Evidence for a Serum Factor That Initiates the Re-calcification of Demineralized Bone* S
|
|
|
- August Barnett
- 5 years ago
- Views:
Transcription
1 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 18, Issue of April 30, pp , by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Evidence for a Serum Factor That Initiates the Re-calcification of Demineralized Bone* S Received for publication, July 21, 2003, and in revised form, January 29, 2004 Published, JBC Papers in Press, February 20, 2004, DOI /jbc.M Paul A. Price, Helen H. June, Nicholas J. Hamlin, and Matthew K. Williamson From the Division of Biology, University of California, San Diego, La Jolla, California The present studies show for the first time that demineralized bone re-calcifies rapidly when incubated at 37 C in rat serum: re-calcification can be demonstrated by Alizarin Red and von Kossa stains, by depletion of serum calcium, and by uptake of calcium and phosphate by bone matrix. Re-calcification is specific for the type I collagen matrix structures that were calcified in the original bone, with no evidence for calcification in periosteum or cartilage. Re-calcification ceases when the amount of calcium and phosphate introduced into the matrix is comparable to that present in the original bone prior to demineralization, and the re-calcified bone is palpably hard. Re-calcified bone mineral is comparable to the original bone mineral in calcium to phosphate ratio and in Fourier transform infrared and x-ray diffraction spectra. The serum activity responsible for re-calcification is sufficiently potent that the addition of only 1.5% serum to Dulbecco s modified Eagle s medium causes bone re-calcification. This putative serum calcification factor has an apparent molecular mass of kda and is inactivated by trypsin or chymotrypsin. The serum calcification factor must act on bone for 12 h before re-calcification can be detected by Alizarin Red or von Kossa staining and before the subsequent growth of calcification will occur in the absence of serum. The speed, matrix-type specificity, and extent of the seruminduced re-calcification of demineralized bone suggest that the serum calcification factor identified in these studies may participate in the normal calcification of bone. This study is a continuation of our investigations into the mechanisms that initiate the normal calcification of bone and the abnormal calcification of arteries and into the mechanisms that inhibit these calcifications. We recently proposed the hypothesis that artery calcification is linked to bone resorption (1, 2) to explain the association between increased bone resorption and increased artery calcification that has been seen in the vitamin D-treated rat (1), in the osteoprotegerin-deficient mouse (3), and in patients with postmenopausal osteoporosis (see Ref. 2 for references). One prediction of the hypothesis that artery calcification is linked to bone resorption is that inhibitors of bone resorption should inhibit artery calcification (2). In previous * This work was supported in part by National Institutes of Health Grant HL The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. S The on-line version of this article (available at contains Supplemental Figs. A D. To whom correspondence should be addressed: Division of Biological Sciences, 0368, University of California, San Diego, La Jolla, CA Tel.: ; Fax: ; pprice@ ucsd.edu. This paper is available on line at studies we tested this prediction using three different types of bone resorption inhibitors, each with an entirely different mode of action on the osteoclast, the amino bisphosphonates alendronate and ibandronate (2), the cytokine osteoprotegerin (4), and the V-H -ATPase inhibitor SB (5). Each bone resorption inhibitor potently inhibited artery calcification. Ibandronate, osteoprotegerin, and SB are each highly specific inhibitors of the osteoclast at the concentrations used in these studies and have no known actions on vascular cells. Their ability to potently inhibit vascular calcification is therefore hard to reconcile with the hypothesis that the process is initiated by the action of vascular cells in the artery wall. We have therefore advanced an alternative hypothesis to explain the relation between bone resorption and artery calcification, the hypothesis that a causative agent for artery calcification arises in bone metabolism, travels in blood, and then induces calcification in the artery wall. If the link between bone resorption and artery calcification is blood, then it follows that blood should contain the causative agent for artery calcification. However, because this putative-calcifying agent apparently arises in bone-metabolic processes, it seems likely that its normal physiological function is to participate in the calcification of bone, not arteries, and that artery calcification itself occurs only when there is a breakdown in the normal mechanisms that inhibit calcification in the artery wall, such as matrix Gla protein (6, 7). The present experiments were carried out to test the hypothesis that blood contains an agent that participates in the calcification of bone. The results of these experiments demonstrate that serum indeed contains a potent agent (or agents 1 ) that initiates the re-calcification of demineralized bone, and additional experiments were accordingly undertaken to characterize this serum agent. EXPERIMENTAL PROCEDURES Materials 42-day-old, 22-day-old (weanling), and newborn albino rats (Sprague-Dawley derived) were purchased from Charles River Laboratories (Wilmington, MD). One calcification medium, termed DMEM, 2 was prepared by supplementing each 500-ml volume of DMEM (Invitrogen) with 5 ml of penicillin-streptomycin (Invitrogen), 1 ml of 10% sodium azide, and 1.1 ml of 0.5 M sodium phosphate buffer, ph 7.4 (final medium P i concentration 2mM). The other calcification medium was a physiological buffer that contained 30 mm NaHCO 3, 120 mm NaCl, 1 mm CaCl 2,3mM sodium phosphate, ph 7.4, and 0.02% azide. Sodium heparin and Alizarin Red S were purchased from Sigma. Rats were killed by exsanguination while under isoflurane anesthetic (40- and 22-day-old rats) or by decapitation (newborn rats). For serum preparation, blood was allowed to clot for 30 min at room temperature and centrifuged at 1,400 g for 10 min in a clinical centrifuge. 1 The use of the singular terms factor and agent is not meant to imply that there exists only one molecule that is involved in the seruminitiated re-calcification of bone. 2 The abbreviations used are: DMEM, Dulbecco s modified Eagle s medium; FTIR, Fourier transform infrared.
2 19170 Re-calcification of Bone in Serum For plasma, blood was drawn into a heparin-containing syringe to achieve a final heparin concentration of 75 g/ml, and plasma was collected by centrifugation at 1,400 g for 10 min. Serum was stored in 1-ml aliquots at 70 C until use, whereas plasma was used immediately after collection. Tibias and calvaria were dissected from newborn rats, demineralized in 0.5 M EDTA, ph 7.5, for 72 h at room temperature using a 300-fold molar excess of EDTA to mineral calcium, washed exhaustively with ultra pure water to remove all traces of EDTA, and stored at 20 C until use. Tendons were obtained from the tails of 40-day-old rats and extracted with EDTA as described above and dried. Dry tendon segments weighing 3 mg were re-hydrated by overnight equilibration in ultra pure water before use. The University of California, San Diego, Animal Subjects Committee approved all animal experiments. Re-calcification Procedures Experiments to examine matrix calcification in rat serum were carried out with 35-mm culture dishes (Falcon 3001) in a humidified incubator at 37 C and 7.5% CO 2 using 2 ml of rat serum, heparin plasma, or physiological buffer, each containing 0.02% sodium azide to prevent bacterial growth. After equilibration with 7.5% CO 2 at 37 C, serum maintained a stable ph value of 7.4 throughout the duration of the 6-day incubation, and the physiological buffer maintained a ph of A single wet tibia or calvarium, or a rehydrated tendon (3 mg of dry weight) was added to 2 ml of rat serum, rat plasma, or physiological buffer and placed in the incubator for 6 days. To assess the progress of matrix calcification, 50- l aliquots were removed at the start of the incubation and at each 24 h thereafter and stored frozen until analysis to determine media calcium levels. The effect of adding different amounts of rat serum on the calcification of tibia and tendon was examined using 10 ml of DMEM in 100-mm culture dishes (Falcon 3803) in a humidified incubator at 37 C and 5% CO 2. A single demineralized tibia or a 3-mg tendon sample (dry weight) was added to each 100-mm plate. The effect of preincubation in serum on the subsequent calcification of demineralized tibias in DMEM was examined by first preincubating each tibia in 2 ml of rat serum at 37 C and 7.5% CO 2, then rinsing the tibia for 15 min each in two 10-ml aliquots of DMEM, and finally incubating the washed tibia in 10 ml of DMEM for 5 days at 37 C and 5% CO 2. Finally, early stages in the serum-induced calcification of demineralized tibia were examined by incubating each tibia in 2 ml of rat serum for the desired time; tibias were then added directly to Alizarin Red stain or fixed in absolute ethanol, embedded in paraffin, sectioned, and stained with von Kossa. Biochemical Analyses The procedures used for Alizarin Red and von Kossa staining have been described (8). For quantitative assessment of calcification, matrices were extracted for 24 h at room temperature with 1 ml of 0.15 M HCl. Calcium levels in serum and in the acid extract of matrices were determined colorimetrically using cresolphthalein complexone (Sigma), and phosphate levels were determined as described (8). To characterize the molecular mass of the serum calcification factor, 1 ml of serum from a 40-day-old rat was fractionated over a 25-ml column of Sephacryl S-100HR equilibrated at room temperature with DMEM using a buffer reservoir that was continuously flushed with 5%CO 2 to maintain a ph of 7.4. The absorbance at 280 nm was determined for each 1-ml fraction, small aliquots of each fraction were set aside for electrophoresis, and each fraction was then placed into separate wells of a 24-well tissue culture plate. A single demineralized newborn rat tibia was then added to each well and incubated for 6 days at 37 C and 5% CO 2. To determine the apparent molecular weight of serum proteins that co-eluted with the serum calcification activity in this experiment, 2- l aliquots of rat serum and of fractions 9, 11, and 14 (Fig. 5) were electrophoresed on a 4 12% SDS-polyacrylamide gel and stained with Coomassie Brilliant Blue. To determine whether the serum factor that initiates bone re-calcification is inactivated by proteases, four 10-ml samples of DMEM were prepared that contained: no serum, 5% rat serum, 5% serum and 1 mg of trypsin, and 5% serum and 1 mg of chymotrypsin. Each solution was preincubated for 24 h at 37 C and 5% CO 2. A single demineralized newborn rat tibia was then added, and the solutions were incubated for 6 days at 37 C. To assess the effect of preincubation with proteases on the integrity of serum proteins, aliquots of each medium were removed after the 24-h preincubation step, electrophoresed on a 4 12% SDSpolyacrylamide gel, and stained with Coomassie Brilliant Blue (load: no protease, 10 l medium; protease, 50 l). Fourier transform infrared spectroscopy (FTIR) was used to analyze the mineral from tibias that were re-calcified by incubation in rat serum or in DMEM containing 5% rat serum and from newborn rat tibia. Ten demineralized tibias were each re-calcified in 2 ml of rat serum for 6 days at 37 C and 7.5% CO 2. Ten demineralized tibias were also each re-calcified by incubation in 10 ml of DMEM containing 5% rat serum for 6 days at 37 C and 5% CO 2. The diaphyseal regions were then obtained by dissection from each re-calcified tibia and from 10 nondemineralized newborn tibias. The resulting 30 diaphyseal regions were dried, pooled according to the three treatment groups, and ground with an agate mortar. The samples were analyzed as KBr pellets (3% w/w), and 256 scans were acquired at 4-cm 1 resolution using a Nicolet Magna IR 550 FTIR spectrometer. The Re-calcification of Weanling Rat Tibias Tibias were dissected from thirty 22-day-old rats and cut to remove the bone ends, and the midshaft 1 cm of cortical bone was freed of marrow and cancellous bone. Ten tibias were saved for later analyses, and 20 were demineralized for 72 h with 0.5 M EDTA, ph 7.5. Ten demineralized tibias were each separately incubated for 3 days at 37 C in 2 ml of rat serum, washed twice for 15 min each with 20 ml of 25 mm HEPES buffer, ph 7.4, containing 1 mm calcium, 3 mm phosphate, 100 mm NaCl, 30 mm NaHCO 3, and 150 g/ml fetuin (Sigma), and then incubated in 20 ml of this buffer in a humidified incubator at 37 C and 5% CO 2. Ten demineralized tibias were placed directly into 20 ml of this buffer without serum pre-treatment. The incubation buffer was exchanged for fresh buffer every other day for 30 days. At the end of this incubation period, all tibias were dried and weighed (Fig. 11), and x-rays of each tibia were obtained using a Hewlett-Packard Model 4380N Faxitron x-ray machine. Five tibias from each group were then separately demineralized in 1 ml of 150 mm HCl, and calcium and phosphate levels were determined in each acid extract. The remaining five tibias from each group were pooled according to treatment group and ground with an agate mortar. The resulting powders were first analyzed by XRD using a Scintag SDF 2000 x-ray diffractometer, and a portion of this powder was then analyzed by FTIR. RESULTS The Calcification of Demineralized Tibias by Incubation in Rat Serum or Plasma The initial test of the hypothesis that blood contains an agent that calcifies bone was carried out using newborn rat tibias that had been first freed of endogenous mineral by 72 h extraction with 0.5 M EDTA, ph 7.5, a procedure that also killed all cells originally found in the bone. The demineralized tibias were placed in 2 ml of serum from 40-day-old rats and incubated for 6 days at 37 C. Fig. 1 shows that demineralized tibias that have been incubated for 6 days in rat serum stain intensely for calcification with Alizarin Red and that the pattern of staining matches that seen in the original tibia prior to demineralization, with intense staining for calcification in the bone in the midshaft region and absence of staining in the cartilaginous tissue at either end of the tibia. Fig. 2 shows that demineralized tibias that have been incubated for 6 days in rat serum also stain for calcification by von Kossa and that the microscopic pattern of calcification is identical to that seen in tibias prior to demineralization, with intense von Kossa staining for calcification in the type I collagen matrix of bone and no evidence for calcification in either the periosteum or in cartilaginous tissue at the bone ends. Examination of the same sections at higher magnification showed that the calcification induced by incubation of demineralized tibias in rat serum is discontinuous, with numerous discrete calcification foci that could reflect individual nucleation events (figure not shown). Identical von Kossa and Alizarin staining patterns were obtained using newborn tibias that had been demineralized in 150 mm HCl for 72 h, or using EDTA-demineralized tibias that had been subsequently extracted with acetone followed by 24-h incubation in 6 M guanidine HCl (data not shown). These results demonstrate that demineralization conditions do not affect the matrix specificity or extent of the serum-induced re-calcification of bone. The possible re-calcification of demineralized tibias during incubation in serum was further assessed by chemical analysis for calcium and phosphate. Table I shows that incubation in rat serum restored about 60% of the calcium and phosphate found in tibias prior to demineralization and that the molar calcium

.")
