MOLECULAR CELL BIOLOGY
|
|
|
- Leona Short
- 5 years ago
- Views:
Transcription
1 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 22 NERVE CELLS Copyright 2013 by W. H. Freeman and Company

2 Figure 22.1 Typical morphology of two types of mammalian neurons.
3 Typical morphology of neurons Neurons are distinguished by their elongated, asymmetric shape, by their highly localized proteins and organelles, and most of all by a set of proteins that controls the flow of ions across the plasma membrane (Figure 22-1). 1. The nucleus is found in a rounded part of the cell called the cell body. 2. Branching cell processes called dendrites are found at one end, and are the main structures where signals are received from other neurons via synapses. 3. When a neuron is first differentiating, the end of the cell opposite the dendrites undergoes dramatic outgrowth to form a long extended arm called the axon, which is essentially a transmission wire. Axons can be meters in length (in giraffe necks, for example), and are often partly covered with electrical insulation called the myelin sheath, which is made up of cells called glia. 4. The short, branched ends of the axon at the opposite end of the neuron from the dendrites are called the axon termini.
4
5 Nerve conduction 1. Axon 2. Synapase
6 Nerve conduction 1. Axon termini from one neuron are closely apposed to dendrites of another, at junctions called chemical synapses or simple synapses. The axon termini of the presynaptic cell contain many small vesicles, termed synaptic vesicles, each of which is filled with a single kind of small molecule known as a neurotransmitter. Arrival of an action potential at a terminus triggers exocytosis of a small number of synaptic vesicles, releasing their content of neurotransmitter molecules. 2. Binding of neurotransmitter triggers opening or closing of specific ion channels in the plasma membrane of post-synaptic cell dendrites, leading to changes in the membrane potential in this localized area of the postsynaptic cell. Generally these changes depolarize the postsynaptic membrane (making the potential less inside negative). The local depolarization, if large enough, triggers an action potential in the axon. Transmission is unidirectional, from the axon termini of the presynaptic cell to dendrites of the postsynaptic cell (Figure 22-3).
7

8 Action potential Experimental Figure The electric potential across the plasma membrane of living cells can be measured.

9 Experimental Figure 22.2 Recording of an axonal membrane potential over time reveals the amplitude and frequency of action potentials.
10 Action potential 1. The membrane voltage in a typical neuron, called the resting potential because it is the state when no signal is in transit, is established by Na+/K+ ion pumps in the plasma membrane. These are the same ion pumps used by other cells to generate a resting potential. Subsequent movement of K+ out of the cell through resting K+ channels results in a net negative charge inside the cell compared with the outside. The typical resting potential of a neuron is about -60mV (Experimental Figure 11-19). 2. The signals take the form of brief local voltage changes, from inside-negative to inside-positive, an event designated depolarization. A powerful surge of depolarizing voltage change, moving from one end of the neuron to the other, is called an action potential. 3. After the action potential passes a sector of a neuron, channel proteins and pumps restore the insidenegative resting potential (repolarization). The restoration process chases the action potential down the axon to the terminus, leaving the neuron ready to signal again (Experimental Figure 22-2).
11


12 Action potential by changes in Na + and K + permeability Figure 22.6 Depolarization of the plasma membrane due to opening of gated Na + channels.
13 Action potential by changes in Na + and K + permeability This resting membrane potential is generated by outward movement of K+ ions through open nongated K+ channels in the plasma membrane, and is driven by the K+ concentration gradient (cytosol > extracellular medium). The entry of Na+ ions into the cytosol from the medium is thermodynamically favored, driven both by the Na+ concentration gradient (extracellular > cytosol) and the inside-negative membrane potential. However, most Na+ channels in the plasma membrane are closed in resting cells, so little inward movement of Na+ ions can occur. In resting neurons, a type of nongated K+ channel is open part of the time, but the more numerous gated Na+ channels are closed. The movement of K+ ions outward establishes the inside-negative membrane potential characteristic of most cells. During an action potential, some of these Na+ channels open, allowing inward movement of Na+ ions, which depolarizes the membrane. Opening of gated Na+ channels permits an influx of sufficient Na+ ions to cause a reversal of the membrane potential. In the depolarized state, voltage-gated K+ channels open and subsequently repolarize the membrane. Note that the flows of ions are too small to have much effect on the overall concentration of either Na+ or K+ in the cytosol or exterior fluid (Figure 22-6).
14

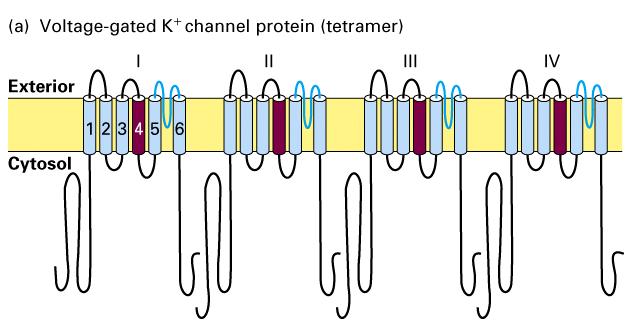
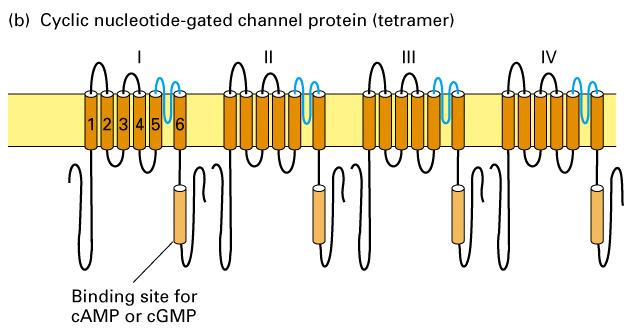
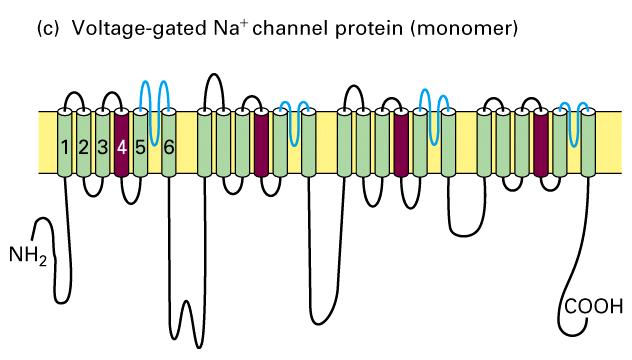
15 Voltage-gated Na + and K + channels. 2 Figure Schematic depictions of the secondary structures of voltage-gated K + and Na + channels.
16 Voltage-gated Na + and K + channels. 1. Voltage-gated K+ channels are composed of four identical subunits, each containing amino acids, and six membrane-spanning α helices, S1 S6. The N-terminus of each subunit, located in the cytosol and labeled N, forms a globular domain essential for inactivation of the open channel. The S5 and S6 helices and the P segment are homologous to those in nongated resting K+ channels, but each subunit contains four additional transmembrane α helices. One of these, S4, is the voltage-sensing α helix and is assisted in this role by forming a stable complex with helices S Voltage-gated Na+ channels are monomers containing amino acids organized into four transmembrane domains (I IV) that are similar to the subunits in voltage-gated K+ channels. The single hydrophobic channel-inactivating segment is located in the cytosol between domains III and IV. Most voltage-gated ion channels also contain regulatory (β) subunits, which are not depicted here (Figure 22-10).
17

18 Figure 22.7 Operational model of the voltage-gated Na + channel.
19 Voltage-gated Na + channel As just discussed, voltage-gated Na+ channels are closed in resting neurons. A small depolarization of the membrane causes a conformational change in these channel proteins that opens a gate on the cytosolic surface of the pore, permitting Na+ ions to pass through the pore into the cell. The greater the initial membrane depolarization, the more voltage-gated Na+ channels that open and the more Na+ ions enter. As Na+ ions flow inward through opened channels, the excess positive charges on the cytosolic face and negative charges on the exoplasmic face diffuse a short distance away from the initial site of depolarization (Figure 22-7). 1. A small depolarization of the membrane triggers movement of positively charged voltage-sensing α helices toward the exoplasmic surface, causing a conformational change in the gate that opens the channel and allows ion flow. 2. After about 1 millisecond, further Na+ influx is prevented by movement of the cytosol-facing channel-inactivating segment into the open channel. As long as the membrane remains depolarized, the channel-inactivating segment remains in the channel opening; during this refractory period, the channel is inactivated and cannot be reopened. A few milliseconds after the insidenegative resting potential is reestablished, the channel-inactivating segment swings away from the pore and the channel returns to the closed resting state, once again able to be opened by depolarization.
20

21 Figure 22.9 Unidirectional conduction of an action potential due to transient inactivation of voltagegated Na + channels. 1. Unidirectional conduction 2. Refractory period
22 Voltage-gated Na + channel As noted earlier, during the refractory period voltage-gated Na+ channels are inactivated for several milliseconds. Such previously opened channels cannot open during this period even if the membrane is depolarized due to passive spread. As illustrated in Figure 22-9, the inability of Na+ channels to reopen during the refractory period ensures that action potentials are propagated only in one direction, from the axon hillock where they originate to the axon terminus. This property of the Na+ channels also limits the number of action potentials per second that a neuron can conduct. Reopening of Na+ channels upstream of an action potential (i.e., closer to the cell body) also is delayed by the membrane hyperpolarization that results from opening of voltage-gated K+ channels (Figure 22-9).
23

24 Voltage-gated K + channel 3 Experimental Figure Experiments with a mutant K + channel lacking the N-terminal globular domains support the ball-and-chain inactivation model. Inactivating segment
25 Voltage-gated K + channel The experimental results shown in Figure demonstrate that inactivation of K+ channels depends on the ball domains, occurs after channel opening, and does not require the ball domains to be covalently linked to the channel protein (Experimental Figure 22-12). 1. In the resting state, the positively charged globular balls at the N-termini of the four subunits in a voltage-gated K+ channel are free in the cytosol. 2. Several milliseconds after the channel is opened by depolarization, one ball moves through an opening ( lateral window ) between two of the subunits and binds in a hydrophobic pocket in the pore s central cavity, blocking the flow of K+ ions. 3. After a few milliseconds, the ball is displaced from the pore, and the protein reverts to the closed, resting state. The ball-and-chain domains in K+ channels are functionally equivalent to the channelinactivating segment in Na+ channels. Structure of a voltage-sensitive K + channel Typically, vertebrate voltage-gated K+ channels are tetramers of four identical subunits arranged as a ring, each contributing to the wall of the trans-membrane K+ pore. Each subunit is composed of six membrane spanning hydrophobic α-helical sequences. Specific domains of the channel subunits have been identified that are responsible for voltage-sensing and converting between the open and closed conformations of the channel. This type of gating is mediated by a voltage-sensing domain that consists of the S4 alpha helix that contains 6 7 positive charges. Changes in membrane potential cause this alpha helix to move in the lipid bilayer. This movement in turn results a conformational change in the adjacent S5 S6 helices that form the channel pore and cause this pore to open or close (Figure 22-11).
26


