Mutations within the pathogenic region of HSV-1 gk signal sequences alter cell surface. expression and neurovirulence.
|
|
|
- Kathryn Allen
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 10 December 2014 J. Virol. doi: /jvi Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 Mutations within the pathogenic region of HSV-1 gk signal sequences alter cell surface expression and neurovirulence Harry H. Matundan 1, Kevin R. Mott 1, Aslam Abbasi Akhtar 2, Joshua J. Breunig 2, and Homayon Ghiasi 1* 1 Center for Neurobiology and Vaccine Development, Ophthalmology Research, Department of Surgery, Cedars-Sinai Medical Center, Los Angeles, CA; and 2 Regenerative Medicine Institute and Dept. of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA *Corresponding author. Center for Neurobiology and Vaccine Development - D2056, Cedars-Sinai Burns and Allen Research Institute, 8700 Beverly Blvd., Los Angeles, CA PHONE: (310) ghiasih@cshs.org Running title: Effect of gk mutation in vitro and in vivo. Key Words: Signal sequence, ocular infection, cell surface expression, primary infection, latency, explant reactivation. 1
2 ABSTRACT To investigate the role of the signal sequences of HSV-1 gk on virus replication and viral pathogenesis, we constructed recombinant viruses with and without mutations within the signal sequences of gk. These recombinant viruses expressed two additional copies of the mutated (MgK) or native form (NgK) of gk gene in place of the latency associated transcript (LAT) with a myc epitope tag, to facilitate detection, at their 3 end. The replication of MgK virus was similar to that of NgK both in vitro and in vivo as well as in the trigeminal ganglia (TG) of latently-infected mice. The levels of gb and gk transcripts in the corneas, TG, and brains of infected mice on days 3 and 5 post infection were markedly virus- and time-dependent as well as tissue-specific. Mutation in the signal sequence of gk in MgK virus blocked cell surface expression of gk-myc in rabbit skin (RS) cells, increased LD 50 and decreased corneal scarring in ocularly infected mice as compared with the NgK or revertant (RgK) virus. MgK and NgK viruses and not the RgK virus had reduced extent of explant reactivation at the lower dose of ocular infection but not at the higher dose. However, time of reactivation was not affected by overexpression of the different forms of gk. Taken together, these results strongly suggest that the 8mer peptide (ITAYGLVL) within the signal sequence of gk promotes cell surface expression of gk in infected cells and ocular pathogenesis in infected mice. 2
3 Significance In this study we have shown for the first time that mutations within the signal sequence of gk blocked cell surface expression of inserted recombinant gk in vitro. Furthermore, this blockage in cell surface expression was correlated with higher LD 50 and lower CS in vivo. Thus, these studies point to a key role for the 8mer within the signal sequence of gk in HSV-1-induced pathogenicity. Downloaded from on January 18, 2019 by guest 3
4 INTRODUCTION HSV-1 infections are among the most frequent serious viral eye infections in the U.S. and are a major cause of viral-induced blindness (1-3). It is estimated that 70-90% of American adults have antibodies to HSV-1 and/or HSV-2 and about 25% of these individuals have clinical symptoms upon routine clinical inquiry. HSV-1 is responsible for >90% of ocular HSV infections (1, 2, 4-7). HSV-1-induced corneal scarring (CS), also broadly referred to as herpes stromal keratitis (HSK) can lead to blindness. Furthermore, HSV-1 is the leading cause of corneal blindness due to an infectious agent in developed countries (1, 2, 4-7). In addition to necrotizing HSK, ocular infection with HSV-1 can cause eye disease ranging in severity from blepharitis, conjunctivitis, and dendritic keratitis, to disciform stromal edema (4, 7-11). In the U.S., approximately 30,000 people per year suffer recurrent ocular HSV episodes, requiring doctor visits, medication, and in severe cases, corneal transplants. Although it is well established that HSV-1-induced CS is due to immune responses to the virus, the exact identity of the immune responses that lead to CS is not yet clearly apparent. The finding that HSV-1 glycoproteins are the major inducers and targets of humoral and cellmediated immune responses following infection (12-16) suggest that they may play a major role in the development of the immune responses that cause CS in infected individuals. We have investigated the role of the HSV-1 glycoproteins in protection from eye disease and have shown that immunization with glycoprotein B (gb), gc, gd, ge or gi completely protected mice against lethal challenge with HSV-1 and eye disease, whereas no significant protection is seen on immunization with gg, gh, gl, or gj (17-21). In marked contrast, immunization with gk leads to exacerbation of eye disease and facial dermatitis (14, 20). The gk-exacerbated CS occurs independently of the mouse strains or virus strains used in the analysis (22) and the pathology is due to the presence of CD8 + CD25 + T cells in the cornea of ocularly-infected mice (23). We also have shown that the titers of anti-gk antibody are higher in the sera of patients with HSK than in the sera of individuals that are positive for HSV-1 but do not have HSK (24). 4
5 gk, one of the 12 known HSV-1 glycoproteins, is a multimembrane-spanning viral glycoprotein encoded by UL53 (16, 25-27). It is expressed on the virions and HSV-1 mutants that lack gk fail to acquire a cytoplasmic envelope efficiently and are unable to efficiently infect and establish latency in neurons (28-32). Deletion of gk has been shown to result in reductions in virus yield, plaque size and translocation of the virus from the cytoplasm to the extracellular space (28). Moreover, viruses with deletion of gk are unable to undergo transport in either retrograde or anterograde directions in neuronal cell cultures (33). Therefore, to analyze the roles of gk in CS, we previously prepared a recombinant HSV-1 that expressed two additional copies of the gk gene. As compared with the wild-type (wt) HSV-1 strain McKrae, this recombinant virus exacerbated CS in two different strains of mice (34). Furthermore, we identified a peptide (8mer; ITAYGLVL) within the signal sequence of gk as being an immunodominant gk T-cell stimulatory region both in vitro and in vivo (35). Administration of this peptide as an eye drop resulted in significantly higher virus replication in the eyes of BALB/c mice, C57BL/6 mice, and NZW rabbits as well as greater viral-induced CS in the ocularly infected mice (36). Moreover, in HSV-infected humanized HLA-A*0201 transgenic mice, this gk 8mer epitope induced strong IFN- -producing cytotoxic CD8 + T-cell responses (36). In the current report, we investigated the effects of mutagenesis of the gk 8mer and overexpression of gk. We performed site-specific mutagenesis within the gk 8mer region and inserted the complete open reading frame (ORF) of the mutated form of the gk gene into both copies of the LAT regions and under the LAT promoter as we described previously (34, 37-40). As suitable antibodies for detection of gk are not available, we inserted the myc epitope tag inframe at the 3 end of the mutated gk ORF for identification of the inserted gk. This virus was designated mutated gk (MgK). As a control and to explore the effects of overexpression, an additional recombinant virus was constructed that expresses two copies of the native form of the gk ORF with a myc tag in-frame at its 3 end. This virus was designated native gk (NgK). This NgK virus is similar to the HSV-gK 3 virus that we described previously (34) except for having the 5
6 myc tag in-frame at its 3 end. Revertant virus, designated RgK, also was used as a control. Both the NgK and MgK viruses expressed the inserted gk in vitro as determined by Western blotting. However, although the NgK virus expressed the inserted gk on the cell surface of infected cells the MgK virus did not. The replication of the NgK, MgK and RgK viruses in mouse tears were similar, as was latency in the TG, but the MgK virus had a higher LD 50 than NgK or RgK with the NgK virus having the lowest LD 50. Ocular infection with NgK resulted in more severe CS than infection with MgK or RgK virus. In summary, the presence of the 8mer within the signal sequence of gk blocked cell surface expression of the inserted form of gk that led to a higher LD 50 and less CS in ocularly-infected mice. Downloaded from on January 18, 2019 by guest 6
7 MATERIALS AND METHODS Viruses, cells, and mice. Triple plaque-purified wt McKrae, dlat2903, HSV-gK 3, MgK, NgK, and RgK viruses were used in this study. Rabbit skin (RS) cells (used for preparation of virus stocks, the culture of mouse tear films, and determination of growth kinetics) were grown in Eagle's minimal essential media (MEM) supplemented with 5% FCS. Female six-week-old inbred BALB/c and C57BL/6 mice were purchased from The Jackson Laboratory (Bar Harbor, Maine). All animal procedures were performed in strict accordance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research and the NIH Guide for the Care and Use of Laboratory Animals (ISBN ). Animal research protocol was approved by the Institutional Animal Care and Use Committee of Cedars-Sinai Medical Center. Construction of gk recombinant viruses. The LAT plasmid (plat) was prepared using the BamHI B fragment of McKrae as we described previously (34, 41). The final construct lacked the 1.6-Kb of LAT fragment and contained 880 bp upstream and 2989 bp downstream of the deleted 1.6-Kb of LAT fragment with a BamHI site to insert the gene of interest. The complete ORF of gk is 338-amino acid long (16). Two forms of full-length gk were synthesized by GenScript (Piscataway, NJ). One, designated native gk (NgK), contained the complete ORF of gk with a myc tag sequence in-frame at its 3 end. The second, designated mutated gk (MgK), was synthesized such that it encoded a form of gk in which five amino acids within the 8mer peptide in the signal region were altered, i.e., ITAYGLVL to AGASGIVI, and had a myc tag sequence in-frame at its 3 end (Fig. 1). For construction of the gk plasmids, the complete ORFs of NgK and MgK were cloned into the BamHI site of the plat expression vector as we described previously (34, 41). The NgK and MgK synthesized genes were inserted into the BamHI site of the plat and the resulting plasmids were designated plat-ngk and plat-mgk, respectively. Each construct contained the 1040 bp gk gene, 33 bp of c-myc (EQKLISEEDL) 7
8 and the gk termination codon bounded by a 880 bp LAT fragment at the 5 end of the gk construct and a 2989 bp LAT fragment at the 3 end. NgK and MgK recombinant viruses were then generated by homologous recombination as we described previously (34, 41). Briefly, plat-ngk or plat-mgk was co-transfected with infectious dlat2903 DNA using the calcium phosphate method. Viruses from the cotransfection were plated and isolated plaques were picked and screened for gk gene insertion using restriction digestion and Southern blot analysis. Selected plaques that contained the inserted gk gene were plaque-purified four times and reanalyzed by restriction digestion and Southern blot analysis to verify that the inserted gk gene was present in the LAT region. For each construct, a single plaque was then chosen for purification and designated NgK or MgK. These viruses contained the gk gene that is in the normal LAT location in the viral genome and are under the control of the LAT promoter. Thus, in addition to the endogenous HSV-1 gk gene, these viruses contained two additional copies of the gk gene under the LAT promoter (one in each viral long repeat). Rescued viruses, in which the inserted gk gene was removed and the original deletion of the LAT gene was restored, were generated by co-transfection and homologous recombination using infectious NgK or MgK DNA and the plat plasmid. These rescued viruses behaved similarly to each other as well as to the parental dlat2903 virus. Thus, the rescued virus, designated RgK, generated from the MgK virus was used as the control virus for the subsequent studies. Southern analyses. RS cells were infected with 10 PFU/cells of McKrae, HSV-gK 3, NgK or MgK. Viral DNA was isolated and digested with EcoRV/EcoRI. Digested DNA was separated by a 0.9% agarose gel and the gel was subsequently blotted onto a positively charged nylon membrane (Amersham Hybond N +, GE Healthcare, Pittsburgh, PA). Following transfer, the membrane was fixed by UV-crosslinking (TL-2000 Ultraviolet Crosslinker). For, DNA hybridization and detection, the Digoxigenin (DIG) nonradioactive nucleic acid labeling and 8
9 detection system (DIG High prime DNA labeling/detection kit for Chemiluminescence, Roche- Diagnostics, Indianapolis, IN) was employed according to the manufacturer s protocol. The complete open reading frame (ORF) of gk was isolated from pac-gk1 construct that we described previously (26), by BamHI digestion. The gk complete ORF along with 7 nucleotides upstream of gk ATG and 15 noncoding nucleotides prior to the stop codon of gk is 1040 bases long and is called gk The gk-1040 was used as probe to identify natural gk and inserted gk in NgK and MgK recombinant viruses. Briefly, the purified gk-1040 fragment was used as template DNA for the hybridization probe, and was random primer labeled with DIG overnight at 37 C. The following day, the membrane was pre-hybridized in the DIG Easy-Hybe buffer (Roche-Diagnostics) for 2 h at 65 C, and subsequently hybridized with the GK-probe overnight in the same solution at 65 C. The membrane was washed twice with a solution of 2x SSC and 0.1% SDS for 5 min at room temperature followed by two additional washes with a solution of 0.5% SSC and 0.1% SDS for 15 min at 65 C. The hybridized DIG-labeled gk probe was immunodetected with anti-digoxigenin-ap conjugate and visualized using the chemiluminescence substrate CSPD Ready-to-Use solution and X-ray film. Virus replication in tissue culture. RS cell monolayers at 70-80% confluency were infected with 0.1 or 0.01 PFU/cell of NgK, MgK, or RgK. Virus was harvested at various times by subjecting the cell monolayers to two freeze-thawing cycles. Virus titers were determined using standard plaque assays on RS cells as described previously (14). Western analysis. RS cell monolayers at 70-80% confluency were infected with 10 PFU/cell of NgK, MgK, RgK or mock-infected. At 16 h post-infection (PI), infected cells were lysed directly into gel sample buffer and protein concentration in the cell lysates were determined by Pierce TM BCA protein assay kit according to the manufacturer s protocol (Thermo Scientific, Rockford, IL). Approximately 40 g of protein was loaded per well, samples were separated by 10% SDS-PAGE, transferred to nitrocellulose, and reacted with anti-myc antibody (Cell Signaling Technology, Danvers, MA). The membrane was incubated with 9
10 horseradish peroxidase-labelled secondary antibody (goat anti-rabbit IgG, 1:2,000) at 37 C for 1 h. The immunoreactive bands were detected using the enhanced chemiluminescence Western blotting system available from Thermo Scientific (Thermo Fisher Scientific, Waltham, MA). Flow Cytometric Analysis. RS cells were infected with 5 PFU/cell of NgK, MgK, or RgK or were mock infected. At 24 h PI, cells were harvested by centrifugation and stained with anti-myc-pe and anti-hsv-1 gc-fitc or anti-annexin-v PE and 7-AAD from BD PharMingen (San Diego, CA) and Biolegend (San Diego, CA). Stained cells were washed twice with FACS buffer (1X PBS with 0.1% sodium azide), resuspended in 4% paraformaldehyde, and analyzed using a multicolor fivelaser LSR II instrument (Applied Biosystems, Foster City, CA). Colocalization of gk expression. RS cells were grown to confluency on Lab-tek chamber slides and infected with 1 PFU/cell of NgK, MgK, RgK, or mock infected for 24 h as we described previously (42). The cells were fixed and permeabilized with 4% paraformaldehyde for 1 h at 4 C followed by 20 min incubation in Serum-Free Protein Block (Dako, Carpentaria, CA) at room temperature. The cells were then incubated with anti-myc-pe and anti-hsv-1 gc- FITC antibodies for 1 h at 4 C. Washed slides were air dried and mounted with 4,6-diamidino- 2-phenylindole (DAPI) prolong Gold (Invitrogen). The fluorophores were imaged in separate channels with a Zeiss ApoTome-equipped Axio Imager Z1 (Carl Zeiss Microimaging) using Nikon AIR laser scan confocal microscope. Ocular infection. Mice were infected ocularly with a 2 l drop of tissue culture media containing 2 X 10 6, 2 X 10 5, 2 X 10 4, 2 X 10 3, or 2 X 10 2 PFU/eye of NgK, MgK, or RgK virus. No corneal scarification was performed on the infected mice. Monitoring corneal scarring (CS). The severity of CS was scored in a masked fashion by examination with a slit lamp biomicroscope. Disease was scored on a 0 to 4 scale (0 = no disease, 1 = 25%, 2 = 50%, 3 = 75%, and 4 = 100% involvement). 10
11 Titration of virus in tears of infected mice. Tear films were collected from both eyes of 20 mice per group on days 1 to 5 post-ocular infection using a Dacron tipped swab. Each swab was placed in 0.5ml tissue culture medium, squeezed, and the amount of virus determined using a standard plaque assay on RS cells (43). Isolation of RNA from cornea, brain and TG of infected mice. C57BL/6 mice were ocularly infected with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus. Corneas, trigeminal ganglia (TG) and brains from infected mice were collected on days 3 and 5 PI. Isolated tissues were immersed in RNAlater RNA stabilization reagent and stored at -20 C until processing. The corneas, TG, or brain from each animal were processed for RNA extraction using TRIzol reagent as we described previously (44). Isolated total RNA was reverse-transcribed with random hexamer primers and murine leukemia virus (MuLV) reverse transcriptase provided in the High Capacity cdna Reverse Transcription Kit (Applied Biosystems, Foster City, CA), according to the manufacturer's recommendations. All isolated corneas were free of contamination from other parts of the mouse eye, vitreous fluid, and tears. DNA extraction for HSV-1 gb and gk DNAs. DNA was isolated from homogenized individual TG on day 28 PI using the commercially available DNeasy Blood &Tissue Kit (Qiagen, Stanford, CA) according to the manufacturer's instructions. TaqMan Real-Time PCR and PCR. The gb and gk custom-made primers and probe set used in this study were as follows: A) gb-specific primers (Forward - 5'- AACGCGACGCACATCAAG-3'; Reverse - 5'-CTGGTACGCGATCAGAAAGC-3'; and Probe - 5'- FAM-CAGCCGCAGTACTACC-3'); and 2) gk specific primers (forward, 5'- GGCCACCTACCTCTTGAACTAC-3'; reverse primer, 5'-CAGGCGGGTAATTTTCGTGTAG-3'; and probe, 5'-FAM-CAGGCCGCATCGTATC-3'). Amplicon length = 82 bp. As an internal control, a set of GAPDH primers from Applied Biosystems (ASSAY I.D. m _g1 - Amplicon Length = 107 bp) was used. 11
12 The expression levels of gb and gk genes, as well as the expression of the endogenous control gene GAPDH, were evaluated using a commercially available TaqMan Gene Expression Assay kit (Applied Biosystems, Foster City, CA) containing optimized primer and probe concentrations. In each experiment, an estimated relative copy number of gb and gk genes was calculated using standard curves generated from pac-gb1 (45) and pgem-gk1040 (26), respectively. Briefly, each plasmid DNA template was serially diluted 10-fold such that 5 l contained from 10 3 to copies of the desired gene and then subjected to TaqMan PCR with the same set of primers as the test samples. By comparing the normalized threshold cycle of each sample to the threshold cycle of the standards, the copy number for each reaction was determined. qpcr and qrt-pcr was performed using an ABI ViiA7 sequence detection system (Applied Biosystems) in 384-well plates. The threshold cycle (CT) values, which represent the PCR cycles at which there is a noticeable increase in the reporter fluorescence above baseline, were determined using ViiA7 RUO software. In vitro explant reactivation assay. Mice were sacrificed at 28 d PI and individual TG were removed and cultured in 1.5ml tissue culture medium as we described previously (46). Briefly, a 10 l aliquot was removed from each culture daily for 20 days and used to infect RS cell monolayers. The RS cells were monitored daily for the appearance of cytopathic effect (CPE) for 5 days to determine the time of first appearance of reactivated virus from each TG. As the media from the explanted TG cultures were plated daily, the time at which reactivated virus first appeared in the explanted TG cultures could be determined. Statistical analysis. Protective parameters were analyzed using the Student's t test and Fisher's exact test using Instat software (GraphPad, San Diego, CA). Results were considered to be statistically significant if the p value was <
13 RESULTS Structure of the HSV-1 NgK and MgK viruses. To examine the effects of the 8mer in the signal region of gk on HSV-1 pathogenesis, we constructed two derivatives of HSV-1 strain McKrae that express two additional copies of the gk gene, with and without mutation, within the 8mer region of gk (Fig. 2). As wt strain McKrae was used as the original parental virus, a LATnull mutant, dlat2903, that was derived from the McKrae strain (41) was used to construct the NgK and MgK viruses. The genomic structure of the wt HSV-1 strain McKrae is shown in Figure 2A and that of dlat2903 in Figure 2B. The location of the TATA box of the LAT promoter, which we described previously (47), is indicated. The NgK and MgK viruses were derived from the dlat2903 strain by the insertion of the NgK and MgK genes and restoration of the LAT promoter so that the inserted NgK (Fig. 2C) or MgK (Fig. 2D) gene is under the control of the endogenous LAT promoter. The genomic structure of the NgK and MgK viruses was confirmed by restriction enzyme analysis, Southern blotting (Fig. 2E), and partial sequencing (not shown). The NgK and MgK viruses, in addition to their endogenous gk gene have two additional copies of NgK and MgK genes, one of which is in each viral long repeat. The rescued viruses for NgK and MgK were found to be identical to each other and are similar to their parental dlat2903, in all parameters examined (not shown). Thus, in all of the subsequent experiments we used the rescued virus derived from MgK, which we designated RgK. Replication of NgK and MgK viruses in tissue culture. To determine if overexpression of gk or mutation within the signal sequence of gk affect viral replication, we quantified virus yield following infection of RS cells with 0.01 or 0.1 PFU/cell of NgK, MgK or RgK. The cell monolayers were freeze-thawed at the indicated time points and the yield of infectious virus quantified using a standard plaque assay. At the lowest MOI (0.01), the yield of viruses was time-dependent. At 48 h PI, the yield of NgK virus was lower than the yield of RgK. The yield of MgK was similar to that of the NgK virus although not at earlier time points (Fig. 3A, P<0.05, student t-test). At an MOI of 0.1, the yields of NgK and MgK viruses were similar at all 13
14 tested time points and were lower than that of RgK virus at 0 to 48 h PI (Fig. 3B, P<0.05, student t-test). At an MOI of 10, no significant differences in the replication of the viruses were detected (data not shown). Western blot analyses of NgK and MgK expression in tissue culture. To verify that the recombinant viruses express the inserted genes, confluent monolayers of RS cells were infected for 24 h with MgK, NgK or RgK at a multiplicity of 5 PFU/cell. Western blot analysis of total protein extracts using an anti-myc antibody revealed a band that was present in NgKinfected cells and MgK-infected cells (Fig. 4, lanes MgK and NgK), which was not detected in the RgK-infected cells (Fig. 4, lane RgK) or mock-infected RS cells (Fig. 4, Mock). In addition to this major band, a smaller and faintly reactive band was detected in the NgK-infected cells and the MgK-infected cells (Fig. 4, Lanes MgK and NgK). Most likely, the major band represents the N-linked glycosylated form of gk whereas the minor band represents the cleaved and partially glycosylated form of gk. Finally, the upper band that was detected in both infected and mockinfected groups may represent the internal myc protein (Fig. 4, upper band). FACS analyses of MgK and NgK infected cells. To further verify that the NgK and MgK virsues expressed the inserted genes, we used FACS analysis. RS cells were infected for 24 h with 5 PFU/cell of NgK-, MgK-, RgK-, or mock-infected. The cells were then surface stained with anti-myc and anti-hsv-1 gc antibodies and subjected to FACS analysis in which stained cells were gated for expression of HSV-1 gc (Fig. 5A) or gk-myc (Fig. 5B). Infection of the cells was verified by the similar levels of expression of HSV-1 gc on cells infected with NgK, MgK or RgK (Fig. 5A). In contrast to gc expression (Fig. 5A), the level of surface expression of gk-myc protein appeared to be more than 50% higher in NgK-infected cells compared with MgK- and RgK-infected cells (Fig. 5B). Thus, presence of low levels of myc expression in the MgK- and RgK-infected cells most likely represent base-line expression of myc which is detected by the internalized anti-myc antibody. 14
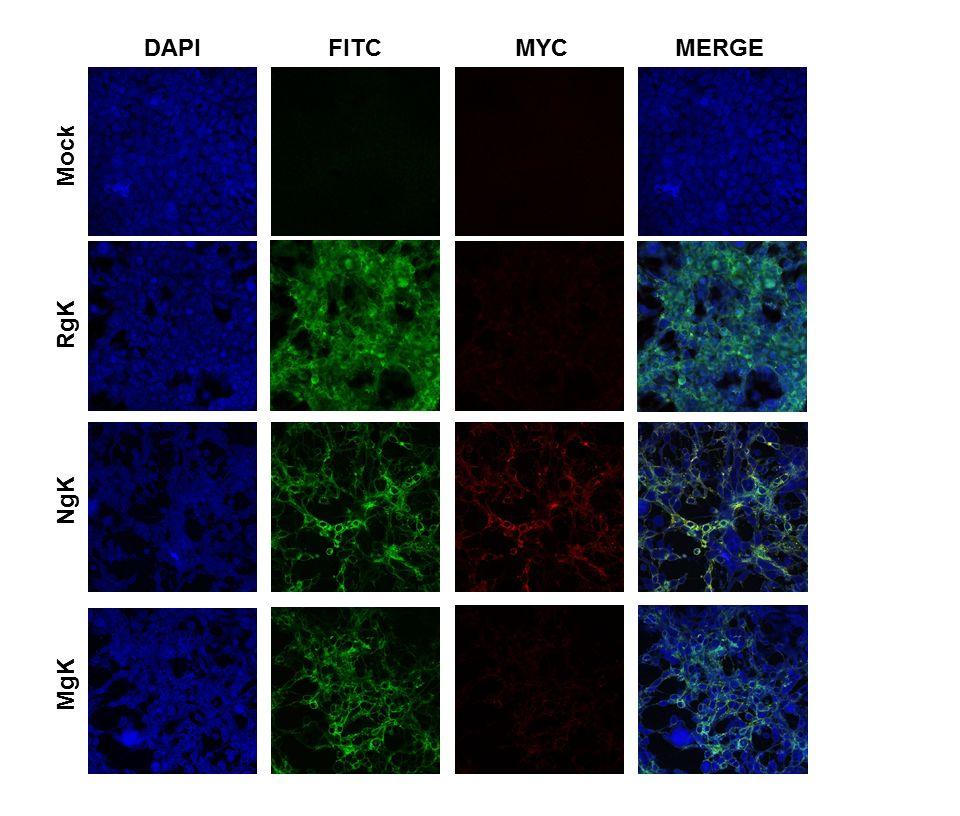
15 Cell surface expression of gc and gk-myc by MgK and NgK in infected cells. Although the results of the Western blotting (Fig. 4) indicated that gk is expressed by both NgK and MgK viruses, the results of the FACS analysis indicated that the two recombinant viruses differed in the cell surface expression of gk-myc (Fig. 5B). We therefore analyzed the surface expression of gk-myc expression using double-label immunofluorescence analysis. RS cells were grown on Lab-Tex chamber slides, infected with 5 PFU/cell of NgK, MgK, RgK or mockinfected for 24 h, then incubated with fluorescently labeled antibodies against HSV-1 gc and the myc epitope. We found strong and similar levels of cell surface expression of gc for the cells infected with NgK, MgK or RgK and, as expected, gc expression was not detected in the mockinfected cells (Fig. 6; gc-fitc). Also as expected, gk-myc expression was not detectable in either the RgK-infected cells or mock-infected cells (Fig. 6; gk-myc). Cell surface expression of gk-myc was detected in the NgK-infected cells but not in the MgK-infected cells. Moreover, on merging the images of gc and gk-myc staining, we found that the cell surface expression of gkmyc and gc colocalized in the NgK infected cells (Fig. 6; Merge). Collectively, these results suggested that mutation in the signal sequence of gk prevented cell surface expression of gk by MgK recombinant virus. This effect could be attributed to the mutation in the signal sequence as the NgK recombinant virus, which also overexpressed the myc sequence, was capable of cell surface expression. Effect of gk overexpression on apoptosis and cell death of infected cells in culture. To investigate the potential effects of gk overexpression and mutagenesis on cell death and apoptosis, we infected RS cells with 5 PFU/cell of NgK, MgK, RgK or mock-infected. At 24 h PI, the infected cells were isolated and FACS analyses performed using the marker of cell death, Propidium iodide, and the apoptosis marker, Annexin V. The percentages of cells staining with Propidium iodide and Annexin V were similar in NgK-, MgK, and RgK-infected cells and were not significantly different from the percentages observed in mock-infected cells (Fig. 7). Thus, we did not find evidence of an association between overexpression of gk and mutagenesis of gk with cell death or apoptosis. 15
16 Replication of NgK and MgK in mouse tear films. Viral replication during ocular infection was assessed by determining the PFU in the tear films of infected mice. C57BL/6 mice were infected ocularly with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus and tear films collected on days 3 and 5 PI. On day 3 PI, the titers of virus were higher in the tear films from RgK- and MgK-infected mice than the NgK-infected mice but these differences were not statistically significant (Fig. 8, P>0.05). On day 5 PI, the titers of virus in tear films of NgK- and MgKinfected mice were similar (Fig. 8, P>0.05) and were significantly lower than the titers of virus in the tear films of RgK-infected mice (Fig. 8, P<0.05). These results were consistent with the reduced virus replication with lower MOI in NgK- and MgK-infected RS cells in tissue culture (Fig. 3, above) and indicated that the presence of the two additional copies of gk significantly reduced the amount of HSV-1 found in the tear films of ocularly infected mice on day 5 PI. Effect of gk overexpression on gb and gk expression during primary ocular infection. To investigate the effect of gk overexpression and mutagenesis on viral transcription in vivo, C57BL/6 mice were infected with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus. The corneas, TG, and brain were collected on days 3 and 5 PI and the total RNA was isolated and subjected to TaqMan RT-PCR to estimate the levels of gk and gb mrna using the level of GAPDH mrna in each sample as an internal control. The results showed differences in expression among the various tissues. On day 3 PI, the levels of gb (Fig. 9A) and gk (Fig. 9B) transcripts were significantly lower in the corneas from MgK-infected mice than in the corneas from NgK- or RgK-infected mice (p<0.05). By day 5 PI, the levels of gb and gk transcripts in the corneas of the MgK-infected mice had increased and were no longer significantly different from the levels in the corneas from NgK- or RgK-infected mice (Fig. 9A and B). On day 3 PI, the levels of both gb (Fig. 9C, p<0.05) and gk (Fig. 9D, p<0.05) transcripts in the TG of mice infected with NgK or MgK were lower than the levels in the TG of mice infected with RgK. Again, by day 5 PI, the levels of both gb and gk in the TG of mice infected with NgK, MgK or RgK were not statistically significant (Fig. 9C, D; p>0.05). On day 3 PI, the levels of gb 16
17 transcripts were similar in the brains of mice infected with either NgK or MgK but were significantly higher in the brains of RgK-infected mice (Fig. 9E; p<0.05). By day 5 PI, the levels of gb transcripts in the brains of mice infected with NgK, MgK or RgK were similar (Fig. 9E; p>0.05). In contrast, the levels of gk transcripts were similar in the brains of mice infected with NgK, MgK or RgK virus on both days 3 and 5 PI (Fig. 9F; p>0.05). These results indicated that the presence of two extra copies of gk influenced viral transcription in a time-dependent and tissue-specific manner and indicated that the levels of gb transcripts were significantly higher than the level of gk transcripts in all three tissues. Collectively, the results described above suggested that virus replication in the tears of infected mice as well as the levels of gb and gk transcripts are affected by the presence two extra copies of the NgK or MgK genes. Viral latency in TG of latently infected mice. To determine if the presence of NgK or MgK genes also influences the level of latency in the TG of latently infected mice, C57BL/6 mice were ocularly infected with NgK, MgK or RgK virus and the TG isolated on day 28 PI. Latency in individual mice was determined by qpcr using primers from the gb and gk regions of the HSV-1 genome. The levels of gb DNA were similar in mice infected with NgK, MgK or RgK (Fig. 10A; p>0.05). Similarly, no significant differences in the levels of gk in the TG of mice infected with NgK, MgK or RgK were detected (Fig. 10B; p>0.05). Thus, the level of latency in TG of infected mice was not affected by the presence of either the NgK or MgK gene. Explant reactivation in TG of latently infected mice. To determine if virus reactivation is affected differentially by NgK or MgK overexpression, C57BL/6 mice were ocularly infected with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus and virus reactivation analyzed by explanting individual TG. The time to reactivation was similar among the NgK, MgK and RgK viruses (5.3 ± 0.2 days vs 5.8 ± 0.4 days; vs 5.4 ± 0.4 days; Fig. 11; 2 X 10 5 PFU/eye; p>0.05). Thus, the time to reactivation was not influenced by the presence of the NgK or MgK gene in mice infected with the recombinant viruses. In contrast, however, the presence of either the NgK gene or the MgK gene affected the numbers of TG showing reactivation. Although all of the TG from RgK-infected mice were 17
18 reactivated during the monitoring period, 50% of TG from NgK-infected mice (6 of 12 TG) and 58% of TG from MgK-infected mice (5 of 12 TG) did not exhibit reactivation of the virus. With respect to the number of TG showing reactivation, the differences between NgK and MgK viruses compared to RgK virus were statistically significant (p<0.002). To determine if a higher infectious dose of virus would increase the numbers of TG showing reactivation of the viruses, we infected C57BL/6 mice with 2 X 10 6 PFU/eye of NgK and MgK viruses. Subsequent monitoring of explant reactivation indicated that the time to reactivation was similar in the TG of mice infected with NgK or MgK viruses (4.5 ± 0.3 days vs 4.7 ± 0.2 days; Fig. 11; 2 X 10 6 PFU/eye; p>0.05). However, the time to reactivation was faster in the TG of mice infected with 2 X 10 6 PFU/eye as compared to those infected with 2 X 10 5 PFU/eye of NgK or MgK virus (Fig. 11; compare the left side with the right side). Furthermore, all of the TG from mice infected with 2 X 10 6 PFU/eye of NgK or MgK virus were reactivated. Thus, at a lower infective dose the presence of the NgK or MgK gene does not affect time to reactivation of the virus but does affect the percentage of TG showing reactivation. Virulence of NgK and MgK viruses in infected BALB/c and C57BL/6 mice. To determine whether there is a difference in the virulence of the NgK and MgK viruses compared to the rescued RgK virus, we used BALB/c and C57BL/6 mice. BALB/c mice are sensitive to HSV-1 infection whereas C57BL/6 mice are resistant. Groups of 5 BALB/c mice were infected ocularly in both eyes with 10-fold serial dilutions of each virus ranging from 2 X 10 2 to 2 X 10 5 PFU/eye and the 50% lethal dose (LD 50 ) was determined for each group (Table 1). Following ocular infection the LD 50 for NgK virus (15,642 PFU) was approximately 2-fold lower than the LD 50 for RgK virus (27,660 PFU) and significantly lower than the LD 50 for MgK virus (2,115,650 PFU) (Table 1). Thus, the MgK virus appeared to be less virulent than either NgK or the marker rescued virus as judged by the ability to kill BALB/c mice following ocular infection. This suggests that the higher mortality in the NgK-infected mice was due to the presence of NgK 18
19 gene and the lower mortality in the MgK-infected mice was due to the mutations in the signal sequence of the gk gene rather than a defect in the recombinant virus. LD 50 studies performed using C57BL/6 mice indicated that at 2 X 10 5 PFU/eye of NgK, MgK, or RgK virus, all the infected mice survived ocular infection (Table 1). Similarly, at a dose of 2 X 10 6 PFU/eye all mice infected with MgK and RgK survived (Table 1). At this dose, one mouse in the NgK group died as the result of infection suggesting that even in C57BL/6 mice NgK is more pathogenic than the MgK or the RgK virus. However, since only 1 mouse died as the result of infection, it was not possible to calculate the LD 50 of the recombinant viruses in C57BL/6 mice. Overall, these results suggested that expression of NgK or MgK affected neurovirulence in infected mice as compared with rescued virus. In addition, NgK gene insertion enhanced neurovirulence in infected mice, while insertion of MgK reduced neurovirulence in infected mice. Effect of NgK and MgK overexpression on CS in infected mice. To determine the effect of NgK or MgK on eye disease, the severity of CS was evaluated following ocular infection of mice described in Table 1. CS was measured in the surviving mice on day 28 PI. BALB/c mice infected with 2 X 10 4, 2 X 10 3 or 2 X 10 2 PFU/eye of NgK, MgK, or RgK virus developed similar levels of CS and the differences were not statistically significant (Table 2; p>0.05). CS also was examined in C57BL/6 mice infected ocularly with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus. C57BL/6 mice infected with 2 X 10 5 PFU/eye of NgK, MgK or RgK virus displayed similar levels of CS (Table 2, C57BL/6, P>0.1). To determine if a higher PFU/eye of virus can increase CS, additional groups of C57BL/6 mice were infected with 2 X 10 6 PFU/eye of each virus. At this dose, CS was higher in the NgK- and MgK- infected mice than in RgK-infected mice (Table 2, 2 X 10 6, C57BL/6). The CS in NgK infected mice was significantly higher than RgK-infected and MgK-infected mice (Table 2). Thus, overexpression of NgK also exacerbates virus-induced CS in infected C57BL/6 mice. 19
20 DISCUSSION Since gk is an essential HSV-1 gene and gk-null viruses are not efficiently propagated in vitro and in vivo (29, 32), to improve our understanding of the role of gk in HSV-1 pathogenesis, we previously constructed and characterized a recombinant HSV-1 that expresses two additional copies of the HSV-1 gk gene (34). This recombinant virus increased pathogenesis in infected mice. More recently, we reported that a 8mer within the signal sequence of gk also contributed to exacerbation of eye disease in different strains of mice and in rabbits (36). Using 33 peptides overlapping the entire 338 amino acids of gk, we established that no other region of gk contributed to this pathogenic effect (35, 36). The results of the current study support and extend our previous observations. Deletion of gk has been shown to affect virus yield and plaque size and translocation of the virus from the cytoplasm to the extracellular space (28) and to play a role in virion entry, cytoplasmic virion envelopment and virus-induced cell fusion (27-30, 48-52). These functions of HSV gk are similar to those reported for the gks of pseudorabies and varicella-zoster virus (53-55). HSV-1 mutants that lack gk fail to acquire a cytoplasmic envelope efficiently. This leads to a drastic reduction in virion egress and spread, thus HSV-1 mutants are unable to efficiently infect and establish latency in neurons (28-32). Our analysis of gk with mutated signal sequence indicated that the titers of the MgK virus in RS cells and the tears of ocularly-infected C57BL/6 mice were similar to the titers of the NgK virus, indicating that the 8mer region of gk does not play a role in the replication of the virus. Despite having similar virus titers in the tear films of C57BL/6 mice, MgK-infected mice had less severe CS than mice infected with the NgK or RgK virus. Previously, we reported that immunization with gk followed by infection with McKrae (or with the avirulent HSV-1 KOS virus) leads to CS in both BALB/c and C57BL/6 mice (22). The severity of CS in NgK-infected mice was similar to that of HSV-gK 3 that lack the myc at its 3 end (34). Overall, NgK virus behaved similar to that of HSV-gK 3 suggesting that inframe expression of the myc tag by NgK had no deleterious effect on gk pathogenesis. 20
21 Previously, we reported that exacerbation of CS in gk immunized mice associated with the presence of CD8 + CD25 + T cells in the cornea of ocularly infected mice (23). In addition, our previous work showed that CD8 + T cells contribute to exacerbation of CS in HSV-gK 3 -infected mice (34) and the pathogenic region of gk is located within the signal sequence of gk (35, 36). The gk 8mer induced CD8 + T cell responses contributed to an increase of eye disease in BALB/c mice, C57BL/6 mice, and NZW rabbits (36). This study is consistent with and extends our previous work by showing that the 8mer within the signal sequence of gk induces a CD8 + T cell response in cornea of ocularly infected mice, which contributes to the exacerbation of eye disease independent of the strain of mice. The gk expressed on virions forms a complex with the membrane-associated UL20 viral protein (52, 56). HSV-1 mutants that lack gk fail to acquire a cytoplasmic envelope efficiently. This leads to a drastic reduction in virion egress and spread and HSV-1 mutants are unable to efficiently infect and establish latency in neurons (28-32). In the current study, NgK was detected on the surface of infected cells when expressed by the LAT promoter. In marked contrast, we found that mutation within the signal sequence of gk prevented cell surface expression of gk by MgK. Thus, mutations in the signal sequence of gk driven by the LAT promoter and in place of LAT blocked transport of gk to the cell surface of infected cells. The lack of cell surface expression of gk in MgK-virus infected cells may be a contributing factor to its reduced pathogenesis in infected mice. The higher LD 50 of the MgK virus in comparison to the NgK virus or rescued virus, as well as the reduced exacerbation of CS may also be a result of the lack of gk cell surface expression. Alternatively the differences in the total protein expression levels between NgK and MgK viruses and not mutation in the signal sequence of MgK may be responsible for the differences in the disease phenotype between NgK and MgK viruses. Taken together with our observation of the exacerbation of CS by overexpression of gk (i.e., NgK virus), these results raise the possibility that cell surface expression of gk and/or proper presentation of the 8mer peptide within the signal sequence of the native form of gk 21
22 plays a key role in the pathogenesis of HSV-1. This would be consistent with a requirement for effective MHC-I presentation and the subsequent generation of CD8 + T cells that is required for exacerbation of CS in infected mice. In addition, we found that the levels of latency in the TG of C57BL/6 mice infected with 2 X 10 5 PFU/eye of each virus was the same. However, at this dose of infection a significant number of TG in both NgK and MgK viruses were not reactivated when compared to the rescued virus, although all TG in NgK and MgK viruses were reactivated when mice were infected with a 10-fold higher dose of each virus. Thus, our results suggest that the presence of extra copies of gk and/or the presence of the myc tag affect the extent of reactivation but not the level of latency. In summary, the results presented here support the hypothesis that overexpression of gk increases corneal scarring in mice. Moreover, the pathogenic role of gk in HSV-1-induced CS appears to be dependent on the cell surface expression of gk. Downloaded from on January 18, 2019 by guest 22
23 ACKNOWLEDGEMENTS This work was supported by NIH grant 1 RO1 EY Downloaded from on January 18, 2019 by guest 23
24 REFERENCES 1. Barron BA, Gee L, Hauck WW, Kurinij N, Dawson CR, Jones DB, Wilhelmus KR, Kaufman HE, Sugar J, Hyndiuk RA, et al Herpetic Eye Disease Study. A controlled trial of oral acyclovir for herpes simplex stromal keratitis. Ophthalmology 101: Wilhelmus KR, Dawson CR, Barron BA, Bacchetti P, Gee L, Jones DB, Kaufman HE, Sugar J, Hyndiuk RA, Laibson PR, Stulting RD, Asbell PA Risk factors for herpes simplex virus epithelial keratitis recurring during treatment of stromal keratitis or iridocyclitis. Herpetic Eye Disease Study Group. Br J Ophthalmol 80: Farooq AV, Shukla D Herpes simplex epithelial and stromal keratitis: an epidemiologic update. Survey of ophthalmology 57: Dawson CR Ocular herpes simplex virus infections. Clin Dermatol 2: Liesegang TJ Classification of herpes simplex virus keratitis and anterior uveitis. Cornea 18: Liesegang TJ Herpes simplex virus epidemiology and ocular importance. Cornea 20: Hill TJ Ocular pathogenicity of herpes simplex virus. Curr Eye Res 6: Binder PS A review of the treatment of ocular herpes simplex infections in the neonate and immunocompromised host. Cornea 3: Streilein JW, Dana MR, Ksander BR Immunity causing blindness: five different paths to herpes stromal keratitis. Immunol Today 18: Thomas J, Rouse BT Immunopathogenesis of herpetic ocular disease. Immunol Res 16: Branco BC, Gaudio PA, Margolis TP Epidemiology and molecular analysis of herpes simplex keratitis requiring primary penetrating keratoplasty. Br J Ophthalmol 88:
25 Burke RL Development of a herpes simplex virus subunit glycoprotein vaccine for prophylactic and therapeutic use. Rev Infect Dis 13:S906-S Burke RL Contemporary approaches to vaccination against herpes simplex virus. Curr Top Microbiol Immunol 179: Ghiasi H, Kaiwar R, Nesburn AB, Slanina S, Wechsler SL Expression of seven herpes simplex virus type 1 glycoproteins (gb, gc, gd, ge, gg, gh, and gi): comparative protection against lethal challenge in mice. J Virol 68: Dix RD, Mills J Acute and latent herpes simplex virus neurological disease in mice immunized with purified virus-specific glycoproteins gb or gd. J Med Virol 17: McGeoch DJ, Dalrymple MA, Davison AJ, Dolan A, Frame MC, McNab D, Perry LJ, Scott JE, Taylor P The complete DNA sequence of the long unique region in the genome of herpes simplex virus type 1. J Gen Virol 69: Ghiasi H, Kaiwar R, Nesburn AB, Wechsler SL Baculovirus-expressed glycoprotein G of herpes simplex virus type 1 partially protects vaccinated mice against lethal HSV-1 challenge. Virology 190: Ghiasi H, Kaiwar R, Nesburn AB, Wechsler SL Baculovirus-expressed glycoprotein H of herpes simplex virus type 1 (HSV-1) induces neutralizing antibody and delayed type hypersensitivity responses, but does not protect immunized mice against lethal HSV-1 challenge. J Gen Virol 73: Ghiasi H, Kaiwar R, Slanina S, Nesburn AB, Wechsler SL Expression and characterization of baculovirus expressed herpes simplex virus type 1 glycoprotein L. Arch Virol 138: Ghiasi H, Bahri S, Nesburn AB, Wechsler SL Protection against herpes simplex virus-induced eye disease after vaccination with seven individually expressed herpes simplex virus 1 glycoproteins. Invest Ophthalmol Vis Sci 36:
26 Ghiasi H, Nesburn AB, Cai S, Wechsler SL The US5 open reading frame of herpes simplex virus type 1 does encode a glycoprotein (gj). Intervirology 41: Ghiasi H, Cai S, Slanina S, Nesburn AB, Wechsler SL Nonneutralizing antibody against the glycoprotein K of herpes simplex virus type-1 exacerbates herpes simplex virus type-1-induced corneal scarring in various virus-mouse strain combinations. Invest Ophthalmol Vis Sci 38: Allen SJ, Mott KR, Ljubimov AV, Ghiasi H Exacerbation of corneal scarring in HSV-1 gk-immunized mice correlates with elevation of CD8+CD25+ T cells in corneas of ocularly infected mice. Virology. 399: Mott KR, Osorio Y, Maguen E, Nesburn AB, Wittek AE, Cai S, Chattopadhyay S, Ghiasi H Role of anti-glycoproteins D (anti-gd) and K (anti-gk) IgGs in pathology of herpes stromal keratitis in humans. Invest Ophthalmol Vis Sci 48: Hutchinson L, Goldsmith K, Snoddy D, Ghosh H, Graham FL, Johnson DC Identification and characterization of a novel herpes simplex virus glycoprotein, gk, involved in cell fusion. J Virol 66: Ghiasi H, Slanina S, Nesburn AB, Wechsler SL Characterization of baculovirus-expressed herpes simplex virus type 1 glycoprotein K. J Virol 68: Foster TP, Rybachuk GV, Kousoulas KG Glycoprotein K specified by herpes simplex virus type 1 is expressed on virions as a Golgi complex-dependent glycosylated species and functions in virion entry. J Virol 75: Foster TP, Kousoulas KG Genetic analysis of the role of herpes simplex virus type 1 glycoprotein K in infectious virus production and egress. J Virol 73: Hutchinson L, Johnson DC Herpes simplex virus glycoprotein K promotes egress of virus particles. J Virol 69:
27 Hutchinson L, Roop-Beauchamp C, Johnson DC Herpes simplex virus glycoprotein K is known to influence fusion of infected cells, yet is not on the cell surface. J Virol 69: David AT, Baghian A, Foster TP, Chouljenko VN, Kousoulas KG The herpes simplex virus type 1 (HSV-1) glycoprotein K(gK) is essential for viral corneal spread and neuroinvasiveness. Curr Eye Res. 33: Jayachandra S, Baghian A, Kousoulas KG Herpes simplex virus type 1 glycoprotein K is not essential for infectious virus production in actively replicating cells but is required for efficient envelopment and translocation of infectious virions from the cytoplasm to the extracellular space. J Virol 71: David AT, Saied A, Charles A, Subramanian R, Chouljenko VN, Kousoulas KG A herpes simplex virus 1 (McKrae) mutant lacking the glycoprotein K gene is unable to infect via neuronal axons and egress from neuronal cell bodies. mbio 3:e Mott KR, Perng GC, Osorio Y, Kousoulas KG, Ghiasi H A Recombinant Herpes Simplex Virus Type 1 Expressing Two Additional Copies of gk Is More Pathogenic than Wild-Type Virus in Two Different Strains of Mice. J Virol. 81: Osorio Y, Mott KR, Jabbar AM, Moreno A, Foster TP, Kousoulas KG, Ghiasi H Epitope mapping of HSV-1 glycoprotein K (gk) reveals a T cell epitope located within the signal domain of gk. Virus Res. 128: Mott KR, Chentoufi AA, Carpenter D, Benmohamed L, Wechsler SL, Ghiasi H The role of a glycoprotein K (gk) CD8+ T-cell epitope of herpes simplex virus on virus replication and pathogenicity. Invest Ophthalmol Vis Sci. 50: Mott KR, Allen SJ, Zandian M, Akbari O, Hamrah P, Maazi H, Wechsler SL, Sharpe AH, Freeman GJ, Ghiasi H Inclusion of CD80 in HSV Targets the Recombinant 27
28 Virus to PD-L1 on DCs and Allows Productive Infection and Robust Immune Responses. PLoS One 9:e Ghiasi H, Osorio Y, Perng GC, Nesburn AB, Wechsler SL Recombinant herpes simplex virus type 1 expressing murine interleukin-4 is less virulent than wildtype virus in mice. J Virol 75: Ghiasi H, Osorio Y, Hedvat Y, Perng GC, Nesburn AB, Wechsler SL Infection of BALB/c mice with a herpes simplex virus type 1 recombinant virus expressing IFN-g driven by the LAT promoter. Virology 302: Ghiasi H, Osorio Y, Perng GC, Nesburn AB, Wechsler SL Overexpression of interleukin-2 by a recombinant herpes simplex virus type 1 attenuates pathogenicity and enhances antiviral immunity. J Virol 76: Perng GC, Dunkel EC, Geary PA, Slanina SM, Ghiasi H, Kaiwar R, Nesburn AB, Wechsler SL The latency-associated transcript gene of herpes simplex virus type 1 (HSV-1) is required for efficient in vivo spontaneous reactivation of HSV-1 from latency. J Virol 68: Allen SJ, Mott KR, Matsuura Y, Moriishi K, Kousoulas KG, Ghiasi H Binding of HSV-1 Glycoprotein K (gk) to Signal Peptide Peptidase (SPP) Is Required for Virus Infectivity. PLoS One 9:e Ghiasi H, Cai S, Slanina SM, Perng GC, Nesburn AB, Wechsler SL The Role of Interleukin (IL)-2 and IL-4 in Herpes Simplex Virus Type 1 Ocular Replication and Eye Disease. J Infect Dis 179: Matundan H, Mott KR, Ghiasi H Role of CD8+ T cells and myeloid DCs in protection from ocular HSV-1 challenge in immunized mice. J Virol 88: Ghiasi H, Kaiwar R, Nesburn AB, Wechsler SL Expression of herpes simplex virus type 1 glycoprotein B in insect cells. Initial analysis of its biochemical and immunological properties. Virus Res 22:
29 Mott KR, Ghiasi H Role of dendritic cells in enhancement of herpes simplex virus type 1 latency and reactivation in vaccinated mice. Clin Vaccine Immunol. 15: Zwaagstra JC, Ghiasi H, Nesburn AB, Wechsler SL Identification of a major regulatory sequence in the latency associated transcript (LAT) promoter of herpes simplex virus type 1 (HSV-1). Virology 182: Debroy C, Pederson N, Person S Nucleotide sequence of a herpes simplex virus type 1 gene that causes cell fusion. Virology 145: Bond VC, Person S Fine structure physical map locations of alterations that affect cell fusion in herpes simplex virus type 1. Virology 132: Little SP, Schaffer PA Expression of the syncytial (syn) phenotype in HSV-1, strain KOS: genetic and phenotypic studies of mutants in two syn loci. Virology 112: Pogue-Geile KL, Spear PG The single base pair substitution responsible for the Syn phenotype of herpes simplex virus type 1, strain MP. Virology 157: Foster TP, Melancon JM, Baines JD, Kousoulas KG The herpes simplex virus type 1 UL20 protein modulates membrane fusion events during cytoplasmic virion morphogenesis and virus-induced cell fusion. J Virol 78: Mo C, Suen J, Sommer M, Arvin A Characterization of Varicella-Zoster virus glycoprotein K (open reading frame 5) and its role in virus growth. J Virol 73: Klupp BG, Baumeister J, Dietz P, Granzow H, Mettenleiter TC Pseudorabies virus glycoprotein gk is a virion structural component involved in virus release but is not required for entry. J Virol 72: Dietz P, Klupp BG, Fuchs W, Kollner B, Weiland E, Mettenleiter TC Pseudorabies virus glycoprotein K requires the UL20 gene product for processing. J Virol 74:
30 Foster TP, Chouljenko VN, Kousoulas KG Functional and physical interactions of the herpes simplex virus type 1 UL20 membrane protein with glycoprotein K. J Virol 82: Downloaded from on January 18, 2019 by guest 30
31 Table 1. Survival of BALB/c and C57BL/6 mice following ocular challenge with gk recombinant viruses. a PFU/eye 2 X X X X x 10 2 Virus C57BL/6 C57BL/6 BALB/c BALB/c BALB/c BALB/c LD 50 (BALB/c), PFU NgK 4/5 12/12 0/5 1/5 5/5 5/5 15,642 MgK 5/5 12/12 4/5 4/5 5/5 5/5 2,115,650 RgK 5/5 10/10 1/5 2/5 5/5 5/5 27, a Mice were challenged ocularly with the specified amount of each virus and survival was 677 determined 28 days after challenge
32 Table 2. Corneal scarring of BALB/c and C57BL/6 mice following ocular infection with gk recombinant viruses. a PFU/eye 2 X 10 6b 2 X X X X 10 2 Virus C57BL/6 C57BL/6 BALB/c BALB/c BALB/c NgK 2.4± ± MgK 1.1± ± ± RgK 0.6± ±0.2 1±1 1.1± ±0.1 a Mice were infected ocularly with the specified amount of each virus and CS was determined 28 days after challenge. b Significantly different from RgK but not MgK using Student s t-test. 32
33 Fig 1. Myc-gK gene constructs used for the generation of NgK and MgK viruses. For construction of recombinant NgK and MgK viruses we synthesized the complete ORF of gk as follows: NgK) The structure of the wild-type gk molecule of 338 aa is shown with an in-frame insertion of c-myc sequence on C terminus. Positions of N-glycosylation sites are indicated at AA residues 48 and 58. The four possible gk transmembrane (TM) regions and the 30 amino acid-signal sequence of wt gk are shown; and MgK) The structure of mutated form of gk is similar to that of NgK, except five of the amino acids within the signal sequences of wt gk were mutated as follows: I16A, T17G, Y19S, L21I, and L23I. The NgK and MgK constructs were inserted into the BamHI site of plat as described in Materials and Methods. Downloaded from on January 18, 2019 by guest 33
34 Fig. 2. Construction and structure of the NgK and MgK mutant viruses. (A) The top schematic diagram shows the wt HSV-1 McKrae genome in the prototypic orientation. TR L and IR L represent the terminal and internal (or inverted) long repeats, respectively, and TR S and IR S represent the terminal and internal (or inverted) short repeats, respectively. U L and U S represent the long and short unique regions, respectively. The solid rectangle represents the very stable 2 kb LAT. The start site for LAT transcription is indicated by the arrow at +1. TATA designates the relative location of the LAT promoter TATA box 28 nt upstream of the start of transcription; (B) dlat2903 has a deletion from LAT nucleotides -161 to in both copies of LAT and makes no LAT RNA; (C) NgK was constructed from dlat2903 by homologous recombination between dlat2903 DNA and a plasmid containing the complete LAT promoter and the entire structural NgK gene (including sequences for myc) signal; (D) MgK was constructed from dlat2903 by homologous recombination between dlat2903 DNA and a plasmid containing the complete LAT promoter and the entire structural MgK gene (including mutated sequences within the gk signal sequences and myc sequences); and (E) NgK and MgK Southern. RS cell monolayers were infected with 5 PFU/cell of NgK, MgK, HSV-gK 3, or McKrae for 24 h. Viral DNAs were isolated, 5 g of DNA/each virus was digested with EcoRV /EcoRI, and hybridized to DIG-labeled HSV-1 gk (HSV-1 nucleotides ). Lanes: McKrae, HSV-gK 3, NgK, and MgK. As expected, only one fragment is seen in McKrae lane. This band also is observed in HSV-gK 3 and is correspondent to HSV-1 nucleotides surrounding the natural occurring gk (HSV-1 nucleotides ). In contrast to McKrae lane, in HSV-gK 3, NgK, and MgK lanes in addition to the natural gk, there are two additional bands corresponding to two extra copies of gk gene inserted within IR L (HSV-1 nucleotides to ) and TR L (HSV-1 nucleotides 13 to 7727) regions of dlat2903. Both regions within the IR L and TR L have a deletion of approximately 1600 bp of LAT. 34
35 Fig. 3. Replication of NgK and MgK viruses in vitro. Subconfluent RS cell monolayers were infected with 0.01 or 0.1 PFU/cell of NgK, MgK, or RgK virus as described in Materials and Methods. Total virus was harvested at the indicated times post-infection by two cycles of freeze-thawing. The amount of virus at each time point was determined by standard plaque assays on RS cells. Each point represents the mean ± SEM from two separate experiments (n=8). Panels: A) 0.01 PFU; and B) 0.1 PFU. Downloaded from on January 18, 2019 by guest 35
36 Fig. 4. Western blot analyses of expressed NgK and MgK proteins. RS cells were infected with the NgK, MgK, or RgK at an MOI of 10 PFU/cell for 16 h. The cells were then lysed directly into gel sample buffer, similar amount of protein was loaded into each well, and samples were separated by 10% SDS-PAGE, transferred to nitrocellulose, and reacted with anti-myc peptide antibody as described in Materials and Methods. Bound antibody was detected with chemoluminiscence detection followed by autoradiography. Lanes: MgK, NgK, RgK, Mock, and M (molecular weight markers). Downloaded from on January 18, 2019 by guest 36
37 Fig. 5. FACS analyses of NgK or MgK infected cells in vitro. RS cells were infected with the NgK, MgK, or RgK at an MOI of 5 PFU/cell for 24 h or were mock infected. At 24 h PI, cells were harvested, stained with anti-hsv-1 gc and anti-myc antibodies, fixed and subjected to FACS analyses as described in Materials and Methods. Stained cells were gated for HSV-1 gc or gk-myc expression and are shown as overlay. Panels: A) gc; and B) gk-myc. Downloaded from on January 18, 2019 by guest 37
38 Fig. 6. Immunostaining of NgK or MgK infected cells in vitro. RS cells were infected with the NgK, MgK, or RgK at an MOI of 5 PFU/cell for 24 h or mock infected. At 24 h PI, cells were fixed, permeabilized, blocked and stained with anti-hsv-1 gc (green), anti-myc (red) and DAPI nuclear stain (blue). Slides were fixed and photomicrographs are shown at 40X direct magnification and colocalization was visualized as yellow. Downloaded from on January 18, 2019 by guest 38
39 Fig. 7. Effect of NgK or MgK on apoptosis and cell death in vitro. RS cells were infected with the NgK, MgK, or RgK at an MOI of 5 PFU/cell for 24 h or mock infected. At 24 h PI, cells were harvested and reacted with Annexin-V and 7-ADD dye to analyze apoptosis and cell death, respectively. Stained cells were fixed and FACS analysis was performed as described in Materials and Methods. The percent of cells positive in each quadrant are shown for each virus or un-infected cells. Downloaded from on January 18, 2019 by guest 39
40 Fig. 8. Virus replication in mouse tears. C57BL/6 mice were ocularly-infected with 2 X 10 5 PFU/eye of NgK, MgK, or RgK virus. Tear films were collected on days 3 and 5 and virus titers were determined by standard plaque assays. Each point represents the mean titers of 20 eyes from two separate experiments Downloaded from on January 18, 2019 by guest 40
41 Fig. 9. Expression of gb and gk in cornea, TG, and brain of infected mice. C57BL/6 mice were ocularly infected with 2 X 10 5 PFU/eye of NgK, MgK, or RgK virus. gb and gk transcripts in cornea, TG and brain were determined on days 3 and 5 PI by qrt-pcr. In each experiment, an estimated relative copy number of the HSV-1 gb and gk was calculated using standard curves generated from pac-gb1 (45) and pac-gk1 (26). Briefly, DNA template was serially diluted 10-fold such that 5 l contained from 10 3 to copies of pac-gb1 or pac-gk1, then subjected to TaqMan PCR with the same set of primers. By comparing the normalized threshold cycle of each sample to the threshold cycle of the standard, the copy number for each reaction was determined. GAPDH expression was used to normalize the relative expression of each transcript in cornea, TG and brain of infected mice. Each bar represents the mean ± SEM from 6 corneas or TG and 3 brains. Panels: A) gb in cornea; B) gk in cornea; C) gb in TG; D) gk in TG; E) gb in brain; and F) gk in brain. 41
42 Fig. 10. Detection of gk and gb DNA in TG of latently infected mice. C57BL/6 mice were ocularly-infected with 2 x 10 5 PFU/eye of NgK, MgK, or RgK virus. TG from individual mice were isolated 28 days PI and TaqMan q-pcr was performed as described in Materials and Methods. GAPDH was used as an endogenous control to normalize the relative expression of gk and gb DNA in infected TGs. The relative copy number of the HSV-1 gb and gk genes were calculated using standard curves as described in figure 9 above. Each point represents the mean ± SEM from 12 TG (6 mice). Panels: A) gb DNA; and B) gk DNA. Downloaded from on January 18, 2019 by guest 42
43 Fig. 11. Effect of NgK and MgK on induced reactivation in latently infected mice. C57BL/6 mice were infected ocularly with 2 X 10 5 PFU/eye of NgK, MgK, or RgK virus, while some mice were infected with 2 X 10 6 PFU/eye of NgK or MgK virus. On day 28 PI, TG from infected mice was isolated and each individual TG incubated in 1.5 ml of tissue culture media at 37 o C. A 100 l aliquot was removed from each culture daily for 15 days and used to infect RS cell monolayers. The RS cells were monitored daily for the appearance of CPE for 5 days to determine the time of first appearance of reactivated virus from each TG. The results are plotted as the number of TG that reactivated daily. Numbers indicate the average time that the TG from each group first showed CPE ± SEM. Each point represents the mean ± SEM from 6 (NgK, 2 X 10 5 PFU/eye), 5 (MgK, 2 X 10 5 PFU/eye), 8 (RgK, 2 X 10 5 PFU/eye), 8 (NgK, 2 X 10 6 PFU/eye), and 8 (MgK, 2 X 10 6 PFU/eye). At 2 X 10 5 PFU/eye, 5 out of 12 TG in NgK infected mice, 7 out of 12 TG in MgK infected mice, and 2 out of 10 TG in RgK infected mice did not reactivated, while 8 out of 8 TG infected with 2 X 10 6 PFU/eye of NgK or MgK were reactivated. 43
44
45
46

47
48
49
following ocular infection of naive mice with a recombinant HSV-1 expressing murine IL-4 Dhong Hyun Lee 1 and Homayon Ghiasi 1,*
 JVI Accepted Manuscript Posted Online 28 February 2018 J. Virol. doi:10.1128/jvi.00051-18 Copyright 2018 American Society for Microbiology. All Rights Reserved. 1 2 An M2 rather than T H 2 response contributes
JVI Accepted Manuscript Posted Online 28 February 2018 J. Virol. doi:10.1128/jvi.00051-18 Copyright 2018 American Society for Microbiology. All Rights Reserved. 1 2 An M2 rather than T H 2 response contributes
Ocular infection with HSV-1 causes eye disease ranging in
 Role of Anti Glycoproteins D (Anti gd) and K (Anti gk) IgGs in Pathology of Herpes Stromal Keratitis in Humans Kevin R. Mott, 1 Yanira Osorio, 1 Ezra Maguen, 1 Anthony B. Nesburn, 2 Alec E. Wittek, 3,4
Role of Anti Glycoproteins D (Anti gd) and K (Anti gk) IgGs in Pathology of Herpes Stromal Keratitis in Humans Kevin R. Mott, 1 Yanira Osorio, 1 Ezra Maguen, 1 Anthony B. Nesburn, 2 Alec E. Wittek, 3,4
The Role of LAT in Increased CD8 T Cell Exhaustion in Trigeminal Ganglia of Mice Latently Infected with Herpes Simplex Virus 1
 JOURNAL OF VIROLOGY, May 2011, p. 4184 4197 Vol. 85, No. 9 0022-538X/11/$12.00 doi:10.1128/jvi.02290-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. The Role of LAT in Increased
JOURNAL OF VIROLOGY, May 2011, p. 4184 4197 Vol. 85, No. 9 0022-538X/11/$12.00 doi:10.1128/jvi.02290-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. The Role of LAT in Increased
Inclusion of CD80 in HSV Targets the Recombinant Virus to PD-L1 on DCs and Allows Productive Infection and Robust Immune Responses
 Inclusion of CD80 in HSV Targets the Recombinant Virus to PD-L1 on DCs and Allows Productive Infection and Robust Immune Responses The Harvard community has made this article openly available. Please share
Inclusion of CD80 in HSV Targets the Recombinant Virus to PD-L1 on DCs and Allows Productive Infection and Robust Immune Responses The Harvard community has made this article openly available. Please share
Overexpression of Interleukin-2 by a Recombinant Herpes Simplex Virus Type 1 Attenuates Pathogenicity and Enhances Antiviral Immunity
 JOURNAL OF VIROLOGY, Sept. 2002, p. 9069 9078 Vol. 76, No. 18 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.18.9069 9078.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Overexpression
JOURNAL OF VIROLOGY, Sept. 2002, p. 9069 9078 Vol. 76, No. 18 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.18.9069 9078.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Overexpression
on November 21, 2018 by guest
 JOURNAL OF VIROLOGY, Oct. 1998, p. 7715 7721 Vol. 72, No. 10 0022-538X/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Local Periocular Vaccination Protects against
JOURNAL OF VIROLOGY, Oct. 1998, p. 7715 7721 Vol. 72, No. 10 0022-538X/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Local Periocular Vaccination Protects against
A Therapeutic Vaccine That Reduces Recurrent Herpes Simplex Virus Type 1 Corneal Disease
 A Therapeutic Vaccine That Reduces Recurrent Herpes Simplex Virus Type 1 Corneal Disease Anthony B. Nesburn, 1 ' 2 Rae Lyn Burke, 5 Homayon Ghiasi, 1 ' 2 Susan M. Slanina, 1 Steven L Wechsler 1 ' 2 and
A Therapeutic Vaccine That Reduces Recurrent Herpes Simplex Virus Type 1 Corneal Disease Anthony B. Nesburn, 1 ' 2 Rae Lyn Burke, 5 Homayon Ghiasi, 1 ' 2 Susan M. Slanina, 1 Steven L Wechsler 1 ' 2 and
Nature Immunology: doi: /ni Supplementary Figure 1. Production of cytokines and chemokines after vaginal HSV-2 infection.
 Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Supplementary Figure 1 Production of cytokines and chemokines after vaginal HSV-2 infection. C57BL/6 mice were (a) treated intravaginally with 20 µl of PBS or infected with 6.7x10 4 pfu of HSV-2 in the
Condition: Herpes Simplex Keratitis
 Condition: Herpes Simplex Keratitis Description: Herpes simplex infection is very common but usually remains latent. When the virus is reactivated it travels along the trigeminal nerve to cause local infection
Condition: Herpes Simplex Keratitis Description: Herpes simplex infection is very common but usually remains latent. When the virus is reactivated it travels along the trigeminal nerve to cause local infection
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Recovery of Herpes Simplex Virus From Oculor Tissues of Latently Infected Inbred Mice
 Investigative Ophthalmology & Visual Science, Vol. 29, No. 2, February 1988 Copyright Association for Research in Vision and Ophthalmology Recovery of Herpes Simplex Virus From Oculor Tissues of Latently
Investigative Ophthalmology & Visual Science, Vol. 29, No. 2, February 1988 Copyright Association for Research in Vision and Ophthalmology Recovery of Herpes Simplex Virus From Oculor Tissues of Latently
Role of the VP16-Binding Domain of vhs in Viral Growth, Host Shutoff Activity, and Pathogenesis
 JOURNAL OF VIROLOGY, Dec. 2004, p. 13562 13572 Vol. 78, No. 24 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.24.13562 13572.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Role
JOURNAL OF VIROLOGY, Dec. 2004, p. 13562 13572 Vol. 78, No. 24 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.24.13562 13572.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Role
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T
 Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
amounts of biologically functional and highly immunogenic glycoproteins (15-21). We vaccinated mice with an equal
. We vaccinated mice with an equal") JOURNAL OF VIROLOGY, Apr. 1994, p. 2118-2126 Vol. 68, No. 4 22-538X/94/$4.+ Copyright 1994, American Society for Microbiology Expression of Seven Herpes Simplex Virus Type 1 Glycoproteins (gb, gc, gd,
JOURNAL OF VIROLOGY, Apr. 1994, p. 2118-2126 Vol. 68, No. 4 22-538X/94/$4.+ Copyright 1994, American Society for Microbiology Expression of Seven Herpes Simplex Virus Type 1 Glycoproteins (gb, gc, gd,
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Animal hosts Natural host Laboratory animals Rabbits Mice Rats Hamsters Newborn or suckling rodents Animal models for viral pathogenesis 4 Growth of v
 Principles of Virology Department of Molecular Genetics & Microbiology Univ ersity of Florida, Gainesv ille, FL 1 Outline Virus cultivation Assay of viruses Virus genetics 2 Virus isolation Evidence of
Principles of Virology Department of Molecular Genetics & Microbiology Univ ersity of Florida, Gainesv ille, FL 1 Outline Virus cultivation Assay of viruses Virus genetics 2 Virus isolation Evidence of
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Oxford Expression Technologies Ltd
 Oxford Expression Technologies Ltd Founded in 2007 as a spin out from Oxford Brookes University and Natural Environment Research Council Technology based on the insect baculovirus expression vectors (BEVs)
Oxford Expression Technologies Ltd Founded in 2007 as a spin out from Oxford Brookes University and Natural Environment Research Council Technology based on the insect baculovirus expression vectors (BEVs)
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
Murine Cytomegalovirus with a Transposon Insertional Mutation at Open Reading Frame M35 Is Defective in Growth In Vivo
 JOURNAL OF VIROLOGY, July 2003, p. 7746 7755 Vol. 77, No. 14 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.14.7746 7755.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Murine Cytomegalovirus
JOURNAL OF VIROLOGY, July 2003, p. 7746 7755 Vol. 77, No. 14 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.14.7746 7755.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Murine Cytomegalovirus
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Identification of Microbes Lecture: 12
 Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
The toll-like receptor 4 ligands Mrp8 and Mrp14 play a critical role in the development of autoreactive CD8 + T cells
 1 SUPPLEMENTARY INFORMATION The toll-like receptor 4 ligands Mrp8 and Mrp14 play a critical role in the development of autoreactive CD8 + T cells Karin Loser 1,2,6, Thomas Vogl 2,3, Maik Voskort 1, Aloys
1 SUPPLEMENTARY INFORMATION The toll-like receptor 4 ligands Mrp8 and Mrp14 play a critical role in the development of autoreactive CD8 + T cells Karin Loser 1,2,6, Thomas Vogl 2,3, Maik Voskort 1, Aloys
Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS
 Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS nucleotide sequences (a, b) or amino acid sequences (c) from
Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS nucleotide sequences (a, b) or amino acid sequences (c) from
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Viral vaccines. Lec. 3 أ.د.فائزة عبد هللا مخلص
 Lec. 3 أ.د.فائزة عبد هللا مخلص Viral vaccines 0bjectives 1-Define active immunity. 2-Describe the methods used for the preparation of attenuated live & killed virus vaccines. 3- Comparison of Characteristics
Lec. 3 أ.د.فائزة عبد هللا مخلص Viral vaccines 0bjectives 1-Define active immunity. 2-Describe the methods used for the preparation of attenuated live & killed virus vaccines. 3- Comparison of Characteristics
The humoral immune responses to IBV proteins.
 The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
INFLUENCE OF HERPES SIMPLEX VIRUS TYPE-1 GLYCOPROTEIN B
 INFLUENCE OF HERPES SIMPLEX VIRUS TYPE-1 GLYCOPROTEIN B EXPRESSION ON VIRAL PATHOGENECITY AND THE CD8 + T CELL RESPONSE by Srividya Ramachandran BSc. Biotechnology, Monash University, 2003 Submitted to
INFLUENCE OF HERPES SIMPLEX VIRUS TYPE-1 GLYCOPROTEIN B EXPRESSION ON VIRAL PATHOGENECITY AND THE CD8 + T CELL RESPONSE by Srividya Ramachandran BSc. Biotechnology, Monash University, 2003 Submitted to
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Supplementary Materials for
 immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
Ali Alabbadi. Bann. Bann. Dr. Belal
 31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
VIRAL TITER COUNTS. The best methods of measuring infectious lentiviral titer
 VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Transcription of the herpes simplex virus, type 1 genome during productive and quiescent. infection of neuronal and non-neuronal cells.
 JVI Accepts, published online ahead of print on 9 April 2014 J. Virol. doi:10.1128/jvi.00516-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 Transcription of the
JVI Accepts, published online ahead of print on 9 April 2014 J. Virol. doi:10.1128/jvi.00516-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 Transcription of the
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Comparison of Herpes Simplex Virus Reactivation in Ganglia In Vivo and in Explants Demonstrates Quantitative and Qualitative Differences
 JOURNAL OF VIROLOGY, July 2004, p. 7784 7794 Vol. 78, No. 14 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.14.7784 7794.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Comparison
JOURNAL OF VIROLOGY, July 2004, p. 7784 7794 Vol. 78, No. 14 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.14.7784 7794.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Comparison
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Supplementary Information Titles Journal: Nature Medicine
 Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Restricted VZV transcription in human trigeminal ganglia
 JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi:10.1128/jvi.01331-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Restricted VZV transcription
JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi:10.1128/jvi.01331-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Restricted VZV transcription
For the rapid, sensitive and accurate measurement of apoptosis in various samples.
 ab14082 500X Annexin V-FITC Apoptosis Detection Reagent Instructions for Use For the rapid, sensitive and accurate measurement of apoptosis in various samples. This product is for research use only and
ab14082 500X Annexin V-FITC Apoptosis Detection Reagent Instructions for Use For the rapid, sensitive and accurate measurement of apoptosis in various samples. This product is for research use only and
The Effect of Immunization with Herpes Simplex Virus Glycoprotein D Fused with Interleukin-2 against Murine Herpetic Keratitis
 The Effect of Immunization with Herpes Simplex Virus Glycoprotein D Fused with Interleukin-2 against Murine Herpetic Keratitis Tomoyuki Inoue*, Yoshitsugu Inoue*, Takao Nakamura*, Atsushi Yoshida*, Yumiko
The Effect of Immunization with Herpes Simplex Virus Glycoprotein D Fused with Interleukin-2 against Murine Herpetic Keratitis Tomoyuki Inoue*, Yoshitsugu Inoue*, Takao Nakamura*, Atsushi Yoshida*, Yumiko
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Clinical Profile of Herpes Simplex Keratitis
 K V Raju MS, Jyothi PT MS, Shimna Iqbal MS Clinical Profile of Herpes Simplex Keratitis Original Article Abstract Aims To document the various clinical presentations and to assess the risk factors contributing
K V Raju MS, Jyothi PT MS, Shimna Iqbal MS Clinical Profile of Herpes Simplex Keratitis Original Article Abstract Aims To document the various clinical presentations and to assess the risk factors contributing
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
HSV LAT AND NEURONAL SURVIVAL
 International Reviews of Immunology, 23: 187 198, 2004 Copyright # Taylor & Francis Inc. ISSN: 0883-0185 print/1563-5244 online DOI: 10.1080=08830180490265592 HSV LAT AND NEURONAL SURVIVAL DAVID C. BLOOM
International Reviews of Immunology, 23: 187 198, 2004 Copyright # Taylor & Francis Inc. ISSN: 0883-0185 print/1563-5244 online DOI: 10.1080=08830180490265592 HSV LAT AND NEURONAL SURVIVAL DAVID C. BLOOM
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
Norgen s HIV proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad icycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells
 The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells Jeremy Herskowitz, Emory University Meagan A. Jacoby, Emory University Samuel Speck, Emory University
The murine gammaherpesvirus 68 M2 gene is required for efficient reactivation from latently infected B cells Jeremy Herskowitz, Emory University Meagan A. Jacoby, Emory University Samuel Speck, Emory University
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Norgen s HIV Proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad T1000 Cycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
CURRICULUM VITAE Shahla Z. Abghari
 Shahla Z. Abghari, Ph. D. Updated March 4, 2014 PAGE 1 CURRICULUM VITAE Shahla Z. Abghari U. S. CITIZENSHIP: April 1992 EDUCATION: 1966-1970 B.S., Biology, Teachers University, Tehran, Iran 1967-1971 B.A.,
Shahla Z. Abghari, Ph. D. Updated March 4, 2014 PAGE 1 CURRICULUM VITAE Shahla Z. Abghari U. S. CITIZENSHIP: April 1992 EDUCATION: 1966-1970 B.S., Biology, Teachers University, Tehran, Iran 1967-1971 B.A.,
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Supplementary information
 Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
With a staggering one billion individuals worldwide
 Immunology and Microbiology A Herpes Simplex Virus Type 1 Human Asymptomatic CD8 þ T-Cell Epitopes-Based Vaccine Protects Against Ocular Herpes in a Humanized HLA Transgenic Rabbit Model Ruchi Srivastava,
Immunology and Microbiology A Herpes Simplex Virus Type 1 Human Asymptomatic CD8 þ T-Cell Epitopes-Based Vaccine Protects Against Ocular Herpes in a Humanized HLA Transgenic Rabbit Model Ruchi Srivastava,
Human Rotavirus A. genesig Advanced Kit. Non structural protein 5 (NSP5) 150 tests. Primerdesign Ltd. For general laboratory and research use only
 150 tests. Primerdesign Ltd. For general laboratory and research use only") TM Primerdesign Ltd Human Rotavirus A Non structural protein 5 (NSP5) genesig Advanced Kit 150 tests For general laboratory and research use only 1 Introduction to Human Rotavirus A Rotavirus is a genus
TM Primerdesign Ltd Human Rotavirus A Non structural protein 5 (NSP5) genesig Advanced Kit 150 tests For general laboratory and research use only 1 Introduction to Human Rotavirus A Rotavirus is a genus
numbe r Done by Corrected by Doctor
 numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
Retro-X qrt-pcr Titration Kit User Manual
 Takara Bio USA Retro-X qrt-pcr Titration Kit User Manual Cat. No. 631453 PT3952-1 (030218) 1290 Terra Bella Avenue, Mountain View, CA 94043, USA U.S. Technical Support: techus@takarabio.com United States/Canada
Takara Bio USA Retro-X qrt-pcr Titration Kit User Manual Cat. No. 631453 PT3952-1 (030218) 1290 Terra Bella Avenue, Mountain View, CA 94043, USA U.S. Technical Support: techus@takarabio.com United States/Canada
CD8α Dendritic Cells Drive Establishment of HSV-1 Latency
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Veterinary and Biomedical Science Veterinary and Biomedical Sciences, Department of 4-2014 CD8α Dendritic Cells
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Veterinary and Biomedical Science Veterinary and Biomedical Sciences, Department of 4-2014 CD8α Dendritic Cells
Received 22 November 1996/Accepted 3 April 1997
 JOURNAL OF VIROLOGY, July 1997, p. 5012 5024 Vol. 71, No. 7 0022-538X/97/$04.00 0 Copyright 1997, American Society for Microbiology Herpes Simplex Virus Type 1 Glycoprotein K Is Not Essential for Infectious
JOURNAL OF VIROLOGY, July 1997, p. 5012 5024 Vol. 71, No. 7 0022-538X/97/$04.00 0 Copyright 1997, American Society for Microbiology Herpes Simplex Virus Type 1 Glycoprotein K Is Not Essential for Infectious
The Severity of Herpes Simplex Viral Keratitis in Mice Does Not Reflect the Severity of Disease in Humans
 Investigative Ophthalmology & Visual Science, Vol. 33, No. 2, February 992 Copyright Association for Research in Vision and Ophthalmology The Severity of Herpes Simplex Viral Keratitis in Mice Does Not
Investigative Ophthalmology & Visual Science, Vol. 33, No. 2, February 992 Copyright Association for Research in Vision and Ophthalmology The Severity of Herpes Simplex Viral Keratitis in Mice Does Not
Human Rotavirus A. genesig Standard Kit. Non structural protein 5 (NSP5) 150 tests. Primerdesign Ltd. For general laboratory and research use only
 150 tests. Primerdesign Ltd. For general laboratory and research use only") TM Primerdesign Ltd Human Rotavirus A Non structural protein 5 (NSP5) genesig Standard Kit 150 tests For general laboratory and research use only 1 Introduction to Human Rotavirus A Rotavirus is a genus
TM Primerdesign Ltd Human Rotavirus A Non structural protein 5 (NSP5) genesig Standard Kit 150 tests For general laboratory and research use only 1 Introduction to Human Rotavirus A Rotavirus is a genus
Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved
 1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
Chronic Infections by Herpes Simplex Viruses and by the Horse and Cat Herpesviruses
 INFECTION AND IMMUNITY, Apr. 70, p. 351-355 Copyright 70 American Society for Microbiology Vol. 1, No. 4 Printed in U.S.A. Chronic Infections by Herpes Simplex Viruses and by the Horse and Cat Herpesviruses
INFECTION AND IMMUNITY, Apr. 70, p. 351-355 Copyright 70 American Society for Microbiology Vol. 1, No. 4 Printed in U.S.A. Chronic Infections by Herpes Simplex Viruses and by the Horse and Cat Herpesviruses
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Disruption of the M2 Gene of Murine Gammaherpesvirus 68 Alters Splenic Latency following Intranasal, but Not Intraperitoneal, Inoculation
 JOURNAL OF VIROLOGY, Feb. 2002, p. 1790 1801 Vol. 76, No. 4 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.4.1790 1801.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Disruption
JOURNAL OF VIROLOGY, Feb. 2002, p. 1790 1801 Vol. 76, No. 4 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.4.1790 1801.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Disruption
General information. Cell mediated immunity. 455 LSA, Tuesday 11 to noon. Anytime after class.
 General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
Received 12 June 2002/Accepted 13 September 2002
 JOURNAL OF VIROLOGY, Dec. 2002, p. 12758 12774 Vol. 76, No. 24 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.24.12758 12774.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. General
JOURNAL OF VIROLOGY, Dec. 2002, p. 12758 12774 Vol. 76, No. 24 0022-538X/02/$04.00 0 DOI: 10.1128/JVI.76.24.12758 12774.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. General
Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains
 Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains Name: Claire Ostertag-Hill Mentor: Dr. Ling Jin Bovine herpesvirus Bovine herpesvirus-1 (BHV-1) Pathogen
Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains Name: Claire Ostertag-Hill Mentor: Dr. Ling Jin Bovine herpesvirus Bovine herpesvirus-1 (BHV-1) Pathogen
Disease caused by herpes simplex virus
 Recurrence of herpes simplex virus in rabbit eyes: Results of a three-year study Peter R. Laibson and Sidney Kibrick Spontaneous reactivation of herpes simplex virus in rabbit ocular tissue was found on
Recurrence of herpes simplex virus in rabbit eyes: Results of a three-year study Peter R. Laibson and Sidney Kibrick Spontaneous reactivation of herpes simplex virus in rabbit ocular tissue was found on
Chapter 5. Virus isolation and identification of measles and rubella in cell culture
 Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Gene Vaccine Dr. Sina Soleimani
 Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Received 29 May 2003/Accepted 24 November 2003
 JOURNAL OF VIROLOGY, Mar. 2004, p. 3184 3189 Vol. 78, No. 6 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.6.3184 3189.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. A Mutation
JOURNAL OF VIROLOGY, Mar. 2004, p. 3184 3189 Vol. 78, No. 6 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.6.3184 3189.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. A Mutation
Received 21 July 2005/Accepted 12 October 2005
 JOURNAL OF VIROLOGY, Jan. 2006, p. 440 450 Vol. 80, No. 1 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.1.440 450.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Point Mutations
JOURNAL OF VIROLOGY, Jan. 2006, p. 440 450 Vol. 80, No. 1 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.1.440 450.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Point Mutations
PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human
 with 5% human") Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
Avian Influenza A H5N8
 TM Primerdesign Ltd Avian Influenza A H5N8 Hemagglutinin (HA) gene & Neuraminidase (NA) gene genesig Standard Kit 150 tests For general laboratory and research use only 1 Introduction to Avian Influenza
TM Primerdesign Ltd Avian Influenza A H5N8 Hemagglutinin (HA) gene & Neuraminidase (NA) gene genesig Standard Kit 150 tests For general laboratory and research use only 1 Introduction to Avian Influenza
For in vitro Veterinary Diagnostics only. Kylt Rotavirus A. Real-Time RT-PCR Detection.
 For in vitro Veterinary Diagnostics only. Kylt Rotavirus A Real-Time RT-PCR Detection www.kylt.eu DIRECTION FOR USE Kylt Rotavirus A Real-Time RT-PCR Detection A. General Kylt Rotavirus A products are
For in vitro Veterinary Diagnostics only. Kylt Rotavirus A Real-Time RT-PCR Detection www.kylt.eu DIRECTION FOR USE Kylt Rotavirus A Real-Time RT-PCR Detection A. General Kylt Rotavirus A products are
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Cytomegalovirus (CMV) End-Point PCR Kit Product# EP36300
 End-Point PCR Kit Product# EP36300") 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cytomegalovirus (CMV) End-Point PCR Kit Product# EP36300 Product
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cytomegalovirus (CMV) End-Point PCR Kit Product# EP36300 Product
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The