CD47 plays a role as a negative regulator in inducing protective immune responses to vaccination. against influenza virus
|
|
|
- Frederica Juliet Foster
- 5 years ago
- Views:
Transcription
1 JVI Accepted Manuscript Posted Online 18 May 216 J. Virol. doi:1.1128/jvi Copyright 216, American Society for Microbiology. All Rights Reserved CD47 plays a role as a negative regulator in inducing protective immune responses to vaccination against influenza virus Young-Tae Lee 1, Eun-Ju Ko 1,2, Youri Lee 1, Yu-Na Lee 1, 3, Zhen Bian 2, Yuan Liu 2 and Sang-Moo Kang 1,2, * 1 Center for Inflammation, Immunity & Infection, Institute for Biomedical Sciences, and 2 Program of Cellular Biology and Immunology, Department of Biology, Georgia State University, Atlanta, GA 333, USA. Short title: The role of CD47 in humoral immunity Key words: Influenza virus, CD47, influenza VLPs, Vaccine *To whom correspondence should be addressed. Sang-Moo Kang, PhD Center for Inflammation, Immunity & Infection, Institute for Biomedical Sciences, Georgia State University, Atlanta, GA 333, USA skang24@gsu.edu (S.M.K.) Telephone:
2 26 ABSTRACT An integrin-associated protein CD47, which is a ligand for the inhibitory receptor signal regulatory protein α, is expressed on B and T cells as well as on most innate immune cells. However, the roles of CD47 in the immune responses to viral infection or vaccination remain unknown. We have investigated the role of CD47 in inducing humoral immune responses after intranasal infection with virus or immunization with influenza virus-like particles (VLP). Virus infection or vaccination with VLP containing hemagglutinin (HA) from A/PR8/34 influenza virus induced higher levels of antigen-specific IgG2c isotype dominant antibodies in CD47-deficient () mice than those in wild type () mice. mice with vaccination showed higher protective efficacy against lethal challenge, as evidenced by no loss in body weight and reduced lung viral titers compared to mice. In addition, inflammatory responses which include cytokine production, leukocyte infiltrates, and interferon gamma (IFN-γ)-producing CD4 + T cells as well as anti-inflammatory cytokine (IL-1) were reduced in the lung of vaccinated mice after challenge with influenza virus. Analysis of lymphocytes indicated that GL7 + germinal center B cells were induced at higher levels in the draining lymph nodes of mice as compared with those in mice. Notably, mice exhibited significant increases in the numbers of antigen-specific memory B cells in spleens and plasma cells in bone marrow despite their lower levels of background IgG antibodies. Thus, these results suggest that CD47 plays a role as a negative regulator in inducing protective immune responses to influenza vaccination
3 49 IMPORTANCE Molecular mechanisms that control B cell activation to produce protective antibodies upon viral vaccination remain poorly understood. The CD47 molecule is known to be a ligand for the inhibitory receptor signal regulatory protein α and expressed on the surfaces of most immune cell types. CD47 was previously demonstrated to play an important role in modulating the migration of monocytes, neutrophils, polymorphonuclear neutrophils, and dendritic cells into the inflamed tissues. Results in this study demonstrate new roles of CD47 in negatively regulating the induction of protective IgG antibodies, germinal center B cells, and plasma cells secreting antigen-specific antibodies as well as macrophages upon influenza vaccination and challenge. As consequences, vaccinated CD47 deficient mice resulted in better control of influenza viral infection and enhanced protection. This study provides insights into understanding the regulatory functions of CD47 in inducing adaptive immunity to vaccination
4 INTRODUCTION Influenza viruses are common pathogens in the respiratory track and highly contagious, which can cause pulmonary diseases. Seasonal influenza virus variants annually cause significant levels of morbidity and mortality which mostly include elderly and sick people (1, 2). Vaccination is the most effective measure to prevent infections with a variety of pathogens, including influenza virus. Virus-like particles (VLPs) are able to effectively stimulate antigen-presenting cells (APCs) which in turn activate T and B cells (3-6). It has been demonstrated that immunization with influenza VLPs is capable of inducing protective humoral responses against seasonal and pandemic influenza virus infections (7-9). However, the mechanisms to evoke long-lasting immune responses are largely unknown. CD47 is a transmembrane protein, which is first identified as the integrin αvβ3. CD47 that is expressed on hematopoietic and non-hematopoietic cells can interact with an inhibitory receptor signal regulatory protein α (SIRPα) (1). SIRPα is also expressed on dendritic cells (DCs) and macrophages, while SIRPα is barely expressed on B and T cells (11, 12). It has been demonstrated that CD47/CD47 and CD47/SIRPα interactions are important for DC and neutrophil migration (13, 14). In addition, CD11b + DCs in the lungs express both CD47 and SIRPα, but CD13 + DCs express only CD47. It was also demonstrated that CD47 helps CD11b + DCs homing to draining lymph nodes during steady and inflammatory conditions (15). The populations of B22 + B cells and CD8 + T cells were reported to remain unchanged in the spleens of SIRPα and mice (16). However, a study reported that CD47-deficient () mice showed a defect in producing IgG antibodies to intravenous antigens (17). Another study demonstrated evidence that antigen-specific antibody responses were lower in mucosal tissues from mice using an allergic airway disease model (15). However, the role of CD47 in inducing specific antibodies in response to vaccination and protective immune responses against infectious viral disease remains largely unknown. Influenza VLP vaccines have been suggested as promising alternative vaccine candidates (18, 19), and also tested in clinical trials (2, 21). Antibody responses to hemagglutinin (HA) after vaccination are the 4
5 major immune correlates in conferring protection against influenza virus infections. Thus, in this study, we investigated the possible roles of CD47 in inducing protective humoral immunity to influenza viral infection or vaccination using influenza VLP containing HA as a vaccine target antigen. We found that vaccination of CD47-deficient mice with influenza VLP displayed robust humoral responses and provided enhanced protection, resulting in reducing inflammatory responses which include reduced production of proinflammatory cytokines and less immune cell infiltration into the lungs after challenge with a lethal dose of influenza virus infection. 5
6 MATERIALS AND METHODS Mice Wild type C57BL/6 (B6 ) mice and CD47 -/- mice () on C57BL/6 background were purchased from the Jackson Laboratory (Bar Harbor, Maine). Mice were bred at the department of animal resources in Georgia State University. Influenza VLP and immunization Influenza VLP was prepared as described (8). Briefly, to obtain influenza VLP, Sf9 cell were coinfected with recombinant baculoviruses (rbvs) expressing M1 and influenza virus hemagglutinin (HA) from A/PR8/34 virus and cultured in SF9-II serum free medium at 27 C for 3 days. Culture supernatants were collected by centrifugation at 2, x g for 2 minutes (min) to remove cells. Collected supernatants containing influenza VLPs were further spun by ultracentrifugation at 1, x g for 6 min. The VLP pellets were resuspended in phosphate-buffered saline (PBS) at 4 C for overnight. Discontinuous sucrose gradient (2%-3%-6%) was used to purify VLPs by ultracentrifugation at 1, x g for 1hour (h) at 4 C. The VLP bands were harvested and the amount of incorporated HA level was analyzed by Western blots and hemagglutination activity assays. Viral infection and mice (6-8 weeks old) were intranasally (i.n.) immunized with influenza VLPs (3µg) at weeks and 4. Immunized mice were anesthetized by isoflurane (Baxter, Deerfield, IL) and i.n. infected with A/PR8/34 virus in 5 µl of PBS per mouse at 6-7 weeks after secondary immunization. Challenge doses were 8 or 33 times the 5% mouse lethal dose (LD 5 ), where 1x LD 5 dose contains approximately 1 plaque forming units (PFU). Mice were daily monitored for 14 days to record body weight changes and survival rates or sacrificed at 6 days post infection (dpi). More than 25% body weight loss was recorded as dead mice. To compare the efficacy of vaccination, unimmunized mice were used as a negative-control group. All animal studies were approved and conducted under the guidelines by Georgia 6
