Acetylcholine again! - thought to be involved in learning and memory - thought to be involved dementia (Alzheimer's disease)
|
|
|
- Theresa Chambers
- 5 years ago
- Views:
Transcription
1 Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function Michael E. Hasselmo * and Bradley P. Wyble Acetylcholine again! - thought to be involved in learning and memory - thought to be involved dementia (Alzheimer's disease)
2 A few words on neuromodulation! Can it be defined? * Spatial distribution: neuromodulators often arise from brain nuclei that project widely to large numbers of brain regions * Time course of action: the actions of neuromodulators are often considered to be slower than those of classic neurotransmitters * Functionality: absence of presence of neuromodulators in given behavioral situations; modulation of existing neural function * Neuromodulators have a large variety of effects: they change intrinsic neural properties; modulate synaptic events; modulate learning and many others. * Some neurotransmitters, like GABA or Acetylcholine can be regarded as neurotransmitters or as neuromodulators depending on the nature of the receptors they act on.


3 Acetylcholine: arises from several nuclei each projecting to targeted brain areas Acts on nicotinic and muscarinic receptors Thought to be important for learning and attentional processing Examples: * in humans, the muscarinic antagonist scopolamine impairs list learning * in rats, certain memory tasks (watermaze, short term memory) are impaired by scopolamine * however, lesions of cholinergic nuceli can often not reproduce these data * loss of cholinergic innervation thought to be implicated in memory loss in Alzheimer s disease


4 Dopamine * Loss of dopaminergic neurons is associated with Parkinson disease * Increase of dopaminergic activity can be associated with Schizophrenia * Activity of dopaminergic neurons in VTA has been shown to increase during behaviorally relevant stimuli, and are thought to be important for reward associations

5 Noradrenaline (NA) * NA neurons in the locus coerulus project all over the brain * NA has been associated with signal-to-noise ratio and signal-detection capabilities
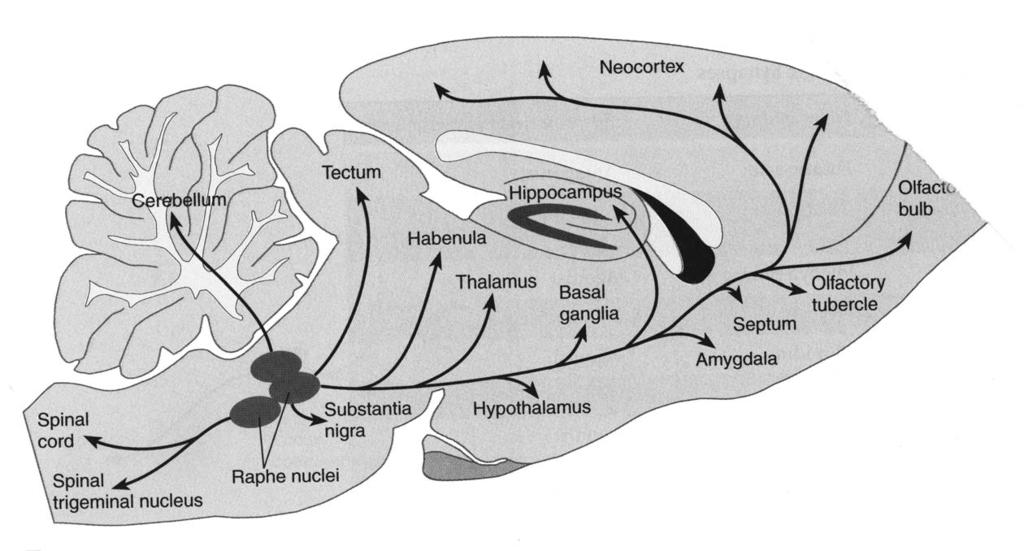

6 Serotonine
 NA present NA")
7 Example: Modulation of signal-to-noise ratio (noradrenaline) NA present NA absent
8 Some commonly observed neuromodulatory effects: * Cholinergic agonists can evoke oscillatory activity in hippocampal slices * Oscillatory activity in the hippocampus of behaving rats depends on cholinergic inputs * Pyramidal cells in hippocampus and cortex are often depolarized by ACh and NA * Synaptic potentials can be modulated (increased or decreased) by ACh or NA * Long term potentiation is modulated by ACh and NA and many others.. * Rats are impaired in long-term and short term memory experiments when certain neuromodulatory effects are blocked * Rats show attentioanl deficits when cholinergic modulation is decreased etc
9 * Most models of human memory function are interpretive they help us understand behavioral data and guide behavioral experiments * The model presented here is mechanistic directly addressing the physiological and anatomical substrates of performance in human memory tasks such as free recall and recognition Adresses these questions by: (1) by simulating specific human memory tasks, such as free recall and recognition (2) by addressing a current issue in human memory modeling the list strength effect (3) by explicitly modeling the effect of the cholinergic antagonist scopolamine on human memory function, and (4) by generating an experimentally testable prediction about the effect of scopolamine on paired associate learning.
10 Why model the effects of drugs? Modeling drug effects on memory function allow us to link effects at a cellular level to effects at a behavioral level
11 Why model the effects of drugs? To test any model, you need to perturb it! One perturbation that can be done in humans is a change in neural and synaptic function due to drugs. You have to: 1) have some idea of how the brain function you are looking at is implemented by a neural circuit 2) know the effect of your drug in that neural circuit 3) be able to correlate the function of your circuit with experimental observations
12 Example we have talked about before Stimulus Behavioral response 1) Observable behavior: gill withdrawal after siphon touch
13 Example we have talked about before Stimulus Behavioral response 1) Observable behavior: gill withdrawal after siphon touch Stimulus 2) Neural circuit underlying the behavior has been identified Behavioral response SN MN
14 Example we have talked about before Stimulus Behavioral response 1) Observable behavior: gill withdrawal after siphon touch Serotonine Stimulus 2) Neural circuit underlying the behavior has been identified Behavioral response SN 3) Manipulation of neural circuit leads to observable change in behavior MN
 Neural circuit underlying the behavior has been identified Behavioral")
 and the")
15 Example we have talked about before Stimulus Behavioral response 1) Observable behavior: gill withdrawal after siphon touch Serotonine Stimulus 2) Neural circuit underlying the behavior has been identified Behavioral response SN MN 3) Manipulation of neural circuit leads to observable change in behavior The manipulation can be used to test how well established the relationship between the neural circuit (or a model thereof) and the behavior is.
16 Acetylcholine acts on two types of receptors in the brain: muscarininc and nicotinic. Scopolamine is a muscarinic antagonist This means that when present, scopolamine binds to the muscarinic receptor without activating it. It prevents the binding of acetylcholine to the receptor and thus the activation of the receptor.
17 Table Car Plant Flower Buddy Hammer Chair...
18 Table Car Plant Flower Buddy Hammer Chair... Prevents ACh from activating muscarinic receptors
19 Table Car Plant Flower Buddy Hammer Chair. List words. on list. Prevents ACh from activating muscarinic receptors Recognize if words were on list
20 Table Car Plant Flower Buddy Hammer Chair. List words. on list. Bed Apple Nail Friend... Prevents ACh from activating muscarinic receptors Recognize if words were on list
21 List words on list Recognize if words were on list Table Car Plant Flower Buddy Hammer Chair. List words. on list. Bed Apple Nail Friend... Prevents ACh from activating muscarinic receptors Recognize if words were on list

22 /or saline control Effect on recall.recognition Effect on learning
23 Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Less feedback excitation (association fibers blocked)
24 Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Pyramidal cells more easily excitable because they are closer to threshold Vm time ACh
25 Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Pyramidal cells spike more when activated time Current injection ACh
26 Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Synaptic plasticity enhanced learning rate is increased
27 Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Overall: 1) Afferent or outside inputs dominate in the presence of ACh 2) Cells are more excitable and respond with more action potential to afferent input 3) Plasticity between pyramidal cells is enhanced
28 Scopalamine prevents all these effects! Cellular effects of ACh in the hippocampus 1) Suppresses synaptic transmission 2) Depolarizes pyramidal cells 3) Suppresses neuronal adaptation 4) Enhances LTP Overall: 1) Afferent or outside inputs dominate in the presence of ACh 2) Cells are more excitable and respond with more action potential to afferent input 3) Plasticity between pyramidal cells is enhanced
29 Effects of scopolamine administration in this model
30 Hippocampal anatomy
31 Hippocampal anatomy and model
32 Hippocampal anatomy and model Medial septum: provides ACh to hippocampus Autoassociative network: stores patterns Self-organization: external input imposed only on input layer (dentate gyrus) but not on target layer (CA3) hetero-associative: association between two patterns (CA1 and CA3)
33 Entorhinal cortex Input pattern (1,0,0,0,1,1,1) Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) Lets look at the details slowly self-organization
34 Entorhinal cortex Input pattern (1,0,0,0,1,1,1) Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) Lets look at the details slowly self-organization CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) hebbian learning rule autoassociation
35 Entorhinal cortex Input pattern (1,0,0,0,1,1,1) Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) Lets look at the details slowly self-organization CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) hebbian learning rule autoassociation association
 CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association")
36 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization
37 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) accompanied by inhibition creates sparse representation in DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization
38 1) Neurons in layer 2 are activated by layer neurons and random synaptic weights Layer 1 Small, random synaptic weights Layer 2 X L2 (i) = Σ w ij *X L1 (j) 2) Inhibitory neuron is activated by all neurons in layer 2 3) Inhibitory neuron inhibits all neurons in layer 2 Inhibitory neuron Layer 2 Inhibitory neuron X IN = 1/N*Σ X L2 (j) X L2 (i) = Σ w ij *X L1 (j) X IN 4) Hebbian learning rule strengthens weights between active layer 1 and layer 2 neurons Layer 1 Strengthened synaptic weights Layer 2 w ij = w ij + X L2 (j)*x L1 (i) 5) Layer 2 neuron responds strongly to layer 1 input Layer 1 Layer 2
39 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) accompanied by inhibition creates sparse representation in DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization stores association between elements of the same pattern
40 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) accompanied by inhibition creates sparse representation in DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization stores association between elements of the same pattern accompanied by inhibition creates sparse representation in CA1
41 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) accompanied by inhibition creates sparse representation in DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization stores association between elements of the same pattern stores association between the pattern in CA3 and the pattern in CA1 accompanied by inhibition creates sparse representation in CA1
42 Lets look at the details slowly Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) accompanied by inhibition creates sparse representation in DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern stores association between elements of the same pattern stores association between the pattern in CA3 and the pattern in CA1 accompanied by inhibition creates sparse representation in CA1
43 Incomplete input pattern Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) existing synaptic weights transmit pattern to DG CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern association fibers recall previously stored pattern CA1 activates completed output pattern in EC
44 How does the model know when to learn??? Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern When input pattern from CA3 and pattern in CA1 (from EC) MATCH the previously stored association, activity level in CA1 is high!
45 How does the model know when to learn??? Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern When input pattern from CA3 and pattern in CA1 (from EC) DON'T match the previously stored association, activity level in CA1 is low!
46 How does the model know when to learn??? Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern inhibitory ACh
47 How does the model know when to learn??? Entorhinal cortex Input pattern (1,0,0,0,1,1,1) self-organization Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) Non-match: ACh neurons have high output: - neurons depolarized - LTP high - association fiber synaptic transmission supressed - learning is ON CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern NO MATCH inhibitory ACh
48 How does the model know when to learn??? Entorhinal cortex Input pattern (1,0,0,0,1,1,1) Dentate gyrus Self-organized pattern (0,0,1,0,0,1,0) CA3 autoassociation of pattern transmitted by CA3 (0,0,1,0,0,1,0) self-organization CA1 comparison between input imposed by EC and pattern in CA3 (0,0,1,0,0,0,1) Entorhinal cortex Input pattern (1,0,0,0,1,1,1) hebbian learning rule autoassociation association self-organization EC: Output pattern recall MATCH Match: ACh neurons have low output: - neurons not depolarized -LTP low - association fiber synaptic transmission not suppressed: recall mode Learning is OFF inhibitory ACh
49 What is being modeled? List learning in humans
50 What is being modeled? List learning in humans neural activation pattern
51 What is being modeled? List learning in humans Context Item neural activation pattern
52 Free recall: input to model is context and model recalls items. This corresponds to telling subject to recall items from a give list Context Item neural activation pattern
53 Recognition: input to model is item and model recalls context. This corresponds to asking subject which list a given item belonged to. Context Item neural activation pattern
54 external input A i association fibers w ij pyramidal cells with activation a i single inhibitory neuron which inhibits all pyramidal neurons in proportion to the average activity
55 external input A i association fibers w ij pyramidal cells with activation a i single inhibitory neuron which inhibits all pyramidal neurons in proportion to the average activity associative memory in region CA3 stores associations between context and items as well as associations between elements of each. Associative weights between context units are stronger because context is repeatedly presented!
 external input")

56 Equations! change in activation (what we call membrane potential) external input excitatory synaptic inputs from other cells in same layer previous activation inhibitory synaptic inputs models adaptation as a function of calcium influx calcium influx as a function of activation
57 More equations! change in synaptic weight slow change in postsynaptic activity slow change in presynaptic activity postsynaptic activity slow change in postsynaptic activity scaling factor threshold slow decay slow change in presynaptic activity presynaptic activity
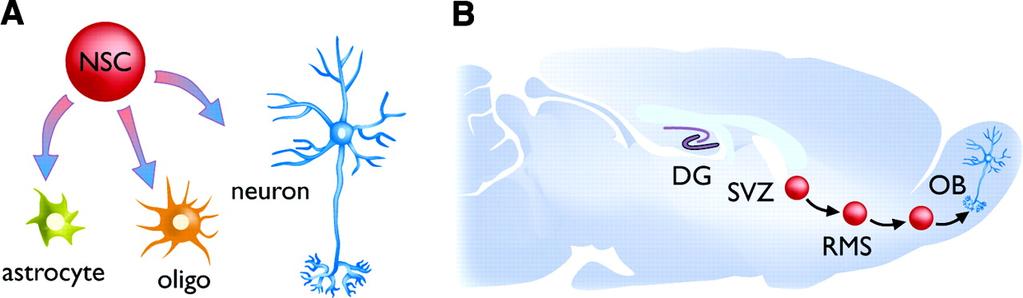
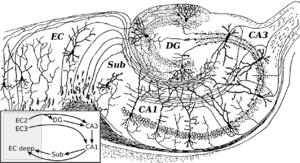
58 More equations! change in synaptic weight slow change in postsynaptic activity slow change in presynaptic activity ACh modulation learning threshold for postsynaptic activity learning threshold for presynaptic activity learning rate
59 More equations! decay of synaptic strength necessary for self-organization learning threshold for postsynaptic activity learning threshold for presynaptic activity learning rate high in CA3 for auto-associative learning low in DG to allow self-organization
60 learning ruled allowed both associative learning and self-organization. for self-organization, plasticity of inhibitory connections was also necessary for self-organization, learning thresholds were lower (because NO external input activates second layer of neurons, thus, activations will be lower)
61
62 EC DG CA3 CA1
63 EC DG CA3 CA1

64 EC II 2 DG CA3 EC IV CA1 EC II
65 EC II DG CA3 EC IV CA1 EC II
66
67 A word on recalling multiple patterns sequentially! context presented - recall pattern 1 - recall pattern 3 - recall pattern 2 - recall pattern 5 -- each pattern has formed an attractor! We learned that once a network has reached an attractor, it cannot move out of it. So how does this work then?
68 1) Context presented 2) Context neurons activate many item neurons 3) because of differences in synaptic weights (due to random initial conditions), one pattern is more activated than others. Global inhibition deactivates neurons belonging to other patterns. 4) spike-adaptation gradually reduces activity of the recalled item, and a different item can be recalled because it is freed from inhibition
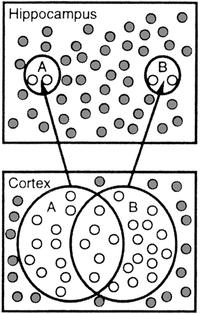
69 Stress, memory and neurogenesis Computational modeling and empirical studies of hippocampal neurogenesis-dependent memory: Effects of interference, stress and depression. Becker S, Macqueen G, Wojtowicz JM.
70 In the dentate gyrus of the young adult rat hippocampus, for example, there are about 1,000,000 granule cells, and about 10,000 new neurons are generated per day, of which about 40% survive to maturation (McDonald and Wojtowicz, 2005). The dentate gyrus is critically situated within the so-called trisynaptic circuit in the hippocampus (see Fig. 1), so that information flows from the rest of the brain, via the entorhinal cortex (EC), through the dentate gyrus and CA3/CA1 subregions. On the other hand, there are also short-circuit connections from the EC to the CA3/CA1 by-passing the dentate gyrus. Thus not all forms of learning and memory may be dependent upon the dentate gyrus, and hence dependent upon neurogenesis. Indeed, empirical evidence from studies using selective ablation and/or genetic manipulation of the dentate gyrus supports a role for the this structure, and hence for the entire trisynaptic circuit, in rapid learning of novel contexts (Nakashiba et al., 2008) and in maintaining distinct representations of similar events such as nearby spatial representations (Gilbert, Kesner and Lee, 2001), while the short-circuit pathways bypassing the dentate gyrus are sufficient to support paired associates learning and incremental spatial learning ( [Nakashiba et al., 2008] and [Gilbert and Kesner, 2003]). Thus, the role of neurogenesis is most likely tied to the dentate-gyrus specific learning and memory functions.
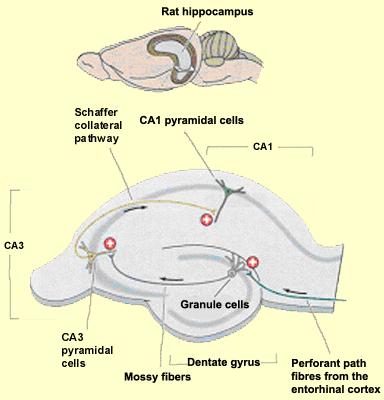
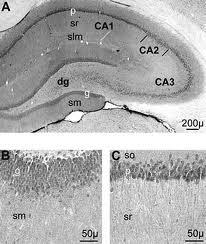
71
72
73
74
75
76
77
78 Nottebohm (2002) suggested that the newly born neurons may be recruited preferentially for storing new memories, thereby protecting old memories from interference. Consistent with this hypothesis,wiskott, Rasch and Kempermann (2006) demonstrated in a simple neural network model that the addition of highly plastic new neurons does effectively prevent new learning from interfering catastrophically with older memories. Conversely, Feng et al. (2001) proposed that neurogenesis is important for clearing out older memories once they are consolidated, and several modelers have demonstrated in abstract neural network models that neuronal turnover improves acquisition by helping to discard older memories

79
80 Spatial learning in the Morris water maze is disrupted by hippocampal lesions (Morris et al., 1990) but not by irradiation (Snyder et al. 2005; Wojtowicz et al., 2008). While irradiated animals learn the water maze at a normal rate, their long-term memory retention of the hidden platform location is greatly impaired relative to controls when they are re-tested four or more weeks later (Snyder et al., 2005). Animals lacking new hippocampal neurons show deficits on trace conditioning (Shors et al., 2001), contextual fear conditioning and delayed non-match to sample (DNMS) at long delays (Winocur et al., 2006; Wojtowicz et al., 2008), while performing normally on corresponding non-hippocampal control tasks, delay conditioning (Shors et al., 2001), cued fear conditioning and DNMS at short delays (Winocur et al., 2006), respectively. Moreover, voluntary running enhances neurogenesis in rodents (van Praag et al., 1999) and correlates weakly with improved performance on contextual fear conditioning (Wojtowicz et al., 2008).
81 These include stress-related reduction in neurogenesis (e.g. Gould et al., 1998; McEwen and Magarinos, 2001), and exercise and environmental enrichment-related increases in neurogenesis (van Praag et al., 1999; Nilsson et al.,1999; Olson et al., 2006; Pereira et al., 2007). In summary, we predict two important functions of hippocampal neurogenesis, namely, (1) to create distinct representations of similar events, thereby minimizing interference between highly similar memories and between memories acquired at different periods of time, and (2) to create a gradually evolving representation of spatial, temporal and other contextual cues that serves to bind together elements of a memory into coherent episodic memory traces.
82
83 The learning equations implement a form of simple Hebbian learning in the perforant path (ECto dentate gyrus, EC-to-CA3 and EC-to-CA1 connections), heteroassociative Hebbian learning in the CA3-to-CA1 connections and temporal associative learning in the CA3 recurrent collaterals. The model is built upon several key assumptions regarding hippocampal coding: (1) there is sparse coding (low activity levels) in all regions; (2) the projection pathway from the DG to CA3 (mossy fiber pathway) is strong (very high synaptic strengths) and sparse (very low probability of connectivity); (3) the CA3 pyramidals are highly interconnected via recurrent collaterals; (4) the trisynaptic pathway involving the dentate gyrus participates in encoding, not recall; and (5) the CA3 collaterals are active during recall but not encoding.
84 1. During encoding, dentate granule cells are active whereas during retrieval they are relatively silent. 2. During encoding, activation of CA3 pyramidals is dominated by the very strong mossy fiber inputs from dentate granule cells. 3. During retrieval, activation of CA3 pyramidals is driven by direct perforant path inputs from the EC combined with timedelayed input from CA3 via recurrent collaterals. 4. During encoding, activation of CA1 pyramidals is dominated by direct perforant path inputs from the EC. 5. During retrieval, CA1 activations are driven by a combination of perforant path inputs from the EC and Shaffer collateral inputs from CA3.
85 New neurons are added only to the DG, and the DG drives activation in the hippocampal circuit only during encoding, not during retrieval. Thus, the new neurons contribute to the formation of distinctive codes for novel events, but not to the associative retrieval of older memories.
86 Paired Associate learning: the learning of syllables, digits, or words in pairs (as in the study of a foreign language) so that one member of the pair evokes recall of the other Each model was trained on a set of 10 paired associates consisting of randomly generated patterns on Day 1, followed by a simulated retention interval of 4 weeks, and then a final cued recall test of the original paired associates. During the retention interval, new unrelated items are learned, a potential source of interference with the original paired associates. 10 pairs Delay with novel pairs Test on original pairs
87 Each of the models simulated had the following architecture: 200 input (EC) neurons, 1000 dentate gyrus neurons, 300 CA3 neurons and 400 CA1 neurons. 200 Twenty repetitions of each model were run, to generate 1000 results of 20 simulated subjects. Three different versions of the model were compared: (1) no neuronal turnover; (2) neural turnover and no preferential bias toward new neurons during encoding; i.e. every neuron in the dentate layer has an equal chance of becoming active when a novel pattern is to be encoded; (3) a model with neural turnover and preferential recruitment of new neurons for new memory formation; that is, new neurons were more likely to be selected for activation when a new memory was to be stored Each model was first trained on a set of paired associates, and then on subsequent weeks, the passage of time and consequent decay of old memories was simulated by exposing the model to a set of random, unrelated items. On each week, the model was then tested for retention of the original set of paired associates. For each of the three models, the rate of neuronal turnover was
88 Neuronal turnover was simulated by randomly selecting a fixed percentage of the dentate layer neurons, and rerandomizing their incoming weights from the EC, and reconnecting them randomly to a different subset of CA3 cells. The percentage of new neurons created in this manner was either 0, 25, 50, 75, or 100.
89
Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function
 Behavioural Brain Research 89 (1997) 1 34 Review article Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function Michael E. Hasselmo
Behavioural Brain Research 89 (1997) 1 34 Review article Free recall and recognition in a network model of the hippocampus: simulating effects of scopolamine on human memory function Michael E. Hasselmo
COGNITIVE SCIENCE 107A. Hippocampus. Jaime A. Pineda, Ph.D.
 COGNITIVE SCIENCE 107A Hippocampus Jaime A. Pineda, Ph.D. Common (Distributed) Model of Memory Processes Time Course of Memory Processes Long Term Memory DECLARATIVE NON-DECLARATIVE Semantic Episodic Skills
COGNITIVE SCIENCE 107A Hippocampus Jaime A. Pineda, Ph.D. Common (Distributed) Model of Memory Processes Time Course of Memory Processes Long Term Memory DECLARATIVE NON-DECLARATIVE Semantic Episodic Skills
Modeling of Hippocampal Behavior
 Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
Modeling of Hippocampal Behavior Diana Ponce-Morado, Venmathi Gunasekaran and Varsha Vijayan Abstract The hippocampus is identified as an important structure in the cerebral cortex of mammals for forming
Lisa M. Giocomo & Michael E. Hasselmo
 Mol Neurobiol (2007) 36:184 200 DOI 10.1007/s12035-007-0032-z Neuromodulation by Glutamate and Acetylcholine can Change Circuit Dynamics by Regulating the Relative Influence of Afferent Input and Excitatory
Mol Neurobiol (2007) 36:184 200 DOI 10.1007/s12035-007-0032-z Neuromodulation by Glutamate and Acetylcholine can Change Circuit Dynamics by Regulating the Relative Influence of Afferent Input and Excitatory
Why do we have a hippocampus? Short-term memory and consolidation
 Why do we have a hippocampus? Short-term memory and consolidation So far we have talked about the hippocampus and: -coding of spatial locations in rats -declarative (explicit) memory -experimental evidence
Why do we have a hippocampus? Short-term memory and consolidation So far we have talked about the hippocampus and: -coding of spatial locations in rats -declarative (explicit) memory -experimental evidence
Ch 8. Learning and Memory
 Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Ch 8. Learning and Memory
 Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory
 Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory Anna Smith Outline 1. 2. 3. 4. 5. Background in Memory Models Models of Consolidation The Hippocampus Competitive Trace Theory
Prior Knowledge and Memory Consolidation Expanding Competitive Trace Theory Anna Smith Outline 1. 2. 3. 4. 5. Background in Memory Models Models of Consolidation The Hippocampus Competitive Trace Theory
Memory Systems II How Stored: Engram and LTP. Reading: BCP Chapter 25
 Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
CRISP: Challenging the Standard Framework of Hippocampal Memory Function
 CRISP: Challenging the Standard Framework of Hippocampal Memory Function Laurenz Wiskott Mehdi Bayati Sen Cheng Jan Melchior Torsten Neher supported by: Deutsche Forschungsgemeinschaft - Sonderforschungsbereich
CRISP: Challenging the Standard Framework of Hippocampal Memory Function Laurenz Wiskott Mehdi Bayati Sen Cheng Jan Melchior Torsten Neher supported by: Deutsche Forschungsgemeinschaft - Sonderforschungsbereich
Psychology 320: Topics in Physiological Psychology Lecture Exam 2: March 19th, 2003
 Psychology 320: Topics in Physiological Psychology Lecture Exam 2: March 19th, 2003 Name: Student #: BEFORE YOU BEGIN!!! 1) Count the number of pages in your exam. The exam is 8 pages long; if you do not
Psychology 320: Topics in Physiological Psychology Lecture Exam 2: March 19th, 2003 Name: Student #: BEFORE YOU BEGIN!!! 1) Count the number of pages in your exam. The exam is 8 pages long; if you do not
Theories of memory. Memory & brain Cellular bases of learning & memory. Epileptic patient Temporal lobectomy Amnesia
 Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Theories of Sensory, short-term & long-term memories Memory & brain Cellular bases
Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Theories of Sensory, short-term & long-term memories Memory & brain Cellular bases
Encoding Spatial Context: A Hypothesis on the Function of the Dentate Gyrus-Hilus System
 Encoding Spatial Context: A Hypothesis on the Function of the Dentate Gyrus-Hilus System A. Minai, ECECS Department, University of Cincinnati, Cincinnati, OH J. Best, Department of Psychology, Miami University,
Encoding Spatial Context: A Hypothesis on the Function of the Dentate Gyrus-Hilus System A. Minai, ECECS Department, University of Cincinnati, Cincinnati, OH J. Best, Department of Psychology, Miami University,
Basics of Computational Neuroscience: Neurons and Synapses to Networks
 Basics of Computational Neuroscience: Neurons and Synapses to Networks Bruce Graham Mathematics School of Natural Sciences University of Stirling Scotland, U.K. Useful Book Authors: David Sterratt, Bruce
Basics of Computational Neuroscience: Neurons and Synapses to Networks Bruce Graham Mathematics School of Natural Sciences University of Stirling Scotland, U.K. Useful Book Authors: David Sterratt, Bruce
CASE 49. What type of memory is available for conscious retrieval? Which part of the brain stores semantic (factual) memories?
 memories?") CASE 49 A 43-year-old woman is brought to her primary care physician by her family because of concerns about her forgetfulness. The patient has a history of Down syndrome but no other medical problems.
CASE 49 A 43-year-old woman is brought to her primary care physician by her family because of concerns about her forgetfulness. The patient has a history of Down syndrome but no other medical problems.
Memory retention the synaptic stability versus plasticity dilemma
 Memory retention the synaptic stability versus plasticity dilemma Paper: Abraham, Wickliffe C., and Anthony Robins. "Memory retention the synaptic stability versus plasticity dilemma." Trends in neurosciences
Memory retention the synaptic stability versus plasticity dilemma Paper: Abraham, Wickliffe C., and Anthony Robins. "Memory retention the synaptic stability versus plasticity dilemma." Trends in neurosciences
Cerebral Cortex. Edmund T. Rolls. Principles of Operation. Presubiculum. Subiculum F S D. Neocortex. PHG & Perirhinal. CA1 Fornix CA3 S D
 Cerebral Cortex Principles of Operation Edmund T. Rolls F S D Neocortex S D PHG & Perirhinal 2 3 5 pp Ento rhinal DG Subiculum Presubiculum mf CA3 CA1 Fornix Appendix 4 Simulation software for neuronal
Cerebral Cortex Principles of Operation Edmund T. Rolls F S D Neocortex S D PHG & Perirhinal 2 3 5 pp Ento rhinal DG Subiculum Presubiculum mf CA3 CA1 Fornix Appendix 4 Simulation software for neuronal
The storage and recall of memories in the hippocampo-cortical system. Supplementary material. Edmund T Rolls
 The storage and recall of memories in the hippocampo-cortical system Supplementary material Edmund T Rolls Oxford Centre for Computational Neuroscience, Oxford, England and University of Warwick, Department
The storage and recall of memories in the hippocampo-cortical system Supplementary material Edmund T Rolls Oxford Centre for Computational Neuroscience, Oxford, England and University of Warwick, Department
Synapse. Structure & Function. Neurotransmitter Sequence. Integration. History: 10/4/12 original version
 Synapse History: 10/4/12 original version Structure & Function (This content is covered in Sinjin's presentation, see link in calendar) Neurotransmitters Synaptic cleft Post-synaptic potential Excitation
Synapse History: 10/4/12 original version Structure & Function (This content is covered in Sinjin's presentation, see link in calendar) Neurotransmitters Synaptic cleft Post-synaptic potential Excitation
A model of the interaction between mood and memory
 INSTITUTE OF PHYSICS PUBLISHING NETWORK: COMPUTATION IN NEURAL SYSTEMS Network: Comput. Neural Syst. 12 (2001) 89 109 www.iop.org/journals/ne PII: S0954-898X(01)22487-7 A model of the interaction between
INSTITUTE OF PHYSICS PUBLISHING NETWORK: COMPUTATION IN NEURAL SYSTEMS Network: Comput. Neural Syst. 12 (2001) 89 109 www.iop.org/journals/ne PII: S0954-898X(01)22487-7 A model of the interaction between
Rolls,E.T. (2016) Cerebral Cortex: Principles of Operation. Oxford University Press.
 Cerebral Cortex: Principles of Operation. Oxford University Press.") Digital Signal Processing and the Brain Is the brain a digital signal processor? Digital vs continuous signals Digital signals involve streams of binary encoded numbers The brain uses digital, all or none,
Digital Signal Processing and the Brain Is the brain a digital signal processor? Digital vs continuous signals Digital signals involve streams of binary encoded numbers The brain uses digital, all or none,
Navigation: Inside the Hippocampus
 9.912 Computational Visual Cognition Navigation: Inside the Hippocampus Jakob Voigts 3 Nov 2008 Functions of the Hippocampus: Memory and Space Short term memory Orientation Memory consolidation(h.m)
9.912 Computational Visual Cognition Navigation: Inside the Hippocampus Jakob Voigts 3 Nov 2008 Functions of the Hippocampus: Memory and Space Short term memory Orientation Memory consolidation(h.m)
Memory, Attention, and Decision-Making
 Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
Systems Neuroscience CISC 3250
 Systems Neuroscience CISC 325 Memory Types of Memory Declarative Non-declarative Episodic Semantic Professor Daniel Leeds dleeds@fordham.edu JMH 328A Hippocampus (MTL) Cerebral cortex Basal ganglia Motor
Systems Neuroscience CISC 325 Memory Types of Memory Declarative Non-declarative Episodic Semantic Professor Daniel Leeds dleeds@fordham.edu JMH 328A Hippocampus (MTL) Cerebral cortex Basal ganglia Motor
Chapter 5: Learning and Behavior Learning How Learning is Studied Ivan Pavlov Edward Thorndike eliciting stimulus emitted
 Chapter 5: Learning and Behavior A. Learning-long lasting changes in the environmental guidance of behavior as a result of experience B. Learning emphasizes the fact that individual environments also play
Chapter 5: Learning and Behavior A. Learning-long lasting changes in the environmental guidance of behavior as a result of experience B. Learning emphasizes the fact that individual environments also play
NERVOUS SYSTEM 1 CHAPTER 10 BIO 211: ANATOMY & PHYSIOLOGY I
 BIO 211: ANATOMY & PHYSIOLOGY I 1 Ch 10 A Ch 10 B This set CHAPTER 10 NERVOUS SYSTEM 1 BASIC STRUCTURE and FUNCTION Dr. Lawrence G. Altman www.lawrencegaltman.com Some illustrations are courtesy of McGraw-Hill.
BIO 211: ANATOMY & PHYSIOLOGY I 1 Ch 10 A Ch 10 B This set CHAPTER 10 NERVOUS SYSTEM 1 BASIC STRUCTURE and FUNCTION Dr. Lawrence G. Altman www.lawrencegaltman.com Some illustrations are courtesy of McGraw-Hill.
Reversed and forward buffering of behavioral spike sequences enables retrospective and prospective retrieval in hippocampal regions CA3 and CA1
 Neural Networks 21 (2008) 276 288 www.elsevier.com/locate/neunet 2008 Special Issue Reversed and forward buffering of behavioral spike sequences enables retrospective and prospective retrieval in hippocampal
Neural Networks 21 (2008) 276 288 www.elsevier.com/locate/neunet 2008 Special Issue Reversed and forward buffering of behavioral spike sequences enables retrospective and prospective retrieval in hippocampal
More dendritic spines, changes in shapes of dendritic spines More NT released by presynaptic membrane
 LEARNING AND MEMORY (p.1) You are your learning and memory! (see movie Total Recall) L&M, two sides of the same coin learning refers more to the acquisition of new information & brain circuits (storage)
LEARNING AND MEMORY (p.1) You are your learning and memory! (see movie Total Recall) L&M, two sides of the same coin learning refers more to the acquisition of new information & brain circuits (storage)
Synaptic plasticityhippocampus. Neur 8790 Topics in Neuroscience: Neuroplasticity. Outline. Synaptic plasticity hypothesis
 Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
Research Article Associative Memory Storage and Retrieval: Involvement of Theta Oscillations in Hippocampal Information Processing
 Neural Plasticity Volume 2011, Article ID 683961, 15 pages doi:10.1155/2011/683961 Research Article Associative Memory Storage and Retrieval: Involvement of Theta Oscillations in Hippocampal Information
Neural Plasticity Volume 2011, Article ID 683961, 15 pages doi:10.1155/2011/683961 Research Article Associative Memory Storage and Retrieval: Involvement of Theta Oscillations in Hippocampal Information
Cogs 107b Systems Neuroscience lec9_ neuromodulators and drugs of abuse principle of the week: functional anatomy
 Cogs 107b Systems Neuroscience www.dnitz.com lec9_02042010 neuromodulators and drugs of abuse principle of the week: functional anatomy Professor Nitz circa 1986 neurotransmitters: mediating information
Cogs 107b Systems Neuroscience www.dnitz.com lec9_02042010 neuromodulators and drugs of abuse principle of the week: functional anatomy Professor Nitz circa 1986 neurotransmitters: mediating information
Memory: Computation, Genetics, Physiology, and Behavior. James L. McClelland Stanford University
 Memory: Computation, Genetics, Physiology, and Behavior James L. McClelland Stanford University A Playwright s Take on Memory What interests me a great deal is the mistiness of the past Harold Pinter,
Memory: Computation, Genetics, Physiology, and Behavior James L. McClelland Stanford University A Playwright s Take on Memory What interests me a great deal is the mistiness of the past Harold Pinter,
9.01 Introduction to Neuroscience Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Working memory short-term
MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Working memory short-term
Chapter 6: Hippocampal Function In Cognition. From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D.
 Chapter 6: Hippocampal Function In Cognition From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Grid Cell The Hippocampus Serves a Role in Multimodal Information Processing and Memory
Chapter 6: Hippocampal Function In Cognition From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Grid Cell The Hippocampus Serves a Role in Multimodal Information Processing and Memory
Synaptic plasticity and hippocampal memory
 Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
Synaptic plasticity and hippocampal memory Tobias Bast School of Psychology, University of Nottingham tobias.bast@nottingham.ac.uk Synaptic plasticity as the neurophysiological substrate of learning Hebb
Different inhibitory effects by dopaminergic modulation and global suppression of activity
 Different inhibitory effects by dopaminergic modulation and global suppression of activity Takuji Hayashi Department of Applied Physics Tokyo University of Science Osamu Araki Department of Applied Physics
Different inhibitory effects by dopaminergic modulation and global suppression of activity Takuji Hayashi Department of Applied Physics Tokyo University of Science Osamu Araki Department of Applied Physics
Notes: Synapse. Overview. PSYC Summer Professor Claffey PDF. Conversion from an signal to a signal - electrical signal is the
 PSYC 170 - Summer 2013 - Professor Claffey Notes: Synapse PDF Overview Conversion from an signal to a signal - electrical signal is the - chemical signal is the Presynaptic - refers to that sends/receives
PSYC 170 - Summer 2013 - Professor Claffey Notes: Synapse PDF Overview Conversion from an signal to a signal - electrical signal is the - chemical signal is the Presynaptic - refers to that sends/receives
Summarized by. Biointelligence Laboratory, Seoul National University
 Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 3 rd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2008. Summarized by H.-S. Seok, K. Kim, and db.-t. TZhang Biointelligence
Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 3 rd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2008. Summarized by H.-S. Seok, K. Kim, and db.-t. TZhang Biointelligence
UNINFORMATIVE MEMORIES WILL PREVAIL
 virtute UNINFORMATIVE MEMORIES WILL PREVAIL THE STORAGE OF CORRELATED REPRESENTATIONS AND ITS CONSEQUENCES Emilio Kropff SISSA, Trieste di Studi Avanzati Internazionale Superiore - e conoscenza seguir
virtute UNINFORMATIVE MEMORIES WILL PREVAIL THE STORAGE OF CORRELATED REPRESENTATIONS AND ITS CONSEQUENCES Emilio Kropff SISSA, Trieste di Studi Avanzati Internazionale Superiore - e conoscenza seguir
From spike frequency to free recall: How neural circuits perform encoding and retrieval.
 Hasselmo et al. 1 From spike frequency to free recall: How neural circuits perform encoding and retrieval. Michael Hasselmo, Bradley P. Wyble, Robert C. Cannon Department of Psychology and Program in Neuroscience
Hasselmo et al. 1 From spike frequency to free recall: How neural circuits perform encoding and retrieval. Michael Hasselmo, Bradley P. Wyble, Robert C. Cannon Department of Psychology and Program in Neuroscience
Computational approach to the schizophrenia: disconnection syndrome and dynamical pharmacology
 Computational approach to the schizophrenia: disconnection syndrome and dynamical pharmacology Péter Érdi1,2, Brad Flaugher 1, Trevor Jones 1, Balázs Ujfalussy 2, László Zalányi 2 and Vaibhav Diwadkar
Computational approach to the schizophrenia: disconnection syndrome and dynamical pharmacology Péter Érdi1,2, Brad Flaugher 1, Trevor Jones 1, Balázs Ujfalussy 2, László Zalányi 2 and Vaibhav Diwadkar
Lecture 12 Todd M. Gureckis, Lila Davachi
 An Introduction to Learning Lecture 12 Todd M. Gureckis, Lila Davachi Department of Psychology New York University {todd.gureckis,lila.davachi}@nyu.edu 1 Interference: Brown-Peterson Paradigm - simple
An Introduction to Learning Lecture 12 Todd M. Gureckis, Lila Davachi Department of Psychology New York University {todd.gureckis,lila.davachi}@nyu.edu 1 Interference: Brown-Peterson Paradigm - simple
Chapter 2: Studies of Human Learning and Memory. From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D.
 Chapter 2: Studies of Human Learning and Memory From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Medium Spiny Neuron A Current Conception of the major memory systems in the brain Figure
Chapter 2: Studies of Human Learning and Memory From Mechanisms of Memory, second edition By J. David Sweatt, Ph.D. Medium Spiny Neuron A Current Conception of the major memory systems in the brain Figure
Anatomy of the Hippocampus
 Anatomy of the Hippocampus Lecture 3.2 David S. Touretzky September, 2015 Human Hippocampus 2 Human Hippocampus 3 Hippocampus Means Seahorse Dissected human hippocampus next to a specimen of hippocampus
Anatomy of the Hippocampus Lecture 3.2 David S. Touretzky September, 2015 Human Hippocampus 2 Human Hippocampus 3 Hippocampus Means Seahorse Dissected human hippocampus next to a specimen of hippocampus
NEURONS COMMUNICATE WITH OTHER CELLS AT SYNAPSES 34.3
 NEURONS COMMUNICATE WITH OTHER CELLS AT SYNAPSES 34.3 NEURONS COMMUNICATE WITH OTHER CELLS AT SYNAPSES Neurons communicate with other neurons or target cells at synapses. Chemical synapse: a very narrow
NEURONS COMMUNICATE WITH OTHER CELLS AT SYNAPSES 34.3 NEURONS COMMUNICATE WITH OTHER CELLS AT SYNAPSES Neurons communicate with other neurons or target cells at synapses. Chemical synapse: a very narrow
Elliot D. Menschik, Shih-Cheng Yen, and Leif H. Finkel
 To appear in Computational Neuroscience: Trends in Research, 1998 (J.M. Bower, ed.) ATTRACTOR DYNAMICS IN REALISTIC HIPPOCAMPAL NETWORKS Elliot D. Menschik, Shih-Cheng Yen, and Leif H. Finkel Institute
To appear in Computational Neuroscience: Trends in Research, 1998 (J.M. Bower, ed.) ATTRACTOR DYNAMICS IN REALISTIC HIPPOCAMPAL NETWORKS Elliot D. Menschik, Shih-Cheng Yen, and Leif H. Finkel Institute
Part 11: Mechanisms of Learning
 Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Bursting dynamics in the brain. Jaeseung Jeong, Department of Biosystems, KAIST
 Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Neurons have cell membranes that separate them from the environment outside the neuron.
 Neural Communication Lecture 11 A. Resting Potential In this section, we will consider the basic unit of the nervous system the neuron and how neurons communicate with each other. The story of neural communication
Neural Communication Lecture 11 A. Resting Potential In this section, we will consider the basic unit of the nervous system the neuron and how neurons communicate with each other. The story of neural communication
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Cell communication. Gated ion channels. Allow specific ions to pass only when gates are open
 increase decrease Cell communication Gated ion channels Allow specific ions to pass only when gates are open Triggered by: potential change, chemical binding, temperature change, stretching 1 Voltage-Gated
increase decrease Cell communication Gated ion channels Allow specific ions to pass only when gates are open Triggered by: potential change, chemical binding, temperature change, stretching 1 Voltage-Gated
Cell communication. Gated ion channels. Voltage-Gated Na + Channel. Allow specific ions to pass only when gates are open
 increase decrease Cell communication Gated ion channels Allow specific ions to pass only when gates are open Voltage-Gated Na + Channel Activation gate ECF Triggered by: change, chemical binding, temperature
increase decrease Cell communication Gated ion channels Allow specific ions to pass only when gates are open Voltage-Gated Na + Channel Activation gate ECF Triggered by: change, chemical binding, temperature
BBS-D _ Mather_ Houghton. The Dentate Gyrus and the Hilar Revised
 BBS-D-15-00899_ Mather_ Houghton The Dentate Gyrus and the Hilar Revised Conor Houghton Department of Computer Science, University of Bristol, UK Department of Computer Science, Merchant Venturers Building,
BBS-D-15-00899_ Mather_ Houghton The Dentate Gyrus and the Hilar Revised Conor Houghton Department of Computer Science, University of Bristol, UK Department of Computer Science, Merchant Venturers Building,
Synfire chains with conductance-based neurons: internal timing and coordination with timed input
 Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Shadowing and Blocking as Learning Interference Models
 Shadowing and Blocking as Learning Interference Models Espoir Kyubwa Dilip Sunder Raj Department of Bioengineering Department of Neuroscience University of California San Diego University of California
Shadowing and Blocking as Learning Interference Models Espoir Kyubwa Dilip Sunder Raj Department of Bioengineering Department of Neuroscience University of California San Diego University of California
Molecular and Circuit Mechanisms for Hippocampal Learning
 Molecular and Circuit Mechanisms for Hippocampal Learning Susumu Tonegawa 1 and Thomas J. McHugh 1 The hippocampus is crucial for the formation of memories of facts and episodes (Scoville and Milner 1957;
Molecular and Circuit Mechanisms for Hippocampal Learning Susumu Tonegawa 1 and Thomas J. McHugh 1 The hippocampus is crucial for the formation of memories of facts and episodes (Scoville and Milner 1957;
DYNAMIC APPROACH to the PATHOLOGICAL BRAIN
 DYNAMIC APPROACH to the PATHOLOGICAL BRAIN Péter Érdi1,2 1 Center for Complex Systems Studies, Kalamazoo College, Kalamazoo, Michigan 2 Dept. Biophysics, KFKI Res. Inst. Part. Nucl. Phys. Hung. Acad. Sci.
DYNAMIC APPROACH to the PATHOLOGICAL BRAIN Péter Érdi1,2 1 Center for Complex Systems Studies, Kalamazoo College, Kalamazoo, Michigan 2 Dept. Biophysics, KFKI Res. Inst. Part. Nucl. Phys. Hung. Acad. Sci.
Synaptic transmission
 Outline Synaptic transmission Sompol Tapechum M.D., Ph.D. Department of Physiology Faculty of Medicine Siriraj Hospital, Bangkok, Thailand. sisth@mahidol.ac.th 2 Structure of synapse Modes of synaptic
Outline Synaptic transmission Sompol Tapechum M.D., Ph.D. Department of Physiology Faculty of Medicine Siriraj Hospital, Bangkok, Thailand. sisth@mahidol.ac.th 2 Structure of synapse Modes of synaptic
Synap&c Plas&city. long-term plasticity (~30 min to lifetime) Long-term potentiation (LTP) / Long-term depression (LTD)
 Long-term potentiation (LTP) / Long-term depression (LTD)") Synap&c Plas&city synaptic connectivity constantly changes in response to activity and other factors During development: provides the basic wiring of the brain s circuits Throughout rest of life: basis
Synap&c Plas&city synaptic connectivity constantly changes in response to activity and other factors During development: provides the basic wiring of the brain s circuits Throughout rest of life: basis
Supplementary Figure 1. Basic properties of compound EPSPs at
 Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Encoding and Retrieval in the CA3 Region of the Hippocampus: A Model of Theta-Phase Separation
 J Neurophysiol 94: 70 82, 2005. First published February 23, 2005; doi:10.1152/jn.00731.2004. Encoding and Retrieval in the CA3 Region of the Hippocampus: A Model of Theta-Phase Separation Steve Kunec,
J Neurophysiol 94: 70 82, 2005. First published February 23, 2005; doi:10.1152/jn.00731.2004. Encoding and Retrieval in the CA3 Region of the Hippocampus: A Model of Theta-Phase Separation Steve Kunec,
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Cholinergic suppression of transmission may allow combined associative memory function and self-organization in the neocortex.
 Cholinergic suppression of transmission may allow combined associative memory function and self-organization in the neocortex. Michael E. Hasselmo and Milos Cekic Department of Psychology and Program in
Cholinergic suppression of transmission may allow combined associative memory function and self-organization in the neocortex. Michael E. Hasselmo and Milos Cekic Department of Psychology and Program in
Sample Lab Report 1 from 1. Measuring and Manipulating Passive Membrane Properties
 Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Evaluating the Effect of Spiking Network Parameters on Polychronization
 Evaluating the Effect of Spiking Network Parameters on Polychronization Panagiotis Ioannou, Matthew Casey and André Grüning Department of Computing, University of Surrey, Guildford, Surrey, GU2 7XH, UK
Evaluating the Effect of Spiking Network Parameters on Polychronization Panagiotis Ioannou, Matthew Casey and André Grüning Department of Computing, University of Surrey, Guildford, Surrey, GU2 7XH, UK
Cellular and Molecular Mechanisms of Learning and Memory
 Cellular/Molecular Mechanisms of Learning and Memory 27 Cellular and Molecular Mechanisms of Learning and Memory 2 Matthew Lattal and Ted Abel The nature of the cellular basis of learning and memory remains
Cellular/Molecular Mechanisms of Learning and Memory 27 Cellular and Molecular Mechanisms of Learning and Memory 2 Matthew Lattal and Ted Abel The nature of the cellular basis of learning and memory remains
TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki
 TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki Rich Turner (turner@gatsby.ucl.ac.uk) Gatsby Unit, 22/04/2005 Rich T. Introduction Interneuron def = GABAergic non-principal cell Usually
TNS Journal Club: Interneurons of the Hippocampus, Freund and Buzsaki Rich Turner (turner@gatsby.ucl.ac.uk) Gatsby Unit, 22/04/2005 Rich T. Introduction Interneuron def = GABAergic non-principal cell Usually
Brain anatomy and artificial intelligence. L. Andrew Coward Australian National University, Canberra, ACT 0200, Australia
 Brain anatomy and artificial intelligence L. Andrew Coward Australian National University, Canberra, ACT 0200, Australia The Fourth Conference on Artificial General Intelligence August 2011 Architectures
Brain anatomy and artificial intelligence L. Andrew Coward Australian National University, Canberra, ACT 0200, Australia The Fourth Conference on Artificial General Intelligence August 2011 Architectures
Cell Responses in V4 Sparse Distributed Representation
 Part 4B: Real Neurons Functions of Layers Input layer 4 from sensation or other areas 3. Neocortical Dynamics Hidden layers 2 & 3 Output layers 5 & 6 to motor systems or other areas 1 2 Hierarchical Categorical
Part 4B: Real Neurons Functions of Layers Input layer 4 from sensation or other areas 3. Neocortical Dynamics Hidden layers 2 & 3 Output layers 5 & 6 to motor systems or other areas 1 2 Hierarchical Categorical
Neurobiology: The nerve cell. Principle and task To use a nerve function model to study the following aspects of a nerve cell:
 Principle and task To use a nerve function model to study the following aspects of a nerve cell: INTRACELLULAR POTENTIAL AND ACTION POTENTIAL Comparison between low and high threshold levels Comparison
Principle and task To use a nerve function model to study the following aspects of a nerve cell: INTRACELLULAR POTENTIAL AND ACTION POTENTIAL Comparison between low and high threshold levels Comparison
Modeling Excitatory and Inhibitory Chemical Synapses
 In review, a synapse is the place where signals are transmitted from a neuron, the presynaptic neuron, to another cell. This second cell may be another neuron, muscle cell or glandular cell. If the second
In review, a synapse is the place where signals are transmitted from a neuron, the presynaptic neuron, to another cell. This second cell may be another neuron, muscle cell or glandular cell. If the second
The synaptic Basis for Learning and Memory: a Theoretical approach
 Theoretical Neuroscience II: Learning, Perception and Cognition The synaptic Basis for Learning and Memory: a Theoretical approach Harel Shouval Phone: 713-500-5708 Email: harel.shouval@uth.tmc.edu Course
Theoretical Neuroscience II: Learning, Perception and Cognition The synaptic Basis for Learning and Memory: a Theoretical approach Harel Shouval Phone: 713-500-5708 Email: harel.shouval@uth.tmc.edu Course
Lateral Inhibition Explains Savings in Conditioning and Extinction
 Lateral Inhibition Explains Savings in Conditioning and Extinction Ashish Gupta & David C. Noelle ({ashish.gupta, david.noelle}@vanderbilt.edu) Department of Electrical Engineering and Computer Science
Lateral Inhibition Explains Savings in Conditioning and Extinction Ashish Gupta & David C. Noelle ({ashish.gupta, david.noelle}@vanderbilt.edu) Department of Electrical Engineering and Computer Science
Chapter 12 Nervous Tissue
 9/12/11 Chapter 12 Nervous Tissue Overview of the nervous system Cells of the nervous system Electrophysiology of neurons Synapses Neural integration Subdivisions of the Nervous System 1 Subdivisions of
9/12/11 Chapter 12 Nervous Tissue Overview of the nervous system Cells of the nervous system Electrophysiology of neurons Synapses Neural integration Subdivisions of the Nervous System 1 Subdivisions of
PSYCHOBIOLOGICAL MODELS OF HIPPOCAMPAL FUNCTION IN LEARNING AND MEMORY
 GLUCK PSYCHOBIOLOGICAL Annu. Rev. & Psychol. MYERS 1997. 48:481 514 MODELS OF LEARNING Copyright 1997 by Annual Reviews Inc. All rights reserved PSYCHOBIOLOGICAL MODELS OF HIPPOCAMPAL FUNCTION IN LEARNING
GLUCK PSYCHOBIOLOGICAL Annu. Rev. & Psychol. MYERS 1997. 48:481 514 MODELS OF LEARNING Copyright 1997 by Annual Reviews Inc. All rights reserved PSYCHOBIOLOGICAL MODELS OF HIPPOCAMPAL FUNCTION IN LEARNING
Neuroscience: Exploring the Brain, 3e. Chapter 4: The action potential
 Neuroscience: Exploring the Brain, 3e Chapter 4: The action potential Introduction Action Potential in the Nervous System Conveys information over long distances Action potential Initiated in the axon
Neuroscience: Exploring the Brain, 3e Chapter 4: The action potential Introduction Action Potential in the Nervous System Conveys information over long distances Action potential Initiated in the axon
arxiv: v1 [q-bio.nc] 25 Apr 2017
![arxiv: v1 [q-bio.nc] 25 Apr 2017](/thumbs/86/93358251.jpg "arxiv: v1 [q-bio.nc] 25 Apr 2017") Neurogenesis and multiple plasticity mechanisms enhance associative memory retrieval in a spiking network model of the hippocampus arxiv:1704.07526v1 [q-bio.nc] 25 Apr 2017 Yansong, Chua and Cheston, Tan
Neurogenesis and multiple plasticity mechanisms enhance associative memory retrieval in a spiking network model of the hippocampus arxiv:1704.07526v1 [q-bio.nc] 25 Apr 2017 Yansong, Chua and Cheston, Tan
Lecture 22: A little Neurobiology
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
episodic memory#recognition#recall#hippocampus#neural network models
 1 Encyclopedia of Cognitive Science Computational Models of Episodic Memory article reference code 444 Kenneth A. Norman Department of Psychology University of Colorado, Boulder 345 UCB Boulder, CO 80309
1 Encyclopedia of Cognitive Science Computational Models of Episodic Memory article reference code 444 Kenneth A. Norman Department of Psychology University of Colorado, Boulder 345 UCB Boulder, CO 80309
Improving Associative Memory in a Network of Spiking
 Improving Associative Memory in a Network of Spiking Neurons Computing Science and Mathematics School of Natural Sciences University of Stirling Scotland FK9 4LA Thesis submitted for the degree of Doctor
Improving Associative Memory in a Network of Spiking Neurons Computing Science and Mathematics School of Natural Sciences University of Stirling Scotland FK9 4LA Thesis submitted for the degree of Doctor
Basal Ganglia General Info
 Basal Ganglia General Info Neural clusters in peripheral nervous system are ganglia. In the central nervous system, they are called nuclei. Should be called Basal Nuclei but usually called Basal Ganglia.
