T HE normal epidermis consists of layers whose
|
|
|
- Carmel Adele Hensley
- 5 years ago
- Views:
Transcription

1 Published Online: 1 August, 1988 Supp Info: Downloaded from jcb.rupress.org on January 17, 2019 Use of Monospecific Antisera and crna Probes to Localize the Major Changes in Keratin Expression during Normal and Abnormal Epidermal Differentiation Andrea Stoler,* Raphael Kopan,* Madeleine Duvic, and Elaine Fuchs* * Departments of Molecular Genetics and Cell Biology and Biochemistry and Molecular Biology, The University of Chicago, Chicago, Illinois 60637; and ~ Department of Dermatology, University of Texas Health Science Center, Houston, Texas Abstract. We report here the isolation and characterization of three antisera, each of which is specific for a single keratin from one of the three different pairs (K1/KI0, K14/K5, K16/K6) that are differentially expressed in normal human epidermis and in epidermal diseases of hyperproliferation. We have used these antisera in conjunction with monospecific crna probes for epidermal keratin mrnas to investigate pathways of differentiation in human epidermis and epidermal diseases in vivo and in epidermal cells cultured from normal skin and from squamous cell carcinomas in vitro. Specifically, our results suggest that: (a) the basalspecific keratin mrnas are down-regulated upon commitment to terminal differentiation, but their encoded proteins are stable, and can be detected throughout the spinous layers; (b) the hyperproliferation-associated keratin mrnas are expressed at a low level throughout normal epidermis when their encoded proteins are not expressed, but are synthesized at high levels in the suprabasal layers of hyperproliferating epidermis, coincident with the induced expression of the hyperproliferation-associated keratins in these cells; and (c) concomitantly with the induction of the hyperproliferation-associated keratins in the suprabasal layers of the epidermis is the down-regulation of the expression of the terminal differentiation-specific keratins. These data have important implications for our understanding of normal epidermal differentiation and the deviations from this process in the course of epidermal diseases of hyperproliferation. T HE normal epidermis consists of layers whose cells possess distinct structure and properties. The cells in the basal layer proliferate; as cells leave this layer and begin to migrate outward towards the skin surface, they cease to divide and undergo a series of biochemical and morphological changes that culminate in the production of a tough, protective outer coat (stratum corneum) of dead, terminally differentiated cells. A number of changes in protein synthesis take place during the process of terminal differentiation. The major changes are in the expression of keratins, a family of proteins (40-67 kd) which assemble into 8-nm structural filaments in the cytoplasm of all epidermal cells. Basal cells express only two keratins, K5 (58 kd) and K14 (50 kd) (Nelson and Sun, 1983). From tissue sectioning and keratin extractions, it is thought that the expression of these keratins may still continue in the suprabasal layers (Fuchs and Green, 1980; Woodcock-Mitchell et al., 1982; Skerrow and Skerrow, 1983). In situ hybridizations on the other hand have indicated that K14 and I(5 mrna expression is predominantly basal (Tyner and Fuchs, 1986; Lersch and Fuchs, 1988). Suprabasal cells express K1 (67 kd), K2 (65 kd), K10 (56.5 kd), and Kll (56 kd) (Viac et al., 1980; Woodcock- Mitchell et al., 1982; Tseng et al., 1982; Skerrow and Sker- row, 1983; Kopan et al., 1987). While K1 and K10 arise from the induction of new mrnas, K2 and K11 appear later and seem to be proteolytic degradation products of K1 and K10, respectively (Fuchs and Green, 1980; Tyner and Fuchs, 1986; Kopan et al., 1987). In humans, expression of large keratins is highly restricted to keratinizing cells of epidermis, exocervix, and anus (Moll et al., 1982a). The shift to the expression of K1 and K10 is one of the earliest biochemical indications that a cell has undergone a commitment towards terminal differentiation. The extent to which the decline in K5 and K14 synthesis and the induction of K1 and K10 synthesis are linked in epidermal differentiation has not yet been fully evaluated. In the course of human epidermal diseases of hyperproliferation, a number of different morphological and biochemical aberrations have been observed. The most significant differences seem to be the reduction in the expression of K1 and K10 (Baden et al., 1978; Hunter and Skerrow, 1981; Thaler et al., 1980; Weiss et al., 1984; McGuire et al., 1984), and the induction of a new set of keratins, K6 (56 kd) and KI6 (48 kd) (Weiss et al., 1984). In normal epidermis, K6 and K16 are only expressed transiently, during wound healing (Lane et al., 1985; Mansbridge and Knapp, 1987) or when skin is placed into tissue culture medium in vitro (Sun The Rockefeller University Press, /88/08/427/20 $2.00 The Journal of Cell Biology, Volume 107, August

2 and Green, 1978; Fuchs and Green, 1980; Tyner and Fuchs, 1986). Most of the documented abnormalities of keratin synthesis in epidermal diseases stem from studies involving protein extraction, electrophoretic separation, or immunoblot analysis. While some immunohistochemical studies have been attempted (Raemakers et al., 1983; Thomas et al., 1984; Leigh et al., 1985; Weiss et al, 1984; Knight et al., 1985; Robinson, 1987), they have been hampered by the lack of monospecific antibodies against specific epidermal keratins. Thus, it has been difficult to assess precisely which cells of the epidermis undergo these changes and the extent to which these biochemical changes might be associated with morphological alterations. In this report, we describe the preparation and characterization of three antisera, each of which is specific for one of the three sets of epidermal keratins (basal, keratinization associated, and hyperproliferation associated). Using three specific epidermal keratin crna probes in conjunction with immunohistochemical localization, we investigate the expression of keratin mrnas and keratin proteins in normal epidermis, and in epidermal diseases of hyperproliferation. We also report the use of an epidermal cell culture method as a model system for studying the morphology and biochemistry of both normal and tumor epidermal cells in vitro. This method involves seeding epidermal cells on a lattice of collagen and fibroblasts, and then floating the artificial skin on a raft to enable epidermal cells to be fed from beneath (Lillie et al., 1980; Karasek et al., 1971; Asselineau et al., 1985; 1986; Kopan et al., 1987). Using this system to culture normal epidermal cells, and the squamous cell carcinoma cell lines SCC-13, SCC-12, and SCC-15, we examine the morphological and biochemical similarities between these cells cultured on rafts and their in vivo counterparts. Materials and Methods Preparation of Synthetic Peptides and Monospecific Antibodies Peptides to K1, K14, and K16 were synthesized, and then coupled to keyhole limpet haemocyanin using glutaraldehyde (K1 and KIt) or m-maleimidobenzoic acid N-hydroxysuccinimide ester (K14) to produce essentially NH2- terminally bound peptide conjugates (Paul Sheppard, Cambridge Research Biochemicals Cambridge, England). At a concentration of 300 ~tg/ml in Freund's complete adjuvant, these conjugates were injected into 7-1b New Zealand White male rabbits whose sera had been pretested by immunofluorescence and immunoblot analysis to make certain it contained no endogenous keratin antibodies. For each peptide, three rabbits were injected. After 3 wk, each rabbit was boosted with 300 ~tg of conjugate (mixed with Freund's incomplete adjuvant), and after a 10-d period, the antisera was tested for the presence of keratin antibodies. If no appreciable titer was obtained, the animals were reboosted with 300 ~tg of conjugate every 14 d until a strong positive reaction was obtained either by immunofluorescence or immunoblot assays. In most cases, antisera reached a maximum titer at approximately the fifth boost. The antiserum with the strongest and most specific titer was chosen for the analyses presented here. These antisera will be referred to as antihk1 (human K1 antiserum), anti-hk14 (human K14 antiserum), and anti-hkl6 (human KI6 antiserum). Preparation of Epidermal Raft Cultures Collagen lattices were prepared and seeded with human keratinocytes from the squamous cell carcinoma lines SCC-12, SCC-13, and SCC-15 (Wu and Rheinwald, 1981) as described previously (Asselineau et al., 1985; Kopan et al., 1987). Cells were grown submerged in culture medium for 1 wk, and then floated on metal rafts for 7-14 d before fixation and sectioning (5 i.tm). Antiserum Staining of 1Issue and Raft Sections Six squamous cell carcinomas, four basal cell carcinomas, one actinic keratosis, and three psoriatic lesions were used for the studies reported here. Unless otherwise indicated, tissue and raft samples were fixed in 4% paraformaldehyde, embedded in paraffin, and sectioned (5 I.tm). Most sections were subjected to immunohistochemistry using the immunogold localization technique as described by the manufacturer, Janssen Life Sciences Products (Piscataway, NJ). Briefly, after incubation of the tissue sections with the rabbit anti-keratin antibodies, samples were exposed to colloidal gold-conjugated goat anti-rabbit secondary antibodies. Antibody staining was then enhanced by silver precipitation. Antisera were used at the following dilutions: anti-hkl4, 1:50; anti-hk1, 1:50; anti-hk16, 1:25. Some of the samples were subjected to immunohistocbemistry using the VECTASTAIN ABC Kit as described by the manufacturer (Vector Laboratories, Inc., Burlingame, CA). Briefly, the section was first exposed to primary antibody, followed by a biotinylated secondary antibody, and then a preformed avidin and biotinylated alkaline phosphatase macromolecular complex. The enzymatic activity was localized through the use of a substrate kit (Vector Laboratories, Inc.), which coupled a soluble product (generated during the hydrolytic reaction) with a capture reagent, producing a blue insoluble precipitate. Epidermal and squamous cell carcinoma cultures from floating rafts were fixed in Carnoy's solution and subjected to immunohistochemistry using classical indirect immunofluorescence. In these cases, fluorescein-conjugated goat anti-rabbit IgG was used as a conjugate. Subcloning of Coding and 3' Noncoding Keratin edna Segments into Riboprobe Vectors K14 edna Subclones. KB-2 is a 1,410-bp edna insert encoding keratin K14 (Hanukoglu and Fuchs, 1983). It has a single TGA stop codon positioned at residue 1,230. A 1,080-bp Bste II/Stu I fragment (nucleotide residues 170-1,250) encompassed a major portion of the coding region of K14 and was subcloned into psp65. K6a edna Subciones. A 1,415-bp Taq I/Kpn I fragment of the 1,680-bp edna KA-1 (Hanukoglu and Fuchs, 1983; residues 20-1,435) containing predominantly the coding region of K6 was subcloned into Escherichia coli plasmid psp64 (Promega Biotec, Madison, WI). Kl6 edna Subclones. The sequence for the complete gene encoding the human keratin K16 has been reported previously (RayChandhury et al., 1986). To prepare a specific cdna clone for this keratin mrna, a 658-bp Pst I/Eco RI fragment containing part of intron VII, exon VIII, and the 3' noncoding portion of the KI6 gene was subcloned in the 3'-5' direction into plasmid pgem 1 (Promega Biotec). Radiolabeling of crna Probes Cloned cdnas were linearized, and Salmonella (SP6) or T7 RNA polymerase was used to prepare 3~S-labeled crna probes. The method of Cox et al. (1984) was followed as described by the manufacturer (Promega Biotec). In Situ Hybridizations In situ hybridizations were performed essentially as described by Cox et al. (1984). Briefly, tissue sections were prepared as described above and treated with 1.0 I.tg/ml proteinase K for 30 min at 37 C and hybridized with 35S- UTP-labeled crna probes that had been hydrolyzed to an average size of 150 bp by sodium carbonate treatment at 60 C. Hybridization conditions were: 50% recrystallized formamide, 0.3 M NaCl, 5 mm EDTA, 20 mm Tris-HCI, ph 8.0, 100 mm dithiothreitol (DTT), 0.02 % BSA, 0.02 % Ficoll, 0.02% polyvinylpyrollidone, and 10% dextran sulfate for 12 h at 41 C. After hybridization, tissue sections were treated with RNase to remove any unhybridized probe and washed with (a) 300 mm NaCl, 30 mm sodium citrate (2x SSC) containing 10 mm DDT at 25 C for 30 min, to quench any nonspecific hybridization and (b) 0.1 SSC containing 10 mm DTT for 1 h at 50 C. Autoradiography was performed using an NTB-2 Kodak emulsion. Samples were exposed for 3-8 d as indicated in the text. Results Design, Preparation, and Characterization of the Antikeratin Antisera Previously, genomic sequences have been reported for the The Journal of Cell Biology, Volume 107,

3 human epidermal keratins K1 (Johnson et al., 1985), K14 (Marchuk et al., 1984, 1985), and K16 (RayChaudhury et al., 1986; Rosenberg et al., 1988). Secondary structure analyses of their predicted amino acid sequences revealed features characteristic of all keratins, namely a central alphahelical region containing hydrophobic repeats indicative of an ability to form a coiled-coil structure, and the presence of an L L E G E sequence at the carboxy-terminal end of the helical region (for review, see Geisler et al., 1982; Fuchs and Hanukoglu, 1983; Steinert et al., 1985). A comparison of amino acid sequences indicated that the most divergent portion of all keratin polypeptides is in the nonhelical domain extending from the L L E G E to the carboxy terminus. These sequences are also thought to protrude along the surface of the keratin filament (Steinert et al., 1983). As haptens for the preparation of monospecific antisera, peptides were chosen to keratin sequences which were located primarily in this nonhelical carboxy-terminal domain. Within this region, segments which either (a) share homology with other known keratin sequences, or (b) contained stretches of a single amino acid residue (e.g., serine) were avoided. Fig. 1 shows the sequences (underlined) which were selected for the production of synthetic peptides and for antibody preparation. Except for the K16 peptide, the entire sequence chosen for peptide synthesis was also contained in the corresponding keratin protein. For anti-hk16, only the carboxy-terminal QSS corresponded to the KI6 sequence (Rosenberg et al., 1988). When used at a concentration of 300 lag/ml of antiserum, a synthetic heptapeptide to the carboxy-terminal sequence FSQGQSS of the K16 protein quantitatively and specifically preadsorbed nearly all of the K16-binding activity of the antiserum. Other proteins (e.g., K14), which have an internal QSS sequence, showed no cross-reactivity with the antiserum (data not shown). Hence, the free carboxyl group of the QSS sequence located at the terminus of the K16 protein seems to be essential to its recognition by this antiserum. The specificity and titers of the antisera were tested by immunoblot analyses of one- and two-dimensional SDS-polyacrylamide gels containing resolved keratins isolated from human epidermis (K1, K2 +, K5 +, K10 +, Kll, K14 ), human breast carcinoma (MCF-7) cells (K8, K18, K19 ), and cultured human epidermal cells (K5 +, K6, K13 +, K14, K15, K16, K17 ) (Fig. 2). Our results indicate that all three antisera are largely monospecific for their respective keratin, with little or no cross-reactivity with any of the other keratins tested. Immunoblot analysis with whole cell extracts further verified the specificity of the antisera (not shown). Peptide Antisera Specific for K1 and K14 Recognize Their Determinants in Human Skin In Vivo To test the usefulness of these antisera for immunohistochemistry, we prepared 4 % paraformaldehyde-fixed, paraffin-embedded sections of human forearm skin and stained them indirectly using the immunogold staining-silver enhancement procedure or the avidin-biotinylated-alkaline phosphatase complex (ABC) staining procedure as described in Materials and Methods (Fig. 3). The anti-hk1 antiserum showed pronounced staining of all suprabasal layers of the epidermis (Fig. 3 A). This staining pattern is consistent with results obtained from tissue sectioning (Fuchs and Green, 1980; Woodcock-Mitchell et al., 1982) and from staining with broadly cross-reacting anti-s, corneum keratin antibodies (Viac et al., 1980; Tseng et al., 1982; Lane et al., 1985). Moreover, the suprabasal expression of the type H keratin K1 coincides with the suprabasal expression of its type I partner K10 (Raemakers et al., 1983; Kopan et al., 1987). In contrast to the pattern of anti-hk1 staining, the staining of anti-hkl4 revealed abundant expression of this keratin in the basal epidermal layer (Fig. 3 B; in particular, note inset). Staining was also observed in the spinous and granular layers of the epidermis, although it was not quite as prominent as in the basal layer. While these data are in agreement with earlier sectioning studies (Fuchs and Green, 1980; Woodcock-Mitchell et al., 1982), they differ from immunohistochemical data obtained with the monoclonal anti-type I keratin antibody AE-1 (Woodcock-Mitchell et al., 1982), which showed only basal staining of the epidermis. Our results demonstrate clearly that K14 persists in the spinous layers, and confirm that the antigenic site for type I keratins that is recognized by AE-1 is masked in these layers. As expected, no staining of normal epidermis was observed with the anti-hk16 antiserum (Fig. 3 C). This finding is consistent with the fact that K16 protein has not been detected in keratin extracts from normal epidermis (Moll et al., 1982a, 1983; Woodcock-Mitchell et al., 1982; Tseng et al., 1982; Tyner and Fuchs, 1986). KI (67 kd): L L E G g g S R M ~_ E ~_ A ~_ N V S V T A S ~_ ~_ ~_ T S I S... K14 C50 kd): C C ~ V V ~_ ~ H ~_ ~_ ~_ L R ~ K ~[-COOIt K16 (48 kd): R P I L ~- ~ S-- g ~. Q - ~ S K R S S 1~ G-- g P F S ~ E ~-COOI~G 9_ ~. ~ S S-COOH Figure 1. Keratin sequences selected for synthetic peptides and antibody preparations. Amino acids sequences chosen for synthetic peptide preparation are shown as underlined residues. Additional sequences are provided for reference and include either the highly conserved L L E G E sequence at the end of the fourth helical domain of all intermediate filament proteins (shown for the human keratin K1) or the carboxy-terminal end of the keratin polypeptide (shown for the human keratins K14 and K16). For hk14, the Cys residue was not present in the K14 sequence, but was added for chemical cross-linking purposes (see Materials and Methods). For hk16, the sequence chosen for antibody production is shown on the line beneath the actual hk16 carboxy-terminal end. Note that only the three terminal residues of hk16 are homologous. Examining the Altered Pattern of Keratin Expression in Hyperproliferating Epidermis Psoriasis is a common disease of epidermal hyperproliferation, characterized by the lack of a well-defined granular layer and the formation of an abnormal, nucleated stratum corneum (referred to as parakeratinization; Fitzpatrick et al., 1979). The degree of parakeratinization has been correlated with the degree of abnormal keratin expression: weakly parakeratinized tissue has a keratin pattern that resembles normal epidermis (K1 +, K2 +, K5 +, K10 +, Kll +, K14+), whereas the pattern seen in heavily parakeratinized tissue is almost indistinguishable from that of cultured keratinocytes (K5 +, K6 +, K14 +, K16 +) (Weiss et al., 1984). Hence, the most striking changes in keratin expression in psoriatic epidermis include a reduction in K1/K10 and an induction of Stoler et al. Keratin Expression in Epidermal Diseases 429
 Human intermediate filament proteins were extracted from autopsy skin (KI +, K2 +, K5 +, K10 +, Kll +, K14+), cultured MCF-7 (breast carcinoma) cells (K8 +, K18 +, K19+), and cultured epidermal")
 or transferred to nitrocellulose paper by electroblotting (Towbin et al., 1979).")
