Involvement of purinergic P2X and P2Y2 receptors in urinary bladder sensation
|
|
|
- Gyles Gibson
- 6 years ago
- Views:
Transcription

 thesis, University of Iowa, 2009. http://ir.uiowa.edu/etd/343. Follow this and additional works at: http://ir.uiowa.edu/etd Part of the Neuroscience and Neurobiology Commons")
1 University of Iowa Iowa Research Online Theses and Dissertations Fall 2009 Involvement of purinergic P2X and P2Y2 receptors in urinary bladder sensation Xiaowei Chen University of Iowa Copyright 2009 Xiaowei Chen This dissertation is available at Iowa Research Online: Recommended Citation Chen, Xiaowei. "Involvement of purinergic P2X and P2Y2 receptors in urinary bladder sensation." PhD (Doctor of Philosophy) thesis, University of Iowa, Follow this and additional works at: Part of the Neuroscience and Neurobiology Commons
2 INVOLVEMENT OF PURINERGIC P2X AND P2Y2 RECEPTORS IN URINARY BLADDER SENSATION by Xiaowei Chen An Abstract Of a thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Neuroscience in the Graduate College of The University of Iowa December 2009 Thesis Supervisor: Professor Emeritus G. F. Gebhart
3 1 ABSTRACT Interstitial cystitis (IC)/painful bladder syndrome (PBS) is a functional visceral disorder characterized by increased bladder activity and chronic pelvic pain in the absence of a pathobiological condition. Enhanced sensory transduction of peripheral bladder afferents is hypothesized to contribute to the pain and mechanical hypersensitivity of IC/PBS patients. The aim of this thesis is to test the hypothesis that purinergic receptors, including ionotropic P2X and metabotropic P2Y, are important for sensory transmission in bladder afferent neurons and may be involved in bladder hypersensitivity after bladder tissue insults. Electrophysiological, single cell RT-PCR and immunohistochemistry techniques were performed in bladder afferent neurons from naïve and bladder inflamed mice to test the hypothesis. In Chapter 2, I characterized the distribution and function of P2X receptors in thoracolumbar (TL) and lumbosacral (LS) dorsal root ganglia (DRG) neurons innervating the urinary bladder, and found that LS and TL bladder neurons have differential purinergic signaling and distinct membrane electrical properties. In Chapter 3, I examined the sensitization of bladder afferent neurons and the plasticity of P2X receptor function in a mouse model of chemical induced bladder inflammation. P2X-mediated signals in LS and TL bladder neurons after bladder inflammation were enhanced compared with those in saline-treated controls, suggesting the importance of P2X in bladder hypersensitivity associated with cystitis. In Chapter 4, the modulation of P2Y on P2X function and the colocalization of P2Y and P2X were examined in bladder sensory neurons. It has been found that P2Y2 receptor enhances bladder sensory neuron excitability and
4 2 facilitates the response of homomeric P2X2 receptor to the purinergic agonist (ATP). The present study provides evidence that LS and TL mouse bladder sensory neurons exhibit distinct P2X signaling, and the function of P2X receptors could be facilitated during bladder inflammation and modulated by activation of P2Y2 receptor, indicating an involvement of P2X and P2Y2 receptors as mechano- and chemosensors in bladder sensory transmission under normal conditions and in bladder hypersensitivity associated with inflammation. Abstract Approved: Thesis Supervisor Title and Department Date
5 INVOLVEMENT OF PURINERGIC P2X AND P2Y2 RECEPTORS IN URINARY BLADDER SENSATION by Xiaowei Chen A thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Neuroscience in the Graduate College of The University of Iowa December 2009 Thesis Supervisor: Professor Emeritus G. F. Gebhart
6 Graduate College The University of Iowa Iowa City, Iowa CERTIFICATE OF APPROVAL PH.D. THESIS This is to certify that the Ph.D. thesis of Xiaowei Chen has been approved by the Examining Committee for the thesis requirement for the Doctor of Philosophy degree in Neuroscience at the December 2009 graduation. Thesis Committee: G. F. Gebhart, Thesis Supervisor Christopher Benson Timothy Brennan Michael O Donnell Yuriy M. Usachev
7 ACKNOWLEDGMENTS I would like to express my thanks to many people who provided assistance in numerous ways to make this thesis possible. It is difficult to overstate my gratitude to my mentor Dr. Gerald F. Gebhart for his invaluable research guidance and friendly support during my graduate study. I greatly appreciated his inspiring input on research and scientific training on my thoughts. With his enthusiasm, his inspiration, and his sense of humor, he has taught me not only science but also a positive attitude to life. To me, he serves as a model of being an outstanding scientist and virtuous person. I would like to thank all the members of the Gebhart Laboratory who provided a great research environment and valuable advice on my research. I wish to thank Khoa Dang, Jun-ho La, Pablo Brumovsky, Masamichi Shinoda and Dave Robinson for assistance with patch-clamp technique, immunostaining, histological assessment and MPO assay. I would like to thank Dr. Derek Molliver for his sound advice on my research project and providing P2Y2 knockout mice used in the experiments described in Chapter 4. I wish to thank Dr. Michael Gold and Liming Fan for their assistance with single cell RT-PCR, Dr. Julie Christianson for her help with MPO assay, Joanne Hirt for secretarial assistance and Michael Burcham for preparation of the figures. Without the help of the people mentioned above, I am not able to complete this thesis. I am grateful to members of the Pain Interest Group at the University of Iowa and the Center for Pain Research at the University of Pittsburgh for their intellectual contribution to work described in this thesis and my scientific ii
8 development. Interactions with these intelligent people have improved my knowledge in the field of pain research as well as the experimental design and interpretation of experimental results. The members of my thesis committee have provided extensive support and effect on my project and their suggestions helped me greatly improve my research work. Lastly, I want to express my gratitude to my husband, Qiyun. I am indebted to him for his understand and sacrifice. His encouragement and comfort helped me through the anxiety and frustration when experiments were not going well. I also wish to thank my family for their selfless love and support of my dream. Without the courage they gave me, I would not be able to pursuit a Ph.D. degree abroad. To them I dedicate this thesis. iii
9 ABSTRACT Interstitial cystitis (IC)/painful bladder syndrome (PBS) is a functional visceral disorder characterized by increased bladder activity and chronic pelvic pain in the absence of a pathobiological condition. Enhanced sensory transduction of peripheral bladder afferents is hypothesized to contribute to the pain and mechanical hypersensitivity of IC/PBS patients. The aim of this thesis is to test the hypothesis that purinergic receptors, including ionotropic P2X and metabotropic P2Y, are important for sensory transmission in bladder afferent neurons and may be involved in bladder hypersensitivity after bladder tissue insults. Electrophysiological, single cell RT-PCR and immunohistochemistry techniques were performed in bladder afferent neurons from naïve and bladder inflamed mice to test the hypothesis. In Chapter 2, I characterized the distribution and function of P2X receptors in thoracolumbar (TL) and lumbosacral (LS) dorsal root ganglia (DRG) neurons innervating the urinary bladder, and found that LS and TL bladder neurons have differential purinergic signaling and distinct membrane electrical properties. In Chapter 3, I examined the sensitization of bladder afferent neurons and the plasticity of P2X receptor function in a mouse model of chemical induced bladder inflammation. P2X-mediated signals in LS and TL bladder neurons after bladder inflammation were enhanced compared with those in saline-treated controls, suggesting the importance of P2X in bladder hypersensitivity associated with cystitis. In Chapter 4, the modulation of P2Y on P2X function and the colocalization of P2Y and P2X were examined in bladder sensory neurons. It has been found that P2Y2 receptor enhances bladder sensory neuron excitability and iv
10 facilitates the response of homomeric P2X2 receptor to the purinergic agonist (ATP). The present study provides evidence that LS and TL mouse bladder sensory neurons exhibit distinct P2X signaling, and the function of P2X receptors could be facilitated during bladder inflammation and modulated by activation of P2Y2 receptor, indicating an involvement of P2X and P2Y2 receptors as mechano- and chemosensors in bladder sensory transmission under normal conditions and in bladder hypersensitivity associated with inflammation. v
11 TABLE OF CONTENTS LIST OF TABLES...ix LIST OF FIGURES... x CHAPTER 1: GENERAL INTRODUCTION... 1 The urinary bladder sensation... 1 A functional bladder disorder of visceral pain: Interstitial cystitis/ painful bladder syndrome... 5 Contribution of purinergic signaling to bladder sensation and pain... 7 Ionotropic P2X receptors... 8 Metabotropic P2Y receptors Thesis objectives CHAPTER 2: CHARACTERIZATION OF PURINERGIC P2X RECEPTORS IN LUMBOSACRAL (LS) AND THORACOLUMBAR (TL) BLADDER SENSORY NEURONS Introduction Results Cell density and size distribution of bladder sensory neurons Characterization of bladder sensory neuron responses to purinergic receptor agonists Characterization of P2X receptor subtypes in bladder sensory neurons Electrophysiological properties of LS and TL bladder sensory neurons P2X receptor expression in bladder sensory neurons Discussion Differences between LS and TL bladder sensory neurons CHAPTER 3: PURINERGIC P2X SIGNALING IN BLADDER SENSORY NEURONS AFTER BLADDER INFLAMMATION Introduction Results Bladder inflammation and tissue damage after CYP treatment Bladder sensory neuron excitability increases after CYP treatment P2X receptor mediated currents after CYP treatment vi
12 P2X receptor expression in bladder sensory neurons after CYP treatment Discussion CYP treatment induces moderate bladder inflammation Bladder sensory neurons exhibit increased cell excitability after bladder inflammation P2X function in bladder afferent neurons is enhanced after bladder inflammation P2X expression in bladder sensory neurons after bladder inflammation CHAPTER 4: EFFECT OF METABOTROPIC P2Y2 RECEPTOR ON BLADDER SENSORY NEURON EXITABILITY AND P2X RECEPTOR FUNCTION Introduction Results UTP increases bladder sensory neuron excitability Effect of UTP on purinergic agonist-evoked responses Metabotropic P2Y2 receptor mediates the effect of UTP P2X and P2Y receptor expression in bladder sensory neurons Discussion CHAPTER 5: GENERAL CONCLUSIONS AND DISCUSSION Overview of experiment results Differential purinergic signaling in LS and TL bladder sensory neurons Contribution of P2X2 and P2X3 receptors to bladder sensory transmission Contribution of P2Y2 receptor to bladder sensory transmission Future directions CHAPTER 6: MATERIALS AND METHODS Animals Bladder neuron retrograde labeling Cell dissociation and culturing Whole cell current- and voltage-patch clamp recording Urinary bladder inflammation Bladder myeloperoxidase (MPO) assay Histological examination of bladder inflammation Single cell RT-PCR vii
13 Multiplex PCR and gene specific nested PCR Immunohistochemistry Data and statistical analysis REFERENCES viii
14 LIST OF TABLES Table 1. Purinergic currents in LS and TL bladder sensory neurons from naïve mice Table 2. Properties of purinergic currents in LS and TL bladder sensory neurons from naive mice Table 3. Passive and active electrical properties of LS and TL bladder sensory neurons Table 4. Summary of P2X3 immunoreactivity in LS and TL bladder DRG neurons from naïve, saline- and CYP-treated mice Table 5. Passive and active electrical properties of LS and TL bladder neurons from saline- and CYP-treated mice Table 6. Purinergic currents in LS and TL bladder sensory neurons from saline- and CYP-treated mice Table 7. Properties of purinergic currents in LS and TL bladder sensory neurons from saline- and CYP-treated mice Table 8. Passive and active electrical properties of LS bladder neurons in the absence and the presence of UTP or UTP and suramin (SUR) application Table 9. Summary of contribution of P2X and P2Y receptors to mouse bladder sensation Table 10. External and internal primers for mouse P2X2, P2X3, P2Y2, P2Y4 and GAPDH cdna ix
15 LIST OF FIGURES Figure 1. The density of bladder pelvic and hypogastric innervations in cultured lumbosacral (LS) and thoracolumbar (TL) mouse DRG neurons Figure 2. Comparison of cell size (capacitance) of LS and TL mouse bladder sensory neurons Figure 3. Examples of principal purinergic currents in LS and TL bladder sensory neurons in response to ATP (30μM) and αβ-met ATP (30μM) Figure 4. Antagonism of purinergic agonist-evoked sustained currents Figure 5. Antagonism of purinergic agonist-evoked slow desensitizing currents Figure 6. Antagonism of purinergic agonist-evoked fast desensitizing currents Figure 7. Examples of principal purinergic current in P2X3-/- mice Figure 8. Examples of bladder sensory neuron responses to current injection and agonist application Figure 9. Single cell nested RT-PCR of P2X2 and P2X3 receptor subunits in LS and TL bladder neurons Figure 10. Immunohistochemistry of P2X3 subunit in bladder sensory neurons from naïve mice Figure 11. Bladder weight and bladder myeloperoxidase (MPO) activity in naïve saline- and CYP-treated mice Figure 12. Histological assessment of bladder inflammation in saline- and CYP- treated mice Figure 13. Examples of responses of bladder sensory neurons to current injection and agonist application in saline- and CYP- treated mice Figure 14. Single cell nested RT-PCR of P2X2 and P2X3 receptor subunits in LS and TL bladder neurons from saline- and CYPtreated mice x
16 Figure 15. Immunohistochemistry of P2X3 subunit in bladder sensory neurons from saline-treated and CYP-treated mice Figure 16. UTP increases the excitability of bladder sensory neurons Figure 17. Examples of the effect of UTP on LS bladder neuron responses to ATP Figure 18. Recovery kinetics of ATP-evoked sustained, slow and fast currents in LS bladder neurons Figure 19. The effect of UTP on ATP-evoked sustained, slow and fast currents in LS and TL bladder neurons Figure 20. The effect of repeated ATP application without UTP on ATPevoked sustained, slow and fast currents in LS and TL bladder neurons Figure 21. The dose-response relationship of ATP evoked-sustained currents in response to UTP in LS bladder neurons Figure 22. The facilitatory effect of UTP on ATP-evoked sustained currents but not fast currents is mediated by G protein-coupled P2Y2 receptor through a PKC dependent pathway Figure 23. The effect of UTP/ATP on bladder sensory neurons containing intracellular GDP-β-S, in presence of PKC inhibitor and from P2Y2 knockout mice Figure 24. Single cell nested RT-PCR of P2X2, P2X3, P2Y2 and P2Y4 receptor subunits in LS and TL bladder neurons xi
17 1 CHAPTER 1: GENERAL INTRODUCTION The urinary bladder sensation The urinary bladder is a hollow, muscular and distensible organ for temporary storage and periodic elimination of urine(alvarez et al., 2008). The sensory input of urinary bladder is requisite for conscious bladder control and normal bladder function, harmonizing compliance and excitation during the urine storage phase and the voiding reflex. Sensory information from urinary bladder travels to the central nervous system through two distinct pathways: the hypogastric/lumbar splanchnic nerves (LSN) and the pelvic nerves (PN). Retrograde tracing studies revealed thoracolumbar (hypogastric) and lumbosacral (pelvic) dorsal root ganglia (DRG) as the primary source of the bladder afferent innervation (Applebaum et al., 1980). These two sensory pathways are suggested to have some similar, but also different functions. Anatomically, the urinary bladder is divided into body and trigone (base) region, consisting of several layers: serosa, muscularis, submucosa (lamina propria), and mucosa (urothelium). Immunohistochemical staining of sensory neuronal markers in the bladder, e.g., calcitonin gene-related peptide (CGRP), substance P (SP) and TRPV1, has demonstrated that the bladder afferent fibers are distributed throughout the bladder wall and the trigone region, and project into the suburothelial lamina propria, urothelium, and detrusor smooth muscle (Gabella and Davis, 1998;Uemura et al., 1975;Uemura et al., 1973) [for review, see (Andersson, 2002)]. It is suggested that the hypogastric and the pelvic bladder afferent nerve terminals may not distribute uniformly in the bladder.
18 2 According to a series studies in the cat, the pelvic nerve axons are evenly distributed throughout the body and trigone regions, whereas axons from the lumbar innervations are more abundant in the trigone region (Uemura et al., 1973;Uemura et al., 1975). A recent study using single-fiber recording technique on bladder-never preparation provides comparable outcomes in the mouse that the pelvic mechanoceptive endings are distributed throughout the bladder, whereas those of hypogastric nerves are concentrated at the base of the bladder (Xu and Gebhart, 2008). The bladder afferents have been revealed to consist of small myelinated Aδ- and unmyelinated C-fiber afferents identified in both PN and LSN by microscopy studies, and the unmyelinated C-fibers are the predominant afferent fibers innervating the bladder (Gabella and Davis, 1998;Uvelius and Gabella, 1998;Floyd and Lawrenson, 1979;Floyd et al., 1976). Three major functional types of bladder afferents have been reported, including mechanosensitive, chemosensitive and silent afferents. The majority (~60%) of bladder pelvic afferents are mechanoreceptors sensing bladder distension (Shea et al., 2000), and their firing activities are correlated with the extent and duration of bladder distension (Andersson and Wein, 2004;Habler et al., 1993). Compared with the mechanosensitive afferents, the bladder chemoreceptive afferents that respond to hypertonic solution, purinergic agonist α,β-methylene ATP, or capsaicin are rare (<10%) in the pelvic nerves (Shea et al., 2000;Moss et al., 1997;Zagorodnyuk et al., 2007). A population (~30%) of bladder afferents that do not respond to either mechanical or chemical stimuli under normal conditions are
19 3 identified as the silent afferent in pelvic nerves (Shea et al., 2000), but the silent fibers could be activated by irritant chemical stimuli or during bladder inflammation (Habler et al., 1990;Yoshimura and de Groat, 1999). Therefore, the bladder silent afferents are presumed to play an important role in bladder afferent hyperexcitability and bladder overactivity associated with urinary urgency, frequency and pain. Although information about the functional properties of hypogastric/lumbar splanchnic bladder afferents is inadequate relative to pelvic bladder afferents, previous studies suggest that bladder hypogastric/lumbar splanchnic and pelvic afferents may be functionally distinct. Moss et al. found that a large proportion of hypogastric bladder afferents are chemosensitive, whereas a smaller proportion of hypogastric bladder afferent fibers than pelvic afferents are mechanoreceptors (Moss et al., 1997). Mitui et al. reported that signal transduction through bladder hypogastric pathway is necessary in micturition hyperreflexia induced by noxious chemical irritation in conscious rats. These findings suggest that hypogastric/lumbar splanchnic afferents differ from pelvic afferents and may signal bladder sensory information of noxious distension and/or chemical irritation to central nervous system. To date, the mechanisms of bladder mechanosensory transmission via PN and LSN afferents are still not well understood. Currently, two main types of mechanosensory transduction have been hypothesized. The direct mechanism relies on activation of mechanically gated ion channels located on afferent terminals. However, the molecular identity of mechanically gated ion channels is
20 4 still controversial. Possible candidates may include the ENaC/ASIC/degenerin Na + channels and transient receptor potential (TRP) channels. The indirect mechanism relies on activation of ionotropic receptors (e.g., P2X3 receptor) expressed on peripheral nerve terminals by chemical mediators (e.g., ATP) released from urothelial or detrusor muscle cells in response to mechanical stimuli. There are abundant reports that the urothelial cells release chemical mediators such as ATP, NO and prostanoids in response to mechanical and chemical stimulations (Birder, 2005;Birder and de Groat, 2007). To date, numerous receptors are identified in bladder sensory neurons and afferent nerve fibers, including TRP channels (TRPV1, TRPA1, and TRPM8), neurotrophic factor receptors (TrkA, TrkB, and GDNF receptor), α and β estrogen receptors, nicotinic and muscarinic receptors, and purinergic receptors (P2X2, P2X3 and P2Y). Activation of these receptors, especially the TRP channels and the purinergic receptors, has been proved to play an essential role in bladder afferent activity, neurotransmitter release and micturition reflex (Bennett et al., 1996;Bennett et al., 2003;Everaerts et al., 2008;Forrest and Keast, 2008;Zhong et al., 2003). Growing evidence on bladder mechano- and chemo-sensation indicates that sensory mechanisms in the urinary bladder are likely to be complicated and involve a coordination of a variety of chemical and mechanical signaling pathways.
