Highly Unsaturated Fatty Acid Synthesis in Marine Fish: Cloning, Functional Characterization and Nutritional. (Gadus morhua L.)
|
|
|
- Loren James
- 5 years ago
- Views:
Transcription
1 Highly Unsaturated Fatty Acid Synthesis in Marine Fish: Cloning, Functional Characterization and Nutritional Regulation of Fatty Acyl Δ6 Desaturase of Atlantic Cod (Gadus morhua L.) Douglas R. Tocher *, Xiaozhong Zheng, Christian Schlechtriem, Nicola Hastings 1, James R. Dick and Alan J. Teale Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, Scotland, United Kingdom Running Title: Δ6 DESATURASE IN ATLANTIC COD Keywords: Highly unsaturated fatty acids; Biosynthesis; 6 Desaturase; Gene expression, Nutritional regulation; Marine fish; Cod. Mailing address: Dr Douglas R Tocher. Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, Scotland, United Kingdom. Tel: ; Fax ; d.r.tocher@stir.ac.uk 1
2 *To whom correspondence should be addressed. Dr Douglas R Tocher. Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, Scotland, United Kingdom. Tel: ; Fax ; d.r.tocher@stir.ac.uk Abbreviations: FO, fish oil; HUFA, highly unsaturated fatty acids (carbon chain length C 20 with 3 double bonds); ORF, open reading frame; Q-PCR, quantitative real-time polymerase chain reaction; RACE, rapid amplification of cdna ends; UTR, untranslated region; VO, vegetable oil. 1 Present address: Roslin Institute, Roslin, Midlothian EH25 9LS, Scotland, United Kingdom 2
3 ABSTRACT: This study reports the cloning, functional characterization, tissue expression and nutritional regulation of a 6 fatty acyl desaturase of Atlantic cod (Gadus morhua). PCR primers were designed based on the sequences of conserved motifs in available fish desaturases and used to isolate a cdna fragment from cod liver, with full-length cdna obtained by Rapid Amplification of cdna Ends. The cdna for the putative desaturase was shown to comprise 1980bp, including a 261bp 5 -UTR, a 375bp 3 -UTR, and an ORF of 1344 bp that specified a protein of 447 amino acids. The protein sequence included three histidine boxes, two transmembrane regions, and an N- terminal cytochrome b 5 domain containing the haem-binding motif HPGG, all characteristics of microsomal fatty acyl desaturases. The cdna displayed Δ6 desaturase activity in a yeast expression system. Quantitative real time PCR assay of gene expression in cod showed that the 6 desaturase gene was highly expressed in brain, followed by liver, kidney, intestine, red muscle and gill, and at much lower levels in white muscle, spleen and heart. The expression of the Δ6 desaturase gene did not appear to be under significant nutritional regulation, with levels in liver and intestine being barely altered in fish fed a vegetable oil blend by comparison with levels in fish fed fish oil. This was reflected in enzyme activity, as hepatocytes or enterocytes showed very little highly unsaturated fatty acid biosynthesis activity irrespective of diet. Further studies are required to determine why the Δ6 desaturase appears to be barely functional in cod under the conditions tested. 3
4 Fish are the only major dietary source for humans of n-3 highly unsaturated fatty acids (HUFA), eicosapentaenoate (EPA; 20:5n-3) and docosahexaenoate (DHA; 22:6n-3) (1) and, with traditional fisheries declining, aquaculture supplies an increasing proportion of the fish in the human food basket (2). However, the current high use of fish oils, derived from feed-grade marine fisheries, in aquaculture feeds is not sustainable, and will constrain growth of aquaculture activities (3,4). Vegetable oils, a sustainable alternative to fish oil, can be rich in C 18 polyunsaturated fatty acids (PUFA) such as linoleic (18:2n-6) and α-linolenic (18:3n-3) acids, but lack the n-3hufa abundant in fish oils (5). The extent to which fish can convert C 18 PUFA to C 20/22 HUFA varies with species, and is associated with their capacity for fatty acyl desaturation and elongation (6). Marine fish are unable to produce EPA and DHA from 18:3n-3 at a physiologically significant rate (5) due to apparent deficiencies in one or more enzymes in the desaturation/elongation pathway (7,8). Thus, flesh fatty acid compositions in marine fish fed vegetable oils are characterized by increased levels of C 18 PUFA and decreased levels of n-3hufa, compromising their nutritional value to the human consumer (9,10). Until depletion of the commercial stocks in the 1990 s, Atlantic cod (Gadus morhua L.) was the most valued food fish obtained from the North Atlantic (11). In recent years significant progress has been made in the culture of cod and the life cycle has been closed, allowing production independently of wild fisheries (12). The large and highly developed market for cod along with the high market price and the decreasing quotas set to preserve wild stocks has greatly increased interest in cod culture in recent years (13-15). However, the establishment of large-scale, sustainable cod culture will require solutions to several nutritional issues including broodstock and larval nutrition and replacement of dietary fish oil with alternatives in ongrowing diets (16-18). Our overall aim is to determine what regulates HUFA biosynthesis in fish and how it can be optimized to enable fish to make effective use of dietary vegetable oil. Recently, fatty acyl desaturases, critical enzymes in the pathways for the biosynthesis of the long-chain C 20/22 HUFA from shorter chain C 18 PUFA, have been cloned from several teleosts (19,20). The cdnas for Δ6 4
5 and Δ5 desaturases have been cloned from Atlantic salmon (Salmo salar L.) (21,22) whereas, in contrast, only Δ6 desaturase cdnas have been cloned from marine fish (23,24). The expression levels of both Δ6 and Δ5 desaturases in salmon were shown to be up regulated in fish fed vegetable oil by comparison with levels in fish fed fish oil (25,26). The specific objectives of the study described here were to investigate fatty acid desaturation and the regulation of the HUFA synthetic pathway in Atlantic cod. Thus, we describe the cdna cloning, functional characterization, and tissue distribution of a fatty acyl desaturase of cod, and report the effects of nutrition on the expression of fatty acyl desaturase and elongase genes in cod fed diets containing either fish oil or a blend of vegetable oils. MATERIALS AND METHODS Cloning of putative fatty acyl desaturase from cod. Liver tissue was obtained from Atlantic cod fed for 3 months with the vegetable oil diet described below. Total RNA was extracted from liver using TRIzol reagent (GibcoBRL, NY, U.S.A.). 3 -RACE cdna was synthesized using MMLV reverse transcriptase (Promega, Madison, WI, U.S.A) primed by the oligonucleotide, Not1polyT, 5 - GATAGCGCCCGCGTTTTTTTTTTTTTTTTV RACE-cDNA was synthesized using the SMART RACE cdna amplification kit (Clontech, NJ, U.S.A). Available fish desaturase sequences were aligned to enable the design of the degenerate forward primer, 5 - CARCAYCAYGCNAARCCNAA-3 and reverse primer, 5 -RAANARRTGYTCDATYTG-3. These were used for PCR isolation of a cod desaturase cdna fragment. PCR amplification was performed using Thermoprime plus DNA polymerase (ABgene, Surrey, UK) under the following touchdown PCR conditions: initial denaturation at 95 o C for 1 min, 10 cycles of denaturation at 95 o C for 15 s, annealing at 62 o C for 30 s (-1.5 o C /cycle), and extension at 72 o C for 90 s, then 6 cycles of denaturation at 95 o C for 15 s, annealing at 52 o C for 30 s and extension at 72 o C for 90 s, followed by 24 cycles of denaturation at 95 o C for 15 s, annealing at 51 o C for 30 s and extension at 72 o C for 90 s. The PCR products were cloned into pbluescript KS II + vector (Stratagene, La Jolla, 5
6 CA, U.S.A.). The nucleotide sequences were determined by standard dye terminator chemistry using a Perkin Elmer ABI-377 DNA sequencer following the manufacturer s protocols (Perkin Elmer, Applied Biosystems). Following isolation of a cdna fragment flanked by the above primers, the specific forward primer 5 -GCCGATGAACATAGACCACG-3 was designed for 3 RACE PCR together with primer Not1PolyT under the following conditions: initial denaturation at 95 o C for 1 min, 25 cycles of denaturation at 95 o C for 20 s, annealing at 56 o C for 30 s, and extension at 72 o C for 2 min. The reverse primer, 5 -GGTGTTGGTGGTGATAGGGCAT-3, was used in conjunction with forward UPM (universal primer mix, long and short), 5' CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT 3' and 5 - CTAATACGACTCACTATAGGGC-3, to perform 5 RACE PCR. Amplification involved an initial step at 95 o C for 1 min and 70 o C for 3 min, and 5 cycles of denaturation at 95 o C for 30s, annealing at 60 o C (-1 o C /cycle) for 30s and extension at 72 o C for 2 min, followed by 24 cycles of denaturation at 95 o C for 30s, annealing at 56 o C for 30s and extension at 72 o C for 2 min. Nucleotide sequences were determined as above and the 3 and 5 RACE PCR fragment sequences aligned to assemble the full nucleotide sequence of the cod putative desaturase cdna using BioEdit version (Tom Hall, Department of Microbiology, North Carolina State University, USA). Heterologous expression of desaturase ORF in Saccharomyces cerevisiae. Expression primers were designed for PCR cloning of the cod putative desaturase cdna ORF. The forward primer, 5 - CGGAATTCAAGCTTAAGATGGGAGGTGGAGGGCA-3 contained a HindIII site (underlined) and the reverse primer, 5 -GCTCTAGACTCGAGTCACTTATGGAGATAAGCATC-3 contained an XhoI site (underlined). PCR was performed using high fidelity DNA polymerase (Roche Diagnostics Ltd., Lewes, East Sussex, UK) following the manufacturer s instructions. Amplification involved an initial denaturation step at 95 o C for 2 min, followed by 30 cycles of denaturation at 95 o C for 30 s, annealing at 58 o C for 30 s, and extension at 72 o C for 90 s followed by a final extension at 72 o C for 10 min. Following PCR, the DNA fragments were restricted and 6
7 ligated into the similarly digested yeast expression vector pyes2 (Invitrogen Ltd, Paisley, UK). Ligation products were then used to transform Top10F E. coli competent cells (Invitrogen Ltd, Paisley, UK) which were screened for the presence of recombinants. Transformation of the yeast S. cerevisiae (strain InvSc1) with the recombinant plasmids was carried out using the S.c.EasyComp Transformation Kit (Invitrogen Ltd, Paisley, UK). Selection of yeast containing the desaturase/pyes2 constructs was on S. cerevisiae minimal medium (SCMM) minus uracil. Culture of the recombinant yeast was carried out in SCMM -uracil broth as described previously (19), using galactose induction of gene expression. Each culture was supplemented with one of the following PUFA substrates; α-linolenic acid (18:3n-3), linoleic acid (18:2n-6), eicosatetraenoic acid (20:4n- 3), dihomo-γ-linolenic acid (20:3n-6), docosapentaenoic acid (22:5n-3) and docosatetraenoic acid (22:4n-6). PUFA were added to the yeast cultures at concentrations of 0.5 mm (C 18 ), 0.75 mm (C 20 ) and 1 mm (C 22 ) as uptake efficiency decreases with increasing chain length. Yeast cells were harvested, washed, dried, and lipid extracted by homogenization in chloroform/methanol (2:1, by vol.) containing 0.01% butylated hydroxytoluene (BHT) as antioxidant as described previously (19). Fatty acid methyl esters (FAME) were prepared, extracted, purified by thin layer chromatography (TLC), and analyzed by gas chromatography (GC), all as described below. The proportion of substrate fatty acid converted to the longer chain fatty acid product was calculated from the gas chromatograms as 100 [product area/(product area + substrate area)]. Unequivocal confirmation of fatty acid products was obtained by GC-mass spectrometry of the picolinyl derivatives as described in detail previously (19). Fish and diets. Atlantic cod were from a stock held at the Institute of Aquaculture and were fed a commercial fishmeal and oil-based diet prior to the trial. Fish with a mean body weight of 87 ± 5 g, were stocked randomly (at 20 fish/tank) into two 1.5 m tanks of 400-L capacity supplied with recirculated seawater (37 ppt) at a constant temperature of 10 o C at 400 L/h and were subjected to a photoperiod regime of 12-h light:12-h dark. Fish were fed at a rate of 1.5% biomass/day in two portions (9 am and 2 pm) for three months before sampling. The diets were manufactured in-house 7
