SUPPLEMENTARY INFORMATION
|
|
|
- Annabella Norman
- 5 years ago
- Views:
Transcription
1 doi: /nature06173 SUPPLEMENTARY INFORMATION Contents of Supplementary Figures Supplementary Figure 1: Purification of mouse EHD2. Supplementary Figure 2: Analytical ultracentrifugation analysis. Supplementary Figure 3: EM analysis of liposome deformation. Supplementary Figure 4: Liposome-dependence of the stimulated ATPase reaction. Supplementary Figure 5: Alignment with secondary structure assignment. Supplementary Figure 6: Conservation plot of the dimerisation interface. Supplementary Figure 7: Interactions of the EH-domain. Supplementary Figure 8: Nucleotide specificity in comparison to Ras. Supplementary Figure 9: Curvature-dependence of liposome binding. Supplementary Figure 10: The phosphate cap. Supplementary Figure 11: Characterisation of mutations in the oligomerisation interface. Supplementary Figure 12: Model of the oligomeric ring. Supplementary Table 1: Data collection statistics. Supplementary Notes: References of Supplementary Material. 1

2 Supplementary Fig. 1: Expression and purification of mouse EHD2. Mouse EHD2 was expressed in Escherichia coli as a His-fusion protein as described in Materials and Methods. NI - Non-induced culture. I - Induced culture. SN - Soluble extract. FT Soluble extract after application to NiNTA Sepharose. E1- EHD2 after elution from NiNTA-Sepharose. E2 EHD2 after dialysis and thrombin cleavage. E3 EHD2 after reapplication and elution from the NiNTA column. This protein was further purified by sizeexclusion chromatography using a Sephadex S200 column (data not shown). 2
3 Supplementary Figure 2: Ultracentrifugation analysis indicates that EHD2 is a dimeric protein. Sedimentation velocity experiments were performed as described in methods at 300 mm NaCl. Selected scans (at equal, ~15 min intervals), and of g(s 20, w ) (the amount of material sedimenting between s 20,w and (s 20,w + s)) and also the residuals for fitting the data with DCDT+, were plotted with the program profit v (Quantum soft, Switzerland). The fitted value is kda which corresponds well with the calculated mass of the dimer of 124 kda. We also observed an EHD2 dimer by size-exclusion chromatography and by dynamic light scattering (data not shown). At 50 µm protein concentration, the hydrodynamic radius did not change in the presence or absence of nucleotides (or in 150 mm versus 300 mm NaCl), as judged by dynamic light scattering experiments (data not shown). 3

4 4

5 5
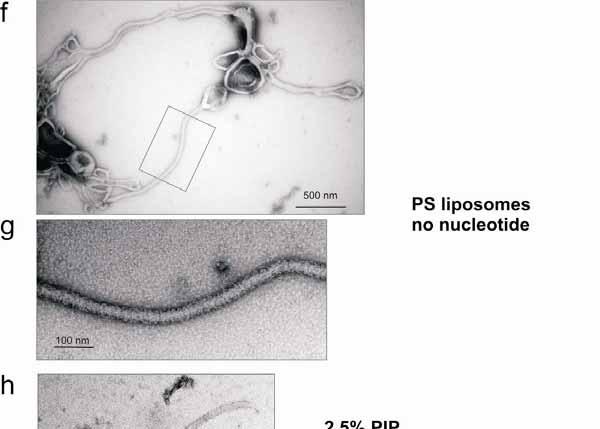
6 Supplementary Figure 3: EHD2 tubulation of liposomes. EHD2 was incubated with the indicated liposomes in the presence and absence of nucleotides and analysed by EM as described in Methods. a, EHD2 deformed PS liposomes into tubular networks, here in the presence of ATP- -S. b, Enlarged views of the indicated area in a. Note the presence of regularly spaced EHD2 rings (some are indicated with arrows). This even spacing may be due to the curvature stress generated by an EHD2 ring along the axis of the lipid tubule which might disfavour binding of the next ring in the direct vicinity (see below). c, Some of the lipid tubules (especially at higher protein concentration) were tightly packed with EHD2 oligomeric rings. d,e In the presence of ADP and in the absence of nucleotide (f,g), EHD2 also tubulated PS liposomes and formed ring-like structures around the tubules. e,g are enlarged views of the indicated areas in d and f, respectively. We did not observe a noticeable change of size of the tubules with the different nucleotide conditions. h,i,j, Under less favourable lipid binding conditions (synthetic liposomes with 87.5% phosphatidyl-choline, 10% cholesterol and only 2.5% PIP 2 ), EHD2 tubulated liposomes in the presence of ATP- -S (h) and ADP (i), but we did not observe tubulation in the absence of nucleotides (j). 6
7 Supplementary Figure 4: a, Membrane binding of 10 M EHD2 to 0.8 m filtered Folch liposomes of the indicated concentration in the presence of 1 mm MgCl 2 and 1 mm ATP- -S. b, ATP hydrolysis, as measured by HPLC analysis, under the same conditions as in a. c, Observed rate and % liposome-bound EHD protein (measured by densitometry). The rate increased with increasing EHD2 bound to liposomes. In all further ATPase experiments 1 mg/ml Folch was used to determine the maximal rate. For liposome binding experiments we used 0.33 mg/ml Folch (unless otherwise indicated) to be in the range where binding differences could be better observed. 7
8 mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : MVFGDVVLSLFIIIFFHFLSLHFLQVLFVCCIPRAVPLG METSSTISIGSCLKEHQKIYKEWFNIADSDGDGRVSGNDATKFFAMSKLSRQELKQVWAVADSKRQGFLGLSEFITAMKLVSLAQEGHEITSDLLKGSID MFSWLKKGG---ARGQRPEAIRTVTSSLKELYRTKLLPLEEHYRFGSFHSPALEDADFDGKPMV MFSWVSKDA---RRKKEPELFQTVAEGLRQLYAQKLLPLEEHYRFHEFHSPALEDADFDNKPMV MFSWLGTDD---RRRKDPEVFQTVSEGLKKLYKSKLLPLEEHYRFHEFHSPALEDADFDNKPMV MFSWMGRQAGGRERAGGADAVQTVTGGLRSLYLRKVLPLEEAYRFHEFHSPALEDADFENKPMI MSRWGRKN-----VKKAPEVIRTVTEGLKSLYRKKLLPLEQYYGFHDFHSPSLEDADFDNKPMV MFSWMGKNE---KTKKSPEVIHTVTEGLKDLYKKKLKPVEDFYRFHDFHSPALEDADFDNKPMV MFSWMGKES----AKGHQDVLQTVTGGLQSLYTGKLLPLEEHYRFHEFHSPALEEADFKNLPMV MFSFLKRE------KNTQEVVENVIGELKKIYRSKLLPLEEHYQFHDFHSPKLEDPDFDAKPMI MSNLFEEGQKKKKTRSMFSWLGGDS---SKKKNKEVLETVSEGLRKIYKQKLLPLEEFHKFHDFHSPALDDPDFDAKPMI MLSALKSS-----KPKDQEAYATVIEGLSKLYFSKLLPLENAYKFHDFHSPPLEKSDFESKPMV MKKLNVQE-----QKETDKLFATSTDALKSLYSSKIKPLEQLTKFGDFFSPTLTDADIAAKPMI MSLYMVERMRKLLY--RTEEETVVYDNVLEGLYSLYKTYILDLEKEFMYYHFYKPLLTSGDFLSKPMI MFGKKKQK------PQMDTSYVSVIDGVKKIYDEKIKKLEADYKYDYLVSPLMRQADFEAKPMV QGDTHARQEKGTNIGRPHVHLLVIVVVRLEIRLSTPQMSSTGVKTATESVAMEPEGLDELIEVLHTNYLKCVKPVEDMYKYDLFRPSWFEETILNQKPFV MSISGAAAPAPLRGRESGGNVPGSMGALIKKLHPLYTQRVRPLEEMYSFDVFRPSWYEETILNERPFI MKSVELPVLEGLENVVSKQKVSKTNVDVEDNVVTKPQVTAKTPWFKSKSIIKPQVNVVTIVDGLKRLYTEKLKPLEVTYRFNDFASPVLTSSDFDAKPMV α1a α1b α2 β1 : 39 : 100 : 61 : 61 : 61 : 64 : 59 : 61 : 60 : 58 : 77 : 59 : 59 : 66 : 58 : 139 : 68 : 200 Supplementary Fig.5 8
9 mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : P loop Switch 1 KPF NPF Switch 2 β1 α3 β2 β2a β2b αe β3 LVAGQYSTGKTSFIQYLLEQEVPGSRVGPEPTTDCFVAVMHGETEGTVPGNALVVDPE-KPFRKLNPFGNTFLNRFMCAQLPNQVLESISIIDTPGILSG LLVGQYSTGKTTFIRHLIEQDFPGMRIGPEPTTDSFIAVMHGPTEGVVPGNALVVDPR-RPFRKLNAFGNAFLNRFMCAQLPNPVLDSISIIDTPGILSG LLVGQYSTGKTTFIRYLLEQDFPGMRIGPEPTTDSFIAVMQGDMEGIIPGNALVVDPK-KPFRKLNAFGNAFLNRFVCAQLPNPVLESISVIDTPGILSG LLVGQYSTGKTTFIRYLLEQDFPGMRIGPEPTTDSFIAVMYGETEGSTPGNALVVDPK-KPFRKLSRFGNAFLNRFMCSQLPNQVLKSISVIDSPGILSG LVVGQYSTGKTTFIKYLLEQDVPGSRIGPEPTTDCFTAIMHGDVEGLIPGNALIVDPN-KPFRKLNPFGNTFLNRFQCAQLPNQVLESISIIDTPGILSG LVVGQYSTGKTTFIKYLLEQDIPGSRIGPEPTTDSFIAVMHGETEGVTPGNALMVDPN-KPFRKLNPFGNTFLNRFMCAHLPNKVLESISLIDTPGILSG LLVGQYSTGKTTFIRYLLEQDFPGMRIGPEPTTDSFIAVMYGEKDGSIPGNALVVDPK-KPFRKLNSFGNAFLNRFMCSQLPNEVLKSISIIDSPGILSG LLVGQYSTGKTTFIRYLLERDFPGIRIGPEPTTDRFIAVMYDDKEGVVPGNALVVDPK-KQFRPLSKYGNAFLNRFQCSSVASPVLNAISIVDTPGILSG LLVGQYSTGKTTFIRYLLESDFPGIRIGPEPTTDRFIAVMHGDEEGSIPGNALVVDAK-KQFRALSGFGNAFLNRFQCSTLPNQVLESVTIVDTPGILSG LLIGQYSTGKTTFIRYLIGEDFPGIRIGPEPTTDSFIAVMNDVRRGIIPGNALVMDHS-KQFRPLSKFGNGFLNRFQCAHMPNGVLEGITFIDTPGILSG LLLGQYSTGKTSFINYLLEKPFVGSNVAVEPSTDRFNAVMHGTDDRILPGNIVCVQSQDFPFKGLEKFGNGFMGRFQCSLSNAPILEKVSFIDTPGVLNI LLLGQYSTGKTTFIKHLIEKEYCGMRIGPEPTTDKFVAVMYNEKEQLIPGNALVSDIT-KPFSQLESFGNSFLSKLECSNTSSEVLKSVTIIDTPGVLSG LFLGQYSTGKTTFINYLLNYDYPGSHIGPEPTTDGFMAIMHGPNSTNIPGNTLCVQSD-KPFTSLSKFGNEFMAKFNGSFCNLPLLEHLTFIDSPGVLSG TFFGPWSSGKSTFINHLLQDNY--LWTGPQPTTAEFTVVLYGEEVGPVSGHVLASAKN-LPFKGLTEFGESFLGKFQGYRVPHELLKRVTLIDTPGILES TLFGPWSAGKTTFINYLLQSND--LWTGPQPTTAEFTVVMYGKEPGPVAGQALANSKH-LPFKGLLDFGESFIRNLKGFQAPHALLERVTLIDTPGVLES MLLGQYSTGKTTFIKHLLGCDYPGAHIGPEPTTDRFVVAMSGPDERTIPGNTMAVQAD-MPFNGLTSFGGAFLSKFECSQMPHPVLDQITLVDTPGVLSG Signature motif NKXD Interaction helix α4 β4 α5 β5 α6 