4 Figure 2. Immunoblot analysis of anti-hkl, anti-hkl4, and anti-hkl6. (.4) Human intermediate filament proteins were extracted from autopsy skin (KI +, K2 +, K5 +, K10 +, Kll +, K14+), cultured MCF-7 (breast carcinoma) cells (K8 +, K18 +, K19+), and cultured epidermal cells (K5 +, K6 +, K13 +, K14 +, K16 +, K17+). Proteins were loaded in this order (lanes 1-3, respectively) and were resolved by one-dimensional PAGE. Proteins were either visualized by staining with Coomassie Blue (marked stain) or transferred to nitrocellulose paper by electroblotting (Towbin et al., 1979). Immunoblots were first stained with fast green to visualize the proteins, and then exposed to either antihkl4 ( t K14), anti-hk16 (ctkl6), or anti-hk1 ( tk1). After exposure to antiserum, blots were washed and then hybridized with IESI-radiolabeled S. aureus protein A to visualize antibody-bound proteins. Hybridized blots were exposed to x-ray film for 48 h. Keratins were identified by number according to their molecular masses and their isoelectric mobilities as indicated by Moll et al. (1982a). Note: K6 at left identifies the band in the cultured cell extract (lane 3). K10, the band seen in the skin keratin extract (lane 1), has similar electrophoretic mobility to K6. (B) Coomassie Blue staining of cultured human epidermal cell keratins, resolved by two-dimensional gel electrophoresis, with a nonequilibrium ph gel in the first dimension and SDS-PAGE in the second (O'Farrell, et al., 1977). The arrow indicates ch~mges in isoelectric mobilities of keratins due to the ability of type I and type II keratins to form complexes, which are not fully resolved in the first dimension. (C) Immunoblot of a two-dimensional gel of cultured epidermal cell keratins resolved as described in B and probed with anti-hk14 as described in A. Note that two spots were selected: one corresponding to the bona fide mobility of KI4, and one corresponding to the complex formed between K14 and its partner K5, which was not resolved in the first dimension (see arrow in B). (D) Immunoblot of a two-dimensional gel of cultured epidermal cell keratins resolved as described in B and probed with anti-hkl6 as described in A. Note that only one spot was selected, since K16 does not form strong complexes with type II keratins (see B). the so called hyperproliferation-associated keratins K6 and K16 (Baden et al., 1978; Thaler et al., 1980; Hunter and Skerrow, 1981; Matoltsy et al., 1983; Bowden et al., 1983; Weiss et al., 1984; McGuire et al., 1984). To examine how individual cells within psoriatic tissue alter their expression of keratin, we used our specific antisera to stain epidermis from samples of strongly parakeratinized psoriatic tissues (Fig. 4). K1 could readily be detected in most suprabasal cells of psoriatic tissue (Fig. 4 A). However, staining was weaker than it was for normal epidermis, particularly in the deeper suprabasal cells. K14 was also detected in psoriatic tissue (Fig. 4 B). Its localization differed slightly from normal epidermis in that anti-hk14 staining was seen in all epidermal cells, extending even to the s. corneum layer. Since no new protein synthesis takes place in this region of the skin, these data suggest that mechanisms that control proteolytic degradation of K14 in the outer layers of normal epidermis do not function in psoriatic epidermis. Interestingly, the pattern of staining observed with antihk16 was very similar to that seen with anti-hk1, with strong staining in the upper spinous and granular layers of psoriatic tissue and none in the basal cells (Fig. 4 C). Hence a correlation seemed to exist between the reduction in the expression of the terminal differentiation-specific keratins and the in- The Journal of Cell Biology, Volume 107,
 was fixed in 4% paraformaldehyde, sectioned (5 gm) and exposed to anti-hk1 (A), anti-hkl4 (B), and anti-hk16 (C).")
5 Figure 3. Anti-keratin staining of normal human epidermis. Normal human skin (from biopsy) was fixed in 4% paraformaldehyde, sectioned (5 gm) and exposed to anti-hk1 (A), anti-hkl4 (B), and anti-hk16 (C). Bound antibodies were then visualized by indirect immunohistochemistry, using the alkaline phosphatase ABC Vectastain method (A-C) or the immunogold labeling and silver enhancement technique (inset, B) as described in Materials and Methods. Sections were examined by photomicroscopy. Note that anti-hkl4 stained the basal epidermal layer more strongly than the suprabasal layers. This is particularly evident in the example shown in the inset of B. Bar, 20 ~m. duction of the hyperproliferation-associated keratins. Moreover, as judged by keratin expression, the major alterations in psoriatic tissue were clearly manifested in the spinous layers, rather than in the basal cells, which still seemed to possess unique structural and biochemical properties characteristic of normal basal epidermal cells. Stoler et al. Keratin Expression in Epidermal Diseases 431
 was fixed in 4% paraformaldehyde, sectioned (5 lam) and exposed to anti-hk1 (A), anti-hkl4 (B), and anti-hkl6 (C).")
6 The Journal of Cell Biology. Volume 107, Figure 4. Anti-keratin staining of psoriatic epidermis. Psoriatic epidermis (from biopsy) was fixed in 4% paraformaldehyde, sectioned (5 lam) and exposed to anti-hk1 (A), anti-hkl4 (B), and anti-hkl6 (C). Bound antibodies were then visualized by indirect immunohistochemistry, using the alkaline phosphatase ABC Vectastain method described in Materials and Methods. Sections were examined by photomicroscopy. Bar, 74 I-tm.
, and exposed to anti-hk1 (A, B, and G), anti-hk14 (Cand D), and antihk16 (E, F, H-J).")
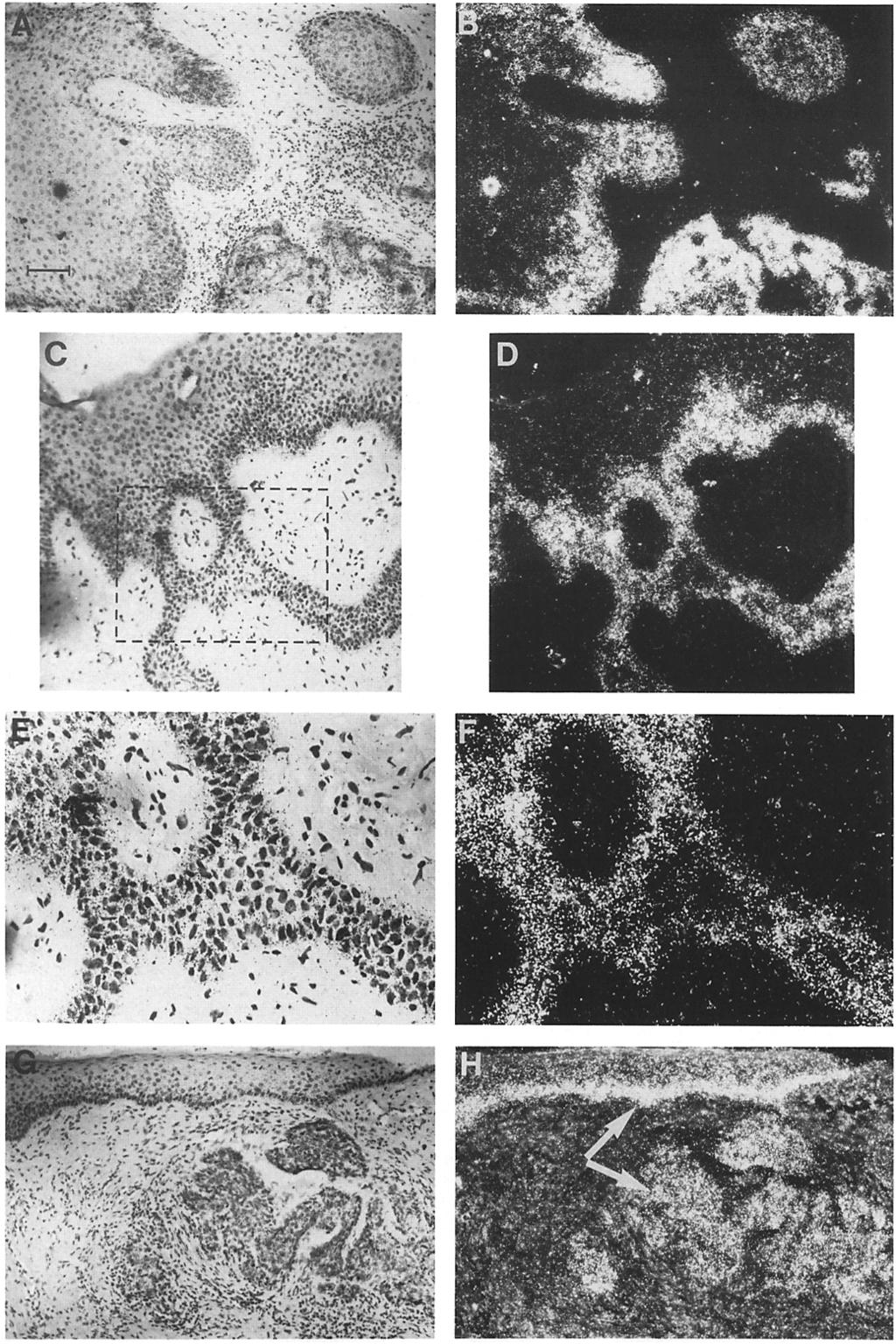
7 Stoler et al. Keratin Expression in Epidermal Diseases 433 Figure 5. Anti-keratin staining of squamous cell carcinomas. Tissue samples from six squamous cell carcinomas were fixed in 4 % paraformaldehyde, sectioned (5 Ixm), and exposed to anti-hk1 (A, B, and G), anti-hk14 (Cand D), and antihk16 (E, F, H-J). Antibody localization was visualized using the immunogold labeling and silver enhancement technique of Janssen Life Sciences Products. Representative samples shown were from a well-differentiated squamous cell carcinoma (A, C, and E), a poorly differentiated squamous cell carcinoma (B, D, and F), a moderately differentiated squamous cell carcinoma (G and H), actinic keratosis (I), and from the epidermis adjacent to a welldifferentiated squamous cell carcinoma (J). Note the presence of keratinizing squames in the well-differentiated and moderately differentiated tumors and the relative lack of differentiation and organization in the poorly differentiated sample. Note the relative absence of basal-like cells in the particular example shown of a moderately differentiated tumor. Note also that the expression of K1 is largely uniform in all of the cells of this tumor, as opposed to that seen in the well-differentiated tumor. Both anti-hk1 and antihk16 correlated well with keratinization in SQCC samples. Note the presence of KI6 in the suprabasal layers of the pre-squamous cell carcinoma sample of actinic keratosis. Also note the abnormal presence of K16 in the epidermis adjacent to a well-differentiated squamous cell carcinoma (J). Bar: (A-H) 30 Ixm; (I and J) 74 ~tm.
 and exposed to anti-hkl (A), anti-hk14 (frame B), and anti-hkl6 (C).")
. The boundaries of the tumor are indicated by the dotted line. Bar, 30 Itm.")
