Hedgehog signaling plays a critical role in directing correct primordial follicle formation in the mouse ovary
|
|
|
- Merry Lang
- 5 years ago
- Views:
Transcription
1 Hedgehog signaling plays a critical role in directing correct primordial follicle formation in the mouse ovary Honors Thesis Presented to the College of Agriculture and Life Sciences, Department of Animal Science of Cornell University in Partial Fulfillment of the Requirements for the Research Honors Program by Heather Brianne Phillips May 2007 Research Mentor, Dr. Susan Quirk
2 Abstract: The Hedgehog (HH) signaling pathway has been identified to play an essential role in cell differentiation, cell fate determination and cell proliferation in both Drosophila and mammalian models. While much is known regarding HH actions in the Drosophila ovary, its role in the developing mammalian ovary remains a mystery. In the days following birth, the mouse ovary transitions from an organ filled with ovigerous cords to one filled with primordial follicles, which are oocytes surrounded by a layer of flattened pre-granulosa epithelial cells, enclosed by a basement membrane, and associated with mesenchymal cells. The purpose of this study was to determine the role HH plays in primordial follicle formation in the mouse ovary. HH signaling was blocked by injecting cyclopamine, a plant alkaloid known to inhibit HH signaling, on days 0, 1, 2 and 3 after birth. Immunohistochemistry using an antibody for laminin, a common basement membrane protein, revealed a significantly higher number of discontinuous basement membranes around primordial follicles in ovaries extracted from cyclopamine-treated mice. H&E staining revealed a significantly lower ratio of mesenchymal cells to oocytes in ovaries from cyclopamine-treated mice, as compared to controls. The reduced HH signaling in this study is thought to have disrupted proper ovigerous cord breakdown and interfered with normal cell communication among oocytes, epithelial cells and mesenchymal cells, suggesting that the HH signaling pathway does play a critical role in directing correct primordial follicle development. This study provides valuable insight into the role HH plays in the developing mouse ovary. 2
3 Acknowledgements: Many thanks to Dr. Quirk for all her guidance and advice throughout the project. Her suggestions, input and editing of the thesis were critical to this project. Many thanks to Bob Cowan for all his help with the methods and laboratory procedures. Without his help this project would not have been possible. Thanks to the Quirk lab for providing the mice, laboratory equipment and supplies for this project. Thanks to the Histology Core Facility, Cornell University, NYS College of Veterinary Medicine for sectioning tissues used in this project. Thanks to the Dextra Undergraduate Research Endowment Fund and the CALS Charitable Trust Grant for providing funding for this project. 3
4 Table of Contents: Abstract. 2 Acknowledgements... 3 Introduction... 5 Literature Review...6 Materials and Methods..12 Results Discussion. 23 Literature Cited. 30 4
5 Introduction: In mice, the formation of ovarian follicles occurs during the first few days of life. At birth, the mouse ovary contains structures referred to as ovigerous cords which are membrane-bound sacs of oocytes and epithelial cells surrounded by mesenchymal cells. Functional and structural organization of the ovary occurs during the first 4 to 8 days postpartum. During this time, ovigerous cords breakdown and somatic cells associate with oocytes to form primordial follicles [1]. Primordial follicles are composed of a quiescent oocyte surrounded by a flattened layer of pre-granulosa cells, enclosed by a basement membrane. Mesenchymal cells in the region surrounding primordial follicles are eventually incorporated into an outer theca layer as the follicle grows and develops [1]. Primordial follicle formation is directed by cell to cell signaling by locally produced autocrine and paracrine growth factors [2]. Inappropriate expression and action of these factors can lead to incomplete cord break down and altered ovarian physiology [2]. Primordial follicles represent the total follicular reserve. This is the total number of follicles available for development and ovulation during a female s lifespan, and as such is a critical indicator of fertility. The HH signaling pathway was first identified in Drosophila, and was shown to play an important role in cell differentiation, cell fate determination and cell proliferation. It was also shown to be essential for proper ovarian function. In addition, the HH pathway was found to play an important role in numerous aspects of development in mammals. Its function in the mammalian ovary is just beginning to be studied [3, 4, 5]. In mammals, the HH signaling pathway utilizes three protein ligands: sonic (SHH), desert (DHH) and Indian (IHH) [6]. These ligands bind to the transmembrane receptor 5
6 patched (PTCH). In the absence of the HH ligand, PTCH blocks the activity of the transmembrane signal transducer protein smoothened (SMO) [7]. The binding of HH to PTCH removes the inhibition of SMO, resulting in activation of downstream transcription factors that then alter gene transcription [7]. Results of several studies have demonstrated that a functional HH pathway is present in the theca and granulosa cells of the mouse ovary [3, 4]. Research to date has focused on the role of HH in developing follicles in the immature and adult ovary [3, 4, 5]. Yet, components of the HH signaling pathway are also expressed in the newborn ovary [3], suggesting a potential role for HH in the formation of primordial follicles. Preliminary data from Dr. Quirk s lab suggest that early follicle development in the neonatal ovary is altered when HH signaling is reduced experimentally. The goal of the research reported in this thesis was to determine the role of the HH signaling pathway in the development of the mouse ovary. The strategy used was to examine the effects of reducing HH signaling in the neonatal mouse ovary on the assembly of primordial follicles. The hypothesis tested is that HH signaling is required to direct correct primordial follicle formation. Literature Review: The mouse ovary is incompletely developed at the time of birth. During the first 4 to 8 days postpartum the ovary undergoes substantial functional and structural organization as it completes development. This process involves the rearrangement of associations among somatic cells and germ cells to form primordial follicles. This is a dynamic process that is just beginning to be fully understood, but is believed to be directed by a variety of cell 6
7 signaling pathways. The focus of the research reported in this thesis is to determine whether the hedgehog (HH) signaling pathway is involved in this early ovarian development. Hedgehog Signaling The HH signaling pathway was first found in Drosophila while screening for mutations that disrupt the larval body plan [6]. Since its discovery, HH has been studied extensively in Drosophila and shown to be important for development of wings, legs, eyes, abdomen, gut, trachea and gonads [6]. In the gonad, it is involved in germ cell migration, cell proliferation and polarization [6]. The HH pathway also plays an important role in mammals. It is essential for proper embryonic development and adult tissue maintenance. In the embryo, HH is involved in the development of multiple organs by regulating cell proliferation and differentiation, as well as directing cell fate [6]. In adult tissue, the HH pathway can be reactivated by tissue damage and stimulates tissue repair [8]. HH signals are received, transmitted and interpreted using pathways that have been highly conserved during the divergent evolution of insects and mammals [7]. While only one HH ligand has been found in Drosophila, three HH ligands exist in the mammal: sonic (SHH), Indian (IHH), and desert (DHH), with the DHH ligand being most closely related to the Drosophila ligand [6]. All three HH ligands are secreted proteins that bind to the transmembrane receptor patched (PTCH). In the absence of the HH ligand, PTCH blocks the activity of the transmembrane signal transducer protein smoothened (SMO) [7]. The binding of HH to PTCH removes the inhibition of SMO. SMO is then able to undergo a 7
8 conformational change that allows it to activate a downstream group of transcription factors which act to alter target gene expression [7]. In Drosophila, the target transcription factor is the segment-polarity gene: cubits interruptus (Ci) [7]. In resting cells, Ci is phosphorylated, which serves to repress transcription of target genes. In the presence of HH signaling, however, Ci is not phosphorylated and is able to activate transcription of HH target genes [7]. In mammals transcription factors include three Ci-related proteins from the GLI-Kruppel family, GLI1, GLI2, and GLI3, which act as either repressors or activators depending on posttranslational processing [6]. GLI1 serves as an activator of HH target gene transcription, while GLI3 acts as a repressor of both HH target gene transcription and Shh expression [6]. HH target genes include Ptch and Gli1 [6], suggesting that the HH signaling pathway works to modulate its own activity. Disruption of HH signaling has been shown to have devastating effects upon an individual. In mice, the absence of HH signaling prenatally results in embryo lethality [9]. Mutation and deregulation of the HH pathway plays a role in tumorigenesis, with over expression of the HH pathway being linked to basal cell carcinomas and other cancers [8]. Furthermore, in male mice in which the Dhh gene has been deleted, abnormal testes development and sterility are observed [10, 11]. In Drosophila, HH signaling stimulates proliferation of ovarian somatic stem cells, promotes polarization of follicle cells, and migration of the primordial germ cells to the gonads in the embryo [6, 12, 13]. Since HH is essential for proper ovarian function in Drosophila, it has been postulated that it is also essential in the mammalian ovary. 8
9 Primordial Follicle Formation in the Mouse Ovary At birth, the mouse ovary consists of structures called ovigerous cords, which are basement-membrane-enclosed sacs of oocytes and epithelial-type somatic cells surrounded by mesenchymal-type cells [1]. The basement membrane surrounding the cords is not thought to be a permanent continuous structure, and as such, intermingling between cells in different regions may occur [1]. The oocytes within the cords are interconnected via intracellular bridges, creating a cyst-like structure [13]. Research suggests this cyst-like structure is the result of synchronous division of a progenitor germ cell during the embryonic time period [13]. Cyst breakdown occurs around the time of birth; however before it occurs mitochondria migrate via intracellular bridges to adjacent oocytes [14]. It has been postulated that this ensures that oocytes destined for future primordial follicle formation acquire functional mitochondria [14]. During the first few days of life, the ovigerous cords become fragmented and tissue remodeling leads to the formation of the first follicles, primordial follicles. Epithelial-type pre-granulosa cells envelop individual oocytes to form primordial follicles. A primordial follicle is composed of a quiescent oocyte surrounded by a flattened layer of epithelial pregranulosa cells and enclosed by a newly synthesized basement membrane [1]. Mesenchymal cells in the region surrounding primordial follicles eventually become incorporated into an outer theca cell layer of the follicle as it grows and develops, in a process that is poorly understood [1]. Recent research suggests that HH signals from granulosa cells may act on the mesenchymal cells to promote development of the theca [4]. Synthesis of the basement membrane around follicles is a poorly understood process. It has been suggested that the basement membrane components (type I collagen, type IV 9