β6 AKQR------VSRGYDFPAVLRWFAERVDLIILLFDAHKLEISDEFSEAIG-ALRGHEDKIRVVLNKADMVETQQLMRVYGALMWALGKVVGTPEVLRVY EKQR------ISRGYDFAAVLEWFAERVDRIILLFDAHKLDISDEFSEVIK-ALKNHEDKIRVVLNKADQIETQQLMRVYGALMWSLGKIINTPEVVRVY EKQR------ISRGYDFAAVLEWFAERVDRIILLFDAHKLDISDEFSEVIK-ALKNHEDKMRVVLNKADQIETQQLMRVYGALMWSLGKIVNTPEVIRVY EKQR------ISRGYDFCQVLQWFAERVDRIILLFDAHKLDISDEFSEAIK-AFRGQDDKIRVVLNKADQVDTQQLMRVYGALMWSLGKVINTPEVLRVY AKQR------VSRGYDFPAVLRWFAERVDRIILLFDAHKLEISDEFSEAIG-ALKGNEDKLRVVLNKADMVDTQQLMRVYGALMWSLGKVFGTPEVLRVY AKQR------VSRGYDFPAVLQWFAERVDRIILLFDAHKLEISDEFSEAIR-ALKGNEDKIRVVLNKADMVETQQLMRVYGALMWSLGKVFNTPEVLRVY EKQR------ISRGYDFCQVLQWFAERVDRIILLFDAHKLDISDEFSEAIK-AFRGQDDKIRVVLNKADQVDTQQLMRVYGALMWSLGKVINTPEVVRVY EKQR------IDRGYDFTGVLEWFAERVDRIILLFDAHKLDISDEFRRSIE-ALKGHDDKIRIILNKADMIDHQQLMRVYGALMWSLGKVLQTPEVARVY EKQR------IDRGYDFTGVLEWFAERVDRIILLFDAHKLDISDEFKRCIE-ALAGNEDKIRIVLNKSDMVDHQQLMRVYGALMWSLGKVFKTPEVSRVY EKQR------VDRGYDFSGVIEWLAERADRIILLFDAHKLDISDEFRRVIE-VLKSNEDKIRIVLNKADMIDSQQLMRVYGALMWSLGKILNTPEVARVY ITHGGNEGKKVGRSYDFPQIVEWFAERSDMILLLFDAHKLDISDEYREAIT-KLKGHDEKIRIVLNKADNINAQQLLRVYGGLMWSLGRVITTPEVKKVT IKQ ISRGYDFEKVIYWFAQRVDLILLIFDAHKLDISDEFRRCIQ-AIKGQDSKIRIILNKADTINTQQLMRVYGSLMWSLGIVINTPEVNRVY EKQR------IGRSYDFNEVVRWFAERADMIVLVFDAHKLDISDEFKGVIE-AVKKHSEKMRLVLNKADSIDSQQLMRVYGALMWSLGKVMQTPECLRVY VKDIR------ERQFDYVKVSRWFAERSDLIFILFDPSKLDAGIELKMMFKHAFRGMEGKVRIVLNKADSINTQELMRVYGSIFWNLSNLINCTEPPRVY AKDIH------QRKYDYVNVCRWFAERSDLIFVFFDPSKLDAGGELRQLFQTSFKGFENRLRLVLNKADTISTQELMRVYGSLFWNLSNFINTTEPPRVY EKQR------MQRSYDFTGVISWFASKCDMILLLFDPHKLDISDEFKRVIT-SLRGNEDKIRVVLNKADQVDTQQLMRVYGALMWSLGKVLNTPEVVRVY : 160 : 160 : 160 : 163 : 158 : 160 : 159 : 157 : 176 : 158 : 159 : 165 : 157 : 236 : 165 : 299 : 253 : 253 : 253 : 256 : 251 : 253 : 252 : 250 : 269 : 251 : 258 : 257 : 250 : 330 : 259 : 392 Supplementary Fig.5 9
10 β6 α7 α8 α9 α10 mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : IGSFWSQPLLVPDN----RRLFELEEQDLFRDIQG-LPRHAALRKLNDLVKRARLVRVHAYIISYLKKEMPTVFGKENKKKQLILKLPVIFAKIQLEHHI IGSFWSHPLLIPDN----RKLFEAEEQDLFKDIQS-LPRNAALRKLNDLIKRARLAKVHAYIISSLKKEMPNVFGKESKKKELVNNLGEIYQKIEREHQI IGSFWSHPLLIPDN----RKLFEAEEQDLFRDIQS-LPRNAALRKLNDLIKRARLAKVHAYIISSLKKEMPSVFGKDNKKKELVNNLAEIYGRIEREHQI IGSFWAQPLQNTDN----RRLFEAEAQDLFRDIQS-LPQKAAVRKLNDLIKRARLAKVHAYIISYLKKEMPSVFGKENKKRELISRLPEIYIQLQREYQI IGSFWSEPLMVTDN----RKLFELEEEDLFADIQN-LPRNAALRKLNDLVKRARLVRVHAHIISYLKQEMPSVFRKDNKKKNLIYQLPVIFSKIQLQHHI IGSFWSEPLMISDN----RRLFELEEQDLFQDIQN-LPRNSALRKLNDLVKRARLVRVHAHIISHLKKEMPSVFGKDNKKKQLINKLPVIFAKIQLEHHI IGSFWSKPLQITEN----RKLFEMEAQDLFRDIQS-LPRNAALRKLNDLIKRARLAKVHAYIISHLKKEMPAVFGKDTKKKELINKLPEIYTQLQREHQI IGSFWDQPLRFDAN----RRLFEDEEQDLFRDLQS-LPRNAALRKLNDLIKRARLAKVHAFIIAELRKDMPSVFGKDSKKKDLIKNLGQVYDRIQREHSI LGSFWDHPLHYDLN----RRLFQDEQHDLFQDLQA-LPRNAALRKLNDLIKRARLAKVHAYIIAELRKQMPSMIGKDKKKKDLIQNLDKIYEQLQREHNI IGSFWDRQLVFDTN----RKLFELEKMDLFRDLAT-LPANGTLRKLNDFIRRTRLAKVHAYVISYLKKEMPSMLGKDSKKKELINNLSKVYDTISRTQKI IGSFWSGPLQNKET----ENLLYSEMVDLIKDILL-LPKNGAIRKVNDLVKRSRLVKVHALILNHLRSEMP-VFGKEKKQAELIANLDKEFQKISLISRI IGSFWDKKLMHDEN----RTIFEEEASDLYKEISK-IPRNSTMVRLNDFIKRCRTLKVHIYLLTHLRKKLP-FFKKFLNKRKIVNSLEKIYEEVSKDYNL VSSFWDQPFKESLF----TGLFEKERDDLMYDLHA-LPKQATVRKVNELCKRARLAKANAYITSYLREQMP-TFGKDKKKAELLKDLNTVFNTVMKRYNL VGSFWDQPYKKGAFT----LLFTEEKTDLLHEIVDVVPQQARDKKVASLIRRAKEVMVHALIVGGMRSDLPLLFGKEKAKRKAIDNLQKTYEIMAAKYKM VGSFWDKPYSPNSFS----RLFAEEKLDLLHELLEVIPQQARDKKVASLIRRAKEVLVHAVILGGIRADLPLLFGKSKAKKKAAEQLPRRYELIGARYKM IGSFNDKPINEVAVGPIGKELFEKEQNDLLADLMD-VPKKACDRKINEFVKRARSAKINAYIMSHLKKEMPAMMGKSKAQQRLMDNLEEEFGKVQREFHL : 348 : 348 : 348 : 351 : 346 : 348 : 347 : 345 : 364 : 346 : 352 : 351 : 344 : 426 : 355 : 491 α11 α12a α12b GPF motif mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : SPGDFPDCQKMQELLMAHDFTKFHSLKPKLLEALDDMLAQDIAKLMPLL-RQEELES------VEAGVQGGAFEGT---RMGPFVER--GPDEAIEDGEE SPGDFPSLRKMQELLQTQDFSKFQALKPKLLDTVDDMLANDIARLMVMV-RQEESLM------PSQVVKGGAFDGT---MNGPFGHG------YGEGAGE SPGDFPNLKRMQDQLQAQDFSKFQPLKSKLLEVVDDMLAHDIAQLMVLV-RQEESQR------PIQMVKGGAFEGT---LHGPFGHG------YGEGAGE SAGDFPEVKAMQEQLENYDFTKFHSLKPKLIEAVDNMLSNKISPLMNLI-SQEETST------PTQLVQGGAFDGT---TEGPFNQG------YGEGAKE SPGDFPDCAKMQEQLMAHDFTKFKALKPNLMNMLDELLSSDIAKLMPLL-RQEEIEAG-----VQPGVQGGAFLGT---RAGPFVEGDPFGEVANENGEV SPGDFPDCAKMQEQLAIHDFKKFHALKPHMIEALDEMLTVDIAKLMPLL-RQEDLET------CDNVVQGGAFDGT---HNGPFIEG------STDGIFE SPGDFPEVKKMQQQLEMYDFSKFHAMKPKLIEAVDNMLANKIAPLMSMI-REEESSK------PTQMVHGGAFEGT---MDGPFGQG------YGEGAGE SPGDFPDIKKMQEVLQHQDFTKFHSLKPHLLDIVDNMLAKDIARLMEMIPQEEMTMV------ADPMVKGGAFEGVIDDHVSPFGYM------KGEGIDA SPGDFPDVNKMREKLQTQDFSKFNPLKPKLLEVVDGMLATDIARLMAQIPKEEAAAPAGSNGSADPTVRGGAFSQT-TEAETPFGFG-RYDARWGEGFDK SIGDFPNINRMQEYLEVHDFKTFSALQPKLIKVVDDMLANEVAKLVQMIPMERDAIVET----HGPAVTGGAFD----TSGTPFSFH------ASEGFEE PMGDFPDVDHYRTVLKVHDFTKFPKINEKMLAQLNEVLAVDFPNLLSRFPIDGTHKP SAYELNPFALD EVDEN PLGDFPPVQFMKEKLLDMDWMRIPKLETKKIDRINKVLNTHIPQLLEMIPKESVTVEVSR FENQEGTIVENKLTPFLEL TSGEIP AAGDFPPIDVYKDRLQNCDFSKFPKLDMRVINAVDDALGTQIPLLLKKFPMEDDLST HTNPFEVA-----SNFEKCCI NWKDFPPVEEYRTFLEKFDLEKFPEIEKAEKDGKIGALQKCIETTLPSMLRPVKSTTAADPRDKVQREKLQQMYLKSIRSQYEGKVGKQGSSDVVQPSHR NHRDFPPVQAYRSFLERFDVAKFPPLQKAEKAGLIRGIQELIDTILPSMLRPVCNTRAANPFEEDKQTGLLSMYRDHVLLQRDGRPGMQGGTDKVASTMG PAGDFPSVEHFREVLGGYNIDKFEKLKPKMIQAVDDMLGYDIPDLLKKFRNPYDN : 436 : 432 : 432 : 435 : 437 : 432 : 431 : 433 : 462 : 432 : 425 : 436 : 417 : 526 : 455 : 546 Supplementary Fig
11 mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : mmehd2 : hsehd1 : hsehd3 : hsehd4 : drehd : xlehd1 : xlehd4 : dmpast1 : cerme1 : sjehd : ddehd : pfehd : ehehd : tcehd : lmehd : atehd : GSEDDAEWVVTKDKSKYDEIFYNLAP-ADGKLSGSKAKTWMVGTKLPNSVLGRIWKLSDVDRDGMLDDEEFALASHLIEAKLEGHGLPTNLPRRLVPPSK G-IDDVEWVVGKDKPTYDEIFYTLSP-VNGKITGANAKKEMVKSKLPNTVLGKIWKLADVDKDGLLDDEEFALANHLIKVKLEGHELPADLPPHLVPPSK G-IDDAEWVVARDKPMYDEIFYTLSP-VDGKITGANAKKEMVRSKLPNSVLGKIWKLADIDKDGMLDDDEFALANHLIKVKLEGHELPNELPAHLLPPSK G-ADEEEWVVAKDKPVYDELFYTLSP-INGKISGVNAKKEMVTSKLPNSVLGKIWKLADCDCDGMLDEEEFALAKHLIKIKLDGYELPSSLPPHLVPPSH G-EEDEEWVVTKDKPKYDEIFYNLAP-NEGKLSGTKAKDWMVSTRLPNSVLGRIWKLSDVDRDGMLDDEEFALASHLIEVKLDGHGLPPELPARLVPPSK G-LDDEEWVVTKDKSKYDEIFFNLAP-TDGKITGTKAKNWMVTTKLPNSVLGKIWKLSDVDRDGMLDDEEFALASHLIEVKLEGHGLPPELPRHLIPPSK G-AEDEEWVVAKDKPVYDELFYTLSP-INGKISGVSAKKEMVNSKLPNSVLGKIWKLADNDKDGMLDDEEFALAKHLIKIKLEGYELPNDLPLHLVPPSQ G-YGEHEWICNKDKPRTDGIFNGLGP-VDGKISGATAKQELIKSKLPNSVLSKIWKLSDVDGDGFLDSDEFALALHLINVKLEGCELPTVLPEHLVPPSK G-ADEAEWVVSRERTTADSTFESLGP-VNGYLSGRAAKEHMVKSKLPNSVLGKVWKLADIDKDGQLDADEFALANYLINLKLEGHEIPSELPKHLIPPSK G-RFENDWIVNRFRQPWDEVFLALNP-MDGKISGEAARKHMLKSGLPNSTLRNIWILGDVDRDGCLDGDEFALVCYLMKLKLEGNELPPTLPAHLIPPSK V-RWTLFENVD--ISAFAPLFNSLNP-MDGKIQGQVAKGPLFQSGLPNNVLAHIWRLSDVDRDGKMDIEEFALAMHLVNVKLKGYELPETLPTTLIPFSK LWVKQKYLLSPIDTSKYSDDFYKLGPDDFGKLSGEQVKQDLIKSKLPSAVLHKIWNLADITEDGYLDLFEYSLARHFIEMKIEGFDLPAKVPKDIIK--- T-LDVSEDEMN---SYQKEFDKLPKN-EVGKVLGKDCFAPLMASGADKKDLQKIWSIADSGKEGALNAHQYILAKALVRTLLQSGGYPDNLPPVE----- ESAVTSSGGEGGNASAAPQQTTGPMTVTNSSSMPDQAQMFMGMMMQMMQQQQQQQQQQQQQQQQQRQQEQRHFVVEPVDEDNVD QGVPDSATTGPSSSVAAAPASGPSSLLGPGPAVTALSASPPFIELPSRAMAASSTAVAASSPPPPPPSSAAATSSPADMQAMMAIMQQMVAYQQQQQQQQ α13 βe1 α14 α15 βe2 α16 RRQKGSAE RRHE RKVAE RKSLPKAD RRQKGSDA RRQKGSAE RKSRLNAE RYD RGEQDPVYPSLNDNDE QNSNQIPRNGYHGD RGGMFGSNPNSSSYQTILSQPQQQQQQYQQQFINNYEQQQQQQQQQQ DRQSSSAPATVNSHGTLSSSSSETSGCLAESSVPVTPQESNLNSEPQPPTVVPQLGLPNSCATQISAWAGGDESKSQH Supplementary Figure 5: Multiple sequence alignment of the EHD2 family and secondary structure assignment. The following sequences were aligned (expasy accession number in brackets): Mus musculus EHD2 (Q8BH64), Homo sapiens EHD1 (Q9H4M9), Homo sapiens EHD3 (Q9NZN3), Homo sapiens EHD4 (Q9H223), Danio rerio EHD (Q6P3J7), Xenopus laevis EHD1 (Q7SYA1), Xenopus laevis EHD4 (Q7ZXE8), Drosophila melanogaster PAST-1 (Q8IGN0), Caenorhabditis elegans RME1 (Q966F0), Schistosoma japonicum EHD (Q5DG45), Dictyostelium discoideum EHD (Q54ST5), Plasmodium falciparum EHD (Q9NLB8), Entamoeba histolytica EHD (Q515L4), Trypanosoma cruzi EHD (Q4DYK9), Leishmania major EHD (Q4QDJ3S), Arabidopsis thaliana EHD (Q3EAA4). Completely conserved residues are boxed in red, conserved residues (> 50% sequence identity) are boxed in green. Helices are indicated as cylinders, -strands as blue arrows with colours according to the domain organisation (see Fig. 1a). Residues mutated in this study are indicated with a diamond ( ). : 543 : 534 : 535 : 541 : 543 : 538 : 537 : 534 : 576 : 544 : 568 : 633 : 535 : 530 : 530 : 533 : 535 : 530 : 529 : 531 : 560 : 530 : 521 : 533 : 507 : 610 :