7 148 State University s IACUC (A1425) Serum preparation and humoral immune responses The antibody titers specific for influenza virus were determined by enzyme-linked immunosorbent assay (ELISA) using inactivated influenza A/PR8/34 virus (ipr8) as a coating antigen (8). Briefly, inactivated PR8 (4µg/ml) was coated on 96-well microtiter plates (Nunc, Rochester, NY) and incubated for overnight at 4 C. The 96-well plates were washed with PBS containing.5 % Tween 2 (PBST) and 3% BSA in PBST was used for blocking reagents for 9 min at 37 C. Collected sera were serially diluted and incubated for 9 min at 37 C. Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG, IgG1, and IgG2c (Southern Biotech, Birmingham, AL) were used as secondary antibodies and then the tetramethybenzidine (TMB) peroxidase substrate (Sigma-Aldrich, St. Louis, Mo) as a substrate. The optical density (OD) values were measured using optical spectrophotometer reader at 45nm. The total antibody concentration was determined with respect to quantitative standard antibody concentration by using purified IgG, IgG1, IgG2c, and IgM. Cytokine assays Cytokines were determined by ELISAs, performed as previously described by Quan et al. (8). Briefly, Ready-Set-Go IL-6, IL-1, and TNF-αkits (ebioscience, San Diego, CA) were used to determine the levels cytokines in the bronchoalveolar lavage fluids (BALF) and lung extracts following the manufacturer s procedures. Cell preparation and flow cytometry Non-adherent cells were collected from the BALF 2 times in PBS. The rest of lung tissues were homogenated and cells were then passed through strainer and spun on 44/67% Percoll gradients at 28 rpm for 2 min. Cell band was harvested and washed with PBS. The frosted glass microscope slides were used to make cell suspensions from the mediastinal lymph node and spleen. Red blood cells were lysed 7
8 with ammonium chloride and lymphocytes were filtered through cell strainers. For flow cytometry analysis, all other markers were stained at 4 C using monoclonal antibodies (mab) specific for CD3, CD4, CD8, CD11b, CD11c, CD19, CD45, CD47, F4/8, and Ly6c (ebioscience or BD Pharmingen, San Diego, CA). The samples were analyzed on a Becton-Dickinson LSR-II/Fortessa flow cytometer (BD, San Diego, CA) and analyzed using the Flowjo software program (Tree Star Inc.). Intracellular cytokine staining analysis Lymphocytes were enriched from the BALF and lungs. Cells from the BALF of the same group were pooled to make enough cell numbers. Lymphocytes were in vitro stimulated in the presence of the synthetic peptide HA (YVQASGRVTVSTRRS) and HA (AELLVALENQHTIDL) (5μg/ml) (32) with Brefeldin A (BFA) (2μg/ml) for 5 h at 37 C incubator. Stimulated lymphocytes were stained with antibodies, including CD4 and CD8. Lymphocytes were then fixed/permeabilized using BD Cytofix/Cytoperm TM Plus kit and stained with IFN-γ cytokine antibodies. Lung viral titers Lung viral titers were determined using the fertilized chicken eggs which were incubated for 1 days at 37 C. Lung homogenates were serially diluted in PBS and injected into the allantoic sacs. Infected embryonated eggs were further incubated for 3 days and left in cold room overnight to euthanized embryos. The allantoic fluids were harvested and transferred into 96-well round bottom plates containing citrated chicken blood. The plates were left at room temperature for 3 mins and viral titers were calculated. Infected wells appeared a uniform reddish color, but negative wells resulted as dots in the center of round bottom plates. Statistics Unless otherwise stated, all results were presented as means ± SEM (standard error of mean). Statistical significance for all experiments was determined using an unpaired two-tailed Student s t test or one-way 8
9 ANOVA. Prism software (GraphPad software Inc, San Diego, CA) was used for all data analysis. The comparisons that were used to generate P values are indicated by horizontal lines (* P <.5, ** P <.1, *** P <.1). 23 9
10 RESULTS mice induce increased virus-specific IgG antibodies after infection despite lower background IgG antibodies. CD47 is expressed by hematopoietic cells, including T and B cells (22). To understand how CD47 deficiency intrinsically influences the production of natural immunoglobulin (Ig) G and IgM, blood samples were collected from naive C57BL/6 wild type () and mice (n = 12) and the total amounts of antigen non-specific background antibodies in sera were measured by ELISA. Both IgG and IgM antibodies were produced in CD47 deficient mice, but serum IgG levels were significantly lower in mice (.5mg/ml) than those in mice (.9 mg/ml) (Fig. 1A). In contrast, similar levels of IgM were observed in sera of (.3mg/ml) and mice (.2mg/ml). To determine whether CD47 molecule influenced lymphocyte homeostasis, the numbers of T and B cells were compared in the spleens of naïve to mice. Similar numbers of CD8 + T and B cells were detected in the spleen of and mice, but reduced CD4 + T cells in numbers were observed in the spleen of CD47 deficient mice (Fig. 1B). Next, we determined whether mice would induce lower levels of virus antigen specific IgG antibodies after influenza virus infection. Naïve and mice (n = 8) were intranasally infected with a sub-lethal low dose of A/PR8/34 virus (.5 x LD 5 ), and then PR8 virus specific IgG antibodies in sera were determined (Fig. 1C). Unexpectedly, mice were found to induce a higher antibody response to influenza virus infection at a sublethal dose than mice (Fig. 1C). To understand the possible correlation of CD47 between antigen presenting cells and T cells, naïve and mice were administered HA-VLPs and activation of antigen presenting cells was monitored. Intranasal administration of VLP antigens induced an increase in numbers of F4/8 + CD8 high and F4/8 + MHCII high activated macrophages in the lungs of mice than those in mice (Fig. 1D). These results suggest that naïve mice maintain lower levels of background IgG antibodies than naïve mice. However, it seems to be paradoxical that mice can raise virus antigen specific IgG 1
11 antibodies at significantly higher levels after infection with influenza virus compared to those in infected mice. Interestingly, mice showed an increase in numbers of MHCII positive activated antigen presenting macrophages in response to intranasal VLP antigen Higher levels of antigen-specific IgG2c antibodies are induced in mice after vaccination To determine the role of CD47 in inducing humoral responses to vaccination, and mice (n = 12) were intranasally immunized with 3µg influenza HA-VLP (A/PR8/34) twice at and 4 weeks. Immune sera were collected 3 weeks after prime and boost immunization. Following prime immunization, antibodies specific for inactivated A/PR8/34 (ipr8) virus were induced at significantly higher levels in mice than those in mice (Fig. 2A and B). Importantly, antibodies specific for ipr8 virus were induced at consistently higher levels in mice than those in mice, although vaccine antigen specific antibodies were increased in mice after boost immunization. The concentration of antibodies showed 4.2 fold increases in mice (34ng/ml) compared to those in mice (8ng/ml) (Fig. 2C). A pattern of antigen-specific IgG1 and IgG2c isotype antibodies is known to be dictated by T helper type 2 (Th2) and Th1-derived cytokines, respectively (24, 25). We compared a pattern of vaccine-specific antibody isotype levels in and mice. mice induced IgG2c isotype antibody predominantly, which was higher than the IgG2c level in mice (Fig. 2D and E). As a result, mice showed a 14-fold increase in the IgG2c/IgG1 ratio compared to mice, indicating that CD47 may play a negative regulator in inducing IgG2c antibody, a Th1 type immune response (Fig. 2F). These results suggest that antigen-specific IgG2c humoral responses are induced more effectively than IgG1 antibody in mice after immunization with influenza VLP vaccine or infection with influenza virus. mice are better protected against influenza after vaccination. 11
12 We determined whether mice with antibody responses at higher levels would be better protected against influenza virus infection. or mice (n = 12) after immunization with influenza VLP were infected with a lethal dose of A/PR8/34 influenza virus (8 x LD 5 ) (Fig. 3A). Naive and mice displayed severe body weight loss over 25% (endpoints) and died or had to be euthanized by day 6 (Fig. 3B). Vaccinated mice lost their weight up to 22-25% resulting in 6% survival until day 8 and gradually recovered from A/PR8 infection. In contrast, immunized mice with influenza VLP did not show obvious weight loss after lethal challenge infection, indicating higher efficacy of protection (Fig. 3A and B). Furthermore, we tested whether HA-VLP vaccination was able to confer protection of mice against a high dose of A/PR8/34 virus challenge. Vaccinated and mice (n = 1) were infected with A/PR8/34 influenza virus (33 x LD 5 ). As expected, all unimmunized mice and immunized mice displayed severe weight loss more than 25% or died 5-8 days after infection (Fig. 3C and D). In contrast, VLP-immunized mice showed around 1% body weight loss 3 days following infection and gradually recovered body weight until 14 days, suggesting that vaccination with HA-VLPs was able to confer protection in CD47 deficient mice even against a high dose of influenza virus infection. Lung viral titers were analyzed to determine the replication of challenge virus. Mice (n = 12) were infected with A/PR8/34 (8 x LD 5 ), BAL fluids and the lung tissues were harvested 6 days post challenge (Fig. 3E and, F). To determine egg infectious titers, the BAL fluids and lung lysates were serially diluted, injected into embryonated chicken eggs, and incubated for 3 days. Thus, these results suggest that mice induce more strong protective immunity of inhibiting viral replication by over 1, folds upon influenza VLP vaccination and infection, which is consistent with higher levels of vaccinespecific antibody responses in mice. mice with influenza VLP vaccination also controlled lung viral replication by approximately 1 folds but with less efficacy compared to mice (Fig. 12