10 collagen, laminin and fibronectin) may all be primarily synthesized by the theca compartment surrounding the outside of the basement membrane of the follicle [1, 15]. In the testis, it has been suggested that epithelial and mesenchymal cells interact to assemble the basement membrane [16]; however the appearance of a similar mechanism in the ovary has yet to be investigated. Laminin appears to be the only protein present in basement membrane surrounding primordial follicles in mice [17]. In primordial follicles, laminin forms a ring around each granulosa cell compartment, which functions to define the basement membrane that separates the primordial follicle from the surrounding area. Each laminin molecule is composed of one α, one β, and one γ chain, yet multiple varieties of each chain occur. The composition of the laminin chain affects the fate of the associated cells, and thus laminin chain composition varies in different tissues [18]. Previous studies suggest that the α1, β1, and γ1 laminin chains are present in the basement membrane surrounding primordial follicles in cows, pigs and rabbits [18, 19]. Primordial follicle formation does not occur in a uniform pattern throughout the ovary. At birth, ovigerous cords are more densely packed in the medulla of the ovary [20]. These cords contain a high number of oocytes with few epithelial cells. As the cords break down, primordial follicles first develop in the medulla [21]. These first primordial follicles are characterized by round, cubodial pre-granulosa cells. Primordial follicles in the cortex appear a little later and differ in the fact that the pre-granulosa cells are flattened, squamous cells [21]. In the first few days postpartum, it is common to see areas of fully enclosed follicles neighboring areas still exhibiting cord-like patterns [20]. Primordial follicle formation is under the control of cell to cell signaling; however which cells are signaling is still debatable. An old idea suggests that signaling between pre- 10
11 granulosa cells and the oocyte directs primordial follicle formation [22]. A second idea suggests that signals from mesenchymal cells direct primordial follicle formation since their presence is thought to be required for the organization of the basement membrane that encloses the primordial follicle [22]. In general, a basement membrane is thought to be organized when epithelial cell and mesenchymal cell interactions occur, suggesting the possibility that epithelial pre-granulosa cells and mesenchymal cells interact to form the basement membrane that surrounds primordial follicles [22]. Recent research suggests both of these ideas may be correct, with cell signaling occurring between the pre-granulosa cells and oocyte, as well as the pre-granulosa cells and the mesenchymal cells. Primordial follicle formation is coordinated by locally produced paracrine and autocrine growth factors [2, 9]. Interactions between the pre-granulosa cells and the oocyte as well as interactions between the pre-granulosa cells and mesenchymal cells may be needed for proper primordial follicle assembly, such that inappropriate expression and actions of these factors may result in abnormal ovarian physiology [2]. Primordial follicles represent the follicular reserve, the total number of follicles available for development during a female s lifespan, and are thus critical for fertility. Primordial follicles become quiescent once formed. Throughout an individual s lifetime, primordial follicles are continuously recruited from this follicle reserve to undergo further growth and development. Previous studies suggest that the overall number of quiescent follicles in the ovary affects the rate at which follicles are recruited into the growing pool [23]. There is an inverse correlation between the number of primordial follicles and the rate at which they are stimulated to grow [23]. A reduction in the size of the primordial stockpile will result in an increase in the proportion of follicles beginning to grow. If too many 11
12 follicles are recruited at once it could have devastating effects on fertility, as the animal may run out of follicles prematurely. A complex network of cell to cell signaling, including the actions of locally produced growth factors, is required for the transition from primordial to a growing primary follicle [2]. If disruption of these signals occur, potentially too many primordial follicles could be recruited into the growing pool leading to depletion of the follicle reserve. Hedgehog s Role in the Mouse Ovary The HH pathway has previously been found to be expressed in the adult mouse ovary. Granulosa cells express all three HH ligands, while granulosa and theca cells express PTCH and SMO [3]. The oocyte is thought to be unable to respond to HH because it doesn t express Smo [4]. Granulosa cells of primary follicles have been found to secrete HH, which can act upon the surrounding pre-theca compartment [4]. Granulosa cells have also been identified as a target for HH signaling, with an increase in HH signaling causing a small increase in granulosa cell proliferation and increased expression of the transcriptional target gene Gli1 [3]. Components of the HH signaling pathway are also expressed in the newborn ovary, with Shh, Ihh and Dhh all being expressed in low levels on day 0 [3]. This suggests a potential role for HH in the formation of primordial follicles. Materials and Methods: Inhibition of HH signaling Cyclopamine is a plant alkaloid and a known pharmacological inhibitor of HH signaling. Cyclopamine (BioMol, Plymouth Meeting, PA) was dissolved in 2-hydroxypropyl- 12
13 β-cyclodextrin solution (Sigma-Aldrich Inc., St. Louis, MO). CD-1 female pups were injected i.p. with two different doses of cyclopamine (2mg/kg or 5mg/kg) or vehicle (2- hydroxypropyl-β-cyclodextrin solution) on days 0 (day of birth), 1, 2 and 3 of age. Ovaries were then isolated on day 4 for analysis. Ovaries were also isolated from uninjected control pups on day 0. In addition, some mice that were treated with either vehicle or high dose cyclopamine from days 0-3 were allowed to reach puberty and were then caged with an adult, fertile CD-1 male. At day 30, ovaries from these mice were collected for analysis. Immunohistochemistry: Day 4 ovaries from vehicle-injected and cyclopamine-injected mice (2mg/kg and 5mg/kg) were fixed in 2% paraformaldehyde. Fixed ovaries were embedded in paraffin wax and 5µM sections were cut by the Histology Core Facility, Cornell University, NYS College of Veterinary Medicine. Sections were deparaffinized in xylene, rehydrated by stepwise incubation in 100% ethanol, 90% ethanol, 70% ethanol, 50% ethanol and then rinsed twice in PBS. Nonspecific binding was blocked at room temperature for 30 minutes in 2% normal goat serum in PBS. Sections were incubated with rabbit polyclonal antiserum (8 µg/ml) raised against laminin-1 isolated from Englebreth Holm-Swarm (EHS) tumors (Sigma- Aldrich Inc., St. Louis, MO) in PBS + 2% BSA in a humidified chamber for 2 hrs at room temperature. Sections were rinsed with PBS four times and then incubated with AlexaFluor555 goat anti-rabbit IgG (0.25 µg/ml; Molecular Probes, Eugene, OR) in PBS + 2% BSA for 1 hr at room temperature. Slides were rinsed 4 times in PBS and then counterstained with Hoechst (Molecular Probes, Eugene, OR). Staining was performed on 13
14 ovary sections from day 4 control and cyclopamine-injected mice, as well as day 0 uninjected control ovaries (n =3 per treatment). Stained sections were evaluated using a fluorescence microscope. Pictures of 4 unique fields were taken from 2 to 4 ovary sections per mouse. The completeness of the laminin rings around primordial follicles was then evaluated in each field, such that each mouse had four fields evaluated. The number of complete and incomplete laminin rings found in the four fields was summed and a percent of incomplete and complete laminin rings was calculated for each mouse. Laminin encapsulation of primordial follicles was considered to be complete if there were no gaps or fragmentation of the laminin ring around the primordial follicle. Laminin encapsulation was scored as incomplete if there were gaps or fragmentation present in the laminin ring. The degree to which the laminin ring was incomplete was then graded on a rubric scale to assign the following scores: 0, presence of a complete laminin ring; 1, presence of a laminin ring around an oocyte that is greater than 50% complete; 2, presence of a laminin ring around an oocyte that is half complete; 3, presence of a laminin ring around an oocyte that is less than 50% complete. Hematoxylin & Eosin staining: Morphology of the developing follicles was evaluated in sections of day 4 ovaries fixed in Bouin s solution and stained with hematoxylin and eosin. Ovaries from three control and three high dose cyclopamine-treated (5mg/kg) mice were evaluated. The number of oocytes, pre-granulosa (epithelial) cells and mesenchymal cells were counted in 4 unique fields per mouse to yield average epithelial: oocyte and mesenchymal: oocyte ratios for each 14
15 mouse. Epithelial cells were identified by their dark, central nucleus and the fact that staining by hematoxylin was lighter than that for mesenchymal cells. Mesenchymal cells were identified by their more flattened shape and darker staining with hematoxylin. Day 30 ovaries from control and high-dose cyclopamine-treated mice (n =3 per treatment) were also stained. The morphology of these ovaries was then observed. RT-PCR for Gli1 expression: In order to determine whether HH signaling had been inhibited through this model, Gli1 expression in the ovary was evaluated. RNA was prepared from whole ovaries from day 4 control and high dose cyclopamine-treated mice (5mg/kg) using a RNeasy MiniKit. Reverse transcription was performed using a High Capacity cdna Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Real time PCR for Gli1 was performed on an ABI Prism 7000 using kits purchased from Applied Biosystems, Foster City, CA (assay number Mm _m1). The assay used spans exons 2 and 3. cdna from RNA will be amplified with this assay, but genomic DNA will not be amplified because of the large introns present in genomic DNA between exons 2 and 3. A standard curve was constructed from cdna prepared from RNA of pooled whole ovaries. 18s rrna concentration was also measured (assay number E). Results were standardized by dividing by the 18s rrna concentration and multiplying by 100. Fertility Evaluation: Some mice that were treated with low and high doses of cyclopamine from days 0-3 were allowed to reach puberty and were then caged with an adult, fertile CD-1 male. 15
16 Litter sizes were recorded for all females that became pregnant. Statistical Analysis: A T-test was used to test the difference in the total number of incomplete laminin rings in control versus cyclopamine-treated mice. A t-test was also used to test differences in the ratios of epithelial cells: oocytes and mesenchymal cells: oocytes in control and cyclopamine-treated mice. The differences in ovarian Gli1 expression in cyclopaminetreated and control mice were tested by a t-test. Results: Quantitative real time RT-PCR was used to evaluate Gli1 expression levels in vehicle-injected and high dose cyclopamine-injected ovaries. This was done to determine if HH signaling had been inhibited by the cyclopamine injections. 18s ribosomal RNA, a housekeeping gene, was used to standardize the values. High dose cyclopamine-treated ovaries had a significantly lower expression level of Gli1 (p= 0.05) (Fig 1). This shows that cyclopamine injections reduced expression of the HH transcriptional target gene Gli1, suggesting that cyclopamine treatment did reduce HH signals. Immunohistochemistry to detect laminin in ovaries of uninjected mice on day 0 revealed the presence of ovigerous cords, as expected. Oocytes were arranged mainly in long 16