8 Figure 6. Anti-keratin staining of basal cell carcinomas. Samples from a superficial l~asal cell carcinoma were fixed in 4% paraformaldehyde, sectioned (5 tim) and exposed to anti-hkl (A), anti-hk14 (frame B), and anti-hkl6 (C). Bound antibodies were visualized by indirect immunohistochemistry, using the immunogold enhancement technique as described in Materials and Methods. Sections were examined by photomicroscopy. Note the abnormal expression of KI6 in the epidermis adjacent to the tumor (C). The boundaries of the tumor are indicated by the dotted line. Bar, 30 Itm. Altered Patterns of Keratin Expression in Specific Cells of Squamous and Basal Cell Carcinomas Squamous cell carcinomas (SQCCs) I are typified by the presence of transformed, squamous-like keratinocytes (for review, see Fitzpatrick et al., 1979). Well-differentiated SQCCs have more organized, basal-like cells at the periphery of the tumor mass, and keratinized "pearls" of differentiated epidermal squames located in the center. Poorly differentiated SQCCS do not show much variation in cell morphology. Cells tend to be squamous-like and irregular, with little evidence of keratinization or of organized basal cells. Previously, it was shown that while KI and KI0 expression is lower in squamous cell carcinoma tissue, the keratinized centers of well-differentiated tumors contain terminal differentiation-specific keratins (Robinson, 1987; Thomas et al., 1984). Indeed, when we stained a section of a well-differentiated SQCC with our anti-hkl antiserum, K1 was readily detected in the keratinized centers (Fig. 5 A). Markedly reduced staining was observed towards the periphery of the tumor mass, and in the unkeratinized areas of the tumor. The association of anti-hkl staining and squamous, keratinized morphology was observed in all samples of squarnous cell carcinoma that we investigated. Thus, even in poorly differentiated squamous cell carcinomas, where the expression of K1 and KI0 has rarely been detected by gel electrophoretic analyses (Weiss et al., 1984; Thomas et al., 1984; Moll et al., 1983, 1984; Breitkreutz et al., 1981), anti-hkl staining could be seen in regions where the squamous morphology of tumor cells was greatest (Fig. 5 B). A particularly interesting example of this was in a moderately differentiated 1. Abbreviation used in this paper: SQCC(s), squamous cell carcinoma(s). squamous cell carcinoma, where most, if not all of the cells of the tumor appeared to be squamous in morphology and stained very brightly with anti-hk1 antiserum (Fig. 5 G). Our antiserum against hk14 consistently showed prominent staining throughout all of the cells of each squamous cell carcinoma (Fig. 5, C and D). In some cases of welldifferentiated SQCCs (such as the example in Fig. 5 C), antihkl4 staining was lower in the basal-like cells than in the central, more squamous regions. In other squamous cell carcinoma samples, wide variation in anti-hkl4 staining was often observed, but no strong correlation with cell morphology could be made. Most SQCCs showed patterns of anti-hk16 staining that were comparable to those seen for psoriatic tissue. The expression of K16 was greatest in well-differentiated tumors and seemed to be most prevalent in the squamous (central) areas of the carcinoma, with markedly reduced levels in the less differentiated (marginal) areas (Fig. 5 E). Poorly differentiated tumors showed very little anti-hk16 staining, with only a few cells staining appreciably above background levels (Fig. 5 F). Moderately differentiated tumors showed variable levels of anti-hkl6 staining: in the example shown (Fig. 5 H), K16 expression was low. These findings are in agreement with previous microdissection analyses of squamous cell carcinomas (Moll et al., 1983, 1984), and further emphasize the correlation between anti-hk16 staining and squamous morphology. Interestingly, K16 expression was readily detected in the suprabasal layers from the lesions of a patient with actinic keratosis, a premalignant hyperproliferative disease which is somewhat similar in appearance to psoriasis, (Fig. 5 I; Fitzpatrick et al., 1979). While anti-hk14 staining was similar to that seen in normal epidermis, anti-hk1 staining appeared to be somewhat reduced, and its location paralleled the anti- The Journal of Cell Biology, Volume 107,
 and hybridized with 35S-UTP-labeled crna probe complementary to human K14 mrna.")
9 Figure 7. In situ localization of K14 mrna in normal epidermis and psoriatic epidermis. Tissue samples were fixed in 4% paraformaldehyde, sectioned (5 ~tm) and hybridized with 35S-UTP-labeled crna probe complementary to human K14 mrna. After hybridization, samples were washed, treated briefly with single-stranded RNase, and exposed to an NTB-2 Kodak autoradiographic emulsion as described in Materials and Methods. Exposed silver grains were developed after 3 d and samples were stained with hematoxylin and eosin to visualize tissue morphology. Sections were examined by photomicroscopy using both brightfield (left) and darkfield (right). Samples were from normal epidermis (A and B) and psoriatic epidermis (C-F), Sample shown in E and F represents a magnified (3.85x) version of an area in frame C. Bar: (A, B, E, and F) 30 gm; (C and D) 74 Ixm. hk16 pattern (data not shown). Moreover, in all of the samples of squamous cell carcinoma that we investigated, abnormal expression of K16 was detected in the adjacent regions of seemingly normal epidermis (Fig. 5 J). This unusual phenomenon has been reported previously using a less specific antibody (Weiss et al., 1983). Whether this abnormal expression occurred as a consequence of some extracellular stimulus released either by the tumor cells or the surrounding damaged tissue was not determined. Thus, K16/K6 expression occurred in epidermal tissue of various and diverse morphologies. The unifying feature of cells expressing these keratins seemed to be their suprabasal location and their association with hyperproliferation or trauma. The patterns of anti-keratin staining of relatively undiffer- Stoler et al. Keratin Expression in Epidermal Diseases 435
10
.")
11 entiated basal cell carcinomas were distinctly different from those of squamous cell carcinomas (Fig. 6). K1 appeared to be extremely low or absent in most tumor cells (Fig. 6 A). In some tumors, K1 was even reduced or absent in two to three suprabasal layers of the adjacent epidermis (data not shown). In contrast, K14 was present in all of the cells of all basal cell carcinomas that we examined (Fig. 6 B). However, the levels of K14 staining were variable, and in some tumors, K14 expression seemed to be markedly reduced when compared with the adjacent basal layer. Surprisingly, while K16 was not detected in any of the basal cell carcinomas that we examined, it was abnormally expressed in the adjacent areas of epidermis (Fig. 6 C). Hence, although the pattern of keratin expression throughout the cells of basal cell carcinomas seemed to be quite uniform and purely basal-like in character, these tumors caused changes in the differentiation processes in adjacent regions of the epidermis. Altered Production of Keratins in Hyperproliferative Epidermal Diseases Appears to Be Largely at the Level of mrna Expression To examine the correlation between mrna and protein expression for the epidermal keratins, we conducted in situ hybridization analyses using radiolabeled crna probes complementary to the mrnas encoding keratins K14, K6a, and K16 (Figs. 7-10). Previously, we had conducted Northern blot analysis and in situ hybridizations to verify the specificity of these probes (Tyner and Fuchs, 1986; RayChaudhury et al., 1986; Rosenberg et al., 1988). When an 35S-UTP-labeled crna probe complementary to a large portion of the coding region of the K14 mrna was hybridized with normal foreskin epidermis, abundant grains were localized over the basal layer of normal epidermis, with low, but detectable hybridization extending into the spinous epidermal layers (Fig. 7, A and B). The dramatic decline in KI4 mrna expression seen during the transit of a normal epidermal cell from the basal to the first suprabasal layer was also observed in psoriatic tissue (C-F). Once again, some labeling was seen in the spinous layers, and even the outermost epidermal layers showed some hybridization. In both normal and psoriatic tissue, the down-regulation of K14 mrna expression in the basal-suprabasal transition seemed to be more striking than the corresponding decrease in K14 protein. The expression of K14 mrna in squamous cell carcinomas was quite variable (Fig. 8, A and B). In well-differentiated regions of the squamous cell carcinomas, clusters of tumor cells frequently showed high grain density (e.g., the lower right cluster of cells in A and B). Leading portions of tumor masses, especially those near the skin surface, also showed high levels of K14 mrna. This pattern of K14 mrna distribution was similar to that observed in normal epidermis, and was frequent in more organized regions of the tu- mor and in cells having the most basal-like appearance (see Fig. 8, C and D, and close-ups in E and F). Some masses of SQCC cells showed levels of KI4 mrnas which were substantially lower than the adjacent regions of normal epidermis (see for example, the small cluster at the upper right in Fig. 8, A and B). This reduction in keratin mrna expression was also observed for cells of some basal cell carcinomas (Fig. 8, G and H; compare basal layer of overlying epidermis with tumor mass in H). These data suggest that the overall level of keratin mrna expression may be significantly down-regulated in some malignant keratinocytes. To determine the extent to which transcriptional and posttranscriptional controls play a role in the induction of the hyperproliferation-associated keratins in different epidermal diseases, we used in situ hybridization with specific radiolabeled K6a and K16 crna probes to examine the level of K6 and K16 mrnas in normal skin, psoriatic skin, and basal and squamous cell carcinomas. After hybridizations with equal concentrations of radiolabeled probes specific for and complementary to (a) the coding portion of the human K6a mrna and (b) the noncoding portion of the human K16 mrna, tissue sections were exposed to photographic emulsion for 8 d (normal epidermis) or 3 d (all samples) before developing the exposed silver grains (Fig. 9 and 10). As had been observed previously (Tyner and Fuchs, 1986), at long exposure, K6a mrna could be detected throughout all of the living layers of normal epidermis at a low, but significant level (Fig. 9, A and B). This was true even when a 3' noncoding probe was used to identify K6a mrna (Tyner and Fuchs, 1986; Lersch and Fuchs, 1988). Upon shorter exposures, the presence of this mrna was more difficult to verify (Fig. 9 C). The expression of K16 mrna closely paralleled that of its partner K6 mrna in normal epidermis (Fig. 9 D). This was also true for other epidermal samples that we examined. In psoriatic tissue, the level of K6a mrna expression was greatly elevated over that seen in normal epidermis (Fig. 9, E and F). This increase in mrna was largely confined to the spinous layers. An increase in the grain density was also observed in the suprabasal layers of psoriatic tissue hybridized with the K16 probe (Fig. 9 G). Hence, the increase in K16 and K6 mrnas correlated well with the appearance of K16 (and presumably K6) protein in the suprabasal cells of psoriatic tissue. The expression of K16 and K6 mrnas was also elevated in some squamous cell carcinoma tissues (Fig. 10). Once again, the localization of these mrnas showed a pattern which was often opposite to that seen for the K14 mrna probe. In well-differentiated tumors, K6 and K16 mrnas were usually concentrated in the inner, more keratinized masses of the tumors, with lower levels in the basal-like cells (Fig. 10, A and B). Lower levels of K6/K16 mrnas were also found in all of the cells of poorly differentiated SQCCs (Fig. 10, C and D). However, the levels of expression of K6 and KI6 mrnas in at least some of the regions of all squa- Figure 8. In situ localization of K14 mrna in squamous and basal cell carcinoma. Tissue samples were fixed and subjected to in situ hybridization with 35S-UTP-labeled crna KI4 probe as described in the legend to Fig. 7. (Left) Brightfield; (right) darkfield. (A and B) A well-differentiated squamous cell carcinoma: note wide variability in hybridization for keratinized cells (lower cluster of cells, with strong hybridization), less keratinized cells (upper cluster, with weak hybridization) and leading edges of the tumor (at left); (C and D) peripheral regions of a well-differentiated squamous cell carcinoma; (E and F) enlargement (3.85 x) of the area contained within the dotted box shown in C; (G and H) a basal cell carcinoma: note that the overall expression of K14 mrna in this basal cell carcinoma was lower than in the basal layer of the epidermis adjacent to the tumor (comparison indicated by the arrows). Bar: (A-D and G and H) 74 gm; (E and F) 30 ~tm. Stoler et al. Keratin Expression in Epidermal Diseases 437