3 Re-calcification of Bone in Serum FIG. 1. Calcification of demineralized rat tibia, calvaria, and tendon by incubation in rat serum: Alizarin Red staining of whole tissue. Newborn rat tibia and calvaria, and tail tendons from 40-day-old rats were extracted for 72 h with 0.5 M EDTA, ph 7.5, rinsed thoroughly with water, and placed in a 35-mm culture dish containing 2 ml of rat serum or 2 ml of a physiological buffer (1 mm Ca, 3mM PO 4, 0.12 M NaCl, 30 mm NaHCO 3, ph 7.4). Tissues were then placed for 6 days in a humidified incubator at 7.5% CO 2 and 37 C (see Experimental Procedures ). The images show the Alizarin Red staining for calcification in newborn rat tibia, calvaria, and tendon prior to EDTA extraction and in EDTA-extracted matrices after incubation for 6 days in the indicated media. FIG. 2. Calcification of demineralized rat tibia, calvaria, and tendon by incubation in rat serum: von Kossa staining of tissue sections. Demineralized rat tibia, calvaria, and tendon were incubated for 6 days in 2 ml of rat serum or in 2 ml of a physiological buffer (1 mm Ca, 3 mm PO 4, 0.12 M NaCl, 30 mm NaHCO 3, ph 7.4) as described in the Fig. 1 legend and then fixed for 24 h in formalin. 5- m-thick sections were stained by von Kossa. to phosphate ratio in re-calcified tibias (1.54) is comparable to the molar ratio in tibias prior to demineralization (1.51). Fig. 3 shows that the re-calcification of bone is accompanied by a 1 mm decline in serum calcium levels over the 6-day incubation period, with most of the decline occurring between the second and third days of incubation. The amount of calcium taken up by the tibia during the incubation (1.94 mol; Table I) accounted for the 2 mol of calcium removed from serum during the incubation (Fig. 3: 1 mm decline in 2 ml volume 2 mol). To determine whether the serum-induced re-calcification of demineralized bone might be a spontaneous process at the calcium and phosphate levels found in rat serum, demineralized tibias were incubated for 6 days in a physiological buffer that contained the same ionic calcium, phosphate, carbonate, and ph as rat serum. This control experiment showed that incubation in physiological buffer did not result in detectable staining for calcification with Alizarin Red (Fig. 1) or von Kossa (Fig. 2) and did not cause an uptake of calcium and phosphate by the bone matrix or a significant decline in medium calcium (data not shown). 3 These results indicate that the calcification of demineralized rat bone during incubation in rat serum is not a spontaneous process driven solely by the ionic calcium, phosphate, temperature, and ph conditions of blood, but rather a process driven by a calcification agent found in rat blood. 3 The physiological buffer did, however, sustain the growth of added hydroxyapatite crystals (data not shown). The failure of demineralized tibia to re-calcify in this buffer therefore shows that the EDTA treatment procedures used to decalcify bone removed all mineral from the matrix of the tibia.
4 19172 Re-calcification of Bone in Serum TABLE I Analysis for calcium and phosphate in tibia, calvaria, and tendon samples after incubation for 6 days in rat serum Demineralized newborn rat calvaria or tibia, or 3 mg of rat tail tendon, were separately incubated for 6dat37 C in 2 ml of rat serum as described in the legends to Figs Tissues were demineralized in acid, and the acid extracts were analyzed for calcium and phosphate as described under Experimental Procedures. The results show the total amount of calcium and phosphate recovered from a single tibia or calvarium, a 1/8 portion of calvarium, or from 3 mg of tail tendon, after the indicated treatments. Tissue Calcium PO 4 mol Tibia Before demineralization After demineralization 0.1 a 0.02 b After incubation in serum Calvaria Before demineralization After demineralization 0.1 a 0.02 b After incubation in serum Calvaria (1/8 portion) Before demineralization After demineralization 0.1 a 0.02 b After incubation in serum Tendon Before incubation in serum 0.1 a 0.02 b After incubation in serum a Detection limit, 0.1 mol of calcium. b Detection limit, 0.02 mol of phosphate. FIG. 3.Calcification of demineralized rat tibia, calvaria, and tendon by incubation in rat serum: changes in serum calcium. Demineralized rat tibia, calvaria, and tendon were incubated for 6 days in 2 ml of rat serum as described in the Fig. 1 legend, and samples of medium were removed at the indicated times and analyzed to determine the levels of calcium. Serum calcium in sample incubated: E, without any tissue; f, with tibia;, with tendon; Œ, with calvarium. Additional experiments were carried out to determine whether the re-calcification activity of rat serum is affected by blood coagulation. In this experiment, serum and heparin plasma samples were obtained from the same rats and tested for their ability to cause the re-calcification of demineralized tibias. The results of these experiments showed that incubation of demineralized tibias for 6 days in heparin plasma resulted in Alizarin Red and von Kossa staining for calcification that were identical to the staining seen in demineralized tibias following incubation in serum from the same rat and caused an uptake of calcium and phosphate by tibias that was comparable to that seen in tibias incubated in serum (data not shown). These results demonstrate that the calcification-inducing activity of blood is not significantly affected by blood coagulation. The Calcification of Demineralized Calvarias by Incubation in Rat Serum or Plasma Additional experiments were carried out to determine whether the membranous bone of newborn rat calvarias also calcifies spontaneously in serum. Newborn rat calvarias were first freed of endogenous mineral and cells by 72-h extraction with 0.5 M EDTA, ph 7.5, and then incubated in 2 ml of rat serum at 37 C. Figs. 1 and 2 show that demineralized calvarias that had been incubated for 6 days in rat serum stained intensely for calcification with Alizarin Red and with von Kossa. Comparable staining was seen for demineralized calvarias that had been incubated for 6 days in heparin plasma (data not shown). Table I shows that incubation in rat serum restored about 11% of the calcium and phosphate found in calvarias prior to demineralization, and Fig. 3 shows that the re-calcification of demineralized calvarias is accompanied by a 1.6 mm decline in serum calcium levels over the 6-day incubation period. The amount of serum calcium taken up by calvarias during the incubation (2.6 mol; Table I) is comparable to the 3.2 mol of calcium removed from serum during the incubation. Demineralized calvarias incubated for 6 days in a physiological buffer that contained the same concentration of ionic calcium, phosphate, and carbonate and the same ph as found in rat serum did not stain for calcification (Figs. 1 and 2) and did not cause a decrease in medium calcium or an uptake of calcium and phosphate by the calvarias (data not shown). Table I shows that incubation in serum restored a far lower percentage of the original mineral to calvarias than to tibias. It seemed likely that the lower extent of calvaria re-calcification was attributable to the limited amount of calcium in the 2-ml serum aliquot, which is over 5-fold lower than the calcium content of calvarias prior to demineralization. To test this hypothesis, the re-calcification experiment was repeated with a 1/8 portion of demineralized calvaria, a portion comparable to a tibia in original mineral content. The extent of re-calcification seen in the 1/8 calvaria sample was comparable to that seen in a newborn tibia (Table I), which shows that there is no significant difference in the extent of re-calcification seen in these two bone matrices when the same amount of bone is used. The Calcification of Tendon by Incubation in Rat Serum or Plasma The above experiments demonstrate that calcification induced by incubation of demineralized tibias and calvarias in rat serum is restricted to those matrix regions that were calcified in the original bone. This pattern of calcification could indicate that agents that remained in the bone structures after demineralization promote the serum-induced calcification. Alternatively, serum-induced calcification could be specific for an extracellular matrix of type I collagen, which is the matrix found in bone, tendon, and other connective tissues. To test the second possibility, tail tendons from 40-day-old rats were incubated in 2 ml of rat serum at 37 C. Figs. 1 and 2 show that tendons that have been incubated for 6 days in rat serum stain intensely for calcification with Alizarin Red and with von Kossa, Table I shows that incubation in rat serum caused a dramatic accumulation of calcium and phosphate in tendons, and Fig. 3 shows that this tendon calcification is accompanied by a corresponding decline in serum calcium levels over the 6-day incubation period. Comparable results were also seen for tendons that had been incubated for 6 days in heparin plasma (data not shown). Tendons incubated for 6 days in a physiological buffer that contained the same concentration of ionic calcium, phosphate, and carbonate and the same ph as found in rat serum did not stain for calcification (Figs. 1 and 2) and did not cause a decrease in medium calcium or an uptake of calcium and phosphate by the tendon (data not shown). The Calcification of Tendons and Demineralized Tibias by Incubation in DMEM Containing Different Amounts of Rat Serum The potency of the serum factor that initiates the re-calcification of demineralized bone was assessed by determining whether demineralized rat tibias would calcify when

