Title. Issue Date Doc URL. Type. File Information. in the medaka. article (author version)
|
|
|
- Ilene Burke
- 6 years ago
- Views:
Transcription
1 Title Gelatinase A and membrane-type matrix metalloprotein in the medaka Author(s)Ogiwara, Katsueki; Takano, Naoharu; Shinohara, Masak CitationProceedings of the National Academy of Sciences of t Issue Date Doc URL Type article (author version) File Information PNAS pdf Instructions for use Hokkaido University Collection of Scholarly and Aca
2 Classification: Biological Sciences (Animal Science or Reproduction) Gelatinase A and membrane-type matrix metalloproteinases 1 and 2 are responsible for follicle rupture during ovulation in the medaka Katsueki Ogiwara *, Naoharu Takano *, Masakazu Shinohara *, Masahiro Murakami, and Takayuki Takahashi * * Division of Biological Sciences, Graduate School of Science, Hokkaido University, Sapporo , Japan; and Amato Pharmaceutical Products, LTD., Fukuchiyama, Kyoto , Japan Correspondence: Takayuki Takahashi, Ph.D., Division of Biological Sciences, Graduate School of Science, Hokkaido University, Sapporo , Japan Tel.: ; Fax: ; ttakaha@sci.hokudai.ac.jp Manuscript information: 24 text pages and 5 figures Word and character counts: Abstract: 149 words; Paper: 46,773 characters Abbreviations: MMP, matrix metalloproteinase; TIMP, tissue inhibitor of metalloproteinase. Data deposition: The sequences reported in this paper have been deposited in the DDBJ database (accession nos. AB185847, AB072928, AB072929, AB185849, AB193468, and AB193469)
3 Identification of the hydrolytic enzymes involved in follicle rupture during vertebrate ovulation remains a central challenge for research in reproductive biology. Here we report a novel approach to this problem using an in vitro ovulation system in the medaka, Oryzias latipes, which is a small freshwater teleost. We found that follicle rupture in the medaka ovary involves the cooperation of at least three matrix metalloproteinases (MMPs), together with the tissue inhibitor of metalloproteinase-2b (TIMP-2b) protein. We determined the discrete roles of each of these proteins during follicle rupture. Our results indicated that gelatinase A induces the hydrolysis of type IV collagen constituting the basement membrane, membrane-type 2 MMP (MT2-MMP) degrades type I collagen present in the theca cell layer, and MT1-MMP and TIMP-2b are involved in the production and regulation of gelatinase A. These findings will help clarify the mechanism of follicle wall degradation during ovulation in mammalian species. Keywords: ovulation; follicle rupture; matrix metalloproteinases; medaka fish
4 Ovulation, which is triggered by a preovulatory surge of luteinizing hormone (LH) released from the pituitary gland, is a dynamic process that results in the liberation of a mature fertilizable ovum from the ovarian follicle. This event, which is known as follicle rupture, has been the subject of intensive investigation over the past century (1-5). Previous studies carried out primarily in mammalian species, including humans, have established that follicle rupture is accomplished by the dissolution of the granulosa cell basement membrane and fragmentation of the collagenous matrix at the apex of the follicular wall, thereby implicating the involvement of proteolytic enzymes. Indeed, a variety of proteases have been proposed as candidates for rupturing the follicle. It is generally believed that follicle rupture during ovulation takes place due to the actions of two proteolytic enzyme systems: the plasminogen activator (PA)/plasmin system (3, 6, 7) and the MMP system (8-11). However, it has been demonstrated that most mouse strains lacking individual proteases retain apparently normal reproductive ability, which is not consistent with this hypothesis (5, 12). A few MMP-deficient mice, including those lacking membrane-type 1 (MT1) MMP (13), and a disintegrin and metalloproteinase domain-17 (ADAM-17) (14), die in utero or shortly after birth; thus, the roles played by these proteases in ovulation remain to be clarified. In addition, the involvement of cathepsin L, and a disintegrin and MMP domain with thrombospondin-like motifs (ADAMTS-1), has been suggested by studies of mice lacking progesterone receptors (15). Mice null for ADAMTS-1 have been shown to develop fewer mature follicles (16), although its relationship to follicle rupture is not clear. A recent study suggested that one function of ADAMTS-1 in ovulation is to cleave versican in the matrix of the expanded cumulus-oocyte complex (17). In short, the proteases that are essential for follicle rupture in ovulation have not yet been
5 identified, despite much effort. In all vertebrates, the growth and proliferation of oogonia, their development to the oocyte stage, and their eventual release from the ovaries are thought to be under similar endocrine regulation (18-20), although the basic ovarian plan has several morphological variants. When searching for the fundamental mechanisms that are common to vertebrate ovaries, the use of the medaka, Oryzias latipes, which is a small freshwater teleost, has several advantages because of its short generation time and the cyclic nature of ovarian activity in mature adults (21, 22): under a constant long photoperiod of 14-h light/10-h dark at 27 C, the medaka usually spawns daily within 1 h of the onset of light for several consecutive days. Thus, we can readily time the successive events of spawning, such as completion of vitellogenesis, breakdown of the germinal vesicle, and ovulation (23). Moreover, using this fish, the process of follicle rupture and oocyte extrusion can be observed in vitro in isolated intact follicles (24, 25). We therefore investigated vertebrate ovulatory processes in the medaka, with the particular aim of identifying the enzymes responsible for the proteolytic degradation of the follicle walls. In the present study, we initially designed an in vitro experimental system for the study of ovulation using dissected ovarian follicles, and we then used this system to examine the effects of various protease inhibitors on the rate of ovulation. The finding that inhibitors of MMPs drastically suppressed in vitro ovulation prompted us to further explore the spatial and temporal expression patterns of most, if not all, of the MMP genes expressed in the follicular tissue of the medaka at both the mrna and protein levels. We also examined the relevance of individual MMPs to the ovulatory process. Materials and Methods
6 Medaka and In Vitro Ovulation Mature female adults of the orange-red variety of medaka were kept in indoor tanks under reproductive conditions (photoperiod, 10-h dark/14-h light; temperature, 27 C). Except where indicated, ovaries were removed at 6 or 3 h of ovulation and were placed in aseptic saline solution (26). Ovarian follicles were immediately isolated using forceps under a dissecting microscope and were then transferred into 90% medium 199 solution (Earle s medium 199; Dainippon Seiyaku), adjusted to ph 7.4 with NaHCO 3. At least 20 follicles per culture dish were used in each experiment. The follicles were cultured at C in 4 ml of culture medium using a mm tissue-culture dish. Ovulation was monitored every hour, and the number of oocytes that had successfully ovulated was counted. The ovulation rate was defined as the percentage of ovulated follicles at a given time. The process of ovulation is indicated in hours relative to the beginning of the light period, set at 0 h. The follicles were incubated with various protease inhibitors, including the MMP inhibitors TAPI-1 (Peptide Institute), TAPI-2 (Peptide Institute), and GM6001 (Chemicon), in order to assess their effects on in vitro ovulation. The concentrations of inhibitors were as follows: EDTA, 2 mm; o-phenanthroline, 1 mm; TAPI-1, 0.1 mm; TAPI-2, 0.1 mm; GM6001, 10 μm; diisopropyl fluorophospate (DFP), 0.2 mm; phenylmethanesulfonyl fluoride (PMSF), 0.2 mm; benzamidine, 0.2 mm; antipain, 0.1 mm; chymostatin, 0.1 mm; leupeptin, 0.1 mm; E-64, 0.2 mm; iodoacetic acid, 50 μm; and pepstatin, 10 μm. For the experiments using antibodies, purified antibody fractions (100 μg) were included in the culture system. IgG fractions prepared from pre-immune rabbit antiserum by means of a protein G-Sepharose column were used as controls. In
7 some experiments, actinomycin D (Sigma), cycloheximide (Sigma), and EDTA were added for various periods of incubation. Isolation of Oocytes and Follicle Layers from Ovulating Follicles Follicles that were about to ovulate were isolated as described above. Before spontaneous in vitro ovulation, the oocytes were mechanically separated from the follicles using forceps. A follicle devoid of its oocyte is composed of a single inner layer of granulosa cells and a single layer of outer theca cells separated by a basement membrane; the oocyte-free follicle thus prepared was referred to as the follicle layer. After ovulation, the oocytes and follicle layers were collected. Preparations of Extracts and Culture Media for Immunoblotting Except where indicated, whole ovaries, dissected follicles, isolated oocytes, and follicle layer tissues were homogenized in phosphate-buffered saline (PBS) and centrifuged at 13,000 g for 10 min to obtain supernatant fractions and precipitates. The supernatants were used directly for blotting, whereas the precipitate fractions were further boiled in 1% SDS and centrifuged at 13,000 g for 10 min, and then the resulting supernatants were used. The in vitro follicle culture media were concentrated before use. Samples or immune precipitates were analyzed by Western blot analysis. The primary antibodies were affinity-purified antibodies produced using blot membranes or anti-human TIMP-2 monoclonal antibody (Santa Cruz Biotechnology, Inc.). Immunoprecipitation and Zymography Ovarian follicle extracts were prepared as described above. Protein G-Sepharose