13 282 3E and F) Vaccinated mice reduce inflammatory responses after infection. Over-induction of proinflammatory cytokines contributes to tissue damages during highly pathogenic influenza virus infections, including H5N1 and H1N1 strains (26, 27). To compare inflammatory responses, the levels of cytokines in BALF and lung extracts of animals (n = 6) immunized with influenza VLP were determined by cytokine ELISA 6 days post infection with A/PR8/34 virus. High levels of interleukin (IL)-6 were detected in the BAL and lungs from naïve and mice as well as immunized mice compared to immunized mice (Fig 4A). Interestingly, the level of tumor necrosis factor-alpha (TNF-α) was lower in both immunized groups than those in unimmunized and mice (Fig. 4B). Similar to IL-6 levels, unimmunized naïve and mice showed highest levels of IL-1 compared to immunized groups. Importantly, immunized mice showed higher amounts of IL-1 than those in immunized mice (Fig. 4C). To better understand lung inflammation, histological analysis was performed 6 days after challenge with A/PR8/34 virus (Fig. 4D). Lung histology results showed severe inflammation in lung tissues from unimmunized and mice, and also substantial inflammation in immunized mice compared to immunized mice (Fig. 4D). Immunized mice exhibited lowest levels of lung inflammation and inflammatory cytokines, as expected by better control of early influenza virus replication (Fig. 2). Therefore, inflammation of lung histopathology appeared to be correlated with high levels of lung viral titers and proinflammatory cytokines. Vaccinated mice show less innate immune cell recruitment upon virus infection. Increased inflammatory responses may induce a large number of cell infiltrates into the lungs in response to viral infection. The cell numbers were analyzed in the BAL and lungs 6 days post infection with a lethal dose of A/PR8/34 virus. The markers for flow cytometry were used to separate populations such as neutrophils and monocytes (28, 29). Importantly, we observed a pattern that the numbers of monocytes 13
14 (CD11b + Ly6c hi F4/8 + ) in the BAL and lungs of immune mice were lower than those in non-immune and mice, but they were higher than those in immune mice (Fig. 5A and B). Similar to monocyte infiltration, lower numbers of neutrophils (CD11b + Ly6c + F4/8 - ) were found in the BAL and lungs of immune mice compared to non-immune mice, but there was no significance (Fig. 5A and C). We observed the significant difference between non-immune and immune mice, because immune mice showed lowest levels of neutrophil recruitment. Thus, the results suggest that high viral load and inflammatory cytokines are correlated with an increase in numbers of innate immune cell recruitment. A previous study has demonstrated that influenza virus infection induces influx of respiratory DCs into the draining LNs (3). We analyzed to compare the number of CD11b + dendritic cells in mediastinal lymph nodes (MLN) 6 days after challenge with A/PR8/34 virus. In consistent with a low level of lung viral loads from vaccinated mice, reduced numbers of respiratory CD11b + DCs were detected in the MLN of vaccinated mice compared to those in vaccinated mice (Fig. 5D). Although similar levels of lung viral loads after challenge with influenza virus were found in unvaccinated naïve and mice, reduced numbers of CD11b + DCs were detected in the MLN of CD47 deficient mice. These results indicate that enhanced protection against influenza virus infection in mice reduces respiratory DC migration into the draining lymph nodes as well as lowers infiltrates of monocytes and neutrophils into the lungs of the mice. Vaccinated mice increase recall T cell recruitment in the lungs Memory T cells rapidly respond to antigen stimulation (31). To determine recall T cell recruitment in the site of infection in vaccinated mice after infection, T cells from the BALF and lungs were stimulated with the synthetic HA (YVQASGRVTVSTRRS) and HA (AELLVALENQHTIDL) peptides specific for CD4 + and CD8 + T cells, respectively (32). Intracellular IFN-γ expression was analyzed by flow 334 cytometry. After influenza virus infection, similar numbers of IFN-γ-producing CD T cells were
15 observed in the lungs of non-immune and mice as well as in immune and mice (Fig. 6A and B). Interestingly, immunized and mice showed increases in numbers of memory specific CD8 + T cells compared to naïve mice day 6 after challenge (Fig. 6A and B). To better understand the role of memory CD8 T cells in protection, CD8 population was depleted in immunized mice before influenza virus challenge. We observed no difference in lung viral clearance between immune and CD8-depleted immune mice (Fig. 6C). These results indicate that vaccination with HA- VLPs induces an increase in numbers of recall T cell recruitment in infected tissues. Importantly virus specific antibodies play a major role in controlling influenza virus infection rather than IFN-γ-producing CD8 + T cells. Intranasal vaccination induces higher levels of mucosal antibodies in mice. Respiratory tracts and lungs are the major sites of entry and replication for influenza virus. To determine whether humoral responses in mucosal tissues would have a correlation with enhanced protection in mice, vaccine-specific antibody levels were determined in BALF 6 days post challenge (Fig. 7). Consistent with serum antibody responses, IgG antibodies were induced at significantly higher levels in the lungs of vaccinated mice than those in naïve, naïve, and vaccinated mice. In addition to IgG antibodies specific for ipr8 virus, and mice with vaccination showed increased levels of IgA in the BALF compared to those in naive and mice, respectively (Fig. 7). Interestingly, the level of specific IgA was approximately 2 fold higher in vaccinated mice than that in vaccinated. These results support that influenza VLP immunization efficiently induces specific IgG and IgA antibodies in mucosal tissues more effectively in mice than in mice. VLP immunization enhances memory B and plasma cell responses in CD47 KO mice. GL7 is used for a marker for germinal center B cells (33). To determine whether CD47 deficiency would 36 influence B cell activation and the formation of germinal center, GL germinal center B cells
16 (CD19 + B22 + ) were analyzed by flow cytometry in MLN of immunized mice (n = 6) 6 days post infection. The frequency of GL7 + B cells in mice was higher than those in mice (Fig. 8A). In addition, the total number of GL7 + MLN B cells in mice was 3 fold higher than that in mice (Fig. 8B). We analyzed in vitro antibody production to determine whether CD47 deficiency would contribute to generating antigen-specific memory B cells and plasmas cells, splenocytes and bone marrow cells were harvested and cultured in 96 well plates coated with ipr8 virus. The higher amounts of specific IgG antibodies were detected in the spleen and bone marrow of vaccinated mice than those of vaccinated mice 1 or 5 days after in vitro incubation (Fig. 8C and D). As expected, very low levels of specific antibodies were observed in naïve and mice compared to vaccinated and mice. More importantly, vaccinated mice showed significantly higher levels of virus specific antibody secreting cell responses. We next determined CD47 expression levels on B and T cells under naïve and infection conditions. The levels of CD47 expression on B cells were similarly observed regardless of infection or immunization of mice (Fig. 8E). B cells are likely to express higher levels of CD47 compared to those in T cells (Fig. 8F). Interestingly, CD47 expression on T cells may be downregulated slightly after viral infection compared to that on T cells in naïve mice although these differences were minor due to low expression levels (Fig. 8F). Therefore, these results suggest that CD47 would have a more significant role in B cell immunity than in T cells and that CD47 may act as a negative regulator in B cell activation and differentiation. 16
17 DISCUSSION In this study, we have shown that CD47 deficiency in mice resulted in generating enhanced levels of antigen-specific IgG antibodies and higher efficacy of protection after vaccination despite low levels of antigen non-specific natural IgG serum antibodies. For the first time, this present study provides evidence that CD47 plays a role as a negative regulator on B cells in inducing antigen-specific antibodies. In addition, CD47 appeared to have a regulatory role in generating germinal center B cells and antibodysecreting plasma cells after VLP vaccination and virus challenge. Reduced CD4 + T cells in numbers were observed in the spleen of either CD47 or SIRPα deficient mice, suggesting that interaction between CD47 and its ligand SIRPα plays an important role in T cell homeostasis (16). In line with a previous study, we found that the numbers of CD4 + T cells but not of CD8 + T and B cells were reduced in the spleens of CD47-deficient mice (Fig. 1B). Also, naïve mice maintain low levels of IgG antibodies. Therefore, it was expected that mice would have defects in inducing protective antibody immune responses after influenza VLP vaccination. Unexpectedly, higher levels of antigen-specific IgG and IgG2c antibodies were observed in VLP-vaccinated mice. It has been shown that dendritic cells and cytokine milieu influence a Th1 immune response. Consistent with Th1 type IgG isotype responses in this study, a previous study demonstrated that CD47 deficiency enhanced Th1 dominant cell response producing IFN-γ and Th1-biased antibody response (34), implying that CD47 is a negative regulator of Th1 immune response. Thus, we expected that vaccination 17