8 and only differed in the oil added to the basal mix (Table 1). In one diet, fish oil (FO) was the only added oil and in the other fish oil was replaced by a vegetable oil (VO) blend containing rapeseed, linseed and palm oils in a 2 : 2 : 1 ratio (Table 2). The diets were formulated to meet all the known nutritional requirements of marine fish (27). The experiment was conducted in accordance with British Home Office guidelines regarding research on experimental animals. After 90 days, the fish weighed 258 ± 21g and there was no significant difference between weights of fish on different dietary treatments. Twelve fish per dietary treatment were sampled, with six used for the preparation of hepatocytes and enterocytes from liver and pyloric caeca, respectively. Tissue samples were collected from the other six fish, with liver and caecal tissue collected from all six fish, and brain, heart, kidney, gill, spleen, adipose tissue, white and red muscle collected from three of these fish for RNA extraction and gene expression studies, and liver, caeca and white muscle collected from four fish for lipid and fatty acid analyses. All tissue samples were frozen immediately in liquid nitrogen and stored at 80 o C prior to further analyses. Cod tissue RNA extraction and quantitative real time PCR (Q-PCR). Total RNA extraction was performed as described above. Five µg of total RNA was reverse transcribed into cdna using M-MLV reverse transcriptase first strand cdna synthesis kit (Promega UK, Southampton, UK). Gene expression of the fatty acyl 6 desaturase and fatty acid elongase (28) genes in tissues from cod fed the different diets was studied by quantitative real-time PCR (Q-PCR). The PCR primers were designed according to the 6 desaturase (accession no. DQ054840), and published elongase (accession no. AY660881) cdna sequences. For the 6 desaturase, the forward and reverse primers were 5 -CCCCAGACGTTTGTGTCAG-3, and 5 -CCTGGATTGTTGCTTTGGAT-3, respectively. For the elongase, the forward and reverse primers were 5 - TGATTTGTGTTCCAAATGGC-3 and 5 -CTCATGACGGGAACCTCAAT-3, respectively. PCR products sizes were 181 and 219 bp, respectively. Amplification of cdna samples and DNA standards was carried out using SYBR Green PCR Kit (Qiagen, Crowley, West Sussex, UK) and the following conditions: 15 min denaturation at 95 o C, 45 cycles of 15 s at 94 o C, 15 s at 55 o C and 8
9 30 s at 72 o C. This was followed by product melt to confirm single PCR products. Thermal cycling and fluorescence detection were conducted in a Rotor-Gene 3000 system (Corbett Research, Cambridge, UK), with expression of target genes quantified and normalized as described previously (29). Lipid extraction and fatty acid analyses. Total lipids of livers, pyloric caeca, flesh (white muscle) and diet samples were extracted by homogenization in chloroform/methanol (2:1, v/v) containing 0.01 % BHT as antioxidant (30). FAME were prepared from total lipid by acid-catalyzed transesterification using 2 ml of 1% H 2 SO 4 in methanol plus 1 ml toluene (31) and FAME extracted and purified as described previously (32). FAME were separated and quantified by gas-liquid chromatography (Fisons GC8600, Fisons Ltd., Crawley, U.K.) using a 30 m x 0.32 mm capillary column (CP wax 52CB; Chrompak Ltd., London, U.K). Hydrogen was used as carrier gas and temperature programming was from 50 o C to 180 o C at 40 o C/min and then to 225 o C at 2 o C/min. Individual methyl esters were identified by comparison to known standards and by reference to published data (33). Activity of the HUFA biosynthesis pathway in hepatocytes and enterocytes. Hepatocytes and enterocytes were prepared from livers and pyloric caeca by collagenase treatment of chopped tissue and sieving through 100 µm nylon gauze essentially as described in detail previously for salmonid and sea bass tissues (26,34). Approximately twice as much cod liver tissue was used for digestion due to the high fat content and consequently lower yield of cells (35). Viability, as assessed by trypan blue exclusion, was > 95% at isolation and decreased by less than 5% over the period of the incubation for both cell types. The enriched enterocyte preparation was predominantly enterocytes although some secretory cells are also present. One hundred µl of the hepatocyte and enterocyte suspensions were retained for protein determination according to the method of Lowry et al. (36) following incubation with 0.4 ml of 0.25% (w/v) SDS/1M NaOH for 45 min at 60 o C. Five ml of each hepatocyte or caecal enterocyte suspension were dispensed into 25 cm 2 tissue culture flasks and incubated at 20 o C for 2 h with 0.3 µci (~ 1 µm) [1-14 C]18:3n-3, added as a BSA 9
10 complex (37). After incubation, cells were harvested, washed, and lipid extracted as described previously (26), and total lipid transmethylated and FAME prepared as described above. The FAME were separated by argentation (silver nitrate) thin-layer chromatography (38), located on the plate by autoradiography for 14 days, and quantified by liquid scintillation after being scraped from the plates as described previously (39). Materials. [1-14 C]18:3n-3 (50-55 mci/mmol) was obtained from NEN (Perkin Elmer LAS (UK) Ltd., Beaconsfield, U.K.). Eicosatetraenoic (20:4n-3), docosapentaenoic (22:5n-3) and docosatetraenoic (22:4n-6) acids (all > 98-99% pure) were purchased from Cayman Chemical Co., Ann Arbor, USA. Linoleic (18:2n-6), α-linolenic (18:3n-3), eicosatrienoic (20:3n-6) acids (all >99% pure), collagenase (type IV), FAF-BSA, BHT and silver nitrate were obtained from Sigma- Aldrich Co. Ltd. (Poole, U.K.). TLC (20 x 20 cm x 0.25 mm) plates pre-coated with silica gel 60 (without fluorescent indicator) were purchased from Merck (Darmstadt, Germany). All solvents were HPLC grade and were from Fisher Scientific (Loughborough, UK). RESULTS Sequence analyses. The full length of the putative cod desaturase cdna (mrna), as determined by 5 and 3 RACE PCR, was shown to be 1980bp which included a 5 -UTR of 261bp and a 3 - UTR of 375bp. Sequencing revealed that the cdna included an ORF of 1344 bp, which specified a protein of 447 amino acids (GenBank accession no. DQ054840). The protein sequence included all the characteristic features of microsomal fatty acyl desaturases, including two transmembrane regions, three histidine boxes and an N-terminal cytochrome b 5 domain containing the haembinding motif, H-P-G-G (Fig.1). Phylogenetic analysis comparing the cod desaturase sequence with desaturases from other fish species and human Δ5 and Δ6 desaturases, clustered the cod most closely with tilapia, sea bream and turbot, and more distantly from carp, zebrafish, and salmonids (Fig.2). A pair-wise comparison between fish and human desaturase sequences showed the amino acid sequence predicted by the cod putative desaturase ORF showed greatest identity to that of 10
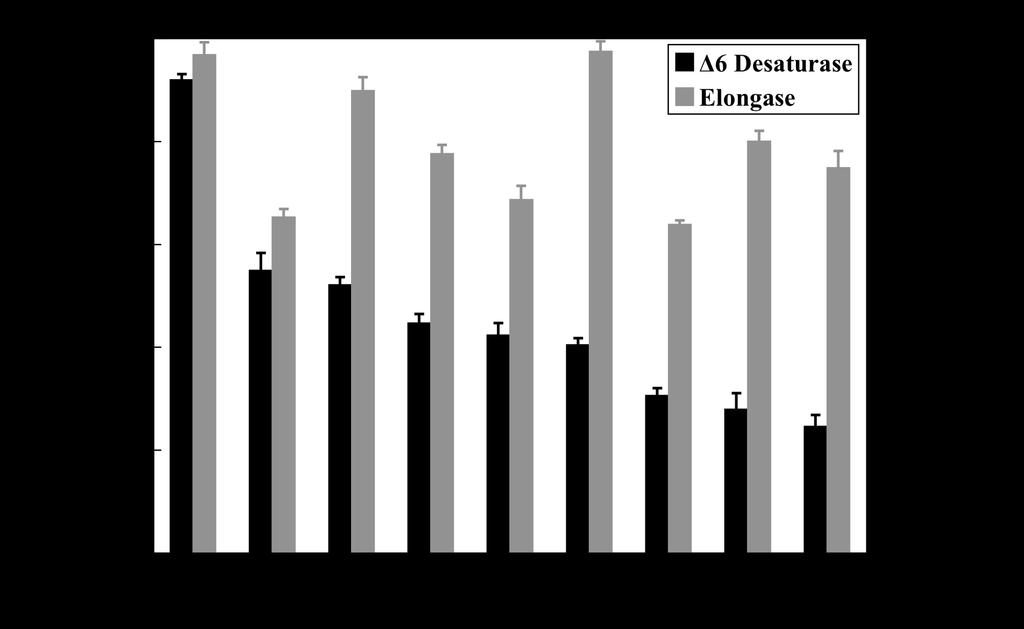
11 gilthead sea bream 6 desaturase (82%), and 64 and 56% identity to the human 6 and 5 cdnas, respectively (Table 3). Functional characterization. The fatty acid composition of the yeast transformed with the vector alone showed the four main fatty acids normally found in S. cerevisiae, namely 16:0, 16:1n-7, 18:0 and 18:1n-9, together with the exogenously derived fatty acids (Fig. 3A and C). The most prominent additional peaks were observed in the profiles of transformed yeast grown in the presence of the 6 desaturase substrates, 18:2n-6 and 18:3n-3 (Fig.3A-D). Based on GC retention time and confirmed by GC-MS, the additional peaks associated with the presence of the cod desaturase cdna were identified as 18:3n-6 (Fig.3B) and 18:4n-3 (Fig.3D), corresponding to the 6 desaturation products of 18:2n-6 and 18:3n-3, respectively. Approximately, 33.5% of 18:3n-3 was converted to 18:4n-3, and 17.5% of 18:2n-6 was converted to 18:3n-6 in yeast transformed with the cod desaturase. No additional peaks representing desaturated fatty acid products were observed in the lipids of transformed S. cerevisiae incubated with 20:3n-6 or 20:4n-3 (Fig.3E and F), indicating no 5 desaturase activity. Similarly, the cod desaturase cdna did not express any 4 desaturase activity as evidenced by the lack of additional peaks representing desaturated products of 22:5n-3 or 22:4n-6 (data not shown). Fatty acyl desaturase and elongase gene expression in cod tissues. The fatty acyl Δ6 desaturase and PUFA elongase genes were expressed in all cod tissues examined. For the desaturase, the abundance of transcript was clearly greatest in brain followed by liver, kidney, intestine, red muscle and gill, and by much lower levels in white muscle, spleen and heart (Fig.4). The abundance of elongase transcript was very high in brain and gill, with lower levels in kidney, spleen, intestine and heart, and a surprisingly low level of expression in liver. Effect of diet on fatty acid compositions of flesh, liver and pyloric caeca. The FO diet contained around 25% total saturates, predominantly 16:0, approximately 46% total monoenes, with 26% as the long chain monoenes 20:1 and 22:1, 3.7% n-6pufa predominantly 18:2n-6, and almost 25% n- 3PUFA, with almost 20% as the n-3hufa, 20:5n-3 and 22:6n-3 (Table 2). The VO diet provided 11
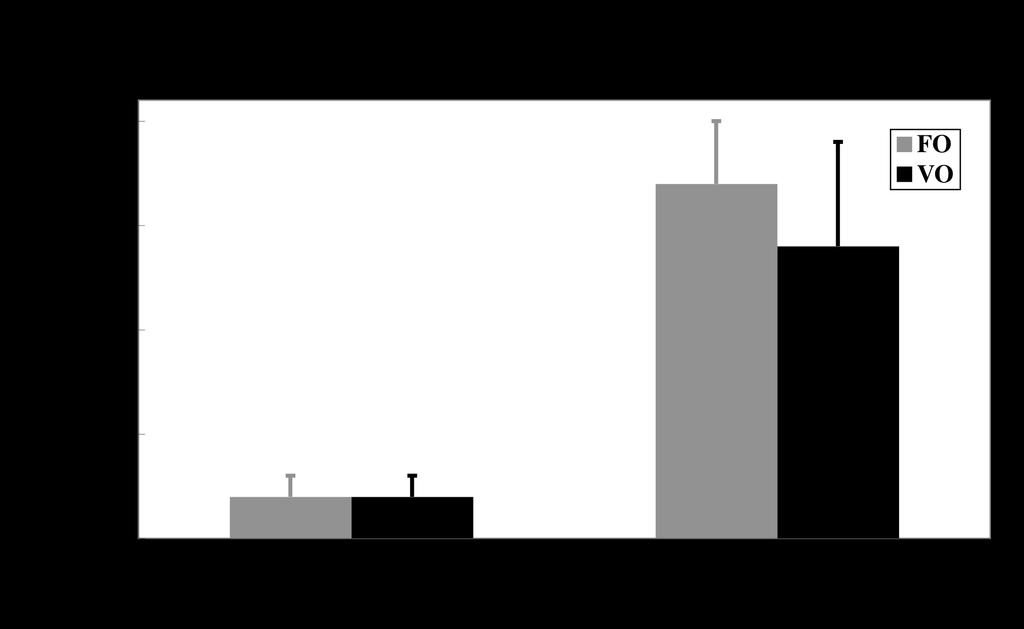
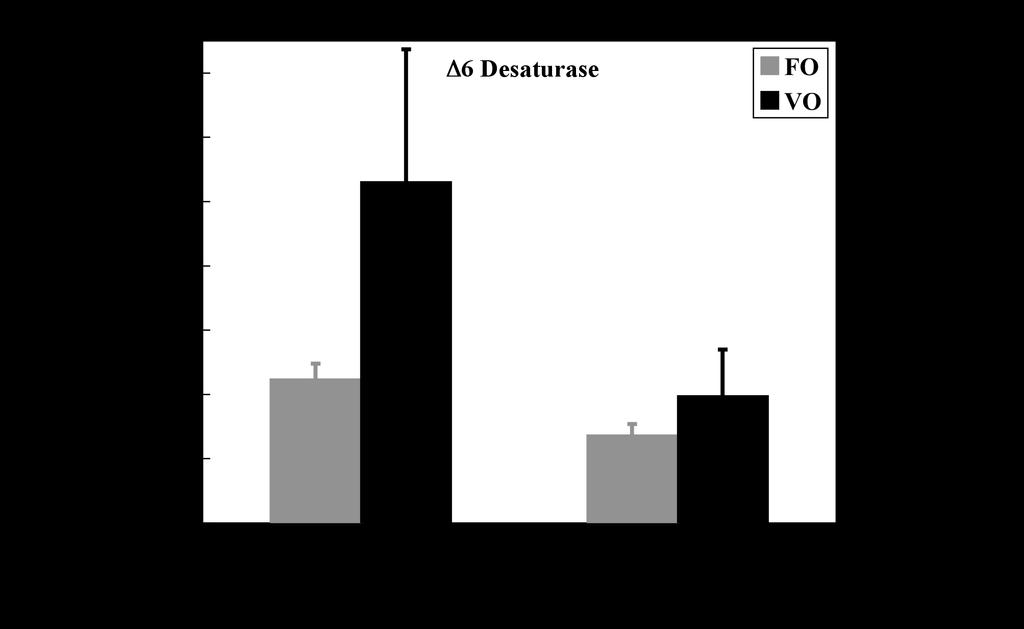
12 slightly lower levels of total saturates and monoenes and a similar level of n-3pufa, but was characterized by increased proportions of 18:1n-9, 18:2n-6 and 18:3n-3 and decreased proportions of n-3hufa and long chain monoenes. Therefore, the VO diet showed levels of 18:1n-9, 18:2n-6 and 18:3n-3 of around 26%, 10% and 13%, respectively, whereas the combined level of 20:5n-3 and 22:6n-3 was reduced by half to 10% (Table 2). The total lipid content of liver was greater than 50% of the wet weight consistent with it being the major lipid storage organ in cod, whereas flesh had a low lipid content of 1% with pyloric caeca containing around 3% total lipid (Table 4). Dietary VO had no significant effect on tissue lipid contents. The rank order of the major fatty acids from total lipid of flesh from cod fed FO was 22:6n-3 > 16:0 > 20:5n-3 > 18:1n-9 > 20:1n-9 (Table 4). The changes in fatty acid composition of the diets in response to replacement of FO with VO, described above, were reflected in the flesh fatty acid compositions with increased proportions of 18:1n-9, 18:2n-6 and 18:3n-3, and decreased proportions of n-3hufa. Pyloric caeca showed a similar fatty acid composition to flesh with dietary VO having a very similar effect. However, consistent with the higher storage (neutral) lipid content, liver fatty acid composition was characterized by higher levels of monounsaturated fatty acids. Thus, the rank order of the major fatty acids from total lipid of liver from fish fed FO was 18:1n-9 > 16:0 > 20:1n-9 > 22:6n-3 > 22:1 > 20:5n-3 > 16:1n-7 (Table 4). However, substitution of FO with VO resulted in similar changes to liver fatty acid composition as in the other tissues, with increased proportions of 18:1n-9, 18:2n-6 and 18:3n-3 and decreased proportions of n-3hufa. Effects of diet on the HUFA biosynthesis pathway in hepatocytes and enterocytes. Total activity of the HUFA biosynthesis pathway as measured by the recovery of radioactivity in the summed desaturated products of [1-14 C]18:3n-3 was around 7-fold higher in enterocytes than in hepatocytes in cod (Fig.5). In both cell types, the rate of synthesis of pathway products was unaffected by dietary treatment. The rate of the HUFA biosynthesis pathway in cod hepatocytes was between 40- and 120-fold lower than that observed in hepatocytes of Atlantic salmon smolts of similar size and assayed at the same time of year and fed similar diets (26) (Fig.6). Similarly, the rate of the 12
13 pathway in cod enterocytes was between 8- and 17-fold lower than that observed in salmon enterocytes. Furthermore, in contrast to the situation in cod, the rate of the HUFA biosynthesis pathway in salmon was significantly higher in both cell types from fish fed a VO blend compared to fish fed FO. Effect of diet on expression of fatty acyl desaturase and elongase genes in liver and intestine. The effect of diet on the normalised expression of the Δ6 desaturase and PUFA elongase genes was determined in liver and intestine (pyloric caeca). The expression of both genes tended to be increased in terms of absolute copy number but the data were characterized by a high degree of variation. Therefore, the only statistically significant effect of diet was increased expression of PUFA elongase in the liver (Fig.7). DISCUSSSION The study reported here revealed that Atlantic cod express a fatty acyl desaturase and functional analysis in yeast confirmed the cod enzyme as a Δ6 desaturase. Comparing the protein sequence with that of a range of other desaturases of fish and human showed the cod Δ6 sequence to be more similar to the human Δ6 than to the human Δ5, but most similar to the Δ6 desaturases previously cloned from other marine fish, gilthead sea bream (Sparus aurata) and turbot (Psetta maximus) (20,22). Phylogenetic analysis of the desaturase sequences reflected classical phylogeny, showing the cod (Paracanthopterygii; Gadiformes) branching from the Acanthopterygia (cichlids, perciformes and pleuronectiformes) line, and further separated from both the carp and zebrafish (Ostariophysi; cyprinids), and salmonids (Salmoniformes; salmonidae) (42). Along with the cloning of Δ6 desaturase cdnas of sea bream and turbot, the work described has confirmed that marine fish have, and express, the gene required for the first step in the HUFA biosynthesis pathway, Δ6 desaturation, suggesting that any deficiencies in this pathway in marine fish would be at a further step such as chain elongation and/or Δ5 desaturation. This is consistent with biochemical data suggesting deficiencies in these steps in turbot and sea bream cell lines (7,8). 13