21 5 A functional bladder disorder of visceral pain: Interstitial cystitis/ painful bladder syndrome Acute pain that we have commonly experienced from environmental stimuli, such as the heat of an iron or the prick of a sewing need, is essential for human body protection. Painful inputs generate conscious and unconscious withdrawal reflexes to avoid sever injury. However, chronic pain that persists longer than the temporal course of natural healing severs no protective function, and is usually associated with a particular type of injury or disease process (Porreca et al., 2002;Shipton and Tait, 2005). Interstitial cystitis (IC), also termed as painful bladder syndrome (PBS), is one of the most perplexing chronic pain condition. It is characterized by chronic pain, pressure and discomfort felt in the lower pelvis or bladder over 6 months and often accompanies with daytime frequency and nocturia in the absence of a urinary tract damage or inflammation. Clinical IC/PBS diagnosis is based on histological examination, cystoscopic observation, and physiologic testing by exclusion of other diseases with similar symptoms (Kusek and Nyberg, 2001;Abrams et al., 2006). Although prevalence of IC/PBS is generally underestimated because it is a symptomatic diagnosis and many patients suffering from urinary frequency, urgency and pain may not be diagnosed with IC, a significant number of adult women patients is estimated to up to 20 million according to a recent study in U.S. population (Ibrahim et al., 2007). IC/PBS patients experience substantial chronic pain, depression and significant impairment of personal quality of life, and consequently associated with an
22 6 enormous socioeconomic costs. The direct medical costs attributable to IC in the U.S. healthcare system have been estimated to over $100 million annually(held et al., 1990). Moreover, the treatment of IC/PBS is based on anecdotal experience and a few controversial clinical trials, and only a part of IC/PBS patients are beneficial from traditional oral medicine and intravesical administration. IC/PBS is referred to a functional visceral disorder along with chronic lower pelvic pain and an alteration of urodynamics in absence of gross pathology. The pathogenesis of IC/PBS is still unclear, and a combination of psychological, physiological and genetic factors has been suggested to contribute to IC/PBS pathogenesis. IC is ten times more commonly diagnosed in women than in men, suggesting a role of sex hormones in epidemiology of IC. It has also been reported that IC patients have a higher incidence of other co-morbid diseases, including allergies (50% of IC patients) (Koziol et al., 1993), inflammatory bowel disease (IBD), irritable bowel syndrome (IBS; 50% of IC patients) (Alagiri et al., 1997), migraine (Theoharides et al., 2008), and fibromyalgia (Erickson et al., 2001). Additionally, some studies have presented that stress, diet habit and smoking can exaggerate the IC/PBS symptoms, indicating that IC/PBS may actually be a systemic disorder with bladder syndromes being the main manifestation (Lutgendorf et al., 2000;Kennedy et al., 2006). There may also be a genetic involvement in interstitial cystitis. First-degree relatives of IC patients have a higher prevalence of IC than people in the general population (Warren et
23 7 al., 2004), consistent with an other genetic study showing a greater concordance of IC among monozygotic than dizygotic twins (Warren et al., 2001). Although IC/PBS patients do not display a robust low urinary tract infection or inflammation(al-hadithi et al., 2005), microscopic changes have been described recently in bladder biopsies from IC/PBS patients. The changes include increased number of bladder nerve ending expressing neuropeptide substance P (Pang et al., 1995), increased number of activated bladder mast cells (Sant et al., 2007) and damage of the urothelial protective layer (Parsons et al., 1991). However, the mechanism of IC/PBS pathogenesis is still poorly understood. Therefore, a further explore of IC is essential in improving management of this perplex syndrome. Contribution of purinergic signaling to bladder sensation and pain Known as the intracellular energy source that is essential for all living cells, ATP can also be released into the extracellular space as a neurotransmitter from nerve terminals or as a signaling molecule from non-neuronal cells, including epithelial, glia and smooth muscle cells (North, 2002). ATP concentration in extracellular environment is maintained and regulated by membrane-bounded or soluble ectonucletidases that hydrolyze ATP into ADP, AMP or camp (Zimmermann, 2006). The role of ATP on transduction of sensory information is first demonstrated by Holton et al. in 1950s, showing a release of ATP from sensory nerves during vasodilation in the rabbit artery(holton and Holton, 1954;Holton,
24 8 1959;North, 2002). Increasing study indicates a potential role of ATP signaling in neural transmission, especially in nociception and urinary bladder function. Direct application of ATP on skin or into skeletal muscle causes pain-related behavior in human subjects (Hamilton et al., 2000;Hilliges et al., 2002;Mork et al., 2003). It has been demonstrated that ATP can be released from urothelial cells in response to bladder distension (Ferguson et al., 1997;Wang et al., 2005). An in vivo study also shows intravesical administration of ATP induces bladder overactivity in conscious rats (Pandita and Andersson, 2002), suggesting a potential role of ATP in afferent control of bladder function via acting on primary afferent terminals. Numerous studies have established that extracellular ATP plays an important role in nociception and bladder sensory transduction via activating ATP-sensitive P2 superfamily, consisting of the ionotropic P2X receptors and the metabotropic P2Y receptors coupled with G-protein (Burnstock, 2006;Burnstock, 2007;North, 2002;von Kugelgen, 2006). Ionotropic P2X receptors P2X receptors are non-selective cation channels permeable to Ca 2+, Na + and K +, and they are reported to be involved in the transduction of various sensory signals including taste, hearing, pain, chemical and visceral sensation (Burnstock, 2006;Burnstock, 2007;Surprenant and North, 2008). P2X mrna transcripts and proteins are abundantly distributed throughout the body, including neurons, glia, epithelium, bone marrow and muscle cells. To date, seven mammalian P2X receptor subunits (P2X1-7) have been identified. They share
25 9 about 40% of sequence identity at the peptide level. Each member of P2X family has intracellular NH 2 and COOH termini containing binding sites of protein kinases. Two hydrophobic trans-membrane motifs are involved in channel gating and formation of the ion pore, and separated by a large extracellular loop containing an ATP-binding site. A modulation site is located close to the pocket of ion pore for binding other cations, including Mg 2+, Ca 2+, Zn + and H +, to regulate channel activities. Functional P2X receptors are either homomultimers or heteromultimers composed of three identical or different P2X receptor subunits, and the composition difference contributes to the pharmacological and kinetic variances among P2X receptors. Among seven members of P2X subunits, P2X2 and P2X3 subunits are proposed to have a key role in mediating the nociceptive effect of ATP. Immunohistochemical studies have shown that P2X2 and P2X3 subunits are predominately localized on small-to-medium size sensory neurons within dorsal root ganglia (DRG) and other sensory ganglia (Brady et al., 2004;Vulchanova et al., 1997;Dunn et al., 2001), on peripheral afferent terminals innervating the urinary bladder(cockayne et al., 2000), and on central afferent terminals projecting to the dorsal horn of the spinal cord (Nakatsuka and Gu, 2001;Nakatsuka et al., 2003;Vulchanova et al., 1998). Numerous studies demonstrate that homomeric P2X3 and heteromeric P2X2/3 receptor antagonists can block pain-related behaviors in animal models of long-lasting, chronic neuropathic and inflammatory pain (Honore et al., 2002b;Jarvis et al., 2001;Jarvis et al., 2002;McGaraughty et al., 2003;Ueno et al., 2003;Wu et al.,
26 ). In addition, reduction of P2X2 and/or P2X3 subunit expression by RNA interference (Barclay et al., 2002;Dorn et al., 2004;Honore et al., 2002a) or genetic elimination (Cockayne et al., 2005;Souslova et al., 2000) has a comparable effect as P2X antagonists on attenuation of pain-related behaviors, indicating the importance of P2X signaling in chronic pain syndromes. P2X receptors have also been revealed to participate in mechanosensory regulation of urinary bladder function. P2X3 immunoreactivity has been identified in primary afferent terminals as well as urothelial cells (Cockayne et al., 2000;Zhong et al., 2003). P2X antagonists abolish the bladder overactivity either induced by intravesical infusion of P2X agonist α, β-methylene ATP (Pandita and Andersson, 2002), or in lower urinary tract obstructed rats (Cova et al., 1999;Velasco et al., 2003) in vivo. A previous in vitro study using bladder-nerve preparation in the rat also showed a comparable inhibitory effect of P2X antagonist on bladder afferent hypersensitivity either induced by ATP or after bladder inflammation (Yu and de Groat, 2008). Transgenic mice have provided instrumental evidence further establishing the role of P2X receptors in bladder function. P2X2 knockout, P2X3 knockout and P2X2/P2X3 double knockout mice exhibit bladder hyporeflexia and decreased afferent nerve activities in response to bladder distension while the ATP release from urothelium during bladder distension in knockout mice is not different compared with wild-type mice (Cockayne et al., 2000;Cockayne et al., 2005;Vlaskovska et al., 2001). These studies indicate the potential role of ATP and P2X receptors in afferent control of bladder function.
27 11 Metabotropic P2Y receptors To date, eight G-protein coupled P2Y receptors (P2Y1, P2Y2, P2Y4 P2Y6, P2Y11, P2Y12, P2Y13, and P2Y14) have been cloned. They are characterized by seven transmembrane-spanning regions that form the ligand binding pocket and share a high level of sequence homology between transmembrane regions. Each P2Y receptor has an extracellular NH 2 terminus and intracellular COOH terminus that contains a binging site of protein kinases. The intracellular loops and COOH terminus among P2Y subtypes display great variation that may contribute to coupling different G-proteins. P2Y receptors can be activated by endogenous purine nucleotides (ATP, ADP) and/or pyrimidine nucleotides (UTP, UDP) based on their differential pharmacological properties. Once activated by ligands, P2Y1, P2Y2, P2Y4, P2Y6 and P2Y11 receptors that are coupled to Gq signal pathway, trigger phospholipase C (PLC) activation, induce the release of endogenous Ca2+ produce, and finally initiate protein kinase C pathway. On the contrary, P2Y12, P2Y13 and P2Y14 receptors that are coupled to G i pathway have been reported to inhibit adenylate cyclase (von Kugelgen, 2006). The contribution of the metabotropic P2Y receptors to sensory transmission has been less well examined compared with P2X receptors. However, the expression of P2Y1, P2Y2, P2Y4 and P2Y6 mrna transcripts has been detected in dorsal root ganglia neurons, implicating a potential role of P2Y receptors in peripheral sensory transduction. Of eight members in P2Y family, P2Y1 and P1Y2 receptors are more abundantly expressed in sensory neurons
28 12 and therefore attract greater attention than other P2Y receptors. P2Y1 receptor has been reported to contribute to innocuous mechanosensory transmission in frog sensory nerve fibers (Nakamura and Strittmatter, 1996). UTP (P2Y2 and P2Y4 agonist) can activate cutaneous afferents, sensitize sensory neurons and induce CREB phosphorylation through the P2Y2 receptor (Molliver et al., 2002;Stucky et al., 2004). P2Y receptors also play an essential role in the enhancement of intracellular calcium and a subsequent CGRP release in response to ATP in sensory neurons (Song et al., 2007;Sanada et al., 2002). These results suggest that metabotropic P2Y receptors are involved in transduction of primary sensory information. In addition, P2Y receptors have been implicated to play an indirect role in mechanosensation and nociception by modulating other receptors or channels. Immunohistochemistry studies have revealed that P2Y1 receptor is co-localized with P2X3 and TRPV1 receptors in rat dorsal root ganglia neurons (Gerevich et al., 2005). Activation of P2Y1 receptor can influence currents through N-type calcium channels (Cav 2.2) and P2X3 receptor in dorsal root ganglia neurons (Gerevich et al., 2004;Gerevich et al., 2005). TRPV1 channel is thought to be important in mechanosensation and nociception (Clapham, 2003). Genetic elimination of TRPV1 causes reduced thermal hypersensitivity and impaired bladder function (Birder et al., 2002;Caterina et al., 2000). It has been shown that P2Y2 receptor potentiates TRPV1 response and reduces its thermal threshold in peripheral afferent neurons (Moriyama et al., 2003;Tominaga et al., 2001). Therefore, the role of P2Y receptors in sensitization of sensory afferents and
29 13 other ion channels implicates an involvement of P2Y signaling in visceral hypersensitivity and chronic pain associated with IC. Thesis objectives The hypothesis of this thesis is that the purinergic signaling mediated by P2X and P2Y receptors plays a role in bladder sensation under normal and pathological conditions, The hypothesis was examined in sensory afferent neurons innervating the urinary bladder from naïve mice or chemical inducedbladder inflamed mice. Majority of previous studies on mechanisms of bladder sensation have been performed on pelvic nerve afferent pathways. Therefore, one aim of this thesis is to characterize purinergic sensitivity in both lumbar splanchnic and pelvic sensory pathway under normal condition (Chapter 2) and in the presence of bladder inflammation (Chapter 3). Chapter 4 examined the influence of metabotropic P2Y2 receptor on the excitability of bladder sensory neurons and the function of P2X receptors. These results provide a substantial support for a peripheral sensory effect of P2X and P2Y2 receptors on bladder function and a contribution of purinergic receptors to bladder hypersensitivity and pain.
30 14 CHAPTER 2: CHARACTERIZATION OF PURINERGIC P2X RECEPTORS IN LUMBOSACRAL (LS) AND THORACOLUMBAR (TL) BLADDER SENSORY NEURONS Introduction Sensory information from the urinary bladder is conveyed to the spinal cord via lumbar splanchinic and pelvic pathways (Vera and Nadelhaft, 1992;Uemura et al., 1975;Uemura et al., 1973). The cell bodies of these afferent pathways are located, respectively, in thoracolumbar (TL) and lumbosacral (LS) dorsal root ganglia (Andersson, 2002;Applebaum et al., 1980). A variety of mechanical and chemical stimuli can activate the peripheral terminals of sensory neurons innervating the urinary bladder, including distension/contraction of the bladder, chemical irritants in urine, endogenous signal molecules released from urothelial cells, and inflammatory mediators produced during tissue insults. However, bladder sensation studies principally focus on the pelvic afferent pathway whereas the contribution of lumbar splanchnic innervation to bladder function is not well investigated. A unique feature of visceral innervation is that each organ is innervated by two nerves (Gebhart and Bielefeldt, 2008), which have some similar but also different functions. For example, the mechanosensitivity and location of receptive endings of the pelvic and lumbar splanchnic innervations of the urinary bladder (Xu and Gebhart, 2008) and colon (Brierley et al., 2005) in the mouse have been directly compared and documented as significantly different. Other studies that
31 15 have examined cell bodies in dorsal root or nodose ganglia of different nerves innervating the same organ have similarly revealed significant differences in gastric (Dang et al., 2005b;Sanada et al., 2002;Sugiura et al., 2005), airway (Undem et al., 2004;Kwong et al., 2008), colon (Brierley et al., 2005) and urinary bladder (Dang et al., 2005a;Xu and Gebhart, 2008) sensory neurons. The importance of these findings relates to resolution of potential mechanisms that may underlie functional visceral disorders (e.g., irritable bowel syndrome, interstitial cystitis [IC], etc.), all of which are characterized by discomfort and pain in the absence of gross pathology. Among potential endogenous mediators of bladder sensation and discomfort, adenosine triphosphate (ATP) and ionotropic purinergic (P2X) receptors have been identified as important in the regulation of micturition and sensation arising from the bladder. ATP is released from bladder urothelium during distension or chemical stimulation (Ferguson et al., 1997;Birder et al., 2003). The micturition reflex in normal conscious animals can be initiated by intravesical ATP application (Pandita and Andersson, 2002); correspondingly, P2X receptor antagonists applied into bladder lumen significantly reduce the bladder contraction and voiding reflex in both normal and bladder obstructed rats (Cova et al., 1999;Velasco et al., 2003). In addition, P2X2, P2X3 and P2X2/P2X3 double knockout mice exhibit reduced bladder reflexes and decreased afferent nerve activity in response to bladder distension (Cockayne et al., 2000;Cockayne et al., 2005;Vlaskovska et al., 2001). Therefore, in this chapter I examined the purinergic sensitivity in bladder sensory neurons and compared the differences
32 16 between the lumbar splanchnic and pelvic innervations of the mouse urinary bladder by study of their cell bodies in TL and LS dorsal root ganglia. Results Cell density and size distribution of bladder sensory neurons To estimate the proportion of bladder sensory neurons contained in LS and TL DRG, I randomly chose 4 LS DRG and 4 TL DRG coverslips from 4 mice, respectively. Each coverslip was divided into four quadrants. Numbers of DiI labeled cells and total cells were counted in a randomly selected viewing field (10X objective) in each quadrant under differential-interference contrast (DIC) mode (Figure 1A) and fluorescence mode (Figure 1B). DiI-labeled cells represented 6.0±0.4% (77/1276) of L6-S2 DRG cells, a proportion significantly greater than the 2.4±0.2% (40/1635) of DiI-labeled T13-L2 DRG cells (Figure 1C). These proportions do not differ from similar data collected in the rat (Dang et al. 2005). Unlike in the rat, however, where TL bladder neuron capacitance was significantly greater than LS bladder neuron capacitance, mean whole cell capacitance (as an index of cell size) did not differ between mouse LS (31.1±1.0 pf) and TL (32.3±1.1 pf) bladder sensory neurons (Figure 2A). The distributions of cell size (Figure 2B) also did not differ; most cells (LS: 71.9%; TL: 69.2%) were medium in size (20-45 pf), as in the rat (Dang et al. 2005). However, in the rat, the proportion of small sized (<20 pf) bladder sensory neurons was significantly
33 17 greater and the number of large sized (>45 pf) cells significantly less in LS than TL DRG (Dang et al, 2008). Characterization of bladder sensory neuron responses to purinergic receptor agonists A total of 205 bladder sensory neurons from naïve mice were studied. ATP (30 μm) was applied to cells as a non-selective P2X receptor agonist; α,βmethylene ATP (α,β-met ATP, 30 μm) was employed as a P2X3 receptorselective agonist (homomeric P2X3 and heteromeric P2X2/3). Agonists were applied for 4 sec at 2 min intervals. Overall, 93.0% (106/114) of LS and 76.9% (70/91) of TL bladder neurons responded to ATP (LS vs TL, P< 0.01); 83.3% (95/114) of the same LS and 76.9% (70/91) of the same TL bladder neurons also responded to α,β-met ATP (Table 1). Among responsive bladder neurons, the overwhelming majority of LS (89.6%, 95/106) and TL (74.3%, 52/70, LS vs TL, P< 0.05, Table 1) bladder sensory neurons exhibited slowly desensitizing currents in response to both ATP and α,β-met ATP (Figure 3A, 3C). The remaining LS bladder neurons (10.4%, 11/106) demonstrated a sustained current without an obvious desensitizing phase during agonist application. Sustained currents in LS bladder neurons were evoked only by ATP and not by α,β-met ATP (Figure 3B). Neither ATP nor α,β-met ATP generated sustained currents in TL bladder sensory neurons; instead, both agonists produced rapidly activating, rapidly inactivating fast currents (25.7%, 28/70, Figure 3D). No correlation between bladder neuron size (whole-cell capacitance) and type of purinergic currents was observed.
34 18 Table 2 summarizes the properties of purinergic agonist-evoked currents in LS and TL bladder sensory neurons from naïve mice. Both agonists produced slowly desensitizing currents in LS and TL neurons. The activation (time to peak) and desensitization (desensitizing time constant) kinetics of these slow currents were not different between LS and TL bladder neurons. However, the current density (current amplitude normalized by whole cell capacitance) of the slow current in LS bladder neurons was significantly greater than in TL neurons in response to ATP (P<0.01) or α,β-met ATP (P<0.05). A sustained current was produced only in LS bladder neurons and a fast current was produced only in TL neurons, suggesting significant differences in P2X receptor subunit composition between LS and TL bladder sensory neurons. Characterization of P2X receptor subtypes in bladder sensory neurons There are seven ionotropic P2X receptor family subunits (P2X1 to P2X7) that have been reported and cloned to date (North, 2002;Surprenant and North, 2008). The P2X2 and P2X3 subunits predominate in dorsal root ganglia and can form functional homomeric P2X2 receptors, heteromeric P2X2/3 receptors or homomeric P2X3 receptors each of which has a distinct pharmacological character (Nicke et al., 1998;Gever et al., 2006). To examine which P2X receptor subunit(s) mediate purinergic currents in mouse bladder sensory neurons, I applied the P2X1, P2X3 and P2X2/3 receptor-selective antagonist 2,3 -O-(2,4,6- trinitrophenyl) adenosine 5 -triphosphate (TNP-ATP, 0.1 μm) or the non-selective P2X receptor antagonist pyridoxal-phosphate-6-azophenyl-2',4'-disulfonate
35 19 (PPADS, 10 μm) for 30 sec before agonist application. Sustained currents evoked by ATP were significantly attenuated by PPADS by a mean 82.0±5.6 %( Figure 4C; an example is given in Fig 4B) but not by TNP-ATP (Figure 4A), suggesting they are mediated by a homomeric P2X2 receptor. As summarized in Figure 5C, TNP-ATP significantly attenuated the slow currents evoked by ATP (by a mean 55.5±2.1%; an example is given in Figure 5A) or α, β-met ATP (by a mean 55.6±3.0%). PPADS similarly attenuated the slow currents evoked by ATP (by a mean 77.7±3.0%; an example is given in Figure 5B) or α, β-met ATP (by a mean 82.0±2.1%), revealing that these slow currents are mediated by heteromeric P2X2/3 receptors. Fast currents are usually associated with homomeric P2X3 receptors, which was confirmed using TNP-ATP (Figure 6A) and PPADS (Figure 6B). Approximately 100% inhibition of ATP-evoked (TNP- ATP: 95.8±1.8%; PPADS: 95.9±0.2%, Figure 6C) and α, β-met ATP-evoked (TNP-ATP: 97.6±0.7%; PPADS: 98.2±0.5%) fast current amplitude were observed using the antagonists and the inhibitory effect was reversible by washout of antagonists. To further evaluate these currents, I applied the same purinergic agonists and antagonists to bladder sensory neurons taken from P2X3 knockout mice. No slow or fast currents were produced by either purinergic agonist; only a sustained current was found. Ten of 12 (83.3%) LS bladder neurons responded to ATP with a sustained current (an example is given in Figure 7A); none responded to α, β- met ATP. TL bladder neurons (n=8) did not respond to either ATP or α, β-met ATP (Figure 7B). The sustained current produced by ATP in LS bladder neurons
36 20 from P2X3 knockout mice was attenuated similarly by TNP-ATP and PPADS as were bladder neurons from wild-type mice (examples are given in Figure 7C, D). These outcomes confirm that activation of homomeric P2X2 receptors is responsible for the sustained current recorded in LS bladder sensory neurons. Moreover, in TL bladder sensory neurons, purinergic currents are principally mediated through heteromeric P2X2/3 receptors and homomeric P2X3 receptors. Electrophysiological properties of LS and TL bladder sensory neurons I also examined the active and passive membrane properties of 32 LS and 30 TL bladder neurons from naïve mice by injecting currents (an example displayed in Figure 8A) and applying agonists (examples of ATP-evoked membrane depolarization and action potential are displayed in Figure 8B) in whole-cell current clamp mode. Input resistance was calculated according to the I/V relationship by injecting a series of hyperpolarizing pulses ranging from 300 to 0 pa (50 ms) in 50pA increments. Oscillation was examined as an index of variability in the resting membrane potential (RMP). To determine rheobase, a series of 10 ms current pulses in 20pA increments (1s apart) was injected. The maximum current (pa) that did not evoke an action potential was taken as rheobase. Action potential (AP) threshold was determined from the inflection point where membrane potential started to dramatically rise and the phase plot slope (the first derivative of membrane potential, dv/dt) reached 10 mv/ms (Naundorf et al., 2006), AP amplitude was measured from peak RMP to the peak of the AP, AP overshoot was the amplitude from 0 mv to the peak of the AP, AP
37 21 duration was determined at 50% of the AP amplitude and the AP falling rate was the velocity of change in potential from the AP peak to RMP. As summarized in Table 3, LS and TL bladder neurons did not differ in input resistance, oscillation, RMP, or AP duration, amplitude, overshoot or failing rate. However, LS neurons had a significantly more negative mean action potential threshold (-34.1±0.5mV) than TL neurons (-28.5±0.8mV, P<0.01), suggesting that LS neurons were generally more easily excited. When 30μM ATP was applied, a significantly greater proportion of LS neurons fired APs than did their TL counterparts (LS: 21/32, 67.5%; TL: 10/30, 33.3%, P<0.05), suggesting that LS bladder neurons are more sensitive to purinergic agonists at the concentration tested. Spontaneous activity was not observed in either LS or TL bladder neurons before or after agonist application. P2X receptor expression in bladder sensory neurons Because the numbers of bladder sensory neurons contained in TL and LS DRG are relatively few, I employed single cell RT-PCR and single cell nested PCR to examine P2X2 and P2X3 expression in bladder sensory neurons. When cdna was harvested after single cell RT-PCR, the mouse GAPDH gene was amplified by conventional PCR as an internal control. Only cells having a thick band of GAPDH amplicon were further processed by nested PCR. Negative results of GAPDH amplification were thought to have an unsuccessful reverse transcription reaction and thus discarded. The remaining bladder neurons were amplified by two rounds of PCR cycles with external primers, then internal primers, respectively.