18 with VLPs might promote an enhanced Th1 dominant response and in turn induce high ratio of IgG2c isotype antibody, although similar numbers of antigen-specific CD4 + and CD8 + T cells were observed in the lungs of non-immune and mice as well as in immune and mice. One possible explanation for low Th1 cell responses in mice is that high viral loads in immune mice might be a driving factor in boosting higher levels of IFN-γ CD4 + T cells primed in immune mice compared to those in immune mice. Results in this study provide evidence that mice vaccinated with influenza VLP are more effective in generating protective immunity against a lethal dose of A/PR8/34 virus, as observed by lower viral titers at day 6 and no loss in body weight after challenge. Consistent with this active immunization and protection, naïve mice were found to induce higher levels of virus-specific IgG antibodies and to be more effectively protected after viral infection compared to mice. Thus, these results suggest that CD47 negatively modulates protective immune responses, consistent with previous findings (34). In contrast to our findings, others have demonstrated that mice have a defect in producing antigen-specific antibodies in response to intra-venous particulate antigens, due to the fact that CD11c + 33D1 + marginal zone DCs were low in the spleen, which is important for up-taking blood-borne antigens (35). However, B cell responses to soluble antigens were normal, implying that T and B cells are not defective (17). Oral immunization induced a significantly lower level of intestinal ovalbumin (OVA)- IgA from mice compared to mice, but similar levels of serum OVA-IgG were detected in and mice (36). We found that viral infection of mice at a sub-lethal dose induced higher levels of virus specific IgG antibodies compared to mice, which is consistent with the results from VLP vaccination. Thus, although it is possible that CD47 may be a key regulator in inducing specific humoral immunity to viral and VLP vaccine antigens, further studies are required to better understand the immunological roles of CD47 in terms of the different routes of vaccination. CD47 helps homing of respiratory CD11b + DCs to the draining LN during steady- and inflammatory-state 18
19 conditions (15). However, no difference was found in expression of costimulatory molecules such as CD4, CD8, or CD86 (17). In consistent with a previous finding, we also observed that reduced numbers of respiratory DCs were detected in the MLN of non-immune mice compared to those in non-immune mice day 6 after influenza virus infection. Delayed DC migration into the MLN may increase duration of antigen stimulation to activate T cells in the infection site. In other words, antibodies can be rapidly induced at the site of T and B cell zones (extrafollicular sites) in the draining LN, but shortlived plasma cells can be generated (37). However, the slower antigen stimulation in the germinal centers (intra- and/or extra-follicular sites) can generate antigen-specific memory B cells and long-lived plasma cells (38, 39). Influenza virus infection induces antigen persistence which can induce poly clonal memory CD4 + T cell responses 3-4 weeks after viral clearance (4), suggesting that sustained CD4 + T cells may contribute to increasing antigen-specific antibody production. In addition, prolonged antigens, which may have adjuvant-like depot effects, are able to continuously stimulate antibody-secreting cells, resulting in high levels of antibody production (41). This hypothesis is further supported by our findings that influenza VLP vaccination of mice induced higher levels of germinal center phenotypic B cells, memory B cells, and antibody secreting plasma cells upon antigen exposure. Importantly, we found that B cells expressed higher levels of CD47 compared to those in T cells and that antigen-specific antibody response was enhanced in the absence of CD47. These results provide evidence that CD47 is a negative regulator in humoral immune responses to viral infection or vaccination. Therefore, it implies that CD47 might have a more significant role in B cell immunity than in T cells. Alternatively, other cell types may be involved in increased antigen-specific antibody production. The SIRPα signal pathway is not blocked in antigen-presenting cells (APCs) in the absence of CD47. Interaction of SIRPα on macrophages with CD47 induces phosphorylation of cytoplasmic immunoreceptor tyrosine-based inhibition motif (ITIM) of SIRPα by Src homology region 2 domain-containing phosphatase-1 (SHP-1) and SHP-2 which can inhibit signaling pathways, including the mitogen-activated protein kinases (MAPK) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) (42). 19
20 Furthermore, a previous study has demonstrated that ligation of CD47 on human dendritic cells with antibodies limited inflammatory cytokine responses but not for their functions (43), which implied enhanced cytokine production during vaccination with influenza VLPs in mice. It is possible that CD47 deficiency may influence antigen presenting cells (APCs) and CD4 + T cell activation pathway as well as B cell activation. In line with previous findings, we detected that mice induced more F4/8 + CD8 high and F4/8 + MHCII high activated macrophages in the lungs than mice after VLP vaccination. It suggests that F4/8 + CD8 high and F4/8 + MHCII high activated macrophages further activate CD4 + T cells and/or induce cytokines, which can provide a biased help to the B cells to induce IgG2c isotype antibodies more effectively. In addition, it was reported that macrophages showed increased phagocytosis of cancer cells in the absence of CD47-mediated signaling (44). Interestingly, SIRPα-Fc treatment interfering with CD47 pathway was able to reduce T helper 2 (Th2)-driven allergic airway inflammation (45), which is in line with the results of IgG2c dominant antibody responses in mice after intranasal VLP vaccination. Our understanding of the mechanisms for triggering respiratory disease is largely unknown. However, there is evidence that inflammatory cytokines and infiltration of leukocytes can cause severe respiratory illness during influenza virus infection. We have demonstrated that unimmunized and mice as well as immunized mice showed high levels of TNF-α, IL-6, and IL-1 as well as infiltration of leukocytes after challenge (Fig. 4 and 5), whereas immunized mice with high amounts of specific antibodies showed low amounts of inflammatory and anti-inflammatory cytokines. In support for our findings, TNF-α and IL-1 are able to up-regulate leukocyte adhesion molecules and IL-6 (46). High levels of TNF-α and IL-6 cytokines therefore indicate severe inflammatory condition upon influenza virus challenge. In addition, an anti-inflammatory cytokine, IL-1, was high in unimmunized group compared to immunized and groups, suggesting that IL-1 may be required to downregulate excessive lung inflammation in naïve host after infection. In support with our observation, a study suggested that a high IL-1 level may be an indicator for disease progression and severity (47). 2
21 Defective neutrophil migration was observed in the absence of CD47 expression (13). Deficiency or blockade of CD47 is protected from lipopolysaccharide (LPS)-induced acute inflammatory disease and Th2-driven airway inflammation by inhibiting the migration of leukocytes and neutrophils, respectively (45, 48). However, in this study, naive mice showed higher or similar cellularity of neutrophils in the BAL and lungs compared with those in naïve mice upon influenza virus infection, suggesting no defect in migration of CD47-deficient neutrophils into the lung. Also, naïve mice were able to induce higher levels of virus specific antibodies, resulting in more resistant to a lower dose of viral infection than mice. It is also possible that the roles of CD47 in modulating monocyte and neutrophil migration into the inflamed tissues might be different between LPS injection and influenza virus infection. Eventually, high levels of virus-specific IgG antibody responses by influenza VLP vaccination of mice resulted in effective control of lung viral loads leading to reduction of inflammation and less leukocyte recruitment. Taken together, our results demonstrate that influenza VLP vaccination can induce higher amounts of virus-specific antibodies and is able to confer enhanced protection against lethal doses of influenza virus infections in the absence of CD47. In addition, mice generated high numbers of germinal center B cells and antibody-secreting memory cells. Thus, these studies provide insights into possible negative regulatory roles of CD47 on B cell activation in developing antigen-specific antibody responses. Further study is required to understand the underlying mechanism by which CD47 negatively regulates B cell activation. 21
22 ACKNOWLEDGMENTS This work was supported by NIH/NIAID grants AI1517 (S.M.K.), AI93772 (S.M.K.), and AI (S.M.K.). 22
