Fas-associated protein with death domain; FITC, Fluorescein isothiocyanate; Mcl-1, Myeloid cell leukemia 1; MOMP, mitochondrial outer membrane permeabilization; NAC, N-acetylcysteine; Nbk/Bik, Natural born killer/bcl-2 interacting killer; NF-κB, nuclear factor-κb; PI, propidium iodide; Puma, p53 up-regulated modulator of apoptosis; SF, sorafenib; Smac, Second mitochondria-derived activator of caspase; STAT3, Signal transducer and activator of transcription 3; TRAIL, Tumor necrosis factor-related apoptosis inducing ligand; XIAP, X- linked inhibitor of apoptosis protein. FIGURE LEGENDS Figure 1: Sorafenib-mediated downregulation of Mcl-1 is not involved in the sensitization toward TRAIL induced apoptosis. A) Sorafenib overcomes TRAIL resistance of RCC cell lines. Indicated RCC cells were incubated with 50 ng/ml TRAIL for 24 h, 20 µm sorafenib for 38 h or pre-incubated for 14 h with sorafenib before addition of TRAIL. Apoptotic (hypodiploid) cells were determined by FACS analysis of relative cellular DNA content. TRAIL did not induce apoptosis in any of the cell lines. Sorafenib did not induce apoptosis in RCC-GW and only marginally in RCC-KP and RCC-26 cells. The combined treatment with sorafenib and TRAIL potently induced apoptosis in all three cell lines. The upper panel shows representative flow cytometry histograms of PI-stained cells. The percentage of hypodiploid, i.e. apoptotic cells is indicated between markers. Lower panel shows mean ± standard deviation (SD) of triplicates. B) Western blot analysis revealed downregulation of Mcl-1 upon sorafenib (SF) treatment in RCC-KW and RCC-26 cells. In contrast, XIAP expression levels were not reduced in any of the cell lines. C) RCC cells were either left untreated or transfected with non-targeting (c) or Mcl-1 sirna and knockdown of Mcl-1 was confirmed by Western blot analysis. D) After Mcl-1 downregulation, cells were treated with TRAIL, cultured for 13
14 additional 24 h and apoptotic cells were assessed by FACS analysis. None of the cell lines were sensitized for TRAIL-induced apoptosis upon Mcl-1 downregulation (mean ± standard deviation (SD) of triplicates). Figure 2: Sorafenib enables TRAIL to activate caspase-8 and the mitochondrial apoptosis signaling pathway. A) After pre-incubation with sorafenib cells were treated with TRAIL for the indicated periods of time and caspase processing, Bid cleavage and release of Smac and cytochrome c were analyzed by immunoblotting. Treatment of cells with sorafenib or TRAIL as single agents for 16 h neither induced caspase processing nor Bid cleavage, or release of SMAC or cytochrome c. In contrast, combined sorafenib/trail treatment resulted in processing of caspase-8 and -3 to the respective active subunits that became detectable after 4 h. This was accompanied by Bid activation, indicated by increased tbid levels, and release of cytochrome c and Smac into the cytosol. B) Cells were treated with sorafenib (SF), TRAIL, or a combination of both and activation of Bax and Bak was analyzed by staining of the cells with conformation-specific antibodies (Bax-NT and Bak-NT, respectively) and subsequent flow cytometry. As single agents sorafenib and TRAIL failed to induce activation of Bax or Bak. However, pre-incubation of cells with sorafenib enables TRAIL to activate Bax as well as Bak. Left panel shows representative flow cytometry histograms of immunostained cells. The percentage of cells positive for conformation-changed Bax or Bak is indicated between markers. Right panel shows mean ± standard deviation (SD) from triplicates. Figure 3: Sorafenib-induced mitochondrial depolarization is accompanied by accumulation of reactive oxygen species. A) RCC cell lines were treated with the indicated concentration of sorafenib and subsequently stained with JC-1 to analyze loss of mitochondrial membrane potential (ΔΨ m ). In control cells JC-1 accumulates in mitochondria and forms red fluorescent J-aggregates reflecting high mitochondrial membrane potential, whereas green cytosolic fluorescence, indicating low mitochondrial membrane potential, increases with the concentration of sorafenib. B) RCC cells were treated with TRAIL and sorafenib as single agents or in combination. Measurement of green JC-1 fluorescence intensity by flow cytometry on a single-cell level revealed that sorafenib but not TRAIL as single agents induces loss of ΔΨ m in each of the RCC cell lines. C) Cells were treated with 20 µm sorafenib alone or in combination with the free radical scavenger NAC. ROS accumulation was assessed by CellRox Green staining and fluorescence microscopy 6 h after treatment. Control cells showed a weak punctate green fluorescence pattern that changed to a bright nuclear fluorescence, demonstrating oxidation of CellRox Green by ROS and binding of the oxidized dye to nuclear DNA. Prevention of ROS accumulation by NAC inhibits nuclear fluorescence staining with CellRox green. In addition, cells with reduced ΔΨ m were determined by JC-1 staining and flow cytometry. Treated cells (open graphs) were compared to controls (gray). In contrast to ROS accumulation, sorafenibinduced loss of ΔΨ m was not inhibited by NAC. Figure 4: Sorafenib-induced depolarization and ROS production is independent of caspase activity. A) In addition to sorafenib (SF) and TRAIL, the caspase-3/7 (z-devd-fmk), caspase-8 (z-letd-fmk) or the pan-caspase inhibitor Q-VD-OPh were added at a final