The role of dystroglycan in the prostate epithelium and prostate cancer
|
|
|
- Barbara Watts
- 5 years ago
- Views:
Transcription

 thesis, University of Iowa, 2011. https://ir.uiowa.edu/etd/2696. Follow this and additional works at: https://ir.uiowa.edu/etd Part of the Biophysics Commons")
1 University of Iowa Iowa Research Online Theses and Dissertations Fall 2011 The role of dystroglycan in the prostate epithelium and prostate cancer Alison K. Esser University of Iowa Copyright 2011 Alison Esser This dissertation is available at Iowa Research Online: Recommended Citation Esser, Alison K.. "The role of dystroglycan in the prostate epithelium and prostate cancer." PhD (Doctor of Philosophy) thesis, University of Iowa, Follow this and additional works at: Part of the Biophysics Commons
2 THE ROLE OF DYSTROGLYCAN IN THE PROSTATE EPITHELIUM AND PROSTATE CANCER by Alison K. Esser An Abstract Of a thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Molecular Physiology and Biophysics in the Graduate College of The University of Iowa December 2011 Thesis Supervisor: Associate Professor Michael D. Henry
3 1 ABSTRACT Interactions between cells and the extracellular matrix (ECM) are essential to the organization and maintenance of tissue architecture and function. ECM receptors serve as a link between the cell and the ECM. Through interactions with various matrix molecules and activation of intracellular signaling pathways, ECM receptors allow cells to sense and respond to their microenvironment. The matrix receptor dystroglycan (DG) has been shown to have roles in tissue morphogenesis including formation of basement membranes, regulation of cell proliferation, differentiation and cell survival. DG is expressed in many tissues but has predominantly been studied in muscle. The function of dystroglycan within epithelia is currently unknown. To gain insight into the role of dystroglycan in the prostate epithelium, we generated individual prostate luminal cell (Probasin Cre) and basal cell (Keratin 5 Cre) specific DG knockout mice. DG was not required for maintenance of the basement membrane, epithelial polarity or cellular homeostasis in the prostate. Furthermore, gland morphology and ability to regenerate following androgen depletion were normal. These studies indicate DG may have more subtle roles within the epithelium. Disruption of cell/ecm interactions is a hallmark of cancer. DG expression is lost in many carcinomas including prostate, yet the molecular mechanism behind loss of expression and the functional consequences remain unclear. To elucidate the molecular mechanism in prostate cancer, we examined DG expression in metastatic prostate cancer cell lines. DG was heterogeneously glycosylated across the cell line panel. Surprisingly, we show that LARGE2 is able to functionally glycosylate DG and loss of LARGE2 expression is a mechanism for DG hypoglycosylation in prostate cancer. Additionally,
4 2 initial results suggest that oncogene expression modulates DG glycosylation status through regulation of LARGE2 expression. This work has shown a novel mechanism for DG hypoglycosylation in prostate cancer. In summary, these studies have contributed new information on the role of DG in the prostate epithelium. Furthermore, we have shown a novel mechanism for loss of DG glycosylation in prostate cancer and have provided initial data suggesting oncogene expression modulates DG glycosylation. These insights may lead to advances in the treatment of prostate cancer. Abstract Approved: Thesis Supervisor Title and Department Date
5 THE ROLE OF DYSTROGLYCAN IN THE PROSTATE EPITHELIUM AND PROSTATE CANCER by Alison K. Esser A thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Molecular Physiology and Biophysics in the Graduate College of The University of Iowa December 2011 Thesis Supervisor: Associate Professor Michael D. Henry
6 Graduate College The University of Iowa Iowa City, Iowa CERTIFICATE OF APPROVAL PH.D. THESIS This is to certify that the Ph.D. thesis of Alison K. Esser has been approved by the Examining Committee for the thesis requirement for the Doctor of Philosophy degree in Molecular Physiology and Biophysics at the December 2011 graduation. Thesis Committee: Michael D. Henry, Thesis Supervisor Kevin Campbell Steven Moore Michael Anderson Mark Stamnes
7 To my parents ii
8 ACKNOWLEDGMENTS I would like to thank all the past and present members of the Henry Lab for their advice, support, and friendship. I would also like to thank Dr. Kevin Campbell for his kind gifts of animals and reagents as well as helpful advice. Thank you to my thesis committee for their suggestions and feedback. Finally, thank you to Dr. Michael Henry for his guidance and mentorship over the years. iii
9 TABLE OF CONTENTS LIST OF FIGURES... vi CHAPTER I. INTRODUCTION...1 The Extracellular Matrix...1 Dystroglycan...2 Structure and Biosynthesis of Dystroglycan...2 Dystroglycan Posttranslational Modifications...3 DG Glycosyltransferases...5 Dystroglycan Interactions...8 Dystroglycan Function in Development...9 DG Function in Muscular Dystrophies...10 DG Function in Non-Epithelial Tissues...11 DG Function in the Epithelium...14 The Extracellular Matrix and Cancer...17 Prostate Cancer...18 DG Function in Cancer...20 Relevance and Focus...22 II. III. DYSTROGLYCAN IS NOT REQUIRED FOR MAINTENANCE OF THE LUMINAL EPITHELIAL BASEMENT MEMBRANE OR CELL POLARITY IN THE MOUSE PROSTATE...24 Introduction...24 Materials and Methods...26 Animals...26 Antibodies...27 Histology and Immunofluorescence Analysis...27 X-gal Histochemistry...28 Transmission Electron Microscopy...29 Protein Isolation and Western Blot...29 Results...30 DG Expression in the Mouse Prostate...30 Normal Morphology, Ultrastructure, Cellular Composition and Function of DG Knockout Prostates...31 Normal Regeneration of DG Knockout Prostates...33 DG Expression in the Castrate-Resistant Basal Cell Population...34 Discussion...35 DYSTROGLYCAN EXPRESSION IN THE PROSTATE BASAL CELL IS NOT REQUIRED FOR MORPHOGENESIS OF THE PROSTATE OR BASEMENT MEMBRANE FORMATION...48 Introduction...48 Materials and Methods...50 Animals...50 Antibodies...51 Histology and Immunofluorescence Analysis...51 X-gal Histochemistry...52 iv
10 Wound Healing...52 Results...53 DG Expression in a Basal Cell Specific Knockout Mouse...53 DG is Not Required for Prostate Development Including the Formation and Maintenance of the Basement Membrane...54 DG Null Prostates are Able to Regenerate Following Ablation and Reintroduction of Androgens DG is Not Required for Morphogenesis of the Ear, Tongue, Stomach or Trachea...56 Wound Closure Rate is Reduced in DG Null Mice...57 Modulation of the Stroma with Loss of DG in a PTEN Deficient Model of Prostate Cancer...58 Discussion...59 IV. LOSS OF LARGE2 EXPRESSION IS A MECHANISM FOR HYPOGLYCOSYLATION OF DYSTROGLYCAN IN PROSTATE CANCER...85 Introduction...85 Materials and Methods...86 Cell Culture...86 shrna Inhibition of LARGE Quantitative PCR...87 Western Blot...88 Flow Cytometry Analysis...88 Laminin-111 Binding...89 Immunofluorescence...89 Results...90 Heterogeneous Functional Glycosylation of DG in Prostate Cancer Cell Lines...90 DG Glycosylation Correlates with LARGE2 mrna Expression...91 LARGE2 Functionally Glycosylates DG in the Prostate...92 Oncogene Expression Modulates DG Glycosylation...94 Reduced Growth and Tumorigenicity with DG knockdown...94 Discussion...95 V. SUMMARY AND FUTURE DIRECTIONS REFERENCES v
11 LIST OF FIGURES Figure 2.1 Dystroglycan localization and expression in the ventral lobe of a prostate specific knockout mouse Dystroglycan localization and expression in all lobes of the prostate Histology and ultrastructure of the prostate Basement membrane and cellular composition of the ventral lobe of the prostate Involution and regeneration of the lateral and ventral lobes of the mouse prostate Expression of DG ligands following involution and regeneration of the mouse prostate DG expression following involution and regeneration of the mouse prostate Expression of DG in the basal cell population of the prostate proximal ducts Models of prostate stem cell lineage Cre recombinase expression in the mouse prostate Dystroglycan expression is lost in DAG1 knockout mice Dystroglycan expression is lost in all lobes of the mouse prostate DG expression is lost in basal cells and some luminal cells DG is lost in the basal cells of the proximal ducts DG knockout mice have no abnormalities in tissue morphology DG null prostates have normal localization of basement membrane proteins and basal cell number DG expression is absent from involuted and regenerated prostates DG expression in keratin 5 negative cells following androgen ablation and regeneration DG KO prostates are able to regenerate following involution DG KO regenerated prostates have normal localization of basement membrane proteins and basal cell number Keratin-5 driven Cre expression deletes DAG1 from many stratified epithelia...76 vi
12 3.14 DG is not required for tissue morphogenesis in some stratified epithelia as assessed by histology DG ligand expression and localization were unchanged in the epidermis The DG null epidermis had normal morphology at the anagen and telogen phases of the hair cycle The progression of the hair cycle was unchanged with loss of DG in the epidermis Loss of DG reduced the rate of wound healing in the epidermis Wound histology in DG WT and KO mice Prostate specific deletion of DAG1 and PTEN in the lateral lobe Modulation of the stroma and vasculature in the lateral prostate of DG PTEN KO mice Functional glycosylation of DG is heterogeneous in prostate cancer LARGE2 expression correlates with hypoglycosylation of DG LARGE2 is expressed in prostate epithelial cell lines LARGE2 functionally glycosylates DG LARGE and LARGE2 expression is dependent on tissue type in cancer cell lines Oncogene expression modulates -dystroglycan glycosylation Generation of DG knockdown cell lines Loss of DG enhances proliferation DG knockdown reduces anchorage to laminin Reduced DG expression enhances anchorage independent growth Tumor growth is not altered with loss of DG vii
13 1 CHAPTER I INTRODUCTION The Extracellular Matrix Interactions between cells and the extracellular matrix are essential to the organization and maintenance of tissue architecture and function. The extracellular matrix (ECM) is composed of an array of molecules including matrix proteins, growth factors and cytokines that give support to and provide information about the microenvironment to cells. Basement membranes are a specialized extracellular matrix that separates the epithelium from the surrounding stroma. Epithelial cells require anchorage to the basement membrane for survival. Basement membranes are primarily composed of laminin, collagen IV, perlecan, entactin and may include growth factors and proteases. The composition of the basement membrane is tightly regulated during development. Its composition varies with tissue type and changes under pathological conditions in order to regulate the proliferation, differentiation and survival of epithelial cells (De Arcangelis and Georges-Labouesse 2000; Gustafsson and Fassler 2000). Extracellular matrix receptors mediate cell/ecm interactions. Through interactions with various ECM molecules and activation of intracellular signaling pathways, ECM receptors allow cells to sense and respond to their microenvironment. In addition, cells can reciprocally utilize inside out signaling to modulate the ECM. Extracellular matrix receptors include integrins, dystroglycan and syndecans. The most studied ECM receptors, integrins, have been shown to modulate cell growth, differentiation, adhesion, motility and apoptosis. Integrins and dystroglycan have been implicated as key mediators of basement membrane formation (Henry and Campbell
14 2 1998; Klass, Couchman et al. 2000; Lohikangas, Gullberg et al. 2001). However, the function of dystroglycan within the epithelium is currently unknown. Dystroglycan The extracellular matrix receptor dystroglycan was discovered as part of the dystrophin-glycoprotein complex (DGC) in skeletal muscle (Ervasti, Ohlendieck et al. 1990; Ervasti and Campbell 1991) where it is required for stability of the sarcolemma. Mutations in the proteins of the DGC result in muscular dystrophies, diseases characterized by muscle wasting and weakness. Thus, dystroglycan has primarily been studied in the skeletal muscle. However, dystroglycan is expressed in many tissues including epithelial, neural and adipose tissues where its function remains largely unknown (Durbeej, Henry et al. 1998). Structure and Biosynthesis of Dystroglycan Dystroglycan (DG) is composed of two non-covalently linked subunits transcribed from a single gene, DAG1. DAG1 is highly conserved across vertebrate species and maps to human chromosome 3p21 and mouse chromosome 9. In humans, DAG1 has two exons and one large intron. The 5.8kb transcript translates to an 895 amino acid, 160kDa, propeptide that undergoes autoproteolysis to generate the 120 kda DG and 43 kda DG subunits (Ibraghimov-Beskrovnaya, Ervasti et al. 1992; Ibraghimov-Beskrovnaya, Milatovich et al. 1993; Holt, Crosbie et al. 2000; Sciandra, Bozzi et al. 2009). Although the significance of this cleavage is not known, the cleavage site is not conserved between vertebrates and invertebrates. Expression of the uncleaved propeptide leads to dystrophic changes in a mouse model indicating cleavage is important for DG function (Jayasinha, Nguyen et al. 2003). Following propeptide cleavage, the
15 3 and subunits form a non-covalent linkage at the C-terminus of DG and N-terminal ectodomain of DG. Following posttranslational processing in the secretory pathway, DG localizes to the plasma membrane. In tissues, localization occurs at the basal side of the cell adjacent to the basement membrane (Ibraghimov-Beskrovnaya, Ervasti et al. 1992; Durbeej, Larsson et al. 1995; Belkin and Smalheiser 1996). Mature dystroglycan peptides are composed of several domains. DG is a peripheral membrane protein with a dumbbell-like shape owing to its two globular domains at the N- and C- termini with an elongated mucin-like domain in the center (Brancaccio, Schulthess et al. 1995; Brancaccio, Schulthess et al. 1997). DG is an integral membrane protein consisting of a transmembrane domain and a PPXY binding motif in the C-terminal cytoplasmic tail. In addition, DG has a nuclear localization signal in the juxtamembrane region of the cytoplasmic domain suggesting it may localize to the nucleus (Oppizzi, Akhavan et al. 2008). Both and DG subunits have been shown to undergo proteolytic cleavage events independent from the proprotein cleavage. DG is cleaved at the N-terminal end by convertase-like activity. It is unclear when in the maturation processes cleavage occurs and what effect it has on the function of DG. DG undergoes proteolytic processing by matrix metalloproteases generating a 30kDa fragment thought to be a degradation product. Proteolysis of DG may disrupt its stability or interaction with and stability of DG resulting in degradation of the protein (Yamada, Saito et al. 2001). Dystroglycan Posttranslational Modifications Proteins undergo many forms of posttranslational modifications. The most common and complex modification is glycosylation as more than one half of proteins are glycosylated (Apweiler, Hermjakob et al. 1999). Protein glycosylation has been shown to have many functions including roles in protein conformation and stability, receptor-
16 4 mediated signaling and protein interactions. There are two types of glycosylation defined by their linkage to amino acid residues. N-glycans have an N-acetylglucosamine (GlcNAc) at the reducing end that links to asparagine (Asn) residues of proteins. O- glycans link to serine (Ser) or Threonine (Thr) residues and have diverse protein-glycan linkages and complex glycan structures. The most common type of O-glycan linkage is the mucin-type consisting of an N-acetylgalactosamine (GalNAc) at the reducing end. Other O-glycosylation types include O-linked fucose, glucose, N-acetylglucosamine (GlcNAc), and mannose. O-mannose glycosylation is rare in vertebrates and so far has only been found in a few glycoproteins in brain, nerve and skeletal muscle (Yuen, Chai et al. 1997; Endo 1999). The biosynthesis for O-glycans occurs after protein folding in the endoplasmic reticulum and golgi apparatus. As proteins move through the secretory pathway carbohydrate moieties are added in a sequential order. Interestingly, there is no template for the glycosylation of proteins and for most O-glycosylation types a recognition consensus sequence has not been determined for the addition of the first sugar residue (Julenius, Molgaard et al. 2005). O-glycans appear to be generated by the specificity of the glycosyltransferases. This specificity can be influenced by the presence of cations, chaperone proteins or the specific structure of the protein substrate itself (Esko and Zhang 1996; Petrova, Koca et al. 2001; Ju and Cummings 2002). The molecular weight of the DG core protein is predicted to be 72 kda, yet in skeletal muscle it appears at 120 kda following electrophoresis as a result of posttranslational modifications including extensive glycosylation (Ibraghimov- Beskrovnaya, Ervasti et al. 1992; Ervasti and Campbell 1993; Ervasti, Burwell et al. 1997). N-linked glycosylation sites are present at the C- and N-termini of DG as well as the ectodomain of DG. N-glycans have been shown to be required for the posttranscriptional processing of DG, particularly for the proteolytic cleavage of the propeptide, but are not required for ligand binding (Holt, Crosbie et al. 2000). The most
17 5 extensive glycosylation occurs at the mucin domain of DG. This region is highly glycosylated with O-linked glycans, some of which are required for ligand binding (Smalheiser and Kim 1995; Smalheiser, Haslam et al. 1998). Phosphorylation is also a common post-translational modification. Recent work has shown that DG is phosphorylated and this phosphorylation is required for ligand binding. Interestingly, the phosphorylation event does not occur on the peptide itself but is instead present on O-mannosyl glycans attached to Thr 317 and Thr 319 of the mucin domain. The putative glycosyltransferase LARGE is responsible for the addition of ligand binding moieties to these phosphorylated O-mannose residues (Yoshida- Moriguchi, Yu et al. 2010; Hara, Kanagawa et al. 2011). DG is also phosphorylated at tyrosine residue 892 in the PPxY domain at the C-terminal tail. This phosphorylation is mediated by c-src tyrosine kinase and may have a role in regulating protein-protein interactions (James, Nuttall et al. 2000; Sotgia, Lee et al. 2001). DG Glycosyltransferases To date, mutations in six known and putative glycosyltransferases (POMT1, POMT2, POMGnT1, Fukutin, FKRP and LARGE) have been implicated in DG glycosylation. These glycosyltransferases function in the biosynthesis of the O-mannosyl glycans required for ligand binding. Mutations in these enzymes cause muscular dystrophies due to hypoglycosylation of DG and loss of laminin binding. The similarity of phenotypes in muscular dystrophy patients with mutations in these glycosyltransferases suggests they act in a converging pathway. The glycosyltransferases POMT1 and POMT2 are type III membrane ER-resident proteins. They have amino acid sequences similarity to yeast protein O- mannosyltransferase, and are thought to initiate the first step of the biosynthetic pathway for O-mannosyl glycans by adding O-linked mannose directly to DG (Willer, Valero et
18 6 al. 2003; Manya, Chiba et al. 2004). POMGnT1 is a known O-linked mannose 1,2-Nacetylglucosaminyltransferase 1 located in the golgi apparatus. It acts downstream of POMT1/2 in the synthetic pathway (Takahashi, Honda et al. 2000; Yoshida, Kobayashi et al. 2001). Fukutin is a golgi apparatus protein and has catalytic domain similarity to phosphoryl-sugar transferases as well as a DXD motif characteristic of glycosyltransferases. Mutation results in hypoglycosylation of DG, however, no enzymatic activity has been confirmed (Kobayashi, Nakahori et al. 1998; Aravind and Koonin 1999; Esapa, Bentham et al. 2003). FKRP (fukutin related protein) was cloned for its similarity to the putative catalytic domain of fukutin. Mutations have been found in muscular dystrophy patients but no enzymatic activity has been shown (Brockington, Blake et al. 2001; Beltran-Valero de Bernabe, Voit et al. 2004). The putative glycosyltransferase LARGE has been of particular interest in the muscular dystrophy field. Studies have shown LARGE expression can bypass mutations in other glycosyltransferases in cells isolated from patients with muscular dystrophy (Barresi, Michele et al. 2004). In addition LARGE has been implicated as the glycosyltransferase necessary for the modification of DG required for ligand binding. LARGE has a wide expression pattern with highest expression levels in brain and cardiac muscle (Peyrard, Seroussi et al. 1999). LARGE was first identified as a common rearrangement in meningioma tumorigenesis (Peyrard, Seroussi et al. 1999). Although glycosyltransferase activity has not been directly demonstrated to date, LARGE is a type II membrane-anchored protein with a short cytoplasmic tail typical of glycosyltransferases. It is unique in its coiled coil motif and two putative catalytic domains (Grewal, Holzfeind et al. 2001). The first catalytic domain contains two DXD motifs, characteristic of glycosyltransferases, and has amino acid similarity to the bacterial WaaJ family of glycosyltransferases (Coutinho, Deleury et al. 2003). The second catalytic domain has one catalytic motif with a single DXD domain and has an amino acid similarity to a -1,3-N-acetylglucosaminyltransferase (ignt), a family of
19 7 glycosyltransferases responsible for part of the erythrocyte I antigen (Grewal and Hewitt 2002). In addition, LARGE is localized to the golgi apparatus, the primary site for protein glycosylation. Proteins with mutations in both DXD motifs of the first catalytic domain localize to the golgi apparatus but are unable to glycosylate DG suggesting both motifs are required for glycosyltransferase activity. Mutation of the DXD motif in the second catalytic domain causes aberrant localization and thereby loss of function leaving the functional effects of this domain unclear. The transmembrane domain and C-termini are also required for correct localization (Brockington, Torelli et al. 2005). In addition to studies showing that gain or loss of LARGE function correlates with DG glycosylation, LARGE interacts with DG and the N-termini of DG is required for this interaction (Kanagawa, Saito et al. 2004). Due to their roles in muscular dystrophies, the glycosyltransferases required for functional glycosylation of DG in muscle have been identified but little is known about glycosyltransferases specific to other tissues. Dystroglycan is widely expressed in neural, endothelial, adipose and epithelial tissues as well as muscle. Variability in the size of DG across tissues indicates it may be differentially glycosylated and therefore have tissue specific functions. Although dystroglycan is widely expressed, its expression levels vary greatly between tissues (Ibraghimov-Beskrovnaya, Ervasti et al. 1992; Ibraghimov- Beskrovnaya, Milatovich et al. 1993; Durbeej, Henry et al. 1998). Additionally, disease phenotypes in the myd mouse and muscular dystrophy patients are restricted to muscle, brain and eye indicating additional glycosyltransferases may compensate in other tissues. Recently, a second member of the LARGE family has been identified. The LARGE2 (LARGE-like, Gyltl1b) gene has 64% amino acid identity to LARGE and is the result of a gene duplication event in the vertebrate lineage (Grewal, McLaughlan et al. 2005). Analogous to LARGE, LARGE2 is a type II membrane-anchored protein with two catalytic domains. However, mammalian LARGE2 lacks a coiled-coil domain and has minimal amino acid similarity to the N-terminal cytoplasmic and transmembrane
20 8 domains of LARGE (Grewal, McLaughlan et al. 2005). LARGE2 has also developed an alternative initial coding exon and is 100 fold smaller than LARGE due to differences in intron size. Although little is known about LARGE2, studies have shown it localizes to the golgi apparatus and has the ability to functionally glycosylate DG in an overexpression system. Glycosyltransferase activity has not yet been demonstrated (Brockington, Torelli et al. 2005). Interestingly, LARGE 2 glycosylated DG runs at a higher molecular weight than LARGE following electrophoresis and these proteins do not cooperate in an overexpression system (Fujimura, Sawaki et al. 2005). LARGE2 also has a more restricted tissue expression profile and is not expressed in brain but is detectable in skeletal muscle at much lower levels than LARGE (Fujimura, Sawaki et al. 2005; Grewal, McLaughlan et al. 2005). Dystroglycan Interactions As an extracellular matrix receptor, DG has the ability to interact with numerous proteins within and outside of the cell. As discussed previously, DG is highly glycosylated in the mucin domain and these phosphorylated O-glycans are required for ligand binding. DG has multiple ligands and its binding partner is dependent on tissue and cell type. In most tissues, DG localizes to the cell surface in an orientation adjacent to the basement membrane where it binds matrix proteins laminin, perlecan and agrin in a Ca 2+ dependent manner (Ibraghimov-Beskrovnaya, Ervasti et al. 1992; Ervasti and Campbell 1993; Bowe, Deyst et al. 1994; Campanelli, Roberds et al. 1994; Gee, Montanaro et al. 1994). DG binds to laminin through interactions with LG domains in the alpha chain (Talts, Andac et al. 1999). DG binds to laminins 111/123, 211 and 511/521. Laminin 332 undergoes cleavage of its LG 4-5 domains during processing and is therefore unable to bind to DG. Dystroglycan also binds to its ligands perlecan, agrin and neurexin through their LG-like domains.
21 9 As part of the dystrophin-glycoprotein complex in muscle, DG interacts with dystrophin thereby linking DG to the cytoskeleton. This interaction occurs through the PPXY motif at the C-terminus of DG and the WW-like domain at the C-terminal end of dystrophin or its homolog utrophin (Ervasti and Campbell 1991; Chung and Campanelli 1999). In addition to serving as a link to the cytoskeleton, DG has also been shown to bind to various signaling molecules. Interestingly, phosphorylation of DG by c-src tyrosine kinase has been shown to disrupt the binding of dystrophin and promote interactions with proteins containing SH2 and SH3 domains, thereby acting as a molecular switch. The adaptor protein Grb2 binds to DG at the C-terminal proline-rich domain (Yang, Jung et al. 1995). In bovine brain DG forms a complex with focal adhesion kinase (FAK) through the SH2 domain of Grb2(Cavaldesi, Macchia et al. 1999). Dystroglycan s binding partners also include Erk and MEK (Spence, Dhillon et al. 2004), a postsynaptic protein rapsyn (Cartaud, Coutant et al. 1998) and members of the caveolin family (Sotgia, Lee et al. 2000). DG has also been shown to act as a key signaling component of the notch pathway (Sirour, Hidalgo et al. 2011) Although DG has been shown to directly interact with a variety of signaling molecules and has been implicated in a number of signaling pathways, the precise role of DG in these pathways and their functional relevance remain unclear. Dystroglycan Function in Development The recent identification of a muscular dystrophy patient with a loss of function mutation in the DAG1 gene indicates the ligand binding function of DG may not be required for development in humans yet studies in mice support a role for DG in embryonic development (Hara, Balci-Hayta et al. 2011). However, embryonic lethality at day 6.5 in Dag1 null mice has been attributed to disruption of a rodent specific extraembryonic basement membrane known as Reichert s membrane (Williamson, Henry