23 FIGURE LEGENDS FIG 1. CD47-deficient mice show increased IgG antibody responses to viral infection despite low natural IgG antibody levels and macrophage activation to vaccination. (A) Natural antibodies in naïve mice. Preimmune sera were collected from wild-type C57BL/6 (n=6) and (n=6) mice. Total immunoglobulin (Ig) G and M levels were determined in the blood by ELISA and measured at OD 45nm. (B) T and B cells in naïve mice. T and B cell numbers were analyzed specific markers such as CD3, CD4, CD8, and CD19 in the spleens of naïve and mice. (C) Virus specific IgG antibodies after infection of naïve mice. Naïve and mice were intranasally infected with a low dose of A/PR8 virus (.5 x LD 5 ). Anti-sera were serially diluted and antigen-specific IgG levels were analyzed by ELISA against inactivated A/PR8/34 virus 3 weeks after infection and measured at OD 45nm. (D) Macrophage responses to vaccination. The cells were collected from the lungs of and mice 6 days after vaccination (3µg of HA-VLPs). The activation markers including CD8 and MHCII were used to analyze the status of macrophage activation. Statistical significance was determined using an unpaired two-tailed Student s t test. Error bars are means ± SEM of concentration from individual animals. *, P<.5; **, P<.1. No significant difference is indicated as NS (not significant). FIG 2. HA-VLP immunization induces enhanced antigen-specific antibody responses in 23
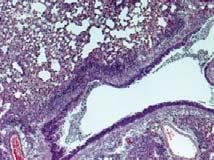
24 mice. and mice were intranasally immunized with HA-VLPs (3µg/mouse) at week and boosted at week 4. Immune sera were collected 3 weeks after prime and boost immunization from animals. (A and B) Anti-sera were serially diluted and antigen-specific IgG levels were analyzed by ELISA against inactivated A/PR8/34 virus and measured at OD 45nm. (C) Total concentration of IgG (ng/ml) against to virus was also calculated based on standard mouse IgG concentration. (D and E) Isotypes of A/PR8/34 specific antibodies were determined using anti-mouse specific isotype antibodies. (F) The ratios of IgG2c relative to IgG1 were measured in and mice. Statistical significance was determined using an unpaired two-tailed Student s t test. Error bars are means ± SEM of concentration or ratios from individual animals. ***, P<.1. FIG 3. Immunization of induces enhanced protection against influenza virus infection. Immunized and mice were challenged with a lethal dose of A/PR8 influenza virus (8 x LD 5 ) (A and B) or (33 x LD 5 ) (C and D) 6 weeks after boost immunization. Body weight changes (A and C) and survival rates (B and D) were monitored daily for 14 day. Lung viral titers in the BAL fluids (E) and lungs (F). Mice were infected with A/PR8/34 (8 x LD 5 ) and the lung tissues were harvested 6 days post challenge. Viral titers were determined by inoculation of chicken eggs with diluted lung samples. EID 5, 5% egg infectious dose. Naïve indicates unimmunized wild-type mice. Statistical significance was determined using an unpaired two-tailed Student s t test or one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; ***, P<.1. FIG 4. Vaccination of mice results in reduction of inflammatory responses. BALF and lung extracts were used to determine inflammatory cytokine levels 6 days after challenge with A/PR8 virus. IL-6 (A), TNF-α (B), and IL-1 (C) were determined in the BALF and lungs of unimmunized and immunized groups. (D) Lung histology. The lungs were harvested from vaccinated and mice 6 days after challenge with a lethal dose of A/PR8 virus (8 x LD 5 ). The tissues were fixed with 1% neutral buffered formalin solution. The lungs were embedded in paraffin and 5μm sections were stained 24
25 with hematoxylin and eosin. Photographs were taken at a magnification of 5. Statistical significance was determined using one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; **, P<.1. No significant difference is indicated as NS (not significant) FIG 5. Vaccinated mice decrease immune cell recruitments upon influenza virus infection. Immune cells were harvested from BALF, lungs and MLN 6 days post infection with A/PR8/34 influenza virus. The frequencies and numbers of monocytes (A and B) and neutrophils (A and C) were analyzed at 6 dpi in the BALF and lung in and mice. (D) Respiratory dendritic cell subsets were analyzed in the MLN using specific markers such as CD11c, CD11b, and MHCII. Statistical significance was determined using one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; **, P<.1; ***, P<.1. No significant difference is indicated as NS (not significant). FIG 6. Enhanced protective immunity in appears to reduce recall T cell responses. Immune cells were harvested from the BAL and lungs of vaccinated mice 6 days post infection (dpi) with A/PR8 influenza virus. (A and B) The cells that are making IFN-γ determined by culture for 5 h in the presence of HA peptides. Antigen-specific T cells were determined by intracellular cytokine staining using anti-ifnγ antibodies. (C) Lung viral titers in the lungs. HA-VLP immune mice received either anti-cd8 depletion antibodies or PBS prior to A/PR8/34 (8 x LD 5 ) infection. The lung tissues were harvested 6 dpi. Viral titers were determined by inoculation of chicken eggs with diluted lung samples. EID 5, 5% egg infectious dose. Statistical significance was determined using one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; **, P<.1. No significant difference is indicated as NS (not significant). FIG 7. Mucosal vaccination induces enhanced antigen-specific IgG and IgA in mice. BAL fluids were used to determine mucosal antibody responses 6 days post infection. Antigen-specific IgG and 25
26 IgA levels were analyzed by ELISA against inactivated A/PR8/34 virus and measured at OD 45nm. Statistical significance was determined using one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; **, P<.1; ***, P< FIG 8. CD47 deficiency enhances memory humoral immune responses. B cell activation and memory antibody responses were determined in the MLN, spleen, and bone marrow 6 days after challenge with influenza virus. The frequency (A) and number (B) of GL7+ germinal center B cells were analyzed in the MLN of immunized mice. The cells from the spleens and bone marrows were collected and cultured for 1 day (C) or 5 days (D) to detect IgG specific to inactivated A/PR8 virus. CD47 expression levels on B cells (E) and T cells (F) were determined under naïve and infection conditions using flow cytometry. Statistical significance was determined using an unpaired two-tailed Student s t test or one-way ANOVA. Error bars are means ± SEM of concentration or ratios from individual animals. *, P<.5; **, P<.1; ***, P<.1. No significant difference is indicated as NS (not significant). 26
27 References: 1. Thompson WW, Shay DK, Weintraub E, Brammer L, Bridges CB, Cox NJ, Fukuda K. 24. Influenza-associated hospitalizations in the United States. JAMA : the journal of the American Medical Association 292: Molinari NA, Ortega-Sanchez IR, Messonnier ML, Thompson WW, Wortley PM, Weintraub E, Bridges CB. 27. The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine 25: Buonaguro L, Tornesello ML, Tagliamonte M, Gallo RC, Wang LX, Kamin-Lewis R, Abdelwahab S, Lewis GK, Buonaguro FM. 26. Baculovirus-derived human immunodeficiency virus type 1 virus-like particles activate dendritic cells and induce ex vivo T- cell responses. Journal of virology 8: Moron VG, Rueda P, Sedlik C, Leclerc C. 23. In vivo, dendritic cells can cross-present viruslike particles using an endosome-to-cytosol pathway. Journal of immunology 171: Song H, Wittman V, Byers A, Tapia T, Zhou B, Warren W, Heaton P, Connolly K. 21. In vitro stimulation of human influenza-specific CD8+ T cells by dendritic cells pulsed with an influenza virus-like particle (VLP) vaccine. Vaccine 28: Sailaja G, Skountzou I, Quan FS, Compans RW, Kang SM. 27. Human immunodeficiency virus-like particles activate multiple types of immune cells. Virology 362: Song JM, Hossain J, Yoo DG, Lipatov AS, Davis CT, Quan FS, Chen LM, Hogan RJ, Donis RO, Compans RW, Kang SM. 21. Protective immunity against H5N1 influenza virus by a single dose vaccination with virus-like particles. Virology 45: Quan FS, Huang C, Compans RW, Kang SM. 27. Virus-like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. Journal of virology 81: Quan FS, Vunnava A, Compans RW, Kang SM. 21. Virus-like particle vaccine protects against 29 H1N1 pandemic influenza virus in mice. PloS one 5:e Brown EJ, Frazier WA. 21. Integrin-associated protein (CD47) and its ligands. Trends in cell biology 11: Matozaki T, Murata Y, Okazawa H, Ohnishi H. 29. Functions and molecular mechanisms of the CD47-SIRPalpha signalling pathway. Trends in cell biology 19: Seiffert M, Brossart P, Cant C, Cella M, Colonna M, Brugger W, Kanz L, Ullrich A, Buhring HJ. 21. Signal-regulatory protein alpha (SIRPalpha) but not SIRPbeta is involved in T-cell activation, binds to CD47 with high affinity, and is expressed on immature CD34(+)CD38(-) hematopoietic cells. Blood 97: Liu Y, O'Connor MB, Mandell KJ, Zen K, Ullrich A, Buhring HJ, Parkos CA. 24. Peptide-mediated inhibition of neutrophil transmigration by blocking CD47 interactions with signal regulatory protein alpha. Journal of immunology 172: Van VQ, Lesage S, Bouguermouh S, Gautier P, Rubio M, Levesque M, Nguyen S, Galibert L, Sarfati M. 26. Expression of the self-marker CD47 on dendritic cells governs their trafficking to secondary lymphoid organs. The EMBO journal 25: Raymond M, Rubio M, Fortin G, Shalaby KH, Hammad H, Lambrecht BN, Sarfati M. 29. Selective control of SIRP-alpha-positive airway dendritic cell trafficking through CD47 is critical for the development of T(H)2-mediated allergic inflammation. The Journal of allergy and clinical immunology 124: e Sato-Hashimoto M, Saito Y, Ohnishi H, Iwamura H, Kanazawa Y, Kaneko T, Kusakari S, Kotani T, Mori M, Murata Y, Okazawa H, Ware CF, Oldenborg PA, Nojima Y, Matozaki T Signal regulatory protein alpha regulates the homeostasis of T lymphocytes in the spleen. Journal of immunology 187: Hagnerud S, Manna PP, Cella M, Stenberg A, Frazier WA, Colonna M, Oldenborg PA. 26. Deficit of CD47 results in a defect of marginal zone dendritic cells, blunted immune 27