concentration of 20 µm to the culture medium of RCC-26 cells After 6 h, cells were harvested and apoptosis was analyzed by annexin V- FITC/PI staining and flow cytometry. Indicated are early (annexin V-FITC+/PI-) and late apoptotic (annexin V-FITC+/PI+) cells. Induction of apoptosis by combined sorafenib/trail treatment is strongly reduced by z-devd-fmk, z-letd-fmk and by Q-VD-OPh. B) Cells were treated with sorafenib in the absence or presence of 20 µm Q-VD-OPh (QVD) and loss of ΔΨ m was analyzed by JC-1 staining and FACS analysis. Treated cells (open graphs) were compared to controls (gray). In contrast to apoptosis, the broad-range pan-caspase inhibitor Q-VD-OPh does not inhibit sorafenib-induced loss of ΔΨ m. C) Cells were treated as in B and accumulation of ROS was analyzed by CellRox Green staining. The pan-caspase inhibitor Q-VD-OPh does not inhibit sorafenib-induced ROS accumulation. 14
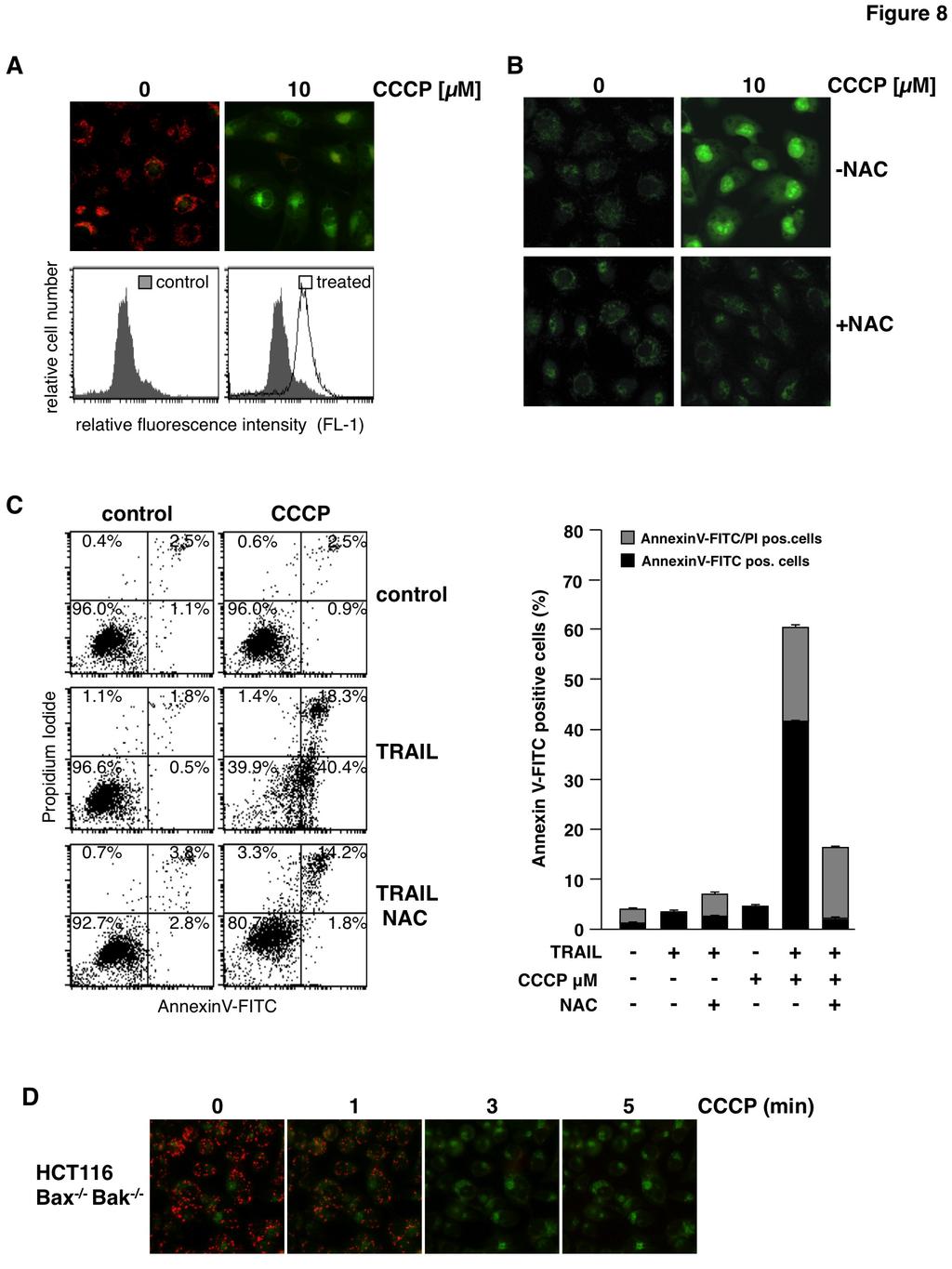
15 Figure 5: Sorafenib-induced ROS accumulation enables TRAIL to activate caspase-8 and is responsible for sensitization toward TRAIL induced cell death. A) RCC cells were treated with sorafenib, TRAIL, and sorafenib/trail in the absence or presence of NAC to prevent ROS accumulation. 8 h post treatment, cells with active caspase-8 were detected by flow cytometry using the FAM-FLICA Caspase-8 Detection Kit. The ROS scavenger NAC blocks sorafenib/trail-induced caspase-8 activation. B) RCC-26 cells were treated as in A and caspase-8 cleavage was analyzed by immunoblotting. Following combined treatment of sorafenib/trailprocaspase-8 is processed to its active subunits. Proteolytic processing of caspase-8 is strongly reduced in the presence of the ROS scavenger NAC. C) Induction of apoptosis was quantified by annexin V-FITC/PI staining and flow cytometry. Cells positive for annexin V-FITC (phosphatidylserine exposure) and negative for PI staining represent early apoptotic cells, whereas late apoptotic cells are positive for both annexin V-FITC and PI. Flow cytometric analysis revealed that induction of apoptosis by combined incubation with sorafenib and TRAIL is strongly reduced upon NAC-mediated inhibition of ROS accumulation. Panels show mean ± standard deviation (SD) of triplicates. Figure 6: Mitochondrial depolarization, ROS accumulation and induction of apoptosis upon sorafenib/trail treatment in cancer cell lines derived from different tissues. A) Mel-2a melanoma and DU145 prostate cancer cells were treated with sorafenib and stained with JC-1 or CellRox Green to analyze loss of ΔΨ m and ROS accumulation, respectively. Similar to RCC cells, sorafenib induced mitochondrial depolarization and ROS accumulation in both cell lines. B) Flow cytometric annexin V-FITC/PI staining revealed that induction of apoptosis by the combined treatment with sorafenib and TRAIL was strongly reduced by NAC in Mel-2a and DU145 cells. Figure 7: Sorafenib-induced mitochondrial depolarization upon sorafenib/trail treatment is independent of Bax and Bak and the MPTP. A) HCT116-wt cells and isogenic HCT116-Bax -/- /Bak -/- cells were treated with sorafenib and stained with JC-1 to analyze loss of mitochondrial membrane potential. Fluorescence microscopy (upper panel) showed decreased red and increased green fluorescence upon treatment with sorafenib in both cell lines, indicating that loss of ΔΨ m is independent of Bax and Bak. B) HCT116-Bax -/- /Bak -/- cells were treated with sorafenib for the indicated time periods and stained with JC-1. Loss of ΔΨ m is detectable 1 to 3 min after treatment in almost all cells. C) In addition to sorafenib, HCT116-wt and HCT116-Bax -/- /Bak -/- cells were pre-incubated with cyclosporin A (CsA) to block MPTP opening. Upon treatment, cells were stained with JC-1 and analyzed by fluorescence microscopy. CsA failed to prevent sorafenib-induced mitochondrial depolarization in HCT116-wt and HCT116-Bax -/- /Bak -/- cells demonstrating that sorafenibinduced depolarization is independent of both, MPTP and Bax/Bak. Figure 8: CCCP-induced mitochondrial depolarization and ROS accumulation sensitizes RCC-26 cells for TRAIL. A) RCC-26 cells were treated with CCCP and stained with JC-1 to analyze loss of mitochondrial membrane potential. Fluorescence microscopy (upper panel) and FACS analysis (lower panel) showed loss of red fluorescence and increase of green fluorescence upon CCCP treatment in all cells, indicating loss of ΔΨ m. B) RCC-26 cells were treated with CCCP alone or in combination with the free radical scavenger NAC. As shown by CellRox Green staining, CCCP-induced ROS accumulation was inhibited by NAC. C) In addition to TRAIL and CCCP, RCC-26 cells were pre-treated with NAC. Induction of apoptosis was quantified by flow cytometric detection of cells positive for annexin V-FITC (phosphatidylserine exposure) and negative for PI staining (intact plasma membrane), i.e., early apoptotic, and detection of annexin V-FITC/PI positive (late apoptotic) cells. FACS measurement revealed that CCCP sensitizes cells for TRAIL-induced apoptosis. Sensitization for TRAIL was inhibited by NAC. Left panel shows representative flow cytometry histograms, right panel shows mean ± standard deviation (SD) of triplicates. D) HCT116-Bax -/- /Bak -/- cells were treated with CCCP for the indicated time periods and stained with JC-1. Loss of ΔΨ m is detectable 1 to 3 min after treatment in almost all cells. 15
