Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice. Deshpande, PhD 1*
|
|
|
- Vivian Lynch
- 5 years ago
- Views:
Transcription
1 Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice Pawan Sharma, PhD 1,2, Roslyn Yi, BSc 1, Ajay P Nayak, PhD 1, Nadan Wang, MSc 1, Francesca Tang, MSc 2, Morgan J Knight, PhD 3, Shi Pan, PhD 1, Brian Oliver, PhD 2, and Deepak A Deshpande, PhD 1* 1 Department of Medicine, Center for Translational Medicine, Thomas Jefferson University, Philadelphia, PA, USA, Respiratory Research Group, Woolcock Institute of Medical Research and School of Life Sciences, University of Technology, Sydney, NSW, Australia, Department of Experimental Therapeutics, University of Texas MD Anderson Cancer Center, Houston, TX, * Address for Correspondence: Deepak A. Deshpande, Ph.D. Associate Professor, Center for Translational Medicine, Jeff Alumni Hall, Rm 543, 1020 Locust Street, Philadelphia, PA Phone: Fax: deepak.deshpande@jefferson.edu
2 ABSTRACT Asthma is characterized by airway inflammation, mucus secretion, remodeling and hyperresponsiveness (AHR). Recent research has established the bronchodilatory effect of bitter taste receptor (TAS2R) agonists in various models. Comprehensive pre-clinical studies aimed at establishing effectiveness of TAS2R agonists in disease models are lacking. Here we aimed to determine the effect of TAS2R agonists on features of asthma. Further, we elucidated a mechanism by which TAS2R agonists mitigate features of asthma. Asthma was induced in mice using intranasal house dust mite or aerosol ova-albumin challenge, and chloroquine or quinine were tested in both prophylactic and treatment models. Allergen challenge resulted in airway inflammation as evidenced by increased immune cells infiltration and release of cytokines and chemokines in the lungs, which were significantly attenuated in TAS2R agonists treated mice. TAS2R agonists attenuated features of airway remodeling including smooth muscle mass, extracellular matrix deposition and pro-fibrotic signaling, and also prevented mucus accumulation and development of AHR in mice. Mechanistic studies using human neutrophils demonstrated that inhibition of immune cell chemotaxis is a key mechanism by which TAS2R agonists blocked allergic airway inflammation and exerted anti-asthma effects. Our comprehensive studies establish the effectiveness of TAS2R agonists in mitigating multiple features of allergic asthma.
3 INTRODUCTION Asthma is a chronic inflammatory disease of the airways affecting over 300 million people worldwide 1 and characterized by airway inflammation, hyperresponsiveness, mucus hypersecretion and remodeling 2-4. The chronic and persistent inflammation leads to structural and functional changes in the resident airway cells 4-7. G protein-coupled receptors (GPCRs) play a vital role in the regulation of airway inflammation, and airway smooth muscle (ASM) contraction, relaxation, and proliferation 8,9. Therefore, GPCR agonists or antagonists comprise mainstay asthma therapies. Identifying a novel class of GPCRs is one of the approaches to developing newer and effective anti-asthma drugs. We recently identified expression of bitter taste receptors (TAS2Rs) on human ASM and demonstrated that stimulation with TAS2R agonists results in relaxation of ASM 10. Other laboratories confirmed the expression of TAS2Rs and the bronchodilatory effect of TAS2R agonists in mouse 11,12, human and guinea pig 16 airways. Administration of TAS2R agonists into the mouse lungs results in bronchodilation in both naïve and allergen-exposed animals 10, and TAS2R expression, signaling, and ASM relaxation and bronchodilatory effects are maintained under inflammatory conditions in human asthmatic ASM cells and intact lung slices 17. TAS2Rs have thus emerged as novel targets in the treatment of obstructive airway diseases such as asthma. Numerous other properties of TAS2Rs including the availability of wellestablished natural and synthetic agonists make TAS2Rs an attractive target (reviewed in 18,19 ). Asthma is a complex disease involving multiple pathogenic features including inflammation, mucus secretion, remodeling and bronchoconstriction. We have previously shown that TAS2R agonists are potent bronchodilators, yet their effect on key features of asthma remains undetermined. Because beta-agonists exhibit little or no anti-inflammatory activity, and
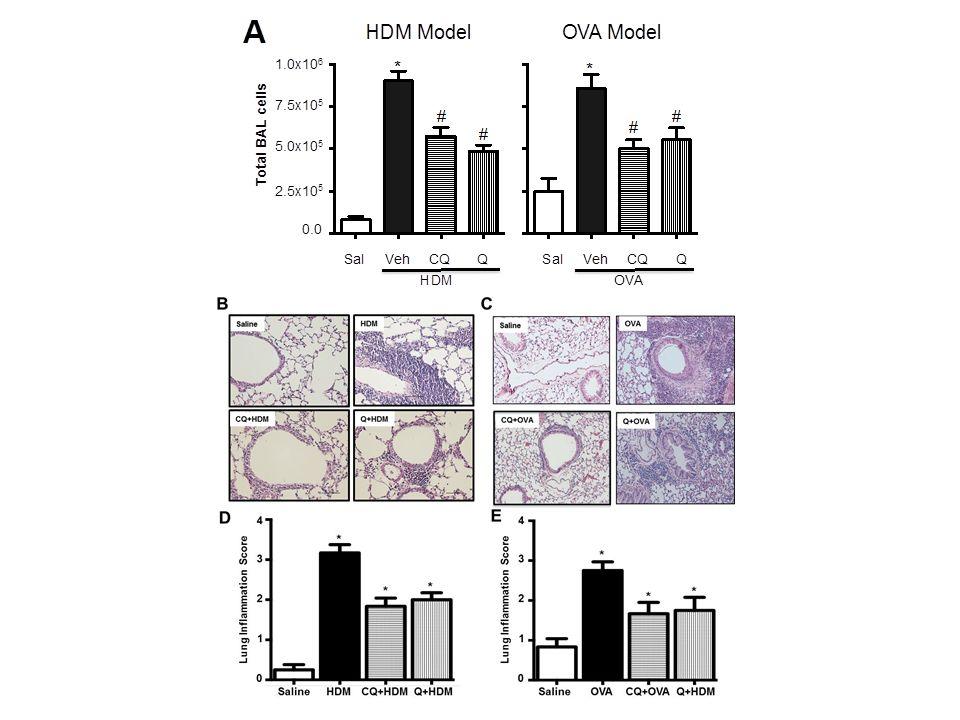
4 no current asthma drugs appear effective in deterring airway remodeling, a multi-tasking asthma drug capable of inhibiting bronchoconstriction, airway inflammation, and airway remodeling would constitute a novel therapeutic addressing an important, unmet clinical need. Here we used two different mouse models of allergic asthma to assess the effect of TAS2R agonists on key features of asthma. Our results demonstrate that TAS2R agonists are effective in not only reducing AHR, but also in preventing allergic airway inflammation and multiple features of airway remodeling. RESULTS Mice were pretreated with TAS2R agonists, chloroquine or quinine by intranasal (HDM model) or aerosol (OVA model) route, and challenged with either HDM or OVA as described in Methods and illustrated in Figure 1. Twenty-four hours post-final challenge, lung function, airway inflammation, and features of airway remodeling were assessed. Aerosol characterization of chloroquine showed more than 60% of aerosolized drug was contained in particles of sufficient size (< 3.3 µm) to reach and deposit in the lower airways (Fig. 1C) 20. Aerosolization of a 1.5 mg/ml chloroquine solution produced a mean aerosol concentration of 2.6 µg/l, which corresponds to a 500 ng dose per 30 minutes treatment. Effect of TAS2R agonists on allergic airway inflammation HDM and OVA challenge resulted in a robust increase in immune cell infiltration in the airways as demonstrated by a significant increase in cell count in the BAL compared to controls. Pretreating mice with TAS2R agonists chloroquine or quinine significantly (p<0.05) reduced infiltration of inflammatory cells (Fig. 2A). This was further confirmed by histological evaluation of H&E stained lung sections (Fig. 2B-E). Differential cell count revealed infiltration of
5 eosinophils, neutrophils and to a lesser extent accumulation of lymphocytes and macrophages in the BAL. TAS2R agonists significantly (p<0.05) inhibited infiltration of eosinophils, neutrophils and macrophages (Table 1). HDM and OVA challenge also caused a significant increase in multiple cytokines and chemokines in the lung, and TAS2R agonists inhibited this induction to different degrees (Table 2). HDM exposure in mice specifically led to significant increases in the levels of pro-eosinophilic-th2 cytokines (IL-4, IL-5, IL-9 and IL-13), pro-inflammatory cytokines eotaxin, KC, TNF-α, IP-10 and RANTES as well as IL-17, a monocyte and neutrophil chemo-attractant to the tissue and the anti-inflammatory cytokine IL-10. TAS2R agonists differentially inhibited the induction of most cytokines and chemokines, with quinine being more effective than chloroquine in global suppression of inflammation. In OVA-exposed mice there was a significant induction of IL-4, IL-5, IL-13, IL-17, eotaxin, KC, all of which were significantly suppressed by TAS2R agonists pretreatment (Table 2). Similarly OVA induced TNF-α, IP-10 and RANTES, yet TAS2R agonist pretreatment failed to significantly repress the induction of these agents. Collectively, these results indicate that TAS2R agonists demonstrate anti-inflammatory activity in the lung, albeit with differential efficacy with respect to specific cytokine suppression and the allergen employed. Effect of TAS2R agonists on features of airway remodeling The effect of TAS2R agonists on allergen-induced airway remodeling was assessed by determining expression of marker proteins in the BAL or lung lysates and by histology. Collagen deposition as assessed by trichrome staining of lung sections was significantly lower in lungs of mice treated with TAS2R agonists (Fig. 3A-D). Secreted collagen was significantly higher in allergen-challenged animals, and chloroquine or quinine pretreatment significantly (p<0.05)
6 inhibited this response (Fig. 3F). Allergen-induced expression of TGF-β1 (a marker of profibrotic remodeling) was markedly inhibited by chloroquine and quinine (Fig. 3G). TGF-β action on airway cells involves phosphorylation of Smad2, and immunoblot analysis of lung lysates revealed inhibition of allergen-induced increase in phospho-smad2 by TAS2R agonists (Fig. 3E). Finally, immunoblot analyses of whole lung lysates revealed increased expression of fibronectin (Fig. 3H-I) in HDM- or OVA-exposed mice, which was inhibited by pretreatment with TAS2R agonists. Immunofluorescence staining of sm-α-actin in lung sections revealed increased smooth muscle around the airways in HDM or OVA challenged animals, and TAS2R agonists significantly (p<0.05) inhibited this induction (Fig. 4A-D). Moreover, the accompanying allergen-induced increase in smmhc, sm-α-actin, and calponin expression in lung lysates was similarly inhibited by chloroquine and quinine pretreatment in HDM (Fig. 4E-H) and OVA (Fig. 4I-L) models of allergic asthma. Increased mucus accumulation is another key feature of asthma. HDM or OVA challenge resulted in excessive accumulation of mucus in the airways (assessed by PAS staining), and pretreatment with TAS2R agonists inhibited this accumulation (Fig. 5A-D). Matrix metalloproteinases (MMPs) have been implicated in asthmatic airway remodeling. HDM or OVA challenge resulted in robust induction of MMP-8 (neutrophil collagenase), pro-mmp-9 (gelatinase) and MMP-12 (macrophage metalloelastase) in BAL fluid, when compared to salinechallenged mice (Fig 5E-J). In the HDM model, TAS2R agonist quinine was more effective than chloroquine in blocking induction of MMP-8 and MMP-12 while both TAS2R agonists blocked pro-mmp-9 induction. In the OVA model, chloroquine was more effective than quinine in blocking induction of MMP-8 and MMP-12, whereas both TAS2R agonists blocked pro-mmp-9
7 induction. These results suggest that TAS2R agonists differentially regulate MMPs induction in models of allergic asthma. Effect of TAS2R agonists on airway hyperresponsiveness Airway responsiveness to MCh challenge was determined using anesthetized, intubated and ventilated mice. HDM challenge resulted in an increased bronchoconstrictor response to MCh, and pretreatment with TAS2R agonists inhibited development of AHR (Fig. 6A). A similar but slightly increased bronchoconstrictor response occurred in OVA-challenged mice, and again this was reversed by pretreatment with chloroquine or quinine (Fig. 6B). Furthermore, aerosolized chloroquine (250 µg/ml) and quinine (150 µg/ml) (administered after the highest concentration of MCh) were able to provide effective bronchorelaxation as seen by significant reduction in airway resistance (R) in both HDM- and OVA- challenged mice (Fig. 6C-D). Effect of TAS2R agonists on allergic airway inflammation in treatment model The ability of TAS2R agonists in attenuating various features of allergic airway inflammation was further evaluated in a treatment model. Allergic inflammation was established in mice via intranasal administration of HDM for 3 weeks and following this, for next 2 weeks mice were treated with TAS2R agonists, chloroquine or quinine by intranasal route, and challenged with HDM (Fig. 7A). Treatment of mice with TAS2R agonists resulted in reduced infiltration of the inflammatory cells in the airways and attenuation of histopathological changes in the lung tissue (Fig. 7B and C). Significant reductions were observed in accumulation of eosinophils, neutrophils and to lesser extent macrophages in the airways following treatment with TAS2R agonists (Table 1). Furthermore, treatment with TAS2R agonists resulted in significant reduction in levels of allergen-induced cytokines such as eotaxin, KC, IP-10 and RANTES in the
8 airways (Table 3). Repeated instillation of HDM in the airways for 5 weeks resulted is extensive collagen deposition around the conducting airways (Fig. 7C), which was attenuated by treatment with TAS2R agonists. Furthermore, soluble collagen levels in lung tissue lysates were also significantly lower following treatment with TAS2R agonists (Fig. 7D). Pro-fibrotic cytokine TGF-β1 expression was prominent in mice exposed with HDM and its expression was attenuated in mice treated with TAS2R agonists (Table 3). In concert, we also observed inhibition of HDMinduced accumulation of phospho-smad2 as well as fibronectin levels by treatment with TAS2R agonists (Fig. 7E). Treatment of mice with quinine also resulted in reduced expression of sm-αactin, smmhc and calponin in the lung tissue compared to untreated mice (Fig. 7F and G). Comparable effects were observed following treatment with chloroquine, although calponin expression was not affected. Treatment with TAS2R agonists also resulted in fewer PAS + mucusproducing cells in the bronchi and bronchioles (Fig. 7H). Finally, treatment with TAS2R agonists resulted inhibition of dose-dependent MCh-induced bronchoconstrictor response in HDMchallenged mice (Fig. 7I). Mechanism of action of TAS2R agonists As TAS2R receptor agonists exhibited protective effects on airway inflammation through attenuated expression of cytokines and chemokines and significant reduction in allergen-induced immune cell influx into the mouse lungs, we investigated the potential mechanism for the broad anti-inflammatory action of TAS2R agonists. Using peripheral blood neutrophils from healthy human volunteers we studied the effect of TAS2R agonists on CXCL8 (IL-8)-mediated chemotaxis of immune cells (neutrophils) (Fig. 8). Our results demonstrate dose-dependent reduction in the immune cell recruitment to a chemotactic gradient (CXCL8, 100 ng/ml, 90 min) by both TAS2R agonists chloroquine and quinine; the effect being more pronounced and
9 significant at 500 µm and 1 mm respectively. Our previous study using human ASM cells suggest that TAS2R agonists inhibit mitogeninduced ASM growth 21. The anti-remodeling effect of TAS2R agonists observed in this study may involve anti-mitogenic effect of TAS2R agonists. TAS2R agonists induce efficacious bronchodilation in murine models and human ASM cells/tissues. The bronchodilatory effect may account for an attenuated AHR in animals treated with TAS2R agonists compared to allergen-challenged animals. DISCUSSION Recent studies have established the expression of TAS2R on extraoral tissues including ASM. Most importantly, stimulation of airways with a variety of TAS2R agonists results in relaxation of airways and bronchodilation. Based on this observation TAS2Rs have emerged as novel, promising targets for treating obstructive lung diseases such as asthma. To further validate the potential use of TAS2Rs as anti-asthma drug targets, we assessed the effectiveness of two different bitter ligands (chloroquine and quinine) delivered by two different routes (intranasal and aerosol) in murine models challenged with two different allergens (OVA and HDM). Aerosol quantification further confirmed the feasibility of delivering chloroquine to lower airways by aerosol route. Our findings suggest that TAS2R agonists mitigate allergen-induced airway inflammation, remodeling, mucus secretion and AHR in murine models of asthma. TAS2Rs are known to be activated by a variety of structurally diverse natural and synthetic compounds. Most of the TAS2Rs are broadly tuned in terms of agonist activation. Some of the commonly used bitter tastants include chloroquine, quinine, denatonium, aristocholic acid, saccharine, and noscapine. We investigated the effect of two TAS2R agonists, chloroquine and quinine on asthma in murine models. The receptor subtype specificity of these two bitter
10 tastants has been established using a heterologous expression system 22. Chloroquine and quinine bind to TAS2R3, 7, 10, and 39, and TAS2R4, 7, 10, 14, 39, 40, 43, 44 and 46, respectively. Although TAS2R expression is found on multiple resident airway cells and on immune cells, the subtype of TAS2Rs expressed on different types of cells is different. Using broad-tuned ligands allowed us to discern the effect of bitter tastants on multiple features of asthma pathogenesis in mice. TAS2R expression was demonstrated in human leukocytes primarily on lymphocytes (11 different subtypes with 3 of them being expressed at significantly high levels in asthmatics) and mast cells (9 different subtypes). Stimulation of blood leukocytes with TAS2R agonists resulted in inhibition of LPS-induced release of IL-13, IL-4, IL-5, TNF-α, IL-1β and IFN-γ, all of which are known to play an important role in asthma 23. TAS2R agonists chloroquine and denatonium inhibited IgE-induced activation of mast cells and degranulation reflected by the inhibition of release of PGD2 and histamine 24. Mast cell activation is known to contribute to asthma pathogenesis and AHR. These two studies demonstrate the potential anti-inflammatory effect mediated via activation of TAS2Rs on immune cells. Using two different murine models of allergic asthma, we demonstrate significant and clinically relevant inhibition of immune cell infiltration as well cytokines/chemokines release in the lung by TAS2R agonists. Our findings also demonstrate a differential regulation of cytokines and chemokines release by HDM and OVA in mice. Of note, the anti-inflammatory effects of TAS2R agonists have been demonstrated in the upper airways and respiratory epithelium in chronic rhinosinusitis 25,26. More importantly our studies using both animal models of asthma, and mechanistic studies in humans in vitro clearly demonstrate that TAS2R agonists dampen global allergen-induced airway inflammation
11 and have broader application in terms of reducing dependence on inhaled steroids, mainstay maintenance therapy for asthma. A genomics study recently demonstrated that TAS2R expression is increased in peripheral blood leukocytes, and the expression level of TAS2R is correlated with severity of the disease in asthmatics (17). Should such an upregulation of TAS2R with asthma extend to other cells/tissues including ASM, this disease feature would further favor the use of TAS2R agonists as asthma therapeutics given receptor upregulation would help protect against receptor desensitization which is believed to limit the therapeutic efficacy of anti-asthma GPCR agonists, including the beta-2-adrenoceptor 27,28. Our findings in both the HDM and OVA models suggest a significant inhibition of mucus accumulation in the airways when treated with TAS2R agonists. Shah et al. 29 demonstrated that TAS2Rs are expressed on ciliary epithelium of airways and stimulation with bitter tastants results in increased ciliary beat frequency and hence might hasten mucus clearance. Gene expression data from total RNA isolated from murine lung revealed inhibition of allergen-induced upregulation of MUC-5 gene by TAS2R ligands. Therefore, inhibition of mucus synthesis and enhancement of clearance represent a means by which bitter tastants mediate the inhibition of mucus accumulation in the lung we observed herein on two different mouse models. Our results demonstrate that TAS2R agonists inhibit allergen-induced airway remodeling characterized by deposition of extracellular matrix proteins such as collagen and fibronectin. Interestingly, ASM mass was also decreased in the animals treated with TAS2R agonists. A recent study from our laboratory demonstrated that TAS2R agonists chloroquine, quinine and saccharine inhibit mitogen-induced growth of cultured human ASM cells in a dose-dependent fashion 21. These observations are valuable considering the fact the current anti-asthma medications are effective in either inhibiting airway inflammation or reversing
12 bronchoconstriction, but have limited or no utility in treating airway remodeling, a important clinical feature in chronic asthmatics 5-7,30. ASM mass is directly correlated to asthma symptoms in humans 31,32. Beta-agonists, the mainstay rescue medications in asthma, modestly inhibit ASM growth in vitro. Activation of PKA is central to ASM relaxation and ASM cell growth inhibition by beta agonists 9. TAS2R agonists on the other hand do not activate camp/pka pathway, yet induce a robust bronchodilatory and anti-proliferative effect on airways 21. These studies suggest TAS2R agonists activate novel signal transduction mechanisms in airways cells to promote anti-asthma effects. Allergen challenge in murine models resulted in the secretion of TGF-β, MMPs and activation of pro-fibrotic signaling (e.g. p-smad); treating animals with TAS2R agonists inhibited all of these features. These findings suggest inhibition of gene expression by TAS2R agonists in airways cells. We recently demonstrated that the anti-mitogenic effect of TAS2R agonists on human ASM cells involves inhibition of gene expression required for cell cycle progression 21. Lung function measurements in allergen-challenged mice demonstrate the ability of TAS2R agonist pretreatment to inhibit the induction of AHR caused by either HDM or OVA. AHR occurs not only due to calcium sensitization of ASM during inflammation, but is also promoted by structural and functional changes in airway cells upon allergen exposure. Herein we demonstrate that not only do TAS2Rs function as effective acute bronchodilators, but can also mitigate features of allergen-induced airway inflammation and remodeling that contribute to AHR. In summary, our studies using two different murine model of asthma demonstrate that TAS2R agonists chloroquine and quinine administered by aerosol or intranasal route inhibit allergen-induced airway inflammation, remodeling, mucus production and AHR, four cardinal
13 features of human asthma. TAS2Rs are known to be ubiquitously expressed and regulate a variety of cellular functions. Although several natural and synthetic compounds with known pharmacological characteristics are known to activate TAS2Rs, future studies are needed to develop TAS2R agonists and antagonists with high affinity and high subtype specificity. Alternately, existing drugs could be repurposed for asthma therapy. Advances in medicinal chemistry and computational modeling should catalyze the drug discovery process in exploiting TAS2Rs as novel anti-asthma therapeutic target. MATERIALS AND METHODS Chemicals and reagents Alexa Fluor-conjugated secondary antibodies were obtained from Life Technologies (Carlsbad, CA), IRDye-conjugated anti-mouse and anti-rabbit secondary antibodies were obtained from LI-COR Biosciences (Lincoln, NE). Mouse anti-smmhc, anti-sm-α-actin, anticalponin, rabbit anti-fibronectin were obtained from Sigma (St. Louis, MO) and anti-phospho Smad2 and anti-smad2 antibodies were purchased from Cell Signaling (Danvers, MA). Sircol collagen assay was obtained from Biocolor Life Sciences, UK. Staining kits (PAS, Trichrome), chloroquine, quinine and saccharine were obtained from Sigma. H&E and Afog staining kits were obtained from Vector Labs. All other chemicals were of analytical grade. Human volunteer recruitment The study was approved by the Human Research Ethics Committee, The University of Sydney prior to commencement. All methods were performed in accordance with the relevant guidelines and regulations of the institution. Participants were required to be over 18 years of age. Exclusion criteria included if they were asthmatic, pregnant, known to faint during venipuncture
14 procedures or had a bloodborne infection or condition. All patients provided written informed consent, and basic demographic information was collected. Mouse models of allergic asthma All animal procedures were approved by the Institutional Animal Care Committee of Thomas Jefferson University and University of Maryland, Baltimore. All methods were performed in accordance with the relevant guidelines and regulations of the institutions. Animal surgeries were performed under tribromoethanol (Avertin, 250 mg/kg) anesthesia. Prophylactic model - Female BALB/c mice at 8 weeks were intranasally challenged five days a week for three consecutive weeks with 25 µg of house dust mite (HDM) extract (Dermatophagoides pteronyssinus, Greer Labs, USA) in 35 µl saline (Fig. 1A). A select set of mice were administered either chloroquine (50 mg/kg), quinine (10 mg/kg) or vehicle (1% ethanol) in 25 µl volume by intranasal route 30 min prior to the HDM challenges. Similarly, mice were sensitized by injecting 2 mg alum in 0.2 ml PBS or alum with 100 µg ovalbumin (OVA) per mouse on day 1 and day 14 intraperitoneally (Fig. 1B). Mice were then challenged with aerosolized sterile PBS with or without 1% OVA on days 19, 21, 23, 25 and 27 for 30 min. Select sets of animals were treated with 1.5 mg/ml chloroquine or quinine for 30 min by aerosol prior to OVA challenge. Twenty-four hours after the last challenge, lung function measurements were performed, bronchoalveolar lavage (BAL) fluid was collected, fresh or formalin-fixed lungs were harvested. Treatment model Female BALB/c mice (8 weeks old) were initially challenged 5 days a week for 3 weeks via intranasal administration of 25 µg of HDM extract in 35 µl saline. Following this, mice were treated with chloroquine (50 mg/kg), quinine (10 mg/kg) or vehicle (1% ethanol) in 25 µl volume by intranasal route 30 min prior to subsequent HDM challenges for additional 2
15 weeks. Twenty-four hours following the last challenge, lung function measurements were performed, bronchoalveolar lavage (BAL) fluid was collected and fresh or formalin-fixed lungs were harvested for downstream analyses. Aerosol characterization and quantification Chloroquine was dissolved in water to a concentration of 1.5 mg/ml. Drug was nebulized by an AirLife Misty Max 10 nebulizer with 10 liter/min of air and captured by an Anderson Cascade Impactor (3M) or glass impinger (Ace) according to standard practice. Collected samples and standards were then separated on a Luna 3u Phenyl-Hexyl column (Phenomenex), peaks were ionized (Acquity TQD) and analyzed using Mass Lynx (v4.1). Assessment of BAL cellularity BAL samples were subjected to centrifugation and cell pellet resuspended in 1 ml PBS. Total cell count in BAL samples was determined by hemocytometer and data expressed as cells/ml. The BAL cells after centrifugation were stained with Hema-3 staining kit (Fisher Scientific) and differential cell count was determined by brightfield microscope. Measurement of BAL cytokines and chemokines The content of cytokines and chemokines in BAL fluids were measured by Multiplexing LASER Bead Technology (Eve Technologies, Calgary, Canada) using a 31-Plex-mouse cytokine/chemokine array (Cat #: MD31) and expressed as pg/ml. Preparation of Lung Lysates Mouse lungs were cut into small pieces in 250 µl of lysis buffer (40 mm Tris, 150 mm NaCl, 1% IgepalCA-630, 1% deoxycholic acid, 1 mm NaF, 5mM β-glycerophosphate, 1 mm Na 3 VO 4, 10 µg/ml aprotinin, 10 µg/ml leupeptin, 7 µg/ml pepstatin A, 1 mm PMSF, ph 8.0).
16 Lung tissues were homogenized using a polytron. The lysate was centrifuged (760 g, 5 min) and the supernatant stored at -80 C for subsequent analyses. Immunoblotting Protein concentration was determined by Pierce BCA assay kit (Thermo Scientific, Rockford, IL) and subjected to immunoblot analysis using protocols described previously 10,33 with primary antibodies noted above (1:1000), followed by incubation with respective secondary antibodies (LI-COR Biosciences) (1:5000). Immunoblots were visualized and bands quantified using the Odyssey infrared imaging system (LI-COR Biosciences). Soluble Collagen Assay Total soluble collagen content in the lung lysates was assessed using Sircol collagen assay (Biocolor, UK) according to the manufacturer's protocol as per 34. Collagen assay was performed by mixing lung homogenates with Sircol Dye reagent and measuring absorbance using a plate reader. Collagen content was quantified using a standard curve generated by reference standards and was normalized to the total lung protein content in each sample. Histopathological analysis Paraffin embedded tissues were used for histological evaluation as described previously 33,35-37, using 5 µm sections mounted on Superfrost Plus slides. Slides were deparaffinized, rehydrated and subjected to heat-induced epitope retrieval. The sections after blocking were stained with the primary antibody (1:100 anti-sm-α-actin) at 4 C overnight followed by staining with 1:250 goat anti-mouse Alexa Fluor conjugated secondary antibody at room temperature for 1 hour. The sections were washed in PBS and stained with Draq5 for 15 minutes. Finally, the sections were washed, mounted with Prolong Antifade (Molecular Probes, USA), and fluorescent imaging performed using an Olympus BX-51 fluorescent microscope. The lung tissues fixed in
17 10% formalin, embedded in paraffin were cut and stained with H&E, trichrome blue, AFOG and PAS staining using a standard histological protocol 36. Image acquisition and analysis was performed using a brightfield microscope. Lung inflammatory changes were graded using a semi-quantitative scoring system based on the following parameters: peribronchial inflammation and degree of total cell infiltration. Peribronchial inflammation was graded on a scale of 0 4 with 0, absent; 1, slight; 2, mild; 3, moderate; and 4, severe. The degree of cell infiltration was scored of 0 3 with 0, no cells; 1, few cells; 2, moderate influx of cells; and 3, extensive influx of cells. The total lung inflammation score was expressed as the sum of the scores for each parameter. Similarly, airway fibrosis in the formalin-fixed lung tissue was analyzed using AFOG or Trichrome stain and scored by using a scoring system for measuring subepithelial fibrosis (increased content of ECM protein such as collagen, stains blue with AFOG and Trichrome). A scoring range of 0-3 was used with 0, none; 1, infrequent; 2, common; and 3, widespread; blue staining in the airway wall. Peripheral blood neutrophil isolation and migration assay Neutrophils were isolated from peripheral blood collected from volunteers as previously described Briefly, 40 ml of blood was mixed with 10 ml acid citrate dextrose (ACD), 10 ml of phosphate buffered saline (PBS) (Gibco, Carlsbad, CA, USA), and 6 ml of 10% dextran (MP Biomedicals, Santa Ana, USA) and left for 20 minutes for sedimentation to occur at room temperature. The top layer was removed, overlaid on Ficoll Paque-PLUS (GE Healthcare, Little Chalfont, UK), and centrifuged at 490 g for 10 minutes. The supernatant was discarded and the cell pellet of granulocytes was resuspended in sterile water for 30 seconds to lyse remaining red blood cells before osmolarity was reestablished with equal parts of 2x PBS. Cells were then incubated for 30 minutes at 4 C with CD16 magnetic beads (Miltenyi Biotec, Bergisch,
18 Germany) before running through a magnetic column as per the manufacturer's instructions. Previous optimization of the protocol showed typical purity was 99% or greater by a haematoxylin and eosin stain. Neutrophils were resuspended in 1% FBS, 1% 1 M HEPES, and 1% penicillin/streptomycin RPMI 1640 at cells/ml and were co-incubated (250,000 cells/well) with TAS2R agonists (chloroquine and quinine; 100 µm, 500 µm and 1mM) in the upper well of the Transwell inserts (Corning, Corning, New York, USA) while CXCL8 (100 ng/ml, R&D Biosciences, USA) was added to stimulate cell chemotaxis in the lower well. Following an incubation period (90 min at 37 C, 5% CO 2 ), the cells that have migrated through the membrane were stained with trypan blue and manually counted. Measurement of Lung Mechanics Lung mechanics was measured using a flexivent system (Scireq, Canada) as described previously 10,35,36,41. Anesthetized and intubated mouse was ventilated with a tidal volume of 250 µl at 150 breaths/minute. A PEEP of 3 cm H 2 O was used for all studies. Mice were subjected to an increased dose of nebulized methacholine (MCh) challenge protocol using an in-line ultrasonic nebulizer. Using low frequency forced oscillation technique, respiratory mechanical input impedance (Zrs) was derived from the displacement of the ventilator's piston and the pressure in its cylinder. By fitting Zrs to the constant phase model, flexivent software calculated airway resistance (R), which was normalized to body weight. Change in lung resistance (R) was recorded for each dose of MCh. Animals were treated with aerosolized chloroquine (250 µg/ml) or quinine (150 µg/ml) at the end of MCh challenge and change in lung resistance recorded. Data analyses Values reported for all data represent means ± standard error of means (SEM). For all studies, replicate data from at least 8-11 mice were obtained. The statistical significance of
19 differences between two means was determined by one-way or two-way ANOVA with Bonferroni's multiple comparison test for comparison between treatments or Tukey s multiple comparison test. Differences were considered to be statistically significant when p < REFERENCES 1 CDC. CDC Vital Signs: Asthma in the US Growing Every Year. Available from: (2011). 2 Dolhnikoff, M. et al. The outer wall of small airways is a major site of remodeling in fatal asthma. The Journal of allergy and clinical immunology 123, , 1097 e1091, doi: /j.jaci (2009). 3 Gerthoffer, W. T., Schaafsma, D., Sharma, P., Ghavami, S. & Halayko, A. J. Motility, survival, and proliferation. Comprehensive Physiology 2, , doi: /cphy.c (2012). 4 Ward, C. et al. Reduced airway distensibility, fixed airflow limitation, and airway wall remodeling in asthma. American journal of respiratory and critical care medicine 164, , doi: /ajrccm (2001). 5 Elias, J. A., Zhu, Z., Chupp, G. & Homer, R. J. Airway remodeling in asthma. The Journal of clinical investigation 104, , doi: /jci8124 (1999). 6 James, A. Airway remodeling in asthma. Current opinion in pulmonary medicine 11, 1-6 (2005). 7 James, A. L. & Wenzel, S. Clinical relevance of airway remodelling in airway diseases. The European respiratory journal 30, , doi: / (2007).
20 8 Deshpande, D. A. & Penn, R. B. Targeting G protein-coupled receptor signaling in asthma. Cell Signal 18, , doi: /j.cellsig (2006). 9 Billington, C. K. & Penn, R. B. Signaling and regulation of G protein-coupled receptors in airway smooth muscle. Respiratory research 4, 2 (2003). 10 Deshpande, D. A. et al. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat Med 16, , doi: /nm.2237 (2010). 11 Zhang, C. H. et al. The cellular and molecular basis of bitter tastant-induced bronchodilation. PLoS Biol 11, e , doi: /journal.pbio (2013). 12 Tan, X. & Sanderson, M. J. Bitter tasting compounds dilate airways by inhibiting airway smooth muscle calcium oscillations and calcium sensitivity. Br J Pharmacol 171, , doi: /bph (2014). 13 Bai, Y. et al. Cryopreserved Human Precision-cut Lung Slices as a Bioassay for Live Tissue Banking: a Viability Study of Bronchodilation with Bitter-taste Receptor Agonists. American journal of respiratory cell and molecular biology, doi: /rcmb ma (2015). 14 Grassin-Delyle, S. et al. The expression and relaxant effect of bitter taste receptors in human bronchi. Respiratory research 14, 134, doi: / (2013). 15 Camoretti-Mercado, B. et al. Pleiotropic Effects of Bitter Taste Receptors on [Ca2+]i Mobilization, Hyperpolarization, and Relaxation of Human Airway Smooth Muscle Cells. PloS one 10, e , doi: /journal.pone (2015). 16 Pulkkinen, V., Manson, M. L., Safholm, J., Adner, M. & Dahlen, S. E. The bitter taste receptor (TAS2R) agonists denatonium and chloroquine display distinct patterns of
21 relaxation of the guinea pig trachea. American journal of physiology 303, L , doi: /ajplung (2012). 17 Robinett, K. S. et al. Bitter taste receptor function in asthmatic and nonasthmatic human airway smooth muscle cells. American journal of respiratory cell and molecular biology 50, , doi: /rcmb rc (2014). 18 Liggett, S. B. Bitter taste receptors on airway smooth muscle as targets for novel bronchodilators. Expert Opin Ther Targets 17, , doi: / (2013). 19 Liggett, S. B. Bitter taste receptors in the wrong place: novel airway smooth muscle targets for treating asthma. Trans Am Clin Climatol Assoc 125, 64-74; discussion (2014). 20 Cushing, I. E. & Miller, W. F. Nebulization therapy. Clin Anesth 1, (1965). 21 Sharma, P. et al. Anti-mitogenic Effect of Bitter Taste Receptor Agonists on Airway Smooth Muscle Cells. American journal of physiology, ajplung , doi: /ajplung (2015). 22 Ji, M. et al. Identification of novel compounds for human bitter taste receptors. Chem Biol Drug Des 84, 63-74, doi: /cbdd (2014). 23 Orsmark-Pietras, C. et al. Transcriptome analysis reveals upregulation of bitter taste receptors in severe asthmatics. The European respiratory journal 42, 65-78, doi: / (2013). 24 Ekoff, M. et al. Bitter taste receptor (TAS2R) agonists inhibit IgE-dependent mast cell activation. The Journal of allergy and clinical immunology 134, , doi: /j.jaci (2014).
22 25 Lee, R. J. & Cohen, N. A. Bitter and sweet taste receptors in the respiratory epithelium in health and disease. J Mol Med (Berl) 92, , doi: /s (2014). 26 Lee, R. J. et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. The Journal of clinical investigation 124, , doi: /jci72094 (2014). 27 Penn, R. B. et al. Effects of inflammation and acute beta-agonist inhalation on beta 2-AR signaling in human airways. Am J Physiol 271, L (1996). 28 Penn, R. B., Panettieri, R. A., Jr. & Benovic, J. L. Mechanisms of acute desensitization of the beta2ar-adenylyl cyclase pathway in human airway smooth muscle. American journal of respiratory cell and molecular biology 19, , doi: /ajrcmb (1998). 29 Bako, G. et al. Follow up of Graves' patients thyroid stimulating antibody activity during treatment. A prospective study. Radiobiol Radiother (Berl) 28, (1987). 30 Warner, S. M. & Knight, D. A. Airway modeling and remodeling in the pathogenesis of asthma. Current opinion in allergy and clinical immunology 8, 44-48, doi: /aci.0b013e3282f3b5cb (2008). 31 Benayoun, L., Druilhe, A., Dombret, M. C., Aubier, M. & Pretolani, M. Airway structural alterations selectively associated with severe asthma. American journal of respiratory and critical care medicine 167, , doi: /rccm oc (2003). 32 Tsurikisawa, N. et al. Bronchial hyperresponsiveness to histamine correlates with airway remodelling in adults with asthma. Respir Med 104, , doi: /j.rmed (2010).
23 33 Sharma, P. et al. beta-dystroglycan binds caveolin-1 in smooth muscle: a functional role in caveolae distribution and Ca2+ release. Journal of cell science 123, , doi: /jcs (2010). 34 Schaafsma, D. et al. The mevalonate cascade as a target to suppress extracellular matrix synthesis by human airway smooth muscle. American journal of respiratory cell and molecular biology 44, , doi: /rcmb oc (2011). 35 Sharma, P. et al. Role of dystrophin in airway smooth muscle phenotype, contraction and lung function. PloS one 9, e102737, doi: /journal.pone (2014). 36 Sharma, P. et al. Epithelium-dependent modulation of responsiveness of airways from caveolin-1 knockout mice is mediated through cyclooxygenase-2 and 5-lipoxygenase. Br J Pharmacol 167, , doi: /j x (2012). 37 Sharma, P. et al. Expression of the dystrophin-glycoprotein complex is a marker for human airway smooth muscle phenotype maturation. American journal of physiology 294, L57-68 (2008). 38 Baines, K. J., Simpson, J. L., Scott, R. J. & Gibson, P. G. Immune responses of airway neutrophils are impaired in asthma. Exp Lung Res 35, (2009). 39 Pillay, J. et al. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. The Journal of clinical investigation 122, , doi: /jci57990 (2012). 40 Tang, F. S. et al. A novel immunomodulatory function of neutrophils on rhinovirusactivated monocytes in vitro. Thorax 71, , doi: /thoraxjnl (2016).
24 41 Sharma, P. et al. Characterization of the dystrophin-glycoprotein complex in airway smooth muscle: role of delta-sarcoglycan in airway responsiveness. Can J Physiol Pharmacol 93, , doi: /cjpp (2015). Authors Contributions P.S., and D.A.D. conception and design of research; P.S., R.Y., A.P.N., N.W., F.T., and S.P. performed experiments; M.J.K. performed aerosol particle measurements; P.S. analyzed data; P.S., B.O., and D.A.D. interpreted results of experiments; P.S. and A.P.N. prepared figures; P.S. and D.A.D. drafted manuscript; P.S., B.O., and D.A.D. edited and revised manuscript; P.S., R.Y., A.P.N., N.W., F.T., M.J.K., S.P., B.O., and D.A.D. approved final version of manuscript. Acknowledgements This study was supported by grants from American Asthma Foundation and National Institute on Aging Grant AG to D. A. Deshpande. Competing financial interests The authors have declared that no conflict of interest exists.
25 FIGURE LEGENDS Figure 1: Allergen sensitization and challenge protocols in mice. (A) Male FVB/N mice (8- weeks) were injected with 2 mg alum in 0.2 ml PBS or alum with 100 mg ovalbumin (OVA) per mouse on day 1 and day 14 intraperitoneally. On days 19, 21, 23, 25 and 27 mice were challenged with aerosolized OVA (1%) or saline for 30 min. A select set of animals were treated with nebulized chloroquine or quinine (1.5 mg/ml) or vehicle for 30 min prior to OVA challenge each day. (B) Female BALB/c mice were challenged with house dust mite (HDM, 25 µg/35µl) allergen intranasally as shown in the above schematic: 5 days/week for 3 weeks. 30 min prior to HDM challenges, a select set of mice were treated with chloroquine (50 mg/kg) or quinine (10 mg/kg) intranasally. 24 h after the last allergen challenge, phenotype was assessed by collecting lung lavage, formalin-fixed or frozen lung samples, and measuring lung function by flexivent. Key features of asthma namely airway inflammation, remodeling and hyperresponsiveness were assessed respectively. (C) Chloroquine aerosol particle size distribution. Aerosolized Chloroquine (1.5 mg/ml) was captured in an Anderson Cascade Impactor and quantified by HPLC-MS. Based on the aerosol quantification, each animal received 0.5 µg per day based on 30 min of aerosol treatment. Figure 2: TAS2R agonists inhibit allergen-induced airway inflammation. (A) Total BAL cell count was performed using a hemocytometer. Histology was performed on formalin-fixed lungs and then stained with H&E for assessing tissue inflammation (B and C). Lung inflammation scoring was performed on H&E stained tissue slides as described in the methods section using multiple images (3 images/mouse) (D and E). Data shown above represent the mean of 8-11
26 animals in each group; *p<0.05 (Saline vs HDM or OVA), #p<0.05 (HDM or OVA vs CQ or Q). CQ: chloroquine; Q: quinine. Figure 3: TAS2R agonists mitigate features of fibrotic airway remodeling. Formalin-fixed lungs were stained with AFOG (A) or Trichrome (B) for assessing airway fibrosis to HDM and OVA. Airway fibrosis scoring was performed as described in the methods for both HDM (C) and OVA (D) models. Immunoblotting was performed using the lung lysates to assess pro-fibrotic signaling for p-smad-2 (E), Fibronectin (H) and β-actin was used as a loading control (I). Total soluble collagen (F) was also quantified using a Sircol-collagen assay and levels of TGF-β1 (G) release in the BAL fluid was measured using multiplex array as described in the methods. Data shown above represent the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM or OVA), #p<0.05 (HDM or OVA vs CQ or Q) using one way ANOVA. HDM: house dust mite; OVA: ovalbumin; CQ: chloroquine; Q: quinine, FN: fibronectin. Figure 4: TAS2R agonists inhibit accumulation of smooth muscle mass upon allergen challenge. Airway smooth muscle mass was determined by immunofluorescence method in both HDM (A) and OVA (B) models. Stained images were then quantified using Image J analysis software to determine sm-α-actin staining in the lungs (C, D). Key smooth muscle marker proteins were assessed by immunoblotting using the lung-lysates in the HDM model. Representative western blots (E) and densitometric analyses for smmhc (F), sm-α-actin (G) and calponin (H) are shown in the figure. Similarly smooth muscle markers were assessed in the OVA model as shown in the representative immunoblots (I) with densitometric analysis for smmhc (J), sm-α-actin (K) and calponin (L). β-actin was used as a loading control. Data shown above represent the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM or OVA),
27 #p<0.05 (HDM or OVA vs CQ or Q) using 1-way ANOVA. HDM: house dust mite; OVA: ovalbumin; CQ: chloroquine; Q: quinine. Figure 5: TAS2R agonists inhibit mucus accumulation and MMP release in the airways. Formalin-fixed airways were stained with PAS to assess mucus accumulation in the airways in both HDM model (A) and OVA model (C). Airway mucus staining was blindly graded using multiple images (3 images/mouse) for both HDM (B) and OVA (D). A variety of MMPs were measured in the BAL namely MMP-8 (E & H), prommp-9 (F & I) and MMP-12 (G & J) in HDM and OVA animals respectively. Results are described as pg/ml. Data shown above represents the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM or OVA), #p<0.05 (HDM or OVA vs CQ or Q) using 1-way ANOVA. CQ: chloroquine; Q: quinine. Figure 6: Allergen-induced AHR is diminished upon pre-treatment with TAS2R agonists. Mice were subjected to an increased dose of nebulized methacholine (MCh) challenge protocol to assess characteristics of respiratory mechanics in HDM (A) and OVA model (B). Animals were treated with aerosolized chloroquine or quinine at the end of the highest concentration of MCh and % change in R was computed in HDM (C) or OVA (D) group. *P < 0.05, was considered significant for HDM or OVA vs saline and #P < 0.05, was considered significant for HDM or OVA vs CQ or Q treated group. Data shown is the mean of 8-11 mice in each group. Figure 7. Effect of TAS2R agonists on features of allergic asthma in the treatment model. A. Female BALB/c mice were initially dosed 5 days a week for 3 weeks with 25 µg of HDM extract. Mice were then treated with either chloroquine (CQ, 50 mg/kg) or quinine (Q, 10 mg/kg) or vehicle (1% ethanol) 30 min prior to subsequent HDM challenges for additional 2 weeks. B. Histopathological assessment of tissue inflammation was performed on formalin-fixed lung tissue
28 using H&E staining. C. Formalin-fixed lungs were stained with Trichrome for assessing airway fibrosis to HDM. D. Total soluble collagen in lung tissue was quantified using a Sircol-collagen assay. E. Immunoblotting was performed using the lung lysates to evaluate pro-fibrotic signaling for p-smad-2 and fibronectin (FN). F. Representative western blots for sm-α-actin, smmhc, and calponin are shown. β-actin was used as a loading control. G. ASM mass was determined by staining formalin-fixed lung tissue for sm-α-actin using immunofluorescence method. H. Fixed lungs were stained with PAS to assess accumulation of mucus producing cells in the bronchioles. I. Mice were subjected to an increased dose of nebulized methacholine (MCh) challenge protocol to assess features of respiratory mechanics. *P < 0.05, was considered significant for HDM or OVA vs saline and #P < 0.05, was considered significant for HDM or OVA vs CQ or Q treated group. Data shown is the mean of 8-11 mice in each group. Figure 8: Effect of TAS2R agonists on neutrophil chemotaxis. The neutrophil chemotaxis or migration to CXCL8 was measured using Transwell inserts. Isolated and purified peripheral human blood neutrophils were plated at desired density in the upper chamber of the Transwell with or without TAS2R agonists (CQ and Q) at increasing concentrations (100 µm, 500 µm and 1 mm) for 90 minutes at 37 o C while a chemotactic gradient was created using IL-8 (100 ng/ml) in the lower chamber of the Transwell plate. The results are expressed as fold change in cell migration from upper well to lower well and expressed as neutrophil migration index relative to control, which is kept as 1. Data shown above represent the mean of 5-human donors in each group; *p<0.05 (Control vs IL-8), #p<0.05 (IL-8vsCQ or Q) using 1-way ANOVA. CQ: chloroquine;q:quinine.
 Allergen-induced asthma models Differential BALF counts (cell numbers ± SEM) Eosinophils Neutrophils Macrophages Lymphocytes")

29 TABLES Table 1. Differential cell counts (prophylactic model and treatment model) Allergen-induced asthma models Differential BALF counts (cell numbers ± SEM) Eosinophils Neutrophils Macrophages Lymphocytes Prophylactic Allergen HDM Saline control 329 ± ± ,725 ± 13,721 21,550 ± 4,800 Vehicle + HDM 301,750 ± 22,605 * 227,583 ± 16,484 * 242,250 ± 20,579 * 131,450 ± 20,437 * CQ + HDM 151,909 ± 17,391 # 153,655 ± 17,520 # 168,227 ± 18,120 # 101,936 ± 13,797 # Q + HDM 121,778 ± 6,984 # 123,133 ± 12,419 # 150,778 ±17,505 # 88,689 ± 15,446 # Allergen - OVA Saline control 129 ± 129 2,137 ± ,888 ± 63,857 71,250 ± 25,706 Vehicle + OVA 228,700 ± 40,250 * 217,900 ± 17,124* 258,200 ± 34, ,590 ± 12,947 CQ + OVA 114,430 ± 6,335 # 113,030 ± 6,824 # 208,080 ± 37,431 67,120 ± 8,401 Q + OVA 134,900 ± 21,007 # 147,330 ± 20,739 # 171,660 ± 29,377 98,680 ± 19,500 Treatment model Allergen - HDM Saline control 353 ± ± ,911 ± 9,907 39,700 ± 3,994 Vehicle + HDM 222,625 ± 18,383 * 274,500 ± 36,029 * 302,250 ± 29,842 * 149,438 ± 17,656 * CQ + HDM 147,300 ± 10,964 # 177,000 ± 14,052 # 240,875 ± 20, ,750 ± 17,465 Q + HDM 149,000 ± 19,431 # 149,375 ± 15,427 # 203,750 ± 17,783 # 154,683 ± 27,903 Data shown above represents the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM or OVA), #p<0.05 (HDM or OVA vs CQ or Q) using 1-way ANOVA. CQ: chloroquine; Q: quinine; OVA: Ovalbumin; HDM: house dust mite; BALF: bronchoalveolar lavage fluid; SEM: standard error of means.
30 Table 2. BALF Cytokines and chemokines release in prophylactic models Treatment groups Cytokines/ Chemokines (pg/ml ± SEM) Allergen - HDM Allergen OVA Saline control Vehicle + HDM CQ + HDM Q + HDM Saline control Vehicle + HDM CQ + HDM Q + HDM IL ± ± 3.70 * 7.15 ± 0.90 # 6.47 ± 1.27 # 0.21 ± ± 1.92 * 4.97 ± 0.65 # 6.76 ± 1.69 # IL ± ± 7.06 * ± 3.92 # ± 4.61 # 0.53 ± ± 6.44 * ± 6.64 # ± 5.67 # IL ± ± 7.53 * ± ± 6.36 # 2.28 ± ± ± ± IL ± ± 3.35 * 7.83 ± 2.50 # 4.40 ± 1.34 # 0.12 ± ± 1.48 * 3.25 ± 0.85 # 6.12 ± 2.56 # IL ± ± 0.25 * 0.82 ± ± ± ± 0.16 * 0.50 ± 0.14 # 0.65 ± 0.17 # IL ± ± * ± ± 0.54 # n.d. Eotaxin ± ± * ± 7.89 # ± 9.41 # ± ± * ± # ± # KC 19.5 ± ± * ± # ± # ± ± * ± # ± # TNF-α 1.24 ± ± 3.41 * ± 2.09 # 6.72 ± 1.36 # 0.81 ± ± 1.18 * 4.14 ± ± 0.67 IP ± ± 4.80 * ± ± 2.50 # 4.48 ± ± 3.36 * ± ± 1.61 RANTES 0.14 ± ± 2.30 * 3.40 ± 0.80 # 2.12 ± 0.42 # 0.31 ± ± 0.83 * 1.23 ± ± 0.26 Data shown above represents the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM or OVA), #p<0.05 (HDM or OVA vs CQ or Q) using 1-way ANOVA. CQ: chloroquine; Q: quinine; OVA: Ovalbumin; HDM: house dust mite extract; SEM: standard error of means; KC: keratinocyte chemoattractant; IP-10: interferon-γ-induced protein 10; RANTES: regulated on activation, normal T cell expressed and secreted; n.d.: not detected.

31 Table 3. Treatment model for HDM-induced allergic airway disease Cytokines/Chemokines (pg/ml ± SEM) Treatment groups (Allergen HDM) Saline control Vehicle + HDM CQ + HDM Q + HDM Eotaxin ± ± * ± 5.71 # ± 5.23 # KC ± ± * ± # ± # IP ± ± 3.26 * 8.80 ± 1.59 # 8.36 ± 1.33 # RANTES 4.98 ± ± 2.24 * 5.70 ± 0.74 # 5.10 ± 0.68 # TGF-β ± ± * ± # ± # Data shown above represents the mean of 8-11 animals in each group; *p<0.05 (Saline vs HDM), #p<0.05 (HDM vs CQ or Q) using 1-way ANOVA. CQ: chloroquine; Q: quinine; HDM: house dust mite extract; SEM: standard error of means; KC: keratinocyte chemoattractant; IP-10: interferon-γ-induced protein 10; RANTES: regulated on activation, normal T cell expressed and secreted; TGF-β1: transforming growth factor beta 1.

32
33

34

35

36

37

38

39
Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice.
 Thomas Jefferson University Jefferson Digital Commons Department of Medicine Faculty Papers Department of Medicine 4-11-2017 Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice.
Thomas Jefferson University Jefferson Digital Commons Department of Medicine Faculty Papers Department of Medicine 4-11-2017 Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice.
Role of Tyk-2 in Th9 and Th17 cells in allergic asthma
 Supplementary File Role of Tyk-2 in Th9 and Th17 cells in allergic asthma Caroline Übel 1*, Anna Graser 1*, Sonja Koch 1, Ralf J. Rieker 2, Hans A. Lehr 3, Mathias Müller 4 and Susetta Finotto 1** 1 Laboratory
Supplementary File Role of Tyk-2 in Th9 and Th17 cells in allergic asthma Caroline Übel 1*, Anna Graser 1*, Sonja Koch 1, Ralf J. Rieker 2, Hans A. Lehr 3, Mathias Müller 4 and Susetta Finotto 1** 1 Laboratory
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic
 Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
10.00 PBS OVA OVA+isotype antibody 8.00 OVA+anti-HMGB1. PBS Methatroline (mg/ml)
") RESEARCH ARTICLE Penh (100% of PBS) 1 PBS 8.00 +anti-hmgb1 6.00 4.00 p=0.054 Cellular & Molecular Immunology advance online publication, PBS 3.12 6.25 Methatroline (mg/ml) Neutrophil isolation and culture
RESEARCH ARTICLE Penh (100% of PBS) 1 PBS 8.00 +anti-hmgb1 6.00 4.00 p=0.054 Cellular & Molecular Immunology advance online publication, PBS 3.12 6.25 Methatroline (mg/ml) Neutrophil isolation and culture
The Effect of BTP on the Development of Allergic Asthma in Mice
 93 The Effect of BTP on the Development of Allergic Asthma in Mice Alana Curry, McNair Scholar, Virginia State University Faculty Advisor: Avery August, Ph.D Associate Professor of Immunology Department
93 The Effect of BTP on the Development of Allergic Asthma in Mice Alana Curry, McNair Scholar, Virginia State University Faculty Advisor: Avery August, Ph.D Associate Professor of Immunology Department
Bitter Taste Receptors: Taking a Bigger Bite of Airway Smooth Muscle Pathophysiology
 Page 1 of 6 Editorial Bitter Taste Receptors: Taking a Bigger Bite of Airway Smooth Muscle Pathophysiology Charles W. Emala, Sr. Department of Anesthesiology Columbia University, New York, New York Contact
Page 1 of 6 Editorial Bitter Taste Receptors: Taking a Bigger Bite of Airway Smooth Muscle Pathophysiology Charles W. Emala, Sr. Department of Anesthesiology Columbia University, New York, New York Contact
Role of C5 in the development of airway inflammation, airway hyperresponsiveness, and ongoing airway response
 Research article Role of C5 in the development of airway inflammation, airway hyperresponsiveness, and ongoing airway response Tao Peng, 1 Liming Hao, 2 Joseph A. Madri, 2 Xiao Su, 1 Jack A. Elias, 3 Gregory
Research article Role of C5 in the development of airway inflammation, airway hyperresponsiveness, and ongoing airway response Tao Peng, 1 Liming Hao, 2 Joseph A. Madri, 2 Xiao Su, 1 Jack A. Elias, 3 Gregory
Potent and Selective CRTh2 Antagonists are Efficacious in Models of Asthma, Allergic Rhinitis and Atopic Dermatitis
 Potent and Selective CRTh2 Antagonists are Efficacious in Models of Asthma, Allergic Rhinitis and Atopic Dermatitis Laura L. Carter, Yoshi Shiraishi, Yooseob Shin, Laurence Burgess, Christine Eberhardt,
Potent and Selective CRTh2 Antagonists are Efficacious in Models of Asthma, Allergic Rhinitis and Atopic Dermatitis Laura L. Carter, Yoshi Shiraishi, Yooseob Shin, Laurence Burgess, Christine Eberhardt,
Impact of Asthma in the U.S. per Year. Asthma Epidemiology and Pathophysiology. Risk Factors for Asthma. Childhood Asthma Costs of Asthma
 American Association for Respiratory Care Asthma Educator Certification Prep Course Asthma Epidemiology and Pathophysiology Robert C. Cohn, MD, FAARC MetroHealth Medical Center Cleveland, OH Impact of
American Association for Respiratory Care Asthma Educator Certification Prep Course Asthma Epidemiology and Pathophysiology Robert C. Cohn, MD, FAARC MetroHealth Medical Center Cleveland, OH Impact of
Airway Inflammation in Asthma Chih-Yung Chiu 1,2, Kin-Sun Wong 2 1 Department of Pediatrics, Chang Gung Memorial Hospital, Keelung, Taiwan.
 REVIEW ARTICLE Chih-Yung Chiu 1,2, Kin-Sun Wong 2 1 Department of Pediatrics, Chang Gung Memorial Hospital, Keelung, Taiwan. 2 Division of Pediatric Pulmonology, Department of Pediatrics, Chang Gung Memorial
REVIEW ARTICLE Chih-Yung Chiu 1,2, Kin-Sun Wong 2 1 Department of Pediatrics, Chang Gung Memorial Hospital, Keelung, Taiwan. 2 Division of Pediatric Pulmonology, Department of Pediatrics, Chang Gung Memorial
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Figure 1
 d f a IL7 b IL GATA RORγt h HDM IL IL7 PBS Ilra R7 PBS HDM Ilra R7 HDM Foxp Foxp Ilra R7 HDM HDM Ilra R7 HDM. 9..79. CD + FOXP + T reg cell CD + FOXP T conv cell PBS Ilra R7 PBS HDM Ilra R7 HDM CD + FOXP
d f a IL7 b IL GATA RORγt h HDM IL IL7 PBS Ilra R7 PBS HDM Ilra R7 HDM Foxp Foxp Ilra R7 HDM HDM Ilra R7 HDM. 9..79. CD + FOXP + T reg cell CD + FOXP T conv cell PBS Ilra R7 PBS HDM Ilra R7 HDM CD + FOXP
Soluble ADAM33 initiates airway remodeling to promote susceptibility for. Elizabeth R. Davies, Joanne F.C. Kelly, Peter H. Howarth, David I Wilson,
 Revised Suppl. Data: Soluble ADAM33 1 Soluble ADAM33 initiates airway remodeling to promote susceptibility for allergic asthma in early life Elizabeth R. Davies, Joanne F.C. Kelly, Peter H. Howarth, David
Revised Suppl. Data: Soluble ADAM33 1 Soluble ADAM33 initiates airway remodeling to promote susceptibility for allergic asthma in early life Elizabeth R. Davies, Joanne F.C. Kelly, Peter H. Howarth, David
Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and
 Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and Thy1 in NH cells derived from the lungs of naïve mice.
Supplementary Figure S1. Flow cytometric analysis of the expression of Thy1 in NH cells. Flow cytometric analysis of the expression of T1/ST2 and Thy1 in NH cells derived from the lungs of naïve mice.
Identification of an IFN-γ/mast cell axis in a mouse model of chronic asthma
 Research article Identification of an IFN-γ/mast cell axis in a mouse model of chronic asthma Mang Yu, 1 Michael R. Eckart, 2 Alexander A. Morgan, 3 Kaori Mukai, 1 Atul J. Butte, 3 Mindy Tsai, 1 and Stephen
Research article Identification of an IFN-γ/mast cell axis in a mouse model of chronic asthma Mang Yu, 1 Michael R. Eckart, 2 Alexander A. Morgan, 3 Kaori Mukai, 1 Atul J. Butte, 3 Mindy Tsai, 1 and Stephen
Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained
 Representative pictures of H&E-stained") 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
Supplemental Table 1. Primer sequences for transcript analysis
 Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
CD4 + and CD8 + T cells play a central role in a HDM driven model of allergic asthma
 Raemdonck et al. Respiratory Research (2016)17:45 DOI 10.1186/s12931-016-0359-y RESEARCH CD4 + and CD8 + T cells play a central role in a HDM driven model of allergic asthma Open Access Kristof Raemdonck
Raemdonck et al. Respiratory Research (2016)17:45 DOI 10.1186/s12931-016-0359-y RESEARCH CD4 + and CD8 + T cells play a central role in a HDM driven model of allergic asthma Open Access Kristof Raemdonck
Defining Asthma: Clinical Criteria. Defining Asthma: Bronchial Hyperresponsiveness
 Defining Asthma: Clinical Criteria Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine, 2nd ed. Murray, Nadel, eds.(saunders:philadelphia)
Defining Asthma: Clinical Criteria Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine, 2nd ed. Murray, Nadel, eds.(saunders:philadelphia)
Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive
 Online Data Supplement: Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive asthma Dan Cheng, Zheng Xue, Lingling Yi, Huimin Shi, Kan Zhang, Xiaorong Huo, Luke R. Bonser,
Online Data Supplement: Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive asthma Dan Cheng, Zheng Xue, Lingling Yi, Huimin Shi, Kan Zhang, Xiaorong Huo, Luke R. Bonser,
Systems Pharmacology Respiratory Pharmacology. Lecture series : General outline
 Systems Pharmacology 3320 2017 Respiratory Pharmacology Associate Professor Peter Henry (Rm 1.34) Peter.Henry@uwa.edu.au Division of Pharmacology, School of Biomedical Sciences Lecture series : General
Systems Pharmacology 3320 2017 Respiratory Pharmacology Associate Professor Peter Henry (Rm 1.34) Peter.Henry@uwa.edu.au Division of Pharmacology, School of Biomedical Sciences Lecture series : General
Immunology of Asthma. Kenneth J. Goodrum,Ph. Ph.D. Ohio University College of Osteopathic Medicine
 Immunology of Asthma Kenneth J. Goodrum,Ph Ph.D. Ohio University College of Osteopathic Medicine Outline! Consensus characteristics! Allergens:role in asthma! Immune/inflammatory basis! Genetic basis!
Immunology of Asthma Kenneth J. Goodrum,Ph Ph.D. Ohio University College of Osteopathic Medicine Outline! Consensus characteristics! Allergens:role in asthma! Immune/inflammatory basis! Genetic basis!
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
IL-13 Augments Compressive Stress-induced Tissue Factor Expression in Human Airway Epithelial Cells
 IL-13 Augments Compressive Stress-induced Tissue Factor Expression in Human Airway Epithelial Cells Jennifer A. Mitchel, Silvio Antoniak, Joo-Hyeon Lee, Sae-Hoon Kim, Maureen McGill, David I. Kasahara,
IL-13 Augments Compressive Stress-induced Tissue Factor Expression in Human Airway Epithelial Cells Jennifer A. Mitchel, Silvio Antoniak, Joo-Hyeon Lee, Sae-Hoon Kim, Maureen McGill, David I. Kasahara,
Defining Asthma: Clinical Criteria. Defining Asthma: Bronchial Hyperresponsiveness
 Defining Asthma: Clinical Criteria Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine, 2nd ed. Murray, Nadel, eds.(saunders:philadelphia)
Defining Asthma: Clinical Criteria Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine, 2nd ed. Murray, Nadel, eds.(saunders:philadelphia)
Defining Asthma: Bronchial Hyperresponsiveness. Defining Asthma: Clinical Criteria. Impaired Ventilation in Asthma. Dynamic Imaging of Asthma
 Defining Asthma: Clinical Criteria Defining Asthma: Bronchial Hyperresponsiveness Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine,
Defining Asthma: Clinical Criteria Defining Asthma: Bronchial Hyperresponsiveness Atopy 34% Recent wheeze 20% Asthma 11% AHR 19% n = 807 From: Woolcock, AJ. Asthma in Textbook of Respiratory Medicine,
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Basis of Immunology and
 Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Safety, PK and PD of ARRY-502, a CRTh2 Antagonist, in Healthy Subjects with a History of Seasonal Allergies
 Safety, PK and PD of ARRY502, a CRTh2 Antagonist, in Healthy Subjects with a History of Seasonal Allergies L. Burgess*, L. Anderson, C. Nugent, N. Klopfenstein, C. Eberhardt, L. Carter, C. Kass, S. RojasCaro,
Safety, PK and PD of ARRY502, a CRTh2 Antagonist, in Healthy Subjects with a History of Seasonal Allergies L. Burgess*, L. Anderson, C. Nugent, N. Klopfenstein, C. Eberhardt, L. Carter, C. Kass, S. RojasCaro,
Supplementary Material
 Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Electrolytes. Summary: (This area will include a brief description of what the protocol is used for and why someone would need to use it.
 Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Searching for Targets to Control Asthma
 Searching for Targets to Control Asthma Timothy Craig Distinguished Educator Professor Medicine and Pediatrics Penn State University Hershey, PA, USA Inflammation and Remodeling in Asthma The most important
Searching for Targets to Control Asthma Timothy Craig Distinguished Educator Professor Medicine and Pediatrics Penn State University Hershey, PA, USA Inflammation and Remodeling in Asthma The most important
Supporting Information
 Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
E-1 Role of IgE and IgE receptors in allergic airway inflammation and remodeling
 E-1 Role of IgE and IgE receptors in allergic airway inflammation and remodeling Ruby Pawankar, MD, Ph.D. FRCP, FAAAAI Prof. Div of Allergy, Dept of Pediatrics Nippon Medical School Tokyo, Japan pawankar.ruby@gmail.com
E-1 Role of IgE and IgE receptors in allergic airway inflammation and remodeling Ruby Pawankar, MD, Ph.D. FRCP, FAAAAI Prof. Div of Allergy, Dept of Pediatrics Nippon Medical School Tokyo, Japan pawankar.ruby@gmail.com
COPD lungs show an attached stratified mucus layer that separate. bacteria from the epithelial cells resembling the protective colonic
 COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
In vitro bactericidal assay Fig. S8 Gentamicin protection assay Phagocytosis assay
 In vitro bactericidal assay Mouse bone marrow was isolated from the femur and the tibia. Cells were suspended in phosphate buffered saline containing.5% BSA and 2 mm EDTA and filtered through a cell strainer.
In vitro bactericidal assay Mouse bone marrow was isolated from the femur and the tibia. Cells were suspended in phosphate buffered saline containing.5% BSA and 2 mm EDTA and filtered through a cell strainer.
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES
 HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Systems Pharmacology Respiratory Pharmacology. Lecture series : General outline
 Systems Pharmacology 3320 2017 Respiratory Pharmacology Associate Professor Peter Henry (Rm 1.34) Peter.Henry@uwa.edu.au Division of Pharmacology, School of Biomedical Sciences Lecture series : General
Systems Pharmacology 3320 2017 Respiratory Pharmacology Associate Professor Peter Henry (Rm 1.34) Peter.Henry@uwa.edu.au Division of Pharmacology, School of Biomedical Sciences Lecture series : General
Evaluation of directed and random motility in microslides Assessment of leukocyte adhesion in flow chambers
 Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Supplementary Information
 Supplementary Information TABLE S1. SUBJECT CHARACTERISTICS* Normal Control Subjects Subjects with Asthma p Value Number 23 48 Age (years) 35±10 35±10 0.75 Sex, M:F (% F) 9:12 (57) 17:26 (60) 0.76 FEV1
Supplementary Information TABLE S1. SUBJECT CHARACTERISTICS* Normal Control Subjects Subjects with Asthma p Value Number 23 48 Age (years) 35±10 35±10 0.75 Sex, M:F (% F) 9:12 (57) 17:26 (60) 0.76 FEV1
Concurrent dual allergen exposure and its effects on airway hyperresponsiveness, inflammation and remodeling in mice
 Disease Models & Mechanisms, 75-8 (9) doi:1.14/dmm.1859 RESEARCH ARTICLE Concurrent dual allergen exposure and its effects on airway hyperresponsiveness, inflammation and remodeling in mice Franco A. DiGiovanni
Disease Models & Mechanisms, 75-8 (9) doi:1.14/dmm.1859 RESEARCH ARTICLE Concurrent dual allergen exposure and its effects on airway hyperresponsiveness, inflammation and remodeling in mice Franco A. DiGiovanni
Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines
 CORRECTION NOTICE Nat. Med. doi:10.1038/nm.3547; corrected online 25 August 2014 Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines Christine Schauer, Christina
CORRECTION NOTICE Nat. Med. doi:10.1038/nm.3547; corrected online 25 August 2014 Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines Christine Schauer, Christina
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Biomerker onderzoek voor isocyanaatgeïnduceerd
 CGC en NVAB meeting, 6 Juni 2013, s-hertogenbosch Biomerker onderzoek voor isocyanaatgeïnduceerd astma Jeroen Vanoirbeek Asthma 1. Chronic airway disease: prevalence: 5-10%, 300.000 people affected world-wide
CGC en NVAB meeting, 6 Juni 2013, s-hertogenbosch Biomerker onderzoek voor isocyanaatgeïnduceerd astma Jeroen Vanoirbeek Asthma 1. Chronic airway disease: prevalence: 5-10%, 300.000 people affected world-wide
Primary Adult Naïve CD4+ CD45RA+ Cells. Prepared by: David Randolph at University of Alabama, Birmingham
 Primary Adult Naïve CD4+ CD45RA+ Cells Prepared by: David Randolph (drdrdr@uab.edu) at University of Alabama, Birmingham Goal: To obtain large numbers of highly pure primary CD4+ CD45RO- CD25- cells from
Primary Adult Naïve CD4+ CD45RA+ Cells Prepared by: David Randolph (drdrdr@uab.edu) at University of Alabama, Birmingham Goal: To obtain large numbers of highly pure primary CD4+ CD45RO- CD25- cells from
IKKα Causes Chromatin Modification on Pro-Inflammatory Genes by Cigarette Smoke in Mouse Lung
 IKKα Causes Chromatin Modification on Pro-Inflammatory Genes by Cigarette Smoke in Mouse Lung Se-Ran Yang, Samantha Valvo, Hongwei Yao, Aruna Kode, Saravanan Rajendrasozhan, Indika Edirisinghe, Samuel
IKKα Causes Chromatin Modification on Pro-Inflammatory Genes by Cigarette Smoke in Mouse Lung Se-Ran Yang, Samantha Valvo, Hongwei Yao, Aruna Kode, Saravanan Rajendrasozhan, Indika Edirisinghe, Samuel
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Dr Rodney Itaki Lecturer Division of Pathology Anatomical Pathology Discipline
 Pathology of Asthma Dr Rodney Itaki Lecturer Division of Pathology Anatomical Pathology Discipline Bronchial Asthma Definition: chronic, relapsing inflammatory lung disorder characterised by reversible
Pathology of Asthma Dr Rodney Itaki Lecturer Division of Pathology Anatomical Pathology Discipline Bronchial Asthma Definition: chronic, relapsing inflammatory lung disorder characterised by reversible
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Mouse Anti-HDM IgG Antibody Assay Kit
 Mouse Anti-HDM IgG Antibody Assay Kit Catalog # 3030 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Asthma is a common chronic inflammatory disease that affects 300 million people of
Mouse Anti-HDM IgG Antibody Assay Kit Catalog # 3030 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Asthma is a common chronic inflammatory disease that affects 300 million people of
Immunology of Asthma. Kenneth J. Goodrum,Ph. Ph.D. Ohio University College of Osteopathic Medicine
 Immunology of Asthma Kenneth J. Goodrum,Ph Ph.D. Ohio University College of Osteopathic Medicine Outline Consensus characteristics/incidence data Immune/inflammatory basis Etiology/Genetic basis Hygiene
Immunology of Asthma Kenneth J. Goodrum,Ph Ph.D. Ohio University College of Osteopathic Medicine Outline Consensus characteristics/incidence data Immune/inflammatory basis Etiology/Genetic basis Hygiene
Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury
 Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury Bastian OW, Koenderman L, Alblas J, Leenen LPH, Blokhuis TJ. Neutrophils contribute to
Neutrophils contribute to fracture healing by synthesizing fibronectin+ extracellular matrix rapidly after injury Bastian OW, Koenderman L, Alblas J, Leenen LPH, Blokhuis TJ. Neutrophils contribute to
Mouse Total IgA Antibody Detection Kit
 Mouse Total IgA Antibody Detection Kit Catalog # 3019 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The total IgA levels in specimens are often determined in mouse disease models involving
Mouse Total IgA Antibody Detection Kit Catalog # 3019 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION The total IgA levels in specimens are often determined in mouse disease models involving
Procaspase-3. Cleaved caspase-3. actin. Cytochrome C (10 M) Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk
 Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk") A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Reviewers' comments: Reviewer #1 Expert in EAE and IL-17a (Remarks to the Author): This study shows that the inducible camp early repressor (ICER) is involved in development of Th17 cells that are pathogenic
Chronic respiratory disease: towards better treatments
 Chronic respiratory disease: towards better treatments Alaina J. Ammit PhD Alaina.Ammit@uts.edu.au Professor of Respiratory Pharmacology, UTS Director, Woolcock Emphysema Centre, Woolcock Institute of
Chronic respiratory disease: towards better treatments Alaina J. Ammit PhD Alaina.Ammit@uts.edu.au Professor of Respiratory Pharmacology, UTS Director, Woolcock Emphysema Centre, Woolcock Institute of
Allergy and Immunology Review Corner: Chapter 75 of Middleton s Allergy Principles and Practice, 7 th Edition, edited by N. Franklin Adkinson, et al.
 Allergy and Immunology Review Corner: Chapter 75 of Middleton s Allergy Principles and Practice, 7 th Edition, edited by N. Franklin Adkinson, et al. Chapter 75: Approach to Infants and Children with Asthma
Allergy and Immunology Review Corner: Chapter 75 of Middleton s Allergy Principles and Practice, 7 th Edition, edited by N. Franklin Adkinson, et al. Chapter 75: Approach to Infants and Children with Asthma
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies
 NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
CONTENTS. STUDY DESIGN METHODS ELISA protocol for quantitation of mite (Dermatophagoides spp.) Der p 1 or Der f 1
 Der p 1 or Der f 1") CONTENTS STUDY DESIGN METHODS ELISA protocol for quantitation of mite (Dermatophagoides spp.) Der p 1 or Der f 1 ELISA protocol for mite (Dermatophagoides spp.) Group 2 ALLERGENS RESULTS (SUMMARY) TABLE
CONTENTS STUDY DESIGN METHODS ELISA protocol for quantitation of mite (Dermatophagoides spp.) Der p 1 or Der f 1 ELISA protocol for mite (Dermatophagoides spp.) Group 2 ALLERGENS RESULTS (SUMMARY) TABLE
SUPPLEMENTARY INFORMATION
 1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
Nishino, Tomoya; Fukushima, Chizu; Asthma and Immunology, 113, 2, (201
 NAOSITE: Nagasaki University's Ac Title Author(s) Citation Clearance of Aspergillus fumigatus inflammation Fukahori, Susumu; Matsuse, Hiroto; Nishino, Tomoya; Fukushima, Chizu; Annals of Allergy, Asthma
NAOSITE: Nagasaki University's Ac Title Author(s) Citation Clearance of Aspergillus fumigatus inflammation Fukahori, Susumu; Matsuse, Hiroto; Nishino, Tomoya; Fukushima, Chizu; Annals of Allergy, Asthma
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Respiratory Pharmacology PCTH 400 Asthma and β-agonists
 Respiratory Pharmacology PCTH 400 Asthma and β-agonists Dr. Tillie-Louise Hackett Department of Anesthesiology, Pharmacology and Therapeutics University of British Columbia Associate Director, Centre of
Respiratory Pharmacology PCTH 400 Asthma and β-agonists Dr. Tillie-Louise Hackett Department of Anesthesiology, Pharmacology and Therapeutics University of British Columbia Associate Director, Centre of
Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-10. Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald,
 Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-1 Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald, James A. Dromey, Bo Han Lee, Junyan Qian, Ralph M Böhmer and Leonard
Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-1 Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald, James A. Dromey, Bo Han Lee, Junyan Qian, Ralph M Böhmer and Leonard
Inhibition of airway remodeling in IL-5 deficient mice
 Inhibition of airway remodeling in IL-5 deficient mice See the related Commentary beginning on page 507. Jae Youn Cho, Marina Miller, Kwang Je Baek, Ji Won Han, Jyothi Nayar, Sook Young Lee, Kirsti McElwain,
Inhibition of airway remodeling in IL-5 deficient mice See the related Commentary beginning on page 507. Jae Youn Cho, Marina Miller, Kwang Je Baek, Ji Won Han, Jyothi Nayar, Sook Young Lee, Kirsti McElwain,
Understanding How Allergic Responses End: The Allergy Resolvome. Lipid mediators
 Understanding How Allergic Responses End: The Allergy Resolvome Lipid mediators Koichiro Asano Tokai University School of Medicine, Kanagawa, JAPAN ko-asano@tokai-u.jp Resolution of granulocytic inflammation
Understanding How Allergic Responses End: The Allergy Resolvome Lipid mediators Koichiro Asano Tokai University School of Medicine, Kanagawa, JAPAN ko-asano@tokai-u.jp Resolution of granulocytic inflammation
Kun Jiang 1, He-Bin Chen 1, Ying Wang 1, Jia-Hui Lin 2, Yan Hu 1, Yu-Rong Fang 1
 Original Article Changes in interleukin-17 and transforming growth factor beta 1 levels in serum and bronchoalveolar lavage fluid and their clinical significance among children with asthma Kun Jiang 1,
Original Article Changes in interleukin-17 and transforming growth factor beta 1 levels in serum and bronchoalveolar lavage fluid and their clinical significance among children with asthma Kun Jiang 1,
The TLR5 ligand flagellin promotes asthma by. priming allergic responses to indoor allergens
 The TLR ligand flagellin promotes asthma by priming allergic responses to indoor allergens Rhonda H. Wilson 1,3, Shuichiro Maruoka 1,3, Gregory S. Whitehead 1, Julie F. Foley 1, Gordon P. Flake 1, Michelle
The TLR ligand flagellin promotes asthma by priming allergic responses to indoor allergens Rhonda H. Wilson 1,3, Shuichiro Maruoka 1,3, Gregory S. Whitehead 1, Julie F. Foley 1, Gordon P. Flake 1, Michelle
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Mouse Anti-OVA IgM Antibody Assay Kit
 Mouse Anti-OVA IgM Antibody Assay Kit Catalog # 3017 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Ovalbumin (OVA) is a widely used antigen for inducing allergic reactions in experimental
Mouse Anti-OVA IgM Antibody Assay Kit Catalog # 3017 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Ovalbumin (OVA) is a widely used antigen for inducing allergic reactions in experimental
Supplementary Figure 1:
 Supplementary Figure 1: (A) Whole aortic cross-sections stained with Hematoxylin and Eosin (H&E), 7 days after porcine-pancreatic-elastase (PPE)-induced AAA compared to untreated, healthy control aortas
Supplementary Figure 1: (A) Whole aortic cross-sections stained with Hematoxylin and Eosin (H&E), 7 days after porcine-pancreatic-elastase (PPE)-induced AAA compared to untreated, healthy control aortas
well for 2 h at rt. Each dot represents an individual mouse and bar is the mean ±
 Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis
 icell Skeletal Myoblasts Application Protocol Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis Introduction The skeletal muscle is one of the
icell Skeletal Myoblasts Application Protocol Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis Introduction The skeletal muscle is one of the
Neonatal Exposure to Microbial Phosphorylcholine Modulates the Development of House Dust Mite Allergy During Adult Life
 Neonatal Exposure to Microbial Phosphorylcholine Modulates the Development of House Dust Mite Allergy During Adult Life J. Sides SEM Preeyam Patel Mentor: Dr John Kearney 015 CAMBAC Research Day 9/18/15
Neonatal Exposure to Microbial Phosphorylcholine Modulates the Development of House Dust Mite Allergy During Adult Life J. Sides SEM Preeyam Patel Mentor: Dr John Kearney 015 CAMBAC Research Day 9/18/15
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Asthma. - A chronic inflammatory disorder which causes recurrent episodes of wheezing, breathlessness, cough and chest tightness.
 Obstructive diseases Asthma - A chronic inflammatory disorder which causes recurrent episodes of wheezing, breathlessness, cough and chest tightness. - Characterized by Intermittent and reversible (the
Obstructive diseases Asthma - A chronic inflammatory disorder which causes recurrent episodes of wheezing, breathlessness, cough and chest tightness. - Characterized by Intermittent and reversible (the
9607 Dr. Perry Road Suite 103. Ijamsville, MD Tel: (301) Fax: (301) Ultrasensitive Samoa Human IL-33 Immunoassay
 Fax: (301) Ultrasensitive Samoa Human IL-33 Immunoassay") AssayGate, Inc. 9607 Dr. Perry Road Suite 103. Ijamsville, MD 21754. Tel: (301)874-0988. Fax: (301)560-8288. Ultrasensitive Samoa Human IL-33 Immunoassay INTRODUCTION Interleukin-33 (IL-33), a cytokine
AssayGate, Inc. 9607 Dr. Perry Road Suite 103. Ijamsville, MD 21754. Tel: (301)874-0988. Fax: (301)560-8288. Ultrasensitive Samoa Human IL-33 Immunoassay INTRODUCTION Interleukin-33 (IL-33), a cytokine
Ligand-mediated cytoplasmic retention of the Ah receptor inhibits. Gulsum E. Muku, Tejas S. Lahoti, Iain A. Murray, Michael A.
 Ligand-mediated cytoplasmic retention of the Ah receptor inhibits macrophage mediated acute inflammatory responses Gulsum E. Muku, Tejas S. Lahoti, Iain A. Murray, Michael A. Podolsky, Kayla Smith, Troy
Ligand-mediated cytoplasmic retention of the Ah receptor inhibits macrophage mediated acute inflammatory responses Gulsum E. Muku, Tejas S. Lahoti, Iain A. Murray, Michael A. Podolsky, Kayla Smith, Troy
Allergic diseases and treatment. Feng Qian ( 钱峰 )
") Allergic diseases and treatment Feng Qian ( 钱峰 ) fengqian@sjtu.edu.cn Hypersensitivity Antigen Host Adaptive responses protective to the host harmful to the host The deleterious consequences of the adaptive
Allergic diseases and treatment Feng Qian ( 钱峰 ) fengqian@sjtu.edu.cn Hypersensitivity Antigen Host Adaptive responses protective to the host harmful to the host The deleterious consequences of the adaptive
Mouse Serum Anti-HDM IgE Antibody Assay Kit
 Mouse Serum Anti-HDM IgE Antibody Assay Kit Catalog # 3037 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Asthma is a common chronic inflammatory disease that affects 300 million people
Mouse Serum Anti-HDM IgE Antibody Assay Kit Catalog # 3037 For Research Use Only - Not Human or Therapeutic Use INTRODUCTION Asthma is a common chronic inflammatory disease that affects 300 million people
SUPPLEMENTARY INFORMATION. CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative. breast cancer patients
 SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Immunological Lung Diseases
 Emphysema and Fibrosis Universitätsklinik für Pneumologie Prof. Thomas Geiser Head Div. of Pulmonary Medicine and Laboratory of Lung Research, MU50 thomas.geiser@insel.ch The healthy lung: The pathway
Emphysema and Fibrosis Universitätsklinik für Pneumologie Prof. Thomas Geiser Head Div. of Pulmonary Medicine and Laboratory of Lung Research, MU50 thomas.geiser@insel.ch The healthy lung: The pathway
Supplemental Figure 1
 Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
Allergic rhinitis (Hay fever) Asthma Anaphylaxis Urticaria Atopic dermatitis
 Asthma Anaphylaxis Urticaria Atopic dermatitis") Hypersensitivity Disorders Hypersensitivity Disorders Immune Response IgE Disease Example Ragweed hay fever IgG Cytotoxic Immune complex T Cell Hemolytic anemia Serum sickness Poison ivy IgE-mediated Diseases
Hypersensitivity Disorders Hypersensitivity Disorders Immune Response IgE Disease Example Ragweed hay fever IgG Cytotoxic Immune complex T Cell Hemolytic anemia Serum sickness Poison ivy IgE-mediated Diseases
"Role for β2ar biased signaling to address the β-agonist controversy in asthma"
 "Role for β2ar biased signaling to address the β-agonist controversy in asthma" Julia K. L. Walker, Ph.D. Professor in Duke University School of Nursing and Pulmonary Medicine Support provided by NHLBI
"Role for β2ar biased signaling to address the β-agonist controversy in asthma" Julia K. L. Walker, Ph.D. Professor in Duke University School of Nursing and Pulmonary Medicine Support provided by NHLBI
EPIGENTEK. EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE
 Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE") EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric)
EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric)
Management of Bronchial Asthma in Adults..emerging role of anti-leukotriene
 Management of Bronchial Asthma in Adults..emerging role of anti-leukotriene Han-Pin Kuo MD, PhD Department of Thoracic Medicine Chang Gung University Chang Gung Memorial Hospital Taipei, Taiwan Bronchial
Management of Bronchial Asthma in Adults..emerging role of anti-leukotriene Han-Pin Kuo MD, PhD Department of Thoracic Medicine Chang Gung University Chang Gung Memorial Hospital Taipei, Taiwan Bronchial
Page 39 of 44. 8h LTA & AT h PepG & AT h LTA
 Page 39 of 44 Fig. S1 A: B: C: D: 8h LTA 8h LTA & AT7519 E: F: 8h PepG G: 8h PepG & AT7519 Fig. S1. AT7519 overrides the survival effects of lipoteichoic acid (LTA) and peptidoglycan (PepG). (A) Human
Page 39 of 44 Fig. S1 A: B: C: D: 8h LTA 8h LTA & AT7519 E: F: 8h PepG G: 8h PepG & AT7519 Fig. S1. AT7519 overrides the survival effects of lipoteichoic acid (LTA) and peptidoglycan (PepG). (A) Human
Minute TM Plasma Membrane Protein Isolation and Cell Fractionation Kit User Manual (v5)
") Minute TM Plasma Membrane Protein Isolation and Cell Fractionation Kit Catalog number: SM-005 Description Minute TM plasma membrane (PM) protein isolation kit is a novel and patented native PM protein
Minute TM Plasma Membrane Protein Isolation and Cell Fractionation Kit Catalog number: SM-005 Description Minute TM plasma membrane (PM) protein isolation kit is a novel and patented native PM protein
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
Topical nanocrystalline silver cream inhibits expression of matrix metalloproteinase-9 in animal models of allergic contact dermatitis.
 Topical nanocrystalline silver cream inhibits expression of matrix metalloproteinase-9 in animal models of allergic contact dermatitis. K C Bhol, P J Schechter NUCRYST Pharmaceuticals Inc, Wakefield, MA
Topical nanocrystalline silver cream inhibits expression of matrix metalloproteinase-9 in animal models of allergic contact dermatitis. K C Bhol, P J Schechter NUCRYST Pharmaceuticals Inc, Wakefield, MA
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration
 Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please