38 22 Figure 9A shows an example of positive single cell RT-PCR amplicons of P2X2 and P2X3 mrna. Product length corresponded with the expected size of the targeted region. 15 LS and 15 TL bladder DRG neurons per mouse (n=3) were collected for single cell PCR assay. The P2X2 subunit transcript was more abundant in LS bladder neurons (88.9%±2.2%) than in TL counterparts (53.3%±3.8%; P<0.01). It is exhibited in Figure 9B that the P2X3 subunit transcript was abundant in both LS (93.3%±3.8) and TL bladder neurons (97.7%±2.2%). Figure 9C shows the proportions of LS and TL bladder sensory neurons that expressed only P2X2, only P2X3 or both P2X2 and P2X3 transcripts. Consistent with the purinergic-evoked currents in LS and TL bladder neurons, P2X2 and P2X3 transcript co-expression were predominant in both LS (88.9%±2.2%) and TL (51.2%±5.9%) bladder neurons. Cells only expressing P2X2 transcripts were found exclusively in LS bladder (8.9%±2.2%), which is consistent with observation of sustained currents in LS bladder neurons. Cells only expressing P2X3 transcripts were more frequently detected in TL (51.2%±5.9%) bladder neurons compared with LS counterparts (2.2%±2.2%; P<0.05). I also examined P2X3 immunoreactivity in bladder sensory neurons retrogradely labeled by Alexa Fluor 488-conjugated Cholera Toxin B subunit (CTB), a hydrophilic and membrane-permeable fluorescent tracing dye. Figure 10 shows example images of P2X3 immunostaining at LS (an example of L6 DRG displayed in Figure 10A) and TL (an example of L1 DRG displayed in Figure 10B) DRGs from CTB labeled mice. Distribution of bladder sensory
39 23 neurons (green) and the immunoreactivity of P2X3 receptor subunits (red) are displayed in the left and middle panel, respectively; combined images of double fluorescent labeling are presented in the right panel. As summarized in Table 4, 27.2% (83/305) of LS DRG neurons exhibited P2X3 immunoreactivity, which was significantly less than 36.4% (156/429, P<0.05) of TL cells showing P2X3 immunoreactivity. Similar with proportion of DiI positive cells, CTB-labeled cells represented 7.2% (22/305) of LS DRG cells, a proportion significantly greater than the 2.8% (12/429, P<0.05) of TL DRG cells. Colocalization of positive P2X3 immunoreactivity and CTB labeling was rare in LS DRG; only 5% of CTB labeled bladder sensory neurons exhibited detectable P2X3 immunoreactivity. On the contrary, positive P2X3 immunoreactivity was ~10 fold more frequently detected in CTB-labeled TL bladder DRG neurons (50%, 6/12; P<0.01) than LS counterparts. Discussion Differences between LS and TL bladder sensory neurons Previous studies suggest that the pelvic and lumbar splanchnic afferent pathways innervating the urinary bladder may serve different functions. Bladder afferent fibers in pelvic and lumbar splanchnic pathways are both involved in chemo- and mechano-sensation, including noxious sensations (Andersson, 2002;Mitsui et al., 2001;Moss et al., 1997;Shea et al., 2000;Sengupta and Gebhart, 1994;Su et al., 1997;Nazif et al., 2007). However, these two bladder afferent pathways differentially respond to mechanical and chemical stimuli
40 24 (Dang et al., 2005a;Xu and Gebhart, 2008). In the present study, it was confirmed that mouse bladder sensory neurons in the pelvic (LS) and splanchnic (TL) pathways exhibit significantly different responses to purinergic agonists based on kinetics of activation/inactivation and pharmacologic antagonism of the inward currents produced. In naïve mice, a greater proportion of LS bladder neurons responded to purinergic agonists (~90%) than did TL bladder neurons (~75%). Three types of purinergic currents were identified based on kinetic parameters and responses to agonists and antagonists: homomeric P2X2 receptors (producing a sustained current), heteromeric P2X2/3 receptors (producing a rapidly activating, slow desensitizing current), and homomeric P2X3 receptors (producing a fast current). The predominant current produced by both purinergic agonists in both LS and TL bladder neurons was a heteromeric P2X2/3 slow current; the slow current densities produced by both purinergic agonists in LS bladder neurons were significantly greater than in TL counterparts. I also measured active and passive membrane properties of LS and TL bladder neurons, and found LS bladder neurons had a significantly lower (more negative) mean action potential threshold and produced action potentials in a greater proportion of LS than TL neurons, suggesting that pelvic bladder afferents more tend to be easily activated by membrane depolarization or ATP when released, for example, from urothelial cells. This interpretation further suggests that pelvic bladder afferents are more sensitive to bladder distension during normal urine filling, which is supported by the greater proportion of stretch
41 25 sensitive bladder pelvic afferent fibers than lumbar splanchnic afferent fibers, and further distinguished by the greater dynamic response of pelvic bladder afferents to mechanical stimulation (Xu and Gebhart, 2008). A P2X2 homomeric sustained current was observed only in LS bladder neurons and only in about 10% of neurons responsive to ATP. A P2X3 homomeric fast current was observed only in TL bladder neurons, occurring in about 25% of the neurons, but produced equally by ATP and α, β-met ATP. These outcomes suggest that [1] all mouse LS bladder neurons responding to purinergic agonists express the P2X2 subunit, with the vast majority also expressing the P2X3 subunit, and [2] all TL bladder neurons responding to purinergic agonists express the P2X3 subunit with the significant majority also expressing the P2X2 subunit. These interpretations are supported by single cell nested PCR, which revealed that the P2X2 subunit transcript predominates in LS bladder neurons whereas the P2X3 subunit transcript predominates in both LS and TL bladder neurons. These results in mice differ from those previously reported in the rat. Dang et al (2008) compared rat LS and TL bladder neuron responses to purinergic agonists. Based on inactivation kinetics, the predominant current in naïve rat LS bladder neurons was a P2X2/3 heteromeric slow current evoked by both ATP and α,β-met ATP (same concentrations as used herein) in 87% of neurons (consistent with the present observation in mouse LS bladder neurons). They noted no P2X2 homomeric sustained currents (present in ~10% of mouse LS bladder neurons studied here), but did observe small percentages (6%) of
42 26 P2X3 homomeric fast currents and of mixed, rapidly activating and mixed desensitizing currents (fit using a double exponential; 7%). No P2X3 homomeric fast currents were found in mouse LS bladder neurons. The predominant (50 60%) purinergic-evoked current in rat TL bladder neurons was the mixed current, which was never noted in either LS or TL mouse bladder neurons, with about one-third of rat TL bladder neurons exhibiting a P2X2/3 heteromeric slow current (the predominant current [75%] in mouse TL bladder neurons). Overall, more LS than TL bladder neurons respond to purinergic agonists in both rat and mouse. P2X2 homomeric sustained currents are present in mouse LS bladder neurons, but not in either rat LS or TL bladder neurons, and P2X3 homomeric fast currents are present only in mouse TL bladder neurons, but in both LS and TL rat bladder neurons. The expression of P2X transcripts and protein in bladder sensory neurons reported here is consistent with other electrophysiological studies of bladder afferents (Rong et al., 2002;Zhong et al., 2003), and immunohistochemical localization in DRG (Vulchanova et al., 1997;Vulchanova et al., 1998), including bladder sensory neurons (Dang et al. 2008), and nerve terminals in the suburothelial nerve plexus (Cockayne et al., 2000;Studeny et al., 2005). Transcription and expression of P2X receptor subunits are variable between species, organs and ganglia (North, 2002;Burnstock, 2006;Grubb and Evans, 1999). Results from single cell nested PCR support the kinetic and pharmacological results reported here in that some bladder neurons expressed only P2X2 transcripts, only P2X3 transcripts, or both P2X2 and P2X3 transcripts.
43 27 P2X2 transcripts are expressed with greater frequency in LS than TL bladder neurons, whereas P2X3 transcripts are highly expressed in both LS and TL bladder neurons (>90%). The frequency of P2X receptor subunit expression in bladder sensory neurons is consistent with the proportions of neurons exhibiting P2X homo- or heteromeric currents based on desensitization kinetics. There is a high frequency of colocalization of P2X2 and P2X3 subunits in both LS and TL bladder neurons, consistent with the predominant P2X2/3 heteromeric slow current evoked by both purinergic agonists in LS and TL bladder neurons. The proportion of LS neurons that express only the P2X2 transcript is low, consistent with the ~10% of LS neurons that exhibited a sustained, homomeric P2X2 inward current to application of ATP. Similarly, the proportion of LS neurons that express only the P2X3 transcript very low, and no P2X3 homomeric fast currents were found in LS neurons. Results from immunohistochemistry staining of P2X3 subunit in LS and TL DRGs support the electrophysiological study: [1] the small proportion of LS and TL bladder sensory in whole dorsal root ganglia; [2] a significantly greater frequency of P2X3 in TL than LS bladder neurons. A lower proportion of bladder sensory neurons expressing P2X3 protein than mrna transcripts may be due to partial translation of P2X3 mrna into protein, internalization and degradation of functional P2X3 receptors, or the sensitivity of immunohistochemistry staining technique. In conclusion, because significantly more LS responded to purinergic agonists, and LS neurons exhibited significantly greater current density of slow
44 28 type (the predominant current type in bladder neurons) than the TL counterparts, it is suggested that purinergic transmission of the urinary bladder in the normal physiological state is principally conveyed through pelvic rather than hypogastric/lumbar splanchnic afferents.
45 29 CHAPTER 3: PURINERGIC P2X SIGNALING IN BLADDER SENSORY NEURONS AFTER BLADDER INFLAMMATION Introduction Bladder inflammation, also termed as cystitis, can occur as a result of bacteria infection, irritant chemicals in the urine, or unidentified causes (IC/PBS). Bladder inflammation is characterized by discomfort/pain in lower pelvic region, bladder hypersensitivity, edema, and inflammatory reaction of numerous cells in the bladder tissue. Because activation of bladder sensory afferents plays a substantial role in transmitting mechanical or chemical signals to central nervous system, considerable attention has been focused on the plasticity of bladder afferent nerves and sensory neurons induced by pathological changes. Accumulating evidence has identified various endogenous molecules serving as potential mediators of bladder inflammation and pain. In addition to an essential role in bladder sensation under physiological conditions, it has been proposed that ATP and ionotropic purinergic (P2X) receptor may also contribute to bladder dysfunction following chronic inflammation. ATP is released from bladder urothelium during distension or chemical stimulation (Ferguson et al., 1997;Birder et al., 2003) and the release is increased in IC patients (Sun et al., 2001a;Sun et al., 2001b). In animal models of cystitis, blockage of P2X receptors can reduce bladder overactivity in vivo and hypersensitivity of bladder afferent fibers in response to mechanical and electrical stimuli in vitro(ito et al., 2008;Yu and de Groat, 2008). Human studies on IC/PBS patients also suggest the
46 30 association between purinergic signaling and functional bladder disorders (Tempest et al., 2004;Ray et al., 2003). As mentioned in Chapter 1, the cell bodies of bladder pelvic and lumbar splanchnic nerves are located in lumbosacral (LS, L6 S2) and thoracolumbar (TL, T13 L2) DRGs. Recent studies have reported that bladder lumbar splanchnic afferents respond more vigorously to chemical stimuli than pelvic nerve counterparts(moss et al., 1997;Mitsui et al., 2001), suggesting that pelvic and lumbar splanchnic nerves may make different contribution to signaling noxious stimuli after bladder inflammation. To test whether P2X plays a role in signaling the urinary bladder sensory information in the bladder-inflamed state, systemic administration of cyclophosphamide (CYP), was used in wild-type C57BL/6 mice. CYP can be metabolized into bladder irritant acrolein (Cox, 1979) that causes hemorrhagic cystitis in humans as an adverse event and produces visceral pain behaviors and a cystitis-like syndrome, including edema, ulceration of the urothelium and hemorrhage in rodents (Bon et al., 2003). Therefore, I used this well-established bladder inflammation model to examine the consequences of inflammation on characters of mouse LS and TL bladder neurons, especially the plasticity of P2X functions. It is hypothesized that bladder inflammation changes the excitability and the purinergic sensitivity of bladder sensory neurons.
47 31 Results Bladder inflammation and tissue damage after CYP treatment A group of C57BL/6 mice were treated intraperitoneally with 100 mg/kg cyclophosphamide dissolved in saline daily for 5 days. Mice treated with same volume of vehicle (saline) were served as controls. Relative to naïve and salinetreated controls, CYP-treated bladders generally had thick walls accompanied by visibly decreased lumen volume. Mean bladder weight (Figure 11A) after CYP treatment was significantly greater (38.6 ± 1.3 mg, n=8) than bladders taken from saline-treated (23.7 ±0.7mg, n=6; P<0.005) and naïve mice (22.9±1.1mg, n=6; P<0.005). A biochemical assay of bladder myeloperoxidase (MPO) activity was applied on naïve, saline and CYP-treated mice to evaluate bladder inflammation induced by CYP treatment. MPO is the most abundant protein in neutrophils and can catalyze hydrogen peroxide (H 2 O 2 ) into hypochlorous acid, which is considered as a powerful antimicrobial agent. MPO activity assay is widely used for quantitative assessment of neutrophil infiltration in inflammatory diseases. However, no significant change of bladder MPO activity after CYP treatment was detected compared with saline-treated or naïve mice (Figure 11B). The bladder MPO activities of naïve, saline- and CYP- treated mice were all relative low. Histological examination of bladders from CYP-treated mice revealed mild submucosal edema and unfolding of the urothelium, neither of which was apparent in bladders from saline-treated mice (Figure 12). Although MPO activity
48 32 did not differ between bladders from CYP- and saline-treated/naive mice, CYP treatment did produce histological insult of the urinary bladder. Bladder sensory neuron excitability increases after CYP treatment Using the same whole-cell current clamp protocols described in Chapter 2, active and passive membrane properties of LS and TL bladder neurons were examined in bladder sensory neurons taken from CYP- and saline-treated mice. As summarized in Table 5, rheobase was significantly lower in both LS (from ±8.7mV to ±9.5mV; P<0.05) and TL (from ±22.6mV to128.2 ±8.5mV; P<0.05) bladder neurons from CYP-treated relative to saline-treated mice (Figure 13A-D). The magnitude of membrane depolarization produced by α,β-met ATP was significantly increased in LS and TL bladder neurons after CYP treatment, but not by ATP (examples are given in Fig 13E-H). Input resistance, AP duration, amplitude, overshoot or failing rate was not changed in both LS and TL balder sensory neurons after inflammation. No bladder neurons from CYPtreated mice exhibited spontaneous activity. P2X receptor mediated currents after CYP treatment As presented in Table 6, 93.3% of LS bladder neurons from CYP-treated mice responded to ATP, similar to the 96.9% of LS neurons from saline-treated mice that responded. However, the proportions of LS bladder neurons that exhibited sustained and slow currents to application of ATP were significantly different in CYP- treated relative to saline-treated mice. A significantly greater proportion (P<0.01) of LS neurons from CYP-treated mice (42.9%) exhibited
49 33 sustained currents than did LS neurons taken from saline-treated mice (9.7%). Correspondingly, the proportion of LS neurons from CYP-treated mice that gave slow currents (57.1%) was significantly reduced (P<0.01) relative to the 90.3% observed in saline-treated mice. α, β-met ATP evokes only slow currents in LS bladder neurons and the proportion of LS neurons from CYP-treated mice (53.3%) that responded to α, β-met ATP was also significantly decreased relative to saline-treated mice (87.5%; P<0.01). The activation (time to peak) and desensitization (desensitizing time constant) kinetics of the slow currents evoked by ATP/α, β-met ATP were not different between neurons from saline- and CYPtreated mice. The current densities of both the sustained and slow currents in LS neurons from CYP-treated mice exhibited a trend to increase, but not significantly greater than those of saline controls (Table 7). These results suggest that the subunit composition of functional P2X receptors is altered by bladder inflammation, with a greater contribution made by homomeric P2X2 receptors in pelvic nerve LS neurons after inflammation. The effect of bladder inflammation on TL bladder neurons was less remarkable than on their LS counterparts. The proportions of TL bladder neurons that responded to either of the purinergic agonists were not significantly different between CYP- and saline-treated mice (Table 6). Although the proportions of responses did not differ, the current density of the fast current evoked by agonists was greater in neurons from CYP-treated mice while current density of the slow response to agonists was significantly less in neurons from CYP-treated mice (both relative to saline-treated mice; Table 7).
50 34 P2X receptor expression in bladder sensory neurons after CYP treatment Because the numbers of bladder sensory neurons contained in TL and LS DRG are relatively few, I employed single cell RT-PCR and single cell nested PCR to examine P2X2 and P2X3 expression in bladder sensory neurons, as described in Chapter 2. Figure 14A shows an example of positive single cell RT-PCR amplicons of P2X2 and P2X3 mrna. Product length corresponded with the expected size of the targeted region. 15 LS and 15 TL bladder DRG neurons per mouse were collected for single cell PCR assay. Both saline- and CYP-treated group consisted of 45 cells taken from 3 mice. The P2X2 subunit transcript was more abundant in LS bladder neurons from saline-treated animals (91.1%±2.2%) than in TL counterparts (46.7%±6.7%; P<0.01). The frequencies of P2X2 transcript expression in TL bladder neurons from CYP-treated mice (73.3%±6.7%) significantly increased relative to saline-treated mice (46.7%±6.7%; P<0.05). The P2X3 subunit transcript was abundant in both LS (saline: 91.1%±5.8%; CYP: 83.7%±10.4%) and TL bladder neurons (saline: 93.3%±3.8%; CYP: 100%). The frequencies of P2X3 transcript expression did not differ between cells taken from CYP- and saline-treated mice (Figure 14A). Figure 14B shows the proportions of LS and TL bladder sensory neurons that expressed only P2X2, only P2X3 or both P2X2 and P2X3 transcripts. Consistent with the purinergic-evoked currents in LS and TL bladder neurons from bladder inflamed and naïve mice, P2X2 and P2X3 transcript co-expression
51 35 were predominant in both LS (saline: 82.2%±2.2%; CYP: 76.8%±9.8%) and TL saline: 51.1%±8.0%; CYP: 73.3%±6.7%) bladder neurons. Cells only expressing P2X2 transcripts were found in LS bladder sensory neurons from saline- (6.7%±3.8%) and CYP-treated (14.0%±7.3) mice. Cells only expressing P2X3 transcripts were more frequently detected in TL (saline: 49.0%±8.0%; CYP: 26.7%±6.7%) bladder neurons compared with LS counterparts (saline: 4.5%±2.2%; CYP: 4.6%±2.3%; both P<0.05). The frequencies of P2X2/P2X3 transcript expression did not differ between cells taken from CYP- and salinetreated mice. P2X3 immunoreactivity in CTB-labeled bladder sensory neuron from CYPand saline-treated mice was also examined, as shown in Figure 15. Distribution of bladder sensory neurons (green) and the immunoreactivity of P2X3 receptor subunits (red) are displayed in the left and middle panel, respectively; combined images of double fluorescent labeling are presented in the right panel. As summarized in Table 4, 23.6% (164/696) of LS DRG neurons from saline-treated mice (Figure 15A) and 19.7% (104/529) of LS DRG neurons from CYP-treated mice (Figure 15C) exhibited positive P2X3 immunoreactivity, which was significantly less than the proportion of TL bladder sensory neurons expressing P2X3 receptor subunits (saline: 36.3% [64/696], Figure 15B; CYP: 37.0% [181/489], Figure 15D, both P<0.05). No significant change of P2X3 expression was detected after CYP treatment relative to saline controls. Consistent with the frequency of bladder sensory neurons detected in naïve mouse DRG, 8.0% (56/696) of LS and 2.3% (11/470) of TL DRG neurons were identified as CTB-
52 36 labeled bladder sensory neurons from saline-treated mice; 6.6% (35/529) of LS and 2.0% (10/489) TL DRG neurons were recognized as bladder neurons from CYP-treated mice. Co-localization of positive P2X3 immunoreactivity and CTB labeling was rare in LS DRG; only 3.5% and 2.8% of CTB labeled bladder sensory neurons from saline- and CYP-treated mice exhibited detectable P2X3 immunoreactivity. On the contrary, positive P2X3 immunoreactivity was fold more frequently detected in TL bladder neurons (saline: 63.6% [7/11]; 50%, 5/10; both P<0.01) than LS counterparts. These results revealed that P2X3 expression was not greatly changed in protein level after CYP-induced bladder inflammation. Discussion CYP treatment induces moderate bladder inflammation After systematic treatment of CYP for 5 days, CYP-treated bladders exhibited a significant weight increase, accompanied by edema in submucosa area and visibly decreased lumen volume of the bladder relative to naive and saline-treated controls. However, bladder tissues from CYP-treated mice did not show an increase of MPO activity, suggesting no sever neutrophil infiltration associated with CYP induced-bladder inflammation in the mice train tested. Because only subtle changes and no obvious inflammatory process are identified in IC/PBS patients, systematic CYP administration in C57BL/6 mice may sever as a feasible animal model to study the human bladder functional disorders.
53 37 Interestingly, the ability of CYP to produce moderate bladder edema without significant bladder inflammation in the mouse contrasts with results of previous work in the rat. In present study, 100mg/kg CYP administration in mice daily for five continuous days did not induce bladder hemorrhage or enhance MPO activity significantly, except the edema in suburothelial region. However, rats treated by a same dosage of CYP for three times in day1, 3 and 5 exhibited great hemorrhage, edema, partial loss of urothelium of the bladders accompanied with a significant increase of bladder MPO activity (Dang et al., 2008). A study of CYP-induced visceral pain in multiple mouse strains also reported differences of pain behavior between mouse strains after CYP treatment. The source of the species and strains variation in the extent of CYP-induced bladder pathology is unclear. It might be as a result of different distribution and/or expression of functional mechano- and chemo-sensors and different transmission pattern of bladder sensory information among species and strains. Bladder sensory neurons exhibit increased cell excitability after bladder inflammation Sensory information from the bladder is transmitted to central nerves system through either the paired pelvic or the lumbar splanchinic nerves, with their cell bodies located in LS and TL DRGs. The active and passive membrane properties of sensory afferent neurons innervating the bladder have been described previously in Chapter 2 using whole-cell current-clamp technique. In general, LS bladder sensory neurons have a significantly more negative action potential threshold and a significantly greater proportion of firing action potential
54 38 in response to the purinergic agonist, suggesting that LS bladder neurons were generally more easily excited and more sensitive to purinergic agonists compared with the TL counterparts. Therefore, the present study was performed to determine whether LS and TL bladder sensory neurons had enhanced cell excitability and purinergic responses after tissue insult. After bladder inflammation induced by CYP, LS and TL bladder neuron excitability increased, as evidenced by a significant decrease in rheobase, and an increased membrane depolarization in responses to the purinergic agonist α, β-methylene ATP, suggesting inflammation-produced alteration in cell excitability and the subunit composition of P2X receptors. These results are consistent with a similar previous study in the rat (Dang et al., 2008), CYP-induced rat bladder inflammation significantly decreased the rheobase and action potential threshold, increased proportions of bladder neuron with spontaneous action potential firing, and also had greater membrane depolarization and more action potential in responses to purinergic agonists in both LS and TL bladder sensory neurons. These results indicate that purinoceptive bladder sensory neurons will be sensitized under conditions of bladder insults and relay sensory information to the spinal cord and higher nervous centre of viscerosensation. P2X function in bladder afferent neurons is enhanced after bladder inflammation As described in Chapter 2, Three types of purinergic currents were identified: sustained (homomeric P2X2) currents were detected only in LS neurons, rapidly activating, slow deactivating (heteromeric P2X2/3) currents
55 39 predominated in both LS and TL neurons, and fast activating/de-activating (homomeric P2X3) currents were detected only in TL neurons. Relative to TL bladder neurons, current density of slow current evoked by either ATP or α, β- methylene ATP was greater in LS neurons than the TL neurons. In addition, a greater proportion of LS (93%) than TL (77%) bladder neurons responded to purinergic agonists, suggesting that LS bladder neurons were generally more sensitive to purinergic agonists. However, there is no related study on purinergic signals in mouse bladder afferent neurons from both pelvic and lumbar splanchinic pathway. Therefore, inflammation-produced alteration of P2X currents in LS and TL mouse bladder neurons were examined by whole-cell voltage clamp technique. In LS bladder neurons, more than 40% exhibited P2X2 homomeric sustained currents after CYP treatment relative to ~10% of neurons from salinetreated mice. There was a corresponding decrease in the proportion of LS bladder neurons exhibiting P2X2/3 heteromeric slow currents (from 90% to 57%). There were no P2X3 homomeric currents in LS neurons from saline-treated (or naïve) mice and none were observed after bladder inflammation. In TL bladder neurons, in contrast, there were no changes in proportions of purinergic-evoked currents after CYP treatment relative to saline treatment; the P2X3 fast current density, however, was significantly increased after bladder inflammation. These results in mice differ from those previously reported in the rat. Dang et al (2008) compared rat LS and TL bladder neuron responses to purinergic agonists after CYP treatment for 3 days. After bladder inflammation in
56 40 the rat, the only current evoked by either ATP or α, β-met ATP in LS bladder neurons was a P2X2/3 heteromeric slow current (increasing from 87% to 100% of neurons studied); fast and mixed currents noted in naïve and saline-treated mice were absent. In rat TL bladder neurons, the proportion of neurons exhibiting P2X3 homomeric fast currents evoked by α, β-met ATP increased significantly from 22% (saline) to 43% after CYP treatment whereas P2X2/3 heteromeric slow currents evoked by both agonists were reduced by about 50%. In the present study, no significant change was observed in purinergic-evoked currents in mouse TL bladder neurons; the principal post-cyp change was a four fold increase in the proportion of LS bladder neurons exhibiting a P2X2 homomeric sustained current, a current not seen in rat LS or TL bladder neurons. The significant increase in P2X2 homomeric currents in LS bladder neurons after CYP treatment suggests that a homomeric P2X2 receptor is involved in sensitization of pelvic nerve bladder afferents, consistent with reduced urinary bladder reflexes, decreased pelvic nerve afferent responses to bladder distension, and decreased nociceptive responses to intraplantar formalin in P2X2 knockout mice (Cockayne et al., 2005). With respect to P2X3, the amplitude of the P2X3 homomeric fast current was significantly enhanced and its desensitizing time constant decreased in TL bladder neurons after CYP treatment, suggesting a role in sensitization of the splanchnic pathway, consistent with reduced urinary bladder reflexes in P2X3 knockout mice (Cockayne et al., 2005).