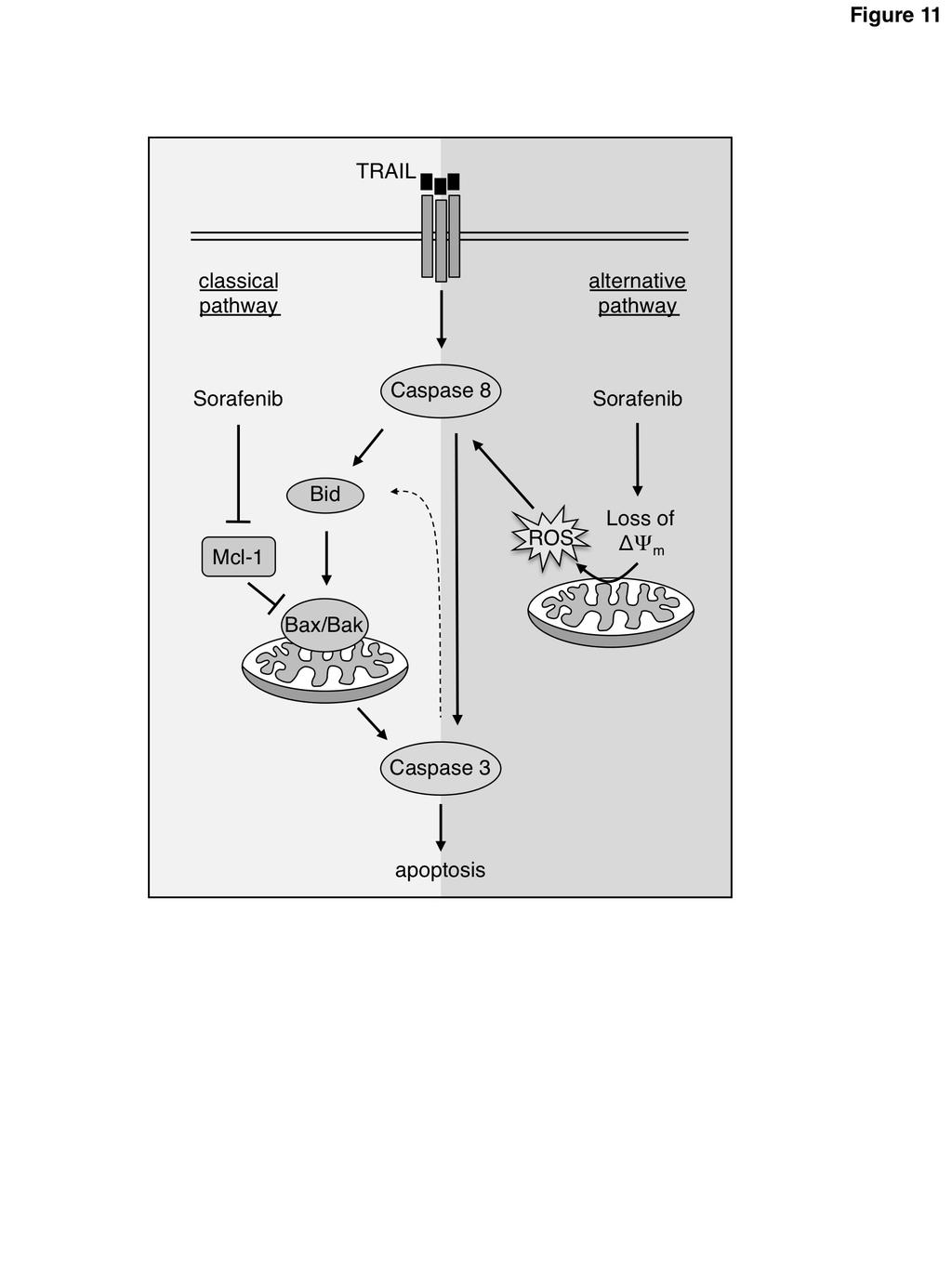
16 Figure 9: Caspase-8 activation and apoptosis upon the combined treatment with sorafenib and TRAIL is independent of Bax and Bak. A) RCC-26 cells were transfected with non-targeting or Bax- and Bak-targeting sirna. After 24 h, protein extracts were prepared and analyzed by Western blot to confirm downregulation of the targeted proteins. B) After downregulation of Bax and Bak, cells were treated with TRAIL and sorafenib. Cells with active caspase-8 were detected by flow cytometry using the FAM-FLICA Caspase-8 Detection Kit. Measurement revealed that knockdown of Bax and Bak did not inhibit caspase-8 activation by the combined treatment of cells with sorafenib and TRAIL. C) Cells were treated as in B) and apoptosis was quantified by flow cytometric detection of annexin V-FITC/PI positive cells. Downregulation of Bax and Bak decreased but did not prevent sorafenib/trail-induced apoptosis. Figure 10: Combined knockdown of Bax, Bak, and Bid does not prevent sorafenib/trail-induced apoptosis. A) RCC-26 cells were transfected with non-targeting or Bax-, Bak- and Bid-targeting sirna. After 24 h, protein extracts were prepared and analyzed by Western blot to confirm the downregulation of the targeted proteins. B) After combined downregulation of Bax, Bak, and Bid cells were treated with TRAIL and sorafenib and proportion of annexin V-FITC/PI positive cells was detected by flow cytometry. Measurement revealed that the combined knockdown of Bax, Bak, and Bid attenuated but did not prevent sorafenib/trail-induced apoptosis. Figure 11: Sensitization of cancer cells toward TRAIL-induced apoptosis by sorafenib. Sensitization of cancer cells to TRAIL-induced apoptosis is mediated by a classical mechanism, including downregulation of Mcl-1, which activates the Bax/Bak-triggered mitochondrial caspase cascade. Alternatively, as shown in this study, sorafenib can overcome TRAIL resistance by Bax/Bak-independent dissipation of the mitochondrial membrane potential, resulting in ROS accumulation and increased activation of caspase-8. 16
17 17
18 18
19 19
20 20
21 21
22 22
23 23
24 24
25 25
26 26
27 27
Supplementary Figures
 Supplementary Figures Figure S1. Validation of kinase regulators of ONC201 sensitivity. Validation and screen results for changes in cell viability associated with the combination of ONC201 treatment (1
Supplementary Figures Figure S1. Validation of kinase regulators of ONC201 sensitivity. Validation and screen results for changes in cell viability associated with the combination of ONC201 treatment (1
Repository of the Max Delbrück Center for Molecular Medicine (MDC) in the Helmholtz Association
 in the Helmholtz Association") Repository of the Max Delbrück Center for Molecular Medicine (MDC) in the Helmholtz Association http://edoc.mdc-berlin.de/10942 Endogenous Bak inhibitors Mcl-1 and Bcl-xL: differential impact on TRAIL
Repository of the Max Delbrück Center for Molecular Medicine (MDC) in the Helmholtz Association http://edoc.mdc-berlin.de/10942 Endogenous Bak inhibitors Mcl-1 and Bcl-xL: differential impact on TRAIL
Apoptotic Pathways in Mammals Dr. Douglas R. Green
 Apoptotic Pathways in Mammals Douglas R. Green 1 Apoptosis A form of cell death that is defined morphologically, and features a number of biochemical events Programmed cell death Cell death that occurs
Apoptotic Pathways in Mammals Douglas R. Green 1 Apoptosis A form of cell death that is defined morphologically, and features a number of biochemical events Programmed cell death Cell death that occurs
Impact factor: Reporter:4A1H0019 Chen Zi Hao 4A1H0023 Huang Wan ting 4A1H0039 Sue Yi Zhu 4A1H0070 Lin Guan cheng 4A1H0077 Chen Bo xuan
 Curcumin Protects Neonatal Rat Cardiomyocytes against High Glucose-Induced Apoptosis via PI3K/Akt Signalling Pathway Wei Yu,1,2 Wenliang Zha,1 Zhiqiang Ke,1 Qing Min,2 Cairong Li,1 Huirong Sun,3 and Chao
Curcumin Protects Neonatal Rat Cardiomyocytes against High Glucose-Induced Apoptosis via PI3K/Akt Signalling Pathway Wei Yu,1,2 Wenliang Zha,1 Zhiqiang Ke,1 Qing Min,2 Cairong Li,1 Huirong Sun,3 and Chao
Signaling Apoptosis. Scott André Oakes, M.D. Dept. of Pathology Univ. of Calif-San Francisco. Cyt c Release BAX/BAK. Apoptosome Formation
 Signaling Apoptosis Cyt c Release BAX/BAK Apoptosome Formation Caspase Activation Scott André Oakes, M.D. Dept. of Pathology Univ. of Calif-San Francisco Why do we need cell death? Sculpt Organs Control
Signaling Apoptosis Cyt c Release BAX/BAK Apoptosome Formation Caspase Activation Scott André Oakes, M.D. Dept. of Pathology Univ. of Calif-San Francisco Why do we need cell death? Sculpt Organs Control
Apoptosis Chapter 9. Neelu Yadav PhD
 Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org 1 Apoptosis: Lecture outline Apoptosis a programmed cell death pathway in normal homeostasis Core Apoptosis cascade is conserved Compare
Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org 1 Apoptosis: Lecture outline Apoptosis a programmed cell death pathway in normal homeostasis Core Apoptosis cascade is conserved Compare
Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108
 37:108") Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108 https://doi.org/10.1186/s13046-018-0774-7 CORRECTION Correction to: Novel smac mimetic APG- 1387 elicits ovarian cancer cell killing
Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108 https://doi.org/10.1186/s13046-018-0774-7 CORRECTION Correction to: Novel smac mimetic APG- 1387 elicits ovarian cancer cell killing
