Cryo EM structure of the ribosome SecYE complex in the
|
|
|
- William Miles
- 5 years ago
- Views:
Transcription
1 Supplementary Information Cryo EM structure of the ribosome SecYE complex in the membrane environment Jens Frauenfeld,, James Gumbart, Eli O. van der Sluis,, Soledad Funes, Marco Gartmann,, Birgitta Beatrix,, Thorsten Mielke, Otto Berninghausen,, Thomas Becker,, Klaus Schulten and Roland Beckmann,, # Gene Center, Department for Biochemistry, University of Munich, Feodor Lynen Str., Munich, Germany Munich Center For Integrated Protein Science (CIPSM), Department of Chemistry and Biochemistry, Butenandtstr., Munich, Germany Department of Physics, Beckman Institute, University of Illinois at Urbana Champaign, Urbana, IL, 0, U Departamento de Genética Molecular, Instituto de Fisiología Celular, Circuito Exterior S/N, Ciudad Universitaria, Universidad Nacional Autónoma de México, Mexico, D.F., 00, Mexico Ultrastrukturnetzwerk, Max Planck Institute for Molecular Genetics, Ihnestr., Berlin, Institut für Medizinische Physik und Biophysik, Charite Universitätsmedizin Berlin, Ziegelstrasse, 0 Berlin, Germany # Corresponding author Roland Beckmann: beckmann@lmb.uni-muenchen.de Tel: Fax: + 0
2 Supplementary Methods MDFF MDFF is a method to flexibly fit atomic models into cryo-em density maps while simultaneously preserving the stereochemical accuracy of model,. In MDFF, the atomic model is simulated using molecular dynamics in the presence of the cryo-em density map, represented through an additional potential in the simulation. From this potential, forces proportional to the gradient of the cryo-em density are derived that then drive atoms into high-density regions of the map. In addition, restraints are applied to maintain the secondary structure of protein and RNA molecules, which otherwise would distort or break under the forces required for fitting. Fitting of the 0S proceeded in stages using an approach employed previously,,. A total simulation time of. ns was used to fit the ribosome. Simulations All MD simulations, including MDFF, were carried out using NAMD.b and the CHARMM force field with CMAP corrections -. Simulation protocols, including multiple time-stepping and particle mesh Ewald, are identical to those used in Gumbart et al.. After completion of modeling and MDFF, the resulting ribosome- Nanodisc model was used for further equilibrium simulations. Water and ions were added in an iterative procedure using VMD. To reduce simulation complexity and to focus on the interactions between the ribosome and SecYE and Nanodisc, the ribosome and nascent chain were truncated just downstream of the L/L constriction point. Any ribosomal backbone atoms within Å of the truncation point
3 were constrained. At the point of closest approach, SecYE was at least Å away from the truncation point. While previous simulations of the ribosome-secy complex required. million atoms, simulation of the truncated ribosome-nanodisc complex required only 00,000 atoms. Equilibration of the system occurred in stages. First, only the lipid tails were allowed to move, permitting them to melt, for 0. ns. Next, water and sidechains were released for an additional. ns. For the next. ns, only the encircling Apo A- proteins of the Nanodisc were constrained; the secondary structure of all proteins and RNA was also enforced during this time, and for a further ns. Finally, after ns of total simulation time, all restraints were released. At all times, a constant temperature of 0 K and a constant pressure of atm were maintained. Figures Densities for the large and small ribosomal subunit, the P-site trna, the nascent FtsQ-chain, the E. coli SecYE and the Nanodisc-Lipid-Bilayer were isolated using the color zone function of Chimera 0. A lower contour level of the ligand densities for surface representation was applied for some figures. This indicates that ligand densities are partially flexible or still under-represented because of incomplete removal of ligand-free ribosomal particles from the final particle subset. Supplemental Fig. a shows the entire electron density filtered at different resolutions using only one contour level for all parts.
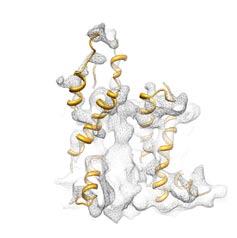
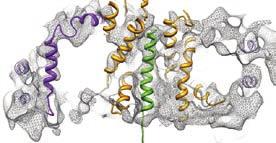
4 Supplementary Figure Legends Supplementary Fig. : Raw data, effect of resolution on TM helices (a) The complete 0S-RNC-Nd-SecYEG density is shown, filtered at different frequencies ranging from -0 Å, as indicated. (b) Close-up of the 0S-Nd-SecYEG density, side view cut perpendicular to the plane of the membrane to show the lateral gate of SecY, filtered from -0 Å, as indicated. Two layers of density are visible (upper membrane interface, UMI and lower membrane interface, LMI), separated by a region of lower density (hydrophobic core, HC), containing rod-like features. (c) Close-ups of the Nanodisc-density, showing different views with the fitted models of SecY (orange), (purple), the signal anchor (green) and the electron density represented in grey mesh. Supplementary Figure : Canonical binding of PCCs to ribosomes (a) Close-up on the interaction of cytosolic loop L/ of the mammalian Sec complex (red, PDB: WWB ) with the eukaryotic 0S ribosome (b) Close-up on the interaction of cytosolic loop L/ of a mixed model of the archeal SecYEβ complex with L/ and L/ replaced by a model of the corresponding E.coli SecY loops (purple, PDB: BO0 ) (c) Close-up on the interaction of cytosolic loop L/ of the E.coli SecYEG complex (orange) with the prokaryotic 0S RNC and an inserted signal anchor
5 (d) Close-up of the map filtered at - Å showing the interaction of cytosolic loop L/ of the E.coli SecYEG complex with the fitted models of the E.coli SecYEG complex (orange) with the prokaryotic 0S RNC and an inserted signal anchor (e) as in (d), but rotated around 0 Supplementary Figure : Fitting of SecY structures into the cryo-em density and comparison with the D crystal structure of the E. coli SecYEG complex (a) Close-up of the SecY density, side view cut perpendicular to the plane of the membrane to show SecY TM helices,, with fitted X-ray structures of SecY M. janaschii (blue, left), T. maritima (yellow, middle) and our E. coli model (orange, right). (b) Close-up of the SecY density, side view cut perpendicular to the plane of the membrane to show the lateral gate with SecY TM helices,,,, with fitted X- ray structures of SecY M. janaschii (blue, left), T. maritima (yellow, middle) and our current E. coli model (orange, right). (c) Cytosolic view of the electron density projection map of the D crystal structure of the E. coli SecYEG complex with the fitted X-ray structure of the SecYEβ from M. jannaschii. SecY TM helices in red and labelled in green, C-terminal helix in grey (figure adapted from ref#). The position of the two additional N-terminal helices of E. coli is labelled in purple, Secβ in grey.
6 (d) Cytosolic view of the electron density map of the cryo-em structure of the open E. coli SecYEG complex. SecY TM helices in orange, TM helices in purple, signal anchor () in green. Note the slightly outward shifted position of the N-terminal density compared to its position in the D-crystal map. The position of the SecG TM helices (red) is according to an alignment of the X-ray structure of the SecYEG complex from T. maritima on our E. coli model. (e) as in (d), with the aligned X-ray structure of the SecYEG complex from T. maritima (red) on our E. coli model. Supplementary Figure : RMSD values of SecYE and of the signal anchor relative to SecYE. The root-mean-square deviation (RMSD) over time is presented for (a) the backbone of SecYE and (b) that of the signal anchor. In both cases, RMSD was calculated after first performing a least-squares fit of SecYE over all frames of the simulation trajectory. Data for the initial. ns of the simulation in which the proteins were restrained are not shown. Supplementary Figure : Formation of H-bonds during simulation. Hydrogen bonds formed between different components of the simulation over time are shown. (a,b) H-bonds between the ribosome and (a) SecY and (b). (c,d) H- bonds between SecY and (c) the nascent chain and (d) the signal anchor. The solid black line denotes a running average of the full data in light grey. Only data from the last 0 ns of the simulation, i.e., the completely unrestrained portion, are shown. H-
7 bonds were counted when the distance between the hydrogen donor and the acceptor was within. Å and the angle formed by the donor, hydrogen, and acceptor was greater than. Supplementary Figure : Surface representation of the all-atom model of a 0S- RNC-Nd-SecYEG complex (a) Surface representation of the all-atom model of a 0S-RNC-Nd-SecYE complex that was used for the free MD simulation, coloured as in Fig.. Phospholipid headgroups are red (oxygen) and blue (nitrogen). Right: close-up of the isolated SecYE complex in the same position within the Nanodisc of the left panel. (b) as in (a), but rotated 0 around the y-axis. (c) as in (b), but rotated 0 around the y-axis. Supplementary Figure : Analysis of ribosomal proteins L, L, L Comparison of X-ray structures and cryo-em densities of an inactive ribosome (PDB: IV) vs. MDFF-models and cryo-em densities of an active ribosome. (a) Conformation of L. Left, isolated density of L in an inactive ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-left, isolated density of L in active ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-right, isolated density of L in an active ribosome with a MDFF-model of L of an active ribosome (light blue). Right, overlay of the X-ray structure of the inactive L with the MDFF-model of L.