28 response to particulate antigen and impairment of skin dendritic cell migration. Journal of immunology 176: Kang SM, Kim MC, Compans RW Virus-like particles as universal influenza vaccines. Expert review of vaccines 11: Kang SM, Pushko P, Bright RA, Smith G, Compans RW. 29. Influenza virus-like particles as pandemic vaccines. Current topics in microbiology and immunology 333: Lopez-Macias C, Ferat-Osorio E, Tenorio-Calvo A, Isibasi A, Talavera J, Arteaga-Ruiz O, Arriaga-Pizano L, Hickman SP, Allende M, Lenhard K, Pincus S, Connolly K, Raghunandan R, Smith G, Glenn G Safety and immunogenicity of a virus-like particle pandemic influenza A (H1N1) 29 vaccine in a blinded, randomized, placebo-controlled trial of adults in Mexico. Vaccine 29: Khurana S, Wu J, Verma N, Verma S, Raghunandan R, Manischewitz J, King LR, Kpamegan E, Pincus S, Smith G, Glenn G, Golding H H5N1 virus-like particle vaccine elicits cross-reactive neutralizing antibodies that preferentially bind to the oligomeric form of influenza virus hemagglutinin in humans. Journal of virology 85: Gresham HD, Dale BM, Potter JW, Chang PW, Vines CM, Lowell CA, Lagenaur CF, Willman CL. 2. Negative regulation of phagocytosis in murine macrophages by the Src kinase family member, Fgr. The Journal of experimental medicine 191: Wilson MS, Feng CG, Barber DL, Yarovinsky F, Cheever AW, Sher A, Grigg M, Collins M, Fouser L, Wynn TA. 21. Redundant and pathogenic roles for IL-22 in mycobacterial, protozoan, and helminth infections. Journal of immunology 184: Takeda K, Tanaka T, Shi W, Matsumoto M, Minami M, Kashiwamura S, Nakanishi K, Yoshida N, Kishimoto T, Akira S Essential role of Stat6 in IL-4 signalling. Nature 38: Snapper CM, Paul WE Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science 236: Szretter KJ, Gangappa S, Lu X, Smith C, Shieh WJ, Zaki SR, Sambhara S, Tumpey TM, Katz JM. 27. Role of host cytokine responses in the pathogenesis of avian H5N1 influenza viruses in mice. Journal of virology 81: Kash JC, Basler CF, Garcia-Sastre A, Carter V, Billharz R, Swayne DE, Przygodzki RM, Taubenberger JK, Katze MG, Tumpey TM. 24. Global host immune response: pathogenesis and transcriptional profiling of type A influenza viruses expressing the hemagglutinin and neuraminidase genes from the 1918 pandemic virus. Journal of virology 78: Smith KA, Harcus Y, Garbi N, Hammerling GJ, MacDonald AS, Maizels RM Type 2 innate immunity in helminth infection is induced redundantly and acts autonomously following CD11c(+) cell depletion. Infection and immunity 8: Dunay IR, Damatta RA, Fux B, Presti R, Greco S, Colonna M, Sibley LD. 28. Gr1(+) inflammatory monocytes are required for mucosal resistance to the pathogen Toxoplasma gondii. Immunity 29: Legge KL, Braciale TJ. 23. Accelerated migration of respiratory dendritic cells to the regional lymph nodes is limited to the early phase of pulmonary infection. Immunity 18: Masopust D, Vezys V, Marzo AL, Lefrancois L. 21. Preferential localization of effector memory cells in nonlymphoid tissue. Science 291: Crowe SR, Miller SC, Brown DM, Adams PS, Dutton RW, Harmsen AG, Lund FE, Randall TD, Swain SL, Woodland DL. 26. Uneven distribution of MHC class II epitopes within the influenza virus. Vaccine 24: Pasare C, Medzhitov R. 25. Control of B-cell responses by Toll-like receptors. Nature 438: Bouguermouh S, Van VQ, Martel J, Gautier P, Rubio M, Sarfati M. 28. CD47 expression on T cell is a self-control negative regulator of type 1 immune response. Journal of immunology 18:
29 Martin F, Kearney JF. 22. Marginal-zone B cells. Nature reviews. Immunology 2: Westlund J, Livingston M, Fahlen-Yrlid L, Oldenborg PA, Yrlid U CD47-deficient mice have decreased production of intestinal IgA following oral immunization but a maintained capacity to induce oral tolerance. Immunology 135: Smith KG, Hewitson TD, Nossal GJ, Tarlinton DM The phenotype and fate of the antibody-forming cells of the splenic foci. European journal of immunology 26: Wolniak KL, Shinall SM, Waldschmidt TJ. 24. The germinal center response. Critical reviews in immunology 24: Chappell CP, Jacob J. 26. Identification of memory B cells using a novel transgenic mouse model. Journal of immunology 176: Jelley-Gibbs DM, Brown DM, Dibble JP, Haynes L, Eaton SM, Swain SL. 25. Unexpected prolonged presentation of influenza antigens promotes CD4 T cell memory generation. The Journal of experimental medicine 22: Stills HF, Jr. 25. Adjuvants and antibody production: dispelling the myths associated with Freund's complete and other adjuvants. ILAR journal / National Research Council, Institute of Laboratory Animal Resources 46: Kong XN, Yan HX, Chen L, Dong LW, Yang W, Liu Q, Yu LX, Huang DD, Liu SQ, Liu H, Wu MC, Wang HY. 27. LPS-induced down-regulation of signal regulatory protein {alpha} contributes to innate immune activation in macrophages. The Journal of experimental medicine 24: Demeure CE, Tanaka H, Mateo V, Rubio M, Delespesse G, Sarfati M. 2. CD47 engagement inhibits cytokine production and maturation of human dendritic cells. Journal of immunology 164: Tseng D, Volkmer JP, Willingham SB, Contreras-Trujillo H, Fathman JW, Fernhoff NB, Seita J, Inlay MA, Weiskopf K, Miyanishi M, Weissman IL Anti-CD47 antibodymediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response. Proceedings of the National Academy of Sciences of the United States of America 11: Raymond M, Van VQ, Rubio M, Welzenbach K, Sarfati M. 21. Targeting SIRP-alpha protects from type 2-driven allergic airway inflammation. European journal of immunology 4: Van Reeth K. 2. Cytokines in the pathogenesis of influenza. Veterinary microbiology 74: Yu X, Zhang X, Zhao B, Wang J, Zhu Z, Teng Z, Shao J, Shen J, Gao Y, Yuan Z, Wu F Intensive cytokine induction in pandemic H1N1 influenza virus infection accompanied by robust production of IL-1 and IL-6. PloS one 6:e Su X, Johansen M, Looney MR, Brown EJ, Matthay MA. 28. CD47 deficiency protects mice from lipopolysaccharide-induced acute lung injury and Escherichia coli pneumonia. Journal of immunology 18:
30 Figure 1 A Concentration (mg/ml) Absorbance (45nm) C Natural antibodies ** IgG IgG to ipr8 IgM /Serum dilution NS (PR8) (PR8) D B # of cells (1x 1 ) 7 # of cells (1x 1 ) * Spleen CD4+ T cells CD8+ T cells CD19+ B cells * CD11c+CD11b- F4/8+CD8hi NS * NS CD11c+CD11b- F4/8+MHCII hi
31 Figure 2 A Absorbance (45nm) D Absorbance (45nm) IgG to ipr8 B IgG to ipr8 C IgG concentration (Preimmune) (Prime) (Boost) /Serum dilution IgG1 IgG2c E Absorbance (45nm) Absorbance (45nm)2.5. (Preimmune) (Prime) (Boost) /Serum dilution IgG1 IgG2c /Serum dilution 1 1/Serum dilution IgG (ng/ml) F IgG2c/IgG1 ratio *** *** Ratio of IgG2c/IgG1
32 Figure 3 % change in body weight A C % change in body weight E Virus titers (log 1 EID5 ) ,P< x LD Days post challenge * P< x LD Days post challenge Virus titers (BAL) *** ** * * *** Survival (%) Survival (%) B 12 1 D F Virus titers (log 1 EID5 ) x LD Days post challenge x LD Days post challenge Virus titers (Lung) *** *** *** *** ***















33 Figure 4 Concentration (pg/ml) D A IL-6 * NS NS BAL * ** NS Lung B Concentration (pg/ml) NS * ** * BAL TNF-α NS * ** ** Lung C Concentration (pg/ml) IL-1 *** * *** ** * NS NS BAL ** Lung Downloaded from on December 23, 218 by guest
34 Figure 5 A 1 5 Monocyte 32% 26% 26% 27% % 34% 38% 24% Ly6c Neutriphil BAL 1 1 # of cells (1x 1 ) 5 # of cells (1x 1 ) 5 B C D # of cells (1x 1 ) % Monocytes in BAL ** Neutrophils in BAL CD11b+ DCs in MLN * 64% *** 18% * # of cells (1x 1 ) 6 # of cells (1x 1 ) 6 Monocytes in Lung 6 *** ** *** *** ** 49% NS NS NS 19% % 17% F4/8 Neutrophils in Lung ** 45% *** ** Lung
 IFN-γ B 15 1 5 C 1 3 1 2 1 1 15 1 1 5 1 4 1 3 1")

35 Figure 6 A % 2.2% 1.7% 1.1% # of cells Virus titers (log 1 EID5 ) IFN-γ B C # of cells CD4 2.3% 4.9% 2.6% 4.3% IFN-γ+ CD4+ T cells in Lung NS ** NS ** Virus titers (Lung) NS CD8X CD8 IFN-γ+ CD8+ T cells in Lung NS * *
36 Figure 7 IgG concentration (ng/ml) IgG to ipr8 in BAL ** IgA to ipr8 in BAL ** *** 1 8 *** IgA concentration (ng/ml) * ***

37 Figure 8 B22 A C E IgG concentration (ng/ml) % of Max GL7.51%.72% *** ****** 1day * * B # of cells D days ** *** 2 ** ****** *** *** ****** Spleen Bone marrow Spleen Bone marrow CD47 Negative Naive B cells B cells (Naive & infection) B cells (HA-VLP & infection) IgG concentration (ng/ml) F % of Max B22+GL7+ Cells * CD47 Negative Naive T cells T cells (Naive & infection) T cells (HA-VLP & infection)
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Heat-killed Lactobacillus casei
 Heat-killed Lactobacillus casei confers broad protection against influenza A virus primary infection and develops heterosubtypic immunity against future secondary infection Yu-Jin Jung, Young-Tae Lee,
Heat-killed Lactobacillus casei confers broad protection against influenza A virus primary infection and develops heterosubtypic immunity against future secondary infection Yu-Jin Jung, Young-Tae Lee,
Supporting Information
 Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
SUPPLEMENTARY INFORMATION
 Supplementary Text Results In vitro activation experiments established that treatment with anti-cd3 plus anti- CD28 was sufficient to promote EBI2 transcript on CD4 T cells (Extended Data Fig. 1d, e).