12 Supplementary Figure 6: The EHD dimer interface. The alignment in Supplementary Fig. 5 was used to create a surface conservation plot of the EHD2 G-domain and helical domain, with conserved residues shown in purple and non-conserved residues shown in cyan. Helix 6 with its invariant W238 and the contiguous loop of the opposing EHD2 monomer are shown in orange interacting with the conserved surface. 12
13 13
14 Supplementary Figure 7: EH-domain interactions. a, The Ca 2+ -binding site of the EH-domain. Residues involved in Ca 2+ -binding and W490 at the bottom of the peptide binding pocket are in dark grey and the bound GPF-peptide from the opposing molecule is in light grey. Five direct ligands for the Ca 2+ -ion which has been maintained during purification could be identified which are the carboxy sidechains of D494, D496, D498, E505 and one main chain oxygen from M500 (mc500). The sixth ligands might be a water molecule not seen at the current resolution. b, Top and side view of the EHD2 monomer with the EH-domain linker shown in red and the NPF containing loop in magenta. Since the EH-domain linker is on the opposite face of the G-domain to the NPF containing loop, we argue that binding of the EH-domain to the NPF motif of the same molecule is sterically restricted. On the bottom, two adjacent monomers from the predicted EHD2 oligomer are shown (compare Fig. 4c). In this case, the EH-domain could bind without steric hindrance to the NPF containing loop of the adjacent monomer. c, Analysis of peptide binding to the EH-domain. The affinity of hepta-peptides from EHD2 containing the indicated xpf motifs to the EH-domain of EHD2 was measured by ITC at 10 C in 100 mm HEPES (ph 7.5), 50 mm NaCl, 200 µm CaCl 2. Peptides 118 PEKPFRKL and 418 RMGPFVER did not show detectable binding whereas binding of peptide 124 KLNPFGNT could be fitted to an affinity of M (n=1.33). This affinity is well within the range of other related EH-domain interactions 18. The weak interaction between the GPF motif and the EH-domain and the C-terminal tail with R536 anchored in the nucleotide binding site could well prevent EHD2 from self-assembling in solution since the local concentration of the EH-domain in the dimer is high. Upon concentration of EHD2 at the lipid surface, however, a switch of the EH-domain to the high affinity side-site NPF motif of the adjacent dimer might be favoured. Furthermore, the EH-domain might be displaced from the top site during oligomerisation since its C-terminal tail with R536 is pointing into the predicted G-domain oligomerisation interface and would clash upon oligomerisation. The presence of two (or more) lowaffinity interaction sites during protein complex assembly on membranes is a common theme also in clathrin-mediated endocytosis - it guarantees stable complex formation with a high degree of reversibility 41. Our data do not exclude the possibility that EHDs are initially recruited to sites of action by NPF motif containing binding partners. However, we did not observe any co-localisation of described EHD binding partners, pacsin1 and 2 (ref. 12) along the length of EHD tubules in vivo (data not shown). The importance of a flexible linker between the helical and the EH-domains is underpinned by a dominant negative mutant in C. elegans Rme1 which is a point mutant of this linker

15 Supplementary Figure 8: Comparison of the nucleotide-binding sites of Ras (pdb code 5p21, left) and EHD2 (right). Residues from the NKxD motif involved in specific nucleotide recognition are shown with hydrogen bonds indicated. Specific guanosine recognition in GTPases is mediated by a highly conserved NKxD motif (called G4) where the asparagine side-chain forms a hydrogen bond to the carbonyl group at carbon6 of the guanosine base and the aspartate side-chain forms a double hydrogen bond to nitrogen1 of the guanosine base and the amine group at carbon2 (left). Aspartate has been shown to be crucial since mutating it to asparagine in Ras reduces nucleotide affinity by more than 1000-fold 42. Directly following the NKxD motif, a large hydrophobic residue (leucine in Ras and dynamin, M223 in EHD2) lines the nucleotide binding pocket. The NKxD motif of EHD2 (starting at residue 219) is also highly conserved in EHD family members (Supplementary Fig. 5). The carboxyamide group of N219 forms a hydrogen bond to the C6 amino group of the adenine base (right). M223, whose side-chain is buttressed by the side chain of H192 (not shown), is closer to the purine base than the corresponding leucine residue in Ras and sterically excludes accommodation of an amino group at C2, thus explaining the inability of EHDs to bind to guanine nucleotides. Instead of forming a double hydrogen bond, the completely invariant D222 of EHD2 surprisingly forms a salt bridge with the conserved R536 which is supplied from the C-terminus of the superjacent EH-domain (right). This arginine is not contributing to nucleotide binding since the EHD without the EH-domain binds with the same affinity to ATP- -S (data not shown). 15

16 Supplementary Figure 9: Top: Electrostatic surface representation of EHD2. Red indicates negative charge and blue positive charge at neutral ph. The orientation of EHD2 is the same as in Fig. 2a. The membrane interaction site is highly curved which is a consequence of EHD2 dimerisation. The diameter of the sphere is approximately 7 nm, but this may not be so extreme if the tips of the loops (a phenylalanine and a valine residue) are inserted into the membrane and if the membrane-binding sites show some flexibility. Bottom: EHD binding to liposomes of different sizes in the presence and absence of 1 mm nucleotides and 1 mm MgCl 2 (P - pelleted fraction, S -supernatant). We used synthetic liposomes containing 87.5% phosphatidyl-choline, 10% cholesterol and 2.5% PIP 2 at the same protein/lipid ratio as in Supplementary Fig. 3. The protein:membrane interaction is more stringent (lower affinity interaction) to these liposomes than to Folch or PS liposomes, and this allows us to look for nucleotide and liposome size dependency (curvature preference) of the interaction. We argue that a protein with a curved dimer interface, like that in the EHD2 dimer, should show curvature preference for small liposomes which have a curvature closely matching the curvature of the membrane-binding interface of the EHD2 dimer. As a result, no or less energy is required to deform the lipid bilayer of small liposomes to the preferred curvature. Preferred binding to small liposomes is precisely what we observed but only in the nucleotide-free state. In the nucleotide-bound state, we previously observed limited oligomerisation of the residual EHD2 protein bound to these liposomes by electron microscopy (Supplementary Fig. 3). In our predicted EHD2 oligomer (see Supplementary Fig. 12), there are two curvatures, the one formed by the lipid-binding interface of the dimer and the other formed by the oligomer. These are orthogonal to each other. Thus, while the EHD2 dimer in the ATP-bound state may still be expected to bind with higher affinity to small liposomes, we observe that the EHD2 oligomer makes tubules and energy is required for this lipid bilayer deformation. An oligomer bound to a vesicle of non-ideal curvature will have an increased offrate, and thus we observed less binding in our experiments. The issue of curvature sensing versus curvature driving of membrane interacting proteins has been previously discussed

17 Supplementary Figure 10: Phosphate cap. The phosphates of the AMP-PNP molecule are covered by residues from switch I and the P-loop. This cap does not allow the insertion of a catalytic residue in trans into the active site. This so-called phosphate cap is also present in GBP1 (ref. 44) whose mechanism of GTP hydrolysis has been shown to involve dimerisation-dependent positioning of the attacking nucleophilic water molecule by a catalytic serine in cis from switch I (ref. 15). 17

18 Supplementary Figure 11: a, Membrane binding of oligomerisation-interface mutants to Folch liposomes (methods as in Fig. 1c). b, The K193D mutant, which in vitro does not display any liposome-stimulated ATPase reaction, showed extensive tubulation with essentially no punctuate staining when over-expressed as EGFP-tagged protein in HeLa cells (100% cells showed a tubular phenotype, n=53). On the contrary, overexpression of the E91Q mutant, which in vitro had only a mildly impaired stimulated ATPase reaction, resulted in a similar phenotype as wild type EHD2, with 82% of the examined low-level expressing cells showing a tubular and punctuate phenotype, 16% showing only punctuate staining and 2% a tubular phenotype (n=43). 18

19 Supplementary Figure 12: a, Top and side view of the EHD2 oligomeric ring model in a surface representation. For better clarity, the EH-domains are not included. The diameter of the embraced lipid tubule is 18 nm and the thickness of the EHD2 ring is 10 nm, in agreement to what is observed in the EM assays. Approximately twenty EHD2 dimers constitute one turn in this model. b, Arrangement of the EHD2 dimers in the oligomer. The high curvature of the membrane interaction site of EHD2 (Supplementary Fig. 9) is oriented perpendicular to the curvature of the lipid tubule. This arrangement is predicted to induce local curvature on the lipid tubule. c, In an alternative model, the oligomer could also form along the length of a tubule (parallel to its long axis) and thus maximise the use of the highly curved membrane interface of the dimer. However, such an arrangement was never observed by EM (Fig. 1d, Supplementary Fig. 3). Furthermore, our ring-based oligomer model is consistent with the absence of any curvature-sensitive membrane binding upon oligomerisation. 19
20 Supplementary Table 1 Data collection statistics Selenomethionine derivative Data collection Space group C2 Cell dimensions a, b, c (Å) 99.9, 134.7, 56.1 (º) 90, 106.5, 90 Wavelength Resolution (Å) * ( ) R sym * 7.8 (36.1) I/ I * (4.4) Completeness (%)* 96.4 (93.8) Redundancy * 3.4 (3.4) *Highest resolution shell is shown in parenthesis. 20
21 Supplementary notes / Supplementary References 23. Lenzen, C., Cool, R. H. & Wittinghofer, A. Analysis of intrinsic and CDC25-stimulated guanine nucleotide exchange of p21ras-nucleotide complexes by fluorescence measurements. Methods Enzymol. 255, (1995). 24. Van Duyne, G. D., Standaert, R. F., Karplus, P. A., Schreiber, S. L. & Clardy, J. Atomic structures of the human immunophilin FKBP-12 complexes with FK506 and rapamycin. J. Mol. Biol. 229, (1993). 25. Kabsch, W. Automatic Processing of Rotation Diffraction Data from Crystals of Initially Unknown Symmetry and Cell Constants. Journal of Applied Crystallography 26, (1993). 26. Sheldrick, G. M. & Schneider, T. R. in Macromolecular Crystallography, Pt B (1997). 27. de la Fortelle, E. & Bricogne, G. in Methods in Enzymology (eds. Carter, C. W., Jr & Sweet, R. M.) (1997). 28. McRee, D. E. XtalView/Xfit--A versatile program for manipulating atomic coordinates and electron density. J Struct Biol 125, (1999). 29. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. Refinement of Macromolecular Structures by the Maximum-Likelihood Method. Acta Cryst. D53, 240 (1997). 30. Laskowski, R. A., Macarthur, M. W., Moss, D. S. & Thornton, J. M. Procheck - a Program to Check the Stereochemical Quality of Protein Structures. Journal of Applied Crystallography 26, (1993). 31. Kraulis, P. J. Molscript - a Program to Produce Both Detailed and Schematic Plots of Protein Structures. Journal of Applied Crystallography 24, (1991). 32. Merritt, E. A. & Murphy, M. E. Raster3D Version 2.0. A program for photorealistic molecular graphics. Acta Crystallogr D Biol Crystallogr 50, (1994). 33. Landau, M. et al. ConSurf 2005: the projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Research 33, W299-W302 (2005). 34. Potterton, E., McNicholas, S., Krissinel, E., Cowtan, K. & Noble, M. The CCP4 moleculargraphics project. Acta Crystallographica Section D-Biological Crystallography 58, (2002). 35. DeLano, W. L. The PyMOL Molecular Graphics System (DeLano Scientific, Palo Alto, CA, USA, 2002). 36. Guex, N. & Peitsch, M. C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 18, (1997). 37. Collaborative Computational Project, N. The CCP4 suite: programs for protein crystallography. Acta Crystallogr. D 50, 760 (1994). 38. Philo, J. S. A method for directly fitting the time derivative of sedimentation velocity data and an alternative algorithm for calculating sedimentation coefficient distribution functions. Analyt. Biochem. 279, (2000). 39. Philo, J. S. Improved methods for fitting sedimentation coefficient distributions derived by time-derivative techniques. Analyt. Biochem. 354, (2006). 40. Laue, T. M., Shah, B. D., Ridgeway, T. M. & Pelletier, S. L. in Analytical Ultracentrifugation in Biochemistry and Polymer Science (eds. Harding, S. E., Rowe, A. J. & Horton, J. C.) (Royal Society of Chemistry, Cambridge, UK, 1992). 41. Schmid, E. M. et al. Role of the AP2 beta-appendage hub in recruiting partners for CCV assembly. PLoS Biol. 4, e262 (2006). 21
22 42. Schmidt, G. et al. Biochemical and biological consequences of changing the specificity of p21ras from guanosine to xanthosine nucleotides. Oncogene 12, (1996). 43. McMahon, H. T. & Gallop, J. L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 438, (2005). 44. Prakash, B., Renault, L., Praefcke, G. J., Herrmann, C. & Wittinghofer, A. Triphosphate structure of guanylate-binding protein 1 and implications for nucleotide binding and GTPase mechanism. EMBO J. 19, (2000). 22
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature07422 SUPPLEMENTARY INFRMATIN K S(P) R S I M Q(L4) R M 7 6 Sp Q I K R 5 4 3 2 1 L(L0) L L S E 0 +1 +2 +3 Figure S1a Difference electron density (mfo DFc) for the peptide (Qpeptide),
doi: 10.1038/nature07422 SUPPLEMENTARY INFRMATIN K S(P) R S I M Q(L4) R M 7 6 Sp Q I K R 5 4 3 2 1 L(L0) L L S E 0 +1 +2 +3 Figure S1a Difference electron density (mfo DFc) for the peptide (Qpeptide),
Introduction to proteins and protein structure
 Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random
. EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random") S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
Amino Acids. Review I: Protein Structure. Amino Acids: Structures. Amino Acids (contd.) Rajan Munshi
 Rajan Munshi") Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
(B D) Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.
 Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.") Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/2/4/e1500980/dc1 Supplementary Materials for The crystal structure of human dopamine -hydroxylase at 2.9 Å resolution Trine V. Vendelboe, Pernille Harris, Yuguang