Supplementary Information
 Supplementary Information Significance of p53 dynamics in regulating apoptosis in response to ionizing radiation, and polypharmacological strategies Bing Liu 1,, Divesh Bhatt 1,, Zoltan N. Oltvai 2, Joel
Supplementary Information Significance of p53 dynamics in regulating apoptosis in response to ionizing radiation, and polypharmacological strategies Bing Liu 1,, Divesh Bhatt 1,, Zoltan N. Oltvai 2, Joel
C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is
 is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is") ' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
Silibinin i activates p53-caspase-2 pathway and causes caspase-mediated cleavage of Cip1/p21 in apoptosis
 Silibinin i activates p53-caspase-2 pathway and causes caspase-mediated cleavage of Cip1/p21 in apoptosis induction in bladder transitional-cell papilloma RT4 cells: Evidence for a regulatory loop between
Silibinin i activates p53-caspase-2 pathway and causes caspase-mediated cleavage of Cip1/p21 in apoptosis induction in bladder transitional-cell papilloma RT4 cells: Evidence for a regulatory loop between
#19 Apoptosis Chapter 9. Neelu Yadav PhD
 #19 Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org Why cells decide to die? - Stress, harmful, not needed - Completed its life span Death stimulation or Stress Cell Survival Death Functions
#19 Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org Why cells decide to die? - Stress, harmful, not needed - Completed its life span Death stimulation or Stress Cell Survival Death Functions
The Annexin V Apoptosis Assay
 The Annexin V Apoptosis Assay Development of the Annexin V Apoptosis Assay: 1990 Andree at al. found that a protein, Vascular Anticoagulant α, bound to phospholipid bilayers in a calcium dependent manner.
The Annexin V Apoptosis Assay Development of the Annexin V Apoptosis Assay: 1990 Andree at al. found that a protein, Vascular Anticoagulant α, bound to phospholipid bilayers in a calcium dependent manner.
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
p53 and Apoptosis: Master Guardian and Executioner Part 2
 p53 and Apoptosis: Master Guardian and Executioner Part 2 p14arf in human cells is a antagonist of Mdm2. The expression of ARF causes a rapid increase in p53 levels, so what would you suggest?.. The enemy
p53 and Apoptosis: Master Guardian and Executioner Part 2 p14arf in human cells is a antagonist of Mdm2. The expression of ARF causes a rapid increase in p53 levels, so what would you suggest?.. The enemy
The effect of elemene reversing the multidurg resistance of A549/DDP lung cancer cells
 213 12 33 12 TUMOR Vol. 33, December 213 www.tumorsci.org 161 Basic Research DOI: 1.3781/j.issn.1-7431.213.12. 5 Copyright 213 by TUMOR A549/DDP 1, 2 2 1 3 4 1 1 1. 3619 2. 355 3. 3611 4. 361 elemene cisplatin
213 12 33 12 TUMOR Vol. 33, December 213 www.tumorsci.org 161 Basic Research DOI: 1.3781/j.issn.1-7431.213.12. 5 Copyright 213 by TUMOR A549/DDP 1, 2 2 1 3 4 1 1 1. 3619 2. 355 3. 3611 4. 361 elemene cisplatin
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 8: Apoptosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Myxoma virus Poxvirus Infects rabbits
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 8: Apoptosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Myxoma virus Poxvirus Infects rabbits
Apoptosome dysfunction in human cancer
 Apoptosis 2004; 9: 691 704 C 2004 Kluwer Academic Publishers Apoptosome dysfunction in human cancer K. M. Hajra and J. R. Liu Department of Obstetrics and Gynecology, University of Michigan Medical School,
Apoptosis 2004; 9: 691 704 C 2004 Kluwer Academic Publishers Apoptosome dysfunction in human cancer K. M. Hajra and J. R. Liu Department of Obstetrics and Gynecology, University of Michigan Medical School,
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Molecular biology :- Cancer genetics lecture 11
 Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
Pro-apoptotic signalling through Toll-like receptor 3 involves TRIF-dependent
 Pro-apoptotic signalling through Toll-like receptor 3 involves TRIF-dependent activation of caspase-8 and is under the control of inhibitor of apoptosis proteins in melanoma cells Arnim Weber, Zofia Kirejczyk,
Pro-apoptotic signalling through Toll-like receptor 3 involves TRIF-dependent activation of caspase-8 and is under the control of inhibitor of apoptosis proteins in melanoma cells Arnim Weber, Zofia Kirejczyk,
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Apoptotic cell signaling in cancer progression and therapyw
 Integrative Biology Dynamic Article Links Cite this: Integr. Biol., 2011, 3, 279 296 www.rsc.org/ibiology REVIEW ARTICLE Apoptotic cell signaling in cancer progression and therapyw Jessica Plati, a Octavian
Integrative Biology Dynamic Article Links Cite this: Integr. Biol., 2011, 3, 279 296 www.rsc.org/ibiology REVIEW ARTICLE Apoptotic cell signaling in cancer progression and therapyw Jessica Plati, a Octavian
Supporting Information
 Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Mechanisms of Cell Death
 Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy August 25, 2008 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT A.
Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy August 25, 2008 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT A.