22 10 et al. 1997). Reicherts membrane forms between the trophoblast and parietal endoderm following implantation. Aberrant localization of matrix proteins collagen IV and laminin as well as dystroglycan in the area of Reicherts membrane indicate disruption of its structure. Additionally, the presence of maternal red blood cells within the yolk sac indicates disruption of its function. Interestingly, a different embryonic basement membrane in these mice appears normal suggesting DG may be required for the formation of some but not all basement membranes. Mice expressing the Mox-2 Cre transgene (MORE-DG null mice) have deletion of DG in all embryonic tissues but maintain DG expression in the extraembryonic membranes including Reichert s membrane (Satz, Barresi et al. 2008). These mice have normal histology at day E7.5 and survive to 48hrs to 4 weeks after birth supporting the disruption of Reichert s membrane as the cause of embryonic lethality in DG null mice. Additional studies in embryoid bodies have indicated a role for DG in the formation of basement membranes (Henry and Campbell 1998). DG is required for laminin-1 binding in mouse embryonic stem cells, further suggesting DG has important roles in basement membrane formation (Henry, Satz et al. 2001). DG Function in Muscular Dystrophies The identification and characterization of the dystrophin-glycoprotein complex in muscular dystrophy patients has provided valuable insight into the function of dystroglycan (Hara, Balci-Hayta et al. 2011). While muscular dystrophies are caused by disruption of any one of several proteins in the DGC, a subgroup of muscular dystrophies, dystroglycanopathies, are defined by disruption of dystroglycan function. Dystroglycanopathies are characterized by hypoglycosylation of DG and loss of laminin binding due to mutations in the glycosyltransferases that function in the biosynthesis of the O-mannosyl glycans required for ligand binding (Chiba, Matsumura et al. 1997;
23 11 Cohn, Henry et al. 2002; Michele, Barresi et al. 2002). To date, mutations in six known and putative glycosyltransferases (POMT1, POMT2, POMGnT1, Fukutin, FKRP and LARGE) have been found in patients with several types of congenital and limb girdle muscular dystrophies. These patients exhibit a broad spectrum of pathologies including defects in muscle, brain and eye. Whether these glycosyltransferases have additional targets remain unknown. However, the overlapping phenotypes in these patients and in mouse models suggest DG may be the only target. The recent identification of a muscular dystrophy patient with a mutation in DAG1 offers new insight into function of DG in humans. This patient has a missense mutation (Thr192Met) that results in hypoglycosylation of DG due to an inability to bind LARGE in a mouse model. This patient presents with limb girdle muscular dystrophy and cognitive impairment and creates a new class of primary dystroglycanopathies (Hara, Balci-Hayta et al. 2011). As was utilized with this patient, mouse models can recapitulate most of the phenotypes seen in muscular dystrophies providing a way to discern the underlying molecular mechanisms. Currently, tissue specific knockout mice provide the best model to define the function of DG in a wide range of tissues. DG Function in Non-Epithelial Tissues Mice with skeletal muscle specific deletion of DG display a mild form of muscular dystrophy similar to that seen in some muscular dystrophy patients (Cohn, Henry et al. 2002). The presence of muscle degeneration, loss of membrane integrity and variations in muscle fiber size indicate loss of DG and thereby localization of the DGC at the sarcolemma membrane can cause muscular dystrophy. DG was not required for basement membrane ultrastructure or laminin deposition in skeletal muscle. DG expression was retained in the satellite cells (stem cells) of the mice resulting in
24 12 regeneration of muscle fibers and prevention of fibrosis and fat replacement. Thus, DG not only has a role in stability of the sarcolemma but also has an important role in satellite cell function. This was confirmed in the MORE-DG null mice that express DG in all extraembryonic tissue but not in embryonic tissues, thereby bypassing embryonic lethality(satz, Barresi et al. 2008). These mice have DG null satellite cells and exhibit severely impaired regeneration of muscle fibers. Mice chimeric for DG expression have a severe muscular dystrophy phenotype that may also result from loss of DG in satellite cells. In addition, these mice have abnormalities in the neuromuscular junctions.(cote, Moukhles et al. 1999). In vitro studies have also indicated a role for DG in the aggregation of acetylcholinesterase receptors (Jacobson, Cote et al. 2001). These studies indicate that in skeletal muscle DG is required for the localization of the DGC, sarcolemma membrane stability, maintenance of satellite cell regenerative capacity and formation of synapse at the neuromuscular junctions. Many patients with muscular dystrophies have associated brain defects. DG is expressed in the central nervous system and loss of DG function may contribute to brain abnormalities in these patients as well as those present in skeletal muscle. DG is normally localized to glial endfeet, which form basement membranes adjacent to the pial surface of the brain and spinal cord (glia limitans) and along cerebral blood vessels. DG is also expressed in neurons. DG deletion in the central nervous system (GFAP Cre) specifically, in the radial glia and astrocytes, results in pathologies similar to those seen in congenital muscular dystrophy (CMD) patients. DG is required for the localization of the DGC to the membrane of astrocyte foot processes as seen in skeletal muscle. Deletion of DG results in fusion of cerebral hemispheres and cerebellar folia, disruption of neuronal layering in the cerebral cortex and aberrant neuronal migration. In addition, the glia limitans is disrupted and laminin binding is significantly reduced indicating DG is required for formation or maintenance of the basement membrane. DG may also have a
25 13 role the central nervous system synapsis similar to its role in neuromuscular junctions (Moore, Saito et al. 2002). Deletion of DG throughout all cell types of the mouse brain due to epiblast specific Cre expression (Mox2-Cre) also results in disruption of the integrity of the glia limitans, loss of laminin binding ability, defects in neuron migration and fusion of the cerebral hemisphere. However these mice also display more severe malformations including hydrocephalus. The pathological variability in these mouse models may result from later expression of the GFAP-Cre transgene during development (E7.5 vs E14.5) or outside effects due to loss of DG from other tissues in the Mox2-Cre mice (Tallquist, Weismann et al. 2000). Studies in the peripheral nervous system have shown DG is important for myelination and sodium channel stabilization in the peripheral nerve (Saito, Moore et al. 2003). Muscular dystrophy patients also often have abnormalities of the eye. DG is localized to the glial endfeet of the inner limiting membrane and adjacent to the vasculature as well as at ribbon synapses of rod and cone photoreceptors in the retina (Koulen, Blank et al. 1998; Blank, Koulen et al. 1999). Deletion of DG in the epiblast causes ocular malformations in the anterior and posterior chambers of the eye in addition to brain abnormalities. Eye defects included microophthalmia, herniated lens and iris tissues and fusion of the lens and corneal tissues. Defects of the eye correlate with disruption of the basement membrane in the cornea and retina indicating DG has an important role in the morphogenesis of the eye (Satz, Barresi et al. 2008). In the retina, DG anchors dystrophin and an inward rectifying K+ channel to the glial endfeet. Disruption results in attenuation of the electroretinogram -wave, a defect seen in CMD patients. Interestingly, deletion of DG is required for the attenuation of the -wave indicating DG intracellular interactions are necessary for retinal physiology but not for laminar structure (Satz, Philp et al. 2009).
26 14 These studies have elucidated important functions for DG in the CNS, PNS, muscle and eye. DG has been shown to have many functions including roles in basement membrane assembly and stabilization as well as tissue morphogenesis. It has additional roles in neuron migration, synaptic plasticity, retinal physiology and myelination and nodal architecture of the peripheral nerve. Interestingly, little is known about its function in tissues including the epithelium. DG Function in the Epithelium At initiation of this thesis work, studies into the function of DG in epithelial tissues consisted of in vitro assays and deletion in Drosophila melanogaster. No epithelial specific knockout mouse models had been published. During the course of this thesis work and following the publication of my probasin cre prostate specific DG knockout mouse, mammary and kidney specific models were published. The phenotypes of these mouse models will be addressed in the summary and future directions section. Compared to muscle, little is known about DG function in epithelial tissues. It is expressed in many non-muscle cells including epithelial, glial and adipose cells which share the property of being in direct contact with basement membranes. In muscle, DG functions as part of the dystrophin-glycoprotein complex (DGC). In epithelial tissues, DGC complexes generally include a combination of family members and isoforms of dystrobrevin, utrophin, syntrophin and dystroglycan (Kachinsky, Froehner et al. 1999). Sarcoglycan and sarcospan are expressed in some epithelia but have not been shown to be complexed with dystroglycan in the DGC of epithelial tissues (Durbeej and Campbell 1999). The function of DG in the epithelium, as a part of the DGC or independently, remains largely unknown. In the epithelium, several studies have implicated DG in basement membrane formation. DG is required for the formation of some basement membranes during
27 15 development (Williamson, Henry et al. 1997; Henry and Campbell 1998). In embryonic stem cells, it is responsible for laminin binding while 1 integrin and perlecan are necessary for matrix formation. Mammary epithelial cells also require DG for initial laminin binding which then enables 1 integrins to polymerize the matrix suggesting a role in basement membrane formation (Weir, Oppizzi et al. 2006). In Xenopus laevis, DGN-1 deletion disrupts laminin deposition and fibronectin organization as well as epithelial differentiation in the epidermis (Sirour, Hidalgo et al. 2011). Extracellular matrix receptors have essential roles in tissue morphogenesis including cell differentiation and the establishment of polarity. Antibody blocking experiments with cultured embryonic kidney have shown a role for DG in epithelial branching morphogenesis (Durbeej, Larsson et al. 1995). This has been extended to lung and salivary gland and is dependent on dystroglycan binding to laminin (Durbeej, Talts et al. 2001). In mammary epithelial cells, DG is required for lactogenic differentiation and survival of mammary epithelial cells (Sgambato, Di Salvatore et al. 2006). Furthermore, DG is necessary for laminin-111 induced polarity and differentiation in mammary epithelial cells independent of the cytoplasmic domain, supporting the idea that DG functions as a co-receptor with 1 integrins (Weir, Oppizzi et al. 2006). Work in intestinal epithelial cells has also demonstrated DG/laminin interactions regulate integrin/laminin binding (Driss, Charrier et al. 2006). One of the best in vivo models for polarity is the development of the epithelium in follicle cells of the Drosophila ovary. In this model, DG was required for planar polarity of basal stress fibers. This phenotype is also seen in dystrophin mutants indicating DG/dystrophin act together in this pathway (Deng, Schneider et al. 2003; Schneider, Khalil et al. 2006; Mirouse, Christoforou et al. 2009). Additional work has shown DG does not function in the apical basal polarity pathway of Drosophila follicle cells. However dual deletion of DG and a gene required for mitochondrial function disrupts apical basal polarity. Disruption of mitochondrial
28 16 function leads to a low energy state within the cell suggesting DG acts in the apical basal polarity pathway under conditions of energetic stress. Previous studies have shown adenosine monophosphate stimulated protein kinase (AMPK) regulates polarity under energetic stress conditions by phosphorylating myosin light change. DG when bound to perlecan is required for AMPK pathway activation and maintenance of polarity demonstrating a role for DG in polarity under energetic stress. Furthermore, loss of DG causes increased mitochondrial oxidative metabolism leading to a greater cold tolerance and a preference for lower temperatures. (Takeuchi, Nakano et al. 2009). Dystroglycan acts as a mechanotransducer in rat lung alveolar epithelial cells (AECs). Following cyclic stretch during normal respiration and during mechanical ventilation, laminin-311 via DG transduces a mechanical stretch signal across the membrane resulting in activation of the MAPK pathway. Activation of this pathway may inhibit cell-cycle arrest and protect against apoptosis thereby protecting the lung epithelium from stretch-induced injury (Jones, Lane et al. 2005). DG is also required for activation of the metabolic sensing AMPK signaling pathway following cyclic mechanical stretch of AECs, potentially protecting against injury and preserving energy stores (Budinger, Urich et al. 2008). These studies support a role for DG in both mechanotransduction and metabolic signaling pathways. As discussed previously, DG interacts with a variety of signaling molecules. In the epithelium, DG activates the STAT5, MAPK and AMPK pathways yet its precise role and the functional outcomes of DG signaling are unclear. Interestingly several studies have shown DG is not required for DG function indicating DG functions outside of direct activation of signaling pathways. DG may indirectly modulate signaling pathways by acting as a co-receptor to integrins and growth factor receptors (Driss, Charrier et al. 2006; Leonoudakis, Singh et al. 2010). Alterations in basement membranes, polarity and signaling pathways are characteristic of cancer development and progression. DG has been implicated in these
29 17 processes. Therefore, aberrant DG function may contribute to cancer progression. Reduced expression of DG has been demonstrated in a variety of cancers yet the molecular mechanism and functional consequences remain unknown. The Extracellular Matrix and Cancer Cancers develop through aberrant expression of genes resulting from genetic mutations, chromosomal rearrangements and/or altered epigenetics. Cancer cells possess specific characteristics known as the hallmarks of cancer that facilitate cancer development and progression. These include uncontrolled growth, resistance to apoptosis, and the ability to invade surrounding tissue. It is becoming clear that altered cell behavior is not only a result of genetic changes within the cell but is also due to regulation by the microenvironment (Hanahan and Weinberg 2000; Hanahan and Weinberg 2011). Therefore, in carcinomas of tissues such as the prostate, these traits may result from altered cell-ecm interactions, composition of the ECM, degradation of matrix proteins and enhanced cell motility. In carcinomas, the ability of the cancer cells to proliferate in an anchorage independent manner, become motile and invade through basement membranes facilitates the development of metastatic disease. Metastasis occurs when a primary tumor disseminates to one or more secondary sites. This process occurs in a sequential set of events known as the metastatic cascade. Initially, cancer cells proliferate and recruit a vasculature at the primary site. This is followed by degradation of the basement membrane and invasion of cells into the surrounding tissue. Following intravasation and extravasation of the vasculature, cells proliferate and recruit a vasculature at a secondary site. Many of these steps involve modulation of the ECM and cell/ecm interactions. Cell/ECM interactions are disrupted during the initial stages of tumor growth when cells no longer require interactions with the ECM for growth. Degradation of the
30 18 epithelial basement membrane and modulation of matrix receptors is required for invasion into and migration across the surrounding extracellular matrix. In addition to removing a structural barrier, basement membrane degradation releases growth factors and other proteins that stimulate the proliferation and migration of the cancer cells. Cells traverse the vasculature by expressing surface molecules specific to the endothelium. Once at the secondary site, cells must again modulate the expression of their matrix receptors to adapt to a new matrix composition. Although the steps of metastasis are fairly well understood, the molecular mechanisms contributing to tumor progression and metastasis remain unclear. Prostate Cancer The prostate is part of the male urogenital system in most mammals and is located at the base of the bladder surrounding the urethra. Its normal function is to store and secret an alkaline fluid that is a portion of the seminal fluid. The development of the prostate is dependent on androgens and reciprocal interactions between the urogenital sinus epithelium and mesenchyme. During development prostatic buds develop from the urogenital sinus and invade into the surrounding urogenital mesenchyme following exposure to androgens. Branching morphogenesis occurs in the second and third trimesters and is completed by birth in humans (Timms, Mohs et al. 1994). In contrast, branching occurs postnatally in rodents (Sugimura, Cunha et al. 1986). Since the prostate is a hormone dependent tissue, androgens are required for maintenance of cell growth and survival in the adult prostate (Donjacour and Cunha 1988). The human prostate is divided into three zones; central, peripheral and transitional. The vast majority of prostate cancers are present in the peripheral zone which is also the largest zone. The prostate is composed of glands surrounded by a dense fibromuscular stroma. Its pseudostratified epithelium is composed of three cell types; basal, luminal and neuroendocrine. Prostate cancers express
31 19 luminal cell markers although both basal and luminal cells have the ability to act as cancer initiating cells in mouse models (Wang, Kruithof-de Julio et al. 2009; Goldstein, Huang et al. 2010; Lawson, Zong et al. 2010). The cancer initiating cell type in prostate cancer remains unclear. Prostate cancer is the most commonly diagnosed noncutaneous cancer in the U.S. It is the second leading cause of cancer-related deaths in men. Prostate cancer is primarily diagnosed in men over the age of 45 with the average age of diagnosis at 70. It is estimated that 1 in 6 men will be diagnosed with prostate cancer in their lifetime. However, the late onset and characteristic slow growth of prostate cancer means many men will die from other health issues. Of the men that progress to advanced metastatic prostate cancer, 30%-60% of those diagnosed, an estimated 90% will develop bone metastases which are the primary cause of pain and morbidity in prostate cancer patients (Bubendorf, Schopfer et al. 2000; Rubin, Putzi et al. 2000). Prostate cancers are characterized by significant histological heterogeneity and the presence of multiple foci within the prostate leading to difficulty in tumor grading and treatment selection. Prostate cancers arise from alterations in gene expression through mutations, chromosomal rearrangements or epigenetic modulations. These alterations transform the glandular epithelium to preneoplastic lesions that eventually progress to invasive carcinoma and metastases. Prostate cancer most often metastasizes to the adrenal glands, liver and bone. However, even metastatic prostate cancer cells retain their androgen dependence providing an avenue for therapy. Treatment of early stage non-metastatic prostate cancer generally consists of prostatectomy and radiation. Due to the common advanced age at diagnosis and the characteristic slow growth of prostate tumors, watchful waiting or monitoring of the tumor without therapeutic intervention, has recently gained support. For patients with advanced metastatic prostate cancer, the primary course of treatment is androgen deprivation therapy (ADT). ADT induces apoptosis and involution of the prostate
32 20 resulting in reduction of tumor size. However, eventually almost all patients will have tumor recurrence and progression to androgen-independent disease state. Androgen independent prostate cancer (AIPC) occurs when cancer cells modulate the androgen signaling pathway, allowing them to survive and proliferate under androgen deprivation conditions. Cells alter the androgen receptor (AR) pathway through up-regulation of AR, mutations that decrease AR ligand specificity or that activates AR in the absence of a ligand. In addition, some cells activate androgen-receptor-independent signaling pathways. While ADT provides patients with additional time and quality of life, it also drives progression of the cancer to an androgen independent state for which there is no cure. The average life expectancy for patients with AIPC is months (Smaletz and Scher 2002). Despite current research efforts, clinicians remain unable to differentiate patients with indolent disease from those who will progress to metastatic disease. In addition, little progress has been made in the treatment of advanced prostate cancer and morbidity rates have remained largely unchanged. DG Function in Cancer Disruption of epithelial/ecm interactions promotes cancer progression through aberrant regulation of cellular processes. As a mediator of cell-matrix interactions, loss of DG has been linked to a variety of cancers. In breast and prostate cancer, loss of DG immunoreactivity with DG specific antibody 8D5 (Novocastra) correlates with increased tumor grade (Henry, Cohen et al. 2001) and has prognostic significance in breast cancer (Sgambato, Migaldi et al. 2003). This has now been extended to include oral, cervical and vulvar squamous cell carcinomas as well as pediatric sarcomas and neural cancers indicating loss of DG could act as a conserved molecular mechanism for cancer progression (Jing, Lien et al. 2004; Sgambato, Tarquini et al. 2006; Martin, Glass et al. 2007; Sgambato, De Paola et al. 2007). It is unclear whether loss of
33 21 immunoreactivity to the antibody IIH6 represents loss of the DG subunit through proteolytic cleavage and shedding of DG or hypoglycosylation of DG (Losasso, Di Tommaso et al. 2000; Singh, Itahana et al. 2004). Loss of DG has also been demonstrated and may act as an additional mechanism for loss of DG function in cancer (Henry, Cohen et al. 2001). In panels of various metastatic cancer cell lines, hypoglycosylated DG and DG were present at the cell surface indicating reduced glycosylation may be a central mechanism for loss of DG function in cancer (Bao, Kobayashi et al. 2009; de Bernabe, Inamori et al. 2009). Loss of DG glycosylation in breast cancer as well as cervical and lung cancers was shown to be due to epigenetic silencing of the glycosyltransferase LARGE. In prostate cancer, DG glycosylation status can be influenced by the glycosyltransferase 3GnT1, which may collaborate with LARGE and LARGE2 to glycosylate DG. However, previous studies in breast and prostate cancer lines have also shown reduced expression or complete loss of the DG subunit and an inability to retain DG at the cell surface with overexpression of DG (Muschler, Levy et al. 2002; Sgambato, Camerini et al. 2004; Sgambato, De Paola et al. 2007). Therefore, loss of DG expression can occur by a number of mechanisms in cancer cell lines. Relatively little is known regarding DG function in cancer progression. Overexpression of DG in normal and breast cancer cell lines inhibited proliferation and reduced tumorigenicity whereas loss of DG increased proliferation (Muschler, Levy et al. 2002). In prostate cancer, loss of DG glycosylation correlated with enhanced cell migration and tumor growth (Bao, Kobayashi et al. 2009). These studies implicate DG as a negative regulator of tumor growth through an unknown mechanism. DG has also been suggested to have roles in cytoskeletal organization, proliferation and polarity in epithelial cells and has been shown to inhibit growth of glioma cell lines (Muschler, Levy et al. 2002; Calogero, Pavoni et al. 2006). These studies suggest DG may function through regulation of cell signaling pathways and reorganization of the cytoskeleton.
34 22 However, the mechanism by which loss of DG promotes cancer progression remains unclear. Relevance and Focus The extracellular matrix (ECM) is composed of an array of molecules including matrix proteins, growth factors and cytokines that give support to and provide information about the microenvironment to cells. Cell/ECM interactions are essential to maintaining cellular homeostasis and are mediated by ECM receptors. Disruption of these receptors results in aberrant communication between epithelial cells and the ECM, a hallmark of cancer. However, the mechanisms behind alterations in matrix receptor function and in what manner these disruptions contribute to prostate cancer progression is poorly understood. Elucidating the underlying molecular mechanisms will contribute to advances in diagnosis and treatment. DG has primarily been studied in muscle but is expressed in many other tissues including the epithelium. Previous studies have indicated roles for DG in epithelial proliferation, differentiation, polarity and basement membrane formation. Loss of DG expression and function has been shown in many cancers including prostate, yet little is known about dystroglycan s role in the epithelium and thus how its disruption promotes prostate cancer progression. Therefore, the focus of my thesis work was to determine the function of DG in the normal prostate epithelium and prostate cancer progression. Specifically, we have generated individual prostate luminal cell and basal cell specific DG knockout mice to determine the function of DG in the prostate epithelium. In addition, we have investigated the molecular mechanism by which DG is hypoglycosylated in prostate cancer. This work reveals a novel mechanism for DG hypoglycosylation in prostate cancer as well as a potentially novel association between
35 23 oncogene expression and DG glycosylation status. These insights may lead to advances in the treatment of prostate cancer.
36 24 CHAPTER II DYSTROGLYCAN IS NOT REQUIRED FOR MAINTENANCE OF THE LUMINAL EPITHELIAL BASEMENT MEMBRANE OR CELL POLARITY IN THE MOUSE PROSTATE Introduction Dystroglycan (DG) was first identified as a component of the dystrophinglycoprotein complex in skeletal muscle (Ervasti, Ohlendieck et al. 1990). It is composed of a highly glycosylated extracellular subunit non-covalently linked to a transmembrane subunit, which are post-translationally derived products of a single gene (Ervasti and Campbell 1991; Ibraghimov-Beskrovnaya, Ervasti et al. 1992). DG binds to several extracellular matrix proteins including various laminins and perlecan which are important for both the structural and signaling functions of basement membranes (Talts, Andac et al. 1999). The cytoplasmic domain of DG is tethered to the actin cytoskeleton via linker proteins such as dystrophin and utrophin (Jung, Yang et al. 1995). This domain has also been shown to interact with various signaling molecules, though a clear signaling function for DG has yet to emerge, e.g. (Yang, Jung et al. 1995; James, Nuttall et al. 2000; Ferletta, Kikkawa et al. 2003). In skeletal muscle, disruption of this linkage via mutations in laminin-211 or dystrophin or by inappropriate carbohydrate modifications on DG which are necessary for extracellular matrix binding lead to various forms of muscular dystrophy indicating that this linkage is critical for the integrity of this tissue (Cohn 2005). DG is expressed in many other non-muscle cells including epithelial, neural and adipose cells that all share the property of being in direct contact with basement membranes (Durbeej, Henry et al. 1998). DG expression has been observed in all