27 Figure Conduction of action potentials in myelinated axons.
28 Conduction of action potentials in myelinated axons The axonal membrane is in direct contact with the extracellular fluid only at the nodes. Moreover, all the voltage-gated Na channels and all the Na/K pumps, which maintain the ionic gradients in the axon, are located in the nodes. As a consequence of this localization, the inward movement of Na ions that generates the action potential can occur only at the myelin-free nodes. The excess cytosolic positive ions generated at a node during the membrane depolarization associated with an action potential spread passively through the axonal cytosol to the next node with very little loss or attenuation, since they cannot cross the myelinated axonal membrane. This causes a depolarization at one node to spread rapidly to the next node, permitting, in effect, the action potential to jump from node to node. This phenomenon explains why the conduction velocity of myelinated neurons is about the same as that of much larger diameter unmyelinated neurons (Figure 22-13).
29

30 Figure Three types of glial cells.

31 Figure Formation and structure of a myelin sheath in the peripheral nervous system.
32 Three types of glial cells and structure of a myelin sheath 1. Figure shows three main types of glial cells present in the nervous system, two of which produce myelin sheaths: oligodendrocytes make sheaths for the central nervous system (CNS), and Schwann cells make them for the peripheral nervous system (PNS). Astrocytes, also shown in the figure, facilitate synapse formation and communication between neurons. 2. The myelin sheath is a stack of specialized plasma membrane sheets produced by a glial cell that wraps itself around the axon. In the peripheral nervous system, these glial cells are called Schwann cells. In the central nervous system, they are called oligodendrocytes. Figure illustrates the formation and basic structure of a myelin sheath, which contains both membrane and cytosolic components (Figure 22-15).
33

34 2. Synapses 4

35 Figure 22.3 A Chemical Synapse

36 Figure Synaptic vesicles in the axon terminus near the region where neurotransmitter is released.


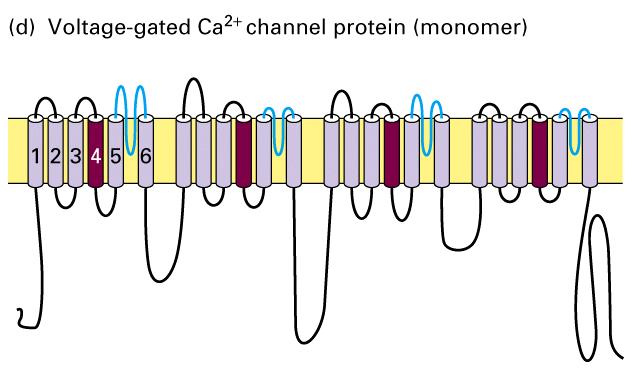
37 Chemical and electric synapses
38 Synapses 1. Synapses generally transmit signals in only one direction: an axon terminal from thepresynaptic cell sends signals that are picked up by the postsynaptic cell. There are two general types of synapse: the relatively rare electric synapse, discussed later, and, the chemical synapse, illustrated in Figure At chemical synapses, impulses are transmitted by the release of neurotransmitters from the axon terminal of the presynaptic cell into the synaptic cleft. Their subsequent binding to specific receptors on the postsynaptic cell causes a change in the ion permeability and thus the potential of the postsynaptic plasma membrane. In an electric synapse, ions move directly from one neuron to another via gap junctions. The membrane depolarization associated with an action potential in the presynaptic cell passes through the gap junctions, leading to a depolarization, and thus an action potential, in the postsynaptic cell. 2. In both cases, the presynaptic neuron was stimulated and the membrane potential was measured in both the presynaptic and postsynaptic cells. Signals are transmitted across an electric synapse within a few microseconds because ions flow directly from the presynaptic cell to the postsynaptic cell through gap junctions. In contrast, signal transmission across a chemical synapse is delayed about 0.5 ms the time required for secretion and diffusion of neurotransmitter and the response of the postsynaptic cell to it (Figure 22-16).
39
40 Figure Cycling of neurotransmitters and of synaptic vesicles in axon termini.
41 Cycling of neurotransmitters and of synaptic vesicles in axon termini Two features critical to synapse function differ from other secretory pathways: (a) secretion is tightly coupled to arrival of an action potential at the axon terminus, and (b) synaptic vesicles are recycled locally to the axon terminus after fusion with the plasma membrane. Figure shows the entire cycle whereby synaptic vesicles are filled with neurotransmitter, release their contents, and are recycled. Depolarization of the plasma membrane cannot, by itself, cause synaptic vesicles to fuse with the plasma membrane. In order to trigger vesicle fusion, an action potential must be converted into a chemical signal namely, a localized rise in the cytosolic Ca 2+ concentration. The transducers of the electric signals are voltagegated Ca 2+ channels localized to the region of the plasma membrane adjacent to the synaptic vesicles. The membrane depolarization due to arrival of an action potential opens these channels, permitting an influx of Ca 2+ ions from the extracellular medium into the axon terminal. This ion flux raises the local cytosolic Ca 2+ concentration near the synaptic vesicles from <0.1 um, characteristic of the resting state, to um. Binding of Ca 2+ ions to proteins that connect the synaptic vesicle with the plasma membrane induces membrane fusion and thus exocytosis of the neurotransmitter. The subsequent export of extra Ca 2+ ions by ATP-powered Ca 2+ pumps in the plasma membrane rapidly lowers the cytosolic Ca 2+ level to that of the resting state, enabling the axon terminus to respond to the arrival of another action potential (Figure 22-19).
42
43
44 Structures of voltage-gated Ca 2+ channels The Shaker K + channel and most other voltage-gated K+ channels that have been identified are tetrameric proteins composed of four identical subunits arranged in the membrane around a central pore. Each subunit is constructed of six membrane-spanning α helixes, designated S1 S6, and a P segment. Voltage-gated Na + channels and Ca 2+ channels are monomeric proteins organized into four homologous domains, I IV. Each of these domains is similar to a subunit of a voltage-gated K + channel. However, in contrast to voltage-gated K + channels, which have four channel inactivating segments, the monomeric voltage-gated channels have a single channel-inactivating segment. Except for this minor structural difference and their varying ion permeabilities, all voltage-gated ion channels are thought to function in a similar manner and to have evolved from a monomeric ancestral channel protein that contained six transmembrane α helices.
45
46 Figure Structures of several small molecules that function as neurotransmitters. 5
47 UN Figure 22.1
48 Neurotransmitters Numerous small molecules function as neurotransmitters at various synapses. With the exception of acetylcholine, the neurotransmitters shown in Figure are amino acids or derivatives of amino acids. Nucleotides such as ATP and the corresponding nucleosides, which lack phosphate groups, also function as neurotransmitters. Each neuron generally produces just one type of neurotransmitter (Figure 22-18). For example, acetylcholine is synthesized from acetyl coenzyme A (acetyl CoA), an intermediate in the degradation of glucose and fatty acids, and choline in a reaction catalyzed by choline acetyltransferase (UN Figure 22-1).
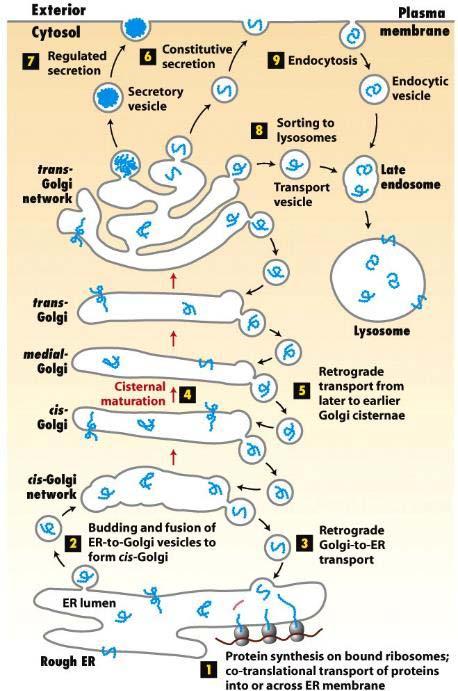
49

50 Figure Sequential activation of gated ion channels at a neuromuscular junction.


51 Figure Three-dimensional structure of the nicotinic acetylcholine receptor.
52 Neuromuscular junction 1. The nicotinic acetylcholine receptor, which is expressed in muscle cells, is a ligand-gated channel that admits both K and Na. The simultaneous increase in permeability to Na and K ions following binding of acetylcholine produces a net depolarization to about 15 mv from the muscle resting potential of 85 to 90 mv. As shown in Figure 22-21, this localized depolarization of the muscle plasma membrane triggers opening of voltage-gated Na channels, leading to generation and conduction of an action potential in the muscle cell surface membrane by the same mechanisms described previously for neurons. When the membrane depolarization reaches T tubules, specialized invaginations of the plasma membrane, it affects Ca2 channels in the plasma membrane apparently without causing them to open. Somehow this causes opening of adjacent Ca2-release channels in the sarcoplasmic reticulum membrane. The subsequent flow of stored Ca2 ions from the sarcoplasmic reticulum into the cytosol raises the cytosolic Ca2 concentration sufficiently to induce muscle contraction (Figure 22-21). 2. The acetylcholine receptor from skeletal muscle is a pentameric protein with a subunit composition of α 2 βγδ. The α, β, γ and δ subunits have considerable sequence homology; on average, about percent of the residues in any two subunits are similar. The complete receptor has fivefold symmetry, and the actual cation channel is a tapered central pore lined by homologous segments from each of the five subunits (Figure 22-22).
53 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 14 Vesicular Traffic, Secretion, and Endocytosis Copyright 2013 by W. H. Freeman and Company

54 Figure 14.1 Overview of the secretory and endocytic pathways of protein sorting.




55 The Nobel Prize in Physiology or Medicine 2013: discoveries of machinery regulating vesicle traffic, a major transport system in our cells
56 The secretory and endocytic pathways of protein sorting 1. The 2013 Nobel Prize honours three scientists who have solved the mystery of how the cell organizes its transport system. Each cell is a factory that produces and exports molecules. Randy Schekman discovered a set of genes that were required for vesicle traffic. James Rothman unravelled protein machinery that allows vesicles to fuse with their targets to permit transfer of cargo. Thomas Südhof revealed how signals instruct vesicles to release their cargo with precision. Through their discoveries, Rothman, Schekman and Südhof have revealed the exquisitely precise control system for the transport and delivery of cellular cargo. ( Nature (1977) 269(5631): ) 2. Figure 14-1 outlines the major routes for protein trafficking in the secretory pathway. 1 Once newly synthesized proteins are incorporated into the ER lumen or membrane, 2 they can be packaged into anterograde (forward-moving) transport vesicles. These vesicles fuse with each other to form a flattened membrane-bounded compartment known as the cis-golgi cisterna. 3 Certain proteins, mainly ER-localized proteins, are retrieved from the cis-golgi to the ER via a different set of retrograde (backward-moving) transport vesicles. 4 A new cis-golgi cisterna with its cargo of proteins physically moves from the cis position (nearest the ER) to the trans position (farthest from the ER), successively becoming first a medial-golgi cisterna and then a trans-golgi cisterna. 5 Proteins in the secretory pathway that are destined for compartments other than the ER or Golgi eventually reach a complex network of membranes and vesicles termed the trans-golgi network (TGN). From this major branch point in the secretory pathway, a protein can be loaded into one of at least three different kinds of vesicles. 6 After budding from the trans-golgi network, the first type of vesicle immediately moves to and fuses with the plasma membrane, releasing its contents by exocytosis. In all cell types, at least some proteins are loaded into such vesicles and secreted continuously in this manner. 7 The second type of vesicle to bud from the trans-golgi network, known as secretory vesicles, are stored inside the cell until a signal for exocytosis causes release of their contents at the plasma membrane. 8 The third type of vesicle that buds from the trans-golgi network is directed to the lysosome, an organelle responsible for the intracellular degradation of macromolecules, and to lysosome-like storage organelles in certain cells. Secretory proteins destined for lysosomes first are transported by vesicles from the trans-golgi network to a compartment usually called the late endosome; proteins then are transferred to the lysosome by a mechanism that is not well understood but may involve direct fusion of the endosome with the lysosomal membrane. 9 The endosome also functions in the endocytic pathway in which vesicles bud from the plasma membrane bringing membrane proteins and their bound ligands into the cell. After being internalized by endocytosis, some proteins are transported to lysosomes, while others are recycled back to the cell surface.
57

