Functional Characterization of the Interaction between Human La and Hepatitis B Virus RNA
|
|
|
- Rosanna Hutchinson
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on August 9, 2004 as Manuscript M Functional Characterization of the Interaction between Human La and Hepatitis B Virus RNA IMKE EHLERS**; SVEN HORKE; KERSTIN REUMANN; ANDREAS RANG#, FRANK GROSSE $, HANS WILL, AND TILMAN HEISE * Heinrich-Pette-Institut für Experimentelle Virologie und Immunologie an der Universität Hamburg, Martinistrasse 52, D Hamburg, Germany ** Present address: Developmental Biology Program Sloan-Kettering Institute, New York, NY # Present address: Institute of Virology, University Hospital Charité, Schumannstr. 20/21, D Berlin, Germany $ Institut für Molekulare Biotechnologie, Abteilung Biochemie, Beutenbergstraße 11, D Jena Running title: HBV RNA stability Key words: Hepatitis B Virus / HBV RNA/ RNA-binding proteins / RNA stability / La protein / La autoantigen * Corresponding Author, Mailing address: Tilman Heise, Dr. Heinrich-Pette-Institut für Experimentelle Virologie und Immunologie, Universität Hamburg Postfach Hamburg Germany Phone: Fax: heise@hpi.uni-hamburg.de 1 Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
2 The La protein is a multifunctional RNA binding protein and has also been suggested to be involved in the stabilization of Hepatitis B Virus (HBV) RNA. Here we demonstrate that antibodies against the human La protein specifically precipitate HBV RNA from HBV RNPcontaining mammalian cell extracts providing evidence for the association between human La and HBV RNA. Moreover we report, that the turnover of HBV RNA depends on structural features and less on the primary sequence of the La binding site on the viral RNA. In addition we show that the interaction between human La and HBV RNA in vitro is modulated by accessory factor/s in a phosphorylation dependent manner. Taken together these data indicate that both structural features, the composition of La/HBV ribonucleoprotein particles as well as interacting cellular factors are critical determinants in the regulation of the stability of the HBV RNA. 2
3 INTRODUCTION RNA metabolism depends on the formation of ribonucleoprotein particles mediating diverse processes such as splicing, polyadenylation, nuclear export, and the regulation of mrna stability (1,2). The formation of RNPs is a tightly controlled process, potentially regulated by several stimuli, including hormones and cytokines. Such stimuli can alter the RNA-binding activity of proteins on the post-translational level by phosphorylation or dephosphorylation and thereby the processing and stability of RNAs (3-5). In addition, RNA processing depends on various cis-acting elements including splice sites, export elements and endoribonucleolytic cleavage sites recognized by RNA binding proteins. In order to fully understand the regulation of processing of a specific RNA, both trans-acting factors and cisacting elements as well as their functions need to be known. The same applies for a detailed understanding of the metabolism of viral RNA. Such studies could lead to the identification of novel cellular targets valuable for the development of innovative antiviral strategies when focussed on the posttranscriptional control of RNAs of viruses with global medical importance. This applies to Hepatitis B Virus (HBV) with more than 300 million chronically infected carriers worldwide which await more effective antiviral therapies. HBV is a noncytopathic, hepatotropic virus with a 3.2-kb circular DNA genome. After conversion into a cccdna this genome serves in the nucleus as template for transcription of all viral RNAs. Synthesis of these transcripts is driven by at least four promoters leading to a large size heterogeneity with many different 5 -ends, while they all have very similar 3 -ends due to processing at the same polyadenylylation site (6). The so called pregenomic RNA (slightly longer than genome length) is encapsidated into nucleocapsids where it is reverse transcribed into viral DNA. This RNA serves also as messenger for synthesis of the viral P.- protein as well as for the core protein, a regulator protein, designated as e-antigen. The viral surface proteins as well as a regulatory protein with a role in hepatocarcinogenesis, designated HBx, are translated from subgenomic mrnas, 2.4 kb, 2.1 kb, and 0.7 kb in length. Recently, several factors and cis-acting elements involved in the posttranscriptional control of HBV gene expression were described. A number of publications highlighted the potential 3
4 of cytokines to induce the posttranscriptional downregulation of HBV RNA (for review see (7)) in a HBV transgenic mouse model (8), in human hepatoma cells (9,10) as well as the inhibition of duck HBV replication (11-13). In the HBV transgenic mouse model, the cytotoxic T lymphocyte (CTL) response to HBV antigens was shown to inhibit HBV replication by a posttranscriptional, non-cytotoxic mechanism leading to effective viral RNA degradation as induced by IFN-γ and TNF-α (8,14,15). While trying to evaluate the intracellular mechanism(s) responsible for the cytokine-mediated posttranscriptional destabilization of HBV RNA, the mouse La autoantigen (homologue to human La autoantigen) has been identified. The La protein interacts with a small cis-acting element located within the viral RNA between position nt (16,17). The tight temporal correlation between the cytokine-mediated down-regulation of HBV RNA and the cytokineinduced fragmentation of full-length La led to the assumption that full-length La stabilizes HBV RNA by interacting with the cis-acting element (18). Recently, it has been shown that HBV RNA is cleaved close to the La binding site by an endoribonucleolytic activity present in nuclear extracts prepared from HBV transgenic mice (19). Upregulation of this activity coincided with the cytokine-induced fragmentation of La and degradation of HBV RNA, supporting the assumption that HBV RNA is more accessible to endoribonucleolytic cleavage after disappearance of full-length La protein. More specifically, we hypothesize that La, in concert with additional trans-acting factors, forms an HBV RNA ribonucleoprotein complex stabilizing HBV RNA. The La protein has been described as important cellular factor involved in the RNA metabolism of a variety of viruses. Most often a function of La was attributed to the translational regulation of viruses like polio and Hepatitis C (20-23). In cultured cells relocalization of the predominantly nuclear La protein into the cytoplasm during infection was frequently observed (24,25). Moreover, La protein was reported to stabilize not only cellular histone mrna (26), but also RNA of hepatitis C virus (27). We studied whether hla is associated with full-length HBV RNA, and evaluated the importance of the La binding site for the half-life of HBV RNA. We show that HBV RNA is co-precipitated with hla, a strong indication for a physical interaction between hla and HBV 4
5 RNA in living cells. In addition, we demonstrate in transient transfection experiments that disruption of the proposed La binding site diminishes HBV RNA half-life. Furthermore we show that recombinant hla binds WT and mutant HBV RNA with the same affinity but with lower specificity arguing for the requirement of accessory factors for a specific interaction. Collectively, these results support the concept that hla contributes to HBV RNA stability. 5
6 EXPERIMENTAL PROCEDURES Plasmid constructs and mutagenesis-the HBV expression plasmid pch-9/3091 (kind gift of H. Schaller, Heidelberg, Germany) referred to as phbv-wt contains a more than full-length HBV genome (subtype ayw) in which synthesis of the pregenomic RNA is under control of a CMV promotor. Mutations were introduced into the plasmid by PCR according to the site directed mutagenesis method (Stratagene, USA) using proof-reading Pwo DNA Polymerase (Roche, Germany). Different nucleotide substitutions were introduced with the following oligonucleotides: mutant phbv-m2: sense primer (5'-CCG ATC CAT ACT AAG GAA CTC CTA GAA GCT TGT TTT GCT C-3'), antisense primer (5'-G AGC AAA ACA AGC TTC TAG GAG TTC CTT AGT ATG GAT CGG-3'); mutant phbv-m7: sense primer (5'-GCC GAT CCA TAC TCG CCA ACT CCT AGG GCG TTG TTT TGC TCG C-3'), antisense primer (5'-G CGA GCA AAA CAA CGC CCT AGG AGT TGG CGA GTA TGG ATC GGC-3'); mutant phbv-m12: sense primer (5'-GCG GAA CTC CTA TAC GCT TGT TTT GCT CG-3'), antisense primer (5'-CGA GCA AAA CAA GCG TAT AGG AGT TCC GC-3'); mutant phbv-m13: sense primer (5'-CCA TAC TGC GGA AAT AAT AGC CGC TTG TTT TGC TGC-3'), antisense primer (5'-CGA GCA AAA CAA GCG GCT ATT ATT TCC GCA GTA TGG-3'). ptethbv contains a more than full-length HBV genome (subtype ayw) in which synthesis of the pregenomic RNA is under control of the Tet promoter (kindly provided by Dr. Christoph Seeger, (28)). To introduce mutation M-2 into ptethbv plasmid, the Dra III and Nco I fragment of phbv-m2 was ligated into the Dra III and Nco I linearized ptethbv using standard methods. The plasmid was designated ptethbv-m2. Correct mutagenesis was confirmed by sequencing using the Taq-cycle sequencing protocol with IRD-800 labeled primers (MWG Biotech, Ebersberg, Germany) and a Licor automated sequencing device (MWG Biotech). 6
