Insulin signaling diverges into Akt dependent and independent signals to regulate the
|
|
|
- Lisa Edwards
- 5 years ago
- Views:
Transcription
1 Insulin signaling diverges into Akt dependent and independent signals to regulate the recruitment/docking and the fusion of GLUT4 vesicles to the plasma membrane. Eva Gonzalez and Timothy E. McGraw* Department of Biochemistry Weill Medical College of Cornell University 1300 York Ave. New York, NY * corresponding author. temcgraw@med.cornell.edu Tel: Fax: Running title: 6 Figures 1
2 Summary Insulin modulates glucose disposal in muscle and adipose tissue by regulating the cellular redistribution of the GLUT4 glucose transporter. Protein kinase Akt/PKB is a central mediator of insulin-regulated translocation of GLUT4; however, the GLUT4 trafficking step(s) regulated by Akt is not known. Here we use acute pharmacologic Akt inhibition to show that Akt is required for insulin-stimulated exocytosis of GLUT4 to the plasma membrane. Our data also suggest that the AS160 Rab GAP protein is not the only Akt target required for insulin-stimulated GLUT4 translocation. Using a total internal reflection microscopy assay to show that Akt activity is specifically required for an insulin-mediated pre-fusion step involving the recruitment and/or docking of GLUT4 vesicles to within 250 nm of the plasma membrane. Moreover, the insulinstimulated fusion of GLUT4 vesicles with the plasma membrane can occur independent of Akt activity, although based on inhibition by wortmannin, it is dependent on PI3-kinase activity. Hence, to achieve full redistribution of GLUT4 into the plasma membrane, insulin signaling bifurcates to independently regulate both fusion and a pre-fusion step(s). 2
3 Introduction Insulin regulates glucose uptake into muscle and fat cells by inducing the reversible redistribution of the GLUT4 glucose transporter from intracellular storage sites to the plasma membrane (Ishiki et al., 2005; Watson and Pessin, 2006). In unstimulated adipocytes GLUT4 is excluded from the plasma membrane (PM) by a dynamic process involving slow exocytosis and rapid endocytosis. Insulin induces a net redistribution of GLUT4 to the PM by accelerating GLUT4 exocytosis and inhibiting its endocytosis (e.g., (Govers et al., 2004; Karylowski et al., 2004; Martin et al., 2006). The exocytic steps regulated by insulin are not known and potentially include: stimulation of the formation of GLUT4 transport vesicles (Xu and Kandror, 2002); mobilization of a pool of GLUT4 vesicles (Bogan et al., 2003); increased movement of GLUT4 vesicles (Bose et al., 2002; Semiz et al., 2003); increased docking at the PM of GLUT4 vesicles and/or increased fusion of GLUT4 vesicles with the PM (Koumanov et al., 2005; Lizunov et al., 2005; van Dam et al., 2005). Substantial evidence supports the serine/threonine protein kinase Akt/PKB as a crucial regulator of insulin-mediated GLUT4 redistribution (for review see (Watson and Pessin, 2006). Overexpression of a constitutively active mutant of Akt leads to the translocation of GLUT4 to the PM (Kohn et al., 1996), and inhibition of Akt by interfering antibodies, substrate peptides, or overexpression of a dominant-negative Akt construct partially blocks insulin-induced GLUT4 translocation (Hill et al., 1999; Wang et al., 1999). Of the three mammalian Akt isoforms, Akt2 is strongly correlated with the regulation of glucose homeostasis. Targeted disruption of Akt2, but not Akt1 or Akt3, resulted in impaired insulin-regulated glucose uptake (Cho et al., 2001; Bae et al., 2003). 3
4 Despite advances in our understanding of GLUT4 intracellular trafficking and strong evidence of Akt being a crucial regulator of insulin-induced GLUT4 redistribution, the specific input of Akt signaling to GLUT4 trafficking is not known. Recently an allosteric Akt1 and Akt2 inhibitor (Akti-1/2) has been developed, this compound has been shown to inhibit Akt activation but not affect other AGC kinase family members (DeFeo-Jones et al., 2005; Zhao et al., 2005). Given the broad range of metabolic actions of Akt signaling, acute inhibition of insulin-induced Akt activation represents a useful tool to address the specific role of Akt signaling on GLUT4 trafficking. In this study we use acute pharmacological inhibition and shrna-mediated Akt knockdown to identify the steps in GLUT4 trafficking modulated by Akt signaling. We found that Akt is a crucial regulator of insulin-induced GLUT4 exocytosis, while insulin-regulated GLUT4 endocytosis is independent of Akt activation. Using total internal reflection fluorescence (TIRF) microscopy and a novel fusion assay, we show that along the exocytic pathway Akt regulates the insulin-mediated pre-fusion recruitment and/or docking of GLUT4 vesicles to the PM. In contrast, insulin-regulation of GLUT4 vesicle fusion with the PM is dependent on phosphatidylinositol 3 kinase (PI3-kinase) activity but independent of Akt activity. 4
5 Materials and Methods Ligands and Chemicals. Fluorescent secondary antibodies were purchased form Jackson Immunolabs, Inc. (West Grove, Pennsylvania). Antibodies against phospho-akt (Ser 473 ), Akt, Akt2, phospho-akt substrate (PAS), phosho-gsk3β (Ser 9 ) and ERK1/2 were from Cell Signaling Technologies (Beverly, MA). Antibodies against Akt1, cebpα, and PPARγ were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The GLUT4 antibody was a gift from Sam Cushman, NIH. The IRAP antibody was a gift from Susanna Keller, University of Virgina. Anti-AS160 antibody was a gift from Gustav Lienhard (Dartmouth Medical School). The isozyme selective Akt inhibitor AI1/2 (Akti-1/2) and the PI3-kinase inhibitor, wortmannin, were purchased from Calbiochem. Mouse anti-ha epitope monoclonal antibody was purified from ascites (Covance, Berkley, CA) using a protein G affinity column (Amersham Bioscience, Uppsala, Sweden). The concentration of HA.11 required to saturate the HA epitope of HA- GLUT4-GFP was determined for each preparation of antibody as described (Karylowski et al., 2004). Cell culture and electroporation. 3T3-L1 fibroblast were cultured, differentiated into adipocytes and electroporated as previously described (Zeigerer et al., 2002). The HA-GLUT4-GFP, IRAP- TR and TR constructs have been previously described (Lampson et al., 2000; Subtil et al., 2000). Stable expression of HA-GLUT4 was achieved using the retroviral expression vector pbabe- Puro (Shewan et al., 2000; Karylowski et al., 2004). Cells were maintained at 37 C in 5% CO 2 /air during growth and differentiation and for all incubations in kinetic experiments. All experiments were conducted in day 6 after differentiation. 5
6 Drug treatment, cell lysates and immunoblots. Cells were serum-starved for 2h in serum-free DMEM with 20 mm of sodium bicarbonate, 20 mm HEPES (ph7.2) (SF-DMEM) at 37 C in 5%CO 2 /air for 2h prior to all experiments. AI1/2 and wortmannin were dissolved in dimethyl sulfoxide and stored at -20 C. For immunoblots, cells were washed with 150 mm NaCl, 20 mm HEPES, 1 mm CaCl 2, 5 mm KCl, 1 mm MgCl 2 (ph 7.2) (Media 1) and lysed in Laemmli buffer. Cells were harvested by scraping and the lysates were sheared through a Q26G5/8 syringe. Proteins were resolved in SDS-PAGE, transferred to nitrocellulose membranes and probed with antibodies using protocols provided by the suppliers. shrna retroviral constructs. The psiren RetroQ system from BD Clontech Bioscience was used according to manufacture s instructions. The targeting sequences used for Akt1 and Akt2 were Akt1: 5 AGAACCGTGTCCTGCAGAA 3, Akt2: 5 CCATGAATGACTTCGATTA 3, the non specific sequence used as negative control for shrna activity and the targeting sequence used for AS160 have been described (Eguez et al., 2005). Hairpins were cloned into the psiren RetroQ vector and AmphoPak-293 cells (BD ClonTech Bioscience) were transfected with the retroviral constructs. Cell culture from the packaging cells were harvested and used to infect 3T3-L1 fibroblast. Infected cells were selected for growth in puromycin (Eguez et al., 2005). Kinetic studies. All live cell incubations were performed in SF-DMEM. Adipocytes were incubated in SF-DMEM for 2h previous to insulin stimulation. Cells were pre-incubated with the Akt inhibitor 1 µm AI1/2 or vehicle for 1h prior to insulin stimulation. For indirect immunofluorescence, cells were fixed with 3.7% formaldehyde, incubated with primary antibody 6
7 in phosphate-saline buffer (PBS) and 5% calf serum for 30 min at 37 C, washed and incubated with fluorescent secondary antibody in PBS-5% calf serum. PBS with 5% calf serum and 250 μg/ml saponin was used for indirect immunofluorescence of permeabilized cells. The methods to measure translocation and kinetics have been described (Lampson et al., 2001; Zeigerer et al., 2002; Karylowski et al., 2004; Zeigerer et al., 2004; Martin et al., 2006). Fluorescence quantification. Fluorescence microscopy was performed using a DMIRB inverted microscope (Leica Microsystems, Deerfield, IL), with a cooled charge-coupled device camera (Princeton Instruments, Trenton, NJ). Images were collected with a 40 x 1.25 numerical aperture oil immersion objective. MetaMorph software (Universal Imaging, West Chester, PA) was used for image processing and quantification as previously described (Lampson et al., 2001; Zeigerer et al., 2002; Karylowski et al., 2004). Total Internal Reflection Fluorescence (TIRF) microscopy. To measure GLUT4 translocation adipocytes were incubated in SF-DMEM medium for 3 h at 37 C in 5% CO 2 /air. Some cells were incubated with Akt inhibitor AI1/2 (1 μm) during the last hour of this incubation or with the PI3-kinase inhibitor wortmannin (100 nm) for 30 min prior to stimulation with insulin (1 nm) for 30 min. Cells were fixed in 3.7% formaldehyde, and the insulin-induced GFP redistribution was determined using a total internal reflection fluorescence microscope. The TIRF microscope has been described previously (Moskowitz et al., 2003). A 60 x 1.45 numerical aperture oil immersion objective (Olympus America, Melville, NY) was used to perform prismless TIRF. The evanescent field decay length was ~250 nm with this objective, with a pixel size of 112x112 nm 2 in the acquired images. Cells expressing HA-GLUT4-GFP were identified by 7