37 25 epithelia examined to date including the prostate (Henry, Cohen et al. 2001). In the normal human prostate, the expression of laminins (111/123, 211, 332, 511/521) and perlecan have been documented (Murdoch, Liu et al. 1994; Brar, Dalkin et al. 2003). Of these, all but laminin-332 have been demonstrated as a DG ligand (Ibraghimov- Beskrovnaya, Ervasti et al. 1992; Talts, Sasaki et al. 2000; Ferletta, Kikkawa et al. 2003). Constitutive disruption of DG in the mouse leads to embryonic lethality around E6.5 (Williamson, Henry et al. 1997). Mutant embryos fail to develop Reichert s membrane, one of the first basement membrane structures formed in the rodent embryo. Moreover, embryonic stem cells lacking DG fail to bind laminin or perlecan on their surface and embryoid bodies fail to develop a sub-endodermal basement membrane (Henry and Campbell 1998; Henry, Satz et al. 2001; Kanagawa, Michele et al. 2005). More recently, Weir et al. have demonstrated that DG is required for laminin assembly on the basal surfaces of mammary epithelial cells (Weir, Oppizzi et al. 2006). Together these findings suggest that DG is important for the assembly of basement membranes. Compared to skeletal muscle, far less is known about DG function in epithelia. A requirement for DG function in epithelial cells has been demonstrated in flies, worms and mammals. Function blocking antibody experiments have shown that DG is required for kidney, lung and salivary gland epithelial morphogenesis in mice (Durbeej, Larsson et al. 1995; Durbeej and Ekblom 1997). In Caenorhabditis elegans mutation of DG disrupts the development of the gonadal epithelium, where it may be involved in the maintenance, but not assembly, of its basement membrane (Johnson, Kang et al. 2006). In addition to its aforementioned role in basement membrane assembly/maintenance, other studies have indicated a role for DG in epithelial polarity. Several studies indicate a requirement for DG in the polarity of the Drosophila melanogaster follicular epithelium as well as in cultured mammary epithelial cells (Deng, Schneider et al. 2003; Weir, Oppizzi et al. 2006; Mirouse, Christoforou et al. 2009). Complementary roles for basement membrane laminins and perlecan in the establishment and/or maintenance of epithelial polarity have
38 26 also been defined (Huang, Hall et al. 2003; Li, Edgar et al. 2003; Schneider, Khalil et al. 2006). Thus, an emerging body of work indicates that DG, through its interactions with its ligands may be critically involved in epithelial polarity in certain contexts. Here we assess the expression and function of DG in the mouse prostatic epithelium. Using Cre-lox technology we generated a prostate-specific DG knockout mouse. We show that DG expressed in luminal epithelial cells is not essential for maintenance of the basement membrane composition or ultrastructure and its expression is not required for prostate cellular homeostasis. Furthermore, we show that DG is expressed in the castration-resistant basal cell population of the prostate and in the proximal ducts, currently implicated as a stem cell niche. DG protein expression increases in involuted prostates in association with a highly-folded basement membrane. Residual expression of DG in both castrate and intact mice is due to the lack of Cre expression in the basal cell compartment. Thus, contrary to expectations, we show that DG is not required for the maintenance of the basement membrane or the polarity associated with luminal epithelial cells in the prostate. Residual expression of DG basal/stem cell population of the prostate may contribute to the lack of expected phenotypes in this organ. Materials and Methods Animals All procedures involving animals were performed according to The University of Iowa Animal Care and Use Committee policies. 129.B6 female DGfl/fl mice (Cohn, Henry et al. 2002); kind gift from Dr. Kevin Campbell) were crossed with male B6.D2 PB-Cre4 mice (NCI Mouse Models of Human Cancer Repository). F1 DGfl/wt; Cre+ males were bred to DGfl/wt; Cre- females. The F2 DGfl/fl; Cre+ males were bred to
39 27 DGfl/fl; Cre- females. F3 males were used for analysis. Herein WT refers to animals with a DGfl/+; Cre+ or DGfl/fl; Cre- genotype and KO refers to animals with DGfl/fl; Cre+ genotype. Female 129-Gt(ROSA)26Sor/J mice (Soriano 1999), (The Jackson Laboratories) were also bred to male B6.D2 PB-Cre4 mice. Beta-galactosidase activity was assayed in the F1 generation. For genotyping, tail DNA was extracted (REDExtract- N-Amp Tissue PCR Kit, Sigma) and PCR was performed. All animals were assessed at 6 months of age unless otherwise noted. For castration/regeneration studies, mice at 6 to 9 months of age were anesthetized (100 mg/kg ketamine, 10 mg/kg xylazine) and castrated. Castrated mice were euthanized by CO2 asphyxiation after 21 days. Remaining castrated mice were implanted with testosterone pellets (Innovative Research of America) at 15 mg/kg for 21 days. Testosterone-treated mice were euthanized after 21 days. Antibodies Anti- DG (IIH6, gift from Dr. Kevin Campbell (Ervasti and Campbell 1991), anti- DG (8D5, Novocastra), CK5 (AF138, Covance), Ki-67 (NCL-Ki67p, Novocastra), p63 (clone 4A4, BD Pharmingen), perlecan (MAB1948, Chemicon), pan-laminin (Durham and Snyder 1995); gift from Dr. Jeanne Snyder), laminin 5 (Mahoney, Stappenbeck et al. 2008), (gift from Jeffrey Miner). Histology and Immunofluorescence Analysis Mouse urogenital organs were removed as a unit and fixed in 4% paraformaldehdye for 4 hours at 4 C and then transferred to 30% ethanol overnight. Prostates were dissected from the seminal vesicles and bladder. Prostates were processed through an ethanol to xylene gradient for paraffin embedding using a Ventana Automated
40 28 Tissue Processor. 5 m sections were cut using a Leica RM2125 rotary microtome. Tissues were deparaffinized and hydrated through a xylene to ethanol gradient followed by citrate buffer or proteinase K antigen retrieval. After 1X phosphate buffered saline (PBS) wash, slides were blocked for 1 hour at room temperature (RT) in blocking solution (2% donkey serum, 1% BSA, 0.05% Tween-20, 0.1% Triton X-100 in 0.01M PBS). Slides were incubated with primary antibody in blocking solution overnight at RT. After two 1X PBS washes, slides were incubated with secondary antibody and nuclear stain 4',6-diamidino-2-phenylindole (DAPI) in blocking solution for 1 hour at RT. Slides were washed twice and coverslipped. Number of keratin 5 (K5)-positive cells per 100 DAPI-positive epithelial nuclei were counted for 3 separate areas of the ventral prostate per mouse. WT (n=3) and KO (n=2) mice were assessed at 6 months of age. Resulting data were averaged and analyzed by t-test. Sections were analyzed independently by two individuals, and similar results were obtained. Ki67 positive cells were quantified as described for K5-positive cells. WT (n=3) and KO (n=3) were assessed at six months of age. For hematoxylin and eosin staining, prostate sections were processed under standard conditions. X-gal Histochemistry Mouse urogenital organs were removed and prostates were dissected from the seminal vesicles and bladder. Prostates were fixed for 1 hour at 4 C in 4% paraformaldehyde followed by incubation in 30% sucrose overnight at 4 C. The tissue was then embedded in Optimal Cutting Temperature embedding medium, frozen briefly in liquid nitrogen and stored at -20 C overnight. Tissues were cut using the Zeiss Micron Model HM505E at 10 m. Slides were stored at -80 C. For staining, slides were washed one time in 1X PBS and incubated for 30 minutes at RT in rinse buffer (100 mm sodium phosphate ph 7.3, 2 mm MgCl2, 0.01% sodium deoxycholate, 0.02% NP-40). Following
41 29 aspiration of the rinse buffer, slides were incubated in staining solution (rinse buffer plus 5 mm potassium ferricyanide, 5 mm potassium ferrocyanide and 1 mg/ml X-gal) for 4 hours at 37 C. Slides were rinsed in PBS, counterstained with eosin, and coverslipped. Transmission Electron Microscopy Mouse prostates were fixed with 2% paraformaldehyde/2% glutaraldehyde in PBS, rinsed three times in 0.1 M sodium cacodylate, then postfixed with a solution of 1% OsO4 and 1.25% potassium ferrocyanide in cacodylate buffer. After three rinses in cacodylate buffer, the samples were exposed to a 1% aqueous tannic acid solution for 20 minutes, dehydrated through a graded series of acetones, and embedded in Eponate-12 (Ted Pelling, Inc., Redding, CA). After embedment and polymerization, 110 nm sections were cut using a Leica UC6 Ultramicrotome, and these were stained with 5% uranyl acetate followed by Reynold s lead citrate. Sections were viewed on a JEOL JEM-1230 transmission electron microscope. Protein Isolation and Western Blot Prostates were removed from intact or castrated mice and lobes were individually dissected. Lobes from each mouse were combined and SDS lysis buffer (50 mm Tris- HCl, 100 mm NaCl, 1 mm EDTA, 1% Triton X, dh 2 0, complete protease inhibitors (Sigma)) were added. After incubation for 5 min on ice, prostate were homogenized for 5 seconds. Sample Buffer (2 M sucrose, 2 M Tris-HCl, 1:20 2-Mercaptoethanol and trace amounts of bromophenol blue) was added to the lysates in a 1:1 ratio. Samples were boiled for 5 minutes and stored at -80 C. Protein quantification was carried out using the BioRad Protein Assay following the manufacturer s instructions. Protein lysates (40 g) were loaded on a 4-20% gradient, Tris-HCl, 10 well, 50 l ReadyGel and electrophoresed
42 30 for 1 h at 150 volts. Protein was then transferred onto a polyvinylidene fluoride membrane at 350 ma for 1 h on ice. Blots were blocked in blotto (50 mm Tris, 100 mm NaCl, 0.1% Tween 20 and 5% dried milk) 1 h at RT. Blots were incubated with a 1:500 dilution of IIH6, 1:200 dilution of 8D5 (NovoCastra) and 1:2000 dilution of beta-actin (Sigma) in blotto overnight at 4 C. Blots were washed 2X5 minutes in blotto at RT. Blots were then incubated with a 1:5,000 dilution of goat anti-mouse IgM peroxidase for DG (Roche) or 1:20,000 donkey anti-mouse peroxidase for DG and actin (Jackson ImmunoResearch Laboratories) for 1 h at RT. Blots were washed in 50 mm Tris, 100 mm NaCl, and 0.1% Tween 20 3X5 minutes. Blots were developed using SuperSignal West Pico Chemiluminescent Substrate (Pierce). Results DG Expression in the Mouse Prostate DG expression has been examined in many tissues but has not been characterized in the mouse prostate. In order to determine the localization of DG in the mature mouse prostate we stained sections with antibodies to both and DG. DG was localized to the basolateral surfaces of epithelial cells in contact with the basement membrane (Figure 2.1A and D). Importantly, these cells reacted with the glycosylation-sensitive antibody IIH6 (Figure 2.1A) indicating that DG is post-translationally modified in prostate epithelial tissue as it is in others such as muscle and nerve. We observed similar DG staining in epithelial cells of all four lobes of the mouse prostate with the ventral lobes expressing the highest amounts of DG (Figure 2.2A to C). In order to explore the function of DG in the mouse prostate, we employed a conditional gene targeting strategy as we have previously used (Cohn, Henry et al. 2002; Moore, Saito et al. 2002). To generate a null DG mutant prostate epithelium, we utilized
43 31 a transgenic mouse line with prostate epithelial-specific expression of Cre recombinase (PB-Cre4) (Wu, Wu et al. 2001). Cre expression is controlled by the modified rat probasin promoter ARR2PB resulting in specific, but variable, expression in the luminal epithelial cells of all prostatic lobes. PB-Cre4 mice were bred to mice bearing the floxed DG allele to generate a prostate epithelium-specific DG knockout mouse. Loss of both and DG from the epithelium was detectable by 3 months of age (Figure 2.1B to E) in all lobes of the prostate (Figure 2.2). This experiment also established the specificity of the prostate epithelial staining with antibodies directed to both and DG. In addition, muscle surrounding the urethra expressed DG indicating prostate-specific loss (Fig. 2.1B to E, insets). Interestingly, the DG KO animals maintained small patches of DG-positive cells (arrow, Figure 2.1B). Incomplete Cre-mediated recombination in the PB-Cre4 strain has been described previously and may reflect variable transgene expression in different prostate lobes and/or the presence of basal cells in which the androgendependent ARR2PB promoter is not expected to be active (Wu, Wu et al. 2001). Normal Morphology, Ultrastructure, Cellular Composition and Function of DG Knockout Prostates In order to assess the consequences of a loss of DG expression in the mouse prostate, we evaluated the histology, ultrastructure and cellular composition of DG knockout prostates compared to littermate controls. Histologic assessment of all four lobes of the prostate in DG WT (n= 8) and KO (n= 8) mice at 12 months of age showed no abnormalities in gland morphology (Figure 2.3A to F) respectively. Additionally, DG knockout prostates were indistinguishable from control littermates at both 6 and 18 months (data not shown). Moreover, although the preceding mice were on a mixed B6.129 background, we have also evaluated prostate-specific knockouts in an extensively
44 32 backcrossed B6 background at 5 months of age (n=2) and also observed no histological differences (data not shown). We have previously shown that DG is involved in the establishment and/or maintenance of basement membranes in some tissues (Henry and Campbell 1998; Moore, Saito et al. 2002). We therefore examined the basement membranes of DG knockout prostates at the level of ultrastructure and immunohistochemistry. Transmission electron microscopy was used for ultrastructure analysis. The basement membrane was present on the basal side of the epithelium in both WT (n=2) and KO (n=2) animals. No apparent changes in continuity, morphology or distance from the epithelium were observed (Figure 2.3G and J). Moreover, on the apical surface of the cells microvilli and intact tight and adherens junctions were observed (Figure 2.3H and I, K and L). These data indicate that DG is not required for polarity of luminal epithelial cells. The basement membrane is composed primarily of laminins, collagen IV, perlecan and nidogen/entactin. DG binds to both laminins and perlecan through conserved G-domains in these molecules (Talts, Andac et al. 1999). Therefore, we assessed the expression and localization of perlecan and laminin by immunofluorescence. Perlecan and laminin were present in the basement membrane of the prostate glands and staining intensity and distribution was unaltered by the status of DG (Figure 2.4A and D, B and E) respectively. Although no changes were evident using a pan-laminin antibody, changes in the expression of particular laminin isoforms in the DG knockout prostates was possible. The expression of laminin isoforms has not yet been extensively evaluated in the mouse prostate basement membrane. We therefore assessed expression of laminin-511/521 by probing with antibodies specific for the 5 chain of laminin. Laminin-511/521 staining was similar between DG KO prostates and WT littermates (Figure 2.4C and F). We did not detect expression of the laminin 1 chain in adult mouse prostate epithelial basement membranes.
45 33 The prostate is composed of two primary cell populations; basal and luminal cells. Unlike human prostatic epithelium, mouse basal cells do not form a continuous layer juxtaposed to the basement membrane and are instead spread discontinuously throughout the epithelium except in the proximal ducts where basal cells are concentrated. Consequently, mouse luminal epithelial cells are in direct contact with the basement membrane. Utilizing basal cell specific marker K5, we assessed the role of DG in maintaining cellular composition of the prostate (Figure 2.4G and I). There was no significant difference in the percentage of K5-positive cells in the epithelial compartment between WT (3.7 ± 2.4) and KO (4.3 ± 2.3) mice at six months of age (p=0.603, t-test). Epithelial cell/extracellular matrix interactions are known to affect cell proliferation and apoptosis. We therefore examined proliferation by Ki-67 staining (Figure 2.4H and J). There was no significant difference in proliferation in the epithelium between WT (0.58 ±0.90) and KO (0.25 ± 0.49) mice at six months of age (p=0.408, t-test). One function of the prostate is to secrete and store prostatic fluid which is a component of semen. We therefore assessed function of the prostate by fertility. We assessed the number of pups per litter for 5 litters sired by each of 3 WT and KO males (15 litters each genotype). Loss of DG from the prostate does not affect fertility as there was no significant difference in number of pups per litter between WT (6.1 ± 1.6) and KO (6.5 ± 2.6) mice (p=0.677, t-test). Normal Regeneration of DG Knockout Prostates The prostate is able to undergo multiple cycles of involution following androgen ablation (castration) and regeneration with reintroduction of androgens. Epithelial cell/extracellular matrix interactions are essential for cell proliferation and differentiation necessary for regeneration. We therefore determined how loss of DG from the prostate epithelium would affect this process. Prostate-specific DG KO and WT littermate mice
46 34 were castrated between 6 and 9 months of age. Twenty one days after castration, prostates were regenerated by reintroduction of testosterone. Prostates were assessed after 21 days of testosterone treatment. As expected, castration resulted in a significant decrease in individual gland size and overall tissue size (Figure 2.5A and B). Following restoration of androgens, DG knockout prostates were able to regenerate comparably to WT littermate controls as assessed by histology (Figure 2.5C and D). Expression and localization of laminin and perlecan were unchanged compared to WT animals (Figure 2.6A and D). DG Expression in the Castrate-Resistant Basal Cell Population Involution of the prostate after androgen ablation results in apoptosis of luminal cells with survival of a basal cell population resistant to the effects of castration. Both and DG expression was significantly enhanced in WT mice after castration compared with intact controls (Figure 2.7 A and C and G). The castrated prostate takes on a pleated appearance due to the delayed degradation of the basement membrane as the glands undergo apoptosis (de Carvalho and Line 1996). Therefore increased DG expression may be associated with the excess basement membrane. Interestingly, DG expression in the KO mice was similar to WT mice indicating DG is expressed in the androgen-independent basal cell population and that this expression is maintained in PB4-Cre expressing mice (Figure 2.7A and D). DG expression was lost after regeneration of the prostate although some patches of cells with expression were evident, similar to intact control animals (Figure 2.7B and E, C and F). Due to the ability of the prostate gland to undergo repetitive cycles of involution and regeneration, an androgen-independent stem cell population must exist to regenerate the glands (Isaacs 1985). Furthermore, this population exists as a part of the larger
47 35 androgen-independent basal cell population and is thought to reside in a stem cell niche located in the proximal prostatic ducts; although recent evidence indicates the existence of a distinct luminal stem cell population (Tsujimura, Koikawa et al. 2002; Goto, Salm et al. 2006; Wang, Kruithof-de Julio et al. 2009). We therefore examined the expression of DG in the proximal ducts (Figure 2.8A and B). DG was expressed in the basal cell population of the proximal ducts, identified by anatomic location K5 and p63 staining, in both WT and KO mice (Figure 2.8C and F) suggesting DG may be expressed in this prostate stem cell population. To confirm the lack of expression of Cre from the ARR2PB promoter in this compartment, we crossed PB-Cre4 mice to the 129- Gt(ROSA)26Sor/J reporter strain (Soriano 1999). As expected, this showed - galactosidase staining in the distal prostate (Figure 2.8G) but not in the proximal ducts penetrating the periurethral muscle (Figure 2.8H) Discussion DG has been most extensively studied in skeletal muscle although it is expressed in many other tissues including epithelia. As mentioned above, accumulating data indicates that DG is involved in the development of epithelial tissues including formation of basement membranes and the establishment of cell polarity. On the other hand, disruption of DG in the mouse epiblast using Mox2-Cre (which creates an embryo constitutively lacking DG); resulted in mice that survived to birth, though they died shortly thereafter (Satz, Barresi et al. 2008). Although epithelial tissues were not extensively examined in these mice, this phenotype suggests that DG may not have a role in epithelial development or may have a role not yet assessed by initial characterization. We therefore sought to determine the role of DG in the prostatic epithelium using a prostate-specific knockout mouse. Utilizing the PB-Cre4 transgenic mouse we deleted DG in the prostate from mice carrying a floxed DG allele. Given the prior work in this
48 36 area, we were surprised to find that loss of DG expression did not lead to any observed phenotype. Specifically we evaluated histology, ultrastructure, expression of DG ligands, cellular composition, proliferative status, regenerative capacity, and fertility in mice in cohorts of mice up to 18 months of age. These data afford several conclusions regarding the role of DG in the prostatic epithelium. Earlier studies support the concept that DG is required for the assembly and/or maintenance of some basement membrane structures in various tissues (Williamson, Henry et al. 1997; Henry and Campbell 1998; Moore, Saito et al. 2002). Because the PB- Cre4 transgene is not extensively expressed during early urogenital development and early branching morphogenesis, we cannot assess DG function during this period when basement membranes are first assembled in the prostate (Wu, Wu et al. 2001). However, our data indicate that DG is not required for the subsequent maintenance of the basement membrane associated with luminal epithelial cells up to 18 months of age. We found no differences in the ultrastructure or molecular composition using pan-laminin, laminin 5 and perlecan antibodies, representing known DG ligands. Residual expression of DG is observed in what are most likely to be K5-positive basal cells, in which the ARR2PB promoter is not active. Because this population comprises <5% of the cells in the distal epithelium of the prostate it would seem unlikely to account for the maintenance of the entire basement membrane structure. However, basal cells extend processes over a large area which could play a role in maintenance of the basement membrane (Soeffing and Timms 1995; Hayward, Brody et al. 1996). Thus, our data indicate that DG in luminal epithelial cells is not required for the maintenance of the basement membrane in the prostate. Perhaps other ECM receptors are involved in this process, or, once assembled, the intrinsic self-assembly properties may act to maintain basement membrane structure (Yurchenco, Tsilibary et al. 1985). DG may still be required for the initial assembly or maintenance of basement membranes in some tissues; such as Reichert s membrane or the pial basement membrane (Williamson, Henry et al. 1997; Moore, Saito et al. 2002).
49 37 Prior studies showing that DG is required for laminin assembly in cultured mammary epithelial cells involves de novo assembly (Weir, Oppizzi et al. 2006). However, a requirement for DG in basement membrane assembly or maintenance is likely to be tissue-specific. The basement membrane between the visceral endoderm and epiblast and in analogous structures in embryoid body preparations that recapitulate the development of the epiblast, as well as the basement membrane in skeletal muscle, persists in the absence of DG function (Cohn, Henry et al. 2002; Li, Harrison et al. 2002). This may reflect a role for DG in the assembly or maintenance of certain types of basement membranes as both Reichert s membrane and the pial basement membrane are relatively thick and/or are not formed between continuous cell layers unlike that between the visceral endoderm and epiblast. Another unexpected finding is that DG is not required for the maintenance of epithelial polarity in mouse luminal prostatic epithelial cells. Luminal epithelial cells in the DG knockout prostate showed polarized features such as apical microvilli and tight junctions. Prior studies in mammary epithelial cells have indicated a role for DG in polarity (Weir, Oppizzi et al. 2006). In this case, the role for DG in polarity may be secondary to its role in laminin assembly as the latter may provide polarizing cues. In Drosophila melanogaster it has become evident that the role of DG in epithelial polarity is context-dependent as the role of DG in polarity is evident only under conditions of energetic stress (Deng, Schneider et al. 2003; Mirouse, Christoforou et al. 2009). It will be interesting to determine if a similar role for DG in maintaining polarity under energetic stress exists for mammalian epithelial cells. Here we also found that DG is expressed in the basal cell compartment of the prostate epithelium, including those in the proximal ducts which are thought to contain prostate stem cells (Tsujimura, Koikawa et al. 2002; Goto, Salm et al. 2006). Since the ARR2PB promoter is not active in these cells, DG is not disrupted in this compartment. This likely explains why prostate regeneration following castration is normal in mice
50 38 with prostate-specific DG knockout. Androgen ablation (castration) results in loss of androgen dependent luminal cells from the prostatic ducts by apoptosis while the androgen independent basal cells survive. Other approaches will be required to test the role of DG in this compartment. Earlier studies showing a role for DG in muscle regeneration suggest that DG may play a role in stem cell function in other tissues (Cohn, Henry et al. 2002). Following castration, prostatic ducts involute, resulting in a folded basement membrane surrounding the atrophied glands. Interestingly, we found that both and DG were elevated in the atrophied glands in WT animals. This suggests that membrane fragments may be associated with the residual basement membrane. This residual basement membrane may be involved in the regeneration of the prostate and suggests that de novo basement membrane assembly may not be required for prostate regeneration. It will be interesting to determine whether DG associated with this residual basement membrane plays a role in maintaining this structure or in subsequent regeneration. Disruption of epithelial cell/extracellular matrix interactions is a hallmark of cancer due to the disruption in regulation of cell proliferation, apoptosis, survival and differentiation. Loss of DG function may be associated with the progression of epithelial cancers including prostate cancer (Henry, Cohen et al. 2001; Sgambato, De Paola et al. 2007). Overexpression of DG in breast cancer cells inhibits cell proliferation and decreases tumorigenesis (Muschler, Levy et al. 2002). These data suggest loss of DG from the epithelium may result in disruption of tissue homeostasis. However, in the normal prostate, tissue architecture remained unchanged by histological assessment of DG KO mice from 3-18 months of age compared with WT animals. We also showed that number and localization of K5-positive basal cells as well as proliferation as assessed by Ki-67 staining remained the same. These data suggest DG does not have a direct role in regulation of proliferation, survival and differentiation of adult mouse prostate luminal epithelial cells. Thus, loss of DG function in luminal cells is insufficient to initiate
51 39 neoplastic progression in the mouse prostate. However, it remains possible that loss of DG function contributes to tumor progression when combined with other molecular or cellular alterations.