57 41 P2X expression in bladder sensory neurons after bladder inflammation The proportion of LS neurons from saline-treated mice that express only the P2X2 transcript is low, consistent with the ~10% of LS neurons that exhibited a sustained, homomeric P2X2 inward current to application of ATP, and doesn t increase significantly after CYP treatment. Similarly, the proportion of LS neurons that express only the P2X3 transcript very low, and no P2X3 homomeric fast currents were found in LS neurons. P2X2 gene transcription in TL bladder neurons was significantly increased after CYP treatment, suggesting enhanced purinergic (e.g., P2X2/3) signaling at the transcriptional level. Expression of P2X3 subunit by the immunohistochemistry staining of LS and TL DRGs from saline- and CYP-treated mice showed a consistent result with the study on naïve mice. Positive P2X3 immunoreactivity in mouse bladder neurons was not significantly altered by CYP treatment, which contrasts with the outcome of P2X3 immunohistochemistry staining in the rat (Dang et al., 2008). Bladder inflammation significantly increased the fraction of rat TL (from 40% to 68%) but not the LS DRG neurons (from 83% to 89%) that exhibited P2X3 immunoreactivity. Because of the very high proportion of P2X mrna detected and unchanged P2X3 expression in bladder neurons after bladder inflammation, post-transcriptional modulation of P2X receptor function and/or interaction with other molecules/signaling pathways may be important in bladder neurons and sensitization of bladder afferents.
58 42 In summary, the kinetic, pharmacological, and expression profiles of purinergic signals in mice bladder neurons under pathological states, in addition to the differences of purinergic transmission between the rat and mouse, suggest a great role of the lumbar splanchinic(tl) and pelvic(ls) sensory pathways in bladder hypersensitivity in the pathological states. Targeting the altered subunit composition of purinergic receptors might be useful for management of bladder inflammation or functional disorders (e.g., internal cystitis).
59 43 CHAPTER 4: EFFECT OF METABOTROPIC P2Y2 RECEPTOR ON BLADDER SENSORY NEURON EXITABILITY AND P2X RECEPTOR FUNCTION Introduction Interstitial cystitis (IC)/painful bladder syndrome is characterized by urge and increased urination frequency accompanied by chronic pelvic pain in the absence of a pathobiological condition to explain symptoms (Burkman, 2004;Nickel, 2004). One of the potential endogenous mediators of bladder discomfort and pain is adenosine triphosphate (ATP), which is released from bladder urothelium during distension or chemical stimulation (Ferguson et al., 1997;Birder et al., 2003) and an increased release of ATP is detected in IC patients (Sun et al., 2001a;Sun et al., 2001b). Both ionotropic P2X and metabotropic P2Y receptors could be activated by extracellular ATP (Ralevic and Burnstock, 1998). To date, seven P2X subunits (P2X1-P2X7) have been reported (North, 2002); among these P2X receptor subtypes, P2X2 and P2X3 subunits are widely expressed in peripheral neurons and are thought to play an important role in the transduction of both bladder sensory information and of nociceptive signals (Dunn et al., 2001;Burnstock, 2006). The P2Y receptor family has eight members (P2Y1, 2, 4, 6, 11-14) to date; They respond to endogenous purine and pyrimidine nucleotides (ATP, ADP, UTP, UDP) released from various tissues (von Kugelgen, 2006;Lazarowski and Boucher, 2001;Lazarowski and Harden, 1999). There is growing evidence that
60 44 metabotropic P2Y receptors are involved in neural signal transduction and modulation. P2Y receptors in sensory neurons have been reported to contribute to innocuous mechanosensory transmission (Nakamura and Strittmatter, 1996), activate cutaneous afferents (Stucky et al., 2004), and mediate internal Ca 2+ release, CREB phosphorylation, and release of CGRP (Molliver et al., 2002;Sanada et al., 2002;Song et al., 2007). P2Y receptors also potentiate TRPV1 channel signaling gated by capsaicin or acid and reduce its thermal threshold in peripheral afferent neurons (Moriyama et al., 2003;Tominaga et al., 2001). Therefore, P2Y signaling is likely to play a role in mechanosensation and/or nociception under normal or pathological conditions. Two different pathways, lumbar splanchnic and pelvic nerve afferent pathways, convey sensory information from the bladder to the spinal cord (Vera and Nadelhaft, 1992;Uemura et al., 1975;Uemura et al., 1973). The cell bodies of these pathways are located, respectively, in thoracolumbar (TL) and lumbosacral (LS) dorsal root ganglia (Andersson, 2002;Applebaum et al., 1980). Based on their differential responses to multiple mechanical and chemical stimuli, these different bladder nerves have some similar but also different functions, suggesting they may exert different influences in the regulation of bladder function under normal or pathological conditions (Dang et al., 2005a;Xu and Gebhart, 2008). In the present report, I examined the effect of the P2Y agonist UTP on bladder neuron excitability and the interaction between P2X and P2Y
61 45 receptors in bladder sensory neurons from both lumbar splanchnic and pelvic pathways. Results A total of 194 LS and 118 TL bladder neurons were studied. Consistent with Chapter 2, the majority of LS and TL bladder neurons responded to P2X receptor agonists. Three types of purinergic currents were identified: (1) sustained currents, mediated through homomeric P2X2 receptors, were detected only in ~10% of LS purinoceptive neurons; (2) rapidly activating, slow deactivating currents, mediated through heteromeric P2X2/3 receptors, predominated in both LS (~90%) and TL (~75%) purinoceptive neurons; and (3) fast activating/de-activating currents, mediated through homomeric P2X3 receptors, were detected only in ~25% of TL purinoceptive neurons. Sustained currents were evoked by the purinergic agonist ATP, but not α,β-methylene ATP, whereas slow and fast deactivating currents were produced by both ATP and α,β-methylene ATP. I found that the effects of UTP were essentially restricted to LS bladder neurons; TL bladder neurons were not greatly affected. Accordingly, data presented here were mainly collected from LS bladder neurons unless indicated otherwise. UTP increases bladder sensory neuron excitability I examined the active membrane properties of 24 LS bladder neurons from naïve mice by injecting currents before and after UTP application (1μM, 40s) in whole-cell current clamp mode (Table 8). Input resistance was calculated according to the I/V relationship by injecting a series of hyperpolarizing pulses
62 46 ranging from 300 to 0 pa (30 ms) in 50pA increments. To determine rheobase, a series of 10 ms current pulses in 20pA increments (1s apart) were injected. The maximum current (pa) that did not evoke an action potential was taken as rheobase. Action potential (AP) threshold was determined from the inflection point where membrane potential started to dramatically rise and the phase plot slope (the first derivative of membrane potential, dv/dt) reached 10 mv/ms (Naundorf et al., 2006). AP amplitude was measured from resting membrane potential (RMP) to the peak of the AP, AP overshoot was the amplitude from 0 mv to the peak of the AP, AP duration was determined at 50% of the AP amplitude between the rising and falling phases and the AP falling rate was the velocity of change in potential from the AP peak to RMP. As summarized in Table 8, UTP application (1μM, 40s) depolarized LS bladder neurons; the RMP (-63.9±0.9 mv before UTP treatment) was significantly less negative (-55.6±1.5; P<0.005) after UTP. The rheobase was also significantly lower after UTP application relative to control (from 142.4±11.6 pa to 82.2±5.2 pa; P<0.01) in bladder sensory neurons (Figure 16A-B). Current injection (2x rheobase, 500 ms) depolarized and evoked AP firing in bladder sensory neurons. UTP significantly increased the number of action potentials in response to 2x rheobase current injection (Figure 16D) relative to before UTP (Figure 16C). In addition, about one-half of the bladder neurons tested (10/21) exhibited sustained spontaneous activity after UTP application (Figure 16E). UTP did not induce changes in input resistance, AP threshold, duration, amplitude, overshoot or falling rate (Table 8).
63 47 To further investigate the effect of UTP on bladder neuron excitability, I applied suramin (50μM), a nonselective P2 antagonist, on total 15 LS bladder DRG neurons 2 minutes before and during UTP application. Suramin prevented the effects of UTP on bladder sensory neuron excitability; the data are summarized in Table 8. Suramin blocked the UTP-produced reduction in membrane depolarization of LS bladder, the decrease in rheobase and the increase in number of action potentials in response to 2x rheobase current injection (all P<0.01). In addition, the spontaneous AP firing of bladder sensory neurons induced by UTP was completely inhibited by suramin (0/15). As UTP does not act as an ionotropic P2X agonist (Burnstock, 2007;von Kugelgen, 2006), these outcomes suggest a role of P2Y receptors in bladder sensory neuron hypersensitivity. Effect of UTP on purinergic agonist-evoked responses Because endogenous ATP could activate both P2X and P2Y receptors (principally P2Y2 and P2Y4 receptors), I hypothesized that there is an interaction between P2X and P2Y receptors in response to purinergic agonists. To examine the contribution of P2Y receptors to purinergic responses in mouse bladder sensory neurons, I applied the P2X/Y agonist ATP or P2X-selective agonist α, β- methylene ATP (30μM, 4s) followed by a 2-minute washout period, then applied external solution containing UTP (1μM) or nothing (control) for 40 sec before repeating the ATP or α, β-methylene ATP application. After another 2-minute washout period, currents induced by a third exposure to ATP or α, β-methylene ATP were recorded. Because ~10% of LS bladder neurons exhibiting ATP
64 48 evoked-sustained currents do not respond to the P2X-selective agonist α, β- methylene ATP, which activates homomeric P2X3 and heteromeric P2X2/3 receptors, and no effective P2X2 agonist is available, I used ATP as a homomeric P2X2 receptor agonist in the following experiments. In addition, ATP application was limited to a relative short duration (4 sec) and immediately followed by a bath solution washout to minimize possible activation of P2Y receptors. I examined the passive membrane properties of bladder neurons in response to P2X agonists after UTP in whole-cell current clamp mode. UTP application did not produce statistically significant differences in the magnitude of membrane depolarization produced by ATP or α, β-methylene ATP, or frequency of bladder neuron AP firing in response to ATP or α, β-methylene ATP (Table 8). However, purinergic responses of some bladder neurons (8/21) were sensitized by UTP. They either exhibited AP firing during application of UTP (Figure 17A) or an increased frequency of APs (Figure 17B) during purinergic agonist application compared with responses before UTP application. Other neurons (13/21) did not exhibit significant changes (Figure 17C-D). Because the P2 antagonist suramin also greatly inhibits P2X responses (data not shown), membrane properties of bladder neurons in response to ATP or α, β-met ATP after suramin treatment can not be examined. I also examined the effect of UTP on P2X agonist-evoked inward currents in whole-cell voltage clamp mode. Because the effect of UTP was evaluated by comparison between first and second application of purinergic agonist, I first
65 49 calculated the recovery kinetics from desensitization of P2X currents. The application interval between two purinergic agonist applications was increased stepwise. The current amplitude of the second application was standardized to the first application and plotted against time using a single-exponential fit. The sustained current evoked by ATP recovered extremely rapidly and did not show obvious decay (Figure 18A-B), whereas the slow current required around 40s for complete recovery (Figure 18C-D) and fast current required a even longer period (~120s) for recovery than slow current (Figure18E-F). Based on the results of curve fitting, the recovery time constant of the ATP-evoked slow current was 16.8s (Figure 18G) and of ATP-evoked fast current was 70.3s (Figure 18H). The time constant of the sustained current could not be determined because the data were not fit to the single exponential equation. All data were collected in LS bladder neurons, except that data of fast currents were collected in TL bladder neurons because no fast currents were observed in LS neurons. The recovery kinetics of the slow and fast current evoked by α, β-methylene ATP was similar to slow current evoked by ATP presented above. UTP application (0.3μM) for 40s significantly facilitated ATP-evoked sustained currents (Figure 19A) and fast currents (Figure 19C), but not ATPevoked slow currents (Figure 19B), suggesting that metabotropic P2Y receptors enhanced the responses of homomeric P2X2 and P2X3 receptors in bladder sensory neurons. However, this effect could instead be due to the repetitive application of ATP because ATP is also an agonist at P2Y receptors, although the application period of ATP (4S) was relatively short. Therefore, I repeated ATP
66 50 applications (30μM, 4s) three times using the same protocol described above but without UTP application. ATP-evoked sustained (Figure 20A) and slow (Figure 20B) currents did not significantly change after repetitive ATP application. However, I still observed a facilitatory effect on the fast current evoked by second ATP application comparing with the current evoked by first ATP application in the same neuron (Figure 20C). Figures 19D and 20D summarize the effect of UTP and repetitive ATP application on ATP-evoked sustained, slow in LS bladder sensory neurons and fast currents in TL bladder sensory neurons. There was no significant effect of UTP application on α, β-met ATP-evoked slow currents in LS bladder neurons. To further confirm the facilitatory effect of UTP on ATP-evoked sustained currents, I determined the dose-response relationship for UTP, increasing UTP concentrations from 30nM to 10μM. To eliminate the variation in current amplitude, I normalized P2X currents under different UTP concentrations to those associated with application of 30nM UTP. As an example given in Figure 21A, the effect of UTP on ATP-evoked sustained currents was concentrationdependent. The EC 50 and 95% confidence interval for UTP was 0.52μM ( μM; n=7, Figure 21B). Because there was no significant effect of UTP on slow currents, I did not determine the dose-response relationship of UTP on slow currents evoked by ATP or α, β-met ATP. These outcomes confirmed that UTP application facilitates homomeric P2X2-mediated sustained currents.
67 51 Metabotropic P2Y2 receptor mediates the effect of UTP UTP is an agonist at P2Y2 and P2Y4 receptors, which are coupled to Gq and activate the phospholipase C (PLC)/protein kinase C (PKC) signaling pathway (von Kugelgen, 2006). To confirm that the UTP effect was mediated via G-protein coupled receptors, an internal solution containing GDP-β-S (100μM), a global G-protein blocker inhibiting all G-protein-mediated events in the cytoplasm, was applied using the same repetitive ATP/UTP application protocol as above. As shown in Figure 22A and 23A, GDP-β-S repressed the facilitatory effect of UTP on the purinergic sustained current, but not the fast current. In addition, the facilitatory effect of repeated ATP on the fast current was also not inhibited by intracellular GDP-β-S, whereas the trend towards increasing sustained currents by repetitive ATP application was absent in the presence of GDP-β-S (Figure 22B and 23B). These outcomes suggest that the effect of UTP on homomeric P2X2 sustained currents in bladder sensory neurons is dependent on a G protein-coupled pathway, but the effects of UTP and ATP on homomeric P2X3 fast currents are not related with G protein-coupled receptors, consistent with previous studies reporting that P2X3 receptor can be sensitized by triphosphate nucleotides (ATP, UTP and GTP) via activation of the ecto-pkc phosphorylation site in the extracellular loop of P2X3 receptor subunit (Stucky et al., 2004;Wirkner et al., 2005). I also applied a PKC inhibitor (myristoylated Protein Kinase C Inhibitor 20-28, 10μM) to LS bladder neurons to further examine whether the effect of UTP on
68 52 P2X2 sustained current is PKC dependent. As shown in Figure 22C and 23C, the facilitation of sustained current by UTP was significantly inhibited by the PKC blocker, but the slow current amplitude after UTP application was not affected by the PKC blocker. Because most P2Y receptor antagonists presently available (e.g., PPADS, suramin, reactive blue, etc.) also block P2X receptors, I applied ATP and UTP as above to bladder sensory neurons taken from P2Y2 knockout mice to confirm whether P2Y2 mediates the facilitatory effect of UTP on P2X currents. Figure 23D shows significant decreases of both ATP-evoked sustained and slow currents after UTP application in LS bladder neurons from P2Y2 knockout mice. Comparison of sustained currents after UTP between wild type and knockout mice (Figure 22D) also revealed a significant facilitatory effect of UTP on purinergic currents in neurons from wild type but not P2Y2 knockout mice, indicating that P2Y2 is required for P2X2 receptor modulation by UTP. P2X and P2Y receptor expression in bladder sensory neurons Because the numbers of bladder sensory neurons contained in LS and TL DRG are relatively few, I employed single cell RT-PCR and single cell nested PCR to examine P2X and P2Y receptor expression in bladder sensory neurons. When cdna was harvested after single cell RT-PCR, the mouse GAPDH gene was amplified by conventional PCR as an internal control. Only cells positive for GAPDH amplicon were further processed by nested PCR. Negative results of GAPDH amplification were thought to have an unsuccessful reverse transcription reaction or a failed collection and thus discarded. The remaining bladder neurons
69 53 were amplified by two rounds of PCR with external primers, then internal primers, respectively. Figure 24A shows an example of positive single cell RT-PCR amplicons of P2X2, P2X3, P2Y2 and P2Y4 mrna. Product length corresponded to the expected size of the targeted region. Fifteen LS and fifteen TL bladder DRG neurons per mouse (total 3 animals) were collected for single cell PCR. The expression pattern of P2X2 and P2X3 subunit transcript has been discussed in a Chapter 2. The P2Y2 transcript was more abundant in LS (46.7±10.1%) and TL (37.8±2.3%) bladder neurons than the P2Y4 transcript (LS: 22.2±5.8%; TL: 13.3±3.8%). The frequencies of P2Y2 and P2Y4 transcript expression did not differ significantly between LS and TL bladder neurons (Figure 24B). The expression of P2Y2 and P2Y4 was also examined in saline- and CYP-treated mice. CYP treatment didn t change the P2Y receptor expression in bladder sensory neurons compared with saline controls. Figure 24C and 24D shows the proportions of P2Y transcripts in P2X transcript-positive bladder sensory neurons. In P2X2 transcript-positive bladder neurons (Figure 24C), 51.2% of LS and 52.4% of TL bladder neurons also expressed P2Y2 mrna, whereas only 22.0% of LS and 19.1% of TL bladder neurons expressed P2Y4 mrna. 17.1% of LS and 14.3% of TL bladder neurons expressing P2X2 mrna were found to express both P2Y2 and P2Y4 transcripts. Similarly, in P2X3 transcript-positive bladder neurons (Figure 24D), 51.2% of LS and 40.5% of TL bladder neurons also expressed P2Y2 mrna, whereas 24.4% of LS and 14.3% of TL bladder neurons expressed P2Y4 mrna. 19.5% of LS
70 54 and 9.5% of TL P2X3 mrna-positive neurons also expressed P2Y2 and P2Y4 transcripts. The frequencies of P2Y2/P2Y4 transcript expression in P2X2/P2X3 transcript-positive cells did not differ between LS and TL bladder sensory neurons. Discussion In the present study, I demonstrate that extracellular UTP sensitizes bladder sensory neurons and increases cell excitability by depolarizing resting membrane potential, decreasing rheobase and inducing action potentials. Our results extend previous findings that UTP evokes action potentials in isolated DRG neurons and identified nociceptors (Molliver et al., 2002;Stucky et al., 2004). About half of bladder sensory neurons are more sensitive to purinergic agonists after UTP application. Furthermore, UTP also facilitates the sustained currents mediated by homomeric P2X2 receptors through metabotropic P2Y2 receptors in LS bladder sensory neurons. Finally, P2Y2 transcripts are expressed in ~50% of LS and ~40% of TL bladder sensory neurons, with similar frequency in P2Xpositive LS and TL bladder sensory neurons, whereas P2Y4 transcripts are expressed in ~20% of LS and ~10% of TL bladder sensory neurons. These outcomes suggest that ATP or UTP released in an extracellular environment can sensitize primary afferent neurons and enhance responses of ionotropic P2X receptors to their endogenous purinergic agonists (e.g., ATP) via P2Y receptors. Both ATP and UTP are released into the extracellular environment under normal conditions, and their release is enhanced when cells are injured or damaged (Cook and McCleskey, 2002;Lazarowski and Harden, 1999),
71 55 suggesting that endogenous purines and pyramidines act as signaling molecules in response to environmental stimuli and play a role in nociceptive transduction (Burnstock, 2006;North, 2002). The present study shows that application of UTP sensitizes bladder sensory neurons via the P2Y2 receptor, which is consistent with other studies that UTP and ATP excite sensory neurons and activate cutaneous afferent fibers by evoking sustained action potential firing or reducing mechanical response thresholds (Lechner and Lewin, 2009;Molliver et al., 2002;Stucky et al., 2004). Our results also indicate that activation of metabotropic P2Y receptors by relatively low concentrations and prolonged application of agonist (e.g., UTP), which does not induce an obvious inward current, can facilitate ionotropic P2X receptor-mediated responses to subsequent purinergic agonist administration. These results suggest that exposure to UTP/ATP in a low concentration may prime subsequent purinergic signaling via metabotropic P2Y receptors and elevate the sensitivity of ionotropic P2X receptors. Studies using P2X2 knockout and P2X2/P2X3 double knockout mice indicate that both P2X2 and P2X3 play important roles in the transduction of noxious sensation from the bladder and in the regulation of normal bladder function (Cockayne et al., 2005). Purinergic currents mediated by different P2X receptor subtypes were previously characterized in Chapter 2. I found in the present study that only homomeric P2X2-mediated sustained currents in LS bladder sensory neurons were facilitated by UTP. The heteromeric P2X2/3- mediated slow currents in both LS and TL bladder sensory neurons were not significantly affected by UTP application. This suggests a selective effect of UTP
72 56 on a prolonged, sustained P2X2 current. In Chapter 3, I found that the frequency of sustained homomeric P2X2 currents increases about 4 fold (from 10% to 40%) in LS bladder sensory neurons from cyclophosphamide-induced bladder inflamed mice, implying that the effect of P2Y activation on P2X2 signaling could be dramatically amplified in pathological bladder disorders such as IC/PBS. Collectively, these findings suggest that homomeric P2X2 receptor contributes to purinergic signal transduction and sensitization of bladder sensory neurons. I examined the effect of UTP on both LS and TL bladder neurons because there is increasing evidence that pelvic and lumbar splanchnic sensory pathways innervating pelvic organs contribute differentially to visceral function and disorders (Brierley et al., 2005;Dang et al., 2005a;Xu and Gebhart, 2008;Sanada et al., 2002;Sugiura et al., 2005). I found a greater effect of UTP on LS than TL bladder neurons, suggesting a role in sensitization of the pelvic pathway innervating the bladder, consistent with increased activity of pelvic afferents by purinergic agonists in bladder-inflamed rat (Yu and de Groat, 2008). Among the P2Y receptor family, P2Y1, P2Y2, P2Y4, P2Y6 and P2Y11 are coupled to the Gq signal pathway, which activates phospholipase C and induces the release of intracellular Ca 2+ stores and protein kinase C activation; P2Y11 can also activate adenylate cyclase but is not expressed in rodents. On the other hand, P2Y12, P2Y13 and P2Y14 are coupled to G i and inhibit adenylate cyclase (von Kugelgen, 2006). Additionally, P2Y1, P2Y2, P2Y4 and P2Y6 receptors are detected in peripheral sensory neurons located in dorsal root and nodose ganglia (Ruan and Burnstock, 2003;Gerevich and Illes, 2004). Because only P2Y2 and
73 57 P2Y4 receptors are activated by UTP, I only examined expression of P2Y2 and P2Y4 in LS and TL bladder sensory neurons. A previous study on distribution of P2Y2 by in situ hybridization demonstrated that P2Y2 mrna is expressed ~90% of small size rat DRG neurons and only one third of large DRG neurons (Molliver et al., 2002); a similar study reported that 24% of rat DRG neurons expressed P2Y2 mrna (Kobayashi et al., 2006). I found in the present study that ~50% of bladder sensory neurons, which are small-to medium in size according to the analysis of cell size described in Chapter 2 but represent only about 5% of the neurons in their respective DRG, expressed P2Y2 mrna. The P2Y4 receptor is less widely expressed than P2Y1 or P2Y2 receptors (Ruan and Burnstock, 2003;Kobayashi et al., 2006), which is consistent with our finding that 10%-20% of bladder neurons express P2Y4 mrna. The present study established that the effect of UTP on ATP-evoked currents is mediated by P2Y2 receptors because the facilitatory effect of UTP is absent in P2Y2 knockout mice. Additional support for a role of P2Y2 over P2Y4 is that the effect of UTP was blocked by suramin, which is an antagonist at P2Y2 but not P2Y4 receptor. Other studies in P2Y2 knockout mice exhibit decreased calcium flux induced by UTP, reduced TRPV1 function and impaired thermal nociception (Malin et al., 2008). P2Y2 knockout mice also exhibit an altered osmotic reabsorption of water which influences volume and concentration of the urine stored in urinary bladder (Zhang et al., 2008), suggesting P2Y2 receptor is involved in regulation of urination mechanism. Interestingly, I found UTP application had an inhibitory effect on both slow and sustained purinergic
74 58 currents in P2Y2 knockout mice, suggesting that UTP may be involved in a P2Y2-independent mechanism to inhibit P2X signaling in the absence of P2Y2. Under normal conditions, ATP released from urothelial cells in response to bladder distension activates P2X receptors in bladder afferents. Activation of P2X3-expressing sensory afferents contributes to bladder reflexes as well as sensation. In the present report, I describe a role for the ATP/UTP-gated receptor P2Y2 in the regulation of bladder neuron excitability and identify an interaction between P2X and P2Y receptors that leads to enhanced P2X signaling. Hyperexcitability and increased firing of bladder afferents are associated with urinary urgency, frequency and pain, all of which are features of interstitial cystitis (Nazif et al., 2007). These findings provide a new dimension to the role of nucleotide signaling in bladder function that may contribute to the pathophysiology of bladder inflammation and injury.