advances.sciencemag.org/cgi/content/full/2/4/e1500980/dc1 Supplementary Materials for The crystal structure of human dopamine -hydroxylase at 2.9 Å resolution Trine V. Vendelboe, Pernille Harris, Yuguang
SDS-Assisted Protein Transport Through Solid-State Nanopores
 Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.
, Purified EpEX and EpEX analyzed on homogenous 12.") Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Secondary Structure North 72nd Street, Wauwatosa, WI Phone: (414) Fax: (414) dmoleculardesigns.com
 Fax: (414) dmoleculardesigns.com") Secondary Structure In the previous protein folding activity, you created a generic or hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously
Secondary Structure In the previous protein folding activity, you created a generic or hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously
Signal Transduction Cascades
 Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Review II: The Molecules of Life
 Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2007 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2007 Outline Introduction Proteins Carbohydrates Lipids
Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2007 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2007 Outline Introduction Proteins Carbohydrates Lipids
Biology 5A Fall 2010 Macromolecules Chapter 5
 Learning Outcomes: Macromolecules List and describe the four major classes of molecules Describe the formation of a glycosidic linkage and distinguish between monosaccharides, disaccharides, and polysaccharides
Learning Outcomes: Macromolecules List and describe the four major classes of molecules Describe the formation of a glycosidic linkage and distinguish between monosaccharides, disaccharides, and polysaccharides
Activities for the α-helix / β-sheet Construction Kit
 Activities for the α-helix / β-sheet Construction Kit The primary sequence of a protein, composed of amino acids, determines the organization of the sequence into the secondary structure. There are two
Activities for the α-helix / β-sheet Construction Kit The primary sequence of a protein, composed of amino acids, determines the organization of the sequence into the secondary structure. There are two
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation
 Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
Supporting Information
 Supporting Information McCullough et al. 10.1073/pnas.0801567105 A α10 α8 α9 N α7 α6 α5 C β2 β1 α4 α3 α2 α1 C B N C Fig. S1. ALIX Bro1 in complex with the C-terminal CHMP4A helix. (A) Ribbon diagram showing
Supporting Information McCullough et al. 10.1073/pnas.0801567105 A α10 α8 α9 N α7 α6 α5 C β2 β1 α4 α3 α2 α1 C B N C Fig. S1. ALIX Bro1 in complex with the C-terminal CHMP4A helix. (A) Ribbon diagram showing
Secondary Structure. by hydrogen bonds
 Secondary Structure In the previous protein folding activity, you created a hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously folds
Secondary Structure In the previous protein folding activity, you created a hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously folds
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine
 Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine") Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
CS612 - Algorithms in Bioinformatics
 Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Supplementary Materials. High affinity binding of phosphatidylinositol-4-phosphate. by Legionella pneumophila DrrA
 Supplementary Materials High affinity binding of phosphatidylinositol-4-phosphate by Legionella pneumophila DrrA Running title: Molecular basis of PtdIns(4)P-binding by DrrA Stefan Schoebel, Wulf Blankenfeldt,
Supplementary Materials High affinity binding of phosphatidylinositol-4-phosphate by Legionella pneumophila DrrA Running title: Molecular basis of PtdIns(4)P-binding by DrrA Stefan Schoebel, Wulf Blankenfeldt,
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
Chemistry 135, First Exam. September 23, Chem 135, Exam 1 SID:
 Chemistry 135, First Exam September 23, 2015 This exam will be worth 15% of your overall grade. Please read all instructions/questions carefully and provide answers in the space provided. There should
Chemistry 135, First Exam September 23, 2015 This exam will be worth 15% of your overall grade. Please read all instructions/questions carefully and provide answers in the space provided. There should
Ras and Cell Signaling Exercise
 Ras and Cell Signaling Exercise Learning Objectives In this exercise, you will use, a protein 3D- viewer, to explore: the structure of the Ras protein the active and inactive state of Ras and the amino
Ras and Cell Signaling Exercise Learning Objectives In this exercise, you will use, a protein 3D- viewer, to explore: the structure of the Ras protein the active and inactive state of Ras and the amino
Chapter 3. Structure of Enzymes. Enzyme Engineering
 Chapter 3. Structure of Enzymes Enzyme Engineering 3.1 Introduction With purified protein, Determining M r of the protein Determining composition of amino acids and the primary structure Determining the
Chapter 3. Structure of Enzymes Enzyme Engineering 3.1 Introduction With purified protein, Determining M r of the protein Determining composition of amino acids and the primary structure Determining the
Previous Class. Today. Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism. Protein Kinase Catalytic Properties
 Previous Class Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism Today Protein Kinase Catalytic Properties Protein Phosphorylation Phosphorylation: key protein modification
Previous Class Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism Today Protein Kinase Catalytic Properties Protein Phosphorylation Phosphorylation: key protein modification
Short polymer. Dehydration removes a water molecule, forming a new bond. Longer polymer (a) Dehydration reaction in the synthesis of a polymer
 Dehydration reaction in the synthesis of a polymer") HO 1 2 3 H HO H Short polymer Dehydration removes a water molecule, forming a new bond Unlinked monomer H 2 O HO 1 2 3 4 H Longer polymer (a) Dehydration reaction in the synthesis of a polymer HO 1 2 3
HO 1 2 3 H HO H Short polymer Dehydration removes a water molecule, forming a new bond Unlinked monomer H 2 O HO 1 2 3 4 H Longer polymer (a) Dehydration reaction in the synthesis of a polymer HO 1 2 3
P450 CYCLE. All P450s follow the same catalytic cycle of;
 P450 CYCLE All P450s follow the same catalytic cycle of; 1. Initial substrate binding 2. First electron reduction 3. Oxygen binding 4. Second electron transfer 5 and 6. Proton transfer/dioxygen cleavage
P450 CYCLE All P450s follow the same catalytic cycle of; 1. Initial substrate binding 2. First electron reduction 3. Oxygen binding 4. Second electron transfer 5 and 6. Proton transfer/dioxygen cleavage
Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL
 Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL For Questions 1-10 choose ONE INCORRECT answer. 1. Which ONE of the following statements concerning the
Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL For Questions 1-10 choose ONE INCORRECT answer. 1. Which ONE of the following statements concerning the
Structural biology of viruses
 Structural biology of viruses Biophysical Chemistry 1, Fall 2010 Coat proteins DNA/RNA packaging Reading assignment: Chap. 15 Virus particles self-assemble from coat monomers Virus Structure and Function
Structural biology of viruses Biophysical Chemistry 1, Fall 2010 Coat proteins DNA/RNA packaging Reading assignment: Chap. 15 Virus particles self-assemble from coat monomers Virus Structure and Function
Bioinformatics for molecular biology
 Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
MBB 694:407, 115:511. Please use BLOCK CAPITAL letters like this --- A, B, C, D, E. Not lowercase!