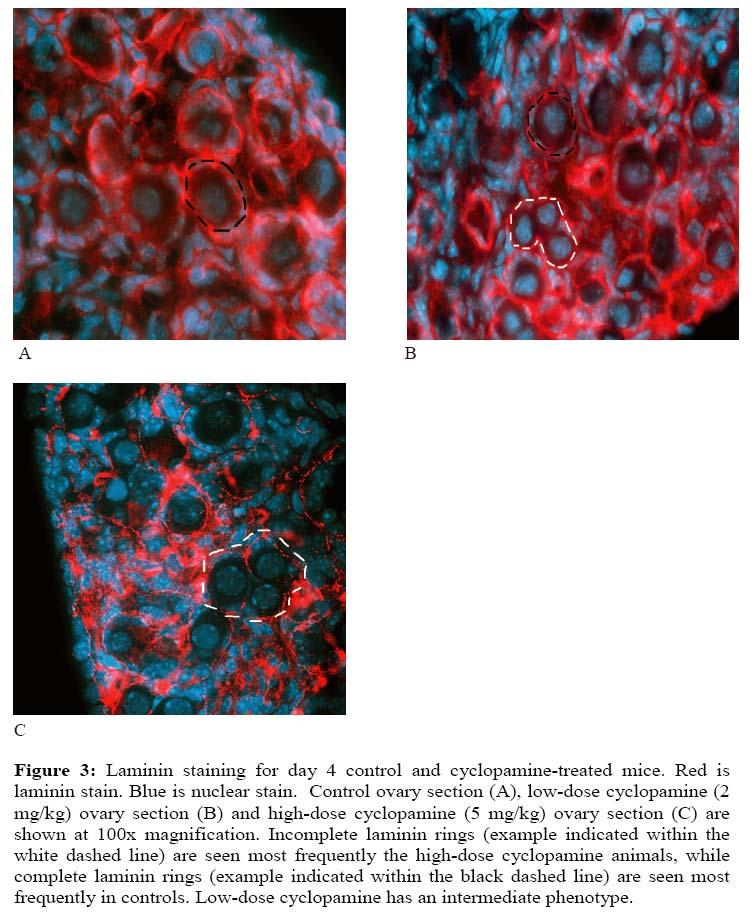
17 chains that were surrounded by somatic cells and enclosed by a laminin-containing basement membrane (Fig 2A). The ovigerous cords varied in size, with sections through some containing only two oocytes and others containing as many as 10 to 15. Ovaries from vehicle-injected mice on day 4 showed expected cord breakdown and primordial follicle development. Single oocytes were surrounded by a flattened ring of somatic cells and a laminin-containing basement membrane (Fig 2B). On average, 69.8% of 17
18 primordial follicles were enclosed by a complete basement membrane, with a score of 0 (Table 1). There were few areas of the ovaries in which basement membranes surrounding primordial follicles were incomplete. Only 3.7% of follicles had basement membranes that were very discontinuous, with a score of 3. Primary follicles were present in day 4 ovaries and were concentrated in the center, medullary region of the ovary. In contrast to control mice, mice treated with low and high doses of cyclopamine had an increased frequency of primordial follicles surrounded by basement membranes that were incomplete. This effect was most pronounced in mice treated with the higher dose of cyclopamine in which 24.9% of follicles had discontinuous membranes with a score of 3 and only 26.1% of follicles had complete basement membranes with a score of 0 (Table 1). As shown in Figure 3C, ovaries consisted mainly of primordial follicles enclosed by incomplete basement membranes with large gaps. In addition, a portion of oocytes seemed to have associations with only a few somatic cells and had minimal amounts of laminin surrounding them. The effect was less pronounced in mice treated with a low dose of cyclopamine (2mg/kg) in which only 10.2% of follicles had discontinuous basement membranes with a score of 3 and 60.8% of follicles had complete basement membranes with a score of 0. As shown in Fig 3B, areas of primordial follicles with complete basement membranes and areas with incomplete basement membranes were both apparent. The phenotype observed in mice treated with the high dose of cyclopamine was variable. Ovaries from two of the mice had the phenotype described above. The ovary from a third mouse had areas of relatively normal primordial follicle development in which oocytes were surrounded by a ring of flattened somatic cells and enclosed by a basement membrane that was often incomplete. However, the same ovary had areas of densely packed 18
19

20 cells containing mostly somatic cells and a few scattered oocytes, surrounded by a basement membrane (Fig 4B). Despite some variation in the phenotype of mice treated with the high dose of cyclopamine, the percentage of follicles with basement membranes that were incomplete was significantly higher than in control mice (p<0.05) (Fig 5). A similar statistical analysis was not possible for mice treated with the low dose of cyclopamine because the sample size was too small (n=2). 20

21 21
 cells (Fig 6).")
22 H&E sections of day 4 ovaries from control and high dose cyclopamine-treated mice (n=3) were evaluated for differences in the ratio of epithelial and mesenchymal cells to oocytes present. In control and cyclopamine-treated mice, primordial follicles were surrounded by a layer of flattened pre-granulosa (epithelial) cells (Fig 6). In controls, a layer of mesenchymal cells is often found surrounding the epithelial layer associated with an individual oocyte (Fig 6A and C). Mesenchymal cells appeared to be interspersed between individual follicles. Contrastingly, in cyclopamine-treated animals mesenchymal cells were most often in long chains or clumps, rather than interspersed between and associating with individual follicles (Fig 6B and D). The epithelial cell to oocyte ratio was not different between controls and cyclopamine-treated mice (p>0.05) (Fig 7). The mesenchymal cell to oocyte ratio was significantly less in cyclopamine-treated mice compared to controls (p<0.05) (Fig 7). Ovaries from day 30 vehicleinjected and high dose cyclopamine injected mice (n =3), stained with H&E, were obersevered to identify any striking morphology abnormalities due to cyclopamine-treatment. Ovaries from both vehicle-injected and high 22
23 dose cyclopamine injected mice had follicles at all stages of development. Some ovaries had corpus luteua present suggesting the mouse had ovulated previously. Neither the number of follicles at each stage of development nor the total number of primordial follicles present was counted. The fertility study conducted found that low dose (n= 6) and high dose (n =1) cyclopamine-treated mice had litter sizes similar to those of vehicle-injected mice (n = 2). Average litter size ranged between 13 and 15 pups for controls, 12 to 15 pups for low dose cyclopamine-treatment and 9 pups for high dose cyclopamine-treatment. These are within the normal range for CD-1 mice suggesting all cyclopamine-treated mice were still fertile and able to become pregnant. Discussion: Primordial follicle formation is a complex process involving multiple cell types and cell signals. The findings of this study suggest that the HH signaling pathway plays an important role in directing correct primordial follicle formation in the mouse. Inhibiting HH signaling through cyclopamine injections resulted in improper primordial follicle formation, as characterized by incomplete basement membranes and altered association of mesenchymal cells with primordial follicles. As expected, control mice showed a normal phenotype, with normal cord breakdown and primordial follicle formation occurring between day 0 and day 4. Day 4 ovaries exhibited proper assembly of primordial follicles. The disappearance of cord structures and the presence of primordial follicles as well as growing primary follicles in 23
24 these mice suggest that the period of primordial follicle formation is almost complete by day 4. An altered phenotype, however, was seen in cyclopamine-treated mice, including poor basement membrane formation around primordial follicles. High dose cyclopaminetreatment resulted in the most severe phenotype, while low-dose cyclopamine treatment resulted in more moderate changes. There was variation in the phenotype in both groups, with the most variation among the high dose cyclopamine-treated mice. The variation in the phenotype in the mice treated with the high dose of cyclopamine suggests that cyclopamine may have affected some animals more drastically than others. The areas of densely packed somatic cells surrounded by basement membranes observed in one mouse may have once been cords in which all the oocytes underwent apoptosis. Further research is needed to determine if cyclopamine treatment causes oocyte apoptosis. This could be accomplished by increasing the number of replicate mice and by studying the time course of the effects after cyclopamine treatment to determine if loss of oocytes occurs. Interestingly, in day 4 ovaries stained for laminin from transgenic mice (Amhr2 Cre/+ smom2) that have a continually activated HH pathway, similar areas of densely-packed somatic cells were observed. In those mice, there are areas containing densely-packed somatic cells with no apparent oocytes enclosed by a basement membrane, as well as areas of relatively normal primordial follicle development. This suggests that altering HH signaling by either depressing it or up-regulating it can have similar effects on follicle development and suggest that precisely regulated HH signaling is required for proper primordial follicle development. 24
25 Since Gli1 expression analysis shows that HH was inhibited in the cyclopaminetreatment model, it can be suggested that reduced HH signaling caused incomplete basement membranes to be formed. While there is no previous study regarding the effects of HH signaling on basement membrane formation in the ovary, prior studies support the idea that Dhh modulates the production of the basal lamina around seminiferous tubules [10]. When HH signaling was inhibited in the testes of Dhh knockout mice the basal lamina around seminiferous tubules contained numerous areas with small gaps, giving the appearance of a discontinuous and incomplete basal lamina [10]. Prior studies have shown all three HH ligands to be present in granulosa cells [3]. Furthermore granulosa and theca cells have been shown to exhibit PTCH and SMO [3], suggesting that HH acts on both granulosa and theca cells. It is thought that theca cells may synthesize basement membrane components [1], when they interact with epithelial cells via cell signaling pathways [22]. The finding that granulosa cells of primary follicles secrete HH, which can then act upon the surrounding mesenchymal-derived theca cells [4], suggests that perhaps HH acts on mesenchymal cells surrounding cords to initiate basement membrane formation around primordial follicles. Reduction of HH signaling in cyclopamine-treated mice may have prevented a signal from being expressed by the pre-granulosa cells, resulting in disrupted communication between the mesenchymal and epithelial cells. The mesenchymal cells may not have been able to synthesize basement membrane components, including laminin, in appropriate quantities due to this blocked cell interaction. Thus a lack of cell communication between epithelial and mesenchymal cells may have caused the high incidence of discontinuous basement membranes around primordial follicles observed in the cyclopamine-treated mice. 25
26 The decreased ratio of mesenchymal cells to oocytes in cyclopamine-treated mice suggests that HH also plays a role in this part of primordial follicle formation. HH has been identified as a key player in regulating cell proliferation in multiple organs, including the ovary [5]. This suggests a potential role for HH in regulating mesenchymal cell proliferation in the ovary. In the developing palate, epithelial-expressed SHH was found to regulate palatal mesenchyme cell proliferation through the maintenance of cyclin D1 and D2 expression [24]. HH also regulates mesenchyme cell proliferation in the gut [25] and in the developing lungs [26] through epithelial cell and mesenchymal cell interactions. In these studies, the absence of HH led to decreased mesenchymal cell proliferation. Since HH is an important regulator of cell proliferation in other organs and a decreased ratio of mesenchymal cells to oocytes was observed in this study, it is possible that reduced HH signaling in the ovary inhibited mesenchymal cell proliferation. Mesenchymal cells proliferate extensively in both the embryonic and neonatal ovary [20]. In the present study, cyclopamine was used to inhibit HH signaling after birth and thus may have prevented mesenchymal cells from proliferating during that period. The ratio of epithelial cells to oocytes was similar between controls and cyclopamine-treated mice. Epithelial cells only appear to proliferate in the embryonic ovary, not after birth [20]. Since HH was blocked by cyclopamine treatment after birth, and epithelial cells are not proliferating at this time, it is not surprising that the epithelial cell to oocyte ratio was not reduced. The reduced mesenchymal cell to oocyte ratio in cyclopamine-treated mice could have also been caused by failure of the mesenchymal cells to localize appropriately around primordial follicles. In the day 0 ovary, mesenchymal cells are arranged in long 26