8 beads were treated with buffer A (20 mm Tris HCl (ph 8.0), 0.15 M NaCl, and 0.05% Tween-20) containing 0.1% BSA for 30 min at 4 C. After washing the beads with buffer A, they were incubated with purified antibodies for 1 h at 4 C and washed with buffer B (50 mm Tris HCl (ph 8.0), 0.15 M NaCl, 1% Triton X-100, and 0.1% SDS). The beads were then incubated at 4 C for 16 h with the extracts or in vitro ovulation culture media that had been added to the same volume of 2 buffer B. The beads were washed four times, incubated with SDS-sample buffer without β-mercaptoethanol for 6 h at room temperature, and analyzed by gelatin zymography. Cloning, RT-PCR, in situ hybridization, recombinant protein preparation, enzyme assays, antibody generation, immunohistochemistry, and the medaka type I collagen isolation procedures are described in Supporting Methods. Results Metalloproteinase Inhibitors Inhibit In Vitro Ovulation. Follicles isolated at various time points from the ovaries of the medaka were cultured at C in order to determine the rate of ovulation. Throughout the study, we defined the time of ovulation as the point at which the fish were expected to ovulate in vivo: this corresponded to the start of the light period and was set to 0 h. Follicles that were isolated 2 6 h before the predicted time of ovulation ovulated in vitro (Fig. 1A). When follicles were isolated 7 h before the predicted onset of ovulation (that is, at 7 h), the ovulation rate was reduced. Ovulated oocytes were readily recognized by the appearance of freed attaching filaments on their surface (Fig. 1B). Under these conditions, breakdown of the germinal vesicle occurred in almost all of the ovulated oocytes examined. Ovulated oocytes were successfully fertilized and could
9 subsequently develop into adults; approximately 75% of the fertilized oocytes developed normally to hatching (data not shown). Based on these observations, the in vitro ovulation experiments in this study were conducted using follicles isolated 3 or 6 h before the onset of the light phase. EDTA, o-phenanthroline, tumor necrosis factor-α protease inhibitor (TAPI-1), TAPI-2, and GM6001, all of which have been shown to be inhibitors of MMPs, drastically reduced the rate of ovulation (Fig. 1C). Follicles cultured in the presence of the inhibitors had apparently normal morphology (data not shown). When EDTA was included in the tissue cultures between 0 and 4 h, ovulation was completely suppressed (Fig. 1D). In addition, ovulation was almost completely suppressed when actinomycin D or cycloheximide was added between -5 and 0 h (Fig. 1E). These results indicated that the synthesis of mrna and protein is required for follicle rupture during ovulation. The results also indicate that MMPs play important roles in the proteolytic events associated with the ovulatory process in the medaka. Medaka Ovarian Follicles Express at Least Seven Species of MMPs. In order to search for candidate MMPs that are involved in follicle rupture during ovulation in the fish, RT-PCR was performed with two degenerate oligonucleotide primers using total RNA isolated from fish ovaries at the time of ovulation. Subcloning of the amplified cdna fragments and subsequent analyses identified seven distinct medaka MMPs: gelatinase A (AB033754), gelatinase B (AB033755), stromelysin-3 (AB055705), MT1-MMP (AB185847), MT2-MMP (AB072928), MT3-MMP (AB072929) and MT5-MMP (AB047650). In situ hybridization analysis revealed that gelatinase A, MT1-MMP, MT3-MMP, MT5-MMP, and stromelysin-3 mrna were
10 expressed in small, growing oocytes, while gelatinase B and MT2-MMP mrna were localized in the tissues ovulated follicles (Fig. 6, Supporting Information). The results regarding gelatinase A, gelatinase B, MT5-MMP, and stromelysin-3 were consistent with the findings of our previous reports (27-29). Effects of the Medaka MMPs on Type I and Type IV Collagens. Active recombinant proteins of the seven MMPs were synthesized as fusion proteins, and the MT-MMPs were produced without cytoplasmic and transmembrane domains. All of the MMP preparations yielded a single protein band on SDS-PAGE under reducing conditions (Fig. 7A, Supporting Information). The gelatin-hydrolyzing activities of the MMPs were detected using gelatin zymography (Fig. 7B, Supporting Information). Follicle rupture during ovulation is thought to result from dissolution of the collagen fibers in the tunica albuginea and the theca externa, and the major species present in the connective tissue matrix are believed to be type I and type IV collagen. Therefore, we examined the effects of the medaka MMPs on medaka type I collagen. MT2-MMP degraded the collagen most rapidly (Fig. 2A), although hydrolysis was also observed with MT5-MMP. Medaka type I collagen was resistant to gelatinases A and B, MT1-MMP, MT3-MMP, and stromelysin-3. Because we were unable to test medaka type IV collagen due to the difficulty of obtaining this protein substrate in sufficient quantities, collagen of mammalian origin was used. Medaka gelatinases A and B were able to hydrolyze bovine type IV collagen (Fig. 2B). We also tested whether gelatinases A and B could act as substrates for MT-MMPs. For this purpose, recombinant proenzymes of the gelatinases were prepared. Incubation
11 of progelatinase A with active MT1-MMP, MT2-MMP, and MT3-MMP resulted in the conversion of the proenzyme (68 kda) to the mature enzyme (52 kda) via a 55-kDa intermediate (Fig. 7C, Supporting Information). MT5-MMP did not show any detectable activity towards progelatinase A. None of the MT-MMPs tested were able to activate progelatinase B (data not shown). Ovulating Follicles Express MMP Proteins in Distinct Temporal Patterns. In order to detect MMPs in the fish ovary, we prepared antibodies against the seven MMPs. Western blot analysis of the fish ovary extracts indicated that the fish ovary expresses gelatinases A and B, MT1-MMP, MT2-MMP, MT5-MMP, and stromelysin-3, whereas MT3-MMP was not detected at any of the levels of significance examined here (Fig. 8, Supporting Information). Immunohistochemical staining with antibodies for gelatinase A, MT1-MMP, MT5-MMP, and stromelysin-3 produced signals in the oocyte cytoplasm of small growing follicles (Fig. 9, Supporting Information). In contrast, anti-gelatinase B and anti-mt2-mmp antibodies revealed the presence of their respective antigens in the follicular tissue just after oocyte release. These results were consistent with those from the in situ hybridization experiments (Fig. 6, Supporting Information). Although we attempted to localize MT3-MMP immunohistochemically in the ovary sections, no clear signals were observed. This finding provided further support for the view that MT3-MMP mrna is not translated sufficiently in the oocyte to permit the detection of the corresponding protein. In order to gain a better understanding of the relationship between ovulation and MMPs, changes in the expression of the six MMPs during the course of in vitro ovulation were investigated using immunoprecipitation/western blotting or Western
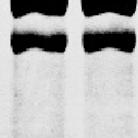
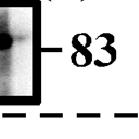
12 blotting with specific antibodies. Using the extracts of 20 isolated follicles that were ready to ovulate, immunoreactive materials were detected with antibodies to gelatinase A, gelatinase B, MT1-MMP, or MT2-MMP (Fig. 3A), but not with antibodies to MT5-MMP or stromelysin-3 (data not shown). Zymographic analyses of the immunoprecipitates revealed that the oocyte extract and the culture medium contained gelatinase A in two forms: 62- and 55-kDa proteins (Fig. 3B, Upper Two Panels). To clarify the relationship between these two forms of gelatinase A, we examined the effects of p-aminophenylmercuric acetate (APMA), which is a well-known activator of secreted MMPs (30), on these proteins. Treatment with this compound resulted in a strong reduction in the 62-kDa band, with concomitant appearance of a 59-kDa band in the oocyte extract and culture medium (Fig. 3C, Upper Panel). In addition, the 55-kDa protein remained present after APMA treatment. These results demonstrated that the 62-, 59-, and 55-kDa polypeptides are the pro-protein, intermediate protein, and mature active forms of gelatinase A, respectively. Immunoprecipitation followed by zymographic analyses for gelatinase B showed that the follicular layers expressed the 83-kDa polypeptide alone intracellularly, whereas the follicle culture medium contained an additional 71-kDa polypeptide (Fig. 3B). The results of APMA treatment confirmed that these were the pro- and active mature enzymes, respectively (Fig. 3C). Based on these results, changes in the intracellular and extracellular levels of proand mature active MMPs during the course of ovulation can be summarized as shown in Figure 3D, assuming that the MT-MMPs that are detectable in the tissue extracts are predominantly in the active form. In fact, the sizes of the polypeptides detected using antibodies specific for MT1-MMP and MT2-MMP were in good agreement with the
13 theoretically predicted sizes. Since gelatinase A, gelatinase B, MT1-MMP, and MT2-MMP were present in an active enzymatic form in the follicles at the time of ovulation, these four MMPs may play a role in follicle rupture during ovulation. Specific Antibodies for Gelatinase A, MT1-MMP, and MT2-MMP Reduce the Rate of In Vitro Ovulation. We found that the addition of rabbit sera to the system caused the complete suppression of ovulation, regardless of whether antisera or non-immune control sera were used (data not shown). This was presumably due to protease inhibitors that were present in the sera in large quantities. Therefore, we purified IgGs and specific antibodies in order to isolate the serum protease inhibitors for further investigation. We confirmed that IgG fractions containing anti-mmp antibodies inhibited the enzyme activities of the respective antigens by more than 70% in vitro, using purified recombinant MMPs and FITC-labeled porcine gelatin type I as a substrate (Fig. 10, Supporting Information). Incubation of the follicles with anti-gelatinase A, anti-mt1-mmp, and anti-mt2-mmp antibodies resulted in a marked reduction of ovulation (Fig. 4A). The addition of antibodies together with the respective antigens to the culture nullified the ovulation-suppressing activities of the antibodies. Anti-gelatinase B and anti-mt3-mmp antibodies were found to exert no effect on the rate of ovulation. The combination of two MMP antibodies inhibited in vitro ovulation, although the extent of inhibition was similar to that produced by only one type of antibody alone. This was also the case for the combined effect of three MMP antibodies. These results indicate that gelatinase A, MT1-MMP, and MT2-MMP play important roles during in vitro ovulation in the medaka.