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Table S1. New colony formation 7 days after stimulation with doxo and VCR in JURKAT cells
 Table S1. New colony formation 7 days after stimulation with and in JURKAT cells drug co + number of colonies 7±14 4±7 48±11 JURKAT cells were stimulated and analyzed as in Table 1. Drug concentrations
Table S1. New colony formation 7 days after stimulation with and in JURKAT cells drug co + number of colonies 7±14 4±7 48±11 JURKAT cells were stimulated and analyzed as in Table 1. Drug concentrations
Index 255. B lymphocytes, 26 Bak, 96 basal condition preparation, See also apoptosis measurement
 Index accurate apoptosis measurement, 23. See also apoptosis measurement activated Bax staining by flow cytometry, 105 106. See also mitochondrial membrane potential active caspase-3, 15 active caspase-3
Index accurate apoptosis measurement, 23. See also apoptosis measurement activated Bax staining by flow cytometry, 105 106. See also mitochondrial membrane potential active caspase-3, 15 active caspase-3
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
The Biochemistry of apoptosis
 The Biochemistry of apoptosis 1 1 The apoptosis is composed of multiple biochemical events 2 2 Biochemical, cellular, and molecular events in Apoptosis 1. Membrane blebbing; phosphatidyl serine exposure
The Biochemistry of apoptosis 1 1 The apoptosis is composed of multiple biochemical events 2 2 Biochemical, cellular, and molecular events in Apoptosis 1. Membrane blebbing; phosphatidyl serine exposure
Supplementary figure legends
 Supplementary figure legends Supplementary Figure 1. Exposure of CRT occurs independently from the apoptosisassociated loss of the mitochondrial membrane potential (MMP). (A) HeLa cells treated with MTX
Supplementary figure legends Supplementary Figure 1. Exposure of CRT occurs independently from the apoptosisassociated loss of the mitochondrial membrane potential (MMP). (A) HeLa cells treated with MTX
Supporting Information. FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce. apoptosis
 1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
#19 Apoptosis Chapter 9. Neelu Yadav PhD
 #19 Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org Why cells decide to die? - Stress, harmful, not needed - Completed its life span Death stimulation or Stress Cell Survival Death Functions
#19 Apoptosis Chapter 9 Neelu Yadav PhD Neelu.Yadav@Roswellpark.org Why cells decide to die? - Stress, harmful, not needed - Completed its life span Death stimulation or Stress Cell Survival Death Functions
RCPA Research Award Final Progress Review
 RCPA Research Award 2010-2011 Final Progress Review Name: Dr Craig Wallington-Beddoe Degree/Institution/Year: PhD, The University of Sydney, Year 2 Research Project Title: New Therapeutic Strategies for
RCPA Research Award 2010-2011 Final Progress Review Name: Dr Craig Wallington-Beddoe Degree/Institution/Year: PhD, The University of Sydney, Year 2 Research Project Title: New Therapeutic Strategies for
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Muse Assays for Cell Analysis
 Muse Assays for Cell Analysis Multiple Assay Outputs for Cell Analysis Cell Health Cell Signalling Immunology Muse Count & Viability Kit Muse Cell Cycle Kit Muse Annexin V & Dead Cell Kit Muse Caspase
Muse Assays for Cell Analysis Multiple Assay Outputs for Cell Analysis Cell Health Cell Signalling Immunology Muse Count & Viability Kit Muse Cell Cycle Kit Muse Annexin V & Dead Cell Kit Muse Caspase
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Getting TRAIL back on track for cancer therapy
 (2014) 21, 1350 1364 & 2014 Macmillan Publishers Limited All rights reserved 1350-9047/14 www.nature.com/cdd OPEN Review Getting TRAIL back on track for cancer therapy J Lemke 1,2, S von Karstedt 1, J
(2014) 21, 1350 1364 & 2014 Macmillan Publishers Limited All rights reserved 1350-9047/14 www.nature.com/cdd OPEN Review Getting TRAIL back on track for cancer therapy J Lemke 1,2, S von Karstedt 1, J
Introduction to pathology lecture 5/ Cell injury apoptosis. Dr H Awad 2017/18
 Introduction to pathology lecture 5/ Cell injury apoptosis Dr H Awad 2017/18 Apoptosis = programmed cell death = cell suicide= individual cell death Apoptosis cell death induced by a tightly regulated
Introduction to pathology lecture 5/ Cell injury apoptosis Dr H Awad 2017/18 Apoptosis = programmed cell death = cell suicide= individual cell death Apoptosis cell death induced by a tightly regulated
Part I Molecular Cell Biology
 1 Part I Molecular Cell Biology RNA Regulation: Advances in Molecular Biology and Medicine, First Edition. Edited by Robert A. Meyers. 2014 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2014 by Wiley-VCH
1 Part I Molecular Cell Biology RNA Regulation: Advances in Molecular Biology and Medicine, First Edition. Edited by Robert A. Meyers. 2014 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2014 by Wiley-VCH
Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival
 Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
http / / cjbmb. bjmu. edu. cn Chinese Journal of Biochemistry and Molecular Biology COX-2 NTera-2 NTera-2 RT-PCR FasL caspase-8 caspase-3 PARP.
 ISSN 1007-7626 CN 11-3870 / Q http / / cjbmb bjmu edu cn Chinese Journal of Biochemistry and Molecular Biology 2012 7 28 7 630 ~ 636 NTera-2 ** ** * 410081 COX-2 NTera-2 MTT NTera-2 NTera-2 Hoechest 33258
ISSN 1007-7626 CN 11-3870 / Q http / / cjbmb bjmu edu cn Chinese Journal of Biochemistry and Molecular Biology 2012 7 28 7 630 ~ 636 NTera-2 ** ** * 410081 COX-2 NTera-2 MTT NTera-2 NTera-2 Hoechest 33258
IMMP8-1. Different Mechanisms of Androg and IPAD on Apoptosis Induction in Cervical Cancer Cells
 IMMP8-1 Different Mechanisms of Androg and IPAD on Apoptosis Induction in Cervical Cancer Cells Assanan Dokmaikaew* Tipaya Ekalaksananan** Dr.Chamsai Pientong** ABSTRACT Androg and IPAD are recently known
IMMP8-1 Different Mechanisms of Androg and IPAD on Apoptosis Induction in Cervical Cancer Cells Assanan Dokmaikaew* Tipaya Ekalaksananan** Dr.Chamsai Pientong** ABSTRACT Androg and IPAD are recently known
A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia cells
 (2004) 11, S73 S85 & 2004 Nature Publishing Group All rights reserved 1350-9047/04 $30.00 www.nature.com/cdd A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia
(2004) 11, S73 S85 & 2004 Nature Publishing Group All rights reserved 1350-9047/04 $30.00 www.nature.com/cdd A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia
Effect of FADD expression during UVC carcinogenesis
 Effect of FADD expression during UVC carcinogenesis Maja Radnić * *, Lidija Kalinić, Igor Matić, Ana Ruso, Inga Marijanović, Ivana Furčić and Biserka Nagy University of Zagreb, Faculty of Science, Department
Effect of FADD expression during UVC carcinogenesis Maja Radnić * *, Lidija Kalinić, Igor Matić, Ana Ruso, Inga Marijanović, Ivana Furčić and Biserka Nagy University of Zagreb, Faculty of Science, Department
The involvement of Bcl-2 family proteins in AKT-regulated cell survival in cisplatin resistant epithelial ovarian cancer
 /, 2017, Vol. 8, (No. 1), pp: 1354-1368 The involvement of Bcl-2 family proteins in AKT-regulated cell survival in cisplatin resistant epithelial ovarian cancer Yan Dai 1, Shiguang Jin 2,3, Xueping Li
/, 2017, Vol. 8, (No. 1), pp: 1354-1368 The involvement of Bcl-2 family proteins in AKT-regulated cell survival in cisplatin resistant epithelial ovarian cancer Yan Dai 1, Shiguang Jin 2,3, Xueping Li
Annals of RSCB Vol. XVI, Issue 1