Supplementary Text Results In vitro activation experiments established that treatment with anti-cd3 plus anti- CD28 was sufficient to promote EBI2 transcript on CD4 T cells (Extended Data Fig. 1d, e).
Supplementary Information:
 Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supporting Information
 Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).
 Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).") Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Adaptive Immunity: Humoral Immune Responses
 MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
SUPPLEMENTARY INFORMATION
 Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES
 CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
Antigen Presentation and T Lymphocyte Activation. Abul K. Abbas UCSF. FOCiS
 1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit
 Hemagglutinin ELISA kit") 2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
Mouse Anti-OVA IgM Antibody Assay Kit
 Mouse Anti-OVA IgM Antibody Assay Kit Catalog # 3017 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Ovalbumin (OVA) is a widely used antigen for inducing allergic reactions in experimental
Mouse Anti-OVA IgM Antibody Assay Kit Catalog # 3017 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Ovalbumin (OVA) is a widely used antigen for inducing allergic reactions in experimental
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Interferon γ regulates idiopathic pneumonia syndrome, a. Th17 + CD4 + T-cell-mediated GvH disease
 Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
The Adaptive Immune Response. B-cells
 The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas
 Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas Chapter 04: Antigen Recognition in the Adaptive Immune System Test Bank MULTIPLE CHOICE 1. Most T lymphocytes
Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas Chapter 04: Antigen Recognition in the Adaptive Immune System Test Bank MULTIPLE CHOICE 1. Most T lymphocytes
Supplemental Table I.
 Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Chapter 24 The Immune System
 Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Role of Tyk-2 in Th9 and Th17 cells in allergic asthma
 Supplementary File Role of Tyk-2 in Th9 and Th17 cells in allergic asthma Caroline Übel 1*, Anna Graser 1*, Sonja Koch 1, Ralf J. Rieker 2, Hans A. Lehr 3, Mathias Müller 4 and Susetta Finotto 1** 1 Laboratory
Supplementary File Role of Tyk-2 in Th9 and Th17 cells in allergic asthma Caroline Übel 1*, Anna Graser 1*, Sonja Koch 1, Ralf J. Rieker 2, Hans A. Lehr 3, Mathias Müller 4 and Susetta Finotto 1** 1 Laboratory
General Overview of Immunology. Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center
 General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland Approved for public release; distribution unlimited
 AD (Leave blank) Award Number: W81XWH-07-1-0345 TITLE: Second-Generation Therapeutic DNA Lymphoma Vaccines PRINCIPAL INVESTIGATOR: Larry W. Kwak, M.D., Ph.D. CONTRACTING ORGANIZATION: University of Texas
AD (Leave blank) Award Number: W81XWH-07-1-0345 TITLE: Second-Generation Therapeutic DNA Lymphoma Vaccines PRINCIPAL INVESTIGATOR: Larry W. Kwak, M.D., Ph.D. CONTRACTING ORGANIZATION: University of Texas
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MyD88 Plays an Essential Role in Inducing B Cells Capable of Differentiating into Antibody-Secreting Cells after Vaccination
 JOURNAL OF VIROLOGY, Nov. 2011, p. 11391 11400 Vol. 85, No. 21 0022-538X/11/$12.00 doi:10.1128/jvi.00080-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. MyD88 Plays an Essential
JOURNAL OF VIROLOGY, Nov. 2011, p. 11391 11400 Vol. 85, No. 21 0022-538X/11/$12.00 doi:10.1128/jvi.00080-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. MyD88 Plays an Essential
Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and
 Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and Thy1 in NH cells derived from the lungs of naïve mice.
Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and Thy1 in NH cells derived from the lungs of naïve mice.
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set
 p24 / Capsid Protein p24 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
B220 CD4 CD8. Figure 1. Confocal Image of Sensitized HLN. Representative image of a sensitized HLN
 B220 CD4 CD8 Natarajan et al., unpublished data Figure 1. Confocal Image of Sensitized HLN. Representative image of a sensitized HLN showing B cell follicles and T cell areas. 20 µm thick. Image of magnification
B220 CD4 CD8 Natarajan et al., unpublished data Figure 1. Confocal Image of Sensitized HLN. Representative image of a sensitized HLN showing B cell follicles and T cell areas. 20 µm thick. Image of magnification
Mouse Total IgA Antibody Detection Kit
 Mouse Total IgA Antibody Detection Kit Catalog # 3019 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The total IgA levels in specimens are often determined in mouse disease models involving
Mouse Total IgA Antibody Detection Kit Catalog # 3019 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The total IgA levels in specimens are often determined in mouse disease models involving
W/T Itgam -/- F4/80 CD115. F4/80 hi CD115 + F4/80 + CD115 +
 F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
Adaptive Immunity: Specific Defenses of the Host
 17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
B cell activation and antibody production. Abul K. Abbas UCSF
 1 B cell activation and antibody production Abul K. Abbas UCSF 2 Lecture outline B cell activation; the role of helper T cells in antibody production Therapeutic targeting of B cells 3 Principles of humoral
1 B cell activation and antibody production Abul K. Abbas UCSF 2 Lecture outline B cell activation; the role of helper T cells in antibody production Therapeutic targeting of B cells 3 Principles of humoral
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic
 Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
Supplementary Figure 1. NAFL enhanced immunity of other vaccines (a) An over-the-counter, hand-held non-ablative fractional laser (NAFL).
 An over-the-counter, hand-held non-ablative fractional laser (NAFL).") Supplementary Figure 1. NAFL enhanced immunity of other vaccines (a) An over-the-counter, hand-held non-ablative fractional laser (NAFL). (b) Depiction of a MTZ array generated by NAFL. (c-e) IgG production
Supplementary Figure 1. NAFL enhanced immunity of other vaccines (a) An over-the-counter, hand-held non-ablative fractional laser (NAFL). (b) Depiction of a MTZ array generated by NAFL. (c-e) IgG production
TCR, MHC and coreceptors
 Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Studying Repeated Immunization in an Animal Model. Kanta Subbarao Laboratory of Infectious Diseases, NIAID
 Studying Repeated Immunization in an Animal Model Kanta Subbarao Laboratory of Infectious Diseases, NIAID Animal models in Influenza Research Commonly used Mice Ferrets Guinea pigs Non human primates Less
Studying Repeated Immunization in an Animal Model Kanta Subbarao Laboratory of Infectious Diseases, NIAID Animal models in Influenza Research Commonly used Mice Ferrets Guinea pigs Non human primates Less
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells
 Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
The Immune System. These are classified as the Innate and Adaptive Immune Responses. Innate Immunity
 The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
T Cell Effector Mechanisms I: B cell Help & DTH
 T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
Immune mechanisms of influenza vaccine adjuvants and respiratory syncytial viral vaccines
 Georgia State University ScholarWorks @ Georgia State University Biology Dissertations Department of Biology 12-15-2016 Immune mechanisms of influenza vaccine adjuvants and respiratory syncytial viral
Georgia State University ScholarWorks @ Georgia State University Biology Dissertations Department of Biology 12-15-2016 Immune mechanisms of influenza vaccine adjuvants and respiratory syncytial viral
B6/COLODR/SPL/11C/83/LAP/#2.006 B6/COLODR/SPL/11C/86/LAP/#2.016 CD11C B6/COLODR/SPL/11C/80/LAP/#2.011 CD11C
 CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
Sipper BK Experimental Animal Co. (Shanghai, China) and bred in a specific. pathogen-free environment. The animal study protocol was approved by the
 and bred in a specific. pathogen-free environment. The animal study protocol was approved by the") Supplementary information, Data S1 Materials and Methods Mice, Ad vectors and reagents Female C57BL/6 mice, 8-10 weeks of age, were purchased from Joint Ventures Sipper BK Experimental Animal Co. (Shanghai,
Supplementary information, Data S1 Materials and Methods Mice, Ad vectors and reagents Female C57BL/6 mice, 8-10 weeks of age, were purchased from Joint Ventures Sipper BK Experimental Animal Co. (Shanghai,
Zheng, BJ; Du, LY; Zhao, GY; Lin, YP; Sui, HY; Chan, C; Ma, S; Guan, Y; Yuen, KY. Citation Hong Kong Medical Journal, 2008, v. 14 suppl. 4, p.