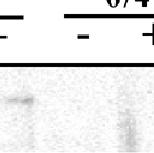
14 Regulation of Gelatinase A Activity by MT1-MMP and TIMP-2b in Cultured Follicles When follicles isolated at 6 h of ovulation were cultured in the presence of GM6001 or anti-mt1-mmp antibody for 5 h, the production of active gelatinase A was markedly reduced (Fig. 4B). The generation of active gelatinase A during the 5-h incubation period was not affected by the addition of anti-mt3-mmp antibody. The results revealed that MT1-MMP plays a role in the activation of progelatinase A in culture. We next investigated how active gelatinase A generated during the pre-ovulation period was secreted into the medium without leading to marked degradation of the basement membrane. One plausible explanation for the absence of degradation was that intrinsic MMP inhibitors might inhibit the activity of gelatinase A. To test this idea, we isolated three TIMP cdna clones TIMP-2a (AB185849), TIMP-2b (AB193468), and TIMP-3 (AB193469) from medaka ovary mrnas. Among these clones, TIMP-2b mrna was found to be the predominant transcript in fully-grown oocytes (Fig. 11, Supporting Information). We therefore focused on the role of TIMP-2b in the regulation of gelatinase A activity. The oocyte extracts and the culture medium of ovulating follicles contained TIMP-2b with an Mr of 28 kda (Fig. 4C, Uupper Panel; Fig. 12, Supporting Information). The maximal TIMP-2b band intensity was detected in the culture medium at 2 h of ovulation, and the signals from subsequent samples examined at 2-h intervals became steadily weaker. These results thus confirmed that the TIMP-2b protein was present in ovulating follicles and was most actively secreted from the oocytes of in vitro-cultured follicles during the pre-ovulation period. We further
15 analyzed the anti-timp-2b immunoprecipitates of the culture medium using Western blot analysis and gelatin zymography. A 65-kDa polypeptide was identified (under reducing conditions) using anti-gelatinase-a antibody (Fig. 4C, Middle Panel), and two polypeptides of 62 and 55 kda were detected (under non-reducing conditions) by examining the gelatinolytic activity (Fig. 4C, Lower Panel). These results demonstrated the presence of gelatinase A (and progelatinase A) in complex with TIMP-2b in the in vitro ovulation culture medium. In summary, our results demonstrated that TIMP-2b is expressed in ovulating follicles and reaches a maximum concentration during the preovulation period. These findings strongly suggest that this intrinsic inhibitor regulates gelatinase A activity at or around the time of ovulation. DISCUSSION In 1916, Schochet (1) first noted that the rupture of the Graafian follicle is due in part to the digestion of the theca folliculi by a proteolytic ferment or enzyme in the liquor folliculi. Since then, intense efforts have been made to identify the proteases that are essential for follicle rupture during ovulation, primarily in mammalian species. Although the identification of proteases responsible for follicle rupture remains to be achieved, previous studies have strongly suggested that this process is accomplished by the cooperative effects of several MMPs and their inhibitors (4, 5). Our current findings strongly suggested that this is indeed the case. The present study indicated that gelatinase A, MT1-MMP, and MT2-MMP play important roles in the process of follicle rupture during ovulation in the medaka. On the basis of our data, we can propose a mechanistic model of this process, implicating these
16 MMPs together with the intrinsic inhibitor TIMP-2b (Fig. 5). Namely, in the follicle that is destined to ovulate, the synthesis of progelatinase A and MT1-MMP begins in the oocyte at some point prior to ovulation. At the same time, the oocyte produces TIMP-2b. When MT1-MMP has been activated intracellularly and is localized on the surface of the oocyte, this membrane-bound MMP may interact with secreted progelatinase A and TIMP-2b to form the progelatinase A/MT1-MMP/TIMP-2b complex, which is well documented as the unique mechanism for progelatinase A activation in mammals (30-32). Progelatinase A activation in the follicles of the medaka ovary appears to occur in a similar manner. The present model is supported by the finding that the activation of progelatinase A in the culture medium of ovarian follicles was noticeably inhibited by the addition of anti-mt1-mmp antibodies. The expression of furin, which is likely to be involved in the activation of MT1-MMP, has recently been reported in the oocytes of ovarian follicles of the medaka (33). Active gelatinase A produced in this manner may be inhibited until the time of ovulation by the formation of a complex with TIMP-2b; this inhibition is thought to be due to a presumed excess of the inhibitor compared to the amount of enzyme (Fig. 5A). At the time of ovulation, the transcription and translation of MT2-MMP and gelatinase B are initiated in the granulosa cells of the ovulating follicle. The former may undergo furin-catalyzed intracellular activation, and may subsequently be distributed over the surface of the cells, while the latter is secreted from the same cells into the extracellular space in an inactive precursor form. Increased production of active gelatinase A and gelatinase B together with a decrease in TIMP-2b at, or around, the time of ovulation may reverse the molar ratio of MMP/TIMP-2b compared with that of the pre-ovulation period, resulting in the release of MMP from inhibition by TIMP-2b. Unlike MT1-MMP and MT2-MMP, gelatinase A is able to
17 degrade type VI collagen in vitro. Therefore, the basement membrane situated between the granulosa and theca cell layers might be a target substrate for this MMP (Fig. 5B). Although medaka gelatinase B was found to hydrolyze type IV collagen more rapidly than gelatinase A, the results of our in vitro ovulation experiments using specific antibodies indicated that the former does not participate in basement-membrane degradation. Basement-membrane degradation resulting from the activity of gelatinase A allows the granulosa cells to come into contact with the ECM in the interstitial space, where the theca cells are localized. As MT2-MMP hydrolyzed medaka collagen type I in vitro (Fig. 3), and because this type of collagen is thought to be a major ECM protein in this region, the MT2-MMP expressed on the surface membrane of the granulosa cells is thought to be responsible for the dissolution of the collagen (Fig. 5C). It should be noted that MT2-MMP may play an additional role in the activation of progelatinase A, as we demonstrated here that it was able to activate the precursor gelatinase in vitro. Follicle rupture during ovulation in the medaka does not require extensive ECM degradation throughout the layer surrounding the oocyte. As is the case in mammalian ovulation (2), hydrolysis of the ECM proteins at a single restricted site is expected to be sufficient for ovulation in the medaka. Once this local hydrolysis has occurred, the subsequent process appears to proceed spontaneously (Fig. 5D). Clearly, the mechanical force generated by the swelling of the ovulating oocyte facilitates ovulation. However, the mechanism and regulation of this restricted hydrolysis is still unknown. We should also note that externally added active recombinant MMPs, including gelatinase A and MT2-MMP, were found to have no accelerating effect on in vitro ovulation. This finding may indicate the importance of well-regulated molecular interactions between the intrinsic MMPs, TIMP-2b, and ECM proteins in follicle rupture. In summary, upon
18 ovulation, gelatinase A is responsible for the hydrolysis of type IV collagen that forms the basement membrane, while MT2-MMP degrades type I collagen present in the theca cell layer. Moreover, MT1-MMP and TIMP-2b are both involved in the activation of progelatinase A and the regulation of its activity. We believe that the present study identifies, for the first time, proteases that are critical for follicle rupture during ovulation in vertebrates. The use of similar experimental systems established for mammalian ovaries (34-37) will most likely aid in the determination of the enzymes that facilitate follicle rupture in mammals. The eventual identification of mammalian ovulatory proteases will provide information that is of potential clinical importance. Our current approach and findings are expected to be useful for future studies in this area. Acknowledgments We would like to thank Dr. Junji Ohnishi (Hokkaido University, Japan) for his valuable comments and discussions. This work was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan ( ), research fellowships from the Japan Society for the Promotion of Science for Young Scientists, and the 21 st COE program. References 1. Schochet, S. S. (1916) Anat. Rec. 10, Espey, L. L. (1967) Am. J. Physiol. 212, Tsafriri, A. (1995) Adv. Exp. Med. Biol. 377, Richards, J. (2001) Endocrinology 142,