8 (b) Conformation of L, side view as in Fig.b. Left, isolated density of L in an inactive ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-left, isolated density of L in active ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-right, isolated density of L in an active ribosome with a MDFF-model of L of an active ribosome (light blue). Right, overlay of the X-ray structure of the inactive L with the MDFF-model of L. (c) Conformation of L, side view as in Fig.c. Left, isolated density of L in an inactive ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-left, isolated density of L in active ribosome with the fitted X-ray structure of L of an inactive ribosome (dark grey). Middle-right, isolated density of L in an active ribosome with a MDFF-model of L of an active ribosome (light blue). Right, overlay of the X-ray structure of the inactive L with the MDFF-model of L. Supplementary Figure : Comparison of L/ conformation within the ribosomal tunnel Close-up of a section through the ribosomal exit tunnel with fitted models of L/ of SecY. (a) A model for an inactive, monomeric SecY bound to a non-translating ribosome (purple, PDB: BO0) was fitted according to the position of ribosomal RNA and superimposed to our model of the translating ribosome with the nascent chain (green). In that position, L/ of the inactive SecY would prevent the exit of the nascent chain.
9 Upper panel: side view, lower panel: view from the inside of the ribosomal tunnel towards the ribosomal exit (b) as in (a), but with a model for an inactive, monomeric SecY with an alternate L/ conformation binding to a non-translating ribosome (ruby, PDB: BO). Also in this position, the exit of the nascent chain is hindered by L/ of the inactive SecY. (c) view as in (a). The model for the translating ribosome bound to an open SecY (orange) within a membrane environment. L/ reaches up along the wall of the ribosomal tunnel and contacts both, the nascent chain and L. The position of L/ within the ribosomal exit tunnel of the hybrid complex allows the exit of the nascent chain (d) view as in (a), but with a model for the mammalian Sec complex bound to a translating wheat germ ribosome (red, PDB: WWB), fitted according to the position of ribosomal RNA and superimposed to our model of the translating ribosome with the nascent chain (green). The position of L/ within the ribosomal exit tunnel of the hybrid complex allows the exit of the nascent chain. (e) Close-up of the density showing the interaction of L/ with the nascent chain in the ribosomal exit tunnel with the fitted models for SecY, 0S subunit and the nascent chain Supplementary Figure : Conformational changes and opening of SecYE. (a) View of the lateral gate of the PCC. Comparison of the membrane-embedded, open ribosome-bound SecYE (orange, purple) with SecYE from the T. maritima SecA-SecYEG complex (grey). Loop movements are indicated with round arrows,
10 helix movements are indicated with small black arrows. in green, the NC has been omitted for better clarity. (b) as in (a), but viewed from the cytoplasmic side with the NC in green. (c) Comparison of SecY structures in different conformations, viewed from the cytoplasmic side. Left, structure of the closed, detergent-solubilised SecY from M. janaschii (PDB: RHZ). Middle left, structure of the pre-open, detergent-solubilised SecY from T. maritima. Middle right, model of the open, membrane-embedded SecY from E. coli. Right, model of the open, membrane-embedded SecY from E.coli with a helix within the lateral gate (d) as in c, but view of the lateral gate Supplementary Figure 0: Horizontal sections of Nd-SecYEG and corresponding models Three consecutive horizontal sections, sliced within the plane of the membrane in the hydrophobic core of the lipid bilayer, as indicated (, upper;, middle;, lower). (a) Sections through the experimental map at - Å with the fitted model for Nd- SecYEG and the signal anchor. Charged lipid headgroups are visible within the slices. The likely position of the SecG TM helices (marked) in the density is according to the X-ray structure of the SecYEG complex from T. maritima. (b) Sections through a density based on the molecular model for SecYE/ within the Nanodisc at - Å. Additional density from charged lipid headgroups are visible, similar to the appearance of the experimental map. Since the model does not include
11 SecG, the density does not display rod-like features in the position where SecG is expected, in contrast to the experimental map. (c) Sections through a density based on the molecular model for SecYE/ without the Nanodisc (no lipids) at - Å. No additional density from charged lipid headgroups is visible. (d) Sections through a density based on the X-ray structure of the SecYEG complex from T. maritima at - Å. Supplementary Figure : Plot of ribosome-lipid contact area during simulation. The surface area of interaction (measured in Å ) vs. time between the membrane and (a) the entire ribosome, (b) L, and (c) L is shown. The blue lines at. ns and ns denote the different stages of equilibration, noted in part (a) and in the MD Simulations section of the Methods. Supplementary Figure : Comparison of the position of the signal anchor with respect to the ribosome in (i) a SRP bound state and (ii) the PCC-inserted state (a) Close-up of the ribosomal exit site. A molecular model of SRP bound to a translating ribosome with a signal anchor (PDB: j). Note the orientation of the signal anchor with respect to ribosomal rrna H. (b) Same view as in (a), but now with the molecular model of the PCC-inserted signal anchor. Note the orientation of the signal anchor with respect to H. (c) As in (a), rotated 0 (d) As in (b), rotated 0 0
12 Supplementary Table : Cross-correlation coefficients Cross-correlation coefficients for different structures. An isolated map of the transmembrane region of SecYE and the signal anchor/nascent chain filtered at - Å was used for all calculations. Simulated maps were generated at a resolution of. Å. Initial and final refer to pre- and post-mdff states, respectively. For the rotated structure, SecY (or SecYE) and nascent chain were rotated about SecY s central axis 0. Supplementary Table : Ribosome-SecY interactions. Interactions between residues in the ribosome and in SecY. Specific residue-residue interactions were calculated over 0. ns of equilibration in which the backbone of all protein and RNA was restrained; thus, the interactions listed represent the fitted structure only. The criteria for H-bonds is given in Supplementary Figure 0; hydrophobic/hydrophilic interactions were counted when hydrophobic/hydrophilic heavy (non-hydrogen) atoms came within.0 Å of each other, respectively. Interactions were only counted when they appeared in at least 0% of frames, i.e., 0 out of the 00 frames taken every ps in the 0. ns simulation. If they appeared in between 0% and 0% of frames, they are denoted as weak. Supplementary Table : Ribosome interactions. Interactions between the ribosome and. See the caption of Supplementary Table for a description. Supplementary Table : NC-ribosome-SecY interactions. Interactions between the nascent chain and SecY and the ribosome. See the caption of Supplementary Table for a description.
13 Supplementary Table : NC-SecY interactions. Interactions between the nascent chain and SecY. See the caption of Supplementary Table for a description. Supplementary Table : -SecY interactions. Interactions between the SecY and the signal anchor. See the caption of Supplementary Table for a description. Supplementary Table : Interactions between H and lipids Figure: Interactions between H of the ribosome and lipids. (a) Ribosome-SecYnanodisc system. H is indicated in red. (b) Direct hydrogen bonding between a backbone phosphate of H and a PE lipid molecule. (c) Mg + -bridged interaction between the phosphates of H and a PE lipid molecule. (d) Mg + -bridged interaction between a phosphate of H and the head group of a PG lipid molecule. Table: Interactions between H of the ribosomal S RNA and lipids during free equilibration of ribosome-secye-nanodisc system (0-ns simulation). Interactions are classified into three types: hydrogen bonds, hydrophilic and ion-bridging. An ionbridging interaction is counted when a Mg + ion is less than Å from negatively charged atoms of both an RNA base and a lipid headgroup. The interaction is considered stable when it persists for at least 00 ps. Interactions primarily involved the RNA backbone on one side and the lipid phosphate or the NH + group of PE on the other side.