")

-specific")

 compared")








52 40 Figure 2.1. Dystroglycan localization and expression in the ventral lobe of a prostate sp pecific knockout mouse. DG is expressed on the basal surface of the epithelium (yellow, arrows) in the prostate gland as shown by -DG (IIH6) and -DG (8D5)-specific antibodies (A,D). DG expression is lost in DG KO mice (B,E) compared with WT (A,D) although patchy areas of expression remain (arrowheads) (B,E). Tissues stained with secondary antibodies only were used as a negative control (C,F).. Muscle surrounding the bladder neck was used as a positive control (inset boxes). Nuclei were stained with DAPI (blue). Scale bar 50 m. Autofluoresc cence of luminal secretions (*).

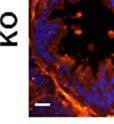




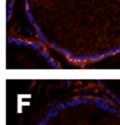
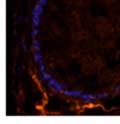


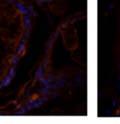
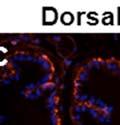

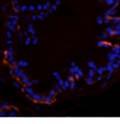
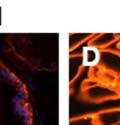




53 41 Figure 2.2. Dystroglycan localization and expression in all lobes of the prostate. Dy ystroglycan (IIH6) is expressed in all lobes of the mouse prostate (A-C). DG expression is lost in the DG KO (E-G)) mice compared with WT (A-C) although patchy areas of expression remain. Muscle surrounding the bladder neck was used as a positive control (D, H) Nuclei were stained with DAPI (blue). Scale bar 50 m.










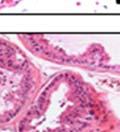
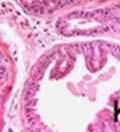
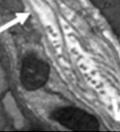

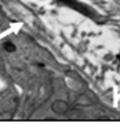








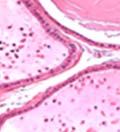





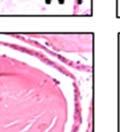
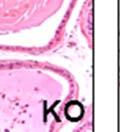

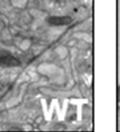

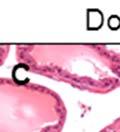
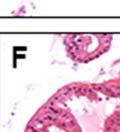



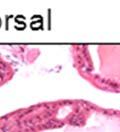

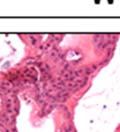

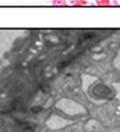
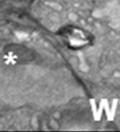


54 42 Figure 2.3. Histology and ultrastructure of thee prostate. Hematoxylin and eosin stain of the ventral (A,D); lateral (B,E) and d dorsal (C,F) prostatic lobes in DG WT (n=8) and KO (n=8) mice were similar at six months of age. Scale bar (A-F); 50 m. Transmission electron microscopy of the basement membrane (arrows) of the lateral lobe of DG WT (G) and KO (J) mice. Epithelial microvilli and tight junctions (arrows)) of the lateral lobe of DG WT (H,I) and KO (K,L) mice, respectively. Two WT and two KO mice were analyzed. Epithelial cell (*). Scale bars (H,I,K,L) 1.0 m (G,J) 0.4 m.





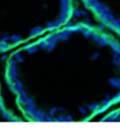

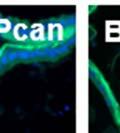

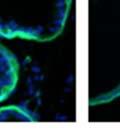


















55 43 Figure 2.4. Basement membrane and cellular composition of the ventral lobe of the pr rostate. Extracellular matrix proteins and DG binding partners perlecan and laminin (green) showed a similar localization and staining intensity in DG WT (A,B) and KO mice (D,E) respectively. Likewise,, laminins 511/521 (red) were also unchanged (C,F). Basal cell number (G,I) as assessed by anti-k5 staining (red) and proliferation rate (H,J) as assessed by Ki-67 staining (green) were similar. Nuclei were stained with DAPI (blue). 3 WT and 3 KO animals were assessed for each protein. Scale bar 500 m.



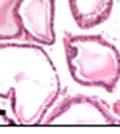
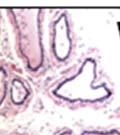
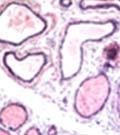
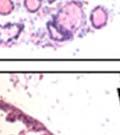
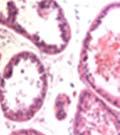
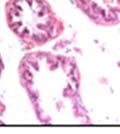

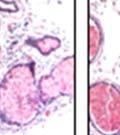


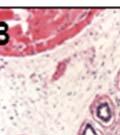
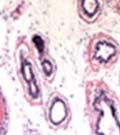











56 44 Figure 2.5. Involution and regeneration of the lateral and ventral lobes of the mouse prostate. DG WT (A) and KO (B) mice were castrated at 6 to 9 months of age. Testosterone was introduced 21 days post castration and regenerated prostates weree analyzed 21 days after reintroduction of testosterone. WT (C) and KO (D) prostates regenerated normally. Scale bar is 150 m.

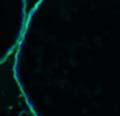







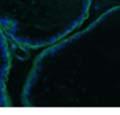







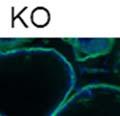



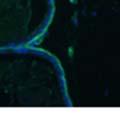
57 45 Figure 2.6. Expression of DG ligands followingg involutionn and regeneration of the mouse mlo prostate. DG binding partner laminin is expressed with normal calization in the regenerated prostatee (A,B). Perlecan is expressed and localized to the basement membrane in both WT (C) and KO (D) regenerated prostates. Nuclei were stained with DAPI (blue). Scale bar is 50 m.








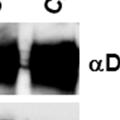
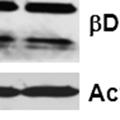






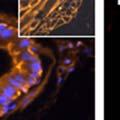
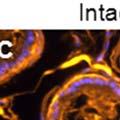
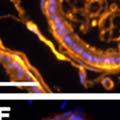

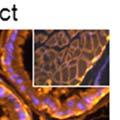
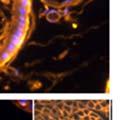
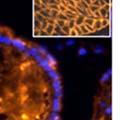
58 46 Figure 2.7. DG expression following involutionn and regeneration of the mouse pr rostate. DG expression is enriched after involution in both WT and KO mice (A,D) and is reduced after restoration of androgens allowed for regeneration in KO mice. Immunofluorescence staining for DG (IIH6) was performed on the ventral lobe of castrated WT (A) and KO (D) mice, regenerated ventral lobe (B,E) and normal ventral prostatee (C,F) respectively. Nuclei were stained with DAPI (blue). Muscle surroundingg the bladder neck was used as a positive control (inset boxes) 6 mice of each genotype were assessed. Scale bar 50 m. (G) DG (IIH6) and DGG (8D5) protein expression was enriched in protein lysates of castrated (C1, C2) prostatess compared with intact (I) prostates as shown by western blot.

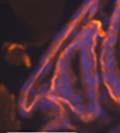

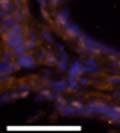



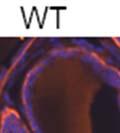




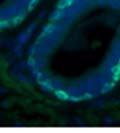


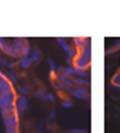


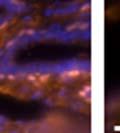


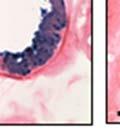








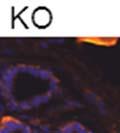






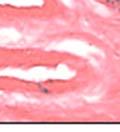
59 47 Figure 2.8 Expression of DG in the basal cell population of the prostate proximal ducts. Immunofluorescence staining for DG (red)) expression in the proximal ducts of WT and KO mice (A,B). Basal cells are indicated by P63, red (C,D,) and Keratin 5, green (E,F) expression.. Nuclei are stained with DAPI (blue). 3 mice of each genotype were assessed. Scale bar 50 m. X-gal staining (blue) detects Cre recombinase activity in the distal ducts (G) but not the proximal ducts (H) of the mouse prostate ventral lobe. Scale bar 50 m.
60 48 CHAPTER III DYSTROGLYCAN EXPRESSION IN THE PROSTATE BASAL CELL IS NOT REQUIRED FOR MAINTENANCE OF TISUE HOMEOSTASIS OR THE EPITHELIAL BASEMENT MEMBRANE Introduction Epithelial cell/extracellular matrix interactions are essential to tissue morphogenesis and homeostasis. Maintenance of cellular homeostasis requires a balance between cell differentiation, proliferation and apoptosis. The prostate epithelium is composed of three cell types; luminal, basal and rare neuroendocrine cells. Although the basal cell population comprises less than 5% of the epithelium, they have long extensions that wrap around the ducts and penetrate between the luminal cells accounting for up to 19% of the ductal surface area (Soeffing and Timms 1995; Hayward, Brody et al. 1996). Basal cells have an essential role in maintaining ductal integrity and survival of luminal cells (Isaacs and Coffey 1981; Kurita, Medina et al. 2004). In addition, the basal population is thought to include the prostate stem cell population, primarily due to their ability to regenerate the prostate following androgen depletion and reintroduction of testosterone. Currently two models have been proposed to describe stem cell fate in the prostate (Figure 3.1). The first states that stem cells become intermediate basal cells which terminally differentiate into luminal cells. The second suggests that two separate stem cells populations exist, a basal stem cell and a luminal stem cell. Recently an androgen independent luminal cell population has been identified supporting the existence of two distinct prostate stem cells (Wang, Kruithof-de Julio et al. 2009; Liu, Pascal et al. 2011). Thus, stem cell lineage in the prostate remains unclear and widely debated.
61 49 We have previously shown DG expression in basal cells of the prostate epithelium (Esser, Cohen et al. 2010). It is also expressed in some stem cell populations including satellite cells of skeletal muscle where deletion of DG leads to exhaustion of the stem cell pool (Cohn, Henry et al. 2002). In embryonic stem cells, DG is required for laminin binding (Henry and Campbell 1998). Thus, DG may have a function in prostate stem cells. DG has roles in tissue morphogenesis, cell growth and survival suggesting that defects in DG may disrupt development of the prostate or maintenance of cellular homeostasis (Durbeej, Larsson et al. 1995; Durbeej, Talts et al. 2001; Weir, Oppizzi et al. 2006). We have previously shown expression in luminal cells is not required for maintenance of the basement membrane composition or ultrastructure, cellular architecture of the glands or regeneration of the prostate (Esser, Cohen et al. 2010). However, we have not yet assessed its function in the prostate basal/stem cell population. Here we assess the expression and function of DG in the basal/stem cell population of the mouse prostate. Utilizing Cre-lox technology we have generated a Keratin 5 specific knockout mouse. Keratin 5 driven expression of cre recombinase deletes DG from the basal cells of the prostate as well as other stratified epithelia. We show that DG is deleted from the majority of the epithelium including some luminal cells. We further show that DG is not required for, maintenance of the basement membrane or cellular architecture. Following involution and regeneration, DG null prostates have no morphological abnormalities or disruption of basement membrane composition or cellular architecture. Interestingly, deletion of DG from the epithelia of the ear, hind-stomach, trachea and tongue does not disrupt normal tissue morphology. However, in the skin, DG may have a role in wound healing. Furthermore, DG may have a role in the modulation of tumor stroma during cancer progression suggesting DG may function in the K5 Cre prostates in ways we have not yet assessed. These data suggest DG may have a more subtle role within these tissues.
62 50 Materials and Methods Animals All procedures involving animals were performed according to The University of Iowa Animal Care and Use Committee policies. C57BL/6 male DG fl/fl mice (Cohn, Henry et al. 2002); (kind gift from Dr. Kevin Campbell) were crossed with female C57BL/6 Keratin5-Cre mice (Ramirez, Page et al. 2004). F 1 DG fl/wt ; Cre + males were bred to DG fl/wt ; Cre - females. The F 2 DG fl/fl ; Cre + males were bred to DG fl/fl ; Cre - females. F 3 males were used for analysis. Herein WT refers to animals with a DG fl/wt Cre + or DG fl/fl ; Cre- genotype and KO refers to animals with DG fl/fl ; Cre + genotype. Female 129- Gt(ROSA)26 Sor/J mice (Soriano 1999), The Jackson Laboratories) were also bred to male C57BL\6 K5-Cre mice. Beta-galactosidase activity was assayed in the F1 generation. For genotyping, tail DNA was extracted (REDExtract-N-Amp Tissue PCR Kit, Sigma) and PCR was performed. All animals were assessed at 6 months of age unless otherwise noted. For castration/regeneration studies, mice at 3 to 4 months of age were anesthetized (100 mg/kg ketamine, 10 mg/kg xylazine) and castrated. Castrated mice were euthanized by CO 2 asphyxiation after 21 days. Remaining castrated mice were implanted with testosterone pellets (Innovative Research of America) at 15 mg/kg for 21 days. Testosterone-treated mice were euthanized after 21 days. 129.B6; D2.B6 DG fl/fl ; PbCre+ male mice (generated as described in chapter 2) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) (Cohn, Henry et al. 2002) were crossed with PTEN fl/fl female mice (Jackson laboratories). F 1 DG fl/wt ; PTEN fl/wt ; Cre + males were bred to DG fl/wt ; PTEN fl/wt ; Cre - females. The F 2
63 51 DG fl/wt ; PTEN fl/fl ; Cre + males were bred to DG fl/fl ; PTEN fl/fl ; Cre - females. F 3 males were used for analysis. All animals were assessed at 4 months of age. Antibodies Anti- DG (IIH6, gift from Dr. Kevin Campbell (Ervasti and Campbell 1991), anti- DG (8D5, Novocastra), CK5 (AF138, Covance), Ki-67 (NCL-Ki67p, Novocastra), perlecan (MAB1948, Chemicon), pan-laminin (Durham and Snyder 1995); (gift from Dr. Jeanne Snyder), PTEN (Zymed Labs), CD31 (BD Pharmigen). Histology and Immunofluorescence Analysis Mouse urogenital organs were removed as a unit and fixed in 4% paraformaldehyde for 4 hours at 4 C and then transferred to 30% ethanol overnight. Prostates were dissected from the seminal vesicles and bladder. Mouse skin was removed from the back and fixed in 4% paraformaldehyde for 4 hours at 4 C and then transferred to 70% ethanol overnight. Tissues were processed through an ethanol to xylene gradient for paraffin embedding using a Ventana Automated Tissue Processor. 5 m sections were cut using a Leica RM2125 rotary microtome. Tissues were deparaffinized and hydrated through a xylene to ethanol gradient followed by citrate buffer or proteinase K antigen retrieval. After 1X phosphate buffered saline (PBS) wash, slides were blocked for 1 hour at room temperature (RT) in blocking solution (2% donkey serum, 1% BSA, 0.05% Tween-20, 0.1% Triton X-100 in 0.01M PBS). Slides were incubated with primary antibody in blocking solution overnight at RT. After two 1X PBS washes, slides were incubated with secondary antibody and nuclear stain 4',6-diamidino-2-phenylindole (DAPI) in blocking solution for 1 hour at RT. Slides were washed twice and coverslipped. Number of keratin 5 (K5)-positive cells per 100 DAPI-positive epithelial
64 52 nuclei were counted for 3 separate areas of the ventral prostate per mouse. WT (n=3) and KO (n=2) mice were assessed at 6 months of age. Resulting data were averaged and analyzed by t-test. Sections were analyzed independently by two individuals, and similar results were obtained. For hematoxylin and eosin staining, prostate sections were processed under standard conditions. X-gal Histochemistry Mouse urogenital organs were removed and prostates were dissected from the seminal vesicles and bladder. Prostates were fixed for 1 hour at 4 C in 4% paraformaldehyde followed by incubation in 30% sucrose overnight at 4 C. The tissue was then embedded in Optimal Cutting Temperature embedding medium, frozen briefly in liquid nitrogen and stored at -20 C overnight. Tissues were cut using the Zeiss Micron Model HM505E at 10 m. Slides were stored at -80 C. For staining, slides were washed one time in 1X PBS and incubated for 30 minutes at RT in rinse buffer (100 mm sodium phosphate ph 7.3, 2 mm MgCl 2, 0.01% sodium deoxycholate, 0.02% NP-40). Following aspiration of the rinse buffer, slides were incubated in staining solution (rinse buffer plus 5 mm potassium ferricyanide, 5 mm potassium ferrocyanide and 1 mg/ml X-gal) for 4 hours at 37 C. Slides were rinsed in PBS, counterstained with eosin, and coverslipped. Wound Healing For wound healing studies, female mice at 6-7 weeks of age were anesthetized (100 mg/kg ketamine, 10 mg/kg xylazine). 4mm biopsy punches were used to create 2 wounds in the epithelium on the dorsal side of the mouse (WT n=22, KO n=22). Photographs were taken at the time of wounding and every other day after for ten days
65 53 following anesthetization with isofluorane. Wound size was quantified using Image J software. Results DG Expression in a Basal Cell Specific Knockout Mouse We have previously shown DG expression in the epithelium of the mouse prostate. In order to understand its role in basal cells, we employed a gene targeting strategy we have previously used. For deletion of DG in the basal cells of the prostate, we utilized a transgenic mouse expressing Cre recombinase driven by the bovine keratin 5 promoter (K5-Cre). Keratin 5 is expressed in many stratified and pseudo-stratified epithelia including the prostate. However, expression of Cre recombinase had not yet been demonstrated in the prostate of this K5-Cre mouse strain (Ramirez, Page et al. 2004). We therefore crossed the K5-Cre strain to the 129-Gt(ROSA)26 Sor/J reporter strain. -galactosidase expression confirmed K5 driven Cre expression in all lobes of the prostate except the anterior (Figure 3.2). Low levels of Cre expression in the anterior prostate may require prolonged staining with X-gal for detection. K5-Cre mice were then bred to mice harboring the floxed Dag1 allele to generate an epithelial basal cell specific knockout mouse. DG expression was lost by three months of age in all lobes of the prostate as indicated by antibodies for glycosylated DG (IIH6) and DG (8D5)(Figure 3.3, Figure 3.4). Deletion of Dag1 was specific to the prostate as expression was maintained in the muscle surrounding the urethra (insets). Interestingly, some glands had complete elimination of DG, indicating loss in both basal and luminal cells, while others maintained small areas of expression. This staining pattern correlated with transgene expression as indicated by -galactosidase activity. To confirm loss of DG from the
66 54 basal cell population and maintained expression in the luminal cell population, we dual labeled DG null prostates with antibodies for keratin 5 and DG (Figure 3.5). K5 positive basal cells were IIH6 negative as expected. Cells that maintained DG expression were K5 negative and are presumably luminal cells. In addition many K5 negative cells were also IIH6 negative, suggesting a significant part of the luminal cell population arises from K5 positive cells. Deletion of DG in the prostate basal cell was further confirmed by immunofluorescence staining in the proximal region of the prostate gland. These regions are enriched in K5+ basal cells and are thought to act as the prostate stem cell niche. Staining with an antibody for DG (IIH6) showed loss of DG expression in the proximal ducts (Figure 3.6). This correlated with expression of Cre as identified by b- galactosidase activity. Staining was specific as WT proximal ducts showed no staining. These data indicate DG is lost in the basal cell population as well as some K5 negative cells. DG is Not Required for Prostate Development Including the Formation and Maintenance of the Basement Membrane The prostate functions to secrete and store fluid that later becomes part of the seminal fluid. To determine whether loss of DG affects fertility, we quantified the number of pups per litter for three and four litters bred with two KO and two DG heterozygous males (seven litters per genotype). Loss of DG from the prostate does not affect fertility as there was no significant difference in number of pups per litter between WT (7.0 ± 1.27) and KO (6.4 ± 0.611) mice (p=0.6927, t-test). The prostate forms from the urogenital sinus through the events of bud formation, branching morphogenesis and canalization of the glands. Keratin 5 has been shown to be
67 55 expressed in the urogenital epithelium during development (Soeffing and Timms 1995). Deletion of DG should therefore occur at the earliest time points of gland formation providing the ability to assess DG function during development. Prostate basal cells maintain ductal integrity and function in the survival of luminal cells. DG has been shown to be required for branching morphogenesis and basement formation in some tissues indicating loss of DG expression may affect gland morphology or formation of the basement membrane. However, no abnormalities were present at 3 and 6 months of age in DG null prostates by histological assessment (Figure 3.7). Immunofluorescence staining of DG binding partner s laminin and perlecan showed they were expressed and localized to the basement membrane as expected (Figure 3.8). Laminin was detected with a pan-laminin antibody and therefore alterations in the expression of specific laminin isoforms were not assessed. DG has also been shown to regulate cell growth and survival. Utilizing a keratin 5 specific antibody, we assessed the localization and number of basal cells. K5+ basal cells were present and localized normally throughout the gland. There was no significant difference in the number of basal cells in WT (0.17±0.07) and KO (0.26±0.10) dorsal lobes (p-value= 0.52, n=2). DG Null Prostates are Able to Regenerate Following Ablation and Reintroduction of Androgens The prostate is able to undergo repeated cycles of involution and regeneration following androgen deprivation and reintroduction. Involution occurs due to apoptosis of androgen dependent luminal cells and subsequent degradation of the basement membrane. Cell/extracellular matrix (ECM) interactions are necessary for cell proliferation, differentiation and survival during regeneration of the glands. Glands are thought to regenerate by activation of stem cells in the androgen independent basal cell population. DG has been shown to be required for stem cell function. We therefore asked
68 56 if DG was required for basal cell regeneration of the prostate epithelium. Twenty one days after surgical castration, involuted prostates did not express DG as expected (Figure 3.9). However, glands with a small number of cells retaining DG expression were present, similar to intact animals (Figure 3.10). Recent work has indicated the existence of an androgen independent luminal cell. We show that the cells retaining DG expression following androgen deprivation are K5 negative suggesting they are androgen independent luminal cells (Figure 3.10). Following involution, prostate were regenerated by reintroduction of androgens for twenty one days. An additional group of mice then underwent a second round of involution and regeneration. Prostates from regenerated and twice cycled DG null mice did not express DG as expected (Figure 3.9) although small areas retained DG expression as seen in the intact prostates (Figure 3.10). Regenerated and cycled prostates had no morphological abnormalities by histological assessment (Figure 3.11). During involution and regeneration of the prostate, the basement membrane is degraded and restored. We therefore evaluated the expression and localization of DG ligands laminin and perlecan in regenerated prostates which had no differences in expression or localization to the basement membrane (Figure 3.12). Again, expression of specific laminin isoforms was not determined. K5+ basal cells were localized as expected and appeared in similar numbers in WT and KO animals. These data indicate DG expression in basal cells in not required for regeneration of the prostate epithelium. DG is Not Required for Morphogenesis of the Ear, Tongue, Stomach or Trachea DG is required for morphogenesis in several tissues in vitro but was not required for prostate morphogenesis in vivo (Durbeej, Larsson et al. 1995; Durbeej, Talts et al. 2001). Keratin 5 is expressed in all stratified epithelia and Cre expression has been
69 57 confirmed in the tongue, trachea, stomach and ear of the K5 Cre mice (Ramirez, Page et al. 2004). DG is also expressed in many epithelial tissues allowing assessment of DG function in many tissues of the K5 Cre mice. We therefore next determined if DG was required for morphogenesis in other tissues. DG localized to the basal surface of the epithelium adjacent to the basement membrane in all tissues assessed from three month old wild type mice (Figure 3.13). Immunofluorescence staining with IIH6 showed complete loss of glycosylated DG from the epithelium of the ear, tongue, hind-stomach and trachea of DG null mice. Histological assessment of ear skin, tongue, hind-stomach and trachea of DG null mice showed no abnormalities in the morphology of the tissues (Figure 3.14). However, a phenotype may be discerned with additional analyses of these tissues. Wound Closure Rate is Reduced in DG Null Mice To further assess the role of DG in the skin, we evaluated DG expression and tissue morphology. DG is expressed at the basal side of the epithelium and immunofluorescence analysis with the DG glycan specific antibody IIH6 confirmed loss of DG in the KO animals (Figure 3.15). Muscle sections and secondary antibody only staining were shown as controls. Laminin and perlecan were expressed and localized to the basement membrane as expected by immunofluorescence staining. To determine if there were disruptions in morphology during the various stages of the hair cycle, skin was harvested during the anagen (growth) and telogen (resting) phases. No alterations in morphology were present by H&E stain (Figure 3.16). We next assessed if loss of DG affected progression of the hair cycle. WT (n=1) and KO (n=1) mice were shaved and remaining hair was removed with a depilatory cream during the telogen phase of the hair cycle (Figure 3.17). Mice were anesthetized with isofluorane and imaged every other day throughout the anagen phase. No differences in hair cycle were observed however,
70 58 further experiments should analyze hair cycle histologically. We next assessed the role of DG in wound healing following removal of the skin with a biopsy punch. Wound areas were quantified using Image J software and displayed as percent wound closure. DG reduced the wound closure rate at days 8 and 10 (Figure 3.18). To elucidate the mechanism behind the reduction in wound closure, a few wounds were removed with a 6mm biopsy punch at days 6 and 10 (Figure 3.19). Due to the amount of variability between wounds, additional samples are needed for a complete histological assessment. Modulation of the Stroma with Loss of DG in a PTEN Deficient Model of Prostate Cancer Cohorts were assessed by histology at 4 months of age (Figure 3.20A to C) Mice that were WT for DG and PTEN (DG WT; PTEN WT) mice had normal prostate morphology. Mice that were heterozygous for DG and homozygous for PTEN deletion (DG Het; PTEN KO) had developed prostatic intraepithelial neoplasia (PIN) and were used as controls. Mice with homozygous loss of DG and PTEN (DG KO; PTEN KO) also had developed PIN and appeared identical to the DG Het controls. Immunofluorescence staining with the DG specific antibody 8D5 (Figure 3.20D and E), showed reduced expression in the DG heterozygous mice and loss of DG staining in the DG KO mice. This was further confirmed with the DG glycosylation specific antibody IIH6. The epithelial cells in PTEN KO mice were no longer in contact with the basement membrane and had lost polarity resulting in expression of DG throughout the plasma membrane. Staining for PTEN also confirmed loss of PTEN in the PTEN KO mice (Figure 3.20J to L). We next assessed expression and localization of DG ligand laminin. Interestingly, laminin was expressed in large quantities and localized throughout the reactive stroma in DG HET; PTEN KO mice (Figure 3.21A to C). However, in DG PTEN KO mice, laminin staining remained thick around the glands but was lost throughout the stroma.
71 59 We also assessed the number of blood vessels in these mice by CD31 immunohistochemistry (Figure 3.21D to E). Overlay images were used to better display alterations in the vasculature between genotypes (Figure 3.21F to H). Surprisingly, loss of PTEN resulted in an increase in the number of blood vessels that was further enhanced with loss of DG. These data indicated loss of DG expression modulates the composition of the tumor stroma. This data further supports a role for DG in the prostate epithelium. Discussion Unlike the human prostate, basal cells of the mouse prostate do not form a continuous layer along the basement membrane. They are instead interspersed throughout the gland allowing both basal and luminal cells to interact with the basement membrane. DG is expressed in both basal and luminal cells of the mouse prostate. Studies have indicated roles for DG ranging from tissue morphogenesis and basement membrane formation to regulation of cell polarity, proliferation and survival. We have previously shown that DG expression in luminal cells is not required for maintenance of the epithelial basement membrane or cell polarity in the mouse prostate (Esser, Cohen et al. 2010). However, its function in basal cells of the prostate is unknown. Prostate basal cells are required for gland architecture and luminal cell survival. Additionally, the K5+ population is thought to include the prostate stem cell population. As addressed previously, DG has roles in embryonic and muscle stem cells. We therefore generated a basal cell specific DG knockout mouse utilizing the keratin 5 promoter to drive Cre expression. We found that DG is not required for maintenance of the epithelial basement membrane or cytoarchitecture. Therefore DG may have a role in the prostate epithelium that we have not yet been able to assess. Keratin-5 is expressed in all stratified epitheliums. Several strains of K5-Cre mice have been generated and primarily utilized for epidermal research. While K5 driven Cre
72 60 expression had been shown in the epidermis as well as several additional tissues, none including the strain we have utilized, has established expression in the prostate. Through generation of a K5-Cre; Rosa Gt(ROSA)26 Sor/J reporter strain, we have shown Cre expression in three month old prostates. Cre was expressed in the ventral, lateral and dorsal lobes of the prostate. Staining was not detected in the anterior lobe probably due to low expression levels as DG staining was lost in the anterior lobes of DG null mice. Cre expression was not limited to the basal cell population but was also expressed in luminal cells as indicated by Cre expression in the majority of cells in some glands. Interestingly, this work has provided an insight into the prostate stem cell(s). Two models have been proposed to describe stem cell differentiation in the prostate (Figure 3.1). The first describes basal cells as an intermediate cell that gives rise to luminal cells as is seen in other stratified epitheliums such as the epidermis. The second proposes two independent stem cell populations. If one prostate K5+ stem cell population exists, DG deletion in the basal cells would result in compete loss of DG staining. Keratin 5 cells are interspersed around the prostate gland in a discontinuous layer. If two independent stem cell populations exist in the prostate, loss of DG expression would be expected to replicate this pattern. Here we have shown that DG is lost throughout the majority of the prostate epithelium. K5+ basal cells as well as numerous K5- (luminal) cells do not express DG. This suggests K5+ stem cells can give rise to both basal and luminal cells. However, DG expression is retained in a small number of K5- cells suggesting the existence of a distinct luminal stem cell population. Furthermore, involuted prostates also retain small areas of DG expression supporting the existence of an androgen independent luminal stem cell. We also show that regeneration of the prostate epithelium does not result in complete restoration of DG expression in all luminal cells. Therefore, these data support a model in which the majority of luminal cells are generated through differentiation of the basal cells while others may arise from a separate luminal stem cell. This androgen
73 61 independent luminal stem cell has been postulated to act under specific conditions such as androgen ablation (Wang, Kruithof-de Julio et al. 2009; Liu, Pascal et al. 2011). We also acknowledge we have not ruled out aberrant transgene expression as a cause of retained DG expression. However, previous studies in the epidermis utilizing this K5-Cre mouse strain have not reported abnormal transgene expression (Ramirez, Page et al. 2004). Therefore, further lineage tracing studies are required to definitively establish stem cell fate. Although we have established Cre expression at 3 months of age in the prostate, the time point at which it is first expressed in the developing tissue of these animals is unknown. However, K5 is expressed in the urogenital sinus epithelium at the earliest points of epithelial budding during prostate development. Therefore, loss of DG expression should occur early in development. We have found that DG null prostates developed without morphological abnormalities. Basement membranes were present and cytoarchitecture of the glands were intact. Regenerated prostates also had normal morphology suggesting DG may not have a role in the development or regeneration of the prostate gland. Assessment of gland structure at 6 months of age also indicates DG is not required for maintenance of the basement membrane or gland architecture. We additionally assessed tissue morphology in other tissues with stratified epithelia. Tissue morphology was unchanged with loss of DG in the epithelium of skin, tongue, stomach and trachea further indicating DG is not required for maintenance of epithelial morphology is some tissues. However, DG may have roles in the epithelium we have not yet assessed. This is supported by reduced wound healing in the DG null mice. Loss of DG expression in the epidermis reduced wound healing at days 6 and 10. Wound healing occurs in stages with the initial formation of a fibrin clot and inflammatory response followed by re-epithelialization and eventually remodeling of the wound. DG null wounds were significantly larger at the end of the re-epithelialization phase suggesting this phase may occur more slowly with loss of DG. Further characterization of
74 62 the wounds will be required to elucidate the mechanism behind a reduction in wound healing. This may include reduced motility of the keratinocytes or disruption of basement membrane formation. To further investigate the role of DG in the epithelium, we generated a Dag1 Pten prostate specific double knockout mouse. Loss of PTEN resulted in the formation of PIN within the prostate glands. However the additional loss of DG did not enhanced tumor progression or result in abnormalities in gland morphology. Compared with the original characterization of the mouse model by Wu et al., the progression of cancer is reduced in our mice as prostates should have areas of invasion and metastases by four months of age (Wang, Gao et al. 2003). The effects of PTEN loss vary greatly with genetic background. Thus the reduction in cancer progression is probably an effect of our mixed genetic background as compared to the C57BL/6xDBA2 background of the original PTEN mice. Loss of PTEN expression results in an inflammatory stromal response as shown by H&E. This reactive stroma has enhanced laminin expression and abnormal localization throughout the stroma. Loss of DG in the PTEN KO mice results in loss of laminin from the stroma, however expression is retained at the basement membrane suggesting DG modulates laminin expression and localization in the stroma but not at the basement membrane. Furthermore, we show that the number of blood vessels is enhanced with loss of PTEN and the formation of the inflammatory stroma. Blood vessel formation is further enhanced with loss of DG, however, these results have not been quantified. Therefore initial results suggest DG has a role in modulation of the stroma as well as in blood vessel formation in the context of an inflammatory tumor stroma. Due to the slow growth of prostate cancer in these mice, no further characterization was completed and they were bred to a C57Bl/6 background for further analysis. We have therefore demonstrated that loss of DG did not alter maintenance of the epithelial basement membrane or tissue homeostasis and cytoarchitecture in the prostate. However DG has a role in the epithelium under specific contexts. This includes a role in
75 63 epidermal wound healing as well as modulation of tumor stroma. Previous work also supports a role for DG in the epithelium under certain conditions. Several studies in Drosophila and in the mouse lung that have shown a role for DG in the AMPK pathway as an energy sensor required for maintenance of epithelial polarity and as a mechanoreceptor (Budinger, Urich et al. 2008; Mirouse, Christoforou et al. 2009; Takawira, Budinger et al. 2011). Therefore, additional studies are needed to determine DG function in epithelial tissues.











