CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS
|
|
|
- Eugene Atkinson
- 5 years ago
- Views:
Transcription

1 University of Kentucky UKnowledge University of Kentucky Doctoral Dissertations Graduate School 2007 CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS Darshini Trivedi University of Kentucky, Click here to let us know how access to this document benefits you. Recommended Citation Trivedi, Darshini, "CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS" (2007). University of Kentucky Doctoral Dissertations This Dissertation is brought to you for free and open access by the Graduate School at UKnowledge. It has been accepted for inclusion in University of Kentucky Doctoral Dissertations by an authorized administrator of UKnowledge. For more information, please contact
2 ABSTRACT OF DISSERTATION Darshini Trivedi The Graduate School University of Kentucky 2007
3 CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS ABSTRACT OF DISSERTATION A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the College of Pharmacy at the University of Kentucky By Darshini Trivedi Lexington, Kentucky Director: Dr. Charles D. Loftin, Assistant Professor of Pharmaceutical Sciences Lexington, Kentucky 2007 Copyright Darshini Trivedi 2007
4 ABSTRACT OF DISSERTATION CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS Transition of the cardiopulmonary circulation at birth requires functional closure of the ductus arteriosus (DA). The DA is an arterial shunt that is vital in the fetus for diverting the pulmonary circulation away from the uninflated lungs. Failure of the vessel to functionally close after birth is known as patent DA, which is the second most common congenital heart disease. Patent DA may seriously compromise neonatal health and current pharmacological treatments are often limited by serious complications or a significant failure rate, thereby increasing the necessity for surgical intervention. Recently, we were the first to show that genetic or pharmacological inactivation of cyclooxygenase (COX) -2 produces postnatal patent DA in mice. We also demonstrated that the DA expresses high levels of COX-2 during normal closure after birth, suggesting novel contractile actions of COX-2-dependent prostanoids in the DA. In humans, patent DA is more common in preterm infants than those born at full-term, however, mechanism(s) responsible for the reduced DA closure have not been identified. In the current studies, we examined COX-1 and COX-2 expression in the DA at multiple stages of gestation to determine whether alterations in the expression of these enzymes contribute to patent DA in preterm mice. Using real-time PCR, analysis of the time-course of COX-2 mrna in the fetal mouse DA indicated that COX-2 expression significantly increased with advancing gestational age. The preterm (day 17.5) neonatal mouse DA showed attenuated COX-2 expression, as compared to the full-term (day 19.5) neonatal DA at 3 hours after birth. Furthermore, the DA of preterm neonatal mice showed incomplete closure after 3 hours of birth, a time-point when the DA of full-term neonates was completely remodeled. These data indicate a correlation between reduced DA closure and attenuated COX-2 expression. Additionally, COX-2 expression was significantly attenuated in the DA of mice deficient in the prostanoid receptor EP4, which also show a patent DA phenotype, suggesting the importance of this receptor for the induction of COX-2 required for DA closure. Overall, these studies suggest that attenuated expression of COX-2 may contribute to increased patent DA at preterm gestation.
5 KEYWORDS: cyclooxygenase-2, ductus arteriosus, prostanoid receptors, congenital heart defects, vascular remodeling Darshini Trivedi July 30, 2007
6 CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS By Darshini Trivedi Director of Dissertation Dr. Charles D. Loftin Director of Graduate Studies Dr. Janice Buss June 5, 2007
7 RULES FOR THE USE OF DISSERTATIONS Unpublished dissertations submitted for the Doctor s degree and deposited in the University of Kentucky Library are as a rule open for inspection, but are to be used only with due regard to the rights of the authors. Bibliographical references may be noted, but quotations or summaries of parts may be published only with the permission of the author, and with the usual scholarly acknowledgements. Extensive copying or publication of the dissertation in whole or in part also requires the consent of the Dean of the Graduate School of the University of Kentucky. A library that borrows this dissertation for use by its patrons is expected to secure the signature of each user. Name Date
8 DISSERTATION Darshini Trivedi The Graduate School University of Kentucky 2007
9 CYCLOOXYGENASE-2-DEPENDENT REMODELING OF THE DUCTUS ARTERIOSUS DISSERTATION A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the College of Pharmacy at the University of Kentucky By Darshini Trivedi Lexington, Kentucky Director: Dr. Charles D. Loftin, Assistant Professor of Pharmaceutical Sciences Lexington, Kentucky 2007 Copyright Darshini Trivedi 2007
10 ACKNOWLEDGMENTS My career in science has been a continuous journey, beginning from the year 1998, when I accepted a student internship at the National Institute of Environmental Health Sciences, NC. The past 9 years have provided me with an extremely educational and fulfilling experience and I would like to acknowledge the numerous people who have made this possible. First and foremost, I express my gratitude towards my long time mentor, Dr. Charles Loftin who has primarily contributed to my development as a scientist. Chuck has been a constant support throughout my scientific career for the past 9 years. He has taught me to be patient, focused and persevering, which has allowed me to get through the inherent frustrations and disappointments that accompany scientific research. He has continuously guided me during this research with his knowledge and expertise, and without his mentorship, this dissertation would have been incomplete. I thank my committee members, Dr. Todd Porter, Dr. Daniel Tai, Dr. Alan Daugherty and Dr. Lisa Cassis, who have guided me throughout my research, and have contributed towards the development of my scientific skills through their powerful inquiries during our meetings. To my fellow colleagues, Lakshmi Vetcha, Carrie Schafer and Jonathan Gitlin, your friendship and support in the lab was always appreciated. I thank all my friends in Lexington, who have made the last 5 years of my life enjoyable. I thank you for your continuous support and good wishes. I thank my partner in life, Brett, who has been patient and unrelenting, and has provided me with all the love and support that I needed to get through my graduate work and dissertation. Last but not the least, to my family, I am deeply grateful for your care, love and encouragement, without which none of this would have been possible. iii
11 TABLE OF CONTENTS Acknowledgements iii List of Tables.vii List of Figures...viii Chapter 1: BACKGROUND THE DUCTUS ARTERIOSUS (DA).. 1 Developmental Origin of the DA...1 Anatomic Closure of the DA.2 Patent DA...3 Current Treatments for Patent DA.4 PROSTAGLANDINS AND CYCLOOXYGENASES...7 COX-1 8 COX-2 8 Inhibition of COX Activity 9 COX-1 and COX-2-Deficient Mice..12 FACTORS REGULATING DA CLOSURE hydroxyprostaglandin dehydrogenase...13 Cyclooxygenases 15 Other Factors Regulating DA Closure...17 PROSTANOID RECEPTORS.18 Dilatory Prostanoid Receptors and the DA 20 Contractile Prostanoid Receptors and the DA...22 METHODS Animals 24 Tissue Collection.24 RNA Isolation..24 cdna Preparation 25 Quantitation of mrna Expression..25 Histology..26 Statistics...26 Chapter 2: CHARACTERIZATION OF THE CELL TYPES IN THE MOUSE DA Introduction..28 iv
12 Results...30 CHAPTER 3: ANALYSIS OF INDOMETHACIN-INDUCED CONSTRICTION OF THE DA IN UTERO Introduction..36 Results..37 Discussion 40 CHAPTER 4: ANALYSIS OF COX-1 AND COX-2 EXPRESSION IN THE FETAL AND NEONATAL MOUSE DA Introduction..42 Results..44 Expression of COX-2 mrna in the fetal DA at multiple stages of gestation. Expression of COX-2 mrna in the fetal DA at multiple stages of gestation Analysis of COX-1 and COX-2 expression in the DA of neonatal mice at fullterm gestation Discussion 52 CHAPTER 5: ANALYSIS OF POSTNATAL DA CLOSURE AND COX EXPRESSION AT PRETERM AND FULL-TERM GESTATION IN MICE Introduction..54 Results..56 Morphological analysis of DA closure at 0 and 3 hours after birth at full-term gestation Histological analysis of DA closure at 0 and 3 hours after birth at full-term gestation Histological analysis of DA closure at 3 hours after birth at preterm gestation Postnatal expression of COX-1 and COX-2 in the DAs of preterm neonatal mice. Discussion 64 CHAPTER 6: PROSTANOID RECEPTORS IN THE MOUSE DA Introduction..67 Results..69 Quantitation of prostanoid receptors in the fetal DA at multiple stages of gestation. v
13 Analysis of COX-2 mrna expression in the DA of EP4-deficient neonatal mice. Discussion 73 CHAPTER 7: GENERAL DISCUSSION Summary and Conclusions.78 Clinical Relevance and Future Directions..79 REFERENCES.85 VITA 102 vi
14 LIST OF TABLES Table 1: Incidence of patent DA with the genetic deficiency or pharmacological inactivation of COX-1 and/or COX-2 in mice 16 Table 2: Prostanoid receptors, signaling and vascular function.19 vii
15 LIST OF FIGURES Figure 1.1: Illustration of the fetal and neonatal cardiopulmonary circulation 5-6 Figure 1.2: Illustration of the cyclooxygenase pathway 11 Figure 1.3: PGE 2 maintains dilation of the DA in utero 14 Figure 2.1: Basement membrane staining.31 Figure 2.2: Smooth muscle cell staining...32 Figure 2.3: Endothelial cell staining.33 Figure 2.4: Elastin staining...34 Figure 2.5: Desmin staining...35 Figure 3: Histological analysis of indomethacin-induced DA constriction...39 Figure 4.1: Expression of COX-2 mrna in the fetal DA at multiple stages of gestation...45 Figure 4.2: Expression of COX-1 mrna in the fetal DA at multiple stages of gestation...47 Figure 4.3: COX-1 and COX-2 expression in the DA of full-term neonatal mice..50 Figure 5.1: Morphological analysis of DA closure...57 Figure 5.2: Histological analysis of DA closure Figure 5.3: COX-1 and COX-2 mrna expression in the DA of preterm neonatal mice 62 Figure 5.4: Immunohistochemical analysis of COX-2 expression...63 Figure 5.5. Proposed model of normal DA closure at full-term gestation and DA patency at preterm gestation.66 Figure 6.1: Expression of prostanoid receptor mrna in the DA at multiple stages of gestation..70 Figure 6.2: Expression of COX-2 mrna in the DA at EP4-deficient mice 72 Figure 6.3: Proposed model for the induction of COX-2 expression following EP4 activation 77 Figure 7: COX-1 inhibitor treatment for patent DA at preterm gestation...81 viii
16 CHAPTER 1 BACKGROUND THE DUCTUS ARTERIOSUS The ductus arteriosus (DA) is a muscular artery in the fetus which connects the main pulmonary artery and the descending aorta and functions in utero to divert greater than 90% of the right ventricular cardiac output to bypass the uninflated lungs. 1 Loss of the maternal oxygen supply at birth necessitates rapid initiation of pulmonary function and transition of the cardiopulmonary circulation in the newborn. An essential component of this postnatal circulatory remodeling is the functional closure of the DA. In the healthy full-term infant, DA constriction begins after birth to initiate functional closure during the first few days of life. 2 Developmental Origin of the DA During embryonic development, the DA originates together with other great arteries, from the pharyngeal arterial system. Even though the cells forming the heart and circulatory system are of mesodermal origin, the arches of the pharyngeal arterial system are populated with cardiac neural crest cells, which also contribute to formation of the vessel wall. Studies in the chick and mammalian embryos have shown that cardiac neural crest derived cells contribute to a variety of cells in the arches, including the arterial smooth muscle cells (SMCs). 3 Even though anatomical diversity exists in the aortic arch system in all vertebrates, the ancestral arterial system from which it is derived consists of 6 aortic arches. It is known that lung-bearing vertebrates including lungfish, amphibians, reptiles, birds, marsupials and mammals, all possess a DA. Despite the diversity in the remodeling of the arterial system, the DA in all air-breathing vertebrates develops from a specific segment of the sixth arch of the pharyngeal arterial system. Throughout development, the DA undergoes a unique differentiation program, which prepares it for closure in the newborn at the stage involving lung respiration. This in utero differentiation of the DA occurs during the final one-third of gestation with full contractile ability occurring only at full-term. Particularly, in all mammals, when the transition to pulmonary respiration occurs, the DA rapidly closes after birth. 4 1
17 Anatomic Closure of the DA. DA closure after birth occurs in two phases. The first phase involves constriction of smooth muscle, which obstructs the DA lumen. The second phase involves anatomical remodeling of the vessel, which results in permanent closure. In humans, permanent closure of the DA is complete within a few days of birth, whereas in mice, complete closure is achieved within 3 hours of birth. Permanent remodeling of the DA involves extensive intimal thickening and intimal cushion formation, which begins with the lifting, infolding and in-growth of the endothelial cells, followed by migration of SMC from the media, into the subendothelial space. 2, 4 Increased production of specific extracellular matrix components such as hyaluronic acid, chondroitin sulfate and fibronectin is also associated with intimal cushion formation. DA remodeling also involves disassembly of the internal elastic lamina and loss of elastin to promote SMC migration. 5 Release of extracellular matrix is also an important component of DA remodeling. The DA smooth muscle is involved in complex interactions with molecules of the extracellular matrix such as laminin, fibronectin, growth factors (e.g. TGF-!), endothelial hyaluronic acid and chondrotin sulfate. 2 Studies by Mason et al. demonstrated that inhibition of fibronectin production inhibited DA remodeling, emphasizing the importance of fibronectin in the intimal cushion formation during DA remodeling. 6 The interaction of the DA smooth muscle to the components of the extracellular matrix is facilitated by the action of specific cell surface receptors. The integrin family of receptors is thought to play an important role in the remodeling of the endothelial and SMCs of the DA. Specifically, several studies in vitro have suggested a role for several members of the integrin family of receptors including, "v!1 and "v!3, in the attachment and migration of SMCs to the extracellular matrix, to produce obliteration of the DA lumen. 7 DA remodeling is also thought to require a significant degree of hypoxia of DA smooth muscle. Studies by Clyman et al. have demonstrated a significant contribution of vascular endothelial growth factor (VEGF) in producing neointimal expansion during hypoxia-induced remodeling of the DA. 8 It was suggested that VEGF-induced cell migration in the DA may be mediated by the "v!3 integrin which is upregulated during DA closure. 7, 9 In addition to the migration of SMCs, inflammatory cell infiltration into 2
18 the vessel may also contribute DA remodeling during closure. A recent study by Waleh et al. proposed that the remodeling of the DA involves processes similar to those observed during pathological remodeling in diseases such as atherosclerosis. 10 These studies showed that postnatal DA remodeling involves an inflammatory response in which monocytes/macrophages and to some extent T-lymphocytes, are recruited in the DA lumen. Following remodeling of DA SMC, significant apoptosis and cytolytic necrosis are thought to be required for complete obliteration of the DA. 11, 12 The inner intimal region of the DA has been shown to significantly express markers of apoptosis during anatomical remodeling of the DA. The SMCs in this region of the DA which are originally highly differentiated during initial DA constriction, then become dedifferentiated during complete DA closure and undergo cytolytic necrosis during the final stage of DA development. 12 The SMCs in the inner and outer media have different apoptotic properties. The outer media has more prominent apoptosis in the beginning, which then changes into apoptosis of the inner media. It is suggested that the cytolytic necrosis or ischemic degeneration observed in the DA is a result of the sustained contraction of the SMCs and occurs following the apoptotic events. Following the apoptotic and necrotic events, there is a sustained and complete loss of blood supply and the vessel degenerates into the ligamentum arteriosum. which in the adult is only a remnant of the DA. Patent Ductus Arteriosus Infants born premature show an increased risk for developing a number of serious complications during the neonatal period, particularly those related to the cardiopulmonary circulation. One of the frequent complications of prematurity is patent DA, which is the second most common congenital heart disease. 13, 14 Patent DA is the delay or complete failure of DA closure during the first few days after birth. The incidence of patent DA increases with the degree of infant prematurity and is often observed in combination with respiratory distress. For infants with symptoms of respiratory distress, patent DA occurs in one-quarter of neonates born from weeks of gestation, whereas the incidence is greater than 75% in extremely premature infants 3
19 born less than 29 weeks of gestation. 15 Alterations in systemic or pulmonary circulation 16, 17 produce three types of cardiovascular manifestations associated with patent DA. First, because of reduced pulmonary vascular resistance following birth, patent DA produces a left-to-right shunting of the circulation from the aorta into the pulmonary arteries. Recirculation of oxygenated blood through the lungs overburdens the heart and predisposes the infant to congestive heart failure. Second, diversion of blood flow in the aorta through a patent DA produces tissue specific deficits in perfusion, which increases susceptibility to intraventricular hemorrhage, necrotizing enterocolitis, cerebral ischemia and renal insufficiency. Third, blood flow through the patent DA into the lungs contributes to pulmonary hypertension and edema, thereby compromising lung compliance and gas exchange, and increasing pneumonia susceptibility. Current Treatments for Patent DA Surgical ligation or indomethacin administration are the most utilized methods of treating patent DA in preterm neonates. 18 Although surgical ligation may be the initial treatment in complicated cases when cardiothoracic teams are available, DA ligation is a major operation which requires thoracotomy and may have complications of its own. 17 Because of these limitations, intravenous administration of indomethacin is most often the first-line treatment for patent DA in preterm neonates. Although indomethacin induces an initial constriction of the DA in a majority of treated neonates, it is significantly less effective in achieving permanent closure, with DA reopening reported to occur in from 30% to greater than 50% of the treated preterm neonates. 17, 19, 20 The use of indomethacin may also compromise neonatal health by reducing intestinal, cerebral and renal blood flow Novel approaches need to be developed that are more safe and effective than indomethacin to reduce the necessity for surgical ligation to permanently close the DA in preterm infants. 4
20 FIGURE 1.1 A. Illustration of the fetal circulation before birth. Arrows indicate the direction of the flow of blood from the right ventricle to the DA, bypassing the pulmonary arteries, thereby allowing majority of the blood to bypass the lungs. 5
21 Figure 1.1 B. Illustration of patent DA 26 Arrows indicate the back-flow of the oxygenated blood from the aorta to the DA, with some of the flow directed back through the pulmonary arteries to the lungs. 6
22 PROSTAGLANDINS and CYCLOOXYGENASES Prostaglandins (PGs) are bioactive lipid mediators derived from arachidonic acid (AA), that were originally extracted from the prostate, semen and seminal vesicles, early in the 1930s. 27 They were first shown to cause contraction of smooth muscle and reduction in blood pressure. It is now known that prostaglandins are formed by most cells in the body and act as autocrine or paracrine lipid mediators. Prostaglandins are not stored but are synthesized de novo from membrane-released arachidonic acid, when cells are stimulated by specific cytokines, grown factors, mechanical trauma, or other stimuli. The release of arachidonic acid from membranes is controlled by a host of phospholipase A 2 enzymes, and type IV cytosolic phospholipase A 2 (cpla 2 ) shows preference for the release of phospholipids containing arachidonic acid. The activity of cpla 2 is tightly regulated by the control of translocation to the nuclear envelope, endoplasmic reticulum and the Golgi apparatus following agonist-dependent cell stimulation At the ER and nuclear membrane, arachidonic acid that is released by cpla2 or other phospholipases, is acted upon by an enzyme known as prostaglandin H synthase (PGHS). The biosynthesis of prostaglandins is dependent upon PGHS, which catalyzes the first committed step in the pathway. 32 The enzyme is also known as cyclooxygenase (COX), which was used to describe the first of two enzymatic activities of the protein, and is present as two isoforms, COX-1 and COX-2. The cyclooxygenase reaction involved in the synthesis of prostaglandins is a two-step reaction. The first step involves the oxygenation of arachidonic acid, which is enzymatically cyclized to yield endoperoxide-containing prostaglandin G 2 (PGG 2 ), by the insertion of two oxygen molecules. The protein also enzymatically reduces PGG 2 to PGH 2 via a separate peroxidase active site. PGH 2 is then converted enzymatically or by non-enzymatic hydrolysis to yield the biologically active PGs, PGE 2, PGF 2 ", PGD 2 and PGI 2 (prostacyclin), as well, as thromboxane A 2, and collectively these are known as prostanoids. These biologically active prostanoids exit the cell to activate G-protein 27, 30, 31, couple receptors to mediate their biological functions. 7
23 COX-1 COX-1 is constitutively expressed and is primarily localized in the endoplasmic reticulum. Traditionally, it was thought that prostanoids produced by this constitutively expressed COX-1 were responsible for mediating physiological housekeeping functions. Particularly high levels of COX-1 are found in platelets, kidney, stomach and skin, tissues and organs in which prostanoids have particularly specialized signaling functions. COX-1 expression increases in cell lines that undergo differentiation, and changes in COX-1 expression are primarily thought to be associated with alterations in the developmental status of the cell. Ram seminal vesicles have long been use as a tissue for isolation of COX-1 protein for enzymatic studies, and in 1989 the COX-1 gene was first cloned from sheep, and later the complete amino acid sequence of the human COX-1 gene was determined The gene for COX-1 is approximately 22 kb in length with 11 exons resulting in the protein consisting of 576 amino acids with a molecular mass of approximately 70 kda. The 5' flanking region of the COX-1 gene does not contain a TATA box, which is characteristic of a housekeeping gene. There are several regulatory elements present in the COX-1 promoter, which include, two Sp1 motifs, two AP2 sites and an NF-IL6 motif. 42 Studies with human umbilical vein endothelial cells (HUVEC), have shown that Sp1 cis-regulatory element, contributes to the constitutive expression of COX-1. It was shown that mutation of either Sp1 sites, resulted in a reduction of the promoter activity by 50%, as compared to the wild-type, whereas mutations in both the 27, 36, 42 Sp1 sites, resulted in reduction of the basal activity by 75%. COX-2 COX-2 was discovered in the early 90s as primary response gene. 38, COX-2 is an inducible enzyme, and is traditionally thought to play an important role in the production of prostanoids that primarily contribute to pathological conditions. The expression of COX-2 is known to increase in response to stimuli in various tissues and cells. COX-2 is primarily localized in the nuclear envelope, and various proinflammatory factors such as IL-1, TNF-alpha, INF-gamma, lipopolysaccharide (LPS), TPA, hormones such as FSH and LH, and several growth factors including EGF, PDFG and FGF are known to stimulate expression of COX-2. The COX-2 gene is located on a 8
24 separate chromosome than COX-1, which for humans is chromosome The COX-2 promoter contains the TATA box, as well as binding sites for several transcription factors such as NF-#B, NF-IL6, as well as the cyclic AMP response element binding protein (CREB). 27, 51, 52 The 3 main MAPK pathways, including ERK1/2, JNK/SAPK and p38 27, 36, 42 are known to contribute to the induction of the COX-2 gene. Inhibition of COX activity The amino acid sequence for COX-1 and COX-2 share a 60% identity. 53 There are 3 distinct domains that comprise the structure of the COX proteins: an N-terminal epidermal growth factor domain, a membrane binding motif, and a C-terminal catalytic domain that consists of the COX and peroxidase active sites. The COX active site is situated at the end of a hydrophobic channel that runs from the membrane-binding 54, 55 surface of the enzyme into the interior of the molecule. Non-steroidal anti-inflammatory drugs (NSAIDs), such as aspirin have been prominent anti-inflammatory, analgesic and antipyretic medications since 1889 when aspirin was first marketed. NSAIDs act on the COX active site in several ways. Aspirin irreversibly inactivates both COX-1 and COX-2 (primarily COX-1) by acetylating an active serine site at residue This is a covalent modification that interferes with the positioning of arachidonic acid into the active site. In contrast, drugs such as ibuprofen are reversible competitive inhibitors of both COX isoforms, that compete with arachidonic acid at the COX active site. 57 Drugs such as flurbiprofen and indomethacin are a third class of NSAIDs that cause a slow, time-dependent irreversible inhibition of 31, 56, 58 COX-1 and COX-2. Traditional NSAIDs such as ibuprofen and indomethacin, which inhibit the activity of both COX-1 and COX-2, provide significant therapeutic efficacy. However, the chronic use of these medications is associated with severe side effects such as gastrointestinal ulcerations and bleeding. Because COX-1 is not up-regulated during inflammatory conditions and was thought to be involved in housekeeping functions such as cytoprotection of the gastric mucosa, the side effects of traditional NSAIDs were attributed to the inhibition of COX-1. Whereas, because COX-2 was known to be upregulated primarily during pathological conditions such as pain and inflammation, the 9
25 therapeutic effects of traditional NSAIDs were attributed to inhibition of COX-2. Because of this rationale, the identification of drugs that selectively inhibit COX-2 activity, became the focus of NSAID development. The crystal structures for COX-1 and COX-2 are remarkably similar, and have provided insight into how specificity for COX-2 is achieved. 55, 59 There is a single amino acid difference within the hydrophobic channel of the COX enzyme at position 523 (isoleucine in COX-1 and valine in COX-2), which has been shown to be critical for the selectivity of several drugs for COX It is thought that the smaller valine molecule in COX-2 creates a larger 'side-pocket' which gives substrate access to COX-2. This results in a 17% increase in NSAID binding by the COX-2 active site, and allows for development of more bulky inhibitors with COX-2 binding capability which are not efficiently bound by COX Celecoxib and rofecoxib are examples of novel inhibitors that were developed from these binding studies which produce selective inhibition of COX-2 activity. 10

26 FIGURE 1.2. Illustration of the cyclooxygenase pathway 11
27 COX-1 and COX-2 deficient mice The biological functions of the COX enzymes were originally hypothesized from in vivo and in vitro studies examining the pharmacological effects of NSAIDs. However, because of activity on non-cox targets, the lack of selectivity for an individual COX isoform, and only partial inhibition of either or both COX isoforms, the pharmacological effects of NSAIDs may not always reflect physiological or pathological functions of the COX isoforms. To better understand the physiological and pathological functions of the COX isoforms, mice deficient in either COX-1 or COX-2 were developed Studies using these mice provided insight into the novel roles of both COX-1 and COX-2. These studies revealed that COX-1 in addition to COX-2 is involved in the development of various pathologies, whereas COX-2, in addition to COX-1 plays a significant role in development and the maintenance of homeostasis. Because COX-1 was traditionally thought to be responsible for maintaining housekeeping functions, it was surprising that the COX-1-deficient mice had very few phenotypic abnormalities , 69 The most unexpected finding was that these mice did not develop spontaneous gastric ulcerations, as it was thought that prostaglandins from the activity of COX-1 were responsible for maintaining the integrity of the gastric mucosa. Furthermore, these mice did not display kidney pathology and showed decreased ex vivo platelet aggregation without prolonged bleeding times in vivo. The most severe phenotype observed in the COX-1-deficient mice was the delayed onset of labor in 100% of the females. Studies by Tiano et al., using the COX-1-deficient mice showed that COX-1 plays an important role in skin tumorigenesis. 70 Therefore, these knockout studies have determined that COX-1 is essential for providing only a limited number of house keeping functions but is capable of playing a significant role in pathological conditions , 66, 69, 70 In contrast to the COX-1-deficient mice, the COX-2-deficient mice exhibit overt phenotypes. 64 All COX-2-deficient mice are born in the expected Mendelian ratios, however only about 65% of these mice survive up to weaning. Furthermore, the kidneys of these mice show a nephropathy, with poorly developed and reduced number of glomeruli, and pale and smaller kidneys. The female mice are infertile and have impaired 12
28 ovulation, implantation and decidualization. 71 Thus, studies using the COX-2-deficient mice revealed that COX-2 plays an important role in normal physiological processes FACTORS REGULATING DA CLOSURE 15-hydroxyprostaglandin dehydrogenase The dilation of the DA in utero is an active process maintained primarily by PGE 2, circulating in the fetus The placenta is a major source of PGE 2 in the fetal circulation 76 and traditionally it is thought that loss of this PGE 2 source following birth, and the resultant decline in PGE 2 levels, initiates contraction of the DA. 2 PGE 2 is catabolized into biologically inactive products by 15-hydroxyprostaglandin dehydrogenase (PGDH) and recently, mice deficient in PGDH were developed, and these mice exhibit a patent DA phenotype. 77 Treatment of PGDH-deficient neonates with indomethacin, in the immediate postnatal period corrects the patent DA. Therefore, it was concluded that patent DA in PGDH-deficient mice resulted from elevated levels of PGE 2 after birth. It is known that PGDH expression increases late in gestation, particularly in fetal lungs. 78 Because a majority of the fetal circulation is shunted away from the lungs through the DA; PGE 2 is thought to be protected from being catabolized by PGDH in utero. However, after birth when the transition to pulmonary respiration occurs and blood flow to the lungs increases, PGE 2 is broken down by PGDH, thereby resulting in a drop in PGE 2 levels, and initiation of DA closure. 13

29 Figure 1.3. PGE 2 maintains dilation of the DA in utero. The dilation of the DA in utero is active process maintained by primarily by PGE 2, circulating in the fetus. The placenta is a major source of PGE 2 in the fetal circulation. Loss of this PGE 2 source following birth, together with induction of PGDH, the enzyme that catabolizes PGE 2, is thought to initiate closure of the DA. 14
30 Cyclooxygenases To better understand the functions of both COX-1 and COX-2, we developed mice that were deficient in both enzymes. These mice were born alive in the expected Mendelian ratios, however 100% of these mice died within hours after birth. After extensive pathological analysis, the only pathology that was found in these animals was a patent DA. 79 Until our recent work, the predominant function of the COX isoforms expressed in the DA, was thought to be the synthesis of PGE 2. 80, 81 Numerous observations in humans and rodents indicate that maternal administration of NSAIDs constrict the DA in late-term fetuses Therefore, the finding of patent DA in the COX-deficient mice was surprising because the lack of PG production in COX-deficient mice was expected to cause premature DA contraction, similar to the effects of NSAIDs. After the finding of patent DA in the COX-1/COX-2 double knockout mice, the COX-2- deficient neonates were re-examined, and it was found that 35% of these mice also have a patent DA. Thus, our work using mice with targeted disruptions in the genes encoding COX-1 and COX-2 was the first report to identify a role for COX-2 in DA closure after birth. (Table 1) 79 We extended these genetic studies to examine the effects of pharmacological inhibition, and found that specific inhibition of COX-2 during pregnancy also dramatically increases the incidence of patent DA in neonatal mice. (Table 1) 86 These findings are highly significant to human health because COX-2 inhibitors are currently being used in pregnant women in the United States and Europe to evaluate the efficacy of these medications for treatment of preterm labor. 87, 88 Our work suggests that if the mechanisms of DA closure are conserved between species, the use of COX-2 inhibitors by women during pregnancy may impede normal closure of the DA after birth, thereby compromising neonatal health. We have also demonstrated that the DA expresses a greater level of COX-2 as compared to the adjacent aorta, suggesting the autocrine synthesis of vasoconstrictor prostanoids that may initiate DA closure after birth. However, to date, the identity of the specific COX-2-derived constrictor prostanoid or that of the specific contractile prostanoid receptor remains elusive. 15
31 Table 1. Incidence of patent DA with the genetic deficiency or pharmacological 79, 86 inactivation of COX-1 and/or COX-2 in mice. Genetic deficiency of COX-2 results in patent DA in mice. This incidence increases when the gene copy number for COX-1 is reduced. Celecoxib treatment significantly increases patent DA incidence in neonates expressing COX-2 (*, P < 0.01, significantly different from vehicle treated). The patent DA incidence in the COX-2-deficient neonates following celecoxib administration does not increase significantly above the spontaneous patent DA incidence, indicating the selectivity of the drug for COX-2. Untreated Celebrex Treated Patent DA Patent DA Offspring Genotype % Incidence % Incidence COX-1(+/+)/COX-2(+/+) 0! COX-1(+/!)/COX-2(+/+) 0! COX-1(!/!)/COX-2(+/+) 0 Patent Da % Incidence 100 COX-1(+/+)/COX-2(+/!) 0! Offspring COX-1(+/!)/COX-2(+/!) Genotype Vehicle 0 Celecoxib 79 COX-1(+/+)/COX-2(+/+) COX-1(!/!)/COX-2(+/!) 0 59* 100 COX-1(+/!)/COX-2(+/+) COX-1(+/+)/COX-2(!/!) * COX-1(!/!)/COX-2(+/+) COX-1(+/!)/COX-2(!/!) * COX-1(+/!)/COX-2(+/!) COX-1(!/!)/COX-2(!/!) * COX-1(!/!)/COX-2(+/!) 0 100* COX-1(+/+)/COX-2(!/!) COX-1(+/!)/COX-2(!/!) COX-1(!/!)/COX-2(!/!) 100! 16
32 Other factors regulating DA closure In addition to vasodilatory prostaglandins, nitric oxide has been known to have an accessory role in dilation of the DA. Nitric oxide is known to be a dilator of DA smooth muscle. Studies in the lamb DA have shown that nitric oxide donors such as sodium nitroprusside and glyceryl trinitrate, both of which increase intracellular concentrations of cgmp, are capable of dilating the DA. 2 It has been shown previously in vitro that treatment of the DA with inhibitors of nitric oxide synthase, caused constriction of the DA, however the effect was minimum. 89 Other studies have shown that treatment of the DA with inhibitors of nitric oxide synthase in combination with a COX inhibitor such as indomethacin, is more effective than either class of agents used alone. 90 Thus, even though nitric oxide is involved in dilating DA in utero, it only plays an accessory role. Physiological increases in oxygen tension have been known to influence closure of the DA. It has traditionally been thought that a decrease in circulating PGE 2 concentrations after birth, together with an increase in arterial oxygen tension that follows the first breath, both trigger postnatal DA closure. 2 Oxygen is known to increase intracellular calcium concentrations and cause contraction of DA smooth muscle. Studies in vitro have shown that treatment of the DA with antagonists for potassium and calcium channels, together with oxygen exposure resulted in inhibition of delayed rectifier potassium channels, causing membrane depolarization and increased entry of calcium, thereby resulting in DA constriction. 2 Although potassium channels are involved in oxygen-dependent DA constriction, a role for this ion channel in contributing to patent DA has not been shown. Furthermore, studies in our laboratory have found that treatment of COX-1/COX-2 or COX-2 -deficient neonates with inhaled oxygen does not enhance DA closure. 86 Thus, even though increased oxygen tension may normally be involved in DA closure, oxygen-dependent stimulation is not sufficient to produce DA closure in the absence of the COX isoforms. The SMCs and endothelial cells in the DA are known to synthesize endothelin-1 (ET-1) and exogenous ET-1 is a potent constrictor of the DA. In vitro studies have shown that fetal DA synthesis of ET-1 is regulated by oxygen tension 2, and ET-1 receptor type 17
33 A antagonists partially dilate DA constriction induced by oxygen. 91 Although there are 92, 93 conflicting reports on the ability of ET-1 receptor antagonists to affect DA closure, the deficiency of the ET-1 receptor type A does not affect normal postnatal DA closure in mice. 94 PROSTANOID RECEPTORS Individual prostanoids act through specific membrane bound receptors to mediate their biological responses To date, eight types of prostanoid receptors have been identified and are highly conserved between mice and humans. These receptors include: the PGD 2 receptors, DP1, DP2 (CRTH2- chemoattractant receptor homologous molecule expressed on Th2 cells); the PGE 2 receptors, EP1, EP2, EP3 and EP4; the PGF 2" receptor FP, the prostacyclin receptor IP; and the TXA 2 receptor TP. 97, 98 All prostanoid receptors are G protein-coupled rhodopsin-type receptors with distinct intracellular signaling pathways (Table 2). Among these prostanoid receptors, DP, EP2, EP4 and IP have been termed as the relaxant receptors that mediate a rise in intracellular camp. Whereas, TP, EP1 and FP are termed the contractile receptors, as they contribute to calcium mobilization. EP3 is also considered a contractile receptor, but it mediates a reduction in camp levels, and thus is termed as an 'inhibitory' receptor. Mice deficient in each of these receptors have been developed and studies using these mice have revealed 96, 98 novel physiological and pathophysiological functions of prostanoids. 18
34 Table 2. Prostanoid receptors, signaling and vascular function. Prostanoid Receptor Subtype Signaling Vascular Function PGD 2 DP DP1 $ camp Dilation DP2 (CRTH2) % camp, Phospholipase C activation Undetermined PGE 2 EP EP1 $ Ca 2+ Constriction EP2 $ camp Dilation EP3 % camp Constriction EP4 $ camp Dilation PGF 2" FP Phospholipase C activation, $ Ca 2+ Constriction PGI 2 IP $ camp Dilation TXA 2 TP Phospholipase C activation, $ Ca 2+ Constriction 19
35 Dilatory prostanoid receptors and the DA It is well known that the dilation of the DA in utero is an active process that is maintained by actions of PGE 2. Therefore, most of the studies to date have focused on investigating the role of the PGE 2 receptors in the regulation of DA dilation. Studies by Smith et al. examined the expression of the dilatory receptors in the rabbit DA using ex vivo techniques. 99 These studies utilized pharmacological agonists and antagonists, to examine the effect of these drugs on dilation of the pre-constricted DA ex vivo. These studies showed that an agonist for the EP4 receptor was equipotent to PGE 2 in dilating the DA, whereas the EP1 and EP3 agonist sulprostone, failed to mediate DA relaxation. Furthermore, an antagonist specific for the EP4 receptor blocked the dilatory effect of PGE 2 on the DA. Therefore, it was concluded that the primary PGE 2 receptor in the rabbit DA was EP4. However, studies by Nguyen et al., and Segi et al., showed that mice 100, 101 deficient in the EP4 receptor surprisingly had a postnatal patent DA phenotype. These findings suggested a role of PGE 2 receptors other than EP4 in maintaining DA dilation and/or a novel previously unidentified function of EP4 that is required for DA closure. Studies by Smith et al., extended their findings to the ovine and baboon DA and found that EP3 and EP4 were expressed, however the expression of EP4 decreased with advancing gestational age. 102 Studies by Bhattacharya et al. examined the expression of the EP receptors in the porcine DA. 103 These studies suggested that EP1 was not expressed in the porcine DA, but EP2, EP3 and EP4 were present. Furthermore, they suggested that the expression of EP3 and EP4 decreased with advancing gestation, whereas the expression of EP2 remained unchanged. From these studies, it was concluded that the primary dilatory receptor in the porcine DA was EP2. This group extended their studies in the ovine DA and found that similar to the porcine DA, EP2, EP2 and EP4 were expressed in the fetal DA, whereas only EP2 was expressed in the newborn DA. Furthermore, stimulation of both EP2 and EP4 caused DA relaxation by mediating a rise in camp levels. They also suggested that although stimulation of the EP3 receptor caused a decline in camp levels, activation of this receptor produced DA relaxation. These studies were the first to suggest a role for the known contractile EP3 receptor in dilation of the ovine DA. The relaxant 20
36 effect of EP3 was attributed in part to the stimulation of K ATP channels. 104 A more recent study by Waleh et al., also examined EP receptor expression in the immature and mature, sheep and baboon DA. 105 They found that in contrast to the studies by Smith et al., 102 the expression of EP4 remained unchanged in the DA, and EP2 was expressed in the DA at all stages of gestation. Thus, from the multitude of reports to date describing the various functions of EP receptor expression in the DA of different species, there have been several discrepancies in the findings. Although the significance of alterations in EP receptor expression has yet to be determined, the most consistent observation in the reports to date is the presence of the EP4 receptor in the DA and the ability of the DA to dilate in response to EP4-specific agonists. We have also recently shown that endogenous treatment of wild-type neonatal mice with an EP4-selective agonist, results in DA patency. 86 So the question as to why the EP4-deficient mice exhibit a patent DA phenotype remains to be answered. A recent report by Yokoyama et al. has shed some light into this paradoxical finding of patent DA in EP4 knockout mice. 106 These studies proposed an additional role for EP4, other that mediating DA dilation. First, they examined the expression of EP4 in the rat DA and found that EP4 expression significantly increased with advancing gestational age. Smooth muscle migration from the vascular media into the endothelial layer is an important vascular remodeling process involved in complete closure of the DA. These studies examined migration of the DA smooth muscle in response to EP4 agonist stimulation and found that similar to the effects of PGE 2, an EP4 agonist successfully resulted in migration of DA smooth muscle cells. Furthermore, they showed that EP4 stimulation resulted in induction of the hyaluronic acid gene. Hyaluronic acid is an important component of the extracellular matrix, and is involved in intimal cushion formation, and smooth muscle cell migration during DA remodeling. The DA of EP4- deficient mice had reduced expression of hyaluronic acid. Thus, it was concluded that hyaluronic acid production is stimulated by EP4 receptor activation and production of this extracellular matrix component is an essential process in DA remodeling, without which, results in patent DA
37 Contractile prostanoid receptors and the DA Few studies have been reported to date, examining the effects of contractile prostanoids, and the characterization of contractile receptors in the DA. The prostanoid receptors known to mediate contraction of vascular smooth muscle are EP1, EP3, FP and TP. Studies by Smith et al., investigated the effects of agonists for the receptors on contraction of the rabbit DA ex vivo. 107 The EP1/EP3 agonist, sulprostone, as well as the TP agonist U46619, caused concentration-dependent contraction of the DA, in the low nanomolar range, as examined by the isometric tension technique. The FP agonist had no effect on DA contraction, but in contrast caused DA relaxation at higher concentrations, which may be due to the non-specific binding of the agonist to other receptors. The endogenous ligand for EP1/EP3 is PGE 2, and the primary effect of this ligand on the DA is known to be dilation. Therefore, it is not clear how the same endogenous ligand can have different effects on ductal tone when binding either the contractile or relaxant receptors. Furthermore, the authors themselves question these findings by saying that the contractile effects of sulprostone on the DA may have a non-specific effect by binding to either TP or other prostanoid receptors. Therefore, in these studies, it was concluded that the TP receptor might have a role in mediating DA contraction. We recently reported the endogenous effects of prostanoid receptor agonists on DA contraction in vivo. Mice that are deficient in both COX-1 and COX-2 show a 100% incidence of patent DA, and thus provide an advantageous model for examining the effects of different prostanoids or analogs on DA closure. 86 COX-1/COX-2 double knockout mice were treated with sulprostone, an FP agonist, an EP4-specific agonist, PGE 2, carbaprostacyclin (IP agonist), U46619 (a TP agonist and a PGH2 mimetic) or IBOP (TP agonist and TXA 2 mimetic). Of all the compounds examined, only the two structurally distinct TP agonists, U46619 and IBOP, induce DA closure in the COX- 1/COX-2 double knockout mice in vivo, which is histologically indistinguishable from normal DA closure that occurs in wild-type mice. Treatment of COX-1/COX-2 double knockout mice with agonists selective for all other prostanoid receptors has no effect on DA closure. Thus, these studies suggest a role for the TP receptor in mediating DA closure. However, genetic deficiency of the only known TP receptor in mice does not alter DA closure 108, suggesting the role for a novel prostanoid receptor in DA closure. 22
38 Furthermore, the possibility of an unidentified TP isoform has been suggested previously. 97, 109, 110 Thus; it is possible that a currently uncharacterized contractile receptor is present, which may be responsible for mediating the actions of COX-2- derived prostanoid(s) on the DA. Copyright Darshini Trivedi
39 METHODS Animals The wild-type mice used in these studies (8 to 10 weeks of age) were on a mixed background of C57BL/6J and 129/Ola and have been maintained by continuous 64, 65, 69 intercrossing with mice from the same colony, as we have reported previously. EP4 wild-type, heterozygous and homozygous -deficient mice were generated by crossing EP4 heterozygous mice that were backcrossed ten times to C57BL/6CrSlc (Japan SLC, Shizuoka, Japan). EP4 mutants were genotyped by PCR as previously described. 101 The morning after pairing was designated as gestation day 0.5 upon detection of a copulation plug. Preterm wild-type mice were delivered by Cesarean section on gestation days 16 (82% gestation) or 17 (87% gestation). Full-term wild-type mice were delivered either by Cesarean section or natural birth on gestation day 19.5 (100% gestation). Mice were housed under barrier conditions with food and water provided ad libitum. Experiments were conducted in accordance with the Institutional Animal Care and Use Committee at the University of Kentucky. Tissue Collection Mice for the 0 hour time point were euthanized immediately following birth whereas all mice that were be analyzed at the 3 hour time point, were placed in an atmosphere of 100% oxygen for the first 10 minutes following birth to improve survival. At the designated time points, animals were be euthanized in a CO 2 chamber followed by thoracotomy, to expose the heart and great vessels. The DA extends from the main pulmonary artery and connects to the proximal descending aorta just after the origin of the left subclavian artery. DA tissue was excised carefully without contamination of surrounding vessels, and lysed immediately in tissue lysis buffer from the RNA isolation kit. RNA Isolation Total RNA was isolated from tissues using the RNAqueous Kit from Ambion, which uses a modified version of the traditional Chomczynski and Sacchi method. 111 The RNAqueous method utilizes the ability of nucleic acids in concentrated chaotropic salt 24
40 solutions, to bind glass fibers. DA tissue was first lysed in a glass micro-mortar and pestle apparatus, using a buffer containing a high concentration of guanidium salt, which also inactivates RNAses. The lysate was then diluted with an ethanol solution, which allowed the RNA to bind the glass fibers in the filter cartridge. The solution was passed through the filter cartridge, and the filter pad, where the RNA remained bound while the other cellular components flowed through. The filter cartridge was washed 3 times using wash buffer to allow removal of contaminating materials. RNA was then eluted using a hot elution solution of low ionic strength. RNA concentrations were measured using the Nano Drop system. cdna Preparation Total RNA (~150 ng per DA) was reverse transcribed using the following components: random hexamers, 10X reverse transcription buffer, deoxynucleotide triphosphate mix (dntps), RNAse inhibitor, and Superscript II reverse transcriptase. The reverse transcription was carried out using a thermal cycler (Eppendorf) with the following conditions: RNA + hexamers + dntps at 65 C for 5 min followed by placing the mixture on ice for 5 min. This was followed by addition of the remaining components to the tube and back into the thermal cycler for the following conditions: 25 C for 10 minutes, 42 C for 50 minutes and 70 C for 15 minutes to terminate the reaction. This cdna was stored at -20 C until utilized for gene expression analysis. Quantitation of mrna expression Gene expression quantitation was performed in a two-step RT-PCR (ABI Prism 7000 system) in which the PCR step is coupled with fluorogenic 5' nuclease chemistry (Taqman chemistry). Primer/probe assays for COX-1, COX-2, EP2, EP3, EP4, and TP were purchased from Applied Biosystems (Foster City, CA). mrna encoding the housekeeping gene, hypoxanthine phospho-ribosyl transferase (HPRT) was also quantitated for an internal normalizing control. 112 There was no significant difference in HPRT mrna levels throughout gestation, when compared to other housekeeping genes (data not shown). A relative standard curve using cdna from TPA-treated skin or lung tissue was run within the same reaction. The quantity of mrna for the gene of interest 25
41 was extrapolated from its respective standard curve, followed by normalization with the HPRT levels. Because the expression analysis for each gene of interest was performed on separate reaction plates, and because the primers for each gene of interest were of different efficiencies, comparison of expression levels between genes of interest was not performed. Histology At the designated time points, neonates were sacrificed followed by fixation of torsos in 10% neutral buffered formalin. As we have described previously 79, 86, upper torsos were transected above the rib cage and processed (paraffin embedded) for histological analysis. Analyses were performed on transverse sections of the upper thorax. The entire length of the DA from the descending aorta to the bifurcation of the pulmonary arteries was serial sectioned, followed by immunohistochemical analysis for COX-2 (primary antibody from Cayman Chemical, Ann Arbor, MI), smooth muscle cell marker "-actin (Dako Cytomation, Carpentaria, CA), endothelial cell marker PECAM (Santa Cruz) and the intermediate filament protein, desmin (Chemicon). Antigen retrieval was carried out in citrate buffer by heating to 96 C in a microwave oven, followed by cooling for 20 min at room temperature. Sections were first blocked with 1% BSA, 1% nonfat dried milk, and 1% normal serum. Incubations with the primary antibodies were carried out overnight at room temperature in a humidity chamber. Antibody binding was detected using the Vectastain Elite ABC kit (Vector laboratories, Burlingame, CA), using the manufacturer's instructions. DA sections were also stained with hematoxylin (H) and eosin (E) for morphological analysis of DA closure. Statistics Data are expressed as mean ± SEM. Data were analyzed using GraphPad Prizm software and tested for normality. Statistically significant differences in the expression of COX-1, COX-2, EP2, EP4 and TP, at multiple stages of gestation were determined by one-way ANOVA. Two-way ANOVA was utilized in the measurement of gene expression differences between the DA and aorta, at different time-points. Unpaired 26
42 Student's t-test was used to determine difference of means in the experiments involving gene expression at 2 different time-points after birth, or two different gestational timepoints. Copyright Darshini Trivedi
43 CHAPTER 2 CHARACTERIZATION OF THE CELL TYPES IN THE MOUSE DA Introduction Morphology of the DA. The wall of the DA is composed of three layers: 1) The intima which is comprised of endothelial cells 2) intimal cushions which consist of cellular and elastic layers between the endothelial cells and the internal elastic lamina and 3) the medial layer which consists of SMCs which are oriented longitudinally, circularly and spirally. At early gestation, the medial SMC layer in the DA is similar to that of the adjacent great vessels. In order to achieve permanent closure after birth, the DA develops in utero such that it acquires a highly muscular phenotype. With advancing gestation, the intimal cushions in the DA progressively become thicker than those in the adjacent arteries. The internal elastic lamina becomes fragmented and prominent and thin layers of elastin and collagen line the layers in the intimal cushions. 4 Differences between the DA and Adjacent Great Vessels. Despite the similarities between the developmental origin of the DA and adjacent arterial beds, the DA unlike its neighboring arteries, becomes highly muscular as development progresses. Even though the internal lamina of the DA consists of an elastin layer, as compared to the adjacent arteries, the DA has significantly less elastin. 4, 5, 113 In normal vascular smooth muscle, two types of smooth muscle type myosin heavy chain isoforms SM1 and SM2, and two other non-muscle type MHC isoforms are expressed. Of the two smooth muscle type isoforms, SM2 is associated with the contractile phenotype of SMCs. Several studies have shown that as compared to the adjacent aorta and pulmonary artery, the DA expresses a greater level of SM2. 4, 114, 115 The increased expression of SM2 in the DA as compared to the adjacent great vessels is thought to contribute to the contractile nature of DA smooth muscle. Furthermore, myofilament structures, indicative of contractile function, were more abundant in the DA as compared to adjacent vessels. 4 Studies in our laboratory (unpublished) as well as by others have shown that expression of the intermediate filament protein desmin, indicative of the 28
44 advanced differentiation and development of contractile function, is greater in the DA of mice, as compared to adjacent aorta. 4 Thus, the DA is a muscular artery, which consists of highly differentiated SMCs and possesses a contractile machinery which is unique to this artery. 29
45 Results We utilized the mouse DA as a model for studying the morphology of this vessel. We employed the periodic acid/schiff's staining method to identify the basement membrane and to distinguish between the layers of cells in the DA. As shown in figure 2.1, the DA is comprised of multiple layers of cells. The media of the DA is primarily formed of smooth muscle cells (Figure 2.2). The inner media appears to be comprised of radially oriented smooth muscles cells, whereas the middle and outer layers of the media contains more closely packed, circularly oriented smooth muscle cells. These medial smooth muscle cells are encompassed with a layer of fibroblast like cells in the adventitia. Endothelial cells comprise the inner lining of the intimal layer, as shown in Figure 2.3. The formation of intimal cushions has been thought to be an important event in remodeling of the DA. When comparing figures 2.2 and 2.3, it appears as if the endothelial cells have separated from the internal elastic lamina, and smooth muscle cells have migrated to occlude the lumen. Progressive development of the intimal cushions is accompanied by fragmentation of the internal elastic lamina, as shown in (Figure 2.4). In contrast, in the patent DA, the internal elastic lamina appears to be intact with apparent distinct layers (Figure 2.5). We also examined expression of the contractile filament protein desmin in the DA and the adjacent vessels. Our studies show that the DA expresses a dramatically greater level of desmin in the SMCs of the DA as compared to SMCs of the aorta and adjacent arteries (Figure 2.6). This increased desmin expression contributes to the contractile nature of the DA, as opposed to the neighboring elastic arteries. 30

46 Figure 2.1 Basement membrane staining This figure represents a completely remodeled DA, from a neonatal mouse at 3 hours after birth. Sections were stained with periodic acid/schiff reagent (Newcomer supply). Basement membrane DA Ao 31

47 Figure 2.2 Smooth muscle cell staining Slides were stained with an antibody against "-actin, which is a marker of smooth muscle cells. The slides are counter stained with periodic acid/schiff's reagent, which allows distinguishing between the different layers of cells. Brown staining (DAB reagent) indicates smooth muscle cells. Basement membrane Smooth muscle cells 32
 have detached from the internal elastic lamina, and have segregated in")
48 Figure 2.3 Endothelial cell staining Slides were stained with an antibody against PECAM, which is a marker of endothelial cells. As shown in the figure, it is apparent that the endothelial cells (as indicated by the brown staining) have detached from the internal elastic lamina, and have segregated in the lumen. Endothelial cells Basement membrane 33


49 Figure 2.4 Elastin staining Closed DA Patent DA 34
 indicates greater desmin expression in the DA as compared to the adjacent aorta. Figures shown are representative of DA from a minimum of 5 mice.")
50 Figure 2.5 Desmin staining Sections were stained with an antibody against the contractile filament protein, desmin. The brown staining (DAB reagent) indicates greater desmin expression in the DA as compared to the adjacent aorta. Figures shown are representative of DA from a minimum of 5 mice. DA, ductus arteriosus, Ao, descending aorta. DA Ao Copyright Darshini Trivedi
51 CHAPTER 3 ANALYSIS OF INDOMETHACIN-INDUCED CONSTRICTION OF THE DA IN UTERO Introduction The role of PGs in the regulation of DA tone was initially determined from the observation that NSAIDs such as indomethacin modulate DA tone following birth. The dilation of the DA in utero is active process maintained by prostaglandins PGs, primarily PGE 2, circulating in the fetus. The placenta is thought to be the major source of the circulating PGE 2 in the fetus. It has been shown that in humans and rodents, maternal administration of indomethacin results in the reduction of the circulating PGE 2 levels, possibly by inhibiting the placental PGE 2 source, thereby resulting in DA constriction. 2 Studies in sheep and rats have shown that maternal administration with indomethacin, late in gestation results in premature constriction of the fetal DA Similar to the effects of indomethacin, we and others have shown previously, that the COX-2-selective inhibitor, celecoxib, when administered acutely to late gestation fetuses in utero, results in premature constriction of the fetal DA. 86, 116 In contrast to the effect of the COX inhibitors on late gestation fetuses, studies have shown that the preterm fetuses are less responsive to the constricting effects of COX inhibitors. 82, 117 This suggests that the development of the DA with advancing gestation may be related to the contractile function of the DA. In the current studies, we utilized maternal indomethacin treatment as a method to investigate the relationship between the ontogeny of the DA and the development of DA contractile function. 36
52 Results It is known that the DA of fetuses remains patent, thereby allowing the blood flow in the heart to bypass the uninflated lungs. We examined DA closure in mouse fetuses on gestation day 18.5 following maternal treatment with vehicle. Similar to our previous findings, the DA of fetuses treated with vehicle in utero, was completely patent. (Figure 3A) As we have previously reported, we utilized indomethacin treatment to examine the ability of the fetal DA to close prematurely. Indomethacin was administered to pregnant female mice on gestation day Four hours after dosing, fetuses were analyzed for premature DA constriction by histology. As compared to the completely patent DA of vehicle-treated controls (Figure 3A), indomethacin treatment induced complete occlusion of the DA of late term fetal mice on gestation day 18.5 (Figure 3B). We also utilized indomethacin treatment to examine the ability of the DA of preterm fetuses to contract in utero. Our results show that in contrast to the DA of late gestation fetuses, indomethacin resulted only in partial constriction of the preterm fetal DA of mice on gestation day 16 (Figure 3C). These studies show that in mice, the preterm gestation fetal DA is less responsive to pharmacologically-induced constriction, and suggest that near full-term maturation is required for complete development of DA contractile function. 37
53 Figure 3 Incomplete indomethacin-induced constriction of the DA in preterm fetal mice. Indomethacin was administered to pregnant dams on gestation day 16 or 18.5 (50 mg/kg, oral gavage 5% gum arabic). Four hours after dosing, fetuses were delivered by Cesarean section and sacrificed for DA analysis. n & 5 per group. DA, ductus arteriosus, Ao, descending aorta. DA of fetal mice treated with vehicle A) or indomethacin B) on gestation day C) DA of fetal mice treated with indomethacin on gestation day 16. Sections are stained with H&E. 38

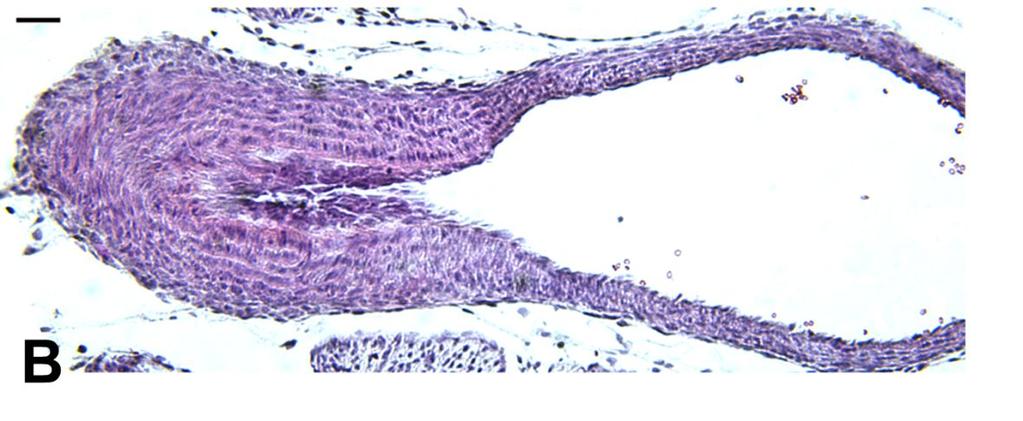

54 Vehicle Indomethacin- day 18.5 Indomethacin- day 16 39
55 Discussion Fetal DA constriction induced by the non-selective COX inhibitor indomethacin has long been used to study pharmacologically-induced contractile responses in the DA. 105, 118 Maternal treatment with the non-selective COX inhibitor indomethacin, is known to induce two paradoxical effects on the DA. In both humans and rodents, maternal administration of indomethacin results in premature constriction of the DA by inhibiting the synthesis of prostanoids important for DA dilation. 2, 85, 117 Alternatively, it has also been shown that indomethacin treatment during pregnancy can increase the risk for postnatal patent DA Our recent report has shown that similar to previous studies with indomethacin, acute administration of a COX-2-selective inhibitor but not a COX-1- selective inhibitor, results in premature constriction of the DA.Whereas chronic administration with a COX-2-selective inhibitor, results in postnatal patent DA. 86 A recent report by Reese et al., replicated our findings in mice and showed similar results. 122 The contrasting effects of COX-2 inhibition that we showed recently, and the paradoxical effects of indomethacin shown by others, suggest opposing actions of different prostanoids on the regulation of DA tone. The dilation of the DA in utero is active process maintained by prostaglandins PGs, primarily PGE 2, circulating in the fetus. The placenta is thought to be the major source of the circulating PGE 2 in the fetus. Therefore, it is possible that the constriction of the DA that is observed following acute administration of indomethacin or a COX-2-selective inhibitor results from the reduction of circulating PGE 2 levels by inhibiting COX activity in the placenta. In contrast, the patent DA observed with indomethacin treatment or with chronic treatment with a COX- 2 inhibitor may be due to sufficient inhibition of synthesis of contractile prostanoids produced by COX-2 in the DA. Although we do not completely understand the two opposing effects on the DA resulting from indomethacin treatment, particularly at the time of peak COX-2 expression, nonetheless we can use this agent as a convenient functional marker for the ability of COX-2 to induce DA closure. Previous studies in rats and humans have shown that the preterm fetal DA is less responsive to indomethacin-induced constriction, in contrast to the DA of full-term fetuses. 82, 117 In the current studies, we examined the effect of gestational age on 40
56 indomethacin-induced fetal DA constriction in mice. Our results show that the preterm fetal DA on gestation day 16 was less responsive to the constricting effects of indomethacin, as compared to the late gestation fetal DA on day Therefore, in mice the contractile nature of the DA develops late in gestation. Copyright Darshini Trivedi
57 CHAPTER 4 ANALYSIS OF COX-1 AND COX-2 EXPRESSION IN THE FETAL AND NEONATAL MOUSE DA Introduction Previous studies in higher species such as pigs and sheep, have examined the developmental regulation of the COX isoforms in the DA. Studies by Guerguerian et al., measured COX-1 and COX-2 expression in fetal and newborn porcine DA and determined that COX-1 expression was constitutive in the fetal and newborn DA. 80 In contrast, COX-2 expression was only detected in the newborn DA. They also examined the relative contribution of the COX isoforms in the synthesis of PGE 2 and found that COX-2 was responsible for synthesizing greater than 90% of PGE 2 in the newborn DA. Based on these findings, Guerguerian et al. concluded that COX-2 does not play a role in regulating DA tone after birth. Another study by Clyman et al., examined the expression of the COX isoforms in the late-gestation fetal lamb DA. 1 These studies suggested that both COX-1 and COX-2 were expressed in the fetal lamb DA. The expression of COX-1 was detected in the endothelial cells lining the DA lumen, as well as the smooth muscle cells of the media. However, COX-2 expression was only detected in the endothelium lining the DA lumen. These studies also examined the relative contributions of COX-1 and COX-2 for PGE 2 synthesis, using selective inhibitors of each isoform, and suggested that both COX isoforms contributed equivalently to the production of PGE 2 in the DA. A more recent study by Baragatti et al. examined the expression of the COX isoforms in the full-term gestation fetal mouse DA. 123 They showed that both COX-1 and COX-2 mrna were expressed in the DA of full-term fetal mice. These studies also suggested that COX-2 co-localized with microsomal PGE 2 synthase-1, an enzyme downstream of the COX isoforms, which converts the intermediate prostanoid PGH 2 into PGE 2. Based on these findings, Baragatti et al. suggested that in the full-term fetal DA, COX-2 is the isoform responsible for synthesizing PGE 2. Recently, studies by Rheinlaender et al. examined COX isoform expression in the human DA and suggested that COX-1 is the predominant isoform expressed in the DA 42
58 throughout gestation, whereas the expression of COX-2 was weak. 124 These studies also suggested that that the cellular expression pattern of the COX isoforms changed with advancing gestation. All these previous reports have focused on the role of the COX enzymes in synthesizing PGE 2 important for DA dilation. Furthermore, there are several discrepancies regarding the expression of the COX enzymes in the fetal DA. The factors involved in ontogenic changes in COX expression seem to vary between tissues and cells, and thus far remain elusive. Our previous studies have shown that about 35% of mice genetically deficient in COX-2 show a postnatal patent DA and resulting mortality. The patent DA incidence increases as the gene copy number for COX-1 and COX-2 decreases, reaching a 100% in mice doubly deficient in both COX-1 and COX-2. 65, 79 We extended these findings and showed that wild-type neonatal mice born after exposure of pregnant mice with a COX-2 selective inhibitor, show an increased postnatal patent DA incidence. 86 These studies indicated the importance of COX-2-derived prostanoids in postnatal closure of the DA. Our studies were the first to suggest that COX-2-derived prostanoid(s), other than PGE 2 play an active role in closure of the DA after birth. Our previous studies also indicated the importance of COX-2 in constriction of the DA in utero. We utilized indomethacin treatment to assess the ability of the fetal DA to constrict. Our studies showed that all fetuses genetically deficient in COX-2 were resistant to indomethacin-induced premature DA constriction. 79 This suggested that the expression of COX-2 is required for the contractile function of the DA in utero. In the current studies, we examined the expression of COX-1 as well as COX-2 in the fetal DA at multiple stages of gestation, as well as the neonatal DA at full-term gestation. These studies will provide a better understand of the role of the COX isoforms in DA closure after birth. 43
59 Results Expression of COX-2 mrna in the fetal DA at multiple stages of gestation. We examined the time-course of COX-2 expression in the DA of fetal mice at multiple stages of gestation. To perform these studies, fetuses were obtained by Cesarean section on days 16, 17 and 19.5 of gestation, and DA tissue was excised immediately (0 hours) for analysis of mrna expression. Comparative quantitative analysis of COX-2 mrna expression showed that COX-2 expression significantly increased with advancing gestation, with the highest expression in the DA on day 19.5 (Figure 4.1). Furthermore, the gestation day 16 time-point with the lowest level of COX-2 mrna coincided with the time at which we observed incomplete indomethacin-induced constriction of the fetal DA (Figure 3C). These data demonstrate a correlation between reduced DA constriction and attenuated COX-2 expression in the DA of preterm mice. 44
 mice were obtained at 0 hr (sacrificed immediately after birth) for analysis of mrna expression for COX-2 in the DA by real-time PCR.")
60 Figure 4.1 Expression of COX-2 increases in the fetal DA with advancing gestation. Preterm (gestation days 16 and 17) and full-term (gestation day 19.5) mice were obtained at 0 hr (sacrificed immediately after birth) for analysis of mrna expression for COX-2 in the DA by real-time PCR. The housekeeping gene HPRT was used as an internal control for normalizing mrna levels. Data represented as mean COX-2 mrna levels +/- SEM. **, significantly different from 0 hour day 16, P < 0.01 (one-way ANOVA), n & 10. ***, significantly different from 0 hour day 16 and 0 hour day 17, P < (one-way ANOVA), n & 10 45
61 Expression of COX-1 mrna in the fetal DA at multiple stages of gestation. We also quantitated COX-1 mrna expression in the DA on gestation days 16, 17 and 19.5 of gestation. We found that in contrast to COX-2 expression, the expression of COX-1 did not significantly change with advancing gestational age (Figure 4.2). This finding suggests that alterations in COX-1 mrna expression are not associated with in utero development of fetal DA contractile function. 46
62 Figure 4.2 Expression of COX-1 remains constitutive in the fetal DA with advancing gestation. Preterm (gestation days 16 and 17) and full-term (gestation day 19.5) mice were obtained at 0 hr (sacrificed immediately after birth) for analysis of mrna expression for COX-1 in the DA by real-time PCR. The housekeeping gene HPRT was used as an internal control for normalizing mrna levels. Data represented as mean COX-1 mrna levels +/- SEM. 47
63 Analysis of COX-1 and COX-2 expression in the DA of neonatal mice at full-term gestation We examined COX-2 mrna expression in the DA of full-term neonates, immediately after birth (0 hours) when the DA was patent, and 3 hours after birth when the DA was completely closed. Quantitative real-time PCR analysis indicated that COX-2 mrna dramatically increased (approximately 10 fold) in the DA when compared between 0 and 3 hours after birth on gestation day 19.5 (Figure 4.3-A). This increase in COX-2 mrna expression from 0 to 3 hours after birth was unique to the DA, as no such increase in COX-2 expression was observed in the adjacent aorta. We also examined COX-2 protein expression by immunohistochemistry and found that significant COX-2 protein was localized in the smooth muscle cells of the DA with evidence of perinuclear expression, but not in the smooth muscle cells of the adjacent aorta (Figure 4.3-B). Therefore, a significant increase in smooth muscle cell expression of COX-2 accompanies closure of the DA after full-term birth. We also compared COX-1 expression in the full-term neonatal mouse DA between 0 and 3 hours of birth. In contrast to COX-2 expression, the expression of COX- 1 mrna was constitutive with no significant difference between the two time-points (Figure 4.3-C). 48
64 Figure 4.3 Induction of COX-2 expression in the DA of full-term neonatal mice. A) Full-term mice were delivered by Cesarean section or natural birth on gestation day 19.5 and sacrificed at 0 or 3 hours after birth for analysis of COX-2 mrna in the DA or aorta. Data represented as mean COX-2 mrna levels +/- SEM. *, significantly different from 0 hour DA and 3 hour Ao. P < (two-way ANOVA), n & 10. B) Immunohistochemical analysis of COX-2 expression. Full-term mice were delivered by Cesarean section or natural birth on gestation day 19.5 and sacrificed at 2 or 3 hours after birth. Brown staining (di-amino benzidine, DAB) indicates COX-2 expression. Figure shown is representative of neonatal DA from a minimum of 5 mice after 2 hours of birth. Inset shows perinuclear staining. DA, ductus arteriosus, Ao, descending aorta. C) Real-time PCR analysis of COX-1 mrna expression in the DA at 0 and 3 hours after birth on gestation day Data represented as mean COX-1 mrna levels +/- SEM. 49

65 A 50

66 C 51
67 Discussion Several groups have examined the developmental regulation of the COX isoforms in the DA in multiple species. There have been significant inconsistencies in these reports, showing that either COX-2 is not expressed, is expressed weakly or increases during gestation. 1, 80, 123 Furthermore all these reports together focus on the role of COX-2 in synthesizing the dilatory PGE 2 responsible for maintaining a dilated DA in utero and suggest that the withdrawal of the dilatory PGE 2 after birth may allow for DA closure. It is also suggested that COX-2 does not play a role in regulating DA closure after birth. These studies by other groups investigating the ontogeny of the COX isoforms in the DAs of other species do not explain our findings of the role of COX-2 in mediating DA closure. The present investigation was carried out to resolve the apparent inconsistencies in previous reports and gain, at the same time, a better insight into the functional organization of the COX enzymes in the DA. We have previously reported that the DA of COX-2-deficient fetuses fails to constrict in response to indomethacin, suggesting the importance of COX-2 expression for in utero contractile function. 79 Therefore, in the current study, we investigated the relationship between the ontogeny of COX-2 expression and the developmental regulation of fetal DA constriction. COX-2 expression in the DA was significantly greater in full-term fetuses than that in preterm fetuses. Our previous report, together with our current study suggest that the contractile function of the DA that develops towards the end of term requires the in utero increase in COX-2 expression with advancing gestational age. The factors responsible for inducing normal closure of the DA after birth have not been clearly defined. Our previous studies using mice deficient in COX-2, clearly demonstrate the importance of this enzyme in postnatal DA closure. In mice, DA closure commences within 30 minutes after full-term birth, and is complete by 3 hours after birth. We have found that that COX-2 expression is induced in the neonatal DA at 3 hours after birth following full-term gestation, which coincides with the time at which the DA is completely remodeled. Thus, the induction of COX-2 in the DA may be required for normal closure of the DA after birth. The induction of COX-2 that we observe in the DA is unique to this vessel, as no 52
68 such increase is observed in the adjacent vessels such as the aorta. This suggests that autocrine production of prostaglandins in DA actively mediates DA closure. The vascular remodeling process involving intimal thickening, that occurs during closure of DA, are similar to the processes that occur in pathological process such as atherosclerosis and restenosis. 125 In such pathological conditions, it has been reported previously that COX-2 is significantly induced in the aorta. This induction of COX-2 in the aorta during atherosclerosis is associated with increase in production of prostanoids such as PGE 2 and TXA 2, together with macrophage accumulation and recruitment of inflammatory cytokines and chemokines A recent study by Waleh et al. showed that similar to atherosclerosis, macrophage accumulation in the vessel wall plays an important role in DA remodeling. 10 They found that this macrophage accumulation is associated with the induction of cytokines and adhesion molecules, and the extent of neointimal remodeling is directly related to the degree of mononuclear cell adhesion. Thus, the DA provides a physiological model of vascular remodeling, and studies on the DA may provide further insight into the development of treatments for vascular pathologies such as atherosclerosis. Copyright Darshini Trivedi
69 CHAPTER 5 ANALYSIS OF POSTNATAL DA CLOSURE AND COX EXPRESSION AT PRETERM AND FULL-TERM GESTATION IN MICE Introduction The initial phase of closure of the DA involves a contraction of its thick muscular wall. This is followed by anatomical remodeling of the vessel, which includes migration of the smooth muscle cells into the sub-endothelial space, creating intimal cushions. Following remodeling of DA SMC, significant apoptosis and cytolytic necrosis are thought to be required for complete obliteration of the DA. DA closure in humans is complete within a few days after birth. However, in other species such as mice, rats and rabbits, DA closure is complete within a few hours after birth. After permanent closure of the DA, a remnant of the vessel remains, which is known as the ligamentum arteriosum, which is formed by fibrosis of the closed vessel. 12 The factors responsible for inducing normal closure of the DA after birth have not been clearly defined. Our previous studies using mice deficient in COX-2, clearly demonstrate the importance of this enzyme in postnatal DA closure. In mice, DA closure commences within 30 minutes after full-term birth and is complete within 3 hours. Our current studies show that COX-2 expression is dramatically induced in the neonatal DA within 3 hours of birth at full-term gestation, which correlates with the time-point when the DA is completely remodeled. This suggests that induction of COX-2 expression in the DA may be required for achieving permanent closure after full-term birth. In humans, the incidence of patent DA is increased at premature gestation, however the mechanisms responsible for this increased incidence are not clear. Our previous study utilized indomethacin treatment as a functional marker of constriction of the DA in utero. We showed that the DA of premature fetuses failed to completely constrict in response to indomethacin, as opposed to the DA of full-term fetuses. We also examined the ontogeny of COX isoforms in the fetal DA at multiple stages of gestation. We found that the time-point of gestation when the DA of preterm fetuses failed to constrict in response to indomethacin, coincided with the time-point at which the 54
70 expression of COX-2 in the DA was the lowest. This suggests that the failure of DA to constrict prematurely may be due to a reduced expression of COX-2. In the current study, we utilized the mouse to establish a model of postnatal patent DA resulting from prematurity in gestation. To date, there has no animal model established to study the mechanisms of DA patency at preterm gestation. Because our previous study showed that the preterm gestation fetuses show incomplete indomethacininduced constriction, we hypothesized that neonatal mice born at preterm gestation may also show a post-natal patent DA. Furthermore, we compared the expression of COX-1 and COX-2 in the DA from preterm and full-term neonatal mice. We hypothesized that similar to our observations in preterm fetal mice, the DA of preterm full-term neonatal mice will show attenuated COX-2 expression. 55
71 Results Morphological analysis of DA closure at 0 and 3 hours after birth at full-term gestation As shown in Figure 5.1, we found that that the DA was completely patent in neonatal mice at 0 hrs after birth at full-term gestation. This was indicated by the blue dye flowing through the widely open DA, with its diameter being similar to that of the adjacent aorta. The DA was completely occluded in neonatal mice at 3 hours after birth on gestation day This was indicated by end of the blue dye at the point where the pulmonary artery ended and the DA began. The opaque white tissue is indicative of a completely closed DA. 56
 The DA of a neonatal mouse, analyzed immediately")
 The DA of a neonatal mouse analyzed 3 hours after birth.")
72 Figure 5.1. Morphological Analysis of DA Closure A) The DA of a neonatal mouse, analyzed immediately after birth. The blue dye passing through the DA is indicative of DA patency. B) The DA of a neonatal mouse analyzed 3 hours after birth. The failure of the blue dye to pass through the DA is indicative of a closed DA. Arrow, DA. Patent DA Closed DA 57
73 Histological analysis of DA closure at 0 and 3 hours after birth at full-term gestation We also analyzed DA closure in mice using histological techniques. When analyzed at 0 hours after birth on gestation day 19.5, the DA was completely open, with no evidence of neointimal thickening, and the diameter of the vessel wall being similar to that of the adjacent aorta. (Figure 5.2A) In contrast, the DA was completely remodeled in all neonates sacrificed at 3 hours after birth at full-term gestation. (Figure 5.2B) This was indicated by significant neointimal formation and complete obliteration of the vessel lumen. Histological analysis of DA closure at 3 hours after birth at preterm gestation Our studies confirmed that the DA of mice born at full-term gestation closes completely at 3 hours after birth. In this study, we wanted to examine the effect of prematurity in gestation on DA closure in mice. In order to successfully accomplish these studies, it was important to determine the survival incidence of mice born at preterm gestation, and to ensure that the survival incidence was comparable to that of full-term gestation mice. Based on our previous studies, we know that the survival incidence of full-term wild-type neonates delivered by Cesarean section on day 19 is approximately 98%. To determine the survival incidence of preterm wild-type neonates, we delivered litters on gestation day 17.5 by Cesarean section. The neonates were placed in an atmosphere of 100% oxygen for the first 10 minutes following birth and exposed to tactile stimulation using a vibrating heating pad and monitored for up to 5 hours after birth. A survival incidence of 83% (19/23) was observed in these preterm neonates. This level of survival was acceptable for the studies to examine the effect of prematurity on DA closure. Our findings indicate that in contrast to the complete DA closure observed in all neonates analyzed at 3 hours after full-term birth on day 19.5, significant lumen was visible in the DAs of all preterm neonates that were analyzed at 3 hours after birth on gestation day 17 (Figure 5.2C). These findings indicate that the DAs of neonatal mice born premature show incomplete closure. 58
74 Figure 5.2 Histological Analysis of DA Closure A) This figure is a representative transverse section of a completely patent DA of neonatal mice immediately after birth at full-term gestation. B) This panel is a representative transverse section of a completely closed DA of neonatal ice, 3 hours after birth at full-term gestation on day C) Representative transverse section of an incompletely closed DA from neonatal mice, 3 hours after birth at preterm gestation on day 17. Sections are stained with H&E. DA, ductus arteriosus, Ao, descending aorta. 59



75 Figure 5.2. Histological analysis of DA closure. A 0 hr-day 19.5 B 3 hr- day 19.5 C 3 hr- day 17 60
76 Postnatal expression of COX-1 and COX-2 in the DAs of preterm neonatal mice. To determine whether the incomplete closure of the DA in preterm neonatal mice that we observed (Figure 5.2) was associated with attenuated expression of COX-2, we compared COX-2 expression in the DA between preterm (day 17) and full-term (day 19.5) neonates at 3 hours after birth. In comparison to the level of expression observed in the DAs of full-term neonates, COX-2 mrna expression in the DAs of preterm neonates, was significantly lower (Figure 5.3A). The attenuated COX-2 expression coincided with the time-point at which we observed incomplete closure of the DA at preterm gestation (Figure 5.2C). In contrast to COX-2 expression, there was no significant difference in COX-1 mrna expression between DAs from preterm and fullterm neonatal mice at 3 hours after birth (Figure 5.3B). Immunohistochemical analysis of COX-2 protein expression indicated that the DA of full-term neonatal mice showed significantly greater number of COX-2-positive cells than the DA of preterm neonatal mice (Figures 5.4C and 5.4D). Furthermore, the COX-2 expression was primarily localized in the smooth muscle cells of the DA (Figure 5.4E). Thus, these data establish a direct correlation between the attenuated COX-2 expression and increased patent DA. 61
 mice were delivered by Cesarean section or natural delivery, and sacrificed at 3 hours after birth for analysis of COX-1 and COX-2 expression.")
77 Figure 5.3 Attenuated COX-2 mrna expression in the DA of preterm neonatal mice. Preterm (gestation day 17) and full-term (gestation day 19.5) mice were delivered by Cesarean section or natural delivery, and sacrificed at 3 hours after birth for analysis of COX-1 and COX-2 expression. A) COX-2 mrna expression is significantly greater in the DA of day 19.5 neonates as compared to the DA of day 17 neonates. *, P < 0.05 (Student's t test), n & 5. B) COX-1 mrna expression is not significantly different between the DA of preterm and full-term neonatal mice. Data are represented as mean COX-1 or COX-2 mrna levels +/- SEM. A B Figure 5B hr day 17 3hr day 19.5 Gestational Age 62
 The DA of a day 17 neonatal mouse at 3 hours after birth shows fewer COX-2- positive cells as compared to D) the DA of a day 19.5 neonatal mouse at 3 hours after birth.")
78 Figures 5.4 Attenuated smooth muscle COX-2 protein expression in the DA of preterm neonatal mice. C) The DA of a day 17 neonatal mouse at 3 hours after birth shows fewer COX-2- positive cells as compared to D) the DA of a day 19.5 neonatal mouse at 3 hours after birth. Significant lumen is visible in C), indicative of partial DA closure due to prematurity. E) DA of a day 19.5 neonatal mouse at 3 hours after birth, stained for smooth muscle "-actin. Brown staining (DAB reagent) indicates COX-2 or actin expression and sections are counterstained with hematoxylin (blue). Figures shown (magnification, X300) are representative of immunohistochemical analysis of neonatal DA from a minimum of 5 mice. 63
79 Discussion In humans, patent DA is more common in premature infants, than those born at full-term, although mechanisms responsible for the increased incidence are poorly understood. We have previously reported that genetic or pharmacological inactivation of COX-2 results in patent DA in mice, indicating the importance of COX-2 in postnatal closure of the DA. 79, 86 The current studies were aimed at better understanding the COX- 2-dependent mechanisms of DA closure and elucidating the factors responsible for patent DA at preterm gestation. Our findings are the first to suggest that reduced expression of COX-2 in the DA at preterm gestation contributes to increased patent DA. Our studies are the first to utilize the mouse as a model to examine patent DA at preterm gestation. Normal DA closure in mice at full-term gestation is complete within 3 hours after birth. With our finding that incomplete development resulted in attenuated fetal DA constriction, we investigated whether prematurity in gestation also compromised normal DA closure after birth. We observed that at 3 hours after birth, DA closure was incomplete in preterm neonates, in contrast to the completely remodeled DA of 3 hour-old neonates born at full-term gestation. This suggests that similar to humans, premature birth compromises postnatal DA closure in mice. We have shown that COX-2 expression is induced in the DA of neonatal mice following full-term birth, and correlates with the time at which the DA is completely remodeled. This suggests that the increased expression of COX-2 is required for the normal closure of the vessel after birth. Therefore, we hypothesized that the incomplete closure of the DA in preterm gestation neonatal mice that we observed, may result from attenuated expression of COX-2. In support of this hypothesis, we found that COX-2 expression was significantly lower in the DAs of neonatal mice delivered at preterm gestation, as compared to that expressed in the DAs of full-term neonates. The reduced COX-2 expression corresponded with the time-point at which we observed incomplete closure of the preterm neonatal DA. Thus, our studies demonstrate a direct correlation between the attenuated expression of COX-2 and the failure of complete closure of the DA at preterm gestation. Previous studies examining developmental regulation of the COX isoforms in the DA have focused on the action of these enzymes in DA dilation. In other species, it has 64
80 been reported that COX-1 is the predominant isoform expressed in the premature fetal DA, whereas COX-2 is primarily expressed in the DA late in gestation. 80 Our current findings in mice are in agreement with previous reports, and show that COX-1 is constitutively expressed in the DA throughout gestation, and COX-2 is induced in the DA from preterm to full-term gestation. It has also been suggested that the DA at preterm gestation is more sensitive to dilation in response to PGE 2 derived from the activity of COX-1, a process which may contribute to increased postnatal patent DA. 80, 129 Our work emphasizes the importance of COX-2 in mediating DA closure. Because of its potent dilatory effect, it is unlikely that PGE 2 is the COX-2-derived prostanoid that induces DA closure. We propose that COX-2 expressed in the DA synthesizes a prostanoid other that PGE 2, that directly mediates DA closure. Therefore, patent DA at preterm gestation may result from constitutive expression of COX-1 together with the attenuated expression of COX-2. 65
; whereas the expression of COX-1 is constitutive in the DA, resulting in the synthesis of")
81 Figure 5.5. Proposed model of normal DA closure at full-term gestation and DA patency at preterm gestation. At full-term gestation, COX-2 expression is induced, resulting in the synthesis of constrictor prostanoid(s); whereas the expression of COX-1 is constitutive in the DA, resulting in the synthesis of the dilatory PGE 2. At this time-point, the effect of the vasoconstrictor overrides the vasodilator, the resultant effect being DA closure. However, at preterm gestation, the COX-2 is not induced, whereas the expression of COX-1 is constitutive, resulting in the predominant effect of the dilatory PGE 2, resulting in patent DA. Copyright Darshini Trivedi
82 CHAPTER 6 PROSTANOID RECEPTORS IN THE MOUSE DA Introduction Individual prostanoids act through specific membrane bound receptors to mediate their biological responses. 97, 98 To date, eight types of prostanoid receptors have been identified and are highly conserved between mice and humans. Prostanoid receptors are G protein-coupled rhodopsin-type receptors with distinct intracellular signaling pathways. Mice deficient in each of these receptors have been developed and studies using these mice have revealed novel physiological and pathophysiological functions of 96, 98 prostanoids. In numerous species including humans, PGE 2 has long been known to be a potent dilator of the DA. There are four known genes that encode the PGE 2 receptors, namely EP1, EP2, EP3 and EP4. Pharmacological studies have suggested that EP1 and EP3 are 104, 105, 110 the contractile receptors, whereas EP2 and EP4 are the dilatory receptors. Because of the known vasodilatory function of PGE 2 on the DA, there has been considerable interest in identifying the major PGE 2 receptor expressed in the DA to better understand the role of PGE 2 in mediating ductal dilation. There have been several studies in a variety of animal species including lamb, baboon, rat, rabbit and pig, examining the expression of the EP receptors in the DA. Of all the EP receptors, EP4 is the one that is thought to be primarily expressed in the DA. However, there have been conflicting results regarding expression of the other EP receptors, as well the expression pattern of all the EP receptors in the DA during the transition from fetal to neonatal life. Studies using the rabbit DA suggest that only the EP4 receptor is present in the DA, whereas in the porcine DA, EP2, EP3 and EP4 are expressed in equivalent proportions. These studies using the porcine DA also suggest that expression of EP3 and EP4 is down-regulated in the newborn. Another study by Smith et al., utilized pharmacological agents to examine ligand-receptor binding, and suggested that expression of the EP4 receptor declined with advancing gestational age, whereas more recent studies using similar techniques have suggested that the expression of this receptor does not change. 102 Thus overall, depending on the species, gene expression in 67
83 the DA for EP2, EP3 and EP4 does not change, decreases, or increases with advancing 99, 102, 103, 105, 107, 110 gestation. Most reports to date have focused on examining expression of prostanoid receptors responsible for maintaining DA dilation. Our recent studies using mice deficient in the COX isoforms indicated the importance of COX-2 expression for DA closure after birth. We also demonstrated that the DA expresses high levels of COX-2 during normal closure after birth, suggesting novel contractile actions of COX-2- dependent prostanoids in the DA. These studies suggested that a prostanoid in addition to the dilatory PGE 2 was synthesized in the DA, which mediated DA closure. The report by Smith et al., for the first to suggest contractile actions of agonists for the TXA 2 receptor TP, as well those for the PGE 2 receptors EP1 and EP The studies were performed using pharmacological techniques to measure ex vivo contractile effects of prostanoid or prostanoid agonists. In our recent study, we utilized a variety of prostanoid analogs to examine the in vivo effects of these agents in mediating DA contraction. Of all the pharmacological agents that we examined, only the PGH 2 analog U46619, and the TXA 2 analog, IBOP that are known to bind the TXA 2 receptor TP, mediated DA contraction in vivo. 86 Previous studies have suggested that the DA does not express TXA 2 synthase, and therefore does not synthesize TXA Based on these studies together with our previous studies, it is possible that PGH 2 may be the prostanoid that actively mediates normal DA closure. Furthermore, these studies implicated a role for the TP receptor in DA closure. The studies by Smith et al., showed that the TP receptor is expressed in the rabbit DA. 107 In the current studies, we utilized mouse DA to examine the expression of the various prostanoid receptors in the DA. In addition to the COX-deficient mice, mice deficient in the EP4 receptor also exhibit a patent DA phenotype, 100, 101 which is a surprising finding based on the known dilatory role for PGE 2. Based on our studies, we believe that an induction in COX-2 expression is important for closure of the DA after birth in mice. However, the mechanisms responsible for the induction in COX-2 expression are not clear. Because of the known role of COX-2 in DA closure, we hypothesized that the failure of DA closure in the EP4-deficient mice may be due to attenuated expression of COX-2. 68
84 Results Quantitation of prostanoid receptors in the fetal DA at multiple stages of gestation. Previous reports have suggested that the prostanoid receptors EP2, EP3, EP4 and TP may be involved in modulating DA tone. In the current studies, we examined mrna expression levels for these prostanoid receptors. As shown in Figure 6.1A, the fetal mouse DA expressed a significant level of TP, however, the level of expression did not significantly change throughout gestation. EP2 mrna expression was nearly undetectable in the fetal DA (data not shown). The expression of EP3 mrna was constitutive in the fetal DA, when examined on days 16, 17 and 19.5 of gestation (Figure 6.1B). In contrast, expression of the EP4 receptor in the fetal mouse DA increased significantly from gestation days 16 to 19.5 (Figure 6.1C). 69
 and full-term (gestation day 19.")
 TP, B) EP3 and C) EP4 mrna levels +/- SEM.")
85 Figure 6.1 Expression of EP4, but not EP3 or TP, increases with advancing gestational age. Preterm (gestation days 16 and 17) and full-term (gestation day 19.5) mice were obtained at 0 hours (sacrificed immediately after birth) for analysis of mrna expression in the DA. Data represented as mean A) TP, B) EP3 and C) EP4 mrna levels +/- SEM. EP4 mrna expression in the DA significantly increased throughout gestation, P < 0.05 (one-way ANOVA), n & 8. A B C 70
86 Reduced COX-2 mrna expression in the DA of EP4-deficient neonatal mice. In addition to the COX-deficient mice, mice deficient in the PGE 2 receptor EP4 also exhibit a patent DA phenotype. 100, 101 This finding is surprising because it is well established that PGE 2 produces dilation of the DA. 2 Furthermore, there is no evidence that EP4 receptor activation is involved in constriction of the smooth muscle of the DA or any other vasculature. Having determined that the pattern of expression of EP4 receptor mrna mimics that of COX-2 expression, we hypothesized that the expression of EP4 may be required for the induction of COX-2 in the mouse DA. We therefore examined COX-2 expression in the DAs of EP4-deficient neonatal mice with a patent DA, as compared to EP4 expressing neonatal mice with a closed DA. Our results indicate that COX-2 expression was significantly attenuated in the DAs of EP4-deficient mice at 3 hours after birth, as compared to that in the DAs of littermate EP4 heterozygous or wildtype mice (Figure 6.2). These data suggest that an interplay between EP4 and COX-2 may be required for the initiation of DA closure after birth. 71
87 Figure 6.2 Reduced COX-2 expression in the DA of EP4-deficient neonatal mice. Full-term gestation (day 19.5) EP4 wild-type (EP4 +/+ ), heterozygous (EP4 +/ ) and homozygous-deficient (EP4 / ) mice were obtained by natural delivery and sacrificed 3 hours after birth for analysis of COX-2 expression in the DA. Data represented as mean COX-2 mrna levels +/- SEM. *, significantly different from EP4 +/+ and EP4 +/ DA, P < 0.01 (one-way ANOVA), n & 8. 72
88 Discussion PGE 2 mediates its biological functions by binding to 4 different receptors, namely EP1, EP2, EP3 and EP4. PGE 2 mediates different biological responses depending on the expression of a specific receptor in a particular tissue. Traditionally, based mainly on pharmacological experiments, both EP2 and EP4 are thought to maintain relaxation responses through a cyclic AMP-dependent mechanism, whereas EP1 and EP3 are thought to be the contractile receptors. 97 A recent study identified an additional contractile function of EP3 in the DA. 104 Because PGE 2 is a known potent dilator of the DA, several studies in numerous species have focused on examining EP receptor expression in the DA to better understand the role of PGE 2 in modulating DA tone. Depending on the species, gene expression in the DA for EP2, EP3 and EP4 does not change, decreases, or increases with advancing gestation. 102, 103, 105 Of the known PGE 2 receptors, EP4 expression in the DA is most conserved between species such as rabbits, mice, lambs, baboon and humans. Therefore, it has been thought that EP4 is the primary receptor through which PGE 2 mediates its dilatory actions on the DA. Because of its known dilatory function on the DA, studies to date have focused on the targeting the EP4 receptor for the regulation of DA tone. A recent study utilized a pharmacological antagonist specific for the EP4 receptor, for causing constriction of the DA in rats. 130 The EP4 receptor antagonist ONO-AE3 208 was administered to fetal rats in utero, or to neonatal rats pre-treated with a PGE 2 analogue or vehicle. The EP4 antagonist caused premature closure of the fetal DA, as well as induced closure of the DA in neonatal rats pre-treated with PGE 2. From this study, the authors concluded the EP4 was the primary dilatory receptor present in the rat DA, and targeting this receptor might be an alternate treatment for patent DA. Another study from the same group also examined the effect of EP4 activation for causing DA dilation in neonatal rats after birth. 131 Similar to mice, DA closure in rats is complete within a few hours, whereas it takes a few days in human beings. There are certain congenital heart malformations in humans, such as coarctation of the aorta, that require keeping the DA patent after birth. Because of the potent dilatory function of PGE 2 on the DA, administration of a PGE 2 analogue is most often used as treatment for such conditions, however it is associated with certain side effects. The rationale of this study 73
89 by Momma et al., was that targeting the EP4 receptor specifically might be better than a PGE 2 analog, for treatment of defects such as coarctation of the aorta. In the study by Momma et al., the EP4 agonist was compared to the PGE 2 analog, as well as to another agonist specific for EP2, and the potential dilatory effects of the drugs were examined on already closed DAs of neonatal rats. 131 These studies showed that the EP4 agonist effectively caused DA dilation, and to a greater extent than that caused by the PGE 2 analog. Similar to their previous studies, the authors concluded that the EP4 receptor is the primary receptor responsible for mediating DA dilation. Furthermore, activation of the EP4 receptor might be a better treatment for congenital heart defects, where the DA is required to be open. Most studies to date have focused on the dilatory function of the EP4 receptor. Our current study proposes an alternate role for the EP4 receptor in the DA. Of the EP receptors that we examined in our current studies, only EP4 mrna expression significantly increased with advancing gestation. Our studies also show that COX-2 expression in the DA significantly increased with advancing gestation, and the induction in COX-2 expression is essential for mediating DA closure. Thus, the gestational expression pattern of EP4 mimicked that which we observed for COX-2. The coregulation of EP4 and COX-2 suggests a relationship between this prostanoid receptor and COX-2, for inducing DA closure. Our studies propose, that in addition to the dilatory role, activation of the EP4 receptor functions as a trigger for the induction of COX-2 expression important for DA closure. However, the mechanism(s) responsible for mediating the induction of COX-2 in the DA are not clear. In other cell types, EP4 activation is known to induce COX-2 expression. 132, 133 Our studies indicate that the genetic deficiency of EP4 results in attenuated COX-2 expression in the DA. Thus, our results provide an explanation for the paradoxical finding of patent DA in the EP4- deficient mice, and suggest that EP4 expression may be important for modulating the normal induction of COX-2 and resultant DA closure after full-term birth. We propose that EP4 receptor activation at full-term gestation initiates a signaling cascade modulating the up-regulation of COX-2 expression and resultant DA closure. The primary signaling pathway following EP4 activation that has been identified involves an increase in intracellular camp, and increased camp levels are known to 74
90 induce COX-2 expression. 31, 42, 97, 134 The increase in camp activates protein kinase A (PKA) which phosphorylates the camp response element binding protein (CREB). CREB is a transcription factor known to induce COX-2 expression by binding to the 31, 42, 52 camp response element (CRE) regulatory site present in the COX-2 promoter. However, increased camp is associated with dilation of the DA. 104 Because we know that increased COX-2 expression is important for DA closure, and because of the potential opposing dilatory action of camp, it is not likely that the camp-pka pathway is the primary mechanism utilized by EP4 for induction of COX-2 in the DA. In addition to the traditional camp-dependent pathway, recently it has been shown that EP4 receptor 132, activation initiates other signaling pathways which may be camp-independent. EP4 receptor stimulation activates phosphatidylinositol (PI) 3-kinase, resulting in activation of the transcription factor, T-cell factor 4 (Tcf-4). 136 Transcriptional activation of Tcf-4 has been shown to up-regulate the expression of COX-2 by binding to a Tcf binding element (TBE) recently discovered in the COX-2 promoter. 138 Therefore, it has been proposed that EP4 activation may be another mechanism by which COX-2 expression is induced in pathological conditions (Figure 6.3). 132, 135 Until our studies, the interplay between EP4 and COX-2-induced DA closure has not been investigated. The idea that PGE 2 may induce the expression of COX-2 has been shown previously in various cell culture systems. 139, 140 However, the EP4-deficient mice provide a unique in vivo model to study endogenous prostanoid signaling mechanisms resulting in the dramatic upregulation of COX-2 that we observe. Overall, these studies explain the mechanism by which an individual prostanoid, PGE 2, may have opposing effects on DA physiology. One of the opposing effects is dilation, which is a well-characterized function of PGE 2 in the DA; whereas in our studies we suggest an indirect effect of PGE 2 leading to the induction of COX-2 and the resulting synthesis of constrictor prostanoid, possibly PGH 2. A recent study examined the gestational expression pattern of the prostanoid receptors in the fetal rat DA. They showed that EP2, EP3 and EP4 were expressed in the fetal DA throughout gestation. However, the expression of EP4 was the greatest towards the end of term. 106 These findings are in accordance with are current results showing that EP4 expression in the fetal DA increases with advancing gestation. Furthermore, these 75
91 studies by Yokoyama et al., also propose an additional active role for the EP4 receptor in mediating DA closure
 3- kinase, resulting in phosphorylation of AKT. This results in translocation of!-catenin (!")
92 Figure 6.3. Proposed model for the induction of COX-2 expression following EP4 activation. Stimulation of the EP4 receptor by PGE 2 activates phosphatidylinositol (PI) 3- kinase, resulting in phosphorylation of AKT. This results in translocation of!-catenin (!cat) to the nucleus and activation of the transcription factor, T-cell factor 4 (Tcf 4). Transcriptional activation of Tcf 4 has been shown to up-regulate the expression of COX- 2 by binding to a Tcf binding element (TBE) recently discovered in the COX-2 promoter. 132, 133, 136 Therefore, we propose that EP4 activation may be another mechanism by which COX-2 expression is induced in the DA. Copyright Darshini Trivedi
93 CHAPTER 7 GENERAL DISCUSSION Summary and Conclusions Previous studies from our laboratory, in mice with the genetic or pharmacological inactivation of COX-2 have emphasized a role for this enzyme in the postnatal closure of the DA. The goal of the current studies was to determine the mechanisms responsible for increased patent DA at preterm gestation. First, the effect of gestational age on indomethacin-induced constriction of the fetal DA or wild-type mice was determined. It was demonstrated that administration of indomethacin to pregnant female mice on gestation day 18.5 resulted in premature constriction of the DA in utero. However, indomethacin induced only a partial constriction of the fetal mouse DA, when administered on gestation day 16. These findings were in accordance with several previous studies in other species. Therefore it was concluded that the contractile function of the DA developed late in gestation. These studies also showed that the expression of COX-2 significantly increased with advancing gestational age, whereas the expression of COX-1 remained constitutive. The time-points at which observed the lowest and greatest level of COX-2 expression in the fetal DA were observed, coincided with the time-points at which attenuated and increased in utero DA constriction were observed respectively. Therefore it was concluded that the expression of COX-2 was important for the in utero development of ductal contractile function. I then wanted to investigate the effect of gestational age on postnatal DA closure in wild-type mice. These studies showed that similar to humans, DA closure was compromised in neonatal mice born at preterm gestation. It was also found that COX-2 expression was attenuated in the DA of neonatal mice born at preterm gestation, whereas the expression of COX-1 did not significantly change. These studies suggested that attenuated COX-2 expression might contribute to increased patent DA at preterm gestation. Thus, these studies also provide a novel animal model for studying the mechanisms of patent DA. 78
94 I also utilized another model of patent DA in mice resulting from the deficiency of the PGE 2 receptor EP4. PGE 2 has long been known to be a potent dilator of the DA; therefore the finding of a patent DA in EP4-deficient mice was surprising. Because of the current studies emphasizing a role for COX-2 in DA closure, it was hypothesized that a failure of induction in COX-2 expression resulting in patent DA in the EP4-deficient mice. In support of this hypothesis, it was found that COX-2 expression was significantly attenuated in neonatal mice deficient in the EP4 receptor. These studies suggested a role for EP4 in the induction of COX-2 in the DA. It was also found that in the fetal DA the expression of EP4 followed a pattern similar to that of COX-2, during advancing gestation. Thus, these studies provided evidence for an additional role for EP4 in the developmental regulation of DA contractile function. These studies have provided a better insight into the mechanisms responsible for patent DA and for the development of novel therapeutics for this serious congenital heart defect. Clinical Relevance and Future Directions Previous studies have demonstrated that in other species COX-1 is constitutively expressed in the DA of both full-term and preterm neonates 80, 123. The current studies also show that COX-1 is constitutively expressed in the DA of fetal and neonatal mice at all stages of gestation and after birth. Even though COX-2 has been suggested to be responsible for synthesizing the majority of PGE 2 in the fetal DA at the end of gestation, COX-1 is thought to be the major contributor of PGE 2 synthesis in the DA of the premature fetus. 80, 141 It has also been suggested that the preterm gestation fetal DA is 105, 142, more sensitive to the dilatory effects of PGE 143 2, which may contribute to the increased patent DA at preterm. We believe that in preterm neonates with reduced expression of COX-2 in the DA, the dilatory effect of COX-1-derived PGE 2 predominates over COX-2-dependent DA constriction, thus resulting in the failure of DA closure. This hypothesis suggests that inhibition of COX-1-derived PGE 2 in preterm neonates will increase postnatal DA closure. Therefore, future studies in the Loftin laboratory will examine the effect of postnatal administration of a COX-1-specific inhibitor on accelerating DA closure in preterm neonatal mice. Recently, a COX-1-79
95 specific inhibitor (SC-560) has been developed and surprisingly, it lacks the gastric toxicity of classical COX inhibitors such as indomethacin, which inhibit both COX-1 and COX We have also recently shown that this COX-1 inhibitor does not adversely affect fetal or neonatal health like nonselective or COX-2-selective inhibitors. 86 Therefore, COX-1-specific inhibition may be more safe and effective than the current pharmacological or surgical treatments for patent DA in preterm infants (Figure 7). 80

96 Figure 7. COX-1 inhibitor treatment for patent DA at preterm gestation. At preterm gestation, the effect of COX-1-derived PGE 2 predominates over the vasoconstrictor synthesized by the reduced COX-2, resulting in DA patency. At this timepoint, administration of a COX-1 inhibitor may accelerate DA closure by inhibiting the PGE 2 and resulting in the vasoconstrictor overriding the vasodilator. 81
EICOSANOID METABOLISM
 1 EICOSANOID METABOLISM EICOSANOIDS C20 polyunsaturated fatty acids e.g. Arachidonic acid Eicosanoids physiologically, pathologically and pharmacologically active compounds PG Prostaglandins TX - Thromboxanes
1 EICOSANOID METABOLISM EICOSANOIDS C20 polyunsaturated fatty acids e.g. Arachidonic acid Eicosanoids physiologically, pathologically and pharmacologically active compounds PG Prostaglandins TX - Thromboxanes
PHM142 Lecture 4: Platelets + Endothelial Cells
 PHM142 Lecture 4: Platelets + Endothelial Cells 1 Hematopoiesis 2 Platelets Critical in clotting - activated by subendothelial matrix proteins (e.g. collagen, fibronectin, von Willebrand factor) and thrombin
PHM142 Lecture 4: Platelets + Endothelial Cells 1 Hematopoiesis 2 Platelets Critical in clotting - activated by subendothelial matrix proteins (e.g. collagen, fibronectin, von Willebrand factor) and thrombin
Patent ductus arteriosus: pathophysiology and management
 (2006) 26, S14 S18 r 2006 Nature Publishing Group All rights reserved. 0743-8346/06 $30 www.nature.com/jp ORIGINAL ARTICLE Patent ductus arteriosus: pathophysiology and management ER Hermes-DeSantis 1
(2006) 26, S14 S18 r 2006 Nature Publishing Group All rights reserved. 0743-8346/06 $30 www.nature.com/jp ORIGINAL ARTICLE Patent ductus arteriosus: pathophysiology and management ER Hermes-DeSantis 1
Cell Signaling (part 1)
") 15 Cell Signaling (part 1) Introduction Bacteria and unicellular eukaryotes respond to environmental signals and to signaling molecules secreted by other cells for mating and other communication. In multicellular
15 Cell Signaling (part 1) Introduction Bacteria and unicellular eukaryotes respond to environmental signals and to signaling molecules secreted by other cells for mating and other communication. In multicellular
Prostaglandins And Other Biologically Active Lipids
 Prostaglandins And Other Biologically Active Lipids W. M. Grogan, Ph.D. OBJECTIVES After studying the material of this lecture, the student will: 1. Draw the structure of a prostaglandin, name the fatty
Prostaglandins And Other Biologically Active Lipids W. M. Grogan, Ph.D. OBJECTIVES After studying the material of this lecture, the student will: 1. Draw the structure of a prostaglandin, name the fatty
By Dr.Asmaa Al sanjary
 By Dr.Asmaa Al sanjary Preterm delivery is defined by a birth occurring before 37 completed weeks of gestation. Prematurity is multifactorial and its incidence has increased during the last decade in most
By Dr.Asmaa Al sanjary Preterm delivery is defined by a birth occurring before 37 completed weeks of gestation. Prematurity is multifactorial and its incidence has increased during the last decade in most
ulcer healing role 118 Bicarbonate, prostaglandins in duodenal cytoprotection 235, 236
 Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3-
 1 2 The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3- Vitelline vein from yolk sac 3 However!!!!! The left
1 2 The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3- Vitelline vein from yolk sac 3 However!!!!! The left
ANSC/NUTR 618 Lipids & Lipid Metabolism
 I. Nonessential fatty acids ANSC/NUTR 618 Lipids & Lipid Metabolism A. Synthesized completely by the fatty acid synthase reaction (e.g., myristic and palmitic acid). B. Produced by the modification of
I. Nonessential fatty acids ANSC/NUTR 618 Lipids & Lipid Metabolism A. Synthesized completely by the fatty acid synthase reaction (e.g., myristic and palmitic acid). B. Produced by the modification of
renoprotection therapy goals 208, 209
 Subject Index Aldosterone, plasminogen activator inhibitor-1 induction 163, 164, 168 Aminopeptidases angiotensin II processing 64 66, 214 diabetic expression 214, 215 Angiotensin I intrarenal compartmentalization
Subject Index Aldosterone, plasminogen activator inhibitor-1 induction 163, 164, 168 Aminopeptidases angiotensin II processing 64 66, 214 diabetic expression 214, 215 Angiotensin I intrarenal compartmentalization
Lecture Outline. Hormones & Chemical Signaling. Communication Basics: Overview. Communication Basics: Methods. Four methods of cell communication
 Lecture Outline Hormones & Chemical Signaling Communication Basics Communication Overview Communication Methods Signal pathways Regulation (modulation) of signal pathways Homeostasis... again Endocrine
Lecture Outline Hormones & Chemical Signaling Communication Basics Communication Overview Communication Methods Signal pathways Regulation (modulation) of signal pathways Homeostasis... again Endocrine
number Done by Corrected by Doctor
 number 9 Done by Nazek Hyasat Corrected by Bahaa Najjar & mohammed AL-shrouf Doctor Alia Shatnawi HOW DO DRUGS WORK??? You know that receptors are targeted by drugs, the question now is how do these drugs
number 9 Done by Nazek Hyasat Corrected by Bahaa Najjar & mohammed AL-shrouf Doctor Alia Shatnawi HOW DO DRUGS WORK??? You know that receptors are targeted by drugs, the question now is how do these drugs
Autocrine signalling. Signals are not always diffusible but may be membrane bound or a part of the extracellular matrix. The inflammatory response
 Paracrine signals Proteins, lipids that diffuse short distances Include growth factors and cytokines Secreted or synthesized as membrane-bound precursor (pro-peptide) that is proteolytically processed
Paracrine signals Proteins, lipids that diffuse short distances Include growth factors and cytokines Secreted or synthesized as membrane-bound precursor (pro-peptide) that is proteolytically processed
Blood Vessels. Types of Blood Vessels Arteries carry blood away from the heart Capillaries smallest blood vessels. Veins carry blood toward the heart
 C H A P T E R Blood Vessels 20 Types of Blood Vessels Arteries carry blood away from the heart Capillaries smallest blood vessels The site of exchange of molecules between blood and tissue fluid Veins
C H A P T E R Blood Vessels 20 Types of Blood Vessels Arteries carry blood away from the heart Capillaries smallest blood vessels The site of exchange of molecules between blood and tissue fluid Veins
Cell Signaling part 2
 15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
Pharmacology - Problem Drill 11: Vasoactive Agents
 Pharmacology - Problem Drill 11: Vasoactive Agents Question No. 1 of 10 1. Vascular smooth muscle contraction is triggered by a rise in. Question #01 (A) Luminal calcium (B) Extracellular calcium (C) Intracellular
Pharmacology - Problem Drill 11: Vasoactive Agents Question No. 1 of 10 1. Vascular smooth muscle contraction is triggered by a rise in. Question #01 (A) Luminal calcium (B) Extracellular calcium (C) Intracellular
Chapter 6 Communication, Integration, and Homeostasis
 Chapter 6 Communication, Integration, and Homeostasis About This Chapter Cell-to-cell communication Signal pathways Novel signal molecules Modulation of signal pathways Homeostatic reflex pathways Cell-to-Cell
Chapter 6 Communication, Integration, and Homeostasis About This Chapter Cell-to-cell communication Signal pathways Novel signal molecules Modulation of signal pathways Homeostatic reflex pathways Cell-to-Cell
Eicosanoid synthesis
 Seminar 12 Eicosanoid synthesis Examples of eicosanoids prostaglandins prostacyclins thromboxanes leukotrienes epoxyeicosatrienoic acids (EET) They have roles in: inflammation fever regulation of blood
Seminar 12 Eicosanoid synthesis Examples of eicosanoids prostaglandins prostacyclins thromboxanes leukotrienes epoxyeicosatrienoic acids (EET) They have roles in: inflammation fever regulation of blood
Cell Injury MECHANISMS OF CELL INJURY
 Cell Injury MECHANISMS OF CELL INJURY The cellular response to injurious stimuli depends on the following factors: Type of injury, Its duration, and Its severity. Thus, low doses of toxins or a brief duration
Cell Injury MECHANISMS OF CELL INJURY The cellular response to injurious stimuli depends on the following factors: Type of injury, Its duration, and Its severity. Thus, low doses of toxins or a brief duration
Pathology of Coronary Artery Disease
 Pathology of Coronary Artery Disease Seth J. Kligerman, MD Pathology of Coronary Artery Disease Seth Kligerman, MD Assistant Professor Medical Director of MRI University of Maryland Department of Radiology
Pathology of Coronary Artery Disease Seth J. Kligerman, MD Pathology of Coronary Artery Disease Seth Kligerman, MD Assistant Professor Medical Director of MRI University of Maryland Department of Radiology
The Patent Ductus Arteriosus (PDA) and the Preterm Baby. Tanya Hatfield, RNC-NIC, MSN Neonatal Outreach Educator
 and the Preterm Baby. Tanya Hatfield, RNC-NIC, MSN Neonatal Outreach Educator") The Patent Ductus Arteriosus (PDA) and the Preterm Baby Tanya Hatfield, RNC-NIC, MSN Neonatal Outreach Educator Objectives Describe normal cardiac physiology and development Understand the unique physiologic
The Patent Ductus Arteriosus (PDA) and the Preterm Baby Tanya Hatfield, RNC-NIC, MSN Neonatal Outreach Educator Objectives Describe normal cardiac physiology and development Understand the unique physiologic
Kofinas Perinatal Providing Care to the Unborn
 Alexander D. Kofinas, MD Director, Kofinas Perinatal Associate Professor, Clinical Obstetrics and Gynecology Cornell University, College of Medicine Patient instructions for the use of Indomethacin and
Alexander D. Kofinas, MD Director, Kofinas Perinatal Associate Professor, Clinical Obstetrics and Gynecology Cornell University, College of Medicine Patient instructions for the use of Indomethacin and
Chapter 5 Control of Cells by Chemical Messengers
 Chapter 5 Control of Cells by Chemical Messengers = How hormones and other signals work Intercellular Communication = Intercellular Signal Transmission Chemical communication Electrical communication Intercellular
Chapter 5 Control of Cells by Chemical Messengers = How hormones and other signals work Intercellular Communication = Intercellular Signal Transmission Chemical communication Electrical communication Intercellular
Chapter 9. Cellular Signaling
 Chapter 9 Cellular Signaling Cellular Messaging Page 215 Cells can signal to each other and interpret the signals they receive from other cells and the environment Signals are most often chemicals The
Chapter 9 Cellular Signaling Cellular Messaging Page 215 Cells can signal to each other and interpret the signals they receive from other cells and the environment Signals are most often chemicals The
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS
 Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Zool 3200: Cell Biology Exam 4 Part I 2/3/15
 Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Close to site of release (at synapse); binds to receptors in
; binds to receptors in") Chapter 18: The Endocrine System Chemical Messengers 1. Neural 2. Endocrine 3. Neuroendocrine 4. Paracrine 5. Autocrine Endocrine System --Endocrine and nervous systems work together --Endocrine vs. Nervous
Chapter 18: The Endocrine System Chemical Messengers 1. Neural 2. Endocrine 3. Neuroendocrine 4. Paracrine 5. Autocrine Endocrine System --Endocrine and nervous systems work together --Endocrine vs. Nervous
Nafith Abu Tarboush DDS, MSc, PhD
 Nafith Abu Tarboush DDS, MSc, PhD natarboush@ju.edu.jo www.facebook.com/natarboush Lipids (cholesterol, cholesterol esters, phospholipids & triacylglycerols) combined with proteins (apolipoprotein) in
Nafith Abu Tarboush DDS, MSc, PhD natarboush@ju.edu.jo www.facebook.com/natarboush Lipids (cholesterol, cholesterol esters, phospholipids & triacylglycerols) combined with proteins (apolipoprotein) in
Anatomy & Physiology
 1 Anatomy & Physiology Heart is divided into four chambers, two atrias & two ventricles. Atrioventricular valves (tricuspid & mitral) separate the atria from ventricles. they open & close to control flow
1 Anatomy & Physiology Heart is divided into four chambers, two atrias & two ventricles. Atrioventricular valves (tricuspid & mitral) separate the atria from ventricles. they open & close to control flow
Exam Key. NROSCI/BIOSC 1070 and MSNBIO 2070 Final Exam December 19, 2015 Total POINTS: % of grade in class
 NROSCI/BIOSC 1070 and MSNBIO 2070 Final Exam December 19, 2015 Total POINTS: 100 20% of grade in class 1) Zollinger-Ellison syndrome results from a gastrin-secreting tumor, producing increased numbers
NROSCI/BIOSC 1070 and MSNBIO 2070 Final Exam December 19, 2015 Total POINTS: 100 20% of grade in class 1) Zollinger-Ellison syndrome results from a gastrin-secreting tumor, producing increased numbers
Signal Transduction: Information Metabolism. Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire
 Signal Transduction: Information Metabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire Introduction Information Metabolism How cells receive, process and respond
Signal Transduction: Information Metabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire Introduction Information Metabolism How cells receive, process and respond
Name: Class: "Pharmacology NSAIDS (1) Lecture
 Lecture") I Name: Class: "Pharmacology NSAIDS (1) Lecture د. احمد الزهيري Inflammation is triggered by the release of chemical mediators from injured tissues and migrating cells. The specific mediators vary with
I Name: Class: "Pharmacology NSAIDS (1) Lecture د. احمد الزهيري Inflammation is triggered by the release of chemical mediators from injured tissues and migrating cells. The specific mediators vary with
Patient Ductus Arteriosus (PDA)
") Patient Ductus Arteriosus (PDA) WINSTON HELEN Author: Paediatric Department Document Number: STHK7026 Version: 5 Review date: 30th June 2021 The Normal Heart The Heart is made up of four chambers: the
Patient Ductus Arteriosus (PDA) WINSTON HELEN Author: Paediatric Department Document Number: STHK7026 Version: 5 Review date: 30th June 2021 The Normal Heart The Heart is made up of four chambers: the
DEVELOPMENT OF THE CIRCULATORY SYSTEM L E C T U R E 5
 DEVELOPMENT OF THE CIRCULATORY SYSTEM L E C T U R E 5 REVIEW OF CARDIAC ANATOMY Heart 4 chambers Base and apex Valves Pericardial sac 3 layers: epi, myo, endo cardium Major blood vessels Aorta and its
DEVELOPMENT OF THE CIRCULATORY SYSTEM L E C T U R E 5 REVIEW OF CARDIAC ANATOMY Heart 4 chambers Base and apex Valves Pericardial sac 3 layers: epi, myo, endo cardium Major blood vessels Aorta and its
Development of Blood Vessels and Fetal Circulation *
 OpenStax-CNX module: m46610 1 Development of Blood Vessels and Fetal Circulation * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the
OpenStax-CNX module: m46610 1 Development of Blood Vessels and Fetal Circulation * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the
1. Which of the following blood vessels has a thin elastic layer? A. Aorta. B. Pulmonary artery. C. Posterior vena cava. D. Mesenteric capillary.
 CIRCULATORY SYSTEM 1. Which of the following blood vessels has a thin elastic layer? A. Aorta. B. Pulmonary artery. C. Posterior vena cava. D. Mesenteric capillary. 2. Capillary beds are equipped with
CIRCULATORY SYSTEM 1. Which of the following blood vessels has a thin elastic layer? A. Aorta. B. Pulmonary artery. C. Posterior vena cava. D. Mesenteric capillary. 2. Capillary beds are equipped with
BIOLOGY. Cell Communication CAMPBELL. Reece Urry Cain Wasserman Minorsky Jackson. Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick
 CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 11 Cell Communication Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Cellular Messaging Cells can signal to
CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 11 Cell Communication Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Cellular Messaging Cells can signal to
Propagation of the Signal
 OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
Cardiovascular System Blood Vessels
 Cardiovascular System Blood Vessels Structure of Blood Vessels The three layers (tunics) Tunica intima composed of simple squamous epithelium Tunica media sheets of smooth muscle Contraction vasoconstriction
Cardiovascular System Blood Vessels Structure of Blood Vessels The three layers (tunics) Tunica intima composed of simple squamous epithelium Tunica media sheets of smooth muscle Contraction vasoconstriction
Spleen. Vertebrate hearts Pericardial cavity division in coelum. Vessel walls. Endocardium = endothelium of blood vessels. Artery elastic tissue
 Spleen White pulp macrophages, monocyte storage Red pulp - (RBC) storage, and prod n (in nonmammals) Vertebrate hearts Pericardial cavity division in coelum Endocardium = endothelium of blood vessels Fig.
Spleen White pulp macrophages, monocyte storage Red pulp - (RBC) storage, and prod n (in nonmammals) Vertebrate hearts Pericardial cavity division in coelum Endocardium = endothelium of blood vessels Fig.
A2b. PDA Management--A Reflection on the Evidence: Does it Help with Management? Session Summary. Session Objectives. References.
 FANNP 23RD NATIONAL NNP SYMPOSIUM: CLINICAL UTE AND REVIEW A2b Management--A Reflection on the Evidence: Does it Help with Management? Alfonso Vargas, MD Neonatologist Pediatrix Medical Group, Tampa, FL
FANNP 23RD NATIONAL NNP SYMPOSIUM: CLINICAL UTE AND REVIEW A2b Management--A Reflection on the Evidence: Does it Help with Management? Alfonso Vargas, MD Neonatologist Pediatrix Medical Group, Tampa, FL
AP Biology Cells: Chapters 4 & 5
 AP Biology Cells: Chapters 4 & 5 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. The was the first unifying principle of biology. a. spontaneous generation
AP Biology Cells: Chapters 4 & 5 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. The was the first unifying principle of biology. a. spontaneous generation
2. capillaries - allow exchange of materials between blood and tissue fluid
 Chapter 19 - Vascular System A. categories and general functions: 1. arteries - carry blood away from heart 2. capillaries - allow exchange of materials between blood and tissue fluid 3. veins - return
Chapter 19 - Vascular System A. categories and general functions: 1. arteries - carry blood away from heart 2. capillaries - allow exchange of materials between blood and tissue fluid 3. veins - return
Warm Up! Test review (already! ;))
)") Warm Up! Test review (already! ;)) Write a question you might find on the Unit 5 test next week! (Multiple choice, matching, fill in, or short answer!) - challenge yourself and be ready to share!!! PowerPoint
Warm Up! Test review (already! ;)) Write a question you might find on the Unit 5 test next week! (Multiple choice, matching, fill in, or short answer!) - challenge yourself and be ready to share!!! PowerPoint
NONSTEROIDAL ANTI- INFLAMMATORY DRUGS
 NONSTEROIDAL ANTI- INFLAMMATORY DRUGS MRS. M.M. HAS A 3 YR. HX OF PROGRESSIVE RIGHT HIP PAIN. THE PAIN INCREASES WITH WEIGHT BEARING ACTIVITY. PT. HAS BEEN ON ACETAMINOPHEN WITHOUT RELIEF. PERTINENT LABS
NONSTEROIDAL ANTI- INFLAMMATORY DRUGS MRS. M.M. HAS A 3 YR. HX OF PROGRESSIVE RIGHT HIP PAIN. THE PAIN INCREASES WITH WEIGHT BEARING ACTIVITY. PT. HAS BEEN ON ACETAMINOPHEN WITHOUT RELIEF. PERTINENT LABS
The Cell Organelles. Eukaryotic cell. The plasma membrane separates the cell from the environment. Plasma membrane: a cell s boundary
 Eukaryotic cell The Cell Organelles Enclosed by plasma membrane Subdivided into membrane bound compartments - organelles One of the organelles is membrane bound nucleus Cytoplasm contains supporting matrix
Eukaryotic cell The Cell Organelles Enclosed by plasma membrane Subdivided into membrane bound compartments - organelles One of the organelles is membrane bound nucleus Cytoplasm contains supporting matrix
Endothelium. A typical endothelial cell is about 30mm long, Accounts for 1% or less of the arterial weight
 Endothelium Discovered in 1845 A typical endothelial cell is about 30mm long, 10mm wide, and 0.2 3 mm thick Accounts for 1% or less of the arterial weight As recently as the late 1960s it was thought of
Endothelium Discovered in 1845 A typical endothelial cell is about 30mm long, 10mm wide, and 0.2 3 mm thick Accounts for 1% or less of the arterial weight As recently as the late 1960s it was thought of
BIOLOGY 111. CHAPTER 3: The Cell: The Fundamental Unit of Life
 BIOLOGY 111 CHAPTER 3: The Cell: The Fundamental Unit of Life The Cell: The Fundamental Unit of Life Learning Outcomes 3.1 Explain the similarities and differences between prokaryotic and eukaryotic cells
BIOLOGY 111 CHAPTER 3: The Cell: The Fundamental Unit of Life The Cell: The Fundamental Unit of Life Learning Outcomes 3.1 Explain the similarities and differences between prokaryotic and eukaryotic cells
Pathophysiology: Left To Right Shunts
 Pathophysiology: Left To Right Shunts Daphne T. Hsu, MD dh17@columbia.edu Learning Objectives Learn the relationships between pressure, blood flow, and resistance Review the transition from fetal to mature
Pathophysiology: Left To Right Shunts Daphne T. Hsu, MD dh17@columbia.edu Learning Objectives Learn the relationships between pressure, blood flow, and resistance Review the transition from fetal to mature
Leen Osama, Lujain Hamdan, Osama Mohd, Razi Kittaneh... Faisal Mohammad
 23 Leen Osama, Lujain Hamdan, Osama Mohd, Razi Kittaneh... Faisal Mohammad Revision of previous lectures G-proteins coupled receptors mechanism: When a hormone binds to G-protein coupled receptor, GTP
23 Leen Osama, Lujain Hamdan, Osama Mohd, Razi Kittaneh... Faisal Mohammad Revision of previous lectures G-proteins coupled receptors mechanism: When a hormone binds to G-protein coupled receptor, GTP
GENERAL SUMMARY Corpus luteum is a transient endocrine structure formed from the ruptured ovarian follicle. Its main function is to secrete P 4, a pro
 Corpus luteum is a transient endocrine structure formed from the ruptured ovarian follicle. Its main function is to secrete P 4, a pro-gestational hormone, essential for establishment and maintenance of
Corpus luteum is a transient endocrine structure formed from the ruptured ovarian follicle. Its main function is to secrete P 4, a pro-gestational hormone, essential for establishment and maintenance of
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression
 Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
The dynamic regulation of blood vessel caliber
 INVITED BASIC SCIENCE REVIEW The dynamic regulation of blood vessel caliber Colleen M. Brophy, MD, Augusta, Ga BACKGROUND The flow of blood to organs is regulated by changes in the diameter of the blood
INVITED BASIC SCIENCE REVIEW The dynamic regulation of blood vessel caliber Colleen M. Brophy, MD, Augusta, Ga BACKGROUND The flow of blood to organs is regulated by changes in the diameter of the blood
CONGENITAL HEART DISEASE (CHD)
") CONGENITAL HEART DISEASE (CHD) DEFINITION It is the result of a structural or functional abnormality of the cardiovascular system at birth GENERAL FEATURES OF CHD Structural defects due to specific disturbance
CONGENITAL HEART DISEASE (CHD) DEFINITION It is the result of a structural or functional abnormality of the cardiovascular system at birth GENERAL FEATURES OF CHD Structural defects due to specific disturbance
Pathophysiology: Left To Right Shunts
 Pathophysiology: Left To Right Shunts Daphne T. Hsu, MD dh17@columbia.edu Learning Objectives Learn the relationships between pressure, blood flow, and resistance Review the transition from fetal to mature
Pathophysiology: Left To Right Shunts Daphne T. Hsu, MD dh17@columbia.edu Learning Objectives Learn the relationships between pressure, blood flow, and resistance Review the transition from fetal to mature
CELL PARTS TYPICAL ANIMAL CELL
 AP BIOLOGY CText Reference, Campbell v.8, Chapter 6 ACTIVITY1.12 NAME DATE HOUR CELL PARTS TYPICAL ANIMAL CELL ENDOMEMBRANE SYSTEM TYPICAL PLANT CELL QUESTIONS: 1. Write the name of the cell part in the
AP BIOLOGY CText Reference, Campbell v.8, Chapter 6 ACTIVITY1.12 NAME DATE HOUR CELL PARTS TYPICAL ANIMAL CELL ENDOMEMBRANE SYSTEM TYPICAL PLANT CELL QUESTIONS: 1. Write the name of the cell part in the
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Hyaline membrane disease. By : Dr. Ch Sarishma Peadiatric Pg
 Hyaline membrane disease By : Dr. Ch Sarishma Peadiatric Pg Also called Respiratory distress syndrome. It occurs primarily in premature infants; its incidence is inversely related to gestational age and
Hyaline membrane disease By : Dr. Ch Sarishma Peadiatric Pg Also called Respiratory distress syndrome. It occurs primarily in premature infants; its incidence is inversely related to gestational age and
Cell Communication. Chapter 11. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 11 Cell Communication PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 11 Cell Communication PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 15: Signal transduction
 Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
Chapter 26 Biochemistry 5th edition. phospholipids. Sphingolipids. Cholesterol. db=books&itool=toolbar
 http://www.ncbi.nlm.nih.gov/sites/entrez? db=books&itool=toolbar 1 The surface of a soap bubble is a bilayer formed by detergent molecules 2 Chapter 26 Biochemistry 5th edition phospholipids Sphingolipids
http://www.ncbi.nlm.nih.gov/sites/entrez? db=books&itool=toolbar 1 The surface of a soap bubble is a bilayer formed by detergent molecules 2 Chapter 26 Biochemistry 5th edition phospholipids Sphingolipids
Development: is the growth of an individual organism from a simple to a more complex or mature level. A slow process of progressive change
 1. Define the following terms (use your own words): development, growth, differentiation, histogenesis, organogenesis, morphogenesis, reproduction, tissue, organ, organ system, and organism. Development:
1. Define the following terms (use your own words): development, growth, differentiation, histogenesis, organogenesis, morphogenesis, reproduction, tissue, organ, organ system, and organism. Development:
PATENT DUCTUS ARTERIOSUS IN THE PRETERM INFANT EVIDENCE FOR & AGAINST TREATMENT
 PATENT DUCTUS ARTERIOSUS IN THE PRETERM INFANT EVIDENCE FOR & AGAINST TREATMENT Dr. Youssef Abou Zanouna, FRCPI, FACC Consultant Pediatric Cardiologist King Fahd Military Medical Complex Dhahran Introduction
PATENT DUCTUS ARTERIOSUS IN THE PRETERM INFANT EVIDENCE FOR & AGAINST TREATMENT Dr. Youssef Abou Zanouna, FRCPI, FACC Consultant Pediatric Cardiologist King Fahd Military Medical Complex Dhahran Introduction
PCTH 400. Endothelial dysfunction and cardiovascular diseases. Blood vessel LAST LECTURE. Endothelium. High blood pressure
 PCTH 400 LAST LECTURE Endothelial dysfunction and cardiovascular diseases. Classic Vascular pharmacology -chronic -systemic Local Vascular pharmacology -acute -targeted High blood pressure Blood pressure
PCTH 400 LAST LECTURE Endothelial dysfunction and cardiovascular diseases. Classic Vascular pharmacology -chronic -systemic Local Vascular pharmacology -acute -targeted High blood pressure Blood pressure
Update on mangement of patent ductus arteriosus in preterm infants. Dr. Trinh Thi Thu Ha
 Update on mangement of patent ductus arteriosus in preterm infants Dr. Trinh Thi Thu Ha Outline 1. Overview of PDA 2. Timing of screening PDA? 3. When to treat PDA? Timing of ductal closure Prenatal
Update on mangement of patent ductus arteriosus in preterm infants Dr. Trinh Thi Thu Ha Outline 1. Overview of PDA 2. Timing of screening PDA? 3. When to treat PDA? Timing of ductal closure Prenatal
Signal Transduction Cascades
 Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
OBJECTIVE. Lipids are largely hydrocarbon derivatives and thus represent
 Paper 4. Biomolecules and their interactions Module 20: Saturated and unsaturated fatty acids, Nomenclature of fatty acids and Essential and non-essential fatty acids OBJECTIVE The main aim of this module
Paper 4. Biomolecules and their interactions Module 20: Saturated and unsaturated fatty acids, Nomenclature of fatty acids and Essential and non-essential fatty acids OBJECTIVE The main aim of this module
CELLS. Cells. Basic unit of life (except virus)
") Basic unit of life (except virus) CELLS Prokaryotic, w/o nucleus, bacteria Eukaryotic, w/ nucleus Various cell types specialized for particular function. Differentiation. Over 200 human cell types 56%
Basic unit of life (except virus) CELLS Prokaryotic, w/o nucleus, bacteria Eukaryotic, w/ nucleus Various cell types specialized for particular function. Differentiation. Over 200 human cell types 56%
General Principles of Endocrine Physiology
 General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
NON STEROIDAL ANTI INFLAMMATORY NSAID
 NON STEROIDAL ANTI INFLAMMATORY DRUGS NSAID inflammation Normal, protective response to tissue injury 1-physical trauma 2-noxious chemical 3-microbial agent NSAIDs act by inhibiting the synth. Of prostaglandins
NON STEROIDAL ANTI INFLAMMATORY DRUGS NSAID inflammation Normal, protective response to tissue injury 1-physical trauma 2-noxious chemical 3-microbial agent NSAIDs act by inhibiting the synth. Of prostaglandins
Cellular Signaling Pathways. Signaling Overview
 Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling. How do cells receive and respond to signals from their surroundings?
 Cell signaling How do cells receive and respond to signals from their surroundings? Prokaryotes and unicellular eukaryotes are largely independent and autonomous. In multicellular organisms there is a
Cell signaling How do cells receive and respond to signals from their surroundings? Prokaryotes and unicellular eukaryotes are largely independent and autonomous. In multicellular organisms there is a
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1
 GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
Cell Communication. Local and Long Distance Signaling
 Cell Communication Cell to cell communication is essential for multicellular organisms Some universal mechanisms of cellular regulation providing more evidence for the evolutionary relatedness of all life
Cell Communication Cell to cell communication is essential for multicellular organisms Some universal mechanisms of cellular regulation providing more evidence for the evolutionary relatedness of all life
C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is
 is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is") ' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
Chapter 16: Endocrine System 1
 Ch 16 Endocrine System Bi 233 Endocrine system Endocrine System: Overview Body s second great controlling system Influences metabolic activities of cells by means of hormones Slow signaling Endocrine glands
Ch 16 Endocrine System Bi 233 Endocrine system Endocrine System: Overview Body s second great controlling system Influences metabolic activities of cells by means of hormones Slow signaling Endocrine glands
LOW DOSE ASPIRIN CARDIOVASCULAR DISEASE FOR PROPHYLAXIS OF FOR BACKGROUND USE ONLY NOT TO BE USED IN DETAILING
 LOW DOSE ASPIRIN FOR PROPHYLAXIS OF CARDIOVASCULAR DISEASE FOR BACKGROUND USE ONLY NOT TO BE USED IN DETAILING Use of Low Dose Aspirin to Treat and Prevent Cardiovascular Disease In recent decades, aspirin
LOW DOSE ASPIRIN FOR PROPHYLAXIS OF CARDIOVASCULAR DISEASE FOR BACKGROUND USE ONLY NOT TO BE USED IN DETAILING Use of Low Dose Aspirin to Treat and Prevent Cardiovascular Disease In recent decades, aspirin
Cell Cell
 Go to cellsalive.com. Select Interactive Cell Models: Plant and Animal. Fill in the information on Plant and Animal Organelles, then Click on Start the Animation Select Plant or Animal Cell below the box.
Go to cellsalive.com. Select Interactive Cell Models: Plant and Animal. Fill in the information on Plant and Animal Organelles, then Click on Start the Animation Select Plant or Animal Cell below the box.
Regulation of cell function by intracellular signaling
 Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
Short-Term Outcome Of Different Treatment Modalities Of Patent Ductus Arteriosus In Preterm Infants. Five Years Experiences In Qatar
 ISPUB.COM The Internet Journal of Cardiovascular Research Volume 7 Number 2 Short-Term Outcome Of Different Treatment Modalities Of Patent Ductus Arteriosus In Preterm Infants. Five Years Experiences In
ISPUB.COM The Internet Journal of Cardiovascular Research Volume 7 Number 2 Short-Term Outcome Of Different Treatment Modalities Of Patent Ductus Arteriosus In Preterm Infants. Five Years Experiences In
Attribution: University of Michigan Medical School, Department of Microbiology and Immunology
 Attribution: University of Michigan Medical School, Department of Microbiology and Immunology License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution
Attribution: University of Michigan Medical School, Department of Microbiology and Immunology License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Cell Quality Control. Peter Takizawa Department of Cell Biology
 Cell Quality Control Peter Takizawa Department of Cell Biology Cellular quality control reduces production of defective proteins. Cells have many quality control systems to ensure that cell does not build
Cell Quality Control Peter Takizawa Department of Cell Biology Cellular quality control reduces production of defective proteins. Cells have many quality control systems to ensure that cell does not build
The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae.
 Endoplasmic reticulum (ER) The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae. Cisternae serve as channels for the transport of
Endoplasmic reticulum (ER) The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae. Cisternae serve as channels for the transport of
Acid-base management during hypothermic CPB alpha-stat and ph-stat models of blood gas interpretation
 Acid-base management during hypothermic CPB alpha-stat and ph-stat models of blood gas interpretation Michael Kremke Department of Anaesthesiology and Intensive Care Aarhus University Hospital, Denmark
Acid-base management during hypothermic CPB alpha-stat and ph-stat models of blood gas interpretation Michael Kremke Department of Anaesthesiology and Intensive Care Aarhus University Hospital, Denmark
- Biosignaling: Signal transduction. References: chapter 8 of Lippincots chapter 1 3 of Lehningers
 Basic concepts of Metabolism Metabolism and metabolic pathway Metabolic Map Catabolism Anabolism - Regulation of Metabolism Signals from within the cell (Intracellular) Communication between cells. - Biosignaling:
Basic concepts of Metabolism Metabolism and metabolic pathway Metabolic Map Catabolism Anabolism - Regulation of Metabolism Signals from within the cell (Intracellular) Communication between cells. - Biosignaling:
The Physiology of the Fetal Cardiovascular System
 The Physiology of the Fetal Cardiovascular System Jeff Vergales, MD, MS Department of Pediatrics Division of Pediatric Cardiology jvergales@virginia.edu Disclosures I serve as the medical director for
The Physiology of the Fetal Cardiovascular System Jeff Vergales, MD, MS Department of Pediatrics Division of Pediatric Cardiology jvergales@virginia.edu Disclosures I serve as the medical director for
Cell Communication. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 11 Cell Communication PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 11 Cell Communication PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Introduction to Emergency Medical Care 1
 Introduction to Emergency Medical Care 1 OBJECTIVES 6.1 Define key terms introduced in this chapter. Slides 11, 15, 17, 26, 27, 31, 33, 37, 40 42, 44, 45, 51, 58 6.2 Describe the basic roles and structures
Introduction to Emergency Medical Care 1 OBJECTIVES 6.1 Define key terms introduced in this chapter. Slides 11, 15, 17, 26, 27, 31, 33, 37, 40 42, 44, 45, 51, 58 6.2 Describe the basic roles and structures
Full file at
 1. This is defined as fully differentiated body part with specialized functions: A) cell B) organ C) tissue D) system 2. Which of the following is characteristic of the plasma membrane? A) contains a single
1. This is defined as fully differentiated body part with specialized functions: A) cell B) organ C) tissue D) system 2. Which of the following is characteristic of the plasma membrane? A) contains a single
numbe r Done by Corrected Docto Alia Shatnawi
 numbe r 9 Done by Nazek Hyasat Corrected Bahaa Najjar & mohammed AL-shrouf Docto Alia Shatnawi HOW DO DRUGS WORK??? You know that receptor targets by the drugs, the question now how these drugs work on
numbe r 9 Done by Nazek Hyasat Corrected Bahaa Najjar & mohammed AL-shrouf Docto Alia Shatnawi HOW DO DRUGS WORK??? You know that receptor targets by the drugs, the question now how these drugs work on
Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research
 Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research Shoude Jin Harbin Medical University, China Background COPD ------ a silent killer Insidious,
Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research Shoude Jin Harbin Medical University, China Background COPD ------ a silent killer Insidious,
Chapter 10. Regulatory Strategy
 Chapter 10 Regulatory Strategy Regulation of enzymatic activity: 1. Allosteric Control. Allosteric proteins have a regulatory site(s) and multiple functional sites Activity of proteins is regulated by
Chapter 10 Regulatory Strategy Regulation of enzymatic activity: 1. Allosteric Control. Allosteric proteins have a regulatory site(s) and multiple functional sites Activity of proteins is regulated by
Chapter 11. Cell Communication
 Chapter 11 Cell Communication Overview: The Cellular Internet Cell-to-cell communication Is absolutely essential for multicellular organisms Concept 11.1: External signals are converted into responses
Chapter 11 Cell Communication Overview: The Cellular Internet Cell-to-cell communication Is absolutely essential for multicellular organisms Concept 11.1: External signals are converted into responses
CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY. Ms. K. GOWRI. M.Pharm., Lecturer.
 CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY Ms. K. GOWRI. M.Pharm., Lecturer. Structure of cell: Human body develops from a single cell zygote which results from fusion of the ovum andd the spermatozoan.
CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY Ms. K. GOWRI. M.Pharm., Lecturer. Structure of cell: Human body develops from a single cell zygote which results from fusion of the ovum andd the spermatozoan.
Receptors Functions and Signal Transduction- L4- L5
 Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Human Anatomy and Physiology - Problem Drill 16: The Endocrine System
 Human Anatomy and Physiology - Problem Drill 16: The Endocrine System Question No. 1 of 10 The endocrine system is made up of a number of organs and glands. Which one of the following is not an organ or
Human Anatomy and Physiology - Problem Drill 16: The Endocrine System Question No. 1 of 10 The endocrine system is made up of a number of organs and glands. Which one of the following is not an organ or