 ENDOPLASMIC RETICULUM INVOLVEMENT IN APOPTOSIS OF NORMAL AND TREATED GINGIVAL FIBROBLASTS Ancuţa Goriuc, Raluca Jipu, Roxana Irina Iancu, M. Costuleanu GR. T. POPA UNIVERSITY OF MEDICINE AND PHARMACY,
ENDOPLASMIC RETICULUM INVOLVEMENT IN APOPTOSIS OF NORMAL AND TREATED GINGIVAL FIBROBLASTS Ancuţa Goriuc, Raluca Jipu, Roxana Irina Iancu, M. Costuleanu GR. T. POPA UNIVERSITY OF MEDICINE AND PHARMACY,
Supplementary Materials
 Supplementary Materials Figure S1. MTT Cell viability assay. To measure the cytotoxic potential of the oxidative treatment, the MTT [3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyl tetrazolium bromide] assay
Supplementary Materials Figure S1. MTT Cell viability assay. To measure the cytotoxic potential of the oxidative treatment, the MTT [3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyl tetrazolium bromide] assay
SUPPLEMENTARY INFORMATION
 Figure S1 Induction of non-apoptotic death of SV40-transformed and primary DKO MEFs, and DKO thymocytes. (A-F) STS-induced non-apoptotic death of DKO MEF. (A, B) Reduced viability of DKO MEFs after exposure
Figure S1 Induction of non-apoptotic death of SV40-transformed and primary DKO MEFs, and DKO thymocytes. (A-F) STS-induced non-apoptotic death of DKO MEF. (A, B) Reduced viability of DKO MEFs after exposure
Annals of Oncology Advance Access published January 10, 2005
 Annals of Oncology Advance Access published January 10, 2005 Original article Annals of Oncology doi:10.1093/annonc/mdi077 Expression of survivin and bax/bcl-2 in peroxisome proliferator activated receptor-g
Annals of Oncology Advance Access published January 10, 2005 Original article Annals of Oncology doi:10.1093/annonc/mdi077 Expression of survivin and bax/bcl-2 in peroxisome proliferator activated receptor-g
APPLICATION NOTE 1850 Millrace Drive, Suite 3A Eugene, Oregon
 APPLICATION NOTE 185 Millrace Drive, Suite 3A Eugene, Oregon 973 In-Cell ELISA (ICE) Assay Platform Monitoring apoptosis in cells: a high-throughput, quantitative cellbased assay. Rev. Introduction: Apoptosis:
APPLICATION NOTE 185 Millrace Drive, Suite 3A Eugene, Oregon 973 In-Cell ELISA (ICE) Assay Platform Monitoring apoptosis in cells: a high-throughput, quantitative cellbased assay. Rev. Introduction: Apoptosis:
GMS 6644: Apoptosis. Introduction
 GMS 6644: Apoptosis Introduction (Feb. 15, 2006) Lei Xiao, Ph.D. Department of Anatomy & Cell Biology UF Shands Cancer Center ARB Rm R4-250, 846-1199, lxiao@ufl.edu Outline of the Lecture Different types
GMS 6644: Apoptosis Introduction (Feb. 15, 2006) Lei Xiao, Ph.D. Department of Anatomy & Cell Biology UF Shands Cancer Center ARB Rm R4-250, 846-1199, lxiao@ufl.edu Outline of the Lecture Different types
Supplementary Figure 1
 Supplementary Figure 1 Expression of apoptosis-related genes in tumor T reg cells. (a) Identification of FOXP3 T reg cells by FACS. CD45 + cells were gated as enriched lymphoid cell populations with low-granularity.
Supplementary Figure 1 Expression of apoptosis-related genes in tumor T reg cells. (a) Identification of FOXP3 T reg cells by FACS. CD45 + cells were gated as enriched lymphoid cell populations with low-granularity.
PROFESSIONAL EXPERIENCE
 EDUCATION Doctor of Philosophy, Division of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, The Ohio State University Master of Science, Division of Organic Chemistry, Chemistry, National Taiwan
EDUCATION Doctor of Philosophy, Division of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, The Ohio State University Master of Science, Division of Organic Chemistry, Chemistry, National Taiwan
Epigonal Conditioned Media from Bonnethead Shark, Sphyrna tiburo, Induces Apoptosis in a T-Cell Leukemia Cell Line, Jurkat E6-1
 Mar. Drugs 2013, 11, 3224-3257; doi:10.3390/md11093224 Article OPEN ACCESS marine drugs ISSN 1660-3397 www.mdpi.com/journal/marinedrugs Epigonal Conditioned Media from Bonnethead Shark, Sphyrna tiburo,
Mar. Drugs 2013, 11, 3224-3257; doi:10.3390/md11093224 Article OPEN ACCESS marine drugs ISSN 1660-3397 www.mdpi.com/journal/marinedrugs Epigonal Conditioned Media from Bonnethead Shark, Sphyrna tiburo,
Imaging of glycolytic metabolism in primary glioblastoma cells with
 63 Chapter 5 Imaging of glycolytic metabolism in primary glioblastoma cells with RIMChip 5.1. Introduction Glioblastoma(GBM) is one of the most common brain tumors 1. It is composed of heterogeneous subpopulations
63 Chapter 5 Imaging of glycolytic metabolism in primary glioblastoma cells with RIMChip 5.1. Introduction Glioblastoma(GBM) is one of the most common brain tumors 1. It is composed of heterogeneous subpopulations
CAN Resubmission SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION CAN-05-0686 Resubmission Protein Immunoblotting. Primary antibodies used in this study include: Casp-9, Casp-3 and Survivin from Novus Biologicals (Littleton, CO); Casp-8 (Ab-3)
SUPPLEMENTAL INFORMATION CAN-05-0686 Resubmission Protein Immunoblotting. Primary antibodies used in this study include: Casp-9, Casp-3 and Survivin from Novus Biologicals (Littleton, CO); Casp-8 (Ab-3)
Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line
 Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line Endah Puji Septisetyani, Muthi Ikawati, Barinta Widaryanti and Edy Meiyanto* ) Cancer Chemoprevention Research Center,
Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line Endah Puji Septisetyani, Muthi Ikawati, Barinta Widaryanti and Edy Meiyanto* ) Cancer Chemoprevention Research Center,
supplementary information
 DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
Supporting Information. Non-thermal plasma with 2-deoxy-D-glucose synergistically induces cell death by targeting glycolysis in blood cancer cells
 Supporting Information Non-thermal plasma with 2-deoxy-D-glucose synergistically induces cell death by targeting glycolysis in blood cancer cells Neha Kaushik, 1 Su Jae Lee, 2 Tae Gyu Choi 3, Ku Youn Baik,
Supporting Information Non-thermal plasma with 2-deoxy-D-glucose synergistically induces cell death by targeting glycolysis in blood cancer cells Neha Kaushik, 1 Su Jae Lee, 2 Tae Gyu Choi 3, Ku Youn Baik,
Berberine Sensitizes Human Ovarian Cancer Cells to Cisplatin Through mir-93/ PTEN/Akt Signaling Pathway
 Chen Accepted: et al.: February Berberine 24, Sensitizes 2015 Ovarian Cancer Cells to Cisplatin www.karger.com/cpb 956 1421-9778/15/0363-0956$39.50/0 Original Paper This is an Open Access article licensed
Chen Accepted: et al.: February Berberine 24, Sensitizes 2015 Ovarian Cancer Cells to Cisplatin www.karger.com/cpb 956 1421-9778/15/0363-0956$39.50/0 Original Paper This is an Open Access article licensed
Samali A Figure S1.
 Deegan S, Saveljeva S, Logue SE, Pakos-Zebrucka K, Gupta S, Vandenabeele P, Bertrand MJ,Samali A. (2014) Deficiency in the mitochondrial apoptotic pathway reveals the toxic potential of autophagy under
Deegan S, Saveljeva S, Logue SE, Pakos-Zebrucka K, Gupta S, Vandenabeele P, Bertrand MJ,Samali A. (2014) Deficiency in the mitochondrial apoptotic pathway reveals the toxic potential of autophagy under
Supporting Information
 Copyright WILEY-VCH Verlag GmbH & Co. KGaA, 69469 Weinheim, Germany, 212. Supporting Information for Adv. Funct. Mater., DOI:.2/adfm.2122233 MnO Nanocrystals: A Platform for Integration of MRI and Genuine
Copyright WILEY-VCH Verlag GmbH & Co. KGaA, 69469 Weinheim, Germany, 212. Supporting Information for Adv. Funct. Mater., DOI:.2/adfm.2122233 MnO Nanocrystals: A Platform for Integration of MRI and Genuine
BL-8040: BEST-IN-CLASS CXCR4 ANTAGONIST FOR TREATMENT OF ONCOLOGICAL MALIGNANCIES. Overview and Mechanism of Action Dr.
 BL-8040: BEST-IN-CLASS CXCR4 ANTAGONIST FOR TREATMENT OF ONCOLOGICAL MALIGNANCIES Overview and Mechanism of Action Dr. Leah Klapper, CSO 88 BL-8040: Novel CXCR4 Antagonist For Hematological Cancers Indications:
BL-8040: BEST-IN-CLASS CXCR4 ANTAGONIST FOR TREATMENT OF ONCOLOGICAL MALIGNANCIES Overview and Mechanism of Action Dr. Leah Klapper, CSO 88 BL-8040: Novel CXCR4 Antagonist For Hematological Cancers Indications:
The death receptors: signaling and modulation
 The death receptors: signaling and modulation 1 1 The extrinsic cell death pathway 2 Nat Rev Drug Discov. 2008 Dec;7(12):1001-12. 2 Death receptors Belong to the tumor necrosis factor (TNF) receptor gene
The death receptors: signaling and modulation 1 1 The extrinsic cell death pathway 2 Nat Rev Drug Discov. 2008 Dec;7(12):1001-12. 2 Death receptors Belong to the tumor necrosis factor (TNF) receptor gene
Sodium selenite induces apoptosis in colon cancer cells via Bax-dependent mitochondrial pathway
 European Review for Medical and Pharmacological Sciences Sodium selenite induces apoptosis in colon cancer cells via Bax-dependent mitochondrial pathway Z. LI 1,2, J. MENG 3, T.-J. XU 4, X.-Y. QIN 5, X.-D.
European Review for Medical and Pharmacological Sciences Sodium selenite induces apoptosis in colon cancer cells via Bax-dependent mitochondrial pathway Z. LI 1,2, J. MENG 3, T.-J. XU 4, X.-Y. QIN 5, X.-D.
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
[Abstract] Objective: Investigate the effect of KLT on the expression of Fas/Apo-1, FasL and PCNA genes in renal carcinoma cell line (GRC-1).
![[Abstract] Objective: Investigate the effect of KLT on the expression of Fas/Apo-1, FasL and PCNA genes in renal carcinoma cell line (GRC-1).](/thumbs/90/104150076.jpg "[Abstract] Objective: Investigate the effect of KLT on the expression of Fas/Apo-1, FasL and PCNA genes in renal carcinoma cell line (GRC-1).") Study on Effect of Kanglaite Injection(KLT) on the Expression of Fas/Apo-1, FasL and PCNA in Renal Carcinoma Cell Lines Wang Junjie, Sun Xinchen, Sheng Wenjiang, Yu Lizhang [Abstract] Objective: Investigate
Study on Effect of Kanglaite Injection(KLT) on the Expression of Fas/Apo-1, FasL and PCNA in Renal Carcinoma Cell Lines Wang Junjie, Sun Xinchen, Sheng Wenjiang, Yu Lizhang [Abstract] Objective: Investigate
Alpna Tyagi 1, Rana P.Singh 1, Chapla Agarwal 1,2 and Rajesh Agarwal 1,2,
 Carcinogenesis vol.27 no.11 pp.2269 2280, 2006 doi:10.1093/carcin/bgl098 Advance Access publication June 15, 2006 Silibinin activates p53-caspase 2 pathway and causes caspase-mediated cleavage of Cip1/p21
Carcinogenesis vol.27 no.11 pp.2269 2280, 2006 doi:10.1093/carcin/bgl098 Advance Access publication June 15, 2006 Silibinin activates p53-caspase 2 pathway and causes caspase-mediated cleavage of Cip1/p21
mirna Dr. S Hosseini-Asl
 mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Mitochondria in apoptosis. Jean-Claude Martinou, MD, Ph.D Department of cell biology University of Geneva Geneva, Switzerland
 Mitochondria in apoptosis Jean-Claude Martinou, MD, Ph.D Department of cell biology University of Geneva Geneva, Switzerland The dual role of mitochondria in life and death QuickTime and a TIFF (Uncompressed)
Mitochondria in apoptosis Jean-Claude Martinou, MD, Ph.D Department of cell biology University of Geneva Geneva, Switzerland The dual role of mitochondria in life and death QuickTime and a TIFF (Uncompressed)
PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells
 PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells C.-G. Sun 1 *, J. Zhuang 1 *, W.-J. Teng 1, Z. Wang 2 and S.-S. Du 3 1 Department of Oncology,
PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells C.-G. Sun 1 *, J. Zhuang 1 *, W.-J. Teng 1, Z. Wang 2 and S.-S. Du 3 1 Department of Oncology,
Introduction. Cancer Biology. Tumor-suppressor genes. Proto-oncogenes. DNA stability genes. Mechanisms of carcinogenesis.
 Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Expression of TRAIL and its receptor DR5 and their significance in acute leukemia cells
 Expression of TRAIL and its receptor DR5 and their significance in acute leukemia cells S.M. Chen, H. Sun, Y.F. Liu, J. Ma, Q.T. Zhang, J. Zhu and T. Li Department of Hematology, The First Affiliated Hospital
Expression of TRAIL and its receptor DR5 and their significance in acute leukemia cells S.M. Chen, H. Sun, Y.F. Liu, J. Ma, Q.T. Zhang, J. Zhu and T. Li Department of Hematology, The First Affiliated Hospital
Problem Set 8 Key 1 of 8
 7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
CD14 + S100A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer
 CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
A redox state-dictated signalling pathway deciphers the malignant cell specificity of CD40-mediated apoptosis
 OPEN Oncogene (2017) 36, 2515 2528 www.nature.com/onc ORIGINAL ARTICLE A redox state-dictated signalling pathway deciphers the malignant cell specificity of CD40-mediated apoptosis CJ Dunnill 1, K Ibraheem
OPEN Oncogene (2017) 36, 2515 2528 www.nature.com/onc ORIGINAL ARTICLE A redox state-dictated signalling pathway deciphers the malignant cell specificity of CD40-mediated apoptosis CJ Dunnill 1, K Ibraheem
Crosstalk between Adiponectin and IGF-IR in breast cancer. Prof. Young Jin Suh Department of Surgery The Catholic University of Korea
 Crosstalk between Adiponectin and IGF-IR in breast cancer Prof. Young Jin Suh Department of Surgery The Catholic University of Korea Obesity Chronic, multifactorial disorder Hypertrophy and hyperplasia
Crosstalk between Adiponectin and IGF-IR in breast cancer Prof. Young Jin Suh Department of Surgery The Catholic University of Korea Obesity Chronic, multifactorial disorder Hypertrophy and hyperplasia
Multi-Parameter Apoptosis Assay Kit
 Multi-Parameter Apoptosis Assay Kit Catalog Number KA1335 5 x 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Multi-Parameter Apoptosis Assay Kit Catalog Number KA1335 5 x 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Deregulation of signal transduction and cell cycle in Cancer
 Deregulation of signal transduction and cell cycle in Cancer Tuangporn Suthiphongchai, Ph.D. Department of Biochemistry Faculty of Science, Mahidol University Email: tuangporn.sut@mahidol.ac.th Room Pr324