14 Supplementary References 0 Trabuco, L.G., Villa, E., Mitra, K., Frank, J., & Schulten, K., Flexible fitting of atomic structures into electron microscopy maps using molecular dynamics. Structure (), - (00). Trabuco, L.G., Villa, E., Schreiner, E., Harrison, C.B., & Schulten, K., Molecular dynamics flexible fitting: a practical guide to combine cryoelectron microscopy and X-ray crystallography. Methods (), -0 (00). Gumbart, J., Trabuco, L.G., Schreiner, E., Villa, E., & Schulten, K., Regulation of the protein-conducting channel by a bound ribosome. Structure (), - (00). Villa, E. et al., Ribosome-induced changes in elongation factor Tu conformation control GTP hydrolysis. Proc Natl Acad Sci U S A 0 (), 0-0 (00). Phillips, J.C. et al., Scalable molecular dynamics with NAMD. J Comput Chem (), -0 (00). MacKerell, A.D. et al., All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins Ä. The Journal of Physical Chemistry B 0 (), - (). Foloppe, N., Mackerell, A., & Jr, All-atom empirical force field for nucleic acids: I. Parameter optimization based on small molecule and condensed phase macromolecular target data. Journal of Computational Chemistry (), - 0 (000). Mackerell, A.D., Jr., Feig, M., & Brooks, C.L., rd, Extending the treatment of backbone energetics in protein force fields: limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J Comput Chem (), 00- (00). Humphrey, W., Dalke, A., & Schulten, K., VMD: visual molecular dynamics. J Mol Graph (), -, - (). Pettersen, E.F. et al., UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem (), 0-. (00). Becker, T. et al., Structure of Monomeric Yeast and Mammalian Sec Complexes Interacting with the Translating Ribosome. Science (00). Menetret, J.-F. et al., Ribosome Binding of a Single Copy of the SecY Complex: Implications for Protein Translocation. Molecular Cell (), 0-0 (00). Collinson, I., The structure of the bacterial protein translocation complex SecYEG. Biochem Soc Trans (Pt ), -0 (00). Halic, M. et al., Following the signal sequence from ribosomal tunnel exit to signal recognition particle. Nature (), 0- (00).








15 c Supplementary Figure a - Å - Å - Å -0 Å b TM TM 0S UMI 0S UMI TM HC TM HC LMI LMI - Å - Å TM TM 0S UMI 0S UMI TM HC TM HC LMI LMI - Å -0 Å c TM TM 0 0





16 Supplementary Figure a b c L La L L L L L/ L/ L/ d e L/ C-term. C-term. L/ L LH



17 Supplementary Figure a b RHZ Methanocaldococcus jannaschii DIN Thermotoga maritima msb Escherichia coli c d e,, 0, 0 SecG SecG
18 Supplementary Figure

19 Supplementary Figure





20 Supplementary Figure a NC 0S L/ SecY C-term. 0S L L H 0- Å 0 b trna NC 0S 0S L/ L/ SecY C-term. H L SecY C-term. 0 c 0S NC L/ SecY C-term. 0S 0- Å L





21 Supplementary Figure inactive map inactive pdb active map inactive pdb active map active pdb active pdb inactive pdb a L b L c L








22 Supplementary Figure a 0S Ribosome SecYEG b 0S Ribosome SecYEG c 0S RNC SecYEG NC L NC L NC L L/ L/ L/ L/ L/ L/ H H H L L L L/ L/ L/ NC NC NC H H H d 0S RNC Sec e NC L NC L L/ L/ L/ L L/ H NC L/ La L L L/ NC NC L/ H









23 Supplementary Figure a b L/ L/ b 0 b c a a a d








24 d c b a 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG, 0 SecG 0 SecG 0 SecG charged lipid headgroups charged lipid headgroups Supplementary Figure 0

25 Supplementary Figure



26 Supplementary Figure a b L L H L L H Cytoplasm.S RNA N SecY G Periplasm 0 0 c d H L.S RNA H L/ L Cytoplasm G Periplasm
27 Supplementary Table : Cross-correlation coefficients Structure Cross-correlation coeff. SecY/ (initial) 0. SecY/ (final) 0. SecY/ (rotated 0 ) 0. RHZ (SecY only) 0. DIN (SecY only) 0. Structure Cross-correlation coeff. SecYE/ (initial) 0.0 SecYE/ (final) 0. SecYE/ (rotated 0 ) 0. RHZ (SecYE only) 0. DIN (SecYE only) 0.
28 Supplementary Table : Ribosome-SecY interactions SecY residue Ribosome residue Interaction Arg SecY L/ Gln (L) H-bond Arg SecY L/ Ura (S) hydrophilic Arg SecY L/ Ade (S) hydrophilic Val SecY L/ Gua (S) H-bond Val SecY L/ Ura (S) H-bond Val SecY L/ Ade (S) hydrophobic Asn SecY L/ Ade (S) H-bond Tyr SecY L/ Lys (L) hydrophobic (weak) Tyr SecY L/ Val (L) hydrophobic Tyr SecY L/ Ade (S) hydrophilic (weak) Arg SecY L/ Ade (S) H-bond Arg SecY L/ Gua (S) H-bond Gln SecY L/ Ade0 (S) H-bond Gln SecY L/ Gua (S) hydrophilic Gln SecY L/ Ade0 (S) H-bond Gln SecY L/ Ade0 (S) hydrophilic Arg SecY L/ Cyt (S) hydrophilic Arg SecY L/ Gln (L) H-bond Arg SecY L/ Ade (S) H-bond Tyr SecY L/ Cyt (S) H-bond Lys SecY L/ Gua (S) H-bond Lys SecY L/ Ura (S) hydrophilic Phe SecY L/ Cyt (S) H-bond Val SecY L/ Ade (S) H-bond Ile SecY L/ Ura (S) H-bond Ile SecY L/ Gua (S) H-bond Ile SecY L/ Ade (S) hydrophobic Arg SecY L/ Ura (S) H-bond Arg SecY L/ Gua (S) H-bond Arg SecY L/ Ade (S) H-bond Glu0 SecY L/ Ade (S) H-bond Tyr SecY L/ Asp (L) hydrophilic Tyr SecY C-term. Ala0 (L) hydrophobic Ser SecY C-term. Cyt0 (S) hydrophilic Lys SecY C-term. Cyt0 (S) H-bond Asn SecY C-term. Cyt (S) H-bond Asn SecY C-term. Cyt0 (S) hydrophilic Lys SecY C-term. Gua (S) hydrophilic Lys SecY C-term. Ura (S) H-bond Lys SecY C-term. Gua (S) H-bond Tyr SecY C-term. Gua (S) H-bond Gly SecY C-term. Ura (S) H-bond Arg SecY L/ Gln (L) H-bond Arg SecY L/ Ura (S) hydrophilic Arg SecY L/ Ade (S) hydrophilic Val SecY L/ Gua (S) H-bond
29 Supplementary Table : Ribosome- interactions residue Ribosome residue Interaction Arg N-term. Glu (L) H-bond Leu N-term. Leu (L) hydrophobic Glu N-term. Asn (L) hydrophilic Glu N-term. Gln (L) hydrophilic Gly amphi. Glu00 (L) H-bond Lys amphi. Glu (L) H-bond Lys amphi. Glu00 (L) H-bond Arg amphi. Glu (L) H-bond/hydrophilic Glu amphi. Gln (L) H-bond Arg amphi. Phe (L) H-bond Thr amphi. Leu (L) H-bond Lys amphi. Gln (L) hydrophilic (weak) Lys amphi. Asp (L) H-bond Trp amphi. Leu (L) hydrophobic
30 Supplementary Table : NC-ribosome-SecY interactions NC residue Ribosome/SecY residue Interaction Gln0 NC Arg (L) H-bond Arg0 NC Cyt (S) hydrophilic Arg0 NC Ade (S) hydrophilic Arg0 NC Ade0 (S) hydrophilic Gln0 NC Ade (S) H-bond Gln0 NC His0 (L) Hydrophilic Glu00 NC Ade0 (S) H-bond Glu00 NC Gln SecY L/ H-bond Ile NC Ade (S) hydrophobic Ile NC Ade (S) H-bond Gln NC Ade (S) H-bond Gln NC Ade (S) hydrophilic Gln NC Gua (S) H-bond Gln NC Ade (S) H-bond Gln NC Gua (S) H-bond Ile NC Ala SecY C-term. hydrophobic Ile NC Tyr SecY L/ hydrophobic Val NC Ala SecY C-term. hydrophobic Val NC Ala0 (L) hydrophobic Asp NC Thr SecY L/ hydrophilic (weak) Asp NC Arg SecY L/ hydrophilic Asp NC Pro (L) H-bond Gln0 NC Glu0 SecY C-term. hydrophilic Met NC Pro SecY L/ hydrophobic Met NC Leu SecY L/ hydrophobic Phe NC Val SecY TM hydrophobic Phe NC Asn0 SecY L/ H-bond Phe NC Val SecY TM hydrophobic Phe NC Phe SecY TM hydrophobic