58 Experimental Figure Electron micrograph of the Golgi complex in an exocrine pancreatic cell reveals secretory and retrograde transport vesicles.
59 Secretory and retrograde transport vesicles At one time it was thought that small transport vesicles carry secretory proteins from the cis- to the medial-golgi and from the medial- to the trans-golgi. Indeed, electron microscopy reveals many small vesicles associated with the Golgi complex that move proteins from one Golgi compartment to another. However, these vesicles most likely mediate retrograde transport, retrieving ER or Golgi enzymes from a later compartment and transporting them to an earlier compartment in the secretory pathway. In this way enzymes that modify secretory proteins come to be localized in the correct compartment (Experimental Figure 14-15).
60

61 Experimental Figure 14.4 Phenotypes of yeast sec mutants identified five stages in the secretory pathway.
62 Five stages in the secretory pathway A large number of yeast mutants initially were identified based on their ability to secrete proteins at one temperature and inability to do so at a higher, nonpermissive temperature. When these temperaturesensitive secretion (sec) mutants are transferred from the lower to the higher temperature, they accumulate secreted proteins at the point in the pathway blocked by the mutation. Analysis of such mutants identified five classes (A E) characterized by protein accumulation in the cytosol, rough ER, small vesicles taking proteins from the ER to the Golgi complex, Golgi cisternae, or constitutive secretory vesicles. To determine the order of the steps in the pathway, researchers analyzed double sec mutants. For instance, when yeast cells contain mutations in both class B and class D functions, proteins accumulate in the rough ER, not in the Golgi cisternae. Since proteins accumulate at the earliest blocked step, this finding shows that class B mutations must act at an earlier point in the secretory pathway than class D mutations do. These studies confirmed that as a secreted protein is synthesized and processed it moves sequentially from the cytosol rough ER ER-to-Golgi transport vesicles Golgi cisternae secretory vesicles and finally is exocytosed (Experimental Figure 14-4).
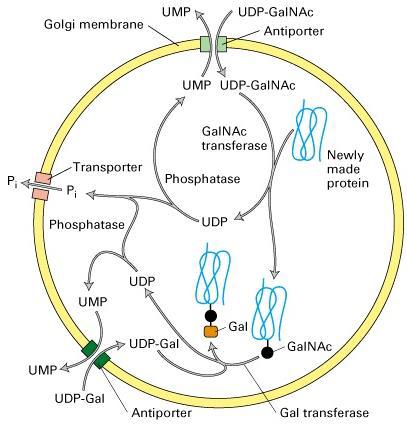
63

64 Figure Molecular Biology of the Cell ( Garland Science 2008)
65 The functional compartmentalization of the Golgi apparatus Proteins exported from the ER enter the first of the Golgi processing compartments (the cis Golgi compartment), after having passed through the cis Golgi network. They then move to the next compartment (the medial compartment, consisting of the central cisternae of the stack) and finally to the trans compartment, where glycosylation is completed. The lumen of the trans compartment is thought to be continuous with the trans Golgi network, where proteins are segregated into different transport packages and dispatched to their final destinations the plasma membrane, lysosomes, or secretory vesicles. he oligosaccharide processing steps occur in a correspondingly organized sequence in the Golgi stack, with each cisterna containing a characteristic abundance of processing enzymes. Proteins are modified in successive stages as they move from cisterna to cisterna across the stack, so that the stack forms a multistage processing unit. This compartmentalization might seem unnecessary, since each oligosaccharide processing enzyme can accept a glycoprotein as a substrate only after it has been properly processed by the preceding enzyme. Nonetheless, it is clear that processing occurs in a spatial as well as a biochemical sequence: enzymes catalyzing early processing steps are concentrated in the cisternae toward the cis face of the Golgi stack, whereas enzymes catalyzing later processing steps are concentrated in the cisternae toward the trans face. The functional compartmentalization of the Golgi apparatus is summarized in diagrammatic form in Figure (
66

67 Figure Processing of N-linked oligosaccharide chains on glycoproteins within cis-, medial-, and trans-golgi cisternae in vertebrate cells.
68 N-glycosylation in Glogi 1. A second way to follow the transport of secretory proteins takes advantage of modifications to their carbohydrate side chains that occur at different stages of the secretory pathway. To understand this approach, recall that many secretory proteins leaving the ER contain one or more copies of the N- linked oligosaccharide Man8(GlcNAc)2, which are synthesized and attached to secretory proteins in the ER. 2. As a protein moves through the Golgi complex, different enzymes localized to the cis-, medial-, and trans-golgi cisternae catalyze an ordered series of reactions to these core Man8(GlcNAc)2 chains. After removal of three mannose residues in the cis-golgi (step 1), the protein moves by cisternal progression to the medial-golgi. Here, three GlcNAc residues are added (steps 2 and 4), two more mannose residues are removed (step 3), and a single fucose is added (step 5). Processing is completed in the trans-golgi by addition of three galactose residues (step 6) and finally by linkage of an N- acetylneuraminic acid residue to each of the galactose residues (step 7). Specific transferase enzymes add sugars to the oligosaccharide, one at a time, from sugar nucleotide precursors imported from the cytosol. This pathway represents the Golgi processing events for a typical mammalian glycoprotein. Variations in the structure of N-linked oligosaccharides can result from differences in processing steps in the Golgi (Figure 14-14).
69
70 O-glycosylation in Golgi
71 O-glycosylation in Golgi Biosynthesis of the O-linked oligosaccharide in glycophorin and similar glycoproteins begins with transfer of N-acetylgalactosamine (GalNAc) from UDP N-acetylgalactosamine to the hydroxyl group of a serine or threonine residue in the protein. This reaction is catalyzed by a GalNAc transferase that is localized to the rough ER or the cis-golgi network. After the protein has moved to the trans-golgi vesicles, a galactose residue is added to the N-acetylgalactosamine by a specific trans-golgi galactosyltransferase. In vertebrate cells biosynthesis of typical O-linked oligosaccharides is completed by the addition of two negatively charged N-acetylneuraminic acid (also called sialic acid) residues from a CMP precursor; these reactions also occur in the trans-golgi or the trans-golgi network. 1 2 All known glycosyltransferases that act on secretory proteins are integral membrane proteins with active sites facing the lumen of the organelle. Each glycosyltransferase is specific for both the donor sugar nucleotide and the acceptor molecule. The immediate precursors used in the biosynthesis of oligosaccharides are nucleoside diphosphate or monophosphate sugars.
72
73
74 All the sugar nucleotides used in the synthesis of glycoproteins and glycolipids are made in the cytosol from nucleoside triphosphates and sugar phosphates. Specific antiport proteins in the membranes of the rough ER and Golgi cisternae catalyze the import of the sugar nucleotides into the lumina of these organelles and the export of free nucleotides (UMP, CMP, and GMP) generated within the organelles. The one-for-one exchanges catalyzed by these antiporters maintain the concentration of sugar nucleotides in the rough ER and Golgi lumina at a constant level, a requirement for oligosaccharide synthesis.
75
")
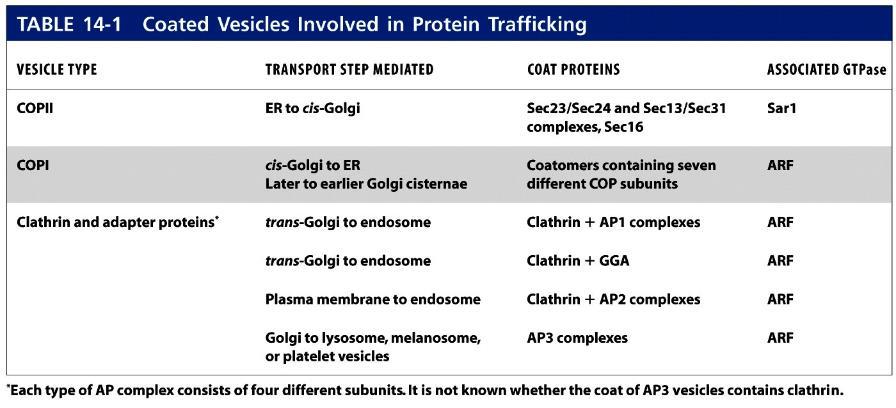
76 Figure 12-7 Molecular Biology of the Cell ( Garland Science 2008) 2
77 Molecular mechanisms of vesicular traffic Small membrane-bounded vesicles that transport proteins from one organelle to another are common elements in the secretory and endocytic pathways. These vesicles bud from the membrane of a particular parent (donor) organelle and fuse with the membrane of a particular target (destination) organelle (Figure 12-7 Molecular Biology of the Cell).

78 Figure 14.6 Overview of vesicle budding and fusion with a target membrane.
79 Vesicle budding and fusion 1. The budding of vesicles from their parent membrane is driven by the polymerization of soluble protein complexes onto the membrane to form a proteinaceous vesicle coat (a). Interactions between the cytosolic portions of integral membrane proteins and the vesicle coat gather the appropriate cargo proteins into the forming vesicle. Thus the coat not only adds curvature to the membrane to form a vesicle but also acts as the filter to determine which proteins are admitted into the vesicle. 2. The integral membrane proteins in a budding vesicle include v-snares, which are crucial to eventual fusion of the vesicle with the correct target membrane. Shortly after formation of a vesicle is completed, the coat is shed exposing its v-snare proteins. The specific joining of v-snares in the vesicle membrane with cognate t-snares in the target membrane brings the membranes into close apposition, allowing the two bilayers to fuse (b) (Figure 14-6).
80
81
")
82 Figure 13-4 Molecular Biology of the Cell ( Garland Science 2008)

83 Experimental Figure 14.9 Coated vesicles accumulate during in vitro budding reactions in the presence of a nonhydrolyzable analog of GTP.
84 Three types of coated vesicles 1. Three types of coated vesicles have been characterized, each with a different type of protein coat and each formed by reversible polymerization of a distinct set of protein subunits. Each type of vesicle, named for its primary coat proteins, transports cargo proteins from particular parent organelles to particular destination organelles (Table 14-1): COPII vesicles transport proteins from the rough ER to the Golgi. COPI vesicles mainly transport proteins in the retrograde direction between Golgi cisternae and from the cis- Golgi back to the rough ER. Clathrin vesicles transport proteins from the plasma membrane (cell surface) and the trans-golgi network to late endosomes. 2. Electron micrographs of in vitro budding reactions often reveal structures that exhibit discrete regions of the parent membrane bearing a dense coat accompanied by the curvature characteristic of a completed vesicle. Such structures, usually called vesicle buds, appear to be intermediates that are visible after the coat has begun to polymerize but before the completed vesicle pinches off from the parent membrane (Figure 13-4 Molecular Biology of the Cell, Experimental Figure 14-9).
85
")
86 Figure 13-5 Molecular Biology of the Cell ( Garland Science 2008)
87 Protein trafficking As cargo proteins move from the cis face to the trans face of the Golgi complex by cisternal progression, modifications to their oligosaccharide chains are carried out by Golgi resident enzymes. The retrograde trafficking of COPI vesicles from later to earlier Golgi compartments maintains sufficient levels of these carbohydrate-modifying enzymes in their functional compartments. Eventually, properly processed cargo proteins reach the trans-golgi network, the most-distal Golgi compartment. Here they are sorted into vesicles for delivery to their final destination (Figure 13-5 Molecular Biology of the Cell).
88