8 GFP in epifluorescence mode of the microscope. Both epifluorescence and TIRF images of cells were acquired. The GFP fluorescence in the TIRF mode was divided by the GFP epifluorescence intensity, normalizing the TIRF fluorescence for the total HA-GLUT4-GFP expressed per cell. All images were corrected for background fluorescence measured in cells that did not express HA-GLUT4-GFP. To measure insulin-induced HA-GLUT4 fusion, adipocytes stably expressing HA- GLUT4 were used. Two samples were prepared per experimental condition and cells were fixed and stained with anti-ha antibody followed by Cy3-labeled secondary antibody in the presence or absence of saponin. Epifluorescence and TIRF images were acquired. All images were corrected for background fluorescence measured in cells stained with Cy3-labeled secondary antibody. Fluorescence in TIRF mode in permeabilized cells was divided by total fluorescence in the epifluorescence mode to measure insulin-induced recruitment and docking of HA-GLUT4. Fluorescence in the TIRF mode in intact non-permeabilized cells was divided by fluorescence in TIRF mode in permeabilized cells to measure insulin-induced HA-GLUT4 vesicle fusion. For each condition in each experiment, images of 10 to 15 fields were acquired (6 to 10 cells per field). The TIRF signal was between 2-8 fold the background TIRF signal depending on the experimental conditions. The variance of total fluorescence in both epifluorescence and TIRF mode for the different fields in each experiment was < 20% of the mean fluorescence value. For live cell TIRF microscopy studies, cells were serum-starved for 2h in SF-DMEM at 37 C in 5% CO 2, and incubated for 1h in 1 μm AI1/2 or vehicle. Cells were allowed to equilibrate in the TIRF microscope equipped with a temperature-controlled objective at 37 C for 30 min. Videos were acquired at a rate of two frames per minute with a cooled CCD camera. Insulin was added to the samples after recording five frames. To quantify the insulin-induced 8
9 recruitment of GLUT4 to the vicinity of the PM, TIR fluorescence in each frame was quantified, background was subtracted and values were normalized to TIR fluorescence at time zero and plotted as a function of time. Hexose transport- Hexose transport activity in 3T3-L1 adipocytes was measured by the uptake of 3 H-2-deoxy-glucose as described (Gibbs et al., 1988). 9
10 Results Akt inhibition impairs insulin-induced GLUT4 translocation and glucose uptake in adipocytes. The effect of acute pharmacological inhibition of Akt activity on insulin regulated GLUT4 trafficking and glucose transport has not been previously explored. Although overexpression of Akt mutants, and down-regulation or knockout of Akt isoforms have all proved the importance of Akt activity on insulin regulated GLUT4 redistribution, when studying the acute effect of insulin on GLUT4 redistribution prolonged Akt inhibition could render confounding effects considering the signaling potential of this kinase. We used the recently developed allosteric Akt1 and Akt2 inhibitor AI1/2 (Akti-1/2) to explore the role of insulin-induced Akt activation on GLUT4 redistribution and glucose uptake in 3T3-L1 adipocytes. Pretreatment of adipocytes for 1 h with 1 μm of AI1/2 completely blocked the phosphorylation of Akt at Ser 473 promoted by 1 nm insulin and partially inhibited Akt phosphorylation in cells stimulated with 170 nm insulin (Fig. 1A & B). 1 μm AI1/2 was used in this study because it was the lowest concentration of AI1/2 required to completely inhibit Akt phosphorylation induced by 1 nm insulin (not shown). Insulin-induced phosphorylation of GSK3β and AS160, two targets of Akt action, were reduced by AI1/2, demonstrating that AI1/2 inhibited Akt activation. Insulin-stimulated phosphorylation of the MAP kinases ERK1/2, which is independent of Akt activation, was not affected by AI1/2, documenting that AI1/2 inhibited Akt signaling without having general inhibitory effects on signaling downstream of the insulin receptor. We used a HA-GLUT4-GFP reporter construct to characterize the role of Akt signaling in insulin-induced GLUT4 translocation. This construct contains an exofacial HA epitope and GFP fused to the carboxyl cytoplasmic domain (Lampson et al., 2000). The increase of HA- GLUT4-GFP in the PM is measured by an increase in HA epitope in the PM. Stimulation of 10
11 adipocytes with insulin induced the redistribution of HA-GLUT4-GFP to the PM. Despite the significant difference in Akt phosphorylation induced by 1 nm and 170 nm insulin (Fig. 1A), both insulin concentrations promoted a similar GLUT4 translocation to the PM, suggesting that Akt activation induced by 1 nm insulin, a physiologic concentration of insulin, is sufficient to promote maximum effects on GLUT4 translocation in 3T3-L1 adipocytes (Fig. 1D). Insulin-induced redistribution of HA-GLUT4-GFP was significantly inhibited by 1 µm AI1/2 (Fig. 1D). GLUT4 translocation stimulated by 1 nm insulin was reduced by 65 ± 6 % and translocation induced by 170 nm insulin was reduced by 30 ± 10%, establishing that acute inhibition of Akt activity by AI1/2 blunts insulin-stimulated GLUT4 translocation to the PM. Importantly, although AI1/2 treatment abolished 1 nm insulin-induced Akt phosphorylation (Fig. 1A), it did not completely block GLUT4 translocation (Fig. 1D), suggesting that signaling pathways other than Akt contribute to the full effect of insulin on GLUT4 redistribution to the PM. AI1/2 inhibited insulin-stimulated 2-deoxy-glucose uptake, demonstrating that Akt activity is required for insulin-regulated glucose transport (Fig. 1E). Thus, both the translocation data and the hexose uptake data support a prominent role for Akt signaling in insulin-regulation of GLUT4 trafficking in adipocytes. However, a comparison of GLUT4 translocation and insulin-stimulated 2-deoxy-glucose uptake at 1 nm insulin revealed an uncoupling of hexose transport from GLUT4 redistribution, because 1 nm insulin is sufficient for maximal translocation yet not for maximal hexose uptake (Fig. 1D & E). Since the GLUT1 transporter also contributes to insulin-stimulated glucose uptake in 3T3-L1 adipocytes, hexose uptake is not a direct measure of GLUT4 translocation. The discordance in GLUT4 in the PM and hexose 11
12 uptake may reflect differences in insulin signaling to GLUT4 and GLUT1 (Yeh et al., 1995; Liao et al., 2006). GLUT4/IRAP trafficking but not TR trafficking is regulated by Akt signaling. The only other protein known to traffic by the GLUT4 pathway is the aminopeptidase IRAP (Ross et al., 1997; Garza and Birnbaum, 2000). To explore whether Akt signaling is specifically required for insulin-induced GLUT4 redistribution or regulates the IRAP/GLUT4 pathway we next determined the effect of AI1/2 inhibition of Akt on the behavior of IRAP. For these studies, we used an IRAP-transferrin receptor chimera (IRAP-TR) previously shown to traffic like IRAP in adipocytes (Subtil et al., 2000). Inhibition of Akt significantly impaired insulin-induced IRAP-TR translocation to the PM, consistent with the effects of AI1/2 on GLUT4 translocation (Fig. 1F). Thus, AI1/2 inhibition of Akt activation affects insulin regulation of the GLUT4/IRAP trafficking pathway rather than specially alerting the behavior of GLUT4. Insulin promotes a 2-fold increase of transferrin receptor (TR) in the PM, documenting that insulin has effects on general membrane trafficking, albeit minor in magnitude compared to the effect on GLUT4 and IRAP. AI1/2 inhibition of Akt signaling did not have a significant effect on insulin-stimulated redistribution of the TR to the PM, establishing that although Akt signaling is involved in the regulation of the GLUT4/IRAP trafficking pathway, it is not required for the effect of insulin on general endocytic trafficking (Fig 1G). Thus, the insulin signaling pathway that affects general membrane recycling does not completely overlap with the signaling mechanism that regulates GLUT4/IRAP translocation. Akt2 knockdown impairs insulin-induced GLUT4 translocation. 12
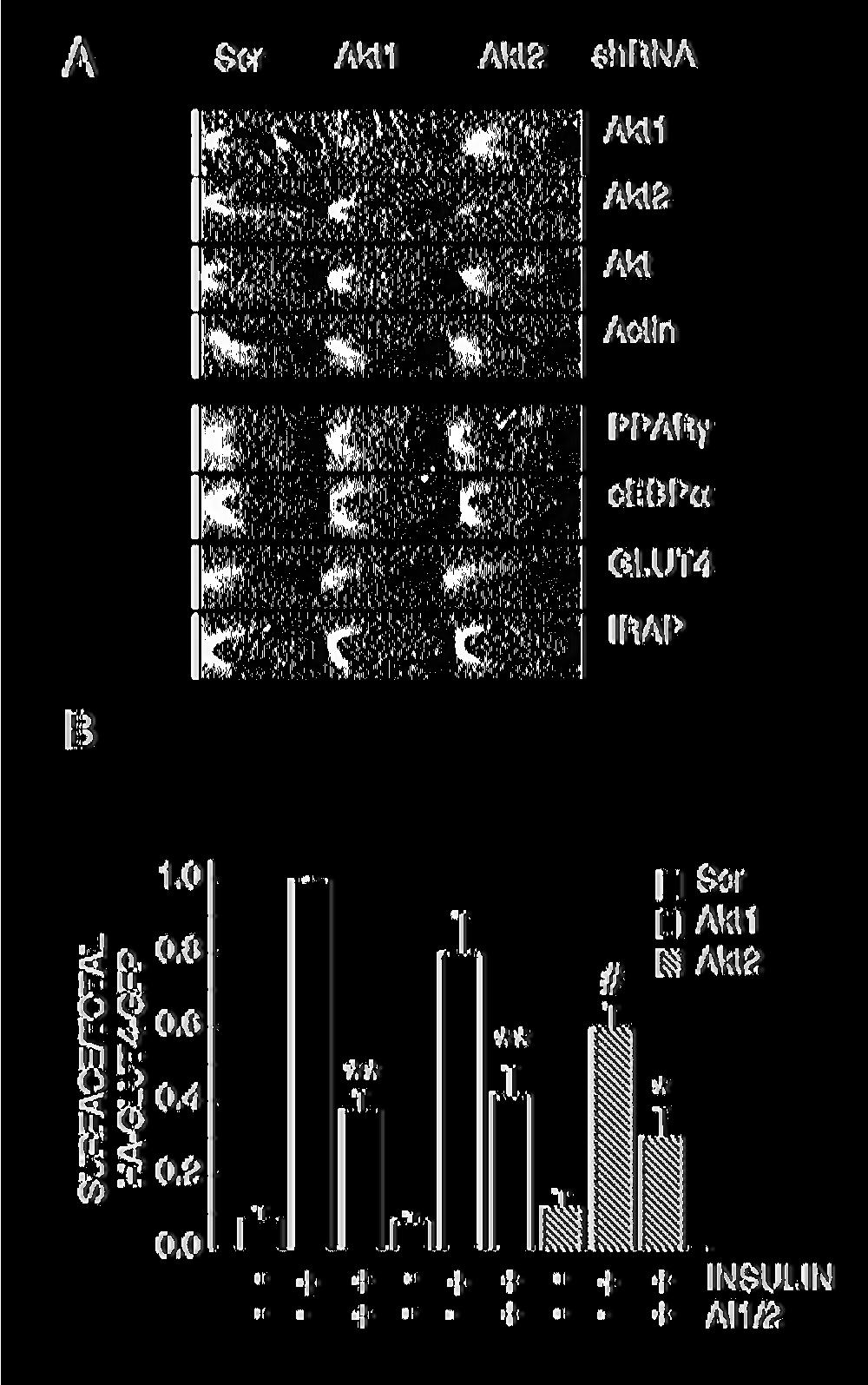
13 To further explore the role of Akt in insulin-regulated GLUT4 translocation, we individually knocked-down Akt1 and Akt2 using retroviral delivered shrna. Akt1 and Akt2 expression were significantly reduced by the isoforms specific shrna (Fig. 2A). Down-regulation of Akt1 or Akt2 did not reproducibly alter the expression level of the other isoform, nor did it alter differentiation into adipocytes, as determined by the expression of the differentiation markers PPARγ, cebpα, GLUT4 and IRAP (Fig 2A). In Akt2 knockdown adipocytes, insulin-induced GLUT4 translocation was inhibited by 40 ± 6% (Fig. 2B). The effect of Akt1 knockdown on GLUT4 translocation was not statistically significant, although the data trended toward a reduction in translocation (Fig. 2B). These results show that Akt2 is the principal Akt isoform regulating GLUT4 trafficking, in agreement with previous studies (Bae et al., 2003; Jiang et al., 2003). Inhibition of Akt activity with AI1/2 further reduced GLUT4 translocation in Akt2 knockdown adipocytes, suggesting that the Akt in these cells, either Akt1 or residual Akt2, contributes to the effect of insulin on GLUT4 distribution. However, even in AI1/2-treated Akt2 knockdown adipocytes, insulin still promoted an ~3-fold redistribution of GLUT4 to the PM (Fig. 2B). These data provide strong support for the hypothesis that insulin signals to GLUT4 by both Akt-dependent and Akt-independent pathways. Akt regulates insulin-induced HA-GLUT4-GFP exocytosis We next determined whether Akt activation is required for insulin stimulation of HA-GLUT4- GFP exocytosis. Adipocytes expressing HA-GLUT4-GFP were stimulated with insulin for 30 min ± AI1/2, followed by incubation in medium containing anti-ha epitope antibody HA.11, insulin ± AI1/2. During the incubation, any HA-GLUT4-GFP that appears on the PM binds 13
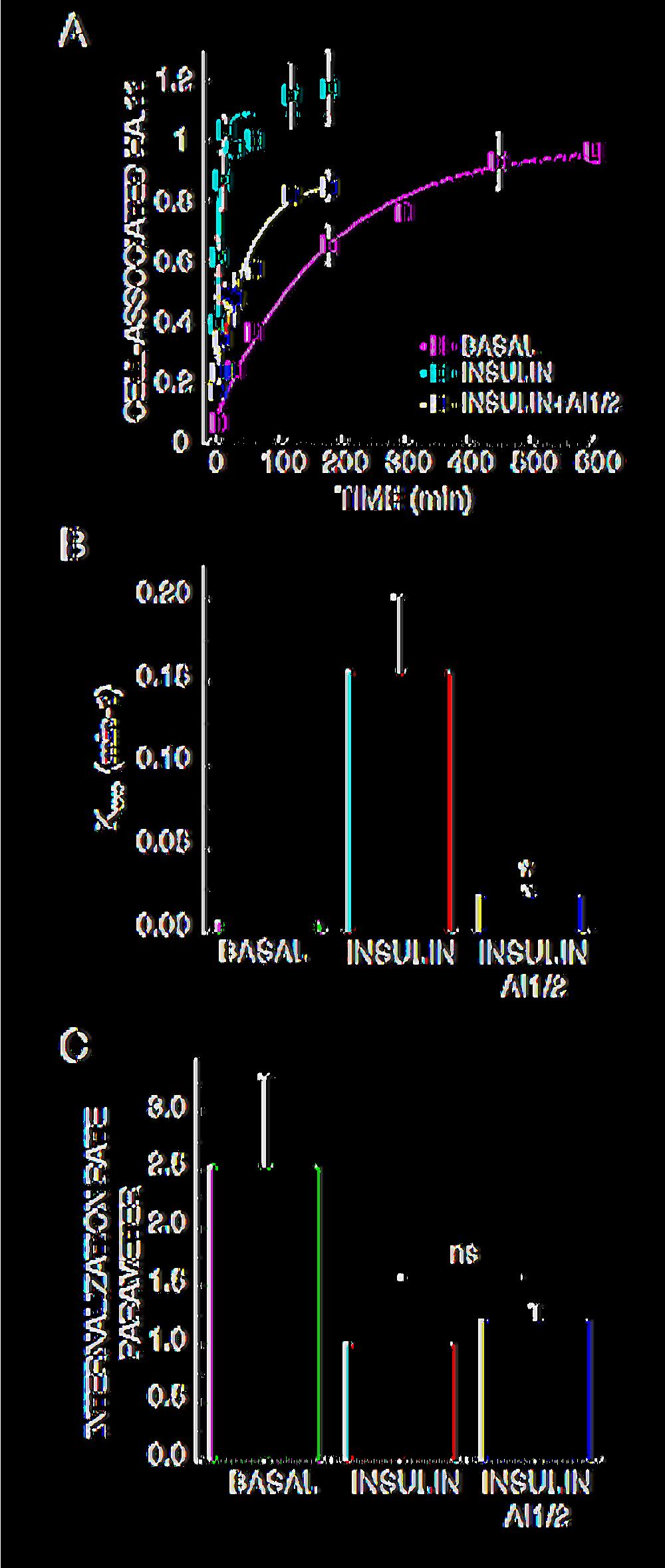
14 HA.11 antibody in the medium, the rate at which cell-associated fluorescence increases over time of incubation is a measure of the exocytosis of HA-GLUT4-GFP (Fig. 3A)(Karylowski et al., 2004). Since the appearance of the HA epitope on the cell surface, which is ultimately dependent on vesicle fusion, is monitored in this exocytosis assay, this assay does not distinguish between regulation of pre-fusion processes (e.g., docking) from regulation of vesicle fusion. AI1/2 treatment slowed insulin-stimulated GLUT4 exocytosis (appearance on the PM) by about 7-fold (Fig. 3A & B). In addition to slowing GLUT4 exocytosis, AI1/2 also reduced the plateau level reached in insulin-stimulated cells, indicating that a portion of intracellular GLUT4 did not cycle to the PM during the time frame of these experiments (Fig. 3A). AI1/2 inhibition of Akt did not completely abrogate the insulin stimulatory effects on GLUT4 exocytosis. In insulin-stimulated AI1/2 treated adipocytes, GLUT4 was exocytosed ~3 times as fast as in basal adipocytes (Fig. 3A & B). These results demonstrate that although Akt signaling is required for insulin regulation of GLUT4 exocytosis, inhibition of Akt signaling is not sufficient to completely block the effects of insulin on GLUT4 exocytosis, and are consistent with the conclusions of translocation experiments in Figs. 1D & 2B. Akt activity is not required for insulin regulation of GLUT4 internalization Insulin-mediated regulation of GLUT4 redistribution to the plasma membrane occurs mainly by accelerating the exocytosis rate of GLUT4, but insulin also have an inhibitory effect on GLUT4 internalization (Jhun et al., 1992; Czech and Buxton, 1993; Zeigerer et al., 2004). We measured the effect of Akt inhibition on insulin regulation of GLUT4 internalization. 3T3-L1 adipocytes expressing HA-GLUT4-GFP were stimulated with insulin ± AI1/2 and incubated with a saturating concentration of HA.11 antibody. The ratio between the amounts of HA.11 14