76 64 Figure 3.1. Models of prostate stem cell lineage. Two models for stem cell lineage have be een proposed in the prostate and are displayed ass a schematic. Nomenclature is as follows keratin 5 (K5) and keratin 8 (K8).


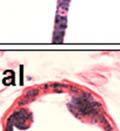



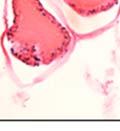
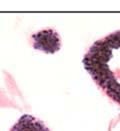



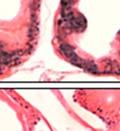



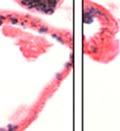


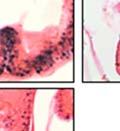
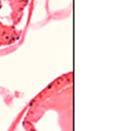
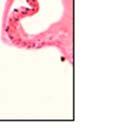
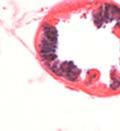








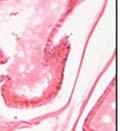
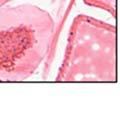
77 65 Figure 3.2. Cre recombinase expression in the mouse prostate. K5-Cre mice weree br red to a129-gt(rosa)26sor/j reporter strain. X-gal stain (blue) of individual prostate lobes shows Cre expression in all lobes of the prostate except the anterior. WT mice were shown as a negative control. Scale bar is 50 m.




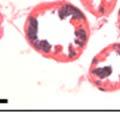
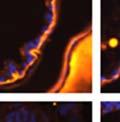


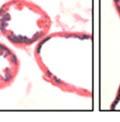

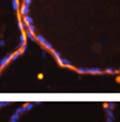






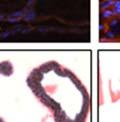






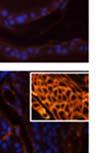


78 66 Figure 3.3. Dystroglycan expression is lost in DAG1 knockout mice. DG (IIH6 an ntibody) is lost with deletion of DAG1 (n=6) compared to WT mice (n=6) by immunofluorescence staining. Small areas of DG expression were present in DG knockout animals (arrows). Tissues stained with secondary antibodies only were used as a negative control. Muscle surrounding the bladder neck was used as a positive control (inset boxes). Nuclei were stained with DAPI (blue). X-gal stain of keratin-5 Cre129-Gt(ROSA) )26Sor/J reporter mice indicates Cre expression in the ventrall and lateral prostate. WT mice weree stained as a control. Scale bars indicate 50 m.

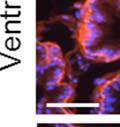


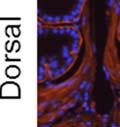

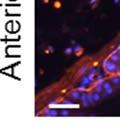
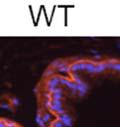

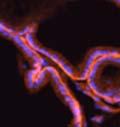









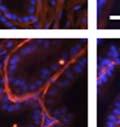
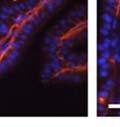




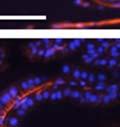
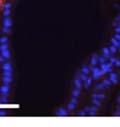
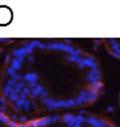



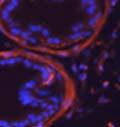
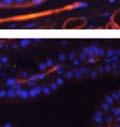

79 67 Figure Dystroglycan expression is lost inn all lobes of the mouse prostate. Im mmunofluorescence staining for DGG (8D5) is lost with deletion of DAG1 in alll lobes of the prostate (n=6) compared to WT mice (n=6). Scale bar indicates 50 m.
 and Keratin 5 (K5) in the dorsal prostate of a DG KO mouse.")
. B) Some K5 negative luminal cells are also DG negative (arrowheads).")
80 68 Figure 3.5. DG expression is lost in basal cells and some luminal cells. Im mmunofluorescence staining for DGG (8D5) and Keratin 5 (K5) in the dorsal prostate of a DG KO mouse. A) K5 negative/ DG negative basal cells are present(arrows) as well as K5 negative/ DG positive luminal cells (arrowheads). B) Some K5 negative luminal cells are also DG negative (arrowheads). Nuclei weree stained with DAPI (blue). Scale bars are 50 m.














81 69 Figure 3.6. DG is lost in the basal cells of the proximal ducts. Immmunofluorescence sta aining of K5 (green) positive basal cells enriched in the proximal regions of the prostate (arrowheads). X-gal staining (black arrows) of prostates from K5 Cre 129-Gt(ROSA)26Sor/ /J reporter strain show Cre expression that correlates with loss of DG expression ( white arrows, red, IIH6). Tissues stained with secondary antibodies onlyy were used d as a negative control. Muscle surrounding the bladder neck was used as a positive control (inset boxes). Nuclei were stained with DAPI (blue). Scale bars are 50 m.





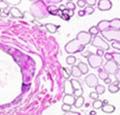





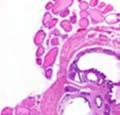


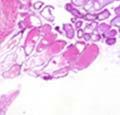


82 70 Figure 3.7. DG knockout mice have no abnormalities in tissue morph He ematoxyllin and eosin stains of DG null (n=6) and WT (n=6) hology. prostates were compared at three and six months of age.



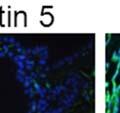

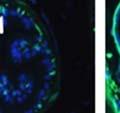

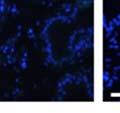
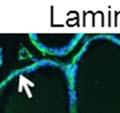
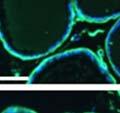
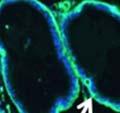
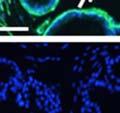




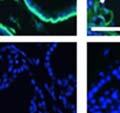


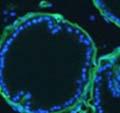

83 71 Figure 3.8. DG null prostates have normal localization of basement membrane pr roteins and basal cell number. DG ligands laminin and perlecan (arrows) are expressed and localized to the basement membrane of the dorsal lobe in KO prostates as indicated by immunofluorescencee staining. Keratin 5 positive (green) are present in similar quantities in DG KO (n=2) and WT (n=2) mice. Tissues stained with secondary antibody only are negative controls. DAPI staining of the nuclei is blue. Scale bars are 50 m.

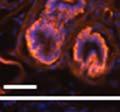
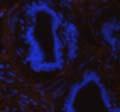
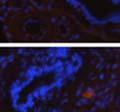
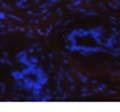



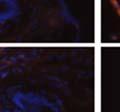
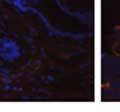


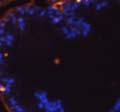
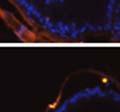






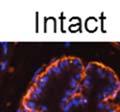
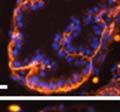
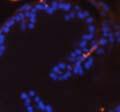
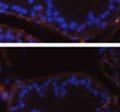
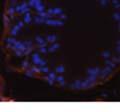
84 72 Figure 3.9. DG expresssion is absent from involuted and regenerated prostates. Im mmunofluorescent staining shows loss of DG (IIH6) from the dorsal lobe of involuted and regenerated DG KO prostates (n=3)) compared to WT mice (n=3). This staining is similar to intactt controls. The antibody controls were incubated with secondary antibody only. DAPI staining of the nuclei is blue. Scale bars are 50 m.


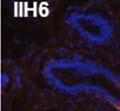




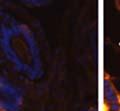

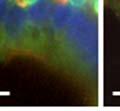

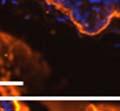

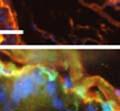







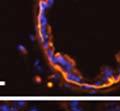
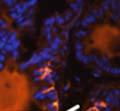


85 73 Figure DG expression in keratin 5 negative cells following androgen ablation and regeneration. Immunofluorescence staining indicates loss of DG (IIH6) in KO involuted and regenerated prostates (n=3). Small areas of IIH6 staining were retained in some glands of involuted and regenerated prostates similar to intact glands (arrowheads) ). K5+ basall cells (green) were 8D5 ( DG, red) negative (arrows) while K5- cells retained DG expression. DAPI stained nuclei are blue. Scale bars are 50 m.






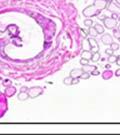

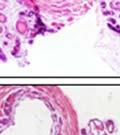

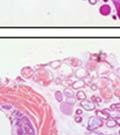



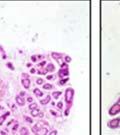
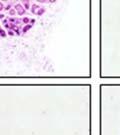
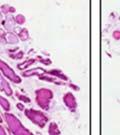





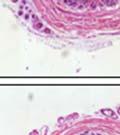
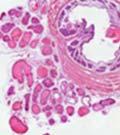

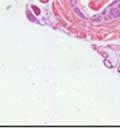



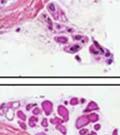



86 74 Figure DG KO prostates are able to regenerate following involution. Hematoxylin and eosin staining of DG KO and WT prostates. The prostates of DG null and WT mice were comparedd at 3 months of age (intact), 21 days post-surgical castration, 21 days post reintroduction of androgens and following an additional cycle of involution and regeneration.





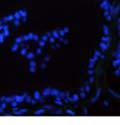
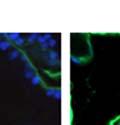

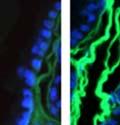



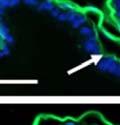


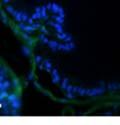
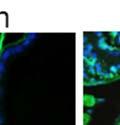


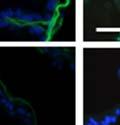





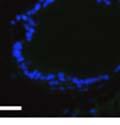



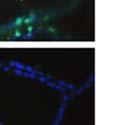
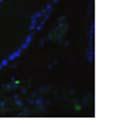
87 75 Figure DG KO regenerated prostates have normal localization of basement membrane proteins and basal cell number. DG ligands laminin (arrowheads) and perlecan (arrows) were expressed and localized to the basement membrane as indicated by immunofluorescence staining. Keratin 5 positive are present in similar quantities in DG KO regenerated prostates (n=3) and WT mice (n=3). Tissues stained with secondary only controls serve as negative controls. Nuclei stained with DAPI are blue. Scale bars are 50 m.












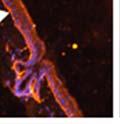

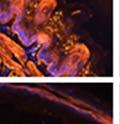




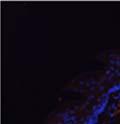





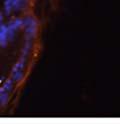
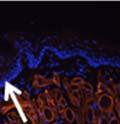




88 76 Figure Keratin-5 driven Cre expression deletes DAG1 from many stratified epithelia. DGG was lost from the stratified epithelia of the ear, tongue, hind- are stomach and trachea as shown by IIH66 immunofluorescence (arrows). Asterisk indicates stroma underlying the epithelium. DAPI stained nuclei blue. Scale bars indicate 50 m.