27 chains surrounding the ovigerous cords, but by day 4 these cells become localized around primordial follicles. The fact that these cells were still arranged in chains in ovaries of cyclopamine-treated mice on day 4 rather than localized around individual primordial follicles suggests that HH signaling is necessary to control the localization of mesenchymal cells around primordial follicles. Preliminary studies were performed in which mice treated with high dose cyclopamine on days 0-3 were examined at 30 days of age. Cyclopamine-treated mice caged with fertile adult males had litter sizes similar to controls, indicating that the cyclopamine-treated mice were fertile. Histology of day 30 ovaries from cyclopaminetreated mice showed that follicles at all stages of development were present. This is as expected, since all cyclopamine-treated mice still had a certain percentage of correctly formed primordial follicles. These primordial follicles would be able to enter the growing pool, develop normally and ovulate. The mice had normal sized litters, suggesting that the number of follicles that ovulated was not affected. This result may be due to a compensatory mechanism within the ovary. The ovary generates the species-specific number of ovulatory follicles, even when the size of the primordial follicle pool is reduced. There is an inverse correlation between the number of primordial follicles and the rate at which they are stimulated to grow, such that a reduction in the size of the primordial stockpile will result in an increase in the proportion of follicles beginning to grow [23]. This occurs to ensure that the correct number of follicles ovulate. A number of cell signaling pathways have been shown to be involved in primordial follicle development [9]. For example, the Notch pathway has been shown to be expressed in oocytes and pre-granulosa cells prior to the assembly of primordial 27
28 follicles [27]. Furthermore, when Notch signaling was suppressed in cultured newborn mouse ovaries, there was a marked reduction in primordial follicles and an increase in the number of germ cells that remained in cords [27]. Thus, multiple pathways may direct primordial follicle formation and the inhibition of one may result in decreased efficiency of primordial follicles formation rather than complete inhibition. Even if HH is effectively blocked, some primordial follicles may form as the other required pathways remain intact. This study is the first to examine the effects of HH signaling on primordial follicle formation in the mouse ovary. While this study offers evidence for an important role of HH signaling in primordial follicle formation, additional research is necessary to support this. Cyclopamine should be administered for 8 days after birth to fully understand HH s role in primordial follicle development. While a majority of primordial follicle formation occurs during the first four days of life, primordial follicle formation is not absolutely complete at this time. This leaves open the possibility that additional follicle formation could occur after termination of cyclopamine treatment on day 3. Inhibition of the HH signaling pathway through day 8 might be expected to generate a more severe reduction in primordial follicle formation. Whether the treatment of newborn mice with cyclopamine resulted in a reduced number of primordial follicles at day 30 was not examined, but ovaries have been collected and fixed for future study. In the future, the number of primordial follicles and follicles at various other stages of development will be assessed to confirm an effect on reduced follicle formation and to determine if inhibition of HH signaling early in life affects the dynamics of follicle development in the mature ovary. Levels of follicle 28
29 atresia in day 30 cyclopamine-treated mice should also be evaluated. Preliminary studies with Amhr cre/+ Smo flox/null mice, a transgenic mouse model in which the Smo gene has been conditionally deleted in the ovary and reproductive tract, show an abnormally high level of atresia in primary follicles. Therefore, it would be interesting to determine whether this effect also occurs in adult mice that had been treated with cyclopamine neonatally. The results of this study are consistent with the hypothesis that HH signaling plays a role in primordial follicle formation in the mouse ovary. It appears to do so by affecting the mesenchymal cell to oocyte ratio as well as basement membrane formation. When HH signaling is reduced, it appears that mesenchymal cells may not proliferate as extensively or that they may not localize normally around developing primordial follicles. This may result in decreased signaling between epithelial and mesenchymal cells, such that primordial follicle formation and basement membrane deposition is disrupted. Future research is necessary to support these findings. 29
30 Literature Cited: 1. Hirshfield AN. Development of follicles in the mammalian ovary. International Review of Cytology 1991; 124: Skinner MK. Regulation of primordial follicle assembly and development. Human Reproduction Update 2005; 11(5): Russell MC, Cowan RG, Harman RM, Walker AL, and Quirk SM. The hedgehog signaling pathway in the mouse ovary. Biology of Reproduction 2007; 77: Wijgerde M, Ooms M, Hoogerbrugge JW, and Grootegoed JA. Hedgehog signaling in mouse ovary: Indian hedgehog and desert hedgehog from granulosa cells induce target gene expression in developing theca cells. Endocrinology 2005; 146(8): Ren Y, Cowan RG, Harman RM, Quirk SM. Dominant activation of the hedgehog signaling pathway in the ovary alters theca development and prevents ovulation. Mol Endocrinol In Press. 6. Ingham PW, McMahon AP. Hedgehog signaling in animal development: paradigms and principles. Genes & Development 2001; 15(23): Hooper JE, and Scott MP. Communicating with hedgehogs. Nature 2005; 6: Rubin LL, and de Sauvage FJ. Targeting the hedgehog pathway in cancer. Nature 2006; 5: Pangas SA. Growth Factors in Ovarian Development. Seminars in reproductive medicine 2007; 25(4): Pierucci-Alves F, Clark AM, Russell LD. A developmental study of the Desert Hedghog-Null Mouse Testis. Biology of Reproduction 2001; 65: Clark AM, Garland KK, Russell LD. Desert hedgehog (Dhh) is required in the mouse testis for formation of adult-type lydig cells and normal development of peritubular cells and seminiferous tubules. Biology of Reproduction 2000; 63: Forbes AJ, Lin H, Ingham PW, and Spradling AC. Hedgehog is required for the proliferation and specification of ovarian somatic cells prior to egg chamber formation in Drosophila. Development 1996; 122:
31 13. Zhang Y, and Kalderon D. Hedgehog acts as a somatic stem cell factor in the Drosophila ovary. Nature 2001; 410: Pepling ME, Spradling AC. Female mouse germ cells form synchronously dividing cysts. Development 1998; 125: Bagvandoss P, Midgley AR Jr, Wicha M. Developmental changes in the ovarian follicular basal lamina detected by immunofluorescence and electron microscopy. J Histochem Cytochem 1983; 31(5) Skinner MK, Tung PS, Fritz IB. Cooperativity between Sertoli cells and testicular peritubular cells in the production and deposition of extracellular matrix components. J Cell Biol 1985; 100(6) Berkholltz CB, Lai BE, Woodruff TK, and Shea LD. Distribution of extracellular matrix proteins type I collagen, type IV collagen, fibronectin and laminin in mouse folliculogenesis. Histochem Cell Biology 2006; 126: Van Wezel LL, Rodgers HF, and Rodgers RJ. Differential localization of laminin chains in bovine follicles. Journal of Reproduction and Fertility 1998; 112: Lee VH, Britt JH, Dunbar BS. Localization of laminin proteins during early follicular development in pig and rabbit ovaries. Journal of reproduction and fertility 1996; 108: Hirshfield AN, DeSanti AM. Patterns of Ovarian cell proliferation in rats during the embryonic period and the first three weeks postpartum. Biology of reproduction 1995; 53: Hirshfield AN. Heterogenetiy of cell populations that contribute to the formation of primordial follicles in rats. Biology of reproduction 1992; 47: Rajah R, Glaser EM, Hirshfield AN. The changing architecture of the neonatal rat ovary during histogenesis. Developmental dynamics 1992; 194: Hirshfield AN. Relationship between the supply of primordial follicles and the onset of follicular growth in rats. Biology of reproduction 1994; 50: Lan Y, Jiang R. Sonic hedgehog signaling regulates reciprocal epithelialmesenchymal interactions controlling palatal outgrowth. Development 2009;136(8): Katoh Y, Katoh M. Hedgehog signaling pathway and gastrointestinal stem cell signaling network (review). Int J Mol Med Dec;18(6):
32 26. Pepicelli CV, Lewis PM, McMahon AP. Sonic hedgehog regulates branching morphogenesis in the mammalian lung. Curr Biol Sep 24;8(19): Trombly DJ, Woodruff TK, Mayo KE. Suppression of Notch signaling in the neonatal mouse ovary decreases primordial follicle formation. Endocrinology 2009; 150(2):
OVARY The surface of the ovary is covered with surface epithelium
 OVARY Cow The ovary, or female gonad, is: 1. an exocrine gland, producing oocytes 2. an endocrine gland, secreting hormones, i.e., estrogen and progesterone OVARY OVARY The surface of the ovary is covered
OVARY Cow The ovary, or female gonad, is: 1. an exocrine gland, producing oocytes 2. an endocrine gland, secreting hormones, i.e., estrogen and progesterone OVARY OVARY The surface of the ovary is covered
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Occurrence of polyovular follicles and its possible significance in the ovary of the bat, Scotophilus heathi
 Biol Res 31: 75-80 (1998) Occurrence of polyovular follicles and its possible significance in the ovary of the bat, Scotophilus heathi UP SINGH, J DOVAL and A KRISHNA* Department of Zoology, Bañaras Hindu
Biol Res 31: 75-80 (1998) Occurrence of polyovular follicles and its possible significance in the ovary of the bat, Scotophilus heathi UP SINGH, J DOVAL and A KRISHNA* Department of Zoology, Bañaras Hindu
Biology 4361 Developmental Biology. October 11, Multiple choice (one point each)
") Biology 4361 Developmental Biology Exam 1 October 11, 2005 Name: ID#: Multiple choice (one point each) 1. Sertoli cells a. surround spermatocytes b. are the structural components of the seminiferous tubules
Biology 4361 Developmental Biology Exam 1 October 11, 2005 Name: ID#: Multiple choice (one point each) 1. Sertoli cells a. surround spermatocytes b. are the structural components of the seminiferous tubules
The role of growth factors in regulating cellular events during ovarian follicular development Leon J. Spicer
 The role of growth factors in regulating cellular events during ovarian follicular development Leon J. Spicer Department of Animal Science, Oklahoma State University, Stillwater, OK USA SESSION #54 EAAP
The role of growth factors in regulating cellular events during ovarian follicular development Leon J. Spicer Department of Animal Science, Oklahoma State University, Stillwater, OK USA SESSION #54 EAAP
Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a,
 Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a, Transverse sections of E17.5 ovary and mesonephros from Gli1-LacZ reporter embryos (n=3) after LacZ staining (blue). The
Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a, Transverse sections of E17.5 ovary and mesonephros from Gli1-LacZ reporter embryos (n=3) after LacZ staining (blue). The
Developing Molecularly Targeted Therapies for Basal Cell Carcinoma. Ivor Caro, MD, FAAD
 Developing Molecularly Targeted Therapies for Basal Cell Carcinoma Ivor Caro, MD, FAAD Disclosures Genentech, Inc Medical Director, Dermatology (employee) Stock holder Hedgehog Signaling Pathway Fundamental
Developing Molecularly Targeted Therapies for Basal Cell Carcinoma Ivor Caro, MD, FAAD Disclosures Genentech, Inc Medical Director, Dermatology (employee) Stock holder Hedgehog Signaling Pathway Fundamental
Biology 4361 Developmental Biology Exam 1 ID#: October 11, 2005
 Biology 4361 Developmental Biology Name: Key Exam 1 ID#: October 11, 2005 Multiple choice (one point each) 1. Primordial germ cells a. are immortal b. produce polar bodies c. are haploid d. are somatic
Biology 4361 Developmental Biology Name: Key Exam 1 ID#: October 11, 2005 Multiple choice (one point each) 1. Primordial germ cells a. are immortal b. produce polar bodies c. are haploid d. are somatic
Gametogenesis. Omne vivum ex ovo All living things come from eggs.
 Omne vivum ex ovo All living things come from eggs. William Harvery, 1651 Gametogenesis This lecture is the preface, so to speak, to embryology; that is, it introduces the development of the specialized
Omne vivum ex ovo All living things come from eggs. William Harvery, 1651 Gametogenesis This lecture is the preface, so to speak, to embryology; that is, it introduces the development of the specialized
SUPPLEMENTARY MATERIAL. Sample preparation for light microscopy
 SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
SUPPLEMENTARY MATERIAL Sample preparation for light microscopy To characterize the granulocytes and melanomacrophage centers, cross sections were prepared for light microscopy, as described in Material
Supplemental Figure 1: Leydig cells are reduced at multiple stages in both male sterile mutants
 SUPPLEMENTAL FIGURE LEGENDS: Supplemental Figure 1: Leydig cells are reduced at multiple stages in both male sterile mutants (Sgpl1 -/- and Plekha1 -/- ). Using an antibody against CYP11a1 to label Leydig
SUPPLEMENTAL FIGURE LEGENDS: Supplemental Figure 1: Leydig cells are reduced at multiple stages in both male sterile mutants (Sgpl1 -/- and Plekha1 -/- ). Using an antibody against CYP11a1 to label Leydig
REPRODUCTIVE CYCLE OF FEMALE MAMMAL
 REPRODUCTIVE CYCLE OF FEMALE MAMMAL Fig. 8-12 Secondary follicles growing follicles increase in number of layers of granulosa cells Tertiary follicles maturing follicles antrum formation fluid filled space
REPRODUCTIVE CYCLE OF FEMALE MAMMAL Fig. 8-12 Secondary follicles growing follicles increase in number of layers of granulosa cells Tertiary follicles maturing follicles antrum formation fluid filled space
Morphometric analysis of ovarian follicles of Black Bengal goats during winter and summer season
 Morphometric analysis of ovarian follicles of Black Bengal goats during winter and summer season MA Bari 1, ME Kabir 1, MB Sarker 1, AHNA Khan 2 and M Moniruzzaman* 1 1 Department of Animal Science, Bangladesh
Morphometric analysis of ovarian follicles of Black Bengal goats during winter and summer season MA Bari 1, ME Kabir 1, MB Sarker 1, AHNA Khan 2 and M Moniruzzaman* 1 1 Department of Animal Science, Bangladesh
Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy
 Kavya Puchhalapalli CALS Honors Project Report Spring 2017 Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy Abstract Malignant brain tumors including medulloblastomas and primitive neuroectodermal
Kavya Puchhalapalli CALS Honors Project Report Spring 2017 Studying The Role Of DNA Mismatch Repair In Brain Cancer Malignancy Abstract Malignant brain tumors including medulloblastomas and primitive neuroectodermal
Strategic delivery: Setting standards Increasing and. Details: Output: Demonstrating efficiency. informing choice.
 Strategic delivery: Setting standards Increasing and informing choice Demonstrating efficiency economy and value Details: Meeting Scientific and Clinical Advances Advisory Committee Agenda item 6 Paper
Strategic delivery: Setting standards Increasing and informing choice Demonstrating efficiency economy and value Details: Meeting Scientific and Clinical Advances Advisory Committee Agenda item 6 Paper
SUMMARY. Keywords: quail, Coturnix japonica, morphology, ovary, oviduct, neurotrophins, immunohistochemistry
 SUMMARY Keywords: quail, Coturnix japonica, morphology, ovary, oviduct, neurotrophins, immunohistochemistry Studies on the development of biological systems have expanded using animal models, always to
SUMMARY Keywords: quail, Coturnix japonica, morphology, ovary, oviduct, neurotrophins, immunohistochemistry Studies on the development of biological systems have expanded using animal models, always to
Analysis on the mechanism of reduced nephron number and the pathological progression of chronic renal failure in Astrin deficient rats
 Analysis on the mechanism of reduced nephron number and the pathological progression of chronic renal failure in Astrin deficient rats Summary of Doctoral Thesis Hidenori Yasuda Graduate School of Veterinary
Analysis on the mechanism of reduced nephron number and the pathological progression of chronic renal failure in Astrin deficient rats Summary of Doctoral Thesis Hidenori Yasuda Graduate School of Veterinary
The Initiation of Follicle and Oocyte. Growth in the Mouse Ovary
 BIOLOGY OF RPRODUCTION 20, 773-778 (1979) The Initiation Follicle and Oocyte Growth in the Mouse Ovary SU LINTRN-MOOR and G.P.M. MOOR School Biological Sciences, Macquarie University, North Ryde, N.S.W.
BIOLOGY OF RPRODUCTION 20, 773-778 (1979) The Initiation Follicle and Oocyte Growth in the Mouse Ovary SU LINTRN-MOOR and G.P.M. MOOR School Biological Sciences, Macquarie University, North Ryde, N.S.W.
Genome Integrity in Mammalian Oocytes
 Genome Integrity in Mammalian Oocytes ESHRE Workshop on mammalian folliculogenesis and oogenesis April 19 21 Stresa Italy 2003 Workshop Lisbon Genome Integrity Structure is chromatin in open or closed
Genome Integrity in Mammalian Oocytes ESHRE Workshop on mammalian folliculogenesis and oogenesis April 19 21 Stresa Italy 2003 Workshop Lisbon Genome Integrity Structure is chromatin in open or closed
Supplementary Materials and Methods
 Supplementary Materials and Methods Whole Mount X-Gal Staining Whole tissues were collected, rinsed with PBS and fixed with 4% PFA. Tissues were then rinsed in rinse buffer (100 mm Sodium Phosphate ph
Supplementary Materials and Methods Whole Mount X-Gal Staining Whole tissues were collected, rinsed with PBS and fixed with 4% PFA. Tissues were then rinsed in rinse buffer (100 mm Sodium Phosphate ph
Early Embryonic Development
 Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Reproductive Endocrinology. Isabel Hwang Department of Physiology Faculty of Medicine University of Hong Kong Hong Kong May2007
 Reproductive Endocrinology Isabel Hwang Department of Physiology Faculty of Medicine University of Hong Kong Hong Kong May2007 isabelss@hkucc.hku.hk A 3-hormone chain of command controls reproduction with
Reproductive Endocrinology Isabel Hwang Department of Physiology Faculty of Medicine University of Hong Kong Hong Kong May2007 isabelss@hkucc.hku.hk A 3-hormone chain of command controls reproduction with
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG)
") In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/-
 Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
A Genetic Program for Embryonic Development
 Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Oogenesis. Key Concepts. Female Reproductive Tract
 Oogenesis 1 Key Concepts Female Reproductive Tract Ovary Oogenesis Follicles Ovulation Corpus Luteum Molecular Activity Primordial Germ Cells (PGCs) 2 Female Reproductive Tract Ovary Oviduct Uterus Vagina
Oogenesis 1 Key Concepts Female Reproductive Tract Ovary Oogenesis Follicles Ovulation Corpus Luteum Molecular Activity Primordial Germ Cells (PGCs) 2 Female Reproductive Tract Ovary Oviduct Uterus Vagina
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression
 Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
Fresh and Frozen Ovary Tissue Transplants: Mechanism of Adult Primordial Follicle Recruitment And Fetal Oocyte Arrest
 Fresh and Frozen Ovary Tissue Transplants: Mechanism of Adult Primordial Follicle Recruitment And Fetal Oocyte Arrest Locking and Unlocking: Oocyte Meiosis and PGC differentiation Yasui et al 2012 Factors
Fresh and Frozen Ovary Tissue Transplants: Mechanism of Adult Primordial Follicle Recruitment And Fetal Oocyte Arrest Locking and Unlocking: Oocyte Meiosis and PGC differentiation Yasui et al 2012 Factors
Bi-potent Gonads. Sex Determination
 יצירת הגונדות Primordial Germ Cells (PGCs) Somatic cells Genital ridge Bi-potent Gonads Sex Determination Testis and Sperm Ovary and Oocyte Migration of Primordial Germ Cells in the Chick Embryo The
יצירת הגונדות Primordial Germ Cells (PGCs) Somatic cells Genital ridge Bi-potent Gonads Sex Determination Testis and Sperm Ovary and Oocyte Migration of Primordial Germ Cells in the Chick Embryo The
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products
 stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
The Beauty of the Skin
 The Beauty of the Skin Rose-Anne Romano, Ph.D Assistant Professor Department of Oral Biology School of Dental Medicine State University of New York at Buffalo The Big Question How do approximately 50 trillion
The Beauty of the Skin Rose-Anne Romano, Ph.D Assistant Professor Department of Oral Biology School of Dental Medicine State University of New York at Buffalo The Big Question How do approximately 50 trillion
A. Major parts 1. Nucleus 2. Cytoplasm a. Contain organelles (see below) 3. Plasma membrane (To be discussed in Cellular Transport Lecture)
 3. Plasma membrane (To be discussed in Cellular Transport Lecture)") Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
Lecture 5: Cellular Biology I. Cell Theory Concepts: 1. Cells are the functional and structural units of living organisms 2. The activity of an organism is dependent on both the individual and collective
Mohammad Sha ban. Basheq Jehad. Hamzah Nakhleh
 11 Mohammad Sha ban Basheq Jehad Hamzah Nakhleh Physiology of the reproductive system In physiology, we are concerned with the mechanisms in which the system functions, and how the system responds to different
11 Mohammad Sha ban Basheq Jehad Hamzah Nakhleh Physiology of the reproductive system In physiology, we are concerned with the mechanisms in which the system functions, and how the system responds to different
Spermatogenesis. What is it and what does it look like? How do hormones regulate spermatogenesis?
 Spermatogenesis What is it and what does it look like? How do hormones regulate spermatogenesis? FSH, androgens, growth factors Animal Physiology (Hill, Wise, Anderson): Ch. 15 435-438 1 Spermatogenesis:
Spermatogenesis What is it and what does it look like? How do hormones regulate spermatogenesis? FSH, androgens, growth factors Animal Physiology (Hill, Wise, Anderson): Ch. 15 435-438 1 Spermatogenesis:
Organelles of the Cell & How They Work Together. Packet #7
 Organelles of the Cell & How They Work Together Packet #7 Introduction Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging from 1 1000 cubic
Organelles of the Cell & How They Work Together Packet #7 Introduction Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging from 1 1000 cubic
Physiology of Male Reproductive System
 Physiology of Male Reproductive System the anterior pituitary gland serves as the primary control of reproductive function at puberty Ant Pituitary secretes FSH & large amounts of LH (ICSH) FSH & LH cause
Physiology of Male Reproductive System the anterior pituitary gland serves as the primary control of reproductive function at puberty Ant Pituitary secretes FSH & large amounts of LH (ICSH) FSH & LH cause
Embryology 3. Spermatogenesis:
 Embryology 3 Spermatogenesis: The 2 testis in males are each divided into lobes and lobules by connective tissue septa forming 250 lobule and in each lobule there are 1 to 4 seminefrous tubule ( so almost
Embryology 3 Spermatogenesis: The 2 testis in males are each divided into lobes and lobules by connective tissue septa forming 250 lobule and in each lobule there are 1 to 4 seminefrous tubule ( so almost
Human height. Length of some nerve and muscle cells. Chicken egg. Frog egg. Most plant and animal cells Nucleus Most bacteria Mitochondrion
 10 m 1 m 0.1 m 1 cm Human height Length of some nerve and muscle cells Chicken egg Unaided eye 1 mm Frog egg 100 µm 10 µm 1 µm 100 nm 10 nm Most plant and animal cells Nucleus Most bacteria Mitochondrion
10 m 1 m 0.1 m 1 cm Human height Length of some nerve and muscle cells Chicken egg Unaided eye 1 mm Frog egg 100 µm 10 µm 1 µm 100 nm 10 nm Most plant and animal cells Nucleus Most bacteria Mitochondrion
LQB383 Testbank. Week 8 Cell Communication and Signaling Mechanisms
 LQB383 Testbank Week 8 Cell Communication and Signaling Mechanisms Terms to learn match the terms to the definitions --------------------------------------------------------------------------------------------------------------------------
LQB383 Testbank Week 8 Cell Communication and Signaling Mechanisms Terms to learn match the terms to the definitions --------------------------------------------------------------------------------------------------------------------------
Animal Science 434 Reproductive Physiology
 Animal Science 434 Reproductive Physiology Development of the Pituitary Gland Lec 5: Embryogenesis of the Pituitary and Sexual Development Stomodeum Brain Infundibulum Rathke s Pouch Germ Cell Migration
Animal Science 434 Reproductive Physiology Development of the Pituitary Gland Lec 5: Embryogenesis of the Pituitary and Sexual Development Stomodeum Brain Infundibulum Rathke s Pouch Germ Cell Migration
Supplemental Data. Wnt/β-Catenin Signaling in Mesenchymal Progenitors. Controls Osteoblast and Chondrocyte
 Supplemental Data Wnt/β-Catenin Signaling in Mesenchymal Progenitors Controls Osteoblast and Chondrocyte Differentiation during Vertebrate Skeletogenesis Timothy F. Day, Xizhi Guo, Lisa Garrett-Beal, and
Supplemental Data Wnt/β-Catenin Signaling in Mesenchymal Progenitors Controls Osteoblast and Chondrocyte Differentiation during Vertebrate Skeletogenesis Timothy F. Day, Xizhi Guo, Lisa Garrett-Beal, and
MBios 401/501: Lecture 12.1 Signaling IV. Slide 1
 MBios 401/501: Lecture 12.1 Signaling IV Slide 1 Pathways that require regulated proteolysis 1. Notch and Delta 2. Wnt/ b-catenin 3. Hedgehog 4. NFk-B Our last topic on cell signaling are pathways that
MBios 401/501: Lecture 12.1 Signaling IV Slide 1 Pathways that require regulated proteolysis 1. Notch and Delta 2. Wnt/ b-catenin 3. Hedgehog 4. NFk-B Our last topic on cell signaling are pathways that
Organelles of the Cell & How They Work Together. Packet #5
 Organelles of the Cell & How They Work Together Packet #5 Developed by Mr. Barrow 2018 1 Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging
Organelles of the Cell & How They Work Together Packet #5 Developed by Mr. Barrow 2018 1 Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Maturation and Freezing of Bovine Oocytes
 Maturation and Freezing of Bovine Oocytes D. Mapes and M. E. Wells Story in Brief Immature bovine oocytes were aspirated from small to medium size follicles of bovine ovaries by needle and syringe. The
Maturation and Freezing of Bovine Oocytes D. Mapes and M. E. Wells Story in Brief Immature bovine oocytes were aspirated from small to medium size follicles of bovine ovaries by needle and syringe. The
Cell Divisions. The autosomes represent the whole body. * Male Sex Chromosomes: XY * Female Sex Chromosomes: XX
 Cell Divisions Each Cell (including gonads) has 46 chromosomes (23 pairs of chromosomes: 22 pairs of autosomes, 1 pair of sex chromosomes) which are located in the nucleus). The autosomes represent the
Cell Divisions Each Cell (including gonads) has 46 chromosomes (23 pairs of chromosomes: 22 pairs of autosomes, 1 pair of sex chromosomes) which are located in the nucleus). The autosomes represent the
Chapter 4: Cell Structure and Function
 Chapter 4: Cell Structure and Function Robert Hooke Fig. 4-2, p.51 The Cell Smallest unit of life Can survive on its own or has potential to do so Is highly organized for metabolism Senses and responds
Chapter 4: Cell Structure and Function Robert Hooke Fig. 4-2, p.51 The Cell Smallest unit of life Can survive on its own or has potential to do so Is highly organized for metabolism Senses and responds
Distribution of type IV collagen, laminin, nidogen and fibronectin in the haemodynamically stressed vascular wall
 Histol Histopath (1 990) 5: 161-1 67 Histology and Histopathology Distribution of type IV collagen, laminin, nidogen and fibronectin in the haemodynamically stressed vascular wall Reinhold Kittelberger,
Histol Histopath (1 990) 5: 161-1 67 Histology and Histopathology Distribution of type IV collagen, laminin, nidogen and fibronectin in the haemodynamically stressed vascular wall Reinhold Kittelberger,
Reproduction and Development. Female Reproductive System
 Reproduction and Development Female Reproductive System Outcomes 5. Identify the structures in the human female reproductive system and describe their functions. Ovaries, Fallopian tubes, Uterus, Endometrium,
Reproduction and Development Female Reproductive System Outcomes 5. Identify the structures in the human female reproductive system and describe their functions. Ovaries, Fallopian tubes, Uterus, Endometrium,
General Biology. Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell division
 General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
General Biology. Overview: The Key Roles of Cell Division. Unicellular organisms
 General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
Animal Science 434 Reproductive Physiology"
 Animal Science 434 Reproductive Physiology" Embryogenesis of the Pituitary and Sexual Development: Part A Development of the Pituitary Gland" Infundibulum" Brain" Rathke s Pouch" Stomodeum" Germ Cell Migration"
Animal Science 434 Reproductive Physiology" Embryogenesis of the Pituitary and Sexual Development: Part A Development of the Pituitary Gland" Infundibulum" Brain" Rathke s Pouch" Stomodeum" Germ Cell Migration"
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
(a) TEM of a plasma. Fimbriae. Nucleoid. Ribosomes. Plasma membrane. Cell wall Capsule. Bacterial chromosome
 TEM of a plasma. Fimbriae. Nucleoid. Ribosomes. Plasma membrane. Cell wall Capsule. Bacterial chromosome") 0 m m 0. m cm mm 00 µm 0 µm 00 nm 0 nm Human height Length of some nerve and muscle cells Chicken egg Frog egg Most plant and animal cells Most bacteria Smallest bacteria Viruses Proteins Unaided eye Light
0 m m 0. m cm mm 00 µm 0 µm 00 nm 0 nm Human height Length of some nerve and muscle cells Chicken egg Frog egg Most plant and animal cells Most bacteria Smallest bacteria Viruses Proteins Unaided eye Light
DRB666 Applied Developmental and Reproductive Biology (Spring 2013)
") DRB666 Applied Developmental and Reproductive Biology (Spring 2013) Director: 651 Ilalo Street, BSB163-3 e-mail: yyamazak@hawaii.edu Phone: (808) 692-1416 Instructors (e-mail): Steve Ward Yusuke Marikawa
DRB666 Applied Developmental and Reproductive Biology (Spring 2013) Director: 651 Ilalo Street, BSB163-3 e-mail: yyamazak@hawaii.edu Phone: (808) 692-1416 Instructors (e-mail): Steve Ward Yusuke Marikawa
Organelles of the Cell & How They Work Together. Packet #5
 Organelles of the Cell & How They Work Together Packet #5 Developed by Mr. Barrow 2018 1 Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging
Organelles of the Cell & How They Work Together Packet #5 Developed by Mr. Barrow 2018 1 Introduction Organization of cells is basically similar in all cells. Additionally, most cells are tiny Ranging
3 cell types in the normal ovary
 Ovarian tumors 3 cell types in the normal ovary Surface (coelomic epithelium) the origin of the great majority of ovarian tumors (neoplasms) 90% of malignant ovarian tumors Totipotent germ cells Sex cord-stromal
Ovarian tumors 3 cell types in the normal ovary Surface (coelomic epithelium) the origin of the great majority of ovarian tumors (neoplasms) 90% of malignant ovarian tumors Totipotent germ cells Sex cord-stromal
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Testicular stem cells
 Testicular stem cells Dirk G. de Rooij Department of Endocrinology Faculty of Biology, Utrecht University 1. Knowledge on the development of the spermatogenic stem cell lineage 2. Principals of the nature
Testicular stem cells Dirk G. de Rooij Department of Endocrinology Faculty of Biology, Utrecht University 1. Knowledge on the development of the spermatogenic stem cell lineage 2. Principals of the nature
DAX1, testes development role 7, 8 DFFRY, spermatogenesis role 49 DMRT genes, male sex differentiation role 15
 Subject Index N-Acetylcysteine, sperm quality effects 71 Ambiguous genitalia, origins 1, 2 Anti-Müllerian hormone function 13 receptors 13 Sertoli cell secretion 10, 38 Apoptosis assays in testes 73, 74
Subject Index N-Acetylcysteine, sperm quality effects 71 Ambiguous genitalia, origins 1, 2 Anti-Müllerian hormone function 13 receptors 13 Sertoli cell secretion 10, 38 Apoptosis assays in testes 73, 74
Web Activity: Simulation Structures of the Female Reproductive System
 differentiate. The epididymis is a coiled tube found along the outer edge of the testis where the sperm mature. 3. Testosterone is a male sex hormone produced in the interstitial cells of the testes. It
differentiate. The epididymis is a coiled tube found along the outer edge of the testis where the sperm mature. 3. Testosterone is a male sex hormone produced in the interstitial cells of the testes. It
MicroRNA and Male Infertility: A Potential for Diagnosis
 Review Article MicroRNA and Male Infertility: A Potential for Diagnosis * Abstract MicroRNAs (mirnas) are small non-coding single stranded RNA molecules that are physiologically produced in eukaryotic
Review Article MicroRNA and Male Infertility: A Potential for Diagnosis * Abstract MicroRNAs (mirnas) are small non-coding single stranded RNA molecules that are physiologically produced in eukaryotic
General Biology 1004 Chapter 11 Lecture Handout, Summer 2005 Dr. Frisby
 Slide 1 CHAPTER 11 Gene Regulation PowerPoint Lecture Slides for Essential Biology, Second Edition & Essential Biology with Physiology Presentation prepared by Chris C. Romero Neil Campbell, Jane Reece,
Slide 1 CHAPTER 11 Gene Regulation PowerPoint Lecture Slides for Essential Biology, Second Edition & Essential Biology with Physiology Presentation prepared by Chris C. Romero Neil Campbell, Jane Reece,
HANDOUT # 1 GAMETOGENESIS
 Gametogenesis 1 HANDOUT # 1 GAMETOGENESIS Anatomy Department R.A. FADEL Gametogenesis 2 بسم هللا الرحمن الرحيم ت هأ م وى }46 } ا نل و و ا زل و ج و ن و و و و و ن ه و أ ل ه ث وى }45{ إ ول ن أ و ة م ن ان
Gametogenesis 1 HANDOUT # 1 GAMETOGENESIS Anatomy Department R.A. FADEL Gametogenesis 2 بسم هللا الرحمن الرحيم ت هأ م وى }46 } ا نل و و ا زل و ج و ن و و و و و ن ه و أ ل ه ث وى }45{ إ ول ن أ و ة م ن ان
Sex Determination and Gonadal Sex Differentiation in Fish
 Sex Determination and Gonadal Sex Differentiation in Fish Yoshitaka Nagahama Okazaki National Research Institutes, Japan This first slide shows the processes of gonadal sex differentiation and gametogenesis
Sex Determination and Gonadal Sex Differentiation in Fish Yoshitaka Nagahama Okazaki National Research Institutes, Japan This first slide shows the processes of gonadal sex differentiation and gametogenesis
Characterization of Anti-Hamster ZP-0 Monoclonal Antibody
 Characterization of Anti-Hamster ZP-0 Monoclonal Antibody K. Ookata (1), K.Takagishi (1), S. Konno (2) and T. Oikawa(1,2) (1) Developmental and Reproductive Biology Center, Yamagata 990, Japan and (2)
Characterization of Anti-Hamster ZP-0 Monoclonal Antibody K. Ookata (1), K.Takagishi (1), S. Konno (2) and T. Oikawa(1,2) (1) Developmental and Reproductive Biology Center, Yamagata 990, Japan and (2)
Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment
 Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development
Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development
Ovary. Ovary is the female gonad or sex gland. Gonad: an organ that produces gametes; a testis or ovary.
 Ovary The ovary is composed of two parts: Outer cortex : houses the follicels. Inner medulla: is the middle region composed of connective tissue, blood vessels and lymphatics. Ovary is the female gonad
Ovary The ovary is composed of two parts: Outer cortex : houses the follicels. Inner medulla: is the middle region composed of connective tissue, blood vessels and lymphatics. Ovary is the female gonad
SUPPLEMENTARY INFORMATION
 a. Smo+/+ b. Smo+/+ 5.63 5.48 c. Lin- d. e. 6 5 4 3 Ter119 Mac B T Sca1 Smo+/+ 25 15 2 o BMT 2 1 5 * Supplementary Figure 1: Deletion of Smoothened does not alter the frequency of hematopoietic lineages
a. Smo+/+ b. Smo+/+ 5.63 5.48 c. Lin- d. e. 6 5 4 3 Ter119 Mac B T Sca1 Smo+/+ 25 15 2 o BMT 2 1 5 * Supplementary Figure 1: Deletion of Smoothened does not alter the frequency of hematopoietic lineages
CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions
 CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions Yi Feng, Peng Cui, Xiaowei Lu, Brian Hsueh, Fredrik Möller Billig, Livia Zarnescu Yanez, Raju Tomer, Derek
CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions Yi Feng, Peng Cui, Xiaowei Lu, Brian Hsueh, Fredrik Möller Billig, Livia Zarnescu Yanez, Raju Tomer, Derek
Animal Science 434! Tonic and Preovulatory Surge of GnRH! Tonic and Preovulatory Surge of GnRH! Lecture 11: The Follicular Phase of the Estrous Cycle!
 Tonic and Preovulatory Surge of GnRH! Animal Science 434! Lecture 11: The Follicular Phase of the Estrous Cycle!! (-)! Hypothalamus! GnRH! Estradiol! (-)! Tonic and Preovulatory Surge of GnRH! Anterior!
Tonic and Preovulatory Surge of GnRH! Animal Science 434! Lecture 11: The Follicular Phase of the Estrous Cycle!! (-)! Hypothalamus! GnRH! Estradiol! (-)! Tonic and Preovulatory Surge of GnRH! Anterior!
Regulation of Gene Expression in Eukaryotes
 Ch. 19 Regulation of Gene Expression in Eukaryotes BIOL 222 Differential Gene Expression in Eukaryotes Signal Cells in a multicellular eukaryotic organism genetically identical differential gene expression
Ch. 19 Regulation of Gene Expression in Eukaryotes BIOL 222 Differential Gene Expression in Eukaryotes Signal Cells in a multicellular eukaryotic organism genetically identical differential gene expression
Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body
 Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body Question No. 1 of 10 A biopsy sample is obtained from a lesion on the right cheek of a male patient. A technician in the histology lab
Human Anatomy and Physiology- Problem Drill 04: Tissues of the Body Question No. 1 of 10 A biopsy sample is obtained from a lesion on the right cheek of a male patient. A technician in the histology lab
Nature Genetics: doi: /ng Supplementary Figure 1. Assessment of sample purity and quality.
 Supplementary Figure 1 Assessment of sample purity and quality. (a) Hematoxylin and eosin staining of formaldehyde-fixed, paraffin-embedded sections from a human testis biopsy collected concurrently with
Supplementary Figure 1 Assessment of sample purity and quality. (a) Hematoxylin and eosin staining of formaldehyde-fixed, paraffin-embedded sections from a human testis biopsy collected concurrently with
11. SEXUAL DIFFERENTIATION. Germinal cells, gonocytes. Indifferent stage INDIFFERENT STAGE
 11. SEXUAL DIFFERENTIATION INDIFFERENT STAGE Early in pregnancy, (within 10-15 % of the pregnancy s expected length) a genital ridge is formed in the sides of the embryonic tissue, ventral to the mesonephros
11. SEXUAL DIFFERENTIATION INDIFFERENT STAGE Early in pregnancy, (within 10-15 % of the pregnancy s expected length) a genital ridge is formed in the sides of the embryonic tissue, ventral to the mesonephros
Germ Cell Transplantation in Fish
 Larvi 2009 Germ Cell Transplantation in Fish Goro Yoshizaki (Tokyo University of Marine Science and Technology, SORST/JST) Tuna Mackerel Body weight; 300 kg 300 g Body length; 3 m 30 cm Scombridae family
Larvi 2009 Germ Cell Transplantation in Fish Goro Yoshizaki (Tokyo University of Marine Science and Technology, SORST/JST) Tuna Mackerel Body weight; 300 kg 300 g Body length; 3 m 30 cm Scombridae family
Cellular compartments
 Cellular compartments 1. Cellular compartments and their function 2. Evolution of cellular compartments 3. How to make a 3D model of cellular compartment 4. Cell organelles in the fluorescent microscope
Cellular compartments 1. Cellular compartments and their function 2. Evolution of cellular compartments 3. How to make a 3D model of cellular compartment 4. Cell organelles in the fluorescent microscope
EVALUATION OF FOLLICULAR ATRESIA AND ELECTROPHORETIC PATTERN OF FOLLICULAR FLUID PROTEINS IN ACYCLIC BUFFALO (Bubalus bubalis)
") Original Article Buffalo Bulletin (December 2013) Vol.32 No.4 EVALUATION OF FOLLICULAR ATRESIA AND ELECTROPHORETIC PATTERN OF FOLLICULAR FLUID PROTEINS IN ACYCLIC BUFFALO (Bubalus bubalis) F.A. Khan 1,*,
Original Article Buffalo Bulletin (December 2013) Vol.32 No.4 EVALUATION OF FOLLICULAR ATRESIA AND ELECTROPHORETIC PATTERN OF FOLLICULAR FLUID PROTEINS IN ACYCLIC BUFFALO (Bubalus bubalis) F.A. Khan 1,*,
Rejuvenation of Gamete Cells; Past, Present and Future
 Rejuvenation of Gamete Cells; Past, Present and Future Denny Sakkas PhD Scientific Director, Boston IVF Waltham, MA, USA Conflict of Interest I have no conflict of interest related to this presentation.
Rejuvenation of Gamete Cells; Past, Present and Future Denny Sakkas PhD Scientific Director, Boston IVF Waltham, MA, USA Conflict of Interest I have no conflict of interest related to this presentation.
The reproductive lifespan
 The reproductive lifespan Reproductive potential Ovarian cycles Pregnancy Lactation Male Female Puberty Menopause Age Menstruation is an external indicator of ovarian events controlled by the hypothalamicpituitary
The reproductive lifespan Reproductive potential Ovarian cycles Pregnancy Lactation Male Female Puberty Menopause Age Menstruation is an external indicator of ovarian events controlled by the hypothalamicpituitary
Reproductive System. Testes. Accessory reproductive organs. gametogenesis hormones. Reproductive tract & Glands
 Reproductive System Testes gametogenesis hormones Accessory reproductive organs Reproductive tract & Glands transport gametes provide nourishment for gametes Hormonal regulation in men Hypothalamus - puberty
Reproductive System Testes gametogenesis hormones Accessory reproductive organs Reproductive tract & Glands transport gametes provide nourishment for gametes Hormonal regulation in men Hypothalamus - puberty
SISTEMA REPRODUCTOR (LA IDEA FIJA) Copyright 2004 Pearson Education, Inc., publishing as Benjamin Cummings
 Copyright 2004 Pearson Education, Inc., publishing as Benjamin Cummings") SISTEMA REPRODUCTOR (LA IDEA FIJA) How male and female reproductive systems differentiate The reproductive organs and how they work How gametes are produced and fertilized Pregnancy, stages of development,
SISTEMA REPRODUCTOR (LA IDEA FIJA) How male and female reproductive systems differentiate The reproductive organs and how they work How gametes are produced and fertilized Pregnancy, stages of development,
Animal Development. Lecture 3. Germ Cells and Sex
 Animal Development Lecture 3 Germ Cells and Sex 1 The ovary of sow. The ovary of mare. The ovary of cow. The ovary of ewe. 2 3 The ovary. A generalized vertebrate ovary. (Wilt and Hake, Ch 2, 2004) 4 The
Animal Development Lecture 3 Germ Cells and Sex 1 The ovary of sow. The ovary of mare. The ovary of cow. The ovary of ewe. 2 3 The ovary. A generalized vertebrate ovary. (Wilt and Hake, Ch 2, 2004) 4 The
Histology = the study of tissues. Tissue = a complex of cells that have a common function
 { EPITHELIAL TISSUE Histology = the study of tissues Tissue = a complex of cells that have a common function The Four Primary Tissue Types: Epithelium (epithelial tissue) covers body surfaces, lines body
{ EPITHELIAL TISSUE Histology = the study of tissues Tissue = a complex of cells that have a common function The Four Primary Tissue Types: Epithelium (epithelial tissue) covers body surfaces, lines body
EFFECT OF IRRADIATION IN INFANCY ON THE MOUSE OVARY
 EFFECT OF IRRADIATION IN INFANCY ON THE MOUSE OVARY A QUANTITATIVE STUDY OF OOCYTE SENSITIVITY HANNAH PETERS and EMILIA LEVY Einsen Laboratory, Copenhagen, Denmark (Received 13th June 1963) Summary. The
EFFECT OF IRRADIATION IN INFANCY ON THE MOUSE OVARY A QUANTITATIVE STUDY OF OOCYTE SENSITIVITY HANNAH PETERS and EMILIA LEVY Einsen Laboratory, Copenhagen, Denmark (Received 13th June 1963) Summary. The
Journal Club. 03/04/2012 Lama Nazzal
 Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
BONE TISSUE. Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology
 BONE TISSUE Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology BONE FUNCTION Support Protection (protect internal organs) Movement (provide leverage system for skeletal muscles, tendons, ligaments
BONE TISSUE Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology BONE FUNCTION Support Protection (protect internal organs) Movement (provide leverage system for skeletal muscles, tendons, ligaments
Chapter 22 The Reproductive System (I)
") Chapter 22 The Reproductive System (I) An Overview of Reproductive Physiology o The Male Reproductive System o The Female Reproductive System 22.1 Reproductive System Overview Reproductive system = all
Chapter 22 The Reproductive System (I) An Overview of Reproductive Physiology o The Male Reproductive System o The Female Reproductive System 22.1 Reproductive System Overview Reproductive system = all
Fertilization depends on mechanisms that help sperm meet eggs of the same species.
 Fertilization depends on mechanisms that help sperm meet eggs of the same species. www.uchsc.edu/ltc/fertilization.html Fertilization union of sperm and egg Is a chain of events. Interruption of any step
Fertilization depends on mechanisms that help sperm meet eggs of the same species. www.uchsc.edu/ltc/fertilization.html Fertilization union of sperm and egg Is a chain of events. Interruption of any step
Ovarian Follicular Development in the Untreated and
 Ovarian Follicular Development in the Untreated and PMSG-treated Cyclic Rat Hajime MIYAMOTO, Goro KATSUURA and Takehiko ISHIBASHI Department of Animal Science, College of Agriculture, Kyoto University,
Ovarian Follicular Development in the Untreated and PMSG-treated Cyclic Rat Hajime MIYAMOTO, Goro KATSUURA and Takehiko ISHIBASHI Department of Animal Science, College of Agriculture, Kyoto University,
Sexual Reproduction. For most diploid eukaryotes, sexual reproduction is the only mechanism resulting in new members of a species.
 Sex Determination Sexual Reproduction For most diploid eukaryotes, sexual reproduction is the only mechanism resulting in new members of a species. Meiosis in the sexual organs of parents produces haploid
Sex Determination Sexual Reproduction For most diploid eukaryotes, sexual reproduction is the only mechanism resulting in new members of a species. Meiosis in the sexual organs of parents produces haploid
Endocrine secretion cells secrete substances into the extracellular fluid
 Animal Hormones Concept 30.1 Hormones Are Chemical Messengers Endocrine secretion cells secrete substances into the extracellular fluid Exocrine secretion cells secrete substances into a duct or a body
Animal Hormones Concept 30.1 Hormones Are Chemical Messengers Endocrine secretion cells secrete substances into the extracellular fluid Exocrine secretion cells secrete substances into a duct or a body
Structure of Ovaries
 Structure of Ovaries What Are Oocyte Accessory Cells? Follicle cells somatic connected by desmosomes, gap junctions, interdigitating microvilli for anchorage and exchange function in yolk transport, egg
Structure of Ovaries What Are Oocyte Accessory Cells? Follicle cells somatic connected by desmosomes, gap junctions, interdigitating microvilli for anchorage and exchange function in yolk transport, egg
Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses
 The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses") Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses using an anti-cre antibody; testes at 1 week (left panel),
Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses using an anti-cre antibody; testes at 1 week (left panel),
Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive
 Online Data Supplement: Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive asthma Dan Cheng, Zheng Xue, Lingling Yi, Huimin Shi, Kan Zhang, Xiaorong Huo, Luke R. Bonser,
Online Data Supplement: Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive asthma Dan Cheng, Zheng Xue, Lingling Yi, Huimin Shi, Kan Zhang, Xiaorong Huo, Luke R. Bonser,
Inner ear development Nervous system development
 Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
Upcoming Sessions April 22: Nervous System Development Lecture April 24: Reviews of Axonal Pathfinding in Sensory Systems April 29: Inner Ear Development Lecture May 1: May 6: May 8: Auditory System Pathfinding
Lab Animal Tissue. LEARNING OBJECTIVES: To understand the relationship between the structure and function of different animal tissues
 Name: Bio A.P. PURPOSE: HYPOTHESIS: NONE Lab Animal Tissue BACKGROUND: In animals, groups of closely related cells specialized to perform the same function are called tissues. There are four general classes
Name: Bio A.P. PURPOSE: HYPOTHESIS: NONE Lab Animal Tissue BACKGROUND: In animals, groups of closely related cells specialized to perform the same function are called tissues. There are four general classes
CASE 41. What is the pathophysiologic cause of her amenorrhea? Which cells in the ovary secrete estrogen?
 CASE 41 A 19-year-old woman presents to her gynecologist with complaints of not having had a period for 6 months. She reports having normal periods since menarche at age 12. She denies sexual activity,
CASE 41 A 19-year-old woman presents to her gynecologist with complaints of not having had a period for 6 months. She reports having normal periods since menarche at age 12. She denies sexual activity,