75 59 CHAPTER 5: GENERAL CONCLUSIONS AND DISCUSSION Overview of experiment results The objective of this thesis is to test the hypothesis that the ionotropic P2X (principally P2X2 and P2X3) and the metabotropic P2Y2 receptors contribute to bladder sensory transmission. The present study of peripheral sensory neurons innervating mouse bladder provides evidence that LS and TL mouse bladder sensory neurons exhibit distinct purinergic signaling, which changes after bladder inflammation and can be modulated by activation of metabotropic P2Y2 receptor (a summary represented in Table 10). These outcomes support the role of P2X and P2Y2 receptors as mechano- and chemo-sensors in peripheral afferent neurons in bladder hypersensitivity following tissue insults. Therefore, some subtypes of purinergic receptor family are able to serve as potential targets for therapeutic intervention in human bladder disorders, e.g. interstitial cystitis/painful bladder syndrome. Differential purinergic signaling in LS and TL bladder sensory neurons The pelvic and lumbar splanchinic innervations of the urinary bladder have some different features in anatomy and function. It has been reported that pelvic nerve axons are uniformly distributed throughout the urinary bladder, whereas axons from the lumbar innervations are more numerous in the trigone region. In this thesis, I injected fluorescent dye (DiI or CTB) in the neck of the bladder (trigone area) and found twice as many DRG neurons projecting through the
76 60 pelvic nerves (PN) as the lumbar splanchnic nerves (LSN). Although it could cause an underestimation of bladder innervations that the bladder was not entirely labeled with retrograde tracing dye, a significantly higher density of the pelvic than the lumbar splanchnic innervation even in the trigone region suggests a greater contribution of pelvic pathway to bladder sensory transmission than lumbar splanchnic pathway. The pelvic and lumbar splanchinic innervations of the urinary bladder may also exert different functions. Recent reports have implicated the importance of P2X receptor in bladder function by in vivo and in vitro methods (Cockayne et al., 2000;Cockayne et al., 2005;Pandita and Andersson, 2002;Velasco et al., 2003;Vlaskovska et al., 2001;Yu and de Groat, 2008). In Chapter 2, I compared the P2X function in mouse bladder sensory neurons in the pelvic (LS) and splanchnic (TL) pathways, including the kinetics of activation/inactivation and pharmacologic antagonism of the inward currents evoked by purinergic agonists. In addition to similarities, I found that, in naïve mice, [1] a greater proportion of LS bladder neurons responded to purinergic agonists than did TL bladder neurons; [2] although the predominant current produced by both purinergic agonists in both LS and TL bladder neurons was a heteromeric P2X2/3 slow current, the slow current densities produced by both purinergic agonists in LS bladder neurons were significantly greater than in TL counterparts; [3] homomeric P2X2 sustained current was only detected in LS bladder neurons, whereas homomeric P2X3 fast current was only detected in TL bladder neurons.
77 61 The electrical properties of LS and TL bladder sensory neurons are also different, based on their active and passive membrane properties in response to current injection and agonist application. LS bladder neurons have a significantly lower action potential threshold than the TL counterparts and a greater proportion of LS neuron raise action potentials in response to ATP than TL neurons do, suggesting that PN bladder afferents more tend to be easily activated by membrane depolarization or ATP when released from urothelial cells, and therefore are more sensitive to bladder distension during the urine filling stage. A previous electrophysiological study of mouse bladder afferent fibers (Xu and Gebhart, 2008) provides comparable results showing a greater proportion of stretch sensitive bladder pelvic afferent fibers than lumbar splanchnic afferent fibers, and a greater dynamic response of PN afferents to mechanical stimulation. In conclusion, the outcomes of the present study supplement the fundamental knowledge of afferent signaling from the urinary bladder in the mouse using electrophysiological approach based on directly comparing the basic properties and purinergic responses of bladder afferent neurons in the pelvic and lumbar splanchnic sensory pathway. Contribution of P2X2 and P2X3 receptors to bladder sensory transmission It has been revealed that P2X2 and P2X3 receptors play a significant role in visceral mechanosensory transduction and hypersensitivity. The findings in knockout mice provide instrumental evidence in support of the importance of P2X receptors in bladder mechanosensation. P2X2, P2X3 and P2X2/P2X3 double
78 62 knockout mice exhibit micturition hyporeflexia and hyposensitivity of bladder afferent nerves in response to bladder distension whereas the ATP release from the urothelial cells evoked by bladder distension is not significantly changed compared with wild type mice (Cockayne et al., 2000;Cockayne et al., 2005;Vlaskovska et al., 2001). Electrophysiological studies indicate that P2X2 and P2X3 subunits account for virtually all ATP-mediated responses in peripheral sensory neurons (Cockayne et al., 2005), and the predominant P2X receptor(s) in dorsal root ganglia neurons are heteromeric P2X2/3 and homomeric P2X3 receptors (Burgard et al., 1999). Consequently, the great majority of bladder mechanosensation studies on purinergic signaling focus on the heteromeric P2X2/3 and homomeric P2X3 receptors. P2X3 antagonist TNP-ATP, which is a 1000-fold more potent in inhibiting homomeric P2X3 and heteromeric P2X2/3 receptors than homomeric P2X2 receptor, abolishes the bladder overactivity induced by intravesical infusion of ATP in conscious rats (Pandita and Andersson, 2002). TNP-ATP is also effective in inhibiting ATP or α, β-methylene ATP induced facilitation of bladder afferent never activity in the rat and mouse (Rong et al., 2002;Yu and de Groat, 2008). Consistent with previous findings, it has been shown in this thesis that the majority of LS and TL mouse bladder neurons exhibit heteromeric P2X2/3 mediated currents in response to ATP or α, β- methylene ATP. In addition, the responses of homomeric P2X3 receptor to ATP and α, β-methylene ATP, which are identified only in TL bladder neurons, is facilitated after bladder inflammation, suggesting that P2X3 receptor is important
79 63 in mechano-hypersensitivity of sensory afferents under pathological conditions of the bladder. Compared with the dominance of heteromeric P2X2/3 and homomeric P2X3 receptors in dorsal root ganglia, homomeric P2X2 receptor is found in a small percentage (~10%) of dorsal root ganglia neurons by patch-clamp studies in P2X3 knockout mice (Cockayne et al., 2005). Our findings of bladder sensory neurons from lumbosacral dorsal root ganglia show a consistent result that the purinergic responses of ~10% of purinoceptive LS bladder dorsal root ganglia neurons are mediated by homomeric P2X2 receptor. Although the homomeric P2X2 receptor is not the predominate P2X receptor in naïve mice, the proportion of LS bladder neurons exhibiting P2X2 receptor mediated-sustained currents dramatically increased 4 fold to ~40% in bladder inflamed mice, indicating a substantial role of P2X2 receptor in mediating mechanosensory transduction during bladder inflammation. Additionally, the activity of homomeric P2X2 receptor can be facilitated by metabotropic P2Y2 receptor through PKC pathway. In consideration of an increased ATP release from urothelium reported in interstitial cystitis patients (Sun et al., 2001a;Sun et al., 2001b) and the ability of ATP to activate both P2X and P2Y receptors, the positive interaction between P2X2 and P2Y2 receptors implies that the function of homomeric P2X2 receptor can be enhanced by the elevated concentration of ATP and modulation via P2Y2 receptor in pathological states of the urinary bladder. In a summary, the present study supports an important role for both P2X2 and P2X3 in viscerosensory transmission by exerting potentially different but also
80 64 overlapping functions. Given the interest in investigating P2X antagonists for conditions of bladder inflammation or functional bladder disorders as interstitial cystitis, the findings in the thesis suggest that selective antagonists of homomeric P2X2 and P2X3 receptors may have therapeutic potential in the treatment of these conditions. Contribution of P2Y2 receptor to bladder sensory transmission In comparison to P2X2 and P2X3 receptors, the contribution of the metabotropic P2Y2 receptor to bladder sensory transduction has not been well examined. Previous studies implicated that P2Y2 may have an indirect effect on bladder sensation by modulation of other receptors, ion channels or signal molecules. It has been reported that P2Y2 receptor potentiates TRPV1 channel activity and reduces its thermal threshold in peripheral afferent neurons (Moriyama et al., 2003;Tominaga et al., 2001), and TRPV1 channel is thought to be an important mediator of bladder mechanosensitivity (Birder et al., 2002;Cockayne et al., 2005;Tominaga et al., 2001). P2Y2 receptor in sensory neurons has also been implicated to mediate the Ca 2+ release from internal stores, CREB phosphorylation, and the release of CGRP (Molliver et al., 2002;Sanada et al., 2002), suggesting P2Y2 signaling is likely to play a role in visceral mechanosensation and/or nociception under normal or pathological conditions. In this thesis, UTP is shown to have an ability to increase cell excitability of bladder sensory neurons. Of the identified pyrimidine-binding P2Y receptors,
81 65 P2Y2 and P2Y4 are potently activated by UTP as well as ATP. The possibility that P2Y4 mediates the effect of UTP on bladder sensory neurons is ruled out by demonstrating that P2Y2 antagonist suramin could completely block the UTPinduced increased bladder neuron excitability. Examination of P2Y2 and P2Y4 expression further revealed a more widespread distribution of P2Y2 mrna than P2Y4 mrna. Therefore, the actions of UTP on bladder neuron activity are presumably mediated by P2Y2 receptor, suggesting P2Y2 receptor may be involved in bladder overactivity and hyperreflexia in functional bladder disorder and cystitis. In addition, electrophysiological studies in knockout mice reveal that P2Y2 receptor potentiates the homomeric P2X2 receptor responses, and the potentiation is G protein dependent and requires PKC activation. In a conclusion, whether direct or indirect, the contribution of P2Y2 receptor to bladder neurons excitability and purinergic signals extends the current understanding of P2Y receptors in bladder sensory transmission, indicating the potential of metabotropic P2Y receptors in visceral mechanosensation. Future directions In this thesis, I have shown that the differential purinergic signaling pathways of bladder pelvic and lumbar splanchnic innervations and the contribution of P2X and P2Y2 receptors to bladder sensory transduction by electrophysiological, single cell RT-PCR and immunohistochemistry methods. The facilitation of purinergic signaling in bladder sensory neurons after bladder inflammation and the influences of P2Y2 receptor on bladder neuron excitability
82 66 and P2X2 function, suggest that P2X and P2Y2 may be the potentially important mediators for bladder mechanosensation and hypersensitivity. Further experiments will identify the expression of P2X and P2Y2 subunit in bladder afferent terminals projecting into urothelium under both physiological and bladder inflammatory conditions, examine P2X and P2Y2 contribution to bladder function by behavior methods in naïve, P2X knockout or knockdown mice by RNA interfering technique, and confirm the facilitatory effect of P2Y2 receptor on activity of bladder afferent nerves using bladder-nerve preparation. These studies will help to elucidate the mechanism of P2X and P2Y2 in peripheral neurons innervating mouse bladder and the role they play in chronic pain and functional disorder (e.g., IC/PBS) in human.
83 67 CHAPTER 6: MATERIALS AND METHODS Animals Male C57BL/6 mice (6-8 weeks; Taconic Labs, Germantown, NY) were used for most experiments; P2X3 and P2Y2 knockout mice were also used [see refs in Cockayne et al., 2002 and Malin et al., 2008 for initial characterization of knockouts].. Mice were housed in polypropylene cages with ad libitum access to food and water. All protocols were reviewed and approved by the Institutional Animal Care & Use Committee, the University of Pittsburgh. Bladder neuron retrograde labeling Mice were anesthetized with 2% inhaled isoflurane (Hospira Inc., Lake Forest, IL), the bladder exposed via a lower abdominal incision ~5 mm in length and 10 ul of 1,1'-dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate (DiIC 18 (3); Molecular Probes, Eugene, OR; 0.2mg/ml in DMSO) was injected into 3-4 sites within the bladder wall and the base around the trigone using a 30 gauge needle. Sterile cotton-tipped applicators were applied to injection sites to absorb any DiI that leaked from injection sites. The incision was closed (6.0 silk, Ethicon Inc., Somerville, NJ) and post-operative analgesia provided after surgery (buprenorphine, 0.05mg/kg, i.p.; Bedford labs, Bedford, OH). Cell dissociation and culturing Lumbosacral (LS; L6-S2) and thoracolumbar (TL; T13-L2) DRG were harvested for electrophysiological whole cell recordings two weeks after retrograde labeling. After dissection of DRG, the pelvic area around the bladder was examined for evidence of DiI leakage; leakage was rarely noted and DRG
84 68 from mice with evidence of leakage were not used. LS and TL DRG were incubated at 37 C, 5% CO 2 for 45 min in DMEM media containing 1% penicillin/streptomycin (Gibco, Invitrogen Corp. Grand Island, NY), 1 mg/ml type 4 collagenase and 1 mg/ml neutral protease (Worthington Biochemical Corp., Lakewood, NJ). Tissue was gently triturated and collected after 5 min centrifugation at 120 X g, washed 3 times and re-suspended in DMEM media containing 0.5 mm L-glutamine, 1% penicillin/streptomycin and 10% fetal bovine serum. The cells were plated on poly-d-lysine-coated coverslips (Becton Dickinson Labware, Bedford, MA) and incubated at 37 C, 5% CO 2 for hrs. Only DiI positive bladder sensory neurons were studied. All recordings were performed within 24 hours after plating. Whole cell current- and voltage-patch clamp recording Coverslips were transferred to a recording chamber perfused with an external solution (in mm) consisting of NaCl 140, KCl 5, MgCl 2 1, CaCl 2 2, glucose 10, and HEPES 10 at ph 7.4, 310 mosm. Glass micropipette tips were heat-polished to resistances ~ 2-3 MΩ and filled with an internal solution (in mm) consisting of KCl 130, NaCl 4, CaCl 2 0.2, HEPES 10, EGTA 10, MgATP 2, and NaGTP0.5 at ph 7.25, 300 mosm. After establishing the whole-cell configuration, membrane voltage was clamped at -70 mv using an Axopatch 200B amplifier (Axon Instruments, Union City, CA), digitized at 1 khz (Digidata 1350; Axon Instruments), and controlled by Clampex software (pclamp 9; Axon Instruments). Cell capacitance was obtained by reading the value from the Axopatch 200B
85 69 amplifier. In current clamp experiments, only neurons that had a resting membrane potential more negative than -50 mv and a distinct action potential overshoot >0 mv were studied. Drugs were applied through a 3-barrel glass pipette placed close (~100 m) to the cell using a fast-step SF-77B superfusion system (Warner Instruments, Hamden, CT). Agonists were applied for 4 sec at an interval of 2 min; antagonists were superfused for 30 sec before the application of agonists. The reagents were prepared fresh from stock solutions on the day of the experiment. ATP, α, β- methylene ATP, Guanosine-5'-triphosphate (GTP), 2,3 -O-(2,4,6- trinitrophenyl) adenosine 5 -triphosphate (TNP-ATP), and pyridoxal-phosphate-6-azophenyl- 2',4'-disulfonate (PPADS) were obtained from Sigma-Aldrich (St. Louis, MO). UTP, suramin, guanosine- 5'- O- (2- thiodiphosphate) (GDP-β-S ) and myristoylated Protein Kinase C Inhibitor were purchased from Calbiochem (La Jolla, CA All experiments were performed at room temperature (21 23 C). Drugs were prepared fresh from stock solutions on the day of the experiment. All experiments were performed at room temperature (21 23 C). Urinary bladder inflammation Two weeks after retrograde labeling, mice were treated intraperitoneally either with saline or cyclophosphamide (CYP; 100 mg/kg, dissolved in saline; Sigma-Aldrich Inc., St. Louis, MO) daily for 5 days; mice were sacrificed by CO 2 inhalation on day 6 and the LS (L6-S2) and TL(T13-L2) dorsal root ganglia (DRG) were removed for electrophysiological study or immunohistochemistry staining. Systemic administration of CYP, which is metabolized to the bladder irritant
86 70 acrolein(cox, 1979), can cause hemorrhagic cystitis in humans as an adverse event and produces a cystitis-like condition in rodents (e.g., (Bon et al., 2003). Bladder myeloperoxidase (MPO) assay Urinary bladders were removed from CYP- and saline-treated mice deeply anesthetized with inhaled isoflurane. Each bladder was longitudinally cut into two parts, one part for histological examination and the other for MPO assay. For the MPO assay, bladder tissue was rinsed with saline, patted dry and homogenized in ice-cold 50 mm phosphate buffer containing 0.5% hexadecyltrimethylammonium bromide. After three repeated freeze (dry ice) - thaw (37 C water bath) cycles, samples were centrifuged at room temperature for 1min at 200 X g. 10 l supernatant was diluted to 300 l in phosphate buffer containing 0.05% hydrogen peroxide and 0.5% o-dianisidine dihydrochloride, a ph-sensitive indicator dye. Absorbance of the diluted supernatant was measured by a spectrophotomer (SpetraMax Plus 384, Molecular Devices, ). All drugs were purchased from Sigma-Aldrich, Inc. Histological examination of bladder inflammation For histological evaluation, bladder tissue from CYP- and saline-treated mice deeply was rinsed in saline and fixed in 4% formaldehyde, embedded in paraffin and cut at 10 m thickness. Tissue sections were stained with 5% hematoxylin and eosin and evaluated by a pathologist (Robert H. Garman, DVM, DACVP, Consultants in Veterinary Pathology, Inc.).