; (D) normal epidermis hybridized with a KI6 probe and exposed for 3 d (brightfield); (E and F) psoriatic epidermis hybridized with a K6a probe and exposed for 3 d (brightfield, E;")
12 Figure 9. In situ localization of K6/K16 mrnas in normal and psoriatic epidermis. Tissue samples were fixed and subjected to in situ hybridization with 3~S-UTP-labeled crna probe to either K6 or to K16 as described in the legend to Fig. 7. Samples were from: (A and B) normal epidermis hybridized with a K6a probe and exposed for 8 d (brightfield, A; darkfield, B); (C) normal epidermis hybridized with a K6a probe and exposed for 3 d (brightfield); (D) normal epidermis hybridized with a KI6 probe and exposed for 3 d (brightfield); (E and F) psoriatic epidermis hybridized with a K6a probe and exposed for 3 d (brightfield, E; darkfield, F); (G) psoriatic tissue hybridized with a KI6 probe and exposed for 3 d (brightfield, magnified 2.5 x over other samples). Note the suprabasal location of K6/K16 mrnas in psoriatic tissue. Bar: (A-D and G) 30 I.tm; (E and F) 74 p.m. The Journal of Cell Biology, Volume 107,

13 Figure 10. In situ localization of K6/K16 mrnas in squamous and basal cell carcinomas. Tissue samples were fixed and subjected to in situ hybridization with 35S-UTP-labeled crna probe to either K6a or to K16 as described in the legend to Fig. 7. Hybridizations were similar for both K6a and K16 probes, and only one of the two hybridizations is shown for each sample. (A, C, and E) Brightfield; (B, D, and F) darkfield. (A and B) A well-differentiated squamous cell carcinoma hybridized with a K6a probe; (C and D) a poorly differentiated squamous cell carcinoma hybridized with a K6a probe; (E and F) a basal cell carcinoma, hybridized with a K16 probe. The tumor boundaries in B and D are indicated by the dotted lines. Note that the expression of K6/K16 mrnas is high in the inner regions of the squamous cell carcinoma, whereas the number of grains in the tumor mass of the basal cell carcinoma was negligible. Note also that the epidermis adjacent to the tumor shows an increase in K16 grain density (indicated by the arrow in F). In E: e, epidermis; d, dermis; t, basal cell tumor. Bar: (A-D) 30 ~tm; (E and F) 74 ~tm. mous cell carcinomas were always higher than in normal epidermis. In addition, high levels of K6/K16 mrna expression were observed for the suprabasal squamous cells of the actinic keratosis sample that we examined (data not shown). Hence an increased expression of these mrnas may precede the malignant transformation leading to the development of squamous cell carcinomas. In striking contrast to squamous cell carcinomas, basal cell carcinomas showed a little or no evidence of K6/K16 mrna expression (Fig. 10, E and F). However, as can be seen by the arrow in Fig. 10 F,, the level of expression of the hyperproliferation-associated mrnas is elevated in the apparently normal epidermis adjacent to the tumor as compared with more distant regions. This increase in K6 and K16 Stoler et al. Keratin Expression in Epidermal Diseases 439
14 Table L Summary of Keratin Protein and mrna Expression in Human Epidermal Diseases K14 K6 KI6 Anti-hK 1 Anti-hK 14 Anti-hK 16 mrna mrna mrna Normal epidermis Basal Spinous S. corneum / + i /+ -/ Psoriatic Epidermis Basal Spinous Parakeratinized Squamous cell carcinoma Well differentiated leading edges -/ inner, keratinized + +/ Moderately differentiated +/ /+ + + Poorly differentiated + + Adjacent epidermis Basal Spinous-inner -/ Spinous-outer S. corneum / /+ -/ /+ +/++++ -/+ -/+ ++/++++ +/ / /+ --/ Basal cell carcinoma All tumor cells Adjacent epidermis Basal Spinous-inner Spinous-outer S. corneum + / / / + - / /+ -/+ --/ /+ + - Wherever slashes are shown, the levels of antibody staining or the expression of mrnas were variable. In these cases the value at the left of the slash indicates the minimum levels detected, while the value at the right of the slash indicates the maximum levels detected. mrna expression in the suprabasal cells of the epidermis adjacent to tumor masses was also observed for squamous cell carcinoma samples. Thus, for all of the tumor samples that we examined, the spinous-like cells which expressed KI6 protein also showed a corresponding increase in K6 and K16 mrnas. A summary of the immunohistochemical and in situ hybridization data obtained for normal, psoriatic, and tumor skin tissue is provided in Table I. K14 mrna and protein expression was most prominent in basal epidermal cells and in less differentiated cells of squamous and basal cell carcinomas. K1 expression was seen in terminally differentiating cells in epidermis and in keratinized regions of SQCCs, but not in basal cell carcinomas. K1 expression was down-regulated in psoriatic tissue and in tissue from actinic keratosis. Expression of the hyperproliferation-associated keratins was limited to the suprabasal cells of psoriatic tissue and actinic keratosis tissue, to the squamous, well-differentiated cells of squamous cell carcinomas, and to the epidermis surrounding a tumor. Basal epidermal cells showed no evidence of K6! K16 expression at the protein level, and only very low levels at the mrna level. K6/K16 induction in suprabasal cells was always accompanied by a corresponding and dramatic increase in mrna expression. Whether the low, base level expression of K6 and K16 mrnas observed in normal epider- mis might functionally contribute to the early stages of wound healing or in the development of epidermal disease remains to be determined. Morphology and Pattern of Expression of Keratins in Normal and Squamous Cell Carcinoma Cells Cultured on Floating CoUagen-Fibroblast Rafts In Vitro Is Parallel to That Seen In Vivo When normal human epidermal cells are cultured on collagen-fibroblast rafts that are subsequently (at confluence) floated at the air-medium interface, they stratify extensively and undergo many of the biochemical features characteristic of terminal differentiation (Fig. 11 A; see also Asselineau et al., 1986; Kopan et al., 1987). These features include the expression of low, but significant levels of KI and K10 in the suprabasal layers (Fig. 11 B; Kopan et al., 1987). The expression of K5/K14 and K16/K17/K6 in epidermal rafts has been reported (Asselineau et al., 1986; Kopan et al., 1987), but it has not been localized. When sections of epidermal rafts were stained with antihk14, K14 was detected throughout the epidermal layers, with expression being greatest in the basal layer, but with marked persistance in the suprabasal layers (Fig. 11 C). When rafts were stained with anti-hk16, K16 was most abun- The Journal of Cell Biology, Volume 107,
.")
15 Figure 11. Morphology and keratin expression in floating rafts of cultured human epidermal cells. Epidermal cells were cultured on floating rafts as described in Materials and Methods. "Rafts" were fixed in Carnoy's fixative and sectioned (5 lam). Sections were either stained with hematoxylin and eosin (A) or subjected to indirect immunofluorescence (B-D) with anti-hk1 (B), anti-hk14 (C), or anti-hk16 (D). Dotted line in D indicates the boundary between the epidermal culture and the artificial dermis which in this case did not detach from the sample. Bar: (A and D) 30 lam; (B and C) 20 ~m. dant in the suprabasal layers (Fig. 11 D). Hence, the pattern of keratin expression in epidermal rafts seemed to be very similar to that observed for psoriatic tissue (compare with Fig. 4), and for epidermis in the vicinity of either squamous or basal cell tumors (compare with Figs. 5 and 6). The finding that raft cultures of normal epidermal cells in vitro are similar to hyperproliferating epidermis in vivo led us to wonder whether SQCC lines grown on rafts might display morphological and biochemical features related to SQCCs in vivo. To test this possibility, the SQCC lines SCC- 12 (facial epidermis), SCC-15 (tongue), and SCC-13 (facial epidermis) were cultured on floating lattices of collagen and fibroblasts for d. Sections of these cultures revealed marked disorganization of the cells within the "tissues" and clear evidence of abnormal keratinization (Fig. 12, A-C). Each tumor line exhibited basal-like, spinous-like, and granular-containing cells (BL, SP, and GR in Fig. 12 C); however, the differentiative process varied greatly for each line. The SCC-12 line was interesting in that it seemed to invade the collagen-fibroblast layer, with epidermal growth extending deep into this lattice (Fig. 12 A). The other two lines showed little or no evidence of this behavior, but nonetheless had morphologies which were clearly abnormal (Fig. 12, B and C). In all three lines, a number of mitotic cells could be observed even in the outermost layers of some of the sections. To examine the pattern of keratin expression, sections of SCC-13 rafts were stained with anti-keratin antisera. SCC rafts were unusual in that they showed only patchy staining with anti-hk1 (Fig. 12, D and E). This keratin was restricted to suprabasal cells, and staining was most abundant in the central regions of the raft (Fig. 12 D). Less-stratified regions showed less K1 expression, which was also restricted to the suprabasal cells located in the centers, rather than the growing edges of these areas (Fig. 12 E). In well-stratified portions of the raft, K14 protein was detected in all layers, with strongest staining in the inner layers (see Fig. 12 F). In less-stratified portions on the raft, K14 expression seemed to be most abundant in the inner and outer layers of the tissue (Fig. 12 G). The pattern of K16 expression in SCC-13 rafts was similar to that obtained for K1 (Fig. 12, H and I). Hence, only suprabasal cells showed K16 staining, and this was most prominent in more stratified regions of the culture (Fig. 12 H). In most cases, poorly stratified areas at the growing edges of the raft cultures showed little or no K16 staining (Fig. 12 I). Stoler et al. Keratin Expression in Epidermal Diseases 441

 or subjected to indirect immunofluorescence (D-l) with anti-hkl (D and E), anti-hkl4 (F and G), or anti-hk16 (H and I).")
16 Figure 12. Morphology and keratin expression in floating rafts of cultured human squarnous cell carcinoma lines. Keratinocytes from squamous cell carcinoma lines of skin, SCC-12 and SCC-13, or tongue, SCC-15, were cultured on floating rafts as described in Materials and Methods. Rafts were fixed in Carnoy's fixative and sectioned (5 I~m). Sections were either stained with hematoxylin and eosin (A-C) or subjected to indirect immunofluorescence (D-l) with anti-hkl (D and E), anti-hkl4 (F and G), or anti-hk16 (H and I). Samples were from rafts of: SCC-12 (A), SCC-13 (Cand D-I), and SCC-15 (B). In D-l, the left portion of each panel shows staining of a well-differentiated portion of the raft; the right portion shows staining of the growing edge of the raft. BL, basal-like cells; SP, spinous-like cells; and GR granulocytes. Bar: (A-C, E, G, and I) 30 Ixm; (D, F, and H) 40 gm. Collectively, our studies indicate that certain morphological and biochemical features of the terminal differentiation process are maintained in squamous cell carcinoma cultures, but that the process is grossly abnormal when compared with primary epidermal cultures (Fig. 11). In all cases, both K1 and K16 were detected, and their expression seemed to be greatest in the squamous, more stratified regions of the rafts. These results are in good agreement with the findings ob- tained for SQCCs in vivo and suggest that the raft system closely approximates the morphological and biochemical aberrations of SQCCs. Whether this culture method will provide a parallel between the degree of differentiation of an SQCC in situ and the morphology of the corresponding tumor line in vitro awaits more careful analyses using primary SQCC cells from a number of different SQCCs. The Journal of Cell Biology. Volume 107,

17 Discussion Do the Monospecific Antisera Detect Their Keratins in All Epidermal Cells Where They Are Present? One of the major concerns with immunohistochemical studies is whether a negative result represents the absence or the masking of a protein. For the keratins, masking of antigenic sites has already been observed in some in vivo studies (Woodcock-Mitchell et al., 1982; Weiss et al., 1984). Physicochemical studies on keratins have suggested that the nonhelical end domains of the polypeptides may protrude along the surface of the keratin filament (Steinert et al., 1983; 1985; Roop et al., 1984; Albers and Fuchs, 1987), indicating that these domains might provide the best antigenic sites to guard against antibody masking. Since the carboxy terminus is also the most highly divergent region of the keratin polypeptide, this region seemed to be most suitable candidate for the production of monospecific anti-keratin antibodies. Our studies involving anti-hk14 have indicated that not only is this antiserum monospecific for K14, but in addition, it seems to detect K14 where it is present. This antiserum demonstrated clearly the abundant expression of K14 in the basal epidermal layer, with extension of expression to the suprabasal cells as well. While tissue-sectioning experiments (Fuchs and Green, 1980; Woodcock-Mitchell et al., 1982) and mrna analyses (Tyner and Fuchs, 1986) have suggested that expression of K14 might continue in the suprabasal layers, previous studies with other antibodies have not been able to verify this notion. In this report, we have shown that our anti-hk1 antiserum is also monospecific, and the K1 expression patterns obtained with this antiserum are consistent with previous mrna studies (Fuchs and Green, 1980), tissue-sectioning experiments (Fuchs and Green, 1980; Woodstock-Mitchell et al., 1982), and immunohistochemical studies (Viac et al., 1980; Woodcock-Mitchell et al., 1982). Although we cannot be unequivocally certain that the anti-hk1 antigenic site is exposed in all cells of epidermal diseases, the patterns of expression which we observed with the antiserum are consistent with our data on normal epidermis and indicate thatk1 is only expressed in terminally differentiating cells. The presence of anti-hk1 staining in all suprabasal layers of human skin, including the s. corneum, is somewhat different from that seen with an anti-mouse K1 peptide antiserum which showed only spinous and granular layer staining of mouse skin (Roop et al., 1984). This difference may be due to the fact that the anti-mouse K1 antiserum was prepared against the extreme carboxy-terminal tridecapeptide of the mouse K1, whereas our anti-human K1 antiserum was prepared against a 17-mer located just to the carboxyl side of the L L E G E alpha-helical sequence (see Fig. 1). Previously, it was observed that posttranslational processing of the 67- kd K1 to a 65-kD by-product (possibly K2) takes place in the outer layers of human skin (Fuchs and Green, 1980). Collectively, the data suggest that at least some of the residues following the L L E G E sequence in K1 are retained in the s. corneum layers, while the carboxy terminus may be lost. Before our study, little was known about the pattern of expression of K6/K16 in cultured epidermal cells or in epider- mal diseases of hyperproliferation. Early studies on keratin expression in cultured keratinocytes indicated that the cells were largely basal-like in character, and led to initial speculation that all four cultured cell keratins (K5, K6, K14, and K16) might be expressed in basal epidermal cells (Fuchs and Green, 1980). More recent studies have indicated that K6/ K16 are not present in normal epidermis and when they are expressed (e.g., during hyperproliferation or in culture), they may have a suprabasal location. In one such study, cell separation of cultured rabbit corneal epithelial cells suggested a suprabasal location (Schermer et al., 1986). Several other studies using the monoclonal antibody AE1 showed only basal staining of normal epidermis (Woodcock-Mitchell et al., 1982), but basal and spinous staining of psoriatic epidermis (Weiss et al., 1984). Since the repertoire of keratins cross-reacting with AE1 include K16 and since K16 is an induced component of the psoriatic cytoskeleton, it was suggested that K16 might be preferentially localized in the suprabasal layers of psoriatic skin (Weiss et al., 1984). However, because AE1 also cross reacts with K10 and K14 in the epidermis, and because the antigenic determinant recognized by AE1 is known to be masked for both K14 and K10 in the suprabasal cells of normal epidermis, but unmasked for K14 in the basal layer, the use of this antibody as a tool to identify the presence or absence of a particular keratin could not be made with certainty. Our finding that the hk16 antiserum cross reacts with the cytoskeleton of suprabasal cells of psoriatic epidermis is consistent with the early predictions by Weiss et al. (1984). While possibilities of masking cannot be excluded with antihk16, they are less likely considering that (a) the antiserum is polyclonal, and (b) the antiserum is against the carboxyterminal end of the polypeptide chain. The increased levels of K16 and K6 mrnas in the suprabasal layers of psoriatic epidermis support the immunohistochemical localization of these proteins. K1 Localization, Cell Morphology, and Terminal Differentiation The association of K1 and its partner K10 with the process of terminal differentiation has been well documented for normal epidermis (for review, see Fuchs et al., 1987). Their localization in SQCC has been more difficult to determine due to the irregular morphology of the tumors, and at present, the data have yielded somewhat conflicting results. While the microdissection studies of Moll et al. (1983) showed no correlation between the presence of K1/K10 and the appearance of highly keratinized pearls within welldifferentiated SQCCs, the immunohistochemical studies of Thomas et al. (1984) and Robinson (1987) indicated that the terminal differentiation-specific keratins are most abundant in SQCC-pearls. Our monospecific antiserum against human K1 (anti-hk1), has provided evidence that supports the previous immunohistochemical results. Moreover, the correlation between K1 expression and squamous-like morphology extends even to a moderately differentiated squamous cell carcinoma where all of the cells of the tumor appeared squamous in character and expressed K1. KI4 Expression and Basal-like Morphology While K1 expression is strongly associated with squamous- Stoler et al. Keratin Expression in Epidermal Diseases 443

18 like morphology and terminal differentiation, the solitary expression of the K14/K5 pair of keratins appears to be a major feature of all basal-like cells, Hence, both in vivo and in vitro, K14 seems to be the only type I protein expressed in the basal cells of both normal and hyperproliferating epidermis, and in the basal-like cells of basal and squamous cell carcinomas. A high level of K14 mrna also seems to correlate with a basal-like character. However, while K14 mrnas are abundant in basal cells of normal and psoriatic tissue, their expression varies considerably in tumor cells. Whether the low level of K14 expression in the basal-like cells of some tumors reflects a less differentiated state than true basal cells of adult epidermis remains to be determined; however, it is interesting to note that some basal cell carcinomas seem to be less differentiated in that islands of tumor cells can develop into hair and glandular-like structures, a capacity that is normally lost early during embryonic development (Fitzpatrick et al., 1979). K6/K16 Expression and Suprabasal Cells A major finding of our studies is that only suprabasal cells express K6 and K16 during hyperproliferation in the epidermis. This was true for psoriatic tissue, actinic keratosis, and regions of epidermis adjacent to tumors. Even in SQCCs, only the more squamous-like cells rather than the basal-like cells, expressed K6/K16. Moreover, the induction of K6/K16 expression in hyperproliferating epidermal cells seemed to coincide with a down-regulation in K1/K10 expression. Our investigations revealing the opposing behavior of K6/K16 and K1/K10 in the suprabasal cells of hyperproliferating epidermis provide direct experimental evidence in support of earlier hypotheses (Weiss et al., 1984; Schermer et al., 1986). The detection of K6/K16 in all and only the suprabasal cells of hyperproliferative diseases such as psoriasis revealed a marked biochemical distinction between the basal and suprabasal cells. This distinction was further illuminated by (a) the uniform decrease in K1 expression in all suprabasal cells, and (b) the presence of high levels of K14 mrnas in only the basal cells. Our data is also in agreement with that of Leigh et al. (1985), who examined other biochemical markers of basal and suprabasal psoriatic cells. It could be that the pattern of keratin gene expression in basal cells is controlled by the particular differentiation-state of an epidermal