31 Supplementary Table : NC-SecY interactions NC residue SecY residue Interaction Glu NC Ile SecY TM H-bond Gly NC Ile SecY TM H-bond Gly NC Asn SecY TM H-bond Leu NC Ile0 SecY TM hydrophobic Leu NC Ile SecY TM hydrophobic Leu NC Ala SecY TM H-bond Leu NC Pro SecY TM H-bond (weak) Leu NC Ile SecY TM hydrophobic Ala0 NC Ile SecY TM hydrophobic Ala0 NC Ile SecY TM H-bond (weak) Ala0 NC Ile SecY TM hydrophobic (weak) Ala0 NC Ile SecY TM hydrophobic Leu NC Ile0 SecY TM0 hydrophobic Leu NC Ile SecY TM H-bond (weak) Leu NC Ile SecY TM hydrophobic Leu NC Ile SecY TM hydrophobic Leu NC Tyr SecY TM hydrophobic Leu NC Ala SecY TM hydrophobic Ile NC Ile SecY TM hydrophobic Ile NC Gly SecY TM H-bond Ser NC Gly SecY TM H-bond (weak) Ser NC Ile SecY TM H-bond Ser NC Ser SecY TM H-bond Ser NC Arg SecY TM H-bond (weak) Gln NC Gly SecY TM H-bond Gln NC Arg SecY TM hydrophilic Gln NC Ser SecY TM hydrophilic Arg NC Arg SecY TM H-bond (weak) Ile NC Pro SecY TM hydrophobic Ile NC Arg SecY TM H-bond Asp NC Ser SecY TM H-bond Asp NC Lys SecY TM H-bond Asp NC Ser SecY TM H-bond (weak) Asn NC Ile SecY TM H-bond (weak) Asn NC Gln SecY TM H-bond
32 Supplementary Table : -SecY interactions residue SecY residue Interaction Thr Val SecY TMb H-bond Thr Val SecY TM H-bond Leu Ile SecY TM hydrophobic Ala Leu SecY TMb hydrophobic Ile Phe SecY TM hydrophobic Ile Tyr SecY TM hydrophobic Leu Ile0 SecY TMb hydrophobic Leu Gln SecY TMb H-bond Leu Ile SecY TM hydrophobic Phe0 Met SecY TMb hydrophobic Phe0 Ile SecY TMb hydrophobic Phe0 Ile0 SecY TMb hydrophobic Leu Ile SecY TM hydrophobic Val Ile SecY TMb hydrophobic Thr Ser SecY TM H-bond Thr Ile SecY TMb H-bond (weak) Leu Ser SecY TM H-bond Leu Phe SecY TM hydrophobic Val0 Phe SecY TM hydrophobic Val0 Phe SecY TM hydrophobic Trp Phe SecY TM hydrophobic Trp Phe SecY TM hydrophobic Trp Phe SecY TM H-bond Trp Ile0 SecY TM hydrophobic Trp Phe SecY TM hydrophobic Val Phe SecY TM hydrophobic Val Gly0 SecY TM H-bond Val Leu SecY TM hydrophobic (weak) Leu Phe SecY TM hydrophobic Trp Asn SecY TM H-bond Trp Ala SecY TM hydrophobic Trp Ala SecY TM H-bond (weak) Trp Leu SecY TM hydrophobic Met Ile SecY TM hydrophobic





33 Supplementary Table : Interactions between H and lipids
Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels. Supplementary Information
 Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels Craig T. Armstrong, Philip E. Mason, J. L. Ross Anderson and Christopher E. Dempsey
Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels Craig T. Armstrong, Philip E. Mason, J. L. Ross Anderson and Christopher E. Dempsey
CS612 - Algorithms in Bioinformatics
 Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site.
 Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Introduction to proteins and protein structure
 Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random
. EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random") S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
Amino Acids. Review I: Protein Structure. Amino Acids: Structures. Amino Acids (contd.) Rajan Munshi
 Rajan Munshi") Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation
 Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is worth 2 points.
 MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
Objective: You will be able to explain how the subcomponents of
 Objective: You will be able to explain how the subcomponents of nucleic acids determine the properties of that polymer. Do Now: Read the first two paragraphs from enduring understanding 4.A Essential knowledge:
Objective: You will be able to explain how the subcomponents of nucleic acids determine the properties of that polymer. Do Now: Read the first two paragraphs from enduring understanding 4.A Essential knowledge:
Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular
 Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular weight markers (M). Supplementary Figure-2. Overlay of
Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular weight markers (M). Supplementary Figure-2. Overlay of
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
SUPPORTING INFORMATION FOR. A Computational Approach to Enzyme Design: Using Docking and MM- GBSA Scoring
 SUPPRTING INFRMATIN FR A Computational Approach to Enzyme Design: Predicting ω- Aminotransferase Catalytic Activity Using Docking and MM- GBSA Scoring Sarah Sirin, 1 Rajesh Kumar, 2 Carlos Martinez, 2
SUPPRTING INFRMATIN FR A Computational Approach to Enzyme Design: Predicting ω- Aminotransferase Catalytic Activity Using Docking and MM- GBSA Scoring Sarah Sirin, 1 Rajesh Kumar, 2 Carlos Martinez, 2
Biological systems interact, and these systems and their interactions possess complex properties. STOP at enduring understanding 4A
 Biological systems interact, and these systems and their interactions possess complex properties. STOP at enduring understanding 4A Homework Watch the Bozeman video called, Biological Molecules Objective:
Biological systems interact, and these systems and their interactions possess complex properties. STOP at enduring understanding 4A Homework Watch the Bozeman video called, Biological Molecules Objective:
Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information.
 Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information. Supplementary Figure 1. Phenotypic analysis of TRBV25-1 + and TRBV25-1 - CD1d-α-GalCerreactive cells.
Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information. Supplementary Figure 1. Phenotypic analysis of TRBV25-1 + and TRBV25-1 - CD1d-α-GalCerreactive cells.
Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al.
 Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al. Supplementary Figure 1 TWIK-1 stability during MD simulations in a phospholipid
Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al. Supplementary Figure 1 TWIK-1 stability during MD simulations in a phospholipid
Lipid Bilayers Are Excellent For Cell Membranes
 Lipid Bilayers Are Excellent For Cell Membranes ydrophobic interaction is the driving force Self-assembly in water Tendency to close on themselves Self-sealing (a hole is unfavorable) Extensive: up to
Lipid Bilayers Are Excellent For Cell Membranes ydrophobic interaction is the driving force Self-assembly in water Tendency to close on themselves Self-sealing (a hole is unfavorable) Extensive: up to
The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation
 www.bioinformation.net Hypothesis Volume 8(9) The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation Zahra Karimi 1 *,
www.bioinformation.net Hypothesis Volume 8(9) The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation Zahra Karimi 1 *,
Modeling and Molecular Dynamics of Membrane Proteins
 Modeling and Molecular Dynamics of Membrane Proteins Emad Tajkhorshid Department of Biochemistry, Center for Biophysics and Computational Biology, and Beckman Institute University of Illinois at Urbana-Champaign
Modeling and Molecular Dynamics of Membrane Proteins Emad Tajkhorshid Department of Biochemistry, Center for Biophysics and Computational Biology, and Beckman Institute University of Illinois at Urbana-Champaign
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.