 MBB 694:407, 115:511 First Test Severinov/Deis Tue. Sep. 30, 2003 Name Index number (not SSN) Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions
MBB 694:407, 115:511 First Test Severinov/Deis Tue. Sep. 30, 2003 Name Index number (not SSN) Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions
Lecture 15. Membrane Proteins I
 Lecture 15 Membrane Proteins I Introduction What are membrane proteins and where do they exist? Proteins consist of three main classes which are classified as globular, fibrous and membrane proteins. A
Lecture 15 Membrane Proteins I Introduction What are membrane proteins and where do they exist? Proteins consist of three main classes which are classified as globular, fibrous and membrane proteins. A
Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site.
 Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Supplementary Table 1. Data collection and refinement statistics (molecular replacement).
.") Supplementary Table 1. Data collection and refinement statistics (molecular replacement). Data set statistics HLA A*0201- ALWGPDPAAA PPI TCR PPI TCR/A2- ALWGPDPAAA PPI TCR/A2- ALWGPDPAAA Space Group P2
Supplementary Table 1. Data collection and refinement statistics (molecular replacement). Data set statistics HLA A*0201- ALWGPDPAAA PPI TCR PPI TCR/A2- ALWGPDPAAA PPI TCR/A2- ALWGPDPAAA Space Group P2
Crystal Structure of the Subtilisin Carlsberg: OMTKY3 Complex
 John Clizer & Greg Ralph Crystal Structure of the Subtilisin Carlsberg: OMTKY3 Complex The turkey ovomucoid third domain (OMTKY3) is considered to be one of the most studied protein inhibitors. 1 Ovomucin
John Clizer & Greg Ralph Crystal Structure of the Subtilisin Carlsberg: OMTKY3 Complex The turkey ovomucoid third domain (OMTKY3) is considered to be one of the most studied protein inhibitors. 1 Ovomucin
The Chemical Building Blocks of Life. Chapter 3
 The Chemical Building Blocks of Life Chapter 3 Biological Molecules Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent
The Chemical Building Blocks of Life Chapter 3 Biological Molecules Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent
This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is worth 2 points.
 MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
Biological Molecules
 The Chemical Building Blocks of Life Chapter 3 Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent bonds. Carbon may
The Chemical Building Blocks of Life Chapter 3 Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent bonds. Carbon may
Proteins consist of joined amino acids They are joined by a Also called an Amide Bond
 Lecture Two: Peptide Bond & Protein Structure [Chapter 2 Berg, Tymoczko & Stryer] (Figures in Red are for the 7th Edition) (Figures in Blue are for the 8th Edition) Proteins consist of joined amino acids
Lecture Two: Peptide Bond & Protein Structure [Chapter 2 Berg, Tymoczko & Stryer] (Figures in Red are for the 7th Edition) (Figures in Blue are for the 8th Edition) Proteins consist of joined amino acids
Biochemistry - I. Prof. S. Dasgupta Department of Chemistry Indian Institute of Technology, Kharagpur Lecture 1 Amino Acids I
 Biochemistry - I Prof. S. Dasgupta Department of Chemistry Indian Institute of Technology, Kharagpur Lecture 1 Amino Acids I Hello, welcome to the course Biochemistry 1 conducted by me Dr. S Dasgupta,
Biochemistry - I Prof. S. Dasgupta Department of Chemistry Indian Institute of Technology, Kharagpur Lecture 1 Amino Acids I Hello, welcome to the course Biochemistry 1 conducted by me Dr. S Dasgupta,
Lecture 10 More about proteins
 Lecture 10 More about proteins Today we're going to extend our discussion of protein structure. This may seem far-removed from gene cloning, but it is the path to understanding the genes that we are cloning.
Lecture 10 More about proteins Today we're going to extend our discussion of protein structure. This may seem far-removed from gene cloning, but it is the path to understanding the genes that we are cloning.
Judy Wieber. Department of Computational Biology. May 27, 2008
 Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 27, 2008 Outline Introduction Proteins Carbohydrates
Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 27, 2008 Outline Introduction Proteins Carbohydrates
Fatty acids and phospholipids
 PYS 4xx Intro 2 1 PYS 4xx Intro 2 - Molecular building blocks We now describe in more detail the nomenclature and composition of several classes of compounds of relevance to the cell, including: membrane
PYS 4xx Intro 2 1 PYS 4xx Intro 2 - Molecular building blocks We now describe in more detail the nomenclature and composition of several classes of compounds of relevance to the cell, including: membrane
3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B
 3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B 3.2.1 Characterization of the Ligand Binding Site at GPR109A Receptor Ligands of GPR109A Receptor are Carboxylic Acids Nicotinic acid (pyridine-3-carboxylic
3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B 3.2.1 Characterization of the Ligand Binding Site at GPR109A Receptor Ligands of GPR109A Receptor are Carboxylic Acids Nicotinic acid (pyridine-3-carboxylic
Lecture Series 2 Macromolecules: Their Structure and Function
 Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
Introduction to Protein Structure Collection
 Introduction to Protein Structure Collection Teaching Points This collection is designed to introduce students to the concepts of protein structure and biochemistry. Different activities guide students
Introduction to Protein Structure Collection Teaching Points This collection is designed to introduce students to the concepts of protein structure and biochemistry. Different activities guide students
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
!"#$%&' (#%) /&'(2+"( /&3&4,, ! " #$% - &'()!% *-sheet -(!-Helix - &'(&') +,(-. - &'()&+) /&%.(0&+(! - &'(1&2%( Basic amino acids
 /&'(2+( /&3&4,, ! #$% - &'()!% *-sheet -(!-Helix - &'(&') +,(-. - &'()&+) /&%.(0&+(! - &'(1&2%( Basic amino acids") Basic amino acids pk ~ 10.5 pk ~ 12.5 pk ~ 6.0 Polar 25!"#$%&' (#%)! " #$% - &'()!% *-sheet -(!-Helix - &'(&') +,(-. - &'()&+) /&%.(0&+(! - &'(1&2%( /&'(2+"( /&3&4,, :++55 ('&.! 6($.(" 40 > 3&4,, ('&.!
Basic amino acids pk ~ 10.5 pk ~ 12.5 pk ~ 6.0 Polar 25!"#$%&' (#%)! " #$% - &'()!% *-sheet -(!-Helix - &'(&') +,(-. - &'()&+) /&%.(0&+(! - &'(1&2%( /&'(2+"( /&3&4,, :++55 ('&.! 6($.(" 40 > 3&4,, ('&.!
Lecture Series 2 Macromolecules: Their Structure and Function
 Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
2.2 Cell Construction
 2.2 Cell Construction Elemental composition of typical bacterial cell C 50%, O 20%, N 14%, H 8%, P 3%, S 1%, and others (K +, Na +, Ca 2+, Mg 2+, Cl -, vitamin) Molecular building blocks Lipids Carbohydrates
2.2 Cell Construction Elemental composition of typical bacterial cell C 50%, O 20%, N 14%, H 8%, P 3%, S 1%, and others (K +, Na +, Ca 2+, Mg 2+, Cl -, vitamin) Molecular building blocks Lipids Carbohydrates
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site.
 Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
PAPER No. : 16, Bioorganic and biophysical chemistry MODULE No. : 22, Mechanism of enzyme catalyst reaction (I) Chymotrypsin
 Chymotrypsin") Subject Paper No and Title 16 Bio-organic and Biophysical Module No and Title 22 Mechanism of Enzyme Catalyzed reactions I Module Tag CHE_P16_M22 Chymotrypsin TABLE OF CONTENTS 1. Learning outcomes 2.
Subject Paper No and Title 16 Bio-organic and Biophysical Module No and Title 22 Mechanism of Enzyme Catalyzed reactions I Module Tag CHE_P16_M22 Chymotrypsin TABLE OF CONTENTS 1. Learning outcomes 2.
Chapter 7. Heme proteins Cooperativity Bohr effect
 Chapter 7 Heme proteins Cooperativity Bohr effect Hemoglobin is a red blood cell protein that transports oxygen from the lungs to the tissues. Hemoglobin is an allosteric protein that displays cooperativity
Chapter 7 Heme proteins Cooperativity Bohr effect Hemoglobin is a red blood cell protein that transports oxygen from the lungs to the tissues. Hemoglobin is an allosteric protein that displays cooperativity
Structure and Activation of the Visual Pigment Rhodopsin
 Seminar 02.12.2011 Structure and Activation of the Visual Pigment Rhodopsin by Steven O.Smith, 2010 Constantin Schneider FB Physik, FU Berlin GPCR: Overview Rhodopsin is a G protein-coupled receptor (GPCR)
Seminar 02.12.2011 Structure and Activation of the Visual Pigment Rhodopsin by Steven O.Smith, 2010 Constantin Schneider FB Physik, FU Berlin GPCR: Overview Rhodopsin is a G protein-coupled receptor (GPCR)
Chapter 3. Protein Structure and Function
 Chapter 3 Protein Structure and Function Broad functional classes So Proteins have structure and function... Fine! -Why do we care to know more???? Understanding functional architechture gives us POWER
Chapter 3 Protein Structure and Function Broad functional classes So Proteins have structure and function... Fine! -Why do we care to know more???? Understanding functional architechture gives us POWER
Chapter 5: Structure and Function of Macromolecules AP Biology 2011
 Chapter 5: Structure and Function of Macromolecules AP Biology 2011 1 Macromolecules Fig. 5.1 Carbohydrates Lipids Proteins Nucleic Acids Polymer - large molecule consisting of many similar building blocks
Chapter 5: Structure and Function of Macromolecules AP Biology 2011 1 Macromolecules Fig. 5.1 Carbohydrates Lipids Proteins Nucleic Acids Polymer - large molecule consisting of many similar building blocks
A. Lipids: Water-Insoluble Molecules
 Biological Substances found in Living Tissues Lecture Series 3 Macromolecules: Their Structure and Function A. Lipids: Water-Insoluble Lipids can form large biological molecules, but these aggregations
Biological Substances found in Living Tissues Lecture Series 3 Macromolecules: Their Structure and Function A. Lipids: Water-Insoluble Lipids can form large biological molecules, but these aggregations
Levels of Protein Structure:
 Levels of Protein Structure: PRIMARY STRUCTURE (1 ) - Defined, non-random sequence of amino acids along the peptide backbone o Described in two ways: Amino acid composition Amino acid sequence M-L-D-G-C-G
Levels of Protein Structure: PRIMARY STRUCTURE (1 ) - Defined, non-random sequence of amino acids along the peptide backbone o Described in two ways: Amino acid composition Amino acid sequence M-L-D-G-C-G
Chymotrypsin Lecture. Aims: to understand (1) the catalytic strategies used by enzymes and (2) the mechanism of chymotrypsin
 the catalytic strategies used by enzymes and (2) the mechanism of chymotrypsin") Chymotrypsin Lecture Aims: to understand (1) the catalytic strategies used by enzymes and (2) the mechanism of chymotrypsin What s so great about enzymes? They accomplish large rate accelerations (10 10-10
Chymotrypsin Lecture Aims: to understand (1) the catalytic strategies used by enzymes and (2) the mechanism of chymotrypsin What s so great about enzymes? They accomplish large rate accelerations (10 10-10
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
Proteins. Amino acids, structure and function. The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka
 Proteins Amino acids, structure and function The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka O O HO N N HN OH Ser65-Tyr66-Gly67 The Nobel prize in chemistry 2008 Osamu Shimomura,
Proteins Amino acids, structure and function The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka O O HO N N HN OH Ser65-Tyr66-Gly67 The Nobel prize in chemistry 2008 Osamu Shimomura,
Biological Chemistry. Is biochemistry fun? - Find it out!
 Biological Chemistry Is biochemistry fun? - Find it out! 1. Key concepts Outline 2. Condensation and Hydrolysis Reactions 3. Carbohydrates 4. Lipids 5. Proteins 6. Nucleic Acids Key Concepts: 1. Organic
Biological Chemistry Is biochemistry fun? - Find it out! 1. Key concepts Outline 2. Condensation and Hydrolysis Reactions 3. Carbohydrates 4. Lipids 5. Proteins 6. Nucleic Acids Key Concepts: 1. Organic
Molecular building blocks
 2.22 Cell Construction Elemental l composition of ftypical lbacterial cell C 50%, O 20%, N 14%, H 8%, P 3%, S 1%, and others (K +, Na +, Ca 2+, Mg 2+, Cl -, vitamin) Molecular building blocks Lipids Carbohydrates
2.22 Cell Construction Elemental l composition of ftypical lbacterial cell C 50%, O 20%, N 14%, H 8%, P 3%, S 1%, and others (K +, Na +, Ca 2+, Mg 2+, Cl -, vitamin) Molecular building blocks Lipids Carbohydrates
Catalysis & specificity: Proteins at work
 Catalysis & specificity: Proteins at work Introduction Having spent some time looking at the elements of structure of proteins and DNA, as well as their ability to form intermolecular interactions, it
Catalysis & specificity: Proteins at work Introduction Having spent some time looking at the elements of structure of proteins and DNA, as well as their ability to form intermolecular interactions, it
PHAR3316 Pharmacy biochemistry Exam #2 Fall 2010 KEY
 1. How many protons is(are) lost when the amino acid Asparagine is titrated from its fully protonated state to a fully deprotonated state? A. 0 B. 1 * C. 2 D. 3 E. none Correct Answer: C (this question
1. How many protons is(are) lost when the amino acid Asparagine is titrated from its fully protonated state to a fully deprotonated state? A. 0 B. 1 * C. 2 D. 3 E. none Correct Answer: C (this question
Table S1. X-ray data collection and refinement statistics
 Table S1. X-ray data collection and refinement statistics Data collection H7.167 Fab-Sh2/H7 complex Beamline SSRL 12-2 Wavelength (Å) 0.97950 Space group I2 1 3 Unit cell parameters (Å, º) a = b = c=207.3,
Table S1. X-ray data collection and refinement statistics Data collection H7.167 Fab-Sh2/H7 complex Beamline SSRL 12-2 Wavelength (Å) 0.97950 Space group I2 1 3 Unit cell parameters (Å, º) a = b = c=207.3,
Name: Multiple choice questions. Pick the BEST answer (2 pts ea)
") Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
The Structure and Function of Macromolecules
 The Structure and Function of Macromolecules Macromolecules are polymers Polymer long molecule consisting of many similar building blocks. Monomer the small building block molecules. Carbohydrates, proteins
The Structure and Function of Macromolecules Macromolecules are polymers Polymer long molecule consisting of many similar building blocks. Monomer the small building block molecules. Carbohydrates, proteins
Supplementary Information Janssen et al.
 Supplementary Information Janssen et al. Insights into complement convertase formation based on the structure of the factor B CVF complex Bert J.C. Janssen 1, Lucio Gomes 1, Roman I. Koning 2, Dmitri I.
Supplementary Information Janssen et al. Insights into complement convertase formation based on the structure of the factor B CVF complex Bert J.C. Janssen 1, Lucio Gomes 1, Roman I. Koning 2, Dmitri I.
Practice Problems 3. a. What is the name of the bond formed between two amino acids? Are these bonds free to rotate?
 Life Sciences 1a Practice Problems 3 1. Draw the oligopeptide for Ala-Phe-Gly-Thr-Asp. You do not need to indicate the stereochemistry of the sidechains. Denote with arrows the bonds formed between the
Life Sciences 1a Practice Problems 3 1. Draw the oligopeptide for Ala-Phe-Gly-Thr-Asp. You do not need to indicate the stereochemistry of the sidechains. Denote with arrows the bonds formed between the
What are the molecules of life?
 Molecules of Life What are the molecules of life? Organic Compounds Complex Carbohydrates Lipids Proteins Nucleic Acids Organic Compounds Carbon- hydrogen based molecules From Structure to Function Ø Carbon
Molecules of Life What are the molecules of life? Organic Compounds Complex Carbohydrates Lipids Proteins Nucleic Acids Organic Compounds Carbon- hydrogen based molecules From Structure to Function Ø Carbon
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB
 Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Biomolecules. Biomolecules. Carbohydrates. Biol 219 Lec 3 Fall Polysaccharides. Function: Glucose storage Fig. 2.2
 Biomolecules Biomolecules Monomers Polymers Carbohydrates monosaccharides polysaccharides fatty acids triglycerides Proteins amino acids polypeptides Nucleic Acids nucleotides DNA, RNA Carbohydrates Carbohydrates
Biomolecules Biomolecules Monomers Polymers Carbohydrates monosaccharides polysaccharides fatty acids triglycerides Proteins amino acids polypeptides Nucleic Acids nucleotides DNA, RNA Carbohydrates Carbohydrates
Interactions of Polyethylenimines with Zwitterionic and. Anionic Lipid Membranes
 Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
a) The statement is true for X = 400, but false for X = 300; b) The statement is true for X = 300, but false for X = 200;
 The statement is true for X = 400, but false for X = 300; b) The statement is true for X = 300, but false for X = 200;") 1. Consider the following statement. To produce one molecule of each possible kind of polypeptide chain, X amino acids in length, would require more atoms than exist in the universe. Given the size of
1. Consider the following statement. To produce one molecule of each possible kind of polypeptide chain, X amino acids in length, would require more atoms than exist in the universe. Given the size of
WHY IS THIS IMPORTANT?
 CHAPTER 2 FUNDAMENTAL CHEMISTRY FOR MICROBIOLOGY WHY IS THIS IMPORTANT? An understanding of chemistry is essential to understand cellular structure and function, which are paramount for your understanding
CHAPTER 2 FUNDAMENTAL CHEMISTRY FOR MICROBIOLOGY WHY IS THIS IMPORTANT? An understanding of chemistry is essential to understand cellular structure and function, which are paramount for your understanding
Midterm 1 Last, First
 Midterm 1 BIS 105 Prof. T. Murphy April 23, 2014 There should be 6 pages in this exam. Exam instructions (1) Please write your name on the top of every page of the exam (2) Show all work for full credit
Midterm 1 BIS 105 Prof. T. Murphy April 23, 2014 There should be 6 pages in this exam. Exam instructions (1) Please write your name on the top of every page of the exam (2) Show all work for full credit
Supplementary information: Binding of N-methylscopolamine to the extracellular domain of muscarinic acetylcholine receptors
 Supplementary information: Binding of N-methylscopolamine to the extracellular domain of muscarinic acetylcholine receptors Jan Jakubík, Alena Randáková, Pavel Zimčík, Esam E. El-Fakahany, and Vladimír
Supplementary information: Binding of N-methylscopolamine to the extracellular domain of muscarinic acetylcholine receptors Jan Jakubík, Alena Randáková, Pavel Zimčík, Esam E. El-Fakahany, and Vladimír
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
Biological Molecules. Carbohydrates, Proteins, Lipids, and Nucleic Acids
 Biological Molecules Carbohydrates, Proteins, Lipids, and Nucleic Acids Organic Molecules Always contain Carbon (C) and Hydrogen (H) Carbon is missing four electrons Capable of forming 4 covalent bonds
Biological Molecules Carbohydrates, Proteins, Lipids, and Nucleic Acids Organic Molecules Always contain Carbon (C) and Hydrogen (H) Carbon is missing four electrons Capable of forming 4 covalent bonds
Human Anatomy & Physiology C H A P T E R
 PowerPoint Lecture Slides prepared by Barbara Heard, Atlantic Cape Community College Ninth Edition Human Anatomy & Physiology C H A P T E R 2 Annie Leibovitz/Contact Press Images 2013 Pearson Education,
PowerPoint Lecture Slides prepared by Barbara Heard, Atlantic Cape Community College Ninth Edition Human Anatomy & Physiology C H A P T E R 2 Annie Leibovitz/Contact Press Images 2013 Pearson Education,
Protein regulation Protein motion
 Lecture 13 Protein regulation Protein motion Antoine van Oijen BCMP201 Spring 2008 04/02 Section IV 04/09 Hands-on methods session / PS 4 due 1 Today s lecture 1) Mechanisms of protein regulation 2) Molecular
Lecture 13 Protein regulation Protein motion Antoine van Oijen BCMP201 Spring 2008 04/02 Section IV 04/09 Hands-on methods session / PS 4 due 1 Today s lecture 1) Mechanisms of protein regulation 2) Molecular
BIOCHEMISTRY. How Are Macromolecules Formed? Dehydration Synthesis or condensation reaction Polymers formed by combining monomers and removing water.