87 71 Single cell RT-PCR Bladder DRG neurons were retrogradely labeled and mice sacrificed as described above. LS and TL DRG were removed and plated as described above for electrophysiological study. After hr incubation, coverslips were placed in the patch clamp recording chamber and perfused with sterilized external solution. DiI positive neurons were collected with glass pipettes (tip diameter ~ m) by gentle suction and expelled into 0.2 ml microcentrifuge tubes containing 2.5 l lysis buffer consisting of 1X first-strand buffer, 2U RNaseOut, 10 M dntp, 0.5% IGEPAL, and 0.05 g/ l Oligo(dT)12-18 primer. Tubes were incubated at 65 C for 1.5 minutes, then held at room temperature for 2 min. Another 2.5 l of RT-PCR buffer consisting of 50 U SuperScript II reverse transcriptase, 1X first-strand buffer, 2U RNaseOut, and 10 M dntp was rapidly added to each tube. Tubes were incubated at 37 C for 20 min, then at 65 C for 10 min to generate first strands of cdna sequence. Negative controls were tubes without labeled neurons or processed with RT-PCR buffer not containing reverse transcriptase. Only RT-PCR products of the batches that passed tests of negative controls underwent further PCR steps. All single cell RT-PCR reagents were purchased from Invitrogen Inc (Carlsbad, CA). Multiplex PCR and gene specific nested PCR P2X2, P2X3, P2Y2 and P2Y4 genes were amplified through two rounds of PCR (multiplex and nested PCR) from the cdna library of individual mouse bladder neurons. In the first round, two external primers of targeted genes were added together into a 25 l volume PCR solution containing 1X PCR buffer, 0.2mM
88 72 dntp, 1.6 M primers of each gene and 1U Taq Polymerase. In the second round, 1 l of first round PCR amplicons served as a template and two internal primers of individual genes were added to the 25 l PCR system. Both multiplex and nested PCR used the following PCR conditions: 1 cycle of 10 min at 95 C; 32 cycles of 94 C/30 sec, 52 C/30 sec, and 72 C/30 sec before a final extension step at 72 C for 10 min, after which 10 l of the nested PCR products was electrophoresed on a 2% (w/v) agarose gel at 100V for 25 min. After electrophoresis, the gel was stained with 0.005% ethidium bromide and bands of PCR products were visualized under UV light. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as an internal positive control. Single cell RT- PCR products that failed to pass internal positive tests were eliminated from the study. All primers were purchased from Integrated DNA Technologies, Inc. (Coralville, IA) and are listed in table 1. Immunohistochemistry Alexa Fluor 488-conjugated Cholera Toxin B subunit (CTB, 2μg/ml; Invitrogen Inc.) was injected into mouse bladder wall as described earlier for DiI labeling. After 5 days, saline or CYP was given daily for day 1 to day 5. On day 6, naïve, saline and CYP-treated mice were deeply anesthetized and perfused transcardially with ice cold PBS then ice cold 4% paraformaldehyde. LS and TL DRGs were rapidly removed, fixed in 4% paraformaldehyde for 4 h, and cryoprotected in 30% sucrose for 3 days. Frozen DRGs were sectioned at 10 µm using a Leica 3M 3050 cryostat; one section was selected every 50 µm to minimize double counting. DRG sections were blocked with 2% donkey serum in
89 M phosphate-buffered saline (PBS, Sigma Aldrich) for 2 h before incubation in primary antibody (rabbit anti-mouse, 1:2000; Neuromics, Edina, MN) for 24 h, 4 C, after which the primary antibody was aspirated and sections were washed three times for 15 min with 0.01 M PBS. Secondary antibody, Rhodamine Red X- conjugated conjugated to donkey anti-rabbit IgG (Jackson Immuno Research, West Grove, PA), was applied for 4 h and washed three times for 15 min with 0.01 M PBS. Sections were mounted with Fluorescence Mount (Sigma Aldrich Inc.) and viewed with Olympus confocal microscope B211 equipped with separate fluorescence filters. Images were captured and processed by Fluoview Software. All cells positive for CTB fluorescence were counted as bladder sensory neurons and the immunoreactivity of P2X3 subunit protein was quantified. For control experiments, the incubation of primary antibody was omitted. No labeling was observed (data not shown). Data and statistical analysis Data are presented as means ± SE. Analyses were performed using the software package GraphPad Prism 5 (GraphPad Software, San Diego, CA). Dose response curves were generated using the following equation: Y=A+ (B-A)/ (1+10^[(LogEC50-X)]), where X is the logarithm of concentration; Y is the response and starts at A and goes to B with a standard Hill slope of 1.0. Desensitization kinetics were fitted with a standard exponential equation: Y = K 0 + K 1 x exp( t/τ), where Y is the current amplitude at time t, K 0 is the amplitude of the sustained component, and τ is the time constant. K 0 and K 1 represent the contribution to current amplitude from the fast and slow components of the
90 74 current, respectively. For dichotomous variables, a Fisher's exact test was used. Unpaired two-tailed t-tests were used for parametric measures if data were collected from different animals or cells; paired t-tests were applied when comparing current amplitudes from the same cell. Comparison of normalized data from independent group was carried by nonparametric Mann-Whitney U-test. Results were considered statistically significant when P<0.05.
91 75 Table 1. Purinergic currents in LS and TL bladder sensory neurons from naïve mice LS neurons TL neurons ATP % responding 93.0%** (106/114) 76.9% (70/91) sustained 10.4% (11/106) na slow 89.6%* (95/106) 74.3% (52/70) fast na 25.7%(28/70) α,β-met ATP % responding 83.3% (95/114) 76.9% (70/91) sustained na na slow 100% (95/95) 74.3% (52/70) fast na 25.7% (28/70) Note: *, P < 0.05, **, P < 0.01 vs. the corresponding TL current type. na, not applicable.
92 76 Table 2. Properties of purinergic currents in LS and TL bladder sensory neurons from naive mice ATP α, β-met ATP current density (pa/pf) time to peak (ms) desensitiz ing tau (ms) current density (pa/pf) time to peak (ms) desensitiz ing tau (ms) sustained 15.5± ±86 na na na na LS slow 25.7±1.9** 641± ± ±1.1* 1497± ±582 fast na na na na na na sustained na na na na na na TL slow 17.0± ± ± ± ± ±662 fast 9.7± ±32 224±18 6.6± ±36 211±25 Note: Data are means ± SEM. *, P < 0.05, **, P<0.01 vs. the corresponding TL current type. na, not applicable
93 77 Table 3. Passive and active electrical properties of LS and TL bladder sensory neurons LS TL RMP (mv) -70.4± ±1.75 Input Resistance (M ) 526.1± ±41.4 Rheobase (pa) ± ± 14.5 Current injection AP threshold (mv) -34.1±0.5 ** -28.5±0.8 AP amplitude (mv) 117.1± ±2.3 AP duration (ms) 3.6± ±0.3 AP overshoot (ms) 51.4± ±1.4 AP falling rate (mv/ms) 19.2± ±1.2 Spontaneous AP (%) 0 0 ATP (30 M) α,β-met ATP (30 M) Depolarization (mv) 25.5± ±4.6 AP firing frequency 21/32 * 10/30 Depolarization (mv) 15.9± ±2.2 AP firing frequency 10/32 5/30 Note: Data are means ± SEM. *, P < 0.05; **, P<0.01 vs. the corresponding TL current type.
94 78 Table 4. Summary of P2X3 immunoreactivity in LS and TL bladder DRG neurons from naïve, saline- and CYP-treated mice LS TL naïve saline CYP naïve saline CYP P2X3 in DRG neurons 27.2%* (83/305) 23.6%* (164/696) 19.7%* (104/529) 36.4% (156/429) 36.3% (171/470) 37.0% (181/489) CTB in DRG neurons 7.2%* (27/305) 8.0%* (56/696) 6.6%* (35/529) 2.8% (12/429) 2.3% (11/470) 2.0% (10/489) P2X3 staining 5.0%** 3.5%** 2.8%** 50.0% 63.6% 50.0% in CTB (1/20) (2/56) (1/35) (6/12) (7/11) (5/10) neurons Note: *, P < 0.05 vs. the corresponding TL current type.
95 79 Table 5. Passive and active electrical properties of LS and TL bladder neurons from saline- and CYP-treated mice LS TL current injection RMP (mv) input resistance(m ) rheobase (pa) saline -70.5± ±2.7 CYP -68.6± ±2.1 saline ± ±45.1 CYP ± ±29.9 saline 144.3± ±22.6 CYP 110.9± ±8.5 AP threshold saline -33.4± ±1.7 (mv) CYP -35.6± ±1.1 AP amplitude saline 121.1± ±4.2 (mv) CYP 115± ±2.3 AP duration (ms) saline 4.3± ±1.7 CYP 4.5± ±1.1 AP overshoot saline 50.6± ±2.3 (ms) CYP 50.5± ±2.0 AP falling rate saline 19.5± ±1.7 (mv/ms) CYP 21.0± ±1.3 ATP (30 M) depolarization saline 25.8± ±3.1 (mv) CYP 32.8± ±2.4 AP firing saline 19/25 * 10/23 frequency CYP 28/33 ** 10/28 α,β-met ATP (30 M) depolarization (mv) AP firing frequency saline 15.0± ±1.8 CYP 26.5± ±1.7 saline 7/25 3/23 CYP 8/33 5/28 Note: Data are means ± SEM. *, P <0.05, **, P<0.01 vs. TL counterparts;, P < 0.05 vs. saline treatment.
96 80 Table 6. Purinergic currents in LS and TL bladder sensory neurons from saline- and CYP-treated mice ATP αβ-metatp saline-treated CYP-treated saline-treated CYP-treated LS TL % responding sustained slow 96.9%* (31/32) 9.7% (3/31) 90.3% (28/31) 93.3% (42/45) 42.9% (18/42) 57.1% (24/42) 87.5% (28/32) na 100% (28/28) 53.3% (24/45) na 100% (24/24) fast na na na na % responding 78.6% (22/28) 85.3% (29/34) 78.6% (22/28) 85.3% (29/34) sustained na na na na slow fast 72.7% (16/22) 27.3% (6/22) 79.3% (23/29) 20.7% (6/29) 72.7% (16.22) 27.3% (6/22) 79.3% (23/29) 20.7% (6/29) Note: *, P < 0.05 vs. the corresponding TL current type., P < 0.05,, P < 0.01, vs. control (saline) treatment. na, not applicable
97 81 Table 7. Properties of purinergic currents in LS and TL bladder sensory neurons from saline- and CYP-treated mice ATP α,β-met ATP Current type current density (pa/pf) time to peak (ms) Treatment Desensitizing tau (ms) current density (pa/pf) time to peak (ms) Desensitizing tau (ms) sustained saline 15.5± ±759 n/a n/a n/a n/a CYP 20.9± ±244 n/a n/a n/a n/a LS slow saline 25.2± ± ± ± ± ±850 CYP 31.6± ± ± ± ± ±1309 fast saline n/a n/a n/a n/a n/a n/a CYP n/a n/a n/a n/a n/a n/a sustained saline n/a n/a n/a n/a n/a n/a CYP n/a n/a n/a n/a n/a n/a TL slow saline 17.4± ± ± ± ± ±4190 CYP 10.9± ± ± ±0.9 * 2202± ±2499 fast saline 9.1± ± ± ± ± ±4.1 CYP 24.2± 5.0 * 242.4± ±6.5 * 14.4±2.3 * 258.0± ±5.0 Note: Data are means ± SEM. *, P < 0.05 vs. the corresponding saline-treated group. na, not applicable
98 82 Table 8. Passive and active electrical properties of LS bladder neurons in the absence and the presence of UTP or UTP and suramin (SUR) application No UTP (n=39) UTP (n=21) UTP+SUR (n=15) RMP (mv) -63.9± ±1.5 *** -64.9±2.8 Input Resistance (M ) 242.0± ± ±17.1 Rheobase (pa) 142.4± ±5.2 ** 146.2±16.5 AP threshold (mv) -32.6± ± ±1.1 Current Injection AP amplitude (mv) 107.3± ± ±4.4 AP duration (ms) 4.6± ± ±0.9 AP overshoot (ms) 44.5± ± ±1.8 AP falling rate (mv/ms) AP frequency, 2X rheobase UTP induced-ap firing 19.3± ± ± ± ±1.0 ** 3.1±0.5 0/39 10/21 ** 0/15 ATP (30 M) Depolarization (mv) 20.3± ±3.4 na Number of APs 1.1± ±0.8 na α,β-met Depolarization (mv) 13.3± ±1.1 na ATP (30 M) Number of APs 0.8± ±0.6 na Note: Data are means ± SEM. *, P < 0.05; **, P<0.01; ***, P<0.005 vs. Naïve., P < 0.05;, P<0.01;, P<0.005 vs. UTP application. na, not applicable
99 83 Table 9. Summary of contribution of P2X and P2Y receptors to mouse bladder sensation Bladder afferent pathway LS bladder neurons Pelvic TL bladder neurons Lumbar splanchnic /hypogastric % of DRG neurons 6% 2% Changes after bladder inflammation Changes after UTP application Cell excitability Cell excitability Cell excitability Cell excitability Purinoceptive neurons % % expressing P2XR transcripts % expressing P2YR transcripts 93% of LS bladder neurons 77% of TL bladder neurons P2X2: 89% ; P2X3: 93% P2X2: 51%; P2X3: 98% P2Y2: 47%; P2Y4: 22% P2Y2: 38%; P2Y4: 13% P2X/P2Y transcripts after bladder inflammation No change No change % co-expressing P2XR and P2YR P2Y2: ~50% of P2X cells P2Y4: ~20% of P2X cells P2Y2: ~45% of P2X cells P2Y4: ~15% of P2X cells Functional P2XR (current type, % of purinoceptive neurons) P2X2 (sustained, 10%) P2X2/3 (slow, 90%) P2X3 (fast, 26%) P2X2/3 (slow, 74%) Changes of P2XR after bladder inflammation Modulation by UTP/ATP % % Function No change Function No change Function No change Modulation by P2Y2 receptor Yes, Gq-PKC pathway No No, GPCR independent No
100 84 Table 10. External and internal primers for mouse P2X2, P2X3, P2Y2, P2Y4 and GAPDH cdna P2X2 (NM_153400) P2X3 (NM_145526) P2Y2 (NM_008773) P2Y4 (NM_020621) GAPDH (NM_008084) External Forward External Reverse Internal Forward Internal Reverse External Forward External Reverse Internal Forward Internal Reverse External Forward External Reverse Internal Forward Internal Reverse External Forward External Reverse Internal Forward Internal Reverse External Forward External Reverse CTCTTCAGTAACCATGTCCACG CCGGAAGACAGCTCTAATTTGG GAAGATAGGCATCTTGCTCTGG GGGATCCTATGAGGAGTTCTGT GCTCCCTAGAAGAAGATGGAGA CTGTGTGACCATGTTAGGGATG TGTCCTAAGAGGATCCTGTACC GGCATCTAGCACATAGAAGTGG GGGAGAGTAGTGTAGCTGATGA GTCCTTGAGATCATGAGGCTTG GTAGATGCCACACCTATCCAAC CCTTGAGATCATGAGGCTTGTC CCCAAGAGTTGGTAGTAGACAC CGTGCTCTTTGGTCTGGTAATC GTTGGTAGTAGACACAAGAGGG CAGCCTGGTCTATAGAGTGAGT GCTGAGTATGTCGTGGAGTCTA CATACTTGGCAGGTTTCTCCAG
101 85 Figure 1. The density of bladder pelvic and hypogastric innervations in cultured lumbosacral (LS) and thoracolumbar (TL) mouse DRG neurons Four lumbosacral DRG and four thoracolumbar DRG coverslips were randomly chosen from four bladder retrogradely labeled mice. An example of a selected view field (10X objective) of DRG neuron coverslip was shown under differential-interference contrast (DIC) mode (A) and fluorescence mode (B). 6.0±0.4% (77/1276) of L6-S2 DRG and 2.4±0.2% (40/1635) of DiI-labeled T13-L2 DRG cells were recognized as DiI-positive neurons. The proportion of LS bladder neuron was significantly greater the proportion of TL bladder neuron (C).

102 86 A 40 m B 40 m C % of bladder neuron LS ** TL
103 87 Figure 2. Comparison of cell size (capacitance) of LS and TL mouse bladder sensory neurons (A) Scatter plot of whole-cell capacitance (an index of cell size) showing similar pattern of cell size distribution in LS and TL DRGs. (B) Frequency distribution of whole-cell capacitance in LS and TL bladder sensory neurons. Cells with capacitance <20pF were considered as small neurons, and ones with capacitance >45pF were considered as larger neurons; medium size neurons had capacitances between 20 to 45pF. No significant difference of capacitance distribution in LS and TL bladder neurons was detected.
104 88 A capacitance (pf) LS (N=114) TL (N=91) B frequency (%) 100 LS TL small medium large
105 89 Figure 3. Examples of principal purinergic currents in LS and TL bladder sensory neurons in response to ATP (30μM) and αβ-met ATP (30μM) Based on current inactivation kinetics and agonist responses, three distinct current types were identified: slow desensitizing currents predominated in LS (~90%, A) and TL (~75%, C) neurons; (B) sustained currents without an obvious inactivation phase were found only in LS neurons (~10%); and (D) fast desensitizing currents were found only in TL neurons (~25%). The duration of agonist application (4 sec) is denoted by the horizontal bar.
106 90 LS TL A ATP (30 M) -Met ATP (30 M) C ATP (30 M) -Met ATP (30 M) B D ATP (30 M) -Met ATP (30 M) ATP (30 M) -Met ATP (30 M)
107 91 Figure 4. Antagonism of purinergic agonist-evoked sustained currents (A) Sustained currents evoked by ATP (30μM) were not inhibited by TNP- ATP (0.1μM). (B) PPADS (10μM) completely antagonized ATP-evoked sustained currents. (C) Summary of antagonism of sustained currents. Amplitude of sustained current was not decreased by TNP-ATP application (n=5), but was significantly attenuated by PPADS by a mean of 82.0±5.6% (n=5). The duration of agonist application (4 sec) is denoted by the horizontal bar.
108 92 ATP (30 M) ATP (30 M) + TNP-ATP (0.1 M) washout B ATP (30 M) ATP (30 M) + PPADS (10 M) washout C Inhibition of current amplitude TNP-ATP PPADS
109 93 Figure 5. Antagonism of purinergic agonist-evoked slow desensitizing currents Slow desensitizing currents evoked by ATP were greatly attenuated by the P2X3 receptor-selective antagonist TNP-ATP (A) and by the non-selective purinergic antagonist PPADS (B). (C) Summary of antagonism of slow currents. TNP-ATP significantly attenuated the slow currents evoked by ATP (by a mean 55.5±2.1%, n=5) or α,β-met ATP (by a mean 55.6±3.0%, n=4). PPADS also significantly attenuated the slow currents evoked by ATP (by a mean 77.7±3.0%, n=5) or α,β-met ATP (by a mean 82.0±2.1%, n=4). The duration of agonist application (4 sec) is denoted by the horizontal bar.
110 94 ATP (30 M) ATP (30 M) + TNP-ATP (0.1 M) washout B ATP (30 M) ATP (30 M) + PPADS (10 M) washout C Inhibition of current amplitude ATP-evoked TNP-ATP PPADS -Met ATP-evoked
111 95 Figure 6. Antagonism of purinergic agonist-evoked fast desensitizing currents Fast desensitizing currents were completely inhibited by the P2X3 receptor-selective antagonist TNP-ATP (A) and by the non-selective purinergic antagonist PPADS (B). (C) Summary of antagonism of fast currents. TNP-ATP significantly attenuated the fast currents evoked by ATP (by a mean 55.5±2.1%, n=4) and PPADS also attenuated the fast currents evoked by ATP (by a mean 77.7±3.0%, n=4). The duration of agonist application (4 sec) is denoted by the horizontal bar.
112 96 ATP (30 M) ATP (30 M) + TNP-ATP (0.1 M) washout B ATP (30 M) ATP (30 M) + PPADS (10 M) washout C Inhibition of current amplitude TNP-ATP PPADS
113 97 Figure 7. Examples of principal purinergic current in P2X3-/- mice Only a sustained current was evoked by ATP in LS bladder neurons (A) which was inhibited by PPADS (C) but not TNP-ATP (D). No obvious response to ATP or α, β-met ATP was observed in TL bladder neurons (B). The duration of agonist application (4 sec) is denoted by the horizontal bar.
114 98 A LS B TL ATP (30 M) -Met ATP (30 M) ATP (30 M) -Met ATP (30 M) 83.33% (10/12) no response (0/8) C ATP (30 M) ATP (30 M) + PPADS (10 M) washout D ATP (30 M) ATP (30 M) + TNP-ATP (0.1 M) washout
115 99 Figure 8. Examples of bladder sensory neuron responses to current injection and agonist application. (A) A series of current pluses (20 pa increment, 1s apart) was injected, and an action potential in a bladder sensory neuron was evoked. The rheobase was determined as the maximum current (pa) that did not evoke an action potential. Action potential (AP) threshold was determined from the inflection point where membrane potential started to dramatically rise and the phase plot slope (the first derivative of membrane potential, dv/dt) reached 10 mv/ms. (B) Examples of agonist (ATP) application induced- membrane depolarization (left) and action potential (right).
116 100 A AP overshoot AP threshold 50mV 10ms RMP 140pA B ATP (30 M) ATP (30 M) 20mV 2s
117 101 Figure 9. Single cell nested RT-PCR of P2X2 and P2X3 receptor subunits in LS and TL bladder neurons (A) An example of positive nested single cell PCR amplification of P2X2 and P2X3 mrna. Product length corresponded with expected size. The frequency of P2X2 and P2X3 transcription in bladder neurons is summarized in (B). Percentage of cells only expressing P2X2, P2X3, or both P2X2 and P2X3 is illustrated in (C). * TL vs. LS, P<0.05; ** TL vs. LS, P<0.01.
118 102 A P2X2 (113bp) P2X3 (141bp) 200bp 100bp B Frequency (%) ** LS TL 0 P2X2 P2X3 C 100 * LS TL Frequency (%) * ** 0 P2X2 ONLY P2X2/3 P2X3 ONLY
119 103 Figure 10. Immunohistochemistry of P2X3 subunit in bladder sensory neurons from naïve mice Examples of immunohistochemical staining of P2X3 receptor subunit in bladder sensory neurons identified by the presence of Cholera Toxin B (CTB) are shown. (A) Representative images of P2X3 immunostaining in mouse L6 DRG. (B) Representative images of P2X3 immunostaining in mouse L1 DRG. Arrows indicated bladder neurons with positive P2X3 immunoreactivity. The scale bar represents 50μm.




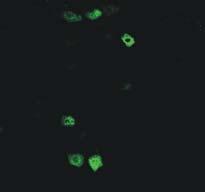

120 104 A CTB P2X3 Merge LS B CTB P2X3 Merge TL
121 105 Figure 11. Bladder weight and bladder myeloperoxidase (MPO) activity in naïve saline- and CYP-treated mice (A) Mean bladder weight after CYP treatment was significantly greater (38.6 ± 1.3 mg, n=8) than that of bladders taken from saline-treated (23.7±0.7mg, n=6; P<0.005) or naïve mice (22.9±1.1mg, n=6; P<0.005). (B) No significant change of bladder MPO activity after CYP treatment (0.0002± Unit/gram, n=8) was detected compared with saline-treated (0.002±0.002 Unit/gram, n=4) or naïve mice (0.0009± Unit/gram, n=4).
122 106 A Bladder weight (mg) *** *** 0 Naive Saline CYP B MPO activity (Unit/gram) Naive saline CYP
123 107 Figure 12. Histological assessment of bladder inflammation in saline- and CYP- treated mice Photomicrographs of representative histological specimen of bladder tissues taken from saline- (A) and CYP- (B) treated mice. Areas identified by boxes are enlarged sequentially from top to bottom. Mild submucosal edema and unfolding of the urothelium were detected in bladders of CYP- but not salinetreated mice. Tissue sections were stained with H&E. Scale bar indicates 100μm.




124 108
125 109 Figure 13. Examples of responses of bladder sensory neurons to current injection and agonist application in saline- and CYP- treated mice The injected current required to evoke an action potential in LS and TL bladder neurons was significantly reduced after CYP treatment (B,D) relative to saline treatment (A,C). The magnitude of membrane depolarization in LS and TL bladder neurons was increased after CYP-treatment compared with salinetreated controls in response to αβ-met ATP (F, H) but not to ATP (E,G).