14 However, despite expressing an apparently active Δ6 desaturase, the activity of the HUFA biosynthesis pathway in both hepatocytes and enterocytes in cod was very low, and considerably lower than the activities measured in salmon hepatocytes and enterocytes (26,43). Indeed, the activities were so low that it was not possible to accurately quantify individual products, although in enterocytes, in which the activity was slightly higher, we were able to confirm that only Δ6 desaturated products (18:4n-3, 20:4n-3 and 22:4n-3) were observed on the autoradiographs. Furthermore, the majority product was 18:4, and so very little HUFA (defined as C 20 and 3 double bonds) was actually produced, and neither EPA nor DHA were detected in cod. In contrast, functional expression in the yeast system had shown that the cod Δ6 had substantial enzymatic activity towards 18:3n-3. This perhaps highlights the known limitations of heterologous expression systems that will be using the endogenous yeast transcription system rather than the cod transcription machinery. It is also clear that it is essential that gene expression at the protein level should also be studied to confirm translation of the Δ6 desaturase gene in cod tissues. However, in the same yeast system, the salmon Δ6 gene gave over 60% conversion of 18:3n-3 (22) compared to 33% for the cod Δ6. Therefore, consistent with the salmon expressing higher HUFA synthesis activities, the salmon Δ6 was more active than the cod Δ6 in a comparative system although this comparison is limited by the considerations mentioned above. Conversely, conversion of 18:2n-6 by the cod Δ6 exceeded that of the salmon Δ6 (18% vs. 14%) in the yeast expression system (22). This was interesting as it had previously been shown that the cod PUFA elongase was unique among the fish desaturase and elongase genes functionally characterized so far by being more active towards the n-6 substrate than the equivalent n-3 substrates (28). The reason for the cod genes being relatively more active towards n-6 substrates is unclear. In addition to being considerably lower than in salmon, the rate of the HUFA biosynthesis pathway observed in cod was even lower than that observed in previous studies in other marine fish. The rate of desaturation of 14 C-18:3n-3 to all products in sea bass (Dicentrarchus labrax) was around 0.2 pmol.h -1.mg protein -1 in hepatocytes and between 0.7 and 1.1 pmol.h -1.mg protein -1 in 14
15 caecal enterocytes, but this latter figure still represented only 2.1% of total radioactivity recovered, with 97.9% of total radioactivity recovered as the substrate [1-14 C]18:3n-3 (34). The activities in cod were around 10-fold lower at 0.02 and 0.15 pmol.h -1.mg protein -1 in hepatocytes and enterocytes, respectively. It was perhaps noteworthy that the HUFA biosynthesis activities were higher in enterocytes than in hepatocytes in both sea bass and cod. This may be a real biological difference as intestine is now acknowledged as a site of significant fatty acid metabolism, at least in salmonids (44). However, it is possible that part of that difference may be due to the practical problems in isolating cells from such a lipid-rich tissue as cod liver. Despite using twice as much liver as caeca (in terms of wet weight), the cell yield of hepatocytes was very low and may have been a contributing factor. Problems of low yield from lipid rich livers and also analyzing autoradiography data from very low activity samples (see above) were encountered in the previous study in sea bass (34). Expression of the cod Δ6 desaturase was greatest in brain followed by liver, kidney and intestine, whereas expression of the PUFA elongase was very high in brain and gill with a low level of expression in liver. In salmon, the expression of both Δ6 and Δ5 desaturases and the PUFA elongase were highest in intestine, liver and brain (22). Mammalian Δ6 and Δ5 desaturases also show relatively high expression in liver and brain, as well as heart and kidney (45,46). A third desaturase gene in humans (FADS3) also shows highest expression in brain, heart and liver (47). In contrast, intestine does not appear to be a site of high desaturase expression in mammals (48). In mouse, a PUFA elongase was expressed to the greatest extent in testis and liver followed by brain and kidney (49). In rat, two elongases were isolated with one, a PUFA elongase (relo1), being expressed highly in lung and brain whereas the other, responsible for elongating saturated fatty acids (relo2), was only expressed in liver (50). The tissue expression analysis in cod also showed that expression of PUFA elongase exceeded Δ6 desaturase expression. This was in contrast with the situation in salmon where both Δ6 and Δ5 desaturase expression exceeded expression of PUFA elongase in all tissues (22). This may be real biological difference between cod and salmon, but 15
16 there may be other contributing factors. For example, in salmon, there are at least two other desaturase gene sequences in the genome (Zheng, unpublished data). The entire DNA gene sequences for these two desaturases are not known, and the cdnas have not been isolated, and so the fatty acid specificities, tissue distributions, and hence possible functions, if any, of these desaturases are not known. The function of the third (FADS3) desaturase gene in humans is also unknown at the present time (47). Mammalian desaturase genes have been demonstrated to be subject to nutritional regulation. The expression of Δ6 desaturase in liver was increased in mice fed triolein (18:1n-9), an EFA-deficient diet, compared to mice fed corn oil, a diet rich in 18:2n-6 (45). Similarly, the expression of both Δ6 and Δ5 desaturases was 4-fold higher in rats fed a fat-free diet or a diet containing triolein compared to that in rats fed either safflower oil (18:2n-6) or menhaden oil (n-3hufa) (46). Similar results have been obtained in salmonids with dietary linseed oil (rich in 18:3n-3) increasing the expression of liver Δ6 desaturase in rainbow trout, and liver Δ5 desaturase and elongase in Atlantic salmon, compared to levels in fish fed fish oil (20,25). In recent studies, expression levels of both Δ6 and Δ5 desaturases were increased in liver of salmon fed a vegetable oil blend (rich in C 18 PUFA) compared to levels in fish fed fish oil, but elongase expression was not increased (22,26). In the present study, the expression of Δ6 desaturase in cod liver and intestine was not significantly increased in fish fed vegetable oil compared to that in fish fed fish oil, although expression of the PUFA elongase was increased in liver by dietary vegetable oil. The expression of the genes was generally reflected in the activity of the HUFA biosynthesis pathway and in the fatty acid compositions of the tissues which showed no evidence of any Δ6 desaturase or PUFA elongase activity, with 18:4n-3 and 20:4n-3 levels not maintained, and 18:3n-6 levels only very low and 20:3n-6 not observed at all, in fish fed vegetable oil. Previously, increased desaturase expression was reflected in higher enzyme activities in both mice (45) and salmon (25,26). In the only other study on a marine fish, expression of Δ6 desaturase in sea bream liver was higher in fish fed a 16
17 HUFA-free diet compared to that in fish fed a HUFA-rich diet, but activities were not measured in that study (23). In conclusion, the study described here has demonstrated that Atlantic cod express a fatty acyl desaturase gene, the product of which shows substantial Δ6 desaturase activity in an heterologous yeast expression system. The Δ6 desaturase was highly expressed in brain followed by liver, kidney, intestine and all tissues examined. These data reinforce the impression that the poor ability of marine fish, such as cod, to synthesise HUFA is not due to lack of a Δ6 desaturase, but rather to deficiencies in other parts of the biosynthetic pathway. However, cod hepatocytes and enterocytes showed very little Δ6 desaturase or HUFA biosynthesis activity. The expression of the Δ6 desaturase and PUFA elongase genes did not appear to be under significant nutritional regulation, being generally similar in livers and intestine of fish fed the VO diet compared to fish fed the FO diet, with this reflected in unchanged enzyme activities in hepatocytes and enterocytes. Further studies are required to determine why the Δ6 desaturase appears to be barely functional in cod. ACKNOWLEDGMENTS XZ and part of this work were supported by the Biotechnology and Biological Sciences Research Council (BBSRC) of the UK (BB/C51237X/1). CS was supported by a Marie Curie Intra-European Fellowship (MEIF-CT ). NH was in receipt of a Natural Environment Research Council (NERC) studentship. We thank Dr. J. Gordon Bell for formulation and manufacture of the experimental diets. REFERENCES 1. Simopoulos, A.P. (2000) Human Requirements for n-3 Polyunsaturated Fatty Acids, Poult. Sci. 79, Tidwell, J.H. and Allan, G.L. (2002) Fish as Food: Aquaculture s Contribution, World Aquaculture 33,
18 3. Tacon, A.G.J. (2004) Use of Fish Meal and Fish Oil in Aquaculture: A Global Perspective, Aquatic Resources, Culture and Development 1, Pike, I. (2005) Eco-efficiency in Aquaculture: Global Catch of Wild Fish Used in Aquaculture, Internat. Aquafeed 8, Sargent, J.R. Tocher, D.R. and Bell, J.G. (2002) The Lipids, in Fish Nutrition, 3 rd edn. (Halver, J.E. and Hardy, R.W., eds.), pp , Academic Press, San Diego. 6. Tocher, D.R. (2003) Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish, Rev. Fisheries Sci. 11, Ghioni, C., Tocher, D.R., Bell, M.V., Dick, J.R. and Sargent, J.R. (1999) Low C 18 to C 20 Fatty Acid Elongase Activity and Limited Conversion of Stearidonic Acid, 18:4n-3, to Eicosapentaenoic Acid, 20:5n-3, in a Cell Line from the Turbot, Scophthalmus maximus, Biochim. Biophys. Acta 1437, Tocher, D.R. and Ghioni, C. (1999) Fatty Acid Metabolism in Marine Fish: Low Activity of 5 Desaturation in Gilthead Sea Bream (Sparus aurata) Cells, Lipids 34 (1999) Izquierdo, M., Obach, A., Arantzamendi, L., Montero, D., Robaina, L. and Rosenlund, G. (2003) Dietary Lipid Sources for Seabream and Seabass: Growth Performance, Tissue Composition and Flesh Quality, Aquacult. Nutr. 9, Regost, C., Arzel, J., Robin, J., Rosenlund, G. and Kaushik, S.J. (2003) Total Replacement of Fish Oil by Soybean or Linseed Oil With a Return to Fish Oil in Turbot (Psetta maxima). 1. Growth Performance, Flesh Fatty Acid Profile, and Lipid Metabolism, Aquaculture 217, Kurlansky, M. (1998) Cod: A Biography of the Fish That Changed the World. Johnathan Cape, London. 12. Brown, J.A., Minkoff, G. and Puvanendran, V. (2003) Larviculture of Atlantic Cod (Gadus morhua): Progress, Protocols and Problems, Aquaculture 227, Morais, S., Bell, J.G., Robertson, D.A., Roy, W.J. and Morris, P.C. (2001) Protein/Lipid Ratios in Extruded Diets for Atlantic Cod (Gadus morhua L.): Effects on Growth, Feed Utilization, Muscle Composition and Liver Histology, Aquaculture 203, Hemre, G.-I., Karlsen, O., Mangor-Jensen, A. and Rosenlund, G. (2003) Digestibility of Dry Matter, Protein, Starch and Lipid by Cod, Gadus morhua: Comparison of Sampling Methods, Aquaculture 225, Hemre, G.-I., Karlsen, O., Eckhoff, K., viet, K., Mangor-Jensen, A. and Rosenlund, G. (2004) Effect of Season, Light Regime and Diet on Muscle Composition and Selected Parameters in Farmed Atlantic Cod, Gadus morhua L., Aquacult. Res. 35, Lall, S.P. and Nanton, D. (2002) Nutrition of Atlantic Cod, Bull. Aquacult. Assoc. Can. 102,
19 17. Pavlov, D., Kjorsvik, E., Refsti, T. and Andersen, O. (2004) Broodstock and Egg Production, in Culture of Cold Water Marine Fish. (Moksness, E. Kjorsvik, E. and Olsen, Y. eds.), Blackwell, Oxford. 18. Salze, G., Tocher, D.R., Roy, W.J. and Robertson, D.A. (2005) Egg Quality Determinants in Cod (Gadus morhua L.): Egg Performance and Lipids in Eggs From Farmed and Wild Broodstock, Aquaculture Res. 36, Hastings, N., Agaba, M., Tocher, D.R., Leaver, M.J., Dick, J.R., Sargent, J.R. and Teale, A.J. (2001) A Vertebrate Fatty Acid Desaturase With 5 and 6 Activities, Proc. Natl. Acad. Sci. U.S.A. 98, Seiliez, I., Panserat, S., Kaushik, S. and Bergot, P. (2001) Cloning, Tissue Distribution and Nutritional Regulation of a Δ6-Desaturase-Like Enzyme in Rainbow Trout, Comp. Biochem. Physiol. 130B, Hastings, N., Agaba, M.K., Tocher, D.R., Zheng, X., Dickson, C.A., Dick, J.R. and Teale, A.J. (2004) Molecular Cloning and Functional Characterization of Fatty Acyl Desaturase and Elongase cdnas Involved in the Production of Eicosapentaenoic and Docosahexaenoic Acids from α-linolenic Acid in Atlantic Salmon (Salmo salar), Mar. Biotechnol, 6, Zheng, X., Tocher, D.R., Dickson, C.A., Dick, J.R., Bell, J.G. and Teale, A.J. (2005) Highly Unsaturated Fatty Acid Synthesis in Vertebrates: New Insights With the Cloning and Characterisation of a Δ6 Desaturase of Atlantic Salmon, Lipids 40, Seiliez, I., Panserat, S., Corraze, G., Kaushik, S. and Bergot, P. (2003) Cloning and Nutritional Regulation of a 6-Desaturase-Like Enzyme in the Marine Teleost Gilthead Seabream (Sparus aurata), Comp. Biochem. Physiol. 135B, Zheng, X., Seiliez, I., Hastings, N., Tocher, D.R., Panserat, S., Dickson, C.A., Bergot, P., Teale, A.J. (2004) Characterisation and Comparison of Fatty Acyl 6 Desaturase cdnas From Freshwater and Marine Teleost Fish Species, Comp. Biochem. Physiol. 139B, Zheng, X., Tocher, D.R., Dickson, C.A., Bell, J.G. and Teale, A.J. (2004) Effects of Diets Containing Vegetable Oil on Expression of Genes Involved in Polyunsaturated Fatty Acid Biosynthesis in Liver of Atlantic Salmon (Salmo salar), Aquaculture 236, Zheng, X., Torstensen, B.E., Tocher, D.R., Dick, J.R., Henderson, R.J. and Bell, J.G. (2005) Environmental and Dietary Influences on Highly Unsaturated Fatty Acid Biosynthesis and Expression of Fatty Acyl Desaturase and Elongase Genes in Liver of Atlantic Salmon (Salmo salar), Biochim. Biophys. Acta 1734, U.S. National Research Council (1993) Nutrient Requirements of Fish, National Academy Press, Washington D.C. 19