Deregulation of signal transduction and cell cycle in Cancer Tuangporn Suthiphongchai, Ph.D. Department of Biochemistry Faculty of Science, Mahidol University Email: tuangporn.sut@mahidol.ac.th Room Pr324
ONCOLOGY LETTERS 7: , 2014
 1738 Fermentation supernatants of Lactobacillus delbrueckii inhibit growth of human colon cancer cells and induce apoptosis through a caspase 3 dependent pathway YING WAN 1, YI XIN 1, CUILI ZHANG 1, DACHANG
1738 Fermentation supernatants of Lactobacillus delbrueckii inhibit growth of human colon cancer cells and induce apoptosis through a caspase 3 dependent pathway YING WAN 1, YI XIN 1, CUILI ZHANG 1, DACHANG
Introduction NEOPLASIA. Qun Liu and Yair Gazitt
 NEOPLASIA Potentiation of dexamethasone-, paclitaxel-, and Ad-p53 induced apoptosis by Bcl-2 antisense oligodeoxynucleotides in drug-resistant multiple myeloma cells Qun Liu and Yair Gazitt Overexpression
NEOPLASIA Potentiation of dexamethasone-, paclitaxel-, and Ad-p53 induced apoptosis by Bcl-2 antisense oligodeoxynucleotides in drug-resistant multiple myeloma cells Qun Liu and Yair Gazitt Overexpression
NOVEMBER 27, 2009 VOLUME 284 NUMBER 48 JOURNAL OF BIOLOGICAL CHEMISTRY 33447
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 48, pp. 33447 33455, November 27, 2009 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Caspase-9 Activation
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 48, pp. 33447 33455, November 27, 2009 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Caspase-9 Activation
Apoptosis Oncogenes. Srbová Martina
 Apoptosis Oncogenes Srbová Martina Cell Cycle Control point Cyclin B Cdk1 Cyclin D Cdk4 Cdk6 Cyclin A Cdk2 Cyclin E Cdk2 Cyclin-dependent kinase (Cdk) have to bind a cyclin to become active Regulation
Apoptosis Oncogenes Srbová Martina Cell Cycle Control point Cyclin B Cdk1 Cyclin D Cdk4 Cdk6 Cyclin A Cdk2 Cyclin E Cdk2 Cyclin-dependent kinase (Cdk) have to bind a cyclin to become active Regulation
SOD1 inhibition reduces non-small cell lung cancer by inducing cell death
 SUPPLEMENTAL INFORMATION SOD1 inhibition reduces non-small cell lung cancer by inducing cell death Andrea Glasauer, Laura A. Sena, Lauren P. Diebold, Andrew P. Mazar & Navdeep S. Chandel Inventory of Supplemental
SUPPLEMENTAL INFORMATION SOD1 inhibition reduces non-small cell lung cancer by inducing cell death Andrea Glasauer, Laura A. Sena, Lauren P. Diebold, Andrew P. Mazar & Navdeep S. Chandel Inventory of Supplemental
The discovery of Bcl-2
 The discovery of Bcl-2 Bcl-2: B-cell lymphoma 2 The pro-survival subfamily of Bcl-2 protein family Cloning of Bcl-2 as the oncogene which is deregulated at t(14;18) lymphomas Pioneer works by Tsujimoto
The discovery of Bcl-2 Bcl-2: B-cell lymphoma 2 The pro-survival subfamily of Bcl-2 protein family Cloning of Bcl-2 as the oncogene which is deregulated at t(14;18) lymphomas Pioneer works by Tsujimoto
In vivo prediction of anti-tumor effect of 3- bromopyruvate in Hepatocellular Carcinoma using Tc-99m labeled annexin-v imaging
 In vivo prediction of anti-tumor effect of 3- bromopyruvate in Hepatocellular Carcinoma using Tc-99m labeled annexin-v imaging Won Kim, Jung-Hwan Yoon*, Gi Jeong Cheon, Tae Sup Lee, Chung Yong Kim Department
In vivo prediction of anti-tumor effect of 3- bromopyruvate in Hepatocellular Carcinoma using Tc-99m labeled annexin-v imaging Won Kim, Jung-Hwan Yoon*, Gi Jeong Cheon, Tae Sup Lee, Chung Yong Kim Department
ORIGINAL ARTICLE. Hang Huang 1,2, Lin-Jin Li 3, Hai-Bo Zhang 4, An-Yang Wei 4. Summary. Introduction
 JBUON 2017; 22(1): 112-118 ISSN: 1107-0625, online ISSN: 2241-6293 www.jbuon.com E-mail: editorial_office@jbuon.com ORIGINAL ARTICLE Papaverine selectively inhibits human prostate cancer cell (PC-3) growth
JBUON 2017; 22(1): 112-118 ISSN: 1107-0625, online ISSN: 2241-6293 www.jbuon.com E-mail: editorial_office@jbuon.com ORIGINAL ARTICLE Papaverine selectively inhibits human prostate cancer cell (PC-3) growth
Apoptosis-based Therapies: Mechanisms and Applications
 Apoptosis-based Therapies: Mechanisms and Applications Perspective Bcl-2 family members, potential usage of BH3 domains as drug targets Bcl-2/xL inhibitors - Antisense, inhibitors of protein-protein interactions,
Apoptosis-based Therapies: Mechanisms and Applications Perspective Bcl-2 family members, potential usage of BH3 domains as drug targets Bcl-2/xL inhibitors - Antisense, inhibitors of protein-protein interactions,
Understanding sensitivity to BH3 mimetics: ABT-737 as a case study to foresee the complexities of personalized medicine
 Stamelos et al. Journal of Molecular Signaling 2012, 7:12 REVIEW Understanding sensitivity to BH3 mimetics: ABT-737 as a case study to foresee the complexities of personalized medicine Vasileios A Stamelos
Stamelos et al. Journal of Molecular Signaling 2012, 7:12 REVIEW Understanding sensitivity to BH3 mimetics: ABT-737 as a case study to foresee the complexities of personalized medicine Vasileios A Stamelos
Cross-Talk in Cell Death Signaling
 Commentary Cross-Talk in Cell Death Signaling By Sophie Roy and Donald W. Nicholson From the Merck Frosst Centre for Therapeutic Research, Merck Research Laboratories, Pointe Claire- Dorval, Quebec H9R
Commentary Cross-Talk in Cell Death Signaling By Sophie Roy and Donald W. Nicholson From the Merck Frosst Centre for Therapeutic Research, Merck Research Laboratories, Pointe Claire- Dorval, Quebec H9R
Key words: apoptosis, beta-amyrin, cell cycle, liver cancer, tritepenoids
 JBUON 2018; 23(4): 965-970 ISSN: 1107-0625, online ISSN: 2241-6293 www.jbuon.com E-mail: editorial_office@jbuon.com ORIGINAL ARTICLE Antitumor effects of beta-amyrin in Hep-G2 liver carcinoma cells are
JBUON 2018; 23(4): 965-970 ISSN: 1107-0625, online ISSN: 2241-6293 www.jbuon.com E-mail: editorial_office@jbuon.com ORIGINAL ARTICLE Antitumor effects of beta-amyrin in Hep-G2 liver carcinoma cells are
ASCO Annual Meeting 2013, May 31 June , Chicago, IL
 Phase I Study of Safety and Pharmacokinetics of GDC-0917, an Antagonist of Inhibitor of Apoptosis Proteins in Patients with Refractory Solid Tumors or Lymphoma 1 A Tolcher, 1 K Papadopoulos, 1 A Patnaik,
Phase I Study of Safety and Pharmacokinetics of GDC-0917, an Antagonist of Inhibitor of Apoptosis Proteins in Patients with Refractory Solid Tumors or Lymphoma 1 A Tolcher, 1 K Papadopoulos, 1 A Patnaik,
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary