ACCEPTED. Revised IAI
|
|
|
- Kristian Bryan
- 6 years ago
- Views:
Transcription
1 IAI Accepts, published online ahead of print on 7 April 2008 Infect. Immun. doi: /iai Copyright 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Revised IAI Acquisition of the vatpase proton pump and phagosome acidification is essential for escape of Francisella tularensis into the macrophage cytosol 1 Marina SantiC, 2 Rexford Asare, 1 Ivana Skrobonja, 2 Snake Jones, and 2 Yousef Abu Kwaik* 1 Department of Microbiology and Parasitology, University of Rijeka, Medical Faculty, Rijeka, Croatia; 2 Department of Microbiology and Immunology, University of Louisville, College of Medicine, Louisville, KY Key words: trafficking, ph, intracellular, Listeria Short title: Acidification of the F. tularensis phagosome Address correspondence to: Yousef Abu Kwaik, Department of Microbiology and Immunology, Room 412, University of Louisville College of Medicine, 319 Abraham Flexner Way 55A, Louisville, KY 40202, USA. Phone: ; Fax: ; abukwaik@louisville.edu
2 ABSTRACT The F. tularensis-containing phagosome (FCP) matures to a late endosome-like phagosome prior to bacterial escape into the cytosol of macrophages, where bacterial proliferation occurs. Our data show that within the first 15 min post-infection of primary human monocyte-derived macrophages (hmdms); ~90% of the FCPs acquire the proton vatpase pump and the lysomotropic dye, LysoTracker, which concentrates in acidic compartments, similar to phagosomes harboring the Listeria monocytogenes control. Acquisition of the proton vatpase pump and the lysomotropic dye is gradually lost by min post-infection, which coincides with bacterial escape into the cytosol of hmdms. Co-localization of phagosomes harboring the igld mutant with the vatpase and the LysoTracker was also transient and the loss of colocalization was faster than the wild type strain, which is consistent with faster escape of the igld mutant into the macrophage cytosol. In contrast, co-localization of both makers with phagosomes harboring the iglc mutant was persistent, which is consistent with fusion to the lysosomes and failure of the iglc mutant to escape into the macrophage cytosol. We have utilized a fluorescence microscopy-based phagosome integrity assay for differential labeling of vacuolar vs. cytosolic bacteria using anti-bacterial antibodies loaded into the cytosol of live hmdms. We show that specific inhibition of the proton vatpase pump by Bafilomycin A1 (BFA) blocks escape of the wild type strain of F. tularensis and the igld mutant into the cytosol of hmdms, similar to L. monocytogenes. The effect of BFA on blocking bacterial escape into the cytosol is completely reversible as the bacteria escape after removal of BFA. We also show that the limited fusion of the FCP to lysosomes is not due to failure to recruit the late endosomal fusion regulator, Rab7. Therefore, within few minutes of its biogenesis, the FCP transiently acquires the proton vatpase pump to acidify the phagosome, and this transient acidification is essential for subsequent bacterial escape into the macrophage cytosol. 2
3 INTRODUCTION Francisella tularensis is a facultative intracellular bacterium that causes tularemia in many mammalian species including humans (7, 10, 29). Due to its high infectivity and morbidity and mortality, F. tularensis has been classified as Category A bioterrorism agent. There are four closely-related subspecies of F. tularensis (tularensis, holarctica, mediasiatica, and novicida), and subspecies tularensis is the most virulent to humans (7, 10-12, 26, 28, 29). F. tularensis subsp. novicida is attenuated in humans, but replicates robustly within primary human and mice macrophages, causes disease in mice, and is an attractive model to study pathogenesis of tularemia (16, 26, 29). Importantly, intracellular trafficking of F. tularensis subsp. novicida within primary human and mice macrophages is indistinguishable from the two virulent subsp. tularensis and holarctica (6, 8, 13, 24, 27). Therefore, F. tularensis subsp. novicida is a useful model to study intracellular trafficking of F. tularensis subsp. Ingested particles are normally processed by macrophages through the default endosomal lysosomal degradation pathway, which is one of the first lines of defense against microbial infection (9, 14, 15, 26). The nascent phagosome matures to an early endosome stage regulated by Rab5, followed by maturation into a late endosome regulated by Rab7. The late endosome becomes acidified upon acquisition of the proton vatpase pump that imports hydrogen protons into the phagosome. The acidified late endosome then fuses to lysosomes and becomes a hydrolase-rich phagolysosome, within which most ingested particles are degraded. This process is very rapid and is completed within minutes of formation of the phagosome (see ref. (26) for a recent review). Therefore, many intracellular pathogens have evolved with idiosyncratic strategies to avoid a fatal fate within the phagolysosomes. For example, Listeria monocytogenes escapes from the acidified late endosome-like phagosome into the cytosol of the host cell. Escape of L. monocytogenes into the cytosol requires the vatpase pump to acidify the 3
4 phagosome, resulting in activation of the pore-forming hemolysin, Listeriolysin O (22). Inhibition of the vatpase pump by Bafilomycin A1 (BFA) (5, 21) blocks escape of L. monocytogenes from the phagosome into the cytosol of the host cell (4). The intracellular fate of three F. tularensis subsp (tularensis, holarctica-derived LVS and novicida) is similar but unique when compared to other intracellular pathogens (6, 8, 13, 24, 27). The Francisella-containing phagosome (FCP) matures into a late endosomal-like stage that acquires the late endosomal marker LAMP-2 followed by bacterial escape into the macrophage cytosol. It is not known whether the FCP acquires the proton vatpase pump and becomes acidified transiently prior to bacterial escape into the cytosol by min post-infection. The Francisella pathogenicity island (FPI) (20) gene iglc, and its regulator MglA are essential for bacterial escape into the cytosol of hmdms (24, 27). In contrast, recent studies have shown that a F. tularensis mutant defective in the FPI gene igld, which is downstream of iglc, escapes into the cytosol of hmdms but fails to replicate within the cytosol (25). Therefore, F. tularensis transduces signals into the macrophage cytosol to render it hospitable for proliferation. Based on earlier ultra structural studies, disruption of the FCP was thought to occur by 4-6h post-infection (8, 13, 24, 27). Based on these observations it was determined that the lysomotropic agent, LysoTracker, which concentrates in acidic compartments, does not concentrate around F. tularensis within macrophages at 4h after infection, but earlier stages of the infection have not been examined (8). However, we and others have recently utilized different fluorescence-based assays for differential labeling of vacuolar vs. cytosolic bacteria to determine more accurately the kinetics of disruption of the FCP in macrophages (17, 25). The data from two independent groups have shown that the FCP becomes disrupted by minutes post-infection (6, 25). Therefore, the earlier study (8) that determined the status of acidification of the FCP presumed the FCP to be intact at 4h post-infection, while the recent more accurate 4
5 studies have shown that the disrupted phagosome equilibrates with the cytosol within minutes post-infection. Importantly, phagosome acidification normally occurs within min after phagosome biogenesis from the plasma membrane. Therefore the objectives of the present study were to determine whether the FCPs acquire the vatpase proton pump and become acidified transiently prior to bacterial escape into the cytosol by min. Our data show that the FCP transiently acquires the proton vatpase pump and becomes acidified within min post-infection and this acidification is essential for escape of F. tularensis into the cytosol of human macrophages, similar to L. monocytogenes. 5
6 MATERIALS AND METHODS Bacteria and macrophages The wild-type F. tularensis subsp. novicida strain U112 and it isogenic iglc and igld mutant have been described previously (16), and grown on buffered-charcoal yeast extract (BCYE) agar plates. The Tetracycline resistant plasmid pkk214 encodes gfp was introduced to the F. tularensis strains (1). Legionella pneumophila strain AA100 expressing GFP has been grown as described previously (18). The hemolytic EGD strain (serovar1/2a) of L. monocytogenes was grown on blood agar at 37 C for 24 hours (2). To prepare hmdms, peripheral blood monocytes were isolated from healthy volunteers with no history of tularemia or Legionnaires disease and hmdms were prepared as we described previously (24). Obtaining blood was approved by the institutional IRB with a consent form according to standard federal laws. Confocal laser scanning microscopy For confocal microscopy, the hmdms were allowed to attach to glass cover slips in 24- well culture plates. At different time points after infection the cells were fixed, permeabilized and blocked as we described previously (24). Co-localization of the Francisella-containing phagosomes (FCP) with the vatpase proton pump was determined using rabbit anti-vatpase antiserum (Chemicon International), as we described previously (3). Anti Rab7 rabbit antiserum was obtained from Sigma and used at 1:100 dilution. Anti- rabbit and anti-mouse secondary antibodies conjugated to Alexa flour-594 were obtained from Molecular Probes (Eugene, Oregon). To load the cytosol of live hmdms with antibacterial antibodies, we used the glass bead loading technique (17, 23) that we have recently adapted for the infection of hmdms by L. 6