5 Re-calcification of Bone in Serum FIG. 4.Calcification of demineralized rat tibia and tendon by incubation in DMEM containing different amounts of rat serum: Alizarin Red staining of whole tissue. Rat tibia and tail tendons were extracted with 0.5 M EDTA, ph 7.5, rinsed with water, and placed in 100-mm culture dishes containing 10 ml of DMEM plus the indicated amount of rat serum. Tissues were incubated for 6 days in a humidified incubator at 37 C and then stained with Alizarin Red (see Experimental Procedures ). incubated in DMEM cell culture medium containing different amounts of rat serum. Fig. 4 shows that demineralized tibias that have been incubated for 6 days in DMEM containing 1.5 or 15% rat serum stain intensely for calcification with Alizarin Red and that the pattern of staining matches that seen in the original tibia prior to demineralization (cf. Fig. 1). Table II shows that demineralized tibias incubated for 6 days in DMEM containing 15% or 1.5% rat serum have dramatically increased levels of calcium and phosphate compared with tibias incubated in DMEM alone, with a 3-fold greater accumulation of calcium and phosphate at 15% serum than at 1.5% serum. Demineralized tibias that have been incubated for 6 days in DMEM containing 15% or 1.5% rat serum also stained for calcification by von Kossa; the microscopic pattern of this calcification was discontinuous, with numerous discrete calcification foci scattered in the extracellular bone matrix (not shown). The number of these foci was markedly lower for tibias recalcified in DMEM with 1.5% serum. Demineralized tibias that had been incubated for 6 days in DMEM alone, or in DMEM containing 0.15% rat serum, did not stain for calcification with Alizarin Red (Fig. 4) or von Kossa (not shown) and did not have a significant uptake of calcium and phosphate (Table II). This experiment has been repeated using fresh and frozen rat serum, and fresh and frozen rat heparin plasma, and in each instance demineralized tibias incubated for 6 days in DMEM containing 1.5% serum or plasma became calcified, whereas tibias incubated in DMEM containing 0.15% serum or plasma did not (not shown). Similar methods were used to assess the potency of the serum factor that initiates calcification of tendon. Tendons that were incubated for 6 days in DMEM containing 15% or 1.5% rat serum stained intensely with Alizarin Red (Fig. 4) and with von Kossa (not shown). Tendons incubated for 6 days in DMEM containing 0.15% serum also stained with Alizarin Red, but the staining was typically restricted to small regions of the tissue. Table II shows that tendons incubated for 6 days in DMEM containing 15%, 1.5%, or 0.15% rat serum have increased levels of calcium and phosphate compared with tendons prior to incubation. Tendons that were incubated for 6 days in DMEM in the absence of serum did not stain for calcification with Alizarin Red (Fig. 4) or with von Kossa (not shown) and did not have a significant uptake of calcium and phosphate (Table II). Evidence That the Serum Factor That Initiates Bone Recalcification Is Macromolecular The apparent size of the serum factor that initiates bone re-calcification was examined by gel filtration over a 25-ml column of Sephacryl S100 HR. DMEM was chosen as the equilibration buffer for this column, because the studies described above (cf. Fig. 4) showed that TABLE II Analysis for calcium and phosphate in tibia and tendon samples after incubation for 6 days in DMEM containing rat serum Demineralized newborn rat tibia or 3 mg of rat tail tendon were separately incubated for 6 d at 37 C in 10 ml of DMEM containing the indicated amount of rat serum, as described in the legend to Fig. 4. Tissues were then demineralized in acid, and the acid extracts were analyzed for calcium and phosphate as described under Experimental Procedures. The results show the total amount of calcium and phosphate recovered from a single tibia or from 3 mg of tail tendon, after the indicated treatments. Tissue Calcium PO 4 mol Tibia Before demineralization After demineralization 0.1 a 0.02 b After incubation in media b After incubation in 0.15% serum in media After incubation in 1.5% serum in media After incubation in 15% serum in media Tendon Before incubation in media 0.1 a 0.02 b After incubation in media b After incubation in 0.15% serum in media After incubation in 1.5% serum in media After incubation in 15% serum in media a Detection limit, 0.1 mol of calcium. b Detection limit, 0.02 mol of phosphate. incubation for 6 days in DMEM alone will not re-calcify demineralized bone but will do so if as little as 1.5% serum is added to it. The use of DMEM as the column buffer allowed the presence of calcification activity in effluent fractions to be assessed by simply incubating a single demineralized tibia in each 1 ml fraction volume for 6 days at 37 C. Fig. 5 shows the 280-nm absorbance profile obtained when 1 ml of rat serum was fractionated by gel filtration over the 25-ml column of Sephacryl S100HR, and Fig. 6 shows the effects of incubating single demineralized newborn rat tibias in the indicated 1-ml fractions from this column for 6 days at 37 C. As can be seen, demineralized tibias incubated for 6 days in fractions 11 and 12 had extensive Alizarin Red staining for calcification, whereas demineralized tibias incubated for 6 days in fractions 9, 10, 13, and 14 (Fig. 6) or in the other fractions examined (7, 8, 15 26) had no Alizarin Red staining. Chemical analyses further showed that there was a dramatic accumulation of calcium and phosphate in demineralized tibias incubated for 6 days in fractions 11 and 12 (Fig. 6), whereas there was a modest increase in calcium and no increase in phosphate for demineralized tibias incubated for 6 days in the other fractions tested (Fig. 6: fractions 9, 10, 13, and 14; not shown,

6 19174 Re-calcification of Bone in Serum FIG. 5. Gel filtration evidence that the serum calcification factor that initiates bone re-calcification is a macromolecule: Protein analyses. A 1-ml sample of serum from a 40-day-old rat was applied to a 25-ml column of Sephacryl S-100 HR equilibrated with DMEM culture medium at room temperature. The graph shows the absorbance at 280 nm for each 1 ml effluent fraction. Inset, 2- l aliquots of rat serum and of fractions 9, 11, and 14 were electrophoresed on a 4 12% SDS-polyacrylamide gel and stained with Coomassie Brilliant Blue. FIG. 6. Gel filtration evidence that the serum calcification factor that initiates bone re-calcification is a macromolecule: Assays for calcification activity. Each 1-ml fraction from the Sephacryl S-100HR fractionation described in Fig. 5 was placed into a well of a 24-well tissue culture plate. A single demineralized newborn rat tibia was added to each fraction and incubated for 6 days at 37 C and 5% CO 2. Tibias were stained with Alizarin Red, photographed, and then analyzed for calcium and phosphate (see Experimental Procedures ). The data show the Alizarin Red staining and the amount of calcium and phosphate recovered from a tibia after incubation for 6 days in the indicated fraction. A demineralized tibia was incubated for 6 days in 1 ml of DMEM culture medium containing 15% rat serum as a positive control. As in previous studies, tibias incubated in DMEM alone showed no evidence for calcification (not shown). fractions 7, 8, 15 26). An identical, modest increase in calcium was also seen in demineralized tibias incubated in column buffer alone and may reflect the presence of calcium binding macromolecules in demineralized bone matrix. If the increase in calcium and phosphate found in demineralized tibias following incubation in fraction 11 is due to the formation of a calcium phosphate mineral phase, the calculated molar ratio of calcium to phosphate in this mineral phase is about 1.6. This experiment has been repeated several times, and in each instance demineralized tibias incubated in fractions 11 and 12 showed extensive Alizarin Red staining for calcification and comparable uptake in calcium and phosphate, whereas tibias incubated in all other effluent fractions showed no evidence for calcification. Fig. 5 also shows the SDS-polyacrylamide gel electrophoresis of selected fractions from this column. As can be seen, the fraction containing the most calcification activity (fraction 11) is in the approximate elution position of albumin, the major protein of serum, and we therefore estimate the molecular mass of the serum calcification factor to be kda. Because demineralized tibias did not re-calcify when incubated in fractions containing proteins with molecular mass above 150 kda (fractions 9 and 10), the serum calcification factor cannot be greater than 150 kda. Because tibias also did not calcify when incubated in the fractions corresponding to lower molecular mass serum components (fractions 13 26), the serum calcification factor cannot have a low molecular mass. (For reference, NaCl and ionic calcium elute in fractions 24 and 25 of this chromatogram). This gel filtration experiment was repeated to assess the potency of the serum calcification factor activity in each fraction. In this experiment, demineralized tibias were incubated for 6 days in 1 ml of medium that consisted of 0.1 ml of each fraction and 0.9 ml of DMEM. Only tibia incubated in this 1/10

7 Re-calcification of Bone in Serum FIG. 7. Effect of trypsin or chymotrypsin on the serum-induced re-calcification of bone. A, 10 ml of DMEM containing 5% rat serum by volume was preincubated for 24 h at 37 C and 5% CO 2 with 100 g/ml trypsin, 100 g/ml chymotrypsin, or no added protease. A fourth sample of DMEM alone was also prepared. After the 24-h preincubation, a single demineralized newborn rat tibia was added to each sample and incubated for 6 days at 37 C and 5% CO 2. Tibias were stained with Alizarin Red, photographed, and then analyzed for calcium and phosphate. The data show the Alizarin Red staining and the amount of calcium and phosphate recovered from each tibia. B, after the 24-h preincubation step, aliquots of each medium were electrophoresed on a 4 12% SDS-polyacrylamide gel and stained with Coomassie Brilliant Blue (see Experimental Procedures ). dose of fractions 11 and 12 proved to be calcified. This result shows that there is sufficient calcification factor activity in fractions 11 and 12 to calcify at least 20 demineralized tibias. Evidence for the Proteinacious Nature of the Serum Calcification Factor That Initiates Bone Re-calcification Additional experiments were carried out to determine whether the serum calcification factor that initiates bone re-calcification is inactivated by proteases. Trypsin (0.01%) or chymotrypsin (0.01%) were added to 10 ml of DMEM containing 5% rat serum, and the solutions were preincubated for 24 h at 37 C. A single demineralized tibia was then added to each 10-ml volume, and the solutions were incubated at 37 C for another 6 days. As can be seen in Fig. 7A, demineralized tibias incubated in solutions containing 5% serum plus 100 g/ml trypsin or 100 g/ml chymotrypsin did not calcify, whereas tibias incubated in solutions containing 5% serum alone calcified extensively. To assess the effect of preincubation with proteases on the integrity of serum proteins, medium obtained at the end of the 24-h preincubation step was subjected to SDS gel electrophoresis. As seen in Fig. 7B, trypsin and chymotrypsin both caused extensive proteolytic cleavage of serum proteins. These results demonstrate that protease treatment cleaves serum proteins and prevents the serum-induced re-calcification of demineralized bone but do not actually prove that the proteases prevent serum-induced bone re-calcification by acting on the serum calcification factor itself rather than on the matrix of the demineralized tibia. To address this question, tibias were incubated for 6 days in DMEM containing 100 g/ml trypsin or in DMEM alone. Tibias were then washed with fresh medium (to remove trypsin) and incubated for another 6 days in DMEM containing 5% rat serum and no added trypsin. This experiment showed that there was comparable Alizarin Red staining for calcification in tibias preincubated for 6 days in DMEM plus trypsin and in tibias preincubated for 6 days in DMEM alone and comparable uptake of calcium and phosphate by the tibias (data not shown). These results demonstrate that trypsin treatment prevents the serum-dependent re-calcification of bone by inactivating the serum calcification factor itself, not by affecting bone matrix. Evidence That the Mineral Formed in the Serum-induced Re-calcification of Demineralized Bone Is Comparable to the Mineral Originally Found in Bone Fig. A (see Supplemental Data) shows the FTIR spectra of newborn rat tibia and of demineralized tibias that had been re-calcified by incubation for 6 days in either rat serum or in DMEM containing 5% rat serum. The spectra show the amide I and II peaks, which characterize the protein component of the bone matrix, and the phosphate and carbonate peaks, which characterize the mineral component. The ratio of the heights of the phosphate and carbonate peaks compared with the heights of the amide peaks is indicative of the extent to which the organic matrix has become mineralized. Although it is clear from an examination of the spectra that the re-calcified samples did not achieve the same degree of calcification as newborn bone, the spectra are very similar in their overall characteristics. The spectra also show that there was more mineral in the bone that was recalcified in DMEM containing 5% rat serum than was present in the matrix that was incubated in serum only. Identification of Initial Events in the Serum-induced Recalcification of Demineralized Tibia The above experiments demonstrate that the presence of serum is critical to calcification in DMEM but do not provide information on when serum acts. Experiments were therefore carried out to determine the effect of pre-treating tibias with serum on their subsequent calcification in DMEM. In the initial experiment, demineralized newborn rat tibias were incubated for 24 h in serum, for 5 days in DMEM, or for 24 h in serum followed by 5 days in DMEM. As can be seen in Fig. 8, there is no accumulation of

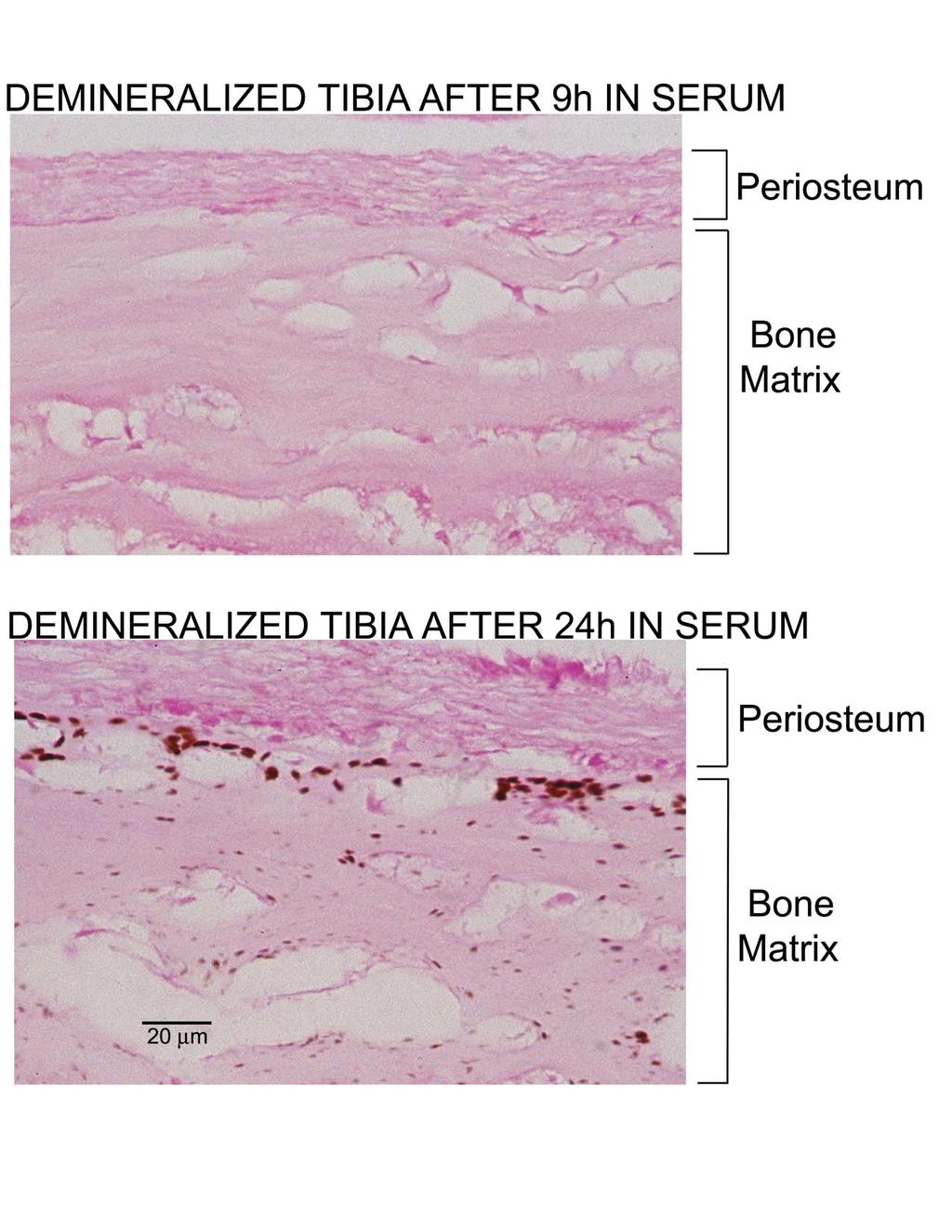
8 19176 Re-calcification of Bone in Serum FIG. 8.Effect of preincubation in rat serum on the subsequent calcification of tibias in DMEM. Single demineralized newborn rat tibias were incubated for 5 days in 10 ml of DMEM, for 24 h in 2 ml of rat serum, or for 24 h in 2 ml of rat serum followed by 5 days in 10 ml of DMEM. Tibias were then demineralized in acid, and the acid extracts were analyzed for calcium and phosphate (see Experimental Procedures ). The data show the amount of calcium and phosphate recovered from each tibia, as well as the amount of calcium and phosphate found in a newborn rat tibia prior to demineralization. calcium phosphate mineral after incubation in DMEM alone, and a small, but significant accumulation of a calcium phosphate mineral after incubation for 24 h in serum alone. In contrast, there is a dramatically increased amount of calcium phosphate accumulation after 24 h in serum followed by 5 days in DMEM, an amount far greater than the sum of the accumulation seen after incubation for 5 days in DMEM plus the accumulation seen after incubation for 24 h in serum. This synergistic effect of serum pre-treatment on calcification suggests that serum pre-treatment commits the demineralized tibia to further calcium and phosphate accumulation during subsequent incubation in DMEM. Interestingly, the final level of calcium and phosphate in tibias incubated for 24 h in serum followed by 5 days in DMEM is comparable to that present in tibias prior to demineralization (Table I), and the re-mineralized tibias are palpably hard. Because incubating tibias in a second 10-ml volume of DMEM did not significantly increase their calcium and phosphate content and did not calcify cartilage or periosteal structures, it appears that the capacity of demineralized bone to take up mineral is equal to the original mineral content of the tibia (data not shown). Additional experiments were carried out to determine the minimum time of serum pre-treatment needed for the subsequent re-calcification of demineralized tibias during the 5 days incubation in DMEM. As seen in Fig. 9, the minimum preincubation time was about 12 h, and further increasing the preincubation time in serum produces a progressive increase in the amount of calcium and phosphate in the tibia after 5 days incubation in DMEM. Surprisingly, there was no accumulation of calcium and phosphate in tibias pre-treated with serum for 9 h or less and then incubated for 5 days in DMEM (Fig. 9). The results of Alizarin Red staining for calcification confirmed the presence of calcification in tibias pre-treated with serum for 24 h and then incubated for 5 days in DMEM, and the absence of calcification in tibias pre-treated with serum for only9hand then incubated for 5 days in DMEM (not shown). To further characterize the initial steps in the serum-induced re-calcification of bone, tibias were examined by Alizarin Red staining after different times of pre-treatment in rat serum. This experiment showed that tibias that were pre-treated in serum for 24 h stained with Alizarin Red, whereas tibias pre-treated for only 9 h did not stain (not shown). This staining pattern was seen in subsequent experiments, and in all instances tibias pre-treated with serum for 12 h or longer stained with Alizarin Red, whereas tibias pre-treated with serum for 9 h or less did not stain with Alizarin Red. These results indicate that formation of a mineral phase that can stain with Alizarin Red during pre-treatment with serum predicts subsequent calcium and phosphate accumulation during incubation in DMEM (Fig. 9). Von Kossa staining was also used to characterize the critical initial steps in the serum-induced re-calcification of bone. As seen in Fig. B (Supplemental Data), pre-treatment with serum for 24 h resulted in numerous minute calcification foci throughout the demineralized bone matrix and the absence of foci in the periosteum and osteocyte lacunae. The largest calcification foci are located in the bone adjacent to the periosteum, which suggests that the growth of the foci could be limited by diffusion of calcium and phosphate into the matrix. In contrast, there was no evidence for calcification foci in the bone matrix of tibias incubated in serum for only 9 h (Supplemental Fig. B). These results indicate that formation of discrete mineral foci that can stain with von Kossa during pre-treatment with serum (Supplemental Fig. B) also predicts subsequent accumulation of calcium and phosphate during subsequent incubation in DMEM (Fig. 9). It seems likely that the accumulation of calcium and phosphate during incubation in DMEM reflects the growth of the initial foci present after the 24-h preincubation in serum. In principle, the failure of pre-treatment in serum for 9 h or less to cause Alizarin Red staining, von Kossa staining, or subsequent mineralization during incubation in DMEM could be explained either by time-dependent changes in the formation of the mineral phase, or time-dependent changes in the activity of the putative calcification factor in serum. Additional experiments were accordingly carried out to see if incubation of serum in the absence of tibias might be necessary to activate the calcifying activity found serum. Freshly obtained serum was incubated at 37 C for 18 h and then used to pre-treat tibias for 9 or 24 h. The tibias were then incubated in DMEM for 5 days. This experiment showed that tibias incubated in pre-treated serum for 24 h calcified, whereas tibias incubated in pre-treated serum for only 9 h did not calcify (not shown). These results show that the failure of pre-treatment for 9 h or less to cause calcification during subsequent incubation for 5 days in DMEM must be due to time-dependent changes in the mineral phase not to changes in the activity of the calcification factor in serum caused by incubation at 37 C. The Total Re-calcification of Weanling Rat Tibia A final experiment was carried out to determine whether the seruminitiated re-calcification of bone could restore the mineral content of bone to its original level prior to demineralization. One-centimeter segments were cut from the midshaft region of tibias from 22-day-old rats and demineralized in EDTA. Each demineralized tibia segment was then incubated separately in 2 ml of rat serum for 3 days at 37 C to initiate the re-calcification reaction. This incubation step resulted in extensive Alizarin Red staining for calcification (not shown) and the incorporation of about 4 mol of calcium and 2.5 mol of phosphate into the bone matrix. Examination of von Kossa-stained sections of these tibias showed the presence of numerous brown calcification foci scattered throughout the demineralized bone matrix; in the representative section shown in Fig. 10 there are about 800 calcification foci, and the foci range from 0.2 to 2 m in size. The counterstain used in this study, nuclear fast red, stains cytoplasm and the demineralized bone matrix pink and cell nuclei red. Osteocyte lacunae, which have a roughly spherical shape with a diameter of about 10 m, appear as 5- to

9 Re-calcification of Bone in Serum FIG. 9. Determination of the minimum preincubation in rat serum required for the subsequent calcification of tibias in DMEM culture medium. Demineralized newborn rat tibias were incubated for the indicated times in 2 ml of rat serum and then placed in 10 ml of DMEM for a period of 5 days. Tibias were then demineralized in acid, and the acid extracts were analyzed for calcium and phosphate (see Experimental Procedures ). The data show the amount of calcium and phosphate recovered from each tibia. FIG. 10. Effect of incubating weanling rat tibias in serum for 3 days on histological staining for calcification. Tibias were dissected from 22-day-old rats, cut to obtain 1-cm sections of the midshaft, freed of marrow and cancellous bone, and demineralized with EDTA. Tibias were then incubated in 2 ml of rat serum for 3 days and fixed in ethanol. 5- m-thick cross sections were stained by von Kossa (stains calcification brown to black) and counterstained by nuclear fast red (stains collagen matrix and cytoplasm pink and nuclei red). BV, blood vessel; OCL, osteocyte lacuna; M, calcification foci. Magnification, 1000; the bar denotes 10 m. 12- m diameter holes in this 5- m-thick section. The size, shape, and number of osteocyte lacunae seen in the present study are in agreement with the results of earlier studies of demineralized rat cortical bone (9). There was no evidence for calcification in the osteocyte lacunae or blood vessels in any of the tibia sections examined.