15 internalized versus HA.11 on the cell surface at different times give us a measure of the endocytosis rate of GLUT4. We observed that the internalization rate of HA-GLUT4-GFP in insulin-stimulated cells was about 2-3 times slower than in basal adipocytes (Fig 3C). Inhibition of Akt signaling did not significantly perturb the internalization rate of insulin-stimulated adipocytes demonstrating that Akt activity is not required for insulin-regulation of GLUT4 internalization. Akt signaling to GLUT4 exocytosis involves several Akt targets. The Akt substrate, AS160, is a Rab GTPase activating protein (GAP) required for basal GLUT4 intracellular retention as well as for insulin-induced GLUT4 redistribution to the PM (Sano et al., 2003; Zeigerer et al., 2004; Eguez et al., 2005). AS160 is proposed to function as a negative regulator of basal GLUT4 exocytosis by maintaining an unknown rab protein in the GDP-bound, inactive state. Akt phosphorylation of AS160 turns off the GAP activity, allowing the rab protein to switch to the GTP-bound, active state. Consistent with this model, knockdown of AS160 results in an increase in basal GLUT4 exocytosis (Eguez et al., 2005). If AS160 is the sole Akt substrate required for Akt-dependent effects on GLUT4 translocation, then inhibition of Akt in AS160 knockdown cells should not have a further effect on insulin-stimulated GLUT4 translocation to the PM. To address this question we examined the effect of AI1/2 on AS160 knockdown adipocytes (Eguez et al., 2005). AS160 was efficiently down regulated by the shrna and we did not detect any insulin-induced AS160 phosphorylation in the knockdown cells (Fig. 4A). AI1/2 inhibited insulin-stimulated exocytosis of GLUT4 in AS160 knockdown adipocytes by about 2-fold, reducing the rate constant from ~0.08 min -1 to ~0.05 min -1 (Fig. 4B & C). Thus, inhibition of Akt impairs GLUT4 exocytosis in AS160-15
16 knockdown adipocytes, suggesting that Akt substrates other than AS160 are involved in insulin regulation of GLUT4 exocytosis. Furthermore, the observation that insulin promotes an acceleration of GLUT4 exocytosis in AI1/2-treated AS160-knockdown adipocytes provides additional evidence for non-akt mediated regulation of GLUT4 exocytosis. Akt signaling regulates insulin-induced recruitment/docking of GLUT4 vesicles to the PM. Insulin regulation of GLUT4 exocytosis might occur at several steps, including the recruitment, docking and/or the fusion of GLUT4 vesicles with the PM. To further explore the role of Akt signaling in regulation of GLUT4 exocytosis, we used total internal reflection fluorescence (TIRF) microscopy and a fixed time point assay to measure insulin-induced redistribution of HA-GLUT4-GFP (Zeigerer et al., 2004). In this assay the effect of insulin on GLUT4 distribution is measured as an increase in GLUT4-containing vesicles within ~250 nm of the dorsal PM (Fig. 5A). Quantification of these data revealed a 2- to 3-fold redistribution of HA- GLUT4-GFP to the TIRF zone (Fig. 5B & C). The fold redistribution measured in the TIRF assay is smaller than in the epitope exposure assay because in the TIRF assay the basal value reflects GLUT4 within 250 nm of the PM, whereas in the epitope exposure assay only GLUT4 that is in the PM is detected (Fig. 5A). Inhibition of PI3-kinase with 100 nm wortmannin blocked insulin-induced redistribution of GLUT4 in the TIRF assay, demonstrating that in these conditions GLUT4 vesicles were not trapped within 250 nm of the dorsal PM, as would be the case if GLUT4 vesicles had docked but had not fused to the PM (Fig. 5C). AI1/2 also inhibited recruitment of GLUT4 in the TIRF microscopy assay (Fig. 5B & C), demonstrating that Akt activity is required for GLUT4 vesicle recruitment and/or docking to the PM and suggesting Akt 16
17 is the downstream effector of the PI3-kinase required for insulin regulation of recruitment and/or docking. We further analyzed the effect of Akt inhibition on insulin-induced GLUT4 redistribution using time-lapse TIRF microscopy. It has been previously shown that GLUT4 vesicles are mobile within the TIRF zone in basal rat primary adipocytes (Lizunov et al., 2005). We also found that GLUT4-containing vesicles in basal 3T3-L1 adipocytes are highly mobile within the TIRF zone (not shown). Insulin stimulation triggers the recruitment of HA-GLUT4-GFP containing vesicles to the TIRF zone, reaching a plateau within 15 min of insulin addition (Fig. 5D). Upon insulin stimulation, the GFP fluorescence in the TIRF zone increases and the fluorescence becomes less punctate and more diffuse, consistent with HA-GLUT4-GFP having fused into the membrane. GLUT4 vesicles in insulin-stimulated AI1/2 pre-treated cells were also highly motile in the TIRF zone; however, HA-GLUT4-GFP accumulation within the TIRF zone was dramatically reduced compared to insulin-treated control adipocytes (Fig. 5D). GLUT4 within the TIRF zone of AI1/2-treated, insulin-stimulated adipocytes plateaued at about 30% of the control level, consistent with the translocation studies in Figs 1 and 2. Although we did not extend the live cell imaging beyond 30 min, we found in fixed cell experiments that the amount of GLUT4 in the PM of AI1/2-treated adipocytes did not increase when incubation with insulin was increased from 30 to 60 min, indicating that the reduced amount of GLUT4 in the PM of AI1/2-treated cells at 30 min reflects a new steady state distribution of GLUT4 between the PM and interior of cells (not shown). Live cell analysis also revealed a 1 to 2 min lag in the recruitment of GLUT4 to the TIRF zone in AI1/2 treated cells compared to control adipocytes as well as a reduction in the rate of accumulation of GLUT4 in the TIRF zone. These data demonstrate that Akt signaling is essential for insulin-induced accumulation of GLUT4 vesicles 17
18 within ~250 nm of the PM, and thereby suggest that Akt regulates recruitment and/or docking of these vesicles to the PM. PI3-kinase but not Akt signaling regulates insulin-induced fusion of GLUT4 vesicles to the PM. We developed a TIRF microscopy fusion assay to determine whether Akt signaling was required for insulin stimulation of GLUT4 vesicle fusion with the PM. For these studies we used adipocytes stably expressing HA-GLUT4 (Govers et al., 2004; Martin et al., 2006). First, we developed a TIRF assay to measure redistribution of HA-GLUT4 to within 250 nm of the PM by adapting the assay described in Fig. 5. Indirect immunofluorescence (IF) of HA-epitope in permeabilized cells using the TIRF microscopy mode was performed to measure HA-GLUT4 within 250 nm of the PM (Fig. 6A). The total amount of HA-GLUT4 in cells was measured in epifluorescence microscopy of the same cells (Fig. 6B). The ratio of the TIRF fluorescence to epifluorescence is a measure of the redistribution of HA-GLUT4 to within 250 nm of the PM, independent of fusion of HA-GLUT4 into the PM. As was the case in the TIRF translocation assay using HA-GLUT4-GFP (Fig. 5C), insulin induced an approximately 2-fold increase in HA- GLUT4 in the TIRF zone, and both wortmannin and AI1/2 inhibited this redistribution (Fig. 6D). These data provide additional support for the hypothesis that Akt and PI3-kinase activity are required for the recruitment and/or docking of GLUT4 vesicles to the PM. Next we developed an assay to measure insulin regulation of the fusion of HA-GLUT4 vesicles with the dorsal PM, independent of insulin s effects on recruitment/docking. HA- GLUT4 in the dorsal PM (vesicle fusion) was measured by IF of HA epitopes in intact cells using TIRF microscopy (Fig. 6C). Only HA-GLUT4 fused into the dorsal PM is detected in the absence of detergent. In parallel samples we measured, using TIRF microscopy, HA-GLUT4 18
19 within 250 nm of the dorsal membrane by IF of detergent permeabilized cells, which is a measure of the total HA-GLUT4 within the TIRF zone (Fig. 6A). The efficiency of HA-GLUT4 fusion with the PM was measured as the ratio of the amount inserted into the PM to the total amount in the TIRF zone. In the insulin-stimulated state about 60% of GLUT4 recruited to the TIRF zone is fused with the PM versus about 25% in the basal state (Fig. 6E). These data demonstrate that insulin stimulates a 2-fold increase in the efficiency of the fusion of HA- GLUT4-containing vesicles with the PM, as well as increasing the accumulation of GLUT4 vesicles near the plasma membrane, as shown in Fig. 5C. Inhibition of Akt with AI1/2 did not significantly reduce insulin-stimulation of the efficiency of GLUT4 vesicle fusion with the PM (Fig. 6E), even though Akt inhibition reduced GLUT4 recruitment to the TIRF zone (Fig. 6D). These data indicate that Akt activity is required for a pre-fusion step involving the recruitment/docking of GLUT4-containing vesicles to the PM but not for efficient fusion of GLUT4 vesicles with the PM, because insulin stimulation of GLUT4 vesicle fusion is not significantly affected by inhibition of Akt, whereas insulinstimulated recruitment/docking of GLUT4 vesicles is affected by Akt inhibition. By contrast, inhibition of PI3-kinase with 100 nm wortmannin reduced the efficiency of GLUT4 vesicle fusion to the PM, demonstrating a requirement for PI3-kinase activity in insulin-stimulated fusion of GLUT4 vesicles (Fig. 6E). These results suggest that insulin regulates GLUT4 exocytosis by increasing both the recruitment/docking and the fusion of GLUT4 vesicles with the PM. Both these processes are dependent on PI3-kinase signaling. However, only the recruitment and/or docking of GLUT4 vesicles is dependent on Akt activation. 19
20 Discussion In this study we have used acute inhibition of Akt activity to dissect the role of Akt signaling on insulin-regulated GLUT4 trafficking. We show that Akt signaling is required for insulinstimulated GLUT4 exocytosis but not for insulin regulation of GLUT4 internalization. Along the exocytic pathway, Akt activity is specifically required for the recruitment/docking of GLUT4 vesicles that precedes insulin-stimulated fusion of these vesicles with the PM. Although the fusion of the vesicles with the PM requires PI3-kinase signaling, it is independent of Akt activity. Thus, our data are consistent with insulin signaling to GLUT4 exocytosis by both Aktdependent and Akt-independent mechanisms. Although Akt activation is specifically required for insulin regulation of the GLUT4/IRAP trafficking pathway, the effects of insulin on TR trafficking do not require Akt activity. Therefore, our data indicate that insulin signaling to GLUT4 trafficking diverges from signaling to the general endocytic pathway (that is, TR trafficking). The effect of insulin on TR behavior may be the Akt-independent arm of signaling to GLUT4. Studies to determine the overlap between insulin signaling to TR and GLUT4 will be the subject of future work. Protein kinase Akt has been established as a crucial regulator of insulin-induced GLUT4 translocation, however a number of observations support the hypothesis that insulin signals to GLUT4 by both Akt-dependent and independent mechanisms. For example, knockdown of both Akt1 and Akt2 in 3T3-L1 adipocytes did not completely inhibit GLUT4 translocation (Jiang et al., 2003), although residual Akt in the knockdown cells could account for the effect of insulin in that study. Here we show that acute inhibition of Akt with 1 µm AI1/2, which completely blocked phosphorylation of Akt-Ser 473 induced by 1 nm insulin (the concentration used for functional studies) significantly reduced, but did not completely block, insulin-stimulated 20
21 GLUT4 translocation in control adipocytes as well as adipocytes in which Akt2 was knocked down. These data reinforce the idea that insulin signals to GLUT4 by Akt-dependent and independent mechanisms, and that full GLUT4 translocation to the PM requires both signaling pathways. What other effectors of insulin action are required for redistribution of GLUT4 to the PM? There is evidence that insulin activation of atypical PKC is required for insulin-stimulated glucose uptake (Kotani et al., 1998; Standaert et al., 1999; Sajan et al., 2006). In addition, insulin activation of the small GTPase TC10, by a mechanism independent of PI3-kinase has a role in GLUT4 redistribution to the PM (Chiang et al., 2001; Inoue et al., 2003; Saltiel and Pessin, 2003). Additional studies are required to determine the relative contributions of these pathways and their individual roles in the redistribution of GLUT4. As a first step toward this goal, in this report we have mapped the role of Akt activation on GLUT4 translocation to the PM. GLUT4 exocytosis is a multi-step regulated process: An Akt dependent, pre-fusion vesicle recruitment/docking step. Here we use a TIRF microscopy assay in intact cells to show that insulin enhances both the recruitment/docking of GLUT4 vesicles and the fusion of GLUT4 vesicles with the PM. The recruitment/docking step is measured as a change in GLUT4 in the TIRF excitation zone (~250 nm of the dorsal PM) and is independent of whether the GLUT4 vesicles have fused to the PM. Inhibition of Akt blocked insulin-stimulated GLUT4 accumulation within 250 nm of the PM (Fig. 5). The TIRF zone is about 3 GLUT4 vesicle diameters in depth and therefore it should encompass vesicles docked to the PM. Hence, these data suggest that Akt signaling is required for the GLUT4 vesicle recruitment/docking, since if vesicles were arrested at a docked state, 21
22 GFP in the TIRF zone would have been increased by insulin. Inhibition of PI3-kinase activation, which is required for activation of Akt, also inhibited insulin-stimulated accumulation of GLUT4 within the TIRF zone. The TIRF microscopy assay cannot distinguish between active recruitment of GLUT4 vesicles to within 250 nm of the PM from regulated docking to the PM. There is some controversy in the literature whether insulin actively recruits GLUT4 vesicles to the PM. For example, a recent live cell TIRF analysis of GLUT4 trafficking in primary rat fat cells concluded that GLUT4 vesicle movement to the PM is not accelerated by insulin, but rather that the docking of these vesicles to the PM is regulated by insulin (Lizunov et al., 2005). Based on those observations, and the results of our studies, we favor the model that Akt is required for the regulated docking of GLUT4 vesicles with the PM. Using live cell TIRF microscopy we observed that Akt inhibition reduced the insulin-stimulated net accumulation of GLUT4 at the PM, slowed the rate of GLUT4 accumulation within the TIRF zone, and induced a lag in the increase of GLUT4 in the TIRF zone following insulin stimulation (Fig. 5D). These observations are all consistent with an inefficient docking of GLUT4 vesicles at the PM when Akt activation is blocked. However, once these vesicles are properly docked, albeit an inefficient process when Akt is inhibited, fusion with the PM is properly stimulated by insulin. Thus, the steady state level of GLUT4 in the PM of insulin-stimulated, AI1/2-treated adipocytes is reduced compared to control cells because the recruitment and/or docking is inefficient. However, GLUT4 in the PM of insulin-stimulated, AI1/2-treated adipocytes is increased relative to basal cells because fusion (and other steps of GLUT4 movement) are regulated by an Akt-independent process(es). Recently Holman and coworkers have developed an vitro system to measure insulininduced GLUT4 vesicle fusion (Koumanov et al., 2005). The results of that study argue that 22
23 insulin regulation of GLUT4 vesicle fusion is a key step in GLUT4 translocation to the plasma membrane, and that fusion is dependent on the recruitment of active Akt to the plasma membrane. However, the in vitro studies do not directly address differences between vesicle docking and fusion. Vesicle docking precedes fusion and therefore unstimulated (or inefficient) docking would be scored as poor fusion in the in vitro assay. Thus, their in vitro data are compatible with our proposal that Akt regulates a pre-fusion step. Our results in intact cells extend their proposal that insulin action on GLUT4 occurs, in part, at the plasma membrane, by showing that principally a pre-fusion step (recruitment and/or docking), and not the fusion, is the Akt regulated step. The best-characterized downstream effector of Akt required for GLUT4 translocation is the rab GAP AS160. Past studies have shown that a dominant-interfering AS160 mutant that cannot be phosphorylated by Akt, blocked insulin-stimulated redistribution of GLUT4 to within 250 nm of the PM (Zeigerer et al., 2004), and we have also observed that AS160 knockdown impairs insulin-induced recruitment of GLUT4 vesicles to the TIRF zone (Supplemental Figure 1). Those results support the model that Akt regulates docking of GLUT4 to the PM, and they suggest that Akt regulates docking at least in part by targeting AS160. Rab proteins are known to regulate docking and fusion of vesicles and it is tempting to speculate that the rab target of AS160 GAP activity has a role in the regulation of docking. At this time the rab target of AS160 is not known. It is important to note, however, that although we favor the model that the pre-fusion step regulated by Akt activity is vesicle docking and not active recruitment of GLUT4 vesicles to the PM, the results of several studies support the hypothesis that insulin signals to regulate GLUT4 vesicle movement along microtubules (Bose et al., 2002; Semiz et al., 2003) and/or the actin 23
24 cytoskeleton (Tong et al., 2001). Thus, these effects of insulin could have a role in the active recruitment of GLUT4 vesicles to within 250 nm of the PM. Regardless of whether insulin actively recruits GLUT4 from intracellular sites and/or promotes docking of vesicles, our data show that PI3-kinase and Akt signaling are necessary for a pre-fusion step resulting in the accumulation of GLUT4 vesicles within ~ 250 nm of the PM. Previous studies have shown that inhibition of PI3-kinase (Bose et al., 2004) or a temperature block of Akt activity (van Dam et al., 2005) promoted the accumulation of GLUT4 vesicles adjacent to the PM, as would be the case if the vesicles were docked but not fused. An interpretation of those data is that insulin regulates docking independent of PI3-kinase and Akt inactivation. We do not, however, observe this effect in the TIRF assay. One possible explanation for this discrepancy is that we measured GLUT4 near the PM with TIRF microscopy and the other studies used confocal microscopy. If the GLUT4 vesicles accumulate beyond 250 nm of the membrane when PI3-kinase or Akt are inhibited, they would not be detected in TIRF assay, but they would appear as a plasma membrane ring in confocal analysis. Thus, those previous studies may have uncovered a regulated tethering (beyond 250 nm) of the PM that is stimulated by insulin in a PI3-kinase/Akt -independent process, but according to our data might not have measured insulin regulated GLUT4 vesicle docking at the PM. GLUT4 exocytosis is a multi-step regulated process: GLUT4 vesicle fusion, a PI3-kinasedependent, Akt-independent process. Although insulin-stimulated recruitment/docking of GLUT4 vesicles to the TIRF zone is reduced when Akt is inhibited, insulin-stimulated fusion of GLUT4 vesicles that are within 250 nm of the PM is not significantly affected by Akt inhibition, indicating that Akt activity does not regulate 24
25 fusion. In contrast, in cells in which PI3-kinase is inhibited, insulin-induced fusion of GLUT4 vesicles is blocked. Therefore, our data document a divergence in insulin signaling to GLUT4 vesicles, with vesicle recruitment/docking requiring Akt activation and fusion requiring PI3- kinase activity but not Akt activation. Previous studies support insulin-regulation of GLUT4 vesicle fusion (Kanda et al., 2005; Koumanov et al., 2005; van Dam et al., 2005). One possible candidate to regulate GLUT4 vesicle fusion is Munc18c, a syntaxin 4 interacting protein (Thurmond et al., 2000; Kanda et al., 2005). Studies of adipocytes derived from Munc18c null mice support the hypothesis that Munc18c is a negative regulator of GLUT4 fusion at the PM and a downstream target of PI3- kinase activation, consistent with out findings that fusion requires PI3-kinase activity (Kanda et al., 2005). It has been proposed that insulin signaling, through PKCζ/λ, promotes disassociation of Munc18c from syntaxin 4, allowing for the assembly of the SNARE fusion complex (Thurmond et al., 2000; Hodgkinson et al., 2005). Those results suggest that PKCζ/λ may be the PI3-kinase effector regulating the fusion. On the other hand, insulin-mediated recruitment of the exocyst complex to the PM has been proposed to regulate GLUT4 translocation, potentially by regulating docking (Inoue et al., 2003). Future studies are required to determine how and whether these pathways converge to regulate GLUT4 vesicle docking and fusion. Insulin regulation of GLUT4 trafficking is a complex process that requires the integration of several signaling pathways downstream of the insulin receptor. Our data support a model in which insulin signals to GLUT4 exocytosis, regulating the recruitment/docking and fusion of GLUT4 vesicles to the PM. Dissecting the specific input of insulin signaling mediators on GLUT4 movement is a requisite if we are to understand how insulin regulates glucose transport in fat and muscles and how this process is disrupted in insulin-resistant states. 25
26 References Bae, S.S., Cho, H., Mu, J., and Birnbaum, M.J. (2003). Isoform-specific regulation of insulindependent glucose uptake by Akt/protein kinase B. J Biol Chem 278, Bogan, J.S., Hendon, N., McKee, A.E., Tsao, T.S., and Lodish, H.F. (2003). Functional cloning of TUG as a regulator of GLUT4 glucose transporter trafficking. Nature 425, Bose, A., Guilherme, A., Robida, S.I., Nicoloro, S.M., Zhou, Q.L., Jiang, Z.Y., Pomerleau, D.P., and Czech, M.P. (2002). Glucose transporter recycling in response to insulin is facilitated by myosin Myo1c. Nature 420, Bose, A., Robida, S., Furcinitti, P.S., Chawla, A., Fogarty, K., Corvera, S., and Czech, M.P. (2004). Unconventional myosin Myo1c promotes membrane fusion in a regulated exocytic pathway. Mol Cell Biol 24, Chiang, S., Baumann, C., Kanzaki, M., Thurmond, D., Watson, R., Neudauer, C., Macara, I., Pessin, J., and Saltiel, A. (2001). Insulin-stimulated GLUT4 translocation requires the CAPdependent activation of TC10. Nature 410, p Cho, H., Mu, J., Kim, J.K., Thorvaldsen, J.L., Chu, Q., Crenshaw, E.B., 3rd, Kaestner, K.H., Bartolomei, M.S., Shulman, G.I., and Birnbaum, M.J. (2001). Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB beta). Science 292, Czech, M.P., and Buxton, J.M. (1993). Insulin action on the internalization of the GLUT4 glucose transporter in isolated rat adipocytes. J Biol Chem 268, DeFeo-Jones, D., Barnett, S.F., Fu, S., Hancock, P.J., Haskell, K.M., Leander, K.R., McAvoy, E., Robinson, R.G., Duggan, M.E., Lindsley, C.W., Zhao, Z., Huber, H.E., and Jones, R.E. (2005). Tumor cell sensitization to apoptotic stimuli by selective inhibition of specific Akt/PKB family members. Mol Cancer Ther 4, Eguez, L., Lee, A., Chavez, J.A., Miinea, C.P., Kane, S., Lienhard, G.E., and McGraw, T.E. (2005). Full intracellular retention of GLUT4 requires AS160 Rab GTPase activating protein. Cell Metab 2, Garza, L., and Birnbaum, M. (2000). Insulin-responsive aminopeptidase trafficking in 3T3-L1 adipocytes. J. Biol. Chem. 275, Gibbs, E.M., Lienhard, G.E., and Gould, G.W. (1988). Insulin-induced translocation of glucose transporters to the plasma membrane precedes full stimulation of hexose transport. Biochemistry 27, Govers, R., Coster, A.C., and James, D.E. (2004). Insulin increases cell surface GLUT4 levels by dose dependently discharging GLUT4 into a cell surface recycling pathway. Mol Cell Biol 24, Hill, M.M., Clark, S.F., Tucker, D.F., Birnbaum, M.J., James, D.E., and Macaulay, S.L. (1999). A role for protein kinase Bbeta/Akt2 in insulin-stimulated GLUT4 translocation in adipocytes. Mol Cell Biol 19, Hodgkinson, C.P., Mander, A., and Sale, G.J. (2005). Protein kinase-zeta interacts with munc18c: role in GLUT4 trafficking. Diabetologia 48, Inoue, M., Chang, L., Hwang, J., Chiang, S.H., and Saltiel, A.R. (2003). The exocyst complex is required for targeting of Glut4 to the plasma membrane by insulin. Nature 422,
27 Ishiki, M., Randhawa, V.K., Poon, V., Jebailey, L., and Klip, A. (2005). Insulin regulates the membrane arrival, fusion, and C-terminal unmasking of glucose transporter-4 via distinct phosphoinositides. J Biol Chem 280, Jhun, B.H., Rampal, A.L., Liu, H., Lachaal, M., and Jung, C.Y. (1992). Effects of insulin on steady state kinetics of GLUT4 subcellular distribution in rat adipocytes. Evidence of constitutive GLUT4 recycling. J Biol Chem 267, Jiang, Z.Y., Zhou, Q.L., Coleman, K.A., Chouinard, M., Boese, Q., and Czech, M.P. (2003). Insulin signaling through Akt/protein kinase B analyzed by small interfering RNA-mediated gene silencing. Proc Natl Acad Sci U S A 100, Kanda, H., Tamori, Y., Shinoda, H., Yoshikawa, M., Sakaue, M., Udagawa, J., Otani, H., Tashiro, F., Miyazaki, J., and Kasuga, M. (2005). Adipocytes from Munc18c-null mice show increased sensitivity to insulin-stimulated GLUT4 externalization. J Clin Invest 115, Karylowski, O., Zeigerer, A., Cohen, A., and McGraw, T.E. (2004). GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes. Mol Biol Cell 15, Kohn, A.D., Summers, S.A., Birnbaum, M.J., and Roth, R.A. (1996). Expression of a constitutively active Akt Ser/Thr kinase in 3T3-L1 adipocytes stimulates glucose uptake and glucose transporter 4 translocation. J Biol Chem 271, Kotani, K., Ogawa, W., Matsumoto, M., Kitamura, T., Sakaue, H., Hino, Y., Miyake, K., Sano, W., Akimoto, K., Ohno, S., and Kasuga, M. (1998). Requirement of atypical protein kinase clambda for insulin stimulation of glucose uptake but not for Akt activation in 3T3-L1 adipocytes. Mol Cell Biol 18, Koumanov, F., Jin, B., Yang, J., and Holman, G.D. (2005). Insulin signaling meets vesicle traffic of GLUT4 at a plasma-membrane-activated fusion step. Cell Metab 2, Lampson, M.A., Racz, A., Cushman, S.W., and McGraw, T.E. (2000). Demonstration of insulinresponsive trafficking of GLUT4 and vptr in fibroblasts. J Cell Sci 113 (Pt 22), Lampson, M.A., Schmoranzer, J., Zeigerer, A., Simon, S.M., and McGraw, T.E. (2001). Insulinregulated release from the endosomal recycling compartment is regulated by budding of specialized vesicles. Mol Biol Cell 12, Liao, W., Nguyen, M.T., Imamura, T., Singer, O., Verma, I.M., and Olefsky, J.M. (2006). Lentiviral short hairpin ribonucleic acid-mediated knockdown of GLUT4 in 3T3-L1 adipocytes. Endocrinology 147, Lizunov, V.A., Matsumoto, H., Zimmerberg, J., Cushman, S.W., and Frolov, V.A. (2005). Insulin stimulates the halting, tethering, and fusion of mobile GLUT4 vesicles in rat adipose cells. J Cell Biol 169, Martin, O.J., Lee, A., and McGraw, T.E. (2006). GLUT4 distribution between the plasma membrane and the intracellular compartments is maintained by an insulin-modulated bipartite dynamic mechanism. J Biol Chem 281, Moskowitz, H.S., Heuser, J., McGraw, T.E., and Ryan, T.A. (2003). Targeted chemical disruption of clathrin function in living cells. Mol Biol Cell 14, Ross, S.A., Herbst, J.J., Keller, S.R., and Lienhard, G.E. (1997). Trafficking kinetics of the insulin-regulated membrane aminopeptidase in 3T3-L1 adipocytes. Biochem Biophys Res Commun 239, Sajan, M.P., Rivas, J., Li, P., Standaert, M.L., and Farese, R.V. (2006). Repletion of Atypical Protein Kinase C following RNA Interference-mediated Depletion Restores Insulin-stimulated Glucose Transport. J Biol Chem 281,
28 Saltiel, A.R., and Pessin, J.E. (2003). Insulin signaling in microdomains of the plasma membrane. Traffic 4, Sano, H., Kane, S., Sano, E., Miinea, C.P., Asara, J.M., Lane, W.S., Garner, C.W., and Lienhard, G.E. (2003). Insulin-stimulated phosphorylation of a Rab GTPase-activating protein regulates GLUT4 translocation. J Biol Chem 278, Semiz, S., Park, J.G., Nicoloro, S.M., Furcinitti, P., Zhang, C., Chawla, A., Leszyk, J., and Czech, M.P. (2003). Conventional kinesin KIF5B mediates insulin-stimulated GLUT4 movements on microtubules. Embo J 22, Shewan, A., Marsh, B., Melvin, D., Martin, S., Gould, G., and James, D. (2000). The cytosolic C-terminus of the glucose transporter GLUT4 contains an acidic cluster endosomal targeting motif distal to the dileucine signal. [Record Supplied By Publisher]. Biochem J 350, p Standaert, M.L., Bandyopadhyay, G., Perez, L., Price, D., Galloway, L., Poklepovic, A., Sajan, M.P., Cenni, V., Sirri, A., Moscat, J., Toker, A., and Farese, R.V. (1999). Insulin activates protein kinases C-zeta and C-lambda by an autophosphorylation-dependent mechanism and stimulates their translocation to GLUT4 vesicles and other membrane fractions in rat adipocytes. J Biol Chem 274, Subtil, A., Lampson, M.A., Keller, S.R., and McGraw, T.E. (2000). Characterization of the insulin-regulated endocytic recycling mechanism in 3T3-L1 adipocytes using a novel reporter molecule. J Biol Chem 275, Thurmond, D., Kanzaki, M., Khan, A., and Pessin, J. (2000). Munc18c function is required for insulin-stimulated plasma membrane fusion of GLUT4 and insulin-responsive amino peptidase storage vesicles. Mol Cell Biol 20, p Tong, P., Khayat, Z.A., Huang, C., Patel, N., Ueyama, A., and Klip, A. (2001). Insulin-induced cortical actin remodeling promotes GLUT4 insertion at muscle cell membrane ruffles. J Clin Invest 108, van Dam, E.M., Govers, R., and James, D.E. (2005). Akt activation is required at a late stage of insulin-induced GLUT4 translocation to the plasma membrane. Mol Endocrinol 19, Wang, Q., Somwar, R., Bilan, P.J., Liu, Z., Jin, J., Woodgett, J.R., and Klip, A. (1999). Protein kinase B/Akt participates in GLUT4 translocation by insulin in L6 myoblasts. Mol Cell Biol 19, Watson, R.T., and Pessin, J.E. (2006). Bridging the GAP between insulin signaling and GLUT4 translocation. Trends Biochem Sci 31, Xu, Z., and Kandror, K.V. (2002). Translocation of small preformed vesicles is responsible for the insulin activation of glucose transport in adipose cells. Evidence from the in vitro reconstitution assay. J Biol Chem 277, Yeh, J.I., Verhey, K.J., and Birnbaum, M.J. (1995). Kinetic analysis of glucose transporter trafficking in fibroblasts and adipocytes. Biochemistry 34, Zeigerer, A., Lampson, M., Karylowski, O., Sabatini, D., Adesnik, M., Ren, M., and McGraw, T. (2002). GLUT4 retention in adipocytes requires two intracellular insulin-regulated transport steps. Mol Biol Cell 13, p Zeigerer, A., McBrayer, M.K., and McGraw, T.E. (2004). Insulin Stimulation of GLUT4 Exocytosis, but Not Its Inhibition of Endocytosis, Is Dependent on RabGAP AS160. Mol Biol Cell 15, Zhao, Z., Leister, W.H., Robinson, R.G., Barnett, S.F., Defeo-Jones, D., Jones, R.E., Hartman, G.D., Huff, J.R., Huber, H.E., Duggan, M.E., and Lindsley, C.W. (2005). Discovery of 2,3,5-28
29 trisubstituted pyridine derivatives as potent Akt1 and Akt2 dual inhibitors. Bioorg Med Chem Lett 15, Acknowledgements. We wish to thank members of the McGraw laboratory, Gus Lienhard and Markus Schober for helpful discussions and suggestions, and Daniel Chung for expert technical assistance. This work was supported by NIH DK52852 (TEM) and an American Heart Association Heritage Affiliate Award (EG). 29
30 Figure legends Figure 1. Inhibition of Akt impairs GLUT4 translocation and glucose uptake in 3T3-L1 adipocytes. A. 3T3-L1 adipocytes were treated with 1 µm AI1/2 or vehicle for 1h prior to insulin stimulation for 15 min. Cell lysates were analyzed in immunoblots using noted antibodies. B. Densitometric analyses from pooled immunoblot data. The means ± SE of 3 independent experiments are shown. C. Immunoblot analyses of adipocytes stimulated with 1 nm insulin for 15 min. As noted, some samples were treated with 1 μm AI1/2 for 1h prior to insulin stimulation. Phosphorylated AS160 (arrow) was detected with the PAS antibody. D. Surface-to-total distribution of HA-GLUT4-GFP. Each bar is the mean ± SE of 3 independent experiments. The surface-to-total HA-GLUT4-GFP distribution was normalized to the distribution in cells stimulated with 1 nm insulin for 30 min. As noted, some samples were treated with 1 μm AI1/2 for 1h prior to 30 min insulin stimulation. E. Glucose uptake in adipocytes. Cells were treated with vehicle or 1 μm AI1/2 for 1h prior to 30 min insulin stimulation. Each bar is the mean ± SE of 4 independent experiments. F & G. Surface-to-total distribution of IRAP-TR chimera (F) and TR (G) in adipocytes. Cells were treated with vehicle or 1 μm AI1/2 for 1h prior to 30 min incubation in insulin. Each bar is the mean ± SE of 2 independent experiments. The surface-to-total distributions were normalized to the distribution in control cells stimulated with 1 nm insulin. * indicates p<0.05, ** indicates p<0.01 versus cells that were not treated with AI1/2 (ANOVA). 30
31 Figure 2. shrna-mediated Akt knockdown impairs insulin-induced GLUT4 translocation. A. Immunoblot analyses of extracts from adipocytes expressing scrambled, Akt1 or Akt2 specific shrna. B. Surface-to-total distribution of HA-GLUT4-GFP in control, Akt1 and Akt2 knockdown adipocytes. 1 nm insulin and 1 µm AI1/2 were used. Each bar is the mean ± SE of 3 independent experiments. In each experiment the surface-to-total HA-GLUT4-GFP distribution was normalized to the distribution in control insulin stimulated cells. * Indicates p<0.05, ** indicates p<0.01 versus cells that were not treated with AI1/2, # indicates p<0.01 versus insulinstimulated control cells (ANOVA). Figure 3. Akt regulates insulin-induced GLUT4 exocytosis but not insulin inhibition of GLUT4 internalization. A. Steady-state basal and insulin-stimulated GLUT4 exocytosis kinetics in 3T3-L1 adipocytes. The data are the mean ± SE of 3 independent experiments for the insulin-stimulated state and 2 experiments for the basal state. 1 nm insulin and 1 µm AI1/2 were used. The lines are a fit of the data to (Cy3/GFP) t = (HA-GLUT4-GFP) plateau [(HA-GLUT4-GFP) intracellular *(e - kexo*t )]; where Kexo is the exocytosis rate constant, t is the time of incubation, Cy3/GFP is cell associated HA.11 at time t, (HA-GLUT4-GFP) plateau is the Cy3-to-GFP plateau level, (HA- GLUT4-GFP) intracellular is the amount of HA-GLUT4-GFP intracellular at time 0. B. Basal and insulin-stimulated GLUT4 K exo in the presence or absence of AI1/2. Each bar is the mean ± SE of 2 or 3 independent experiments. * indicates p<0.05 compared to insulinstimulated state in the absence of the Akt inhibitor (ANOVA). 31
32 C. Basal and insulin-stimulated GLUT4 internalization rate parameter in the presence or absence of AI1/2. Each bar is the mean ± SE of 2 to 4 independent experiments. The data are normalized to the internalization rate parameter measured for control cells in the insulin stimulated state. The internalization rate parameter is proportional to the internalization rate constant but is not the rate constant since different fluorophores are used to measure surface and internal HA-GLUT4-GFP. Figure 4. Akt regulates GLUT4 exocytosis by AS160 dependent and independent mechanisms. A. Immunoblot analyses of cell extracts from adipocytes expressing scrambled or AS160 shrna. Phosphorylated AS160 (arrow) was detected with the PAS antibody. B. Steady-state basal and insulin-stimulated GLUT4 exocytosis kinetics in AS160 knockdown 3T3-L1 adipocytes. The data are the mean ± SE of 3 independent experiments in the insulin-stimulated state and 2 independent experiments in the basal state. 1 nm insulin and 1 µm AI1/2 were used. C. Basal and insulin-stimulated GLUT4 K exo in the presence or absence of AI1/2 in AS160 knockdown adipocytes. Each data point is the mean ± SE of 2 to 3 independent experiments. * Indicates p<0.05 compared to insulin-stimulated state in the absence of the Akt inhibitor (ANOVA). Figure 5. Akt signaling is required for insulin-induced recruitment/docking of GLUT4 vesicles to the plasma membrane. 32
33 A. TIRF translocation assay. HA-GLUT4-GFP fluorescence in basal or insulinstimulated cells was measured in the epifluorescence mode and in the TIRF mode. In the epifluorescence mode, translocation is measured as the exposure of the HA-epitope of HA- GLUT4-GFP on the surface of intact adipocytes, which is measured by indirect immunofluorescence (IF). The HA fluorescence is normalized to GFP in cells also detected by epifluorescence (Lampson et al., 2000). In the TIRF assay, redistribution of HA-GLUT4-GFP to within 250 nm of the membrane is measured by GFP fluorescence. The TIRF GFP signal is normalized to the total amount of HA-GLUT4-GFP per cell measured in the epifluorescence mode of the microscope. The TIRF assay measures redistribution to within 250 nm of the PM independent of whether the GLUT4 is inserted into the PM (that is, independent of vesicle fusion). B. Representative images of HA-GLUT4-GFP in the basal state and following 1 nm insulin stimulation for 30 min ± 1 µm AI1/2. Total HA-GLUT4-GFP was determined by GFP fluorescence in the epifluorescence mode, HA-GLUT4-GFP inserted in the PM was determined by indirect immunofluorescence of the HA epitope, and HA-GLUT4-GFP in the TIRF zone was determined by GFP fluorescence in the TIRF mode. C. Quantification of GLUT4 redistribution measured using TIRF microscopy. For each experiment the data are normalized to the insulin stimulated state. The values shown are the means ± SE of 3 independent experiments. 1 µm AI1/2, 100 nm wortmannin, and 1 nm insulin were used. * Indicates p<0.05 versus the insulin stimulated state (ANOVA). D. Quantification of time-lapsed TIRF microscopy of HA-GLUT4-GFP in the TIRF zone. Images were acquired every 30 sec for 30 min. 1 nm insulin or vehicle (mock) were added 33
34 after the 5 th frame (noted by the arrow). 1 µm AI1/2 was added 1h prior to recording. Data shown are the mean ± SE from 6-9 cells per condition, derived from 3 independent experiments. Figure 6. PI3-kinase but not Akt signaling regulates insulin-induced GLUT4 vesicle fusion to the PM. A. HA-GLUT4 redistribution to within 250 nm of the dorsal PM was measured by IF of permeabilized cells using TIRF microscopy. Although all the anti-ha antibodies in the cells are bound by fluorescently labeled secondary antibody, only the fluorescent secondary antibody bound to anti-ha-epitope antibodies within the TIRF zone are excited (red stars). B. The total amount of HA-GLUT4 in permeabilized cells was determined by epifluorescence microscopy. The ratio of the TIRF to epifluorescence is a measure of the redistribution of HA-GLUT4 to within 250 nm of the dorsal PM (panel D). C. HA-GLUT4 inserted into the PM was determined IF of intact (non-permeabilized cells) by TIRF microscopy. The ratio of the TIRF fluorescence in intact cells to TIRF fluorescence on permeabilized cells (panel A) is proportional to the efficiency of HA-GLUT4 vesicle fusion with the PM since it reflects the fraction of HA-GLUT4 within 250 nm of the membrane that is inserted in the PM. D. Measurement of insulin-induced GLUT4 vesicle recruitment/docking in 3T3-L1 adipocytes stably expressing HA-GLUT4. The data are the ratio of the HA-GLUT4 within 250 nm of the PM of permeabilized cells, as determined by IF (anti-ha) in TIRF mode (panel A), divided by the total HA-GLUT4 in cells determined in epifluorescence mode of the same samples (panel B). The data are normalized to the insulin-stimulated state of control cells. The 34
35 data are the means ± SE of 3 to 6 independent experiments. 1 nm insulin, 100 nm wortmannin and 1 µm AI1/2 were used. E. Efficiency of GLUT4 vesicle fusion with PM. The data are the ratio of the HA- GLUT4 in the PM, as determined by IF in TIRF mode of intact cells (panel C) divided by the HA-GLUT4 within the TIRF zone, as determined in TIRF mode of permeabilized cells (panel A). The means ± SE of 3 to 6 independent experiments are shown. 1 nm Insulin, 100 nm wortmannin and 1 µm AI1/2 were used. The difference between insulin-stimulated AI1/2 treated cells and insulin stimulated control cells was not statistically different. * Indicates p<0.001 versus the insulin stimulated control cells, # indicates p<0.05 versus AI1/2, insulinstimulated cells (ANOVA). 35

36
37
38

39

40

41
Insulin Stimulation of GLUT4 Exocytosis, but Not Its Inhibition of Endocytosis, Is Dependent on RabGAP AS160
 Molecular Biology of the Cell Vol. 15, 4406 4415, October 2004 Insulin Stimulation of GLUT4 Exocytosis, but Not Its Inhibition of Endocytosis, Is Dependent on RabGAP AS160 Anja Zeigerer,* Mary Kate McBrayer,
Molecular Biology of the Cell Vol. 15, 4406 4415, October 2004 Insulin Stimulation of GLUT4 Exocytosis, but Not Its Inhibition of Endocytosis, Is Dependent on RabGAP AS160 Anja Zeigerer,* Mary Kate McBrayer,
Rab10, a Target of the AS160 Rab GAP, Is Required for Insulin-Stimulated Translocation of GLUT4 to the Adipocyte Plasma Membrane
 Article Rab10, a Target of the AS160 Rab GAP, Is Required for Insulin-Stimulated Translocation of GLUT4 to the Adipocyte Plasma Membrane Hiroyuki Sano, 1,5 Lorena Eguez, 2,5 Mary N. Teruel, 3 Mitsunori
Article Rab10, a Target of the AS160 Rab GAP, Is Required for Insulin-Stimulated Translocation of GLUT4 to the Adipocyte Plasma Membrane Hiroyuki Sano, 1,5 Lorena Eguez, 2,5 Mary N. Teruel, 3 Mitsunori
GLUT4 Retention in Adipocytes Requires Two Intracellular Insulin-regulated Transport Steps
 Molecular Biology of the Cell Vol. 13, 2421 2435, July 2002 GLUT4 Retention in Adipocytes Requires Two Intracellular Insulin-regulated Transport Steps Anja Zeigerer,* Michael A. Lampson,* Ola Karylowski,*
Molecular Biology of the Cell Vol. 13, 2421 2435, July 2002 GLUT4 Retention in Adipocytes Requires Two Intracellular Insulin-regulated Transport Steps Anja Zeigerer,* Michael A. Lampson,* Ola Karylowski,*
GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
 MBC in Press, published on October 31, 2003 as 10.1091/mbc.E03-07-0517 GLUT4 retention by intracellular cycling GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
MBC in Press, published on October 31, 2003 as 10.1091/mbc.E03-07-0517 GLUT4 retention by intracellular cycling GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
GLUT4 traffic and insulin signaling. Molecular Mechanisms Controlling GLUT4 Intracellular Retention. Vincent Blot and Timothy E.
 Molecular Mechanisms Controlling GLUT4 Intracellular Retention. Vincent Blot and Timothy E. McGraw 1 Department of Biochemistry Weill Cornell Medical College 1 Corresponding author 1300 York Ave. New York,
Molecular Mechanisms Controlling GLUT4 Intracellular Retention. Vincent Blot and Timothy E. McGraw 1 Department of Biochemistry Weill Cornell Medical College 1 Corresponding author 1300 York Ave. New York,
IRAP is a key regulator of GLUT4 trafficking by controlling the sorting of GLUT4 from endosomes to specialized insulinregulated
 IRAP is a key regulator of GLUT4 trafficking by controlling the sorting of GLUT4 from endosomes to specialized insulinregulated vesicles Ingrid Jordens *, Dorothee Molle *, Wenyong Xiong *, Susanna R.
IRAP is a key regulator of GLUT4 trafficking by controlling the sorting of GLUT4 from endosomes to specialized insulinregulated vesicles Ingrid Jordens *, Dorothee Molle *, Wenyong Xiong *, Susanna R.
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
A pre-docking role for microtubules in insulin-stimulated glucose transporter 4 translocation
 A pre-docking role for microtubules in insulin-stimulated glucose transporter 4 translocation Yu Chen*, Yan Wang*, Wei Ji*, Pingyong Xu and Tao Xu National Key Laboratory of Biomacromolecules, Institute
A pre-docking role for microtubules in insulin-stimulated glucose transporter 4 translocation Yu Chen*, Yan Wang*, Wei Ji*, Pingyong Xu and Tao Xu National Key Laboratory of Biomacromolecules, Institute
/07/$15.00/0 Molecular Endocrinology 21(12): Printed in U.S.A.
: Printed in U.S.A.") 0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
phosphatidylinositol 4,5-bisphosphate (PI(4,5)P 2 ) and increases intracellular phosphatidylinositol 3,4,5-trisphosphate
P 2 ) and increases intracellular phosphatidylinositol 3,4,5-trisphosphate") THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 10, pp. 6991 6999, March 2, 2012 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. Regulation of Insulin
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 10, pp. 6991 6999, March 2, 2012 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. Regulation of Insulin
Demonstration of insulin-responsive trafficking of GLUT4 and vptr in fibroblasts
 Journal of Cell Science 113, 4065-4076 (2000) Printed in Great Britain The Company of Biologists Limited 2000 JCS1716 4065 Demonstration of insulin-responsive trafficking of GLUT4 and vptr in fibroblasts
Journal of Cell Science 113, 4065-4076 (2000) Printed in Great Britain The Company of Biologists Limited 2000 JCS1716 4065 Demonstration of insulin-responsive trafficking of GLUT4 and vptr in fibroblasts
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Requires Signaling though Akt2 Independent of the. Transcription Factors FoxA2, FoxO1, and SREBP1c
 Cell Metabolism, Volume 14 Supplemental Information Postprandial Hepatic Lipid Metabolism Requires Signaling though Akt2 Independent of the Transcription Factors FoxA2, FoxO1, and SREBP1c Min Wan, Karla
Cell Metabolism, Volume 14 Supplemental Information Postprandial Hepatic Lipid Metabolism Requires Signaling though Akt2 Independent of the Transcription Factors FoxA2, FoxO1, and SREBP1c Min Wan, Karla
One of the primary actions of insulin on adipose and
 DIABETES-INSULIN-GLUCAGON-GASTROINTESTINAL Rab5 Activity Regulates GLUT4 Sorting Into Insulin- Responsive and Non-Insulin-Responsive Endosomal Compartments: A Potential Mechanism for Development of Insulin
DIABETES-INSULIN-GLUCAGON-GASTROINTESTINAL Rab5 Activity Regulates GLUT4 Sorting Into Insulin- Responsive and Non-Insulin-Responsive Endosomal Compartments: A Potential Mechanism for Development of Insulin
Signaling pathways in insulin action: molecular targets of insulin resistance
 Signaling pathways in insulin action: molecular targets of insulin resistance Jeffrey E. Pessin, Alan R. Saltiel J Clin Invest. 2000;106(2):165-169. https://doi.org/10.1172/jci10582. Perspective Insulin
Signaling pathways in insulin action: molecular targets of insulin resistance Jeffrey E. Pessin, Alan R. Saltiel J Clin Invest. 2000;106(2):165-169. https://doi.org/10.1172/jci10582. Perspective Insulin
Characterization of AS160 activity and its regulation by Akt
 Chapter 2 Characterization of AS160 activity and its regulation by Akt Introduction Insulin stimulated glucose transport plays a central role in maintaining whole-body glucose homeostasis [1, 2]. The hormone
Chapter 2 Characterization of AS160 activity and its regulation by Akt Introduction Insulin stimulated glucose transport plays a central role in maintaining whole-body glucose homeostasis [1, 2]. The hormone
Development of a new model system to dissect isoform specific Akt signalling in adipocytes
 Biochem. J. (2015) 468, 425 434 doi:10.1042/bj20150191 425 Development of a new model system to dissect isoform specific Akt signalling in adipocytes Esi Kajno*, Timothy E. McGraw* and Eva Gonzalez* 1
Biochem. J. (2015) 468, 425 434 doi:10.1042/bj20150191 425 Development of a new model system to dissect isoform specific Akt signalling in adipocytes Esi Kajno*, Timothy E. McGraw* and Eva Gonzalez* 1
THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS
 Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Gapex-5, a Rab31 Guanine Nucleotide Exchange Factor that Regulates Glut4 Trafficking in Adipocytes
 Article Gapex-5, a Rab31 Guanine Nucleotide Exchange Factor that Regulates Glut4 Trafficking in Adipocytes Irfan J. Lodhi, 1,3 Shian-Huey Chiang, 1 Louise Chang, 1 Daniel Vollenweider, 4 Robert T. Watson,
Article Gapex-5, a Rab31 Guanine Nucleotide Exchange Factor that Regulates Glut4 Trafficking in Adipocytes Irfan J. Lodhi, 1,3 Shian-Huey Chiang, 1 Louise Chang, 1 Daniel Vollenweider, 4 Robert T. Watson,
G(alpha)11 signaling through ARF6 regulates F- actin mobilization and GLUT4 glucose transporter translocation to the plasma membrane
11 signaling through ARF6 regulates F- actin mobilization and GLUT4 glucose transporter translocation to the plasma membrane") University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 7-5-2001 G(alpha)11 signaling through ARF6 regulates F- actin mobilization and
University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 7-5-2001 G(alpha)11 signaling through ARF6 regulates F- actin mobilization and
The clathrin adaptor Numb regulates intestinal cholesterol. absorption through dynamic interaction with NPC1L1
 The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
Project report October 2012 March 2013 CRF fellow: Principal Investigator: Project title:
 Project report October 2012 March 2013 CRF fellow: Gennaro Napolitano Principal Investigator: Sergio Daniel Catz Project title: Small molecule regulators of vesicular trafficking to enhance lysosomal exocytosis
Project report October 2012 March 2013 CRF fellow: Gennaro Napolitano Principal Investigator: Sergio Daniel Catz Project title: Small molecule regulators of vesicular trafficking to enhance lysosomal exocytosis
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway
 Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Supplementary Material and Methods
 Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
What would you observe if you fused a G1 cell with a S cell? A. Mitotic and pulverized chromosomes. B. Mitotic and compact G1 chromosomes.
 What would you observe if you fused a G1 cell with a S cell? A. Mitotic and pulverized chromosomes. B. Mitotic and compact G1 chromosomes. C. Mostly non-compact G1 chromosomes. D. Compact G1 and G2 chromosomes.
What would you observe if you fused a G1 cell with a S cell? A. Mitotic and pulverized chromosomes. B. Mitotic and compact G1 chromosomes. C. Mostly non-compact G1 chromosomes. D. Compact G1 and G2 chromosomes.
Supplemental information contains 7 movies and 4 supplemental Figures
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
Running title: Trafficking kinetics of Glut4 and LRP1 in fibroblasts and adipocytes
 INSULIN-REGULATED GLUT4 TRANSLOCATION: MEMBRANE PROTEIN TRAFFICKING WITH SIX DISTINCTIVE STEPS * Paul Duffield Brewer, Estifanos N. Habtemichael, Irina Romenskaia, Cynthia Corley Mastick, and Adelle C.F.
INSULIN-REGULATED GLUT4 TRANSLOCATION: MEMBRANE PROTEIN TRAFFICKING WITH SIX DISTINCTIVE STEPS * Paul Duffield Brewer, Estifanos N. Habtemichael, Irina Romenskaia, Cynthia Corley Mastick, and Adelle C.F.
How many signals impinge on GLUT4 activation by insulin?
 Cellular Signalling 19 (2007) 1 7 www.elsevier.com/locate/cellsig Review How many signals impinge on GLUT4 activation by insulin? Aibin He, Xiaojun Liu, Lizhong Liu, Yongsheng Chang, Fude Fang National
Cellular Signalling 19 (2007) 1 7 www.elsevier.com/locate/cellsig Review How many signals impinge on GLUT4 activation by insulin? Aibin He, Xiaojun Liu, Lizhong Liu, Yongsheng Chang, Fude Fang National
Nature Immunology: doi: /ni.3631
 Supplementary Figure 1 SPT analyses of Zap70 at the T cell plasma membrane. (a) Total internal reflection fluorescent (TIRF) excitation at 64-68 degrees limits single molecule detection to 100-150 nm above
Supplementary Figure 1 SPT analyses of Zap70 at the T cell plasma membrane. (a) Total internal reflection fluorescent (TIRF) excitation at 64-68 degrees limits single molecule detection to 100-150 nm above
Samuel W. Cushman 2 and Karin G. Stenkula 2,4. Kidney Diseases; National Institutes of Health, Bethesda, MD, U.S.A.
 Page 1 of 25 1 Impaired Tethering and Fusion of GLUT4 Vesicles in Insulin-resistant Human Adipose Cells Vladimir A. Lizunov 1, Jo-Ping Lee 2,*, Monica C. Skarulis 3, Joshua Zimmerberg 1, Samuel W. Cushman
Page 1 of 25 1 Impaired Tethering and Fusion of GLUT4 Vesicles in Insulin-resistant Human Adipose Cells Vladimir A. Lizunov 1, Jo-Ping Lee 2,*, Monica C. Skarulis 3, Joshua Zimmerberg 1, Samuel W. Cushman
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
Rip11 is a Rab11- and AS160-RabGAP-binding protein required for insulin-stimulated glucose uptake in adipocytes
 Research Article 4197 Rip11 is a Rab11- and AS160-RabGAP-binding protein required for insulin-stimulated glucose uptake in adipocytes Gavin I. Welsh 1, *, Sophie E. Leney 1, *, Bethan Lloyd-Lewis 1, Matthew
Research Article 4197 Rip11 is a Rab11- and AS160-RabGAP-binding protein required for insulin-stimulated glucose uptake in adipocytes Gavin I. Welsh 1, *, Sophie E. Leney 1, *, Bethan Lloyd-Lewis 1, Matthew
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras
-based studies of receptor dynamics in living cells with Berthold s Mithras") Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson
 Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
Maintaining a normal blood glucose level is essential for
 pubs.acs.org/biochemistry Endocytosis, Recycling, and Regulated Exocytosis of Glucose Transporter 4 Kevin Foley,, Shlomit Boguslavsky, and Amira Klip*,, Cell Biology Program, The Hospital for Sick Children,
pubs.acs.org/biochemistry Endocytosis, Recycling, and Regulated Exocytosis of Glucose Transporter 4 Kevin Foley,, Shlomit Boguslavsky, and Amira Klip*,, Cell Biology Program, The Hospital for Sick Children,
The elements of G protein-coupled receptor systems
 The elements of G protein-coupled receptor systems Prostaglandines Sphingosine 1-phosphate a receptor that contains 7 membrane-spanning domains a coupled trimeric G protein which functions as a switch
The elements of G protein-coupled receptor systems Prostaglandines Sphingosine 1-phosphate a receptor that contains 7 membrane-spanning domains a coupled trimeric G protein which functions as a switch
Anti-diabetic Activity of a Polyphenol-rich Extract from Phellinus. igniarius in KK-Ay Mice with Spontaneous Type 2 Diabetes Mellitus
 Electronic Supplementary Material (ESI) for Food & Function. This journal is The Royal Society of Chemistry 2017 Anti-diabetic Activity of a Polyphenol-rich Extract from Phellinus igniarius in KK-Ay Mice
Electronic Supplementary Material (ESI) for Food & Function. This journal is The Royal Society of Chemistry 2017 Anti-diabetic Activity of a Polyphenol-rich Extract from Phellinus igniarius in KK-Ay Mice
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/10/471/eaah5085/dc1 Supplementary Materials for Phosphorylation of the exocyst protein Exo84 by TBK1 promotes insulin-stimulated GLUT4 trafficking Maeran Uhm,
www.sciencesignaling.org/cgi/content/full/10/471/eaah5085/dc1 Supplementary Materials for Phosphorylation of the exocyst protein Exo84 by TBK1 promotes insulin-stimulated GLUT4 trafficking Maeran Uhm,
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
 A novel role for myosin II in insulin-stimulated glucose uptake in 3T3-L1 adipocytes By: Paul A. Steimle, F. Kent Fulcher, and Yashomati M. Patel Steimle, P.A., Fulcher, K.F., Patel, Y.M. (2005) A novel
A novel role for myosin II in insulin-stimulated glucose uptake in 3T3-L1 adipocytes By: Paul A. Steimle, F. Kent Fulcher, and Yashomati M. Patel Steimle, P.A., Fulcher, K.F., Patel, Y.M. (2005) A novel
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
KEY CONCEPT QUESTIONS IN SIGNAL TRANSDUCTION
 Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Insulin stimulated glucose uptake in adipocytes requires the co-localization of
 STALL JR., RICHARD, M.S. Investigation of the Interaction Between Myosin IIA and Filamentous Actin During Insulin Stimulated Glucose Uptake in 3T3-L1 Adipocytes. (2012) Directed by Dr. Yashomati M. Patel
STALL JR., RICHARD, M.S. Investigation of the Interaction Between Myosin IIA and Filamentous Actin During Insulin Stimulated Glucose Uptake in 3T3-L1 Adipocytes. (2012) Directed by Dr. Yashomati M. Patel
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
* * 11. GnRH. Molecular Biology of the Cell 5th Ed. Alberts et al., Garland Science, 2008
 I e-mail: okay@bs.s.u-tokyo.ac.jp http://www.bs.s.u-tokyo.ac.jp/~naibunpi/lab.html Molecular Biology of the Cell 5th Ed. Alberts et al., Garland Science, 2008 Ion Channels of Excitable Membranes 3rd Ed.
I e-mail: okay@bs.s.u-tokyo.ac.jp http://www.bs.s.u-tokyo.ac.jp/~naibunpi/lab.html Molecular Biology of the Cell 5th Ed. Alberts et al., Garland Science, 2008 Ion Channels of Excitable Membranes 3rd Ed.
THE MECHANISMS USED by insulin to regulate the
 0013-7227/00/$03.00/0 Vol. 141, No. 11 Endocrinology Printed in U.S.A. Copyright 2000 by The Endocrine Society Effects of Adenoviral Gene Transfer of Wild-Type, Constitutively Active, and Kinase-Defective
0013-7227/00/$03.00/0 Vol. 141, No. 11 Endocrinology Printed in U.S.A. Copyright 2000 by The Endocrine Society Effects of Adenoviral Gene Transfer of Wild-Type, Constitutively Active, and Kinase-Defective
CONTRACTING ORGANIZATION: Rush University Medical Center Chicago, IL 60612
 AD Award Number: W81XWH-11-1-0302 TITLE: Yin and Yang of Heparanase in breast Tumor Initiation PRINCIPAL INVESTIGATOR: Xiulong Xu, Ph.D. CONTRACTING ORGANIZATION: Rush University Medical Center Chicago,
AD Award Number: W81XWH-11-1-0302 TITLE: Yin and Yang of Heparanase in breast Tumor Initiation PRINCIPAL INVESTIGATOR: Xiulong Xu, Ph.D. CONTRACTING ORGANIZATION: Rush University Medical Center Chicago,
PHSI3009 Frontiers in Cellular Physiology 2017
 Overview of PHSI3009 L2 Cell membrane and Principles of cell communication L3 Signalling via G protein-coupled receptor L4 Calcium Signalling L5 Signalling via Growth Factors L6 Signalling via small G-protein
Overview of PHSI3009 L2 Cell membrane and Principles of cell communication L3 Signalling via G protein-coupled receptor L4 Calcium Signalling L5 Signalling via Growth Factors L6 Signalling via small G-protein
L6 GLUT4myc Cell Growth Protocol
 L6 GLUT4myc Cell Growth Protocol Background: Parental L6 cells selected for high fusion (2, 3) were stably transfected with a rat GLUT4 cdna carrying a myc epitope (recognized by the commercially available
L6 GLUT4myc Cell Growth Protocol Background: Parental L6 cells selected for high fusion (2, 3) were stably transfected with a rat GLUT4 cdna carrying a myc epitope (recognized by the commercially available
Homework Hanson section MCB Course, Fall 2014
 Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
Vesicle Transport. Vesicle pathway: many compartments, interconnected by trafficking routes 3/17/14
 Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus
 Emerging Science Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus George Wolf Insulin resistance is defined as the reduced responsiveness to normal circulating
Emerging Science Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus George Wolf Insulin resistance is defined as the reduced responsiveness to normal circulating
04_polarity. The formation of synaptic vesicles
 Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
3 The abbreviations used are: GAP, GTPase-activating protein; AICAR, 5 -aminoimidazole-4-carboxamide
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 14, pp. 9187 9195, April 4, 2008 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Inhibition of GLUT4 Translocation
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 14, pp. 9187 9195, April 4, 2008 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Inhibition of GLUT4 Translocation
Insulin-induced GLUT4 Movements in C2C12 Myoblasts: Evidence against a Role of Conventional Kinesin Motor Proteins
 Kobe J. Med. Sci., Vol. 54, No. 1, pp. E14-E22, 2008 Insulin-induced GLUT4 Movements in C2C12 Myoblasts: Evidence against a Role of Conventional Kinesin Motor Proteins KAZUO TAKAZAWA, TETSUYA NOGUCHI,
Kobe J. Med. Sci., Vol. 54, No. 1, pp. E14-E22, 2008 Insulin-induced GLUT4 Movements in C2C12 Myoblasts: Evidence against a Role of Conventional Kinesin Motor Proteins KAZUO TAKAZAWA, TETSUYA NOGUCHI,
Patrick G. P. Foran, Laura M. Fletcher, Paru B. Oatey, Nadiem Mohammed, J. Oliver Dolly, and Jeremy M. Tavaré
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 40, Issue of October 1, pp. 28087 28095, 1999 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Protein Kinase
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 40, Issue of October 1, pp. 28087 28095, 1999 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Protein Kinase
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human
 and HeLa (bottom) cells expressing for 24 h human") Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Integrin CD11b negatively regulates TLR-triggered inflammatory responses by. activating Syk and promoting MyD88 and TRIF degradation via cbl-b
 Integrin CD11b negatively regulates TLR-triggered inflammatory responses by activating Syk and promoting MyD88 and TRIF degradation via cbl-b Chaofeng Han, Jing Jin, Sheng Xu, Haibo Liu, Nan Li, and Xuetao
Integrin CD11b negatively regulates TLR-triggered inflammatory responses by activating Syk and promoting MyD88 and TRIF degradation via cbl-b Chaofeng Han, Jing Jin, Sheng Xu, Haibo Liu, Nan Li, and Xuetao
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
7.06 Spring of PROBLEM SET #6
 7.6 Spring 23 1 of 6 7.6 PROBLEM SET #6 1. You are studying a mouse model of hypercholesterolemia, a disease characterized by high levels of cholesterol in the blood. In normal cells, LDL particles in
7.6 Spring 23 1 of 6 7.6 PROBLEM SET #6 1. You are studying a mouse model of hypercholesterolemia, a disease characterized by high levels of cholesterol in the blood. In normal cells, LDL particles in
Chapter 13: Vesicular Traffic
 Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
7.06 Cell Biology EXAM #3 April 24, 2003
 7.06 Spring 2003 Exam 3 Name 1 of 8 7.06 Cell Biology EXAM #3 April 24, 2003 This is an open book exam, and you are allowed access to books and notes. Please write your answers to the questions in the
7.06 Spring 2003 Exam 3 Name 1 of 8 7.06 Cell Biology EXAM #3 April 24, 2003 This is an open book exam, and you are allowed access to books and notes. Please write your answers to the questions in the
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Edens and Levy, http://www.jcb.org/cgi/content/full/jcb.201406004/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Nuclear shrinking does not depend on the cytoskeleton
Supplemental material Edens and Levy, http://www.jcb.org/cgi/content/full/jcb.201406004/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Nuclear shrinking does not depend on the cytoskeleton
CaMKII-Mediated Phosphorylation of the Myosin Motor Myo1c Is Required for Insulin-Stimulated GLUT4 Translocation in Adipocytes
 Article CaMKII-Mediated Phosphorylation of the Myosin Motor Myo1c Is Required for Insulin-Stimulated GLUT4 Translocation in Adipocytes Ming Fai Yip, 1 Georg Ramm, 1 Mark Larance, 1,2 Kyle L. Hoehn, 1 Mark
Article CaMKII-Mediated Phosphorylation of the Myosin Motor Myo1c Is Required for Insulin-Stimulated GLUT4 Translocation in Adipocytes Ming Fai Yip, 1 Georg Ramm, 1 Mark Larance, 1,2 Kyle L. Hoehn, 1 Mark
Role of the Protein Kinase TBK1 in Insulin-Stimulated Glucose Transport. Maeran Uhm
 Role of the Protein Kinase TBK1 in Insulin-Stimulated Glucose Transport by Maeran Uhm A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Molecular
Role of the Protein Kinase TBK1 in Insulin-Stimulated Glucose Transport by Maeran Uhm A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Molecular
High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart.
 in the heart.") High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart. Daniel Aston, Rebecca A. Capel, Kerrie L. Ford, Helen C. Christian,
High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart. Daniel Aston, Rebecca A. Capel, Kerrie L. Ford, Helen C. Christian,
COORDINATION OF CELL SIGNALING AND TRANSPORT MACHINERY DURING INSULIN-STIMULATED GLUCOSE TRANSPORT. Xiao-wei Chen
 COORDINATION OF CELL SIGNALING AND TRANSPORT MACHINERY DURING INSULIN-STIMULATED GLUCOSE TRANSPORT By Xiao-wei Chen A dissertation submitted in partial fulfillment of the requirements for the degree of
COORDINATION OF CELL SIGNALING AND TRANSPORT MACHINERY DURING INSULIN-STIMULATED GLUCOSE TRANSPORT By Xiao-wei Chen A dissertation submitted in partial fulfillment of the requirements for the degree of
Cell Cycle, Mitosis, and Microtubules. LS1A Final Exam Review Friday 1/12/07. Processes occurring during cell cycle
 Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic
 Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
genome edited transient transfection, CMV promoter
 Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
Supplementary Figure 1. In the absence of new protein translation, overexpressed caveolin-1-gfp is degraded faster than caveolin-1-gfp expressed from the endogenous caveolin 1 locus % loss of total caveolin-1-gfp
Molecular Trafficking
 SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
endomembrane system internal membranes origins transport of proteins chapter 15 endomembrane system
 endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
Abstract Introduction
 Decreased colocalization of synapsin I and Munc13 within presynaptic axon terminals of the earthworm neuromuscular junction when stimulated could help determine how the two proteins interact during neurotransmitter
Decreased colocalization of synapsin I and Munc13 within presynaptic axon terminals of the earthworm neuromuscular junction when stimulated could help determine how the two proteins interact during neurotransmitter
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Biochimie 93 (2011) 697e709. Contents lists available at ScienceDirect. Biochimie. journal homepage:
 697e709. Contents lists available at ScienceDirect. Biochimie. journal homepage:") Biochimie 93 (2011) 697e709 Contents lists available at ScienceDirect Biochimie journal homepage: www.elsevier.com/locate/biochi Research paper Dimethyl sulfoxide enhances GLUT4 translocation through a
Biochimie 93 (2011) 697e709 Contents lists available at ScienceDirect Biochimie journal homepage: www.elsevier.com/locate/biochi Research paper Dimethyl sulfoxide enhances GLUT4 translocation through a
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
The rabbit femoral artery was prepared and each arterial ring was permeabilized
 Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Supplemental Materials Molecular Biology of the Cell
 Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
Supplemental Materials. STK16 regulates actin dynamics to control Golgi organization and cell cycle
 Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
STEIN IN-TERM EXAM -- BIOLOGY FEBRUARY 16, PAGE
 STEIN IN-TERM EXAM -- BIOLOGY 3058 -- FEBRUARY 16, 2017 -- PAGE 1 of 9 There are 25 questions in this Biology 3058 exam. All questions are "A, B, C, D, E, F, G, H" questions worth one point each. There
STEIN IN-TERM EXAM -- BIOLOGY 3058 -- FEBRUARY 16, 2017 -- PAGE 1 of 9 There are 25 questions in this Biology 3058 exam. All questions are "A, B, C, D, E, F, G, H" questions worth one point each. There
The SUMO Conjugating Enzyme Ubc9 is a Regulator of GLUT4 Turnover and Targeting to the Insulin-Responsive Storage Compartment in 3T3-L1 Adipocytes
 Diabetes In Press, published online May 29, 2007 The SUMO Conjugating Enzyme Ubc9 is a Regulator of GLUT4 Turnover and Targeting to the Insulin-Responsive Storage Compartment in 3T3-L1 Adipocytes Received
Diabetes In Press, published online May 29, 2007 The SUMO Conjugating Enzyme Ubc9 is a Regulator of GLUT4 Turnover and Targeting to the Insulin-Responsive Storage Compartment in 3T3-L1 Adipocytes Received
TRANSPORT ACROSS THE plasma membrane is the initial
 0013-7227/02/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 87(2):716 723 Printed in U.S.A. Copyright 2002 by The Endocrine Society PKC- Mediates Insulin Effects on Glucose Transport in Cultured
0013-7227/02/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 87(2):716 723 Printed in U.S.A. Copyright 2002 by The Endocrine Society PKC- Mediates Insulin Effects on Glucose Transport in Cultured
SUPPLEMENTARY INFORMATION
 In the format provided by the authors and unedited. 2 3 4 DOI: 10.1038/NMAT4893 EGFR and HER2 activate rigidity sensing only on rigid matrices Mayur Saxena 1,*, Shuaimin Liu 2,*, Bo Yang 3, Cynthia Hajal
In the format provided by the authors and unedited. 2 3 4 DOI: 10.1038/NMAT4893 EGFR and HER2 activate rigidity sensing only on rigid matrices Mayur Saxena 1,*, Shuaimin Liu 2,*, Bo Yang 3, Cynthia Hajal
Supplementary Figure S1 (a) (b)
 (b)") Supplementary Figure S1: IC87114 does not affect basal Ca 2+ level nor nicotineinduced Ca 2+ influx. (a) Bovine chromaffin cells were loaded with Fluo-4AM (1 μm) in buffer A containing 0.02% of pluronic
Supplementary Figure S1: IC87114 does not affect basal Ca 2+ level nor nicotineinduced Ca 2+ influx. (a) Bovine chromaffin cells were loaded with Fluo-4AM (1 μm) in buffer A containing 0.02% of pluronic
BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented
 Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or
 Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Figure S1. Western blot analysis of clathrin RNA interference in human DCs Human immature DCs were transfected with 100 nm Clathrin SMARTpool or control nontargeting sirnas. At 90 hr after transfection,
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Lecture 36: Review of membrane function
 Chem*3560 Lecture 36: Review of membrane function Membrane: Lipid bilayer with embedded or associated proteins. Bilayers: 40-70% neutral phospholipid 10-20% negative phospholipid 10-30% cholesterol 10-30%
Chem*3560 Lecture 36: Review of membrane function Membrane: Lipid bilayer with embedded or associated proteins. Bilayers: 40-70% neutral phospholipid 10-20% negative phospholipid 10-30% cholesterol 10-30%
Signal Transduction Pathways. Part 2
 Signal Transduction Pathways Part 2 GPCRs G-protein coupled receptors > 700 GPCRs in humans Mediate responses to senses taste, smell, sight ~ 1000 GPCRs mediate sense of smell in mouse Half of all known
Signal Transduction Pathways Part 2 GPCRs G-protein coupled receptors > 700 GPCRs in humans Mediate responses to senses taste, smell, sight ~ 1000 GPCRs mediate sense of smell in mouse Half of all known
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
Supplemental information
 Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental
Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental