A DISSERTATION SUBMITTED TO THE PROGRAM IN CANCER BIOLOGY AND THE COMMITTEE ON GRADUATE STUDIES OF STANFORD UNIVERSITY
|
|
|
- Jason Fisher
- 6 years ago
- Views:
Transcription
1 LGR5 + INTESTINAL STEM CELLS AND THEIR MAINTENANCE A DISSERTATION SUBMITTED TO THE PROGRAM IN CANCER BIOLOGY AND THE COMMITTEE ON GRADUATE STUDIES OF STANFORD UNIVERSITY IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY Luis Alberto Chia August 2012
2 2012 by Luis Alberto Chia. All Rights Reserved. Re-distributed by Stanford University under license with the author. This dissertation is online at: ii
3 I certify that I have read this dissertation and that, in my opinion, it is fully adequate in scope and quality as a dissertation for the degree of Doctor of Philosophy. Calvin Kuo, Primary Adviser I certify that I have read this dissertation and that, in my opinion, it is fully adequate in scope and quality as a dissertation for the degree of Doctor of Philosophy. Anson Lowe I certify that I have read this dissertation and that, in my opinion, it is fully adequate in scope and quality as a dissertation for the degree of Doctor of Philosophy. Julien Sage I certify that I have read this dissertation and that, in my opinion, it is fully adequate in scope and quality as a dissertation for the degree of Doctor of Philosophy. Marius Wernig Approved for the Stanford University Committee on Graduate Studies. Patricia J. Gumport, Vice Provost Graduate Education This signature page was generated electronically upon submission of this dissertation in electronic format. An original signed hard copy of the signature page is on file in University Archives. iii
4 ABSTRACT The intestinal epithelium is one of the most rapidly proliferating tissues in the body. A complete turnover of the epithelium occurs every 3-5 days in the mouse, a process that is maintained by a small population of intestinal stem cells (ISCs) that reside in the crypt bases. The signals that regulate the behavior of these ISCs are still poorly understood. However, the recent identification of genes that mark functional stem cells has yielded insights into how ISCs are regulated and maintained. Here, we demonstrate that Bmi1 and Lgr5 mark two functionally distinct ISCs in vivo. Lgr5 marks mitotically active ISCs that exhibit exquisite sensitivity to canonical Wnt modulation, contribute robustly to homeostatic regeneration, and are quantitatively ablated by irradiation. In contrast, Bmi1 marks quiescent ISCs that are insensitive to Wnt perturbations, contribute weakly to homeostatic regeneration, and are resistant to high dose radiation injury. Post-irradiation, however, the normally quiescent Bmi1 + ISCs dramatically proliferate to clonally repopulate multiple contiguous crypts and villi. Clonogenic culture of isolated single Bmi1 + ISCs yields long-lived self-renewing spheroids of intestinal epithelium that produce Lgr5-expressing cells, thereby establishing a lineage relationship between these two populations in vitro. Taken together, these data provide direct evidence that Bmi1 marks quiescent, injury-inducible reserve ISCs that exhibit striking functional distinctions from Lgr5 + ISCs, and support a model whereby distinct ISC populations facilitate homeostatic versus injury-induced regeneration. How these ISCs maintain their stemness remains unclear. Paneth cells have been suggested to serve as niche cells for the Lgr5 + ISCs, perhaps through the secretion of IV
5 essential paracrine factors, but recent reports clearly demonstrate that Paneth cells are not required for Lgr5 + ISC maintenance. Recently, the G-protein-coupled receptors Lgr4-6 were reported to associate with Wnt receptors to mediate R-spondin signaling. Given the importance of Wnt/R-spondin signaling in intestinal crypt maintenance, we tested the in vivo function of Lgr5 by adenovirus-mediated overexpression of the soluble ligandbinding Lgr5 extracellular domain. Circulating Lgr5 ectodomain induced the migration of Paneth cells from the crypts, their eventual loss, and a concomitant disappearance of Lgr5 + ISCs. Paneth cell migration was associated with downregulation of Wnt signaling and its target EphB3. The loss of Lgr5 + ISCs did not affect maintenance of the intestinal epithelium, nor did Lgr5 + stem cells disappear from non-intestinal organs. Together, these findings characterize an easily tractable experimental model for the in vivo deletion of Lgr5 + ISCs, and suggest that Lgr receptors function to actively maintain Lgr5 + ISCs in vivo. V
6 DEDICATION This thesis is dedicated to my and, who encouraged me to pursue my passions, instilled in me a hunger for knowledge and truth, and supported me every step of the way. This thesis is also dedicated to my friends, who not only motivated me over the years, but also inspired and reminded me to stay hungry. VI
7 ACKNOWLEDGEMENTS This work could not have been accomplished without the support, help, and encouragement from numerous individuals. To my thesis advisor Calvin Kuo for providing me with an excellent atmosphere to do research and the freedom to follow my instincts. His passion, enthusiasm, patience, guidance, and unwavering support of my scientific pursuits truly distinguish him from other advisors. To Akifumi Ootani, mentor and friend, for planting the seeds for what would eventually evolve into my thesis project, and for taking me around his hometown of Saga, Japan to indulge in the best sushi and ramen I have ever had. To Kelley Yan, friend, mentor, and queen of the FACS machine, for thoughtful discussions, sound advice, and a timely introduction to Mount Veeder and Howell Mountain. She has played an instrumental role in my development as a scientist and in the completion of this dissertation. Kelley s pure love for science and unwavering belief in the good of science continues to inspire me. She is truly one of the most talented individuals that I know. I am forever thankful that she let me shadow her and learn from her. It has been a privilege to work with her. To my committee members Anson Lowe, Julien Sage, and Marius Wernig for their encouraging words, insightful questions, and thoughtful criticisms. To Amato Giaccia for enthusiastically agreeing to chair my thesis defense and for his great advice. To the members of the Kuo Lab, past and present, for their friendship and expertise. I want to especially thank Michael Mancuso, Lisa McGinnis, and Kevin Wei VII
8 for paving the way, and Terry Reyes for finishing strong. I also want to acknowledge Carol Chan, David Corney, Frank Kühnert, Valerie Hilliard, Cindy Kosinski, Xingnan Li, Lincoln Nadauld, James Neal, and Mario Vallon, for their knowledge and expertise, and for ensuring that I did not reinvent the wheel. I additionally want to recognize the technical contributions of Lauren Tracy and Jenny Yuan. Lastly, I want to thank Sebastian Lim and Rachel Wolfson for reminding me about how inspiring science can be. To the Cancer Biology Program, Dali Ma, Grace Kolar, Anita Blanco, Tenea Nelson, Anika Green, Kelly Haehnel, and Jenny Hu for everything that you do to make sure that I would graduate. To Rolando, Will, and Reynaldo for letting me keep my harem. To Chelsey Simmons and Cody Simmons for believing in me and being awesome partners in crime. To my friends for allowing me to make mistakes and loving me anyway. I especially want to acknowledge Brian Chi, Lindsay Docto, Aaron Hoo, Andrew Huang, Elaine Khoong, Christina Li and Annie Oh for always being there. To my mom, dad, and Lucia for their unconditional love and support even though they didn t understand what it was that I was doing. We came to the United States with nothing but a suitcase and a dream, and somehow, with a little bit of luck, I found myself here. Last but not least, to Ms. Fordham, my fourth grade teacher, for believing in my potential and starting it all. VIII
9 PUBLICATION OF CHAPTERS AND ROLE OF AUTHOR Chapter 1 Previously published review: Chia LA, Kuo CJ. The Intestinal Stem Cell. Prog Mol Biol Transl Sci. 2010;96: Chapter 2 Previously published paper: Yan KS, Chia LA, Li X, Ootani A, Su J, Lee JY, Su N, Luo Y, Heilshorn SC, Amieva MR, Sangiorgi E, Capecchi MR, Kuo CJ. The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations. Proc Natl Acad Sci U S A Jan 10;109(2): Chapter 3 This chapter, written by Luis A. Chia is unpublished. IX
10 TABLE OF CONTENTS Abstract... IV Acknowledgements...VII Publication of chapters and role of author... IX Table of Contents...X List of Illustrations... XI Chapter 1: Introduction Abstract The Intestinal epithelium Monoclonality of intestinal crypts The identity and localization of intestinal stem cells Lgr5 marks CBC cells that function as ISCs Bmi1 marks + 4 cells that function as ISCs Other putative intestinal stem cell markers The intestinal stem cell niche Wnt signaling Notch signaling BMP signaling Hedgehog signaling Epidermal Growth Factor signaling Evidence for a non-mesenchymal niche Intestinal stem cells as the cell origin of colorectal cancer...20 Chapter 2: The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations Abstract Introduction Results Discussion Methods Figures Figure legends Supplementary figures Supplementary figure legends...55 Chapter 3: Functional loss of Lgr5 + ISCs and Paneth cells by the overexpression of the Lgr5 extracellular domain Abstract Introduction Results Discussion Methods Figures Figure legends Supplementary figures Supplementary figure legends...82 References...83 X
11 LIST OF ILLUSTRATIONS Chapter 1 Figure 1: Architecture of the intestinal epithelium...4 Chapter 2 Figure 1: Basal proliferation status and response to canonical Wnt pathway modulation of Lgr5 + versus Bmi1 + ISCs...42 Figure 2: Differential responses of Lgr5 + versus Bmi1 + ISCs to acute radiation injury...43 Figure 3: Differential responses of Lgr5 + versus Bmi1 + lineages to acute radiation... injury...44 Figure 4: Clonogenic culture of single FACS-isolated Bmi1-YFP + ISCs...45 Supplemental Figure S1: Quantitative effects of canonical Wnt pathway modulation on Lgr5 + versus Bmi1 + ISCs...50 Supplemental Figure S2: Bmi1 + lineage proliferates in response to irradiation injury...51 Supplemental Figure S3: Serial passaging of Bmi1-YFP + clonogenic spheroids...52 Supplemental Figure S4: Lgr5-expressing cells arise within Bmi1-YFP + clonogenic Spheroids...53 Supplemental Figure S5: Chapter 3 Figure 1: In vivo overexpression of the Lgr5 ectodomain induces migration and eventual loss of Paneth cells by inhibition of EphB3 expression in Paneth cells...74 Figure 2: In vivo overexpression of Lgr5 ECD induces functional loss of Lgr5 + ISCs...75 Figure 3: Lgr5 ECD-mediated Lgr5 + ISC ablation induces activation and de novo generation of Bmi1 + ISCs in vivo...76 Supplemental Figure S1: Lgr4 ECD treatment does not affect Lgr5 expression...79 Supplemental Figure S2: Lgr4 ECD treatment does not affect Paneth cell localization...80 Supplemental Figure S3: Lineage of preexisting Bmi1 + ISCs decreases after Ad Lgr5 ECD treatment...81 XI
12 CHAPTER 1 INTRODUCTION 1
13 1.1 ABSTRACT The intestinal epithelium is one of the most rapidly proliferating tissues in the body. A complete turnover of the epithelium occurs every 3-5 days in the mouse, a process that is maintained by a small population of intestinal stem cells (ISCs) that reside in the crypt bases. The signals that regulate the behavior of these ISCs are still unknown. This has been due, until recently, to the singular lack of definitive ISC markers. The recent identification of genes that mark functional stem cells has yielded insights into how ISCs are regulated and maintained by their surrounding niche. Herein, we examine the body of literature regarding the precise identity and location of the ISCs, the role of the surrounding niche in ISC maintenance and regulation, as well as the hypothesis that the ISCs are the cells of origin in colorectal cancer. 2
14 1.2 The Intestinal Epithelium The adult intestinal epithelium has a well-defined organizational structure that can be divided into a functional (villus) and proliferative (crypt) region that is comprised of differentiated and undifferentiated cells, respectively. It is one of the most rapidly proliferating organs and complete turnover of the epithelium occurs every 5-7 days in an orderly fashion along the crypt-villus axis. This process is ultimately sustained by a small population of crypt-residing intestinal stem cells (ISCs) that give rise to a pool of multipotent progenitors, which do not differentiate immediately, but instead rapidly proliferate to create a pool of transit amplifying (TA) cells. These TA cells occupy the crypt length and subsequently give rise to the differentiated lineages of the intestine: the absorptive enterocytes, the mucin secreting goblet cells, the peptide hormone secreting enteroendocrine cells, the Paneth cells, M cells, tuft cells, and cup cells. The intestinal crypts are enclosed within a fenestrated sheath of intestinal subepithelial myofibroblasts (ISEMFs) that have features of both fibroblasts and smooth muscle cells. Originally, these ISEMFs were largely thought of as a sheath of fibroblasts that were denser near the crypts than at the surface of the colon or in the villi of the intestine. However, it is now clear that they exist as a syncytium that extends throughout the lamina propia where it merges with the pericytes of the blood vessels that are found throughout the tissue. ISEMFs are!-smooth muscle actin (!-SMA) positive and can be distinguished from the!-sma-positive smooth muscle of the muscularis mucosa by their negative staining for desmin. The development of the intestinal mucosa largely takes place in two broad steps: 1) organogenesis, during which interactions between the embryonic endoderm and 3