89 Figure 14.8 Model for the role of Sar1 in the assembly and disassembly of COPII coats. Figure Three-dimensional structure of the ternary complex comprising the COPII coat proteins Sec23 and Sec24 and Sar1 GTP.
90 Assembly and disassembly of COPII coats 1. Although most of the coat proteins differ considerably from one type of vesicle to another, the coats of all three vesicles contain a small GTP-binding protein that acts as a regulatory subunit to control coat assembly. For both COPI and clathrin vesicles, this GTP-binding protein is known as ARF. A different but related GTP-binding protein known as Sar1 is present in the coat of COPII vesicles. ARF and Sar1 proteins, like Ras, belong to the GTPase superfamily of switch proteins that cycle between inactive GDP-bound and active GTP-bound forms. The cycle of GTP binding and hydrolysis by ARF and Sar1 are thought to control the initiation of coat assembly as schematically depicted for the assembly of COPII vesicles (Figure 14-8) First, an ER membrane protein known as Sec12 catalyzes release of GDP from cytosolic Sar1 GDP and binding of GTP. The Sec12 guanine nucleotide exchange factor apparently receives and integrates multiple, as yet unknown signals, probably including the presence of cargo proteins in the ER membrane that are ready to be transported. Binding of GTP causes a conformational change in Sar1 that exposes its hydrophobic N-terminus, which then becomes embedded in the phospholipid bilayer and tethers Sar1 GTP to the ER membrane. The membrane-attached Sar1 GTP drives polymerization of cytosolic complexes of COPII subunits on the membrane, eventually leading to formation of vesicle buds. Once COPII vesicles are released from the donor membrane, the Sar1 GTPase activity hydrolyzes Sar1 GTP in the vesicle membrane to Sar1 GDP with the assistance of one of the coat subunits. This hydrolysis triggers disassembly of the COPII coat. Thus Sar1 couples a cycle of GTP binding and hydrolysis to the formation and then dissociation of the COPII coat. 2. As described previously, formation of COPII vesicles is triggered when Sec12, a guanine nucleotide exchange factor, catalyzes the exchange of bound GDP for GTP on Sar1. This exchange induces binding of Sar1 to the ER membrane followed by binding of a complex of Sec23 and Sec24 proteins. The resulting ternary complex formed between Sar1 GTP, Sec23, and Sec24 (Figure 14-12).
91


92 Figure Model for docking and fusion of transport vesicles with their target membranes.
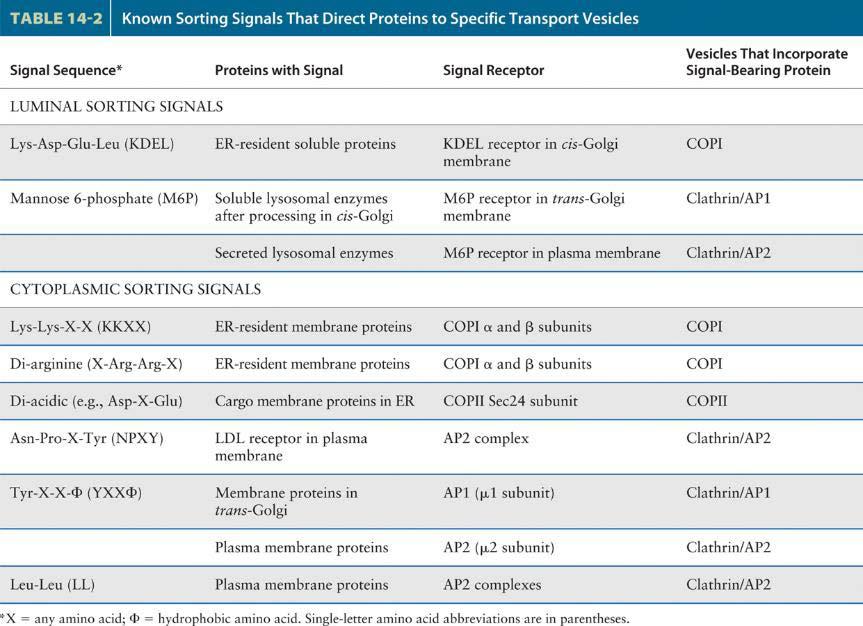
93 Docking and fusion of transport vesicles 1. A second set of small GTP-binding proteins, known as Rab proteins, participate in the targeting of vesicles to the appropriate target membrane. Like Sar1 and ARF, Rab proteins belong to the GTPase superfamily of switch proteins. Conversion of cytosolic Rab GDP to Rab GTP, catalyzed by a specific guanine nucleotide exchange factor, induces a conformational change in Rab that enables it to interact with a surface protein on a particular transport vesicle and insert its isoprenoid anchor into the vesicle membrane. Once Rab GTP is tethered to the vesicle surface, it is thought to interact with one of a number of different large proteins, known as Rab effectors, attached to the target membrane. Binding of Rab GTP to a Rab effector docks the vesicle on an appropriate target membrane (step 1). After vesicle fusion occurs, the GTP bound to the Rab protein is hydrolyzed to GDP, triggering the release of Rab GDP, which then can undergo another cycle of GDP-GTP exchange, binding, and hydrolysis. As noted previously, shortly after a vesicle buds off from the donor membrane, the vesicle coat disassembles to uncover a vesicle-specific membrane protein, a v-snare. Likewise, each type of target membrane in a cell contains t-snare membrane proteins. After Rab-mediated docking of a vesicle on its target (destination) membrane, the interaction of cognate SNAREs brings the two membranes close enough together that they can fuse. One of the best-understood examples of SNAREmediated fusion occurs during exocytosis of secreted proteins (steps 2 and 3). 2. In this case, the v-snare, known as VAMP (vesicle-associated membrane protein), is incorporated into secretory vesicles as they bud from the trans-golgi network. The t-snares are syntaxin, an integral membrane protein in the plasma membrane, and SNAP-25, which is attached to the plasma membrane by a hydrophobic lipid anchor in the middle of the protein (Figure 14-10).
94

95 Figure Vescle-mediated protein trafficking between the ER and cis-golgi. 3
96 Protein trafficking between the ER and cis-golgi Recall that anterograde transport from the ER to Golgi, the first step in the secretory pathway, is mediated by COPII vesicles, whereas the reverse retrograde transport from the cis-golgi to the ER is mediated by COPI vesicles. This retrograde vesicle transport serves to retrieve v-snare proteins and the membrane itself back to the ER to provide the necessary material for additional rounds of vesicle budding from the ER. COPI-mediated retrograde transport also retrieves missorted ERresident proteins from the cis-golgi to correct sorting mistakes. Proteins that have been correctly delivered to the Golgi advance through successive compartments of the Golgi by cisternal progression (Figure 14-11).
97

98 Figure Role of the KDEL receptor in retrieval of ER-resident luminal proteins from the Golgi.
99 Retrieval of ER-resident luminal proteins from the Golgi 1. Most soluble ER-resident proteins carry a Lys-Asp-Glu-Leu (KDEL in the one-letter code) sequence at their C-terminus. Several experiments demonstrated that this KDEL sorting signal is both necessary and sufficient for retention in the ER. For instance, when a mutant protein disulfide isomerase lacking these four residues is synthesized in cultured fibroblasts, the protein is secreted. Moreover, if a protein that normally is secreted is altered so that it contains the KDEL signal at its C-terminus, the protein is retained in the ER. 2. The KDEL sorting signal is recognized and bound by the KDEL receptor, a transmembrane protein found primarily on small transport vesicles shuttling between the ER and the cis-golgi and on the cis-golgi reticulum. In addition, soluble ER-resident proteins that carry the KDEL signal have oligosaccharide chains with modifications that are catalyzed by enzymes found only in the cis-golgi or cis-golgi reticulum; thus at some time these proteins must have left the ER and been transported at least as far as the cis-golgi network. These findings indicate that the KDEL receptor acts mainly to retrieve soluble proteins containing the KDEL sorting signal that have escaped to the cis-golgi network and return them to the ER. 3. This retrieval system prevents depletion of ER luminal proteins such as those needed for proper folding of newly made secretory proteins. The binding affinity of the KDEL receptor is very sensitive to ph. The small difference in the ph of the ER and Golgi favors binding of KDEL-bearing proteins to the receptor in Golgi-derived vesicles and their release in the ER (Figure 14-13).
100
101
102
103 Sorting signals In order for transport vesicles to move specific proteins from one compartment to the next, vesicle buds must be able to discriminate among potential membrane and soluble cargo proteins, accepting only those cargo proteins that should advance to the next compartment and excluding those that should remain as residents in the donor compartment. In addition to sculpting the curvature of a donor membrane, the vesicle coat also functions in selecting specific proteins as cargo. The primary mechanism by which the vesicle coat selects cargo molecules is by directly binding to specific sequences, or sorting signals, in the cytosolic portion of membrane cargo proteins. The polymerized coat thus acts as an affinity matrix to cluster selected membrane cargo proteins into forming vesicle buds. Soluble proteins within the lumen of parent organelles can in turn be selected by binding to the luminal domains of certain membrane cargo proteins, which act as receptors for luminal cargo proteins. The properties of several known sorting signals in membrane and soluble proteins are summarized in Table 14-2.
104

105 Table 13-1 Molecular Biology of the Cell ( Garland Science 2008)
106 Rab proteins The Rab family of proteins is a member of the Ras superfamily of monomeric G proteins. There are approximately 70 different Rabs that have been identified in humans thus far. They are mostly involved in vesicle trafficking. Their complexity can be understood if thought of as address labels for vesicle trafficking, defining the identity and routing of vesicles (Table 13-1 Molecular Biology of the Cell).
107

108 Figure Vesicle-mediated protein trafficking from the trans-golgi network. 4
109 Protein trafficking from the trans-golgi network COPI vesicles mediate retrograde transport within the Golgi (1). Proteins that function in the lumen or in the membrane of the lysosome are first transported from the trans-golgi network via clathrin-coated vesicles (3); after uncoating, these vesicles fuse with late endosomes, which deliver their contents to the lysosome. The coat on most clathrin vesicles contains additional proteins (AP comlexes) not indicated here. Some vesicles from the trans-golgi carrying cargo destined for the lysosome fuse with the lysosome directly (2), bypassing the endosome. These vesicles are coated with a type of AP comlexe; it is unknown whether these vesicles also contain clathrin. The coat proteins surrounding constitutive (4) and regulated (5) secretory vesicles are not yet characterized; these vesicles carry secreted proteins and plasma-membrane proteins from the trans-golgi network to the cell surface (Figure 14-17).
110