, Purified EpEX and EpEX analyzed on homogenous 12.") Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Structure of the measles virus hemagglutinin bound to the CD46 receptor. César Santiago, María L. Celma, Thilo Stehle and José M.
 Supporting Figures and Table for Structure of the measles virus hemagglutinin bound to the CD46 receptor César Santiago, María L. Celma, Thilo Stehle and José M. Casasnovas This PDF file includes: Supplementary
Supporting Figures and Table for Structure of the measles virus hemagglutinin bound to the CD46 receptor César Santiago, María L. Celma, Thilo Stehle and José M. Casasnovas This PDF file includes: Supplementary
3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B
 3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B 3.2.1 Characterization of the Ligand Binding Site at GPR109A Receptor Ligands of GPR109A Receptor are Carboxylic Acids Nicotinic acid (pyridine-3-carboxylic
3.2 Ligand-Binding at Nicotinic Acid Receptor Subtypes GPR109A/B 3.2.1 Characterization of the Ligand Binding Site at GPR109A Receptor Ligands of GPR109A Receptor are Carboxylic Acids Nicotinic acid (pyridine-3-carboxylic
Supplementary Information
 Supplementary Information Two common structural motifs for TCR recognition by staphylococcal enterotoxins Karin Erica Johanna Rödström 1, Paulina Regenthal 1, Christopher Bahl 2, Alex Ford 2, David Baker
Supplementary Information Two common structural motifs for TCR recognition by staphylococcal enterotoxins Karin Erica Johanna Rödström 1, Paulina Regenthal 1, Christopher Bahl 2, Alex Ford 2, David Baker
1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled
 Protein Targeting Objectives 1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled As a protein is being synthesized, decisions
Protein Targeting Objectives 1. to understand how proteins find their destination in prokaryotic and eukaryotic cells 2. to know how proteins are bio-recycled As a protein is being synthesized, decisions
(B D) Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.
 Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.") Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine
 Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine") Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
Lecture 33 Membrane Proteins
 Lecture 33 Membrane Proteins Reading for today: Chapter 4, section D Required reading for next Wednesday: Chapter 14, sections A and 14.19 to the end Kuriyan, J., and Eisenberg, D. (2007) The origin of
Lecture 33 Membrane Proteins Reading for today: Chapter 4, section D Required reading for next Wednesday: Chapter 14, sections A and 14.19 to the end Kuriyan, J., and Eisenberg, D. (2007) The origin of
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and in a
 and in a") Supplementary Figure S1. Distribution of atoms along the bilayer normal. Normalized density profiles obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and
Supplementary Figure S1. Distribution of atoms along the bilayer normal. Normalized density profiles obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and
Review II: The Molecules of Life
 Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2007 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2007 Outline Introduction Proteins Carbohydrates Lipids
Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2007 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2007 Outline Introduction Proteins Carbohydrates Lipids
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION TITLE: Structural Basis of Signal Sequence Surveillance and Selection by the SRP-SR Complex AUTHORS and AFFILIATIONS Ottilie von Loeffelholz 1,2, Kèvin Knoops 1,2,6, Aileen Ariosa
SUPPLEMENTARY INFORMATION TITLE: Structural Basis of Signal Sequence Surveillance and Selection by the SRP-SR Complex AUTHORS and AFFILIATIONS Ottilie von Loeffelholz 1,2, Kèvin Knoops 1,2,6, Aileen Ariosa
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Design of isolated protein and RNC constructs, and homogeneity of purified RNCs. (a) Schematic depicting the design and nomenclature used for all the isolated proteins and RNCs used
Supplementary Figure 1 Design of isolated protein and RNC constructs, and homogeneity of purified RNCs. (a) Schematic depicting the design and nomenclature used for all the isolated proteins and RNCs used
Interactions of Polyethylenimines with Zwitterionic and. Anionic Lipid Membranes
 Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
Introduction to Protein Structure Collection
 Introduction to Protein Structure Collection Teaching Points This collection is designed to introduce students to the concepts of protein structure and biochemistry. Different activities guide students
Introduction to Protein Structure Collection Teaching Points This collection is designed to introduce students to the concepts of protein structure and biochemistry. Different activities guide students
SUPPLEMENTARY INFORMATION. Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity
 SUPPLEMENTARY INFORMATION Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity Christopher Woods 1, Maturos Malaisree 1, Ben Long 2, Simon McIntosh-Smith
SUPPLEMENTARY INFORMATION Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity Christopher Woods 1, Maturos Malaisree 1, Ben Long 2, Simon McIntosh-Smith
in-silico Design of Nanoparticles for Transdermal Drug Delivery Application
 Electronic Supplementary Material (ESI) for Nanoscale. This journal is The Royal Society of Chemistry 2018 in-silico Design of Nanoparticles for Transdermal Drug Delivery Application Rakesh Gupta and Beena
Electronic Supplementary Material (ESI) for Nanoscale. This journal is The Royal Society of Chemistry 2018 in-silico Design of Nanoparticles for Transdermal Drug Delivery Application Rakesh Gupta and Beena
The Basics: A general review of molecular biology:
 The Basics: A general review of molecular biology: DNA Transcription RNA Translation Proteins DNA (deoxy-ribonucleic acid) is the genetic material It is an informational super polymer -think of it as the
The Basics: A general review of molecular biology: DNA Transcription RNA Translation Proteins DNA (deoxy-ribonucleic acid) is the genetic material It is an informational super polymer -think of it as the
Molecular Dynamics Simulation of Membrane Channels
 Molecular Dynamics Simulation of Membrane Channels Part II. Structure-Function Relationship and Transport in Aquaporins Emad Tajkhorshid Beckman Institute, UIUC Summer School on Theoretical and Computational
Molecular Dynamics Simulation of Membrane Channels Part II. Structure-Function Relationship and Transport in Aquaporins Emad Tajkhorshid Beckman Institute, UIUC Summer School on Theoretical and Computational
2. Which of the following amino acids is most likely to be found on the outer surface of a properly folded protein?
 Name: WHITE Student Number: Answer the following questions on the computer scoring sheet. 1 mark each 1. Which of the following amino acids would have the highest relative mobility R f in normal thin layer
Name: WHITE Student Number: Answer the following questions on the computer scoring sheet. 1 mark each 1. Which of the following amino acids would have the highest relative mobility R f in normal thin layer
Properties of amino acids in proteins
 Properties of amino acids in proteins one of the primary roles of DNA (but far from the only one!!!) is to code for proteins A typical bacterium builds thousands types of proteins, all from ~20 amino acids
Properties of amino acids in proteins one of the primary roles of DNA (but far from the only one!!!) is to code for proteins A typical bacterium builds thousands types of proteins, all from ~20 amino acids
Cells. Variation and Function of Cells
 Cells Variation and Function of Cells Plasma Membrane= the skin of a cell, it protects and nourishes the cell while communicating with other cells at the same time. Lipid means fat and they are hydrophobic
Cells Variation and Function of Cells Plasma Membrane= the skin of a cell, it protects and nourishes the cell while communicating with other cells at the same time. Lipid means fat and they are hydrophobic
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
Bioinformatics for molecular biology
 Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
Conformational Flexibility of the Peptide Hormone Ghrelin in Solution and Lipid Membrane Bound: A Molecular Dynamics Study
 Open Access Article The authors, the publisher, and the right holders grant the right to use, reproduce, and disseminate the work in digital form to all users. Journal of Biomolecular Structure & Dynamics,
Open Access Article The authors, the publisher, and the right holders grant the right to use, reproduce, and disseminate the work in digital form to all users. Journal of Biomolecular Structure & Dynamics,
paper and beads don t fall off. Then, place the beads in the following order on the pipe cleaner:
 Beady Pipe Cleaner Proteins Background: Proteins are the molecules that carry out most of the cell s dayto-day functions. While the DNA in the nucleus is "the boss" and controls the activities of the cell,
Beady Pipe Cleaner Proteins Background: Proteins are the molecules that carry out most of the cell s dayto-day functions. While the DNA in the nucleus is "the boss" and controls the activities of the cell,
Short polymer. Dehydration removes a water molecule, forming a new bond. Longer polymer (a) Dehydration reaction in the synthesis of a polymer
 Dehydration reaction in the synthesis of a polymer") HO 1 2 3 H HO H Short polymer Dehydration removes a water molecule, forming a new bond Unlinked monomer H 2 O HO 1 2 3 4 H Longer polymer (a) Dehydration reaction in the synthesis of a polymer HO 1 2 3
HO 1 2 3 H HO H Short polymer Dehydration removes a water molecule, forming a new bond Unlinked monomer H 2 O HO 1 2 3 4 H Longer polymer (a) Dehydration reaction in the synthesis of a polymer HO 1 2 3
Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL
 Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL For Questions 1-10 choose ONE INCORRECT answer. 1. Which ONE of the following statements concerning the
Multiple-Choice Questions Answer ALL 20 multiple-choice questions on the Scantron Card in PENCIL For Questions 1-10 choose ONE INCORRECT answer. 1. Which ONE of the following statements concerning the
Proteins are sometimes only produced in one cell type or cell compartment (brain has 15,000 expressed proteins, gut has 2,000).