15 mesoderm lead to the formation of the forgut, midgut, and hindgut; and 2) cytodifferentiation, in which the ISCs proliferate and differentiate into the four lineages of the intestinal epithelium. Both of these processes are complex, and there is accumulating evidence that the ISEMFs, as well as the epithelium, play an active role in the development of the intestinal mucosa. Until recently, progress in understanding how the ISCs are maintained and regulated has been hampered by the lack of definitive ISC markers and in vitro culture methodologies. Despite this, work in the last 40 to 50 years has made significant headway in terms of identifying the actual ISCs and yielded many insights into how the ISCs are maintained and regulated by their surrounding niche. In fact, the recent identification of genes that functionally mark ISCs and the development of robust tissue culture methodologies has allowed for the emergence of evidence that challenges established paradigms regarding ISC maintenance and regulation.!"!"!"!"!"!"!"!" /*(&+9"3&..8" "C6"3&..8"!"!"!"!"!"!"!"!"!"!"!"!"!"!"!"!" #$%&'&()*+&,"-$..$" 6(0$7$8" "CDE"3&..8"!"!"!"!"!"!"!"!" "F(+&'0&(,0G'$(&"3&..8" H0I.&+"3&..8" 6I80'+5)2&"F(+&'0G4+&8" /'0.$1&'*)2&"3'45+" J40KI'0I.*8+8" Figure 1: Architecture of the intestinal epithelium. 4
16 1.3 Monoclonality of Intestinal Crypts Numerous experiments in both mice and humans have shown that intestinal crypts are clonal populations, which are ultimately derived during development from a single cell. By using mice heterozygous for a defective glucose-6-phosphate dehydrogenase (G6pd) gene, Griffiths et al. demonstrated that the colonic crypts from adult mice were monoclonal. They showed that when compared to intestine from mice homozygous for the defective G6pd gene, which uniformly expressed low enzyme activity, heterozygous mice had large patches of intestine with high and low enzyme activity. Since they found no crypts that exhibited mixed enzyme activity, these results indicated that all epithelial cells in a crypt were derived from a single cell(1). Similarly, Bjerknes and Cheng also confirmed the clonality of crypts by using somatic mutagenesis following the treatment of mice with N-nitroso-N-ethylurea. Specifically, N-nitroso-N-ethylurea induces the random expression of Dolichos biflorus agglutinin (DBA) binding sites on mutated cells and allows for the binding of lectin. In these studies, mutated long-lived cells gave rise to ribbons of lectin-positive cells that migrated up the villus. Some ribbons contained all of the differentiated lineages of the epithelium, suggesting that these cells originated from mutated intestinal stem cells. However, some ribbons only contained columnar or mucinous cells, indicating that these cells originated from committed progenitors(2). Together, these findings support the existence of a hierarchy of pluripotent and committed progenitors within the intestinal crypt. As in the mouse, analysis of intestinal tissues from an XO/XY mosaic individual of male phenotype by nonisotopic in situ hybridization showed that human intestinal 5
17 crypts were also clonal. In other words, the crypts from the XO/XY individual were composed almost exclusively of either XY or XO cells. Indeed, the examination of crypts at patch borders revealed no mixed XO/XY crypts(3). Thus, the evidence implies that both mouse and human crypts are clonal populations. However, these conclusions are only fitting for adult animals since intestinal crypts from neonatal mice have been found to be polyclonal(4). In other words, they show a mixed phenotype, but become monophenotypic within 2 weeks through a process of crypt purification, though the precise mechanisms regulating this process are still unclear. Importantly, the establishment that crypts are clonal does not imply that crypts only contain a single stem cell. In fact, the precise number of stem cells per crypt and what controls their numbers is heavily debated. For example, it has been suggested that as many as 4-16 actual stem cells and potential stem cells may exist in the small intestinal crypt(5). In contrast, colonic crypts may only contain as few as three or four actual stem cells. 1.4 The Identity and Localization of Intestinal Stem Cells Despite a lack of distinctive ISC markers, it was realized early on that ISCs must locate to specific sites, such as the origin of cell efflux. Since the intestinal epithelium is divided into discrete units of proliferation (crypts) and differentiation (villi), and because migration begins at the base of the crypt, it was believed that basal crypt cells, or a subset thereof, were candidate ISCs. In 1974, Cheng and Leblond demonstrated in a series of landmark papers that the four differentiated lineages of the adult intestinal epithelium originated from cells that are located between and immediately above the Paneth cells, which they named crypt-base columnar (CBC) cells. After injecting mice with 3 H- 6
18 thymidine ([ 3 H]dT), which becomes incorporated with the DNA of replicating cells and induces local irradiation; Cheng and Leblond observed that some CBC cells phagocytosed neighboring cells(6-10). As a result, these CBC cells subsequently contained phagosomes that were then used as a cytosolic marker to follow their evolution. The subsequent identification of phagocytic fragments in all four differentiated lineages supported their common origin as well as the Unitarian theory of epithelial cell formation in the mouse intestine. Notably, this hypothesis agreed with the hierarchical organization of cell populations in other tissues, such as the hematopoeitic system. At the time however, the data supporting this hypothesis was equivocal. For example, data presented actually showed that more cell death occurred at cell positions 4-6 than at the crypt base, which was consistent with observations in crypts from mice that were externally irradiated. In 1975, Cairns first proposed the immortal strand hypothesis in which stem cells repeatedly segregate their chromosomes asymmetrically(11). It was argued that such a mechanism would have evolved to minimize the risk of incurring DNA replicationinduced errors in cells that are long lived, since such mutations would be passed on to daughter cells that would terminally differentiate. The inability of cells at cell position 4-5 to retain [ 3 H]dT or bromodeoxyuridine (BrdU) label through many rounds of division (10-15) during normal homeostasis, and the ability to generate label-retaining cells (LRCs) when mice were exposed to label while making new stem cells, lends support to the immortal strand hypothesis and suggests that a population of + 4 cells are the actual ISCs(12). In fact, in double label experiments, it was shown that [ 3 H]dT-LRCs have the ability to incorporate BrdU(13). Analysis of such double-labeled cells showed that both 7
19 labels were differentially segregated. Specifically, BrdU disappeared from the LRCs after the second round of cell division, while the [ 3 H]dT persisted through many rounds of cell division. Of note, whereas these LRCs were found most frequently at the + 4 region, they in fact consisted of cells located between the Paneth cells and as high as cell positions above the crypt base. 1.5 Lgr5 marks CBC cells that function as ISCs The recent identification of the leucine-rich G protein-coupled receptor 5 (Lgr5), a Wnt target gene that encodes a G protein coupled receptor of unknown function, as a putative marker of ISCs strengthened the hypothesis that CBC cells were the actual stem cells of the intestinal epithelium. These cycling cells are located between the Paneth cells in the crypt bases, have been convincingly shown to mark cells that give rise to the four differentiated lineages of the intestinal epithelium, and possess the ability to regenerate the intestinal epithelium for extended periods of time (>12 months)(14). Recently, Sato and Clevers demonstrated that over a two-week period, single Lgr5-positive cells can be cultured, undergo proliferation and crypt fission, and produce organoids that contain several crypt-like structures when supplemented with numerous growth factors including the Wnt agonist R-spondin1, EGF, Jagged peptide, and Noggin (15). One interpretation of these results is that the proliferation and organization of the Lgr5-positive-derived organoids does not require the presence of a mesenchymal niche and potentially suggests the presence of an epithelial niche for the ISCs. However, it is also possible that the numerous supplementary factors in the Sato cultures, which regulate the Wnt, Notch, and BMP pathways, mimic signals presented by the mesenchyme. Further, the growth of Sato cultures in Matrigel, which is a poorly defined matrix that contains laminin, collagen IV, 8
20 EGF, insulin-like growth factor, tissue plasminogen activator, as well as TGF-", all of which are expressed by myofibroblasts, also suggests that although the physical presence of mesenchymal niche cells may not be essential, the products they secrete may be required. Interestingly, a recent study by Sato and colleagues supports the role of Paneth cells as essential epithelial components of the ISC niche. Since every Lgr5- expressing cell is intimately associated with a Paneth cell at the crypt base, these investigators co-cultured CD24 + Paneth cells with Lgr5 + ISCs with a 10-fold increased efficacy of organoid growth compared to that of single Lgr5 + ISCs. Furthermore, gene expression profiling of CD24 + Paneth cells showed high-level expression of growth factors known to be important for in vitro organoid growth including Wnt3a and EGF. These results support the epithelial cell niche role of Paneth cells. 1.6 Bmi1 marks + 4 cells that function as ISCs At the same time, there is growing evidence that cells at cell position + 4, abutting the uppermost Paneth cell at a position 4 cells above the crypt base (defined as + 1) are also putative ISCs. Bmi1, a member of the Polycomb group family of transcriptional repressors, was recently identified as a marker of ISCs using a lineage tracing strategy and further strengthened the notion that cells at the + 4 position also serve as the actual stem cells of the small intestine. Bmi1 + cells primarily localize to the + 4 position and similarly have the ability to expand, self-renew, and give rise to all of the differentiate lineages of the small intestine(16). It was unknown whether Bmi1 + cells give rise to Lgr5 + or vice versa. One possibility is that Lgr5 and Bmi1 identify two distinct populations of ISCs. The distinct expression pattern of these two genes certainly supports this hypothesis; Lgr5 is 9
21 expressed throughout the small and large intestine(14), while Bmi1 expression is restricted to the proximal small intestine(16). Moreover, emerging evidence reveals that both quiescent and active stem cell subpopulations may coexist within a tissue(17). For example, in the hair follicle, CD34 + LRC bulge cells have been functionally defined as hair follicle stem cells by their ability to reconstitute the entire hair follicle(18, 19). These CD34 + LRCs retain label for many months, suggesting that they exist in a quiescent state and are not actively involved in hair regeneration(20). Recently, it was shown that these bulge stem cells do not directly generate transit-amplifying cells, but rather give rise to an intermediate stem cell population located at the hair germ, which expresses Lgr5, is actively cycling, and can also reconstitute the entire hair follicle(21, 22). These findings demonstrate that the hair follicle contains both quiescent and actively cycling populations of stem cells that reside in separate yet adjacent locations. Studies of HSC also demonstrate that both active and quiescent populations exist in the hematopoietic system(18, 23). Thus, it is possible that Bmi1 + cells, though unknown whether they can retain label, and Lgr5 + cells represent quiescent and actively cycling ISCs, respectively. Clearly, the relationship between these two cell populations requires further investigation. 1.7 Other Putative Intestinal Stem Cell Markers Musashi-1 (Msi-1) was proposed as a putative ISC marker prior to the identification of Lgr5 and Bmi1 as bona fide stem cell markers. Msi-1 encodes an RNAbinding protein that was initially found to be associated with early asymmetric division in sensory organ precursor cells in Drosophila and is believed to modulate Notch signaling by suppressing the expression of Numb mrna synthesis(24, 25). Analysis of Msi-1 protein expression by immunohistochemistry revealed that a small number of cells at cell 10
22 position 4-5 in the adult small intestine and a few cells at the base of the crypt in the large intestine were positive for Msi-1(26). Additionally, strong Msi-1 expression was observed in developing crypts, post-irradiation regenerating crypts, and in early adenomas(26). Together these data suggested that Msi-1 could mark ISCs. However, the ability of these Msi-1 cells to give rise to all of the differentiated lineages of the intestine has not yet been demonstrated. Likewise, the microtubule-associated kinase DCAMKL-1 (Doublecortin and CaM Kinase-like1) was also proposed as a candidate ISC marker(27). DCAMKL-1 is expressed in cells near position 4 in normal intestinal crypts, and these cells were found to be Msi-1 positive(26). DCAMKL-1 + crypt epithelial cells were shown to retain BrdU in a modified label retention assay. One group even demonstrated the ability of these cells to self-renew and form spheroids in suspension culture(28). However, robust lineage tracing experiments have not been performed and it remains unclear whether these cells can give rise to all of the differentiated lineages of the intestinal epithelium. Another putative ISC marker is mouse telomerase reverse transcriptase (mtert), which is a gene that is downregulated upon differentiation in most somatic cells. In the intestine, the analysis of mtert-gfp mice showed that GFP specifically marks long-term BrdU-retaining intestinal crypt cells(29). Though its ability to functionally mark ISCs has not yet been determined, the availability of the mtert-gfp mice and methodology to culture single ISCs via the Sato method, suggests that mtert + ISCs can be prospectively isolated and assayed for its stem cell activity in vitro. Other putative ISC markers include: BMPR1A, phosphorylated-pten, phosphorylated-"-catenin, Wip1 phosphatase, phosphorylated-akt, Apex1, Diap3, 11
23 Gemin4, Rhobtb3, and Wdrl2(30). Whether these genes mark ISCs that can give rise to all of the differentiated lineages of the intestine requires further investigation. Importantly, the lineage tracing methodologies used to identify Lgr5 as a stem cell marker, have enabled the identification of additional markers such as Olmf4 and Ascl2(31), though these markers have not been themselves confirmed through lineage tracing studies. Regardless, the association of the expression of these genes with + 4 cells or cells in the lower regions of the crypt have yielded some interesting insights. For example, the Wnt target Achaete scute-like 2 (Ascl2) controls the fate of Lgr5 + ISCs. Specifically, conditional deletion of Ascl2 in the adult small intestine induces the disappearance of the Lgr5 + ISCs. Accordingly, the transgenic overexpression of Ascl2 throughout the intestinal epithelium induces crypt hyperplasia, as well as the appearance of ectopic crypt like structures in the villi(31). These reports are in accordance with findings about the importance of Wnt signaling in maintaining the ISC niche and the ISCs. 1.8 The Intestinal Stem Cell Niche In 1978, Schofield introduced the idea of a stem cell niche when he proposed a hypothesis in which hematopoietic stem cells (HSCs) are seen in association with other cells, which confer HSCs with their stem cell-like behavior. Schofield initially described the stem cell as a fixed tissue cell, whose maturation and continued proliferation as a stem cell is respectively prevented and assured(32). According to this hypothesis, the progeny of HSCs that do not occupy a similar stem cell niche are considered first generation colony-forming cells that proliferate, mature, and differentiate. 12
24 In the intestine, the stem cell niche is likely comprised of epithelial, subepithelial, and luminal components. Mucosal constituents include the permeable basement membrane and the cells that are located beneath it, such as mesenchymal cells (myofibroblasts, fibroblasts and smooth muscle cells), endothelial cells, neural cells, and immune cells. All of these cells may regulate the function of stem cells via secreted proteins or their direct interaction with the stem cells themselves. For example, it is known that Wnt signaling is important for the maintenance of the intestinal stem cells and that the epithelial and mesenchymal cells differentially express specific components of the Wnt signaling pathway, suggesting that mesenchymal-epithelial crosstalk is required for the maintenance of the intestine(33-35). On the other hand, the niche may also be comprised of luminal components, which may be derived from epithelial cells or from the large number of bacteria present in the lumen. For example, the mucin glycoprotein Muc2 appears to play a crucial role in stem cell maintenance as Muc2-deficient mice have been reported to exhibit aberrant crypt morphology and altered cell maturation and migration(36). Though considerable knowledge about the niche has been gained in the past few years, the precise components of the niche largely remain unknown because the identification and exact location of the intestinal stem cell is still under intense investigation. Still it has emerged that several signaling pathways likely play a role in the maintenance and regulation of the ISCs and their differentiated progeny, such as the Wnt, Notch, BMP, Hh, and EGFR signaling pathways. 13
25 1.9 Wnt Signaling The Wnt signaling pathway is critical during embryonic development and organogenesis in many species. Wnts are evolutionarily conserved, cysteine-rich glycoproteins that signal in both a paracrine and autocrine fashion. There are 19 known murine Wnt genes, which bind to the seven-pass transmembrane protein receptors of the frizzled (Fz) family, 10 of which have been identified in mice, and to the single-span low-density lipoprotein receptor-related proteins (LRP). When the Wnt ligand binds to the Fz/LRP complex, this triggers the release of "-catenin from a so-called destruction complex, which is comprised of glycogen synthase kinase 3-" (GSK3-"), axin, and the adenomatous polyposis coli (APC) tumor-suppressor protein. The free "-catenin then accumulates and translocates to the nucleus, where it associates with DNA-binding factors of the T-cell factor/lymphocyte enhancer factor (TCF/LEF) family to activate the transcription of target genes including c-myc, cyclin D1, CD44, c-jun, and Lgr5. Importantly, Wnts also stimulate cellular responses independently of "-catenin and TCF through the so-called non-canonical pathway, which involves the intracellular release of Ca2 + and/or the planar polarity pathway. Previous studies have demonstrated the importance of the Wnt signaling pathway in the maintenance of the proliferative compartment of the intestinal epithelium. For example, the systemic expression of Dkk1, a Wnt inhibitor, by adenoviral infection results in crypt loss and the subsequent degeneration of the intestinal epithelium(35). Likewise, transgenic expression of Dkk1 under the control of the villin promoter (37) or disruption of Tcf7l2 (Tcf4), a downstream effector of Wnt signaling, in mice, specifically depletes the epithelial stem cell compartment of the small intestine without disrupting the 14
26 induction of epithelial cells from endoderm, as demonstrated by the presence of differentiated enterocytes and goblet cells in Tcf7l2 -/- embryos(34). In colorectal cancer cells, disruption of "-catenin/tcf signaling results in rapid cell cycle arrest. Furthermore, induced deletion of the Wnt target gene c-myc in adult mice results in rapid loss of intestinal crypts(38). Accordingly, the injection of human R-spondin 1, a Wnt agonist, into mice results in the rapid onset of crypt cell proliferation that is accompanied by "- catenin stabilization, as well as increases in the number of crypt based columnar cells in the crypts (unpublished results). These data suggest that Wnt signaling is required for the maintenance of ISCs. In addition to maintaining homeostasis, the Wnt signaling pathway also plays a key role in malignant transformation. Mutations of the APC tumor suppressor gene are found in approximately 60-80% of human sporadic colorectal tumors(39, 40). This mutation inhibits the cytosolic degradation of "-catenin by the GSK3-"/axin/APC destruction complex, which results in the accumulation of nuclear "-catenin and the subsequent upregulation of pro-proliferation genes that are induced by TCF/LEF transcription Notch signaling In other systems, the Notch signaling pathway has been shown to be crucial in regulating cell fate decisions. In the intestine, Notch receptors, the ligand Delta, and the associated Hes transcription factors are chiefly expressed at the base of the intestinal crypts. The Notch genes encode transmembrane receptors that interact with ligands, which are also transmembrane proteins that are located on adjacent cells. In vertebrates, there are four receptors and five ligands. Receptor-ligand interaction results in cleavage 15