111 Figure Cycling of neurotransmitters and of synaptic vesicles in axon termini.

112 Figure Synaptotagmin-mediated fusion of synaptic vesicles with the plasma membrane.
113 Cycling of neurotransmitters Neurotransmitters are released by exocytosis, a process in which neurotransmitter-filled synaptic vesicles fuse with the axonal membrane, releasing their contents into the synaptic cleft. The exocytosis of neurotransmitters from synaptic vesicles involves vesicle-targeting and fusion events similar to those that occur during the intracellular transport of secreted and plasma-membrane proteins (Figure 22-19). Synaptotagmin-mediated fusion of synaptic vesicles with the plasma membrane The signal that triggers exocytosis of docked synaptic vesicles is a rise in the Ca 2+ concentration in the cytosol near vesicles from < 0.1 um, characteristic of resting cells, to um following arrival of an action potential in stimulated cells. A Ca 2+ -binding protein called synaptotagmin, located in the membrane of synaptic vesicles, is thought to be a key component of the vesicle fusion machinery that triggers exocytosis in response to Ca 2+. Fusion of synaptic vesicles with the plasma membrane of axon terminals depends on the same proteins that mediate membrane fusion of other regulated secretory vesicles. The principal v-snare in synaptic vesicles (VAMP) tightly binds syntaxin and SNAP-25, the principal t-snares in the plasma membrane of axon terminals, to form four-helix SNARE complexes. After fusion, SNAP proteins and NSF within the axon terminal promote disassociation of VAMP from t-snares, as in the fusion of secretory vesicles depicted previously. In molecular biology, complexin (also known as synaphin), a eukaryotic specific protein, is a cytoplasmic neuronal protein which binds to the SNARE protein complex (SNAREpin) with a high affinity. In the presence of Ca2+, the transport vesicle protein synaptotagmin displaces complexin, allowing the SNARE protein complex to bind the transport vesicle to the presynaptic membrane. Complexin acts as both an inhibitor and a facilitator of synaptic vesicle fusion and neurotransmitter release (Figure 22-20).
114



115 Figure Molecular Biology of the Cell ( Garland Science 2008) Figure Proteolytic processing of proproteins in the constitutive and regulated secretion pathways.
116 The constitutive and regulated secretory pathways All cells require this constitutive secretory pathway. Specialized secretory cells, however, have a second secretory pathway in which soluble proteins and other substances are initially stored in secretory vesicles for later release. This is the regulated secretory pathway, found mainly in cells specialized for secreting products rapidly on demand such as hormones, neurotransmitters, or digestive enzymes (Figure 13-63). The two pathways diverge in the trans Golgi network. The constitutive secretory pathway operates in all cells. Many soluble proteins are continually secreted from the cell by this pathway, which also supplies the plasma membrane with newly synthesized lipids and proteins. Specialized secretory cells also have a regulated secretory pathway, by which selected proteins in the trans Golgi network are diverted into secretory vesicles, where the proteins are concentrated and stored until an extracellular signal stimulates their secretion. The regulated secretion of small molecules, such as histamine, occurs by a similar pathway; these molecules are actively transported from the cytosol into preformed secretory vesicles. There they are often complexed to specific macromolecules (proteoglycans, for histamine), so that they can be stored at high concentration without generating an excessively high osmotic pressure. ( The pro-proteins of most constitutively secreted proteins (e.g., albumin) are cleaved only once at a site C-terminal to a dibasic recognition sequence such as Arg-Arg or Lys-Arg (Figure 14-24a). Proteolytic processing of proteins whose secretion is regulated generally entails additional cleavages. In the case of proinsulin, multiple cleavages of the single polypeptide chain yields the N-terminal B chain and the C- terminal A chain of mature insulin, which are linked by disulfide bonds, and the central C peptide, which is lost and subsequently degraded (Figure 14-24b).
117

118 Figure Trafficking of soluble lysosomal enzymes from the trans-golgi network and cell surface to lysosomes. 5
119 Mannose 6-phosphate residues target soluble proteins to lysosomes 1. As shown in Figure 14-22, the segregation of M6P-bearing lysosomal enzymes from secreted and membrane proteins occurs in the trans-golgi network. Here transmembrane mannose 6-phosphate receptors bind the M6P residues on lysosome-destined proteins very tightly and specifically. Clathrin/AP1 vesicles containing the M6P receptor and bound lysosomal enzymes then bud from the trans-golgi network, lose their coats, and subsequently fuse with the late endosome by mechanisms described previously. Because M6P receptors can bind M6P at the slightly acidic ph ( 6.5) of the trans-golgi network but not at a ph less than 6, the bound lysosomal enzymes are released within late endosomes, which have an internal ph of Furthermore, a phosphatase within late endosomes usually removes the phosphate from M6P residues on lysosomal enzymes, preventing any rebinding to the M6P receptor that might occur in spite of the low ph in endosomes. Vesicles budding from late endosomes recycle the M6P receptor back to the trans- Golgi network or, on occasion, to the cell surface. Eventually, mature late endosomes fuse with lysosomes, delivering the lysosomal enzymes to their final destination. 2. The sorting of soluble lysosomal enzymes in the trans-golgi network (steps 1 4 ) shares many of the features of trafficking between the ER and cis-golgi compartments mediated by COPII and COPI vesicles (Figure 14-22) First, mannose 6-phosphate acts as a sorting signal by interacting with the luminal domain of a receptor protein in the donor membrane. Second, the membrane-embedded receptors with their bound ligands are incorporated into the appropriate vesicles in this case, AP1-containing clathrin vesicles by interacting with the vesicle coat. Third, these transport vesicles fuse only with one specific organelle, here the late endosome, as the result of interactions between specific v-snares and t-snares. And finally, intracellular transport receptors are recycled after dissociating from their bound ligand.
120
")

121 Figure Formation of mannose 6-phosphate (M6P) residues that target soluble enzymes to lysosomes. Figure Molecular Biology of the Cell ( Garland Science 2008)
122 Mannose 6-phosphate (M6P) residues 1. Most of the sorting signals that function in vesicular trafficking are short amino acid sequences in the targeted protein. In contrast, the sorting signal that directs soluble lysosomal enzymes from the trans- Golgi network to the late endosome is a carbohydrate residue, mannose 6-phosphate (M6P), which is formed in the cis-golgi. The addition and initial processing of one or more preformed N-linked oligosaccharide precursors in the rough ER is the same for lysosomal enzymes as for membrane and secreted proteins, yielding core Man 8 (GlcNAc) 2 chains. In the cis-golgi, the N-linked oligosaccharides present on most lysosomal enzymes undergo a two-step reaction sequence that generates M6P residues (Figure 14-21, Figure Molecular Biology of the Cell). 2. The M6P residues that direct proteins to lysosomes are generated in the cis-golgi by two Golgiresident enzymes (Figure 14-21). 1 2 Step 1: An Nacetylglucosamine (GlcNAc) phosphotransferase transfers a phosphorylated GlcNAc group to carbon atom 6 of one or more mannose residues. Because only lysosomal enzymes contain sequences (red) that are recognized and bound by this enzyme, phosphorylated GlcNAc groups are added specifically to lysosomal enzymes. Step 2: After release of a modified protein from the phosphotransferase, a phosphodiesterase removes the GlcNAc group, leaving a phosphorylated mannose residue on the lysosomal enzyme.
123

124 Figure Model for dynamin-mediated pinching off of clathrin/ap-coated vesicles.
125 Clathrin/AP-coated vesicles A fundamental step in the formation of a transport vesicle that we have not yet considered is how a vesicle bud is pinched off from the donor membrane. In the case of clathrin/ap-coated vesicles, a cytosolic protein called dynamin is essential for release of complete vesicles. At the later stages of bud formation, dynamin polymerizes around the neck portion and then hydrolyzes GTP. The energy derived from GTP hydrolysis is thought to drive contraction of dynamin around the vesicle neck until the vesicle pinches off. Interestingly, COPI and COPII vesicles appear to pinch off from donor membranes without the aid of a GTPase such as dynamin (Figure 14-19).
126

127 Figure Structure of clathrin coats.
128 Structure of clathrin coats The best-characterized vesicles that bud from the trans-golgi network (TGN) have a two-layered coat: an outer layer composed of the fibrous protein clathrin and an inner layer composed of adapter protein (AP) complexes. Purified clathrin molecules, which have a three-limbed shape, are called triskelions from the Greek for threelegged. Each limb contains one clathrin heavy chain (180,000 MW) and one clathrin light chain ( 35,000 40,000 MW). Triskelions polymerize to form a polygonal lattice with an intrinsic curvature (Figure 14-18).
129

130 Experimental Figure Proteolytic cleavage of proinsulin occurs in secretory vesicles after they have budded from the trans-golgi network.
131 In the case of soluble lysosomal enzymes, the proproteins are called proenzymes, which are sorted by the M6P receptor as catalytically inactive enzymes. In the late endosome or lysosome a proenzyme undergoes a proteolytic cleavage that generates a smaller but enzymatically active polypeptide. Delaying the activation of lysosomal proenzymes until they reach the lysosome prevents them from digesting macromolecules in earlier compartments of the secretory pathway. Normally, mature vesicles carrying secreted proteins to the cell surface are formed by fusion of several immature ones containing proprotein. Proteolytic cleavage of proproteins, such as proinsulin, occurs in vesicles after they move away from the trans-golgi network (Figure 14-23).
132
")
133 Figure 13-1 Molecular Biology of the Cell ( Garland Science 2008) 6

134 Figure Molecular Biology of the Cell ( Garland Science 2008)
135 Exocytosis and endocytosis 1. After budding from the trans-golgi network, the first type of vesicle immediately moves to and fuses with the plasma membrane, releasing its contents by exocytosis. In all cell types, at least some proteins are loaded into such vesicles and secreted continuously in this manner (Figure 13-1 Molecular Biology of the Cell). 2. Although transport proteins in the plasma membrane mediate the movement of ions and small molecules across the lipid bilayer, proteins and some other soluble macromolecules in the extracellular milieu are internalized by endocytosis. In this process, a segment of the plasma membrane invaginates into a coated pit, whose cytosolic face is lined by a specific set of proteins including clathrin. Endocytosis is a way for cells to take up nutrients that are in macromolecular form for example, cholesterol in the form of lipoprotein particles and iron complexed with the serum protein transferrin. Endocytosis also can function as a regulatory mechanism to decrease signaling activity by withdrawing receptors for a particular signaling molecule from the cell surface (Figure 13-1, Molecular Biology of the Cell ).
136

137 Figure Endocytic pathway for internalizing low-density lipoprotein (LDL).

138 Experimental Figure The initial stages of receptor-mediated endocytosis of low-density lipoprotein (LDL) particles are revealed by electron microscopy.

139 Figure Molecular Biology of the Cell ( Garland Science 2008)

140 Figure Model of low-density lipoprotein (LDL).
141 Figure Model for ph-dependent binding of LDL particles by the LDL receptor.
142 Receptor-mediated endocytosis Most mammalian cells produce cell-surface receptors that specifically bind to apob-100 and internalize LDL particles by receptor-mediated endocytosis. After endocytosis, the LDL particles are transported to lysosomes via the endocytic pathway and then are degraded by lysosomal hydrolases. LDL receptors, which dissociate from their ligands in the late endosome, recycle to the cell surface. The major features of the LDL endocytic pathway as currently understood are depicted in Figure The LDL receptor is an 839-residue glycoprotein with a single transmembrane segment; it has a short C-terminal cytosolic segment and a long Nterminal exoplasmic segment that contains a β-propeller domain and a ligand-binding domain. Seven cysteine-rich imperfect repeats form the ligand-binding domain, which interacts with the apob-100 molecule in a LDL particle (Figure 14-29, Experimental Figure 14-26). low-density lipoprotein (LDL) 1. A lipoprotein particle has a shell composed of proteins (apolipoproteins) and a cholesterol-containing phospholipid monolayer. The shell is amphipathic because its outer surface is hydrophilic, making these particles water soluble, and its inner surface is hydrophobic (Figure 14-27). 2. Most receptors, including the M6P receptor and cell-surface receptors for LDL particles and asialoglycoprotein, bind their ligands tightly at neutral ph but release their ligands if the ph is lowered to 6.0 or below. The late endosome is the first vesicle encountered by receptor-ligand complexes whose luminal ph is sufficiently acidic to promote dissociation of most endocytosed receptors from their tightly bound ligands. The mechanism by which the LDL receptor releases bound LDL particles is now understood in detail. At the endosomal ph of , histidine residues in the β-propeller domain of the receptor become protonated, forming a site that can bind with high affinity to the negatively charged repeats in the ligandbinding domain. This intramolecular interaction sequesters the repeats in a conformation that cannot simultaneously bind to apob-100, thus causing release of the bound LDL particle (Figure 14-30).
143 LDL As will be discussed in detail in the next chapter, low density lipoprotein (LDL) is one of several complexes that carry cholesterol through the bloodstream. A LDL particle, a sphere nm in diameter, has an outer phospholipid shell containing a single molecule of a large protein known as apob-100; the core of a particle is packed with cholesterol in the form of cholesteryl esters. Among the common macromolecules that vertebrate cells internalize by receptor-mediated endocytosis are cholesterol containing particles called low-density lipoprotein (LDL); the iron-binding protein transferrin; many protein hormones (e.g., insulin); and certain glycoproteins. Receptor-mediated endocytosis of such ligands generally occurs via clathrin/ap2- coated pits and vesicles in a process similar to the packaging of lysosomal enzymes by mannose 6-phosphate (M6P) in the trans-golgi network (Figure Molecular Biology of the Cell).
144

145 Figure The transferrin cycle, which operates in all growing mammalian cells.
146 The transferrin cycle 1. An exception to the general theme of ph-dependent receptor-ligand dissociation in the late endosome occurs in the endocytic pathway that delivers transferrin-bound iron to cells. A major glycoprotein in the blood, transferrin transports iron to all tissue cells from the liver (the main site of iron storage in the body) and from the intestine (the site of iron absorption) (Figure 14-31) The iron-free form, apotransferrin, binds two Fe 3+ ions very tightly to form ferrotransferrin. All mammalian cells contain cell-surface transferrin receptors that avidly bind ferrotransferrin at neutral ph, after which the receptor-bound ferrotransferrin is subjected to endocytosis. Like the components of a LDL particle, the two bound Fe 3+ atoms remain in the cell, but the apotransferrin part of the ligand does not dissociate from the receptor and is secreted from the cell within minutes after being endocytosed. Although apotransferrin remains bound to the transferrin receptor at the low ph of late endosomes, changes in ph are critical to functioning of the transferrin endocytic pathway. 4 At a ph below 6.0, the two bound Fe 3+ atoms dissociate from ferrotransferrin, are reduced to Fe 2+ by an unknown mechanism, and then are exported into the cytosol by an endosomal transporter specific for divalent metal ions. 5 6 The receptor-apotransferrin complex remaining after dissociation of the iron atoms is recycled back to the cell surface. Although apotransferrin binds tightly to its receptor at a ph of 5.0 or 6.0, it does not bind at neutral ph. Hence the bound apotransferrin dissociates from the transferrin receptor when the recycling vesicles fuse with the plasma membrane and the receptor-ligand complex encounters the neutral ph of the extracellular interstitial fluid or growth medium. The recycled receptor is then free to bind another molecule of ferrotransferrin, and the released apotransferrin is carried in the bloodstream to the liver or intestine to be reloaded with iron.
147

148 Figure Sorting of proteins destined for the apical and basolateral plasma membranes of polarized cells.
149 Sorting of proteins destined for the apical and basolateral plasma membranes 1. The plasma membrane of polarized epithelial cells is divided into two domains, apical and basolateral; tight junctions located between the two domains prevent the movement of plasmamembrane proteins between the domains. 2. Several sorting mechanisms direct newly synthesized membrane proteins to either the apical or basolateral domain of epithelial cells, and any one protein may be sorted by more than one mechanism (Figure 14-25) Microscopic and cell-fractionation studies indicate that proteins destined for either the apical or the basolateral membranes are initially located together within the membranes of the trans-golgi network. In some cases, proteins destined for the apical membrane are sorted into their own transport vesicles that bud from the trans-golgi network and then move to the apical region, whereas proteins destined for the basolateral membrane are sorted into other vesicles that move to the basolateral region. The GPI anchor is the only apical-basolateral sorting signal identified so far. In hepatocytes and some other polarized cells, all plasma-membrane proteins are directed first to the basolateral membrane. Apically destined proteins then are endocytosed and moved across the cell to the apical membrane (transcytosis).
150

151 Figure Molecular Biology of the Cell ( Garland Science 2008)
Chapter 1: Vesicular traffic. Biochimica cellulare parte B 2017/18
 Chapter 1: Vesicular traffic Biochimica cellulare parte B 2017/18 Major Protein-sorting pathways in eukaryotic cells Secretory and endocytic pathways Unifying principle governs all protein trafficking
Chapter 1: Vesicular traffic Biochimica cellulare parte B 2017/18 Major Protein-sorting pathways in eukaryotic cells Secretory and endocytic pathways Unifying principle governs all protein trafficking
Protein Trafficking in the Secretory and Endocytic Pathways
 Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
endomembrane system internal membranes origins transport of proteins chapter 15 endomembrane system
 endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
Summary of Endomembrane-system
 Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic
 Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
Vesicle Transport. Vesicle pathway: many compartments, interconnected by trafficking routes 3/17/14
 Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Mechanism of Vesicular Transport
 Mechanism of Vesicular Transport Transport vesicles play a central role in the traffic of molecules between different membrane-enclosed enclosed compartments. The selectivity of such transport is therefore
Mechanism of Vesicular Transport Transport vesicles play a central role in the traffic of molecules between different membrane-enclosed enclosed compartments. The selectivity of such transport is therefore
Homework Hanson section MCB Course, Fall 2014
 Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
In the previous chapter we explored how proteins are targeted
 17 VESICULAR TRAFFIC, SECRETION, AND ENDOCYTOSIS Electron micrograph of clathrin cages, like those that surround clathrin-coated transport vesicles, formed by the in vitro polymerization of clathrin heavy
17 VESICULAR TRAFFIC, SECRETION, AND ENDOCYTOSIS Electron micrograph of clathrin cages, like those that surround clathrin-coated transport vesicles, formed by the in vitro polymerization of clathrin heavy
Molecular Trafficking
 SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
Intracellular Compartments and Protein Sorting
 Intracellular Compartments and Protein Sorting Intracellular Compartments A eukaryotic cell is elaborately subdivided into functionally distinct, membrane-enclosed compartments. Each compartment, or organelle,
Intracellular Compartments and Protein Sorting Intracellular Compartments A eukaryotic cell is elaborately subdivided into functionally distinct, membrane-enclosed compartments. Each compartment, or organelle,
Intracellular vesicular traffic. B. Balen
 Intracellular vesicular traffic B. Balen Three types of transport in eukaryotic cells Figure 12-6 Molecular Biology of the Cell ( Garland Science 2008) Endoplasmic reticulum in all eucaryotic cells Endoplasmic
Intracellular vesicular traffic B. Balen Three types of transport in eukaryotic cells Figure 12-6 Molecular Biology of the Cell ( Garland Science 2008) Endoplasmic reticulum in all eucaryotic cells Endoplasmic
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Zool 3200: Cell Biology Exam 4 Part I 2/3/15
 Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Chapter 13: Vesicular Traffic
 Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
1. endoplasmic reticulum This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Practice Exam 2 MCBII
 1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine
 Dr. Diala Abu-Hsasan School of Medicine") Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine dr.abuhassand@gmail.com An overview of cellular components Endoplasmic reticulum (ER) It is a network of membrane-enclosed
Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine dr.abuhassand@gmail.com An overview of cellular components Endoplasmic reticulum (ER) It is a network of membrane-enclosed
Outline. Neuron Structure. Week 4 - Nervous System. The Nervous System: Neurons and Synapses
 Outline Week 4 - The Nervous System: Neurons and Synapses Neurons Neuron structures Types of neurons Electrical activity of neurons Depolarization, repolarization, hyperpolarization Synapses Release of
Outline Week 4 - The Nervous System: Neurons and Synapses Neurons Neuron structures Types of neurons Electrical activity of neurons Depolarization, repolarization, hyperpolarization Synapses Release of
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
1. This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Chapter 4 Neuronal Physiology
 Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins
 Lecture 6: Membranes and Cell Transport Biological Membranes I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins 1. Characteristics a. Phospholipids form bilayers
Lecture 6: Membranes and Cell Transport Biological Membranes I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins 1. Characteristics a. Phospholipids form bilayers
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001
 BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting
 Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
ANATOMY AND PHYSIOLOGY OF NEURONS. AP Biology Chapter 48
 ANATOMY AND PHYSIOLOGY OF NEURONS AP Biology Chapter 48 Objectives Describe the different types of neurons Describe the structure and function of dendrites, axons, a synapse, types of ion channels, and
ANATOMY AND PHYSIOLOGY OF NEURONS AP Biology Chapter 48 Objectives Describe the different types of neurons Describe the structure and function of dendrites, axons, a synapse, types of ion channels, and
Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation
 Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 2/21/17 glycosylation is a non-template derived phenomenon - the presence of
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 2/21/17 glycosylation is a non-template derived phenomenon - the presence of
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation
 Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 3/8/2011 glycosylation is a non-template derived phenomenon - the presence of
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 3/8/2011 glycosylation is a non-template derived phenomenon - the presence of
Branches of the Nervous System
 The Nervous System Branches of the Nervous System There are 2 main branches of the nervous system Central Nervous System Brain Spinal Cord Peripheral Nervous System All nerves leading to rest of body Anatomy
The Nervous System Branches of the Nervous System There are 2 main branches of the nervous system Central Nervous System Brain Spinal Cord Peripheral Nervous System All nerves leading to rest of body Anatomy
Intracellular Vesicular Traffic Chapter 13, Alberts et al.
 Intracellular Vesicular Traffic Chapter 13, Alberts et al. The endocytic and biosynthetic-secretory pathways The intracellular compartments of the eucaryotic ell involved in the biosynthetic-secretory
Intracellular Vesicular Traffic Chapter 13, Alberts et al. The endocytic and biosynthetic-secretory pathways The intracellular compartments of the eucaryotic ell involved in the biosynthetic-secretory
The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae.
 Endoplasmic reticulum (ER) The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae. Cisternae serve as channels for the transport of
Endoplasmic reticulum (ER) The endoplasmic reticulum is a network of folded membranes that form channels through the cytoplasm and sacs called cisternae. Cisternae serve as channels for the transport of
PROTEIN TRAFFICKING. Dr. SARRAY Sameh, Ph.D
 PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
What is Anatomy and Physiology?
 Introduction BI 212 BI 213 BI 211 Ecosystems Organs / organ systems Cells Organelles Communities Tissues Molecules Populations Organisms Campbell et al. Figure 1.4 Introduction What is Anatomy and Physiology?
Introduction BI 212 BI 213 BI 211 Ecosystems Organs / organ systems Cells Organelles Communities Tissues Molecules Populations Organisms Campbell et al. Figure 1.4 Introduction What is Anatomy and Physiology?
Nervous System. Master controlling and communicating system of the body. Secrete chemicals called neurotransmitters
 Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Zool 3200: Cell Biology Exam 4 Part I 2/3/15
 Name: Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
AP Biology Unit 6. The Nervous System
 AP Biology Unit 6 The Nervous System Branches of the Nervous System There are 2 main branches of the nervous system Central Nervous System Brain Spinal Cord Peripheral Nervous System All nerves leading
AP Biology Unit 6 The Nervous System Branches of the Nervous System There are 2 main branches of the nervous system Central Nervous System Brain Spinal Cord Peripheral Nervous System All nerves leading
2013 John Wiley & Sons, Inc. All rights reserved. PROTEIN SORTING. Lecture 10 BIOL 266/ Biology Department Concordia University. Dr. S.
 PROTEIN SORTING Lecture 10 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University Introduction Membranes divide the cytoplasm of eukaryotic cells into distinct compartments. The endomembrane
PROTEIN SORTING Lecture 10 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University Introduction Membranes divide the cytoplasm of eukaryotic cells into distinct compartments. The endomembrane
Chapter 2: Cellular Mechanisms and Cognition
 Chapter 2: Cellular Mechanisms and Cognition MULTIPLE CHOICE 1. Two principles about neurons were defined by Ramón y Cajal. The principle of connectional specificity states that, whereas the principle
Chapter 2: Cellular Mechanisms and Cognition MULTIPLE CHOICE 1. Two principles about neurons were defined by Ramón y Cajal. The principle of connectional specificity states that, whereas the principle
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline
 Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
TRANSPORT PROCESSES. 1b. moving proteins into membranes and organelles
 1b. moving proteins into membranes and organelles SLIDE 1 A typical mammalian cell contains up to 10,000 different kinds of proteins. The vast majority of these proteins are synthesized by cytosolic ribosomes,
1b. moving proteins into membranes and organelles SLIDE 1 A typical mammalian cell contains up to 10,000 different kinds of proteins. The vast majority of these proteins are synthesized by cytosolic ribosomes,
Portions from Chapter 6 CHAPTER 7. The Nervous System: Neurons and Synapses. Chapter 7 Outline. and Supporting Cells
 CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
CHAPTER 7 The Nervous System: Neurons and Synapses Chapter 7 Outline Neurons and Supporting Cells Activity in Axons The Synapse Acetylcholine as a Neurotransmitter Monoamines as Neurotransmitters Other
The Nervous System. Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output
 The Nervous System Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output The Nervous System 2 Parts of the Nervous System 1. central
The Nervous System Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output The Nervous System 2 Parts of the Nervous System 1. central
CELL BIOLOGY - CLUTCH CH INTRACELLULAR PROTEIN TRANSPORT.
 !! www.clutchprep.com CONCEPT: MEMBRANE ENCLOSED ORGANELLES Table of eukaryotic organelles and their functions Organelle Function % volume of cell Cytosol Aqueous fluid where metabolic pathways and chemical
!! www.clutchprep.com CONCEPT: MEMBRANE ENCLOSED ORGANELLES Table of eukaryotic organelles and their functions Organelle Function % volume of cell Cytosol Aqueous fluid where metabolic pathways and chemical
Posttranslational Modification and Targeting of Proteins
 Posttranslational Modification and Targeting of Proteins Graduate Biochemistry Term 2/2016 Assist. Prof. Dr. Panida Khunkaewla School of Chemistry, Institute of Science Suranaree University of Technology
Posttranslational Modification and Targeting of Proteins Graduate Biochemistry Term 2/2016 Assist. Prof. Dr. Panida Khunkaewla School of Chemistry, Institute of Science Suranaree University of Technology
17/01/2017. Protein trafficking between cell compartments. Lecture 3: The cytosol. The mitochondrion - the power plant of the cell
 ell biology 2017 version 13/1 2017 ote endosome vs lysosome handout Lecture 3: Text book Alberts et al.: hapter 12-14 (Topics covered by the lecture) A lot of reading! Focus on principles ell Biology interactive
ell biology 2017 version 13/1 2017 ote endosome vs lysosome handout Lecture 3: Text book Alberts et al.: hapter 12-14 (Topics covered by the lecture) A lot of reading! Focus on principles ell Biology interactive
Chapter 3 Neurotransmitter release
 NEUROPHYSIOLOGIE CELLULAIRE CONSTANCE HAMMOND Chapter 3 Neurotransmitter release In chapter 3, we proose 3 videos: Observation Calcium Channel, Ca 2+ Unitary and Total Currents Ca 2+ and Neurotransmitter
NEUROPHYSIOLOGIE CELLULAIRE CONSTANCE HAMMOND Chapter 3 Neurotransmitter release In chapter 3, we proose 3 videos: Observation Calcium Channel, Ca 2+ Unitary and Total Currents Ca 2+ and Neurotransmitter
Introduction to Neurobiology
 Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
BIOLOGY 2050 LECTURE NOTES ANATOMY & PHYSIOLOGY I (A. IMHOLTZ) FUNDAMENTALS OF THE NERVOUS SYSTEM AND NERVOUS TISSUE P1 OF 5
 FUNDAMENTALS OF THE NERVOUS SYSTEM AND NERVOUS TISSUE P1 OF 5") P1 OF 5 The nervous system controls/coordinates the activities of cells, tissues, & organs. The endocrine system also plays a role in control/coordination. The nervous system is more dominant. Its mechanisms
P1 OF 5 The nervous system controls/coordinates the activities of cells, tissues, & organs. The endocrine system also plays a role in control/coordination. The nervous system is more dominant. Its mechanisms
Molecular Cell Biology 5068 In Class Exam 1 October 3, 2013
 Molecular Cell Biology 5068 In Class Exam 1 October 3, 2013 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your number
Molecular Cell Biology 5068 In Class Exam 1 October 3, 2013 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your number
The Nervous System -The master controlling and communicating system of the body
 The Nervous System -The master controlling and communicating system of the body Functions: -Sensory input -Integration -Motor output Organization of the Nervous System Central nervous system (CNS) -Brain
The Nervous System -The master controlling and communicating system of the body Functions: -Sensory input -Integration -Motor output Organization of the Nervous System Central nervous system (CNS) -Brain
Neurons, Synapses, and Signaling
 Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Molecular Cell Biology 5068 In Class Exam 1 September 29, Please print your name:
 Molecular Cell Biology 5068 In Class Exam 1 September 29, 2015 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 1 September 29, 2015 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Zool 3200: Cell Biology Exam 4 Part II 2/3/15
 Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Multiple choice questions. Pick the BEST answer (2 pts ea)
") Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
Signal Transduction Cascades
 Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Chapter 3 subtitles Action potentials
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 3 subtitles Action potentials Introduction (3:15) This third chapter explains the calcium current triggered by the arrival of the action potential in
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 3 subtitles Action potentials Introduction (3:15) This third chapter explains the calcium current triggered by the arrival of the action potential in
Cell Membrane and Transport
 Cell Membrane and Transport 29/06/2015 11:08 AM Describe the Characteristics of the phospholipid Bilayer. The Phospholipid bilayer is made up of a double layer of membrane lipids that have a hydrophobic
Cell Membrane and Transport 29/06/2015 11:08 AM Describe the Characteristics of the phospholipid Bilayer. The Phospholipid bilayer is made up of a double layer of membrane lipids that have a hydrophobic
Lecture 6 - Intracellular compartments and transport I
 01.25.10 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
01.25.10 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
Chapter 11: Nervous System and Nervous Tissue
 Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM
 STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
What effect would an AChE inhibitor have at the neuromuscular junction?
 CASE 4 A 32-year-old woman presents to her primary care physician s office with difficulty chewing food. She states that when she eats certain foods that require a significant amount of chewing (meat),
CASE 4 A 32-year-old woman presents to her primary care physician s office with difficulty chewing food. She states that when she eats certain foods that require a significant amount of chewing (meat),
Lecture Series 4 Cellular Membranes
 Lecture Series 4 Cellular Membranes Reading Assignments Read Chapter 11 Membrane Structure Review Chapter 21 pages 709-717 717 (Animal( Cell Adhesion) Review Chapter 12 Membrane Transport Review Chapter
Lecture Series 4 Cellular Membranes Reading Assignments Read Chapter 11 Membrane Structure Review Chapter 21 pages 709-717 717 (Animal( Cell Adhesion) Review Chapter 12 Membrane Transport Review Chapter
Advanced Cell Biology. Lecture 33
 Advanced Cell Biology. Lecture 33 Alexey Shipunov Minot State University April 22, 2013 Shipunov (MSU) Advanced Cell Biology. Lecture 33 April 22, 2013 1 / 38 Outline Questions and answers Intracellular
Advanced Cell Biology. Lecture 33 Alexey Shipunov Minot State University April 22, 2013 Shipunov (MSU) Advanced Cell Biology. Lecture 33 April 22, 2013 1 / 38 Outline Questions and answers Intracellular
MOLECULAR AND CELLULAR NEUROSCIENCE
 MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
Chapter 11: Functional Organization of Nervous Tissue
 Chapter 11: Functional Organization of Nervous Tissue I. Functions of the Nervous System A. List and describe the five major nervous system functions: 1. 2. 3. 4. 5. II. Divisions of the Nervous System
Chapter 11: Functional Organization of Nervous Tissue I. Functions of the Nervous System A. List and describe the five major nervous system functions: 1. 2. 3. 4. 5. II. Divisions of the Nervous System
Lecture 36: Review of membrane function
 Chem*3560 Lecture 36: Review of membrane function Membrane: Lipid bilayer with embedded or associated proteins. Bilayers: 40-70% neutral phospholipid 10-20% negative phospholipid 10-30% cholesterol 10-30%
Chem*3560 Lecture 36: Review of membrane function Membrane: Lipid bilayer with embedded or associated proteins. Bilayers: 40-70% neutral phospholipid 10-20% negative phospholipid 10-30% cholesterol 10-30%
3) Most of the organelles in a neuron are located in the A) dendritic region. B) axon hillock. C) axon. D) cell body. E) axon terminals.
 Most of the organelles in a neuron are located in the A) dendritic region. B) axon hillock. C) axon. D) cell body. E) axon terminals.") Chapter 48 Neurons, Synapses, and Signaling Multiple-Choice Questions 1) A simple nervous system A) must include chemical senses, mechanoreception, and vision. B) includes a minimum of 12 ganglia. C) has
Chapter 48 Neurons, Synapses, and Signaling Multiple-Choice Questions 1) A simple nervous system A) must include chemical senses, mechanoreception, and vision. B) includes a minimum of 12 ganglia. C) has
Neurophysiology scripts. Slide 2
 Neurophysiology scripts Slide 2 Nervous system and Endocrine system both maintain homeostasis in the body. Nervous system by nerve impulse and Endocrine system by hormones. Since the nerve impulse is an
Neurophysiology scripts Slide 2 Nervous system and Endocrine system both maintain homeostasis in the body. Nervous system by nerve impulse and Endocrine system by hormones. Since the nerve impulse is an
Neurons, Synapses, and Signaling
 Neurons, Synapses, and Signaling The Neuron is the functional unit of the nervous system. Neurons are composed of a cell body, which contains the nucleus and organelles; Dendrites which are extensions
Neurons, Synapses, and Signaling The Neuron is the functional unit of the nervous system. Neurons are composed of a cell body, which contains the nucleus and organelles; Dendrites which are extensions
Cells communicate with each other via signaling ligands which interact with receptors located on the surface or inside the target cell.
 BENG 100 Frontiers of Biomedical Engineering Professor Mark Saltzman Chapter 6 SUMMARY In this chapter, cell signaling was presented within the context of three physiological systems that utilize communication
BENG 100 Frontiers of Biomedical Engineering Professor Mark Saltzman Chapter 6 SUMMARY In this chapter, cell signaling was presented within the context of three physiological systems that utilize communication
Chapt. 10 Cell Biology and Biochemistry. The cell: Student Learning Outcomes: Describe basic features of typical human cell
 Chapt. 10 Cell Biology and Biochemistry Cell Chapt. 10 Cell Biology and Biochemistry The cell: Lipid bilayer membrane Student Learning Outcomes: Describe basic features of typical human cell Integral transport
Chapt. 10 Cell Biology and Biochemistry Cell Chapt. 10 Cell Biology and Biochemistry The cell: Lipid bilayer membrane Student Learning Outcomes: Describe basic features of typical human cell Integral transport
Cell Membranes and Signaling
 5 Cell Membranes and Signaling Concept 5.1 Biological Membranes Have a Common Structure and Are Fluid A membrane s structure and functions are determined by its constituents: lipids, proteins, and carbohydrates.
5 Cell Membranes and Signaling Concept 5.1 Biological Membranes Have a Common Structure and Are Fluid A membrane s structure and functions are determined by its constituents: lipids, proteins, and carbohydrates.
Neurons. Pyramidal neurons in mouse cerebral cortex expressing green fluorescent protein. The red staining indicates GABAergic interneurons.
 Neurons Pyramidal neurons in mouse cerebral cortex expressing green fluorescent protein. The red staining indicates GABAergic interneurons. MBL, Woods Hole R Cheung MSc Bioelectronics: PGEE11106 1 Neuron
Neurons Pyramidal neurons in mouse cerebral cortex expressing green fluorescent protein. The red staining indicates GABAergic interneurons. MBL, Woods Hole R Cheung MSc Bioelectronics: PGEE11106 1 Neuron
BIOL 158: BIOLOGICAL CHEMISTRY II
 BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
Renata Schipp Medical Biology Department
 Renata Schipp Medical Biology Department Deffinition of cell The cell is the smallest structural and functional unit of all known living organisms The cell was discovered by Robert Hooke in 1665 and also
Renata Schipp Medical Biology Department Deffinition of cell The cell is the smallest structural and functional unit of all known living organisms The cell was discovered by Robert Hooke in 1665 and also
Membrane Structure and Membrane Transport of Small Molecules. Assist. Prof. Pinar Tulay Faculty of Medicine
 Membrane Structure and Membrane Transport of Small Molecules Assist. Prof. Pinar Tulay Faculty of Medicine Introduction Cell membranes define compartments of different compositions. Membranes are composed
Membrane Structure and Membrane Transport of Small Molecules Assist. Prof. Pinar Tulay Faculty of Medicine Introduction Cell membranes define compartments of different compositions. Membranes are composed
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed.,
 Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by B.-W. Ku,
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by B.-W. Ku,
Electrical Properties of Neurons. Steven McLoon Department of Neuroscience University of Minnesota
 Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Unit Three. I. General Functions of the Nervous System. I. General Functions of the Nervous System
 10 Refer to the following URLs. It is a good idea to print them and bring them to class. Be sure to study these along with your book. http://www.sirinet.net/~jgjohnso/nervous.html http://faculty.washington.edu/chudler/ap.html
10 Refer to the following URLs. It is a good idea to print them and bring them to class. Be sure to study these along with your book. http://www.sirinet.net/~jgjohnso/nervous.html http://faculty.washington.edu/chudler/ap.html
THE NERVOUS SYSTEM. Neurons & Impulses
 THE NERVOUS SYSTEM Neurons & Impulses Organization of the Nervous System: Two Major Portions: The central nervous system (CNS) and the peripheral nervous system (PNS). CNS = Brain/Spinal Cord PNS = Nerves-provide
THE NERVOUS SYSTEM Neurons & Impulses Organization of the Nervous System: Two Major Portions: The central nervous system (CNS) and the peripheral nervous system (PNS). CNS = Brain/Spinal Cord PNS = Nerves-provide
Introduction and protein sorting
 Introduction and protein sorting Membrane proteins Major components of cells Nucleic acids Carbohydrates Proteins Lipids (50% of mass of plasma membranes, 30% of mitochondrial membranes, 80% of myelin
Introduction and protein sorting Membrane proteins Major components of cells Nucleic acids Carbohydrates Proteins Lipids (50% of mass of plasma membranes, 30% of mitochondrial membranes, 80% of myelin
Ameen Alsaras. Ameen Alsaras. Mohd.Khatatbeh
 9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
Major Structures of the Nervous System. Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors
 Major Structures of the Nervous System Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors Nervous System Divisions Central Nervous System (CNS) consists
Major Structures of the Nervous System Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors Nervous System Divisions Central Nervous System (CNS) consists
A. Major parts 1. Nucleus 2. Cytoplasm a. Contain organelles (see below) 3. Plasma membrane (To be discussed in Cellular Transport Lecture)
 3. Plasma membrane (To be discussed in Cellular Transport Lecture)") Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
The axon and the nerve impulse, Generation and propagation of the nerve impulse, Ionic channels, Synaptic transmission.
 The axon and the nerve impulse, Generation and propagation of the nerve impulse, Ionic channels, Synaptic transmission Mitesh Shrestha Structure of the neuron the neuron consists of: the cell body, or
The axon and the nerve impulse, Generation and propagation of the nerve impulse, Ionic channels, Synaptic transmission Mitesh Shrestha Structure of the neuron the neuron consists of: the cell body, or
Cells: The Living Units
 Chapter 3 Part B Cells: The Living Units Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 3.4 Active Membrane Transport Two major
Chapter 3 Part B Cells: The Living Units Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 3.4 Active Membrane Transport Two major
Cell Quality Control. Peter Takizawa Department of Cell Biology
 Cell Quality Control Peter Takizawa Department of Cell Biology Cellular quality control reduces production of defective proteins. Cells have many quality control systems to ensure that cell does not build
Cell Quality Control Peter Takizawa Department of Cell Biology Cellular quality control reduces production of defective proteins. Cells have many quality control systems to ensure that cell does not build
Enzymes Part III: regulation II. Dr. Mamoun Ahram Summer, 2017
 Enzymes Part III: regulation II Dr. Mamoun Ahram Summer, 2017 Advantage This is a major mechanism for rapid and transient regulation of enzyme activity. A most common mechanism is enzyme phosphorylation
Enzymes Part III: regulation II Dr. Mamoun Ahram Summer, 2017 Advantage This is a major mechanism for rapid and transient regulation of enzyme activity. A most common mechanism is enzyme phosphorylation
Renáta Schipp Gergely Berta Department of Medical Biology
 The cell III. Renáta Schipp Gergely Berta Department of Medical Biology Size and Biology Biology is a visually rich subject many of the biological events and structures are smaller than the unaided human
The cell III. Renáta Schipp Gergely Berta Department of Medical Biology Size and Biology Biology is a visually rich subject many of the biological events and structures are smaller than the unaided human
Chapter 7: Membranes
 Chapter 7: Membranes Roles of Biological Membranes The Lipid Bilayer and the Fluid Mosaic Model Transport and Transfer Across Cell Membranes Specialized contacts (junctions) between cells What are the
Chapter 7: Membranes Roles of Biological Membranes The Lipid Bilayer and the Fluid Mosaic Model Transport and Transfer Across Cell Membranes Specialized contacts (junctions) between cells What are the
6.5 Nerves, Hormones and Homeostasis
 6.5 Nerves, Hormones and Homeostasis IB Biology SL Part 1 - Nerves Outcomes Part 1 6.5.1State that the nervous system consists of the central nervous system (CNS) and peripheral nerves, and is composed
6.5 Nerves, Hormones and Homeostasis IB Biology SL Part 1 - Nerves Outcomes Part 1 6.5.1State that the nervous system consists of the central nervous system (CNS) and peripheral nerves, and is composed
10.1: Introduction. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial cells) Dendrites.
 Dendrites.") 10.1: Introduction Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial
10.1: Introduction Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Cell types in neural tissue: Neurons Neuroglial cells (also known as neuroglia, glia, and glial
LECTURE STRUCTURE ASC171 NERVOUS SYSTEM PART 1: BACKGROUND 26/07/2015. Module 5
 LECTURE STRUCTURE PART 1: Background / Introduction PART 2: Structure of the NS, how it operates PART 3: CNS PART 4: PNS Why did the action potential cross the synaptic junction? To get to the other side
LECTURE STRUCTURE PART 1: Background / Introduction PART 2: Structure of the NS, how it operates PART 3: CNS PART 4: PNS Why did the action potential cross the synaptic junction? To get to the other side
Chapter 2. The Cellular and Molecular Basis of Cognition
 Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by B.-W. Ku,
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by B.-W. Ku,
Cell Membranes Valencia college
 6 Cell Membranes Valencia college 6 Cell Membranes Chapter objectives: The Structure of a Biological Membrane The Plasma Membrane Involved in Cell Adhesion and Recognition Passive Processes of Membrane
6 Cell Membranes Valencia college 6 Cell Membranes Chapter objectives: The Structure of a Biological Membrane The Plasma Membrane Involved in Cell Adhesion and Recognition Passive Processes of Membrane
Summarized by B.-W. Ku, E. S. Lee, and B.-T. Zhang Biointelligence Laboratory, Seoul National University.
 Chapter 2. The Cellular l and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 3 rd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2008. Summarized by B.-W. Ku,
Chapter 2. The Cellular l and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 3 rd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2008. Summarized by B.-W. Ku,
Lecture Series 5 Cellular Membranes
 Lecture Series 5 Cellular Membranes Cellular Membranes A. Membrane Composition and Structure B. Animal Cell Adhesion C. Passive Processes of Membrane Transport D. Active Transport E. Endocytosis and Exocytosis
Lecture Series 5 Cellular Membranes Cellular Membranes A. Membrane Composition and Structure B. Animal Cell Adhesion C. Passive Processes of Membrane Transport D. Active Transport E. Endocytosis and Exocytosis
A. Membrane Composition and Structure. B. Animal Cell Adhesion. C. Passive Processes of Membrane Transport. D. Active Transport
 Cellular Membranes A. Membrane Composition and Structure Lecture Series 5 Cellular Membranes B. Animal Cell Adhesion E. Endocytosis and Exocytosis A. Membrane Composition and Structure The Fluid Mosaic
Cellular Membranes A. Membrane Composition and Structure Lecture Series 5 Cellular Membranes B. Animal Cell Adhesion E. Endocytosis and Exocytosis A. Membrane Composition and Structure The Fluid Mosaic
Learning expectations for BIOL 131. Chapters 11, Nervous System Overview Read Chapter 11. You should be able to:
 NOTE The quiz will have question ONLY from the material we get through on Tuesday. The first midterm will cover all material from day one until the lecture before the second midterm. Learning expectations
NOTE The quiz will have question ONLY from the material we get through on Tuesday. The first midterm will cover all material from day one until the lecture before the second midterm. Learning expectations