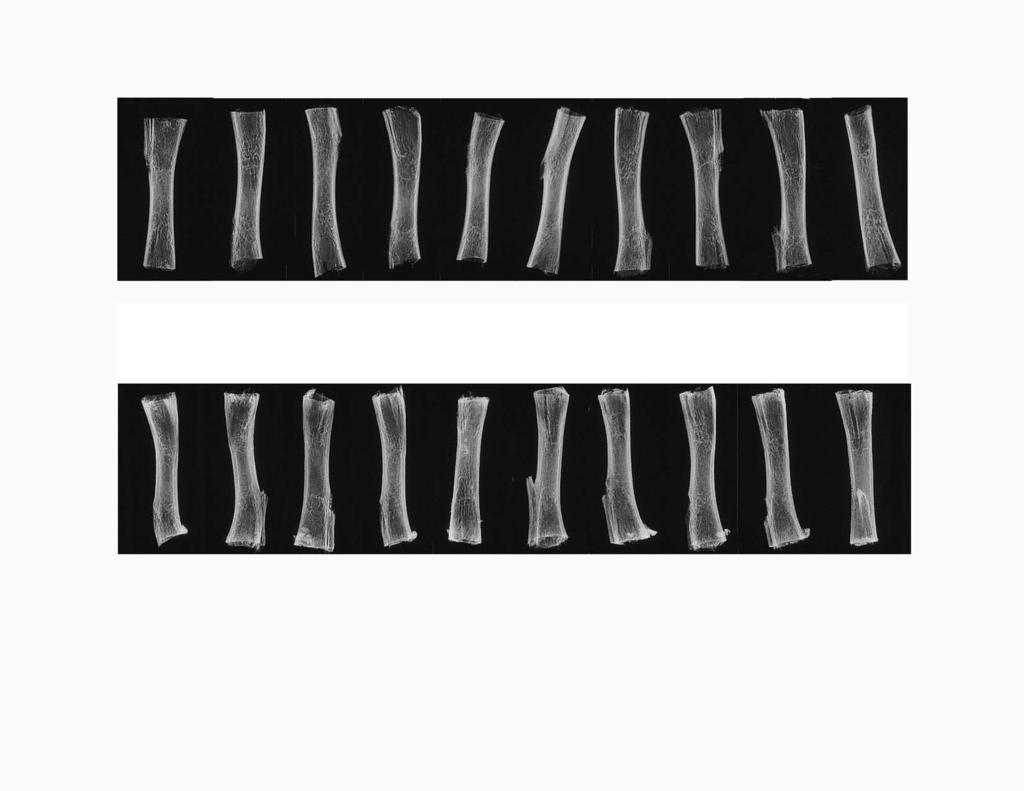
10 19178 Re-calcification of Bone in Serum most of the weight originally found in the tibia and restored somewhat more calcium and phosphate than originally found in the tibia. The molar calcium to phosphate ratio of the recalcified tibias was 1.61, which is higher than the 1.50 ratio found here for the original tibias prior to demineralization but comparable to the 1.65 ratio found in other studies for the bone of adult mammals (10, 11). Supplemental Fig. C shows that the radiographs of the re-calcified weanling rat tibias were comparable to radiographs of tibias prior to demineralization. This result demonstrates that the distribution and density of recalcification match that of the original tibia prior to demineralization. The powder x-ray diffraction spectrum of the recalcified tibias is that expected for hydroxyapatite (Fig. D, Supplemental Data), and the FTIR spectrum of the re-calcified tibias is comparable to the FTIR spectrum of the original tibias prior to demineralization (not shown). FIG. 11. Chemical analysis of weanling rat tibia midshafts before demineralization and after demineralization followed by serum-initiated re-calcification. All of the 30 tibia from the experiment shown in Fig. C (Supplemental Data) were dried and weighed. Five tibias from each group were then demineralized in acid, and the resulting extracts were analyzed for calcium and phosphate. The graph shows the mean S.D. for the weights of the 10 tibias in each group and for the calcium and phosphate in a subset of five tibias from each group. Initial Tibia, tibias that were not demineralized. Re-calcified Tibia, demineralized tibias that were pre-treated with serum for 3 days and then incubated in buffer for 30 days. Demineralized Tibia, demineralized tibias that were incubated for 30 days in buffer without serum pre-treatment. To foster further growth of the calcification foci introduced into the bone matrix by incubation in serum, each tibia was then placed into 20 ml of 25 mm HEPES buffer, ph 7.4, containing 1 mm calcium, 3 mm phosphate, 100 mm NaCl, and 30 mm NaHCO 3, and the tibias were incubated at 37 C. Medium was exchanged for fresh medium every other day for 1 month, and the progress of the re-calcification reaction was monitored by depletion in medium calcium and uptake of calcium by the tibias. Calcium uptake by the tibias was initially rapid, diminished with time, and could no longer be demonstrated by 30 days. 4 Fig. 11 shows that the re-calcification process restored 4 This experiment has been repeated using bovine cortical bone sand that was m in size and was demineralized with EDTA. Substitution of 4 mg of this demineralized bone sand for the 1-cm segments of the weanling tibia midshaft resulted in a similar time course of re-calcification, with cessation of calcium uptake by the matrix after 30-day incubation in the HEPES buffer. The final amount of calcium and phosphate recovered in the bone sand was, within error, the same as that found in the equivalent amount of bone sand prior to demineralization. These observations indicate that the size of the bone matrix particles has no appreciable effect on the extent of the recalcification process. DISCUSSION Evidence for a Factor in Rat Serum That Initiates the Recalcification of Demineralized Bone The present studies show for the first time that demineralized newborn rat bone calcifies rapidly when incubated in rat serum or heparin plasma at the temperature, carbonate concentration, and ph of rat blood. The re-calcification of demineralized bone is remarkably specific for those regions of bone matrix that were calcified prior to demineralization, and we have not seen an instance in which regions of the demineralized newborn rat tibia or calvaria that were calcified in the original bone did not re-calcify when incubated in serum, nor have we seen an instance in which regions that were not calcified in the original bone became calcified during incubation in serum. Identical re-calcification was seen in newborn rat tibias that were demineralized with 0.5 M EDTA, ph 7.5, or with 150 mm HCl, and in demineralized tibias that were further extracted with acetone followed by 6 M guanidine HCl. Efficient re-calcification was also seen with demineralized tibias from weanling rats (Figs. 10 and 11) and with demineralized bovine cortical bone sand (personal observations). There is therefore nothing unique about the species of animal from which the bone matrix was obtained, the age of the animal, the physical structure of the bone, or the method of demineralization. The matrix of demineralized bone calcifies in serum. The potency of the serum agent (or agents 1 ) that initiates the re-calcification of demineralized bone matrix was assessed by examining the effects of diluting serum into DMEM culture medium. This experiment showed that the re-calcification of demineralized bone occurs during 6-day incubation in DMEM containing as little as 1.5% rat serum, but not in DMEM containing 0.15% rat serum or in serum-free DMEM. Because the addition of only 1.5% serum to DMEM has a trivial impact on the calcium and phosphate content of the final solution yet causes dramatic calcification of bone, the serum-induced recalcification of demineralized bone cannot be caused by an effect of serum on the ionic milieu of DMEM but must instead be caused by a potent calcification agent that is present in serum. The histological appearance of the re-calcification of demineralized bone following incubation in either serum alone, or in medium containing 1.5% or 15% serum, is invariably discontinuous, with numerous calcification foci scattered throughout the bone matrix. The density of these foci in bone is in the order 100% serum 15% serum 1.5% serum, which suggests that each foci may reflect a discrete nucleation event initiated by the putative serum calcification factor. The activity of the serum calcification factor does not dilute in DMEM in a regular, monotonic fashion, however, as might have been expected for a single calcification factor acting alone. This is shown by the observation that de-
11 Re-calcification of Bone in Serum mineralized newborn rat bone matrices incubated for 6 days in DMEM containing 1.5% rat serum have innumerable calcification foci in von Kossa stained sections, whereas demineralized bone matrices incubated in the same media containing 0.15% serum have no calcification foci at all, not simply 10% of the number seen at 1.5% serum as would be anticipated from simple dose dilution of the putative serum calcification factor. The same result has also been seen with other demineralized bone matrices, such as ground calf bone and weanling rat tibia, and in each instance there are numerous discrete calcification foci in bone following incubation for 6 days in DMEM containing 1.5% serum and none after incubation in DMEM containing 0.15% serum (personal observations). Evidence That the Putative Serum Calcification Factor May Be Specific for the Calcification of a Type I Collagen Matrix The observation that the re-calcification of demineralized bone in serum, or in medium containing small amounts of serum, is invariably specific for those regions of the bone matrix that were originally calcified could indicate that agents remain in the bone structures after demineralization that promote serum-induced calcification. Alternatively, serum-induced calcification could be specific for the chemical nature of the extracellular matrix of bone, type I collagen. In the present study, we have tested the second hypothesis by examining the possible calcification of rat tail tendon, a matrix that also consists of type I collagen but does not calcify in vivo, during incubation in rat serum and plasma. Because tendon consistently calcified after incubation in rat serum and plasma and in physiological media containing 1.5% rat serum, we tentatively conclude that the specificity of serum-induced re-calcification of demineralized tibia and calvaria may be related to the distribution of type I collagen matrix in these tissues. If this hypothesis is true, the pattern of type I collagen calcification in an animal may be explained by exclusion of serum calcification factor and/or the local production of calcification inhibitors within those matrices that do not normally calcify (e.g. tendon), and by the local production of high levels of calcification factor within those matrices that do calcify (e.g. bone). Relationship between the Serum-dependent Re-calcification of Demineralized Bone and Earlier Studies Showing That Bone Can Be Re-calcified in the Absence of Serum Previous studies have shown that the type I collagen matrices of demineralized bone and tendon will calcify when incubated in suitable test solutions at physiological ph and temperature (12 14), and these observations have led to the conclusion that type I collagen matrices are predisposed to calcify. Because the test solutions used in these early experiments contained no serum, the fact that the matrices of tendon and demineralized bone clearly calcified might appear to invalidate the present conclusion that serum plays a critical role in matrix calcification. These earlier studies utilized a 3.75 mm calcium concentration (12 14), however, which is far higher than calcium concentrations in the buffers used in the present study (1.8 and 1 mm) and far higher than the concentration of ionic calcium in serum (1 mm). In addition, the calcium phosphate ionic product used in these earlier studies was 6.3 mm 2 (12 14), which is well above that in DMEM (3.6 mm 2 ) or rat serum (3 mm 2 ). There is, to our knowledge, no report that demineralized bone can be re-calcified in the absence of serum by incubation in buffers that contain physiological levels of calcium, phosphate, and ph. The high calcium concentration and high calcium phosphate ion product conditions used in these earlier serum-free studies resulted in the occasional precipitation of mineral in the absence of the addition of matrix (12, 13), a precipitation never observed in the present study with DMEM. This observation indicates that the high calcium phosphate ion product conditions used for serum-free re-calcification of demineralized bone in these early studies are at or near the threshold for the spontaneous precipitation of a mineral phase. We have here shown that demineralized bone matrices do not re-calcify when incubated in serum-free solutions that contain physiological levels of calcium, phosphate, and ph and that, therefore, have a calcium phosphate ion product that is well below the threshold for the spontaneous precipitation of a mineral phase. The novel discovery of the present studies is that solutions that contain physiological levels of calcium, phosphate, and ph can be made to support the re-calcification of demineralized bone by the addition of as little as 1.5% rat serum. Our working hypothesis is that the role of the serum calcification factor in re-calcification is to lower the ion product necessary for the formation of a mineral phase within a type I collagen matrix from the 6.26 mm 2 value found in the absence of serum, to the 3.6 mm 2 value found in DMEM containing as little as 1.5% serum. Biochemical Characterization of the Serum Calcification Factor That Initiates the Re-calcification of Bone The present studies provide information on the chemical nature of the serum calcification factor that initiates the re-calcification of demineralized bone. Gel filtration experiments show that the serum calcification factor is a macromolecule that emerges from a Sephacryl S-100HR column in the position of serum proteins with an apparent molecular mass of kda. Additional experiments show that the activity of the serum calcification factor is completely abolished by treatment with trypsin or chymotrypsin. These results strongly suggest that the serum calcification factor that initiates the re-calcification of bone consists of one or more proteins of kda apparent molecular mass. Experiments carried out using different times of pre-treatment in serum followed by incubation for 5 days in serum-free DMEM show that the unique activity of the serum calcification factor is its ability to generate numerous discrete calcification foci throughout the bone matrix and that the subsequent growth of these nuclei occurred rapidly in DMEM in the complete absence of serum. The introduction of nucleation centers within the matrix during incubation in serum does not, however, follow the typical first order kinetics that might be anticipated for a single agent catalyzing a rapid transition to a single mineral phase. There is, instead, a puzzling 9-h lag between the contact of serum with the tibia and the initiation of the first mineral that can be detected by Alizarin Red or von Kossa staining, and the first mineral phase that is competent to grow during subsequent incubation in DMEM. The simplest explanation for this observation is that the one or more putative serum calcification factors must remain associated with a single nucleation event for a prolonged period before this calcification center has acquired the properties of staining and ability to grow in DMEM. The continued uptake of calcium and phosphate by a demineralized tibia ceases when the amount of calcium and phosphate introduced into the matrix is comparable to that present in the original bone prior to demineralization (Fig. 11). 4 In addition, radiographs of the re-calcified tibias show that bone mineral density is identical to that seen in the original bone prior to demineralization (Supplemental Fig. C). These observations suggest that the extent of serum-induced bone re-calcification and normal bone mineralization may be subject to similar physical constraints. Because previous studies have shown that most bone mineral lies within the collagen fiber (15), this constraint could be the finite space that exists within the collagen fibers of the organic bone matrix. Summary The present studies show for the first time that
12 19180 Re-calcification of Bone in Serum serum contains a potent calcification factor that initiates the re-calcification of demineralized bone and provide evidence that this factor is a protein of kda molecular mass. The matrix specificity of serum-initiated bone re-calcification exactly duplicates the gross and microscopic location of mineral found in the original bone. The extent of re-calcification is finite, and the final mineralization density is comparable to that of the original bone. The nature of the mineral phase introduced in the serum-initiated re-calcification of bone is comparable in XRD and FTIR spectra to that originally found in the bone prior to demineralization. Taken together, these observations strongly suggest that serum-initiated re-calcification of demineralized bone could be identical to, or closely similar to, the process by which bone matrix is normally mineralized. Many questions remain, however. Although the present assay employs an arguably physiologically relevant fluid, serum, or plasma, the calcifiable matrix is a demineralized bone that lacks cells as well as some non-collagenous bone proteins. It is not yet clear that this assay system is a reasonable model for matrix mineralization in living bone, and one must therefore be cautious in extrapolating the present findings to normal bone mineralization. Future studies will clearly be needed to identify the protein (or proteins) that are involved in serum-initiated bone re-calcification and to determine whether this protein (or proteins) participates in normal bone mineralization, ectopic calcifications, and/or other calcification processes. REFERENCES 1. Price, P. A., Faus, S. A., and Williamson, M. K. (2000) Arterioscler. Thromb. Vasc. Biol. 20, Price, P. A., Faus, S. A., and Williamson, M. K. (2001) Arterioscler. Thromb. Vasc. Biol. 21, Bucay, N., Sarosi, I., Dunstan, C. R., Morony, S., Tarpley, J., Capparelli, C., Scully, S., Tan, H. L., Xu, W., Lacey, D. L., Boyle, W., and Simonset, W. S. (1998) Genes Dev. 12, Price, P. A., June, H. H., Buckley, J. R., and Williamson, M. K. (2001) Arterioscler. Thromb. Vasc. Biol. 21, Price, P. A., June, H. H., Buckley, J. R., and Williamson, M. K. (2002) Circ. Res. 91, Luo, G., Ducy, P., McKee, M. D., Pinero, G. J., Loyer, E., Behringer, R. R., and Karsenty, G. (1997) Nature 386, Price, P. A., Faus, S. A., and Williamson, M. K. (1998) Arterioscler. Thromb. Vasc. Biol. 18, Chen, P. S., Toribara, T. Y., and Warner, H. (1956) Anal. Chem. 28, Hamaya, M., Mizoguchi, I., Sakakura, Y., Yajima, T., and Abiko, Y. (2002) Calcif. Tissue Int. 70, Elliott, J. C. (1994) Structure and Chemistry of the Apatites and Other Calcium Orthophosphates. Studies in Inorganic Chemistry, p. 260, Elsevier, The Netherlands 11. Driessens, F. C. M., and Verbeeck, R. M. H. (1990) in Biominerals (Driessens, F. C. M., and Verbeeck, R. M. H., eds) p. 183, CRC Press, Boca Raton 12. Bachra, B. N. (1972) Calcif. Tissue Res. 8, Bachra, B. N., and Fischer, H. R. A. (1968) Calcif. Tissue Res. 2, DeSteno, C. V., Feagin, F., and Butler, W. T. (1975) Calcif. Tissue Res. 17, Currey, J. D. (2002) Bones: Structure and Mechanics, 1st Ed., pp. 4 11, Princeton University Press, Princeton and Oxford
13 SUPPLEMENTAL DATA Figure A. FTIR spectra of newborn rat tibias and of demineralized rat tibias after recalcification in rat serum or in DMEM containing 5% rat serum. Ten demineralized newborn rat tibias were each re-calcified by incubation for 6d in 2ml of rat serum, and 10 demineralized newborn rat tibias were each re-calcified by incubation for 6d in 10ml of DMEM containing 5% rat serum. The diaphyseal bone was removed from each re-calcified tibia and from 10 non-demineralized newborn rat tibias and immediately dried. The dried diaphyseal bone was ground and examined by FTIR. Figure B. Effect of incubating newborn rat tibias in serum for 9 and 24h on histological staining for calcification. Demineralized newborn rat tibias were incubated in 2ml of rat serum for 9 or 24h and then fixed in ethanol. Five micron thick longitudinal sections were stained by von Kossa (stains calcification brown to black) and counter stained by nuclear fast red (stains collagen matrix and cytoplasm pink and nuclei red). Magnification, 400X; the bar denotes 20 microns. Osteocyte lacunae, which appear as approximately 10 micron holes in this section, are free of calcification. Figure C. Radiographs of weanling rat tibia midshafts before demineralization, and after demineralization followed by serum-initiated re-calcification. Tibias were dissected from 22 day old rats, cut to obtain 1cm sections of the midshaft, and freed of marrow and cancellous bone. Top row: radiographs of 10 tibias before demineralization. Bottom row: radiograph of 10 tibias after demineralization in EDTA followed by incubation for 3d in rat serum to initiate calcification and subsequent incubation for 30d in 20ml of 25mM HEPES buffer ph 7.4 containing 1mM calcium, 3mM phosphate, 100mM NaCl, and 30mM NaHCO 3 (with medium change every other day). As a control, 10 demineralized tibias were also incubated in the HEPES
14 buffer for 30d but were not pre-treated with serum; these tibias could not be detected on the radiograph at this setting and are therefore not shown. Figure D. Powder X ray diffraction spectrum of weanling rat tibia midshafts before demineralization, and after demineralization followed by serum-initiated re-calcification. Five tibias prior to demineralization and 5 tibias after demineralization followed by seruminitiated re-calcification (see Figure 10) were separately pooled and ground in an agate mortar. The X ray diffraction spectrum of each powder was determined with a Scintag SDF 2000 X ray diffractometer.
15
16
17
Two major types of arterial calcification have been observed
 The Elastic Lamellae of Devitalized Arteries Calcify When Incubated in Serum Evidence for a Serum Calcification Factor Paul A. Price, Wai Si Chan, Dawn M. Jolson, Matthew K. Williamson Downloaded from
The Elastic Lamellae of Devitalized Arteries Calcify When Incubated in Serum Evidence for a Serum Calcification Factor Paul A. Price, Wai Si Chan, Dawn M. Jolson, Matthew K. Williamson Downloaded from
The Inhibition of Calcium Phosphate Precipitation by Fetuin Is Accompanied by the Formation of a Fetuin-Mineral Complex*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 24, Issue of June 13, pp. 22144 22152, 2003 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Inhibition
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 24, Issue of June 13, pp. 22144 22152, 2003 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Inhibition
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
BONE LABORATORY DEMONSTRATIONS. These demonstrations are found on the bulletin boards outside the MCO Bookstore.
 BONE LABORATORY DEMONSTRATIONS These demonstrations are found on the bulletin boards outside the MCO Bookstore. COMPACT & TRABECULAR BONE - LM When viewed under the polarizing light microscope, the layering
BONE LABORATORY DEMONSTRATIONS These demonstrations are found on the bulletin boards outside the MCO Bookstore. COMPACT & TRABECULAR BONE - LM When viewed under the polarizing light microscope, the layering
Supporting information
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Supporting information Experimental Section Synthesis and modification of SBA-15. In a typical
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Supporting information Experimental Section Synthesis and modification of SBA-15. In a typical
BONE TISSUE. Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology
 BONE TISSUE Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology BONE FUNCTION Support Protection (protect internal organs) Movement (provide leverage system for skeletal muscles, tendons, ligaments
BONE TISSUE Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology BONE FUNCTION Support Protection (protect internal organs) Movement (provide leverage system for skeletal muscles, tendons, ligaments
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid Jun Gong, Xin Lu, Xueqin An*
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid Jun Gong, Xin Lu, Xueqin An*
JBC Papers in Press. Published on June 11, 2007 as Manuscript M
 JBC Papers in Press. Published on June 11, 2007 as Manuscript M700591200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m700591200 THE SIZE EXCLUSION CHARACTERISTICS OF TYPE I COLLAGEN:
JBC Papers in Press. Published on June 11, 2007 as Manuscript M700591200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m700591200 THE SIZE EXCLUSION CHARACTERISTICS OF TYPE I COLLAGEN:
Decalcification and Clearing of Bone eg. Fibula. Salem Kharwa Clinical Anatomy Medical School
 Decalcification and Clearing of Bone eg. Fibula Salem Kharwa Clinical Anatomy Medical School Bone composition Bone consists of cells (osteocytes) surrounded by a calcified matrix containing collagen fibres.
Decalcification and Clearing of Bone eg. Fibula Salem Kharwa Clinical Anatomy Medical School Bone composition Bone consists of cells (osteocytes) surrounded by a calcified matrix containing collagen fibres.
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
BONE HISTOLOGY SLIDE PRESENTATION
 BONE HISTOLOGY SLIDE PRESENTATION PRESENTED BY: SKELETECH, INC. Clients and Friends: SkeleTech invites you to use these complimentary images for your own presentations or as teaching slides for bone biology.
BONE HISTOLOGY SLIDE PRESENTATION PRESENTED BY: SKELETECH, INC. Clients and Friends: SkeleTech invites you to use these complimentary images for your own presentations or as teaching slides for bone biology.
The Skeletal System:Bone Tissue
 The Skeletal System:Bone Tissue Dynamic and ever-changing throughout life Skeleton composed of many different tissues cartilage, bone tissue, epithelium, nerve, blood forming tissue, adipose, and dense
The Skeletal System:Bone Tissue Dynamic and ever-changing throughout life Skeleton composed of many different tissues cartilage, bone tissue, epithelium, nerve, blood forming tissue, adipose, and dense
CHAPTER 6 LECTURE OUTLINE
 CHAPTER 6 LECTURE OUTLINE I. INTRODUCTION A. Bone is made up of several different tissues working together: bone, cartilage, dense connective tissue, epithelium, various blood forming tissues, adipose
CHAPTER 6 LECTURE OUTLINE I. INTRODUCTION A. Bone is made up of several different tissues working together: bone, cartilage, dense connective tissue, epithelium, various blood forming tissues, adipose
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Cholesterol determination using protein-templated fluorescent gold nanocluster probes
 Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Deposition of Bone by the Osteoblasts. Bone is continually being deposited by osteoblasts, and it is continually being resorbed where osteoclasts are
 Bone remodeling Deposition of Bone by the Osteoblasts. Bone is continually being deposited by osteoblasts, and it is continually being resorbed where osteoclasts are active. This mechanism is always is
Bone remodeling Deposition of Bone by the Osteoblasts. Bone is continually being deposited by osteoblasts, and it is continually being resorbed where osteoclasts are active. This mechanism is always is
Chapter PURIFICATION OF ALKALINE PROTEASES
 Chapter PURIFICATION OF ALKALINE PROTEASES E /xtracellular alkaline proteases produced by Bacillus sp. K 25 and bacillus pumilus K 242, were purified and the homogeneity was examined by electrophoresis.
Chapter PURIFICATION OF ALKALINE PROTEASES E /xtracellular alkaline proteases produced by Bacillus sp. K 25 and bacillus pumilus K 242, were purified and the homogeneity was examined by electrophoresis.
Fig.S1 ESI-MS spectrum of reaction of ApA and THPTb after 16 h.
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Experiment Cleavage of dinucleotides Dinucleotides (ApA, CpC, GpG, UpU) were purchased from
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Experiment Cleavage of dinucleotides Dinucleotides (ApA, CpC, GpG, UpU) were purchased from
Nano-analytical electron microscopy reveals fundamental insights into human cardiovascular tissue calcification
 Nano-analytical electron microscopy reveals fundamental insights into human cardiovascular tissue calcification Supplementary table 1: Aortic valve donor information (n=32). Patient age (yr) 57±11 (range:
Nano-analytical electron microscopy reveals fundamental insights into human cardiovascular tissue calcification Supplementary table 1: Aortic valve donor information (n=32). Patient age (yr) 57±11 (range:
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Improve Protein Analysis with the New, Mass Spectrometry- Compatible ProteasMAX Surfactant
 Improve Protein Analysis with the New, Mass Spectrometry- Compatible Surfactant ABSTRACT Incomplete solubilization and digestion and poor peptide recovery are frequent limitations in protein sample preparation
Improve Protein Analysis with the New, Mass Spectrometry- Compatible Surfactant ABSTRACT Incomplete solubilization and digestion and poor peptide recovery are frequent limitations in protein sample preparation
Engineering the Growth of TiO 2 Nanotube Arrays on Flexible Carbon Fibre Sheets
 Engineering the Growth of TiO 2 Nanotube Arrays on Flexible Carbon Fibre Sheets Peng Chen, a Li Gu, b Xiudong Xue, a Mingjuan Li a and Xuebo Cao* a a Key Lab of Organic Synthesis of Jiangsu Province and
Engineering the Growth of TiO 2 Nanotube Arrays on Flexible Carbon Fibre Sheets Peng Chen, a Li Gu, b Xiudong Xue, a Mingjuan Li a and Xuebo Cao* a a Key Lab of Organic Synthesis of Jiangsu Province and
B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small cell lung cancer
, NCI-H460 (human non-small cell lung cancer") Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Tivadar Orban, Beata Jastrzebska, Sayan Gupta, Benlian Wang, Masaru Miyagi, Mark R. Chance, and Krzysztof Palczewski
 Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Bone Apatite Composition Of Osteonecrotic Trabecular Bone From The Femoral Head Of Immature Pigs
 Bone Apatite Composition Of Osteonecrotic Trabecular Bone From The Femoral Head Of Immature Pigs Olumide O. Aruwajoye 1,2, Harry KW Kim, MD 1, Pranesh B. Aswath, PhD 2. 1 Texas Scottish Rite Hospital,
Bone Apatite Composition Of Osteonecrotic Trabecular Bone From The Femoral Head Of Immature Pigs Olumide O. Aruwajoye 1,2, Harry KW Kim, MD 1, Pranesh B. Aswath, PhD 2. 1 Texas Scottish Rite Hospital,
Sansom & Manston, 1963) and rats (Payne & Sansom, 1963). It appeared
 and rats (Payne & Sansom, 1963). It appeared") J. Physiol. (1964), 170, pp. 613-620 613 Printed in Great Britain THE RELATIVE TOXICITY IN RATS OF DISODIUM ETHYLENE DIAMINE TETRA-ACETATE, SODIUM OXALATE AND SODIUM CITRATE BY J. M. PAYNE AND B. F. SANSOM
J. Physiol. (1964), 170, pp. 613-620 613 Printed in Great Britain THE RELATIVE TOXICITY IN RATS OF DISODIUM ETHYLENE DIAMINE TETRA-ACETATE, SODIUM OXALATE AND SODIUM CITRATE BY J. M. PAYNE AND B. F. SANSOM
Chapter 7. Skeletal System
 Chapter 7 Skeletal System 1 Introduction: A. Bones are very active, living tissues B. Each bone is made up of several types of tissues and so is an organ. C. Bone functions include: muscle attachment,
Chapter 7 Skeletal System 1 Introduction: A. Bones are very active, living tissues B. Each bone is made up of several types of tissues and so is an organ. C. Bone functions include: muscle attachment,
Protein Dephosphorylation Methods
 Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Total Phosphatidic Acid Assay Kit
 Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
UV Tracer TM Maleimide NHS ester
 UV Tracer TM Maleimide HS ester Product o.: 1020 Product ame: UV-Tracer TM Maleimide-HS ester Chemical Structure: Chemical Composition: C 41 H 67 5 18 Molecular Weight: 1014.08 Appearance: Storage: Yellow
UV Tracer TM Maleimide HS ester Product o.: 1020 Product ame: UV-Tracer TM Maleimide-HS ester Chemical Structure: Chemical Composition: C 41 H 67 5 18 Molecular Weight: 1014.08 Appearance: Storage: Yellow
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Recombinant Trypsin, Animal Origin Free
 Recombinant Trypsin, Animal Origin Free PRODUCT INFORMATION: BioGenomics r-trypsin powder is ready to use, animal origin free optimized for cell culture applications. It is derived by r-dna technology.
Recombinant Trypsin, Animal Origin Free PRODUCT INFORMATION: BioGenomics r-trypsin powder is ready to use, animal origin free optimized for cell culture applications. It is derived by r-dna technology.
Organic Molecule Composition of Milk: Lab Investigation
 Name: Organic Molecule Composition of Milk: Lab Investigation Introduction & Background Milk & milk products have been a major food source from earliest recorded history. Milk is a natural, nutritionally
Name: Organic Molecule Composition of Milk: Lab Investigation Introduction & Background Milk & milk products have been a major food source from earliest recorded history. Milk is a natural, nutritionally
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
E.Z.N.A. SQ Blood DNA Kit II. Table of Contents
 E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Note: During 30 minute incubation; proceed thru appropriate sections below (e.g. sections II, III and V).
.") LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
Skeletal Tissues. Skeletal tissues. Frame; muscles, organs and CT attach. Brain, spinal cord, thoracic organs; heart and lungs.
 Skeletal Tissues Functions 1) support 2) protection 3) movement Skeletal tissues Frame; muscles, organs and CT attach. Brain, spinal cord, thoracic organs; heart and lungs. Aids muscle contraction; generate
Skeletal Tissues Functions 1) support 2) protection 3) movement Skeletal tissues Frame; muscles, organs and CT attach. Brain, spinal cord, thoracic organs; heart and lungs. Aids muscle contraction; generate
Supporting Information
 Supporting Information The Effects of Spacer Length and Composition on Aptamer-Mediated Cell-Specific Targeting with Nanoscale PEGylated Liposomal Doxorubicin Hang Xing +, [a] Ji Li +, [a] Weidong Xu,
Supporting Information The Effects of Spacer Length and Composition on Aptamer-Mediated Cell-Specific Targeting with Nanoscale PEGylated Liposomal Doxorubicin Hang Xing +, [a] Ji Li +, [a] Weidong Xu,
INSTRUCTIONS Pierce Primary Cardiomyocyte Isolation Kit
 INSTRUCTIONS Pierce Primary Cardiomyocyte Isolation Kit 88281 Number Description 88281 Pierce Primary Cardiomyocyte Isolation Kit, contains sufficient reagents to isolate cardiomyocytes from 50 neonatal
INSTRUCTIONS Pierce Primary Cardiomyocyte Isolation Kit 88281 Number Description 88281 Pierce Primary Cardiomyocyte Isolation Kit, contains sufficient reagents to isolate cardiomyocytes from 50 neonatal
FORMATION OF BONE. Intramembranous Ossification. Bone-Lec-10-Prof.Dr.Adnan Albideri
 FORMATION OF BONE All bones are of mesodermal origin. The process of bone formation is called ossification. We have seen that formation of most bones is preceded by the formation of a cartilaginous model,
FORMATION OF BONE All bones are of mesodermal origin. The process of bone formation is called ossification. We have seen that formation of most bones is preceded by the formation of a cartilaginous model,
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
COPD lungs show an attached stratified mucus layer that separate. bacteria from the epithelial cells resembling the protective colonic
 COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
Chapter 6: Skeletal System: Bones and Bone Tissue
 Chapter 6: Skeletal System: Bones and Bone Tissue I. Functions A. List and describe the five major functions of the skeletal system: 1. 2. 3.. 4. 5.. II. Cartilage A. What do chondroblasts do? B. When
Chapter 6: Skeletal System: Bones and Bone Tissue I. Functions A. List and describe the five major functions of the skeletal system: 1. 2. 3.. 4. 5.. II. Cartilage A. What do chondroblasts do? B. When
Chapter 6: SKELETAL SYSTEM
 Chapter 6: SKELETAL SYSTEM I. FUNCTIONS A. Support B. Protection C. Movement D. Mineral storage E. Lipid storage (Fig. 6.8b) F. Blood cell production (Fig. 6.4) II. COMPONENTS A. Cartilage 1. Hyaline 2.
Chapter 6: SKELETAL SYSTEM I. FUNCTIONS A. Support B. Protection C. Movement D. Mineral storage E. Lipid storage (Fig. 6.8b) F. Blood cell production (Fig. 6.4) II. COMPONENTS A. Cartilage 1. Hyaline 2.
Module 2:! Functional Musculoskeletal Anatomy A! Semester 1! !!! !!!! Hard Tissues, Distal Upper Limb & Neurovascular Supply of Upper Limb!
 Functional Musculoskeletal Anatomy A Module 2: Hard Tissues, Distal Upper Limb & Neurovascular Supply of Upper Limb Semester 1 1 18. Bone Tissue & Growth of Bones 18.1 Describe the structure of bone tissue
Functional Musculoskeletal Anatomy A Module 2: Hard Tissues, Distal Upper Limb & Neurovascular Supply of Upper Limb Semester 1 1 18. Bone Tissue & Growth of Bones 18.1 Describe the structure of bone tissue
Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples:
 Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
DELFIA Tb-DTPA ITC Chelate & Terbium Standard
 AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
Rescue IVF protocol for legacy stock
 Rescue IVF protocol for legacy stock Sperm thawing/ivf protocol for MTG sperm samples (80ul per straw) from straw and conventional CPA from Vial (100ml per vial) This protocol is based on methods developed
Rescue IVF protocol for legacy stock Sperm thawing/ivf protocol for MTG sperm samples (80ul per straw) from straw and conventional CPA from Vial (100ml per vial) This protocol is based on methods developed
Bone Tissue- Chapter 5 5-1
 Bone Tissue- Chapter 5 5-1 Bone Functions Support Protection Assistance in movement Mineral storage and release Blood cell production Triglyceride storage 5-2 Bone Chemistry Water (25%) Organic Constituent
Bone Tissue- Chapter 5 5-1 Bone Functions Support Protection Assistance in movement Mineral storage and release Blood cell production Triglyceride storage 5-2 Bone Chemistry Water (25%) Organic Constituent
Chapter CHAPTER 7. ELECTRICAL PROPERTIES OF ZnO DOPED MAGESIUM ALUMIUM SILICATE GLASS-CERAMICS
 Chapter 7 102 CHAPTER 7 ELECTRICAL PROPERTIES OF ZnO DOPED MAGESIUM ALUMIUM SILICATE GLASS-CERAMICS Chapter 7 103 CHAPTER 7 ELECTRICAL PROPERTIES OF ZnO DOPED MAGNESIUM ALUMINUM SILICATE GLASS-CERAMICS
Chapter 7 102 CHAPTER 7 ELECTRICAL PROPERTIES OF ZnO DOPED MAGESIUM ALUMIUM SILICATE GLASS-CERAMICS Chapter 7 103 CHAPTER 7 ELECTRICAL PROPERTIES OF ZnO DOPED MAGNESIUM ALUMINUM SILICATE GLASS-CERAMICS
Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival
 Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB
 Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
SKELETAL TISSUES CHAPTER 7 INTRODUCTION TO THE SKELETAL SYSTEM TYPES OF BONES
 SKELETAL TISSUES CHAPTER 7 By John McGill Supplement Outlines: Beth Wyatt Original PowerPoint: Jack Bagwell INTRODUCTION TO THE SKELETAL SYSTEM STRUCTURE Organs: Bones Related Tissues: Cartilage and Ligaments
SKELETAL TISSUES CHAPTER 7 By John McGill Supplement Outlines: Beth Wyatt Original PowerPoint: Jack Bagwell INTRODUCTION TO THE SKELETAL SYSTEM STRUCTURE Organs: Bones Related Tissues: Cartilage and Ligaments
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/2/12/e1601838/dc1 Supplementary Materials for General and programmable synthesis of hybrid liposome/metal nanoparticles Jin-Ho Lee, Yonghee Shin, Wooju Lee, Keumrai
advances.sciencemag.org/cgi/content/full/2/12/e1601838/dc1 Supplementary Materials for General and programmable synthesis of hybrid liposome/metal nanoparticles Jin-Ho Lee, Yonghee Shin, Wooju Lee, Keumrai
Electronic Supplementary Information
 Electronic Supplementary Material (ESI) for Chemical Communications. This journal is The Royal Society of Chemistry 2015 Electronic Supplementary Information Enzymatic Synthesis and Post-Functionalization
Electronic Supplementary Material (ESI) for Chemical Communications. This journal is The Royal Society of Chemistry 2015 Electronic Supplementary Information Enzymatic Synthesis and Post-Functionalization
Development of a near-infrared fluorescent probe for monitoring hydrazine in serum and living cells
 Supporting Information for Development of a near-infrared fluorescent probe for monitoring hydrazine in serum and living cells Sasa Zhu, Weiying Lin,* Lin Yuan State Key Laboratory of Chemo/Biosensing
Supporting Information for Development of a near-infrared fluorescent probe for monitoring hydrazine in serum and living cells Sasa Zhu, Weiying Lin,* Lin Yuan State Key Laboratory of Chemo/Biosensing
Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms
 Supporting Information for Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms Submitted by Yong Feng, Deli
Supporting Information for Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms Submitted by Yong Feng, Deli
Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury
 Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury Bastian OW, Koenderman L, Alblas J, Leenen LPH, Blokhuis TJ. Neutrophils contribute to
Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury Bastian OW, Koenderman L, Alblas J, Leenen LPH, Blokhuis TJ. Neutrophils contribute to
DELFIA Tb-N1 DTA Chelate & Terbium Standard
 AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
THE EFFECT OF FLUORINE UPON THE PHOSPHATASE CONTENT OF PLASMA, BONES, AND TEETH OF ALBINO RATS
 THE EFFECT OF FLUORINE UPON THE PHOSPHATASE CONTENT OF PLASMA, BONES, AND TEETH OF ALBINO RATS BY MARGARET CAMMACK SMITH AND EDITH M. LANTZ (From the Department oj Nutrition, Agricultural Experiment Station,
THE EFFECT OF FLUORINE UPON THE PHOSPHATASE CONTENT OF PLASMA, BONES, AND TEETH OF ALBINO RATS BY MARGARET CAMMACK SMITH AND EDITH M. LANTZ (From the Department oj Nutrition, Agricultural Experiment Station,
Optimal Differentiation of Tissue Types Using Combined Mid and Near Infrared Spectroscopy
 Optimal Differentiation of Tissue Types Using Combined Mid and Near Infrared Spectroscopy Mugdha V. Padalkar, M.S. 1, Cushla M. McGoverin, Ph.D. 1, Uday P. Palukuru, M.S. 1, Nicholas J. Caccese 1, Padraig
Optimal Differentiation of Tissue Types Using Combined Mid and Near Infrared Spectroscopy Mugdha V. Padalkar, M.S. 1, Cushla M. McGoverin, Ph.D. 1, Uday P. Palukuru, M.S. 1, Nicholas J. Caccese 1, Padraig
Organic Chemistry Diversity of Carbon Compounds
 Organic Chemistry Diversity of Carbon Compounds Hydrocarbons The Alkanes The Alkenes The Alkynes Naming Hydrocarbons Cyclic Hydrocarbons Alkyl Groups Aromatic Hydrocarbons Naming Complex Hydrocarbons Chemical
Organic Chemistry Diversity of Carbon Compounds Hydrocarbons The Alkanes The Alkenes The Alkynes Naming Hydrocarbons Cyclic Hydrocarbons Alkyl Groups Aromatic Hydrocarbons Naming Complex Hydrocarbons Chemical
Pressure Modulation of the Enzymatic Activity of. Phospholipase A2, a Putative Membraneassociated
 SUPPORTING INFORMATION Pressure Modulation of the Enzymatic Activity of Phospholipase A2, a Putative Membraneassociated Pressure Sensor Saba Suladze, Suleyman Cinar, Benjamin Sperlich, and Roland Winter*
SUPPORTING INFORMATION Pressure Modulation of the Enzymatic Activity of Phospholipase A2, a Putative Membraneassociated Pressure Sensor Saba Suladze, Suleyman Cinar, Benjamin Sperlich, and Roland Winter*
PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System
 Application Note LC/MS PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System Purpose This application note describes an automated workflow
Application Note LC/MS PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System Purpose This application note describes an automated workflow
Appendix A: Preparation of Media and Chemicals. Malt Extract Agar (MEA) weighing g was dissolved in 400 ml of distilled water
 weighing g was dissolved in 400 ml of distilled water") Appendix A: Preparation of Media and Chemicals Preparation of Malt Extract Agar (MEA) Malt Extract Agar (MEA) weighing 13.44 g was dissolved in 400 ml of distilled water in an Erlenmeyer flask using a
Appendix A: Preparation of Media and Chemicals Preparation of Malt Extract Agar (MEA) Malt Extract Agar (MEA) weighing 13.44 g was dissolved in 400 ml of distilled water in an Erlenmeyer flask using a
ab Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions.
 ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
Trypsin Mass Spectrometry Grade
 058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
CHAPTER 5 CHARACTERIZATION OF ZINC OXIDE NANO- PARTICLES
 88 CHAPTER 5 CHARACTERIZATION OF ZINC OXIDE NANO- PARTICLES 5.1 INTRODUCTION This chapter deals with the characterization of ZnO nano-particles using FTIR, XRD, PSA & SEM. The results analysis and interpretations
88 CHAPTER 5 CHARACTERIZATION OF ZINC OXIDE NANO- PARTICLES 5.1 INTRODUCTION This chapter deals with the characterization of ZnO nano-particles using FTIR, XRD, PSA & SEM. The results analysis and interpretations
SUPPLEMENTARY MATERIAL. Sample preparation for light microscopy
 SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Heparin Sodium ヘパリンナトリウム
 Heparin Sodium ヘパリンナトリウム Add the following next to Description: Identification Dissolve 1 mg each of Heparin Sodium and Heparin Sodium Reference Standard for physicochemical test in 1 ml of water, and
Heparin Sodium ヘパリンナトリウム Add the following next to Description: Identification Dissolve 1 mg each of Heparin Sodium and Heparin Sodium Reference Standard for physicochemical test in 1 ml of water, and
ab Human Citrate Synthase (CS) Activity Assay Kit
 Activity Assay Kit") ab119692 Human Citrate Synthase (CS) Activity Assay Kit Instructions for Use For the measurement of mitochondrial citrate synthase (CS) activity in Human samples This product is for research use only and
ab119692 Human Citrate Synthase (CS) Activity Assay Kit Instructions for Use For the measurement of mitochondrial citrate synthase (CS) activity in Human samples This product is for research use only and
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION DOI: 10.1038/NNANO.2012.80 Protein-Inorganic Hybrid Nanoflowers Jun Ge, Jiandu Lei, and Richard N. Zare Supporting Online Material Materials Proteins including albumin from bovine
SUPPLEMENTARY INFORMATION DOI: 10.1038/NNANO.2012.80 Protein-Inorganic Hybrid Nanoflowers Jun Ge, Jiandu Lei, and Richard N. Zare Supporting Online Material Materials Proteins including albumin from bovine
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric*
 SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
Skeletal System worksheet
 Skeletal System worksheet Name Section A: Intro to Skeletal System The skeletal system performs vital functions that enable us to move through our daily lives. Support - The skeleton provides support and
Skeletal System worksheet Name Section A: Intro to Skeletal System The skeletal system performs vital functions that enable us to move through our daily lives. Support - The skeleton provides support and
BabyBio IMAC columns DATA SHEET DS
 BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
Table S1b. TLC conditions for each drug (standard or in milk matrix) tested.
 tested.") List of Supporting Information Paragraph on SPE clean-up of OTET and THIA samples. Table S1a. Solvent conditions for compound storage and incubations. Table S1b. TLC conditions for each drug (standard
List of Supporting Information Paragraph on SPE clean-up of OTET and THIA samples. Table S1a. Solvent conditions for compound storage and incubations. Table S1b. TLC conditions for each drug (standard
Functions of the Skeletal System. Chapter 6: Osseous Tissue and Bone Structure. Classification of Bones. Bone Shapes
 Chapter 6: Osseous Tissue and Bone Structure Functions of the Skeletal System 1. Support 2. Storage of minerals (calcium) 3. Storage of lipids (yellow marrow) 4. Blood cell production (red marrow) 5. Protection
Chapter 6: Osseous Tissue and Bone Structure Functions of the Skeletal System 1. Support 2. Storage of minerals (calcium) 3. Storage of lipids (yellow marrow) 4. Blood cell production (red marrow) 5. Protection
OpenStax-CNX module: m Bone Structure * Ildar Yakhin. Based on Bone Structure by OpenStax. Abstract
 OpenStax-CNX module: m63474 1 Bone Structure * Ildar Yakhin Based on Bone Structure by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 4.0 By
OpenStax-CNX module: m63474 1 Bone Structure * Ildar Yakhin Based on Bone Structure by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 4.0 By
EFFECTS OF NICOTINE ON HUMAN MESENCHYMAL STEM CELLS. Connor McNeil Central Catholic HS
 EFFECTS OF NICOTINE ON HUMAN MESENCHYMAL STEM CELLS Connor McNeil Central Catholic HS Purpose To determine whether nicotine causes any effects on human Mesenchymal Stem Cell (hmsc) proliferation or migration
EFFECTS OF NICOTINE ON HUMAN MESENCHYMAL STEM CELLS Connor McNeil Central Catholic HS Purpose To determine whether nicotine causes any effects on human Mesenchymal Stem Cell (hmsc) proliferation or migration
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Biochemical Analysis of Plant Enzymes
 EDVOTEK P.O. Box 1232 West Bethesda, MD 20827-1232 The Biotechnology Biochemical Analysis of Plant Enzymes 904 EDVO-Kit # Storage: Store entire experiment in the refrigerator. Experiment Objective: In
EDVOTEK P.O. Box 1232 West Bethesda, MD 20827-1232 The Biotechnology Biochemical Analysis of Plant Enzymes 904 EDVO-Kit # Storage: Store entire experiment in the refrigerator. Experiment Objective: In
ARTESUNATE TABLETS: Final text for revision of The International Pharmacopoeia (December 2009) ARTESUNATI COMPRESSI ARTESUNATE TABLETS
 ARTESUNATI COMPRESSI ARTESUNATE TABLETS") December 2009 ARTESUNATE TABLETS: Final text for revision of The International Pharmacopoeia (December 2009) This monograph was adopted at the Forty-fourth WHO Expert Committee on Specifications for Pharmaceutical
December 2009 ARTESUNATE TABLETS: Final text for revision of The International Pharmacopoeia (December 2009) This monograph was adopted at the Forty-fourth WHO Expert Committee on Specifications for Pharmaceutical
OSSEOUS TISSUE & BONE STRUCTURE PART I: OVERVIEW & COMPONENTS
 OSSEOUS TISSUE & BONE STRUCTURE PART I: OVERVIEW & COMPONENTS The Skeletal System Skeletal system includes: bones of the skeleton, cartilages, ligaments, and connective tissues What are the functions of
OSSEOUS TISSUE & BONE STRUCTURE PART I: OVERVIEW & COMPONENTS The Skeletal System Skeletal system includes: bones of the skeleton, cartilages, ligaments, and connective tissues What are the functions of
TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
 Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
BIOH111. o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system
 BIOH111 o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 TEXTBOOK AND REQUIRED/RECOMMENDED
BIOH111 o Cell Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 TEXTBOOK AND REQUIRED/RECOMMENDED
Biology. Dr. Khalida Ibrahim
 Biology Dr. Khalida Ibrahim BONE TISSUE Bone tissue is a specialized form of connective tissue and is the main element of the skeletal tissues. It is composed of cells and an extracellular matrix in which
Biology Dr. Khalida Ibrahim BONE TISSUE Bone tissue is a specialized form of connective tissue and is the main element of the skeletal tissues. It is composed of cells and an extracellular matrix in which
Isolation and Molecular Characterization of Local Goat Milk Casein for Nutraceutical Value
 Isolation and Molecular Characterization of Local Goat Milk Casein for Nutraceutical Value Mohd Akmal Azhar 1,*, and Norshafiqa Salim 2 1 Faculty of Engineering Technology, UMP Gambang, Lebuhraya Tun Razak,
Isolation and Molecular Characterization of Local Goat Milk Casein for Nutraceutical Value Mohd Akmal Azhar 1,*, and Norshafiqa Salim 2 1 Faculty of Engineering Technology, UMP Gambang, Lebuhraya Tun Razak,
Homopolymers as Structure-Driving Agents in Semicrystalline Block Copolymer Micelles
 Supporting information for: Homopolymers as Structure-Driving Agents in Semicrystalline Block Copolymer Micelles Georgios Rizis, Theo G. M. van de Ven*, Adi Eisenberg* Department of Chemistry, McGill University,
Supporting information for: Homopolymers as Structure-Driving Agents in Semicrystalline Block Copolymer Micelles Georgios Rizis, Theo G. M. van de Ven*, Adi Eisenberg* Department of Chemistry, McGill University,
NOTES: Skeletal System (Ch 5, part 1)
") NOTES: Skeletal System (Ch 5, part 1) Individual bones are the organs of the skeletal system. A bone contains very active tissues. BONE STRUCTURE: *Bone structure reflects its function. Parts of a long
NOTES: Skeletal System (Ch 5, part 1) Individual bones are the organs of the skeletal system. A bone contains very active tissues. BONE STRUCTURE: *Bone structure reflects its function. Parts of a long
LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade
 AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
Calcineurin Cellular Activity Assay Kit, Colorimetric Cat. No
 User Protocol 207007 Rev. 27-July-04 JSW Page 1 of 9 Calcineurin Cellular Activity Assay Kit, Colorimetric Cat. No. 207007 Introduction Calcineurin (CaN) is a neuronal form of the widely distributed Ca
User Protocol 207007 Rev. 27-July-04 JSW Page 1 of 9 Calcineurin Cellular Activity Assay Kit, Colorimetric Cat. No. 207007 Introduction Calcineurin (CaN) is a neuronal form of the widely distributed Ca
Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine
 Agric. Biol. Chem., 46 (6), 1565~1569, 1982 1565 Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine Minoru Noda, Thanh Vo Van, Isao Kusakabe
Agric. Biol. Chem., 46 (6), 1565~1569, 1982 1565 Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine Minoru Noda, Thanh Vo Van, Isao Kusakabe
Due in Lab. Due next week in lab - Scientific America Article Select one article to read and complete article summary
 Due in Lab 1. Skeletal System 33-34 2. Skeletal System 26 3. PreLab 6 Due next week in lab - Scientific America Article Select one article to read and complete article summary Cell Defenses and the Sunshine
Due in Lab 1. Skeletal System 33-34 2. Skeletal System 26 3. PreLab 6 Due next week in lab - Scientific America Article Select one article to read and complete article summary Cell Defenses and the Sunshine
SalvinOss Xenograft Bone Graft Material In Vivo Testing Summary
 SalvinOss Xenograft Bone Graft Material In Vivo Testing Summary Summary of In Vivo Use Of Bioresorbable Xenograft Bone Graft Materials In The Treatment Of One-Walled Intrabony Defects In A Canine Model
SalvinOss Xenograft Bone Graft Material In Vivo Testing Summary Summary of In Vivo Use Of Bioresorbable Xenograft Bone Graft Materials In The Treatment Of One-Walled Intrabony Defects In A Canine Model
For more information about how to cite these materials visit
 Author(s): University of Michigan Medical School, Department of Cell and Developmental Biology License: Unless otherwise noted, the content of this course material is licensed under a Creative Commons
Author(s): University of Michigan Medical School, Department of Cell and Developmental Biology License: Unless otherwise noted, the content of this course material is licensed under a Creative Commons
Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus
 University of Groningen Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus Published in: Chemistry DOI: 10.1002/chem.201001816
University of Groningen Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus Published in: Chemistry DOI: 10.1002/chem.201001816