20 28. Agaba, M.K., Tocher, D.R., Dickson, C.A., Zheng, X., Dick, J.R. and Teale, A.J. (2005) Cloning and Functional Characterisation of Polyunsaturated Fatty Acid Elongases from Marine and Freshwater Teleost Fish, Comp. Biochem. Physiol. 142B, Whelan, J.A., Russell, N.B. and Whelan, M.A. (2003) A Method for Absolute Quantification of cdna Using Real-Time PCR, J. Immun. Method. 278, Folch, J., Lees, M. and Sloane-Stanley, G.H. (1957) A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues, J. Biol. Chem. 226, Christie, W.W. (2003) Preparation of Derivatives of Fatty Acids, in Lipid Analysis: Isolation, separation and Structural Analysis of Lipids, vol. 15. pp , J. Barnes and Associates, Bridgewater, UK, 32. Tocher, D.R. and Harvie, D.G. (1988) Fatty Acid Compositions of the Major Phosphoglycerides from Fish Neural Tissues: (n-3) and (n-6) Polyunsaturated Fatty Acids in Rainbow Trout (Salmo gairdneri, L.) and Cod (Gadus morhua) Brains and Retinas, Fish Physiol. Biochem. 5, Ackman, R.G. (1980) Fish Lipids, part 1, in Advances in Fish Science and Technology. (Connell, J.J., ed.), pp , Fishing News Books, Farnham. 34. Mourente, G., Dick, J.R., Bell, J.G. and Tocher, D.R. (2005) Effect of Partial Substitution of Dietary Fish Oil by Vegetable Oils on Desaturation and Oxidation of [1-14 C]18:3n-3 and [1-14 C]20:5n-3 in Hepatocytes and Enterocytes of European Sea Bass (Dicentrarchus labrax L.), Aquaculture 248, Bell, J.G., Strachan, F., Good, J. and Tocher, D.R. (2006) Effect of Echium Oil on Growth, Fatty Acid Composition and Metabolism, Gill Prostaglandin Production and Macrophage Activity in Atlantic Cod (Gadus morhua L.), Aquaculture Res. 37, Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J. (1951) Protein Measurement With the Folin Phenol Reagent, J. Biol. Chem. 193, Ghioni, C., Tocher, D.R. and Sargent, J.R. (1997) The Effect of Culture on Morphology, Lipid and Fatty Acid Composition, and Polyunsaturated Fatty Acid Metabolism of Rainbow Trout (Oncorhynchus mykiss) Skin Cells, Fish Physiol. Biochem. 16, Wilson, R. and Sargent, J.R. (1992) High Resolution Separation of Polyunsaturated Fatty Acids by Argentation Thin-Layer Chromatography, J. Chromatogr. 623, Stubhaug, I., Tocher, D.R., Bell, J.G., Dick, J.R. and Torstensen, B.E. (2005) Fatty Acid Metabolism in Atlantic Salmon (Salmo salar L.) Hepatocytes, and Influence of Dietary Vegetable Oil, Biochim. Biophys. Acta 1734, Saitou, N. and Nei, M. (1987) The Neighbor-Joining Method. A New Method for Reconstructing Phylogenetic Trees, Mol. Biol. Evol. 4,
21 41. Zar, J.H. (1984) Biostatistical Analysis, 2 nd edn. Prentice-Hall, Englewood Cliffs. 42. Nelson, J.S. (1994) Fishes of the World. 3 rd edn. John Wiley and Sons, New York. 43. Tocher, D.R., Fonseca-Madrigal, J., Bell, J.G., Dick, J.R., Henderson, R.J. and Sargent, J.R. (2002) Effects of Diets Containing Linseed Oil on Fatty Acid Desaturation and Oxidation in Hepatocytes and Intestinal Enterocytes in Atlantic Salmon (Salmo salar), Fish Physiol. Biochem. 26, Bell, M.V., Dick, J.R. and Porter, A.E.A. (2003) Pyloric Ceca are a Major Site of 22:6n-3 Synthesis in Rainbow Trout (Oncorhynchus mykiss), Lipids 39, Cho, H.P., Nakamura, M.T. and Clarke, S.D. (1999) Cloning Expression and Nutritional Regulation of the Human Δ6 Desaturase, J. Biol. Chem. 274, Cho, H.P., Nakamura, M.T., Clarke, S.D. (1999) Cloning Expression and Nutritional Regulation of the Human Δ5 Desaturase, J. Biol. Chem. 274, Marquardt, A., Stöhr, H., White, K. and Weber, B.H.F. (2000) cdna Cloning, Genomic Structure, and Chromosomal Localization of Three Members of the Human Fatty Acid Desaturase Family, Genomics 66, Leonard, A.E., Bobik, E.G., Dorado, J., Kroeger, P.E., Chuang, L.-T., Thurmond, J.M., Parker Barnes, J.M., Das, T., Huang, Y.-S. and Murkerji, P. (2000) Cloning of a Human cdna Encoding a Novel Enzyme Involved in the Elongation of Long Chain Polyunsaturated fatty acids, Biochem. J. 350, Leonard, A.E., Kelder, B., Bobik, E.G., Chuang, L.-T., Lewis, C.J., Kopchick, J.J., Murkerji, P. and Huang, Y.-S. (2002) Identification and Expression of Mammalian Long-Chain PUFA Elongation Enzymes, Lipids 37, Inagaki, K., Aki, T., Fukuda, Y., Kawamoto, S., Shigeta, S., Ono, K. and Suzuki, O. (2002) Identification and Expression of a Rat Fatty Acid Elongase Involved the Biosynthesis of C18 Fatty Acids, Biosci. Biotechnol. Biochem. 66,
22 Legends to Figures FIG. 1. Comparison of the deduced amino acid sequence of the 6 polyunsaturated fatty acyl desaturase from Atlantic cod with that of Δ6 and Δ5 desaturases from Atlantic salmon and human, and the zebrafish Δ6/Δ5 bifunctional desaturase. Deduced amino acid sequences were aligned using ClustalX. Identical residues are shaded black and similar residues are shaded grey. Identity/similarity shading was based on the BLOSUM62 matrix and the cut off for shading was 75%. The cytochrome b 5 -like domain is dot-underlined, the two transmembrane regions are dash underlined, the three histidine-rich domains are solid underlined and asterisks on the top mark the haem-binding motif, H-P-G-G. FIG. 2. Phylogenetic tree of cod 6 desaturase and desaturases from other fish species (Atlantic salmon, zebrafish, cherry salmon, rainbow trout, seabream, common carp, turbot and tilapia), mammals (mouse and human), fungus (Mortierella alpina) and nematode (Caenorhabditis elegans). The tree was constructed using the Neighbour Joining method (40) using CLUSTALX and NJPLOT. The horizontal branch length is proportional to amino acid substitution rate per site. The numbers represent the frequencies with which the tree topology presented here was replicated after 1000 bootstrap iterations. Sequences marked with an asterisk are not functionally characterized. FIG. 3. Functional expression of the Atlantic cod putative fatty acyl desaturase in transgenic yeast (Saccharomyces cerevisiae). Panels A and C show the fatty acids extracted from yeast transformed with pyes vector without insert and grown in the presence of 18:2n-6 and 18:3n-3. Panels B, D, E and F show the fatty acid compositions of yeast transformed with pyes vector containing the putative desaturase insert and grown in the presence of 18:2n-6, 18:3n-3, 20:3n-6 and 20:4n-3, respectively. The first four peaks in all panels are the main endogenous fatty acids of S. cerevisiae, namely 16:0 (1), 16:1n-7 (2), 18:0 (3) and 18:1n-9 (with 18:1n-7 as shoulder) (4). Peak 5 in panels A and B, and peak 7 in panels C and D are the exogenously added substrate fatty acids, 18:2n-6 and 18:3n-3, respectively. Peaks 6 and 8 in panels B and D were identified as the resultant desaturated products, namely 18:4n-3 and 18:3n-6, respectively. Peaks 9 and 10 in panels E and F are the exogenously added substrate fatty acids, 20:3n-6 and 20:4n-3, respectively. Vertical axis, FID response; horizontal axis, retention time. FIG. 4. Tissue distribution of fatty acyl 6 desaturase and PUFA elongase genes in Atlantic cod. Transcript (mrna) copy number was determined by quantitative real-time PCR (Q-PCR) and normalized as described in the Materials and Methods Section. Results are expressed as means ± 22
23 SEM (n = 4). L, liver; H, heart; G, gill; WM, white muscle; RM, red muscle; I, intestine; B, brain; A, adipose; S, spleen; K, kidney. FIG. 5. Effect of diet on highly unsaturated fatty acid (HUFA) synthesis in hepatocytes and caecal enterocytes after feeding the experimental diets for 90 days. Results are means ± S.D. (n = 4) and represent the rate of conversion (pmol.h -1.mg protein -1 ) by desaturation/elongation of [1-14 C]18:3n- 3 to all desaturated products. There was no significant effect of diet as determined by the Student t- test (41). FIG. 6. Comparison of highly unsaturated fatty acid (HUFA) synthesis in Atlantic salmon hepatocytes and cod hepatocytes and enterocytes. Presentation of results as described in legend to Fig.5. The salmon data were obtained in salmon smolts of similar size (220g) to the cod in the present trial and assayed at the same time of year (July). The salmon were fed similar FO and VO blend (26). An asterisk denotes a significant effect of diet as determined by the Student t-test (P < 0.05) (41). FIG. 7. Effect of dietary vegetable oil on the expression of fatty acyl 6 desaturase and elongase genes in liver and intestine (pyloric caeca) from Atlantic cod. Transcript (mrna) copy number was determined by quantitative real-time PCR (Q-PCR) and normalized as described in the Materials and Methods Section. Results are expressed as means ± SEM (n = 4). An asterisk denotes a significant effect of diet as determined by the Student t-test (P < 0.05) (41). 23
24 TABLE 1 Formulations and proximate compositions of experimental diets a Fish oil Vegetable oil Fishmeal b Soyabean meal c Wheat d Vitamin mix e Mineral mix f Carboxymethylcellulose Fish oil g Rapeseed oil h Linseed oil i Palm oil j Choline chloride Antioxidant mix k Proximate composition Protein 50.0 ± ± 0.2 Lipid 15.7 ± ± 0.1 Ash 10.4 ± ± 0.1 Fibre 1.6 ± ± 0.3 Moisture 12.1 ± ± 0.1 a Formulation in g.kg -1. Proximate compositions are percentages of total diet and are means ± S.D. (n=3). b LT94, Norsemeal Ltd., London, UK. c Cargill, Swinderbury, UK. d J.D. Martin, Tranent, UK. e Supplied (per kg diet): KH 2 PO 4, 22g; FeSO 4.7H 2 O, 1.0g; ZnSO 4.7H 2 O, 0.13g; MnSO 4.4H 2 O, 52.8 mg; CuSO 4.5H 2 O, 12 mg; CoSO 4.7H 2 O, 2 mg. f Supplied (mg/kg diet): ascorbic acid, 1000; myo-inositol, 400; nicotinic acid, 150; calcium pantothenate, 44; all-rac-a-tocopheryl acetate, 40; riboflavin, 20; pyridoxine hydrochloride, 12; menadione, 10; thiamine hydrochloride, 10; retinyl acetate, 7.3; folic acid, 5; biotin, 1; cholecalciferol, 0.06; cyanocobalamin, g FOSOL, Seven Seas Ltd., Hull, UK. h Tesco, Cheshunt, UK. i Croda, Hull, UK. j United Plantations Bhd, Jenderata Estate, Teluk Intan, Malaysia. k Dissolved in propylene glycol and contained (g/l): butylated hydroxy anisole, 60; propyl gallate, 60; citric acid, 40. All the other ingredients were obtained from Sigma Chemical Co. Ltd., Poole, Dorset, UK. 24
25 TABLE 2 Fatty acid composition of experimental diets for Atlantic cod (Gadus morhua) a Fish oil Vegetable oil 14: : : Total saturated b :1n-7 c :1n :1n :1n-9 d :1n-11 e Total monoenes :2n :4n Total n-6 PUFA f :3n :4n :4n :5n :5n :6n Total n-3 PUFA g Total PUFA h a Results are presented as percentages of total fatty acids by weight and are means of analyses of two samples. b Totals contain 15:0, 17:0, 20:0 and 22:0 at up to 0.4%. c Contains n-9 isomer present at up to 0.1%. d Contains n-11 and n-7 isomers present at up to 0.4%. e Contains n-9 and n-7 isomers at up to 1.0%. f Totals contain 20:2n-6, 20:3n-6 and 22:5n-6 at up to 0.2%. g Totals contain 20:3n-3 present at up to 0.1%. h Contains C 16 PUFA present at up to 0.4%. 25
26 TABLE 3 Identity matrix showing the results of a pair-wise comparison between the amino acid sequences of fish and human fatty acyl desaturases a Gilthead Rainbow Atlantic Atlantic Zebrafish seabream Δ6 Turbot Δ6 trout Δ6 salmon Δ6 salmon Δ5 Δ6/Δ5 Human Δ6 Human Δ5 Atlantic cod Δ Gilthead seabream Δ Turbot Δ Rainbow trout Δ Atlantic salmon Δ Atlantic salmon Δ Zebrafish Δ6/Δ Human Δ6 61 a Percentage of identical amino acid residues 26
27 TABLE 4 Total lipid content and fatty acid composition of total lipid from liver, pyloric caeca and flesh (white muscle) of Atlantic cod fed diets containing fish oil (FO) or a vegetable oil blend (VO) a Liver Pyloric caeca Flesh Fatty acid FO VO FO VO FO VO Lipid content 58.3 ± ± ± ± ± ± :0 4.5 ± ± 0.2 * 2.9 ± ± 0.2 * 2.0 ± ± 0.1 * 16: ± ± 0.5 * 17.7 ± ± 0.3 * 18.2 ± ± 0.9 * 18:0 2.6 ± ± ± ± ± ± 0.1 * Total saturated b 22.2 ± ± 0.6 * 24.4 ± ± 0.6 * 23.5 ± ± 0.9 * 16:1n-7 c 6.6 ± ± 0.4 * 3.2 ± ± ± ± 0.3 * 18:1n ± ± 1.3 * 11.5 ± ± 2.3 * 11.4 ± ± 1.7 * 18:1n ± ± ± ± ± ± :1n-9 d 12.1 ± ± 0.7 * 6.7 ± ± 0.2 * 4.7 ± ± 0.4 * 22:1 8.4 ± ± 0.4 * 5.7 ± ± ± ± :1n ± ± ± ± ± ± 0.0 Total monoenes 50.5 ± ± ± ± ± ± :2n ± ± 0.2 * 2.3 ± ± 1.1 * 2.9 ± ± 0.6 * 20:3n ± ± ± ± ± ± :4n ± ± 0.0 * 1.7 ± ± 0.2 * 1.2 ± ± 0.1 * Total n-6 PUFA e 5.1 ± ± 0.5 * 4.7 ± ± 0.9 * 4.7 ± ± 0.6 * 18:3n ± ± 0.9 * 0.8 ± ± 1.6 * 1.0 ± ± 1.2 * 18:4n ± ± 0.1 * 1.1 ± ± 0.1 * 1.2 ± ± 0.0 * 20:4n ± ± 0.0 * 0.5 ± ± 0.0 * 0.5 ± ± :5n ± ± 0.2 * 11.8 ± ± 0.9 * 13.7 ± ± 0.6 * 22:5n ± ± 0.0 * 1.2 ± ± 0.1 * 1.5 ± ± 0.1 * 22:6n ± ± 0.3 * 22.5 ± ± 3.6 * 28.7 ± ± 2.4 * Total n-3 PUFA f 22.2 ± ± ± ± 3.1 * 46.5 ± ± 2.1 Total PUFA 27.3 ± ± 1.1 * 42.8 ± ± ± ± 1.8 a Results are expressed as percentage of wet weight (lipid content) and percentage of total fatty acids (fatty acid composition) and are means ± S.D. (n=4). Asterisks denote a significant effect of diet on the fatty acid composition of each tissue as determined by the Student's t-test (P < 0.05) (41). b Totals include 15:0 and 20:0 present at at up to 0.3%. c Includes 16:1n-9 at up to 0.2%. d Includes 20:1n-7 at up to 0.4%. e Totals include 18:3n-6, 20:2n-6 and 22:5n-6 present at up to 0.4%. f Totals include 20:n-3 present at up to 0.2%. 27