19 5. Ny, T., Wahlberg, P. & Brändström, I. J. M. (2002) Mol. Cell. Endocr. 187, Beers, W. H. (1975) Cell 6, Murdoch, W. J. & McDonnel, A. C. (2002) Reproduction 123, Curry, T. E. Jr., Dean, D. D., Woessner, J. F. Jr., & LeMaire, W. J. (1985) Biol. Reprod. 33, Liu, K., Wahlberg, P. & Ny, T. (1998) Endocrinology 139, Hagglund, A. C., Ny, A., Leonardsson, G. & Ny, T. (1999) Endocrinology 140, Jo, M. & Curry, T. E. Jr. (2004) Biol. Reprod. 71, Pendás, A. M., Folgueras, A. R., Llano, E., Caterina, J., Frerard, F., Rodríguez, F., Astudillo, A., Noël, A., Birkedal-Hansen, H. & López-Otín, C. (2004) Mol. Cell Biol. 24, Holmbeck, K., Bianco, P., Caterina, J., Yamada, S., Kromer, M., Kuznetsov, S. A., Mankani, M., Robey, P. G., Poole, A. R., Piodoux, I. et al. (1999) Cell 99, Peschon, J. J., Slack, J. L., Reddy, P., Stocking, K. L., Sunnarborg, S. W., Lee, D. C., Russell, W. E., Nelson, N., Kozlosky, C. J., Wolfson, M. F. et al. (1998) Science 282, Robker, R. L., Russell, D. L., Espey, L. L., Lydon, J. P., O Malley, B. W. & Richards, J. S. (2000) Proc. Natl. Acad. Sci. USA 97, Shindo, T., Kurihara, H., Kuno, K., Yokoyama, H., Wada, T., Kurihara, Y., Imai, T., Wang, Y., Ogata, M, Nishimatsu, H. et al. (2000) J. Clin. Invest. 105, Russell, D. L., Doyle, K. M., Ochsner, S. A., Sandy, J. D. & Richards, J. S. (2003) J. Biol. Chem. 278, Schroeder, P. C. & Talbot, P. (1985) Gamete Res. 11,
20 19. Richards, J. (1994) Endocr. Rev. 15, Nagahama, Y., Yoshikuni, M., Yamashita, M. & Tanaka, M. (1994) in Fish Physiology, eds. Sherwood, N. M. & Hew, C. L. (Academic, New York), pp Ishikawa, Y. (2000) BioEssays 22, Wittbrodt, J., Shima, A. & Schartl, M. (2002) Nat. Rev. Genet. 3, Iwamatsu, T. (1978) J. Exp. Zool. 206, Hirose, K. & Donaldson, E. (1972) Bull. Jap. Soc. Scient. Fish. 38, Pendergrass, P. & Schroeder, P. (1976) J. Reprod. Fert. 47, Shibata, Y., Iwamatsu, T., Oba, Y., Kobayashi, D., Tanaka, M., Nagahama, Y., Suzuki, N. & Yoshikuni, M. (2000) J. Biol. Chem. 275, Matsui, H., Ogiwara, K., Ohkura, R., Yamashita, M. & Takahashi, T. (2000) Eur. J. Biochem. 267, Kimura, A., Shinohara, M., Ohkura, R. & Takahashi, T. (2001) Biochim. Biophys. Acta 1518, Ogiwara, K., Matsui, H., Kimura, A. & Takahashi, T. (2002) Mol. Reprod. Develop. 61, Nagase, H. (1997) Biol. Chem. 378, Sato, H., Takino, T., Okada, Y., Cao, J., Shinagawa, A., Yamamoto, E. & Seiki, M. (1994) Nature 370, Brew, K., Dinakarpandian, D. & Nagase, H. (2000) Biochim. Biophys. Acta 1477, Ogiwara, K., Shinohara, M. & Takahashi, T. (2004) Gene 337, Baranczuk, R. J. & Fainstat, T. (1976) Am. J. Obstet. Gynecol. 124,
21 35. Talbot, P. (1983) J. Exp. Zool. 225, Osman, P. & Lieuwma-Noordanus, C. (1980) J. Reprod. Fert. 59, Rose, U. M., Hanssen, R. G. J. M. & Kloosterboer, H.J. (1999) Biol. Reprod. 61,
22 Figure legends Fig. 1. In vitro ovulation in medaka and the effects of various chemicals. (A) Follicles obtained from mature medaka ovaries at various pre-ovulation times were incubated. The ovulation rates were determined, and the results are shown as the means of at least five experiments. (B) Follicles collected at 6 h were incubated. Photographs were taken at 2 and 4 h of ovulation. (C) Follicles collected at 3 h of ovulation were incubated with various protease inhibitors, and the ovulation rates were determined at 4 h. The results are shown as the means (± SD) of at least five experiments. (D) Follicles were collected at 3 h for in vitro ovulation. The timing of EDTA treatments (2 mm) is indicated by the thick bars in the left panel. The initiation and termination of the treatments were accomplished by rapid exchanges of the culture medium. The ovulation rates were determined at 4 h of ovulation. The results are presented as the means (± SD) of at least five experiments. (E) Follicles were collected and incubated at 5 h. Actinomycin D (1 μg/ml) or cycloheximide (1 μg/ml) was added to the culture. The timing of the treatments is indicated by the thick bars in the left panel. The other details remained as described above. Fig. 2. Hydrolysis of collagens by recombinant medaka MMPs. (A) Type I collagen purified from the whole body of the medaka fish was incubated with purified MMPs, and degradation of the collagen was analyzed by SDS-PAGE (reducing conditions). The results of control experiments without enzymes, but with medaka collagen (None) and bovine type I collagen, are also shown. Asterisks indicate the bands of recombinant enzymes that were added to the incubations. (B) FITC-labeled bovine type IV collagen was used as substrate for the enzymatic activity of the purified MMPs.
23 Fig. 3. Expression of MMPs in ovarian follicles. (A) Mature ovarian follicles were isolated at 6 h of ovulation and cultured in vitro. At the indicated times, the oocytes, follicular layers, and medium were obtained separately. The oocyte extracts were immunoprecipitated with rabbit anti-medaka gelatinase A antibodies, and the resulting precipitates were analyzed by Western blot analysis with rat anti-medaka gelatinase A antibodies. Extracts of the oocytes and follicle layer, and the concentrated media were used directly for the Western blot analysis. The sizes (kda) of the proteins are indicated. (B) The oocyte extracts and media were immunoprecipitated with rabbit anti-gelatinase A antibodies and the immunoprecipitated materials were analyzed using gelatin zymography (Upper Two Panels). Extracts of the follicular layers and medium were immunoprecipitated with rabbit anti-gelatinase B antibodies, and the immunoprecipitated materials were analyzed by gelatin zymography (Lower Two Panels). (C) The in vitro follicle culture medium at 2 h of ovulation was concentrated, treated with 1 mm APMA at 4 C overnight, and immunoprecipitated with anti-gelatinase A antibodies. The immunoprecipitated materials were then subjected to gelatin zymographic analysis (Upper Panel). Extracts of the follicular layers at 2 h of ovulation were treated with APMA, as described above, and were then immunoprecipitated with anti-gelatinase B antibodies. The immunoprecipitates were analyzed using gelatin zymography (Lower Panel). The sizes of the proteins are indicated. (D) The relative amounts of inactive proenzyme and active mature enzyme at each time point were calculated, based on the results shown in (A) and (B).
24 Fig. 4. Roles of MMPs and TIMP-2b in follicle rupture. (A) Follicles were incubated with pre-immune rabbit IgG (control) or specific antibodies or specific antibodies + antigens. The results are presented as the means (± SD) of five separate experiments. (B) Follicles isolated at 6 h were incubated with GM6001, anti-mt1-mmp, or anti-mt3-mmp antibodies. The medium collected at 0 h was immunoprecipitated with anti-gelatinase A antibodies, and the precipitates were analyzed using gelatin zymography (Upper Panel). Progelatinase A (62 kda) and activated gelatinase A (55 kda) are indicated. The intensities of the bands shown in the upper panel were quantified by densitometry, and the ratio of gelatinase A to progelatinase A in each treatment was determined (Lower Panel). The results are presented as the means (± SD) of three experiments. (C) Follicles isolated at 6 h were incubated. Culture media derived from the pre-ovulation period between 5 and 0 h ( 5/0) and from the post-ovulation period between 0 and 4 h (0/4) were obtained. Immunoprecipitation was conducted using anti-medaka TIMP-2b antibodies, and the precipitates were analyzed by Western blotting with mouse anti-human TIMP-2 monoclonal antibody (Upper Panel) or anti-gelatinase A antibody (Middle Ppanel), or by gelatin zymography (Lower Ppanel). Fig. 5. A Model of Follicle Rupture during Ovulation in the Medaka. (A) In the follicle, a few hours before ovulation, progelatinase A is activated by MT1-MMP on the surface of the oocyte, whereas activated gelatinase A is immediately inactivated by TIMP-2b. (B) At the time of ovulation, the hydrolysis of basement membrane type IV collagen is initiated by active gelatinase A at the follicle ovarian surface contact site. (C) MT2-MMP, which is now expressed on the surface of the
25 granulosa cells, can degrade the type I collagen that is present in the theca cell layer. (D) As a result, the oocyte is exposed at the contact site, leading to ovulation. BM, basement membrane; EC, epithelial cell; GC, granulosa cell; OC, oocyte; TC, theca cell.
















































