7 Cell Culture and Transfection Procedure- Per 60 mm dish, 1.3 x 10 6 HuH7 human hepatoma cells were grown as monolayers in 4 ml Dulbecco s modified Eagle s medium supplemented with 10% fetal calf serum. All plasmids used for transfection were purified using the Maxiprep kit (Qiagen). Transfections were performed using FuGENE 6 Transfection Reagent (Roche, Germany) according to the manufacturer s protocol. HBV expression plasmid (1.5 µg) was co-transfected with 0.5 µg pcmv/seap (kindly provided by J. Taylor, Philadelphia, USA) or, if indicated, with 0.07 µg pegfp-n1. To monitor transfection efficiency, Northern blot membranes were hybridized with a GFP-mRNA specific probe or the amount of SEAP activity secreted into the medium of transfected cells was determined as described previously (9). In vitro Transcription for Northern Blot Analysis- HBV-specific antisense RNA-probe (referred to as HBV probe 1, spanning nucleotides ) was generated by in vitro transcription with T3 RNA polymerase (Promega, USA) using the XhoI linearized plasmid pbsk c/x (kindly provided by G. Sommer, Hamburg, Germany). GAPDH-mRNA antisense RNA probes were generated by in vitro transcription of the Sac I- linearized plasmid pbsk- GAPDH (kindly provided by K. Harbers, Hamburg, Germany) using T7 RNA Polymerase (Promega). DNA templates used for the in vitro transcription to generate antisense RNA probes specific for HBV pregenomic RNA, histone 2A mrna and GFP-mRNA were raised by PCR. pwa 175 (containing the Histone H2A/a gene, kindly provided by W. Albig, Göttingen, Germany) was used for the production of DNA templates for the generation of histone 2A antisense transcripts. Two primers were used for PCR: the antisense primer (5'- gga tcc taa tac gac tca cta tag gga ACA TTG AGA TTT CAG GC-3') contained a restriction site for ClaI (shown in italic), the T7 RNA polymerase promoter sequence (shown in bold) and the histone 2A sequence spanning nt 829 to 847 (capitol letters) and the sense primer contained histone 2A sequences from nt 19 to 37 (5 -CAG CAG TGA GAA TGA 7
8 ACG C-3 ). Plasmid pegfp-n1 was used for the production of DNA templates for the generation of GFP antisense transcripts. Two primers were used: the antisense primer (5'-gga tcc taa tac gac tca cta tag ggt CCA TGC CGA GAG TGA TCC C-3') contained a restriction site for ClaI (shown in italic), the T7 RNA polymerase promoter sequence (shown in bold), the GFP sequence (capitol letters) and the sense primer contained GFP sequences (5 -CCT GGT CGA GCT GGA CGG C-3 ). PCRs for the templates were performed with 1 ng plasmid and the mixture contained 80 pmol of each primer in 1 X PCR buffer, 0.2 mm of GTP, ATP, TTP and CTP, and 2.5 U of Taq DNA polymerase (Roche). PCR was performed as follows: 5 min at 95 C, followed by 35 cycles of 1 min at 95 C, 1 min at 56 C, 1 min at 72 C and finally once 5 min 72 C. The PCR products were purified by size exclusion using Microspin G-25 columns (Amersham Bioscience, Germany) according to the manufacturer`s protocol, ethanol precipitated and used as templates for in vitro transcription. Transcription reactions were carried out with 0.5 to 1.0 µg linearized plasmid or PCR product in a final volume of 20 µl in transcription buffer (Promega) containing 0.31 mm ATP, CTP, GTP, 0.25 µm UTP, and 5.0 µm [α- 32 P]UTP (800 Ci/mmol) (Hartmann Analytic, Braunschweig, Germany), 5 mm DTT, and 20 U RNasin (Promega). The reaction was started by addition of 20 U T7 RNA polymerase (Promega). After incubation for 45 min at 37 C, another 20 U of T7 RNA polymerase were added, and the reaction was continued for 45 min at 37 C. The reaction was terminated by adding 10 µg of yeast trna and 1 U of DNaseI (Promega) and incubated for 15 min at 37 C. Unincorporated nucleotides were removed by using Microspin G-25 columns. Northern Blot Analysis-HuH7 cells were harvested 48 h or as indicated after transfection. Total RNA was prepared using TriPure Isolation Reagent (Roche) according to the manufacturer s protocol. 10 µg of total RNA was separated on a 1.2 % agarose- 8
9 formaldehyde gel. RNA was blotted onto a nylon membrane (Osmonics, Westborough, USA) and hybridized with 32 P-labeled in vitro transcribed RNA probes overnight at 68 C. To standardize transfection efficiency and RNA loading, Northern blots were hybridized with 32 P-labeled in vitro transcribed antisense GFP probe and antisense glycerinaldehyde-3- phosphate dehydrogenase (GAPDH) or antisense histone 2A probes, respectively. Blots were exposed to Fuji imaging screens, and signals were quantified by Fujix BAS 2000 bio-imaging analyzer (Fuji, Japan) and by TINA software (Raytest, Germany). Determination of pregenomic HBV-RNA half life using the Tet-Off-System-HuH7 cells were transfected with 1.5 µg ptethbv-wt or ptethbv-m2 and 0.5 µg puhd-ta, expressing tta, the tetracycline-controlled transactivator. Synthesis of pregenomic RNA was blocked by addition of 500 ng/ml Doxycycline to the culture medium 24 h or 40 h after transfection. Cells were harvested directly upon addition of Doxcycline or 3, 6, and 8 h thereafter. Total RNA was prepared and analyzed by Northern blot analysis. Co-immunoprecipitation and RT-PCR-For immunoprecipitation of endogenous hla-rna complexes about 10 7 HepG cells were harvested and cell lysates prepared as previously described (29). Briefly, cells were washed twice with ice cold PBS, collected in 500 µl ice cold polysomal lysis buffer containing 10 mm HEPES, ph 7.0, 100 mm KCl, 5 mm MgCl 2, 1 mm DTT and 0.5 % Nonidet P U RNAsin (Promega), 4 µl Ribonucleosid vanadyl complex (200 µg/µl, Sigma, USA) and protease inhibitor mix (Roche) were freshly added. Cells were pipetted up and down and incubated for 10 min on ice. The lysate was centrifuged at g for 10 min at 4 C, supernatant was removed and centrifuged again. The final supernatant was stored on ice until use. Protein A sepharose 4 fast flow beads (Amersham Biosciences) were blocked with BSA and pre-coated with specific antibodies as described previously (29). Mouse anti-la antibody (about 7 µg SW5, kindly provided by M. 9
10 Bachmann) or a purified mouse Igg (7.5 µg per reaction, Sigma) were used. The antibody coated beads were washed 6 times with 900 µl NT2 buffer containing 50 mm Tris, ph 7.4, 150 mm NaCl, 1 mm MgCl 2, 0.05% NP-40. Finally the pellets were resuspended in 850 µl NT2 buffer, supplemented with 4 µl vanadyl ribonucleosid complex, 160 units RNasin and 20 mm EDTA and 100 µl cell lysate. The immunoprecipitation reactions were rotated (head over head) at 4 C over night. Beads were collected by centrifugation at 800 g for 2 min and pellets were washed 6 times with ice cold NT2 buffer. Pellets for western blot analysis were resolved in 100 µl SDS denaturing buffer and pellets for RNA analysis were resuspended in 100 µl NT2-buffer supplemented with 0.01 % SDS, 40 µg proteinase K (Roche) and incubated at 50 C for 30 min in a thermomixer (1000 rpm), subsequently RNA was extracted using the TriPure Isolation Reagent (Roche) and precipitated in presence of 20 µg Gycogen. For northern blot analysis the complete RNA pellet was solved in 40 µl loading buffer and loaded onto a 1.2 % agarose-formaldehyde gel and processed as described above. Western blot analysis was performed by standard methods, using 12.5 % polyacrylamig gels and nitrocellulose membranes. For RT PCR analysis the RNA pellets (starting material, pellets, supernatants) were solved in 43 µl DEPC treated water, subsequently 5 µl RNase free DNase reaction buffer (Promega) and 2 units RNAse free DNAse (Promega) were added and incubated for 30 min at 37 C. DNAse was inactivated by heating the sample for 10 min at 70 C. 2 µl of DNase treated RNA was used as template for RT PCR reaction. RT PCR was conducted using the Titan One-Tube RT PCR kit (Roche) and following primer pairs were used: HBV Sense 5 - GGACCCCTTCTCGTGTTACAG-3, antisense 5 -GCTGGTTGTTGAGGATCCTGG-3 ; L-37 sense 5 -ACGAAGGGAACGTCATCGT-3`, antisense 5 - CATTCGACCAGTTCCGGT-3 ; hnrnp E2 sense 5 -GACACCGGTGTGATTGAAGG-3, antisense 5 -CTCAGGACAATTCCCTTCTGAG-3. 10
11 Expression, purification, phosphorylation, dephosphorylation of recombinant hla protein and preparation of nuclear extracts- Recombinant hla protein was prepared as described previously (17). Phosphorylation of recombinant hla was performed for 3 h at 37 C in 30 µl reactions containing 1 µg hla, 0.2 µl Casein Kinase II (New England Biolabs, Germany), 3 µl 10x reaction buffer and 6 µl 1 mm ATP. Dephosphorylation occurred with shrimpsalkaline phosphatase (Roche) or λ-protein phosphatase (New England Biolabs) according to the manufacturer s instructions. All reactions were controlled by 2-D-SDS-PAGE and efficient hyperphosphorylation or dephosphorylation was verified (data not shown). For preparation of nuclear extracts Huh7 cells were plated in 15 cm dishes, grown until confluence, washed twice with 1 x PBS, harvested in 1 ml 1x PBS and collected by centrifugation at 500 x g for 5 min. Cells were resuspended in 1 ml 1 x PBS and pelleted by centrifugation for 10 sec at g. Subsequently the cell pellet was suspended in 600 µl buffer A containing 20 mm HEPES, ph 7.9, 1.5 mm MgCl 2, 10 mm KCl, 0.2 mm EDTA, 0.1% Nonidet 40, at time of use 0.5 mm DTT and 40 µl/ml proteaseinhibitor Mix (Roche) were added. Samples were placed on ice for 10 min. Cell lysis was controlled by light microscopy. Nuclei were collected by centrifugation for 1 min at x g and resuspended in 150 µl buffer B containing 20 mm HEPES, ph 7.9, 1,5 mm MgCl 2, 10 mm KCl, 420 mm NaCl, 0.2 mm EDTA, 25% glycerol and freshly added 0.5 mm DTT and 40 µl/ml proteaseinhibitor Mix. Samples were placed on ice for 20 min and cell lysate was cleared by centrifugation at x g for 10 min and stored at -70 C. Gel Retardation Assay-The standard binding reaction was carried out in a final volume of 40 µl with 200 ng, or as indicated, purified recombinant hla protein and about 200,000 cpm or molar concentrations as indicated, of 32 P-radiolabeled HBV RNA.B in binding buffer containing 10 mm Tris-HCl ph 7.4, 3 mm MgCl 2, 200 mm NaCl, 0.5 mm EDTA and 0.5% 11
12 NP40 (unless otherwise stated). The in vitro transcribed 32 P-labeled RNA.B was denatured at 75 C for 10 min and renatured by cooling slowly to RT prior to addition to the reaction mixture. Samples were incubated for 30 min at RT. After addition of 5 µl of electrophoresis buffer containing 10% glycerol and 0.01% bromphenol blue, reaction mixtures were separated on an 8% native polyacrylamide gel (18 x18 cm) for 3 4 h at 200 Volt at room temperature. The gels were prerun at 240 volt for 1 h in 1 X TBE containing 45 mm Tris, 45 mm boric acid, 1 mm EDTA. Gels were dried (for 1.5 h at 80 C) on Whatman paper (Whatman, UK) using a Bio-Rad Slab Dryer Model 483 (Bio-Rad, Germany) and signals were evaluated using a FUJIX BAS 2000 Phosphorimager System (Fuji, Germany) or documented via exposure to X-ray films. For supershift analysis, 100 ng of NiNTA purified hla was pre-incubated with 5 µl antibody (as indicated in the figures) and 20 U RNasin (Promega) in a 20 µl volume of 10 mm Tris- HCl ph 7.4 at 4 C for 1 h, before the nuclear extracts and the other components of the standard reaction and the labeled RNA were added. Analysis of RNA-protein-interactions with surface plasmon resonance (SPR) technology- Real-time affinity determination of recombinant human La protein with different RNAs was achieved using a BIAcore-2000 device (BIAcore, Germany) producing the sensorgram. All experiments were performed at 25 C with a nitrilotriacetic acid (NTA)- sensor chip (Biacore) and the Histidin-tagged recombinant hla proteins and gel-purified in vitro transcribed HBV RNA.B-WT, -M2 and M7. The chip contains 4 flow cells that were treated and detected simultaneously. All four flow cells were loaded with 25 µl loading-buffer (500 µm NiSO 4 in running-buffer (see below)) at 10 µl min -1 which resulted in a stabile average increase of response units (RUs). After washing flow cells for 150 s at 10 µl min -1 with running buffer (10 mm HEPES ph 7.4, 150 mm NaCl, 50 µm EDTA, 0.005% Tween-20), proteins 12
13 were injected from 0.2 µm solutions with 5 µl min -1 into flow cells 1 to 3. Protein attachment to the chip surface was manually restricted to yield a mean difference of RUs, where 1000 RUs corresponds to approximately 1.0 ng/mm 2 (30). Afterwards, RNA was injected into all four flow cells, thereby obtaining triple values (flow cells 1 to 3) each with one negative control (flow cell 4) reflecting potential unspecific binding of RNA to the Nicoated chip surface. Application of RNAs resulted in RU differences of , depending on the amount loaded. To exclude that any RNA potentially still bound to the fixed protein after a study could interfere with subsequent analysis, the chip surface was regenerated after measurement by applying 30 µl regeneration buffer (10 mm HEPES ph 7.4, 150 mm NaCl, 100 mm EDTA, 0.005% Tween-20) with 10 µl min -1, before the chip was loaded and prepared for successive measurements. RNAs were supplied in at least 4 different concentrations, which resulted in consistent data where the level of response units after RNA loading increased with the amount of RNA loaded. Data were evaluated with the BIAevaluation software (Biacore), curves of flow cell 4 without protein were substracted from the corresponding protein containing flow cells 1 to 3 and results were fitted into Langmuir s model of 1:1 binding. As for the fit, software internal Chi 2 standards never exceeded values above 0.8, R max values were in the expected ranges and the residuals had a mean deviation from fitted graphs of less than 1.0 (not shown). Estimated half-life of a given RNA-protein complex was calculated as t ½ = -ln(0,5)/k d. 13
14 RESULTS The hla protein is associated with HBV RNA- Recently it has been shown that the cytokineinduced posttranscriptional degradation of HBV RNA tightly correlates with the cytokinedependent processing of the mouse La protein (18). In addition it has been shown that recombinant hla interacts with in vitro transcribed HBV RNA.B (17). These data suggest an association of La and HBV RNA also cell culture. Accordingly, we asked whether it would be possible to co-precipitate HBV RNA with hla specific antibodies for immunoprecipitations (IP). For this purpose HepG cells, stable expressing HBV RNA (31), were harvested, lysed, and subsequently the cleared cell lysate was incubated with either anti-la antibodies or mouse IgG 2A coated protein A sepharose beads. As shown in figure 1A and B hla was specifically precipitated and not detectable in the pellet of the control IP. To detect viral RNA in the IPs pellets, RNA was extracted from additional pellets immunoprecipitated with hla specific as well as unspecific antibodies and analyzed by RT- PCR (Fig. 1 A) and northern blotting (Fig. 1 B). RT-PCR was performed with specific primers for HBV RNA, TOP RNA L-37 and the hnrnp E2 RNA. Recently the coprecipitation of L-37 mrna but not of the hnrnp E2 with La was shown (32) and were used as positive and negative controls in our study, respectively. As shown by RT-PCR HBV RNA and L-37 mrna were specifically amplified in the La immunoprecipitation pellet but not in the IgG control pellet. In contrast hnrnp E2 RNA was not co-precipitated. This analysis strongly suggests an association between hla and HBV RNA. To further verify the association between hla and HBV RNA, we performed an additional Coimmunoprecipitation experiment (Fig. 1 B) and analyzed the RNA extracted from the IP pellets for HBV RNA by Northern blot analysis. This experiment reveals that full-length HBV pregenomic RNA was co precipitated with hla but not with IgG s although to a very small extent. Taken together both experiments clearly show the association between hla and HBV RNA and suggest a physical contact between hla and HBV RNA in living cells. Mutagenesis of the La binding site reduces HBV RNA levels- It has been reported earlier that mouse La present in nuclear extracts binds a predicted stem loop structure with high affinity 14
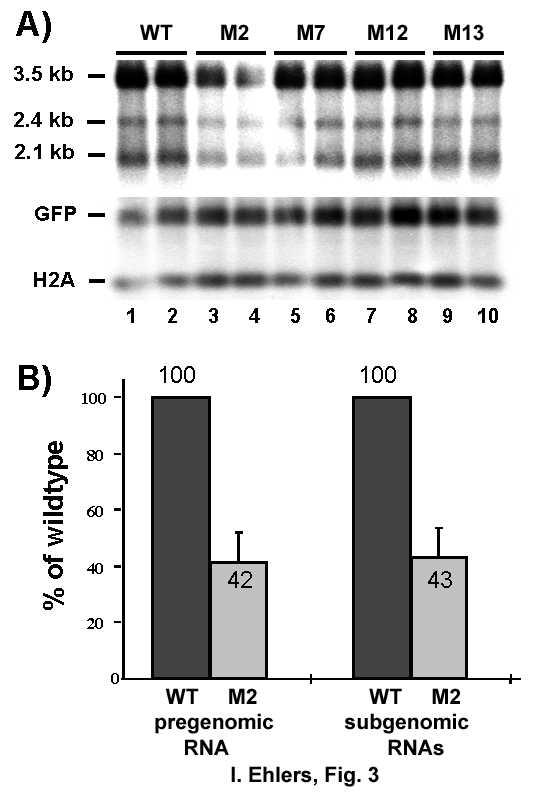
15 (approx. K D of 1 nmol) and with high specificity (16) in vitro. In contrast highly purified recombinant hla binds HBV RNA.B with high affinity but with low specificity (17). It was concluded that accessory factors are required to specify the interaction between La and HBV RNA.B. In competitive RNA binding studies the La binding site was mapped at nucleotide positions of the HBV genome. Computer based analysis of the secondary structure of HBV RNA.B revealed that the proposed binding site forms a stem loop structure (Fig. 2A). Mutagenesis of different nucleotides of this structure revealed that nucleotides at position 1275/76 and 1288/89 were important for a stable interaction between mouse La and in vitro transcribed HBV RNA.B (16). These nucleotide substitutions, referred to as HBV- M2, were introduced to disrupt the stem and thereby change the overall structure of RNA.B unfavorably for La recognition in vitro. However, we have to stress that the current structural information of this region is based on computer calculations and that this structure should be confirmed by alternative methods. If the La binding site is of major importance for viral gene expression the region should be conserved between different HBV genotypes and isolates. Comparison of this region revealed a substantial conservation of this sequence element throughout all HBV genotypes analyzed (Fig. 2B). Only one genotype B isolate contained two nucleotide substitutions which would lead to two mismatches within the stem, inducing a critical destabilization of the predicted structure. The most prominent substitution was located in one of the stem base pairs (C1289A(G)) and was found in 50 % of all analyzed viral sequences. We believe that this striking conservation emphasize the importance of this region for viral gene expression. In an attempt to investigate in greater detail whether the La binding site in the HBV RNA mediates stabilization of the viral transcripts, we designed cell culture experiments using the HBV expression plasmids phbv-wt. phbv-wt encodes the pregenomic RNA under the control of a CMV promoter, whereas the 2.4 and 2.1 kb mrnas encoding the surface proteins were controlled by endogenous promoters. This plasmid was used to introduce several mutations into the La binding site referred to as phbv-m2, phbv-m7, phbv-m12 and phbv-m13 (Fig. 2C). To study the influence of these mutations on HBV RNA levels, wild type and mutant HBV expression plasmids were transiently transfected into HuH-7 cells 15