 BIOCHEMISTRY Organic compounds Compounds that contain carbon are called organic. Inorganic compounds do not contain carbon. Carbon has 4 electrons in outer shell. Carbon can form covalent bonds with as
BIOCHEMISTRY Organic compounds Compounds that contain carbon are called organic. Inorganic compounds do not contain carbon. Carbon has 4 electrons in outer shell. Carbon can form covalent bonds with as
Lab 5: Proteins and the small molecules that love them (AKA Computer Modeling with PyMol #2)
") Lab 5: Proteins and the small molecules that love them (AKA Computer Modeling with PyMol #2) Goals: The objective of this lab is to provide you with an understanding of: 1. Catalysis 2. Small molecule
Lab 5: Proteins and the small molecules that love them (AKA Computer Modeling with PyMol #2) Goals: The objective of this lab is to provide you with an understanding of: 1. Catalysis 2. Small molecule
Lecture Series 2 Macromolecules: Their Structure and Function
 Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
Lecture Series 2 Macromolecules: Their Structure and Function Reading Assignments Read Chapter 4 (Protein structure & Function) Biological Substances found in Living Tissues The big four in terms of macromolecules
Proteins and their structure
 Proteins and their structure Proteins are the most abundant biological macromolecules, occurring in all cells and all parts of cells. Proteins also occur in great variety; thousands of different kinds,
Proteins and their structure Proteins are the most abundant biological macromolecules, occurring in all cells and all parts of cells. Proteins also occur in great variety; thousands of different kinds,
Table S1: Kinetic parameters of drug and substrate binding to wild type and HIV-1 protease variants. Data adapted from Ref. 6 in main text.
 Dynamical Network of HIV-1 Protease Mutants Reveals the Mechanism of Drug Resistance and Unhindered Activity Rajeswari Appadurai and Sanjib Senapati* BJM School of Biosciences and Department of Biotechnology,
Dynamical Network of HIV-1 Protease Mutants Reveals the Mechanism of Drug Resistance and Unhindered Activity Rajeswari Appadurai and Sanjib Senapati* BJM School of Biosciences and Department of Biotechnology,
Supplementary Information
 Supplementary Information Using the pimeloyl-coa synthetase adenylation fold to synthesise fatty acid thioesters Menglu Wang 1a ; Lucile Moynié 2a, Peter J. Harrison 1, Van Kelly 1, Andrew Piper 1, James
Supplementary Information Using the pimeloyl-coa synthetase adenylation fold to synthesise fatty acid thioesters Menglu Wang 1a ; Lucile Moynié 2a, Peter J. Harrison 1, Van Kelly 1, Andrew Piper 1, James
FCC2 5CY7 FCC1 5CY8. Actinonin 5CVQ
 Table S1. Data collection and refinement statistics Data collection Apo 5E5D MA 5CPD MAS 5CP0 Actinonin 5CVQ FCC1 5CY8 FCC2 5CY7 FCC 5CWY FCC4 5CVK FCC5 5CWX FCC6 5CVP X-ray source 17A-KEK PAL4A PAL4A
Table S1. Data collection and refinement statistics Data collection Apo 5E5D MA 5CPD MAS 5CP0 Actinonin 5CVQ FCC1 5CY8 FCC2 5CY7 FCC 5CWY FCC4 5CVK FCC5 5CWX FCC6 5CVP X-ray source 17A-KEK PAL4A PAL4A
Protein Structure and Function
 Protein Structure and Function Protein Structure Classification of Proteins Based on Components Simple proteins - Proteins containing only polypeptides Conjugated proteins - Proteins containing nonpolypeptide
Protein Structure and Function Protein Structure Classification of Proteins Based on Components Simple proteins - Proteins containing only polypeptides Conjugated proteins - Proteins containing nonpolypeptide
Biological Molecules
 Chemical Building Blocks of Life Chapter 3 Biological Molecules Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent
Chemical Building Blocks of Life Chapter 3 Biological Molecules Biological molecules consist primarily of -carbon bonded to carbon, or -carbon bonded to other molecules. Carbon can form up to 4 covalent
Student name ID # 2. (4 pts) What is the terminal electron acceptor in respiration? In photosynthesis?
 What is the terminal electron acceptor in respiration? In photosynthesis?") 1. Membrane transport. A. (4 pts) What ion couples primary and secondary active transport in animal cells? What ion serves the same function in plant cells? 2. (4 pts) What is the terminal electron acceptor
1. Membrane transport. A. (4 pts) What ion couples primary and secondary active transport in animal cells? What ion serves the same function in plant cells? 2. (4 pts) What is the terminal electron acceptor
Role of Soybean Ecto-apyrase in Nodulation
 PAG XX Engineering NUE Workshop W248 January 14, 2012 Role of Soybean Ecto-apyrase in Nodulation Kiwamu Tanaka (Gary Stacey Lab) Division of Plant Sciences University of Missouri Legume-Rhizobium symbiotic
PAG XX Engineering NUE Workshop W248 January 14, 2012 Role of Soybean Ecto-apyrase in Nodulation Kiwamu Tanaka (Gary Stacey Lab) Division of Plant Sciences University of Missouri Legume-Rhizobium symbiotic
SUPPLEMENTAL MATERIAL. UNC119 is required for G protein trafficking in sensory neurons
 1 SUPPLEMENTAL MATERIAL UNC119 is required for G protein trafficking in sensory neurons Houbin Zhang, Ryan N. Constantine, Sergey Vorobiev, Yang Chen, Jayaraman Seetharaman, Yuanpeng Janet Huang, Rong
1 SUPPLEMENTAL MATERIAL UNC119 is required for G protein trafficking in sensory neurons Houbin Zhang, Ryan N. Constantine, Sergey Vorobiev, Yang Chen, Jayaraman Seetharaman, Yuanpeng Janet Huang, Rong
Biology. Chapter 3. Molecules of Life. Concepts and Applications 9e Starr Evers Starr
 Biology Concepts and Applications 9e Starr Evers Starr Chapter 3 Molecules of Life 2015 3.1 What Are the Molecules of Life? The molecules of life contain a high proportion of carbon atoms: Complex carbohydrates
Biology Concepts and Applications 9e Starr Evers Starr Chapter 3 Molecules of Life 2015 3.1 What Are the Molecules of Life? The molecules of life contain a high proportion of carbon atoms: Complex carbohydrates
Lecture 33 Membrane Proteins
 Lecture 33 Membrane Proteins Reading for today: Chapter 4, section D Required reading for next Wednesday: Chapter 14, sections A and 14.19 to the end Kuriyan, J., and Eisenberg, D. (2007) The origin of
Lecture 33 Membrane Proteins Reading for today: Chapter 4, section D Required reading for next Wednesday: Chapter 14, sections A and 14.19 to the end Kuriyan, J., and Eisenberg, D. (2007) The origin of
Lesson 2. Biological Molecules. Introduction to Life Processes - SCI 102 1
 Lesson 2 Biological Molecules Introduction to Life Processes - SCI 102 1 Carbon in Biological Molecules Organic molecules contain carbon (C) and hydrogen (H) Example: glucose (C 6 H 12 O 6 ) Inorganic
Lesson 2 Biological Molecules Introduction to Life Processes - SCI 102 1 Carbon in Biological Molecules Organic molecules contain carbon (C) and hydrogen (H) Example: glucose (C 6 H 12 O 6 ) Inorganic
Carbon. Isomers. The Chemical Building Blocks of Life
 The Chemical Building Blocks of Life Carbon Chapter 3 Framework of biological molecules consists primarily of carbon bonded to Carbon O, N, S, P or H Can form up to 4 covalent bonds Hydrocarbons molecule
The Chemical Building Blocks of Life Carbon Chapter 3 Framework of biological molecules consists primarily of carbon bonded to Carbon O, N, S, P or H Can form up to 4 covalent bonds Hydrocarbons molecule
Supplementary Information for Structural basis for the inhibition of Polo-like kinase 1
 Supplementary Information for Structural basis for inhibition of Polo-like kinase 1 Jun Xu, Chen Shen, Tao Wang & Junmin Quan Correspondence authors: Tao Wang, taowang@pkusz.edu.cn; Junmin Quan, quanjm@szpku.edu.cn
Supplementary Information for Structural basis for inhibition of Polo-like kinase 1 Jun Xu, Chen Shen, Tao Wang & Junmin Quan Correspondence authors: Tao Wang, taowang@pkusz.edu.cn; Junmin Quan, quanjm@szpku.edu.cn
Structure and Function of Macromolecules Chapter 5 Macromolecules Macromolecules Multiple Units Synthesis of Dimers and Polymers
 Structure and Function of Macromolecules Chapter 5 Macromolecules Giant molecules weighing over 100,000 daltons Emergent properties not found in component parts Macromolecules Multiple Units meris = one
Structure and Function of Macromolecules Chapter 5 Macromolecules Giant molecules weighing over 100,000 daltons Emergent properties not found in component parts Macromolecules Multiple Units meris = one