cell. Alternatively, the pattern of keratins might be determined by the close association of basal cells either with the basement membrane or with the dermis. In either case, the increased population of cells in psoriatic epidermis does not seem to express markers typical of basal epidermal cells, and yet they differ from normal spinous cells in their expression of K6/K16 and in their reduced expression of K1/K10. Whether these biochemical differences contribute to the increased proliferative rate of the suprabasal population in psoriasis (Lavker and Sun, 1983; Weinstein and Van Scott, 1965; Leigh et al., 1985) remains to be determined. If basal epidermal cells do not produce K6/K16, then a priori, we would not expect undifferentiated basal cell carcinomas to express these keratins. In fact, none of the four samples of solid or superficial basal cell carcinomas investigated in our study showed appreciable staining with our anti-hk16 antiserum, nor did they express significant levels of K6 or K16 mrnas. Thus, it seems most likely that the occasional report of either significant or trace amounts of K6 and K16 in basal cell carcinomas (Moll et al., 1982b, 1983, 1984; Breitkreutz et al., 1981; Kubilus et al., 1980; Weiss et al., 1984) reflects either (a) the detection of induced K16 and K6 in adjacent epidermis contaminating the tumor sample, or (b) a basal cell tumor capable of undergoing some, perhaps limited or altered, form of differentiation. More comprehensive studies using our antisera and crna probes to investigate additional basal cell carcinomas should help to clarify the extent to which basal cell carcinoma cells might be able to express K6/K16. K6/K16 Expression May Be Regulated at Both the Transcriptional and the Posttranscriptional Levels Previously, we had shown that K6 mrnas are present at a low level throughout normal epidermis even though no K6 protein is detected (Tyner and Fuchs, 1986). These RNAs seem to be translatable in a reticulocyte lysate system in vitro, and they are also of the proper size (Tyner and Fuchs, 1986). Thus, posttranscriptional controls seem to be operating to prevent the synthesis of K6/K16 in normal epidermis. However, when epidermis is placed into tissue culture medium, an increase of up to ninefold is seen in K6 mrna levels, indicating that additional (possibly transcriptional) controls might also be important in regulating the expression of these keratins (Tyner and Fuchs, 1986). Our initial studies led us to wonder whether the induction of hyperproliferation-associated keratins in psoriasis and in squamous cell carcinomas is accompanied by an increase in their mrnas, or alternatively, whether changes in the translatability of existing mrnas might dominate. Our results have clearly demonstrated that in both diseases, the levels of K16 and K6 mrnas are greatly elevated in the cells that are induced to express these keratins. Thus, both in normal skin placed in culture medium and in epidermal diseases of hyperproliferation, a substantial increase in K6/K16 mrna levels takes place concomitantly with the induction of K6/K16 expression. Since these changes take place only in the suprabasal epidermal cells, it is likely that only suprabasal cells have the capacity to sense the external stimulus that triggers the hyperproliferative response. We do not yet know whether the low levels of K16 and K6 mrnas observed in normal epidermis are functional in vivo. Posttranscriptional controls might enable the hyperproliferative response to occur more rapidly than if regulation is strictly at the transcriptional level. If expression of K6 and K16 is important, either to create a cytoskeleton more compatible with cell division, or to enable a greater number of divisions to occur before terminal differentiation, then recruiting premade mrnas to produce these keratins might facilitate the response. However, even if a base level of mrna expression is important to the initial events leading to hyperproliferation, the maintenance of the response seems to be fulfilled by a mechanism that leads to a large increase in the pool size of K6/K16 mrnas. Further studies using both in vivo and in vitro models of epidermal hyperproliferation should help to elucidate the molecular mechanisms underlying this complex and fundamentally important biological response. We would like to extend a special thank you to Ms. Grazina Traska for her expert assistance in tissue culture, Ms. Linda Foster for embedding and sec- The Journal of Cell Biology, Volume 107,

19 tioning our tissue samples, and Dr. Maria Medenica for her helpful discussions of the pathology of epidermal diseases. We x~auld also like to thank Mr. Chuck Weber for his expert printing of the photographs and Mr. Philip Galiga for his artful presentation of the data. Finally, a special thank you goes to Dr. Angela Tyner (Princeton University, Princeton, N J) for her help in in situ hybridizations given while she was still a member of the laboratory, and to Dr. James Rheinwald (Sydney Farber Cancer Institute, Boston, MA) for his gift of the squamous cell carcinoma cells and for his valuable advice regarding these tumor lines. This work was funded by grants from the National Institutes of Health and the Cystic Fibrosis Foundation (CF). E. Fuchs is a Presidential Young Investigator. A. Stoler is a CF postdoctoral trainee. R. Kopan is a predoctoral candidate funded by a grant from the Cancer Research Training Center at the University of Chicago. Received for publication 8 February 1988, and in revised form 9 May References Albers, K., and E. Fuchs The expression of mutant epidermal keratin cdnas transfected in simple epithelial and squamous cell carcinoma lines. J. Cell Biol. 105: Asselinean, D., B. Bernhard, C. Ballly, and M. Darmon Epidermal morphogenesis and induction of the 67kd keratin polypeptide by culture of human keratinocytes at the liquid-air interface. Exp. Cell Res. 159: Asselineau, D., B. A. Bernard, C. Bailly, M. Darmon, and M. Prunieras Human epidermis reconstructed by culture: is it "normal? ~ J. Invest. Dermatol. 86: Baden, H. P., N. McGilvray, C. K. Cbeng, L. D. Lee, and J. Kobilus The keratin polypeptides of psoriatic epidermis. J. Invest. Dermatol. 70: Bowden, P. E., E. J. Wood, and W. J. Cunliffe Comparison of prekeratin polypeptides in normal and psoriatic human epidermis. Biochim. Biophys. Acta. 743: Breitkreutz, D., W. Tilgen, P. Boakamp, and N. Fusenig Correlation of prekeratin peptides and ultrastracture in epithelial cells of human skin tumors in vivo and in vitro. Anticancer Res. 1: Cox, K. H., D. V. DeLeon, L. M. Angerer, and R. C. Angerer Detection of mrnas in sea urchin embryos by in situ hybridization using asymmetric RNA probes. Dev. Biol Fitzpatrick, T. B., A. Z. Eisen, K. Wolff, I. M. Freedberg, and K. F. Austen Dermatology in General Medicine. McGraw-Hill Inc., New York pp. Fuchs, E., and H. Green Changes in keratin gene expression during terminal differentiation of the keratinocyte. Cell. 19: Fuchs, E., and I. Hanukoglu Unraveling the structure of the intermediate filaments. Cell. 34: Fuchs, E., A. L. Tyner, G. J. Giudice, D. Marchuk, A. RayChaudhury, and M. Rosenberg The human genes and their differential expression. Dev. Biol. 22:5-34. Geisler, N., E. Kaufmann, and K. Weber Protein chemical characterization of three structurally distinct domains along the protofilament unit ofdesmin 10 nm filaments. Cell. 30: Hanukoglu, I., and E. Fuchs The cdna sequence of a type II cytoskeletal keratin reveals constant and variable structural domains among keratins. Cell. 33: Hunter, I., and D. Skerrow The proteins of living psoriatic epidermis. Biochim. Bioiphys. Acta. 714: Johnson, L., W. Idler, X-M. Zhou, D. Roop, and P. Steinert Structure of a gene for the human epidermal 67-kda keratin. Proc. Natl. Acad. Sci. USA. 82: Karasek, M. A., and M. E. Charlton Growth of postembryonic skin epithelial cells on collagen gels. J. Invest. Dermatol. 56: Knight, J., B. Gusterson, R. R. Jones, W. Landells, and P. Wilson Monoclonal antibodies of epidermal keratins: biochemical and immunocytochemical characterization: applications in pathology and cell culture. J. Pathol. 145: Kopan, R., G. Traska, and E. Fuchs Retinoids as important regulators of terminal differentiation: examining keratin expression in individual epidermal cells at various stages of keratinization. J. Cell Biol. 105: Kubilus, Z., H. P. Baden, and N. McGilvray Filamentous protein of basal cell epithelioma: characteristics in vivo and in vitro. J. Nat. Cancer Inst. 5: Lane, E. B., J. Bartek, P. E. Purkis, and L. M. Leigh Keratin antigens in differentiating skin. NY Acad. Sci. 455: Lavker, R. M., and T.-T. Sun Epidermal stem cells. J. Invest. Dermatol. 83:121s-127s. Leigh, I. M., K. A. Pulford, F. C. S. Ramaekers, and E. B. Lane Psoriasis: maintenance of an intact monolayer basal cell differentiation compartment in spite of hyperproliferation. Br. J. Dermatol. 113: Lersch, R., and E. Fuchs Sequence and expression of a type I1 keratin, K5, in human epidermal cells. Mol. Cell. Biol. 8: Lillie, J. H., D. K. MacCallum, and A. Jepsen Fine structure of subcultivated stratified squamous epithelium grown on collagen rafts. Exp. Cell Res. 125: McGuire, J., M. Osber, and L. Lightfoot Two keratins MW 50,000 and 56,000 are synthesized by psoriatic epidermis. Br. J. Dermatol. 111 (Suppl.) 27: Mansbridge, J. N., and A. M. Knapp Changes in keratinocyte maturation during wound healing. J. Invest. Dermatol. 89: Marchuk, D., S. McCrohon, and E. Fuchs Remarkable conservation among intermediate filament genes. Cell. 39: Marchuk, D., S. McCrohon, and E. Fuchs Complete sequence of a type I human keratin gene: presence of enhancer-like elements in the regulatory region of the gene. Proc. Natl. Acad. Sci. USA. 82: Matoltsy, A. G., M. N. Matoltsy, and P. J. Cliffel Characterization of keratin polypeptides of normal and psoriatic horny cells. J. Invest. Dermatol. 80: Moll, R., W. Franke, D. Schiller, B. Geiger, and R. Krepler. 1982a. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell. 31: Moll, R., W. W. Franke, B. Volc-Platzer, and R. Krepler. 1982b. Different keratin polypeptides in epidermis and other epithelia of human skin: a specific cytokeratin of molecular weight 46,000 in epithelia of the pilosebaceous tract and basal cell epitheliomas. J. Cell Biol. 95: Moll, R., R. Krepler, and W. W. Franke Complex cytokeratin polypeptide patterns observed in certain human carcinomas. Differentiation. 23: Moll, R., I. Moll, and W. W. Franke Differences of expression of cytokeratin polypeptides in various epithelial skin tumors. Dermatol. Res. 276: Nelson, W., and T.