7 pneumophila (19) and F. tularensis (25). Briefly, the cytoplasm of live hmdms was loaded with antibacterial antibodies and the glass beads to permeabilize the plasma membrane. Infections were carried out using an MOI of 10 and the infection was synchronized by centrifugation at 500 xg for 5 min. After 1 h infection, cells were washed three times with PBS and 400 µl of antibacterial monoclonal antibodies was added on the top of the cover slips, along with an aliquot of 0.5 g of acid washed sterile glass beads (Sigma; microns). The beads were rolled over the cells 12 times, which had no detectable effect on the viability of cells as confirmed by Trypan Blue exclusion. The glass beads were washed off immediately with PBS and the cells were incubated at 37 C for 1 h to allow a sufficient time for the antibodies to bind. The cells were then fixed and processed with conjugated secondary antibodies. Controls were either cells subjected to the same treatment without the glass beads or cells were fixed and permeabilized with 0.05% Triton X-100 for 15 min on ice. Acidification of the F. tularensis-containing phagosomes was determined using the lysomotropic LysoTracker red DND-99 (Molecular Probes). Briefly, hmdms were grown on glass cover slips in 24-well plates and then used for subsequent invasion assays with live or heatkilled bacteria as described above. Thirty minutes prior to the time point, the cells were washed and incubated with 1 µm LysoTracker/ml, washed three times with PBS, fixed with 4% paraformaldehyde and then mounted on glass slides for confocal microscopy analysis. Inhibition of the vatpase pump was accomplished by pre-treating the cells for 1h with 25nM BFA Bafilomycin A1 (BFA), which was maintained during infection. All confocal microscopy analyses were performed on 100 infected cells from three different cover slips for each time point in each experiment, and all experiments were performed three times. The images shown in the figures are stacks of µm-thick z series of sections. 7
8 RESULTS AND DISCUSSION Early acquisition of the proton vatpase pump by the Francisella-containing phagosome Previous studies have demonstrated that upon entry of F. tularensis into macrophages, the FCP matures into a late endosome-like stage that co-localizes with the late endosomal markers LAMPs followed by phagosomal disruption during the first few hours of phagosome biogenesis (8). However, since recent and more accurate studies have shown that the FCPs are disrupted by min post-infection (6, 25), examination of intact vacuoles for any functional or cellular assay should be examined prior to its disruption. It is not known whether the FCP acquires the proton vatpase pump, and whether this acquisition is required for disruption of the phagosome and bacterial escape into the cytosol. Since phagosome acdification is exhibited within min of phagosome biogenesis, we determined whether the FCP acquires the proton vatpase pump within the first min post-infection. Since L. pneumophila excludes the vatpase proton pump from its phagosome, it was used as a negative control (3). Since phagosomes harboring Listeria monocytogenes acquire the proton vatpase to acidify the phagosome (22) prior to bacterial escape into the host cell cytosol, L. monocytogenes was used as a positive control. Our data showed that the Legionella-containing phagosomes excluded the proton vatpase pump (Fig. 1) while the Listeria-containing phagosomes acquired the proton vatpase pump (Fig. 1 and 2). Approximately ~90% of the wild type strain FCPs acquired the proton vatpase pump by 15 min post-infection. Co-localization of the FCP with the vatpase decreased to ~55% by 60 min post-infection (Student t-test, p<0.002), which coincided with disruption of the FCP and bacterial escape into the cytosol by this time point (Fig. 1 and 2). We conclude that 8
9 the FCPs acquire the proton vatpase pump transiently within the first 15 min post-tinfection and this acquisition is gradually and rapidly lost, which coincides with bacterial escape into the macrophage cytosol. Persistent acquisition of the vatpase by phagosomes harboring the iglc mutant and rapid loss by phagosomes harboring the igld mutant The F. tularensis bicistronic intracellular growth locus iglcd is located within the F. tularensis pathogenicity island (27) and both genes are essential for intracellular proliferation. While the iglc mutant fails to escape from the phagosomes, which fuse to lysosomes (27), the igld mutant bacteria escape into the macrophage cytosol faster than the parental strain but fail to replicate in the cytosol of hmdms (25). When the igld mutant-containing phagosomes were examined for acquisiton of the vatpase, our data showed that ~80% of them acquired the proton vatpase pump by 15 min post-infection (Fig. 1 and 2), followed by a rapid decrease to ~ 40% at 30 min after infection (Fig. 1 and 2), which was significantly faster than the wild type strain (student t-test, p<0.01). This is consistent with rapid escape of this mutant into the cytosol compared to the wild type strain (25). In contrast, 85% of the iglc mutant-containing phagosomes acquired and retained the proton vatpase pump at min (Fig. 1 and 2). We conclude that the FCPs of the igld mutant acquire the proton vatpase pump that is lost faster than the parental strain, consitent with the faster escape of the igld mutant into the macrophage cytosol (25). However, the FCPs of the iglc mutant acquire and retain the vatpase consistent with fusion of its phagosomes to lysosomes and failure of the iglc mutant to escape into the cytosol (25). 9
10 F. tularensis is trafficked into acidified vacules during early stages of phagosome biogenesis Direct measurements of acidity of the region surrounding F. tularensis within macrophages at 4 h after infection have shown it to be neutral and not to acquire the lysomotropic agent LysoTracker, suggesting equilibration of the phagosomal environment with that of the macrophage cytosol (8). However, more recent studies have shown that the FCPs become disrupted within min post-infection and >90% of the FCPs are disrupted by 4h postinfection (25). Importantly, phagosome acidification of internalized particles is exhibited within min after formation of the phagosome. Therefore, we monitored acidification of the FCP at minutes using the lysomotropic agent LysoTracker Red DND-99, which concentrates in acidified vesicles and compartments. We used L. monocytogenes as our positive control since its phagosome acquires the vatpase pump (5, 21) to acidify the phagosome, resulting in activation of the pore-forming hemolysin, Listeriolysin O that disrupts the phagosome to allow bacterial escape into the cytosol (22). Our results showed that 80% of the wild type strain FCPs colocalized with the LysoTracker red DND-99 at 15 and 30 min post infection (Fig. 3 and 4). At 1 h after infection of hmdms with the wild type strain, most of the FCPs lost colocalization with the LysoTracker (Fig. 3 and 4), consistent with the gradual decline of colocalization with the vatpase (Fig. 1 and 2), and bacterial escape into the cytosol. Bafilomycin A1 (BFA), a specific pharmacological inhibitor of the vatpase was used to inhibit phagosomal acidification. Inhibition of the vatpase pump by BFA (5, 21) blocks phagosomal acidification and escape of L. monocytogenes from the phagosome into the cytosol of the host cell (4). Therefore, we examined whether BFA blocks acidification of the FCP, using L. monocytogenes as a control. Our data showed that treatment of hmdms with BFA 10
11 dramatically reduced co-localization of the LysoTracker red DND-99 with the FCP; similar to the L. monocytogenes control (Fig. 3 and 4). Therefore, the wild type strain of F. tularensis resides transiently in acidified vacuoles during the first ~130 min after infection and this acidification is blocked by BFA, which is a specific inhibitor of the vatpase pump. Persistent acidification of phagosomes harboring the iglc mutant but rapid loss of acidification of phagosomes harboring the igld mutant Since phagosomes harboring the iglc and igld mutants acquired the vatpase pump, which was rapidly lost by the igld mutant-containing phagosomes, we examined whether the iglc mutant was trafficked into an acidified compartment while the igld mutant rapidly escaped from the acidified compartment. The data showed that similar to the wild type strain, the igld mutant containing phagosome co-localized with the LysoTracker red DND-99 at 15 min after infection but rapidly lost this co-localization at minutes (Fig. 3 and 4). In contrast, most of the iglc mutant-containing phagosomes persistently acquired and retained the LysoTracker red DND-99 (colocalization 80%) during the min time points after infection (Fig. 3 and 4). These data show that while the iglc mutant FCPs are persistently acidified by 15 min postinfection, the igld mutant FCPs are transiently acidified but the mutant bacteria rapidly escape this acidic compartment. These observations are consistent with the rapid escape of the igld mutant from the phagosome into the macrophage cytosol (6, 25). The proton vatpase pump is essential for bacterial escape into the cytosol It has recently been shown that by minutes after infection the FCP becomes disrupted and its integrity is compromised, and is followed by bacterial escape into the cytosol (6, 11
12 25). Inhibition of the vatpase pump by the specific inhibitor Bafilomycin A1 (BFA) (5) blocks escape of L. monocytogenes from the phagosome into the cytosol of the host cell (4). Therefore, we examined whether acqusition of the vatpase pump to acidify the FCP was required for the ability of F. tularensis to escape into the macrophage cytosol. We utilized BFA as the gold standard specific inhibitor of the proton vatpase pump (5) to block acidification of the late endosomal and late endosomal-like phagosomal compartments to determine the effect on bacterial escape into the macrophage cytosol. The hmdms were pre-treated with 25nM BFA for 1h, infected as described above, and examined for integrity of the FCP using the glass bead loading technique. In this assay, the macrophage cytosol is loaded with specific anti-bacterial antibodies after selective permeabilization of the plasma memberane. The infected cells are exameind by fluorescence microscopy for integrity of the phagosome (17), since bacteria in intact phagosomes do not bind the antibodies while cytosolic bacteria/ or bacteria in compromised phagosomes bind the antibodies (25). For a negative control, infected cells were treated with monoclonal antibody without permeabilization of the plasma membrane by the glass beads, which showed no binding of the antibody to intracellular bacteria for any of the strains tested (Fig. 5 and 6). For the positive control, the cells were permeabilized by Triton-X100, which allowed the antibodies to bind to all intracellular bacteria for all the strains (Fig. 5 and 6). L. monocytogenes was used as a control strain (Fig. 5 and 6). The data showed that BFA blocked escape of L. monocytogenes into the cytosol. In contrast to untreated cells, the wild type F. tularensis failed to escape from the phagosome in BFA-treated hmdms at all time points up to 6h postinfection (Student t-test, p<0.001) (Fig. 5 and 6). Pre-treatment of wild type F. tularensis with BFA for 6 h had no effect on viability, escape from the FCP, or intracellular replication within un-treated hmdms (data not shown). Importantly, the effect of BFA was completely reversible (Fig. 5 and 6), since subsequent incubation of BFA-treated infected hmdms in BFA- 12
13 free medium for 1h reversed the effect of BFA on inhibition of bacterial escape, as evident by subsequent escape of F. tularensis from the FCP into the cytosol after removal of the BFA inhibitior (Fig. 5 and 6). Similar results to the BFA treatment were obtained using NH 4 Cl to neutralize the cytosolic vesicles and compartments during the first 60 min post-infection of hmdms (data not shown). Therefore, we conclude that functional activity of the proton vatpase pump to acidify the FCP within the first 15 min post-infection is essential for disruption of the FCP and subsequent escape of F. tularensis from the phagosome into the cytosol of hmdms. Future studies should determine the role of acidification in the biochemical mechanisms that trigger disruption of the phagosomal membrane and bacterial escape into the cytosol. Inhibiton of the vatpase proton pump blocks rapid escape of the igld mutant into the macrophage cytosol Since the igld mutant escapes from the FCP faster than the parental strain but fails to replicate in the cytosol of hmdms (25), we examined whether the rapid escape of the igld mutant from the phagosome was blocked by inhibiting the vatpase proton pump by BFA. Since the iglc mutant does not escape from the phagosome, it was included as a control. The data showed that inhibition of the vatpase proton pump by BFA blocked escape of the igld mutant into the cytosol of hmdms when examined up to 6h post-infection (Fig. 5 and 6). As expected, there was no detectable effect of inhibition of the vatpase proton pump on failure of the iglc mutant control to escape from the phagosome (Fig. 5 and 6). Taken together, we conclude the functional activity of the vatpase proton pump to acidify the FCP is essential for escape of F. tularensis into the macrophage cytosol. 13
14 Rab7 is recruite transiently to the FCP The main regulator of fusion of late endosome to lysosomes is the Rab7 GTPase. Whether the limited fusion of the FCP to lysosomes is due to failure to recruite Ra7 is not known. We utilized confocal microscopy using anti-rab7 antiserum to examine whether Rab was recruited to the FCP prior to bacterial release to the cytoplasm. Our data show that similar to the kinetics of recruitment of other late endosomal markers, such as LAMP1 and LAMP2, Rab7 was was transiently recruited to the FCP and was lost by 60 min post-infection, which conicides with bacterial escape to the cytosol (Fig. 7). The FCP harboing the iglc mutant positive control, which fuses to lysosomes acquired Rab7 at a much higher frequency (>90%) by 30 min post-infection, but the marker was lost by 60 min post-infection., consistent with fusion to the lysosomes that do not have Rab7 (Fig. 7). Phagosomes harboring the L. pneumphila strain AA100 negative control showed minimal acquistion of Rab7 (Fig. 7). Disruption of the FCP was thought to occur by 4-6h post-infection, when examined in earlier ultrastructural studies that can only detect major structural alterations in integrity of the phagosome (8, 13, 24, 27). Therefore, the lysomotropic agent, LysoTracker, has been found not to concentrate around F. tularensis within macrophages at 4h after infection, but earlier stages of the infection have not been examined in these reported studies (8). However, acidification of the phagosome is expected to occur within the first minutes after biogenesis of the phagosome from the plasma membrane. Data from two independent groups utilizing fluorescence microscopy-based assays for differential labeling of vacuolar vs. cytosolic bacteria have shown that the FCP becomes disrupted by minutes post-infection (6, 25). Therefore, the earlier study (8) that has determined the status of acidification of the FCP presumed the FCP to be intact at 4h post-infection, while the recent more accurate studies have shown that the disrupted phagosome equilibrates with the cytosol within minutes post-infection (6, 25). Our current 14
15 data show that within the first 15 min post-infection, the FCPs acquire the proton vatpase pump transiently and co-localize with the lysotracker dye that concentrates in acidic compartments. Both markers are gradually lost from the FCP by min post-infection, which coincides with disruption of the FCPs and bacterial egress into the cytosol within min post-infection. Inhibition of the proton vatpase pump by the specific inhibitor BFA abolishes the ability of the wild type strain of F. tularensis and its igld isogenic mutant to escape into the cytosol of hmdms, and this inhibition is completely reversible. Thus, acquition of the proton vatpase pump and acidification of the FCP is essential for disruption of the FCP and escape of F. tularensis into the macrophage cytosol. Our data also show that the limited fusion of the FCP to lysosomes is not due to failure to acquire the Rab7 GTPase, which controls fusion of late endosomes to lysosomes. We speculate that the FCP does not block fusion to the lysosomes but rather most of the oragansims escape into the cytosol upon phagosome acidification and prior to lysosomal fusion. Our data suggest that at the cellular level, trafficking of F. tularensis and its subsequent escape into the cytosol may resemble that of L. monocytogenes but the biochemical mechanisms involved are likely to be different. Interestingly, genomic analyses of F. tularensis have indicated the absence of potential hydrolytic enzymes involved in lysis of eukaryotic membranes. Whether IglC and/or IglD are directly involved in lysis of the FCP membrane is not known. We speculate that the proton vatpase pump is essential for activation of a bacterial protein that disrupts the FCP, or that the acidic environment within the FCP is essential to trigger specific bacterial gene expression involved in disruption of the FCP. Future studies are directed at dissecting the molecular and biochemical mechanisms involved in disruption of the acidified FCP. 15
16 Acknowledgments YAK is supported by Public Health Service Awards R01AI and R01AI from NIAID and by the commonwealth of Kentucky Research Challenge Trust Fund. MS is supported by the Ministry of Science, Education and Sports of Republic of Croatia ( ). 16
17 References 1. Abd, H., T. Johansson, I. Golovliov, G. Sandstrom, and M. Forsman Survival and growth of Francisella tularensis in Acanthamoeba castellanii. Appl Environ Microbiol 69: Abram, M., and M. Doric Primary Listeria monocytogenes infection in gestating mice. Folia Microbiol (Praha) 42: Asare, R., and Y. Abu Kwaik Early trafficking and intracellular replication of Legionella longbeachaea within an ER-derived late endosome-like phagosome. Cell Microbiol 9: Beauregard, K. E., K. D. Lee, R. J. Collier, and J. A. Swanson ph-dependent perforation of macrophage phagosomes by listeriolysin O from Listeria monocytogenes. J Exp Med 186: Bowman, E. J., A. Siebers, and K. Altendorf Bafilomycins: a class of inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells. Proc Natl Acad Sci U S A 85: Checroun, C., T. D. Wehrly, E. R. Fischer, S. F. Hayes, and J. Celli Autophagy-mediated reentry of Francisella tularensis into the endocytic compartment after cytoplasmic replication. Proc Natl Acad Sci U S A 103: Choi, E Tularemia and Q fever. Med Clin North Am 86: Clemens, D. L., B. Y. Lee, and M. A. Horwitz Virulent and avirulent strains of Francisella tularensis prevent acidification and maturation of their phagosomes and escape into the cytoplasm in human macrophages. Infect Immun 72: Duclos, S., and M. Desjardins Subversion of a young phagosome: the survival strategies of intracellular pathogens. Cell Microbiol 2: Ellis, J., P. C. Oyston, M. Green, and R. W. Titball Tularemia. Clin Microbiol Rev 15: Forsman, M., G. Sandstrom, and B. Jaurin Identification of Francisella species and discrimination of type A and type B strains of F. tularensis by 16S rrna analysis. Appl Environ Microbiol 56: Forsman, M., G. Sandstrom, and A. Sjostedt Analysis of 16S ribosomal DNA sequences of Francisella strains and utilization for determination of the phylogeny of the genus and for identification of strains by PCR. Int J Syst Bacteriol 44: Golovliov, I., V. Baranov, Z. Krocova, H. Kovarova, and A. Sjostedt An attenuated strain of the facultative intracellular bacterium Francisella tularensis can escape the phagosome of monocytic cells. Infect Immun 71: Hackstadt, T Redirection of host vesicle trafficking pathways by intracellular parasites. Traffic 1: Kahn, R. A., H. Fu, and C. R. Roy Cellular hijacking: a common strategy for microbial infection. Trends Biochem Sci 27: Lauriano, C. M., J. R. Barker, F. E. Nano, B. P. Arulanandam, and K. E. Klose Allelic exchange in Francisella tularensis using PCR products. FEMS Microbiol Lett 229: McNeil, P. L., and E. Warder Glass beads load macromolecules into living cells. J Cell Sci 88 ( Pt 5): Molmeret, M., D. Bitar, L. Han, and Y. Abu Kwaik Disruption of the phagosomal membrane and egress of Legionella pneumophila into the cytoplasm during 17
18 late stages of the intracellular infection of macrophages and Acanthamoeba polyphaga. Infect Immun 72: Molmeret, M., M. Santic, R. Asare, R. A. Carabeo, and Y. Abu Kwaik Rapid escape of the dot/icm mutants of Legionella pneumophila into the cytosol of mammalian and protozoan cells. Infect Immun 75: Nano, F. E., N. Zhang, S. C. Cowley, K. E. Klose, K. K. Cheung, M. J. Roberts, J. S. Ludu, G. W. Letendre, A. I. Meierovics, G. Stephens, and K. L. Elkins A Francisella tularensis pathogenicity island required for intramacrophage growth. J Bacteriol 186: Poole, B., and S. Ohkuma Effect of weak bases on the intralysosomal ph in mouse peritoneal macrophages. J Cell Biol 90: Portnoy, D. A., T. Chakraborty, W. Goebel, and P. Cossart Molecular deteminants of Listeria monocytogene pathogenesis. Infect.Immun. 60: Reddy, A., E. V. Caler, and N. W. Andrews Plasma membrane repair is mediated by Ca(2+)-regulated exocytosis of lysosomes. Cell 106: Santic, M., M. Molmeret, and Y. Abu Kwaik Modulation of biogenesis of the Francisella tularensis subsp novicida-containing phagosome in quiescent human macrophages and its maturation into a phagolysosome upon actiavtion by IFN-gamma. Cell. Microbiol. 7: Santic, M., M. Molmeret, J. R. Barker, K. E. Klose, A. Dekanic, M. Doric, and Y. Abu Kwaik A Francisella tularensis pathogenicity island protein essential for bacterial proliferation within the host cell cytosol. Cell Microbiol 9: Santic, M., M. Molmeret, K. E. Klose, and Y. Abu Kwaik Francisella tularensis travels a novel, twisted road within macrophages. Trends Microbiol 14: Santic, M., M. Molmeret, K. E. Klose, S. Jones, and Y. Abu Kwaik The Francisella tularensis pathogencity islad protein IglC and its regulator MglA are essential for modulating phagosome biogenesis and subsequent bacterial escape into the cytoplasm. Cell. Microbiol. 7: Saslaw, S., H. T. Eigelsbach, J. A. Prior, H. E. Wilson, and S. Carhart Tularemia vaccine study. II. Respiratory challenge. Archives of Internal Medicine 107: Tarnvik, A., M. Eriksson, G. Sandstrom, and A. Sjostedt Francisella tularensis- -a model for studies of the immune response to intracellular bacteria in man. Immunology 76:
19 Figure legends Fig. 1. Transient acquisition of the vatpase proton pump by the FCP during earlyl stages of infection. Representative confocal microscopy images of acquisition of the vatpase proton pump by the FCP within hmdms. Co-localization of the FCP with the vatpase proton pump by the GFP-expressing wild type (wt) strain and its isogenic iglc or igld mutant was examined at 15 (not shown), 30 and 60 min post-infection. The wild type strain AA100 of L. pneumophila expressing GFP was used as a negative control and L. monocytogenes (Lm) was used as a positive control. The images are representatives of 100 infected cells examined from three different coverslips. The results shown are representative of three independent experiments. Fig. 2. Quantitative analyses of transient acquisition of the vatpase proton pump by the FCP. Quantification of co-localization of the FCPs of wild type (wt) strain and its isogenic iglc or igld mutant with the vatpase at 15, 30 and 60 min post-infection and compared to L. pneumophila strain AA100 and L. monocytogenes (Lm) controls. Analyses were based on examination of 100 infected cells from three different coverslips. The results shown are representative of three independent experiments and the error bars represent standard deviations of triplicate samples. Fig. 3. Representative confocal microscopy images of transient co-localization of the FCPs with the LysoTracker. Co-localization of the FCP with the LysoTracker DND-99 by the GFPexpressing wild type (wt) strain and its isogenic iglc or igld mutant, or by the L. monocytogenes (Lm) positive control at 15, 30 and 60 min post-infection. In some infections the cells were 19
20 treated by Bafilomycin A1 (BFA). Uninfected (UI) cells were used as negative control while L. monocytogenes served as a positive control. The images are representatives of 100 infected cells examined from three different coverslips. The results shown are representative of three independent experiments. Fig. 4. F. tularensis resides transiently in acidified compartments within hmdms during early time points of infection. Quantification of co-localization of the LysoTracker DND-99 with the FCPs of the wild type (wt) strain and its isogenic iglc or igld mutant, or by the L. monocytogenes (Lm) positive control at 15, 30 and 60 min post-infection. In some infection the cells were treated by Bafilomycin A1 (BFA). Analyses were based on examination of 100 infected cells from three different coverslips. The results shown are representative of three independent experiments and the error bars represent standard deviations of triplicate samples. Fig. 5. Transient acquistion of the proton vatpase pump by the FCP during early stages of infection is essential for disruption of the FCP and bacterial escape into the macrophage cytosol. Representative confocal microscopy images of the phagosome integrity assay by determining differential binding of specific anti-bacterial antibodies loaded into the cytosol of live un-infected hmdms or infected by the wild type (wt) strain and its isogenic iglc or igld mutant, or by L. monocytogenes (Lm) that was used as a positive control. After 15 min of infection at an MOI 10, the cytosol of live hmdms was loaded with specific anti-bacterial Mabs (red) after the 15, 30, 60 min and 6h incubation follwing the 15-min synchronized infection by GFP-expressing bacteria. Inhibition of the vatpase pump was accomplished by pre-treating the cells for 1h with Bafilomycin A1 (BFA), which was maintained during infection. For the BFA reversal, the infected treated cells were incubated for 1h after removal of BFA then bacteria were 20
21 labled and examined after 6h of total incubation. The images are representatives of 100 infected cells examined from three different coverslips. The results shown are representative of three independent experiments. Fig. 6. Quantitative analyses for the role of the vatpase proton pump in escape of F. tularensis from the phagosome into the cytosol of hmdms. The fluorescen microscopy-based phagosome integrity assay was used to differentiate between vacuolar and cytosolic bacteria in untreated cells or cells treated with BFA and infected by the wild type (wt) strain and its isogenic iglc or igld mutant, or by the L. monocytogenes (Lm) positive control. The cells were examiend at 15, 30 and 60 min and 6h incubation follwing the 15-min synchronized infection. In some BFA-treated infected cells, BFA was removed and the cells were incubated for 1h to examine whether the effect of BFA treatment on blocking bacterial escape from the phagosome was reversible (BFA reversal). Quantification was based on examination of 100 infected cells from three different coverslips for accessibility of intracellular bacteria to the antibodies loaded into the macrophage cytosol. The error bars represent standard deviations of triplicate samples and the results shown are representative of three independent experiments. Fig. 7. Quantitative analyses of Rab7 recruitment ot the FCP within hmdms. Colocalization of the FCP with Rab7 was determined by labeling with anti-rab7 rabbit antisera at min post-infectin of hmdms at an MOI of 10. Quantification was based on examination of 100 infected cells from three different coverslips 21




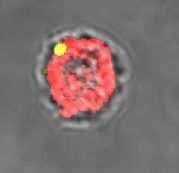





22 30 min wt igld iglc Lm GFP anti-vatpase Merge GFP anti-vatpase Merge wt igld 60 min iglc Lm AA100 Fig.1.
23 % colocalization min 30min 1h 0 wt igld iglc Lm AA100 Fig. 2










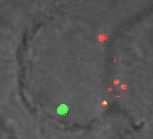








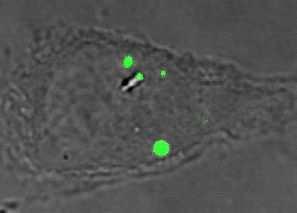

24 30 min GFP LysoTrackerDND Merge GFP LysoTrackerDND Merge GFP LysoTrackerDND Merge wt wt+bfa iglc wt+bfa iglc+bfa igld igld+bfa 60 min wt wt+bfa iglc iglc+bfa igld igld+bfa 30 min UI UI+BFA Lm Lm+BFA 60 min GFP LysoTrackerDND Merge UI UI+BFA Lm Lm+BFA Fig. 3
25 % colocalization wt wt+bfa igld igld+bfa iglc iglc+bfa Lm Lm+BFA 15min 30min 1h Fig. 4















26 30 min GFP mab Merge wt wt+bfa iglc iglc+bfa 60 min wt wt+bfa iglc iglc+bfa GFP mab Merge GFP mab Merge Controls No beads Triton BFA reversal Lm Lm+BFA igld+bfa igld igld Fig. 5
27 % cytoplasmic bacteria min 30min 1h 6h wt wt+bfa igld igld+bfa iglc iglc+bfa Lm Lm+BFA BFA reversal Fig. 6
28 % Colocalization with Rab WT IglC AA100 Strains 15 min 30 min 60 min Fig. 7
Received 11 February 2008/Returned for modification 10 March 2008/Accepted 13 March 2008
 INFECTION AND IMMUNITY, June 2008, p. 2671 2677 Vol. 76, No. 6 0019-9567/08/$08.00 0 doi:10.1128/iai.00185-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Acquisition of the
INFECTION AND IMMUNITY, June 2008, p. 2671 2677 Vol. 76, No. 6 0019-9567/08/$08.00 0 doi:10.1128/iai.00185-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Acquisition of the
Rapid Escape of the dot/icm Mutants of Legionella pneumophila into the Cytosol of Mammalian and Protozoan Cells
 INFECTION AND IMMUNITY, July 2007, p. 3290 3304 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00292-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Rapid Escape of the
INFECTION AND IMMUNITY, July 2007, p. 3290 3304 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00292-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Rapid Escape of the
Maturation of the Legionella pneumophila-containing Phagosome into a Phagolysosome within Gamma Interferon-Activated Macrophages
 INFECTION AND IMMUNITY, May 2005, p. 3166 3171 Vol. 73, No. 5 0019-9567/05/$08.00 0 doi:10.1128/iai.73.5.3166 3171.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Maturation
INFECTION AND IMMUNITY, May 2005, p. 3166 3171 Vol. 73, No. 5 0019-9567/05/$08.00 0 doi:10.1128/iai.73.5.3166 3171.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Maturation
Genetic Susceptibility and Caspase Activation in Mouse and Human Macrophages Are Distinct for Legionella longbeachae and L.
 INFECTION AND IMMUNITY, Apr. 2007, p. 1933 1945 Vol. 75, No. 4 0019-9567/07/$08.00 0 doi:10.1128/iai.00025-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Genetic Susceptibility
INFECTION AND IMMUNITY, Apr. 2007, p. 1933 1945 Vol. 75, No. 4 0019-9567/07/$08.00 0 doi:10.1128/iai.00025-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Genetic Susceptibility
Role of cholesterol of membrane microdomain in the uptake of Francisella tularensis by mouse macrophages
 223,,,,,,, 200433 : LVS pfnltp6 gro-gfp LVS ;, -1 Alexa 594 ; Z LVS ; - -,,,, -1 ;, -1 : ; ; ; -1; Role of cholesterol of membrane microdomain in the uptake of Francisella tularensis by mouse macrophages
223,,,,,,, 200433 : LVS pfnltp6 gro-gfp LVS ;, -1 Alexa 594 ; Z LVS ; - -,,,, -1 ;, -1 : ; ; ; -1; Role of cholesterol of membrane microdomain in the uptake of Francisella tularensis by mouse macrophages
Legionella pneumophila: an intracellular pathogen of phagocytes Prof. Craig Roy
 an intracellular pathogen of phagocytes Section of Microbial Pathogenesis, Yale University School of Medicine 1 Legionella pneumophila Gram-negative bacterium Facultative intracellular pathogen Protozoa
an intracellular pathogen of phagocytes Section of Microbial Pathogenesis, Yale University School of Medicine 1 Legionella pneumophila Gram-negative bacterium Facultative intracellular pathogen Protozoa
THE EFFECT OF INTERFERON-γ AND LIPOPOLYSACCHARIDE ON THE GROWTH OF FRANCISELLA TULARENSIS LVS IN MURINE MACROPHAGE-LIKE CELL LINE J774
 ORIGINAL ARTICLE THE EFFECT OF INTERFERON-γ AND LIPOPOLYSACCHARIDE ON THE GROWTH OF FRANCISELLA TULARENSIS LVS IN MURINE MACROPHAGE-LIKE CELL LINE J774 Monika Holická, Jakub Novosad, Martina Loudová, Manuela
ORIGINAL ARTICLE THE EFFECT OF INTERFERON-γ AND LIPOPOLYSACCHARIDE ON THE GROWTH OF FRANCISELLA TULARENSIS LVS IN MURINE MACROPHAGE-LIKE CELL LINE J774 Monika Holická, Jakub Novosad, Martina Loudová, Manuela
and human macrophages is distinct from L. pneumophila ACCEPTED Department of Microbiology, Immunology, University of Louisville College of Medicine,
 IAI Accepts, published online ahead of print on 29 January 2007 Infect. Immun. doi:10.1128/iai.00025-07 Copyright 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
IAI Accepts, published online ahead of print on 29 January 2007 Infect. Immun. doi:10.1128/iai.00025-07 Copyright 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
General aspects of this review - specific examples were addressed in class.
 General aspects of this review - specific examples were addressed in class. 1 Exam 1 Lecture 2: Discussed intracellular killing mechanisms Important maturation steps Rapid development into a microbicidal
General aspects of this review - specific examples were addressed in class. 1 Exam 1 Lecture 2: Discussed intracellular killing mechanisms Important maturation steps Rapid development into a microbicidal
Received 29 July 2009/Accepted 29 September 2009
 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 2009, p. 7488 7500 Vol. 75, No. 23 0099-2240/09/$12.00 doi:10.1128/aem.01829-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Francisella
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 2009, p. 7488 7500 Vol. 75, No. 23 0099-2240/09/$12.00 doi:10.1128/aem.01829-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Francisella
THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS
 Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Role of Listeriolysin O in Cell-to-Cell Spread of Listeria monocytogenes
 INFECTION AND IMMUNITY, Feb. 2000, p. 999 1003 Vol. 68, No. 2 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Role of Listeriolysin O in Cell-to-Cell Spread
INFECTION AND IMMUNITY, Feb. 2000, p. 999 1003 Vol. 68, No. 2 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Role of Listeriolysin O in Cell-to-Cell Spread
Lysosomes, Peroxisomes and Centrioles. Hüseyin Çağsın
 Lysosomes, Peroxisomes and Centrioles Hüseyin Çağsın Lysosomes Outline Endosomes Molecule transport to the lysosomes Endocytosis Exocytosis Autophagy Vacuoles Peroxisomes Centrioles Lysosomes Lysosomes
Lysosomes, Peroxisomes and Centrioles Hüseyin Çağsın Lysosomes Outline Endosomes Molecule transport to the lysosomes Endocytosis Exocytosis Autophagy Vacuoles Peroxisomes Centrioles Lysosomes Lysosomes
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Vesicle Transport. Vesicle pathway: many compartments, interconnected by trafficking routes 3/17/14
 Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
SUPPLEMENTARY INFORMATION In format provided by JAATTELA (NOVEMBER 2005)
") Box S1: Methods for studying lysosomal function and integrity Volume and distribution of the acidic compartment. Acridine orange is a metachromatic fluorochrome and a weak base that accumulates in the
Box S1: Methods for studying lysosomal function and integrity Volume and distribution of the acidic compartment. Acridine orange is a metachromatic fluorochrome and a weak base that accumulates in the
Current status of vaccine development for tularemia preparedness
 Review article CLINICAL EXPERIMENTAL VACCINE RESEARCH Current status of vaccine development for tularemia preparedness Clin Exp Vaccine Res 2013;2:34-39 pissn 2287-3651 eissn 2287-366X Kee-Jong Hong 1,
Review article CLINICAL EXPERIMENTAL VACCINE RESEARCH Current status of vaccine development for tularemia preparedness Clin Exp Vaccine Res 2013;2:34-39 pissn 2287-3651 eissn 2287-366X Kee-Jong Hong 1,
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Intracellular survival mechanisms of Francisella tularensis, a stealth pathogen
 Microbes and Infection 8 (2006) 561 567 www.elsevier.com/locate/micinf Review Intracellular survival mechanisms of Francisella tularensis, a stealth pathogen Anders Sjöstedt * Department of Clinical Microbiology,
Microbes and Infection 8 (2006) 561 567 www.elsevier.com/locate/micinf Review Intracellular survival mechanisms of Francisella tularensis, a stealth pathogen Anders Sjöstedt * Department of Clinical Microbiology,
BACTERIAL PATHOGENESIS
 BACTERIAL PATHOGENESIS A pathogen is a microorganism that is able to cause disease. Pathogenicity is the ability to produce disease in a host organism. Virulence a term which refers to the degree of pathogenicity
BACTERIAL PATHOGENESIS A pathogen is a microorganism that is able to cause disease. Pathogenicity is the ability to produce disease in a host organism. Virulence a term which refers to the degree of pathogenicity
Host-Pathogen Interactions in Tuberculosis
 Host-Pathogen Interactions in Tuberculosis CNRS - Toulouse, France My presentation will focus on host-cell pathogen interactions in tuberculosis. However, I would first like offer a brief introduction
Host-Pathogen Interactions in Tuberculosis CNRS - Toulouse, France My presentation will focus on host-cell pathogen interactions in tuberculosis. However, I would first like offer a brief introduction
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Croatian International Publications > Croat Med J. 2008;49:450-4
 Croatian International Publications > Croat Med J. 2008;49:450-4 Šantić M, Asare R, Škrobonja I, Jones S, Abu Kwaik Y. Acquisition of the vacuolar ATPase proton pump and phagosome acidification are essential
Croatian International Publications > Croat Med J. 2008;49:450-4 Šantić M, Asare R, Škrobonja I, Jones S, Abu Kwaik Y. Acquisition of the vacuolar ATPase proton pump and phagosome acidification are essential
Evidence that Dot-dependent and -independent factors isolate the Legionella pneumophila phagosome from the endocytic network in mouse macrophages
 Cellular Microbiology (2001) 3(2), 99±114 Evidence that Dot-dependent and -independent factors isolate the Legionella pneumophila phagosome from the endocytic network in mouse macrophages Amrita D. Joshi,
Cellular Microbiology (2001) 3(2), 99±114 Evidence that Dot-dependent and -independent factors isolate the Legionella pneumophila phagosome from the endocytic network in mouse macrophages Amrita D. Joshi,
BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented
 Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Legionella pneumophila Is Internalized by a Macropinocytotic Uptake Pathway Controlled by the Dot/Icm System and the Mouse Lgn1 Locus
 Legionella pneumophila Is Internalized by a Macropinocytotic Uptake Pathway Controlled by the Dot/Icm System and the Mouse Lgn1 Locus Masahisa Watarai, 1, 2 Isabelle Derre, 2 James Kirby, 2 Joseph D. Growney,
Legionella pneumophila Is Internalized by a Macropinocytotic Uptake Pathway Controlled by the Dot/Icm System and the Mouse Lgn1 Locus Masahisa Watarai, 1, 2 Isabelle Derre, 2 James Kirby, 2 Joseph D. Growney,
RESEARCH ARTICLE. Lewis G. Tilney 1 *, Omar S. Harb 1, Patricia S. Connelly 1, Camenzind G. Robinson 2, and Craig R. Roy 2 SUMMARY
 RESEARCH ARTICLE 4637 How the parasitic bacterium Legionella pneumophila modifies its phagosome and transforms it into rough ER: implications for conversion of plasma membrane to the ER membrane Lewis
RESEARCH ARTICLE 4637 How the parasitic bacterium Legionella pneumophila modifies its phagosome and transforms it into rough ER: implications for conversion of plasma membrane to the ER membrane Lewis
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or
 Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
IP: anti-gfp VPS29-GFP. IP: anti-vps26. IP: anti-gfp - + +
 FAM21 Strump. WASH1 IP: anti- 1 2 3 4 5 6 FAM21 Strump. FKBP IP: anti-gfp VPS29- GFP GFP-FAM21 tail H H/P P H H/P P c FAM21 FKBP Strump. VPS29-GFP IP: anti-gfp 1 2 3 FKBP VPS VPS VPS VPS29 1 = VPS29-GFP
FAM21 Strump. WASH1 IP: anti- 1 2 3 4 5 6 FAM21 Strump. FKBP IP: anti-gfp VPS29- GFP GFP-FAM21 tail H H/P P H H/P P c FAM21 FKBP Strump. VPS29-GFP IP: anti-gfp 1 2 3 FKBP VPS VPS VPS VPS29 1 = VPS29-GFP
Mycobacterium tuberculosis and Legionella pneumophila Phagosomes Exhibit Arrested Maturation despite Acquisition of Rab7
 INFECTION AND IMMUNITY, Sept. 2000, p. 5154 5166 Vol. 68, No. 9 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Mycobacterium tuberculosis and Legionella pneumophila
INFECTION AND IMMUNITY, Sept. 2000, p. 5154 5166 Vol. 68, No. 9 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Mycobacterium tuberculosis and Legionella pneumophila
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1. (A) Left, western blot analysis of ISGylated proteins in Jurkat T cells treated with 1000U ml -1 IFN for 16h (IFN) or left untreated (CONT); right, western
SUPPLEMENTARY FIGURES Supplementary Figure 1. (A) Left, western blot analysis of ISGylated proteins in Jurkat T cells treated with 1000U ml -1 IFN for 16h (IFN) or left untreated (CONT); right, western
Phagocytosis: An Evolutionarily Conserved Mechanism to Remove Apoptotic Bodies and Microbial Pathogens
 Phagocytosis of IgG-coated Targets by s Phagocytosis: An Evolutionarily Conserved Mechanism to Remove Apoptotic Bodies and Microbial s 3 min 10 min Mast Cells Can Phagocytose Too! Extension of an F-actin-rich
Phagocytosis of IgG-coated Targets by s Phagocytosis: An Evolutionarily Conserved Mechanism to Remove Apoptotic Bodies and Microbial s 3 min 10 min Mast Cells Can Phagocytose Too! Extension of an F-actin-rich
Francisella tularensis. Patricia Bolivar MS., CLS, PHM
 Francisella tularensis Patricia Bolivar MS., CLS, PHM Case A 42 year old male hunter presents with a painful, purulent conjunctivitis. Ulcerations were present on the conjunctiva. Cervical lymphadenopathy
Francisella tularensis Patricia Bolivar MS., CLS, PHM Case A 42 year old male hunter presents with a painful, purulent conjunctivitis. Ulcerations were present on the conjunctiva. Cervical lymphadenopathy
Intracellular life of Francisella and Legionella within amoebae cells
 Mini-review/Kratki pregledni članak Intracellular life of Francisella and Legionella within amoebae cells Unutarstanični život bakterija Francisella i Legionella u amebama Mateja Ožanič, Valentina Marečić,
Mini-review/Kratki pregledni članak Intracellular life of Francisella and Legionella within amoebae cells Unutarstanični život bakterija Francisella i Legionella u amebama Mateja Ožanič, Valentina Marečić,
Supplemental Information. Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection. Cell Host & Microbe, Volume 15
 Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Cell Host & Microbe, Volume 15 Supplemental Information Gut Microbiota Promotes Hematopoiesis to Control Bacterial Infection Arya Khosravi, Alberto Yáñez, Jeremy G. Price, Andrew Chow, Miriam Merad, Helen
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson
 Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
Chapter 6. Antigen Presentation to T lymphocytes
 Chapter 6 Antigen Presentation to T lymphocytes Generation of T-cell Receptor Ligands T cells only recognize Ags displayed on cell surfaces These Ags may be derived from pathogens that replicate within
Chapter 6 Antigen Presentation to T lymphocytes Generation of T-cell Receptor Ligands T cells only recognize Ags displayed on cell surfaces These Ags may be derived from pathogens that replicate within
JCB. The road less traveled: transport of Legionella to the endoplasmic reticulum. Mini-Review. Craig R. Roy 1 and Lewis G.
 JCB Published Online: 29 July, 2002 Supp Info: http://doi.org/10.1083/jcb.200205011 Downloaded from jcb.rupress.org on November 6, 2018 Mini-Review The road less traveled: transport of Legionella to the
JCB Published Online: 29 July, 2002 Supp Info: http://doi.org/10.1083/jcb.200205011 Downloaded from jcb.rupress.org on November 6, 2018 Mini-Review The road less traveled: transport of Legionella to the
From the Laboratory of Cellular Physiology and Immunology, The Rockefeller University, New York 10021
 Published Online: 1 February, 1981 Supp Info: http://doi.org/10.1084/jem.153.2.398 Downloaded from jem.rupress.org on April 7, 2018 INTERACTION OF THE LEGIONNAIRES' DISEASE BACTERIUM (LEGIONELLA PNEUMOPHILA)
Published Online: 1 February, 1981 Supp Info: http://doi.org/10.1084/jem.153.2.398 Downloaded from jem.rupress.org on April 7, 2018 INTERACTION OF THE LEGIONNAIRES' DISEASE BACTERIUM (LEGIONELLA PNEUMOPHILA)
Supplemental information contains 7 movies and 4 supplemental Figures
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
6. TNF-α regulates oxidative stress, mitochondrial function and autophagy in neuronal cells
 6. TNF-α regulates oxidative stress, mitochondrial function and autophagy in neuronal cells 6.1 TNF-α induces mitochondrial oxidative stress in SH-SY5Y cells. The dysregulation of mitochondria and oxidative
6. TNF-α regulates oxidative stress, mitochondrial function and autophagy in neuronal cells 6.1 TNF-α induces mitochondrial oxidative stress in SH-SY5Y cells. The dysregulation of mitochondria and oxidative
INFECTION AND IMMUNITY, May 2000, p Vol. 68, No. 5. Copyright 2000, American Society for Microbiology. All Rights Reserved.
 INFECTION AND IMMUNITY, May 2000, p. 2671 2684 Vol. 68, No. 5 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Deviant Expression of Rab5 on Phagosomes Containing
INFECTION AND IMMUNITY, May 2000, p. 2671 2684 Vol. 68, No. 5 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Deviant Expression of Rab5 on Phagosomes Containing
J. Cell Sci. 129: doi: /jcs : Supplementary information
 Movie 1. AgLDL is contained in small sub-regions of the lysosomal synapse that are acidic. J774 cells were incubated with agldl dual labeled with a ph sensitive and a ph insensitive fluorophore for 1 hr.
Movie 1. AgLDL is contained in small sub-regions of the lysosomal synapse that are acidic. J774 cells were incubated with agldl dual labeled with a ph sensitive and a ph insensitive fluorophore for 1 hr.
BEH.462/3.962J Molecular Principles of Biomaterials Spring 2003
 Lecture 16: Intracellular drug delivery Last time: Today: nano- and micro-particle drug carriers Delivery to tissues from systemic circulation Intracellular drug delivery Reading: A.S. Hoffman et al.,
Lecture 16: Intracellular drug delivery Last time: Today: nano- and micro-particle drug carriers Delivery to tissues from systemic circulation Intracellular drug delivery Reading: A.S. Hoffman et al.,
General information. Cell mediated immunity. 455 LSA, Tuesday 11 to noon. Anytime after class.
 General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
Viral Genetics. BIT 220 Chapter 16
 Viral Genetics BIT 220 Chapter 16 Details of the Virus Classified According to a. DNA or RNA b. Enveloped or Non-Enveloped c. Single-stranded or double-stranded Viruses contain only a few genes Reverse
Viral Genetics BIT 220 Chapter 16 Details of the Virus Classified According to a. DNA or RNA b. Enveloped or Non-Enveloped c. Single-stranded or double-stranded Viruses contain only a few genes Reverse
Anti-apoptotic signalling by the Dot/Icm secretion system of L. pneumophila
 Cellular Microbiology (27) 9(1), 246 264 doi:1.1111/j.1462-5822.26.785.x First published online 15 August 26 Anti-apoptotic signalling by the Dot/Icm secretion system of L. pneumophila Alaeddin Abu-Zant,
Cellular Microbiology (27) 9(1), 246 264 doi:1.1111/j.1462-5822.26.785.x First published online 15 August 26 Anti-apoptotic signalling by the Dot/Icm secretion system of L. pneumophila Alaeddin Abu-Zant,
/01/$ DOI: /IAI Received 21 July 2000/Returned for modification 26 September 2000/Accepted 13 October 2000
 INFECTION AND IMMUNITY, Jan. 2001, p. 194 203 Vol. 69, No. 1 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.1.194 203.2001 Susceptibility to Secondary Francisella tularensis Live Vaccine Strain Infection in
INFECTION AND IMMUNITY, Jan. 2001, p. 194 203 Vol. 69, No. 1 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.1.194 203.2001 Susceptibility to Secondary Francisella tularensis Live Vaccine Strain Infection in
Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells.
 Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells. a. Schematic of the V-ATPase proton pump macro-complex structure. The V1 complex is composed of
Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells. a. Schematic of the V-ATPase proton pump macro-complex structure. The V1 complex is composed of
lysosomes Ingested materials Defective cell components Degrades macromolecules of all types:
 lysosomes Digests Ingested materials Defective cell components Degrades macromolecules of all types: Proteins Nucleic acids Carbohydrates Lipids Single membrane bound vesicle, contains up to 50 digestive
lysosomes Digests Ingested materials Defective cell components Degrades macromolecules of all types: Proteins Nucleic acids Carbohydrates Lipids Single membrane bound vesicle, contains up to 50 digestive
A Bacterial Virulence Protein Suppresses Host Innate Immunity to Cause Plant Disease
 A Bacterial Virulence Protein Suppresses Host Innate Immunity to Cause Plant Disease Nomura, K., Debroy, S., Lee, Y.H., Pumplin, N., Jones, J., and He, S.Y. (2006). Science 313, 220-223. Presented by:
A Bacterial Virulence Protein Suppresses Host Innate Immunity to Cause Plant Disease Nomura, K., Debroy, S., Lee, Y.H., Pumplin, N., Jones, J., and He, S.Y. (2006). Science 313, 220-223. Presented by:
genome edited transient transfection, CMV promoter
 Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
Differential infection of mononuclear phagocytes by Francisella tularensis: role of the macrophage mannose receptor
 Differential infection of mononuclear phagocytes by Francisella tularensis: role of the macrophage mannose receptor Grant S. Schulert*, and Lee-Ann H. Allen*,,,1 *Inflammation Program and Departments of
Differential infection of mononuclear phagocytes by Francisella tularensis: role of the macrophage mannose receptor Grant S. Schulert*, and Lee-Ann H. Allen*,,,1 *Inflammation Program and Departments of
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
Rhesus Rotavirus Entry into a Polarized Epithelium Is Endocytosis Dependent and Involves Sequential VP4 Conformational Changes
 JOURNAL OF VIROLOGY, Mar. 2011, p. 2492 2503 Vol. 85, No. 6 0022-538X/11/$12.00 doi:10.1128/jvi.02082-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. Rhesus Rotavirus Entry into
JOURNAL OF VIROLOGY, Mar. 2011, p. 2492 2503 Vol. 85, No. 6 0022-538X/11/$12.00 doi:10.1128/jvi.02082-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. Rhesus Rotavirus Entry into
The Involvement of IL-17A in the Murine Response to Sub-Lethal Inhalational Infection with Francisella tularensis
 The Involvement of IL-17A in the Murine Response to Sub-Lethal Inhalational Infection with Francisella tularensis Gal Markel 1, Erez Bar-Haim 1, Eran Zahavy 2, Hila Cohen 1, Ofer Cohen 1, Avigdor Shafferman
The Involvement of IL-17A in the Murine Response to Sub-Lethal Inhalational Infection with Francisella tularensis Gal Markel 1, Erez Bar-Haim 1, Eran Zahavy 2, Hila Cohen 1, Ofer Cohen 1, Avigdor Shafferman
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Antigen presenting cells
 Antigen recognition by T and B cells - T and B cells exhibit fundamental differences in antigen recognition - B cells recognize antigen free in solution (native antigen). - T cells recognize antigen after
Antigen recognition by T and B cells - T and B cells exhibit fundamental differences in antigen recognition - B cells recognize antigen free in solution (native antigen). - T cells recognize antigen after
16 Innate Immunity: M I C R O B I O L O G Y. Nonspecific Defenses of the Host. a n i n t r o d u c t i o n
 ninth edition TORTORA FUNKE CASE M I C R O B I O L O G Y a n i n t r o d u c t i o n 16 Innate Immunity: Nonspecific Defenses of the Host PowerPoint Lecture Slide Presentation prepared by Christine L.
ninth edition TORTORA FUNKE CASE M I C R O B I O L O G Y a n i n t r o d u c t i o n 16 Innate Immunity: Nonspecific Defenses of the Host PowerPoint Lecture Slide Presentation prepared by Christine L.
Chapter 3. Expression of α5-megfp in Mouse Cortical Neurons. on the β subunit. Signal sequences in the M3-M4 loop of β nachrs bind protein factors to
 22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
PhysicsAndMathsTutor.com. Question Number. 1. prevents viruses attaching to {uninfected / eq} host cells / eq ; 2. by binding to receptors / eq ;
 1(a) 1. prevents viruses attaching to {uninfected / eq} host cells / eq ; 2. by binding to receptors / eq ; 3. (therefore) preventing virus from entering cell / eq ; 4. (therefore) viruses cannot replicate
1(a) 1. prevents viruses attaching to {uninfected / eq} host cells / eq ; 2. by binding to receptors / eq ; 3. (therefore) preventing virus from entering cell / eq ; 4. (therefore) viruses cannot replicate
Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death
 www.impactjournals.com/oncotarget/ Oncotarget, Supplementary Materials 2016 Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death Supplementary
www.impactjournals.com/oncotarget/ Oncotarget, Supplementary Materials 2016 Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death Supplementary
Supplementary table and figures
 3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
Supplemental Figures:
 Supplemental Figures: Figure 1: Intracellular distribution of VWF by electron microscopy in human endothelial cells. a) Immunogold labeling of LC3 demonstrating an LC3-positive autophagosome (white arrow)
Supplemental Figures: Figure 1: Intracellular distribution of VWF by electron microscopy in human endothelial cells. a) Immunogold labeling of LC3 demonstrating an LC3-positive autophagosome (white arrow)
Laboratory 10 Factors Controlling Phagocytosis in Tetrahymena,
 BIO354: Cell Biology Laboratory 1 Laboratory 1 Factors Controlling Phagocytosis in Tetrahymena, I. Introduction A characteristic feature of all eukaryotic cells is the ability to pinch off portions of
BIO354: Cell Biology Laboratory 1 Laboratory 1 Factors Controlling Phagocytosis in Tetrahymena, I. Introduction A characteristic feature of all eukaryotic cells is the ability to pinch off portions of
Structure. Lysosomes are membrane-enclosed organelles. Hydrolytic enzymes. Variable in size & shape need
 Lysosomes Structure Lysosomes are membrane-enclosed organelles Hydrolytic enzymes Variable in size & shape need Degrade material taken up from outside and inside the cell Variable in size and shape Lysosomal
Lysosomes Structure Lysosomes are membrane-enclosed organelles Hydrolytic enzymes Variable in size & shape need Degrade material taken up from outside and inside the cell Variable in size and shape Lysosomal
CytoPainter Lysosomal Staining Kit - Blue Fluorescence
 ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
Characterization of Amoebae Interactions with Four Non-Pneumophila Legionella Species
 Clemson University TigerPrints All Theses Theses 5-2016 Characterization of Amoebae Interactions with Four Non-Pneumophila Legionella Species Allison Margaret Palmer Clemson University, ampalme@g.clemson.edu
Clemson University TigerPrints All Theses Theses 5-2016 Characterization of Amoebae Interactions with Four Non-Pneumophila Legionella Species Allison Margaret Palmer Clemson University, ampalme@g.clemson.edu
4b. Innate (nonspecific) Immunity
 Immunity") 4b. Innate (nonspecific) Immunity Chapter 16: Innate (nonspecific) Immunity! Some terms:! Susceptibility: Lack of immunity to a disease.! Immunity: Ability to ward off disease.! Innate immunity: Defenses
4b. Innate (nonspecific) Immunity Chapter 16: Innate (nonspecific) Immunity! Some terms:! Susceptibility: Lack of immunity to a disease.! Immunity: Ability to ward off disease.! Innate immunity: Defenses
Summary and Discussion antigen presentation
 Summary and Discussion antigen presentation 247 248 Summary & Discussion Summary and discussion: antigen presentation For a cell to communicate information about its internal health and status to the immune
Summary and Discussion antigen presentation 247 248 Summary & Discussion Summary and discussion: antigen presentation For a cell to communicate information about its internal health and status to the immune
Department of Microbial Pathogenesis, Yale University School of Medicine, New Haven, Connecticut, United States of America
 Effector Protein Translocation by the Coxiella burnetii Dot/Icm Type IV Secretion System Requires Endocytic Maturation of the Pathogen-Occupied Vacuole Hayley J. Newton*, Justin A. McDonough, Craig R.
Effector Protein Translocation by the Coxiella burnetii Dot/Icm Type IV Secretion System Requires Endocytic Maturation of the Pathogen-Occupied Vacuole Hayley J. Newton*, Justin A. McDonough, Craig R.
Protein Trafficking in the Secretory and Endocytic Pathways
 Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Modulation of Rab5 and Rab7 Recruitment to Phagosomes by Phosphatidylinositol 3-Kinase
 MOLECULAR AND CELLULAR BIOLOGY, Apr. 2003, p. 2501 2514 Vol. 23, No. 7 0270-7306/03/$08.00 0 DOI: 10.1128/MCB.23.7.2501 2514.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved.
MOLECULAR AND CELLULAR BIOLOGY, Apr. 2003, p. 2501 2514 Vol. 23, No. 7 0270-7306/03/$08.00 0 DOI: 10.1128/MCB.23.7.2501 2514.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved.
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Epithelial cell death is an important contributor to oxidant-mediated acute lung injury SUPPORTING INFORMATION 60611, USA
 Epithelial cell death is an important contributor to oxidant-mediated acute lung injury SUPPORTING INFORMATION G.R. Scott Budinger 1,2 *, Gökhan M. Mutlu 1 *, Daniela Urich 2, Saul Soberanes 1, Leonard
Epithelial cell death is an important contributor to oxidant-mediated acute lung injury SUPPORTING INFORMATION G.R. Scott Budinger 1,2 *, Gökhan M. Mutlu 1 *, Daniela Urich 2, Saul Soberanes 1, Leonard
Chlamydia Species-Dependent Differences in the Growth Requirement for Lysosomes
 Chlamydia Species-Dependent Differences in the Growth Requirement for Lysosomes Scot P. Ouellette 1, Frank C. Dorsey 2, Simon Moshiach 3, John L. Cleveland 2, Rey A. Carabeo 1 * 1 Centre for Molecular
Chlamydia Species-Dependent Differences in the Growth Requirement for Lysosomes Scot P. Ouellette 1, Frank C. Dorsey 2, Simon Moshiach 3, John L. Cleveland 2, Rey A. Carabeo 1 * 1 Centre for Molecular
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Significance of the MHC
 CHAPTER 8 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
CHAPTER 8 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
Serafino et al. Thymosin α1 activates complement receptor-mediated phagocytosis in human monocyte-derived macrophages. SUPPLEMENTARY FIGURES
 Supplementary Fig. S1. Evaluation of the purity and maturation of macrophage cultures tested by flow cytometry. The lymphocytic/monocytic cellular fraction was isolated from buffy coats of healthy donors
Supplementary Fig. S1. Evaluation of the purity and maturation of macrophage cultures tested by flow cytometry. The lymphocytic/monocytic cellular fraction was isolated from buffy coats of healthy donors
Project report October 2012 March 2013 CRF fellow: Principal Investigator: Project title:
 Project report October 2012 March 2013 CRF fellow: Gennaro Napolitano Principal Investigator: Sergio Daniel Catz Project title: Small molecule regulators of vesicular trafficking to enhance lysosomal exocytosis
Project report October 2012 March 2013 CRF fellow: Gennaro Napolitano Principal Investigator: Sergio Daniel Catz Project title: Small molecule regulators of vesicular trafficking to enhance lysosomal exocytosis
Bacterial outer membrane vesicles (OMVs)
") Bacterial outer membrane vesicles (OMVs) PhD student: Wei TAN Supervisor: Prof. Guoping Zhao Deparment of Microbiology Faculty of Medicine The Chinese University of Hong Kong Date: 1 Dec 2016 Outline Introduction
Bacterial outer membrane vesicles (OMVs) PhD student: Wei TAN Supervisor: Prof. Guoping Zhao Deparment of Microbiology Faculty of Medicine The Chinese University of Hong Kong Date: 1 Dec 2016 Outline Introduction
Rapid parallel measurements of macroautophagy and mitophagy in
 Supplemental Figures Rapid parallel measurements of macroautophagy and mitophagy in mammalian cells using a single fluorescent biosensor Sargsyan A, Cai J, Fandino LB, Labasky ME, Forostyan T, Colosimo
Supplemental Figures Rapid parallel measurements of macroautophagy and mitophagy in mammalian cells using a single fluorescent biosensor Sargsyan A, Cai J, Fandino LB, Labasky ME, Forostyan T, Colosimo
IFNγ Inhibits the Cytosolic Replication of Shigella flexneri via the Cytoplasmic RNA Sensor RIG-I
 IFNγ Inhibits the Cytosolic Replication of Shigella flexneri via the Cytoplasmic RNA Sensor RIG-I The Harvard community has made this article openly available. Please share how this access benefits you.
IFNγ Inhibits the Cytosolic Replication of Shigella flexneri via the Cytoplasmic RNA Sensor RIG-I The Harvard community has made this article openly available. Please share how this access benefits you.
Effect of Vaccine, Route, and Schedule on Antibody
 APPUED MICROBIOLOGY, Mar. 1969, p. 355-359 Copyright 1969 American Society for Microbiology Vol. 17, No. 3 Printed in U.S.A. Effect of Vaccine, Route, and Schedule on Antibody Response of Rabbits to Pasteurella
APPUED MICROBIOLOGY, Mar. 1969, p. 355-359 Copyright 1969 American Society for Microbiology Vol. 17, No. 3 Printed in U.S.A. Effect of Vaccine, Route, and Schedule on Antibody Response of Rabbits to Pasteurella
Cryptococcus neoformans Is a Facultative Intracellular Pathogen in Murine Pulmonary Infection
 INFECTION AND IMMUNITY, July 2000, p. 4225 4237 Vol. 68, No. 7 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Cryptococcus neoformans Is a Facultative Intracellular
INFECTION AND IMMUNITY, July 2000, p. 4225 4237 Vol. 68, No. 7 0019-9567/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Cryptococcus neoformans Is a Facultative Intracellular
Fluorescence Microscopy
 Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Supplementary Information. Tissue-wide immunity against Leishmania. through collective production of nitric oxide
 Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
ACCEPTED CFU * * CFU CCM PVT. VacA (+) VacA (-) Time (h) Terebiznik et al. Fig 1
 VacA (-) Time (h) Terebiznik et al. Fig 1") IAI Accepts, published online ahead of print on 25 September 2006 Infect. Immun. doi:10.1128/iai.01085-06 Copyright 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
IAI Accepts, published online ahead of print on 25 September 2006 Infect. Immun. doi:10.1128/iai.01085-06 Copyright 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
A Tour of the Cell. Chapter 6. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 6 A Tour of the Cell PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 6 A Tour of the Cell PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Microbiology of Atypical Pneumonia. Dr. Mohamed Medhat Ali
 Microbiology of Atypical Pneumonia Dr. Mohamed Medhat Ali Pneumonia P n e u m o n i a i s a n infection of the lungs that can be caused by viruses, bacteria, and fungi. Atypical! Pneumonia Symptoms. X-ray
Microbiology of Atypical Pneumonia Dr. Mohamed Medhat Ali Pneumonia P n e u m o n i a i s a n infection of the lungs that can be caused by viruses, bacteria, and fungi. Atypical! Pneumonia Symptoms. X-ray
Dr. Ahmed K. Ali Attachment and entry of viruses into cells
 Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
In vitro and Intracellular Activities of Peptide Deformylase. Inhibitor GSK against Legionella pneumophila Isolates
 AAC Accepts, published online ahead of print on 27 October 2014 Antimicrob. Agents Chemother. doi:10.1128/aac.04006-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 In vitro
AAC Accepts, published online ahead of print on 27 October 2014 Antimicrob. Agents Chemother. doi:10.1128/aac.04006-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 In vitro
Anti-infectious Immunity
 Anti-infectious Immunity innate immunity barrier structures Secretory molecules Phagocytes NK cells Anatomical barriers 1. Skin and mucosa barrier 2.hemo-Spinal Fluid barrier 3. placental barrier Phagocytic
Anti-infectious Immunity innate immunity barrier structures Secretory molecules Phagocytes NK cells Anatomical barriers 1. Skin and mucosa barrier 2.hemo-Spinal Fluid barrier 3. placental barrier Phagocytic
Tobias Welte Diagnosis and treatment of community acquired pneumonia the German perspective Summary
 Tobias Welte Diagnosis and treatment of community acquired pneumonia the German perspective Current concepts of diagnosis and treatment of CAP are risk stratified and adapted to the national resistances
Tobias Welte Diagnosis and treatment of community acquired pneumonia the German perspective Current concepts of diagnosis and treatment of CAP are risk stratified and adapted to the national resistances
endomembrane system internal membranes origins transport of proteins chapter 15 endomembrane system
 endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
FORMATION OF A NOVEL PHAGOSOME BY THE LEGIONNAIRES' DISEASE BACTERIUM (LEGIONELLA
 Published Online: 1 October, 1983 Supp Info: http://doi.org/10.1084/jem.158.4.1319 Downloaded from jem.rupress.org on August 31, 2018 FORMATION OF A NOVEL PHAGOSOME BY THE LEGIONNAIRES' DISEASE BACTERIUM
Published Online: 1 October, 1983 Supp Info: http://doi.org/10.1084/jem.158.4.1319 Downloaded from jem.rupress.org on August 31, 2018 FORMATION OF A NOVEL PHAGOSOME BY THE LEGIONNAIRES' DISEASE BACTERIUM
MISSION: understanding the mechanisms of therapeutic strategies
 Telethon Institute of Genetics and Medicine MISSION: understanding the mechanisms of genetic diseases to develop preventive and therapeutic strategies G E N O T Y P E G E Researcher N 1 3 5 O T Y P E 8
Telethon Institute of Genetics and Medicine MISSION: understanding the mechanisms of genetic diseases to develop preventive and therapeutic strategies G E N O T Y P E G E Researcher N 1 3 5 O T Y P E 8
Supplemental Materials Molecular Biology of the Cell
 Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
Live cell imaging of trafficking of the chaperone complex vaccine to the ER. BMDCs were incubated with ER-Tracker Red (1 M) in staining solution for
 in staining solution for") Live cell imaging of trafficking of the chaperone complex vaccine to the ER. BMDCs were incubated with ER-Tracker Red (1 M) in staining solution for 15 min at 37 C and replaced with fresh complete medium.
Live cell imaging of trafficking of the chaperone complex vaccine to the ER. BMDCs were incubated with ER-Tracker Red (1 M) in staining solution for 15 min at 37 C and replaced with fresh complete medium.