Basal Ganglia General Info Neural clusters in peripheral nervous system are ganglia. In the central nervous system, they are called nuclei. Should be called Basal Nuclei but usually called Basal Ganglia.
Basal Ganglia Anatomy, Physiology, and Function. NS201c
 Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Activity-Dependent Development II April 25, 2007 Mu-ming Poo
 Activity-Dependent Development II April 25, 2007 Mu-ming Poo 1. The neurotrophin hypothesis 2. Maps in somatic sensory and motor cortices 3. Development of retinotopic map 4. Reorganization of cortical
Activity-Dependent Development II April 25, 2007 Mu-ming Poo 1. The neurotrophin hypothesis 2. Maps in somatic sensory and motor cortices 3. Development of retinotopic map 4. Reorganization of cortical
Portions from Chapter 6 CHAPTER 7. The Nervous System: Neurons and Synapses. Chapter 7 Outline. and Supporting Cells
 CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
Synaptic Transmission: Ionic and Metabotropic
 Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Plasticity and the NMDA Receptor
 Synaptic Plasticity and the NMDA Receptor Lecture 4.2 David S. Touretzky November, 2015 Long Term Synaptic Plasticity Long Term Potentiation (LTP) Reversal of LTP Long Term Depression (LTD) Reversal of
Synaptic Plasticity and the NMDA Receptor Lecture 4.2 David S. Touretzky November, 2015 Long Term Synaptic Plasticity Long Term Potentiation (LTP) Reversal of LTP Long Term Depression (LTD) Reversal of
CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems
 CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems Compare Chap 31 of Purves et al., 5e Chap 24 of Bear et al., 3e Larry Wittie Computer Science, StonyBrook University http://www.cs.sunysb.edu/~cse511
CSE511 Brain & Memory Modeling Lect 22,24,25: Memory Systems Compare Chap 31 of Purves et al., 5e Chap 24 of Bear et al., 3e Larry Wittie Computer Science, StonyBrook University http://www.cs.sunysb.edu/~cse511
Neurogenesis in Adult Central Nervous System: Death of a Dogma
 Aristotle University of Thessaloniki, Greece, Nov. 2007 Neurogenesis in Adult Central Nervous System: Death of a Dogma Anton B. Tonchev Division of Cell Biology, Varna University of Medicine, Bulgaria
Aristotle University of Thessaloniki, Greece, Nov. 2007 Neurogenesis in Adult Central Nervous System: Death of a Dogma Anton B. Tonchev Division of Cell Biology, Varna University of Medicine, Bulgaria
Mode Shifting Between Storage and Recall Based on Novelty Detection in Oscillating Hippocampal Circuits
 Mode Shifting Between Storage and Recall Based on Novelty Detection in Oscillating Hippocampal Circuits M. Meeter,* J.M.J. Murre, and L.M. Talamini HIPPOCAMPUS 14:722 741 (2004) ABSTRACT: It has been suggested
Mode Shifting Between Storage and Recall Based on Novelty Detection in Oscillating Hippocampal Circuits M. Meeter,* J.M.J. Murre, and L.M. Talamini HIPPOCAMPUS 14:722 741 (2004) ABSTRACT: It has been suggested
Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization
 Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization 1 7.1 Overview This chapter aims to provide a framework for modeling cognitive phenomena based
Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization 1 7.1 Overview This chapter aims to provide a framework for modeling cognitive phenomena based
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline
 Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Beyond Vanilla LTP. Spike-timing-dependent-plasticity or STDP
 Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode
Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons
 Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
Neurons: Structure and communication
 Neurons: Structure and communication http://faculty.washington.edu/chudler/gall1.html Common Components of a Neuron Dendrites Input, receives neurotransmitters Soma Processing, decision Axon Transmits
Neurons: Structure and communication http://faculty.washington.edu/chudler/gall1.html Common Components of a Neuron Dendrites Input, receives neurotransmitters Soma Processing, decision Axon Transmits
Learning and Memory. The Case of H.M.
 Learning and Memory Learning deals with how experience changes the brain Memory refers to how these changes are stored and later reactivated The Case of H.M. H.M. suffered from severe, intractable epilepsy
Learning and Memory Learning deals with how experience changes the brain Memory refers to how these changes are stored and later reactivated The Case of H.M. H.M. suffered from severe, intractable epilepsy
Na + K + pump. The beauty of the Na + K + pump. Cotransport. The setup Cotransport the result. Found along the plasma membrane of all cells.
 The beauty of the Na + K + pump Na + K + pump Found along the plasma membrane of all cells. Establishes gradients, controls osmotic effects, allows for cotransport Nerve cells have a Na + K + pump and
The beauty of the Na + K + pump Na + K + pump Found along the plasma membrane of all cells. Establishes gradients, controls osmotic effects, allows for cotransport Nerve cells have a Na + K + pump and
An attractor network is a network of neurons with
 Attractor networks Edmund T. Rolls An attractor network is a network of neurons with excitatory interconnections that can settle into a stable pattern of firing. This article shows how attractor networks
Attractor networks Edmund T. Rolls An attractor network is a network of neurons with excitatory interconnections that can settle into a stable pattern of firing. This article shows how attractor networks
Serotonergic Control of the Developing Cerebellum M. Oostland
 Serotonergic Control of the Developing Cerebellum M. Oostland Summary Brain development is a precise and crucial process, dependent on many factors. The neurotransmitter serotonin is one of the factors
Serotonergic Control of the Developing Cerebellum M. Oostland Summary Brain development is a precise and crucial process, dependent on many factors. The neurotransmitter serotonin is one of the factors
Cellular Neurobiology / BIPN 140
 SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
Neuromorphic computing
 Neuromorphic computing Robotics M.Sc. programme in Computer Science lorenzo.vannucci@santannapisa.it April 19th, 2018 Outline 1. Introduction 2. Fundamentals of neuroscience 3. Simulating the brain 4.
Neuromorphic computing Robotics M.Sc. programme in Computer Science lorenzo.vannucci@santannapisa.it April 19th, 2018 Outline 1. Introduction 2. Fundamentals of neuroscience 3. Simulating the brain 4.