28 FIG.1 * * * * Atlantic cod D6 MGGGGQLTEPVETS------ACGGR-AASVYTWDEVQKHCHRNDQWLVINRKVYNVTQWAKRHPGGLRVI 63 Atlantic salmon D6 MGGGGQQNDSGEPAKGDRGGPGGGLGGSAVYTWEEVQRHSHRGDQWLVIDRKVYNITQWAKRHPGGIRVI 70 Atlantic salmon D5 MGGGGQQTESSEPAKGDGLEPDGGQGGSAVYTWEEVQRHSHRSDQWLVIDRKVYNITQWAKRHPGGIRVI 70 Zebrafish D6/D5 MGGGGQQTDRITDT NGR--FSSYTWEEVQKHTKHGDQWVVVERKVYNVSQWVKRHPGGLRIL 60 Human D6 MGKGGNQGEGAAER EVSVPTFSWEEIQKHNLRTDRWLVIDRKVYNITKWSIQHPGGQRVI 60 Human D5 MAPDPLAAETAAQG LTPRYFTWDEVAQRSGCEERWLVIDRKVYNISEFTRRHPGGSRVI 59 Atlantic cod D6 SHYAGEDATEAFLAFHPNPKLVQKFLKPLLIGELAVTEPSQDRNKNAAVVEDFQALRTRAEGLGLFQAQP 133 Atlantic salmon D6 SHFAGEDATDAFVAFHPNPNFVRKFLKPLLIGELAPTEPSQDHGKNAVLVQDFQALRNRVEREGLLRARP 140 Atlantic salmon D5 SHFAGEDATEAFSAFHLDANFVRKFLKPLLIGELAPTEPSQDHGKNAALVQDFQALRDHVEREGLLRARL 140 Zebrafish D6/D5 GHYAGEDATEAFTAFHPNLQLVRKYLKPLLIGELEASEPSQDRQKNAALVEDFRALRERLEAEGCFKTQP 130 Human D6 GHYAGEDATDAFRAFHPDLEFVGKFLKPLLIGELAPEEPSQDHGKNSKITEDFRALRKTAEDMNLFKTNH 130 Human D5 SHYAGQDATDPFVAFHINKGLVKKYMNSLLIGELSPEQPSFEPTKNKELTDEFRELRATVERMGLMKANH 129 Atlantic cod D6 LFFCLHLGHILLLELLAWMSVWLWGTGWRTTLLCSFILAVAQAQAGWLQHDFGHLSVFKLSRWNHIFHKF 203 Atlantic salmon D6 LFFSLYLGHILLLEALALGLLWVWGTSWSLTLLCSLMLATSQSQAGWLQHDYGHLSVCKKSSWNHVLHKF 210 Atlantic salmon D5 LFFSLYLGHILLLEALALGLLWVWGTSWSLTLLCSLMLATSQAQAGWLQHDYGHLSVCKKSSWNHKLHKF 210 Zebrafish D6/D5 LFFALHLGHILLLEAIAFMMVWYFGTGWINTLIVAVILATAQSQAGWLQHDFGHLSVFKTSGMNHLVHKF 200 Human D6 VFFLLLLAHIIALESIAWFTVFYFGNGWIPTLITAFVLATSQAQAGWLQHDYGHLSVYRKPKWNHLVHKF 200 Human D5 VFFLLYLLHILLLDGAAWLTLWVFGTSFLPFLLCAVLLSAVQAQAGWLQHDFGHLSVFSTSKWNHLLHHF 199 Atlantic cod D6 IIGHLKGASGNWWNHRHFQHHAKPNVFSKDPDVNMLH-VFVVGDIQPVEYGIKKIKYMPYHHQHQYFFLV 272 Atlantic salmon D6 VIGHLKGASANWWNHRHFQHHAKPNVLSKDPDVNMLH-VFVLGDKQPVEYGIKKLKYMPYHHQHQYFFLI 279 Atlantic salmon D5 VIGHLKGASANWWNHRHFQHHAKPNVFRKDPDINSLP-VFVLGDTQPVEYGIKKLKYMPYHHQHQYFFLI 279 Zebrafish D6/D5 VIGHLKGASAGWWNHRHFQHHAKPNIFKKDPDVNMLN-AFVVGNVQPVEYGVKKIKHLPYNHQHKYFFFI 269 Human D6 VIGHLKGASANWWNHRHFQHHAKPNIFHKDPDVNMLH-VFVLGEWQPIEYGKKKLKYLPYNHQHEYFFLI 269 Human D5 VIGHLKGAPASWWNHMHFQHHAKPNCFRKDPDINMHPFFFALGKILSVELGKQKKNYMPYNHQHKYFFLI 269 Atlantic cod D6 GPPLLIPVYFHIQILRAMFSRRDWVDLAWSMSYYLRYFCCYAPFYGLLGSVALISFVRFLESHWFVWVTQ 342 Atlantic salmon D6 GPPLLIPVFFTIQIFQTMFSQRNWVDLAWSMTFYLRFFCSYYPFFGFFGSVALITFVRFLESHWFVWVTQ 349 Atlantic salmon D5 GPPLIVPVFFNIQIFRTMFSQRDWVDLAWSMSFYLRFFCCYYPFFGFFGSVALISFVRFLESHWFVWVTQ 349 Zebrafish D6/D5 GPPLLIPVYFQFQIFHNMISHGMWVDLLWCISYYVRYFLCYTQFYGVFWAIILFNFVRFMESHWFVWVTQ 339 Human D6 GPPLLIPMYFQYQIIMTMIVHKNWVDLAWAVSYYIRFFITYIPFYGILGALLFLNFIRFLESHWFVWVTQ 339 Human D5 GPPALLPLYFQWYIFYFVIQRKKWVDLAWMITFYVRFFLTYVPLLGLKAFLGLFFIVRFLESNWFVWVTQ 339 Atlantic cod D6 MNHLPMNIDHEKQQDWLSMQLSATCNIEQSCFNDWFSGHLNFQIEHHLFPTMPRHNYQVLAPLVRALCEK 412 Atlantic salmon D6 MNHLPMEIDHERHQDWLTMQLSGTCNIEQSTFNDWFSGHLNFQIEHHLFPTMPRHNYHLVAPLVRTLCEK 419 Atlantic salmon D5 MNHLPMEMDHERHQDWLTMQLSATCNIEQSTFNDWFSGHLNFQIEHHLFPTMPRHNYHLVAPLVRTLCEK 419 Zebrafish D6/D5 MSHIPMNIDYEKNQDWLSMQLVATCNIEQSAFNDWFSGHLNFQIEHHLFPTVPRHNYWRAAPRVRALCEK 409 Human D6 MNHIVMEIDQEAYRDWFSSQLTATCNVEQSFFNDWFSGHLNFQIEHHLFPTMPRHNLHKIAPLVKSLCAK 409 Human D5 MNHIPMHIDHDRNMDWVSTQLQATCNVHKSAFNDWFSGHLNFQIEHHLFPTMPRHNYHKVAPLVQSLCAK 409 Atlantic cod D6 HSIPYQEKTLWRGVADVVRSLKNSGDLWMDAYLHK 447 Atlantic salmon D6 HGIPYQVKTLQKAIIDVVRSLKKSGDLWLDAYLHK 454 Atlantic salmon D5 HGVPYQVKTLQKGMTDVVRSLKKSGDLWLDAYLHK 454 Zebrafish D6/D5 YGVKYQEKTLYGAFADIIRSLEKSGELWLDAYLNK 444 Human D6 HGIEYQEKPLLRALLDIIRSLKKSGKLWLDAYLHK 444 Human D5 HGIEYQSKPLLSAFADIIHSLKESGQLWLDAYLHQ
29 FIG Atlantic Salmon 5, AF Cherry Salmon Des1*, AB Cherry Salmon Des2*,AB Rainbow Trout 6, AF Atlantic Salmon 6, AY Atlantic Cod 6, DQ Nile Tilapia Des*, AB Turbot 6, AY Gilthead Seabream 6, AY Common Carp 6, AF Zebrafish 5/ 6, AF Human 6, AF Mouse 6, AF Human 5, AF Mouse 5, AB Mortierella Alpina 5, AF Mortierella Alpina 6, AF C. Elegans 6, AF C. Elegans 5, AF
30 FIG.3 30
31 31

32 FIG. 4 32
33 FIG. 5 33
34 FIG.6 34

35 FIG.7 35
D. R. Tocher*, M. Agaba, N. Hastings, Bell, J.G., J. R. Dick and A. J. Teale
 Nutritional regulation of hepatocyte fatty acid desaturation and polyunsaturated fatty acid composition in zebrafish (Danio rerio) and tilapia (Oreochromis niloticus) D. R. Tocher*, M. Agaba, N. Hastings,
Nutritional regulation of hepatocyte fatty acid desaturation and polyunsaturated fatty acid composition in zebrafish (Danio rerio) and tilapia (Oreochromis niloticus) D. R. Tocher*, M. Agaba, N. Hastings,
Nicola Hastings, Morris K. Agaba, Douglas R. Tocher, Xiaozhong Zheng, Cathryn A. Dickson, James R. Dick, and Alan J. Teale
 Mar. Biotechnol. 6, 463 474, 2005 DOI: 10.1007/s10126-004-3002-8 Ó 2004 Springer Science+Business Media, Inc. Molecular Cloning and Functional Characterization of Fatty Acyl Desaturase and Elongase cdnas
Mar. Biotechnol. 6, 463 474, 2005 DOI: 10.1007/s10126-004-3002-8 Ó 2004 Springer Science+Business Media, Inc. Molecular Cloning and Functional Characterization of Fatty Acyl Desaturase and Elongase cdnas
A vertebrate fatty acid desaturase with Δ5 and Δ6 activities
 Classification: Biological Sciences, Biochemistry A vertebrate fatty acid desaturase with Δ5 and Δ6 activities Nicola Hastings, Morris Agaba, Douglas R.Tocher, Michael J. Leaver, James R. Dick, John R.
Classification: Biological Sciences, Biochemistry A vertebrate fatty acid desaturase with Δ5 and Δ6 activities Nicola Hastings, Morris Agaba, Douglas R.Tocher, Michael J. Leaver, James R. Dick, John R.
* Author for correspondence. Tel.: ; fax: address:
 Effect of partial substitution of dietary fish oil by vegetable oils on desaturation and β-oxidation of [1-14 C]18:3n-3 (LNA) and [1-14 C]20:5n-3 (EPA) in hepatocytes and enterocytes of European sea bass
Effect of partial substitution of dietary fish oil by vegetable oils on desaturation and β-oxidation of [1-14 C]18:3n-3 (LNA) and [1-14 C]20:5n-3 (EPA) in hepatocytes and enterocytes of European sea bass
This is the peer reviewed version of this article.
 Hepatocyte fatty acid desaturation and polyunsaturated fatty acid composition of liver in salmonids: Effects of dietary vegetable oil by D. R. Tocher, J. G. Bell, P. MacGlaughlin, F. McGhee, and J. R.
Hepatocyte fatty acid desaturation and polyunsaturated fatty acid composition of liver in salmonids: Effects of dietary vegetable oil by D. R. Tocher, J. G. Bell, P. MacGlaughlin, F. McGhee, and J. R.
DIETARY LIPIDS, IMMUNE FUNCTION AND PATHOGENESIS OF DISEASE IN FISH
 DIETARY LIPIDS, IMMUNE FUNCTION AND PATHOGENESIS OF DISEASE IN FISH Santosh P. Lall and Joyce E. Milley National Research Council Canada, Institute for Marine Biosciences, 1411 Oxford Street, Halifax,
DIETARY LIPIDS, IMMUNE FUNCTION AND PATHOGENESIS OF DISEASE IN FISH Santosh P. Lall and Joyce E. Milley National Research Council Canada, Institute for Marine Biosciences, 1411 Oxford Street, Halifax,
Molecular basis for differential elongation of omega-3 docosapentaenoic acid by the rat Elovl5 and Elovl2
 Molecular basis for differential elongation of omega-3 docosapentaenoic acid by the rat Elovl5 and Elovl2 Melissa K. Gregory, 1 Leslie G. Cleland, and Michael J. James Rheumatology Unit, Royal Adelaide
Molecular basis for differential elongation of omega-3 docosapentaenoic acid by the rat Elovl5 and Elovl2 Melissa K. Gregory, 1 Leslie G. Cleland, and Michael J. James Rheumatology Unit, Royal Adelaide
Fatty Acid Metabolism in Marine Fish: Low Activity of Fatty Acyl
 Fatty Acid Metabolism in Marine Fish: Low Activity of Fatty Acyl Δ5 Desaturation in Gilthead Sea Bream (Sparus aurata) Cells Douglas R. Tocher*, and Cristina Ghioni Unit of Aquatic Biochemistry, Institute
Fatty Acid Metabolism in Marine Fish: Low Activity of Fatty Acyl Δ5 Desaturation in Gilthead Sea Bream (Sparus aurata) Cells Douglas R. Tocher*, and Cristina Ghioni Unit of Aquatic Biochemistry, Institute
Fish Meal and Fish Oil Replacement with Landbased Ingredients in Hybrid Striped Bass Feeds
 Fish Meal and Fish Oil Replacement with Landbased Ingredients in Hybrid Striped Bass Feeds Jesse T. Trushenski and Jonah M. May, Center for Fisheries, Aquaculture, and Aquatic Sciences, Southern Illinois
Fish Meal and Fish Oil Replacement with Landbased Ingredients in Hybrid Striped Bass Feeds Jesse T. Trushenski and Jonah M. May, Center for Fisheries, Aquaculture, and Aquatic Sciences, Southern Illinois
A Zebrafish cdna Encoding a Multifunctional Fatty Acid Elongase. Involved in the Production of Eicosapentaenoic (20:5n-3) and
 and") A Zebrafish cdna Encoding a Multifunctional Fatty Acid Elongase Involved in the Production of Eicosapentaenoic (20:5n-3) and Docosahexaenoic (22:6n-3) Acids Morris Agaba, Douglas R. Tocher*, Cathryn A.
A Zebrafish cdna Encoding a Multifunctional Fatty Acid Elongase Involved in the Production of Eicosapentaenoic (20:5n-3) and Docosahexaenoic (22:6n-3) Acids Morris Agaba, Douglas R. Tocher*, Cathryn A.
Nutrient requirement of marine fish larvae for essential fatty acids and phospholipids
 Nutrient requirement of marine fish larvae for essential fatty acids and phospholipids 266071 S96 A 1000-096200611-0075-07 20% n- n- HUFAn-6 n-6 HUFA 1 1.1 1 [1] 1.2 n- HUFA n-9 n-6 n- n-6 AA204n-6 n-
Nutrient requirement of marine fish larvae for essential fatty acids and phospholipids 266071 S96 A 1000-096200611-0075-07 20% n- n- HUFAn-6 n-6 HUFA 1 1.1 1 [1] 1.2 n- HUFA n-9 n-6 n- n-6 AA204n-6 n-
Keywords: Atlantic salmon; Rapeseed oil; Polyunsaturated fatty acids (PUFA); β-oxidation
; β-oxidation") 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 Influence of dietary protein / lipid ratio and fish oil substitution on fatty acid composition and metabolism
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 Influence of dietary protein / lipid ratio and fish oil substitution on fatty acid composition and metabolism
FATTY ACIDS COMPOSITION OF FISH, LINSEED AND RAPESEED OILS
 Short Communication FATTY ACIDS COMPOSITION OF FISH, LINSEED AND RAPESEED OILS S. Ezhil Valavan 1, B Mohan, P Selvaraj, S. C. Edwin, K. Mani, R. Amutha and A. Bharathidhasan Directorate of Distance Education
Short Communication FATTY ACIDS COMPOSITION OF FISH, LINSEED AND RAPESEED OILS S. Ezhil Valavan 1, B Mohan, P Selvaraj, S. C. Edwin, K. Mani, R. Amutha and A. Bharathidhasan Directorate of Distance Education
INFLUENCE OF DIETARY MICRONUTRIENTS ON GROWTH AND DEVELOPMENT OF DIPLOID AND TRIPLOID ATLANTIC SALMON
 1 2 INFLUENCE OF DIETARY MICRONUTRIENTS ON GROWTH AND DEVELOPMENT OF DIPLOID AND TRIPLOID ATLANTIC SALMON (Salmo salar) PARR Douglas Tocher (UoS) San Seastian, 15 Octoer 214 John F. Taylor 1, Christian
1 2 INFLUENCE OF DIETARY MICRONUTRIENTS ON GROWTH AND DEVELOPMENT OF DIPLOID AND TRIPLOID ATLANTIC SALMON (Salmo salar) PARR Douglas Tocher (UoS) San Seastian, 15 Octoer 214 John F. Taylor 1, Christian
THE ROLE OF RENDERED PRODUCTS IN AQUACULTURE FEEDS Dr. Jesse Trushenski
 THE ROLE OF RENDERED PRODUCTS IN AQUACULTURE FEEDS Dr. Jesse Trushenski Center for Fisheries Aquaculture & Aquatic Sciences Southern Illinois University Carbondale Carbondale, Illinois USA saluski@siu.edu
THE ROLE OF RENDERED PRODUCTS IN AQUACULTURE FEEDS Dr. Jesse Trushenski Center for Fisheries Aquaculture & Aquatic Sciences Southern Illinois University Carbondale Carbondale, Illinois USA saluski@siu.edu
Fatty acid metabolism in Atlantic salmon (Salmo salar L.) hepatocytes, and influence of dietary vegetable oil
 hepatocytes, and influence of dietary vegetable oil") Fatty acid metabolism in Atlantic salmon (Salmo salar L.) hepatocytes, and influence of dietary vegetable oil Ingunn Stubhuag a1, Douglas R. Tocher b, J. Gordon Bell b, James R. Dick b, Bente E. Torstensen
Fatty acid metabolism in Atlantic salmon (Salmo salar L.) hepatocytes, and influence of dietary vegetable oil Ingunn Stubhuag a1, Douglas R. Tocher b, J. Gordon Bell b, James R. Dick b, Bente E. Torstensen
ALTERNATIVE SOURCES OF OMEGA-3 OILS FOR BARRAMUNDI, Lates calcarifer, AQUACULTURE
 ALTERNATIVE SOURCES OF OMEGA-3 OILS FOR BARRAMUNDI, Lates calcarifer, AQUACULTURE By Ramez Alhazzaa B.Sc., Grad. Dip. Animal Husbandry A thesis submitted in fulfilment of the requirements for the degree
ALTERNATIVE SOURCES OF OMEGA-3 OILS FOR BARRAMUNDI, Lates calcarifer, AQUACULTURE By Ramez Alhazzaa B.Sc., Grad. Dip. Animal Husbandry A thesis submitted in fulfilment of the requirements for the degree
Recent Developments in the Essential Fatty Acid Nutrition of Fish Abstract. I. Introduction
 Recent Developments in the Essential Fatty Acid Nutrition of Fish John Sargent, Gordon Bell, Lesley McEvoy, Douglas Tocher and Alicia Estevez Institute of Aquaculture, University of Stirling, Stirling
Recent Developments in the Essential Fatty Acid Nutrition of Fish John Sargent, Gordon Bell, Lesley McEvoy, Douglas Tocher and Alicia Estevez Institute of Aquaculture, University of Stirling, Stirling
FISH NUTRITION 101 Feeds & Feeding Strategies for Aquaculture Dr. Jesse Trushenski
 FISH NUTRITION 101 Feeds & Feeding Strategies for Aquaculture Dr. Jesse Trushenski Center for Fisheries Aquaculture & Aquatic Sciences Southern Illinois University Carbondale Carbondale, Illinois USA saluski@siu.edu
FISH NUTRITION 101 Feeds & Feeding Strategies for Aquaculture Dr. Jesse Trushenski Center for Fisheries Aquaculture & Aquatic Sciences Southern Illinois University Carbondale Carbondale, Illinois USA saluski@siu.edu
Is the extended use of fatty acid percentage in fish studies adequate and justified?
 Session 24 - nafsika@aua.gr Is the extended use of fatty acid percentage in fish studies adequate and justified? Nafsika Karakatsouli Department of Applied Hydrobiology Faculty of Animal Science and Aquaculture
Session 24 - nafsika@aua.gr Is the extended use of fatty acid percentage in fish studies adequate and justified? Nafsika Karakatsouli Department of Applied Hydrobiology Faculty of Animal Science and Aquaculture
ブリ幼魚用飼料における魚油精製副産物の利用
 Journal of Fisheries Technology, 7(2), 69-74, 2015 水産技術,7(2), 69-74, 2015 ブリ幼魚用飼料における魚油精製副産物の利用 古板博文 *1 杉田毅 *2 山本剛史 *3 風直樹 *4 *4 山本浩志 Evaluation of nutritional value of a fish oil by-product in a diet
Journal of Fisheries Technology, 7(2), 69-74, 2015 水産技術,7(2), 69-74, 2015 ブリ幼魚用飼料における魚油精製副産物の利用 古板博文 *1 杉田毅 *2 山本剛史 *3 風直樹 *4 *4 山本浩志 Evaluation of nutritional value of a fish oil by-product in a diet
Dietary neutral lipid level and source affect food intake, nutrient absorption, gut structure, enzymatic activity and growth in marine fish larvae
 Dietary neutral lipid level and source affect food intake, nutrient absorption, gut structure, enzymatic activity and growth in marine fish larvae S. Morais 1, L.E.C. Conceição 1, W. Koven 2, I. Rønnestad
Dietary neutral lipid level and source affect food intake, nutrient absorption, gut structure, enzymatic activity and growth in marine fish larvae S. Morais 1, L.E.C. Conceição 1, W. Koven 2, I. Rønnestad
Optimising lipid nutrition in early developing flatfish larvae
 Optimising lipid nutrition in early developing flatfish larvae Dr Gordon Bell, Nutrition Group, Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, Scotland, U.K. What are essential fatty
Optimising lipid nutrition in early developing flatfish larvae Dr Gordon Bell, Nutrition Group, Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, Scotland, U.K. What are essential fatty
Santhosh Karanth*, Prakash Sharma, Asim K. Pal and G. Venkateshwarlu Central Institute of Fisheries Education, Versova, Mumbai , India
 565 Asian-Aust. J. Anim. Sci. Vol. 22, No. 4 : 565-575 April 2009 www.ajas.info Effect of Different Vegetable Oils on Growth and Fatty Acid Profile of Rohu (Labeo rohita, Hamilton); Evaluation of a Return
565 Asian-Aust. J. Anim. Sci. Vol. 22, No. 4 : 565-575 April 2009 www.ajas.info Effect of Different Vegetable Oils on Growth and Fatty Acid Profile of Rohu (Labeo rohita, Hamilton); Evaluation of a Return
British Journal of Nutrition. q The Authors 2010
 (2010), page 1 of 14 q The Authors 2010 doi:10.1017/s0007114510004605 Influence of the dietary protein:lipid ratio and fish oil substitution on fatty acid composition and metabolism of Atlantic salmon
(2010), page 1 of 14 q The Authors 2010 doi:10.1017/s0007114510004605 Influence of the dietary protein:lipid ratio and fish oil substitution on fatty acid composition and metabolism of Atlantic salmon
Evaluating dietary fish oil replacement in juvenile Florida pompano
 FEED SUSTAINABILITY (/ADVOCATE/CATEGORY/FEED-SUSTAINABILITY) Evaluating dietary fish oil replacement in juvenile Florida pompano Tuesday, 2 January 2018 By Artur Nishioka Rombenso, Ph.D., Jesse T. Trushenski,
FEED SUSTAINABILITY (/ADVOCATE/CATEGORY/FEED-SUSTAINABILITY) Evaluating dietary fish oil replacement in juvenile Florida pompano Tuesday, 2 January 2018 By Artur Nishioka Rombenso, Ph.D., Jesse T. Trushenski,
General Overview Usage of Animal Products in Aqua Feed
 General Overview Usage of Animal Products in Aqua Feed Eric De Muylder CreveTec eric@crevetec.be www.crevetec.be Aquaculture overview More than 300 species Fish, crustaceans, shellfish, algae Extensive
General Overview Usage of Animal Products in Aqua Feed Eric De Muylder CreveTec eric@crevetec.be www.crevetec.be Aquaculture overview More than 300 species Fish, crustaceans, shellfish, algae Extensive
Sadasivam Kaushik. UR1067, INRA, Nutrition, Metabolism & Aquaculture Unit St-Pée-sur-Nivelle, France
 Issues and achievements Sadasivam Kaushik UR1067, INRA, Nutrition, Metabolism & Aquaculture Unit 64310 St-Pée-sur-Nivelle, France kaushik@st-pee.inra.fr Grant number: FP7-288925 Jan 2012 Dec 2016 Kick
Issues and achievements Sadasivam Kaushik UR1067, INRA, Nutrition, Metabolism & Aquaculture Unit 64310 St-Pée-sur-Nivelle, France kaushik@st-pee.inra.fr Grant number: FP7-288925 Jan 2012 Dec 2016 Kick
Comparative Study of Fat (Total Cholestrol and Fatty acids) Profile in Farm cultivated and river water fishes communities of Labeo rohita
 Profile in Farm cultivated and river water fishes communities of Labeo rohita") International Journal of Scientific and Research Publications, Volume 7, Issue 7, July 2017 763 Comparative Study of Fat (Total Cholestrol and Fatty acids) Profile in Farm cultivated and river water fishes
International Journal of Scientific and Research Publications, Volume 7, Issue 7, July 2017 763 Comparative Study of Fat (Total Cholestrol and Fatty acids) Profile in Farm cultivated and river water fishes
One fish, two fish, feed fish, food fish Meeting nutritional challenges in aquaculture and aquatic natural resources management
 Southern Illinois University Carbondale One fish, two fish, feed fish, food fish Meeting nutritional challenges in aquaculture and aquatic natural resources management Jesse T. Trushenski Fish Culture
Southern Illinois University Carbondale One fish, two fish, feed fish, food fish Meeting nutritional challenges in aquaculture and aquatic natural resources management Jesse T. Trushenski Fish Culture
British Journal of Nutrition
 (2008), 100, 992 1003 q The Authors 2008 doi:10.1017/s0007114508966071 High levels of vegetable oils in plant protein-rich diets fed to gilthead sea bream (Sparus aurata L.): growth performance, muscle
(2008), 100, 992 1003 q The Authors 2008 doi:10.1017/s0007114508966071 High levels of vegetable oils in plant protein-rich diets fed to gilthead sea bream (Sparus aurata L.): growth performance, muscle
ANSC/NUTR 618 LIPIDS & LIPID METABOLISM. Fatty Acid Elongation and Desaturation
 ANSC/NUTR 618 LIPIDS & LIPID METABOLISM I. Fatty acid elongation A. General 1. At least 60% of fatty acids in triacylglycerols are C18. 2. Free palmitic acid (16:0) synthesized in cytoplasm is elongated
ANSC/NUTR 618 LIPIDS & LIPID METABOLISM I. Fatty acid elongation A. General 1. At least 60% of fatty acids in triacylglycerols are C18. 2. Free palmitic acid (16:0) synthesized in cytoplasm is elongated
Fatty Acid Methylation Kits
 Methyl esterification kit for fatty acids analysis Fatty Acid Methylation Kits Below are two methods for efficiently preparing fatty acid samples for GC analysis. Neither method requires high temperatures,
Methyl esterification kit for fatty acids analysis Fatty Acid Methylation Kits Below are two methods for efficiently preparing fatty acid samples for GC analysis. Neither method requires high temperatures,
OPPORTUNITIES FOR RENDERED PRODUCTS IN AQUACULTURE Advancing science & industry through partnership
 OPPORTUNITIES FOR RENDERED PRODUCTS IN AQUACULTURE Advancing science & industry through partnership Jesse T. Trushenski CENTER FOR FISHERIES, AQUACULTURE, & AQUATIC SCIENCES THE WORLD IS HUNGRY CGIAR CCAFS
OPPORTUNITIES FOR RENDERED PRODUCTS IN AQUACULTURE Advancing science & industry through partnership Jesse T. Trushenski CENTER FOR FISHERIES, AQUACULTURE, & AQUATIC SCIENCES THE WORLD IS HUNGRY CGIAR CCAFS
Describing Nutritional Requirements of Fish
 Lesson C3 3 Describing Nutritional Requirements of Fish Unit C. Animal Wildlife Management Problem Area 3. Fish Management Lesson 3. Describing Nutritional Requirements of Fish New Mexico Content Standard:
Lesson C3 3 Describing Nutritional Requirements of Fish Unit C. Animal Wildlife Management Problem Area 3. Fish Management Lesson 3. Describing Nutritional Requirements of Fish New Mexico Content Standard:
Analysis of Omega 3 and Omega 6 FAMEs in Fish Oil and Animal Fat Using an Agilent J&W DB-FATWAX Ultra Inert GC Column
 Application Note Food Analysis of Omega 3 and Omega 6 FAMEs in Fish Oil and Animal Fat Using an Agilent J&W DB-FATWAX Ultra Inert GC Column Authors Ingrid Van Der Meer, Yun Zou, and Gustavo Serrano Agilent
Application Note Food Analysis of Omega 3 and Omega 6 FAMEs in Fish Oil and Animal Fat Using an Agilent J&W DB-FATWAX Ultra Inert GC Column Authors Ingrid Van Der Meer, Yun Zou, and Gustavo Serrano Agilent
EFFECTS OF THE ß-CAROTENE ON THE GROWTH PERFORMANCE AND SKIN PIGMENTATION OF RAINBOW TROUT (ONCORHYNCHUS MYKISS, W. 1792)
") EFFECTS OF THE ß-CAROTENE ON THE GROWTH PERFORMANCE AND SKIN PIGMENTATION OF RAINBOW TROUT (ONCORHYNCHUS MYKISS, W. 1792) Gülüzar TUNA KELEŞTEMUR *, Özlem EMİR ÇOBAN Fisheries Faculty, Firat University,
EFFECTS OF THE ß-CAROTENE ON THE GROWTH PERFORMANCE AND SKIN PIGMENTATION OF RAINBOW TROUT (ONCORHYNCHUS MYKISS, W. 1792) Gülüzar TUNA KELEŞTEMUR *, Özlem EMİR ÇOBAN Fisheries Faculty, Firat University,
V. Karalazos, E.Å. Bendiksen, J.G. Bell. S (10) DOI: doi: /j.aquaculture Reference: AQUA
 DOI: doi: /j.aquaculture Reference: AQUA") Accepted Manuscript Interactive effects of dietary protein / lipid level and oil source on growth, feed utilisation and nutrient and fatty acid digestibility of Atlantic salmon V. Karalazos, E.Å. Bendiksen,
Accepted Manuscript Interactive effects of dietary protein / lipid level and oil source on growth, feed utilisation and nutrient and fatty acid digestibility of Atlantic salmon V. Karalazos, E.Å. Bendiksen,
THE ESSENTIAL FATTY ACID REQUIREMENTS OF BROILERS 1
 2001 Poultry Science Association, Inc. THE ESSENTIAL FATTY ACID REQUIREMENTS OF BROILERS 1 W. O. ZORNIG, G. M. PESTI 2, and R. I. BAKALLI Department of Poultry Science The University of Georgia Athens,
2001 Poultry Science Association, Inc. THE ESSENTIAL FATTY ACID REQUIREMENTS OF BROILERS 1 W. O. ZORNIG, G. M. PESTI 2, and R. I. BAKALLI Department of Poultry Science The University of Georgia Athens,
REP15/FO Appendix III 1. PROPOSED DRAFT CODEX STANDARD FOR FISH OILS (at Step 5 of the Procedure)
") REP15/FO Appendix III 1 PROPOSED DRAFT CODEX STANDARD FOR FISH OILS (at Step 5 of the Procedure) Appendix III 1. Scope This Standard applies to the fish oils described in section 2 that are presented in
REP15/FO Appendix III 1 PROPOSED DRAFT CODEX STANDARD FOR FISH OILS (at Step 5 of the Procedure) Appendix III 1. Scope This Standard applies to the fish oils described in section 2 that are presented in
Substituting De-hulled Soybean Meal for Fish Meal in Diets for Hybrid Tilapia Rearing in Water Re-circulating System
 Substituting De-hulled Soybean Meal for Fish Meal in Diets for Hybrid Tilapia Rearing in Water Re-circulating System 2004 Principle investigator: Chen-Huei Huang, Ph.D. Professor of Aquaculture Nutrition
Substituting De-hulled Soybean Meal for Fish Meal in Diets for Hybrid Tilapia Rearing in Water Re-circulating System 2004 Principle investigator: Chen-Huei Huang, Ph.D. Professor of Aquaculture Nutrition
: Overview of EFA metabolism
 Figure 1 gives an overview of the metabolic fate of the EFAs when consumed in the diet. The n-6 and n-3 PUFAs when consumed in the form of dietary triglyceride from various food sources undergoes digestion
Figure 1 gives an overview of the metabolic fate of the EFAs when consumed in the diet. The n-6 and n-3 PUFAs when consumed in the form of dietary triglyceride from various food sources undergoes digestion
P.P. SRIVASTAVA AND K.K. JAIN
 Journal of the Indian Fisheries Association 24, 1994 (Proc. Nat. Symp. Aqu~crQps), 139ul43 139 EFFECT OF VITAMIN E (a.. TOCOPHEROL) ON PROTEIN CONTENT OF LABEO ROHITA (HAM.).FRY P.P. SRIVASTAVA AND K.K.
Journal of the Indian Fisheries Association 24, 1994 (Proc. Nat. Symp. Aqu~crQps), 139ul43 139 EFFECT OF VITAMIN E (a.. TOCOPHEROL) ON PROTEIN CONTENT OF LABEO ROHITA (HAM.).FRY P.P. SRIVASTAVA AND K.K.
Recent developments in the essential fatty acid nutrition of fish
 Ž. Aquaculture 177 1999 191 199 Recent developments in the essential fatty acid nutrition of fish John Sargent ), Gordon Bell, Lesley McEvoy, Douglas Tocher, Alicia Estevez Institute of Aquaculture, UniÕersity
Ž. Aquaculture 177 1999 191 199 Recent developments in the essential fatty acid nutrition of fish John Sargent ), Gordon Bell, Lesley McEvoy, Douglas Tocher, Alicia Estevez Institute of Aquaculture, UniÕersity
Changes in Tissue Lipid and Fatty Acid Composition of Farmed Rainbow Trout in Response to Dietary Camelina Oil as a Replacement of Fish Oil
 Lipids (2014) 49:97 111 DOI 10.1007/s11745-013-3862-7 ORIGINAL ARTICLE Changes in Tissue Lipid and Fatty Acid Composition of Farmed Rainbow Trout in Response to Dietary Camelina Oil as a Replacement of
Lipids (2014) 49:97 111 DOI 10.1007/s11745-013-3862-7 ORIGINAL ARTICLE Changes in Tissue Lipid and Fatty Acid Composition of Farmed Rainbow Trout in Response to Dietary Camelina Oil as a Replacement of
Chemistry 107 Exam 4 Study Guide
 Chemistry 107 Exam 4 Study Guide Chapter 10 10.1 Recognize that enzyme catalyze reactions by lowering activation energies. Know the definition of a catalyst. Differentiate between absolute, relative and
Chemistry 107 Exam 4 Study Guide Chapter 10 10.1 Recognize that enzyme catalyze reactions by lowering activation energies. Know the definition of a catalyst. Differentiate between absolute, relative and
An overview of lipid nutrition with emphasis on alternative lipid sources in tilapia feeds
 An overview of lipid nutrition with emphasis on alternative lipid sources in tilapia feeds Wing-Keong Ng & Cheong-Yew Chong Fish Nutrition Laboratory School of Biological Sciences Universiti Sains Malaysia
An overview of lipid nutrition with emphasis on alternative lipid sources in tilapia feeds Wing-Keong Ng & Cheong-Yew Chong Fish Nutrition Laboratory School of Biological Sciences Universiti Sains Malaysia
Spawning advanced and delayed control photoperiod control temperature control
 Spawning advanced and delayed control photoperiod control temperature control Photoperiod control Day Length and Temperature 30 25 Water Temperature (C) 20 15 10 Day Length (h) 5 0 Advancing or delaying
Spawning advanced and delayed control photoperiod control temperature control Photoperiod control Day Length and Temperature 30 25 Water Temperature (C) 20 15 10 Day Length (h) 5 0 Advancing or delaying
Polyunsaturated fatty acid metabolism in a cell culture model of essential fatty acid deficiency in a freshwater fish, carp (Cyprinus carpio)
") Polyunsaturated fatty acid metabolism in a cell culture model of essential fatty acid deficiency in a freshwater fish, carp (Cyprinus carpio) Douglas R. Tocher and James R. Dick Unit of Aquatic Biochemistry,
Polyunsaturated fatty acid metabolism in a cell culture model of essential fatty acid deficiency in a freshwater fish, carp (Cyprinus carpio) Douglas R. Tocher and James R. Dick Unit of Aquatic Biochemistry,
SUBMITTED VERSION.
 SUBMITTED VERSION Wei-Chun Tu, Beverly S. Mühlhäusler, Michael J. James, David A.J. Stone, Robert A. Gibson Dietary alpha-linolenic acid does not enhance accumulation of omega-3 long-chain polyunsaturated
SUBMITTED VERSION Wei-Chun Tu, Beverly S. Mühlhäusler, Michael J. James, David A.J. Stone, Robert A. Gibson Dietary alpha-linolenic acid does not enhance accumulation of omega-3 long-chain polyunsaturated
NORDIC NATURALS NORDIC PET
 NORDIC NATURALS NORDIC PET Introducing The Nordic Pet Range Exceptional Products for Your Exceptional Pet Nordic Naturals achieves exceptional products by using fish oils that surpass stringent quality
NORDIC NATURALS NORDIC PET Introducing The Nordic Pet Range Exceptional Products for Your Exceptional Pet Nordic Naturals achieves exceptional products by using fish oils that surpass stringent quality
Effects of increasing dietary vitamin C on serum lysozyme activity and antioxidant ability of tissues in Pampus argenteus
 9 4 2 0 1 3 8 South China Fisheries Science Vol. 9, No. 4 Aug., 2013 doi: 10. 3969/ j. issn. 2095-0780. 2013. 04. 003,,, ( 200090) : C( ) ( Pampus argenteus), 104. 21 mg kg - 1 455. 33 mg kg - 1 800. 54
9 4 2 0 1 3 8 South China Fisheries Science Vol. 9, No. 4 Aug., 2013 doi: 10. 3969/ j. issn. 2095-0780. 2013. 04. 003,,, ( 200090) : C( ) ( Pampus argenteus), 104. 21 mg kg - 1 455. 33 mg kg - 1 800. 54
Part 4. Polyunsaturated Fatty Acids
 MASS SPECTRA OF PYRROLIDIE DERIVATIVES Part 4. Polyunsaturated Fatty Acids As was described for dienes, the interpretation of mass spectra of pyrrolidides is similar to that for monoenes in that it is
MASS SPECTRA OF PYRROLIDIE DERIVATIVES Part 4. Polyunsaturated Fatty Acids As was described for dienes, the interpretation of mass spectra of pyrrolidides is similar to that for monoenes in that it is
TECHNICAL INFORMATION
 MP Biomedicals, LLC 29525 Fountain Parkway Solon, Ohio 44139 TECHNICAL INFORMATION Telephone: 440/337-1200 Toll Free: 800/854-0530 Fax: 440/337-1180 mailto: biotech@mpbio.com web: http://www.mpbio.com
MP Biomedicals, LLC 29525 Fountain Parkway Solon, Ohio 44139 TECHNICAL INFORMATION Telephone: 440/337-1200 Toll Free: 800/854-0530 Fax: 440/337-1180 mailto: biotech@mpbio.com web: http://www.mpbio.com
Total lipid in the broodstock diet did not affect fatty acid composition and quality of eggs from sea bass (Dicentrarchus labrax L.
 SCI. MAR., 65 (1): 11-19 SCIENTIA MARINA 2001 Total lipid in the broodstock diet did not affect fatty acid composition and quality of eggs from sea bass (Dicentrarchus labrax L.)* JOSÉ Mª. NAVAS 1, MARK
SCI. MAR., 65 (1): 11-19 SCIENTIA MARINA 2001 Total lipid in the broodstock diet did not affect fatty acid composition and quality of eggs from sea bass (Dicentrarchus labrax L.)* JOSÉ Mª. NAVAS 1, MARK
Brett D. Glencross 1, Douglas R. Tocher 2, Chessor Matthew 3, J. Gordon Bell 2
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Effects of graded dietary docosahexaenoic acid in combination with other long-chain polyunsaturated fatty acids in post-smolt Atlantic salmon (Salmo
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Effects of graded dietary docosahexaenoic acid in combination with other long-chain polyunsaturated fatty acids in post-smolt Atlantic salmon (Salmo
Soy Protein Concentrate as a Substitute for Fishmeal in the Feed for Black Carp
 Key Words: Black carp, soy protein concentrate, soybean meal, 80:20 pond technology, China Soy Protein Concentrate as a Substitute for Fishmeal in the Feed for Black Carp INTRODUCTION Results of the 2008
Key Words: Black carp, soy protein concentrate, soybean meal, 80:20 pond technology, China Soy Protein Concentrate as a Substitute for Fishmeal in the Feed for Black Carp INTRODUCTION Results of the 2008
ALTERATIONS IN NEURAL FATTY ACID METABOLISM CAUSED BY VITAMIN E DEFICIENCY
 ALTERATIONS IN NEURAL FATTY ACID METABOLISM CAUSED BY VITAMIN E DEFICIENCY Bonnie Buckingham Faculty Mentor: Dr. Maret Traber Linus Pauling Institute HHMI Summer 2011 Outline Significance Background Hypothesis
ALTERATIONS IN NEURAL FATTY ACID METABOLISM CAUSED BY VITAMIN E DEFICIENCY Bonnie Buckingham Faculty Mentor: Dr. Maret Traber Linus Pauling Institute HHMI Summer 2011 Outline Significance Background Hypothesis
BASF Canola oil: Convenient Healthy Food for Your Heart and Brain. Andy Beadle UK Parliamentary Food and Health Forum 16th March 2010
 e BASF Canola oil: Convenient Healthy Food for Your Heart and Brain Andy Beadle UK Parliamentary Food and Health Forum 16th March 2010 Plant biotechnology at BASF BASF Plant Science Plant Science Sweden
e BASF Canola oil: Convenient Healthy Food for Your Heart and Brain Andy Beadle UK Parliamentary Food and Health Forum 16th March 2010 Plant biotechnology at BASF BASF Plant Science Plant Science Sweden
Biomolecules. Unit 3
 Biomolecules Unit 3 Atoms Elements Compounds Periodic Table What are biomolecules? Monomers vs Polymers Carbohydrates Lipids Proteins Nucleic Acids Minerals Vitamins Enzymes Triglycerides Chemical Reactions
Biomolecules Unit 3 Atoms Elements Compounds Periodic Table What are biomolecules? Monomers vs Polymers Carbohydrates Lipids Proteins Nucleic Acids Minerals Vitamins Enzymes Triglycerides Chemical Reactions
Effects of Yeast Products on Immune Function and Disease Resistance of Hybrid Striped Bass
 Effects of Yeast Products on Immune Function and Disease Resistance of Hybrid Striped Bass Peng Li and Delbert M. Gatlin, III Department of Wildlife and Fisheries Sciences and Faculty of Nutrition, Texas
Effects of Yeast Products on Immune Function and Disease Resistance of Hybrid Striped Bass Peng Li and Delbert M. Gatlin, III Department of Wildlife and Fisheries Sciences and Faculty of Nutrition, Texas
Effects of dietary lipid source on growth, digestibility and tissue fatty acid composition of Heterobranchus longifilis fingerlings
 Journal of Agriculture and Rural Development in the Tropics and Subtropics Vol. 113 No. 1 (2012) 1 11 urn:nbn:de:hebis:34-2012061541299 ISSN: 1612-9830 journal online: www.jarts.info Effects of dietary
Journal of Agriculture and Rural Development in the Tropics and Subtropics Vol. 113 No. 1 (2012) 1 11 urn:nbn:de:hebis:34-2012061541299 ISSN: 1612-9830 journal online: www.jarts.info Effects of dietary
Songpu Variety Common Carp Exhibit Rapid Growth on Soy-Based Diet in Harbin Feeding Trial
 Key Words: Common carp, soybean meal, 80:20 pond technology, China Songpu Variety Common Carp Exhibit Rapid Growth on Soy-Based Diet in Harbin Feeding Trial Michael C. Cremer, Zhou Enhua and Zhang Jian
Key Words: Common carp, soybean meal, 80:20 pond technology, China Songpu Variety Common Carp Exhibit Rapid Growth on Soy-Based Diet in Harbin Feeding Trial Michael C. Cremer, Zhou Enhua and Zhang Jian
The time course of fish oil wash-out follows a simple dilution model in gilthead sea
 * Manuscript 1 1 2 The time course of fish oil wash-out follows a simple dilution model in gilthead sea bream (Sparus aurata L.) fed graded levels of vegetable oils 3 4 5 Laura Benedito-Palos a, Juan C.
* Manuscript 1 1 2 The time course of fish oil wash-out follows a simple dilution model in gilthead sea bream (Sparus aurata L.) fed graded levels of vegetable oils 3 4 5 Laura Benedito-Palos a, Juan C.
Grass Carp Exhibit Excellent Growth and Feed Conversion on Cost Efficient, Soy-Based Diet
 Key Words: Grass carp, soybean meal, soy hulls, 80:20 pond technology, China Grass Carp Exhibit Excellent Growth and Feed Conversion on Cost Efficient, Soy-Based Diet Michael C. Cremer, Zhou Enhua and
Key Words: Grass carp, soybean meal, soy hulls, 80:20 pond technology, China Grass Carp Exhibit Excellent Growth and Feed Conversion on Cost Efficient, Soy-Based Diet Michael C. Cremer, Zhou Enhua and
Requirements of Prawn, Penaeus japonicus, for Essential Fatty Acids
 Mem. Fac. Fish., Kagoshima Univ. Vol. 28 pp. 27-33 (1979) Requirements of Prawn, Penaeus japonicus, for Essential Fatty Acids Akio Kanazawa, Shin-ichi Teshima and Minoru Endo* Abstract Feeding trials using
Mem. Fac. Fish., Kagoshima Univ. Vol. 28 pp. 27-33 (1979) Requirements of Prawn, Penaeus japonicus, for Essential Fatty Acids Akio Kanazawa, Shin-ichi Teshima and Minoru Endo* Abstract Feeding trials using
Rapid Analysis of 37 FAMEs with the Agilent 8860 Gas Chromatograph
 Application Note Food Rapid Analysis of 37 FAMEs with the Agilent 88 Gas Chromatograph Author Youjuan Zhang Agilent Technologies (Shanghai) Co. Ltd., Shanghai 131 P. R. China Abstract An Agilent 88 GC
Application Note Food Rapid Analysis of 37 FAMEs with the Agilent 88 Gas Chromatograph Author Youjuan Zhang Agilent Technologies (Shanghai) Co. Ltd., Shanghai 131 P. R. China Abstract An Agilent 88 GC
INTRODUCING NEW PUPPY & KITTEN FORMULAS GLUTEN FREE. For a lifetime of happiness
 INTRODUCING NEW PUPPY & KITTEN FORMULAS GLUTEN FREE For a lifetime of happiness NOURISH YOUR LITTLE ONE FROM HEAD TO TAIL! Specially selected shape for smaller jaws. Product not to scale. PU P DRY FOR
INTRODUCING NEW PUPPY & KITTEN FORMULAS GLUTEN FREE For a lifetime of happiness NOURISH YOUR LITTLE ONE FROM HEAD TO TAIL! Specially selected shape for smaller jaws. Product not to scale. PU P DRY FOR
Comparative Effects of Dietary Fat Manipulation on Fatty Acid Composition of Rat Stomach, Jejunum, and Colon Phospholipids
 J. Clin. Biochem. Nutr., 22, 101-111, 1997 Comparative Effects of Dietary Fat Manipulation on Fatty Acid Composition of Rat Stomach, Jejunum, and Colon Phospholipids Manohar GARG,* Robert BLAKE, and Brenda
J. Clin. Biochem. Nutr., 22, 101-111, 1997 Comparative Effects of Dietary Fat Manipulation on Fatty Acid Composition of Rat Stomach, Jejunum, and Colon Phospholipids Manohar GARG,* Robert BLAKE, and Brenda
Pallab K. Sarker, Ph.D. 1*, Anne R. Kapuscinski, Ph.D. 1, Alison J. Lanois 1, Erin D. Livesey 1, Katie P. Bernhard 1, Mariah L.