126 110 A saline B CYP LS 160pA 120pA C D 50mV 10ms TL 180pA 120pA E ATP -Met ATP (30 M) saline CYP saline CYP F LS G H 10mV 2s TL
127 111 Figure 14. Single cell nested RT-PCR of P2X2 and P2X3 receptor subunits in LS and TL bladder neurons from saline- and CYP-treated mice The frequency of P2X2 and P2X3 transcription in bladder neurons is summarized in (A). Percentage of cells only expressing P2X2, P2X3, or both P2X2 and P2X3 is illustrated in (B). (* TL vs. LS, P<0.05; ** P<0.01; saline vs. CYP, P<0.05).
128 112 A 100 ** LS-SAL LS-CYP TL-SAL TL-CYP frequency (%) P2X2 P2X3 B frequency (%) * * LS-SAL LS-CYP TL-SAL TL-CYP 0 P2X2 ONLY P2X2/3 P2X3 ONLY
129 113 Figure 15. Immunohistochemistry of P2X3 subunit in bladder sensory neurons from saline-treated and CYP-treated mice Examples of immunohistochemical staining of P2X3 subunit in bladder sensory neurons identified by Cholera Toxin B (CTB) labeling were presented here. Representative images of P2X3 immunostaining in mouse LS and TL DRGs from saline-treated mice were shown in (A: S1 DRG) and (B: L1 DRG). Representative images of P2X3 immunostaining in mouse LS and TL DRGs from CYP-treated mice were shown in (C: S1 DRG) and (D: L1 DRG). The scale bar represents 50μm.



130 114
131 115 Figure 16. UTP increases the excitability of bladder sensory neurons Examples of responses of lumbosacral bladder neurons to current injection before and after UTP application are shown here. (A, B) The injected current required to evoke an action potential in bladder neurons was significantly reduced after UTP application from142.4±11.6 pa to 82.2±5.2 pa (n=24; P<0.01). (C, D) Current injection at 2X rheobase (500ms) evoked a significantly greater number of APs after UTP application (8.7±1.0) relative to before UTP application (3.3±0.5; n=24, P<0.01). (E) An example of sustained AP firing during and after UTP application.
132 116 A control B UTP 120pA 20mV 10ms 80pA C D 20mV 100ms 240pA 160pA E UTP 1 M 20mV 5S
133 117 Figure 17. Examples of the effect of UTP on LS bladder neuron responses to ATP A fraction (8/21) of bladder neurons was sensitized by UTP, showing evoked AP firing (A) or increased frequency of APs (B) relative to the response to ATP before UTP application to the same cell. Other neurons (13/21) did not exhibit changes in response to ATP after UTP application (C, D).
134 118 A ATP (30 M) B UTP (1 M, 40s) UTP (1 M, 40s) ATP (30 M) 20mV 4s C UTP (1 M, 40s) D UTP (1 M, 40s) ATP (30 M) ATP (30 M)
135 119 Figure 18. Recovery kinetics of ATP-evoked sustained, slow and fast currents in LS bladder neurons Examples of sustained (A, B), slow (C, D) and fast (E, F) currents at different application intervals of ATP (30μM) are shown here. Recovered current fractions were fitted with a single exponential (E: sustained and slow current; F: fast current). The time constant of the sustained current is not available because it completely recovered very rapidly. The slow current recovered with a time constant of 16.8s, and the fast current recovered with a time constant of 70.3s.
136 120 A 2s B 5s 0.2nA 5s C 5s D 60s 0.2nA 5s E 40s F 120s 0.5nA 5s
137 121 Figure 18- continued G H normalized ATP-evoked current normalized ATP-evoked current Fast Sustained Slow time (s) time (s)
138 122 Figure 19. The effect of UTP on ATP-evoked sustained, slow and fast currents in LS and TL bladder neurons ATP (30μM) was applied to LS or TL bladder neurons for 4 sec (the left traces), followed by a 2-minute washout period using external solution, and then applied external solution containing UTP (1μM) for 40 sec before repeating the ATP application (the middle traces). After another 2-minute washout period, currents induced by a third exposure to ATP were recorded (the right traces). UTP significantly facilitated ATP-evoked sustained (A) and fast (C) currents but not slow currents (B). (D) Summary of the effect of UTP on ATP-evoked sustained (n=16), slow (n=24) and fast (n=5) currents.
139 A ATP (30 M) UTP (0.3 M, 40s) nA B ATP (30 M) UTP (0.3 M, 40s) 0.2nA C ATP (30 M) UTP (0.3 M, 40s) 0.2nA 2s D normalized ATP-evoked current ATP before UTP ATP with UTP ** sustained slow fast *
140 124 Figure 20. The effect of repeated ATP application without UTP on ATP-evoked sustained, slow and fast currents in LS and TL bladder neurons ATP (30μM) was applied to LS or TL bladder neurons for 4 sec (the left traces), followed by a 2-minute washout period using external solution, and then applied nothing (a control of UTP application) for 40 sec before repeating the ATP application (the middle traces). After another 2-minute washout period, currents induced by a third exposure to ATP were recorded (the right traces). No change of sustained (A) or slow (B) currents after repeated ATP application in LS bladder sensory neurons was observed. ATP-evoked fast (C) current in TL bladder neurons show significant differences in response to repetitive ATP application. (D) Summary of repetitive ATP application on ATP-evoked sustained (n=11), slow (n=20) and fast (n=5) currents.
141 125 A ATP (30 M) 0.5nA B ATP (30 M) 0.2nA C ATP (30 M) 0.5nA 2s D normalized ATP-evoked current st ATP 2nd ATP sustained slow fast *
142 126 Figure 21. The dose-response relationship of ATP evoked-sustained currents in response to UTP in LS bladder neurons (A) An example of the influences of different concentration of UTP on ATP evoked-sustained current in lumbosacral bladder neurons. (B) Normalized doseresponse curve of ATP evoked-sustained currents in response to UTP. The EC 50 and 95% confidence interval for UTP was 0.52μM ( μM; n=7).
143 127 A ATP (30 M) 0.2nA 2s Before UTP UTP 0.03 M UTP 0.1 M UTP 0.3 M UTP 1.0 M UTP 3.0 M UTP 10.0 M B normalized ATP-evoked current log [UTP]M
144 128 Figure 22. The facilitatory effect of UTP on ATP-evoked sustained currents but not fast currents is mediated by G protein-coupled P2Y2 receptor through a PKC dependent pathway. Although the facilitatory effect of UTP on sustained currents was blocked by intracellular GDP-β-s (A), the increased fast currents induced by UTP/repeated ATP application were not inhibited by GDP-β-s (A and B). No significant change of either sustained or slow currents in response to repeated ATP application was found in bladder neurons containing GDP-β-s compared with controls (B). The PKC inhibitor (myristoylated Protein Kinase C Inhibitor 20-28, 10μM) prevented the effect of UTP on P2X2 sustained current in lumbosacral bladder neurons (C). No significant UTP effect on slow current was observed in presence or absence of the PKC blocker in lumbosacral bladder neurons (C). Neurons from P2Y2 knockout mice did not show a facilitatory effect of UTP on sustained currents in lumbosacral bladder neurons compared with wild type bladder neurons (D).
145 ATP with UTP ATP with UTP+GDP- -S * sustained slow fast nd ATP 2nd ATP + GDP- -S sustained slow fast normalized ATP-evoked current normalized ATP-evoked current A C * ATP with UTP ATP with UTP+PKC inhibitor sustained slow B D * ATP with UTP in P2Y2+/+ ATP with UTP in P2Y2-/- sustained slow normalized ATP-evoked current normalized ATP-evoked current
146 130 Figure 23. The effect of UTP/ATP on bladder sensory neurons containing intracellular GDP-β-S, in presence of PKC inhibitor and from P2Y2 knockout mice. The UTP/ATP effect was examined by comparing current amplitude evoked by ATP before and with UTP/repeated ATP application in the same bladder neuron. UTP (A) and repeated ATP (B) application did not elicit significant changes of sustained (n=7 for both UTP and ATP) or slow (UTP: n=7; ATP n=9) currents in lumbosacral bladder neurons containing GDP-β-s (100μM), but did induce significant enhancement of fast current (n=5 for both UTP and ATP). No significant UTP effect on sustained (n=4) or slow (n=5) current was observed in lumbosacral bladder neurons with PKC inhibitor (C). Both ATPevoked sustained (n=6) and slow (n=10) currents were inhibited by UTP in lumbosacral bladder neurons from P2Y2 knockout mice (D).
147 ATP + GDP- -S ATP with UTP+GDP- -S * sustained slow fast st ATP + GDP- -S 2nd ATP + GDP- -S * sustained slow fast normalized ATP-evoked current normalized ATP-evoked current A C ATP before UTP ATP with UTP+PKC inhibitor sustained slow B D ATP before UTP in P2Y2-/- ATP with UTP in P2Y2-/- * * sustained slow normalized ATP-evoked current normalized ATP-evoked current
148 132 Figure 24. Single cell nested RT-PCR of P2X2, P2X3, P2Y2 and P2Y4 receptor subunits in LS and TL bladder neurons (A) Examples of positive nested single cell PCR amplification of P2X and P2Y mrnas. Product length corresponded with expected size. The frequency of P2X and P2Y transcription in bladder neurons is summarized in (B). The P2X2 subunit transcript was significantly more abundant in LS than TL bladder neurons (P<0.01). The percentage of cells expressing P2Y2 and P2Y4 transcripts in P2X2- (C) or P2X3- (D) positive neurons is illustrated in pie charts.


 TL")
 TL")
149 133 A P2X2 P2X3 P2Y2 P2Y4 300bp 200bp 100bp B frequency (%) 100 ** LS TL C P2Y4 0 P2X2 P2X3 P2Y2 P2Y4 P2X2(+) LS cells P2Y2 P2Y4 P2X2(+) TL cells P2Y2 D P2X3(+) LS cells P2Y4 P2Y2 P2X3(+) TL cells P2Y4 P2Y2
Victor Chaban, Ph.D., MSCR
 Hormonal Modulation of Pain : Estrogen Modulation of Visceral Nociception Victor Chaban, Ph.D., MSCR Department of Internal Medicine Charles R. Drew University of Medicine and Science Department of Medicine
Hormonal Modulation of Pain : Estrogen Modulation of Visceral Nociception Victor Chaban, Ph.D., MSCR Department of Internal Medicine Charles R. Drew University of Medicine and Science Department of Medicine
Neurotransmitter Systems II Receptors. Reading: BCP Chapter 6
 Neurotransmitter Systems II Receptors Reading: BCP Chapter 6 Neurotransmitter Systems Normal function of the human brain requires an orderly set of chemical reactions. Some of the most important chemical
Neurotransmitter Systems II Receptors Reading: BCP Chapter 6 Neurotransmitter Systems Normal function of the human brain requires an orderly set of chemical reactions. Some of the most important chemical
What is on the Horizon in Drug Therapy for OAB?
 What is on the Horizon in Drug Therapy for OAB? K-E Andersson, MD, PhD Wake Forest Institute for Regenerative Medicine Wake Forest University School of Medicine Winston Salem, North Carolina Disclosures
What is on the Horizon in Drug Therapy for OAB? K-E Andersson, MD, PhD Wake Forest Institute for Regenerative Medicine Wake Forest University School of Medicine Winston Salem, North Carolina Disclosures
Sensory coding and somatosensory system
 Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
Definitions of IC: U.S. perspective. Edward Stanford MD MS FACOG FACS Western Colorado
 Definitions of IC: U.S. perspective Edward Stanford MD MS FACOG FACS Western Colorado PURPOSE OF A DEFINITION? Identifies with specificity those patients who are most likely to have the disease. Identifies
Definitions of IC: U.S. perspective Edward Stanford MD MS FACOG FACS Western Colorado PURPOSE OF A DEFINITION? Identifies with specificity those patients who are most likely to have the disease. Identifies
Autonomic Nervous System. Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry
 Autonomic Nervous System Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry Peripheral Nervous System A. Sensory Somatic Nervous System B. Autonomic Nervous System 1. Sympathetic Nervous
Autonomic Nervous System Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry Peripheral Nervous System A. Sensory Somatic Nervous System B. Autonomic Nervous System 1. Sympathetic Nervous
Neurological control of the bladder in health and disease
 Section 1 Chapter 1 Neurological control of pelvic organ functions Neurological control of the bladder in health and disease Derek J. Griffiths and Apostolos Apostolidis Peripheral control of micturition
Section 1 Chapter 1 Neurological control of pelvic organ functions Neurological control of the bladder in health and disease Derek J. Griffiths and Apostolos Apostolidis Peripheral control of micturition
Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor
 Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor NOCICEPTORS ARE NOT IDENTICAL PEPTIDE SubP/CGRP Trk A NON-PEPTIDE IB4 P2X 3 c-ret Snider and McMahon
Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor NOCICEPTORS ARE NOT IDENTICAL PEPTIDE SubP/CGRP Trk A NON-PEPTIDE IB4 P2X 3 c-ret Snider and McMahon
W11: Functional Pain Syndromes: Bidirectional Influences in Visceral Nociceptive Processing Aims of course/workshop
 W11: Functional Pain Syndromes: Bidirectional Influences in Visceral Nociceptive Processing Workshop Chair: Matthew Fraser, United States 27 August 2013 09:00-10:30 Start End Topic Speakers 09:00 09:25
W11: Functional Pain Syndromes: Bidirectional Influences in Visceral Nociceptive Processing Workshop Chair: Matthew Fraser, United States 27 August 2013 09:00-10:30 Start End Topic Speakers 09:00 09:25
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline
 Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Physiological processes in the GI tract:
 Gastrointestinal physiology for medical students General principal of gastrointestinal function Motility, nervous control and blood circulation Physiological processes in the GI tract: Motility Secretion
Gastrointestinal physiology for medical students General principal of gastrointestinal function Motility, nervous control and blood circulation Physiological processes in the GI tract: Motility Secretion
211MDS Pain theories
 211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
The Pain Pathway. dorsal root ganglion. primary afferent nociceptor. TRP: Transient Receptor Potential
 Presented by Issel Anne L. Lim 1 st Year PhD Candidate Biomedical Engineering Johns Hopkins University 580.427/580.633 Ca Signals in Biological Systems Outline The Pain Pathway TRP: Transient Receptor
Presented by Issel Anne L. Lim 1 st Year PhD Candidate Biomedical Engineering Johns Hopkins University 580.427/580.633 Ca Signals in Biological Systems Outline The Pain Pathway TRP: Transient Receptor
Pharmacology of Pain Transmission and Modulation
 Pharmacology of Pain Transmission and Modulation 2 Jürg Schliessbach and Konrad Maurer Nociceptive Nerve Fibers Pain is transmitted to the central nervous system via thinly myelinated Aδ and unmyelinated
Pharmacology of Pain Transmission and Modulation 2 Jürg Schliessbach and Konrad Maurer Nociceptive Nerve Fibers Pain is transmitted to the central nervous system via thinly myelinated Aδ and unmyelinated
Detrusor overactivity induced by intravesical application of adenosine 5 -triphosphate under different delivery conditions in rats
 Medicine Urology fields Okayama University Year 2007 Detrusor overactivity induced by intravesical application of adenosine 5 -triphosphate under different delivery conditions in rats Jun Nishiguchi M.D.,
Medicine Urology fields Okayama University Year 2007 Detrusor overactivity induced by intravesical application of adenosine 5 -triphosphate under different delivery conditions in rats Jun Nishiguchi M.D.,
Neural Control of Lower Urinary Tract Function. William C. de Groat University of Pittsburgh Medical School
 Neural Control of Lower Urinary Tract Function William C. de Groat University of Pittsburgh Medical School Disclosures Current funding: NIH Grants, DK093424, DK-091253, DK-094905, DK-090006. Other financial
Neural Control of Lower Urinary Tract Function William C. de Groat University of Pittsburgh Medical School Disclosures Current funding: NIH Grants, DK093424, DK-091253, DK-094905, DK-090006. Other financial
Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels. Vahri Beaumont and Robert S. Zucker
 Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels Vahri Beaumont and Robert S. Zucker Background I h channels discovered in 1976 (Noma A. and Irisawa H.) Voltage-gated
Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels Vahri Beaumont and Robert S. Zucker Background I h channels discovered in 1976 (Noma A. and Irisawa H.) Voltage-gated
EE 791 Lecture 2 Jan 19, 2015
 EE 791 Lecture 2 Jan 19, 2015 Action Potential Conduction And Neural Organization EE 791-Lecture 2 1 Core-conductor model: In the core-conductor model we approximate an axon or a segment of a dendrite
EE 791 Lecture 2 Jan 19, 2015 Action Potential Conduction And Neural Organization EE 791-Lecture 2 1 Core-conductor model: In the core-conductor model we approximate an axon or a segment of a dendrite
CHAPTER 10 THE SOMATOSENSORY SYSTEM
 CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could
CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could
Committee 3. Neural Control. Chair L. BIRDER (USA) Co-Chairman M. DRAKE (UK)
 Co-Chairman M. DRAKE (UK)") Committee 3 Neural Control Chair L. BIRDER (USA) Co-Chairman M. DRAKE (UK) Members W. DE GROAT (USA), C. FOWLER (U.K), E. MAYER (USA), J. MORRISON (UAE), J. PATON (U.K) Consultants D. GRIFFITHS (Canada),
Committee 3 Neural Control Chair L. BIRDER (USA) Co-Chairman M. DRAKE (UK) Members W. DE GROAT (USA), C. FOWLER (U.K), E. MAYER (USA), J. MORRISON (UAE), J. PATON (U.K) Consultants D. GRIFFITHS (Canada),
Outline. Neuron Structure. Week 4 - Nervous System. The Nervous System: Neurons and Synapses
 Outline Week 4 - The Nervous System: Neurons and Synapses Neurons Neuron structures Types of neurons Electrical activity of neurons Depolarization, repolarization, hyperpolarization Synapses Release of
Outline Week 4 - The Nervous System: Neurons and Synapses Neurons Neuron structures Types of neurons Electrical activity of neurons Depolarization, repolarization, hyperpolarization Synapses Release of
Mechanism of Pain Production
 Mechanism of Pain Production Pain conducting nerve fibers are small myelinated (A-delta) or unmyelinated nerve fibers (C-fibers). Cell bodies are in the dorsal root ganglia (DRG) or sensory ganglia of
Mechanism of Pain Production Pain conducting nerve fibers are small myelinated (A-delta) or unmyelinated nerve fibers (C-fibers). Cell bodies are in the dorsal root ganglia (DRG) or sensory ganglia of
MOLECULAR AND CELLULAR NEUROSCIENCE
 MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
CHAPTER 15 LECTURE OUTLINE
 CHAPTER 15 LECTURE OUTLINE I. INTRODUCTION A. The autonomic nervous system (ANS) regulates the activity of smooth muscle, cardiac muscle, and certain glands. B. Operation of the ANS to maintain homeostasis,
CHAPTER 15 LECTURE OUTLINE I. INTRODUCTION A. The autonomic nervous system (ANS) regulates the activity of smooth muscle, cardiac muscle, and certain glands. B. Operation of the ANS to maintain homeostasis,
ANATOMY & PHYSIOLOGY - CLUTCH CH. 6 - CELL COMMUNICATION.
 !! www.clutchprep.com CONCEPT: CELL-TO-CELL CONNECTIONS AND SIGNALING Gap and Tight Junctions: Adjacent cells communicate and hold on to each other via junctions. Two important kinds: Gap Junctions are
!! www.clutchprep.com CONCEPT: CELL-TO-CELL CONNECTIONS AND SIGNALING Gap and Tight Junctions: Adjacent cells communicate and hold on to each other via junctions. Two important kinds: Gap Junctions are
VEGF induces sensory and motor peripheral plasticity, alters bladder function, and promotes visceral sensitivity
 Malykhina et al. BMC Physiology 2012, 12:15 RESEARCH ARTICLE Open Access VEGF induces sensory and motor peripheral plasticity, alters bladder function, and promotes visceral sensitivity Anna P Malykhina
Malykhina et al. BMC Physiology 2012, 12:15 RESEARCH ARTICLE Open Access VEGF induces sensory and motor peripheral plasticity, alters bladder function, and promotes visceral sensitivity Anna P Malykhina
Introduction to Neurobiology
 Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
Supporting Information
 ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
Signal transduction underlying the control of urinary bladder smooth muscle tone Puspitoayu, E.
 UvA-DARE (Digital Academic Repository) Signal transduction underlying the control of urinary bladder smooth muscle tone Puspitoayu, E. Link to publication Citation for published version (APA): Puspitoayu,
UvA-DARE (Digital Academic Repository) Signal transduction underlying the control of urinary bladder smooth muscle tone Puspitoayu, E. Link to publication Citation for published version (APA): Puspitoayu,
NEURONS Chapter Neurons: specialized cells of the nervous system 2. Nerves: bundles of neuron axons 3. Nervous systems
 NEURONS Chapter 12 Figure 12.1 Neuronal and hormonal signaling both convey information over long distances 1. Nervous system A. nervous tissue B. conducts electrical impulses C. rapid communication 2.
NEURONS Chapter 12 Figure 12.1 Neuronal and hormonal signaling both convey information over long distances 1. Nervous system A. nervous tissue B. conducts electrical impulses C. rapid communication 2.
number Done by Corrected by Doctor
 number 13 Done by Tamara Wahbeh Corrected by Doctor Omar Shaheen In this sheet the following concepts will be covered: 1. Divisions of the nervous system 2. Anatomy of the ANS. 3. ANS innervations. 4.
number 13 Done by Tamara Wahbeh Corrected by Doctor Omar Shaheen In this sheet the following concepts will be covered: 1. Divisions of the nervous system 2. Anatomy of the ANS. 3. ANS innervations. 4.