 Title Studies of SARS virus vaccines Author(s) Zheng, BJ; Du, LY; Zhao, GY; Lin, YP; Sui, HY; Chan, C; Ma, S; Guan, Y; Yuen, KY Citation Hong Kong Medical Journal, 2008, v. 14 suppl. 4, p. 39-43 Issued
Title Studies of SARS virus vaccines Author(s) Zheng, BJ; Du, LY; Zhao, GY; Lin, YP; Sui, HY; Chan, C; Ma, S; Guan, Y; Yuen, KY Citation Hong Kong Medical Journal, 2008, v. 14 suppl. 4, p. 39-43 Issued
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs
 Western blot analysis of p38 isoform expression in BM cells, immature DCs") Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs (idcs) and mature DCs (mdcs). A myeloma cell line expressing
Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs (idcs) and mature DCs (mdcs). A myeloma cell line expressing
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented
 Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set
 gp120 / Glycoprotein 120 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
Adaptive Immunity. Jeffrey K. Actor, Ph.D. MSB 2.214,
 Adaptive Immunity Jeffrey K. Actor, Ph.D. MSB 2.214, 500-5344 Lecture Objectives: Understand role of various molecules including cytokines, chemokines, costimulatory and adhesion molecules in the development
Adaptive Immunity Jeffrey K. Actor, Ph.D. MSB 2.214, 500-5344 Lecture Objectives: Understand role of various molecules including cytokines, chemokines, costimulatory and adhesion molecules in the development
NF-κB p65 (Phospho-Thr254)
") Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Determinants of Immunogenicity and Tolerance. Abul K. Abbas, MD Department of Pathology University of California San Francisco
 Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity
 Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Influenza B Hemagglutinin / HA ELISA Pair Set
 Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Principles of Adaptive Immunity
 Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia
 Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
The Immune System is the Third Line of Defense Against Infection. Components of Human Immune System
 Chapter 17: Specific Host Defenses: The Immune Response The Immune Response Immunity: Free from burden. Ability of an organism to recognize and defend itself against specific pathogens or antigens. Immune
Chapter 17: Specific Host Defenses: The Immune Response The Immune Response Immunity: Free from burden. Ability of an organism to recognize and defend itself against specific pathogens or antigens. Immune
Blocking Interhost Transmission of Influenza Virus by Vaccination in the Guinea Pig Model
 JOURNAL OF VIROLOGY, Apr. 2009, p. 2803 2818 Vol. 83, No. 7 0022-538X/09/$08.00 0 doi:10.1128/jvi.02424-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Blocking Interhost Transmission
JOURNAL OF VIROLOGY, Apr. 2009, p. 2803 2818 Vol. 83, No. 7 0022-538X/09/$08.00 0 doi:10.1128/jvi.02424-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Blocking Interhost Transmission
H5N1 ( Avian Flu ) Hemagglutinin ELISA Pair Set
 Hemagglutinin ELISA Pair Set") H5N1 ( Avian Flu ) Hemagglutinin ELISA Pair Set Catalog Number : SEK002 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
H5N1 ( Avian Flu ) Hemagglutinin ELISA Pair Set Catalog Number : SEK002 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature775 4 O.D. (595-655) 3 1 -ζ no antibody isotype ctrl Plated Soluble 1F6 397 7H11 Supplementary Figure 1 Soluble and plated anti- Abs induce -! signalling. B3Z cells stably expressing!
doi: 1.138/nature775 4 O.D. (595-655) 3 1 -ζ no antibody isotype ctrl Plated Soluble 1F6 397 7H11 Supplementary Figure 1 Soluble and plated anti- Abs induce -! signalling. B3Z cells stably expressing!
There are 2 major lines of defense: Non-specific (Innate Immunity) and. Specific. (Adaptive Immunity) Photo of macrophage cell
 and. Specific. (Adaptive Immunity) Photo of macrophage cell") There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
Putting it Together. Stephen Canfield Secondary Lymphoid System. Tonsil Anterior Cervical LN s
 Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Supplemental Figure 1
 Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
CHAPTER-VII IMMUNOLOGY R.KAVITHA, M.PHARM, LECTURER, DEPARTMENT OF PHARMACEUTICS, SRM COLLEGE OF PHARMACY, SRM UNIVERSITY, KATTANKULATHUR.
 CHAPTER-VII IMMUNOLOGY R.KAVITHA, M.PHARM, LECTURER, DEPARTMENT OF PHARMACEUTICS, SRM COLLEGE OF PHARMACY, SRM UNIVERSITY, KATTANKULATHUR. The Immune Response Immunity: Free from burden. Ability of an
CHAPTER-VII IMMUNOLOGY R.KAVITHA, M.PHARM, LECTURER, DEPARTMENT OF PHARMACEUTICS, SRM COLLEGE OF PHARMACY, SRM UNIVERSITY, KATTANKULATHUR. The Immune Response Immunity: Free from burden. Ability of an
Microbiology 204: Cellular and Molecular Immunology
 Microbiology 204: Cellular and Molecular Immunology Class meets MWF 1:00-2:30PM (*exceptions: no class Fri Sept 23, Fri Oct 14, Nov 11, or Wed Nov 23) Lectures are open to auditors and will be live-streamed
Microbiology 204: Cellular and Molecular Immunology Class meets MWF 1:00-2:30PM (*exceptions: no class Fri Sept 23, Fri Oct 14, Nov 11, or Wed Nov 23) Lectures are open to auditors and will be live-streamed
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Chapter 3, Part A (Pages 37-45): Leukocyte Migration into Tissues
: Leukocyte Migration into Tissues") Allergy and Immunology Review Corner: Chapter 3, Part A (pages 37-45) of Cellular and Molecular Immunology (Seventh Edition), by Abul K. Abbas, Andrew H. Lichtman and Shiv Pillai. Chapter 3, Part A (Pages
Allergy and Immunology Review Corner: Chapter 3, Part A (pages 37-45) of Cellular and Molecular Immunology (Seventh Edition), by Abul K. Abbas, Andrew H. Lichtman and Shiv Pillai. Chapter 3, Part A (Pages
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
Nature Medicine: doi: /nm.3922
 Title: Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells Authors: Il-Kyu Kim, Byung-Seok Kim, Choong-Hyun
Title: Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells Authors: Il-Kyu Kim, Byung-Seok Kim, Choong-Hyun
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL IL-1 signaling modulates activation of STAT transcription factors to antagonize retinoic acid signaling and control the T H 17 cell it reg cell balance Rajatava Basu 1,5, Sarah K.
SUPPLEMENTARY MATERIAL IL-1 signaling modulates activation of STAT transcription factors to antagonize retinoic acid signaling and control the T H 17 cell it reg cell balance Rajatava Basu 1,5, Sarah K.
Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces
 Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces Department of Biologic & Materials Sciences School of Dentistry University of Michigan Ann Arbor, Michigan 48109-1078 1 Image quality
Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces Department of Biologic & Materials Sciences School of Dentistry University of Michigan Ann Arbor, Michigan 48109-1078 1 Image quality
Supporting Information Table of Contents
 Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder I Nakaya, Kousik
 SUPPLEMENTARY FIGURES 1-19 T H 2 response to cysteine-proteases requires dendritic cell-basophil cooperation via ROS mediated signaling Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder
SUPPLEMENTARY FIGURES 1-19 T H 2 response to cysteine-proteases requires dendritic cell-basophil cooperation via ROS mediated signaling Hua Tang, Weiping Cao, Sudhir Pai Kasturi, Rajesh Ravindran, Helder
Supplemental Figure 1. Cell-bound Cetuximab reduces EGFR staining intensity. Blood
 Antibody-mediated depletion of CD19-CAR T cells Supplemental 1 Supplemental Materials Supplemental Figure 1. Supplemental Figure 1. Cell-bound Cetuximab reduces EGFR staining intensity. Blood cells were
Antibody-mediated depletion of CD19-CAR T cells Supplemental 1 Supplemental Materials Supplemental Figure 1. Supplemental Figure 1. Cell-bound Cetuximab reduces EGFR staining intensity. Blood cells were
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured
 Naïve CD4 + T cells were cultured") Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Supplemental Figure 1. Signature gene expression in in vitro differentiated Th0, Th1, Th2, Th17 and Treg cells. (A) Naïve CD4 + T cells were cultured under Th0, Th1, Th2, Th17, and Treg conditions. mrna
Blood and Immune system Acquired Immunity
 Blood and Immune system Acquired Immunity Immunity Acquired (Adaptive) Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated
Blood and Immune system Acquired Immunity Immunity Acquired (Adaptive) Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated
Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK40103 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK40103 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Foundations in Microbiology
 Foundations in Microbiology Fifth Edition Talaro Chapter 15 The Acquisition of Specific Immunity and Its Applications Chapter 15 2 Chapter Overview 1. Development of the Dual Lymphocyte System 2. Entrance
Foundations in Microbiology Fifth Edition Talaro Chapter 15 The Acquisition of Specific Immunity and Its Applications Chapter 15 2 Chapter Overview 1. Development of the Dual Lymphocyte System 2. Entrance
T cell maturation. T-cell Maturation. What allows T cell maturation?
 T-cell Maturation What allows T cell maturation? Direct contact with thymic epithelial cells Influence of thymic hormones Growth factors (cytokines, CSF) T cell maturation T cell progenitor DN DP SP 2ry
T-cell Maturation What allows T cell maturation? Direct contact with thymic epithelial cells Influence of thymic hormones Growth factors (cytokines, CSF) T cell maturation T cell progenitor DN DP SP 2ry