.") Lecture 2: Principles of Protein Structure: Amino Acids Why study proteins? Proteins underpin every aspect of biological activity and therefore are targets for drug design and medicinal therapy, and in
Lecture 2: Principles of Protein Structure: Amino Acids Why study proteins? Proteins underpin every aspect of biological activity and therefore are targets for drug design and medicinal therapy, and in
Biomolecules: amino acids
 Biomolecules: amino acids Amino acids Amino acids are the building blocks of proteins They are also part of hormones, neurotransmitters and metabolic intermediates There are 20 different amino acids in
Biomolecules: amino acids Amino acids Amino acids are the building blocks of proteins They are also part of hormones, neurotransmitters and metabolic intermediates There are 20 different amino acids in
Maha AbuAjamieh. Tamara Wahbeh. Mamoon Ahram
 12 Maha AbuAjamieh Tamara Wahbeh Mamoon Ahram - - Go to this sheet s last page for definitions of the words with an asterisk above them (*) - You should memorise the 3-letter abbreviations, of all the
12 Maha AbuAjamieh Tamara Wahbeh Mamoon Ahram - - Go to this sheet s last page for definitions of the words with an asterisk above them (*) - You should memorise the 3-letter abbreviations, of all the
Protein Structure Monday, March
 Michael Morales ffice: 204/202 ary; knock hard if you visit as my office is some door. Lab: 238 & 205 ary ffice hours Either make an appointment or just drop by Phone: 829-3965 E-Mail: moralesm@buffalo.edu
Michael Morales ffice: 204/202 ary; knock hard if you visit as my office is some door. Lab: 238 & 205 ary ffice hours Either make an appointment or just drop by Phone: 829-3965 E-Mail: moralesm@buffalo.edu
BIOCHEMISTRY REVIEW. Overview of Biomolecules. Chapter 4 Protein Sequence
 BIOCHEMISTRY REVIEW Overview of Biomolecules Chapter 4 Protein Sequence 2 3 4 Are You Getting It?? A molecule of hemoglobin is compared with a molecule of lysozyme. Which characteristics do they share?
BIOCHEMISTRY REVIEW Overview of Biomolecules Chapter 4 Protein Sequence 2 3 4 Are You Getting It?? A molecule of hemoglobin is compared with a molecule of lysozyme. Which characteristics do they share?
CHAPTER 21: Amino Acids, Proteins, & Enzymes. General, Organic, & Biological Chemistry Janice Gorzynski Smith
 CHAPTER 21: Amino Acids, Proteins, & Enzymes General, Organic, & Biological Chemistry Janice Gorzynski Smith CHAPTER 21: Amino Acids, Proteins, Enzymes Learning Objectives: q The 20 common, naturally occurring
CHAPTER 21: Amino Acids, Proteins, & Enzymes General, Organic, & Biological Chemistry Janice Gorzynski Smith CHAPTER 21: Amino Acids, Proteins, Enzymes Learning Objectives: q The 20 common, naturally occurring
Practice Problems 3. a. What is the name of the bond formed between two amino acids? Are these bonds free to rotate?
 Life Sciences 1a Practice Problems 3 1. Draw the oligopeptide for Ala-Phe-Gly-Thr-Asp. You do not need to indicate the stereochemistry of the sidechains. Denote with arrows the bonds formed between the
Life Sciences 1a Practice Problems 3 1. Draw the oligopeptide for Ala-Phe-Gly-Thr-Asp. You do not need to indicate the stereochemistry of the sidechains. Denote with arrows the bonds formed between the
Supporting material. Membrane permeation induced by aggregates of human islet amyloid polypeptides
 Supporting material Membrane permeation induced by aggregates of human islet amyloid polypeptides Chetan Poojari Forschungszentrum Jülich GmbH, Institute of Complex Systems: Structural Biochemistry (ICS-6),
Supporting material Membrane permeation induced by aggregates of human islet amyloid polypeptides Chetan Poojari Forschungszentrum Jülich GmbH, Institute of Complex Systems: Structural Biochemistry (ICS-6),
Point total. Page # Exam Total (out of 90) The number next to each intermediate represents the total # of C-C and C-H bonds in that molecule.
 The number next to each intermediate represents the total # of C-C and C-H bonds in that molecule.") This exam is worth 90 points. Pages 2- have questions. Page 1 is for your reference only. Honor Code Agreement - Signature: Date: (You agree to not accept or provide assistance to anyone else during this
This exam is worth 90 points. Pages 2- have questions. Page 1 is for your reference only. Honor Code Agreement - Signature: Date: (You agree to not accept or provide assistance to anyone else during this
During the last half century, much effort has been devoted
 Membranes serve as allosteric activators of phospholipase A 2, enabling it to extract, bind, and hydrolyze phospholipid substrates Varnavas D. Mouchlis a,b,1, Denis Bucher b, J. Andrew McCammon a,b,c,1,
Membranes serve as allosteric activators of phospholipase A 2, enabling it to extract, bind, and hydrolyze phospholipid substrates Varnavas D. Mouchlis a,b,1, Denis Bucher b, J. Andrew McCammon a,b,c,1,
Structural analysis of fungus-derived FAD glucose dehydrogenase
 Structural analysis of fungus-derived FAD glucose dehydrogenase Hiromi Yoshida 1, Genki Sakai 2, Kazushige Mori 3, Katsuhiro Kojima 3, Shigehiro Kamitori 1, and Koji Sode 2,3,* 1 Life Science Research
Structural analysis of fungus-derived FAD glucose dehydrogenase Hiromi Yoshida 1, Genki Sakai 2, Kazushige Mori 3, Katsuhiro Kojima 3, Shigehiro Kamitori 1, and Koji Sode 2,3,* 1 Life Science Research
Biomolecules Amino Acids & Protein Chemistry
 Biochemistry Department Date: 17/9/ 2017 Biomolecules Amino Acids & Protein Chemistry Prof.Dr./ FAYDA Elazazy Professor of Biochemistry and Molecular Biology Intended Learning Outcomes ILOs By the end
Biochemistry Department Date: 17/9/ 2017 Biomolecules Amino Acids & Protein Chemistry Prof.Dr./ FAYDA Elazazy Professor of Biochemistry and Molecular Biology Intended Learning Outcomes ILOs By the end
Biology. Lectures winter term st year of Pharmacy study
 Biology Lectures winter term 2008 1 st year of Pharmacy study 3 rd Lecture Chemical composition of living matter chemical basis of life. Atoms, molecules, organic compounds carbohydrates, lipids, proteins,
Biology Lectures winter term 2008 1 st year of Pharmacy study 3 rd Lecture Chemical composition of living matter chemical basis of life. Atoms, molecules, organic compounds carbohydrates, lipids, proteins,
7.012 F 04 Problem Set 1 September 10 th 2004
 MIT Biology Department 7.012: Introductory Biology Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. laudette Gardel ame Question 1 TA Section 7.012 F 04 Problem Set 1 September
MIT Biology Department 7.012: Introductory Biology Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. laudette Gardel ame Question 1 TA Section 7.012 F 04 Problem Set 1 September
Analysis of Molecular Dynamics Simulations of Biomolecules
 Simulating Membrane Channels Part II. Structure-Function Relationship and Transport in Aquaporins Theoretical and Computational Biophysics Dec 2004, Boston, MA http://www.ks.uiuc.edu/training/ Analysis
Simulating Membrane Channels Part II. Structure-Function Relationship and Transport in Aquaporins Theoretical and Computational Biophysics Dec 2004, Boston, MA http://www.ks.uiuc.edu/training/ Analysis
Modeling and Molecular Dynamics of Membrane Proteins
 Modeling and Molecular Dynamics of Membrane Proteins Emad Tajkhorshid Department of Biochemistry, Center for Biophysics and Computational Biology, and Beckman Institute University of Illinois at Urbana-Champagin
Modeling and Molecular Dynamics of Membrane Proteins Emad Tajkhorshid Department of Biochemistry, Center for Biophysics and Computational Biology, and Beckman Institute University of Illinois at Urbana-Champagin
Supplementary Information: A Critical. Comparison of Biomembrane Force Fields: Structure and Dynamics of Model DMPC, POPC, and POPE Bilayers
 Supplementary Information: A Critical Comparison of Biomembrane Force Fields: Structure and Dynamics of Model DMPC, POPC, and POPE Bilayers Kristyna Pluhackova,, Sonja A. Kirsch, Jing Han, Liping Sun,
Supplementary Information: A Critical Comparison of Biomembrane Force Fields: Structure and Dynamics of Model DMPC, POPC, and POPE Bilayers Kristyna Pluhackova,, Sonja A. Kirsch, Jing Han, Liping Sun,
Protein Secondary Structure
 Protein Secondary Structure Reading: Berg, Tymoczko & Stryer, 6th ed., Chapter 2, pp. 37-45 Problems in textbook: chapter 2, pp. 63-64, #1,5,9 Directory of Jmol structures of proteins: http://www.biochem.arizona.edu/classes/bioc462/462a/jmol/routines/routines.html
Protein Secondary Structure Reading: Berg, Tymoczko & Stryer, 6th ed., Chapter 2, pp. 37-45 Problems in textbook: chapter 2, pp. 63-64, #1,5,9 Directory of Jmol structures of proteins: http://www.biochem.arizona.edu/classes/bioc462/462a/jmol/routines/routines.html
List of Figures. List of Tables
 Supporting Information for: Signaling Domain of Sonic Hedgehog as Cannibalistic Calcium-Regulated Zinc-Peptidase Rocio Rebollido-Rios 1, Shyam Bandari 3, Christoph Wilms 1, Stanislav Jakuschev 1, Andrea
Supporting Information for: Signaling Domain of Sonic Hedgehog as Cannibalistic Calcium-Regulated Zinc-Peptidase Rocio Rebollido-Rios 1, Shyam Bandari 3, Christoph Wilms 1, Stanislav Jakuschev 1, Andrea
Secondary Structure North 72nd Street, Wauwatosa, WI Phone: (414) Fax: (414) dmoleculardesigns.com
 Fax: (414) dmoleculardesigns.com") Secondary Structure In the previous protein folding activity, you created a generic or hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously
Secondary Structure In the previous protein folding activity, you created a generic or hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously
Amino acids. Side chain. -Carbon atom. Carboxyl group. Amino group
 PROTEINS Amino acids Side chain -Carbon atom Amino group Carboxyl group Amino acids Primary structure Amino acid monomers Peptide bond Peptide bond Amino group Carboxyl group Peptide bond N-terminal (
PROTEINS Amino acids Side chain -Carbon atom Amino group Carboxyl group Amino acids Primary structure Amino acid monomers Peptide bond Peptide bond Amino group Carboxyl group Peptide bond N-terminal (
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
Secondary Structure. by hydrogen bonds
 Secondary Structure In the previous protein folding activity, you created a hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously folds
Secondary Structure In the previous protein folding activity, you created a hypothetical 15-amino acid protein and learned that basic principles of chemistry determine how each protein spontaneously folds
Chemical Nature of the Amino Acids. Table of a-amino Acids Found in Proteins
 Chemical Nature of the Amino Acids All peptides and polypeptides are polymers of alpha-amino acids. There are 20 a- amino acids that are relevant to the make-up of mammalian proteins (see below). Several
Chemical Nature of the Amino Acids All peptides and polypeptides are polymers of alpha-amino acids. There are 20 a- amino acids that are relevant to the make-up of mammalian proteins (see below). Several
Dynamic contact network between ribosomal subunits enables rapid large-scale rotation during spontaneous translocation
 Published online 24 June 2015 Nucleic Acids Research, 2015, Vol. 43, No. 14 6747 6760 doi: 10.1093/nar/gkv649 Dynamic contact network between ribosomal subunits enables rapid large-scale rotation during
Published online 24 June 2015 Nucleic Acids Research, 2015, Vol. 43, No. 14 6747 6760 doi: 10.1093/nar/gkv649 Dynamic contact network between ribosomal subunits enables rapid large-scale rotation during
Molecular Biology. general transfer: occurs normally in cells. special transfer: occurs only in the laboratory in specific conditions.
 Chapter 9: Proteins Molecular Biology replication general transfer: occurs normally in cells transcription special transfer: occurs only in the laboratory in specific conditions translation unknown transfer:
Chapter 9: Proteins Molecular Biology replication general transfer: occurs normally in cells transcription special transfer: occurs only in the laboratory in specific conditions translation unknown transfer:
Organic molecules are molecules that contain carbon and hydrogen.
 Organic Chemistry, Biochemistry Introduction Organic molecules are molecules that contain carbon and hydrogen. All living things contain these organic molecules: carbohydrates, lipids, proteins, and nucleic
Organic Chemistry, Biochemistry Introduction Organic molecules are molecules that contain carbon and hydrogen. All living things contain these organic molecules: carbohydrates, lipids, proteins, and nucleic
Page 8/6: The cell. Where to start: Proteins (control a cell) (start/end products)
 (start/end products)") Page 8/6: The cell Where to start: Proteins (control a cell) (start/end products) Page 11/10: Structural hierarchy Proteins Phenotype of organism 3 Dimensional structure Function by interaction THE PROTEIN
Page 8/6: The cell Where to start: Proteins (control a cell) (start/end products) Page 11/10: Structural hierarchy Proteins Phenotype of organism 3 Dimensional structure Function by interaction THE PROTEIN
Life Sciences 1a. Practice Problems 4
 Life Sciences 1a Practice Problems 4 1. KcsA, a channel that allows K + ions to pass through the membrane, is a protein with four identical subunits that form a channel through the center of the tetramer.
Life Sciences 1a Practice Problems 4 1. KcsA, a channel that allows K + ions to pass through the membrane, is a protein with four identical subunits that form a channel through the center of the tetramer.
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
Acta Crystallographica Section D
 Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: A conformational landscape for alginate secretion across the outer membrane of Pseudomonas aeruginosa
Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: A conformational landscape for alginate secretion across the outer membrane of Pseudomonas aeruginosa
Practice Exam 2 MCBII
 1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
The Binding Mode of by Electron Crystallography
 The Binding Mode of Epothilone A on α,β-tubulin by Electron Crystallography James H. Nettles, Huilin Li, Ben Cornett, Joseph M. Krahn, James P. Snyder, Kenneth H. Downing Science, Volume 305, August 6,
The Binding Mode of Epothilone A on α,β-tubulin by Electron Crystallography James H. Nettles, Huilin Li, Ben Cornett, Joseph M. Krahn, James P. Snyder, Kenneth H. Downing Science, Volume 305, August 6,
SDS-Assisted Protein Transport Through Solid-State Nanopores
 Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Judy Wieber. Department of Computational Biology. May 27, 2008
 Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 27, 2008 Outline Introduction Proteins Carbohydrates
Review II: The Molecules of Life Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 27, 2008 Outline Introduction Proteins Carbohydrates
Four Classes of Biological Macromolecules. Biological Macromolecules. Lipids
 Biological Macromolecules Much larger than other par4cles found in cells Made up of smaller subunits Found in all cells Great diversity of func4ons Four Classes of Biological Macromolecules Lipids Polysaccharides
Biological Macromolecules Much larger than other par4cles found in cells Made up of smaller subunits Found in all cells Great diversity of func4ons Four Classes of Biological Macromolecules Lipids Polysaccharides
Chemistry 121 Winter 17
 Chemistry 121 Winter 17 Introduction to Organic Chemistry and Biochemistry Instructor Dr. Upali Siriwardane (Ph.D. Ohio State) E-mail: upali@latech.edu Office: 311 Carson Taylor Hall ; Phone: 318-257-4941;
Chemistry 121 Winter 17 Introduction to Organic Chemistry and Biochemistry Instructor Dr. Upali Siriwardane (Ph.D. Ohio State) E-mail: upali@latech.edu Office: 311 Carson Taylor Hall ; Phone: 318-257-4941;
Copyright 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
 Concept 5.4: Proteins have many structures, resulting in a wide range of functions Proteins account for more than 50% of the dry mass of most cells Protein functions include structural support, storage,
Concept 5.4: Proteins have many structures, resulting in a wide range of functions Proteins account for more than 50% of the dry mass of most cells Protein functions include structural support, storage,
Biochemistry 2 Recita0on Amino Acid Metabolism
 Biochemistry 2 Recita0on Amino Acid Metabolism 04-20- 2015 Glutamine and Glutamate as key entry points for NH 4 + Amino acid catabolism Glutamine synthetase enables toxic NH 4 + to combine with glutamate
Biochemistry 2 Recita0on Amino Acid Metabolism 04-20- 2015 Glutamine and Glutamate as key entry points for NH 4 + Amino acid catabolism Glutamine synthetase enables toxic NH 4 + to combine with glutamate
The Structure and Function of Macromolecules
 The Structure and Function of Macromolecules Macromolecules are polymers Polymer long molecule consisting of many similar building blocks. Monomer the small building block molecules. Carbohydrates, proteins
The Structure and Function of Macromolecules Macromolecules are polymers Polymer long molecule consisting of many similar building blocks. Monomer the small building block molecules. Carbohydrates, proteins
Break Desired Compound into 3 Smaller Ones
 Break Desired Compound into 3 Smaller nes A B C Indole ydrazine Phenol When creating a covalent link between model compounds move the charge on the deleted into the carbon to maintain integer charge (i.e.
Break Desired Compound into 3 Smaller nes A B C Indole ydrazine Phenol When creating a covalent link between model compounds move the charge on the deleted into the carbon to maintain integer charge (i.e.
Insights into the Giardia intestinalis Enolase and Human Plasminogen interaction
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2017 Supplementary Information Insights into the Giardia intestinalis Enolase and Human
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2017 Supplementary Information Insights into the Giardia intestinalis Enolase and Human
G protein coupled receptor interactions with cholesterol deep in the membrane
 G protein coupled receptor interactions with cholesterol deep in the membrane Samuel Genheden a,b, Jonathan W. Essex a,c, & Anthony G. Lee c,d * a School of Chemistry, University of Southampton, Southampton,
G protein coupled receptor interactions with cholesterol deep in the membrane Samuel Genheden a,b, Jonathan W. Essex a,c, & Anthony G. Lee c,d * a School of Chemistry, University of Southampton, Southampton,
BIO 311C Spring Lecture 15 Friday 26 Feb. 1
 BIO 311C Spring 2010 Lecture 15 Friday 26 Feb. 1 Illustration of a Polypeptide amino acids peptide bonds Review Polypeptide (chain) See textbook, Fig 5.21, p. 82 for a more clear illustration Folding and
BIO 311C Spring 2010 Lecture 15 Friday 26 Feb. 1 Illustration of a Polypeptide amino acids peptide bonds Review Polypeptide (chain) See textbook, Fig 5.21, p. 82 for a more clear illustration Folding and
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature22394 Supplementary Table 1 Observed intermolecular interactions within the GLP-1:GLP-1R TMD interface. Superscripts refer to the Wootten residue numbering system
SUPPLEMENTARY INFORMATION doi:10.1038/nature22394 Supplementary Table 1 Observed intermolecular interactions within the GLP-1:GLP-1R TMD interface. Superscripts refer to the Wootten residue numbering system
Lecture 15. Membrane Proteins I
 Lecture 15 Membrane Proteins I Introduction What are membrane proteins and where do they exist? Proteins consist of three main classes which are classified as globular, fibrous and membrane proteins. A
Lecture 15 Membrane Proteins I Introduction What are membrane proteins and where do they exist? Proteins consist of three main classes which are classified as globular, fibrous and membrane proteins. A
Macromolecules of Life -3 Amino Acids & Proteins
 Macromolecules of Life -3 Amino Acids & Proteins Shu-Ping Lin, Ph.D. Institute of Biomedical Engineering E-mail: splin@dragon.nchu.edu.tw Website: http://web.nchu.edu.tw/pweb/users/splin/ Amino Acids Proteins
Macromolecules of Life -3 Amino Acids & Proteins Shu-Ping Lin, Ph.D. Institute of Biomedical Engineering E-mail: splin@dragon.nchu.edu.tw Website: http://web.nchu.edu.tw/pweb/users/splin/ Amino Acids Proteins
Green Segment Contents
 Green Segment Contents Parts Reference Guide Green Segment 1 8 2 6 3 4 5 7 1. Amino Acid Side Chain Chart shows the properties and atomic structure of side chains. 2. Amino Acid Side Chains affect protein
Green Segment Contents Parts Reference Guide Green Segment 1 8 2 6 3 4 5 7 1. Amino Acid Side Chain Chart shows the properties and atomic structure of side chains. 2. Amino Acid Side Chains affect protein
The effect of orientation dynamics in melittin as antimicrobial peptide in lipid bilayer calculated by free energy method
 Journal of Physics: Conference Series PAPER OPEN ACCESS The effect of orientation dynamics in melittin as antimicrobial peptide in lipid bilayer calculated by free energy method To cite this article: Sri
Journal of Physics: Conference Series PAPER OPEN ACCESS The effect of orientation dynamics in melittin as antimicrobial peptide in lipid bilayer calculated by free energy method To cite this article: Sri
Catalysis & specificity: Proteins at work
 Catalysis & specificity: Proteins at work Introduction Having spent some time looking at the elements of structure of proteins and DNA, as well as their ability to form intermolecular interactions, it
Catalysis & specificity: Proteins at work Introduction Having spent some time looking at the elements of structure of proteins and DNA, as well as their ability to form intermolecular interactions, it
Metal Species for Amide Hydrolysis. Literature Seminar, Kiyomichi SHINODA (M1)
") Metal Species for Amide ydrolysis Literature Seminar, 20130511 Kiyomichi SIDA (M1) Table of Contents 1 Introduction 2 Residue- or Sequence-Selective ydrolysis of Amides 3 Protein-Selective ydrolysis of
Metal Species for Amide ydrolysis Literature Seminar, 20130511 Kiyomichi SIDA (M1) Table of Contents 1 Introduction 2 Residue- or Sequence-Selective ydrolysis of Amides 3 Protein-Selective ydrolysis of
Coarse Grained Molecular Dynamics Simulations of Transmembrane Protein-Lipid Systems
 Int. J. Mol. Sci. 2010, 11, 2393-2420; doi:10.3390/ijms11062393 OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Article Coarse Grained Molecular Dynamics
Int. J. Mol. Sci. 2010, 11, 2393-2420; doi:10.3390/ijms11062393 OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Article Coarse Grained Molecular Dynamics
Lipids: diverse group of hydrophobic molecules
 Lipids: diverse group of hydrophobic molecules Lipids only macromolecules that do not form polymers li3le or no affinity for water hydrophobic consist mostly of hydrocarbons nonpolar covalent bonds fats
Lipids: diverse group of hydrophobic molecules Lipids only macromolecules that do not form polymers li3le or no affinity for water hydrophobic consist mostly of hydrocarbons nonpolar covalent bonds fats