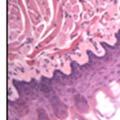
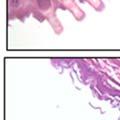









89 77 Figure DG is not required for tissue morphogenesis in some stratified epithelia as assessed by histology. Hematoxylin and eosin stains of the ear, tongue, hind-stomach and trachea. Scale bar iss 50mm.



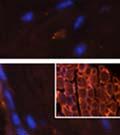

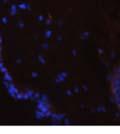
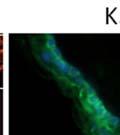




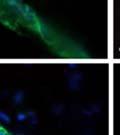
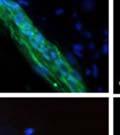


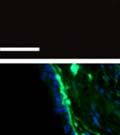
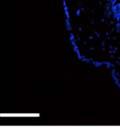


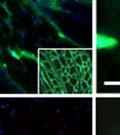


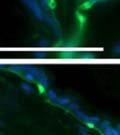
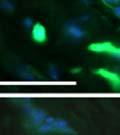
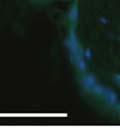
90 78 Figure DG ligand expression and localization were unchanged in the epidermis. A) Immunofluorescence staining of DG with the IIH6 antibody in WT and DG KO mice. B) Keratin 5 staining off the epidermis. C and D) Laminin and perlecan are expressed and localized to the basement membrane in DG KO mice. Muscle and secondary antibody only (control) staining is shown as controls. Nuclei were stained with DAPI (blue). Scale bars are 50 m.



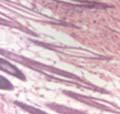

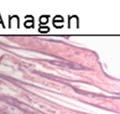



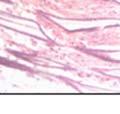
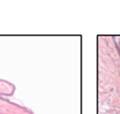



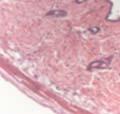





91 79 Figure The DG null epidermis had normal morphology at the anagen and telogen phases of the hair cycle. H& &E of WT and KO skin at the anagenn and telogen phase of the hair cycle.

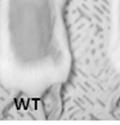

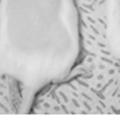
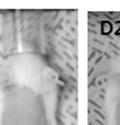













92 80 Figure The progression of the hair cycle was unchanged with loss of DG in the epidermis. WT and KO animals weree shaved at the telogen (resting) phase of the hair cycle. Hair regrowth was observed through the anagen (growth) phase.

93 81 Figure Loss of DG reduced the rate of wound healing in the epidermis. Wounds were generated at day 0 in DG WT and KO mice. Images were taken every two days until wound closure at day 10. Percent wound closure was quantified with Image J software.
















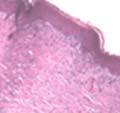

94 82 Figure Wound histology in DG WT and KO mice. Wounds were harvested at days six and ten and stained by H&E.









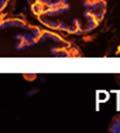
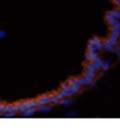



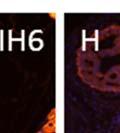


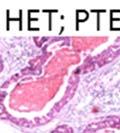
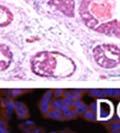
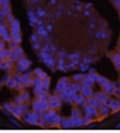
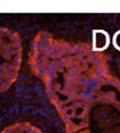





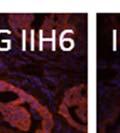

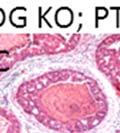

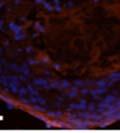



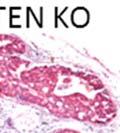



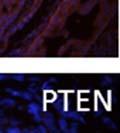

95 83 Figure Prostate specific deletion of DAG11 and PTEN in the lateral lobe. Loss of PTEN results in prostate cancer with a significantt stromal response in the lateral lobe (A-C). DG and PTEN protein is absent in the KO mice (F, I and K,,L) compared to controls (D, E and G) respectively. PTEN loss does not affect DG glycosylation (G,H). 100X magnification (A-C), bar indicates 100 M. 200X magnification (G-I), bar indicates 50 m. 400X magnification (D-F, J-L), bar indicates 20 m.




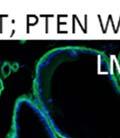
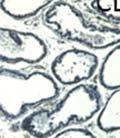





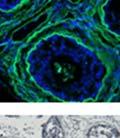





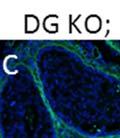



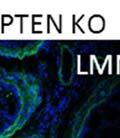


96 84 Figure Modulaton of the stroma and vasculature in the lateral prostate of DG PTEN KO mice. Lamininn staining is reduced and blood vessel density increased in the stroma of DG PTEN KO mice (C and F, I) compared with DG HET PTEN KO mice (B and E, H). 100X magnification (D-F), bar indicates 100 M. 200X magnification (A-C), bar indicates 50 M.
97 85 CHAPTER IV LOSS OF LARGE2 EXPRESSION IS A MECHANISM FOR HYPOGLYCOSYLATION OF DYSTROGLYCAN IN PROSTATE CANCER Introduction DG is expressed in many epithelial tissues where it is thought to mediate cellmatrix interactions (Durbeej, Henry et al. 1998). Disruption of cell-matrix interactions contributes to cancer progression. Reduced expression of DG, DG and/or DG, has been shown in a variety of adenocarcinomas including prostate and is associated with increased tumor aggressiveness and loss of extracellular matrix integrity (Henry, Cohen et al. 2001). The mechanism behind loss of DG expression and the resulting effect on tumor progression are poorly understood. DG is glycosylated by six known and putative glycosyltransferases (POMT1, POMT2, POMGnT1, Fukutin, FKRP, LARGE). Mutations in these enzymes result in hypoglycosylation of DG and loss of ligand binding (van Reeuwijk, Grewal et al. 2007). Many of these glycosyltransferases have been implicated in diseases of muscle and neural tissue in mice and humans. Recently, the posttranslational modification of DG necessary for ligand binding was shown to be a unique phosphorylated O-mannosyl glycan (Yoshida-Moriguchi, Yu et al. 2010; Hara, Kanagawa et al. 2011). This modification is mediated by the putative glycosyltransferase LARGE. Overexpression of LARGE has been shown to functionally compensate for hypoglycosylation of DG in cells from patients harboring mutations in other DG glycosyltransferases (Barresi, Michele et al. 2004). In addition, a mutation in LARGE is responsible for the myodystrophy mouse (Large myd ) phenotype caused by hypoglycosylation of DG
98 86 indicating LARGE has an important role in the functional glycosylation of DG (Grewal, Holzfeind et al. 2001). In cancer, recent work by de Bernabé, et al. found that DG is hypoglycosylated as shown by reduced staining with glycosylation-sensitive DG specific antibodies, in a panel of metastatic cancer cell lines due to epigenetic silencing of LARGE (de Bernabe, Inamori et al. 2009). In prostate cancer cells, DG glycosylation status can be influenced by the glycosyltransferase 3GnT1, which may collaborate with LARGE in DG glycosylation (Bao, Kobayashi et al. 2009). Therefore LARGE may have an important role in DG function in cancer. Here we evaluate DG glycosylation status and the molecular mechanisms contributing to loss of DG glycosylation in prostate cancer cell lines. Surprisingly, we show that expression of LARGE2, a homolog of LARGE, correlates with DG glycosylation status in prostate cancer cell lines. We further demonstrate that endogenous LARGE2 can functionally glycosylate DG. In the mouse prostate, LARGE is not required for glycosylation of DG. We also show that LARGE2 mediated glycosylation of DG correlates with oncogene expression. This work shows for the first time that loss of LARGE2 expression is a molecular mechanism for DG hypoglycosylation in prostate cancer and that oncogene expression may modulate DG glycosylation status. Materials and Methods Cell Culture 22Rv1 (American Type Culture Collection, ATCC), human prostate carcinoma epithelial cell line derived from CWR22, PC-3, metastatic prostate adenocarcinoma from bone (ATCC), LNCaP (Michael Cohen), PC3E and TEM4-18 (PC3 derivatives, Henry Lab), PANC-1, PxPC-3, HT-29, ZR-75, MDA-MB-231 and GP2-293 packaging cells
99 87 (BD Biosciences) were grown with 10% FBS (HyClone, Logan, Utah) and 1% nonessential amino acids. Cultures were maintained at 37oC in 95% air and 5% CO2 and subcultured as needed. shrna Inhibition of LARGE2 In order to knockdown LARGE2 expression, plko1 lentiviral vectors targeting human LARGE2 were purchased from Open Biosystems (TRC collection, RHS4533- NM_152312). Five different constructs along with lentiviral second generation plasmids pspax2 and VSVg (kind gifts from the Trono Lab) were transfected into 293FT cells and culture supernatant from these cells was used to infect PC3E and 22Rv1 cells for 8 hours followed by puromycin selection (1.0 μg/ml). We were able to achieve 70%-80% knockdown of LARGE2 using hairpin construct sequence TRCN CCGGGA TCCACTGGAACTCACCAAACTCGAGTTTGGTGAGTTCCAGTGGATCTTTTTG and TRCN CCGGTGCCTATTCTCTCTACGACTACTCGAGTAGTCGTA GAGAGAATAGGCATTTTTG. Quantitative PCR qpcr analysis was performed as described previously (Svensson et al., 2007). Relative expression values were calculated using the comparative Ct method (Pfaffl, 2001). Primer pairs are as follows: human LARGE N3 forward primer: 5 - aatcttctgaggcgggaact-3, reverse primer 5 -ccggaactcatagcacaggt -3 ; human LARGE2 N2 forward primer 5 - gacctcaaggtgataccactg-3, reverse primer 5 - ctgctgcaactgctcagcac-3 ; 3GnT1 N1 5 - aacctgccggaagagagcttg-3, reverse primer 5 - ctcgcaggcctggctgattcg-3 ; POMT1 N1 forward primer 5 - ccacctgtgcaggtcccagc-3, reverse primer 5 - gatgtcccagctgtctttcc-3 ; POMT2 N1 forward primer 5 -
100 88 atgagaggattctgtgcatt-3, reverse primer 5 - agagtgaggcatcccgtgtc-3 ; POMGnT1 N1 forward primer 5 -gcggagattctgtcagacag-3, reverse primer 5 - tcatcatagtcttgctctgg-3 ; Fukutin N2 forward primer 5 -gggagtaccattctggctga-3, reverse primer 5 - ggaagtcctgcatcctgaaa-3 ; FKRP N2 forward primer 5 -gaactcccccagcctacaac-3, reverse primer 5 -gtctctccagtccccacaaa-3. Western Blot Protein lysates were prepared from sub-confluent cultures following scrape harvest in RIPA buffer (150 mm NaCl2, 1% NP40, 10 mm deoxycholate-na, 0.1% SDS, 50 mm Tris, ph 8.0; rocked at 4 C for 1 h, boiled 5 min) with protease inhibitors. Protein samples (50 mg, Lowery protein analysis) were electrophoresed (4-20% Tris-HCl gels) and transferred (PVDF) prior to antibody incubation. Antibodies were used as follows; IIH6 glycosylation sensitive DG, 1:500 overnight at 4 C in low salt buffer (50mM Tris, 100mM NaCl2,0.1% Tween 20), donkey anti-mouse IgM HRP, 1:2000 for 1h at room temperature (RT); sheep 5 core DG (Kevin Campbell), 1:100 overnight at 4 C, donkey anti-mouse IgG HRP, 1:2000 for 1h at RT; DG specific 8D5 (Novocastra/Leica) for 1:200 overnight at 4 C, donkey anti-mouse IgG, 1:2000 for 1hr at RT. b-actin (Sigma), 1:5000 for 1h at RT, goat anti-rabbit IgG HRP, 1:10,000 for 1h at RT, was utilized as a marker for protein loading. All secondary antibodies are from Jackson Immunoresearch. Flow Cytometry Analysis Cells grown to sub-confluency were detached (Versene, Invitrogen) and washed prior antibody incubation. The cells were then resuspended at 5 x 105 cells in 50 l FACS buffer (PBS % sodium azide + 5% BSA) + IIH6 antibody (1:100, Kevin Campbell) for 45 min on ice. Next, cells were washed and incubated with secondary antibody
101 89 (100 l FACS buffer plus 1:100, donkey anti-mouse IgM Dylight 488, Jackson Immunoresearch) for 30 minutes on ice in the dark. Cells were washed again and resuspended in 400 l of FACs buffer, and transferred to polystyrene FACs tube (BD Biosciences). Samples were analyzed using the Becton Dickinson FACScan at The University of Iowa Flow Cytometry Core Facility. Laminin-111 Binding Assay Cells grown to sub-confluency were detached (Versene, Invitrogen). 5 X 105 cells were pelleted and resuspended in LMN-111 (100 nmol, BD Biosciences) in FACS+ buffer (1% BSA, 1mM CaCl 2, 1mM MgCl 2 ) for 60 min. Cells were then washed in FACS+ buffer, resuspended in 100 l Laminin-111 antibody (Sigma) at 1:100 and incubated for 30 min. Cells were next washed in FACS+ buffer, resuspended in 100 l donkey anti-rabbit IgG Dylight 488 secondary (Jackson Immunoresearch) for 30 min. After an additional washing, cells were resuspended in 400 l FACS+ buffer and analyzed using the Becton Dickinson FACScan at The University of Iowa Flow Cytometry Core Facility. To inhibit laminin-111 binding, some samples were first incubated with DG IIH6 antibody (Kevin Campbell) at 1:100 in 100 l FACS+ buffer. All steps were done on ice. Immunofluorescence Prostate tissue was sectioned (5-7 µm), deparaffinized and rehydrated. Antigen retrieval was performed by 20 min boiling at 95 C followed by 20 min of cooling at RT in citrate buffer (82mM sodium citrate, 1.8mM citric acid in PBS, ph 6) Sections were blocked in blocking buffer (2% donkey serum, 1% BSA, 0.1% Triton X-100, 0.05% Tween in PBS) followed by 8D5 (1:100, Novocastra) primary antibody incubation
102 90 overnight at 25 C. Tissues were washed in blocking buffer and incubated with donkey anti-mouse IgG Cy3 (Jackson Immunoresearch) at 1:200 and DAPI counterstain (Sigma) for 1hr at RT. Results Heterogeneous Functional Glycosylation of DG in Prostate Cancer Cell Lines To determine if DG is expressed in metastatic prostate cancer cell lines, we examined DG expression by western blot analysis with antisera that detects the DG core protein. Staining for the core DG protein revealed that DG was detectable in all prostate cancer cell lines examined (Figure 4.1A). The observed molecular weight of the bands was within the range of ~80 to 150kDa, with 22Rv1 and PC-3 showing higher molecular weight species than LNCaP. The heterogeneous staining pattern is consistent with the fact that DG has a number of carbohydrate modifications, some of which are involved in IIH6 and ligand binding. Full length DG (43kDa) staining with 8D5 showed equivalent levels of expression in 22Rv1 and PC-3 cells and lower levels in LNCaP cells (Figure 4.1A). A previously described 31kDa proteolytic fragment is also detectable in 22Rv1 and PC-3 cells. This data indicates and DG are expressed in the metastatic prostate cancer cell lines examined. Proper glycosylation of DG at the cell surface is required for ligand binding. Flow cytometry analysis with the laminin binding glycan specific antibody IIH6 revealed heterogeneous DG glycosylation heterogeneous across prostate cancer cell lines (Figure 4.1B). Among the cell lines examined, 22Rv1 and PC-3E+ cells were positive for IIH6 staining while PC-3 cells have IIH6 positive (IIH6+) and IIH6 negative (IIH6-) subpopulations. TEM4-18 and LNCaP cell lines are negative for IIH6 staining. Core DG is present in LNCaP lysates indicating expression of the protein. Interestingly,
103 91 TEM4-18 cells, an aggressive subpopulation of PC-3 cells that were isolated following migration through an endothelial cell monolayer, are IIH6- (Drake, Strohbehn et al. 2009). PC-3E+ cells, a less aggressive subpopulation of PC-3 cells, are IIH6+ suggesting loss of DG is associated with a more aggressive phenotype. These data confirm that DG glycosylation is heterogeneous in prostate cancer lines. We next assessed whether DG glycosylation status correlates with laminin binding ability. IIH6+ 22Rv1, PC-3 and PC-3E+ cells were able to bind exogenous laminin-1 at the cell surface whereas IIH6- LNCaP and TEM4-18 cells lines were not (Figure 4.1C). Laminin binding was specific to DG as prior incubation with IIH6 inhibited all exogenous laminin-111 binding. Laminin staining was also detected in cells without addition of exogenous laminin-111 and is likely endogenous laminin bound during tissue culture. Therefore, functional glycosylation of DG in metastatic prostate cancer cell lines is variable. DG Glycosylation Correlates with LARGE2 mrna Expression To elucidate the molecular mechanism behind loss of functional DG glycosylation in prostate cancer cells, we compared mrna expression levels of the six known and putative DG glycosyltransferases in the PC-3 derived cell lines PC-3E+ (IIH6+) and TEM4-18 (IIH6-). The glycosyltransferase 3GnT1 may be required for LARGE function and was therefore included in the panel (Bao, Kobayashi et al. 2009). Across the glycosyltransferase panel, only a single glycosyltransferase, LARGE2, was significantly reduced. LARGE2 mrna was decreased 100 fold in TEM4-18 cells relative to PC-3E+ cells (Figure 4.2A). We next compared LARGE2 mrna levels relative to PC-3 cells (Figure 4.2B). Across the prostate cancer cell line panel, LARGE2 mrna is significantly reduced in TEM4-18 cells but not IIH6- LNCaP cells indicating
104 92 loss of LARGE2 expression may be one of several mechanisms for loss of functional glycosylation in prostate cancer. PC-3E and TEM4-18 cells had low levels of LARGE mrna (Ct = 30) while LNCaP cells had high expression levels (Ct=27). Therefore, LARGE mrna expression levels did not correlate with DG glycosylation status (Figure 4.2C). These data suggest loss of LARGE2 expression may be a mechanism for loss of DG function in prostate cancer. We next assessed whether LARGE2 expression also correlated with DG glycosylation in the prostate epithelium. We quantified mrna expression in nontransformed immortalized prostate epithelial cell lines compared with TEM4-18 cells that express low levels of LARGE and LARGE2 (Figure 4.2B and C). DG was glycosylated in the prostate epithelial lines PWR-1E and RWPE-1 by flow cytometry analysis (Figure 4.3A). LARGE2 mrna was expressed at greater levels in these lines than TEM4-18 cells (IIH6-) (Figure 4.3B). LARGE was expressed at levels less than TEM4-18 cells (Figure 4.3C). These data indicate LARGE2 is expressed at significantly higher levels than LARGE in prostate epithelial cell lines and may act in place of LARGE to functionally glycosylate DG in the prostate. LARGE2 Functionally Glycosylates DG in the Prostate LARGE is required for proper glycosylation of DG in skeletal muscle. Therefore, we next evaluated DG glycosylation status of prostate tissue from the myodystrophy (myd) mouse (Large myd, MDC1D) harboring a mutation in LARGE(Grewal, Holzfeind et al. 2001). Immunofluorescence staining with the glycosylation specific IIH6 antibody in a wild type mouse showed positive staining in the muscle surrounding the urethra and at the basal side of the prostate epithelium (Figure 4.4A). The myd mouse had loss of staining in the muscle as expected yet staining was present in the prostate epithelium indicating LARGE is not required for DG
105 93 glycosylation in the prostate. Tissue from a prostate specific dystroglycan knockout mouse was shown as a control and maintains IIH6 staining in the muscle with loss in the prostate gland. LARGE2 has been shown to functionally glycosylate DG in an overexpression system (Fujimura, Sawaki et al. 2005). To assess whether endogenous LARGE2 functionally glycosylates DG, we generated PC-3E+ (IIH6+) cells that express LARGE2 targeted shrna s. DG glycosylation (IIH6) was reduced by 77% with expression of shrna #147 and 83% with expression of shrna #148 compared with a vector control (Figure 4.4B). To confirm loss of ligand binding ability we assessed laminin-111 binding by flow cytometry. LARGE2 shrna control cells bound exogenous laminin-111 (Figure 4.4C). Laminin binding was reduced in the shlarge2 #147 cell line and was completely absent in the shlarge2 #148 cells. Prior incubation with the IIH6 antibody inhibited all laminin-111 binding indicating binding is specific to dystroglycan. Laminin was detected without the addition of exogenous laminin and likely represents endogenous laminin bound during tissue culture. These data show that LARGE2 is required for the functional glycosylation of DG. Alterations in LARGE2 expression may also act as a mechanism for loss of DG glycosylation in other epithelial cancers. We therefore assessed DG glycosylation status and glycosyltransferase mrna expression in breast, colon and pancreatic cancer cell lines. DG glycosylation was heterogeneous across the cell lines panel (Figure 4.5A). LARGE2 mrna levels correlated with glycosylation status. Specifically, the highly glycosylated breast cancer cell line ZR-75 and colon cancer line HT-20 expressed the greatest levels of LARGE 2 while pancreatic cancer lines BxPC3 and PANC-1 levels were reduced. The IIH6- MDA.MB231 breast cancer cells had very low levels of LARGE2 mrna expression (Figure 4.5B). Interestingly, LARGE mrna expression also correlates with DG glycosylation status (Figure 4.5C). However, in prostate and pancreatic lines LARGE2 is expressed at higher levels whereas in the colon and breast
106 94 cancer cell lines LARGE was expressed at higher levels as assessed by Ct value. Therefore, the utilization of LARGE or LARGE2 for DG glycosylation is dependent on tissue type. Oncogene Expression Modulates DG Glycosylation Although DG glycosylation is lost in many cancers, the manner in which transformation relates to loss of glycosylation is unknown. We assessed DG glycosylation status in a panel of immortalized prostate epithelial cell lines with expression of various oncogenes. PrEC-LH cells are immortalized and expressed glycosylated DG by IIH6 flow cytometry analysis (Figure 4.6A). PrEC-LHMK cells express myc and PI3K and also expressed glycosylated DG at greater levels than PrEC- LHMK cells. PrEC-LHSR cell express ras and small t-antigen. These cells do not stain for IIH6 greater than background levels although background staining was very high in this line. This data suggests oncogene expression may regulate glycosylation status. Quantitative PCR analysis of LARGE2 expression correlated with glycosylation status (Figure 4.6B). Furthermore LARGE expression did not correlate with glycosylation status (Figure 4.6C). Ligand binding experiments showed enhanced laminin-111 binding in PrEC-LHMK and reduced binding in PrEC-LHSR cells compared to PrEC-LH cells (Figure 4.6D). Laminin-111 binding was specific to dystroglycan as prior incubation with IIH6 inhibited all binding. These data suggest oncogene expression modulates DG glycosylation through regulation of LARGE2 expression in the prostate. Reduced Growth and Tumorigenicity with DG knockdown DG expression is reduced in high grade prostate cancer. We therefore assessed the functional consequences of loss of DG expression in prostate cancer cell lines. 22Rv1
107 95 DAG1 shrna1 and shrna4 expressing cell lines had a 50% and 70% reduction, respectively, by IIH6 flow cytometry analysis (Figure 4.7). Growth assays were performed over five days and cell growth was quantified by bioluminescence imaging (BLI). Loss of DG expression significantly enhanced growth in the cell line expressing DG shrna4 whereas DG shrna1 trended in that direction but did not reach significance (Figure 4.8). Cells were next plated onto laminin-111 coated plates for 1 hour, rinsed several times and quantitated by BLI. DG knockdown reduced adhesion in both cell lines (Figure 4.9). We next assessed anchorage independent growth in a soft agar assay. Loss of DG expression resulted in enhanced anchorage independent growth in both DG shrna cell lines (Figure 4.10). To determine DG function in vivo, DG vector and DG shrna 4 knockdown lines were injected subcutaneously into nude mice. Tumor growth was monitored by BLI (Figure 4.11A) and caliper measurements (Fig 4.11B) for 5 weeks, the time point at which the first animal was taken down due to tumor size. DG knockdown had no effect on tumor growth in vivo. However, tumors trended toward enhanced growth suggesting isolation and injection of the cells with the greatest level of DG knockdown may result in significantly enhanced tumor growth in vivo. Discussion Reduced DG glycosylation and/or DG expression has been shown in many adenocarcinomas including prostate, however, the mechanism(s) behind loss of DG expression and/or glycosylation as well as the resulting functional consequences remain unclear (Henry, Cohen et al. 2001). Here we have shown DG glycosylation is heterogeneous in a panel of metastatic prostate cancer cell lines and expression of the putative glycosyltransferase LARGE2 correlates with DG glycosylation. Furthermore, LARGE is not required for DG glycosylation in the mouse prostate. Knockdown of endogenous LARGE2 expression decreased DG glycosylation resulting in loss of
108 96 laminin binding. Furthermore, we have shown that LARGE2 mediated glycosylation of DG correlates with oncogene expression. Therefore, in the prostate, LARGE2 mediates DG glycosylation and acts as a mechanism for hypoglycosylation of DG in prostate cancer. Surprisingly, modulation of LARGE2 expression and DG glycosylation may be a response to oncogene activation. Loss of DG expression occurs in many cancers, however, whether this occurs through loss of the protein or hypoglycosylation of DG remains unclear. Similar to work in other cancers, we have shown that expression of / DG is retained in prostate cancer cell lines and DG glycosylation status was heterogeneous. Previous work has shown that loss of DG glycosylation correlates with aggressiveness in cancer cell lines (Bao, Kobayashi et al. 2009; de Bernabe, Inamori et al. 2009). Our lab has previously published work on two cell lines, PC3E+ and TEM4-18. PC-3E+ cells are a subpopulation of PC-3 cells sorted for E-cadherin expression while TEM4-18 cells were isolated for their ability to more efficiently migrate through an endothelial monolayer (Drake, Strohbehn et al. 2009). TEM4-18 cells have characteristics of an epithelial-tomesenchymal transition including loss of E-cadherin expression and up-regulation of Zeb1 and N-Cadherin. In addition, these cells are more aggressive than PC-3E+ cells in a murine metastatic colonization model. Here we have shown that PC-3E+ cells are IIH6+ and TEM4-18 cells are IIH6- supporting the idea that loss of DG glycosylation correlates with a more aggressive phenotype and potentially a mesenchymal phenotype. Due to the difference in IIH6 status, we utilized these two cell lines to determine the mechanism behind loss of functional glycosylation in prostate cancer. Interestingly, we found that the greatest difference in DG glycosyltransferase mrna expression between these cell lines was a one hundred fold reduction in LARGE2 expression in TEM4-18 cells. LARGE2 is a homolog of LARGE resulting from a gene duplication event (Grewal, McLaughlan et al. 2005). LARGE2 has the same catalytic domains as LARGE
109 97 but has a narrower expression profile in tissues. Therefore, LARGE2 has been predicted to function as a glycosyltransferase and may have a redundant function to LARGE. Further analysis showed LARGE2 expression correlates with DG glycosylation status in our prostate cancer cell line panel with the exception of the LNCaP cell line. These cells were hypoglycosylated yet still expressed LARGE 2 and LARGE. Since DG appears at a lower molecular weight (Figure 4.1A), the N-terminal end of DG could potentially be cleaved at an early time point, inhibiting LARGE or LARGE2 binding necessary for glycosylation (Kanagawa, Saito et al. 2004). This is supported by the fact that expression of LARGE in LNCaP cells did not restore DG glycosylation and no mutations were present in a LARGE2 sequence analysis (data not shown). Additionally, a mutation in dystroglycan could prevent LARGE and LARGE2 binding. This type of mutation has been shown in a muscular dystrophy patient (Hara, Balci-Hayta et al. 2011). However, LNCaP hypoglycosylation could also occur due to mutations in or decreased expression of glycosyltransferases preceding LARGE2 in the glycosylation pathway. We have shown that LARGE is not required for DG glycosylation in the mouse prostate suggesting the utilization of LARGE2 for DG glycosylation in the prostate. In addition, LARGE2 expression was significantly greater in prostate epithelial cell lines when compared to IIH6 negative TEM4-18 cells. LARGE expression was reduced in PWR-1E and RWPE-1 cells compared to TEM4-18 cells. Thus, LARGE2 may act as a prostate specific DG glycosyltransferase in place of LARGE. LARGE2 has been shown to functionally glycosylate DG in an overexpression system. We have demonstrated for the first time that endogenous LARGE2 can functionally glycosylate DG in prostate cancer cells. This data indicates a redundant function for LARGE and LARGE2. However, differences in tissue expression profiles suggest different tissues may utilize one over the other. We have shown in cancer cell lines that LARGE verses LARGE2 glycosylation of DG varies by tissue type. LARGE2
110 98 may predominate in the prostate and pancreas whereas LARGE is principally expressed in colon and breast cancers. Furthermore, LARGE is clearly required for DG glycosylation in skeletal muscle and brain. The redundant roles of LARGE and LARGE2 further explain the lack of involvement of tissues besides muscle and brain with loss of LARGE function (Grewal, Holzfeind et al. 2001). It would be interesting to determine if LARGE and LARGE2 glycosylation of DG is identical or if variations in glycans confer a functional difference. Future work will further elucidate the functional consequences of LARGE2 mediated DG hypoglycosylation. We have shown that LARGE2 mediates DG glycosylation in the prostate and loss of LARGE2 expression is a mechanism for hypoglycosylation of DG in prostate cancer. However, the association between loss of LARGE2 expression and DG hypoglycosylation with transformation and cancer progression remains unknown. We have shown for the first time that LARGE2 mediated glycosylation of DG may be modulated with oncogene expression. DG glycosylation was modulated with myc and PI3K expression as well as ras and small T-antigen expression. Future studies will determine which oncogene or combination of oncogenes is required for modulation of DG glycosylation. Previous work in breast cancer cell lines has shown DG to have roles in proliferation, laminin binding and migration with overexpression of LARGE (CITE). One study in prostate cancer has indicated a role for DG, following LARGE knockdown, in cell migration (CITE). However, none of these studies have directly assessed DG function through knockdown of the protein or the utilization of an in vivo tumor model. We have shown loss of DG in 22Rv1 prostate cancer cells enhanced growth on plastic and anchorage independent growth in soft agar. DG knockdown also reduced laminin- 111 adhesion, potentially resulting in enhanced cell motility. Although roles for DG have been demonstrated in vitro, these results do not always translate to enhanced growth in vivo. We show subcutaneous growth in vivo is not altered with loss of DG. Interestingly,
111 99 DG knockdown tumors trended towards enhanced growth and appeared to grow in two distinct groups. This suggests a certain level of DG knockdown may need to be reached to alter growth in vivo. It would be of interest to sort cells for the greatest level of DG knockdown in the DG shrna4 cell line and assess these cells in an additional subcutaneous tumor growth experiment. The current results suggest DG does not have a role in tumor growth, however, additional experiments are need to further assess its role. It would also be of interest compare the functional consequences of LARGE2 specific hypoglycosylation of DG with DG knockdown to assess laminin dependent verses independent functions of DG. We have shown loss of LARGE2 expression is a molecular mechanism for DG hypoglycosylation in prostate cancer. This work also underscores the importance of understanding the expression pattern and function of LARGE and LARGE2 in a tissue specific context. We have further discovered a relationship between oncogene expression and DG glycosylation status. Loss of dystroglycan enhances prostate cancer cell growth in vitro and may growth in a subcutaneous tumor model. Determining the molecular mechanisms behind DG hypoglycosylation and loss of DG function in cancer is essential for understanding the progression of prostate cancer and for the development of treatments.



 D DG core")
")




.")


 22Rv1,")







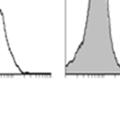




112 100 Figure 4.1. Functional glycosylation of DG iss heterogeneous in prostate cancer. A) D DG core protein (80-100kDa) was expressed in 22Rv1, PC-3 and LNCaP cells (sheep 5) ). DG (43 and 31kDA) was expressed in all prostate cancer cell lines (8D5). -actin was used as a loading controll (50ug). B) 22Rv1, PC-3 and PC-3E+ cell lines are positive for IIH66 staining while LNCaP and TEM4-18 cell lines are negative by FACS analysis. C) 22Rv1, PC-3 and PC-3E+ cells are positive for laminin-111 staining by FACS analysis following incubation with exogenous laminin-111 (100nM). LNCaP and TEM4-18 cells are negative. Priorr Incubationn with the antibody IIH6 inhibits laminin-111 binding. An isotype control was used for all FACS experiments (grey).











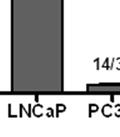


113 101 Figure 4.2. LARGE2 expression correlates with hypoglycosylation of DG. A) mr RNA analysis by qrt-pcr shows reduced LARGE2 expression in TEM4-18 cells compared to PC-3E+ cells. B) ) LARGE2 mrna is detectable in the prostate cancer cell line panel and is greatly reduced in TEM4-18 cells. C) LARGE mrna is detectable in the cell line panell and is reduced in PC-3E+ and TEM4-18 cells. Ct values are shown above the bars as GAPDH Ct/gene of interest (n= =3).






 LARGE2 mrna is")





114 102 Figure 4.3. LARGE2 is expressed in prostate epithelial cell lines. A) PWR-1E and RW WPE-1 prostate cell lines are positive for IIH6 staining by FACS. An iso otype control was used (grey). B) LARGE2 mrna is detectable and is expressed at significantly higher levels than TEM4-18 cells. C) LARGE mrna is detectable and is expressed at lower levels than TEM4-18 cells. Ct values are displayed GAPDH/gene of interest (n= =3).

 Immunoflouresence")
")

")






 or #148")



.")
 control")



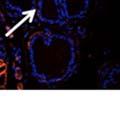
115 103 Figure 4.4. LARGE2 functionally glycosylates DG. A) Immunoflouresence staining sh hows DG glycosylation (IIH6) is present in a WT mouse prostate and the mydystrophy mouse (Largemyd, MDC1D) prostate (arrows). Staining is not present in a prostate specific DG knockout mouse. Muscle staining (asterisks) indicates tissue specific deletion. Scale bars are 50 m. B) PC-3E+ cells transfected with a LARGE2 targeted shrna #147 (green) or #148 (blue) reduced IIH6 staining by 86% and 77% respectively by FACS analysis. A vector (plko) expressing cell line was generated as a control (red). C) PC- 3E+ shlarge2 plko (red) control cells bind exogenous laminin-111. Incubation with IIH6 priorr to laminin-111 inhibited binding. Laminin antibody only was used as a background control. PC-3E+ shlarge2 #147 cells (green) have reducedd laminin-111 binding. PC-3E+ shlarge2 #148 cells (blue) were unable to bind laminin-111. All FACS experiments had an isotype control (grey).



 DG")






 qrt-pcr of")
")


.")
116 104 Figure 4.5. LARGE and LARGE2 expression is dependent on tissue type in cancer ce ell lines. A) DG glycosylation status (IIH6 staining) is heterogeneous across pancreatic, colon and breast cancer cell lines by FACS analysis. An isotype control was used (grey). B) qrt-pcr of LARGE2 mrna levels in pancreatic, colon and breast cancer cell lines. C) qrt-pcr of LARGE mrna expression in pancreatic, colon and breast cancer cell lines. Ct values are displayed as GAPDH/LAR RGE2 (n=3).







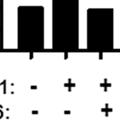








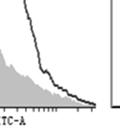






117 105 Figure 4.6. Oncogene expression modulates -dystroglycan glycosylation. A) Flow cy ytometry using the DG glycan specific antibody IIH6. PrEC-LH and PrEC- LHMK lines were positive. PrEC-LHSR cells were negative for IIH6 expression above background levels B) Quantitative PCR of LARGE2 mrna expression in PrEC-LH, PrEC-LHMK and PrEC-LHSR cells relative to TEM4-18 cells. C) Quantitative PCR of LARGE mrna expression in PrEC- are LH, PrEC-LHMK and PrEC-LHSR relative to TEM4-18 cells. Ct values displayed as GAPDH/gen e of interest (n=3).

 Flow cytometry")


.")

118 106 Figure 4.7. Generation of DG knockdown cell lines. A) Flow cytometry using the DG gly ycosylation specific antibody IIH6. Secondary antibody only is shown as a control (grey). DG was reduced 50% with DG shrna1 and 70% with DG shrna 4 compared to vector controls.








119 107 Figure 4.8. Loss of DG enhances proliferation.. Cells were plated at 6000 cells/well and qu uantitated by BLI at days 1 and 5. Data is displayed as fold growth. Growth was significantly increased in the DG shrna#4 cell line (pvalue 0.01).

 we ere")




120 108 Figure 4.9. DG knockdown reduces anchoragee to laminin-111. Cells (100,000/well) we ere added to Laminin-111 (10nM) coated plates for 1 hour. Adhered cells were quantitated following several rinses by BLI. Adhesion was significantly reduced in both DG shrna 1 and 4 knockdown cell lines (pvalue 0.01, pvalue respectively).








121 109 Figure Reduced DG expression enhancess anchorage independent growth. Cells were plated at 5000 cells/well in soft agar. Colonies were quantified after 3 weeks. Growth was enhanced in both DG shrna 1 and 4 lines (pvalue 0.01, pvalue 0.01 respectively).



122 110 Figure Tumor growth is not altered with loss of DG.. Mice were injected with 1 million 22Rv1 vector and DG shrna 4 cells expressing luciferase subcutaneously in nude mice. A and B) Growth was monitored by caliper measurement weekly.
Effects and regulation of dystroglycan glycosylation in cancer
 University of Iowa Iowa Research Online Theses and Dissertations Spring 2015 Effects and regulation of dystroglycan glycosylation in cancer Michael Raymond Miller University of Iowa Copyright 2015 Michael
University of Iowa Iowa Research Online Theses and Dissertations Spring 2015 Effects and regulation of dystroglycan glycosylation in cancer Michael Raymond Miller University of Iowa Copyright 2015 Michael
Histology = the study of tissues. Tissue = a complex of cells that have a common function
 { EPITHELIAL TISSUE Histology = the study of tissues Tissue = a complex of cells that have a common function The Four Primary Tissue Types: Epithelium (epithelial tissue) covers body surfaces, lines body
{ EPITHELIAL TISSUE Histology = the study of tissues Tissue = a complex of cells that have a common function The Four Primary Tissue Types: Epithelium (epithelial tissue) covers body surfaces, lines body
Dr. Heba Kalbouneh. Dr. Heba Kalbouneh. Dr. Heba Kalbouneh
 Dr. Heba Kalbouneh Dr. Heba Kalbouneh Dr. Heba Kalbouneh Basement membrane: What is the basement membrane? - It is a layer of ECM separating the epithelial cells from the underlying connective tissue Basement
Dr. Heba Kalbouneh Dr. Heba Kalbouneh Dr. Heba Kalbouneh Basement membrane: What is the basement membrane? - It is a layer of ECM separating the epithelial cells from the underlying connective tissue Basement
MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
Enzyme-coupled Receptors. Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors
 Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Protein Trafficking in the Secretory and Endocytic Pathways
 Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Signaling. Dr. Sujata Persad Katz Group Centre for Pharmacy & Health research
 Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
G-Protein Signaling. Introduction to intracellular signaling. Dr. SARRAY Sameh, Ph.D
 G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
Cells & Tissues. Chapter 3
 Cells & Tissues Chapter 3 Cell Theory Cell is structural and functional unit of life Activity of an organism is dependent upon its cells Principle of Complementarity functions of cells are dependent upon
Cells & Tissues Chapter 3 Cell Theory Cell is structural and functional unit of life Activity of an organism is dependent upon its cells Principle of Complementarity functions of cells are dependent upon
Mechanisms of Disease: congenital muscular dystrophies glycosylation takes center stage
 Mechanisms of Disease: congenital muscular dystrophies glycosylation takes center stage Paul T Martin SUMMARY Recent studies have defined a group of muscular dystrophies, now termed the dystroglycanopathies,
Mechanisms of Disease: congenital muscular dystrophies glycosylation takes center stage Paul T Martin SUMMARY Recent studies have defined a group of muscular dystrophies, now termed the dystroglycanopathies,
Tissues. tissue = many cells w/ same structure and function. cell shape aids its function tissue shape aids its function
 Tissues tissue = many cells w/ same structure and function cell shape aids its function tissue shape aids its function Histology = study of tissues 4 types of tissues Epithelial coverings contact openings
Tissues tissue = many cells w/ same structure and function cell shape aids its function tissue shape aids its function Histology = study of tissues 4 types of tissues Epithelial coverings contact openings
Tissue: The Living Fabric: Part A
 PowerPoint Lecture Slides prepared by Janice Meeking, Mount Royal College C H A P T E R 4 Tissue: The Living Fabric: Part A Tissues Groups of cells similar in structure and function Types of tissues Epithelial
PowerPoint Lecture Slides prepared by Janice Meeking, Mount Royal College C H A P T E R 4 Tissue: The Living Fabric: Part A Tissues Groups of cells similar in structure and function Types of tissues Epithelial
Cells and Tissues. Lesson 2.1: Molecules of Life Lesson 2.2: Cells Lesson 2.3: Tissues
 2 Cells and Tissues Lesson 2.1: Molecules of Life Lesson 2.2: Cells Lesson 2.3: Tissues Chapter 2: Cells and Tissues Lesson 2.1 Molecules of Life Molecules of Life carbohydrates proteins lipids nucleic
2 Cells and Tissues Lesson 2.1: Molecules of Life Lesson 2.2: Cells Lesson 2.3: Tissues Chapter 2: Cells and Tissues Lesson 2.1 Molecules of Life Molecules of Life carbohydrates proteins lipids nucleic
Lecture 3: Cells and Tissues. Bio 219 Dr. Adam Ross
 Lecture 3: Cells and Tissues Bio 219 Dr. Adam Ross Cell Physiology Cell Physiology Brief review of organelles Should be mostly review Cell surrounded by plasma membrane Lipid bilayer Also surrounds organelles
Lecture 3: Cells and Tissues Bio 219 Dr. Adam Ross Cell Physiology Cell Physiology Brief review of organelles Should be mostly review Cell surrounded by plasma membrane Lipid bilayer Also surrounds organelles
Histology Notes -Part 1: Epithelial Tissues
 Introduction Group of cells w/ similar structure & function = TISSUE Four Basic Tissue Types 1. Epithelial-covers 2. Connective-supports 3. Muscular*-produces movement (will discuss in the muscular system
Introduction Group of cells w/ similar structure & function = TISSUE Four Basic Tissue Types 1. Epithelial-covers 2. Connective-supports 3. Muscular*-produces movement (will discuss in the muscular system
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Significance and Functions of Carbohydrates. Bacterial Cell Walls
 Biochemistry 462a - Carbohydrate Function Reading - Chapter 9 Practice problems - Chapter 9: 2, 4a, 4b, 6, 9, 10, 13, 14, 15, 16a, 17; Carbohydrate extra problems Significance and Functions of Carbohydrates
Biochemistry 462a - Carbohydrate Function Reading - Chapter 9 Practice problems - Chapter 9: 2, 4a, 4b, 6, 9, 10, 13, 14, 15, 16a, 17; Carbohydrate extra problems Significance and Functions of Carbohydrates
Synapse Formation. Steven McLoon Department of Neuroscience University of Minnesota
 Synapse Formation Steven McLoon Department of Neuroscience University of Minnesota 1 Course News Midterm Exam Monday, Nov 13 9:30-11:30am Bring a #2 pencil!! 2 Course News Lecture schedule: Mon (Oct 31)
Synapse Formation Steven McLoon Department of Neuroscience University of Minnesota 1 Course News Midterm Exam Monday, Nov 13 9:30-11:30am Bring a #2 pencil!! 2 Course News Lecture schedule: Mon (Oct 31)
Cell and Tissue Types. Epithelial, Connective, Muscle, Nerve
 Cell and Tissue Types Epithelial, Connective, Muscle, Nerve Objectives Explain the major stages of the cell cycle and cellular division (mitosis). Describe specific events occurring in each of the phases
Cell and Tissue Types Epithelial, Connective, Muscle, Nerve Objectives Explain the major stages of the cell cycle and cellular division (mitosis). Describe specific events occurring in each of the phases
Hole s Human Anatomy and Physiology
 Hole s Human Anatomy and Physiology 1 Chapter 5 Tissues Four major tissue types 1. Epithelial 2. Connective 3. Muscle 4. Nervous 2 Epithelial Tissues General characteristics - cover organs and the body
Hole s Human Anatomy and Physiology 1 Chapter 5 Tissues Four major tissue types 1. Epithelial 2. Connective 3. Muscle 4. Nervous 2 Epithelial Tissues General characteristics - cover organs and the body
Lecture Overview. Chapter 4 Epithelial Tissues Lecture 9. Introduction to Tissues. Epithelial Tissues. Glandular Epithelium
 Visual Anatomy & Physiology First Edition Martini & Ober Chapter 4 Lecture 9 Lecture Overview Introduction to Tissues Location General characteristics Functions Classification Glandular Epithelium 2 Where
Visual Anatomy & Physiology First Edition Martini & Ober Chapter 4 Lecture 9 Lecture Overview Introduction to Tissues Location General characteristics Functions Classification Glandular Epithelium 2 Where
Dr. Heba Kalbouneh. Dr. Heba Kalbouneh. Dr. Heba Kalbouneh
 Dr. Heba Kalbouneh Dr. Heba Kalbouneh Dr. Heba Kalbouneh Tissue: is a group of cells that serve the same function, they are surrounded by extra cellular matrix. The 4 basic types of tissue: 1. epithelial
Dr. Heba Kalbouneh Dr. Heba Kalbouneh Dr. Heba Kalbouneh Tissue: is a group of cells that serve the same function, they are surrounded by extra cellular matrix. The 4 basic types of tissue: 1. epithelial
Epithelial Lecture Test Questions
 Epithelial Lecture Test Questions 1. Which of the following free surfaces lack(s) epithelia: a. lung alveoli (air sacs) b. hard palate c. joint cavities d. abdominal cavity e. salivary gland ducts 2. Which
Epithelial Lecture Test Questions 1. Which of the following free surfaces lack(s) epithelia: a. lung alveoli (air sacs) b. hard palate c. joint cavities d. abdominal cavity e. salivary gland ducts 2. Which
CHAPTER 05 Histology: EPITHELIUM
 BIO 211: ANATOMY & PHYSIOLOGY I 1 CHAPTER 05 Histology: EPITHELIUM Part 01: Brief Introduction Part 02: Survey of Types Dr. Lawrence G. G. Altman www.lawrencegaltman.com Some illustrations are courtesy
BIO 211: ANATOMY & PHYSIOLOGY I 1 CHAPTER 05 Histology: EPITHELIUM Part 01: Brief Introduction Part 02: Survey of Types Dr. Lawrence G. G. Altman www.lawrencegaltman.com Some illustrations are courtesy
TISSUES TYPES. CHAPTER 05 Histology: EPITHELIUM BIO 211: ANATOMY & PHYSIOLOGY I. HISTOLOGY = the study of tissues
 BIO 211: ANATOMY & PHYSIOLOGY I 1 CHAPTER 05 Histology: EPITHELIUM Part 01: Brief Introduction Part 02: Survey of Types Dr. Lawrence G. G. Altman www.lawrencegaltman.com Some illustrations are courtesy
BIO 211: ANATOMY & PHYSIOLOGY I 1 CHAPTER 05 Histology: EPITHELIUM Part 01: Brief Introduction Part 02: Survey of Types Dr. Lawrence G. G. Altman www.lawrencegaltman.com Some illustrations are courtesy
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
Muscular Dystrophy. Biol 405 Molecular Medicine
 Muscular Dystrophy Biol 405 Molecular Medicine Duchenne muscular dystrophy Duchenne muscular dystrophy is a neuromuscular disease that occurs in ~ 1/3,500 male births. The disease causes developmental
Muscular Dystrophy Biol 405 Molecular Medicine Duchenne muscular dystrophy Duchenne muscular dystrophy is a neuromuscular disease that occurs in ~ 1/3,500 male births. The disease causes developmental
Unit I Problem 9 Histology: Basic Tissues of The Body
 Unit I Problem 9 Histology: Basic Tissues of The Body - What is the difference between cytology and histology? Cytology: it is the study of the structure and functions of cells and their contents. Histology:
Unit I Problem 9 Histology: Basic Tissues of The Body - What is the difference between cytology and histology? Cytology: it is the study of the structure and functions of cells and their contents. Histology:
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION. Androgen deprivation therapy is the most used treatment of de novo or recurrent
 CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
Src-INACTIVE / Src-INACTIVE
 Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Essentials of Anatomy and Physiology, 9e (Marieb) Chapter 3 Cells and Tissues. Short Answer. Figure 3.1
 Chapter 3 Cells and Tissues. Short Answer. Figure 3.1") Essentials of Anatomy and Physiology, 9e (Marieb) Chapter 3 Cells and Tissues Short Answer Figure 3.1 Using Figure 3.1, match the following: 1) The illustration of simple cuboidal epithelium is. Answer:
Essentials of Anatomy and Physiology, 9e (Marieb) Chapter 3 Cells and Tissues Short Answer Figure 3.1 Using Figure 3.1, match the following: 1) The illustration of simple cuboidal epithelium is. Answer:
CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY. Ms. K. GOWRI. M.Pharm., Lecturer.
 CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY Ms. K. GOWRI. M.Pharm., Lecturer. Structure of cell: Human body develops from a single cell zygote which results from fusion of the ovum andd the spermatozoan.
CHAPTER II PDL 101 HUMAN ANATOMY & PHYSIOLOGY Ms. K. GOWRI. M.Pharm., Lecturer. Structure of cell: Human body develops from a single cell zygote which results from fusion of the ovum andd the spermatozoan.
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products
 stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
Biology. Dr. Khalida Ibrahim
 Dr. Khalida Ibrahim Biology Histology: Histology: is the study of the tissues of the body. Tissue: group of similar cells combined to perform a common function. The human body is composed of only 4 basic
Dr. Khalida Ibrahim Biology Histology: Histology: is the study of the tissues of the body. Tissue: group of similar cells combined to perform a common function. The human body is composed of only 4 basic
Tissue renewal and Repair. Nisamanee Charoenchon, PhD Department of Pathobiology, Faculty of Science
 Tissue renewal and Repair Nisamanee Charoenchon, PhD Email: nisamanee.cha@mahidol.ac.th Department of Pathobiology, Faculty of Science Topic Objectives 1. Describe processes of tissue repair, regeneration
Tissue renewal and Repair Nisamanee Charoenchon, PhD Email: nisamanee.cha@mahidol.ac.th Department of Pathobiology, Faculty of Science Topic Objectives 1. Describe processes of tissue repair, regeneration
Tissues. Definition. A group of similar cells and their intercellular substances specialized to perform a specific function.
 Chapter 4 - Tissues Tissues Definition A group of similar cells and their intercellular substances specialized to perform a specific function. Tissues Epithelial covers exposed surfaces, lines internal
Chapter 4 - Tissues Tissues Definition A group of similar cells and their intercellular substances specialized to perform a specific function. Tissues Epithelial covers exposed surfaces, lines internal
The Beauty of the Skin
 The Beauty of the Skin Rose-Anne Romano, Ph.D Assistant Professor Department of Oral Biology School of Dental Medicine State University of New York at Buffalo The Big Question How do approximately 50 trillion
The Beauty of the Skin Rose-Anne Romano, Ph.D Assistant Professor Department of Oral Biology School of Dental Medicine State University of New York at Buffalo The Big Question How do approximately 50 trillion
Histology 101! !! Name:! Block: Identify and describe the functions of major tissue types including their subclasses and varieties!
 Histology 101 Identify and describe the functions of major tissue types including their subclasses and varieties Name: Block: "1 Introduction to Tissues Histology Notes Tissue (living fabric) : groups
Histology 101 Identify and describe the functions of major tissue types including their subclasses and varieties Name: Block: "1 Introduction to Tissues Histology Notes Tissue (living fabric) : groups
Computational Systems Biology: Biology X
 Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#5:(October-18-2010) Cancer and Signals Outline 1 2 Outline 1 2 Cancer is a disease of malfunctioning cells. Cell Lineage: Adult
Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#5:(October-18-2010) Cancer and Signals Outline 1 2 Outline 1 2 Cancer is a disease of malfunctioning cells. Cell Lineage: Adult
Lecture Overview. Marieb s Human Anatomy and Physiology. Chapter 4 Tissues: The Living Fabric Epithelial Tissues Lecture 9. Introduction to Tissues
 Marieb s Human Anatomy and Physiology Marieb Hoehn Chapter 4 Tissues: The Living Fabric Epithelial Tissues Lecture 9 Lecture Overview Introduction to Tissues Epithelial Tissues Location General characteristics
Marieb s Human Anatomy and Physiology Marieb Hoehn Chapter 4 Tissues: The Living Fabric Epithelial Tissues Lecture 9 Lecture Overview Introduction to Tissues Epithelial Tissues Location General characteristics
Outline. Bio 105: Tissues Laboratory. Organization of the Human Body. Tissue - Epithelium. Tissues 3/2/ Copyright 2009 Pearson Education, Inc
 Outline Bio 105: Tissues Laboratory Laboratory 5 Reading: Chapter 4 I. Cell to cell contact II. Body Cavities III. Membranes IV. Homeostasis V. Integumentary System I. Includes skin, hair and nails 1 2
Outline Bio 105: Tissues Laboratory Laboratory 5 Reading: Chapter 4 I. Cell to cell contact II. Body Cavities III. Membranes IV. Homeostasis V. Integumentary System I. Includes skin, hair and nails 1 2
Summary of Endomembrane-system
 Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Development: is the growth of an individual organism from a simple to a more complex or mature level. A slow process of progressive change
 1. Define the following terms (use your own words): development, growth, differentiation, histogenesis, organogenesis, morphogenesis, reproduction, tissue, organ, organ system, and organism. Development:
1. Define the following terms (use your own words): development, growth, differentiation, histogenesis, organogenesis, morphogenesis, reproduction, tissue, organ, organ system, and organism. Development:
BIOL212 Biochemistry of Disease. Metabolic Disorders - Obesity
 BIOL212 Biochemistry of Disease Metabolic Disorders - Obesity Obesity Approx. 23% of adults are obese in the U.K. The number of obese children has tripled in 20 years. 10% of six year olds are obese, rising
BIOL212 Biochemistry of Disease Metabolic Disorders - Obesity Obesity Approx. 23% of adults are obese in the U.K. The number of obese children has tripled in 20 years. 10% of six year olds are obese, rising
Structure & Function of Cells
 Anatomy & Physiology 101-805 Unit 4 Structure & Function of Cells Paul Anderson 2011 Anatomy of a Generalised Cell Attached or bound ribosomes Cilia Cytosol Centriole Mitochondrion Rough endoplasmic reticulum
Anatomy & Physiology 101-805 Unit 4 Structure & Function of Cells Paul Anderson 2011 Anatomy of a Generalised Cell Attached or bound ribosomes Cilia Cytosol Centriole Mitochondrion Rough endoplasmic reticulum
Intercellular indirect communication
 Intercellular indirect communication transmission of chemical signals: sending cell signal transmitting tissue hormone medium receiving cell hormone intercellular fluid blood neurocrine neurotransmitter
Intercellular indirect communication transmission of chemical signals: sending cell signal transmitting tissue hormone medium receiving cell hormone intercellular fluid blood neurocrine neurotransmitter
A Dystroglycan Mutation Associated with Limb-Girdle Muscular Dystrophy
 T h e n e w e ngl a nd j o u r na l o f m e dic i n e brief report A Dystroglycan Mutation Associated with Limb-Girdle Muscular Dystrophy Yuji Hara, Ph.D., Burcu Balci-Hayta, Ph.D., Takako Yoshida-Moriguchi,
T h e n e w e ngl a nd j o u r na l o f m e dic i n e brief report A Dystroglycan Mutation Associated with Limb-Girdle Muscular Dystrophy Yuji Hara, Ph.D., Burcu Balci-Hayta, Ph.D., Takako Yoshida-Moriguchi,
Tissue = groups of cells that are similar in structure and function
 Tissue = groups of cells that are similar in structure and function Types Epithelial - covering Connective - support Muscle - movement Nervous - control Membranes line body cavities and hold organs together
Tissue = groups of cells that are similar in structure and function Types Epithelial - covering Connective - support Muscle - movement Nervous - control Membranes line body cavities and hold organs together
The addition of sugar moiety determines the blood group
 The addition of sugar moiety determines the blood group Sugars attached to glycoproteins and glycolipids on the surfaces of red blood cells determine the blood group termed A, B, and O. The A and B antigens
The addition of sugar moiety determines the blood group Sugars attached to glycoproteins and glycolipids on the surfaces of red blood cells determine the blood group termed A, B, and O. The A and B antigens
Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine
 Dr. Diala Abu-Hsasan School of Medicine") Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine dr.abuhassand@gmail.com An overview of cellular components Endoplasmic reticulum (ER) It is a network of membrane-enclosed
Protein sorting (endoplasmic reticulum) Dr. Diala Abu-Hsasan School of Medicine dr.abuhassand@gmail.com An overview of cellular components Endoplasmic reticulum (ER) It is a network of membrane-enclosed
Propagation of the Signal
 OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
VIII Curso Internacional del PIRRECV. Some molecular mechanisms of cancer
 VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
CELL BIOLOGY - CLUTCH CH CELL JUNCTIONS AND TISSUES.
 !! www.clutchprep.com CONCEPT: CELL-CELL ADHESION Cells must be able to bind and interact with nearby cells in order to have functional and strong tissues Cells can in two main ways - Homophilic interactions
!! www.clutchprep.com CONCEPT: CELL-CELL ADHESION Cells must be able to bind and interact with nearby cells in order to have functional and strong tissues Cells can in two main ways - Homophilic interactions
Chapter 1: Cells and Tissues
 Chapter 1: Cells and Tissues Cells and Tissues Carry out all chemical activities needed to sustain life Cells are the building blocks of all living things Tissues are groups of cells that are similar in
Chapter 1: Cells and Tissues Cells and Tissues Carry out all chemical activities needed to sustain life Cells are the building blocks of all living things Tissues are groups of cells that are similar in
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Effects of Second Messengers
 Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Chapter 4 :Organization & Regulation of Body Systems
 Chapter 4 :Organization & Regulation of Body Systems 4.1 Types of tissues What is a tissue? A collection of cells of the same type that perform a common function There are 4 major tissue types in the body:
Chapter 4 :Organization & Regulation of Body Systems 4.1 Types of tissues What is a tissue? A collection of cells of the same type that perform a common function There are 4 major tissue types in the body:
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG)
") In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
Epithelial Tissues. Types of Epithelial Tissues: Lining of Kidney
 Epithelial Tissues Covers the entire body surface and most of the body s inner cavities Outer epidermis (skin) protects from injury and drying out Inner epidermal tissue (on internal surfaces) often serves
Epithelial Tissues Covers the entire body surface and most of the body s inner cavities Outer epidermis (skin) protects from injury and drying out Inner epidermal tissue (on internal surfaces) often serves
Phospholipase C γ Prof. Graham Carpenter
 Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Development of the Nervous System 1 st month
 Development of the Nervous System 1 st month day 1 - fertilization of egg day 6 - uterine implantation day 18 - trilaminar (3-layered) disc (blastoderm, embryo) ectoderm (dorsal) - nervous system and skin
Development of the Nervous System 1 st month day 1 - fertilization of egg day 6 - uterine implantation day 18 - trilaminar (3-layered) disc (blastoderm, embryo) ectoderm (dorsal) - nervous system and skin
Major Structures of the Nervous System. Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors
 Major Structures of the Nervous System Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors Nervous System Divisions Central Nervous System (CNS) consists
Major Structures of the Nervous System Brain, cranial nerves, spinal cord, spinal nerves, ganglia, enteric plexuses and sensory receptors Nervous System Divisions Central Nervous System (CNS) consists
2013 W. H. Freeman and Company. 12 Signal Transduction
 2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
4. A phospholipid is an example of organization at the level.
 1. Physiology is the study of a. the structures of anatomical features. b. cellular metabolism. c. processes that allow organisms to function. d. how organ systems develop from the embryo. 2. Mary spends
1. Physiology is the study of a. the structures of anatomical features. b. cellular metabolism. c. processes that allow organisms to function. d. how organ systems develop from the embryo. 2. Mary spends
C) You find that the Raf kinase is not constitutively active. What was necessary in the previous assay to show any Raf kinase activity?
 You find that the Raf kinase is not constitutively active. What was necessary in the previous assay to show any Raf kinase activity?") PROBLEM SET 3 1. You have obtained immortalized liver cells from a patient who died of Wilson s disease, an inherited disorder of copper metabolism marked by neuronal degeneration and hepatic cirrhosis.
PROBLEM SET 3 1. You have obtained immortalized liver cells from a patient who died of Wilson s disease, an inherited disorder of copper metabolism marked by neuronal degeneration and hepatic cirrhosis.
Dr. Abeer.c.Yousif. Histology -2 nd stage. What is histology?
 What is histology? Histology is the science of microscopic anatomy of cells and tissues, in Greek language Histo= tissue and logos = study and it's tightly bounded to molecular biology, physiology, immunology
What is histology? Histology is the science of microscopic anatomy of cells and tissues, in Greek language Histo= tissue and logos = study and it's tightly bounded to molecular biology, physiology, immunology
Tissues. Definition. A group of similar cells and their intercellular substances specialized to perform a specific function.
 Chapter 4 - Tissues Tissues Definition A group of similar cells and their intercellular substances specialized to perform a specific function. Tissues Epithelial covers exposed surfaces, lines internal
Chapter 4 - Tissues Tissues Definition A group of similar cells and their intercellular substances specialized to perform a specific function. Tissues Epithelial covers exposed surfaces, lines internal
Epithelium-1. Hanan Jafar BDS.MSc.PhD
 Epithelium-1 Hanan Jafar BDS.MSc.PhD General features Epithelium is an avascular tissue composed of cells that cover the exterior body surfaces and line internal closed cavities and tubes. It also forms
Epithelium-1 Hanan Jafar BDS.MSc.PhD General features Epithelium is an avascular tissue composed of cells that cover the exterior body surfaces and line internal closed cavities and tubes. It also forms
Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body
 Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body Question No. 1 of 10 A biopsy sample is obtained from a lesion on the right cheek of a male patient. A technician in the histology lab
Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body Question No. 1 of 10 A biopsy sample is obtained from a lesion on the right cheek of a male patient. A technician in the histology lab
Anatomy PHL 212. Dr. Dina A. A. Hassan. -
 Anatomy PHL 212 Dr. Dina A. A. Hassan Associate Professor College of Pharmacy (Female Section) Sattam Bin Abdulaziz University Al kharj / Kingdom of Saudi Arabia Email :- da.hassan@psau.edu.sa 1 Anatomy
Anatomy PHL 212 Dr. Dina A. A. Hassan Associate Professor College of Pharmacy (Female Section) Sattam Bin Abdulaziz University Al kharj / Kingdom of Saudi Arabia Email :- da.hassan@psau.edu.sa 1 Anatomy
Index. derm.theclinics.com. Note: Page numbers of article titles are in boldface type.
 Note: Page numbers of article titles are in boldface type. A Adhesion and migration, the diverse functions of the laminin a3 subunit, 79 87 Alopecia in epidermolysis bullosa, 165 169 Amblyopia and inherited
Note: Page numbers of article titles are in boldface type. A Adhesion and migration, the diverse functions of the laminin a3 subunit, 79 87 Alopecia in epidermolysis bullosa, 165 169 Amblyopia and inherited
Tumor microenvironment Interactions and Lung Cancer Invasiveness. Pulmonary Grand Rounds Philippe Montgrain, M.D.
 Tumor microenvironment Interactions and Lung Cancer Invasiveness Pulmonary Grand Rounds Philippe Montgrain, M.D. February 26, 2009 Objectives Review epithelial mesenchymal transition (EMT), and its implications
Tumor microenvironment Interactions and Lung Cancer Invasiveness Pulmonary Grand Rounds Philippe Montgrain, M.D. February 26, 2009 Objectives Review epithelial mesenchymal transition (EMT), and its implications
Epithelial Tissue. Functions include: 1. Protection 4. Absorption 2. Secretion 5. Filtration 3. Sensory reception
 Tissues There are 4 primary tissue types in the human body: 1. Epithelial (covering/lining) 2. Connective (support) 3. Muscle (movement) 4. Nervous (control) Epithelium Epithelial Tissue Covers the surface
Tissues There are 4 primary tissue types in the human body: 1. Epithelial (covering/lining) 2. Connective (support) 3. Muscle (movement) 4. Nervous (control) Epithelium Epithelial Tissue Covers the surface
5 Dr. Heba Kalbouneh
 5 Dr. Heba Kalbouneh Glandular epithelium Gland: Is a collection of epithelial cells the secrets a certain product, like: proteins, lipids and carbohydrates. Secretion : A certain material that is produced
5 Dr. Heba Kalbouneh Glandular epithelium Gland: Is a collection of epithelial cells the secrets a certain product, like: proteins, lipids and carbohydrates. Secretion : A certain material that is produced
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting
 Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
PROTEIN TRAFFICKING. Dr. SARRAY Sameh, Ph.D
 PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
Tissues. Group of cells that are similar in structure and function. 4 primary types. Epithelium (covering) Connective (support) Nervous(control)
 Connective (support) Nervous(control)") Tissues Tissues Group of cells that are similar in structure and function 4 primary types Epithelium (covering) Connective (support) Nervous(control) Epithelial tissue (epithelium) Lining, covering, and
Tissues Tissues Group of cells that are similar in structure and function 4 primary types Epithelium (covering) Connective (support) Nervous(control) Epithelial tissue (epithelium) Lining, covering, and
Skeletal Muscle and the Molecular Basis of Contraction. Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry
 Skeletal Muscle and the Molecular Basis of Contraction Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry Like neurons, all muscle cells can be excited chemically, electrically, and
Skeletal Muscle and the Molecular Basis of Contraction Lanny Shulman, O.D., Ph.D. University of Houston College of Optometry Like neurons, all muscle cells can be excited chemically, electrically, and
Unit II: Tissues and Integumentary System
 Unit II: Tissues and Integumentary System 2.1 - Tissues Chapter 4 Written Response #1 1. What is a tissue? 2. What are four major types of tissues? Tissue Definition: a group or mass of similar cells working
Unit II: Tissues and Integumentary System 2.1 - Tissues Chapter 4 Written Response #1 1. What is a tissue? 2. What are four major types of tissues? Tissue Definition: a group or mass of similar cells working
1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled
 Protein Targeting Objectives 1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled As a protein is being synthesized, decisions
Protein Targeting Objectives 1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled As a protein is being synthesized, decisions
Animal Tissue Culture SQG 3242 Biology of Cultured Cells. Dr. Siti Pauliena Mohd Bohari
 Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
KEY CONCEPT QUESTIONS IN SIGNAL TRANSDUCTION
 Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Tissues Review 4 type
 Tissues Review 4 type Tissues Definition: a group of closely associated cells that perform related functions and are similar in structure Between cells: nonliving extracellular material Four basic types
Tissues Review 4 type Tissues Definition: a group of closely associated cells that perform related functions and are similar in structure Between cells: nonliving extracellular material Four basic types
JBC Papers in Press. Published on February 24, 2009 as Manuscript C
 JBC Papers in Press. Published on February 24, 2009 as Manuscript C900007200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.c900007200 LARGE repression in epithelium-derived cancer LOSS
JBC Papers in Press. Published on February 24, 2009 as Manuscript C900007200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.c900007200 LARGE repression in epithelium-derived cancer LOSS
Chapter Skeletal Muscle Structure and Function
 Chapter 10.2 Skeletal Muscle Structure and Function Introduction to Muscle Physiology Movement is a fundamental characteristic of all living things All muscle cells (skeletal, cardiac, and smooth) are
Chapter 10.2 Skeletal Muscle Structure and Function Introduction to Muscle Physiology Movement is a fundamental characteristic of all living things All muscle cells (skeletal, cardiac, and smooth) are
A rare case of muscular dystrophy with POMT2 and FKRP gene mutation. Present by : Ghasem Khazaei Supervisor :Dr Mina Mohammadi Sarband
 A rare case of muscular dystrophy with POMT2 and FKRP gene mutation Present by : Ghasem Khazaei Supervisor :Dr Mina Mohammadi Sarband Index : Congenital muscular dystrophy (CMD) Dystroglycanopathies Walker-Warburg
A rare case of muscular dystrophy with POMT2 and FKRP gene mutation Present by : Ghasem Khazaei Supervisor :Dr Mina Mohammadi Sarband Index : Congenital muscular dystrophy (CMD) Dystroglycanopathies Walker-Warburg
Lab 1 ANIMAL TISSUES
 Lab 1 ANIMAL TISSUES Levels of Organization Animals are multicellular heterotrophs whose cells lack cell walls. Most animals exhibit a hierarchical level of organization: Cells are organized into tissues
Lab 1 ANIMAL TISSUES Levels of Organization Animals are multicellular heterotrophs whose cells lack cell walls. Most animals exhibit a hierarchical level of organization: Cells are organized into tissues
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Inner ear development Nervous system development
 Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
A. Major parts 1. Nucleus 2. Cytoplasm a. Contain organelles (see below) 3. Plasma membrane (To be discussed in Cellular Transport Lecture)
 3. Plasma membrane (To be discussed in Cellular Transport Lecture)") Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
TRANSPORT PROCESSES. 1b. moving proteins into membranes and organelles
 1b. moving proteins into membranes and organelles SLIDE 1 A typical mammalian cell contains up to 10,000 different kinds of proteins. The vast majority of these proteins are synthesized by cytosolic ribosomes,
1b. moving proteins into membranes and organelles SLIDE 1 A typical mammalian cell contains up to 10,000 different kinds of proteins. The vast majority of these proteins are synthesized by cytosolic ribosomes,
Basic Histology. By Mrs. Bailey
 Basic Histology By Mrs. Bailey Primary Tissues 1. Epithelial Tissue 2. Connective Tissue 3. Muscle Tissue 4. Nervous Tissue Very cellular Supported by underlying connective tissue Epithelial & connective
Basic Histology By Mrs. Bailey Primary Tissues 1. Epithelial Tissue 2. Connective Tissue 3. Muscle Tissue 4. Nervous Tissue Very cellular Supported by underlying connective tissue Epithelial & connective
Signal Transduction Cascades
 Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Chapter 4 - Epithelial Tissues
 Chapter 4 - Epithelial Tissues Tissues Definition A group of closely associated cells that work together to perform a specific function Types Epithelial - covering Connective - support Muscle - movement
Chapter 4 - Epithelial Tissues Tissues Definition A group of closely associated cells that work together to perform a specific function Types Epithelial - covering Connective - support Muscle - movement
Regulation of cell function by intracellular signaling
 Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
LQB383 Testbank. Week 8 Cell Communication and Signaling Mechanisms
 LQB383 Testbank Week 8 Cell Communication and Signaling Mechanisms Terms to learn match the terms to the definitions --------------------------------------------------------------------------------------------------------------------------
LQB383 Testbank Week 8 Cell Communication and Signaling Mechanisms Terms to learn match the terms to the definitions --------------------------------------------------------------------------------------------------------------------------
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001
 BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
Tissues. tissue = many cells w/ same structure and function. cell shape aids function tissue shape aids function. Histology = study of tissues
 Tissues tissue = many cells w/ same structure and function cell shape aids function tissue shape aids function Histology = study of tissues 4 types of tissues Epithelial coverings contact openings Connective
Tissues tissue = many cells w/ same structure and function cell shape aids function tissue shape aids function Histology = study of tissues 4 types of tissues Epithelial coverings contact openings Connective
Cell Biology (BIOL 4374 and BCHS 4313) Third Exam 4/24/01
 Third Exam 4/24/01") Cell Biology (BIOL 4374 and BCHS 4313) Third Exam 4/24/01 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. For multiple choice questions,
Cell Biology (BIOL 4374 and BCHS 4313) Third Exam 4/24/01 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. For multiple choice questions,