Synaptic Transmission: Ionic and Metabotropic
 Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Receptors and Drug Action. Dr. Subasini Pharmacology Department Ishik University, Erbil
 Receptors and Drug Action Dr. Subasini Pharmacology Department Ishik University, Erbil Receptors and Drug Action Receptor Receptor is defined as a macromolecule or binding site located on the surface or
Receptors and Drug Action Dr. Subasini Pharmacology Department Ishik University, Erbil Receptors and Drug Action Receptor Receptor is defined as a macromolecule or binding site located on the surface or
Lecture 9: Cell Communication I
 02.05.10 Lecture 9: Cell Communication I Multicellular organisms need to coordinate cellular functions in different tissues Cell-to-cell communication is also used by single celled organisms to signal
02.05.10 Lecture 9: Cell Communication I Multicellular organisms need to coordinate cellular functions in different tissues Cell-to-cell communication is also used by single celled organisms to signal
Chapter 11: Nervous System and Nervous Tissue
 Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007)
") Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007) Introduction Adrian s work on sensory coding Spinal cord and dorsal root ganglia Four somatic sense modalities Touch Mechanoreceptors
Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007) Introduction Adrian s work on sensory coding Spinal cord and dorsal root ganglia Four somatic sense modalities Touch Mechanoreceptors
themes G. F. GEBHART Department of Pharmacology, The University of Iowa, College of Medicine, Iowa City, Iowa 52242
 Am J Physiol Gastrointest Liver Physiol 278: G834 G838, 2000. themes Pathobiology of Visceral Pain: Molecular Mechanisms and Therapeutic Implications IV. Visceral afferent contributions to the pathobiology
Am J Physiol Gastrointest Liver Physiol 278: G834 G838, 2000. themes Pathobiology of Visceral Pain: Molecular Mechanisms and Therapeutic Implications IV. Visceral afferent contributions to the pathobiology
Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients
 Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients Lu-Yuan Lee, Ph.D. Airway Sensory Neurobiology Laboratory Department of Physiology University of Kentucky Medical Center BACKGROUND
Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients Lu-Yuan Lee, Ph.D. Airway Sensory Neurobiology Laboratory Department of Physiology University of Kentucky Medical Center BACKGROUND
Module H NERVOUS SYSTEM
 Module H NERVOUS SYSTEM Topic from General functions of the nervous system Organization of the nervous system from both anatomical & functional perspectives Gross & microscopic anatomy of nervous tissue
Module H NERVOUS SYSTEM Topic from General functions of the nervous system Organization of the nervous system from both anatomical & functional perspectives Gross & microscopic anatomy of nervous tissue
Portions from Chapter 6 CHAPTER 7. The Nervous System: Neurons and Synapses. Chapter 7 Outline. and Supporting Cells
 CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
Part 11: Mechanisms of Learning
 Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Lecture 14. Insect nerve system (II)
") Lecture 14. Insect nerve system (II) Structures (Anatomy) Cells Anatomy How NS functions Signal transduction Signal transmission Overview More on neurons: ions, ion channel, ligand receptor Signal transduction:
Lecture 14. Insect nerve system (II) Structures (Anatomy) Cells Anatomy How NS functions Signal transduction Signal transmission Overview More on neurons: ions, ion channel, ligand receptor Signal transduction:
NIH Public Access Author Manuscript Neurourol Urodyn. Author manuscript; available in PMC 2010 June 23.
 NIH Public Access Author Manuscript Published in final edited form as: Neurourol Urodyn. 2010 ; 29(1): 63 76. doi:10.1002/nau.20761. Changes in Afferent Activity After Spinal Cord Injury William C. de
NIH Public Access Author Manuscript Published in final edited form as: Neurourol Urodyn. 2010 ; 29(1): 63 76. doi:10.1002/nau.20761. Changes in Afferent Activity After Spinal Cord Injury William C. de
Neuroscience: Exploring the Brain, 3e. Chapter 4: The action potential
 Neuroscience: Exploring the Brain, 3e Chapter 4: The action potential Introduction Action Potential in the Nervous System Conveys information over long distances Action potential Initiated in the axon
Neuroscience: Exploring the Brain, 3e Chapter 4: The action potential Introduction Action Potential in the Nervous System Conveys information over long distances Action potential Initiated in the axon
Chapter 4 Neuronal Physiology
 Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
I. Neural Control of Involuntary Effectors. Chapter 9. Autonomic Motor Nerves. Autonomic Neurons. Autonomic Ganglia. Autonomic Neurons 9/19/11
 Chapter 9 I. Neural Control of Involuntary Effectors The Autonomic Nervous System Lecture PowerPoint Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Autonomic
Chapter 9 I. Neural Control of Involuntary Effectors The Autonomic Nervous System Lecture PowerPoint Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Autonomic
Pharmacology of P2X channels
 Pflugers Arch - Eur J Physiol (2006) 452: 513 537 DOI 10.1007/s00424-006-0070-9 INVITED REVIEW Joel R. Gever. Debra A. Cockayne. Michael P. Dillon. Geoffrey Burnstock. Anthony P. D. W. Ford Pharmacology
Pflugers Arch - Eur J Physiol (2006) 452: 513 537 DOI 10.1007/s00424-006-0070-9 INVITED REVIEW Joel R. Gever. Debra A. Cockayne. Michael P. Dillon. Geoffrey Burnstock. Anthony P. D. W. Ford Pharmacology
Lecture 22: A little Neurobiology
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
Receptors Families. Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia
 Receptors Families Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia Receptor Families 1. Ligand-gated ion channels 2. G protein coupled receptors 3. Enzyme-linked
Receptors Families Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia Receptor Families 1. Ligand-gated ion channels 2. G protein coupled receptors 3. Enzyme-linked
Basic Neuroscience. Sally Curtis
 The Physiology of Pain Basic Neuroscience Sally Curtis sac3@soton.ac.uk The behaviour of humans is a result of the actions of nerves. Nerves form the basis of Thoughts, sensations and actions both reflex
The Physiology of Pain Basic Neuroscience Sally Curtis sac3@soton.ac.uk The behaviour of humans is a result of the actions of nerves. Nerves form the basis of Thoughts, sensations and actions both reflex
Chapter 14 The Autonomic Nervous System Chapter Outline
 Chapter 14 The Autonomic Nervous System Chapter Outline Module 14.1 Overview of the Autonomic Nervous System (Figures 14.1 14.3) A. The autonomic nervous system (ANS) is the involuntary arm of the peripheral
Chapter 14 The Autonomic Nervous System Chapter Outline Module 14.1 Overview of the Autonomic Nervous System (Figures 14.1 14.3) A. The autonomic nervous system (ANS) is the involuntary arm of the peripheral
Cellular Messengers. Intracellular Communication
 Cellular Messengers Intracellular Communication Most common cellular communication is done through extracellular chemical messengers: Ligands Specific in function 1. Paracrines Local messengers (neighboring
Cellular Messengers Intracellular Communication Most common cellular communication is done through extracellular chemical messengers: Ligands Specific in function 1. Paracrines Local messengers (neighboring
Chapter 7 Nerve Cells and Electrical Signaling
 Chapter 7 Nerve Cells and Electrical Signaling 7.1. Overview of the Nervous System (Figure 7.1) 7.2. Cells of the Nervous System o Neurons are excitable cells which can generate action potentials o 90%
Chapter 7 Nerve Cells and Electrical Signaling 7.1. Overview of the Nervous System (Figure 7.1) 7.2. Cells of the Nervous System o Neurons are excitable cells which can generate action potentials o 90%
Special Issue on Pain and Itch
 Special Issue on Pain and Itch Title: Recent Progress in Understanding the Mechanisms of Pain and Itch Guest Editor of the Special Issue: Ru-Rong Ji, PhD Chronic pain is a major health problem world-wide.
Special Issue on Pain and Itch Title: Recent Progress in Understanding the Mechanisms of Pain and Itch Guest Editor of the Special Issue: Ru-Rong Ji, PhD Chronic pain is a major health problem world-wide.
Drugs Affecting The Autonomic Nervous System(ANS)
") Drugs Affecting The Autonomic Nervous System(ANS) ANS Pharmacology Lecture 1 Dr. Hiwa K. Saaed College of Pharmacy, University of Sulaimani 2018-2019 AUTOMATIC NERVOUS SYSTEM (ANS) The ANS is the major
Drugs Affecting The Autonomic Nervous System(ANS) ANS Pharmacology Lecture 1 Dr. Hiwa K. Saaed College of Pharmacy, University of Sulaimani 2018-2019 AUTOMATIC NERVOUS SYSTEM (ANS) The ANS is the major
2.4 Autonomic Nervous System
 2.4 Autonomic Nervous System The ANS regulates visceral activities normally outside the realm of consciousness and voluntary control: Circulation. Digestion. Sweating. Pupillary size. The ANS consists
2.4 Autonomic Nervous System The ANS regulates visceral activities normally outside the realm of consciousness and voluntary control: Circulation. Digestion. Sweating. Pupillary size. The ANS consists
Lojayn Salah. Razan Aburumman. Faisal Muhammad
 20 Lojayn Salah Razan Aburumman Faisal Muhammad Note: I tried to include everything that's important from the doctor's slides but you can refer back to them after studying this sheet.. After you read this
20 Lojayn Salah Razan Aburumman Faisal Muhammad Note: I tried to include everything that's important from the doctor's slides but you can refer back to them after studying this sheet.. After you read this
Lujain Hamdan. Ayman Musleh & Yahya Salem. Mohammed khatatbeh
 12 Lujain Hamdan Ayman Musleh & Yahya Salem Mohammed khatatbeh the last lecture, we have studied the differences between the two divisions of the ANS: sympathetic and parasympathetic pathways which work
12 Lujain Hamdan Ayman Musleh & Yahya Salem Mohammed khatatbeh the last lecture, we have studied the differences between the two divisions of the ANS: sympathetic and parasympathetic pathways which work
Chapter 7. The Nervous System: Structure and Control of Movement
 Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
Chapter 7. Objectives
 Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
BIOH111. o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system
 BIOH111 o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook and required/recommended
BIOH111 o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook and required/recommended
By the name of Allah
 By the name of Allah Receptors function and signal transduction ( Hormones and receptors Types) We were talking about receptors of the neurotransmitters; we have 2 types of receptors: 1- Ionotropic receptors
By the name of Allah Receptors function and signal transduction ( Hormones and receptors Types) We were talking about receptors of the neurotransmitters; we have 2 types of receptors: 1- Ionotropic receptors
Ligand-Gated Ion Channels
 Ligand-Gated Ion Channels The Other Machines That Make It Possible... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics II
Ligand-Gated Ion Channels The Other Machines That Make It Possible... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics II
Pathophysiology of Pain
 Pathophysiology of Pain Wound Inflammatory response Chemical mediators Activity in Pain Path PAIN http://neuroscience.uth.tmc.edu/s2/chapter08.html Chris Cohan, Ph.D. Dept. of Pathology/Anat Sci University
Pathophysiology of Pain Wound Inflammatory response Chemical mediators Activity in Pain Path PAIN http://neuroscience.uth.tmc.edu/s2/chapter08.html Chris Cohan, Ph.D. Dept. of Pathology/Anat Sci University
Synaptic transmission
 Outline Synaptic transmission Sompol Tapechum M.D., Ph.D. Department of Physiology Faculty of Medicine Siriraj Hospital, Bangkok, Thailand. sisth@mahidol.ac.th 2 Structure of synapse Modes of synaptic
Outline Synaptic transmission Sompol Tapechum M.D., Ph.D. Department of Physiology Faculty of Medicine Siriraj Hospital, Bangkok, Thailand. sisth@mahidol.ac.th 2 Structure of synapse Modes of synaptic
TMC9 as a novel mechanosensitive ion channel
 TMC9 as a novel mechanosensitive ion channel Mechanical forces play numerous roles in physiology. When an object contacts our skin, it exerts a force that is encoded as touch or pain depending on its intensity.
TMC9 as a novel mechanosensitive ion channel Mechanical forces play numerous roles in physiology. When an object contacts our skin, it exerts a force that is encoded as touch or pain depending on its intensity.
Nervous System. 2. Receives information from the environment from CNS to organs and glands. 1. Relays messages, processes info, analyzes data
 Nervous System 1. Relays messages, processes info, analyzes data 2. Receives information from the environment from CNS to organs and glands 3. Transmits impulses from CNS to muscles and glands 4. Transmits
Nervous System 1. Relays messages, processes info, analyzes data 2. Receives information from the environment from CNS to organs and glands 3. Transmits impulses from CNS to muscles and glands 4. Transmits
Nervous System. Master controlling and communicating system of the body. Secrete chemicals called neurotransmitters
 Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS
 Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Chapter Nervous Systems
 The Nervous System Chapter Nervous Systems Which animals have nervous systems? (Which do not) What are the basic components of a NS? What kind of fish performs brain operations? What differentiates one
The Nervous System Chapter Nervous Systems Which animals have nervous systems? (Which do not) What are the basic components of a NS? What kind of fish performs brain operations? What differentiates one
11/8/16. Cell Signaling Mechanisms. Dr. Abercrombie 11/8/2016. Principal Parts of Neurons A Signal Processing Computer
 Cell Signaling Mechanisms Dr. Abercrombie 11/8/2016 Principal Parts of Neurons A Signal Processing Computer A Multitude of Synapses and Synaptic Actions Summation/Synaptic Integration 1 The Synapse Signal
Cell Signaling Mechanisms Dr. Abercrombie 11/8/2016 Principal Parts of Neurons A Signal Processing Computer A Multitude of Synapses and Synaptic Actions Summation/Synaptic Integration 1 The Synapse Signal
10.1: Introduction. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial cells) Dendrites.
 Dendrites.") 10.1: Introduction Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial
10.1: Introduction Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial
Communication within a Neuron
 Neuronal Communication, Ph.D. Communication within a Neuron Measuring Electrical Potentials of Axons The Membrane Potential The Action Potential Conduction of the Action Potential 1 The withdrawal reflex
Neuronal Communication, Ph.D. Communication within a Neuron Measuring Electrical Potentials of Axons The Membrane Potential The Action Potential Conduction of the Action Potential 1 The withdrawal reflex
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature588 SUPPLEMENTARY INFORMATION Supplemental Information Sensory neuron sodium channel Na v 1.8 is essential for pain at cold temperatures Katharina Zimmermann*, Andreas Leffler*, Alexandru
doi: 1.138/nature588 SUPPLEMENTARY INFORMATION Supplemental Information Sensory neuron sodium channel Na v 1.8 is essential for pain at cold temperatures Katharina Zimmermann*, Andreas Leffler*, Alexandru
Intracellular signalling cascades associated with TRP channels
 Gerald Thiel Intracellular signalling cascades associated with TRP channels Current state of investigations and potential applications Saarland University, Germany Campus Saarbrücken Department of Medical
Gerald Thiel Intracellular signalling cascades associated with TRP channels Current state of investigations and potential applications Saarland University, Germany Campus Saarbrücken Department of Medical
Psych 181: Dr. Anagnostaras
 Psych 181: Dr. Anagnostaras Lecture 5 Synaptic Transmission Introduction to synaptic transmission Synapses (Gk., to clasp or join) Site of action of most psychoactive drugs 6.5 1 Synapses Know basic terminology:
Psych 181: Dr. Anagnostaras Lecture 5 Synaptic Transmission Introduction to synaptic transmission Synapses (Gk., to clasp or join) Site of action of most psychoactive drugs 6.5 1 Synapses Know basic terminology:
Drug Receptor Interactions and Pharmacodynamics
 Drug Receptor Interactions and Pharmacodynamics Dr. Raz Mohammed MSc Pharmacology School of Pharmacy 22.10.2017 Lec 6 Pharmacodynamics definition Pharmacodynamics describes the actions of a drug on the
Drug Receptor Interactions and Pharmacodynamics Dr. Raz Mohammed MSc Pharmacology School of Pharmacy 22.10.2017 Lec 6 Pharmacodynamics definition Pharmacodynamics describes the actions of a drug on the
Cancer-induced bone pain
 Cancer-induced bone pain Common Prevalent in particular cancers: breast (73%), prostate (68%), thyroid (42%), lung (36%), renal (35%), colon (5%) Correlates with an increased morbidity Reduced performance
Cancer-induced bone pain Common Prevalent in particular cancers: breast (73%), prostate (68%), thyroid (42%), lung (36%), renal (35%), colon (5%) Correlates with an increased morbidity Reduced performance
Psychophysical laws. Legge di Fechner: I=K*log(S/S 0 )
") Psychophysical laws Legge di Weber: ΔS=K*S Legge di Fechner: I=K*log(S/S 0 ) Sensory receptors Vision Smell Taste Touch Thermal senses Pain Hearing Balance Proprioception Sensory receptors Table 21-1 Classification
Psychophysical laws Legge di Weber: ΔS=K*S Legge di Fechner: I=K*log(S/S 0 ) Sensory receptors Vision Smell Taste Touch Thermal senses Pain Hearing Balance Proprioception Sensory receptors Table 21-1 Classification
Chp. 16: AUTONOMIC N.S. (In Review: Peripheral N. S.)
") Chp. 16: AUTONOMIC N.S. (In Review: Peripheral N. S.) Peripheral nerves contain both motor and sensory neurons Among the motor neurons, some of these are somatic and innervate skeletal muscles while some
Chp. 16: AUTONOMIC N.S. (In Review: Peripheral N. S.) Peripheral nerves contain both motor and sensory neurons Among the motor neurons, some of these are somatic and innervate skeletal muscles while some
Somatosensation. Recording somatosensory responses. Receptive field response to pressure
 Somatosensation Mechanoreceptors that respond to touch/pressure on the surface of the body. Sensory nerve responds propotional to pressure 4 types of mechanoreceptors: Meissner corpuscles & Merkel discs
Somatosensation Mechanoreceptors that respond to touch/pressure on the surface of the body. Sensory nerve responds propotional to pressure 4 types of mechanoreceptors: Meissner corpuscles & Merkel discs
Pain Pathways. Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH
 Pain Pathways Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH Objective To give you a simplistic and basic concepts of pain pathways to help understand the complex issue of pain Pain
Pain Pathways Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH Objective To give you a simplistic and basic concepts of pain pathways to help understand the complex issue of pain Pain
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Investigating afferent mechanisms involved in bladder hypersensitivity
 Investigating afferent mechanisms involved in bladder hypersensitivity PhD Thesis Natasja Barki Thesis submitted in accordance with the requirements of the University of Sheffield for the degree of the
Investigating afferent mechanisms involved in bladder hypersensitivity PhD Thesis Natasja Barki Thesis submitted in accordance with the requirements of the University of Sheffield for the degree of the
Functions of the Nervous System. Fundamentals of the Nervous System & Nervous Tissue
 Fundamentals of the Nervous System & Nervous Tissue Overview Structure cell types & structures Neurophysiology membrane potential Synapse, neurotransmitters & receptors Functions of the Nervous System
Fundamentals of the Nervous System & Nervous Tissue Overview Structure cell types & structures Neurophysiology membrane potential Synapse, neurotransmitters & receptors Functions of the Nervous System
Receptors and Neurotransmitters: It Sounds Greek to Me. Agenda. What We Know About Pain 9/7/2012
 Receptors and Neurotransmitters: It Sounds Greek to Me Cathy Carlson, PhD, RN Northern Illinois University Agenda We will be going through this lecture on basic pain physiology using analogies, mnemonics,
Receptors and Neurotransmitters: It Sounds Greek to Me Cathy Carlson, PhD, RN Northern Illinois University Agenda We will be going through this lecture on basic pain physiology using analogies, mnemonics,
Electrical Properties of Neurons. Steven McLoon Department of Neuroscience University of Minnesota
 Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Chapter 12 Nervous Tissue. Copyright 2009 John Wiley & Sons, Inc. 1
 Chapter 12 Nervous Tissue Copyright 2009 John Wiley & Sons, Inc. 1 Terms to Know CNS PNS Afferent division Efferent division Somatic nervous system Autonomic nervous system Sympathetic nervous system Parasympathetic
Chapter 12 Nervous Tissue Copyright 2009 John Wiley & Sons, Inc. 1 Terms to Know CNS PNS Afferent division Efferent division Somatic nervous system Autonomic nervous system Sympathetic nervous system Parasympathetic
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
The Autonomic Nervous
 Autonomic Nervous System The Autonomic Nervous Assess Prof. Fawzia Al-Rouq System Department of Physiology College of Medicine King Saud University LECTUR (1) Functional Anatomy & Physiology of Autonomic
Autonomic Nervous System The Autonomic Nervous Assess Prof. Fawzia Al-Rouq System Department of Physiology College of Medicine King Saud University LECTUR (1) Functional Anatomy & Physiology of Autonomic
Modulation of TRP channels by resolvins in mouse and human
 July 9, 2015, Ion Channel Retreat, Vancouver Ion Channel and Pain Targets Modulation of TRP channels by resolvins in mouse and human Ru-Rong Ji, PhD Pain Research Division Department of Anesthesiology
July 9, 2015, Ion Channel Retreat, Vancouver Ion Channel and Pain Targets Modulation of TRP channels by resolvins in mouse and human Ru-Rong Ji, PhD Pain Research Division Department of Anesthesiology
Cell Membrane and Transport
 Cell Membrane and Transport 29/06/2015 11:08 AM Describe the Characteristics of the phospholipid Bilayer. The Phospholipid bilayer is made up of a double layer of membrane lipids that have a hydrophobic
Cell Membrane and Transport 29/06/2015 11:08 AM Describe the Characteristics of the phospholipid Bilayer. The Phospholipid bilayer is made up of a double layer of membrane lipids that have a hydrophobic
Chapter 16. APR Enhanced Lecture Slides
 Chapter 16 APR Enhanced Lecture Slides See separate PowerPoint slides for all figures and tables pre-inserted into PowerPoint without notes and animations. Copyright The McGraw-Hill Companies, Inc. Permission
Chapter 16 APR Enhanced Lecture Slides See separate PowerPoint slides for all figures and tables pre-inserted into PowerPoint without notes and animations. Copyright The McGraw-Hill Companies, Inc. Permission
The Nervous System. Dr. ZHANG Xiong Dept. of Physiology ZJU School of Medicine.
 The Nervous System Dr. ZHANG Xiong Dept. of Physiology ZJU School of Medicine Http://10.10.10.151 Part 1. Summary of the nervous system The Nervous System Central Nervous System Brain + Spinal Cord Peripheral
The Nervous System Dr. ZHANG Xiong Dept. of Physiology ZJU School of Medicine Http://10.10.10.151 Part 1. Summary of the nervous system The Nervous System Central Nervous System Brain + Spinal Cord Peripheral
Name Biology 125 Midterm #2 ( ) Total Pages: 9
 Total Pages: 9") Name - 1 - Biology 125 Midterm #2 (11-15-07) Part 1: (30 pts) Part 2: (40 pts) Part 3: (6 pts) Part 4: (12 pts) Part 5: (12 pts) Total Pages: 9 Total Score: (100 pts) 1 Name - 2 - Part 1: True or False.
Name - 1 - Biology 125 Midterm #2 (11-15-07) Part 1: (30 pts) Part 2: (40 pts) Part 3: (6 pts) Part 4: (12 pts) Part 5: (12 pts) Total Pages: 9 Total Score: (100 pts) 1 Name - 2 - Part 1: True or False.
BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1
 BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1 Terms you should know: synapse, neuromuscular junction (NMJ), pre-synaptic, post-synaptic, synaptic cleft, acetylcholine (ACh), acetylcholine
BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1 Terms you should know: synapse, neuromuscular junction (NMJ), pre-synaptic, post-synaptic, synaptic cleft, acetylcholine (ACh), acetylcholine
G-Protein Coupled Receptors GPCRs. GPCRs
 2 type of ligands 1 G-Protein Coupled Receptors GPCRs One of the largest protein families: > 1000 type of GPCRs in mammals >3% of the human genes Major drug targets: ~ 60 % of approved drugs interact with
2 type of ligands 1 G-Protein Coupled Receptors GPCRs One of the largest protein families: > 1000 type of GPCRs in mammals >3% of the human genes Major drug targets: ~ 60 % of approved drugs interact with
Applied Neuroscience. Conclusion of Science Honors Program Spring 2017
 Applied Neuroscience Conclusion of Science Honors Program Spring 2017 Review Circle whichever is greater, A or B. If A = B, circle both: I. A. permeability of a neuronal membrane to Na + during the rise
Applied Neuroscience Conclusion of Science Honors Program Spring 2017 Review Circle whichever is greater, A or B. If A = B, circle both: I. A. permeability of a neuronal membrane to Na + during the rise
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus