FACE REPRESENTATION IN THE HUMAN PRIMARY AND SECONDARY SOMATOSENSORY CORTEX. Binh Thi Nguyen DOCTOR OF PHILOSOPHY
|
|
|
- Everett Wade
- 5 years ago
- Views:
Transcription
1 FACE REPRESENTATION IN THE HUMAN PRIMARY AND SECONDARY SOMATOSENSORY CORTEX Binh Thi Nguyen DOCTOR OF PHILOSOPHY Department of Physiological Sciences, School of Life Sciences, The Graduate University for Advanced Studies 2005
2 Contents 1 Abbreviations 3 2 Abstract 4 3 Introduction 6 4 Methods Experiment Subjects Stimulation Stimulus area MEG recording Data analysis Statistical analysis Experiment Subject Stimulation MEG recording Data analysis 15 5 Results Face representation in the human primary somatosensory cortex (SI) Waveform Latency and dipole moment Source localization Face representation in the human secondary somatosensory cortex (SII) Waveform Latency and dipole moment Source localization 21 6 Discussion Face representation in the human primary somatosensory cortex (SI) 24 1
3 6.2 Face representation in the human secondary somatosensory cortex (SII) 27 7 General discussion and conclusion 32 8 Acknowledgements 35 9 References Tables Figure Legends Figures Publications 61 2
4 1. Abbreviations ECD: Equivalent current dipole fmri: Functional magnetic resonance imaging GOF: Goodness of fit MEG: Magnetoencephalography PET: Positron emission tomography SI: Primary somatosensory cortex SII: Secondary somatosensory cortex SEF: Somatosensory evoked magnetic fields 3
5 2. Abstract To investigate the somatotopic organization of the facial skin area in the primary and secondary somatosensory cortex (SI and SII), we recorded magnetic fields evoked by air pressure induced tactile stimulation applied to the face areas. For the SI study, we stimulated six points on the face (two points in each part of forehead, cheek and chin), lower lip and thumb. The dipole locations for SI activities were compared among eight stimulation points. The thumb area in SI was located more superior and medial to the lip area, which was consistent with Penfield s homunculus. The face area was located between the thumb and the lip. There was no somatotopic arrangement among three parts of the face as forehead, cheek and chin (represent for three branches of the trigeminal nerve). The lips occupy a large area of the face representation in SI, whereas only a small area located between the thumb and lip areas is devoted to skin covered surfaces. For the SII study, somatosensory evoked magnetic fields were recorded following stimulation of five body sites: the foot, lip and three facial skin points (forehead, cheek and mandibular angle point). We focused on the activities in SII and compared the dipole locations among them. There was a clear somatotopic organization in SII with the lip in the most lateral area, the foot in the most medial area and the face in the 4
6 intermediate area between the lip and the foot, closed to the lip area. However, there was no significant difference of dipole localization in SII among the three areas of the facial skin. The facial skin areas are considered to occupy a small area in SII with insufficient spatial separation to differentiate each area of the facial skin. 5
7 3. Introduction The somatosensory system mediates the processing of cutaneous, kinesthetic, visceral and noxious information; mainly consists of primary somatosensory cortex (SI), secondary somatosensory cortex (SII) and posterior parietal cortex (PPC). SI is located in the postcentral gyrus, beginning at the bottom of the central sulcus and extending posteriorly to the postcentral and intraparitetal sulci. SI receives its main inputs from the sensory thalamus and has a role in the initial process of tactual functions such as encoding the type and intensity of the sensory inputs. There are four distinct areas in SI: Brodmann s areas 3a, 3b, 1 and 2, each of which has a complete body surface map (Kaas et al., 1979). Tactile signals are processed serially through areas 3, 1 and 2. Area 3b is the first step of tactile processing for information from the skin, while area 3a is the first step for information mainly from the deep tissue (Iwamura., 1998). The cortical organization of neurons in SI was first described by Penfield and Boldrey (1937), since then many studies have clarified the somatotopic representation of the body surface in SI (Baumgartner et al., 1992; Hari et al., 1993; Kakigi et al., 1995; Hoshiyama et al., 1995, 1996; Nakamura et al., 1998; Nihashi et al., 2003). Regarding the representation of the face in SI, the face area drawn by Penfield and Boldrey (1937) is organized along 6
8 the central sulcus with the forehead in the superior-medial region adjacent to the hand area, and the chin in the inferio-lateral region. Although there are some studies reporting the locations in SI of the lip (Nakamura et al., 1998; Hoshiyama et al., 1995; Mogilner et al., 1994), tongue (Karhu et al., 1991; Altenmuller et al., 1990; Nakamura et al., 1998), oral cavity (Hari et al., 1993), and ear (Nihashi et al., 2001, 2002, 2003), there are only a few reports (Servos et al., 1999; Yang et al., 1994) on skin covered areas of the face such as the forehead, cheek and chin in humans. Interestingly, some results showed the representation of an inverted face along the central sulcus of the human brain (Servos et al., 1999; Yang et al., 1994; Pons et al., 1991), which is not consistent with the homunculus map drawn by Penfield and Boldrey (1937). In the first study, we sought to determine the topography of the whole face in SI using magnetoencephalography (MEG). MEG with a good spatial and temporal resolution is suitable to study cortical activities whose orientation is tangential to the brain surface. Since the main activities in the postcentral gyrus following somatosensory stimulation occur in the posterior wall of the central sulcus in area 3b (Kakigi et al., 2000) therefore create a tangential dipole, and MEG is suited to our purpose. The SII area is located on the upper bank of the sylvian fissure. SII is involved in complex tactile functions and processes high-order features of the stimulus such as 7
9 attention (Burton et al, 1997; Mima et al, 1997), learning (Diamond et al, 2002), and memory (Diamond et al, 2002; Ridley and Ettinger, 1976). SII receives main inputs from area 3b. Like SI, SII has a somatotopic representation of the body. Unitary recordings in monkeys have revealed several distinct somatotopic maps of the body surface in the SII region (Burton et al., 1995; Friedman et al., 1980; Krubitzer et al., 1995; Robinson and Burton., 1980; Whitsel et al., 1969). These maps have a medio-lateral organization in which the face is represented laterally near the lip, followed in a latero-medial sequence for hand and foot representations, respectively. In humans, a somatotopic organization in the SII region has been identified by direct cortical stimulation (Penfield and Ramussen, 1950; Woolsey et al., 1979), MEG (Hari et al., 1993; Maeda et al., 1999) and functional magnetic resonance imaging (fmri) (Del Gratta et al., 2000; Disbrow et al., 2000; Ruben et al., 2001; Iannetti et al., 2003) studies. For example, the face, hand, trunk and foot are represented in a latero-medial sequence in SII in both monkeys and humans, though the sequence is not very clear compared with that in SI, the so-called homunculus (Penfield and Ramussen, 1950). However, a map in SII of the facial skin area has not been investigated in detail. In the second study, we sought to clarify the somatotopic organization of the face in SII using MEG. 8
10 4. Methods 4.1. Experiment 1: Subjects Eleven healthy subjects, including five females and six males, participated in this study. Their age ranged from 27 to 35 (mean ± S.D: 30.0± 4.0) years. The study was approved in advance by the Ethical Committee of our Institute (The National Institute for Physiological Sciences), and informed consent was obtained from all the subjects Stimulation A sensory output device, which consisted of a small balloon (1 cm in diameter) attached to the stimulation site, was used for stimulation (Figure. 1). The air pressure needed to inflate the small balloon was 0.15 MPa and induced a mechanical stimulation of 750 g/cm2 at the stimulated site. A clear tactile sensation was elicited by the air pressure without a pain sensation. The air pressure, stimulus duration (50 ms), and rate 9
11 of the stimulus (2 Hz or ISI 0.5 s) were controlled by a tactile stimulator system (Nihon Kohden, Tokyo, Japan) Stimulus area We stimulated the glabrous side of the right thumb about 2 cm from the tip, the middle part of the lower lip, and six points (two points each on the forehead, cheek and chin) on the right side of the face. We defined the six points on the face as follows (see Figure. 1): (1, forehead 1) 1 cm lateral to the middle line and 3 cm superior to the eyebrow; (2, forehead 2) lateral point of the forehead region, 4 cm lateral to forehead 1; (3, nose) the top of the nose; (4, cheek) the top of the cheek-bone point; (5, chin 1) 2 cm inferior to the lower lip on the middle line; and (6, chin 2) mandibular angle point MEG recordings Somatosensory-evoked magnetic fields (SEFs) were recorded with a 204-channel helmet-shaped planar neuromagnetometer covering the whole head (Vectorview, ELEKTA Neuromag Yo, Helsinki, Finland) (Figure. 2), which had 102 pairs of planar 10
12 gradiometers oriented orthogonally. The planar gradiometers detect the largest signal just above the source current (Hämäläinen et al., 1993). The signals were recorded with a Hz bandpass filter and digitized at a sampling rate of 900 Hz. The period of analysis was from 50 ms before to 350 ms after the stimulation. The pre-stimulus period was used as the DC baseline. In each session, two points were randomly stimulated. Four sessions were conducted for each subject, three sessions for two points of each region (forehead, cheek and chin) of the face, and one session for the thumb and lip. The order of the sessions was randomized across subjects. For each stimulated point, 200 responses were averaged. White noise was simultaneously delivered during the period of stimulation at a sufficient level to mask the sound made by the air pressure stimulator Data analysis By single equivalent current dipole (ECD) analysis, we estimated a dipole at the peak latency of a constant component around ms following the stimulation. To select the ECD source, we first identified 3 4 channels, which detected large responses around the rolandic region (Hari and Imada, 1999), then selected channels around 11
13 those channels with the large responses. This method has been used previously (Avikainen et al., 2002; Forss et al., 1999; Forss and Jousmäki, 1998). The subset of channels was fixed for each subject. The ECDs were estimated using the signals from these channels. The peak latency was defined by selected one channel among the subset, the channel with the biggest response amplitude. The latency point of the waveform of this channel with the biggest amplitude was defined as the peak latency. The goodness of-fit was calculated. Goodness-of-fit here refers to how well the model signal explains the original signal. In the present study, GOF was defined as: g (%)= [1- n i= 1 2 b bˆ ) / ( i i n b i= 1 2 i ] x 100 where b1,..., bn are the experimental data and ˆb1,...,ˆbn are the value given by the equivalent dipole. Only the ECDs with a GOF over 85% (Avikainen et al., 2002; Forss et al., 1999; Forss and Jousmäki, 1998) were used for further analysis. The locations of dipoles were expressed with x, y and z coordinates where the x-axis indicates the mid-sagittal plane with positive values toward the right preauricular point, the y-axis indicates the coronal plane with positive values in the anterior direction, and the z-axis lies on the plane perpendicular to the x y plane, with positive values toward the upper side (see Figure 3). The 3-D location, orientation and strength of the ECD were superimposed on MR images of each subject with the head assumed to be a sphere. 12
14 For a comparison of the ECD location among the eight stimulus points, we employed a standardized coordinate to reduce the inter-individual difference in head shape. For this purpose, we measured the maximum brain length from the y z plane (x-axis), x z plane (y-axis) and x y plane (z-axis), and on MR images of each subject. The mean value for each axis among seven subjects was calculated and used as a standard. Then, the original x, y and z coordinates for ECDs of each subject were corrected using the ratio of the subject s maximum brain length to the standard. For example, if the mean maximum length of the y axis (standard) was 100 mm, and the maximum length and original ECD value of the y-axis for subject A were 120 and 20 mm, respectively, the corrected y value of subject A was 20*100/120 = 16.7 mm. The data were expressed as the mean ± standard deviation (SD) Statistical analysis The differences in latency, dipole moment and ECD location (x, y and z values) among the eight stimulated points were assessed with a one-way analysis of variance (ANOVA). The level of statistical significance difference was set at 5% (P < 0.05). 13
15 4.2. Experiment 2 The methods of the Experiment 2 were almost the same as Experiment 1; the different points of Experiment 2 will be reported below Subject Ten healthy subjects, 3 females and 7 males, participated in this study. Their age were ranged from 25 to 35 (mean 28.4 years) Stimulation We used the method similar to the Experiment 1. Interstimulus interval (ISI, rate of stimulus) was random from 2-4 s for this study, since the SII response is recorded with a long ISI. We stimulated the dorsal part of the right foot, lateral right part of the upper lip and 3 right facial skin points (By using a long ISI, it took a long time for recording. Therefore we stimulated only three facial skin points in this experiment); (1) the forehead, 3 cm lateral to the mid-line and 3 cm superior to the interior line of the 14
16 eyebrow; (2) the cheek, the top of the cheek-bone point; and (3) the mandibular angle point MEG recordings The period of analysis was from 50 ms before and 250ms after the stimulation. The prestimulus period was used as the DC baseline. Two sessions were conducted for each subject, one for the foot and lip and another for the 3 facial skin points. The order of the sessions was randomized across subjects. During the recording, subjects were asked to count the number of all stimulations. Artifacts caused by blinking and large eye movements were discarded in the off-line analysis by recording electrooculograms. Sequentially, responses were averaged for each stimulated point Data analysis Since the field distribution of the SII activity analyzed in the present study was clearly different from that of the SI activity (Experiment 1), we considered that contribution of SI activity to evoked magnetic fields in these channels was minimum at 15
17 the peak latency of the SII activity. Therefore, we used a single dipole model to obtain ECDs in the SII area. 16
18 5. Results 5.1. Face representation in the human primary somatosensory cortex (SI) Among 11 subjects tested, clear responses to the face stimulation could not be recorded in four, probably due to a small signal-to-noise ratio. Therefore, only results from seven subjects were used in the analyses Waveform Although there were some earlier components, they were small in amplitude and rarely showed GOF values larger than 85%. Therefore, we chose the first consistent component for the analyses. This main deflection, occurring at a peak latency of approximately ms, was termed M45, and had a source orientation toward the posterior regardless of the stimulus point (Figure. 4, Tables 1 and 2) Latency and dipole moment 17
19 An ANOVA showed that the latency of M45 component did not differ among the eight stimulus points (F= 0.42, P= 0.89), but the source strength was significantly different (F= 4.12, P= ). The source strength tended to be larger for the thumb (23.8 nam) or lip (23.0) than for the six facial points ( ) Source localization As shown in Figure. 5, the ECD for the thumb was located in the posterior wall of the central sulcus, and was medial (mean: 9.1 mm), posterior (mean: 9 mm) and superior (mean: 11.8 mm) to the ECD for lip. Results of an ANOVA for the ECD location showed no significant difference among the eight stimulus points (P = 0.07, 0.43, and 0.3 for the x, y and z coordinate, respectively). Most of the ECDs for the six points on the face were located anterior, lateral and inferior to the ECD for thumb, and posterior, medial and superior to the ECD for lip (Figures. 5 and 6). This meant that their locations were between the thumb and lip areas. To investigate the arrangement of representations in SI of three trigeminal branch areas, the six facial points were grouped into three subgroups (V1, V2 and V3), and the ECD location was compared among five groups (V1, V2, V3, thumb and lower lip) with a 18
20 one-way ANOVA. However, we could not find any differences (P = 0.16, 0.2, and 0.11 for the x, y and z coordinate, respectively). Next, the six facial points were grouped into medial (points 1, 3, 5) and lateral (points 2, 4, 6) areas, and the ECD location was compared among four groups (medial facial area, lateral facial area, thumb and lower lip). The results of an ANOVA showed a significant difference in the x coordinate among the four groups (P = 0.02, F = 3.6). The ECD for the thumb, the lateral facial area (3.7 mm lateral to the thumb), the medial facial area (6.7 mm lateral to the thumb), and the lip (9 mm lateral to the thumb) were arranged in this order laterally in the SI Face representation in the human secondary somatosensory cortex (SII) All 10 subjects showed a clear middle-latency component, which was considered to be generated in SII. However, only results of six out of 10 subjects met our criteria for dipole localization, and were used for further analysis Waveform Figure 8 shows the magnetic response of two subjects to the air pressure stimuli 19
21 applied to the foot, lip and three facial skin points. An early response appeared at around 40 ms following stimulation (Figure 7), were consistent with ECDs generated in SI considering the dipole location and antero-posterior direction, which were analyzed in detail in experiment 1 (Experiment 1). At a later latency around ms, responses appeared in the bilateral hemispheres, whose ECDs were located more inferiorly than SI responses and showed a vertical orientation. A single-dipole analysis indicated that the major contributors for early and late responses were a source in the contralateral SI and sources in the upper bank of the Sylvian fissure of both hemispheres corresponding to SII, respectively. Since SII responses recorded in the hemisphere ipsilateral to the stimulation were small in amplitude with a low signal-to-noise ratio in two subjects, we focused on the SII response recorded from the hemisphere contralateral to the stimulation in the present study Latency and dipole moment The peak latency was significantly different among the five stimulation points by a one-way ANOVA (F=5.6, P=0.0025). The peak latency for foot stimulation was longer than those following stimulation of the lip and three facial skin points (Table 3 and 20
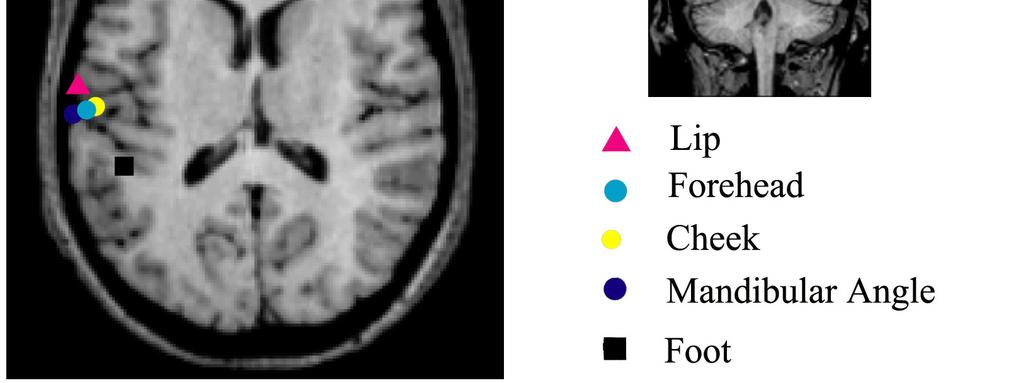
22 Figure. 8). The dipole moment of ECDs showed no significant difference among five stimulus conditions (P=0.47, Table 3). In general, all ECDs directed toward superiorly. The orientation of the ECD was not significantly different among five stimulation points (P=0.14, 0.93 and 0.9 for x, y and z axis vector) Source localization Figure 9 shows the location of ECDs following stimulation of the lip, foot and three facial skin points superimposed on the MR images in one subject. In this case, the ECDs for the foot and lip were located most medially and laterally in SII, respectively, and the ECDs for the facial skin area were located in an intermediate area close to the ECDs generated following stimulation of the lip. Table 4 shows the mean coordinates of each ECD. Figure 10 is a schematic drawing of spatial relationships of ECDs in SII following each stimulus indicating of the coordinates mean and one standard deviation of all subjects. To certify the difference in localization of stimulus points in SII, first we compared the locations of ECDs among the five stimulus points using an ANOVA. The results showed a significant different of medio-lateral coordinate among five stimulation 21
23 groups (F=3.165, P <0.05) with the foot is most medial and the lip is most lateral, the forehead and cheek is also lateral between lip and foot (P=0.003, 0.014, and 0.037, respectively). However, there were no significant difference of the anterior posterior and inferior superior (P=0.64 and 0.92 for y and z values, respectively). As a second step, ECDs for the forehead, cheek and mandibular angle were grouped as facial skin area, since they were located very close to each other. We compared the ECD locations among 3 stimulation groups, the lip, foot and facial skin area using an ANOVA to determine whether there was actually a somatotopic organization of the body surface in the SII region. The results showed that the medio-lateral coordinate was significantly different among these 3 groups (F=5.83, P<0.05). The location of the ECD was most medial for foot (-42.7 mm), most lateral for lip (-54.9), and intermediate between foot and lip for facial skin area (-51.1). Although the ECD location for the facial skin area was also located between the lip and foot both for antero-posterior and infero-superior directions like for the x coordinate but the difference was not significant (P=0.41 and 0.71, respectively). Subsequently we tried to find an orderly arrangement in SII among the 3 facial skin areas, the forehead (V1 territory of the trigeminal nerve), cheek (V2) and mandible angle point (V3) using a one-way ANOVA. The results showed no difference among 3 22
24 stimulation points (P=0.6, 0.9 and 0.52 for x, y and z coordinates, respectively). 23
25 6. Discussion 6.1. Face representation in the human primary somatosensory cortex The study showed that the thumb s location in SI was more superior, medial and posterior than the lower lip s location along the central sulcus, which is consistent with the homunculus drawn by Penfield and Boldrey (1937) and other studies in humans using electrocorticography (McCarthy et al., 1993; Baumgartner et al., 1992) and MEG (Nakamura et al., 1998; Hoshiyama et al., 1996, 1995; Hari et al., 1993) as well as studies in monkeys (Nelson et al., 1998; McCarthy and Allison, 1995; Carlson et al., 1986). With regard to the spatial relationship between the thumb and lip areas in the SI, the ECDs for the facial points were located more inferior and lateral than the ECD for the thumb, and in turn, the lip area was located inferior and lateral to the six facial points, suggesting that the skin-covered areas of the face are represented between the thumb and lip in the SI, which is consistent with results of a single unit study in monkeys (Nelson et al., 1998). However, we could not find a consistent spatial arrangement of the six facial points in SI except that the lateral parts of the facial skin tended to be represented more medial than the medial parts. These results were 24
26 consistent with those of intracranial SEP studies by McCarthy and colleagues in humans (McCarthy et al., 1993) and monkeys (McCarthy and Allison, 1995), demonstrating that only a small region of SI between the thumb and upper lip areas was devoted to the upper face. Pons et al. (1991) also found in monkeys, that the cortical representations of chin and jaw were located adjacent to the hand representation. We considered that the reason we could not clearly separate the six points on the face was that the thumb and lip positions occupied a large area of the SI, and there was a relatively short distance between them. Since the thumb has an important role in prehensile function, its area in the cortex is extensive, and the facial area of SI is mostly occupied by the representation for lips as the lips have very sensitive particularity, a great mechanoreceptor density (Stöhr and Petruch, 1979), play roles in speaking and eating, and a very important part of tactile sensation. In contrast, the tactile function of the face is more modest. Therefore, it was reasonable to conclude that the remaining facial area devoted to facial skin was small. The present results showing that the source strength was significantly greater for the lip and thumb than for the six facial points are consistent with this notion. The result, however, should be interpreted with caution, because it cannot be excluded that the spatial separation of sources in the distal-proximal somatotopy was beyond the resolving capacity of MEG, or due to a large inter-individual difference. Of course, one 25
27 can clearly recognize which part of the face is stimulated in daily life. Therefore, we consider that somatotopy of the face must be present in SI, but we could not significantly separate them in the present MEG study. A relatively large inter-individual difference might be another reason for the lack of statistical significance. In monkeys, the representation of the face in the somatosensory cortex is upside down, that is, the forehead representation is relatively inferior to the chin representation, and the cortical representations of chin and jaw are located adjacent to the hand representation (Pons et al., 1991), suggesting an inverted representation of the face in SI. Similar findings were reported in an fmri study in humans (Servos et al., 1999). In an MEG study of two human subjects, Yang and colleagues (Yang et al., 1994) reported that ECDs for lower facial areas in the SI were located closer to those for fingers as compared to the ECDs for upper facial regions. In this study, we stimulated six points in three areas and as described above the locations of each area in the SI were not differentiated. In monkeys, representations of medial parts of the face were located relatively posterior to those of lateral parts of the face in the SI (Nelson et al., 1998; Carlson et al., 1986; Cusick et al., 1986). In the present study, the results showed that the ECD for the medial parts of the face is located lateral to that for the lateral parts of the face. The 26
28 different pattern of medial and lateral representation of the face in the SI between humans and monkeys may result from a species difference. However, both results suggested segregated representations in the SI of the medial and lateral parts of the face. In conclusion, our results implied that the lips occupy a large area of the facial representation in the SI, whereas only a small area located between the thumb and lips areas is devoted to the skin-covered surface of the face. This is the first study showing that the facial skin areas in the SI are located between the thumb and lower lip areas and close to each other in humans Face representation in the human secondary somatosensory cortex The responses from the SII area were successfully recorded following stimulation of the facial skin points, although results in some subjects did not meet our criteria for a reliable ECD location due to a small S/N ratio, since mechanical air-puff stimulation is not sharp and strong with poor time-locking compared to electrical stimulation. The magnetic response from the SII region could be clearly differentiated from the response originating from SI by the response latency and ECD orientation. As compared with the SI response peaking around 40 ms, the SII activity had much longer response latency, 27
29 which was consistent with previous studies (Hoshiyama et al., 1996; Kakigi et al., 2000; Maeda et al., 1999). The ECD for the SII response oriented upward, which was clearly different from the ECD for SI activity with an anterior posterior orientation (Results session of Experiment 1). In addition, the SII response was found in the bilateral hemispheres, which was in line with previous SII studies using MEG (Forss and Jousmaki, 1998; Hari et al., 1993; Hoshiyama et al., 1996; Inui et al., 2003; Kakigi et al., 1995; Maeda et al., 1999; Makela et al., 2003). The location of ECDs for SII is more lateral, inferior and posterior than that for SI (Results session of Experiment 1). The significantly different medio-lateral locations of the ECDs for foot, lip and facial skin stimulations in the present study indicated a somatotopical organization within SII, confirming previous findings in humans (Disbrow et al., 2000; Maeda et al., 1999). Usually, the foot area of SII is located at the most medial site and the lip and face area have been reported to be located on the lateral side (Disbrow et al., 2000; Maeda et al., 1999). A similar order of somatotopic organization in SII has been identified in animals, for example, the foot area is located most caudal, near the fundus of the lateral sulcus then the hand, trunk, and face are located rostrally, respectively (Burton and Carlson, 1986; Cusick et al., 1989; Krubitzer et al., 1986). Although the difference did not reach the significant level, the present results also implied the existence of an 28
30 antero-posterior and infero-superior arrangement of the body surface in SII. The present results showed that the lip was located most laterally, anteriorly and inferior, the foot was located most medially, posteriorly and superiorly and the facial skin points were located in intermediate regions close to that for lip stimulation. Such arrangements have been reported in both humans (Del Gratta et al., 2000; Disbrow et al., 2000; Hari et al., 1993; Maeda et al., 1999; Ruben et al., 2001) and monkeys (Burton and Carlson, 1986; Cusick et al., 1989; Krubitzer et al., 1986). Although we confirmed the somatotopic arrangement in the SII region, the present study failed to find differences in location in SII of three different facial skin points. This finding was consistent with a previous fmri study (Iannetti et al., 2003), in which activations in the SII region following stimulations of V1 and V3 regions overlapped each other. As reported in Experiment 1, the facial skin areas in SI were just between the thumb and lip areas and overlapped in a small region with no significant difference in location. The present findings in SII were consistent with those made previously in SI. That should explain why we could localize the face area in SI and SII but could not separate each part of the face. Though the lip is a part of the face but it s function is more various compares with the facial skin as lip has an important role in speaking, sensation, eating and there are a large number of neurons responding to lip stimulation 29
31 or lip occupies large area in both SII and SI (McCarthy et al., 1993) Unitary recording studies in monkeys reported that neurons in SII have large, contralateral or bilateral receptive fields though some neurons do not have bilateral or ipsilateral receptive fields (Cusick and Manning, 1998; Cusick et al., 1989; Krubitzer et al., 1986; Pons et al., 1987; Robinson and Burton, 1980). Robinson and Burton (1980) reported that in monkeys, only one-third of SII neurons have bilateral or ipsilateral receptive fields. In humans, Hari et al. (1993) reported that the SEF field patterns of the ipsilateral response were not stable or changeable. This might explain why we could not record a prominent ipsilateral response at all stimulated points of some subjects. The SII location in this study is approximately consistent with those in previous studies that used active discriminative tasks (Ledberg et al., 1995; Roland et al., 1998), while apparently more lateral to those reported in studies that used passive stimulation (Burton et al., 1993, Frot and Mauguière, 1999). Since subjects were instructed to count the number of the stimulus in this study, the present results appear to confirm that attention or discriminative tasks affect the location of active areas around Sylvian fissure (see discussions of Frot and Mauguière, 1999). These results imply that a deeper source other than SII is active simultaneously, probably the insula. Previous studies using intracranial recordings (Frot and Mauguière, 1999), PET (Ledberg et al., 1995; 30
32 Roland et al., 1998) and MEG (Inui et al., 2003) in humans as well as unitary recording studies in monkeys (Augustine., 1996; Burton et al., 1993; Friedman et al., 1980) showed that the insula is involved in tactile processing. In this study, the location of the foot response in two subjects was quite medially (x value) and the latency for foot was relatively longer compared to those of facial skin points. Therefore, it seems possible that the activity in the insula contributed to form the magnetic fields around Sylvian fissure in such cases. In conclusion, our MEG studies indicated that neurons responding to somatosensory stimulation applied to facial skin areas in SII as well as SI were located very close to each other but clearly separated from those responding to stimulation of the lip. This finding should be redrawn after more thorough electrophysiological and neuroimaging studies. 31
33 7. General discussion and conclusion The facial skin areas are represented in SI in a region between the thumb and the lip areas, and are represented in SII in a region between the lip and the foot areas closed to the lip area. Therefore, we could confirm somatotopic arrangements of the body surface including the face both in SI and SII by use of MEG. In SI, the medial part of the face is located more lateral than the lateral part with no somatotopic arrangement among the three branches of the trigeminal nerve, indicating that distinct segmental areas of the face have distinct representations in SI. On the other hand, we could not find any clear arrangement of the facial skin areas in SII, suggesting that the face regions in SII are located close to each other, probably overlapped in part. The results are consistent with the clearer somatotopy in SI than in SII in both monkeys (Robinson and Burton 1980; Sinclair and Burton 1993) and humans (Hari et al., 1993; Ruben et al., 2001; Iannetti et al., 2003). Although we found a statistically significant difference in location for facial skin areas in SI, it was sometimes difficult to find a clear spatial arrangement in a single subject as compared with the very clear separation between the lip and thumb areas in SI. Since the thumb and lip have important roles in daily sensory discrimination and 32
34 speaking, respectively, they should occupy large areas in SI. By contrast, the facial skin areas occupy only a small region in SI, probably reflecting that the tactile function of the face is more modest as compared with the lip and thumb. Therefore, we consider that the spatial resolution of MEG is not high enough for detecting the precise arrangement of the facial skin areas in a very small region. In addition, a unitary recording study in monkeys (Manger et al., 1995) showed that the cortical fields responding to stimulation of different face areas have an irregular shape and lie intermingled in SI. Given similar properties for human SI neurons responding to the face stimulation, it would be expected to be very difficult to find a clear somatotopic arrangement in humans in a noninvasive study. Neurons in SII have complex and large receptive fields reflecting its higher level in tactile processing than SI. As a result, the somatotopic organization in SII is less clear than that in SI though many previous studies in animals (Burton et al., 1995; Friedman et al., 1980; Krubitzer et al., 1995; Robinson and Burton., 1980) and humans (Hari et al., 1993; Maeda et al., 1999; Del Gratta et al., 2000; Disbrow et al., 2000; Ruben et al., 2001; Iannetti et al., 2003) found a rough somatotopy in this region. Given that the facial skin area is represented in a small region in SII like in SI, our failure to find a clear somatotopic organization in SII for the facial skin areas seems reasonable. 33
35 In conclusion, we could localize the face area both in SI and SII using MEG. Neurons responding to somatosensory stimulation applied to facial skin areas in SI as well as in SII were located very close to each other but clearly separated from those responding to stimulation of the lip. From a clinical interest, localization of the body in the cortex could be useful in surgical procedures. 34
36 7. Acknowledgement I would like to express my deep gratitude to my professor, professor Ryusuke Kakigi for his supporting during the time I ve been studied in the Department of Integrative Physiology, National Institute for Physiological sciences in Okazaki, for his encouragement in my studying, for giving me the opportunity so I could come to Japan for studying. I wish to express my sincerely thankful to Dr. Koji Inui and Dr. Minoru Hoshiyama for their guidance and advice in creating the experiment; analysing the data and their invaluable contributed suggestion to writing this thesis. Without their helps, this thesis would not be accomplished. I am grateful to Dr. Hiroki Nakata, Dr. Tuan Diep Tran for their comments and suggestions during the data processing. My thanks are also extended to Mr. Yasuyuki Takeshima and Mr. Osamu Nagata for their technical assistant. Finally, I am greatly appreciating the kindness of all members in the Department of Integrative Physiology, National Institute for Physiological Sciences for their assistant during my time in Japan. 35
37 8. References Altenmuller E, Cornelius CP, Buettner UW. Somatosensory evoked potentials following tongue stimulation in normal subjects and patients with lesions of the afferent trigeminal system. Electroencephalogr. Clin. Neurophysiol 1990; 77: Augustine JR. Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res Rev 1996; 22: Avikainen S, Forss N, Hari R. Modulated activation of the human SI and SII cortices during observation of hand actions. Neuroimage 2002; 15: Baumgartner C, Barth D, Levesque M, Sutherling W. Human hand and lip sensorimotor cortex as studied on electrocorticography. Electroencephalogr. Clin. Neurophysiol 1992; 84: Burton H, Carlson M. Second somatic sensory cortical area (SII) in a prosimian primate Galago crassicaudatus. J Comp Neurol 1986;274: Burton H, Videen TO, Raichle ME. Tactile-vibration-activated foci in insular and parietal opercular cortex studied with positron emission tomography: mapping the second somatosensory area in humans. Somatosens Mot Res 1993;10: Burton H, Fabri M, Alloway K. Cortical areas within the lateral sulcus connected to cutaneous representations in areas 3b and 1: a revised interpretation of the second somatosensory area in macaque monkeys. J Comp Neurol 1995; 355: Burton H, Abend NS, MacLeod AM, Sinclair RJ, Snyder AZ, Raichle ME. Tactile attention tasks enhance activation in somatosensory regions of parietal cortex: a positron emission tomography study. Cereb Cortex 1999; 9(7): Carlson M, Huerta MR, Cusick CG, Kaas JH. Studies on the evolution of multiple somatosensory representations in primates: the organization of anterior parietal cortex in the new word callitrichid, saguinus. J. Comp. Neurol 1986; 246: Cusick CG,Wall JT, Kaas JH. Representations of the face, teeth and oral cavity in areas 3b and 1 of somatosensory cortex in squirrel monkeys. Brain Res 1986; 370:
38 Cusick CG, Wall JT, Felleman DJ, Kaas JH. Somatotopic organization of the lateral sulcus of owl monkey: area 3b, S-II and a ventral somatosensory area. J Comp Neurol 1989; 282: Cusick CG, Manning MM. Somatotopic organization and thalamic connections of the second somatosensory area, SII, in squirrel monkeys. Soc Neurosci Abstr 1998;14:222. Del Gratta C, Della Penna S, Tartaro A, Ferretti A, Torquati K, Bonomo L, Romani GL, Rossini PM. Topographic organization of the human primary and secondary somatosensory areas: an fmri study. Neuroreport 2000; 11: Diamond ME, Harris JA, Petersen RS. Sensory learning and the brain's body map In: R.J. Nelson, Editors, The Somatosensory System, CRC Press, Boca Raton, USA 2002, pp Disbrow E, Roberts T, Krubitzer L. Somatotopic organization of cortical fields in the lateral sulcus of Homo sapiens: evidence for SII and PV. J Comp Neurol 2000; 418:1 21. Ferretti A, Babiloni C, Del Gratta C, Caulo M, Tartaro A, Bonomo L, Maria Rossini P, Luca Romani G. Functional topography of the secondary somatosensory cortex for nonpainful and painful stimuli: an fmri study. NeuroImage 2003;20(3): Ferretti A, Del Gratta C, Babiloni C, Caulo M, Arienzo D, Tartaro A, Maria Rossini P, Luca Romani G. Functional topography of the secondary somatosensory cortex for nonpainful and painful stimulation of median and tibial nerve: an fmri study. NeuroImage 2004;23: Forss N, Jousmäki V. Sensorimotor integration in human primary and secondary somatosensory cortices. Brain Res 1998; 781: Forss N, Hietanen M, Salonen O, Hari R. Modified activation of somatosensory cortical network in patients with right-hemisphere stroke. Brain 1999; 122: Friedman DP, Jones EG, Burton H. Representation pattern in the second somatic sensory area of the monkey cerebral cortex. J Comp Neurol 1980;192: Frot M, Mauguière F. Timing and Spatial Distribution of Somatosensory Responses Recorded in the Upper Bank of the Sylvian Fissure (SII Area) in Humans. Cerebral Cortex 1999; 9 (8):
39 Hämäläinen M, Hari R, Ilmonienmi RJ, Knuutila J, Lounasmaa OV. Magnetoencephalography-theory, instrumentation and applications to noninvasive studies of the working human brain. Rev. Mod. Phys 1993; 65: Hari R, Karhu M, Hämäläinen J, Knuutila O, Salonen M, Sams M, Vilkman V. Functional organization of the human first and second somatosensory cortices: a neuromagnetic study. Eur. J. Neurosci : Hari R, Imada T. Ipsilateral movement-evoked fields reconsidered. Neuroimage 1999; 10: Hoshiyama M, Kakigi R, Koyama S, Kitamura Y, Shimojo M, Watanabe S. Somatosensory evoked magnetic fields after mechanical stimulation of the scalp in humans. Neurosci. Lett 1995; 195: Hoshiyama M, Kakigi R, Koyama S, Kitamura Y, Shimojo M, Watanabe S. Somatosensory evoked magnetic fields following stimulation of the lip in humans. Electroencephalogr. Clin. Neurophysiol 1996; 100: Iannetti GD, Porro CA, Pantano P, Romanelli PL, Galeotti F, Cruccu G. Representation of different trigeminal divisions within the primary and secondary human somatosensory cortex. NeuroImage 2003;19: Inui K, Tran TD, Qiu Y, Wang X, Hoshiyama M, Kakigi R. A comparative magnetoencephalographic study of cortical activations evoked by noxious and innocuous somatosensory stimulations. Neuroscience 2003;120(235):248. Iwamura Y. Hierarchical somatosensory processing. Curr Opin Neurobiol 1998; 8(4): Kaas JH, Nelson RJ, Sur M, Lin CS, Merzenich MM. Multiple representations of the body within the primary somatosensory cortex of primates. Science 1979; 204: Kakigi R, Koyama S, Hoshiyama M, Shimojo M, Kitamura Y, Watanabe S. Topography of somatosensory evoked magnetic fields following posterior tibial nerve stimulation. Electroenceph Clin Neurophysiol 1995;95: Kakigi R, Hoshiyama M, Shimojoa M, Yamasaki H, Watanabe S, Xiang J, Maeda K, Lam K, Itomi K, Nakamura A. The somatosensory evoked magnetic fields. Prog. Neurobiol 2000; 5:
40 Karhu J, Hari R, Lu ST, Paetau R, Rif J. Cerebral magnetic fields to lingual stimulation. Electroencephalogr. Clin. Neurophysiol 1991; 80: Krubitzer LA, Sesma MA, Kaas JH. Microelectrode maps, myeloarchitecture, and cortical connections of three somatotopically organized representations of the body surface in the parietal cortex of squirrels. J Comp Neurol 1986;250(4): Krubitzer LA, Clarey J, Tweedale R, Elston G, Calford M. A redefinition of somatosensory areas in the lateral sulcus of macaque monkeys. J Neurosci 1995;15: Ledberg A, O Sullivan BT, Kinomura S, Roland PE. Somatosensory activations of the parietal operculum of man. A PET study. Eur J Neurosci 1995;7: Maeda K, Kakigi R, Hoshiyama M, Koyama S. Topography of the secondary somatosensory cortex in humans: a magnetoencephalographic study. Neuroreport 1999;10: Makela JP, Illman M, Jousmaki V, Numminen J, Lehecka M, Salenius S, Forss N, Hari R. Dorsal penile nerve stimulation elicits left-hemisphere dominant activation in the second somatosensory cortex. Human Brain Mapp 2003;18:90 9. McCarthy G, Allison T, Spencer DD. Localization of the face area of human sensorimotor cortex by intracranial recording of somatosensory evoked potentials. J Neurosurg 1993;79: McCarthy G, Allison T. Trigeminal evoked potentials in somatosensory cortex of the Macaca mulatta. J. Neurosurg 1995; 82: Mima T, Ikeda A, Nagamine T, Yazawa S, Kunieda T, Mikuni N, Taki W, Kimura J, Shibasaki H.. Human second somatosensory area: subdural and magnetoencephalographic recording of somatosensory evoked responses. J Neurol Neurosurg Psychiatry 1997; 63(4): Mogilner A, Nomura M, Ribary U, Jagow R, Lado F, Rusinek H, Llinas R. Neuromagnetic studies of the lip area of primary somatosensory cortex in humans evidence for an oscillotopic organization. Exp. Brain Res 1994; 99: Nakamura A, Yamada T, Goto A, Kato T, Ito K, Abe Y, Kachi T, Kakigi R. Somatosensory homunculus as drawn by MEG. Neuroimage 1998; 7: Nelson RJ, Sur M, Felleman DJ, Kaas JH. Representations of the body surface in 39
41 postcentral parietal cortex of macaca fascicularis. J. Comp. Neurol 1998; 192: Nguyen TB, Tran DT, Inui K, Hoshiyama M, Kakigi R. Face representation in the human primary somatosensory cortex. Neurosci Res 2004;50: Nguyen TB, Inui K, Hoshiyama M, Nakata H, Tran DT, Kakigi R. Face representation in the human secondary somatosensory cortex. Clin Neurophysiol 2005 (in press) Nihashi T, Kakigi R, Kawakami O, Hoshiyama M, Itomi K, Nakanishi H, Kajita Y, Inao S, Yoshida J. Representation of the ear in human primary somatosensory cortex. Neuroimage 2001; 13: Nihashi T, Kakigi R, Okada T, Sadato N, Kashikura K, Kajita Y, Yoshida J. Functional magnetic resonance imaging evidence for a representation of the ear in human primary somatosensory cortex: comparison with magnetoencephalography study. Neuroimage 2002; 17: Nihashi T, Kakigi R, Hoshiyama M, Miki K, Kajita Y, Yoshida J, Yatsuya H. Effect of tactile interference stimulation of the ear in human primary somatosensory cortex: a magnetoencephalographic study. Electroencephalogr. Clin. Neurophysiol 2003; 114: Nishitani N, Uutela K, Shibasaki H, Hari R. Cortical visuomotor integration during eye pursuit and eye-finger pursuit. J Neurosci 1999; Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937; 60: Penfield W, Ramussen T. The cerebral cortex of man: a clinical study of localization of function. New York: Macmillan; Pons TP, Garraghty PE, Friedman DP, Mishkin M. Physiological evidence for serial processing in somatosensory cortex. Science 1987;237: Pons TP, Garraghty PE, Ommaya AK, Kaas JH, Taub E, Mishkin M. Massive cortical reorganization after sensory deafferentation in adult macaques. Science 1991; 252: Ridley RM, Ettlinger G, Impaired tactile learning and retention after removals of the second somatic sensory projection cortex (SII) in the monkey, Brain Res (3):
42 Robinson CJ, Burton H. Somatotopographic organization in the second somatosensory area of M. fascicularis. J Comp Neurol 1980; 192: Roland PE, O Sullivan B, Kawashima R. Shape and roughness activate different somatosensory areas in the human brain. Proc Natl Acad Sci USA 1998;95: Ruben J, Schwiemann J, Deuchert M, Meyer R, Krause T, Curio G, Villringer K, Kurth R, Villringer A. Somatotopic organization of human secondary somatosensory cortex. Cereb Cortex 2001;11: Servos P, Engel SA, Gati J, Menon R. FMRI evidence for an inverted face representation in human somatosensory cortex. Neuroreport 1999; 10: Sinclair RJ, Burton H. Neuronal activity in the second somatosensory cortex of monkeys (Macaca mulatto) during active touch of gratings. J Neurophysiol 1993; 70: Stöhr M, Petruch F. Somatosensory evoked potentials following stimulation of the trigeminal nerve in man. J. Neurol 1979; 220: Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. 3- Dimensional proportional system: an approach to cerebral imaging. Stuttgart: Thieme Verlag; Whitsel BL, Petrucelli LM, Werner G. Symmetry and connectivity in the map of the body surface in somatosensory area II of primates. J Neurophysiol 1969;32: Woolsey CN, Erickson TC, Gilson WE. Localization in somatic sensory and motor areas of human cerebral cortex as determined by direct recording of evoked potentials and electrical stimulation. J Neurosurg 1979;51: Yang TT, Gallen CC, Schawartz BJ, Bloom FE. Noninvasive somatosensory homunculus mapping in humans by using a large-array biomagnetometer. Proc. Natl. Acad. Sci. U.S.A 1994; 90:
43 Table 1. The mean latency, source strength and GOF value for each ECD (Experiment 1) Stimulation point Latency (ms) Source strength (Q/nAm) GOF (%) (1) Forehead ± ± ±3.5 (2) Forehead ± ± ±4.9 (3) Nose 39.8± ± ±3.1 (4) Cheek 40.9± ± ±4.3 (5) Chin ± ± ±1.9 (6) Chin ± ± ±6.5 Thumb 41.4± ± ±1.4 Lower lip 39.3± ± ±4.7 ECD, Equivalent current dipole; GOF, Goodness of fit. 42
44 Table 2. The mean ECD location of six face points, thumb and lip (Experiment 1) Stimulation point x (mm) y (mm) z (mm) (1)Forehead ± ± ±7.0 (2)Forehead ± ± ±10.4 (3)Nose -51.7± ± ±6.7 (4)Cheek -45.0± ± ±7.3 (5)Chin ± ± ±7.2 (6)Chin ± ± ±11.9 Thumb -42.0± ± ±5.3 Lower lip -52.0± ± ±4.0 The mean standardized x, y and z values (±SD) of each ECD location. X, medio-lateral; Y, rostro-caudal; Z, dorso-ventral To calculate the standardized x, y, z values we measured the maximum brain length from a y-z plane (x axis), x-z plane (y axis) and x-y plane (z axis) on MR image of each subject. The mean value for each axis among 7 subjects was calculated and used as a standard. Then, original x, y and z coordinate values for ECDs of each subject were 43
45 corrected by the ratio of the subject s maximum brain length to the standard (see the methods for more details) 44
46 Table 3. The mean and standard deviation of peak latency and source strength following stimulation of the foot, lip and three face points (Experiment 2) Stimulation Latency Source strength Point (ms) (nam) Lip 97.0± ±4.6 M angle 100.0± ±7.0 Cheek 95.8± ±6.0 Forehead 93.2± ±7.6 Foot 129.0± ±4.6 45
47 Table 4. The mean Talairach coordinates of ECD locations following stimulation of the foot, lip and three face points (Experiment 2) Stimulation x y z Point (mm) (mm) (mm) Lip -58.0± ± ±6.8 M angle -51.3± ± ±5.6 Cheek -53.8± ± ±4.5 Forehead -56.1± ± ±3.4 Foot -45.1± ± ±5.4 The mean standardized Talairach x, y and z values ± SD of each ECD location. X, medio-lateral; Y, antero-posterior; Z, infero-superior 46
48 Figure legends Figure 1. (a) Schematic drawing of the points stimulated on the face in Experiment 1; point 1: forehead 1; point 2: forehead 2; point 3: nose; point 4: cheek; point 5: chin 1; point 6: chin 2. (b) The tip of the stimulator. Figure 2. Whole-scalp 306 channel neuromagnetometer (Elektra Neuromag Yo machine). Left: Helmet- shape with the arrangement of all sensors cover whole head. Right: MEG machine. Figure 3. X, Y, Z coordinates system. X axis goes through LPA and RPA points, with positive direction toward the RPA. Y axis goes through Nasion point, which is a positive side of axis. Z axis is perpendicular to X and Y axis, with the positive axis is superior. Figure 4. Evoked magnetic fields following stimulation of the thumb, lip and facial skin areas in Experiment 1. Left, SEFs following stimulation of the right thumb (black color), nose (red color) and lower lip (blue color) in subject 1. Upper panel, the head viewed from top, illustrates signals recorded by the two orthogonal gradiometers (G1 and G2) of each single sensor unit. A, B and C are selected channels over the peak response 47
49 (enlarged in the lower panel and right). Lower panel, enlargement of the upper panel's insert. Right, responses following stimulation of the eight points over the left rolandic area in subject 1. Vertical line indicates the stimulus onset. Figure 5. Locations of the eight ECDs in subject 1 of Experiment 1. The locations of all sources are superimposed on a single axial and coronal MRI slice, with which the ECD for stimulation of the lip was estimated, to show their relative relationships. The ECD location for thumb is illustrated by a square, for lip by a triangle, and for the six points of facial skin by dots. Figure 6. Spatial relationship of the group-mean ECD locations following stimulation of the eight points in Experiment 1. The location of each ECD is expressed by x, y and z coordinates relative to the ECD location for the thumb. (A) Medial lateral direction (x value) and superior inferior direction (z value); (B) medial lateral direction (x value) and anterior posterior direction (y value). Figure 7. Different field distributions for activities from the primary (SI) and secondary (SII) somatosensory cortex following forehead stimulation in Experiment 2. Figure 8. Evoked magnetic fields following stimulation of the foot, lip and three facial 48
50 skin points recorded in representative subjects in Experiment 2. Left figure: Magnetic responses recorded at all sensors produced by forehead stimulation. Sensors selected for analysis around SII are shown on a circle (A). Right figure: Enlarged waveforms of a single channel showing the largest response in the contralateral hemisphere for all stimulation points from two subjects. Figure 9. ECD locations for foot, lip and three facial points superimposed on MR images in one representative subject in Experiment 2. All ECDs were overlaid on the slice on which the cheek was estimated. The ECD for lip is located most lateral, anterior and inferior, while the ECD for foot is located most medial, posterior and superior. ECDs for 3 facial skin areas are located very close to each other between the lip and foot regions, but closer to the lip region. Since the location for foot stimulation is located posterior to the sources of face. We show a coronal slice for foot. Black square for foot, pink triangle for lip, blue dot for forehead, yellow dot for cheek, navy dot for mandibular angle. Figure 10. Schematic drawing of spatial relationships of ECDs for SII among each stimulation point in Experiment 2. The mean standardized Talairach coordinates of the 49
51 ECDs for foot, lip and facial points. A black square is the ECD of foot, a pink dot is the ECD of lip and small circles indicate the ECD location of each facial point (green for forehead, yellow for cheek and navy for mandibular angle). A large circle indicates their group averages. Bars indicate ± standard deviation. (A) Medio-lateral and infero-superior direction. (B) Medio-lateral and antero-posterior direction. 50

52 Figure 1 51

53 Figure 2 52

54 Figure 3 53
55 Figure 4 54

56 Figure 5 55
57 Figure 6 56
58 Figure 7 57
59 Figure 8 58


60 Binh Thi Nguyen Figure 9 59
Clinical Material and Methods
 J Neurosurg 85:255 262, 1996 Differentiation of receptive fields in the sensory cortex following stimulation of various nerves of the lower limb in humans: a magnetoencephalographic study MOTOKO SHIMOJO,
J Neurosurg 85:255 262, 1996 Differentiation of receptive fields in the sensory cortex following stimulation of various nerves of the lower limb in humans: a magnetoencephalographic study MOTOKO SHIMOJO,
Serial Processing in the Human Somatosensory System
 Serial Processing in the Human Somatosensory System Koji Inui, Xiaohong Wang, Yohei Tamura, Yoshiki Kaneoke and Ryusuke Kakigi Department of Integrative Physiology, National Institute for Physiological
Serial Processing in the Human Somatosensory System Koji Inui, Xiaohong Wang, Yohei Tamura, Yoshiki Kaneoke and Ryusuke Kakigi Department of Integrative Physiology, National Institute for Physiological
Sensorimotor integration in human primary and secondary somatosensory cortices
 Ž. Brain Research 781 1998 Research report Sensorimotor integration in human primary and secondary somatosensory cortices Nina Forss ), Veikko Jousmaki Brain Research Unit, Low Temperature Laboratory,
Ž. Brain Research 781 1998 Research report Sensorimotor integration in human primary and secondary somatosensory cortices Nina Forss ), Veikko Jousmaki Brain Research Unit, Low Temperature Laboratory,
Lateral view of human brain! Cortical processing of touch!
 Lateral view of human brain! Cortical processing of touch! How do we perceive objects held in the hand?! Touch receptors deconstruct objects to detect local features! Information is transmitted in parallel
Lateral view of human brain! Cortical processing of touch! How do we perceive objects held in the hand?! Touch receptors deconstruct objects to detect local features! Information is transmitted in parallel
Sensorimotor Integration in S2, PV, and Parietal Rostroventral Areas of the Human Sylvian Fissure
 J Neurophysiol 97: 1288 1297, 2007. First published November 22, 2006; doi:10.1152/jn.00733.2006. Sensorimotor Integration in S2, PV, and Parietal Rostroventral Areas of the Human Sylvian Fissure Leighton
J Neurophysiol 97: 1288 1297, 2007. First published November 22, 2006; doi:10.1152/jn.00733.2006. Sensorimotor Integration in S2, PV, and Parietal Rostroventral Areas of the Human Sylvian Fissure Leighton
SOMATOSENSORY SYSTEMS
 SOMATOSENSORY SYSTEMS Schematic diagram illustrating the neural pathways that convey somatosensory information to the cortex and, subsequently, to the motor system. Double arrows show reciprocal connections.
SOMATOSENSORY SYSTEMS Schematic diagram illustrating the neural pathways that convey somatosensory information to the cortex and, subsequently, to the motor system. Double arrows show reciprocal connections.
Cortical Control of Movement
 Strick Lecture 2 March 24, 2006 Page 1 Cortical Control of Movement Four parts of this lecture: I) Anatomical Framework, II) Physiological Framework, III) Primary Motor Cortex Function and IV) Premotor
Strick Lecture 2 March 24, 2006 Page 1 Cortical Control of Movement Four parts of this lecture: I) Anatomical Framework, II) Physiological Framework, III) Primary Motor Cortex Function and IV) Premotor
Water immersion modulates sensory and motor cortical excitability
 Water immersion modulates sensory and motor cortical excitability Daisuke Sato, PhD Department of Health and Sports Niigata University of Health and Welfare Topics Neurophysiological changes during water
Water immersion modulates sensory and motor cortical excitability Daisuke Sato, PhD Department of Health and Sports Niigata University of Health and Welfare Topics Neurophysiological changes during water
How strong is it? What is it? Where is it? What must sensory systems encode? 9/8/2010. Spatial Coding: Receptive Fields and Tactile Discrimination
 Spatial Coding: Receptive Fields and Tactile Discrimination What must sensory systems encode? How strong is it? What is it? Where is it? When the brain wants to keep certain types of information distinct,
Spatial Coding: Receptive Fields and Tactile Discrimination What must sensory systems encode? How strong is it? What is it? Where is it? When the brain wants to keep certain types of information distinct,
Spatial Coding: Receptive Fields and Tactile Discrimination
 Spatial Coding: Receptive Fields and Tactile Discrimination What must sensory systems encode? How strong is it? What is it? Where is it? When the brain wants to keep certain types of information distinct,
Spatial Coding: Receptive Fields and Tactile Discrimination What must sensory systems encode? How strong is it? What is it? Where is it? When the brain wants to keep certain types of information distinct,
A Redefinition of Somatosensory Areas in the Lateral Sulcus of Macaque Monkeys
 The Journal of Neuroscience, May 1995, 15(5): 3821-3839 A Redefinition of Somatosensory Areas in the Lateral Sulcus of Macaque Monkeys Leah Krubitzer, Janine Clarey, Rowan Tweedale, Guy Elston, and Mike
The Journal of Neuroscience, May 1995, 15(5): 3821-3839 A Redefinition of Somatosensory Areas in the Lateral Sulcus of Macaque Monkeys Leah Krubitzer, Janine Clarey, Rowan Tweedale, Guy Elston, and Mike
The Topography of Tactile Learning in Humans
 The Journal of Neuroscience, February 1, 2001, 21(3):1056 1061 The Topography of Tactile Learning in Humans Justin A. Harris, Irina M. Harris, and Mathew E. Diamond Cognitive Neuroscience Sector, International
The Journal of Neuroscience, February 1, 2001, 21(3):1056 1061 The Topography of Tactile Learning in Humans Justin A. Harris, Irina M. Harris, and Mathew E. Diamond Cognitive Neuroscience Sector, International
Homework Week 2. PreLab 2 HW #2 Synapses (Page 1 in the HW Section)
") Homework Week 2 Due in Lab PreLab 2 HW #2 Synapses (Page 1 in the HW Section) Reminders No class next Monday Quiz 1 is @ 5:30pm on Tuesday, 1/22/13 Study guide posted under Study Aids section of website
Homework Week 2 Due in Lab PreLab 2 HW #2 Synapses (Page 1 in the HW Section) Reminders No class next Monday Quiz 1 is @ 5:30pm on Tuesday, 1/22/13 Study guide posted under Study Aids section of website
Title. CitationClinical Neurophysiology, 125(2): Issue Date Doc URL. Type. File Information.
: Issue Date Doc URL. Type. File Information.") Title Evaluation of lip sensory disturbance using somatose Author(s)Maezawa, Hitoshi; Matsuhashi, Masao; Yoshida, Kazuya CitationClinical Neurophysiology, 125(2): 363-369 Issue Date 2014-02 Doc URL http://hdl.handle.net/2115/54879
Title Evaluation of lip sensory disturbance using somatose Author(s)Maezawa, Hitoshi; Matsuhashi, Masao; Yoshida, Kazuya CitationClinical Neurophysiology, 125(2): 363-369 Issue Date 2014-02 Doc URL http://hdl.handle.net/2115/54879
Optical imaging of nociception in primary somatosensory cortex of nonhuman
 664 Acta Physiologica Sinica, October 25, 2008, 60 (5): 664-668 http://www.actaps.com.cn Review Optical imaging of nociception in primary somatosensory cortex of nonhuman primates Li-Min CHEN 1,3,*, Robert
664 Acta Physiologica Sinica, October 25, 2008, 60 (5): 664-668 http://www.actaps.com.cn Review Optical imaging of nociception in primary somatosensory cortex of nonhuman primates Li-Min CHEN 1,3,*, Robert
Anatomical Substrates of Somatic Sensation
 Anatomical Substrates of Somatic Sensation John H. Martin, Ph.D. Center for Neurobiology & Behavior Columbia University CPS The 2 principal somatic sensory systems: 1) Dorsal column-medial lemniscal system
Anatomical Substrates of Somatic Sensation John H. Martin, Ph.D. Center for Neurobiology & Behavior Columbia University CPS The 2 principal somatic sensory systems: 1) Dorsal column-medial lemniscal system
Contribution of the human superior parietal lobule to spatial selection process: an MEG study
 Brain Research 897 (2001) 164 168 www.elsevier.com/ locate/ bres Short communication Contribution of the human superior parietal lobule to spatial selection process: an MEG study * Tadahiko Shibata, Andreas
Brain Research 897 (2001) 164 168 www.elsevier.com/ locate/ bres Short communication Contribution of the human superior parietal lobule to spatial selection process: an MEG study * Tadahiko Shibata, Andreas
Mechanosensation. Central Representation of Touch. Wilder Penfield. Somatotopic Organization
 Mechanosensation Central Representation of Touch Touch and tactile exploration Vibration and pressure sensations; important for clinical testing Limb position sense John H. Martin, Ph.D. Center for Neurobiology
Mechanosensation Central Representation of Touch Touch and tactile exploration Vibration and pressure sensations; important for clinical testing Limb position sense John H. Martin, Ph.D. Center for Neurobiology
Note: Waxman is very sketchy on today s pathways and nonexistent on the Trigeminal.
 Dental Neuroanatomy Thursday, February 3, 2011 Suzanne Stensaas, PhD Note: Waxman is very sketchy on today s pathways and nonexistent on the Trigeminal. Resources: Pathway Quiz for HyperBrain Ch. 5 and
Dental Neuroanatomy Thursday, February 3, 2011 Suzanne Stensaas, PhD Note: Waxman is very sketchy on today s pathways and nonexistent on the Trigeminal. Resources: Pathway Quiz for HyperBrain Ch. 5 and
Somatosensation. Recording somatosensory responses. Receptive field response to pressure
 Somatosensation Mechanoreceptors that respond to touch/pressure on the surface of the body. Sensory nerve responds propotional to pressure 4 types of mechanoreceptors: Meissner corpuscles & Merkel discs
Somatosensation Mechanoreceptors that respond to touch/pressure on the surface of the body. Sensory nerve responds propotional to pressure 4 types of mechanoreceptors: Meissner corpuscles & Merkel discs
 Peripheral facial paralysis (right side). The patient is asked to close her eyes and to retract their mouth (From Heimer) Hemiplegia of the left side. Note the characteristic position of the arm with
Peripheral facial paralysis (right side). The patient is asked to close her eyes and to retract their mouth (From Heimer) Hemiplegia of the left side. Note the characteristic position of the arm with
Tactile processing in human somatosensory and auditory cortices
 University of Helsinki Department of Radiology Tactile processing in human somatosensory and auditory cortices Yevhen Hlushchuk Brain Research Unit Low Temperature Laboratory, and Advanced Magnetic Imaging
University of Helsinki Department of Radiology Tactile processing in human somatosensory and auditory cortices Yevhen Hlushchuk Brain Research Unit Low Temperature Laboratory, and Advanced Magnetic Imaging
The Central Nervous System
 The Central Nervous System Cellular Basis. Neural Communication. Major Structures. Principles & Methods. Principles of Neural Organization Big Question #1: Representation. How is the external world coded
The Central Nervous System Cellular Basis. Neural Communication. Major Structures. Principles & Methods. Principles of Neural Organization Big Question #1: Representation. How is the external world coded
Left-hemisphere dominance for processing of vowels: a whole-scalp neuromagnetic study
 Auditory and Vestibular Systems 10, 2987±2991 (1999) BRAIN activation of 11 healthy right-handed subjects was studied with magnetoencephalography to estimate individual hemispheric dominance for speech
Auditory and Vestibular Systems 10, 2987±2991 (1999) BRAIN activation of 11 healthy right-handed subjects was studied with magnetoencephalography to estimate individual hemispheric dominance for speech
POSTERIOR SLOW WAVES of youth (PSWy) Source localization of posterior slow waves of youth using dipole modeling. Regular Article
 Source localization of posterior slow waves of youth using dipole modeling. Regular Article") Psychiatry and Clinical Neurosciences 2012; 66: 582 586 doi:10.1111/j.1440-1819.2012.02398.x Regular Article Source localization of posterior slow waves of youth using dipole modeling Keiko Ohoyama, MD,
Psychiatry and Clinical Neurosciences 2012; 66: 582 586 doi:10.1111/j.1440-1819.2012.02398.x Regular Article Source localization of posterior slow waves of youth using dipole modeling Keiko Ohoyama, MD,
Cerebral Cortex 1. Sarah Heilbronner
 Cerebral Cortex 1 Sarah Heilbronner heilb028@umn.edu Want to meet? Coffee hour 10-11am Tuesday 11/27 Surdyk s Overview and organization of the cerebral cortex What is the cerebral cortex? Where is each
Cerebral Cortex 1 Sarah Heilbronner heilb028@umn.edu Want to meet? Coffee hour 10-11am Tuesday 11/27 Surdyk s Overview and organization of the cerebral cortex What is the cerebral cortex? Where is each
V1-ophthalmic. V2-maxillary. V3-mandibular. motor
 4. Trigeminal Nerve I. Objectives:. Understand the types of sensory information transmitted by the trigeminal system.. Describe the major peripheral divisions of the trigeminal nerve and how they innervate
4. Trigeminal Nerve I. Objectives:. Understand the types of sensory information transmitted by the trigeminal system.. Describe the major peripheral divisions of the trigeminal nerve and how they innervate
Gum Chewing Maintains Working Memory Acquisition
 International Journal of Bioelectromagnetism Vol. 11, No. 3, pp.130-134, 2009 www.ijbem.org Gum Chewing Maintains Working Memory Acquisition Yumie Ono a, Kanako Dowaki b, Atsushi Ishiyama b, Minoru Onozuka
International Journal of Bioelectromagnetism Vol. 11, No. 3, pp.130-134, 2009 www.ijbem.org Gum Chewing Maintains Working Memory Acquisition Yumie Ono a, Kanako Dowaki b, Atsushi Ishiyama b, Minoru Onozuka
Spatiotemporal separability in the human cortical response to visual motion speed: a magnetoencephalography study
 Neuroscience Research 47 (2003) 109/116 www.elsevier.com/locate/neures Spatiotemporal separability in the human cortical response to visual motion speed: a magnetoencephalography study Lihong Wang, Yoshiki
Neuroscience Research 47 (2003) 109/116 www.elsevier.com/locate/neures Spatiotemporal separability in the human cortical response to visual motion speed: a magnetoencephalography study Lihong Wang, Yoshiki
SUPPLEMENTARY MATERIAL. Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome.
 SUPPLEMENTARY MATERIAL Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome. Authors Year Patients Male gender (%) Mean age (range) Adults/ Children
SUPPLEMENTARY MATERIAL Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome. Authors Year Patients Male gender (%) Mean age (range) Adults/ Children
Cerebrum-Cerebral Hemispheres. Cuneyt Mirzanli Istanbul Gelisim University
 Cerebrum-Cerebral Hemispheres Cuneyt Mirzanli Istanbul Gelisim University The largest part of the brain. Ovoid shape. Two incompletely separated cerebral hemispheres. The outer surface of the cerebral
Cerebrum-Cerebral Hemispheres Cuneyt Mirzanli Istanbul Gelisim University The largest part of the brain. Ovoid shape. Two incompletely separated cerebral hemispheres. The outer surface of the cerebral
Medical Neuroscience Tutorial
 Pain Pathways Medical Neuroscience Tutorial Pain Pathways MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. NCC3. Genetically determined circuits are the foundation
Pain Pathways Medical Neuroscience Tutorial Pain Pathways MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. NCC3. Genetically determined circuits are the foundation
SOMATIC SENSATION PART I: ALS ANTEROLATERAL SYSTEM (or SPINOTHALAMIC SYSTEM) FOR PAIN AND TEMPERATURE
 FOR PAIN AND TEMPERATURE") Dental Neuroanatomy Thursday, February 3, 2011 Suzanne S. Stensaas, PhD SOMATIC SENSATION PART I: ALS ANTEROLATERAL SYSTEM (or SPINOTHALAMIC SYSTEM) FOR PAIN AND TEMPERATURE Reading: Waxman 26 th ed, :
Dental Neuroanatomy Thursday, February 3, 2011 Suzanne S. Stensaas, PhD SOMATIC SENSATION PART I: ALS ANTEROLATERAL SYSTEM (or SPINOTHALAMIC SYSTEM) FOR PAIN AND TEMPERATURE Reading: Waxman 26 th ed, :
Serial activation of distinct cytoarchitectonic areas of the human S1 cortex after posterior tibial nerve stimulation
 NEUOPHYSIOOGY, BASIC AND CINICA NEUOEPOT Serial activation of distinct cytoarchitectonic areas of the human S1 cortex after posterior tibial nerve stimulation Isao Hashimoto, 1,2,CA Kenji Sakuma, 2 Tomoaki
NEUOPHYSIOOGY, BASIC AND CINICA NEUOEPOT Serial activation of distinct cytoarchitectonic areas of the human S1 cortex after posterior tibial nerve stimulation Isao Hashimoto, 1,2,CA Kenji Sakuma, 2 Tomoaki
MEG localization of rolandic spikes with respect to SI and SII cortices in benign rolandic epilepsy
 NeuroImage 20 (2003) 2051 2061 www.elsevier.com/locate/ynimg MEG localization of rolandic spikes with respect to SI and SII cortices in benign rolandic epilepsy Y.Y. Lin, a,b,c, * Y.H. Shih, b,c K.P. Chang,
NeuroImage 20 (2003) 2051 2061 www.elsevier.com/locate/ynimg MEG localization of rolandic spikes with respect to SI and SII cortices in benign rolandic epilepsy Y.Y. Lin, a,b,c, * Y.H. Shih, b,c K.P. Chang,
Large-Scale Reorganization in the Somatosensory Cortex and Thalamus after Sensory Loss in Macaque Monkeys
 11042 The Journal of Neuroscience, October 22, 2008 28(43):11042 11060 Behavioral/Systems/Cognitive Large-Scale Reorganization in the Somatosensory Cortex and Thalamus after Sensory Loss in Macaque Monkeys
11042 The Journal of Neuroscience, October 22, 2008 28(43):11042 11060 Behavioral/Systems/Cognitive Large-Scale Reorganization in the Somatosensory Cortex and Thalamus after Sensory Loss in Macaque Monkeys
Serial and Parallel Processing in the Human Auditory Cortex: A Magnetoencephalographic Study
 Cerebral Cortex January 2006;16:18--30 doi:10.1093/cercor/bhi080 Advance Access publication March 30, 2005 Serial and Parallel Processing in the Human Auditory Cortex: A Magnetoencephalographic Study Koji
Cerebral Cortex January 2006;16:18--30 doi:10.1093/cercor/bhi080 Advance Access publication March 30, 2005 Serial and Parallel Processing in the Human Auditory Cortex: A Magnetoencephalographic Study Koji
The Somatosensory System
 The Somatosensory System Reading: BCP Chapter 12 cerebrovortex.com Divisions of the Somatosensory System Somatosensory System Exteroceptive External stimuli Proprioceptive Body position Interoceptive Body
The Somatosensory System Reading: BCP Chapter 12 cerebrovortex.com Divisions of the Somatosensory System Somatosensory System Exteroceptive External stimuli Proprioceptive Body position Interoceptive Body
The Human Parietal Operculum. I. Cytoarchitectonic Mapping of Subdivisions
 Cerebral Cortex February 26;6:254-267 doi:.93/cercor/bhi5 Advance Access publication May, 25 The Human Parietal Operculum. I. Cytoarchitectonic Mapping of Subdivisions Simon B. Eickhoff,2, Axel Schleicher
Cerebral Cortex February 26;6:254-267 doi:.93/cercor/bhi5 Advance Access publication May, 25 The Human Parietal Operculum. I. Cytoarchitectonic Mapping of Subdivisions Simon B. Eickhoff,2, Axel Schleicher
LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS
 LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS WHAT WE DO Present studies and future directions Our laboratory is currently involved in two major areas of research. The first
LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS WHAT WE DO Present studies and future directions Our laboratory is currently involved in two major areas of research. The first
The Representation of the Human Oral Area in the Somatosensory Cortex: a Functional MRI Study
 Cerebral Cortex May 2006;16:669-675 doi:10.1093/cercor/bhj012 Advance Access publication August 3, 2005 The Representation of the Human Oral Area in the Somatosensory Cortex: a Functional MRI Study Jun
Cerebral Cortex May 2006;16:669-675 doi:10.1093/cercor/bhj012 Advance Access publication August 3, 2005 The Representation of the Human Oral Area in the Somatosensory Cortex: a Functional MRI Study Jun
Skin types: hairy and glabrous (e.g. back vs. palm of hand)
") Lecture 19 revised 03/10 The Somatic Sensory System Skin- the largest sensory organ we have Also protects from evaporation, infection. Skin types: hairy and glabrous (e.g. back vs. palm of hand) 2 major
Lecture 19 revised 03/10 The Somatic Sensory System Skin- the largest sensory organ we have Also protects from evaporation, infection. Skin types: hairy and glabrous (e.g. back vs. palm of hand) 2 major
Supporting Information
 Supporting Information Moriguchi and Hiraki 10.1073/pnas.0809747106 SI Text Differences in Brain Activation Between Preswitch and Postswitch Phases. The paired t test was used to compare the brain activation
Supporting Information Moriguchi and Hiraki 10.1073/pnas.0809747106 SI Text Differences in Brain Activation Between Preswitch and Postswitch Phases. The paired t test was used to compare the brain activation
Auditory and Vestibular Systems
 Auditory and Vestibular Systems Objective To learn the functional organization of the auditory and vestibular systems To understand how one can use changes in auditory function following injury to localize
Auditory and Vestibular Systems Objective To learn the functional organization of the auditory and vestibular systems To understand how one can use changes in auditory function following injury to localize
Clinical Neurophysiology
 Clinical Neurophysiology 119 (8) 238 2388 Contents lists available at ScienceDirect Clinical Neurophysiology journal homepage: www.elsevier.com/locate/clinph Central processing of tactile and nociceptive
Clinical Neurophysiology 119 (8) 238 2388 Contents lists available at ScienceDirect Clinical Neurophysiology journal homepage: www.elsevier.com/locate/clinph Central processing of tactile and nociceptive
Representation of the Body Surface in Somatic Koniocortex in the Prosimian Galago
 JOURNAL OF COMPARATIVE NEUROLOGY 189:381-402 (1980) Representation of the Body Surface in Somatic Koniocortex in the Prosimian Galago MRIGANKA SUR, R.J. NELSON, AND J.H. KAAS Departments of Psychology
JOURNAL OF COMPARATIVE NEUROLOGY 189:381-402 (1980) Representation of the Body Surface in Somatic Koniocortex in the Prosimian Galago MRIGANKA SUR, R.J. NELSON, AND J.H. KAAS Departments of Psychology
Analysis of brain activity immediately before conscious teeth clenching using magnetoencephalographic method
 Journal of Oral Rehabilitation 2007 34; 487 496 Analysis of brain activity immediately before conscious teeth clenching using magnetoencephalographic method T. IIDA*,, P. B. C. FENWICK & A. A. IOANNIDES
Journal of Oral Rehabilitation 2007 34; 487 496 Analysis of brain activity immediately before conscious teeth clenching using magnetoencephalographic method T. IIDA*,, P. B. C. FENWICK & A. A. IOANNIDES
Sensory information processing, somato-sensory systems
 mm? Sensory information processing, somato-sensory systems Recommended literature 1. Kandel ER, Schwartz JH, Jessel TM (2000) Principles of Neural Science, McGraw-Hill, Ch. xx. 2. Berne EM, Levy MN, Koeppen
mm? Sensory information processing, somato-sensory systems Recommended literature 1. Kandel ER, Schwartz JH, Jessel TM (2000) Principles of Neural Science, McGraw-Hill, Ch. xx. 2. Berne EM, Levy MN, Koeppen
Exploring the pulvinar path to visual cortex
 C. Kennard & R.J. Leigh (Eds.) Progress in Brain Research, Vol. 171 ISSN 0079-6123 Copyright r 2008 Elsevier B.V. All rights reserved CHAPTER 5.14 Exploring the pulvinar path to visual cortex Rebecca A.
C. Kennard & R.J. Leigh (Eds.) Progress in Brain Research, Vol. 171 ISSN 0079-6123 Copyright r 2008 Elsevier B.V. All rights reserved CHAPTER 5.14 Exploring the pulvinar path to visual cortex Rebecca A.
Sensory coding and somatosensory system
 Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
Double Representation of the Body Surface within Cytoarchitectonic Areas 3b and 1 in SI in the Owl Monkey (Aotus trivirgatus)
") Double Representation of the Body Surface within Cytoarchitectonic Areas 3b and 1 in SI in the Owl Monkey (Aotus trivirgatus) MICHAEL M. MERZENICH,I JON H. KAAS, MRIGANKA SUR AND CHIA-SHENG LIN Coleman
Double Representation of the Body Surface within Cytoarchitectonic Areas 3b and 1 in SI in the Owl Monkey (Aotus trivirgatus) MICHAEL M. MERZENICH,I JON H. KAAS, MRIGANKA SUR AND CHIA-SHENG LIN Coleman
CISC 3250 Systems Neuroscience
 CISC 3250 Systems Neuroscience Levels of organization Central Nervous System 1m 10 11 neurons Neural systems and neuroanatomy Systems 10cm Networks 1mm Neurons 100μm 10 8 neurons Professor Daniel Leeds
CISC 3250 Systems Neuroscience Levels of organization Central Nervous System 1m 10 11 neurons Neural systems and neuroanatomy Systems 10cm Networks 1mm Neurons 100μm 10 8 neurons Professor Daniel Leeds
Somatic Sensory System I. Background
 Somatic Sensory System I. Background A. Differences between somatic senses and other senses 1. Receptors are distributed throughout the body as opposed to being concentrated at small, specialized locations
Somatic Sensory System I. Background A. Differences between somatic senses and other senses 1. Receptors are distributed throughout the body as opposed to being concentrated at small, specialized locations
Elissaios Karageorgiou Æ Ioannis G. Koutlas Æ Aurelio A. Alonso Æ Arthur C. Leuthold Æ Scott M. Lewis Æ Apostolos P. Georgopoulos
 Exp Brain Res (2008) 189:311 321 DOI 10.1007/s00221-008-1425-6 RESEARCH ARTICLE Cortical processing of tactile stimuli applied in quick succession across the fingertips: temporal evolution of dipole sources
Exp Brain Res (2008) 189:311 321 DOI 10.1007/s00221-008-1425-6 RESEARCH ARTICLE Cortical processing of tactile stimuli applied in quick succession across the fingertips: temporal evolution of dipole sources
Gross Organization I The Brain. Reading: BCP Chapter 7
 Gross Organization I The Brain Reading: BCP Chapter 7 Layout of the Nervous System Central Nervous System (CNS) Located inside of bone Includes the brain (in the skull) and the spinal cord (in the backbone)
Gross Organization I The Brain Reading: BCP Chapter 7 Layout of the Nervous System Central Nervous System (CNS) Located inside of bone Includes the brain (in the skull) and the spinal cord (in the backbone)
An Overview of BMIs. Luca Rossini. Workshop on Brain Machine Interfaces for Space Applications
 An Overview of BMIs Luca Rossini Workshop on Brain Machine Interfaces for Space Applications European Space Research and Technology Centre, European Space Agency Noordvijk, 30 th November 2009 Definition
An Overview of BMIs Luca Rossini Workshop on Brain Machine Interfaces for Space Applications European Space Research and Technology Centre, European Space Agency Noordvijk, 30 th November 2009 Definition
SOMATOSENSORY SYSTEMS: Pain and Temperature Kimberle Jacobs, Ph.D.
 SOMATOSENSORY SYSTEMS: Pain and Temperature Kimberle Jacobs, Ph.D. Sensory systems are afferent, meaning that they are carrying information from the periphery TOWARD the central nervous system. The somatosensory
SOMATOSENSORY SYSTEMS: Pain and Temperature Kimberle Jacobs, Ph.D. Sensory systems are afferent, meaning that they are carrying information from the periphery TOWARD the central nervous system. The somatosensory
Medical Neuroscience Tutorial Notes
 Medical Neuroscience Tutorial Notes Finding the Central Sulcus MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. LEARNING OBJECTIVES After study of the assigned learning
Medical Neuroscience Tutorial Notes Finding the Central Sulcus MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. LEARNING OBJECTIVES After study of the assigned learning
PARIETAL LOBE. Vasilios A. Zerris MD, MPH, MSc, FAANS
 PARIETAL LOBE Vasilios A. Zerris MD, MPH, MSc, FAANS Diplomate of the American Board of Neurological Surgery Fellow of the American Association of Neurological Surgeons Professor of Neurosurgery, European
PARIETAL LOBE Vasilios A. Zerris MD, MPH, MSc, FAANS Diplomate of the American Board of Neurological Surgery Fellow of the American Association of Neurological Surgeons Professor of Neurosurgery, European
Representations of the Body Surface. Monkeys: Comparisons With Other Primates
 THE JOURNAL OF COMPARATIVE NEUROLOGY 211:177-192 (1982) Representations of the Body Surface in Cortical Areas 3b and 1 of Squirrel Monkeys: Comparisons With Other Primates MRIGANKA SUR, R. J. NELSON, AND
THE JOURNAL OF COMPARATIVE NEUROLOGY 211:177-192 (1982) Representations of the Body Surface in Cortical Areas 3b and 1 of Squirrel Monkeys: Comparisons With Other Primates MRIGANKA SUR, R. J. NELSON, AND
STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM
 STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM STRUCTURAL ORGANIZATION OF THE BRAIN The central nervous system (CNS), consisting of the brain and spinal cord, receives input from sensory neurons and directs
STRUCTURAL ORGANIZATION OF THE NERVOUS SYSTEM STRUCTURAL ORGANIZATION OF THE BRAIN The central nervous system (CNS), consisting of the brain and spinal cord, receives input from sensory neurons and directs
Representations of the Body Surface in Postcentral Parietal Cortex of Macaca fascicularis
 THE JOURNAL OF COMPARATIVE NEUROLOGY 192~611-643 (1980) Representations of the Body Surface in Postcentral Parietal Cortex of Macaca fascicularis R.J. NELSON, M. SUR, D.J. FELLEMAN, AND J.H. KAAS Departments
THE JOURNAL OF COMPARATIVE NEUROLOGY 192~611-643 (1980) Representations of the Body Surface in Postcentral Parietal Cortex of Macaca fascicularis R.J. NELSON, M. SUR, D.J. FELLEMAN, AND J.H. KAAS Departments
Source Imaging in Mapping Eloquent Cortex December 6 th 2011
 Source Imaging in Mapping Eloquent Cortex December 6 th 2011 Samden Lhatoo, MD FRCP University Hospitals Case Medical Center American Epilepsy Society Annual Meeting Disclosure Name of Commercial Interest
Source Imaging in Mapping Eloquent Cortex December 6 th 2011 Samden Lhatoo, MD FRCP University Hospitals Case Medical Center American Epilepsy Society Annual Meeting Disclosure Name of Commercial Interest
Neurophysiology of systems
 Neurophysiology of systems Motor cortex (voluntary movements) Dana Cohen, Room 410, tel: 7138 danacoh@gmail.com Voluntary movements vs. reflexes Same stimulus yields a different movement depending on context
Neurophysiology of systems Motor cortex (voluntary movements) Dana Cohen, Room 410, tel: 7138 danacoh@gmail.com Voluntary movements vs. reflexes Same stimulus yields a different movement depending on context
The Physiology of the Senses Chapter 8 - Muscle Sense
 The Physiology of the Senses Chapter 8 - Muscle Sense www.tutis.ca/senses/ Contents Objectives... 1 Introduction... 2 Muscle Spindles and Golgi Tendon Organs... 3 Gamma Drive... 5 Three Spinal Reflexes...
The Physiology of the Senses Chapter 8 - Muscle Sense www.tutis.ca/senses/ Contents Objectives... 1 Introduction... 2 Muscle Spindles and Golgi Tendon Organs... 3 Gamma Drive... 5 Three Spinal Reflexes...
Cortical Organization. Functionally, cortex is classically divided into 3 general types: 1. Primary cortex:. - receptive field:.
 Cortical Organization Functionally, cortex is classically divided into 3 general types: 1. Primary cortex:. - receptive field:. 2. Secondary cortex: located immediately adjacent to primary cortical areas,
Cortical Organization Functionally, cortex is classically divided into 3 general types: 1. Primary cortex:. - receptive field:. 2. Secondary cortex: located immediately adjacent to primary cortical areas,
Neural Correlates of Human Cognitive Function:
 Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Thalamus and Sensory Functions of Cerebral Cortex
 Thalamus and Sensory Functions of Cerebral Cortex I: To describe the functional divisions of thalamus. II: To state the functions of thalamus and the thalamic syndrome. III: To define the somatic sensory
Thalamus and Sensory Functions of Cerebral Cortex I: To describe the functional divisions of thalamus. II: To state the functions of thalamus and the thalamic syndrome. III: To define the somatic sensory
Title. Author(s)Maezawa, Hitoshi; Tojyo, Itaru; Yoshida, Kazuya; Fuj. CitationJournal of Oral and Maxillofacial Surgery. Issue Date
Maezawa, Hitoshi; Tojyo, Itaru; Yoshida, Kazuya; Fuj. CitationJournal of Oral and Maxillofacial Surgery. Issue Date") Title Recovery of Impaired Somatosensory Evoked Fields aft Reconstruction Author(s)Maezawa, Hitoshi; Tojyo, Itaru; Yoshida, Kazuya; Fuj CitationJournal of Oral and Maxillofacial Surgery Issue Date 2016-07
Title Recovery of Impaired Somatosensory Evoked Fields aft Reconstruction Author(s)Maezawa, Hitoshi; Tojyo, Itaru; Yoshida, Kazuya; Fuj CitationJournal of Oral and Maxillofacial Surgery Issue Date 2016-07
Competing Streams at the Cocktail Party
 Competing Streams at the Cocktail Party A Neural and Behavioral Study of Auditory Attention Jonathan Z. Simon Neuroscience and Cognitive Sciences / Biology / Electrical & Computer Engineering University
Competing Streams at the Cocktail Party A Neural and Behavioral Study of Auditory Attention Jonathan Z. Simon Neuroscience and Cognitive Sciences / Biology / Electrical & Computer Engineering University
Body Organizations Flashcards
 1. What are the two main regions of the body? 2. What three structures are in the Axial Region? 1. Axial Region (Goes down midline of the body) 2. Appendicular Region (limbs) 3. Axial Region (Goes down
1. What are the two main regions of the body? 2. What three structures are in the Axial Region? 1. Axial Region (Goes down midline of the body) 2. Appendicular Region (limbs) 3. Axial Region (Goes down
Chapter 5. Summary and Conclusions! 131
 ! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
Functional topography of the low postcentral area
 J Neurosurg 97:388 395, 2002 Functional topography of the low postcentral area WARREN BOLING, M.D., DAVID C. REUTENS, M.D., AND ANDRÉ OLIVIER, M.D., PH.D. Montreal Neurological Institute and McConnell
J Neurosurg 97:388 395, 2002 Functional topography of the low postcentral area WARREN BOLING, M.D., DAVID C. REUTENS, M.D., AND ANDRÉ OLIVIER, M.D., PH.D. Montreal Neurological Institute and McConnell
Retinotopy & Phase Mapping
 Retinotopy & Phase Mapping Fani Deligianni B. A. Wandell, et al. Visual Field Maps in Human Cortex, Neuron, 56(2):366-383, 2007 Retinotopy Visual Cortex organised in visual field maps: Nearby neurons have
Retinotopy & Phase Mapping Fani Deligianni B. A. Wandell, et al. Visual Field Maps in Human Cortex, Neuron, 56(2):366-383, 2007 Retinotopy Visual Cortex organised in visual field maps: Nearby neurons have
Location of the Sensorimotor Cortex: Functional and Conventional MR Compared
 Location of the Sensorimotor Cortex: Functional and Conventional MR Compared F. Zerrin Yetkin, R. Anne Papke, Leighton P. Mark, David L. Daniels, Wade M. Mueller, and Victor M. Haughton PURPOSE: To determine
Location of the Sensorimotor Cortex: Functional and Conventional MR Compared F. Zerrin Yetkin, R. Anne Papke, Leighton P. Mark, David L. Daniels, Wade M. Mueller, and Victor M. Haughton PURPOSE: To determine
Clinically Available Optical Topography System
 Clinically Available Optical Topography System Clinically Available Optical Topography System 18 Fumio Kawaguchi Noriyoshi Ichikawa Noriyuki Fujiwara Yûichi Yamashita Shingo Kawasaki OVERVIEW: Progress
Clinically Available Optical Topography System Clinically Available Optical Topography System 18 Fumio Kawaguchi Noriyoshi Ichikawa Noriyuki Fujiwara Yûichi Yamashita Shingo Kawasaki OVERVIEW: Progress
The Topography of Tactile Working Memory
 The Journal of Neuroscience, October 15, 2001, 21(20):8262 8269 The Topography of Tactile Working Memory Justin A. Harris, Irina M. Harris, and Mathew E. Diamond Cognitive Neuroscience Sector, International
The Journal of Neuroscience, October 15, 2001, 21(20):8262 8269 The Topography of Tactile Working Memory Justin A. Harris, Irina M. Harris, and Mathew E. Diamond Cognitive Neuroscience Sector, International
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing
 Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
IJPHY ASYMMETRY OF SOMATOSENSORY CORTICAL PLASTICITY IN PATIENT WITH BILATERAL CARPAL TUNNEL SYNDROME ABSTRACT
 Int J Physiother. Vol 4(1), 1-5, February (2017) ISSN: 2348-8336 CASE STUDY IJPHY ABSTRACT ASYMMETRY OF SOMATOSENSORY CORTICAL PLASTICITY IN PATIENT WITH BILATERAL CARPAL TUNNEL SYNDROME *1 Hikmat Hadoush
Int J Physiother. Vol 4(1), 1-5, February (2017) ISSN: 2348-8336 CASE STUDY IJPHY ABSTRACT ASYMMETRY OF SOMATOSENSORY CORTICAL PLASTICITY IN PATIENT WITH BILATERAL CARPAL TUNNEL SYNDROME *1 Hikmat Hadoush
Human cortical gustatory areas: A review of functional neuroimaging data
 Brain imaging NeuroReport 10, 7±14 (1999) IN an effort to de ne human cortical gustatory areas we reviewed functional neuroimaging data for which coordinates standardized in Talairach proportional space
Brain imaging NeuroReport 10, 7±14 (1999) IN an effort to de ne human cortical gustatory areas we reviewed functional neuroimaging data for which coordinates standardized in Talairach proportional space
Voluntary Movements. Lu Chen, Ph.D. MCB, UC Berkeley. Outline. Organization of the motor cortex (somatotopic) Corticospinal projection
 Corticospinal projection") Voluntary Movements Lu Chen, Ph.D. MCB, UC Berkeley 1 Outline Organization of the motor cortex (somatotopic) Corticospinal projection Physiology of motor neurons Direction representation, population coding
Voluntary Movements Lu Chen, Ph.D. MCB, UC Berkeley 1 Outline Organization of the motor cortex (somatotopic) Corticospinal projection Physiology of motor neurons Direction representation, population coding
Shape and roughness activate different somatosensory areas in the human brain (positron-emission tomography intraparietal cortex shape perception
 Proc. Natl. Acad. Sci. USA Vol. 95, pp. 3295 3300, March 1998 Physiology Shape and roughness activate different somatosensory areas in the human brain (positron-emission tomography intraparietal cortex
Proc. Natl. Acad. Sci. USA Vol. 95, pp. 3295 3300, March 1998 Physiology Shape and roughness activate different somatosensory areas in the human brain (positron-emission tomography intraparietal cortex
Accepted to MICCAI Sources of Variability in MEG
 Accepted to MICCAI 27 Sources of Variability in MEG Wanmei Ou, Polina Golland, and Matti Hämäläinen 2 Department of Computer Science and Artificial Intelligence Laboratory, MIT, USA 2 Athinoula A. Martinos
Accepted to MICCAI 27 Sources of Variability in MEG Wanmei Ou, Polina Golland, and Matti Hämäläinen 2 Department of Computer Science and Artificial Intelligence Laboratory, MIT, USA 2 Athinoula A. Martinos
Neural Plasticity: Merzenich,Taub, and Greenough
 16 Neural Plasticity: Merzenich,Taub, and Greenough BY ERIN CLIFFORD Introduction REVIEW The study of neural plasticity has important implications for psychological development. Plasticity refers to the
16 Neural Plasticity: Merzenich,Taub, and Greenough BY ERIN CLIFFORD Introduction REVIEW The study of neural plasticity has important implications for psychological development. Plasticity refers to the
Cerebellum. Steven McLoon Department of Neuroscience University of Minnesota
 Cerebellum Steven McLoon Department of Neuroscience University of Minnesota 1 Anatomy of the Cerebellum The cerebellum has approximately half of all the neurons in the central nervous system. The cerebellum
Cerebellum Steven McLoon Department of Neuroscience University of Minnesota 1 Anatomy of the Cerebellum The cerebellum has approximately half of all the neurons in the central nervous system. The cerebellum
Pathways of proprioception
 The Autonomic Nervous Assess Prof. Fawzia Al-Rouq Department of Physiology College of Medicine King Saud University Pathways of proprioception System posterior column& Spinocerebellar Pathways https://www.youtube.com/watch?v=pmeropok6v8
The Autonomic Nervous Assess Prof. Fawzia Al-Rouq Department of Physiology College of Medicine King Saud University Pathways of proprioception System posterior column& Spinocerebellar Pathways https://www.youtube.com/watch?v=pmeropok6v8
Seeing through the tongue: cross-modal plasticity in the congenitally blind
 International Congress Series 1270 (2004) 79 84 Seeing through the tongue: cross-modal plasticity in the congenitally blind Ron Kupers a, *, Maurice Ptito b www.ics-elsevier.com a Center for Functionally
International Congress Series 1270 (2004) 79 84 Seeing through the tongue: cross-modal plasticity in the congenitally blind Ron Kupers a, *, Maurice Ptito b www.ics-elsevier.com a Center for Functionally
Human Paleoneurology and the Evolution of the Parietal Cortex
 PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
Regional and Lobe Parcellation Rhesus Monkey Brain Atlas. Manual Tracing for Parcellation Template
 Regional and Lobe Parcellation Rhesus Monkey Brain Atlas Manual Tracing for Parcellation Template Overview of Tracing Guidelines A) Traces are performed in a systematic order they, allowing the more easily
Regional and Lobe Parcellation Rhesus Monkey Brain Atlas Manual Tracing for Parcellation Template Overview of Tracing Guidelines A) Traces are performed in a systematic order they, allowing the more easily
Benefit of Simultaneous Recording of EEG and MEG in Dipole Localization
 Epilepsia, 43(8):924 928, 2002 Blackwell Publishing, Inc. International League Against Epilepsy Benefit of Simultaneous Recording of EEG and MEG in Dipole Localization *Harumi Yoshinaga, *Tomoyuki Nakahori,
Epilepsia, 43(8):924 928, 2002 Blackwell Publishing, Inc. International League Against Epilepsy Benefit of Simultaneous Recording of EEG and MEG in Dipole Localization *Harumi Yoshinaga, *Tomoyuki Nakahori,
CEREBRUM. Dr. Jamila EL Medany
 CEREBRUM Dr. Jamila EL Medany Objectives At the end of the lecture, the student should be able to: List the parts of the cerebral hemisphere (cortex, medulla, basal nuclei, lateral ventricle). Describe
CEREBRUM Dr. Jamila EL Medany Objectives At the end of the lecture, the student should be able to: List the parts of the cerebral hemisphere (cortex, medulla, basal nuclei, lateral ventricle). Describe
Motor Functions of Cerebral Cortex
 Motor Functions of Cerebral Cortex I: To list the functions of different cortical laminae II: To describe the four motor areas of the cerebral cortex. III: To discuss the functions and dysfunctions of
Motor Functions of Cerebral Cortex I: To list the functions of different cortical laminae II: To describe the four motor areas of the cerebral cortex. III: To discuss the functions and dysfunctions of
CEREBRUM Dr. Jamila Elmedany Dr. Essam Eldin Salama
 CEREBRUM Dr. Jamila Elmedany Dr. Essam Eldin Salama Objectives At the end of the lecture, the student should be able to: List the parts of the cerebral hemisphere (cortex, medulla, basal nuclei, lateral
CEREBRUM Dr. Jamila Elmedany Dr. Essam Eldin Salama Objectives At the end of the lecture, the student should be able to: List the parts of the cerebral hemisphere (cortex, medulla, basal nuclei, lateral
Exam 1 PSYC Fall 1998
 Exam 1 PSYC 2022 Fall 1998 (2 points) Briefly describe the difference between a dualistic and a materialistic explanation of brain-mind relationships. (1 point) True or False. George Berkely was a monist.
Exam 1 PSYC 2022 Fall 1998 (2 points) Briefly describe the difference between a dualistic and a materialistic explanation of brain-mind relationships. (1 point) True or False. George Berkely was a monist.
Department of Cognitive Science UCSD
 Department of Cognitive Science UCSD Verse 1: Neocortex, frontal lobe, Brain stem, brain stem, Hippocampus, neural node, Right hemisphere, Pons and cortex visual, Brain stem, brain stem, Sylvian fissure,
Department of Cognitive Science UCSD Verse 1: Neocortex, frontal lobe, Brain stem, brain stem, Hippocampus, neural node, Right hemisphere, Pons and cortex visual, Brain stem, brain stem, Sylvian fissure,
Selective Attention to Face Identity and Color Studied With fmri
 Human Brain Mapping 5:293 297(1997) Selective Attention to Face Identity and Color Studied With fmri Vincent P. Clark, 1 * Raja Parasuraman, 2 Katrina Keil, 1 Rachel Kulansky, 1 Sean Fannon, 2 Jose Ma.
Human Brain Mapping 5:293 297(1997) Selective Attention to Face Identity and Color Studied With fmri Vincent P. Clark, 1 * Raja Parasuraman, 2 Katrina Keil, 1 Rachel Kulansky, 1 Sean Fannon, 2 Jose Ma.
Differential Activation in Somatosensory Cortex for Different Discrimination Tasks
 The Journal of Neuroscience, January 1, 2000, 20(1):446 450 Differential Activation in Somatosensory Cortex for Different Discrimination Tasks Christoph Braun, 1 Renate Schweizer, 1 Thomas Elbert, 2 Niels
The Journal of Neuroscience, January 1, 2000, 20(1):446 450 Differential Activation in Somatosensory Cortex for Different Discrimination Tasks Christoph Braun, 1 Renate Schweizer, 1 Thomas Elbert, 2 Niels
b. The groove between the two crests is called 2. The neural folds move toward each other & the fuse to create a
 Chapter 13: Brain and Cranial Nerves I. Development of the CNS A. The CNS begins as a flat plate called the B. The process proceeds as: 1. The lateral sides of the become elevated as waves called a. The
Chapter 13: Brain and Cranial Nerves I. Development of the CNS A. The CNS begins as a flat plate called the B. The process proceeds as: 1. The lateral sides of the become elevated as waves called a. The
Neuroimaging. BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT. Human Brain Mapping
 11/8/2013 Neuroimaging N i i BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT 2 Human Brain Mapping H Human m n brain br in m mapping ppin can nb
11/8/2013 Neuroimaging N i i BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT 2 Human Brain Mapping H Human m n brain br in m mapping ppin can nb
CHAPTER 10 THE SOMATOSENSORY SYSTEM
 CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could
CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could