 Towards sustainable aquafeeds: Evaluating three marine microalgae for replacement of fish oil and fishmeal in aquaculture diets for Nile tilapia (Oreochromis niloticus) and rainbow trout (Oncorhynchus
Towards sustainable aquafeeds: Evaluating three marine microalgae for replacement of fish oil and fishmeal in aquaculture diets for Nile tilapia (Oreochromis niloticus) and rainbow trout (Oncorhynchus
Effects of dietary levels of 20:5n 3 and 22:6n 3 on tissue lipid composition in juvenile Atlantic salmon, Salmo salar, with emphasis on brain and eye
 Aquaculture Nutrition 1997 3; 175 187 Effects of dietary levels of 20:5n 3 and 22:6n 3 on tissue lipid composition in juvenile Atlantic salmon, Salmo salar, with emphasis on brain and eye T. BRODTKORB
Aquaculture Nutrition 1997 3; 175 187 Effects of dietary levels of 20:5n 3 and 22:6n 3 on tissue lipid composition in juvenile Atlantic salmon, Salmo salar, with emphasis on brain and eye T. BRODTKORB
Growth Performance of Tilapia Fed Soy-Based Feed in Low. Volume, High Density Cages on Phu Long Reservoir, Dalai, Ninh
 Growth Performance of Tilapia Fed Soy-Based Feed in Low Volume, High Density Cages on Phu Long Reservoir, Dalai, Ninh Binh, Yen Khanh District, Vietnam Results of ASA-IM/Soy-in-Aquaculture 2006 Feeding
Growth Performance of Tilapia Fed Soy-Based Feed in Low Volume, High Density Cages on Phu Long Reservoir, Dalai, Ninh Binh, Yen Khanh District, Vietnam Results of ASA-IM/Soy-in-Aquaculture 2006 Feeding
STANDARD FOR FISH OILS CODEX STAN Adopted in 2017.
 STANDARD FOR FISH OILS CODEX STAN 329-2017 Adopted in 2017. CODEX STAN 329-2017 2 1. SCOPE This Standard applies to the fish oils described in section 2 that are presented in a state for human consumption.
STANDARD FOR FISH OILS CODEX STAN 329-2017 Adopted in 2017. CODEX STAN 329-2017 2 1. SCOPE This Standard applies to the fish oils described in section 2 that are presented in a state for human consumption.
NEW! 200 m GC Columns for Detailed Analysis of cis/trans FAME Isomers
 Order: 00--00 (U.S.) -- (Global) NEW! 00 m GC Columns for Detailed Analysis of cis/trans FAME Isomers Leonard M. Sidisky, R&D Manager; and Michael D. Buchanan, Product Manager mike.buchanan@sial.com Over
Order: 00--00 (U.S.) -- (Global) NEW! 00 m GC Columns for Detailed Analysis of cis/trans FAME Isomers Leonard M. Sidisky, R&D Manager; and Michael D. Buchanan, Product Manager mike.buchanan@sial.com Over
Iranian Journal of Fisheries Sciences 13(4)
") Iranian Journal of Fisheries Sciences 13(4) 796-809 2014 Impacts of different dietary lipid sources on growth performance, fatty acid composition and antioxidant enzyme activity of juvenile Black Sea bream,
Iranian Journal of Fisheries Sciences 13(4) 796-809 2014 Impacts of different dietary lipid sources on growth performance, fatty acid composition and antioxidant enzyme activity of juvenile Black Sea bream,
This is the peer reviewed version of this article
 Betancor M, Howarth FJE, Glencross BD & Tocher DR (2014) Influence of dietary docosahexaenoic acid in combination with other long-chain polyunsaturated fatty acids on expression of biosynthesis genes and
Betancor M, Howarth FJE, Glencross BD & Tocher DR (2014) Influence of dietary docosahexaenoic acid in combination with other long-chain polyunsaturated fatty acids on expression of biosynthesis genes and
Linseed oils with different fatty acid patterns in the diet of broiler chickens
 Linseed oils with different fatty acid patterns in the diet of broiler chickens J. ZELENKA, D. SCHNEIDEROVÁ, E. MRKVICOVÁ Faculty of Agronomy, Mendel University of Agriculture and Forestry, Brno, Czech
Linseed oils with different fatty acid patterns in the diet of broiler chickens J. ZELENKA, D. SCHNEIDEROVÁ, E. MRKVICOVÁ Faculty of Agronomy, Mendel University of Agriculture and Forestry, Brno, Czech
Do pigs benefit from omega-3 fatty acids?
 Do pigs benefit from omega-3 fatty acids? Denise Beaulieu Assistant Professor Animal & Poultry Science Introduction What are omega-3 fatty acids? Outline Why would we consider augmenting the diet of growing
Do pigs benefit from omega-3 fatty acids? Denise Beaulieu Assistant Professor Animal & Poultry Science Introduction What are omega-3 fatty acids? Outline Why would we consider augmenting the diet of growing
The lipid profile of the pallid emperor moth Cirina forda Westwood (Lepidoptera: Saturniidae) caterpillar
 caterpillar") BIOKEMISTRI 13: 37-41 (January 2003) Printed in Nigeria The lipid profile of the pallid emperor moth Cirina forda Westwood (Lepidoptera: Saturniidae) caterpillar Adeolu.T. ANDE Department of Biological
BIOKEMISTRI 13: 37-41 (January 2003) Printed in Nigeria The lipid profile of the pallid emperor moth Cirina forda Westwood (Lepidoptera: Saturniidae) caterpillar Adeolu.T. ANDE Department of Biological
Is it possible to influence European sea bass (Dicentrarchus labrax) juvenile metabolism by a nutritional conditioning during larval stage?
 juvenile metabolism by a nutritional conditioning during larval stage?") Please note that this is an author-produced PDF of an article accepted for publication following peer review. The definitive publisher-authenticated version is available on the publisher Web site Aquaculture
Please note that this is an author-produced PDF of an article accepted for publication following peer review. The definitive publisher-authenticated version is available on the publisher Web site Aquaculture
Does dietary n 3 highly unsaturated fatty acids level influence the European sea bass (Dicentrachus labrax) capacity to adapt to a high temperature?
 capacity to adapt to a high temperature?") hor-produced PDF of an article accepted for publication following peer review. The definitive publisher-authenticated version is available on the publisher Web site Please note that this is an aut Aquaculture
hor-produced PDF of an article accepted for publication following peer review. The definitive publisher-authenticated version is available on the publisher Web site Please note that this is an aut Aquaculture
Illinois Soybean Association-Supported Aquaculture Research at SIUC
 Illinois Soybean Association-Supported Aquaculture Research at SIUC Project Updates for Trushenski Research Team CHALLENGES AND OPPORTUNITIES IN AQUACULTURE 1 Aquaculture, the rearing of aquatic plants
Illinois Soybean Association-Supported Aquaculture Research at SIUC Project Updates for Trushenski Research Team CHALLENGES AND OPPORTUNITIES IN AQUACULTURE 1 Aquaculture, the rearing of aquatic plants
Forages, fat, fitness and flavour
 Forages, fat, fitness and flavour Richard Dewhurst and Nigel Scollan Fatty acids in grasses Fatty acids in grass silage Fatty acids and beef Effects of fatty acids on meat appearance and flavour Richard
Forages, fat, fitness and flavour Richard Dewhurst and Nigel Scollan Fatty acids in grasses Fatty acids in grass silage Fatty acids and beef Effects of fatty acids on meat appearance and flavour Richard
Innovations in Soy for Aquaculture. Michael C. Cremer, Ph.D. Aquaculture Utilization Director
 Innovations in Soy for Aquaculture Michael C. Cremer, Ph.D. Aquaculture Utilization Director Forecast: Need to double aquaculture production in a decade Challenge: Identify economically viable and sustainable
Innovations in Soy for Aquaculture Michael C. Cremer, Ph.D. Aquaculture Utilization Director Forecast: Need to double aquaculture production in a decade Challenge: Identify economically viable and sustainable
NUTRITIONAL QUALITY OF EGGS FROM HENS FED WITH DDGS
 NUTRITIONAL QUALITY OF EGGS FROM HENS FED WITH Yan Zhang National Corn to Ethanol Research Center Southern Illinois University Edwardsville and Jill K. Winkler-Moser National Center for Agricultural Utilization
NUTRITIONAL QUALITY OF EGGS FROM HENS FED WITH Yan Zhang National Corn to Ethanol Research Center Southern Illinois University Edwardsville and Jill K. Winkler-Moser National Center for Agricultural Utilization
LABORATORY REPORT SAMPLE. Summary of Deficient Test Results. Vitamin B12 Pantothenate Biotin Spectrox Immunidex
 LABORATORY REPORT Account Number: Name: Jon Doe Gender: Male DOB: 12/28/1984 Requisition Number: Summary of Deficient Test Results Testing determined the following functional deficiencies: Oleic Acid Borderline
LABORATORY REPORT Account Number: Name: Jon Doe Gender: Male DOB: 12/28/1984 Requisition Number: Summary of Deficient Test Results Testing determined the following functional deficiencies: Oleic Acid Borderline
Effects of Dietary Arachidonic Acid (20:4n-6) Levels on Growth Performance and Fatty Acid Composition of Juvenile Eel, Anguilla japonica
 Levels on Growth Performance and Fatty Acid Composition of Juvenile Eel, Anguilla japonica") 508 Asian-Aust. J. Anim. Sci. Vol. 23, No. 4 : 508-514 April 2010 www.ajas.info Effects of Dietary Arachidonic Acid (20:4n-6) Levels on Growth Performance and Fatty Acid Composition of Juvenile Eel, Anguilla
508 Asian-Aust. J. Anim. Sci. Vol. 23, No. 4 : 508-514 April 2010 www.ajas.info Effects of Dietary Arachidonic Acid (20:4n-6) Levels on Growth Performance and Fatty Acid Composition of Juvenile Eel, Anguilla
Growth Performance of Common Carp Fed Soy-Maximized Feed in. Low Volume, High Density Cages on Lake Maninjau, Indonesia
 Growth Performance of Common Carp Fed Soy-Maximized Feed in Low Volume, High Density Cages on Lake Maninjau, Indonesia Results of ASA/Soy-in-Aquaculture 2004 Feeding Trial Lukas Manomaitis and Michael
Growth Performance of Common Carp Fed Soy-Maximized Feed in Low Volume, High Density Cages on Lake Maninjau, Indonesia Results of ASA/Soy-in-Aquaculture 2004 Feeding Trial Lukas Manomaitis and Michael
ESSENTIAL FATTY ACIDS - RED CELL
 Healthscope Functional Pathology 1868 Dandenong Road, Clayton, Victoria 3168, Australia www.functionalpathology.com.au ABN 62 006 823 089 T 1300 55 44 80 F 03 8540 5555 infofp@healthscope.com.au LAB No:
Healthscope Functional Pathology 1868 Dandenong Road, Clayton, Victoria 3168, Australia www.functionalpathology.com.au ABN 62 006 823 089 T 1300 55 44 80 F 03 8540 5555 infofp@healthscope.com.au LAB No:
Improving the Analysis of 37 Fatty Acid Methyl Esters
 Application Note Food Testing Improving the Analysis of 37 Fatty Acid Methyl Esters Using Three Types of Capillary GC columns Authors Yun Zou Agilent Technologies (Shanghai) Co.Ltd, Shanghai 200131 P.R.China
Application Note Food Testing Improving the Analysis of 37 Fatty Acid Methyl Esters Using Three Types of Capillary GC columns Authors Yun Zou Agilent Technologies (Shanghai) Co.Ltd, Shanghai 200131 P.R.China
and alcohols in caecal enterocytes of Atlantic salmon (Salmo
 Utilization and metabolism of palmityl and oleoyl fatty acids and alcohols in caecal enterocytes of Atlantic salmon (Salmo salar L.) A.S. BOGEVIK 1, 3*, R.E. OLSEN 1 and D.R. TOCHER 2 1 Institute of Marine
Utilization and metabolism of palmityl and oleoyl fatty acids and alcohols in caecal enterocytes of Atlantic salmon (Salmo salar L.) A.S. BOGEVIK 1, 3*, R.E. OLSEN 1 and D.R. TOCHER 2 1 Institute of Marine
Differentiating omega-3 fatty acids from SPMs (specialized pro-resolving lipid mediators)
") NIHR Southampton Biomedical Research Centre in nutrition Differentiating omega-3 fatty acids from SPMs (specialized pro-resolving lipid mediators) Philip Calder Professor of Nutritional Immunology University
NIHR Southampton Biomedical Research Centre in nutrition Differentiating omega-3 fatty acids from SPMs (specialized pro-resolving lipid mediators) Philip Calder Professor of Nutritional Immunology University
Guangzhou Pond Feeding Trial Demonstrates Channel Catfish Production with Soy-Based Feed
 Guangzhou Pond Feeding Trial Demonstrates Channel Catfish Production with Soy-Based Feed INTRODUCTION Michael C. Cremer, Zhou Enhua and Zhang Jian ASA-IM/China Aquaculture Program A feeding trial was conducted
Guangzhou Pond Feeding Trial Demonstrates Channel Catfish Production with Soy-Based Feed INTRODUCTION Michael C. Cremer, Zhou Enhua and Zhang Jian ASA-IM/China Aquaculture Program A feeding trial was conducted
COMPLETE FOOD FOR DOGS. LOVING your pet starts with nutrition!
 COMPLETE FOOD FOR DOGS LOVING your pet starts with nutrition! Health vitality and a long life do not solely depend on an ideal nutrition. At TRISANA we firmly believe: Keep as close as possible to Nature!
COMPLETE FOOD FOR DOGS LOVING your pet starts with nutrition! Health vitality and a long life do not solely depend on an ideal nutrition. At TRISANA we firmly believe: Keep as close as possible to Nature!
Pet Food Kibble Preservation 2018 Jim Mann
 Pet Food Kibble Preservation 2018 Jim Mann KEMIN IS ASSURANCE Kemin s Involvement in Pet Food Supply Chain Slaughter Houses Renderers Field Fresh/Frozen Meat Collectors Crops KIBBLE STABILITY PALATABILITY
Pet Food Kibble Preservation 2018 Jim Mann KEMIN IS ASSURANCE Kemin s Involvement in Pet Food Supply Chain Slaughter Houses Renderers Field Fresh/Frozen Meat Collectors Crops KIBBLE STABILITY PALATABILITY
de La Laguna, La Laguna, 38206, Spain Key Words: Sparus aurata, development, Echium oil, GLA, SDA, body lipids.
 Aquaculture Nutrition Effect of dietary substitution of fish oil by Echium oil on growth, plasma parameters and body lipid composition in gilthead seabream (Sparus aurata L.). 1 Mercedes Díaz-López, 1
Aquaculture Nutrition Effect of dietary substitution of fish oil by Echium oil on growth, plasma parameters and body lipid composition in gilthead seabream (Sparus aurata L.). 1 Mercedes Díaz-López, 1
Feed Efficiency, Fish Performance. Rick Decker AquaMax Fish Diets
 Feed Efficiency, Fish Performance & Water Clarity Rick Decker AquaMax Fish Diets Fish Nutrition Why feed fish? Why is nutrition important? What nutrients are essential? How are nutrients determined in
Feed Efficiency, Fish Performance & Water Clarity Rick Decker AquaMax Fish Diets Fish Nutrition Why feed fish? Why is nutrition important? What nutrients are essential? How are nutrients determined in
Fish oils and human diet
 British Journal of Nutrition (1997), 78, Suppl. 1, S5-SI3 Fish oils and human diet BY J. R. SARGENT NERC Unit of Aquatic Biochemistry, Department of Biological and Molecular Sciences, University of Stirling,
British Journal of Nutrition (1997), 78, Suppl. 1, S5-SI3 Fish oils and human diet BY J. R. SARGENT NERC Unit of Aquatic Biochemistry, Department of Biological and Molecular Sciences, University of Stirling,
Performance of Pompano Fed Soy-Optimized, Extruded Feed Using. ASA-IM Low Volume High Density Cages in Vung Tau Province, Vietnam
 Performance of Pompano Fed Soy-Optimized, Extruded Feed Using ASA-IM Low Volume High Density Cages in Vung Tau Province, Vietnam Results of ASA-IM/Soy-in-Aquaculture 2007 Feeding Demonstration Project
Performance of Pompano Fed Soy-Optimized, Extruded Feed Using ASA-IM Low Volume High Density Cages in Vung Tau Province, Vietnam Results of ASA-IM/Soy-in-Aquaculture 2007 Feeding Demonstration Project
Insects in fish feeding. Erik-Jan Lock
 Insects in fish feeding Erik-Jan Lock Finding new feed resources Volumes/price ABP Plant Krill Sustainability/responsible sourcing/ Microalgae Seaweeds Insects SCP Blue mussels Approach Insect as natural
Insects in fish feeding Erik-Jan Lock Finding new feed resources Volumes/price ABP Plant Krill Sustainability/responsible sourcing/ Microalgae Seaweeds Insects SCP Blue mussels Approach Insect as natural