26 Figure 1 Ogiwara et al.



















































27 Figure 2 Ogiwara et al.

























































































28 Figure 3 Ogiwara et al.












































29 Figure 4 Ogiwara et al.




















































































30 Figure 5 Ogiwara et al.
31 Supporting Information Supporting Methods Cloning of Various Medaka cdnas RNA was isolated from the medaka ovary (Nippon Gene). The first-strand cdna synthesis was performed using a SuperScript First-Strand Synthesis System (Life Technologies, Inc.). Full-length cdnas were obtained by initial PCR amplification of fragments using two degenerate oligonucleotide primers followed by library screening or 5 /3 -RACE. Primers and cloning strategies are described in Tables 1 and 2. RT-PCR Total RNAs were prepared from various tissues and treated with DNAse (Roche Molecular Biochemicals). Aliquots of 2 μg of the treated RNAs were used for reverse transcription. PCR was performed for 25 or 30 cycles using Ex Taq DNA polymerase (Takara). The primers and amplified products are summarized in Table 3. In Situ Hybridization In situ hybridization was performed using frozen ovary sections (15 μm) as described previously (1). RNA probes were prepared by in vitro transcription of reverse-transcriptase fragments of cdnas with T3 or T7 RNA polymerase using a digoxigenin (DIG) RNA-labeling kit (Boehringer-Mannheim). The regions and sizes of the specific probes used are described in Table 3. The hybridization was conducted at 50 C for 18 h in 50% formamide, 5 Denhardt s solution, 6 SSPE, and 0.5 mg/ml
32 yeast transfer RNA (for gelatinase A, gelatinase B, MT1-MMP, MT3-MMP, and stromelysin-3) or 50% formamide, 0.5 M NaCl, 10 mm Tris-HCl (ph 8.0), 1 mm EDTA, 10% dextran sulfate (Wako), 1 Denhardt s solution, 0.25% SDS, and 0.2 mg/ml yeast transfer RNA (for MT2-MMP, MT5-MMP, and TIMP-2b). The sections were washed at 50 C in 50% formamide/2 SSC for 30 min, washed once in 2 SSC for 20 min and twice in 0.2 SSC for 20 min at 50 C. The hybridization probes were detected using a Dig Nucleic Acid Detection Kit (Roche Molecular Biochemicals). Preparation of Recombinant Proteins Using the pet30a expression vector (Novagen) in the Escherichia coli expression system, recombinant proteins were produced as a fusion protein with an extra amino-acid sequence of residues (varying with different proteins) at its NH 2 -terminus; the vector-derived amino-terminal stretch contained a His-tag and an S-protein sequence. Harvested cells were lysed and the insoluble materials were dissolved in a solubilization buffer containing 6 M urea, 50 mm Tris HCl (ph 7.8), and 0.5 M NaCl. Solubilized proteins were subjected to affinity chromatography on Ni 2+ -Sepharose (Amersham Pharmacia Biotech). Eluted recombinant proteins were renatured by dialysis against 50 mm Tris HCl (ph 8.0). The proteins were further purified in a Mono Q 5/50 GL column (Amersham Pharmacia Biotech). The strategies used for recombinant protein preparation are summarized in Table 4. The quantities of active enzyme in the purified recombinant MMPs were determined by titrating the samples with GM6001 (Chemicon) according to a previously described method (2). TIMP-2 active-site titration was performed using medaka recombinant gelatinase A, as described previously (3).
33 Enzyme Assays Enzyme samples were incubated in a volume of 100 μl with FITC-labeled bovine type IV collagen (USBiological) (25 μg) in buffer A (50 mm Tris HCl buffer (ph 7.5) containing 5 mm CaCl 2, 50 μm ZnSO 4, and 0.02% NaN 3 ) for 16 h at 27 C. The reactions were terminated by the addition of 300 μl of stopping reagent (42% EtOH, 50 mm Tris HCl (ph 9.5) and 0.2 M NaCl) and the samples were centrifuged. Aliquots of the resulting supernatants were diluted to a volume of 3.0 ml with distilled water, and the fluorescence was measured at an excitation wavelength of 459 nm and an emission wavelength of 520 nm. Enzyme activities in the presence of native medaka type I collagen were assessed using SDS-PAGE analysis (reducing conditions) of samples incubated with the enzymes in buffer A for 16 h at 27 C. To determine the activity of the FITC-labeled porcine type I gelatin (Molecular Probe), the enzyme samples were incubated at 27 C with the substrate (25 μg) in 100 μl of buffer A. After 16 h of incubation, the mixtures were filtered using a Viva Spin 0.5-ml Concentrator (molecular weight cutoff <10,000) (Viva Science). Aliquots of 50 μl of the filtrate were diluted with 3 ml of distilled water for fluorescence measurement. All incubations were performed at an enzyme:substrate ratio of 1/20 1/100. Gelatin zymography was performed as described previously (1). Isolation of Medaka Type I Collagen Type I collagen was isolated from the whole body of a medaka fish according to a previously described method (4). SDS-PAGE of the purified collagen under reducing conditions produced a similar pattern to those of bovine (USBiological) and rainbow trout type I collagens (5), indicating the validity of our isolation procedure.
34 Antibody Generation The protein antigens were produced using the bacterial expression system with pet30a. Recombinant proteins eluted from a Ni 2+ -Sepharose column were injected into rabbits or rats. IgGs were purified by affinity chromatography on a protein G-Sepharose column (Amersham Pharmacia Biotech). Specific antibodies were purified by affinity chromatography on a column of antigen coupled to CNBr-activated Sepharose 4B (Pharmacia) according to the manufacturer s protocol. Specific antibodies were also affinity purified using membranes onto which pure antigens were blotted. Immunohistochemistry Ovary sections (15 μm) were cut on a cryostat and thaw-mounted onto slides coated with silan. Sections on slides that were fixed with 4% paraformaldehyde in PBS or 100% methanol for at least 15 min were treated with 3% H 2 O 2 in PBS. After being blocked with BlockAce (Dainippon Seiyaku) for 1 h at room temperature, each section was incubated with purified primary antibodies for 1 h at room temperature, and was then washed with PBS. Bound antibodies were detected using anti-rabbit IgG (DakoCytomatin EnVision + System-labeled polymer-hrp anti-rabbit, Dako), or anti-rat IgG horseradish peroxidase-linked whole antibody (Pharmacia), according to the manufacturer s instructions. Immunocomplexes were detected using an AEC kit (Vector Laboratories). References 1. Matsui, H., Ogiwara, K., Ohkura, R., Yamashita, M. & Takahashi, T. (2000) Eur. J. Biochem. 267,
35 2. Park, H. I., Turk, B. E., Gerkema, F. E., Cantley, L. C. & Sang, Q. X. (2002) J. Biol. Chem. 277, Willenbrock, F., Crabbe, T., Slocombe, P. M., Sutton, C. W., Docherty, A. J., Cockett, M. I., O Shea, M., Brocklehurst, K., Phillips, I. R. & Murphy, G. (1993) Biochemistry 32, Bornstein, P. & Piez, K. A. (1964) J. Clin. Invest. 43, Saito, M., Sato, K., Kunisaki, N. & Kimura, S. (2000) Eur. J. Biochem. 267, Fig. 6. In situ detection of MMP mrnas in the medaka ovary. The distributions of seven MMP mrnas in the medaka ovary were determined by in situ-hybridization. Samples were stained with antisense (Left) and sense probes (Right). The areas or oocytes that showed positive staining are indicated by arrows. Scale bar = 500 μm. Fig. 7. Preparation of active recombinant MMPs. (A) Purified active MMPs were analyzed by SDS-PAGE under reducing conditions. (B) The MMPs were analyzed by gelatin zymography. (C) Progelatinase A incubated with MT1-MMP, MT2-MMP, MT3-MMP, and MT5-MMP was analyzed using gelatine zymography. The positions of progelatinase A (68 kda), the intermediate (55 kda), and the activated form (52 kda) are shown. Zymography of progelatinase A and gelatinase A was conducted without incubation in order to determine their mobilities under these conditions (Right Panel). Asterisks indicate nonspecifically proteolyzed bands. Fig. 8. Expression of MMP proteins in the medaka ovary. (A) Anti-MMP antibodies
36 were purified and tested for their specificities by Western blot analysis using a membrane loaded with seven recombinant MMP proteins. The S-protein was used to detect all recombinant MMPs. (B) Extracts of medaka ovaries taken 1 h after ovulation were analyzed by Western blot analysis using specific MMP antibodies. The sizes of the proteins specifically detected with the antibodies are shown. Fig. 9. Inmmunohistochemical analysis of MMPs in the medaka ovary. MMPs were immunohistochemically detected in the medaka ovary using purified primary antibodies. The controls were stained with primary antibodies previously treated with the respective antigens (for gelatinase B, MT2-MMP, MT5-MMP, and stromelysin-3), or with diluted preimmune sera (for gelatinase A and MT1-MMP). The areas or oocytes that showed positive staining are indicated by arrows. Scale bar = 500 μm. Fig. 10. Inhibition of recombinant MMP activity by specific antibodies. IgG fractions containing specific antibodies purified through a Protein G-Sepharose 4B column were included in the MMP activity-assay mixtures. The quantity of IgG antibody added amounted to five times (by weight) that of the purified enzymes. The substrate was FITC-labeled porcine type I gelatin. The values given are the average of triplicate assays. Fig. 11. Expression of TIMP-2b mrna in the medaka ovary. (A) RT-PCR for TIMP-2a, TIMP-2b, and TIMP-3 was performed using total RNAs isolated from various tissues of the medaka. Twenty-five PCR cycles were performed. (B) RT-PCR for TIMP-2a, TIMP-2b, and TIMP-3 was performed using total RNAs isolated from oocytes of
37 ovulating follicles at various periovulation time points. Twenty-five PCR cycles were performed. (C) In situ hybridization analysis of TIMP-2b mrna was conducted using the specific probes indicated in Table 3. Staining was performed with antisense (Left Panel) and sense probes (Right Panel). Specific signals are indicated by arrows. Scale bar = 200 μm. Fig. 12. Detection of TIMP-2b protein using ovulating follicles of the medaka. Follicles isolated at 6 h were incubated in the in vitro culture system. At 2, 0, 2, and 4 h of ovulation, the culture medium was replaced with fresh medium. Oocytes were collected at 2, 0, 2, and 4 h of ovulation. Oocyte extracts were immunoprecipitated with anti-medaka TIMP-2b antibodies, and the resulting precipitates were analyzed by Western blotting with anti-human TIMP-2 monoclonal antibody (Upper Panel). In vitro culture media were collected from four samples: those derived from a 3-h incubation between 5 and 2 h ( 5/ 2 in the Lower Panel), and the subsequent 2-h incubations ( 2/0, 0/2, and 2/4 in the Lower Panel). Immunoprecipitation and Western blot analyses of the culture media were conducted as described above. Medaka TIMP-2b protein (28 kda) is shown.
38 Table 1. Degenerate Oligonucleotide Primers Used for Initial PCR Amplification of cdna Fragments Gene Degenerate primer sequence MMPs Sense 5 -CCCAGATG(C/T)GG(A/G)GT(T/G)CCTGA(C/T)-3 Antisense 5 -TGGCC(A/C/G)A(A/G)TTCATGAGC(A/C/T)GC(A/C)AC-3 TIMPs Sense 5 -CA(C/T)CCICA(A/G)(C/G)AIGCITT(C/T)TG(C/T)AA-3 Antisense 5 -C(A/C)(A/T)(A/G)TCC(A/G)TCCAIA(A/G/T)(A/G)CA(C/T)TC-3
39 Table 2. Cloning Strategies for Target Genes Gene Regions used for Cloning method GenBank degenerate PCR accession number primer design MT1-MMP(MMP-14) 635-1,098 5 /3 -RACE AB MT2-MMP (MMP-15) 635-1,089 Screening AB MT3-MMP (MMP-16) Screening AB Gelatinase A (MMP-2) 398-1,318 Screening, 5 -RACE AB Gelatinase B (MMP-9) 439-1,369 Screening AB Stromelysin-3 (MMP-11) Screening, 5 /3 -RACE AB MT5-MMP (MMP-24) Screening, 5 /3 -RACE AB TIMP-2a RACE AB TIMP-2b /3 -RACE AB TIMP /3 -RACE AB193469
40 Table 3. PCR Primers Used for the Amplification of Specific DNA Fragments Gene Amplified regions Probes for in situ GenBank in RT-PCR hybridization accession number analysis MT1-MMP(MMP-14) 1,827 2,207 2,803 3,211 AB MT2-MMP (MMP-15) 2,036 2,590 1,983 2,409 AB MT3-MMP (MMP-16) ,695 2,160 AB Gelatinase A (MMP-2) 2,374 2,683 2,374 2,683 AB Gelatinase B (MMP-9) 2,191 1,482 2,191 2,760 AB Stromelysin-3 (MMP-11) 969 1, ,482 AB MT5-MMP (MMP-24) 1,228 1,966 1,228 1,966 AB TIMP-2a AB TIMP-2b AB TIMP AB OLCA-1 1,529 1,840 D89627
41 Table 4. Strategy for Recombinant Protein Synthesis Gene Expressed regions Corresponding amino acids Restriction sites and GenBank (nucleotide numbers) (residue numbers) expression vectors accession number MT1-MMP 704 2, (active form*) EcoRI-HindIII/pET30a AB MT2-MMP 698 2, (active form*) EcoRI-HindIII/pET30a AB MT3-MMP 572 1, (active form*) EcoRI-HindIII/pET30a AB Gelatinase A 178 2, (proenzyme form) EcoRV-HindIII/pET30a AB Gelatinase A 427 2, (active form) EcoRV-HindIII/pET30a AB Gelatinase B 469 2, (active form) EcoRI-XhoI/pET30a AB Stromelysin , (active form) EcoRI-HindIII/pET30a AB MT5-MMP 407 1, (active form*) EcoRI-HindIII/pET30a AB TIMP-2b (active form) NdeI-XhoI/pET30a AB *Expressed without the transmembrane and cytoplasmic domains.