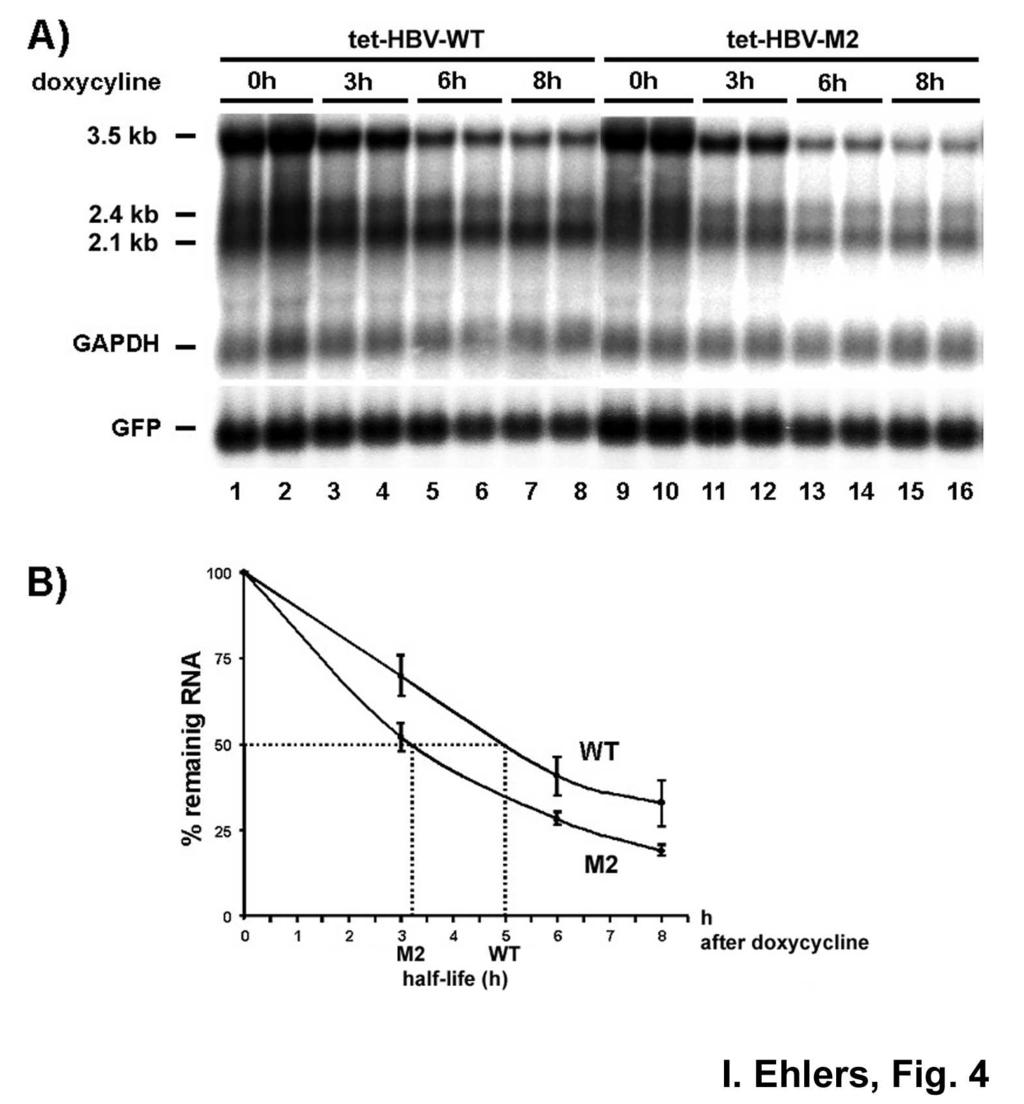
16 and RNA was prepared at indicated time points thereafter and analyzed by Northern blotting. Quantified HBV mrna levels were normalized against transfection efficiency controlled by co-expression of GFP or serum alkaline phosphatase and RNA loading was standardized by detection of GAPDH or Histone 2A mrnas. First we monitored HBV RNA levels transcribed from phbv-wt, phbv-m2, phbv-m7, phbv-m12 and phbv-m13 at 16 h and 24 h post transfection. Differences in the levels of the HBV RNA in cells transfected with phbv-m13, phbv-m7, phbv-m12, or phbv-wt were not detectable (data not shown), indicating that the respective mutations did not effect mrna levels. In contrast, lower HBV RNA levels compared to phbv-wt transfected cells were observed at both time points when phbv-m2 was transfected (data not shown). Very similar results were obtained when the RNA levels transcribed from phbv-wt, phbv-m2, phbv-m7, phbv-m12 and phbv-m13 were prepared 48 h post transfection and analyzed by Northern blotting, again only RNA levels transcribed from phbv-m2 were lower compared to HBV RNA levels transcribed from phbv-wt (Fig. 3A, lanes 1, 2 versus 3, 4). Quantification of 6 unrelated experiments performed in duplicate and with different plasmid preparations revealed an average decrease both of the pregenomic RNA and of the 2.4 kb and 2.1 kb subgenomic RNAs by approximately 60% when comparing transcript levels in phbv- WT and phbv-m2 transfected cells (Fig. 3B), indicating a substantial importance of this cisacting element for HBV RNA expression. Mutagenesis of the La binding site reduces HBV RNA half-life- Next we asked whether reduced HBV RNA-M2 levels compared to HBV wt RNA were due to an accelerated turn over rate. Therefore, the half-life of wild type and phbv-m2 pregenomic RNA was determined using a tetracycline (tet) controlled HBV expression system. Mutation M2 was introduced into the tetracyclin-regulated HBV expression vector (ptethbv-wt, (28)) referred to as ptethbv-m2. This system allows the specific shut-off of pregenomic HBV RNA transcription without affecting the general cellular transcription machinery which would be blocked by the use of inhibitors such as actinomycin D. Hence, potential ambiguous results due to secondary effects caused by inhibiting general transcription were 16
17 prevented. In this system synthesis of the pregenomic RNA was under the control of a CMV promoter inactivated after addition of the tetracycline analog doxycycline. Huh-7 cells were transiently transfected with ptethbv-wt and ptethbv-m2 plasmids, and HBV RNA halflife was determined by Northern blot analysis. A substantial amount of pregenomic RNA levels was raised 24 hours after transfection at which time doxycycline was added to abolish further HBV RNA synthesis (Fig. 4A, lanes 0h). Three, six and eight hours thereafter cells were harvested, RNA prepared and analyzed. Fig. 4A shows a representative Northern blot analysis of the half-life determination of pregenomic RNA. As expected from the reduced HBV-M2 RNA levels observed in our previous experiments, the pregenomic RNA levels of WT HBV were higher than that of HBV-M2 24 h post transfection immediately before doxycyclin addition to the cell culture medium (Fig. 4A, lanes 1, 2 versus 9, 10). Three hours after doxycyline treatment, pregenomic RNA levels were decreased to about 30% for WT HBV and to about 50% for HBV M2 (Fig. 4A, lanes 0h versus lanes 3h; and Fig. 4B). The pregenomic RNA levels decrease further to about 30% and 20% during the following observation period up to 8h post addition of doxycyclin (Fig. 4A, lanes 6h and 8h; and Fig. 4B). By comparing the kinetics of the decline of the pregenomic RNA levels a half-life of ptethbv-m2 pregenomic RNA which is shorter by about 2h compared to HBV WT became apparent (Fig. 4B). The quantification is based on phosphoimaging of the signals from four independent experiments and each performed with duplicate values, by setting the pregenomic RNA levels at time 0 h to 100% and by normalizing transfection efficiency as well as RNA loading. From this careful quantitative analysis of the half-life of HBV RNA two major conclusions can be drawn: first, pregenomic RNA of HBV M2 has a higher turnover than that of HBV WT and, therefore, is considerably less stable; second, the half-life of the pregenomic RNA of WT HBV, so far not reported elsewhere, is about 5h. This data imply that nucleotide substitutions GC and CC at positions 1275/76 and 1287/88, respectively, by adenosines, which weaken the base pairing of the predicted stem (Fig. 1B, HBV-M2) lead to reduced HBV RNA levels and a shorter half-life of the corresponding pregenomic RNA. Therefore, this region appears to contribute to pregenomic HBV RNA stability. 17
18 Recombinant hla interacts with HBV RNA.B-WT and -M2 with similar specificity and affinity in vitro- Next we compared the binding specificity of recombinant hla to in vitro transcribed HBV RNA.B-WT and HBV RNA.B-M2 in gel shift analysis. Competitive gel retardation assays revealed that both cold competitors HBV RNA.B-WT (Fig. 5A; lanes 3-5) and -M2 (Fig. 5A, lanes 7-9) were similarly efficient in competing for the binding of recombinant hla to labeled HBV RNA.B WT. Moreover, no difference in complex formation was observed when direct binding of recombinant hla to labeled HBV RNA.B-WT and -M2 was compared (data not shown). Taken together, in gel retardations assay no significant differences between the binding specificity of hla for HBV RNA.B-WT and that of M2 RNA was observed. To define the specific affinities of the interaction between recombinant hla and HBV RNA.B-WT, -M2 and -M7 we measured binding parameters using surface plasmon resonance (SPR) technology. In these experiments, recombinant His-tagged hla was attached to the NiNTA-chip surface and RNAs were supplied in different concentrations to monitor kinetics of association and dissociation. This procedure allows the accurate determination of binding affinities (Fig. 4B, C and D). As summarized in Figure 4D, recombinant hla binds HBV RNA.B-WT (K D (M) = 27 (±0.1) x 10-9 ), -M2 (K D (M) = 28 (±1) x 10-9 ) and M7 (K D (M) = 34 (±0.7) x 10-9 ) with similar affinities, which confirms our previous determined high affinity interaction between mouse La (K D ~ 1.0 x 10-9 (16)) and recombinant hla (K D ~ 0.8 x 10-9 ; (17)) for HBV RNA.B-WT as acquired by UV-crosslinks or gel retardation experiments, respectively. In conclusion, mouse La in nuclear extracts binds HBV RNA.B both with high affinity and high specificity while highly purified recombinant hla binds HBV RNA.B with high affinity but with low specificity emphasizing our previous results suggesting that accessory factors specifying the binding of La to HBV RNA. Modulation of the hla-hbv RNA.B interaction by nuclear factors and phosphorylation-to test whether additional cellular factors modulate the interaction between highly purified recombinant hla and HBV RNA.B we analyzed the binding in the presence of nuclear 18
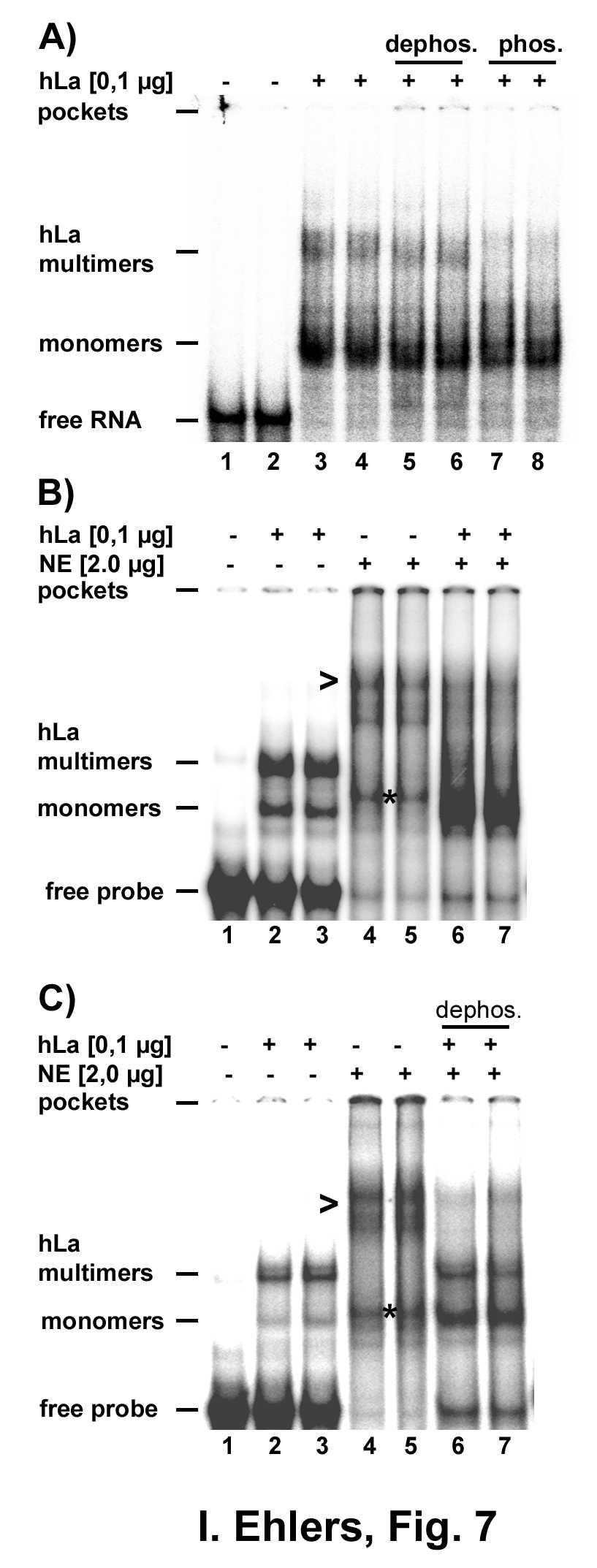
19 extracts prepared from Huh7 cells. Different amounts of recombinant hla or nuclear extracts were incubated with labeled HBV RNA.B-WT and analysed by EMSA. As shown in Figure 6, 50 and 100 ng recombinant hla without nuclear extract forms two major complexes with HBV RNA.B indicated as monomers and multimers (Fig. 6A, lanes 2-7). Next different amounts of nuclear extracts were tested revealing the formation of a major complex of low mobility (indicated by a arrow heads, Fig. 6B, lanes 2-7) and a minor complex of higher mobility (indicated by a star, Fig. 6B, lanes 2-7). The nature of these complexes is unknown but it might be possible that the complex indicated by a star represents a complex between endogenous La and HBV RNA.B because this band disappeared in super shift experiments (Fig. 6C, lanes 4, 5 versus 10, 11). Addition of increasing amounts of nuclear extract to 50 ng recombinant hla led to reduced signals of the multimeric and to increased signals of the monomeric hla/hbv RNA.B complex (Fig. 6A, lanes 4, 5 versus 8 to 13). The predominant formation of the monomeric La/HBV RNA.B complex in presence of nuclear extracts was confirmed if increasing amounts of recombinant hla were added to 2.0 µg nuclear extract (Fig. 6B, compare lanes 8 to 13 with 14, 15). To verify that hla specific RNPs were formed super shift experiments were performed. As shown in Fig. 6C La specific antibodies shifted the multimeric/monomeric hla/hbv RNA.B complexes (lanes 2, 3 versus 8, 9) as well as the dominant monomeric hla/hbv RNA.B complex formed in presence of nuclear extracts (lanes 6, 7 versus 12, 13). In contrast an unrelated antibody was unable to shift the hla/hbv RNA.B complexe (Fig. 6C, lanes 14, 15). These data demonstrate that components present in nuclear extracts modulate the binding behavior by preferentially supporting the formation of monomeric hla HBV RNA.B RNPs. These findings indicate that additional factors are involved in specific fromation of hla/hbv RNA RNPs. As shown earlier, dephosphorylation of mouse nuclear extracts strongly diminished the ability of mouse La to bind to HBV RNA.B (16). In that study it was not possible to discriminate whether the complex formation was abolished due to the dephosphorylation of endogenous mouse La or of accessory factors. La is mainly phosphorylated by casein kinase II at serine 366, and it was shown earlier that this modification did not alter the binding of hy1-rna (33). We now tested whether phosphorylation of recombinant hla by casein 19
20 kinase II or dephosphorylation of hla by alkaline phosphatase affected binding of HBV RNA.B. As shown in Fig. 7A untreated, dephosphorylated, and phosphorylated hla bound HBV RNA.B with similar efficiency, indicating that recombinant hla interacts with HBV RNA.B in a phosphorylation-independent manner. Although, in this experiment a decrease in multimers was observed with phophorylated hla (compare lanes 3,4 and 6, 8), this effect was not reproducible. 2-D gelelectrophoresis analysis of the different hla preparations verified successful phosphorylation and dephosphorylation of recombinant hla used in the gel retardation assays (not shown). These results suggest that the strong reduction in HBV RNA.B binding of mouse La after dephosphorylation of nuclear extracts (16) was due to a phosphorylation-dependent activity present in nuclear extract used. Next we asked whether dephosphorylated nuclear extract still led to the formation predominantly of a monomeric hla/hbv RNA.B complex as visualized by gel retardation assays. Addition of untreated nuclear extracts led almost exclusively to formation of a monomeric complex (Fig. 7B) while after addition of phosphatase treated nuclear extracts multimeric complexes were still clearly visible and the monomers much less abundant (Fig. 7C). In summary we show by gel retardation assays and with surface plasmon resonance technology that hla binds HBV RNA.B-WT and -M2 with comparable affinity but with low specificity, that nuclear extracts modulating the complex formation between hla and HBV RNA.B in a phosphorylationdependent manner suggesting that accessory factors increase phosphorylation-dependently the specificity of the interaction between La and HBV RNA. 20
21 DISCUSSION HBV gene expression is not only regulated at the transcriptional but also at the posttranscriptional level. The cytokine-induced processing of the La protein that coincides with the cytokine-induced degradation of HBV RNA suggests that the La protein stabilizes HBV RNA by a posttranscriptional mechanism. Recently, a predicted stem loop structure was reported to be bound by the La protein which is thought to confer HBV RNA stability. The present study was conducted to gain experimental evidence for the interaction of human La with HBV RNA in cell culture, the requirement of the predicted La binding site for HBV RNA stability and the role of accessory host factors in regulating the interaction between human La and HBV RNA. Herein for the first time we provide experimental data suggesting an association between hla and HBV RNA in human hepatoma cells. The co-precipitation of full-length HBV RNA with hla as shown in this report strongly suggests that La physically interacts with HBV RNA in living cells. Although, we can not rule out the formal possibility that the interaction between hla and HBV RNA is mediated by a bridging factor, the co-precipitation of hla and HBV RNA would still mean that hla is part of HBV RNPs, presumably stabilizing the viral RNA. The association of hla with HBV RNA or HBV stabilizing RNPs, combined with previous results showing that HBV RNA degradation coincides with mouse La fragmentation and that mouse La interacts specifically with the predicted stem-loop structure (16,18) strongly support the hypothesis that hla is involved in the posttranscriptional regulation of HBV RNA. Notably, the putative regulatory element bound by mouse and human La is located in the 3 -region of all viral RNAs targeted by the cytokine-induced posttranscriptional mechanism. To prove the significance of the La binding site for viral RNA stability several mutations were introduced and the half-life of the pregenomic RNAs was determined. These experiments revealed that only mutations disrupting the four base pairs in the stem diminished HBV RNA half-life. Conversely, changing the nucleotides in the stem but 21
22 maintaining base pairing, do not affect HBV RNA levels. These findings indicate that probably structural rather than sequential features of the RNA element are important to uphold HBV RNA half-life. In this context it is important to note, that the sequence of the stem-loop is highly conserved between a diversity of HBV genotypes and isolates (Fig. 1, B). Our study raises the interesting question whether the nucleotide substitutions within the predicted structure involved in La protein binding as present in various HBV genotypes and isolates has positive or negative consequences for the stability of the viral RNAs and efficiency of virus propagation. It was shown that mutation M2 diminishes pregenomic RNA half-life by approximately 2 hours, indicating that HBV RNA stability dependents in part on structural features of this element. Note, with the current data we can not exclude the possibility that the half-life of pregenomic RNA M2 was reduced by La-independent factors. Therefore, it is reasonable to consider that additional regulatory elements within the viral RNA and cellular factors interacting with HBV RNA and/or the La protein have to be integrated to fully understand the decay of HBV RNA under steady state conditions like in our experiments. Furthermore, the variety and remarkable changes in gene expression profiles observed after CTL-injection into HBV transgenic mice (34,35) probably inducing the inhibition of viral replication and inducing HBV RNA decay, let suggest that additional cellular factors and cellular events have to identify to understand the induced decay mechanism specific for the HBV RNA. It is reasonable to speculate that structural features of the La binding site determine the interaction with trans-acting factors and it might therefore be possible that mutation M2 leads to disruption of of hla containing RNPs normally protecting HBV RNA against endoribonucleolytic cleavage. However, changing the structure of this RNA element and so the HBV RNP composition might be not enough to induce efficient degradation as observed after CTL-injection in the mouse system in which HBV RNA degradation was accompanied by the processing of mouse La and the augmentation of an endoribonuclease activity (19). 22
23 Hence, the identification of factors involved in La-HBV RNP formation is of great importance and might shed light on the mechanism regulating HBV RNA decay under different cellular conditions. This view concurs with current accepted pathways of mrna degradation. In general it is believed that the fate of an mrna is regulated by trans-acting factors interacting with cisacting elements within an RNA molecule thereby protecting or destabilizing the RNA (for review (36)). Among the best characterized RNA destabilizing elements are AU-rich regions first described to regulate mrna stability of short lived RNAs (37). The AU-rich regions are recognized by a group of proteins called AU-rich binding proteins (38). In most cases these proteins destabilize RNAs by interacting with the AU-rich regions, but stabilizing effects have also been reported (39-41). To our knowledge, it is not known how exactly this interaction destabilizes/stabilizes RNA. The stable alpha-globin mrna has been described to interact with several proteins forming the so called alpha-complex associated with the 3'- untranslated region. Changes in the composition of this complex lead to endoribonucleolytic cleavage of this RNA (42,43), demonstrating that a variety of trans-acting factors are involved. The requirement of several trans-acting factors was also described for IL-2 mrna stability (44). The transferrin receptor mrna is stabilized by the iron regulatory protein, which interacts with the 3'-end of the RNA if cellular iron levels are low thereby protecting the RNA against endoribonucleolytic cleavage (45). An increase in cellular iron concentration leads to reduced affinity of the iron regulatory protein to the transferrin receptor mrna and subsequently to the degradation of the RNA (reviewed in (46)). Furthermore, intensive studies on the posttranscriptional control of albumin and vitellogenin mrna has shown that albumin mrna is destabilized after estrogen treatment in parallel with the activation of the polysomal ribonuclease 1 (PMR-1; (47,48)), whereas vitellogenin mrna is stabilized by binding of vigilin, protecting the mrna against PMR-1 cleavage (49). These reports clearly demonstrate and support our assumption that a complex regulatory 23
24 mechanism determines HBV RNA stability by integrating the action of regulatory transacting factors interacting with mrna stability determinants. Notably, in competitive UVcrosslinking experiments the binding of mouse La in nuclear extracts was highly specific for HBV RNA.B-WT but not for the mutant M2 (16), indicating that under such experimental settings mouse La was able to discriminate between WT and mutant HBV RNA-M2. In contrast highly purified, recombinant hla binds HBV RNA.B WT and M2 in an unspecific manner, suggesting that additional components, which were present in mouse nuclear extracts, are required for a specific interaction. Until those factors are characterized, it is probably not feasible to explore a difference in binding specificity of recombinant hla to HBV RNA.B-WT versus M2 in vitro and to show more directly that the half-life of pregenomic RNA dependents on the association with hla. We recently identified a hla interacting protein and preliminary data show that this RNA binding protein interacts specifically with HBV RNA.B (Tettweiler G. and Heise T. unpublished data), suggesting that this protein might modulates the association between HBV RNA.B and hla. Previously it has been shown that binding of mouse La to HBV RNA.B in vitro was abolished after dephosphorylation of the La containing nuclear extract (16). In this previous study, it was not possible to distinguish, whether impaired RNA binding activity was due to dephosphorylation of the phosphoprotein La or of other factors present in the nuclear extracts. Our observation that phosphorylation or dephosphorylation of recombinant hla had no effect on binding HBV RNA.B, whereas dephosphorylation of nuclear extracts partially inactivated the activity promoting formation of the monomeric La complex showed that a phosphorylation-dependent activity modulates hla binding to HBV RNA.B. We believe that this initial data support our view that the binding specificity of hla to HBV RNA.B is mediated by phosphorylation-dependent host factors. In the absence of those factors recombinant hla binds HBV RNA.B with low specificity. In this context it is important to note that the newly identified hla interacting protein is also a phosphoprotein and that it will 24
25 of interest to study whether the interaction between hla and this protein might be regulated by phosphorylation. Furthermore, it will be interesting to find out which cellular signaling pathways affect the proposed phosphorylation-dependent composition of HBV RNPs thereby potentially regulating the turnover of viral RNAs. Importantly, it has been shown recently that janus kinase activity is required for the antiviral affect of interferons on HBV replication (35), indicating that cell signaling pathways and thereby phosphorylation/dephosphorylation events are required. This might also applies for cytokine-induced degradation of HBV RNA decay as supported by our finding that binding of mouse and human La to HBV RNA is modulated by accessory factors in a phosphorylation-dependent manner. Previously it has been shown that a variety of cellular factors involved in the regulation of mrna stability, like RNA binding proteins were regulated by phosphorylation/dephosphorylation (50,51) and that ribonucleases activities were elevated by hormones (48,52,53), indicating that posttranslational modification and signal transduction pathways are important cellular mechanism determining mrna levels.as a model, we assume that cytokine-induced HBV RNA degradation is initiated at the La binding site and this concept is supported by the following findings. First, the cytokine-induced HBV RNA degradation occurring in the mouse system in parallel with La processing is associated with increased endoribonuclease activity which leads to cleavage of the HBV RNA close to the La binding site (18,19). Second, mutagenesis of the La binding site reduces HBV RNA half-life from 5 h to 3 h in cell culture experiments and third, binding of the mouse as well as of the human La protein to HBV RNA.B can be modulated by de/phosphorylastion of accessory factors. The cytokineinduced HBV RNA decay might be initiated by the loss of protective factors (e.g. La) inducing structural changes and/or exposing the cleavage site located close to the predicted stem loop to endoribonuclease (19), thereby initiating HBV RNA degradation. We conclude that the La binding site together with the cleavage sites compose a destabilizing element leading to HBV RNA degradation under specific cellular conditions. It will be of 25
26 major importance to identify host factors interacting with hla and/or HBV RNA in a phosphorylation-dependent manner to understand in more detail the initiation of HBV RNA degradation. 26
27 Acknowledgements We thank Christian Schulze (Center for Molecular Neurobiology, Hamburg, Germany) for help and advice in analyzing the SPR data. We are grateful to E. Chan for the prokaryotic expression plasmid pet-human La, M. Bachmann for La monoclonal antibody SW5. This work was supported by the Deutsche Forschungsgemeinschaft HE 2814/2-3 (TH), WI 664/9-1, (HW) and the BMBF (NGFN and HepNet). The Heinrich-Pette-Institut is supported by the Bundesministerium für Gesundheit und soziale Sicherung and the Freie und Hansestadt Hamburg. 27
28 Abbreviations hla = human La autoantigen; RNP = ribonucleinparticle; RRM = RNA recognition motif; WT = wild type; HBV = Hepatitis B Virus; CTL = cytotoxic T lymphocytes 28
29 REFERENCES 1. Keene, J. D. (2001) Proc. Natl. Acad. Sci. U. S. A. 98, Jensen, T. H., Dower, K., Libri, D., and Rosbash, M. (2003) Mol. Cell 11, Kracht, M., and Saklatvala, J. (2002) Cytokine 20, Bevilacqua, A., Ceriani, M. C., Capaccioli, S., and Nicolin, A. (2003) J. Cell Physiol. 195, Shim, J., and Karin, M. (2002) Mol. Cells 14, Seeger, C., and Mason, W. S. (2000) Microbiol. Mol. Biol. Rev. 64, Heise, T. D., M. Petersen J., Rang A, Burda M. R. Will H. (2000) in Chronic Hepatitis. New Concepts of Pathogenesis, Diagnosis and Treatment (Dienes, H. D. S. P. B. C. O. K., ed), pp , Kluwer Academic Publishers 8. Guidotti, L. G., Ishikawa, T., Hobbs, M. V., Matzke, B., Schreiber, R., and Chisari, F. V. (1996) Immunity 4, Rang, A., Gunther, S., and Will, H. (1999) J. Hepatol. 31, Rang, A., Bruns, M., Heise, T., and Will, H. (2002) J. Biol. Chem. 277, Klocker, U., Schultz, U., Schaller, H., and Protzer, U. (2000) J. Virol. 74, Schultz, U., and Chisari, F. V. (1999) J. Virol. 73, Schultz, U., Summers, J., Staeheli, P., and Chisari, F. V. (1999) J. Virol. 73, Guidotti, L. G., Ando, K., Hobbs, M. V., Ishikawa, T., Runkel, L., Schreiber, R. D., and Chisari, F. V. (1994) Proc. Nat. Acad. Sci. U. S. A. 91, Chisari, F. V. (1997) J. Clin. Invest. 99, Heise, T., Guidotti, L. G., and Chisari, F. V. (1999) J. Virol. 73, Horke, S., Reumann, K., Rang, A., and Heise, T. (2002) J. Biol. Chem. 277, Heise, T., Guidotti, L. G., Cavanaugh, V. J., and Chisari, F. V. (1999) J. Virol. 73, Heise, T., Guidotti, L. G., and Chisari, F. V. (2001) J. Virol. 75, Ali, N., and Siddiqui, A. (1997) Proc. Natl. Acad. Sci. U. S. A. 94, Shimazaki, T., Honda, M., Kaneko, S., and Kobayashi, K. (2002) Hepatology 35, Meerovitch, K., Svitkin, Y. V., Lee, H. S., Lejbkowicz, F., Kenan, D. J., Chan, E. K., Agol, V. I., Keene, J. D., and Sonenberg, N. (1993) J. Virol. 67, Pudi, R., Abhiman, S., Srinivasan, N., and Das, S. (2003) J. Biol. Chem. 278, Shiroki, K., Isoyama, T., Kuge, S., Ishii, T., Ohmi, S., Hata, S., Suzuki, K., Takasaki, Y., and Nomoto, A. (1999) J. Virol. 73, Bachmann, M., Falke, D., Schroder, H. C., and Muller, W. E. (1989) J. Gen. Virol. 70, McLaren, R. S., Caruccio, N., and Ross, J. (1997) Mol. Cell. Biol. 17, Spangberg, K., Wiklund, L., and Schwartz, S. (2001) J. Gen. Virol. 82, Rang, A., and Will, H. (2000) Nucleic Acids Res. 28, Tenenbaum, S. A., Lager, P. J., Carson, C. C., and Keene, J. D. (2002) Methods 26, Johnsson, B., Lofas, S., and Lindquist, G. (1991) Anal. Biochem. 198, Sells, M. A., Chen, M. L., and Acs, G. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,
30 32. Intine, R. V., Tenenbaum, S. A., Sakulich, A. L., Keene, J. D., and Maraia, R. J. (2003) Mol. Cell 12, Fan, H., Sakulich, A. L., Goodier, J. L., Zhang, X., Qin, J., and Maraia, R. J. (1997) Cell 88, Wieland, S. F., Vega, R. G., Muller, R., Evans, C. F., Hilbush, B., Guidotti, L. G., Sutcliffe, J. G., Schultz, P. G., and Chisari, F. V. (2003) J. Virol. 77, Robek, M. D., Boyd, B. S., Wieland, S. F., Chisari, F. V., Vega, R. G., Muller, R., Evans, C. F., Hilbush, B., Guidotti, L. G., Sutcliffe, J. G., and Schultz, P. G. (2004) Proc. Natl. Acad. Sci. U. S. A. 101, Epub 2004 Feb Guhaniyogi, J., and Brewer, G. (2001) Gene 265, Shaw, G., and Kamen, R. (1986) Cell 46, Wagner, B. J., DeMaria, C. T., Sun, Y., Wilson, G. M., and Brewer, G. (1998) Genomics 48, Laroia, G., Cuesta, R., Brewer, G., and Schneider, R. J. (1999) Science 284, Chen, C. Y., and Shyu, A. B. (1995) Trend. Biochem. Sci. 20, Sela-Brown, A., Silver, J., Brewer, G., and Naveh-Many, T. (2000) J. Biol. Chem. 275, Wang, Z., and Kiledjian, M. (2000) Mol. Cell. Biol. 20, Wang, Z., and Kiledjian, M. (2000) EMBO J. 19, Chen, C. Y., Gherzi, R., Andersen, J. S., Gaietta, G., Jurchott, K., Royer, H. D., Mann, M., and Karin, M. (2000) Genes Dev. 14, Binder, R., Horowitz, J. A., Basilion, J. P., Koeller, D. M., Klausner, R. D., and Harford, J. B. (1994) EMBO J.l 13, Rouault, T., and Klausner, R. (1997) Curr. Top. Cell. Regul. 35, Chernokalskaya, E., Dompenciel, R., and Schoenberg, D. R. (1997) Nucleic Acids Res. 25, Dompenciel, R. E., Garnepudi, V. R., and Schoenberg, D. R. (1995) J. Biol. Chem. 270, Cunningham, K. S., Dodson, R. E., Nagel, M. A., Shapiro, D. J., and Schoenberg, D. R. (2000) Proc. Natl. Acad. Sci. U. S. A. 97, Eisenstein, R. S., Tuazon, P. T., Schalinske, K. L., Anderson, S. A., and Traugh, J. A. (1993) J. Biol. Chem. 268, Malter, J. S., and Hong, Y. (1991) J. Biol. Chem. 266, Pastori, R. L., Moskaitis, J. E., and Schoenberg, D. R. (1991) Biochemistry 30, Heise, T., Krones, A., Nath, A., Jungermann, K., and Christ, B. (1998) Biol. Chem. Hoppe-Seyler 379,
31 LEGENDS Fig. 1: Co-precipitation of hla and HBV RNA. Cell lysates of HepG cells were applied for immunoprecipitation using the La specific mouse monoclonal antibody SW5 or mouse IgG controls. A) Pellets of immunoprecipitations were analyzed by western blot (WB) analysis using the La specific antibody SW5. SM = starting material (10%); P = Pellet (100%); SU = supernatant (5%); * indicates heavy chains of the antibodies used for immunoprecipitation. RT-PCR analysis of RNA prepared from IP pellets and treated with DNAse. Oligos for HBV RNA, L-37 and hnrnp E2 mrna are given under Experimental Procedures. PCR products were separated on a 2% agrose gel and DNA was visualized by ethidium bromide staining. Con indicates RT-PCR reaction without templates. B) Pellets of immunoprecipitations were analyzed by western blot (WB) analysis using the La specific antibody SW5. SM = starting material (10%); P = Pellet (100%); SU = supernatant (5%); < indicates La degradation products and * indicates heavy and light chains of the antibodies used for immunoprecipitation. For Northern blot analysis RNA was extracted from IP pellets, separated on a 1.2 % agarose/formaldehyde gel and blotted onto nylon membrane. HBV RNA was detected using a 32 P-labeld in vitro transcribed HBV probe 1. TR = 2µg of total RNA prepared from HepG cells. Fig. 2: A) Predicted secondary structure of HBV RNA.B (16). B) The sequence of the proposed La binding site is highly conserved. For sequence alignments of the stem loop sequence the MacVector software was used and following sequences applied: genotype A IDs: 59455, 59418, 59416, 60429, , ; genotype B IDs: , , , ; genotype C IDs: 59585, , , , , , 59404, , 59402, , 59408, , ; genotype D IDs: 59429, 62280, , 59434, , , , 59439, 59448; genotype E IDs: , ; genotype 31
32 F IDs: , , Nucleotide substitutions are shown in bold. Mutation/sequence indicates in how many of the sequences analyzed the indicated subsituion was found C) Outline of nucleotide substitution introduced into the predicted stem loop region. Nucleotide substitutions are shown in bold. Fig. 3: Northern blot analysis of HBV RNA transcribed from wild type and mutated HBV expression plasmids revealed that M2-RNA levels were reduced significantly. A) HuH7 cells were co-transfected with 1.5 µg of plasmids phbv-wt, phbv-m2, phbv-m13, phbv-m7 or phbv-m12. To monitor transfection efficiency 0.07 µg pegfp-n1 were cotransfected. 48 h thereafter RNA was prepared and 10 µg total RNA was analyzed by northern blotting. The pregenomic HBV RNA (3.5 kb) and the subgenomic HBV RNAs (2.4/2.1 kb) were detected using HBV probe 1. To normalize RNA loading and transfection efficiency the same blots were hybridized with histone H2A and GFP mrna specific probes, respectively. B) The pregenomic and subgenomic HBV RNA levels were quantified by phosphorimaging. Six independent experiments in performed in duplicates were quantified and HBV RNA levels were normalized against RNA loading and transfection efficiency. The standard deviations are indicated (+/- SD; n=6). Fig. 4: Mutation M2 diminishes the half-life time of HBV pregenomic RNA. HuH7 cells were co-transfected with 1.5 µg of plasmids tethbv-wt or tethbv-m2 and 0.5 µg puhd- TA and to monitor transfection efficiency 0.07 µg pegfp-n1. 24 h or 40 h thereafter cells were harvested (timepoint 0 h), or doxycyclin was added and cells were harvested at 3 h, 6 h or 8 h thereafter. RNA was prepared and 10 µg total RNA analyzed by northern blotting using HBV probe 1 to detected pregenomic (3.5 kb) and subgenomic HBV RNAs (2.4/2.1 kb). To normalize RNA loading and transfection efficiency the same blots were hybridized 32
33 with a GAPDH and GFP mrna specific probes, respectively. B) The half-life of the pregenomic HBV RNA expressed from phbv-wt and phbv-m2 were determined by northern blotting and quantified by phosphorimaging. Four independent experiments performed in duplicates were analyzed and HBV RNA levels were normalized against RNA loading and transfection efficiency. The standard deviations are indicated (+/- SD; n=4). Fig. 5. A) recombinant hla binds with similar specificity to HBV RNA.B-WT or M2. Standard gel retardation assay was performed under conditions described under "Experimental procedures". 200 ng recombinant hla purified by NiNTA spin columns was analyzed for binding of 50 fmol HBV RNA.B-WT (lane 2 to 9). Reaction without hla (lane 1). For competition experiments, unlabeled competitor HBV RNA.B-WT (lanes 3, 4, 5) or HBV RNA.B-M2 RNA (lanes 7, 8, 9) was used in 10, fold molar excess. B-E) Recombinant hla binds HBV RNA.B-WT, -M2, -M7 with similar affinities. BIAcore experiments were performed to determine the binding affinity of recombinant hla protein to HBV RNA.B-WT, -M2 or -M7 and evaluated according to the procedure described under Experimental procedures. Representative sensorgrams are shown for HBV RNA.B-WT (B), HBV RNA.B-M2 (C) and HBV RNA.B-M7 (D). Molar concentrations of applied RNAs are indicated. Summary of evaluated data is shown in (E). Standard deviations (±) are given for each rate of k a, k d and calculated K D, n gives number of individual values / repetitions. For the fitting according to Langmuirs 1:1 binding, software-internal Chi 2 standards never exceeded values above 0.78 and mean deviation of residual from fitted graphs were less than 1.0 (not shown). Fig. 6. Modulation of hla/hbv RNA.B complexes by nuclear factors. A, B) Standard gel retardation assay was performed under conditions described under "Experimental 33
34 procedures". Indicated amounts of recombinant hla purified by NiNTA spin columns (A, lanes 2 7) and of nuclear extracts separately (B, lanes 2-7) or in combinations (A and B, lanes 8-13) were analyzed for binding of 70 fmol HBV RNA.B-WT. Reaction without hla (lane 1). C) For supershift analysis, 100 ng of NiNTA purified hla was pre-incubated with 5 µl mouse monoclonal La specific antibody SW5 (αla, lanes 8-13) or with 5 µl of an unrelated mouse monoclonal antibody (con, lanes 14, 15)) and 20 U RNasin in a 20 µl volume of 10 mm Tris-HCl ph 7.4 at 4 C for 1 h, before 2 µg of nuclear extracts and the other components of the standard reaction and labeled RNA were added. < and * indicating RNPs formed between HBV RNA.B and nuclear extracts. Fig. 7. A) Binding of recombinant hla is independent of its phosphorylation state. Recombinant hla purified by NiNTA spin columns was phosphorylated by Casein Kinase II, dephosphorylated by shrimps alkaline phosphatase or untreated and applied for gel retardation assays. Efficient phosphorylation/dephosphorylation was verified by 2-D gelelectrophoresis (data not shown). Standard gel retardation assay was performed under conditions described under "Experimental procedures". 100 ng untreated (lanes 3, 4), dephosphorylated (lanes 5, 6) or phosphorylated (lanes 7, 8) recombinant hla was analyzed for binding of cpm HBV RNA.B-WT. Reaction without hla (lane 1, 2). B, C) Nuclear factors affecting the formation of hla/hbv RNA.B complexes are phosphorylation-dependent. B) Standard gel retardation assay was performed under conditions described under "Experimental procedures". 100 ng of recombinant hla (lanes 2, 3) purified by NiNTA spin columns and 2 µg of nuclear extracts (lanes 4, 5) were analyzed separately or in combinations (lanes 6, 7) for binding of 140 fmol HBV RNA.B-WT. Reaction without hla (lane 1). C) 100 ng of recombinant hla (lanes 2, 3) purified by NiNTA spin columns and 2 µg of nuclear extracts (lanes 4, 5) were separately or in combinations 34
35 (lanes 6, 7) analyzed for binding of 140 fmol HBV RNA.B-WT. Prior to the addition of HBV RNA.B-WT sample were treated for 30 min at 37 C with 10 units of alkaline phosphatase. Reaction without hla (lane 1). < and * indicating RNPs formed between HBV RNA.B and nuclear extracts. 35
36
37
38
39
40

41
42
c Tuj1(-) apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP
 apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP") Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Table 3. 3 UTR primer sequences. Primer sequences used to amplify and clone the 3 UTR of each indicated gene are listed.
 Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53
 Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Supplementary Document
 Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplemental Data. Shin et al. Plant Cell. (2012) /tpc YFP N
 /tpc YFP N") MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
Supplementary Materials
 Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at
 were treated with 4NQO at") Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Figure S1. Analysis of genomic and cdna sequences of the targeted regions in WT-KI and
 Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most
 Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 a
 Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
A smart acid nanosystem for ultrasensitive. live cell mrna imaging by the target-triggered intracellular self-assembly
 Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
CD31 5'-AGA GAC GGT CTT GTC GCA GT-3' 5 ' -TAC TGG GCT TCG AGA GCA GT-3'
 Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards
 overnight at 4 C, afterwards") Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Abbreviations: P- paraffin-embedded section; C, cryosection; Bio-SA, biotin-streptavidin-conjugated fluorescein amplification.
 Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Table S1. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9. Assay (s) Target Name Sequence (5 3 ) Comments
 Target Name Sequence (5 3 ) Comments") SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
Supplementary Table 2. Conserved regulatory elements in the promoters of CD36.
 Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because
 Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Citation for published version (APA): Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.
: Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.") University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
 www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
*To whom correspondence should be addressed. This PDF file includes:
 www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL
 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL") 1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
Culture Density (OD600) 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)
 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)") A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
Nature Immunology: doi: /ni.3836
 Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
BIOLOGY 621 Identification of the Snorks
 Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Supplementary Figure 1
 Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 2, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 2, 2006
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Supplemental Information. Th17 Lymphocytes Induce Neuronal. Cell Death in a Human ipsc-based. Model of Parkinson's Disease
 Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Product Manual. Omni-Array Sense Strand mrna Amplification Kit, 2 ng to 100 ng Version Catalog No.: Reactions
 Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
SUPPLEMENTARY DATA. Supplementary Table 1. Primer sequences for qrt-pcr
 Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Antibodies: LB1 buffer For 50 ml For 10ml For 30 ml Final 1 M HEPES, ph 2.5 ml 0.5 ml 1.5 ml 50mM. 5 M NaCl 1.4 ml 280 µl 0.
 Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Cross-talk between mineralocorticoid and angiotensin II signaling for cardiac
 ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
Beta Thalassemia Sami Khuri Department of Computer Science San José State University Spring 2015
 Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast
 Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
A basic helix loop helix transcription factor controls cell growth
 A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
Supplementary Figure 1
 Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Beta Thalassemia Case Study Introduction to Bioinformatics
 Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Supplemental Figures: Supplemental Figure 1
 Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Description of Supplementary Files. File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables
 Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
SUPPORTING INFORMATION
 SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
Nuclear Extraction Kit
 Nuclear Extraction Kit Catalog Number KA1346 50 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay... 3 General Information... 4
Nuclear Extraction Kit Catalog Number KA1346 50 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay... 3 General Information... 4
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
CIRCRESAHA/2004/098145/R1 - ONLINE 1. Validation by Semi-quantitative Real-Time Reverse Transcription PCR
 CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
Protocol for purification of recombinant protein from 300 ml yeast culture
 Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Characterization of Nuclear RNases That Cleave Hepatitis B Virus RNA near the La Protein Binding Site
 JOURNAL OF VIROLOGY, Aug. 2001, p. 6874 6883 Vol. 75, No. 15 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.15.6874 6883.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Characterization
JOURNAL OF VIROLOGY, Aug. 2001, p. 6874 6883 Vol. 75, No. 15 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.15.6874 6883.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Characterization
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
TetR repressor-based bioreporters for the detection of doxycycline using Escherichia
 Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies
 Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Nuclear Extraction Kit
 Nuclear Extraction Kit Item No. 10009277 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION 3 Materials
Nuclear Extraction Kit Item No. 10009277 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION 3 Materials
Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health
 Write your name here Surname Other names Edexcel GCE Centre Number Candidate Number Biology Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health Thursday 8 January 2009 Morning Time: 1 hour
Write your name here Surname Other names Edexcel GCE Centre Number Candidate Number Biology Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health Thursday 8 January 2009 Morning Time: 1 hour
Supporting Information
 Supporting Information Molecular Recognition Based DNA Nanoassemblies on the Surfaces of Nanosized Exosomes Shuo Wan,, Liqin Zhang,,, Sai Wang, Yuan Liu,, Cuichen Wu, Cheng Cui, Hao Sun, Muling Shi, Ying
Supporting Information Molecular Recognition Based DNA Nanoassemblies on the Surfaces of Nanosized Exosomes Shuo Wan,, Liqin Zhang,,, Sai Wang, Yuan Liu,, Cuichen Wu, Cheng Cui, Hao Sun, Muling Shi, Ying
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer. Application Note
 Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Journal of Cell Science Supplementary information. Arl8b +/- Arl8b -/- Inset B. electron density. genotype
 J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10
 J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
Supplementary Information. Bamboo shoot fiber prevents obesity in mice by. modulating the gut microbiota
 Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Lezione 10. Sommario. Bioinformatica. Lezione 10: Sintesi proteica Synthesis of proteins Central dogma: DNA makes RNA makes proteins Genetic code
 Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Supporting Information
 Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Supplementary Figure 1a
 Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Journal of Microbes and Infection,June 2007,Vol 2,No. 2. (HBsAg)2 , (PCR) 1762/ 1764
2 , (PCR) 1762/ 1764") 68 2007 6 2 2 Journal of Microbes and Infection,June 2007,Vol 2,No. 2 2 S 1 1 1 2 2 3 1 (HBsAg)2 ( YIC) S 5 30g 60g YIC ( HBV) DNA > 2 log10 e (HBeAg), 6 DNA, 1 YIC 1, (PCR) (0 ) (44 ) HBV DNA S 2, S a
68 2007 6 2 2 Journal of Microbes and Infection,June 2007,Vol 2,No. 2 2 S 1 1 1 2 2 3 1 (HBsAg)2 ( YIC) S 5 30g 60g YIC ( HBV) DNA > 2 log10 e (HBeAg), 6 DNA, 1 YIC 1, (PCR) (0 ) (44 ) HBV DNA S 2, S a
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help.
 SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness
 World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
Supplementary Materials and Methods
 DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
Astaxanthin prevents and reverses diet-induced insulin resistance and. steatohepatitis in mice: A comparison with vitamin E
 Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplemental Information. Cancer-Associated Fibroblasts Neutralize. the Anti-tumor Effect of CSF1 Receptor Blockade
 Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Product Contents. 1 Specifications 1 Product Description. 2 Buffer Preparation... 3 Protocol. 3 Ordering Information 4 Related Products..
 INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15
 Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
University of Groningen. Vasoregression in incipient diabetic retinopathy Pfister, Frederick
 University of Groningen Vasoregression in incipient diabetic retinopathy Pfister, Frederick IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from
University of Groningen Vasoregression in incipient diabetic retinopathy Pfister, Frederick IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Supplementary Information
 Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Influenza viruses are classified as members of the family
 Rewiring the RNAs of influenza virus to prevent reassortment Qinshan Gao a and Peter Palese a,b,1 Departments of a Microbiology and b Medicine, Mount Sinai School of Medicine, New York, NY 10029 Contributed
Rewiring the RNAs of influenza virus to prevent reassortment Qinshan Gao a and Peter Palese a,b,1 Departments of a Microbiology and b Medicine, Mount Sinai School of Medicine, New York, NY 10029 Contributed
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supporting Information
 Translation of DNA into Synthetic N-Acyloxazolidines Xiaoyu Li, Zev. J. Gartner, Brian N. Tse and David R. Liu* Department of Chemistry and Chemical Biology, Harvard University, Cambridge, Massachusetts
Translation of DNA into Synthetic N-Acyloxazolidines Xiaoyu Li, Zev. J. Gartner, Brian N. Tse and David R. Liu* Department of Chemistry and Chemical Biology, Harvard University, Cambridge, Massachusetts
Product Contents. 1 Specifications 1 Product Description. 2 Buffer Preparation... 3 Protocol. 3 Ordering Information 4
 INSTRUCTION MANUAL Quick-RNA Midiprep Kit Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
INSTRUCTION MANUAL Quick-RNA Midiprep Kit Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
Department of Molecular Genetics and Microbiology, School of Medicine, State University of New York at Stony Brook, Stony Brook, New York 11794
 JOURNAL OF VIROLOGY, May 2003, p. 5152 5166 Vol. 77, No. 9 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.9.5152 5166.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Functional
JOURNAL OF VIROLOGY, May 2003, p. 5152 5166 Vol. 77, No. 9 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.9.5152 5166.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Functional
Characterizing intra-host influenza virus populations to predict emergence
 Characterizing intra-host influenza virus populations to predict emergence June 12, 2012 Forum on Microbial Threats Washington, DC Elodie Ghedin Center for Vaccine Research Dept. Computational & Systems
Characterizing intra-host influenza virus populations to predict emergence June 12, 2012 Forum on Microbial Threats Washington, DC Elodie Ghedin Center for Vaccine Research Dept. Computational & Systems
SUPPLEMENTARY RESULTS
 SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and Gerard D. Wright.
 Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
Mechanisms of alternative splicing regulation
 Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.