27 of the intracellular domain of the Notch receptor, its transduction to the nucleus, and transcriptional activation. It is believed that the cell contact dependent nature of Notch signaling enables the coordination of proliferative/differentiative decisions and cell fates in a group of otherwise uncommitted cells. In the intestine, maintenance of the proliferative compartment requires the constitutive activation of the Notch signaling cascade. Inhibition of the pathway by the conditional inactivation of the downstream transcription factor CSL/RBP-J or the injection of a #-secretase inhibitor, results in the massive conversion of proliferative crypts into post-mitotic goblet cells(41). Accordingly, the requirement for Notch signaling in the maintenance of stem cell proliferation during intestinal tumorigenesis was also demonstrated; #-secretase inhibition in Apc Min mice caused widespread Goblet cell conversion within adenomas(41). Furthermore, the analysis of mice deficient for the basic helix-loop-helix proteins Hes-1, Math-1, and neurogenin-3, all of which are transcriptional targets of Notch signaling in other tissues, have indirectly implicated the Notch signaling pathway in the regulation of the earliest intestinal cell fate decisions(42-44). For example, deletion of Math-1 results in the absence of goblet, Paneth, and enteroendocrine cell lineages, with enterocytes being the only differentiated cell type in the murine small intestine(45) BMP Signaling Bone morphogenetic proteins (BMPs) belong to the transforming growth-" superfamily of proteins whose signaling is initiated by binding to BMP receptor types I or II (BMPR1 or BMPR2). This interaction leads to the phosphorylation of SMAD1, 5, or 8, which heterodimerizes with SMAD4, translocating to the nucleus where they act as 16
28 transcriptional activators. Active BMP signaling, as indicated by the presence of phosphorylated SMADs, is found predominantly in differentiated intestinal epithelial cells. Accordingly, expression of BMPs is highest at the top of intestinal crypts, whereas BMP antagonists, such as Gremlin 1 and 2 and Noggin, are most strongly expressed at the crypt base. BMPs appear to antagonize Wnt signaling, via the PTEN tumor-suppressor protein, and so permit and restrict differentiation and proliferation, respectively. Conditional mutation of BMPR1A resulted in de novo crypt formation and a juvenile polyposis phenotype(46). Correspondingly, there was also reduced differentiation of the intestinal epithelium. As expected, inhibition of BMP signaling by conditional ectopic expression of the BMP-antagonist Noggin also increased proliferation, and led to de novo crypt formation and polyposis(47). Likewise, Gremlin 1, a BMP antagonist, was shown to partially inhibit cell differentiation in vitro and promote proliferation. Consistent with these observations, stimulation of colon cancer cell lines by BMP2, a BMP agonist, inhibited growth(48, 49) Hedgehog signaling The hedgehog (Hh) signaling pathway has also been implicated in the maintenance of the intestinal epithelium. Mice deficient in Sonic (Shh) or Indian (Ihh) hedgehog exhibit intestinal abnormalities. For example, Ihh -/- mice die perinatally and exhibit reduced proliferation in the intervillus region as well as a depleted progenitor cell compartment, suggesting that Ihh is important in the maintenance of ISCs. In contrast, Shh -/- mice display overgrown duodenal villi and stomach epithelium. These findings suggest that Shh may inhibit rather than stimulate proliferation in these regions. 17
29 Nonetheless, both models exhibit reduced smooth muscle, suggesting that Shh and Ihh have partially redundant functions (50). Of note, unlike the other previously described pathways, the direction of hedgehog signaling is reversed. In the intestine, both Sonic (Shh) and Indian (Ihh) hedgehog are initially expressed throughout the intestinal epithelium. After villus formation, both proteins are redistributed becoming concentrated in the epithelial cells of the intervillus region. By using villin-hhip mice, transgenic mice expressing the pan-hedgehog inhibitor, Hhip (hedgehog interacting protein), under the control of the villin promoter, Madison et al. showed that Hedgehog signaling occurs in a paracrine fashion from the epithelium to the Ptch1-expressing ISEMFs and smooth muscle cells(51). Consequently, Hh signaling is not directly involved in the fate of the epithelial cells but is important in the overall organization of the crypts and villi of the intestinal mucosa. Moreover, it has been demonstrated that Hh signaling is an antagonist of Wnt signaling, as the transfection of Indian hedgehog and TCF4 resulted in the downregulation of TCF4 and restoration of Ihh, respectively(52). Thus, it appears that hedgehog signaling functions to restrict Wnt signaling to the crypt base allowing for the indirect regulation of the ISCs Epidermal Growth Factor Signaling EGFR appears to be required for epithelial homeostasis in the mouse GI tract, and EGFR ligands are required for the development and maintenance of the intestine. For example, mice lacking functional EGFR have been shown to develop disorganized crypts(53). In Drosophila, inactivation of EGFR inhibits the growth and division of ISCs(54). These studies demonstrate the essential role that EGFR signaling plays in intestinal homeostasis. Interestingly, genetic studies have shown that ectopic activation 18
30 of the EGFR pathway can accelerate tumor progression in the APC min/+ background(55-57). Accordingly, partial loss of function of EGFR has also been reported to significantly reduce adenoma formation in Apc min/+ mice(58). Furthermore, antibodies targeting EGFR have also been shown to be effective in treating colorectal cancer provided there are no activating mutations in downstream signaling components, such as KRAS or BRAF(59, 60) Evidence for a Non-mesenchymal Niche A model whereby stem cells receive signals from a mesenchymal niche and a stem cell migrates away from its niche to differentiate appears to hold true in the hematopoietic and hair follicle system. However, in the intestine, emerging evidence suggests that novel mechanisms may exist for ISC regulation. Work in Drosophila has revealed for example, that ISCs actively select the fate of their progeny independently of a detectable stem cell niche. In this model, the selective expression of the Notch ligand Delta, by the ISCs, functions to activate Notch signaling in adjacent daughter cells to determine cell fate(61). In fact, studies have indicated that during intestinal organogenesis, a founder adult midgut progenitor (AMP) undergoes asymmetric division and signals via the Notch signaling pathway to direct its first daughter cell to become a peripheral cell, which then act as a niche for AMPs to keep them undifferentiated until metamorphosis. During metamorphosis, the peripheral cells break down and allow the AMPs to respond to Notch signaling and differentiate into the differentiated lineages of the intestinal epithelium(62). Accordingly, studies have shown that the Drosophila EGFR pathway is essential for ISC proliferation during both normal midgut homeostasis and regeneration after 19
31 damage. The analysis of EGFR ligand expression demonstrated that the EGFR ligand Vn was expressed in the visceral muscle during gut regeneration, suggesting that the visceral muscle might serve as part of the ISC niche. However, the specific downregulation of Vn in the visceral muscle did not affect ISC proliferation, demonstrating that visceral muscle-derived Vn is probably not essential for the maintenance of ISCs(54). Interestingly, the expression of Spi and Krn, two other EGFR ligands, in midgut epithelial cells suggests that these epithelial-derived EGFR ligands may be responsible for activating EGFR signaling in the ISCs. Together these findings support the hypothesis that the epithelium itself may represent a critical component of the ISC niche. Therefore, it is tempting to speculate that epithelial cell types may be a critical component of the mouse intestinal niche. In murine small intestine, Paneth cells have been shown to specifically secrete the canonical Wnt ligands, Wnt-3 and Wnt-9b(33). Since nuclear "-catenin is observed in Paneth cells, it is possible that these Wnts operate in an autocrine fashion to induce canonical Wnt signaling or signal to nearby ISC or progenitors to drive their proliferation. However, such signals originating from Paneth cells or other epithelial cell types may be important but not sufficient for intestinal growth, since pure epithelial intestinal cultures appear to absolutely require exogenous growth factor supplementation (R-spondin1, EGF, Noggin, and Jagged) (15) which may reflect signals either provided in vivo by the mesenchyme, or present endogenously in explant cultures that contain mesenchymal elements(63) Intestinal Stem Cells as the Cell of Origin of Colorectal Cancer The self-renewing capabilities of stem cells may be relevant to cancer whereby tumors may initiate from a small subpopulation of cells, which possesses the bulk of 20
32 long-term proliferative activity. The examination of adenomas from XO/XY patients with FAP showed that 76% of adenomas were polyclonal(3). This was despite observations that monocryptal adenomas showed either the XO or XY genotype, and no mixed XO/XY genotype. One possible explanation is that XO/XY adenomas may actually be XY adenomas that have focally lost their Y chromosome. For example, it is known that the Y chromosome is often lost in a variety of carcinomas, although this usually occurs after P53 loss, or during late stages of adenoma development in the intestine. Another possible explanation is that random collisions occur between independently transformed neighboring crypts. However, it appears that neither of these explanations can account for the high proportion of polyclonality that is observed in these patients. Regardless, there is accumulating evidence that the earliest lesion in colorectal tumorigenesis occurs in the stem cells. Recently, Barker et al. showed that the deletion of APC in Lgr5 + cells leads to the transformation of epithelial cells within days(64). These transformed Lgr5 + cells rapidly generate microadenomas, which exhibit unimpeded growth, and develop into macroscopic adenomas within 3-5 weeks. In contrast, the deletion of APC in short-lived transit-amplifying cells results in stalled growth of the induced microadenomas. In these mice, even after 30 weeks, few large adenomas were observed. Furthermore, although a direct comparison was not performed, Barker et al. also reported that the conditional activation of a "-catenin allele (loxp(ex3)) in Bmi1 + cells also resulted in adenoma formation within 3-4 weeks after cre induction(64). In both cases, these observations support the hypothesis that the transformation of ISC constitutes the principal route towards intestinal cancer. 21
33 Moreover, other recent studies have also shown that cancer-initiating cells can be identified in colon carcinomas. By using renal capsule transplantation assays in NOD/SCID mice, O Brien et al. found that a fraction of the CD133 + cells were able to maintain themselves as well as differentiate and re-establish tumor heterogeneity upon serial transplantation(65). Independently, Ricci-Vitiani et al. also established that CD133 + colon cancer cells could be subcutaneously injected into immunodeficient mice where they readily reproduced the original tumor(66). Both studies demonstrated that CD133 - cells did not form tumors. Together, these findings suggest that cancers themselves are maintained by a population of cancer cells, and there is accumulating evidence that tumorigenesis is initiated in the ISCs. An increasing body of knowledge is accumulating regarding the biology of intestinal stem cells, aided by molecular isolation and lineage tracing techniques. It is now clear that single ISCs can indeed give rise to all of the differentiated lineages of the intestinal epithelium. Though the precise identity of the actual ISC is still debated, studies clearly demonstrate that the Lgr5 + CBC cells and the Bmi1 + cells at cell position + 4 both have the ability to self-renew, to undergo multi-lineage differentiation, and to initiate polyposis. However, many questions still remain unanswered, including the potential for distinct populations of ISCs, their functional interrelationships, the control of their numbers, and ultimate relevance to carcinogenesis and cancer stem cells/tumorinitiating cells. Similarly, the precise location and cellular composition of the niches in which ISCs reside remain to be elucidated, along with the relative contributions of mesenchymal versus epithelial niches. Regardless, better understanding of the physiological mechanisms that regulate stem cell maintenance potentially offers new 22
34 strategies to promote tissue regeneration after injury, to maintain stem cell activity during aging, or to sensitize cancer stem cells to therapy. Clearly, translation of these basic biological discoveries will have many implications in the fields of stem cell biology, regenerative medicine, cancer, aging, and therapeutics. 23
35 CHAPTER 2 THE INTESTINAL STEM CELL MARKERS BMI1 AND LGR5 IDENTIFY TWO FUNCTIONALLY DISTINCT POPULATIONS 24
36 2.1 ABSTRACT The small intestine epithelium undergoes rapid and continuous regeneration supported by crypt intestinal stem cells (ISCs). Bmi1 and Lgr5 have been independently identified to mark long-lived multipotent ISCs by lineage tracing in mice, however the functional distinctions between these two populations remain undefined. Here, we demonstrate that Bmi1 and Lgr5 mark two functionally distinct ISCs in vivo. Lgr5 marks mitotically active ISCs that exhibit exquisite sensitivity to canonical Wnt modulation, contribute robustly to homeostatic regeneration, and are quantitatively ablated by irradiation. In contrast, Bmi1 marks quiescent ISCs that are insensitive to Wnt perturbations, contribute weakly to homeostatic regeneration, and are resistant to high dose radiation injury. Post-irradiation, however, the normally quiescent Bmi1 + ISCs dramatically proliferate to clonally repopulate multiple contiguous crypts and villi. Clonogenic culture of isolated single Bmi1 + ISCs yields long-lived self-renewing spheroids of intestinal epithelium that produce Lgr5-expressing cells, thereby establishing a lineage relationship between these two populations in vitro. Taken together, these data provide direct evidence that Bmi1 marks quiescent, injury-inducible reserve ISCs that exhibit striking functional distinctions from Lgr5 + ISCs, and support a model whereby distinct ISC populations facilitate homeostatic versus injury-induced regeneration. 25
37 2.2 INTRODUCTION The G protein-coupled receptor Lgr5 and the Polycomb group protein Bmi1 are two recently described molecular markers of self-renewing and multipotent adult stem cell populations residing in the crypt of the small intestine, capable of supporting regeneration of the intestinal epithelium(14, 16). Despite their similar ability to functionally repopulate the intestinal epithelium as demonstrated by independent in vivo lineage tracing experiments in reporter mice, the ISCs identified by these two molecular markers are spatially distinct. Whereas Lgr5 + ISCs are crypt base columnar (CBC) cells (8, 14) interspersed between Paneth cells and expressed throughout the intestine, Bmi1 + ISCs are mostly restricted to the +4 cell position abutting the uppermost Paneth cell in proximal small intestine crypts(16). Lgr5 + ISCs are actively cycling(14), equipotent and contribute to intestinal homeostasis by neutral drift competition(67-69). By comparison, Bmi1 + ISCs are less well characterized, and due to lack of direct evidence, their cell cycle status is variably ascribed to be rapidly(31) versus slowly cycling(70). It has been suggested that Bmi1 and Lgr5 mark an overlapping and possibly identical or redundant population of ISCs(31, 68, 71), however no direct exploration of their functional similarities and differences has been performed. Further, it is unknown how Bmi1 + and Lgr5 + ISCs relate to a proposed model in which the intestine differentially uses an actively cycling ISC population during homeostasis and a distinct quiescent, injuryinduced ISC population (17, 72) during epithelial repair. We therefore conducted a systematic comparison of Bmi1 + and Lgr5 + ISC function during homeostasis and injuryrepair to investigate whether Lgr5 and Bmi1 mark identical, similar or distinct ISC populations. 26
38 2.3 RESULTS Bmi1 marks quiescent ISCs that contribute minimally to intestinal homeostasis. Given the spatial localization of Bmi1 + ISCs at the + 4 position, where a DNA label-retaining cell has also been described (13, 73), we postulated that Bmi1 marks a quiescent ISC. Lgr5-eGFP-IRES-CreERT2 and Bmi1-CreER; Rosa26-YFP mice were used to compare the basal proliferation status of Lgr5 + versus Bmi1 + ISCs during homeostasis. We used short-term tamoxifen exposure, for induction of Cre-mediated recombination, to selectively mark Bmi1 + ISCs in vivo. Accordingly, Bmi1-CreER; Rosa26-YFP mice were treated with tamoxifen 1 to 2 d prior to sacrifice to genetically label Bmi1 + cells with YFP, revealing one to two YFP + cells at approximately the + 4 cell position (ranging from + 1 to + 6) within ~10% of proximal small intestine crypts, in agreement with previous reports(16). To determine basal proliferation status, labeling of actively cycling S phase cells was performed using the thymidine analog EdU (5- ethynyl-2 -deoxyuridine). Under steady state conditions, histological examination of small intestine revealed 31 ± 5.2% EdU incorporation among Lgr5 + ISCs, identified as GFP + CBC cells in Lgr5-eGFP-IRES-CreERT2 mice. In contrast, only 1.7 ± 0.30 % of Bmi1 + ISCs, identified by the crypt Rosa-YFP + signal following 1.5 d tamoxifen exposure in Bmi1-CreER; Rosa26-YFP mice, incorporated EdU (Fig. 1 A-F, N). To examine the relative contribution of Lgr5 versus Bmi1 ISCs to tissue regeneration under steady-state conditions, lineage tracing was induced by tamoxifen administration in Cre reporter mice to mark the ISCs and their respective progeny. Upon tamoxifen-mediated lineage tracing of Lgr5 + and Bmi1 + ISCs in Lgr5-eGFP-IRES- CreERT2; Rosa26-TdTomato and Bmi1-CreER; Rosa26-YFP mice, Lgr5 + ISCs were 27
39 markedly more efficient at generating progeny than Bmi1 + ISCs by 7 d of lineage tracing with 95 ± 1.7% vs. 18 ± 5.1% lineage stripe generation, respectively (Fig. 1 G-I). This method also likely underestimates their substantial relative difference in progeny generation due to the more qualitatively vigorous nature of Lgr5 striping. Overall, these differences in basal proliferation and lineage-forming efficiency reflect a much greater functional contribution of Lgr5 + ISCs to homeostatic small intestine regeneration compared to Bmi1 + ISCs. Differential responses of Bmi1 + versus Lgr5 + ISCs to canonical Wnt modulation. Since Lgr5 + and Bmi1 + ISCs reside in spatially distinct crypt locations, we explored whether they exhibited differential responses to global modulation of the canonical Wnt pathway, which is required to maintain adult intestine epithelial proliferation and crypt architecture (35, 37, 38). Gain- and loss-of-function manipulation of the canonical Wnt signaling pathway was achieved in mice using adenoviral expression of the soluble, secreted factors R-Spondin1 (Rspo1)(63, 74) and Dickkopf-1 (Dkk1)(35), respectively. A single intravenous injection of adenovirus encoding either the Wnt agonist Rspo1 or antagonist Dkk1 results in hepatic infection and transduction, secretion of the recombinant factor into the systemic circulation, and leads to profound histological changes in the intestinal epithelium within 5 d postinfection(35). In Lgr5-eGFP-IRES-CreERT2 and Bmi1-CreER; Rosa26-YFP mice, canonical Wnt signaling was potently induced by systemic administration of an adenovirus encoding Rspo1 fused to an IgG2! Fc fragment (Ad RSpo1-Fc), causing marked crypt hypertrophy and hyperproliferation. By 5 d post-infection, Ad RSpo1-Fc markedly expanded Lgr5-eGFP + cells, as well as expression of the surrogate marker 28
40 Olfm4 (31), which was not seen with a control adenovirus encoding IgG2! Fc (Ad Fc) (Fig. 1 J, K, U, V and Fig. S1 A, B). Electron microscopy of Ad Rspo1-Fc-treated small intestine confirmed expansion of multiple consecutive slender CBC cells between Paneth cells, consistent with substantially increased numbers of Lgr5 + ISCs, compared to only single CBC cells between Paneth cells with Ad Fc treatment (Fig. 1 O, P). In contrast, Ad RSpo1-Fc treatment did not significantly alter either the relative abundance or the mitotic index of Bmi1 + ISCs labeled with 1 or 2 d tamoxifen exposure in Bmi1-CreER; Rosa26-YFP mice (Fig. 1 L-N and Fig. S1 D, E). Further, Ad RSpo1-Fc did not enhance the basal level of infrequent lineage stripes arising from Bmi1 + ISCs despite dramatic concurrent expansion of the crypt compartment (Fig. S1 G-I) and Lgr5-eGFP + cells (Fig 1 K). Conversely, systemic Wnt loss-of-function studies were performed in these reporter mice using adenovirus encoding Dkk1 (Ad Dkk1), which has been reported to induce rapid crypt loss and destruction of the small intestine epithelial architecture(35) (Fig. 1 Q-T). Correspondingly, Ad Dkk1 induced a profound loss of Lgr5-eGFP and Olfm4 expression in the small intestine crypts (Fig. 1 Q, R, U, W and Fig. S1 A, C). In contrast to the dramatic effect on Lgr5 + ISCs, Ad Dkk1 treatment did not significantly diminish 1 or 2 d tamoxifen-labeled Bmi1 + ISCs in Bmi1-CreER; Rosa26-YFP mice, which in fact persisted despite Ad Dkk1-mediated crypt loss (Fig. 1 S, T and Fig. S1 D, F). Thus, the Lgr5-eGFP + but not the Bmi1 + ISC population exhibited exquisite sensitivity to global gain- and loss-of-function Wnt signaling modulation mediated by RSpo1 and Dkk1, respectively, highlighting substantial functional differences between the response of these two ISC populations to extracellular Wnt signals. 29
41 Differential responses of Bmi1 + versus Lgr5 + ISCs to radiation injury. We further probed the functional differences between Lgr5 + and Bmi1 + ISCs using a radiation injury model. Lgr5-eGFP-IRES-CreERT2 and Bmi1-CreER; Rosa26- YFP mice were treated with 12 Gy whole body irradiation. By 2 d post-irradiation, Lgr5-eGFP + ISCs as well as Olfm4 expression were completely lost from small intestine crypts (Fig. 2 A, C, I, J, K, L), while there were no discernible quantitative effects on Bmi1-YFP + ISCs labeled with 1 d tamoxifen treatment (Fig. 2 B, D). By 4.5 and 7 d post-irradiation, rare Lgr5-eGFP + cells re-emerged, scattered sporadically throughout the small intestine at a frequency of ~1/180 total crypts, but these were still severely diminished compared to unirradiated littermate controls (Fig. 2 E, G). In contrast, irradiation induced a strong proliferative response in 1 d tamoxifen-treated Bmi1 + ISCs, 17 ± 1.5% of which were robustly labeled with EdU by 2 d post-irradiation, compared to 1.7 ± 0.30% during homeostasis (Fig. 2 O-R); this was accompanied by a 5-fold expansion in Bmi1-YFP + ISCs/progeny upon fluorescence activated cell sorting (FACS) analysis by 4.5 d post-irradiation versus unirradiated littermate controls (Fig. 2 M, N). We also examined the functional effects of irradiation on the ability of Lgr5 + versus Bmi1 + ISCs to generate downstream progeny. Two serial tamoxifen injections in Lgr5-eGFP-IRES-CreERT2; Rosa26-YFP mice, 1 d before and 1 d after irradiation were used to irreversibly mark the Lgr5 + lineage with YFP, in a manner independent of concurrent Lgr5 expression. Accordingly, both YFP-marked Lgr5 + cells and their downstream progeny were eradicated by 4.5 and 7 d post-irradiation (Fig. 3 A, B). Similarly, a single tamoxifen injection in Bmi1-CreER; Rosa26-YFP mice was used to irreversibly mark the Bmi1 + lineage, followed 2 d later by 12 Gy irradiation and tissue 30
42 harvest at 7 d post-irradiation. As opposed to the quantitative eradication of Lgr5 + ISCderived progeny, irradiation substantially induced expansion of the Bmi1 + lineage. Indeed, by 7 d post-irradiation in regenerating small intestine, confluent Bmi1 + ISCderived YFP + lineage stripes were seen along multiple adjacent crypts and villi, which were much more extensive than the comparatively atretic Bmi1 + lineage tracing present during homeostasis (Fig. 3 C, D and Fig. S2 A-F). Strikingly, the Bmi1 + lineage showed post-irradiation extension into multiple adjacent crypts and villi emanating from a single crypt as revealed by 3-dimensional (3D) confocal reconstruction (Fig. 3 E-G and Supporting Information Video 1). We also treated Bmi1-CreER; Rosa26-Confetti mice with tamoxifen 2 d prior to 12 Gy irradiation to stochastically label individual Bmi1 + ISCs with one of four possible fluorescent colors (67) and trace their fate in response to injury. Using this multicolor reporter to visualize the dramatic expansion of the Bmi1 + lineage, the progeny arising from the marked clones were noted to be exclusively labeled with a single color at 7 d post-irradiation, attesting to their monoclonal origin despite their extension into contiguous crypts and villi (Fig. 3 H-K and Supporting Information Video 2). Thus, compared to the radiosensitive, actively cycling Lgr5-eGFP + ISCs, the quiescent Bmi1 + ISCs exhibit radioresistance and are rapidly mobilized to proliferate upon injury with significant contribution to epithelial regeneration, and pronounced induction of Bmi1 + lineage tracing. Taken together, these data suggest that Bmi1 + ISCs are quiescent at baseline but actively contribute to injuryassociated repair upon quantitative loss of Lgr5 + population or crypt injury, and suggest that Bmi1 + ISCs play a larger role during epithelial repair than during basal homeostasis. 31
43 Isolated Bmi1 + ISCs are multipotent and give rise to Lgr5-expressing cells in vitro. Single Lgr5-eGFP + ISCs can generate in vitro spheroids in clonogenic culture without requiring a mesenchymal niche (15, 75). To determine whether Bmi1 + ISCs can also form in vitro spheroids, we FACS-isolated single YFP + small intestine epithelial cells, representing Bmi1 + ISCs, from 1 or 2 d tamoxifen-treated Bmi1-CreER; Rosa26- YFP mice. These purified single Bmi1-YFP + cells generated spheroids with similar morphology to Lgr5-eGFP-derived spheroids upon clonogenic culture in Matrigel with previously reported exogenous factors including Epidermal Growth Factor, Noggin, Jagged and RSpo1(15) (Fig. 4 A-D, I and Fig. S3). Consistent with their in vivo stem cell function, the clonogenic spheroids grown from isolated Bmi1-YFP + single cells exhibited multipotency (Fig. 4 E-H), continued proliferation (Fig. 4 J) and maintenance of pan-yfp expression upon serial passage (>8 months with weekly passages) (Fig. S3). Notably, numerous Lgr5 + cells were detected by Lgr5 mrna fluorescence in situ hybridization (FISH) within the Bmi1 + clonally-derived spheroids (Fig. 4 K and Fig. S4), whose clonogenicity was confirmed by the genetic signature of pan-yfp expression seen by both intrinsic YFP fluorescence and immunodetection (Fig. 4 D, K), indicating that the Bmi1 + ISC lineage can generate Lgr5 + cells in vitro. 2.4 DISCUSSION Our findings reveal that under both homeostatic and injury-induced conditions, Bmi1 and Lgr5 mark functionally distinct ISC populations in vivo. While Lgr5 + ISCs are extremely sensitive to RSpo1-mediated Wnt stimulation and Dkk1-mediated Wnt inhibition, Bmi1 + ISCs are relatively refractory to Wnt manipulation. Further, while Lgr5 + ISCs are actively cycling and quantitatively ablated by irradiation injury, the 32
44 normally quiescent Bmi1 + ISCs are instead induced to proliferate upon irradiation, and in fact give rise to progeny that clonally repopulate multiple contiguous crypt-villus axes during subsequent intestinal regeneration. Our results thus provide direct evidence that Bmi1 + ISCs represent a quiescent, injury-inducible reserve ISC population, consistent with a proposed model for co-existence of distinct ISCs active during homeostasis versus regeneration (17, 30, 72). During preparation of this manuscript, Tian and colleagues reported an elegant diphtheria toxin receptor (dtr) knock-in genetic strategy to selectively ablate Lgr5 + ISCs in vivo using diphtheria toxin, revealing that Lgr5 + ISCs are dispensable for intestinal homeostasis(71). Lgr5 + ISC ablation was accompanied by expansion of the Bmi1 + lineage, which is capable of giving rise to Lgr5-expressing cells in vivo(71). Their findings parallel and support our overall conclusions that the Bmi1 + lineage expands upon quantitative loss of the Lgr5 + population and of their lineage inter-relationship. Notably, dtr-mediated genetic ablation of Lgr5 + ISCs differs from our radiation injury model due to lack of crypt loss observed upon diphtheria toxin ablation. Moreover, the mediation of epithelial reconstitution by Bmi1 + ISCs following Lgr5 + ISC ablation by either dtr or radiation injury does not distinguish between models in which these two populations are either functionally redundant or alternatively possess distinct functions. Our data, which reveal profound differences between Bmi1 + and Lgr5 + ISCs in baseline quiescence, cell-cycle entry post-injury, effects of Wnt gain- and loss-of-function, and radiosensitivity, strongly argue for the latter model. The functional differences we describe therefore resolve the fundamental question of whether Bmi1 + and Lgr5 + ISCs are redundant or distinct populations, and indicate that Bmi1 + ISC recruitment post-injury 33
45 marks the utilization of a functionally discrete ISC class. Finally, our findings of Bmi1 + ISC baseline quiescence and inducible proliferation following crypt injury provide the first functional evidence for Bmi1 + ISC as a postulated quiescent, injury-mobilized population, and further underscore the heterogeneity of ISC populations contributing to tissue regeneration. It is certainly possible that Bmi1 may only mark a subset of quiescent stem cells, and our results do not exclude overlapping expression with populations identified by other putative ISC molecular markers(26-28, 46, 70, 76-81), including those that may also function as quiescent and injury-mobilized ISCs. Additional proliferating cells not marked by Bmi1 are present in 2 d post-irradiated crypts using our tamoxifenlabeling strategy, suggesting either variegated Bmi1 expression in our reporter system or the contribution of other ISCs populations to regeneration that perhaps also mediate postinjury regional repair of the colon and distal small intestine. The relative scarcity of Bmi1 + ISCs may be insufficient to repair the entire intestinal epithelium after irradiation, and Bmi1 + ISC are not present in colon(16, 71). Other ISCs markers have been proposed for identification of + 4 position quiescent cells including DCAMKL1(28), mtert(70) and Hopx(82), and these cells themselves may exhibit heterogeneity as there are numerous cells occupying this crypt position within the annulus of the 3D crypt. Notably, mtert has been described to mark an ISC population at the + 4 position mobilized after radiation injury, and which overlaps in expression with both Lgr5 and Bmi1(70, 83). Certainly, the potential overlap or inter-relatedness of Bmi1 + and mtert + ISCs and other + 4 position markers such as DCAMKL1 and Hopx warrants further 34
46 investigation. Further, the significance of overlapping Lgr5 co-expression within Bmi1 + ISCs(71) remains to be determined. Clonogenic cultures derived from isolated single Bmi1-YFP + cells give rise to all differentiated intestinal lineages and Lgr5 + cells, supporting a lineage relationship where a quiescent ISC can give rise to an actively cycling ISC, and parallels in vivo observations of Lgr5 + cell generation from Bmi1 + ISCs(71). This is the first demonstration of clonogenic culture of the Bmi1 + ISC population. Intriguingly, upon removal from the native tissue microenvironment and FACS isolation, the normally quiescent Bmi1 + ISCs generate clonally-derived intestinal spheroids with similar kinetics, morphology and histology to those derived from single Lgr5 + ISCs(15). Interestingly, the self-renewal and proliferation of the Bmi1-derived spheroids, like those derived from Lgr5, is Rspo1-dependent, consistent with prior results with Bmi1 + lineage tracing in airliquid interface organotypic cultures(63), whereas Bmi1 + ISCs are relatively insensitive to RSpo1-Fc in vivo. These results are potentially consistent with a model where Bmi1 + ISC are subject to considerable in vivo repression within the ISC niche, which does not appear to be recapitulated by current in vitro culture systems. Further, the clonogenic culture conditions employed here, which were initially reported for Lgr5 + ISCs(15), may actually select for an actively cycling state. Despite their functional differences in vivo, we cannot completely exclude potential concomitant overlapping Lgr5 co-expression within Bmi1 + ISCs. It also remains to be determined whether the observed differences between the in vivo and in vitro properties are cell autonomous or due to differences in stem cell niche interactions. 35
47 Isolated Bmi1 + ISCs can give rise to Lgr5 + cells in vitro, and in vivo under homeostasis or dtr-mediated Lgr5 + cell ablation(71), though the frequency of this occurrence is unknown. This lineage could occur infrequently during homeostasis in vivo, given the relative paucity of Bmi1 + lineage contribution to basal regeneration. This lineage relationship could possibly also be bi-directional with Lgr5 + ISCs giving rise to Bmi1 + ISCs, paralleling the Lgr5/Hopx bi-directional relationship(82), with superimposed regulatory mechanisms to control the total number of ISCs, to regulate the balance of active and quiescent ISCs within the total stem cell pool, and to restrain homeostatic Bmi1 + ISC proliferation. Overall, our findings of multiple functional distinctions between Bmi1 + and Lgr5 + ISCs provide direct evidence to support a proposed model of separate but cooperative functional roles of multiple and distinct ISC populations (17, 30, 72) residing in adjacent niches that contribute to homeostatic versus injury-induced regeneration, with Bmi1 + ISCs representing a quiescent, injuryinducible reserve ISC population. Further, the demonstration of Bmi1 + ISCs giving rise to Lgr5-expressing cells underscores the potentially complex interplay between these two populations. 36
48 2.5 METHODS Mice. Lgr5-eGFP-IRES-CreERT2 mice were crossed with Rosa26-TdTomato or Rosa26- YFP (JAX) mice to generate Lgr5-eGFP-IRES-CreERT2; Rosa26-TdTomato or Lgr5- egfp-ires-creert2; Rosa26-YFP compound heterozygous mice, respectively. Likewise, Bmi1-CreER mice were crossed to Rosa26-YFP or Rosa26-Confetti mice to generate Bmi1-CreER; Rosa26-YFP or Bmi1-CreER; Rosa26-Confetti compound heterozygous mice, respectively. Adult mice between 4-12 weeks of age were administered IP tamoxifen (9 mg per 40 g body weight) to label Bmi1 ISCs, and to label the Bmi1 + and Lgr5 + lineages using the various Rosa26 Cre reporter strains. Proliferation Assay and Cell Quantitation. Bmi1 + ISCs were labeled in Bmi1-CreER; Rosa26-YFP double heterozygous mice with tamoxifen (IP) 24 hrs prior to sacrifice. Both Lgr5-eGFP-IRES-CreERT2 mice and tamoxifen treated Bmi1-CreER; Rosa26-YFP mice were administered a single 1 mg injection (IP) of the thymidine analogue EdU 4 hrs prior to sacrifice. EdU incorporation was detected according to the manufacturer s instructions with the Click-iT EdU Imaging Kit (Invitrogen) on OCT (Tissue Tek) frozen sections. All intestinal tissue was prepared as OCT frozen sections. Briefly, mouse intestine was fixed in 4% paraformaldehyde for 2-4 hrs, equilibrated into 30% sucrose then embedded in OCT. Cryosections were cut at 8!m. Proliferation indices are expressed as percentage GFP + or YFP + epithelial cells that incorporated EdU over total number of GFP + or YFP + epithelials cells within proximal small intestine crypts. At least total ISCs per mouse were counted from multiple fields. Statistical analysis was performed using n=3 mice for each condition. The mean, standard errors are 37
49 represented. Two-tailed p values were calculated using unpaired Student s t-test. Lineage generation efficiency was quantified by calculating the percentage of GFP + or YFP + crypts or villi with 3 or more cells arising from Lgr5 + or Bmi1 + ISCs following 7 d tamoxifen treatment in Lgr5-eGFP-IRES-CreERT2; Rosa26-TdTomato or Bmi1-CreER; Rosa26-YFP mice, respectively. At least 100 total crypts and villi were counted from multiple fields. Statistical analysis was performed using n=3 mice for each condition. The mean, standard error are represented. Two-tailed p values were calculated using unpaired Student s t-test. Wnt Modulation. Modulation of canonical Wnt signaling was achieved by 5x10 8 p.f.u. injection (IV) of either Ad Fc, Ad Rspo1-Fc or Ad Dkk1. Systemic infection was confirmed by histologic evaluation of the small intestine and immunoblot of the mouse sera for the presence of HA-epitope tagged soluble Rspo1-Fc and Dkk1 proteins. Mice were sacrificed 5 d post-infection. The small intestine was resected en bloc for histologic analysis as above. Radiation Injury. Mice were administered 12 Gy of whole body irradiation and the tissue was harvested at multiple time points following irradiation and tamoxifen administration to label ISCs/ their progeny. n=3 mice or more were used for each condition. Tissue was fixed with 4% PFA as above and processed in OCT for frozen sections, paraffin or prepared as whole mount. 38
50 Flow Cytometry. FACS experiments were performed using fresh small intestine epithelial preparations. A standardized 3 cm segment of proximal jejunum was used for quantitative FACS analysis of ISC populations. Intestinal epithelial cells were extracted from en bloc resected small intestine with 10 mm EDTA and manual shaking, followed by enzymatic dissociation with collagenase/dispase (Roche) to generate a single cell suspension(84). Singlet discrimination was sequentially performed using plots for FSC (FSC-A vs. FSC-H) and SSC (SSC-W vs. SSC-H). Dead cells were excluded by scatter characteristics and propidium iodide. Epithelial cells were identified by CD45 - and EpCAM + immunostaining. ISCs were identified by their endogenous GFP or YFP expression. All FACS experiments were performed on an Aria II sorter (BD) at the Stanford University Shared FACS Facility and FACS data were analyzed using FlowJo software (Treestar). Clonogenic Culture. For Bmi1-derived clonogenic cultures, Bmi1-CreER; Rosa26-YFP mice were treated with tamoxifen as described above. Tissue was dissociated for FACS isolation as above. Singlet discrimination was sequentially performed using plots for FSC (FSC-A vs. FSC-H) and SSC (SSC-W vs. SSC-H). Dead cells were excluded by scatter characteristics. Antibody staining was performed with anti-cd45, anti-cd31 and anti-epcam (ebiosciences). CD31 - CD45 - EpCAM + YFP + cells were isolated by FACS. These isolated cells were cultured and passaged once per week as previously described(15, 75). Lgr5-eGFP + ISCs were also isolated from Lgr5-eGFP-IRES- CreERT2 mice using the FACS gating scheme and culture methods previously reported by Sato et al(15, 75). 39
51 Immunofluorescence. Paraffin-embedded sections were stained using the following primary antibodies: anti-muc2 (Santa Cruz), lysozyme (Dako), chromogranin A (Dako), E-cadherin (BD Biosciences) and sucrase-isomaltase (generated in house at Stanford University). All primary antibodies were used at 1:500. Cy3 and Cy5 conjugated secondary antibodies (Santa Cruz) were used at 1:2000. Alexa Fluor 594- conjugated Phalloidin (Invitrogen) was used at 1:500. Images were captured on a Zeiss Axio-Imager Z1 with ApoTome attachment or Leica CTR 6500 confocal microscope. Lgr5 mrna in situ hybridization. In situ hybridization was performed on 4% paraformaldehyde-fixed Bmi1-derived clonogenic intestinal spheroids using 6!m paraffin-embedded sections. For FISH, Lgr5 digoxigenin-labeled probes were synthesized as previously described(63). Sense strand probes were generated and served as negative controls. Simultaneous anti-yfp immunodetection was performed using anti-gfp primary antibody (Aves) at 1:500, a FITC-conjugated secondary antibody (Aves) at 1:2000 followed by further amplification using the TSA-Biotin system (Perkin Elmer). Olfm4 mrna in situ hybridization by RNAscope. In situ hybridization for Olfm4 RNA was performed using the RNAscope kit (Advanced Cell Diagnostics, Inc., Hayward, CA) according to the manufacturer s instructions. Briefly, 5!m formalin fixed, paraffin embedded tissue sections or 8!m OCT frozen sections were pretreated with heat and protease prior to hybridization with a target probe to Olfm4 mrna. An HRP-based 40
52 signal amplification system was then hybridized to the target probes followed by colorimetric development with DAB. Positive staining was identified as brown, punctate dots present in the nucleus and/or cytoplasm. Negative control probes for the bacterial gene DapB were also included for each slide. 3D Confocal Reconstruction. Small intestine tissue samples were processed for immunofluorescence microscopy as previously described(85). Briefly, samples were fixed with 2% paraformaldehyde for 1 hr and permeabilized in phosphate buffered saline with 1% Triton X-100, 1% saponin and 3% BSA. Bmi1-CreER; Rosa26-YFP samples were stained with DAPI and Phalloidin-Alexa Fluor 594 (Molecular Probes) and antilysozyme (Dako) for visualization of the nuclei, actin cytoskeleton and Paneth cells, respectively. Bmi1-CreER; Rosa26-Confetti samples were not counter stained. For whole mount visualization of intestine, samples were mounted as intact tissue blocks in Vectashield mounting medium (Vector Laboratories) and imaged with a Zeiss LSM 700 or Leica CTR 6500 confocal microscope. Z-stacks were reconstructed into 3D using Volocity (Improvision). Electron Microscopy. Small intestine tissue samples were fixed with 2.5% glutaraldehyde and post-fixed in 1% osmium tetroxide in 100mM phosphate buffer. Tissue was dehydrated, embedded in epoxy resin, and visualized with a JEOL transmission electron microscope at 120kV (model JEM-1210). 41

53 2.6 FIGURES Figure 1 42

54 Figure 2 43

55 Figure 3 44

56 Figure 4 45
57 2.7 FIGURE LEGENDS Figure 1. Basal proliferation status and response to canonical Wnt pathway modulation of Lgr5 + versus Bmi1 + ISCs. (A-F, N) Lgr5-eGFP + ISCs in Lgr5-eGFP- IRES-CreERT2 duodenum are actively cycling and incorporate EdU under homeostasis (A-C). In contrast, Bmi1 + ISCs labeled with 1.5 d tamoxifen exposure in Bmi1-CreER; Rosa26-YFP duodenum are slowly cycling and do not incorporate EdU (D-F, **p = versus Lgr5; DAPI colored in blue). (G-I) Lgr5 + ISCs in Lgr5-eGFP-IRES- CreERT2; Rosa26-TdTomato duodenum generate progeny much more efficiently than Bmi1 + ISCs in Bmi1-CreER; Rosa26-YFP duodenum by 7 d following tamoxifenmediated lineage tracing (*p = , bar = 50!m). TdTomato is shown in red (G). Phalloidin is colored in red and DAPI in blue (H). (J-N) Consequences of canonical Wnt stimulation with Ad Rspo1-Fc on Lgr5 + versus Bmi1 + ISCs. Marked expansion of Lgr5-eGFP + ISCs and the transit-amplifying compartment are seen by direct fluorescence microscopy whereas Bmi1-YFP + ISCs are less responsive to Ad Rspo1-Fc stimulation. Notably, Ad Rspo1-Fc results in enhanced EdU incorporation in Lgr5-eGFP + ISCs (*p = 0.04) but not in Bmi1-YFP + ISCs (p = 0.13) compared to Ad Fc control. (J-M, bar = 20!m). (O, P) Electron microscopy reveals expansion of crypt base columnar (CBC) cells (*) consistent with Lgr5 + ISC morphology in between Paneth cells (Pa) following Ad Rspo1-Fc treatment. Bar = 5!m. (Q, R) Canonical Wnt inhibition with Ad Dkk1 results in complete loss of Lgr5-eGFP + ISCs. (S, T) In contrast, Bmi1-YFP + cells persist despite systemic Wnt blockade with Ad Dkk1. (Q-T, Phalloidin is colored in red and DAPI in blue). (U-W) In situ hybridization (ISH) analysis reveals enhanced expression of Olfm4, a surrogate marker for Lgr5, in response to Ad Rspo1-Fc 46
58 stimulation of canonical Wnt signaling (V), whereas Ad Dkk1 results in complete loss of Olfm4 expression (W) compared to Ad Fc control (U). (Q-W, bar = 50!m). Figure 2. Differential responses of Lgr5 + versus Bmi1 + ISCs to acute radiation injury. (A-H) Direct fluorescence detection of Lgr5-eGFP + versus Bmi1-YFP + ISCs. Phalloidin is depicted in red and DAPI in blue. (A, B) Unirradiated duodenum with Lgr5-eGFP + cells in the crypt bases and rare Bmi1-YFP + ISCs labeled with 1 d tamoxifen exposure in Bmi1-CreER; Rosa-YFP mice. (C-F) Lgr5-eGFP + ISCs are completely lost (C, E) whereas Bmi1-YFP + ISCs (D, F) persist by 2 and 4.5 d post-12 Gy whole body irradiation. (G, H) Lgr5-eGFP + ISCs return in sporadic (~1/180) crypt bases (G) and Bmi-YFP + ISCs are quantitatively unchanged by 7 d post-12 Gy whole body irradiation (H). (I, J) Olfm4 ISH demonstrates loss of Olfm4 expression in crypt bases by 2 d post-12 Gy irradiation confirming loss of Lgr5 expression. (K-N) FACS analysis demonstrates loss of Lgr5-eGFP + epithelial cells by 2 d (K, L), in contrast to expansion of Bmi1-YFP + cells/progeny by 4.5 d following 12 Gy whole body irradiation (M, N). (O-R) Enhanced proliferation of Bmi1-YFP + ISCs by 2 d post-12 Gy irradiation revealed by EdU incorporation (*p = , n = 3 mice, unpaired Student s t-test). Bar = 50!m. Figure 3. injury. Differential responses of Lgr5 + versus Bmi1 + lineages to acute radiation (A, B) Tamoxifen-mediated lineage trace of Lgr5-eGFP + progeny in Lgr5- egfp-ires-creert2; Rosa-YFP mouse duodenum demonstrates prolific lineage generation under homeostasis by 8 d (A). Lgr5-eGFP + ISCs and their lineage are 47
59 ablated at 7 d following 12 Gy whole body irradiation (B). (C, D) Bmi1 + lineage in Bmi1-CreER; Rosa-YFP duodenum marked by 9 d tamoxifen treatment exhibits marked proliferative regenerative response with progeny observed in adjacent crypts and villi at 7 d post-irradiation. (A-D, bar = 100!m; Phalloidin is shown in red and DAPI in blue). (E-G) 3D confocal reconstruction of Bmi1 + lineage regenerative response in the jejunum at 7 d post-12 Gy irradiation highlights confluent patches of Bmi1 + -derived cells in multiple adjacent crypts/villi. Expanded view from regenerated jejunum immunostained with lysozyme, illustrating Paneth cells within a budding crypt (F). Phalloidin is colored in red and DAPI in blue. (H-K) Monoclonal repopulation of 7 d post-irradiated jejunum from Bmi1-CreER; Rosa-Confetti compound heterozygotes, as suggested by monochromatic patches of regenerated crypts/villi illustrated in RFP (H), GFP (I), CFP (J) and YFP (K). Insets represent serial cross-sections through lineage traces indicating involvement of multiple contiguous crypts and villi (G, K). Figure 4. Clonogenic culture of single FACS-isolated Bmi1-YFP + ISCs. (A-D) Single sorted Bmi1-YFP + cells marked by 1 or 2 d tamoxifen administration in Bmi1- CreER; Rosa-YFP mice form intestinal epithelial spheroids in clonogenic culture that demonstrate continued proliferation and self-renewal. Brightfield images of spheroids by 3 d (A), 10 d (B), and 14 d (C) in culture with pan-yfp expression visualized by direct fluorescence (D), consistent with clonal-derivation from a genetically marked Bmi1-expressing cell. Bar = 100!m. (E-H) Bmi1-YFP + ISCs are multipotent in vitro by immunofluorescence antibody detection of lineage markers within clonogenic spheroids including sucrase-isomaltase for enterocytes (E), Muc2 for goblet cells (F), 48
60 lysozyme for Paneth cells (G), and chromogranin A for enteroendocrine cells (H). E- cadherin is colored green and DAPI blue. Bar = 50!m. (I) H&E staining of Bmi1- YFP + spheroid. Bar = 50!m. (J) proliferation of Bmi1-YFP + spheroid by EdU incorporation. E-cadherin is colored green and DAPI blue. Bar = 25!m. (K) Clonogenically-cultured Bmi1-YFP + spheroid from FACS-isolated single cells gives rise to multiple Lgr5 + cells in vitro by Lgr5 FISH. Simultaneous Lgr5 FISH was performed with immunodetection of YFP, demonstrating that Lgr5 + cells are clonally-derived from a cell genetically marked with YFP expression in a Bmi1-CreER; Rosa-YFP mouse. DAPI is shown in blue. Bar = 50!m. 49


61 2.8 SUPPLEMENTAL FIGURES Figure S1. Ad Fc Ad Rspo1-Fc Ad Dkk1 A B C 1.5% 3.6% 0.030% Bmi1-CreER; Rosa-YFP Lgr5-eGFP- IRES-CreER FSC GFP FSC D E F 0.24% 0.24% 0.26% YFP Ad Rspo1-Fc G H I YFP Phalloidin Bmi1-CreER; Rosa-YFP Tamoxifen 7 d; Ad Rspo1-Fc 5 d 50
62 Figure S2. 7 d post-12 Gy A Bmi1 Lineage B C Bmi1-CreER; Rosa-YFP D E F Duodenum and Jejunum - Tamoxifen 9 d 51
63 Figure S3. Brightfield YFP Merged Bmi1 2 d TAM Lgr5-GFP Hi Bmi1 1 d TAM Brightfield GFP Merged oids. Representative spheroids derived from F 52
64 Figure S4. Lgr5 YFP Lgr5 YFP Lgr5 YFP Lgr5 YFP Lgr5 sense Lgr5 anti-sense Lgr5 FISH 53
65 Figure S5. 54
66 2.9 SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1. Quantitative effects of canonical Wnt pathway modulation on Lgr5 + versus Bmi1 + ISCs. (A-C) FACS analysis of Lgr5-eGFP + ISCs in response to 5 d adenovirus treatment with Ad Fc, Ad Rspo1-Fc or Ad Dkk1. Ad Rspo1-Fc expands whereas Ad Dkk1 ablates the Lgr5-eGFP + ISC population by FACS. (D-F) In contrast, Bmi1 + ISCs labeled with 1.5 d tamoxifen treatment in Bmi1-CreER; Rosa26- YFP mice at 5 d post-adenovirus injection, are relatively insensitive to Wnt modulation with Ad Rspo1-Fc and Ad Dkk1 versus Ad Fc. The proportion of Bmi1-YFP + ISCs by FACS does not change relative to total number of CD45 - and EpCAM + intestinal epithelial cells with Wnt gain- and loss-of-function. (G-I) Effects of Ad Rspo1-Fc treatment on Bmi1 + lineage in vivo. Bmi1-CreER; Rosa26-YFP mice were treated with tamoxifen for 1.5 d then injected with adenovirus. Tissue was harvested 5 d postinfection. No increase in Bmi1-derived lineage striping frequency was observed relative to Ad Fc, consistent with slow cycling of Bmi1 + ISCs that is not enhanced by Rspo1- mediated Wnt stimulation. Bmi1 lineage is colored in green and DAPI in blue. Bar = 100!m. Supplemental Figure S2. Bmi1 + lineage proliferates in response to irradiation injury. (A-F) Bmi1-CreER; Rosa26-YFP mice were treated with tamoxifen for 2 d then given 12 Gy whole body irradiation. By 7 d post-irradiation injury (9 d total of tamoxifen exposure), there is marked Bmi1-derived proliferation in response to 55
67 irradiation throughout the proximal small intestine that involves multiple contiguous crypts and villi. Bar = 50!m. Supplemental Figure S3. Serial passaging of Bmi1-YFP + clonogenic spheroids. Representative spheroids derived from FACS-isolated Bmi1-YFP + single cells, marked by tamoxifen administration for 1 or 2 d in Bmi1-CreER; Rosa26-YFP mice, after 8 months of clonogenic culture and serial passaging. The spheroids were serially passaged weekly for 8 months and maintained their morphology and pan-yfp expression. Lgr5 + ISCs with the highest GFP expression (denoted Lgr5-eGFP Hi ) were isolated by FACS from Lgr5-eGFP-IRES-CreERT2 mouse small intestine and clonogenically cultured per previous reports(15) for direct comparison. Bar = 100!m. Supplemental Figure S4. Lgr5-expressing cells arise within Bmi1-YFP + clonogenic spheroids. Multiple Lgr5-expressing cells were detected by Lgr5 mrna FISH within FACS-isolated and clonally-derived Bmi1-YFP + spheroid, as revealed by simultaneous YFP immunodetection. The sense Lgr5 mrna probe was used as a negative control. Bar = 50!m. Supplemental Figure S5. 3D confocal reconstruction of Bmi1 + lineage expansion post-irradiation. Bmi1-CreER; Rosa26-YFP mice were induced with 2 d tamoxifen exposure to irreversibly label Bmi1 + ISCs with YFP then subjected to 12 Gy whole body irradiation. By 7 d post-irradiation, multiple contiguous crypts and villi of the jejunum 56
68 were repopulated by the Bmi1 + lineage (colored in green). Phalloidin is shown in red and DAPI in blue. Supporting Information Video 2. 3D confocal reconstruction of monoclonal expansion of Bmi1 + lineage post-irradiation. Bmi1-CreER; Rosa26-Confetti mice were induced with 2 d tamoxifen exposure to stochastically label Bmi1 + ISCs with 1 of 4 possible fluorescent colors prior to 12 Gy irradiation. By 7 d post-irradiation, progeny arising from the marked clones were exclusively labeled with a single color suggesting a monoclonal origin. Here the contiguous patch of crypts and villi of the repopulated jejunum arose from a clone stochastically marked with YFP (colored in yellow). 57
69 CHAPTER 3 FUNCTIONAL LOSS OF LGR5 + ISCs AND PANETH CELLS BY OVEREXPRESSION OF THE LGR5 EXTRACELLULAR DOMAIN 58
70 3.1 ABSTRACT The leucine-rich repeat containing G protein-coupled receptor 5 (Lgr5) marks actively cycling crypt base columnar cells that are multipotent, long-lived, and largely responsible for homeostatic regeneration of the intestinal epithelium. Recently, the G- protein-coupled receptors Lgr4-6 have been reported to associate with Wnt receptors to mediate R-spondin signaling. Although Paneth cells have been suggested to serve as niche cells for Lgr5 + intestinal stem cells (ISCs), perhaps through the secretion of essential paracrine factors, the precise mechanisms regulating the maintenance of Lgr5 + ISCs remain poorly understood. In this study, adenovirus expressing the soluble ligandbinding Lgr5 extracellular domain was used in vivo to examine the role of the Lgr5 receptor in the maintenance of Lgr5 + ISCs. Circulating Lgr5 ectodomain induced the migration of Paneth cells from the crypts, their eventual loss, and a concomitant disappearance of Lgr5 + ISCs. Paneth cell migration was associated with downregulation of Wnt signaling and its target EphB3. The loss of Lgr5 + ISCs did not affect maintenance of the intestinal epithelium, nor did Lgr5 + stem cells disappear from nonintestinal organs. Together, these findings characterize an easily tractable experimental model for the in vivo deletion of Lgr5 + ISCs, and suggest that Lgr receptors function to actively maintain Lgr5 + ISCs in vivo. 59
71 3.2 INTRODUCTION The gastrointestinal tract is a highly organized system of tissues and organs that functions to break down food into simple molecules and compounds that can be absorbed into the bloodstream via the small intestine. The cells lining the small intestine therefore experience significant mechanical and chemical stress requiring constant homeostatic renewal. In the mouse, the entire epithelium is regenerated every 3-5 days. The cells that are responsible for this activity are the ISCs, which reside in the crypts of Lieberkühn. These ISCs give rise to a pool of multipotent progenitors, which do not differentiate immediately, but instead rapidly proliferate to generate a pool of transit amplifying (TA) cells. These TA cells occupy the crypt length and subsequently give rise to the differentiated lineages of the intestine: the absorptive enterocytes, the mucin secreting goblet cells, the peptide hormone secreting enteroendocrine cells, the Paneth cells, M cells, tuft cells, and cup cells. How these stem cells coordinate their proliferation and differentiation is still poorly understood, but the recent identification of Lgr5 as a marker of CBCs that are long-lived and have the capacity to self-renew and differentiate into all of the adult lineages of the intestinal epithelium has yielded some insights into the evolutionarily conserved pathways that regulate their maintenance(14). Lgr4, Lgr5, and Lgr6 belong to a family of G-protein couple receptors that are related to the glycoprotein hormone receptors(86). This family of receptors possesses a large N-terminal extracellular domain comprised of a long series of leucine-rich repeats. In other organs, such as the hair follicle(21) and stomach(87), Lgr5 similarly marks a population of rapidly proliferating cells that are Wnt responsive and function as stem cells. However, Lgr5 expression is not restricted to functional stem cells of the 60
72 intestine, stomach, and hair follicle. Lgr5 also marks cells in the mammary gland, brain, and upon injury pancreas and liver(88). Therefore, the question remains whether the Lgr receptors play a functional role in stem cells and their maintenance. Until recently, the Lgr receptors were designated as orphan receptors due to the absence of a known ligand. Expression profiling from Lgr4 and Lgr5 double knockout (DKO) and APC, a negative regulator of the Wnt signaling pathway, deleted intestinal crypts revealed that their gene programs were inversely correlated. The apparent Wnt connection resulted in the independent identification by several groups of the R-spondin family of secreted proteins as the ligand for the Lgr receptors(89-92). From this work, it was further revealed that the Lgr receptors physically interact with Wnt pathway components including Frizzled5/7 and Lrp5/6(89). In vitro gain of function and loss of function studies further confirmed that all four R-spondins bind to Lgr4, Lgr5, and Lgr6, and enhance Wnt signaling(89). Here, we investigated the role of the Lgr5 receptor in the maintenance of the intestinal epithelium by in vivo overexpression of the soluble ectodomain (ECD). 3.3 RESULTS In vivo overexpression of the Lgr5 ectodomain induces migration and eventual loss of Paneth cells by inhibition of EphB3 expression in Paneth cells. Homozygous disruption of Lgr5 in mice leads to ankyloglossia, a condition in which the tongue is fused to the floor of the oral cavity in the newborn, thus resulting in neonatal lethality(93). The lethal phenotype precluded the investigation of the role of Lgr5 in adult tissues such as the intestine. However, the role of Lgr5 in the developing intestine was examined. Homozygous deletion of the Lgr5 locus resulted in the 61
73 accelerated maturation of Paneth cells(94), which normally do not develop until weaning. To investigate the role of Lgr5 in the adult intestine, we sought to downregulate Lgr5- mediated signaling by in vivo overexpression of the soluble ECD, as has been shown previously for other receptors including VEGFR(95). Therefore, we hypothesized that adenovirus-mediated overexpression of the Lgr5 ECD would result in increased numbers of Paneth cells in the adult intestine. To test this hypothesis, C57bl/6J mice were intravenously injected with 1x10 9 plaque forming units of adenovirus expressing the Lgr5 ECD (Ad Lgr5 ECD) or the control adenovirus expressing the IgG2! Fc fragment (Ad Fc), and their intestine were examined histologically. In contrast to the embryonic phenotype(94), overexpression of the soluble ECD resulted in Paneth cell migration upward from the crypt bases to the villi from d 1-d 3 and their eventual loss by d 5 postinfection (Figure 1A). Staining using the Paneth-specific marker Lysozyme at d 7 postinfection revealed sustained pan-intestinal ablation of the Paneth cells (Figure 1B). TUNEL, Annexin V, and Cleaved Caspase 3 staining did not reveal an increase in the apoptotic index of the Ad Lgr5 ECD-treated mice. In addition to producing a variety of antimicrobial factors, Paneth cells also express Wnt3!, an activator of the Wnt signaling cascade. Wnt3! activates Wnt signaling in both the Paneth cells and neighboring crypts to actively maintain the intestinal epithelium. Naturally, we examined whether overexpression of the Lgr5 ECD affected proliferation and/or Wnt signaling in the intestine. Staining of the Lgr5 ECDtreated intestine using PCNA (Figure 1C) and Ki67 revealed a marked decrease in the number of proliferative cells in the crypt compartment, though the overall architecture of the intestine remained intact. The transcription factor "-catenin is normally sequestered 62
74 in the cytoplasm of the cell, where it is targeted for proteasomal degradation. When Wnt signaling is activated, the "-catenin destruction complex is inhibited and results in "- catenin accumulation and its subsequent translocation into the nucleus to activate Wnt target genes. To determine the Wnt status of the migrating Paneth cells in Ad Lgr5 ECD-treated mice, we examined "-catenin localization in the granule-containing Paneth cells and found that on d 2 post-infection, when the Paneth cells begin to exit the crypt, migrating Paneth cells lack nuclear "-catenin staining (Figure 1D). This is in contrast to the Ad Fc-treated Paneth cells and the Ad Lgr5 ECD-treated Paneth cells that remain localized to the crypt bases, both of which continue to express nuclear "-catenin. EphB2 and EphB3 are Wnt target genes that have roles in actively maintaining the crypt/villus axis(96). EphB2 and EphB3 double knock-out intestine exhibit abnormal Paneth cells that are aberrantly localized and not restricted to the crypt base. To determine if downregulation of Wnt signaling in the migrating Paneth cells was accompanied by a specific downregulation of the EphB receptors, we examined EphB3 expression. Likewise, migrating Paneth cells from Ad Lgr5 ECD-treated mice exhibited decreased EphB3 expression compared to the Ad Fc-treated Paneth cells and the Ad Lgr5 ECD-treated Paneth cells that remain localized in the crypt bases. Examination of the other differentiated lineages revealed no apparent phenotypes. Overexpression of Lgr5 ECD did not significantly affect the number or the distribution of Chromogranin A + enteroendocrine cells and Muc2 + goblet cells (data not shown). Thus, overexpression of the Lgr5 ECD resulted in the specific migration and eventual loss of the Paneth cells from the intestinal epithelium via a Wnt/EphB3- dependent mechanism, consistent with previous reports that Paneth cells, though 63
75 sufficient in providing many ISC niche signals, are not required for maintenance of the intestinal epithelium (Kim et al, 2012; Fong et al, 2011). In vivo overexpression of Lgr5 ECD induces functional loss of Lgr5 + ISCs. Since the Lgr5 + ISCs are intimately associated with the Paneth cells, we next explored the effects of Lgr5 ECD-mediated ablation of Paneth cells on the Lgr5 + ISCs. Similarly, Lgr5-EGFP-IRES-CreERT2 mice were treated with 1x10 9 p.f.u. of Ad Fc or Ad Lgr5 ECD and examined histologically over time. Direct fluorescence detection of the Lgr5-GFP reporter revealed complete ablation of GFP expressing cells throughout the intestinal epithelium from d 3 to d 15 post-infection (Figure 2A). This loss of GFP was confirmed quantitatively by fluorescence activated cell sorting of Lgr5-eGFP + cells (Figure 2B). The complete ablation of GFP throughout the intestinal epithelium suggests that the Lgr5-GFP + ISCs had disappeared. However, it is also formally possible that CBC cells remain and continue to function as stem cells, but that they no longer express Lgr5 or GFP, if Lgr5 ECD treatment simply downregulates Lgr5-GFP transcription. Thus, to distinguish between a downregulation of Lgr5-GFP signal and a functional loss of Lgr5 + ISCs, we took advantage of well-characterized lineage tracing approaches to permanently mark Lgr5 ISCs. To this end, Lgr5-eGFP-IRES-CreERT2 mice were crossed to either Rosa26-LSL-Confetti or Rosa26-LSL-LacZ mice to generate Lgr5-eGFP-IRES- CreERT2;Rosa26-LSL-Confetti or Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-LacZ compound heterozygotes. These mice were injected with a single dose of Tamoxifen (9mg/40g) to induce Cre-mediated recombination of the LSL cassette and subsequent expression of a reporter cassette under the control of the Rosa26 promoter. Specifically, 64
76 this results in the constitutive expression of the Confetti or LacZ expression cassettes in the Lgr5 expressing cell and all of their subsequent progeny. Tamoxifen was administered before the adenovirus treatment, allowing for permanent labeling of all Lgr5-expressing cells and their progeny, regardless of the Lgr5 expression status at the time of tissue harvest that resulted from adenovirus treatment. Specifically, Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-Confetti mice were injected with a single dose of tamoxifen and 7 d later, the mice were treated with either Ad Fc or Ad Lgr5 ECD. The mice were then harvested 4 d post-infection (11 d posttamoxifen exposure), when Lgr5 expression is gone, but before complete turnover of the intestinal epithelium. Ad Lgr5 ECD-treated intestine showed lineage stripes emanating from villi, rather than from crypt bases (Figure 2C). The absence of both GFP signal and lineage arising from crypts suggests that a non Lgr5-expressing ISC is able to maintain the intestine in the absence of Lgr5 + ISC. This is in contrast to the Ad Fctreated mice, which exhibit robust lineage stripes emanating from crypt bases and extending continuously into the villus tips (Figure 2C). A similar experimental design was employed for the Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-LacZ mice, except that lineages were established for 30 d post-tamoxifen treatment (representing 4-6 complete turnovers of the epithelium) and the mice were harvested 10 d (1-2 complete turnovers of the epithelium) post-adenovirus treatment. Like the Ad Lgr5 ECD-treated intestine from the Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-Confetti mice, the intestine from the Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-LacZ mice lacked any lineage stripes (Figure 2D). Together, these findings suggest that Lgr5 may function to actively maintain Lgr5 + ISCs. 65
77 Lgr5 ECD-mediated Lgr5 + ISC ablation induces activation and de novo generation of Bmi1 + ISCs in vivo. Despite the functional loss of both the Paneth cells and Lgr5 + ISCs, the intestinal epithelium retains its overall architecture, suggesting that a reserve population of ISCs is responsible for maintaining the intestinal epithelium in the absence of Lgr5 + ISCs. Recently, three groups have reported on the lineage relationship between the Lgr5 + ISC and a quiescent + 4 ISC(71, 82, 97). Yan et al. and Tian et al. described the non-essential function of Lgr5 + ISCs in the maintenance of the intestinal epithelial maintenance. Yan et al. induced Lgr5 ablation through total body irradiation, while Tian et al. used a genetic approach to induce Lgr5 ablation by diphtheria toxin. In both cases, Lgr5 loss is associated with high levels of apoptosis and the activation of the reserve Bmi1 + ISCs. We therefore examined whether this non-injury, Lgr5 ECD-mediated model of Lgr5 + ISC ablation similarly activated Bmi1 + ISCs. To this end, Bmi1-CreER mice were crossed to Rosa26-LSL-LacZ or Rosa26- LSL-YFP to generate Bmi1-CreER;Rosa26-LSL-LacZ and Bmi1-CreER;Rosa26-LSL- YFP compound heterozygotes. To assess the Lgr5 ECD-mediated effects on Bmi1 + ISCs, we used short-term tamoxifen exposure to induce Cre-mediated recombination and selectively mark Bmi1 + ISCs in vivo. Accordingly, Bmi1-CreER;Rosa26-LSL-YFP mice were treated with Ad Lgr5 ECD or Ad Fc for 5 d to ablate the Lgr5 + ISCs. The mice were subsequently treated with tamoxifen and harvested 7 d later. FACS sorting of YFP + cells, or Bmi1 + ISCs, revealed a 2-3 fold increase in the total number of Bmi1 + ISCs in the Ad Lgr5 ECD-treated mice compared to the Ad Fc-treated mice (Figure 3A). 66
78 To examine whether the increased number of Bmi1 + cells functioned as stem cells, we used a lineage tracing strategy whereby tamoxifen was injected before or after adenovirus treatment. Tamoxifen injection before adenovirus treatment would only label pre-existing Bmi1 + ISCs and their lineage, while injection after adenovirus treatment would label both preexisting and new Bmi1 + ISCs and their progeny. In the former strategy, Bmi1-CreER;Rosa26-LSL-LacZ were injected with tamoxifen 24 hrs before treatment with Ad Lgr5 ECD or Ad Fc and harvested after 7 d, when Lgr5 + ISCs are ablated. Surprisingly, the Bmi1 lineage decreased with Lgr5 ECD treatment (Figure 3B). Histological analysis revealed that Bmi1 lineages were restricted to the crypts in the Lgr5 ECD-treated mice. This was in contrast to the Ad Fc-treated intestine which exhibited crypt and villus expression of the LacZ reporter (Figure 3C). When the Bmi1- CreER;Rosa26-LSL-LacZ mice were pre-treated with Ad Lgr5 ECD or Ad Fc, injected with tamoxifen at d 7, and harvested at d11 post-infection, the Lgr5 ECD mice exhibited a 2-3 fold increase in lineage tracing (Figure 3C), consistent with the increase observed in the total number of Bmi1 + ISCs post-lgr5 ECD-mediated Lgr5 + ISC ablation. The difference in the lineage tracing efficiency observed suggests that de novo Bmi1 + are generated from non-bmi1 + ISC populations. It further suggests that during homeostasis, Bmi1 + ISCs repopulate the intestine via Lgr5, so that Lgr5 ECD treatment results in decreased lineage striping because Lgr5 + ISCs (formerly Bmi1 + ISCs) are now rendered inactive. In the absence of Lgr5 + ISCs, pre-existing and de novo Bmi1 + ISCs can bypass Lgr5-mediated pathway and give rise to all of the differentiated lineages, with the exception of the Paneth cells, which are absent as a result of the Lgr5 ECD treatment. 67
79 3.4 DISCUSSION Our use of the Lgr5 ECD as a tool to probe the function of Lgr5 in the intestinal epithelium suggests that Lgr5 plays a critical role in the maintenance of the actively cycling Lgr5 + ISCs and their adjacent Paneth cells. Upon exposure to circulating Lgr5 ECD, the normally crypt localized Paneth cells migrate out of the crypt and are eventually lost, presumably through the same mechanism that allows for the renewal of the entire epithelium during homeostatic regeneration. This migration and eventual loss is accompanied by a concomitant functional loss of the Lgr5 + ISC. As in other models where Lgr5 + ISCs are ablated(71), we show that Bmi1 + ISCs can compensate for the loss of Lgr5 + ISCs in vivo. Interestingly, the use of various labeling strategies revealed a novel relationship between the Lgr5 + and Bmi1 + ISCs. It seems that during homeostasis, Bmi1 + ISCs as the reserve ISC population, preferentially differentiates through an Lgr5-dependent pathway. Under conditions where Lgr5 + ISCs are ablated, Bmi1 + ISCs can bypass this preferred differentiation pathway and give rise to differentiated progeny without becoming an Lgr5 + ISC. This is in accordance with a recent report by Yan et al. showing that after whole body irradiation, Bmi1 + ISCs can regenerate the intestinal epithelium in the absence of Lgr5 + ISCs(97). Tian et al. similarly showed that Lgr5 + ISC ablation, via a diphtheria toxin receptor (dtr) knock-in genetic strategy, was accompanied by an expansion of the Bmi1 + lineage(71). In their system, continued injection of the diphtheria toxin was required to ablate existing and newly forming Lgr5 + ISCs, making it impossible to determine the differentiation pathway of the Bmi1 + ISCs. In contrast, our system offers 68
80 an easily tractable experimental model for the sustained in vivo deletion of Lgr5 + ISCs, clearly revealing a novel relationship between these two ISC populations. The labeling of Bmi1 + ISCs before adenovirus treatment further revealed that de novo Bmi1 + ISCs do not arise from pre-existing Bmi1 + ISCs since prelabeling results in decreased lineage tracing despite and increase in the number of Bmi1 + ISCs. Therefore, Bmi1 expression is induced after Ad Lgr5 ECD treatment in cells that did not previously express Bmi1. Whether these cells are other + 4 ISCs, such as those expressing Hopx, or early progenitor cells is unknown. Furthermore, these findings suggest that differences exist between pre-existing and de novo Bmi1 + ISCs. Pre-existing Bmi1 + ISCs are unable to bypass Lgr5-mediated differentiation, while de novo Bmi1 + ISCs have the ability to differentiate in an Lgr5-independent mechanism. Interestingly, the Lgr5 + lineage, upon Lgr5 ECD overexpression, was lost even when the lineage was allowed to trace for 30 d. Over 30 d, the intestinal epithelium experiences, on average, 4-6 generations of complete intestinal turnover. The absence of tracing therefore suggests that the conversion during homeostasis from Lgr5 + ISCs to other reserve ISC populations, such as Bmi1, which is not sensitive to Lgr5 ECD treatment, is rare. Further studies are required to clarify whether the Lgr5 + ISC phenotype is primary or secondary to the Paneth cell phenotype. However, numerous studies offer evidence to support the hypothesis that the Lgr5 ECD likely acts on ISCs and that the Paneth cell phenotype is a secondary effect. Durand et al. demonstrated that deletion of the transcription factor Math1 (Atoh1), an essential driver of secretory cell differentiation, was sufficient to similarly ablate the Paneth cell lineage(98). In contrast with our 69
81 findings, the maintenance and proliferation of Lgr5 + ISCs continued in the absence of Paneth cells. If Lgr5 ECD only inhibited Wnt signaling in the Paneth cells, the Lgr5 + ISCs should retain the ability to function as stem cells. Their concomitant disappearance therefore suggests that Lgr5 ECD affects Lgr5 + ISCs directly. Sato et al. first demonstrated the requirement of R-spondins in the maintenance of sorted Lgr5 + ISCs in vitro(15). The requirement of R-spondins in vivo has not yet been determined. However it is known that Lgr5 + ISCs are exquisitely sensitive to Wnt signaling modulation(97), suggesting the in vivo requirement of these Wnt agonists. The recent deorphanization of the Lgr family of receptors with the identification of the R- spondin family of secreted proteins as the putative ligands for Lgr4-6(89-92) further supports our hypothesis that Lgr5 ECD acts directly on Lgr5 + ISCs by inhibiting endogenous R-spondin interaction with Lgr5. Our findings are additionally interesting given the recent identification of recurrent R-spondin fusions in colon cancer that occur in about 10% of colon tumors(99). These R-spondin fusions are mutually exclusive with APC mutations, retain the ability to potentiate Wnt signaling, and likely play a key role in tumorigenesis. Overall, Lgr5 ECD treatment represents an easily tractable experimental model for the in vivo deletion of Lgr5 + ISCs, and possibly, Lgr5 + CSCs, and therefore may represent a novel method for therapeutic intervention in colon cancer. 70
82 3.5 METHODS Animals. All mice in a mixed genetic background were used between the ages of 2-6 months and housed on a 12-h light/dark cycle in a pathogen-free animal facility. Lgr5- egfp-ires-creert2(barker 2007) mice were crossed to Rosa26-LSL-LacZ, Rosa26- LSL-TdTomato, or Rosa26-Confetti (all purchased from Jackson Laboratory) mice to generate Lgr5-eGFP-IRES-CreER;Rosa26-LSL-LacZ, Lgr5-eGFP-IRES-CreER;Rosa26- LSL-TdTomato, and Lgr5-eGFP-IRES-CreER;Rosa26-LSL-Confetti compound heterozygotes. Likewise, Bmi1-CreER(Sangiorgi 2008) mice were crossed to Rosa26- LSL-LacZ or Rosa26-LSL-YFP (purchased from Jackson Laboratory) mice to generate Bmi1-CreER;Rosa26-LSL-LacZ, and Bmi1-CreER;Rosa26-LSL-YFP compound heterozygotes. To label Bmi1 ISCs, and to label the Lgr5 and Bmi1 lineages, mice were administered a single dose of intraperitoneal tamoxifen (9mg/40g). Mice were injected with adenovirus expressing the Lgr5 ECD or adenovirus expressing Fc through the tail vein with 1x10 9 plaque forming unites (p.f.u) in 0.1 ml PBS. All animal studies were performed in accordance with the National Institutes of Health guidelines for use and care of live animals and were approved by the Institutional Animal Care and Use Committee. Adenovirus. The extracellular domain of Lgr5 was amplified from human cdna by PCR, sequenced, and cloned into the E1 region of E1-E20 Ad strain 5 by homologous recombination followed by adenovirus production in 293 cells and CsCl gradient 71
83 purification of virus as described. The construction of Ad Fc (encoding IgG2! Fc) has been previously described. Immunofluoresence. Paraffin-embedded sections were stained by using the following primary antibodies: anti-muc2 (Santa Cruz), lysozyme (Dako), chromogranin A (Dako), E-cadherin (BD Biosciences), sucrase-isomaltase (generated in house at Stanford University), b-catenin, and EphB3. All primary antibodies were used at 1:500. Cy3 and Cy5 conjugated secondary antibodies (Santa Cruz) were used at 1:2,000. Alexa Fluor 594-conjugated Phalloidin (Invitrogen) was used at 1:500. Images were captured on a Zeiss AxioImager Z1 with ApoTome attachment or Leica CTR 6500 confocal microscope. X-gal staining. Tissues were fixed for 2 hours 4% PFA rinsed twice in PBS and then incubated for O/N hours at 4C in the staining solution [2mM MgCl2, 5mM K3Fe(CN)6, 5mM K4Fe(CN)6, 0.01% NP40, 0.01% sodium deoxycholate and 1mg/ml X-gal in PBS]. Flow Cytometry. FACS experiments were performed by using fresh small intestine epithelial preparations. A standardized 3-cm segment of proximal jejunum was used for quantitative FACS analysis of ISC populations. Intestinal epithelial cells were extracted from en bloc resected small intestine with 10 mm EDTA and manual shaking, followed by enzymatic dissociation with collagenase/dispase (Roche) to generate a single-cell suspension (1). Singlet discrimination was sequentially performed by using plots for FSC 72
84 (FSC-A vs. FSC-H) and SSC (SSC-W vs. SSC-H). Dead cells were excluded by scatter characteristics and propidium iodide. Epithelial cells were identified by CD45! and EpCAM + immunostaining. ISCs were identified by their endogenous GFP or YFP expression. All FACS experiments were performed on an Aria II sorter (BD Biosciences) at the Stanford University Shared FACS Facility, and FACS data were analyzed by using FlowJo software (Treestar). 73








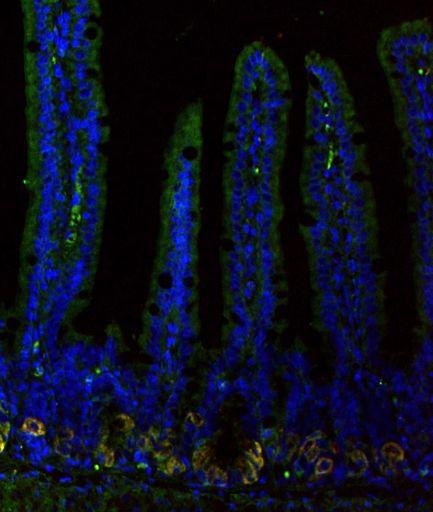



85 3.6 FIGURES Figure 1 $=( $>( $7( -$(./( -$(4567(89!( -$(./( -$34567(89!(( 4:;#<:'%(!"#$%&"'( )%*"&"'( +,%"'( +,%"'( +,%"'( -$(./(0!12( 8EF?=( -$34567(89!(0!12( 4:;#<:'%( C9D-(?3/@A%&B&( G%65%( 74






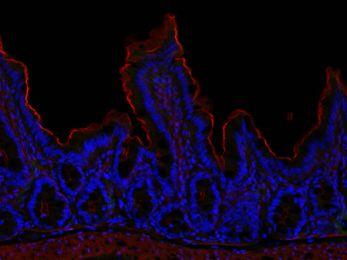







86 Figure 2! N. ' '12"3415# 67315# 8,29:5;7#.2724# *+,(<6=->.,3-=?# *+,(<6=->.,3-=?# '0# 'M# '//# G!H#!"# I;,J3KL# '0# 'E0# 'F0# G!H#!"# I;,J3KL# 75


#*+,#,-#,.")

87 Figure 3!# 9#!"#$%#,-#,.#,/# 0!1#!"# 23(4567# +#!"#$%#!"#&'()#*+,#,-#,.#,/#,8#!"# 0!1# 23(4567#!"#&'()#*+,#!"#$%#!"#&'()#*+,# 76
88 3.7 FIGURE LEGENDS Figure 1. In vivo overexpression of the Lgr5 ectodomain induces migration and eventual loss of Paneth cells by inhibition of EphB3 expression in Paneth cells. (A) H&E staining of Ad Fc or Ad Lgr5 ECD treated mice. Paneth cells migrate out of the crypt bases and disappear by d 5 post-infection. (B) Lysozyme + Paneth cells completely disappear from the crypts throughout the intestinal epithelium at d 7 post-infection. (C) Decreased number of proliferating cells in the duodenum by PCNA. (D) Absent nuclear "-catenin staining, and therefore decreased Wnt signaling, in migrating Paneth cells. (E) Colocalization (yellow) of EphB3 (green) and Lysozyme (red) in cryptlocalized Paneth cells in both Ad Fc and Ad Lgr5 ECD treated intestine. Migrating Paneth cells (red) lose EphB3 expression. Figure 2. In vivo overexpression of Lgr5 ECD induces functional loss of Lgr5 + ISCs. (A) Lgr5-eGFP-IRES-CreERT2 mice were treated with Ad Fc or Ad Lgr5 ECD. Direct fluorescence detection of Lgr5-eGFP +, Phalloidin (red), and DAPI (blue). Ad Lgr5 ECD treated intestine exhibit complete loss of GFP signal at d 5 and d 10 post-infection. (B) FACS analysis of Ad Fc- or Ad Lgr5-treated intestine reveal complete loss of Lgr5- egfp + ISCs. (C) Lgr5-eGFP-IRES-CreERT2;Rosa26-LSL-Confetti mice were treated with tamoxifen at d 0 and Ad Fc or Ad Lgr5 ECD at d 7 and harvest at d 11. Lineage trace emerges from mid villus, revealing functional loss of Lgr5 + ISCs. (D) Lgr5-eGFP- IRES-CreERT2;Rosa26-LSL-LacZ mice were treated with tamoxifen at d 0 and Ad Fc or Ad Lgr5 ECD at d 30 and the intestine was harvested at d 40. No lineage is detected in Ad Lgr5-treated intestine. 77
89 Figure 3. Lgr5 ECD-mediated Lgr5 + ISC ablation induces activation and de novo generation of Bmi1 + ISCs in vivo. (A) Bmi1-CreER;Rosa26-LSL-YFP mice were infected with Ad Fc or Ad Lgr5 ECD at d 0, injected with tamoxifen at d 6, and harvested at d 7 for FACS analysis. Ad Lgr5 ECD-treated mice exhibited a 2-3 fold increase in the number of Bmi1 + ISCs. (B) Bmi1-CreER;Rosa26-LSL-LacZ mice were treated with tamoxifen at d 0, infected with adenovirus at d1, and harvested at d4. There is decreased X-gal staining in Ad Lgr5 ECD-treated intestine. (C) Bmi1-CreER;Rosa26- LSL-LacZ mice were treated with Ad Fc or Ad Lgr5 ECD at d 1, injected with tamoxifen at d 4, and harvested at d 8. Ad Lgr5 ECD-treatment increased the amount of lineage tracing from the Bmi1 locus. 78

90 3.8 SUPPLEMENTAL FIGURES Supplemental Figure S1. 79

91 Supplemental Figure S2. 80


92 Supplemental Figure S3. 81
CHAPTER 6 SUMMARIZING DISCUSSION
 CHAPTER 6 SUMMARIZING DISCUSSION More than 20 years ago the founding member of the Wnt gene family, Wnt-1/Int1, was discovered as a proto-oncogene activated in mammary gland tumors by the mouse mammary
CHAPTER 6 SUMMARIZING DISCUSSION More than 20 years ago the founding member of the Wnt gene family, Wnt-1/Int1, was discovered as a proto-oncogene activated in mammary gland tumors by the mouse mammary
Summary and Concluding Remarks
 Summary and Concluding Remarks Chapter 6 The intestinal epithelium provides an excellent model system for investigating molecular mechanisms regulating cell lineage establishment, stem cell proliferation,
Summary and Concluding Remarks Chapter 6 The intestinal epithelium provides an excellent model system for investigating molecular mechanisms regulating cell lineage establishment, stem cell proliferation,
From crypt stem cell to colorectal cancer
 19 3 2007 6 Chinese Bulletin of Life Sciences Vol. 19, No. 3 Jun., 2007 1004-0374(2007)03-0321-05 ( 510405) Wnt Notch BMP R735.35; R730.21 A From crypt stem cell to colorectal cancer WEN Bin*, CHEN Weiwen
19 3 2007 6 Chinese Bulletin of Life Sciences Vol. 19, No. 3 Jun., 2007 1004-0374(2007)03-0321-05 ( 510405) Wnt Notch BMP R735.35; R730.21 A From crypt stem cell to colorectal cancer WEN Bin*, CHEN Weiwen
SUPPLEMENTARY INFORMATION
 b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
Neoplasia 18 lecture 6. Dr Heyam Awad MD, FRCPath
 Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Molecular Regulation of Intestinal Stem Cells
 Molecular Regulation of Intestinal Stem Cells Review Tulay Bayram Biomedical Sciences University of Utrecht Table of contents Abstract... 3 1. INTRODUCTION... 4 1.1 Structure of the small intestine and
Molecular Regulation of Intestinal Stem Cells Review Tulay Bayram Biomedical Sciences University of Utrecht Table of contents Abstract... 3 1. INTRODUCTION... 4 1.1 Structure of the small intestine and
ANAT3231: lectures overview
 ANAT3231: lectures overview Stem Cell Biology Stem Cell Technology Resources: http://php.med.unsw.edu.au/cell biology/ Essential Cell Biology 3 rd edition Alberts Dr Annemiek Beverdam School of Medical
ANAT3231: lectures overview Stem Cell Biology Stem Cell Technology Resources: http://php.med.unsw.edu.au/cell biology/ Essential Cell Biology 3 rd edition Alberts Dr Annemiek Beverdam School of Medical
RECENT ADVANCES IN BASIC SCIENCE STEM CELL IN GASTROINTESTINAL STRUCTURE AND NEOPLASTIC DEVELOPMENT
 See end of article for authors affiliations Correspondence to: M Brittan, Histopathology Unit, Cancer Research UK, 44 Lincoln s Inn Fields, London WC2A 3PX, UK; mairi.brittan@cancer.org.uk RECENT ADVANCES
See end of article for authors affiliations Correspondence to: M Brittan, Histopathology Unit, Cancer Research UK, 44 Lincoln s Inn Fields, London WC2A 3PX, UK; mairi.brittan@cancer.org.uk RECENT ADVANCES
ANAT3231: lectures overview
 ANAT3231: lectures overview Stem Cell Biology Stem Cell Technology Resources: http://php.med.unsw.edu.au/cell biology/ Essential Cell Biology 3 rd edition Alberts Dr Annemiek Beverdam School of Medical
ANAT3231: lectures overview Stem Cell Biology Stem Cell Technology Resources: http://php.med.unsw.edu.au/cell biology/ Essential Cell Biology 3 rd edition Alberts Dr Annemiek Beverdam School of Medical
Hematopoiesis. - Process of generation of mature blood cells. - Daily turnover of blood cells (70 kg human)
") Hematopoiesis - Process of generation of mature blood cells - Daily turnover of blood cells (70 kg human) 1,000,000,000,000 total cells 200,000,000,000 red blood cells 70,000,000,000 neutrophils Hematopoiesis
Hematopoiesis - Process of generation of mature blood cells - Daily turnover of blood cells (70 kg human) 1,000,000,000,000 total cells 200,000,000,000 red blood cells 70,000,000,000 neutrophils Hematopoiesis
Cancer and Oncogenes Bioscience in the 21 st Century. Linda Lowe-Krentz October 11, 2013
 Cancer and Oncogenes Bioscience in the 21 st Century Linda Lowe-Krentz October 11, 2013 Just a Few Numbers Becoming Cancer Genetic Defects Drugs Our friends and family 200 180 160 140 120 100 80 60 40
Cancer and Oncogenes Bioscience in the 21 st Century Linda Lowe-Krentz October 11, 2013 Just a Few Numbers Becoming Cancer Genetic Defects Drugs Our friends and family 200 180 160 140 120 100 80 60 40
CHAPTER 3. Expression pattern of Wnt signaling components in the adult intestine. Gastroenterology 23: (2005)
") CHAPTER 3 Expression pattern of Wnt signaling components in the adult intestine Gastroenterology 23:1825-1833 (2005) Chapter 3 Expression pattern of Wnt signaling components in the adult intestine Alex
CHAPTER 3 Expression pattern of Wnt signaling components in the adult intestine Gastroenterology 23:1825-1833 (2005) Chapter 3 Expression pattern of Wnt signaling components in the adult intestine Alex
Stem cells in homeostasis and cancer of the gut
 Heijden and Vermeulen Molecular Cancer (2019) 18:66 https://doi.org/10.1186/s12943-019-0962-x REVIEW Stem cells in homeostasis and cancer of the gut Maartje van der Heijden and Louis Vermeulen * Open Access
Heijden and Vermeulen Molecular Cancer (2019) 18:66 https://doi.org/10.1186/s12943-019-0962-x REVIEW Stem cells in homeostasis and cancer of the gut Maartje van der Heijden and Louis Vermeulen * Open Access
Meeting Report. From December 8 to 11, 2012 at Atlanta, GA, U.S.A
 Meeting Report Affiliation Department of Transfusion Medicine and Cell Therapy Name Hisayuki Yao Name of the meeting Period and venue Type of your presentation Title of your presentation The 54 th Annual
Meeting Report Affiliation Department of Transfusion Medicine and Cell Therapy Name Hisayuki Yao Name of the meeting Period and venue Type of your presentation Title of your presentation The 54 th Annual
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Introduction. Cancer Biology. Tumor-suppressor genes. Proto-oncogenes. DNA stability genes. Mechanisms of carcinogenesis.
 Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
A class of genes that normally suppress cell proliferation. p53 and Rb..ect. suppressor gene products can release cells. hyperproliferation.
 Tumor Suppressor Genes A class of genes that normally suppress cell proliferation. p53 and Rb..ect Mutations that inactivate the tumor suppressor gene products can release cells from growth suppression
Tumor Suppressor Genes A class of genes that normally suppress cell proliferation. p53 and Rb..ect Mutations that inactivate the tumor suppressor gene products can release cells from growth suppression
Characteristics of Cancer Stem Cells (CSCs)
") GENReports: Market & Tech Analysis Characteristics of Cancer Stem Cells (CSCs) > Enal Razvi, Ph.D. Biotechnology Analyst, Managing Director Select Biosciences, Inc. enal@selectbio.us! Topic,IntroducEon,and,Scope!
GENReports: Market & Tech Analysis Characteristics of Cancer Stem Cells (CSCs) > Enal Razvi, Ph.D. Biotechnology Analyst, Managing Director Select Biosciences, Inc. enal@selectbio.us! Topic,IntroducEon,and,Scope!
Quantification of early stage lesions for loss of p53 should be shown in the main figures.
 Reviewer #1 (Remarks to the Author): Expert in prostate cancer The manuscript "Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers" uses a number of novel genetically engineered
Reviewer #1 (Remarks to the Author): Expert in prostate cancer The manuscript "Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers" uses a number of novel genetically engineered
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Biochemistry of Carcinogenesis. Lecture # 35 Alexander N. Koval
 Biochemistry of Carcinogenesis Lecture # 35 Alexander N. Koval What is Cancer? The term "cancer" refers to a group of diseases in which cells grow and spread unrestrained throughout the body. It is difficult
Biochemistry of Carcinogenesis Lecture # 35 Alexander N. Koval What is Cancer? The term "cancer" refers to a group of diseases in which cells grow and spread unrestrained throughout the body. It is difficult
MBios 401/501: Lecture 12.1 Signaling IV. Slide 1
 MBios 401/501: Lecture 12.1 Signaling IV Slide 1 Pathways that require regulated proteolysis 1. Notch and Delta 2. Wnt/ b-catenin 3. Hedgehog 4. NFk-B Our last topic on cell signaling are pathways that
MBios 401/501: Lecture 12.1 Signaling IV Slide 1 Pathways that require regulated proteolysis 1. Notch and Delta 2. Wnt/ b-catenin 3. Hedgehog 4. NFk-B Our last topic on cell signaling are pathways that
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer and Oncogenes Bioscience in the 21 st Century. Linda Lowe-Krentz
 Cancer and Oncogenes Bioscience in the 21 st Century Linda Lowe-Krentz December 1, 2010 Just a Few Numbers Becoming Cancer Genetic Defects Drugs Our friends and family 25 More mutations as 20 you get older
Cancer and Oncogenes Bioscience in the 21 st Century Linda Lowe-Krentz December 1, 2010 Just a Few Numbers Becoming Cancer Genetic Defects Drugs Our friends and family 25 More mutations as 20 you get older
Animal Tissue Culture SQG 3242 Biology of Cultured Cells. Dr. Siti Pauliena Mohd Bohari
 Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
Cell Communication and Cell Signaling
 Cell Communication and Cell Signaling Why is cell signaling important? Why is cell signaling important? Allows cells to communicate and coordinate functions/activities of the organism Usually involves
Cell Communication and Cell Signaling Why is cell signaling important? Why is cell signaling important? Allows cells to communicate and coordinate functions/activities of the organism Usually involves
609G: Concepts of Cancer Genetics and Treatments (3 credits)
") Master of Chemical and Life Sciences Program College of Computer, Mathematical, and Natural Sciences 609G: Concepts of Cancer Genetics and Treatments (3 credits) Text books: Principles of Cancer Genetics,
Master of Chemical and Life Sciences Program College of Computer, Mathematical, and Natural Sciences 609G: Concepts of Cancer Genetics and Treatments (3 credits) Text books: Principles of Cancer Genetics,
Genetics and Cancer Ch 20
 Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
number Done by Corrected by Doctor Maha Shomaf
 number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
Colonic polyps and colon cancer. Andrew Macpherson Director of Gastroentology University of Bern
 Colonic polyps and colon cancer Andrew Macpherson Director of Gastroentology University of Bern Improtance of the problem of colon cancers - Epidemiology Lifetime risk 5% Incidence/10 5 /annum (US Detroit
Colonic polyps and colon cancer Andrew Macpherson Director of Gastroentology University of Bern Improtance of the problem of colon cancers - Epidemiology Lifetime risk 5% Incidence/10 5 /annum (US Detroit
Journal Club. 03/04/2012 Lama Nazzal
 Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
Development of Carcinoma Pathways
 The Construction of Genetic Pathway to Colorectal Cancer Moriah Wright, MD Clinical Fellow in Colorectal Surgery Creighton University School of Medicine Management of Colon and Diseases February 23, 2019
The Construction of Genetic Pathway to Colorectal Cancer Moriah Wright, MD Clinical Fellow in Colorectal Surgery Creighton University School of Medicine Management of Colon and Diseases February 23, 2019
Stem Cells. Induced Stem Cells
 Induced Stem Cells Stem Cells Mouse and human somatic cells can either be reprogrammed to a pluripotent state or converted to another lineage with a combination of transcription factors suggesting that
Induced Stem Cells Stem Cells Mouse and human somatic cells can either be reprogrammed to a pluripotent state or converted to another lineage with a combination of transcription factors suggesting that
Cell Death & Renewal (part 2)
") 17 Cell Death & Renewal (part 2) Programmed Cell Death A major signaling pathway that promotes cell survival is initiated by the enzyme PI 3-kinase, which phosphorylates PIP2 to form PIP3, which activates
17 Cell Death & Renewal (part 2) Programmed Cell Death A major signaling pathway that promotes cell survival is initiated by the enzyme PI 3-kinase, which phosphorylates PIP2 to form PIP3, which activates
oncogenes-and- tumour-suppressor-genes)
") Special topics in tumor biochemistry oncogenes-and- tumour-suppressor-genes) Speaker: Prof. Jiunn-Jye Chuu E-Mail: jjchuu@mail.stust.edu.tw Genetic Basis of Cancer Cancer-causing mutations Disease of aging
Special topics in tumor biochemistry oncogenes-and- tumour-suppressor-genes) Speaker: Prof. Jiunn-Jye Chuu E-Mail: jjchuu@mail.stust.edu.tw Genetic Basis of Cancer Cancer-causing mutations Disease of aging
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
Contents. Preface XV Acknowledgments XXI List of Abbreviations XXIII About the Companion Website XXIX
 Contents Preface XV Acknowledgments XXI List of Abbreviations XXIII About the Companion Website XXIX 1 General Aspects of Signal Transduction and Cancer Therapy 1 1.1 General Principles of Signal Transduction
Contents Preface XV Acknowledgments XXI List of Abbreviations XXIII About the Companion Website XXIX 1 General Aspects of Signal Transduction and Cancer Therapy 1 1.1 General Principles of Signal Transduction
Modeling colorectal cancer as a 3-dimensional disease in a dish: the case for drug screening using organoids, zebrafish, and fruit flies
 University of Massachusetts Amherst From the SelectedWorks of Michele Markstein 2013 Modeling colorectal cancer as a 3-dimensional disease in a dish: the case for drug screening using organoids, zebrafish,
University of Massachusetts Amherst From the SelectedWorks of Michele Markstein 2013 Modeling colorectal cancer as a 3-dimensional disease in a dish: the case for drug screening using organoids, zebrafish,
Targeting the cgmp Pathway to Treat Colorectal Cancer
 Thomas Jefferson University Jefferson Digital Commons Department of Pharmacology and Experimental Therapeutics Faculty Papers Department of Pharmacology and Experimental Therapeutics 29 Targeting the cgmp
Thomas Jefferson University Jefferson Digital Commons Department of Pharmacology and Experimental Therapeutics Faculty Papers Department of Pharmacology and Experimental Therapeutics 29 Targeting the cgmp
Wnt signaling. Ramray Bhat.
 Wnt signaling Ramray Bhat ramray@mrdg.iisc.ernet.in Starting with animal biology and viral infections The discovery of certain laboratory murine strains that were highly susceptible to mammary gland cancer.
Wnt signaling Ramray Bhat ramray@mrdg.iisc.ernet.in Starting with animal biology and viral infections The discovery of certain laboratory murine strains that were highly susceptible to mammary gland cancer.
Foxm1 Transcription Factor is Required for Lung Fibrosis and Epithelial to Mesenchymal Transition.
 Manuscript EMBO-2012-82682 Foxm1 Transcription Factor is Required for Lung Fibrosis and Epithelial to Mesenchymal Transition. David Balli, Vladimir Ustiyan, Yufang Zhang, I-Ching Wang, Alex J. Masino,
Manuscript EMBO-2012-82682 Foxm1 Transcription Factor is Required for Lung Fibrosis and Epithelial to Mesenchymal Transition. David Balli, Vladimir Ustiyan, Yufang Zhang, I-Ching Wang, Alex J. Masino,
Multistep nature of cancer development. Cancer genes
 Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
A. Incorrect! All the cells have the same set of genes. (D)Because different types of cells have different types of transcriptional factors.
Because different types of cells have different types of transcriptional factors.") Genetics - Problem Drill 21: Cytogenetics and Chromosomal Mutation No. 1 of 10 1. Why do some cells express one set of genes while other cells express a different set of genes during development? (A) Because
Genetics - Problem Drill 21: Cytogenetics and Chromosomal Mutation No. 1 of 10 1. Why do some cells express one set of genes while other cells express a different set of genes during development? (A) Because
Early cell death (FGF) B No RunX transcription factor produced Yes No differentiation
 B No RunX transcription factor produced Yes No differentiation") Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Cancer Stem Cells & Glioblastoma
 Cancer Stem Cells & Glioblastoma JP Hugnot «Brain plasticity, Neural stem cells and Glial tumors» INSERM U1051-UM2 Institut des Neurosciences de Montpellier Montpellier 1-Stem cells and Brain Stem Cells
Cancer Stem Cells & Glioblastoma JP Hugnot «Brain plasticity, Neural stem cells and Glial tumors» INSERM U1051-UM2 Institut des Neurosciences de Montpellier Montpellier 1-Stem cells and Brain Stem Cells
Enterprise Interest Nothing to declare
 Enterprise Interest Nothing to declare Update of mixed tumours of the GI tract, the pancreas and the liver Introduction to the concept of mixed tumours and clinical implication Jean-Yves SCOAZEC Surgical
Enterprise Interest Nothing to declare Update of mixed tumours of the GI tract, the pancreas and the liver Introduction to the concept of mixed tumours and clinical implication Jean-Yves SCOAZEC Surgical
The Wnt target MASH-2 regulates expression of the putative intestinal specific stemcell markers Sox4 and Sfrp5.
 The Wnt target MASH-2 regulates expression of the putative intestinal specific stemcell markers Sox4 and Sfrp5. Submitted for publication: Irma M. Oving *, Marielle E. van Gijn *, Johan H. van Es, Marc
The Wnt target MASH-2 regulates expression of the putative intestinal specific stemcell markers Sox4 and Sfrp5. Submitted for publication: Irma M. Oving *, Marielle E. van Gijn *, Johan H. van Es, Marc
TITLE: A PSCA Promoter Based Avian Retroviral Transgene Model of Normal and Malignant Prostate
 AD Award Number: DAMD17-03-1-0163 TITLE: A PSCA Promoter Based Avian Retroviral Transgene Model of Normal and Malignant Prostate PRINCIPAL INVESTIGATOR: Robert Reiter, M.D. CONTRACTING ORGANIZATION: The
AD Award Number: DAMD17-03-1-0163 TITLE: A PSCA Promoter Based Avian Retroviral Transgene Model of Normal and Malignant Prostate PRINCIPAL INVESTIGATOR: Robert Reiter, M.D. CONTRACTING ORGANIZATION: The
FOR REVIEW. BMB Reports - Manuscript Submission. Manuscript Draft. Manuscript Number: BMB
 BMB Reports - Manuscript Submission Manuscript Draft Manuscript Number: BMB-18-095 Title: Insulin Receptor Substrate 2:A Bridge between Hippo and AKT Pathways Article Type: Perspective (Invited Only) Keywords:
BMB Reports - Manuscript Submission Manuscript Draft Manuscript Number: BMB-18-095 Title: Insulin Receptor Substrate 2:A Bridge between Hippo and AKT Pathways Article Type: Perspective (Invited Only) Keywords:
Problem Set 8 Key 1 of 8
 7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
Haematopoietic stem cells
 Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Molecular biology :- Cancer genetics lecture 11
 Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
Molecular biology :- Cancer genetics lecture 11 -We have talked about 2 group of genes that is involved in cellular transformation : proto-oncogenes and tumour suppressor genes, and it isn t enough to
icamp: Cancer biology tutorial II: recent developments in tumor biology, experimental methodology, and reference identification
 icamp: Cancer biology tutorial II: recent developments in tumor biology, experimental methodology, and reference identification Stem cells and the environment in the adenoma-carcinoma sequence (Medema,
icamp: Cancer biology tutorial II: recent developments in tumor biology, experimental methodology, and reference identification Stem cells and the environment in the adenoma-carcinoma sequence (Medema,
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Introduction to Genetics
 Introduction to Genetics Table of contents Chromosome DNA Protein synthesis Mutation Genetic disorder Relationship between genes and cancer Genetic testing Technical concern 2 All living organisms consist
Introduction to Genetics Table of contents Chromosome DNA Protein synthesis Mutation Genetic disorder Relationship between genes and cancer Genetic testing Technical concern 2 All living organisms consist
Principles of cell signaling Lecture 4
 Principles of cell signaling Lecture 4 Johan Lennartsson Molecular Cell Biology (1BG320), 2014 Johan.Lennartsson@licr.uu.se 1 Receptor tyrosine kinase-induced signal transduction Erk MAP kinase pathway
Principles of cell signaling Lecture 4 Johan Lennartsson Molecular Cell Biology (1BG320), 2014 Johan.Lennartsson@licr.uu.se 1 Receptor tyrosine kinase-induced signal transduction Erk MAP kinase pathway
Prof. R. V. Skibbens. BIOS 10 and BIOS 90: BioScience in the 21 st Century. Cell Cycle, Cell Division and intro to Cancer.
 Prof. R. V. Skibbens August 31, 2015 BIOS 10 and BIOS 90: BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell Cycle Why a cell cycle? What is the goal? trauma growth development
Prof. R. V. Skibbens August 31, 2015 BIOS 10 and BIOS 90: BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell Cycle Why a cell cycle? What is the goal? trauma growth development
Cover Page. The handle holds various files of this Leiden University dissertation.
 Cover Page The handle http://hdl.handle.net/1887/22278 holds various files of this Leiden University dissertation. Author: Cunha Carvalho de Miranda, Noel Filipe da Title: Mismatch repair and MUTYH deficient
Cover Page The handle http://hdl.handle.net/1887/22278 holds various files of this Leiden University dissertation. Author: Cunha Carvalho de Miranda, Noel Filipe da Title: Mismatch repair and MUTYH deficient
mirna Dr. S Hosseini-Asl
 mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
Src-INACTIVE / Src-INACTIVE
 Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
 Rath, N., and Olson, M. (2016) Regulation of pancreatic cancer aggressiveness by stromal stiffening. Nature Medicine, 22(5), pp. 462-463. There may be differences between this version and the published
Rath, N., and Olson, M. (2016) Regulation of pancreatic cancer aggressiveness by stromal stiffening. Nature Medicine, 22(5), pp. 462-463. There may be differences between this version and the published
VIII Curso Internacional del PIRRECV. Some molecular mechanisms of cancer
 VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Neuroepithelial Cells and Neural Differentiation
 Neuroepithelial Cells and Neural Differentiation Neurulation The cells of the neural tube are NEUROEPITHELIAL CELLS Neural crest cells migrate out of neural tube Neuroepithelial cells are embryonic stem
Neuroepithelial Cells and Neural Differentiation Neurulation The cells of the neural tube are NEUROEPITHELIAL CELLS Neural crest cells migrate out of neural tube Neuroepithelial cells are embryonic stem
ulcer healing role 118 Bicarbonate, prostaglandins in duodenal cytoprotection 235, 236
 Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:
 into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:") Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Lecture 10. G1/S Regulation and Cell Cycle Checkpoints. G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint
 Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
- is a common disease - 1 person in 3 can expect to contract cancer at some stage in their life -1 person in 5 can expect to die from it
 MBB157 Dr D Mangnall The Molecular Basis of Disease CANCER Lecture 1 One of the simpler (and better) definitions of cancer comes from the American Cancer Society, who define cancer as; 'Cancer is a group
MBB157 Dr D Mangnall The Molecular Basis of Disease CANCER Lecture 1 One of the simpler (and better) definitions of cancer comes from the American Cancer Society, who define cancer as; 'Cancer is a group
C. elegans Embryonic Development
 Autonomous Specification in Tunicate development Autonomous & Conditional Specification in C. elegans Embryonic Development Figure 8.36 Bilateral Symmetry in the Egg of the Tunicate Styela partita Fig.
Autonomous Specification in Tunicate development Autonomous & Conditional Specification in C. elegans Embryonic Development Figure 8.36 Bilateral Symmetry in the Egg of the Tunicate Styela partita Fig.
UNC-Duke Biology Course for Residents Fall Cell Cycle Effects of Radiation
 UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
Protein Kinase C Modulates Wnt Signaling In Colon Tumoral Cell Lines
 Protein Kinase C Modulates Wnt Signaling In Colon Tumoral Cell Lines Dra Martha Robles Flores Department of Biochemistry Facultad de Medicina Universidad Nacional Autónoma de México Tissue anatomy of the
Protein Kinase C Modulates Wnt Signaling In Colon Tumoral Cell Lines Dra Martha Robles Flores Department of Biochemistry Facultad de Medicina Universidad Nacional Autónoma de México Tissue anatomy of the
5.1. KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. 68 Reinforcement Unit 2 Resource Book
 5.1 THE CELL CYCLE KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. Cells have a regular pattern of growth, DNA duplication, and division that is called the cell cycle.
5.1 THE CELL CYCLE KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. Cells have a regular pattern of growth, DNA duplication, and division that is called the cell cycle.
Supplemental Figure 1: Lrig1-Apple expression in small intestine. Lrig1-Apple is observed at the crypt base and in insterstial cells of Cajal, but is
 Supplemental Figure 1: Lrig1-Apple expression in small intestine. Lrig1-Apple is observed at the crypt base and in insterstial cells of Cajal, but is not co-expressed in DCLK1-positive tuft cells. Scale
Supplemental Figure 1: Lrig1-Apple expression in small intestine. Lrig1-Apple is observed at the crypt base and in insterstial cells of Cajal, but is not co-expressed in DCLK1-positive tuft cells. Scale
Supplemental Experimental Procedures
 Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Mitosis and the Cell Cycle
 Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Alan G. Casson FRCSC Professor of Surgery & VP Integrated Health Services University of Saskatchewan & Saskatoon Health Region
 Identification and Characterization of Stem-like Cells in Human Esophageal Adenocarcinoma and Normal Epithelial Cell Lines No disclosures / conflict of interest Alan G. Casson FRCSC Professor of Surgery
Identification and Characterization of Stem-like Cells in Human Esophageal Adenocarcinoma and Normal Epithelial Cell Lines No disclosures / conflict of interest Alan G. Casson FRCSC Professor of Surgery
Combinations of genetic mutations in the adult neural stem cell compartment determine brain tumour phenotypes
 Manuscript EMBO-2009-71812 Combinations of genetic mutations in the adult neural stem cell compartment determine brain tumour phenotypes Thomas S. Jacques, Alexander Swales, Monika J. Brzozowski, Nico
Manuscript EMBO-2009-71812 Combinations of genetic mutations in the adult neural stem cell compartment determine brain tumour phenotypes Thomas S. Jacques, Alexander Swales, Monika J. Brzozowski, Nico
Cancer Cells. It would take another 20 years and a revolution in the techniques of biological research to answer these questions.
 Cancer Cells Cancer, then, is a disease in which a single normal body cell undergoes a genetic transformation into a cancer cell. This cell and its descendants, proliferating across many years, produce
Cancer Cells Cancer, then, is a disease in which a single normal body cell undergoes a genetic transformation into a cancer cell. This cell and its descendants, proliferating across many years, produce
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION. Androgen deprivation therapy is the most used treatment of de novo or recurrent
 CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
General Structure of Digestive Tract
 Dr. Nabil Khouri General Structure of Digestive Tract Common Characteristics: Hollow tube composed of a lumen whose diameter varies. Surrounded by a wall made up of 4 principal layers: Mucosa Epithelial
Dr. Nabil Khouri General Structure of Digestive Tract Common Characteristics: Hollow tube composed of a lumen whose diameter varies. Surrounded by a wall made up of 4 principal layers: Mucosa Epithelial
Cutting to the chase: How pathogenic mutations cause Alzheimer s
 Cutting to the chase: How pathogenic mutations cause Alzheimer s The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published
Cutting to the chase: How pathogenic mutations cause Alzheimer s The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published
CONTRACTING ORGANIZATION: University of Michigan Ann Arbor, Michigan
 AD AWARD NUMBER: W81XWH-04-1-0471 TITLE: Chemo Resistance of Breast Cancer Stem Cells PRINCIPAL INVESTIGATOR: Max S. Wicha, M.D. CONTRACTING ORGANIZATION: University of Michigan Ann Arbor, Michigan 48109-1274
AD AWARD NUMBER: W81XWH-04-1-0471 TITLE: Chemo Resistance of Breast Cancer Stem Cells PRINCIPAL INVESTIGATOR: Max S. Wicha, M.D. CONTRACTING ORGANIZATION: University of Michigan Ann Arbor, Michigan 48109-1274
Test Bank for Robbins and Cotran Pathologic Basis of Disease 9th Edition by Kumar
 Link full download:https://getbooksolutions.com/download/test-bank-for-robbinsand-cotran-pathologic-basis-of-disease-9th-edition-by-kumar Test Bank for Robbins and Cotran Pathologic Basis of Disease 9th
Link full download:https://getbooksolutions.com/download/test-bank-for-robbinsand-cotran-pathologic-basis-of-disease-9th-edition-by-kumar Test Bank for Robbins and Cotran Pathologic Basis of Disease 9th
HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
MIT OpenCourseWare http://ocw.mit.edu HST.161 Molecular Biology and Genetics in Modern Medicine Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
Cell signalling pathways in the HSC niche. Dr. Abdullah Aljedai
 Cell signalling pathways in the HSC niche Dr. Abdullah Aljedai 31-10-2009 Learning objectives & resources 1- To introduce the concept of cell signaling process in the haemopoietic system. 2- To outline
Cell signalling pathways in the HSC niche Dr. Abdullah Aljedai 31-10-2009 Learning objectives & resources 1- To introduce the concept of cell signaling process in the haemopoietic system. 2- To outline
Inner ear development Nervous system development
 Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
Prof. R. V. Skibbens
 Prof. R. V. Skibbens September 8, 2017 BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell growth and division What are the goals? I Cell Cycle what is this? response to
Prof. R. V. Skibbens September 8, 2017 BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell growth and division What are the goals? I Cell Cycle what is this? response to
THE EFFECTS OF UNDEGRADED GLYCOSAMINOGLYCANS FROM MUCOPOLYSACCHARIDOSES ON OSTEOBLAST DIFFERENTIATION AND MINERALISATION IN VITRO
 THE EFFECTS OF UNDEGRADED GLYCOSAMINOGLYCANS FROM MUCOPOLYSACCHARIDOSES ON OSTEOBLAST DIFFERENTIATION AND MINERALISATION IN VITRO Nathan Rout-Pitt B.Sc. (Hons) Department of Paediatrics The University
THE EFFECTS OF UNDEGRADED GLYCOSAMINOGLYCANS FROM MUCOPOLYSACCHARIDOSES ON OSTEOBLAST DIFFERENTIATION AND MINERALISATION IN VITRO Nathan Rout-Pitt B.Sc. (Hons) Department of Paediatrics The University
Snai1 regulates cell lineage allocation and stem cell maintenance in the mouse intestinal epithelium
 Article Snai1 regulates cell lineage allocation and stem cell maintenance in the mouse intestinal epithelium Katja Horvay 1, Thierry Jardé 1, Franca Casagranda 2, Victoria M Perreau 2, Katharina Haigh
Article Snai1 regulates cell lineage allocation and stem cell maintenance in the mouse intestinal epithelium Katja Horvay 1, Thierry Jardé 1, Franca Casagranda 2, Victoria M Perreau 2, Katharina Haigh
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is
 is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is") ' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
Cancer and sornat ic evolution
 Chapter 1 Cancer and sornat ic evolution 1.1 What is cancer? The development and healthy life of a human being requires the cooperation of more than ten million cells for the good of the organism. This
Chapter 1 Cancer and sornat ic evolution 1.1 What is cancer? The development and healthy life of a human being requires the cooperation of more than ten million cells for the good of the organism. This
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Chapt 15: Molecular Genetics of Cell Cycle and Cancer
 Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
CHANTALE T. GUY, M5C. A Thesis. for the Degree. Doctor of Philosophy. McMaster University. (c) Copyright by Chantale T.
 Copyright by Chantale T.") ROLE AND MECHANISM OF ACTION OF TYROSINE KINASES IN MAMMARY TUMORIGENESIS By CHANTALE T. GUY, M5C. A Thesis Submitted to the School of Graduate Studies in Partial Fulfilment of the Requirements for the
ROLE AND MECHANISM OF ACTION OF TYROSINE KINASES IN MAMMARY TUMORIGENESIS By CHANTALE T. GUY, M5C. A Thesis Submitted to the School of Graduate Studies in Partial Fulfilment of the Requirements for the
The Biology and Genetics of Cells and Organisms The Biology of Cancer
 The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
Biochemistry of Cancer
 4/6/15 Biochemistry of Cancer Tumor Suppressor Genes 1 Tumor Suppressor Genes Recessive phenotype Oncogenes (viral or cellular) dominate a phenotype i.e. acfvafon of cancer like behaviors. Ras, Myc, Erb
4/6/15 Biochemistry of Cancer Tumor Suppressor Genes 1 Tumor Suppressor Genes Recessive phenotype Oncogenes (viral or cellular) dominate a phenotype i.e. acfvafon of cancer like behaviors. Ras, Myc, Erb
Cancer genetics
 Cancer genetics General information about tumorogenesis. Cancer induced by viruses. The role of somatic mutations in cancer production. Oncogenes and Tumor Suppressor Genes (TSG). Hereditary cancer. 1
Cancer genetics General information about tumorogenesis. Cancer induced by viruses. The role of somatic mutations in cancer production. Oncogenes and Tumor Suppressor Genes (TSG). Hereditary cancer. 1