42

43
44

45
46

47
Ovulation, which is triggered by a preovulatory surge of luteinizing
 Gelatinase A and membrane-type matrix metalloproteinases 1 and 2 are responsible for follicle rupture during ovulation in the medaka Katsueki Ogiwara*, Naoharu Takano*, Masakazu Shinohara*, Masahiro Murakami,
Gelatinase A and membrane-type matrix metalloproteinases 1 and 2 are responsible for follicle rupture during ovulation in the medaka Katsueki Ogiwara*, Naoharu Takano*, Masakazu Shinohara*, Masahiro Murakami,
Characterization of Luteinizing Hormone and Luteinizing Hormone Receptor and Their Indispensable Role in the Ovulatory Process of the Medaka
 Characterization of Luteinizing Hormone and Luteinizing Hormone Receptor and Their Indispensable Role in the Ovulatory Process of the Medaka Katsueki Ogiwara 1, Chika Fujimori 1, Sanath Rajapakse 2, Takayuki
Characterization of Luteinizing Hormone and Luteinizing Hormone Receptor and Their Indispensable Role in the Ovulatory Process of the Medaka Katsueki Ogiwara 1, Chika Fujimori 1, Sanath Rajapakse 2, Takayuki
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Collagenase Assay Kit
 Collagenase Assay Kit Catalog # 31 and 32 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Collagenases are members of the matrix metalloproteinase (MMP) family and degrade collagen types
Collagenase Assay Kit Catalog # 31 and 32 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Collagenases are members of the matrix metalloproteinase (MMP) family and degrade collagen types
Collagenase Assay Kit
 Collagenase Assay Kit Catalog # 31 and 32 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The collagenases are members of the matrix metalloproteinase (MMP) family and degrade collagen
Collagenase Assay Kit Catalog # 31 and 32 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The collagenases are members of the matrix metalloproteinase (MMP) family and degrade collagen
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
SUPPLEMENTARY MATERIAL. Sample preparation for light microscopy
 SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
ABSTRACT. Key words: ovulation, ovary, human, follicle, collagen, MMP and TIMP. ISBN-10: ISBN-13:
 HUMAN OVULATION Studies on collagens, gelatinases and tissue inhibitors of metalloproteinases Anna Karin Lind Department of Obstetrics and Gynecology Institute of Clinical Sciences Sahlgrenska University
HUMAN OVULATION Studies on collagens, gelatinases and tissue inhibitors of metalloproteinases Anna Karin Lind Department of Obstetrics and Gynecology Institute of Clinical Sciences Sahlgrenska University
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
OVARY The surface of the ovary is covered with surface epithelium
 OVARY Cow The ovary, or female gonad, is: 1. an exocrine gland, producing oocytes 2. an endocrine gland, secreting hormones, i.e., estrogen and progesterone OVARY OVARY The surface of the ovary is covered
OVARY Cow The ovary, or female gonad, is: 1. an exocrine gland, producing oocytes 2. an endocrine gland, secreting hormones, i.e., estrogen and progesterone OVARY OVARY The surface of the ovary is covered
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved
 1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
PRODUCT INFORMATION & MANUAL
 PRODUCT INFORMATION & MANUAL 0.4 micron for Overall Exosome Isolation (Cell Media) NBP2-49826 For research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 303.730.1950 - P:
PRODUCT INFORMATION & MANUAL 0.4 micron for Overall Exosome Isolation (Cell Media) NBP2-49826 For research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 303.730.1950 - P:
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Title. Author(s)Ogiwara, Katsueki; Takahashi, Takayuki. CitationMolecular and cellular endocrinology, 450: Issue Date
Ogiwara, Katsueki; Takahashi, Takayuki. CitationMolecular and cellular endocrinology, 450: Issue Date") Title Involvement of the nuclear progestin receptor in LHrequired for follicle rupture during ovulation in th Author(s)Ogiwara, Katsueki; Takahashi, Takayuki CitationMolecular and cellular endocrinology,
Title Involvement of the nuclear progestin receptor in LHrequired for follicle rupture during ovulation in th Author(s)Ogiwara, Katsueki; Takahashi, Takayuki CitationMolecular and cellular endocrinology,
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
The Distribution of Collagenous Connective Tissue in Rat Ovarian Follicles
 BIOLOGY OF REPRODUCTION 14, 502-506 (1976). The Distribution of Collagenous Connective Tissue in Rat Ovarian Follicles LAWRENCE L ESPEY Reproductive Physiology Laboratory, Trinity University, San Antonio,
BIOLOGY OF REPRODUCTION 14, 502-506 (1976). The Distribution of Collagenous Connective Tissue in Rat Ovarian Follicles LAWRENCE L ESPEY Reproductive Physiology Laboratory, Trinity University, San Antonio,
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Procaspase-3. Cleaved caspase-3. actin. Cytochrome C (10 M) Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk
 Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk") A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
Green Cathepsin B Kit. For Research Use Only
 Green Cathepsin B Kit For Research Use Only ICT9151-25 Tests Component Storage Conditions Quantity 1 vial Rhodamine 110-(RR) 2-20 o C 25 tests 1 vial Hoechst 33342 (200 µg) 2-8 o C 1 ml 1 bottle 10x Cellular
Green Cathepsin B Kit For Research Use Only ICT9151-25 Tests Component Storage Conditions Quantity 1 vial Rhodamine 110-(RR) 2-20 o C 25 tests 1 vial Hoechst 33342 (200 µg) 2-8 o C 1 ml 1 bottle 10x Cellular
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Effects of Catecholamines and Dibenamine on Ovulation in the Perfused Fowl Ovary
 Effects of Catecholamines and Dibenamine on Ovulation in the Perfused Fowl Ovary Tomoki HIGUCHI, Tomoki SOH, Frank HERTELENDY* and Kousaku TANAKA Faculty of Agriculture, Kyushu University, Higashi-ku,
Effects of Catecholamines and Dibenamine on Ovulation in the Perfused Fowl Ovary Tomoki HIGUCHI, Tomoki SOH, Frank HERTELENDY* and Kousaku TANAKA Faculty of Agriculture, Kyushu University, Higashi-ku,
Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard. Product Number: AD0014
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3
 Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS
: DEVELOPMENT OF ANTIVESICANT DRUGS") STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS Xiannu Jin 1, Radharaman Ray 2, Guang Xu 1 and Prabhati Ray 1 1 Department
STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS Xiannu Jin 1, Radharaman Ray 2, Guang Xu 1 and Prabhati Ray 1 1 Department
Exosomes Immunocapture and Isolation Tools: Immunobeads
 Product Insert Exosomes Immunocapture and Isolation Tools: Immunobeads HBM-BOLF-##/##-# HBM-BOLC-##/##-# HBM-BTLF-##/##-# Overall Exosome capture from Human Biological fluids Overall Exosome capture from
Product Insert Exosomes Immunocapture and Isolation Tools: Immunobeads HBM-BOLF-##/##-# HBM-BOLC-##/##-# HBM-BTLF-##/##-# Overall Exosome capture from Human Biological fluids Overall Exosome capture from
Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit
 Protein Quantity Microplate Assay Kit") PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab)
 Protocol (Mirmira Lab)") Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade
 AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Supplemental Information
 Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Note: During 30 minute incubation; proceed thru appropriate sections below (e.g. sections II, III and V).
.") LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature
 PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard. Product Number: AD0013
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
RNA/DNA Stabilization Reagent for Blood/Bone Marrow
 For general laboratory use. Not for use in diagnostic procedures. FOR IN VITRO USE ONLY. RNA/DNA Stabilization Reagent for Blood/Bone Marrow For simultaneous cell lysis and stabilization of nucleic acids
For general laboratory use. Not for use in diagnostic procedures. FOR IN VITRO USE ONLY. RNA/DNA Stabilization Reagent for Blood/Bone Marrow For simultaneous cell lysis and stabilization of nucleic acids
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
For the rapid, sensitive and accurate quantification of Ras in various samples
 ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
Mouse C-peptide EIA. Cat. No. YII-YK013-EX FOR LABORATORY USE ONLY
 Mouse C-peptide EIA Cat. No. YII-YK013-EX FOR LABORATORY USE ONLY TOYO 2CHOME, KOTO-KU, TOKYO, 135-0016, JAPAN http://www.cosmobio.co.jp e-mail : export@cosmobio.co.jp Phone : +81-3-5632-9617 FAX : +81-3-5632-9618
Mouse C-peptide EIA Cat. No. YII-YK013-EX FOR LABORATORY USE ONLY TOYO 2CHOME, KOTO-KU, TOKYO, 135-0016, JAPAN http://www.cosmobio.co.jp e-mail : export@cosmobio.co.jp Phone : +81-3-5632-9617 FAX : +81-3-5632-9618
Tivadar Orban, Beata Jastrzebska, Sayan Gupta, Benlian Wang, Masaru Miyagi, Mark R. Chance, and Krzysztof Palczewski
 Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
A ph-dependent Charge Reversal Peptide for Cancer Targeting
 Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
Mouse Cathepsin B ELISA Kit
 GenWay Biotech, Inc. 6777 Nancy Ridge Drive San Diego, CA 92121 Phone: 858.458.0866 Fax: 858.458.0833 Email: techline@genwaybio.com http://www.genwaybio.com Mouse Cathepsin B ELISA Kit Catalog No. GWB-ZZD154
GenWay Biotech, Inc. 6777 Nancy Ridge Drive San Diego, CA 92121 Phone: 858.458.0866 Fax: 858.458.0833 Email: techline@genwaybio.com http://www.genwaybio.com Mouse Cathepsin B ELISA Kit Catalog No. GWB-ZZD154
LH (Bovine) ELISA Kit
 ELISA Kit") LH (Bovine) ELISA Kit Catalog Number KA2280 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
LH (Bovine) ELISA Kit Catalog Number KA2280 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression. through inhibition of Hsp90
 Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
Reduction in Serum Lecithin:Cholesterol Acyltransferase Activity Prior to the Occurrence of Ketosis and Milk Fever in Cows
 FULL PAPER Biochemistry Reduction in Serum Lecithin:Cholesterol Acyltransferase Activity Prior to the Occurrence of Ketosis and Milk Fever in Cows Hisami NAKAGAWA-UETA 1) and Norio KATOH 2) * 1) Ishikawa
FULL PAPER Biochemistry Reduction in Serum Lecithin:Cholesterol Acyltransferase Activity Prior to the Occurrence of Ketosis and Milk Fever in Cows Hisami NAKAGAWA-UETA 1) and Norio KATOH 2) * 1) Ishikawa
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Adjust to ph=7 with some HCl if too basic, or NaOH if too acid (but it will be quite close to neutrality even without adding any of these)
") BIG VOLUME SOLUTIONS Solutions should be prepared with distilled and autoclaved water unless otherwise indicated. If the solutions have to be left unused for long periods (> 1 week), it is recommended
BIG VOLUME SOLUTIONS Solutions should be prepared with distilled and autoclaved water unless otherwise indicated. If the solutions have to be left unused for long periods (> 1 week), it is recommended
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Supporting Information for:
 Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
Endocrinology laboratory Department of Zoology Kalyani University Kalyani, West Bengal India
 Epidermal growth factor (EGF) promotes ovarian steroidogenesis and epidermal growth factor receptor (EGFR) signaling is required for gonadotropin-induced steroid production in common carp Cyprinus carpio
Epidermal growth factor (EGF) promotes ovarian steroidogenesis and epidermal growth factor receptor (EGFR) signaling is required for gonadotropin-induced steroid production in common carp Cyprinus carpio
Influenza B Hemagglutinin / HA ELISA Pair Set
 Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Supplementary Figure 1. mrna targets were found in exosomes and absent in free-floating supernatant. Serum exosomes and exosome-free supernatant were
 Supplementary Figure 1. mrna targets were found in exosomes and absent in free-floating supernatant. Serum exosomes and exosome-free supernatant were separated via ultracentrifugation and lysed to analyze
Supplementary Figure 1. mrna targets were found in exosomes and absent in free-floating supernatant. Serum exosomes and exosome-free supernatant were separated via ultracentrifugation and lysed to analyze
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
ab SREBP-2 Translocation Assay Kit (Cell-Based)
") ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
Matrix metalloproteinases
 Matrix metalloproteinases,, 1. 2. 2-1. Domain 2-2. TIMPs 2-3. MMPs, 3. 4. 1. Collagens, proteoglycans (Extracellular matrix, ECM),,. matrixins matrix metalloproteinases (MMPs) [1, 2]. MMPs,, cytokines
Matrix metalloproteinases,, 1. 2. 2-1. Domain 2-2. TIMPs 2-3. MMPs, 3. 4. 1. Collagens, proteoglycans (Extracellular matrix, ECM),,. matrixins matrix metalloproteinases (MMPs) [1, 2]. MMPs,, cytokines
Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/-
 Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
About the Kits...2 Description 2 Components 3 Storage 3. Factors That Influence Factor Xa Activity... 4
 Novagen User Protocol TB205 Rev. C 0107 1 of 9 Factor Xa Kits Table of Contents About the Kits...2 Description 2 Components 3 Storage 3 Factors That Influence Factor Xa Activity... 4 Factor Xa Cleavage...5
Novagen User Protocol TB205 Rev. C 0107 1 of 9 Factor Xa Kits Table of Contents About the Kits...2 Description 2 Components 3 Storage 3 Factors That Influence Factor Xa Activity... 4 Factor Xa Cleavage...5
Effect of Bovine Follicular Fluid Added to the Maturation Medium on Sperm Penetration in Pig Oocytes Matured In Vitro
 Article Effect of Bovine Follicular Fluid Added to the Maturation Medium on Sperm Penetration in Pig Oocytes Matured In Vitro Abstract Naoki ISOBE Research Associate Graduate School for International Development
Article Effect of Bovine Follicular Fluid Added to the Maturation Medium on Sperm Penetration in Pig Oocytes Matured In Vitro Abstract Naoki ISOBE Research Associate Graduate School for International Development
Rat Leptin-HS ELISA FOR LABORATORY USE ONLY YANAIHARA INSTITUTE INC AWAKURA, FUJINOMIYA - SHI SHIZUOKA, JAPAN
 YK051 Rat Leptin-HS ELISA FOR LABORATORY USE ONLY YANAIHARA INSTITUTE INC. 2480-1 AWAKURA, FUJINOMIYA - SHI SHIZUOKA, JAPAN 418 0011 Contents Introduction 2 Characteristics 3 Composition 4 Method 5-6 Notes
YK051 Rat Leptin-HS ELISA FOR LABORATORY USE ONLY YANAIHARA INSTITUTE INC. 2480-1 AWAKURA, FUJINOMIYA - SHI SHIZUOKA, JAPAN 418 0011 Contents Introduction 2 Characteristics 3 Composition 4 Method 5-6 Notes
FBA Translation (New Series) No. 136
 No. 136") Freshwater Biological Association FBA Translation (New Series) No. 136 Title. Modulation of the effectiveness of 17 α-hydroxy-20 β-dihydroprogesterone or of a gonadotrophic extract on the in vitro intrafollicular
Freshwater Biological Association FBA Translation (New Series) No. 136 Title. Modulation of the effectiveness of 17 α-hydroxy-20 β-dihydroprogesterone or of a gonadotrophic extract on the in vitro intrafollicular
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPORTING MATREALS. Methods and Materials
 SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
20S Proteasome Activity Assay Kit
 20S Proteasome Activity Assay Kit For 100 Assays Cat. No. APT280 FOR RESEARCH USE ONLY NOT FOR USE IN DIAGNOSTIC PROCEDURES USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23
20S Proteasome Activity Assay Kit For 100 Assays Cat. No. APT280 FOR RESEARCH USE ONLY NOT FOR USE IN DIAGNOSTIC PROCEDURES USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23
Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine
 Agric. Biol. Chem., 46 (6), 1565~1569, 1982 1565 Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine Minoru Noda, Thanh Vo Van, Isao Kusakabe
Agric. Biol. Chem., 46 (6), 1565~1569, 1982 1565 Substrate Specificity and Salt Inhibition of Five Proteinases Isolated from the Pyloric Caeca and Stomach of Sardine Minoru Noda, Thanh Vo Van, Isao Kusakabe
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
SensoLyte Generic MMP Assay Kit *Colorimetric*
 SensoLyte Generic MMP Assay Kit *Colorimetric* Revision#1.2 Catalog # Kit Size Last updated: May2017 AS-72095 100 Assays (96-well plate) Optimized Performance: This kit is optimized to detect MMP activity
SensoLyte Generic MMP Assay Kit *Colorimetric* Revision#1.2 Catalog # Kit Size Last updated: May2017 AS-72095 100 Assays (96-well plate) Optimized Performance: This kit is optimized to detect MMP activity
REPRODUCTIVE CYCLE OF FEMALE MAMMAL
 REPRODUCTIVE CYCLE OF FEMALE MAMMAL Fig. 8-12 Secondary follicles growing follicles increase in number of layers of granulosa cells Tertiary follicles maturing follicles antrum formation fluid filled space
REPRODUCTIVE CYCLE OF FEMALE MAMMAL Fig. 8-12 Secondary follicles growing follicles increase in number of layers of granulosa cells Tertiary follicles maturing follicles antrum formation fluid filled space
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
PURIFICATION AND ACTION SITES OF A FOLLICLE STIMULATING HORMONE INHIBITOR FROM BOVINE FOLLICULAR FLUID t
 PURIFICATION AND ACTION SITES OF A FOLLICLE STIMULATING HORMONE INHIBITOR FROM BOVINE FOLLICULAR FLUID t E. Sato, T. Ishibashi and A. Iritani Kyoto university 2, Kyoto 606, Japan Summary The purification
PURIFICATION AND ACTION SITES OF A FOLLICLE STIMULATING HORMONE INHIBITOR FROM BOVINE FOLLICULAR FLUID t E. Sato, T. Ishibashi and A. Iritani Kyoto university 2, Kyoto 606, Japan Summary The purification
Problem-solving Test: The Mechanism of Protein Synthesis
 Q 2009 by The International Union of Biochemistry and Molecular Biology BIOCHEMISTRY AND MOLECULAR BIOLOGY EDUCATION Vol. 37, No. 1, pp. 58 62, 2009 Problem-based Learning Problem-solving Test: The Mechanism
Q 2009 by The International Union of Biochemistry and Molecular Biology BIOCHEMISTRY AND MOLECULAR BIOLOGY EDUCATION Vol. 37, No. 1, pp. 58 62, 2009 Problem-based Learning Problem-solving Test: The Mechanism
Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained
 Representative pictures of H&E-stained") 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
Plasma Membrane Protein Extraction Kit
 ab65400 Plasma Membrane Protein Extraction Kit Instructions for Use For the rapid and sensitive extraction and purification of Plasma Membrane proteins from cultured cells and tissue samples. This product
ab65400 Plasma Membrane Protein Extraction Kit Instructions for Use For the rapid and sensitive extraction and purification of Plasma Membrane proteins from cultured cells and tissue samples. This product
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Rabbit Polyclonal antibody to NFkB p65 (v-rel reticuloendotheliosis viral oncogene homolog A (avian))
)") Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Human Cathepsin D ELISA Kit
 GenWay Biotech, Inc. 6777 Nancy Ridge Drive San Diego, CA 92121 Phone: 858.458.0866 Fax: 858.458.0833 Email: techline@genwaybio.com http://www.genwaybio.com Human Cathepsin D ELISA Kit Catalog No. GWB-J4JVV9
GenWay Biotech, Inc. 6777 Nancy Ridge Drive San Diego, CA 92121 Phone: 858.458.0866 Fax: 858.458.0833 Email: techline@genwaybio.com http://www.genwaybio.com Human Cathepsin D ELISA Kit Catalog No. GWB-J4JVV9