-T. Sun The 50- and 58-kdalton keratin classes as molecular markers for stratified squamous epithelia: cell culture studies. J. Cell Biol. 97: O'Farrell, P. Z., H. M. Goodman, and P. H. O'Farrell High resolution two-dimensional electrophoresis of basic as well as acidic proteins. Cell. 12: Ramaekers, F. C. S., J. J. G. Puts, O. Moesker, A. Kant, A. Huysmans, D. Haag, P. H. K. Jap, C. J. Herman, and G. P. Vooijs Antibodies to intermediate filament proteins in the immunohistochemical identification of human tumours: an overview. Histochem. 15: RayChaudhury, A., D. Marchuk, M. Lindhurst, and E. Fuchs Three tightly linked genes encoding human type I keratins: conservation of sequence in the 5' untranslated leader and 5' upstream regulatory regions. Mol. Cell. Biol. 6: Robinson, J. K Expression of keratin proteins in deeply invasive basal and squamous cell carcinoma: an immunohistochemical study. J. Dermatol. Surg. Oncol. 13: Roop, D R., C. K. Cheng, L. Titterington, C. A. Meyers, J. R. Stanley, P. M. Steinert, and S. H. Yuspa Synthetic peptides corresponding to keratin subunits elicit highly specific antibodies. J. BioL Chem. 259: Rosenberg, M., A. RayChaudhury, T. B. Shows, M. M. Le Beau, and E. Fuchs A group of type I keratin genes on human chromosome 17: characterization and expression. Mol. Cell. BioL 8: Schermer, A., S. Galvin, and T.-T. Sun Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 103: Skerrow, D., and C. J. Skerrow Tonofilament differentiation in human epidermis: isolation and polypeptide chain composition of keratinocyte subpopulation. Exp. Cell Res. 143: Steinert, P. M., R. H. Rice, D. R. Roop, B. L. Trus, and A. C. Steven Complete amino acid sequence of a mouse epidermal keratin subunit and implications for the structure of intermediate filaments. Nature (Lond.). 302: Steinert, P. M., A. C. Steven, and D. R. Roop The molecular biology of intermediate filaments. Cell. 42: Sun, T.-T., and H. Green Keratin filaments in cultured human epidermal cells. J. BioL Chem. 253: Thaler, M., K. Fukuyama, W. L. Epstein, and K. A. Fisher Comparative studies of keratins isolated from psoriasis and atopic dermatitis. J. Invest. Dermatol. 75: Thomas, P., J. W. Said, G. Nash, and S. Banks-Schlegel Profiles of keratin proteins in basal and squamous cell carcinomas of the skin. An immunohistocbemical study. Lab. Invest. 50: Towbin, H., T. Staehelin, and J. Gordon Electrophoretic transfer of proteins from polyacrylamide gels onto nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA. 81: Tseng, S. C. G., M. J. Jarvinen, W. G. Nelson, J.-W. Huang, J. Woodcock- Mitchell, and T.-T. Sun Correlation of specific keratins with different types of epithelial differentiation: monoclonal antibody studies. Cell. 30: Tyner, A. L., and E. Fuchs Evidence for posttranscriptional regulation of the keratins expressed during hyperproliferation and malignant transformation in human epidermis. J. Cell Biol. 103: Viac, J., M. J. Staquet, J. Thivolet, and C. Goujon Experimental Stoler et al. Keratin Expression in Epidermal Diseases 445
SHORT COMMUNICATION. Human Papillomavirus Type 11 E1 Ú E4 and L1 Proteins Colocalize in the Mouse Xenograft System at Multiple Time Points
 VIROLOGY 214, 259 263 (1995) SHORT COMMUNICATION Human Papillomavirus Type 11 E1 Ú E4 and L1 Proteins Colocalize in the Mouse Xenograft System at Multiple Time Points DARRON R. BROWN,*,,1 JANINE T. BRYAN,
VIROLOGY 214, 259 263 (1995) SHORT COMMUNICATION Human Papillomavirus Type 11 E1 Ú E4 and L1 Proteins Colocalize in the Mouse Xenograft System at Multiple Time Points DARRON R. BROWN,*,,1 JANINE T. BRYAN,
T HE epidermis consists of layers whose cells
 Retinoids as Important Regulators of Terminal Differentiation: Examining Keratin Expression in Individual Epidermal Cells at Various Stages of Keratinization Raphael Kopan, Grazina Traska, and Elaine Fuchs
Retinoids as Important Regulators of Terminal Differentiation: Examining Keratin Expression in Individual Epidermal Cells at Various Stages of Keratinization Raphael Kopan, Grazina Traska, and Elaine Fuchs
Histopathology: skin pathology
 Histopathology: skin pathology These presentations are to help you identify, and to test yourself on identifying, basic histopathological features. They do not contain the additional factual information
Histopathology: skin pathology These presentations are to help you identify, and to test yourself on identifying, basic histopathological features. They do not contain the additional factual information
Suprabasal change and subsequent formation of disulfide-stabilized homo- and hetero-dimers of keratins during esophageal epithelial differentiation
 Journal of Cell Science 104, 727-740 (1993) Printed in Great Britain The Company of Biologists Limited 1993 727 Suprabasal change and subsequent formation of disulfide-stabilized homo- and hetero-dimers
Journal of Cell Science 104, 727-740 (1993) Printed in Great Britain The Company of Biologists Limited 1993 727 Suprabasal change and subsequent formation of disulfide-stabilized homo- and hetero-dimers
3 CHAPTER 3: RESULTS
 3 CHAPTER 3: RESULTS 3.1 Histopathology 3.1.1 Normal Squamous Epithelium The squamous epithelium that covers the ectocervix of the uterus is composed of different layers starting at the basement membrane
3 CHAPTER 3: RESULTS 3.1 Histopathology 3.1.1 Normal Squamous Epithelium The squamous epithelium that covers the ectocervix of the uterus is composed of different layers starting at the basement membrane
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Meibomian Gland Dysfunction
 Investigative Ophthalmology & Visual Science. Vol. 30. No. 5. May 1989 Copyright Association for Research in Vision and Ophthalmology Meibomian Gland Dysfunction //. The Role of Kerarinizarion in o Rabbit
Investigative Ophthalmology & Visual Science. Vol. 30. No. 5. May 1989 Copyright Association for Research in Vision and Ophthalmology Meibomian Gland Dysfunction //. The Role of Kerarinizarion in o Rabbit
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Amelio et al., http://www.jcb.org/cgi/content/full/jcb.201203134/dc1 Figure S1. mir-24 regulates proliferation and by itself induces
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Amelio et al., http://www.jcb.org/cgi/content/full/jcb.201203134/dc1 Figure S1. mir-24 regulates proliferation and by itself induces
T HE epidermis, the epithelial component of skin, undergoes
 Abnormal Expression and Processing of Keratins in Pupoid Fetus (pf/pf) and Repeated Epilation (Er/Er) Mutant Mice Chris Fisher,* Alma Jones,* and Dennis R. Roopw Departments of *Biological Structure and
Abnormal Expression and Processing of Keratins in Pupoid Fetus (pf/pf) and Repeated Epilation (Er/Er) Mutant Mice Chris Fisher,* Alma Jones,* and Dennis R. Roopw Departments of *Biological Structure and
'Abbreviations used in this paper: BCC, basal cell carcinoma; IEF, isoelectric focusing ; kd, kilodaltons.
 Monoclonal Antibody Analysis of Keratin Expression in Epidermal Diseases : A 48- and 56-kdalton Keratin as Molecular Markers for Hyperproliferative Keratinocytes ROBERT A WEISS,* RIVA EICHNER,*5 and TUNG-TIEN
Monoclonal Antibody Analysis of Keratin Expression in Epidermal Diseases : A 48- and 56-kdalton Keratin as Molecular Markers for Hyperproliferative Keratinocytes ROBERT A WEISS,* RIVA EICHNER,*5 and TUNG-TIEN
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
ELECTRON MICROSCOPY OF A SMALL PIGMENTED CUTANEOUS LESION*
 ELECTRON MICROSCOPY OF A SMALL PIGMENTED CUTANEOUS LESION* The description of the lesion in the title of this rcport is intentionally non-committal. Diagnosed clinically as a lentigo, it was removed as
ELECTRON MICROSCOPY OF A SMALL PIGMENTED CUTANEOUS LESION* The description of the lesion in the title of this rcport is intentionally non-committal. Diagnosed clinically as a lentigo, it was removed as
t Defined as the period of time it takes for the AND PROTEIN SYNTHESIS IN PIG EPIDERMIS* 9) and by the location of mitoses (11), it
 and by the location of mitoses (11), it") THE JOURNAL OF INVETIGATIVE DERMATOLOGY Copyright 1965 by The Williams & Wilkins Co. Vol., No. 6 Printed in U..A. AUTORADIOGRAPHIC TUDIE OF TURNOVER TIME AND PROTEIN YNTHEI IN PIG EPIDERMI* The epidermis
THE JOURNAL OF INVETIGATIVE DERMATOLOGY Copyright 1965 by The Williams & Wilkins Co. Vol., No. 6 Printed in U..A. AUTORADIOGRAPHIC TUDIE OF TURNOVER TIME AND PROTEIN YNTHEI IN PIG EPIDERMI* The epidermis
Basal cell carcinoma 5/28/2011
 Goal of this Presentation A practical approach to the diagnosis of cutaneous carcinomas and their mimics Thaddeus Mully, MD University of California San Francisco To review common non-melanoma skin cancers
Goal of this Presentation A practical approach to the diagnosis of cutaneous carcinomas and their mimics Thaddeus Mully, MD University of California San Francisco To review common non-melanoma skin cancers
Protein MultiColor Stable, Low Range
 Product Name: DynaMarker Protein MultiColor Stable, Low Range Code No: DM670L Lot No: ******* Size: 200 μl x 3 (DM670 x 3) (120 mini-gel lanes) Storage: 4 C Stability: 12 months at 4 C Storage Buffer:
Product Name: DynaMarker Protein MultiColor Stable, Low Range Code No: DM670L Lot No: ******* Size: 200 μl x 3 (DM670 x 3) (120 mini-gel lanes) Storage: 4 C Stability: 12 months at 4 C Storage Buffer:
5 Identification of Binding Partners of the Annexin A2 / P11 Complex by Chemical Cross-Linking
 5 Identification of Binding Partners of the Annexin A2 / P11 Complex by Chemical Cross-Linking In the quest of the omics sciences for holistic schemes, the identification of binding partners of proteins
5 Identification of Binding Partners of the Annexin A2 / P11 Complex by Chemical Cross-Linking In the quest of the omics sciences for holistic schemes, the identification of binding partners of proteins
Expression of mrnas from Two Multigene Families
 MOLECULAR AND CELLULAR BIOLOGY, Apr. 1983, p. 495-502 0270-7306/83/040495-08$02.00/0 Copyright 0 1983, American Society for Microbiology Vol. 3, No. 4 Tissue Specificity of Epithelial Keratins: Differential
MOLECULAR AND CELLULAR BIOLOGY, Apr. 1983, p. 495-502 0270-7306/83/040495-08$02.00/0 Copyright 0 1983, American Society for Microbiology Vol. 3, No. 4 Tissue Specificity of Epithelial Keratins: Differential
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
CMB621: Cytoskeleton. Also known as How the cell plays with LEGOs to ensure order, not chaos, is temporally and spatially achieved
 CMB621: Cytoskeleton Also known as How the cell plays with LEGOs to ensure order, not chaos, is temporally and spatially achieved Lecture(s) Overview Lecture 1: What is the cytoskeleton? Membrane interaction
CMB621: Cytoskeleton Also known as How the cell plays with LEGOs to ensure order, not chaos, is temporally and spatially achieved Lecture(s) Overview Lecture 1: What is the cytoskeleton? Membrane interaction
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Type IV collagen and laminin staining patterns in benign
 J Clin Pathol 1989;42:1173-1177 Type IV collagen and laminin staining patterns in benign and malignant cutaneous lesions RONA M MacKIE, D B CLELLAND, CHRISTINE J SKERROW From the Department ofdermatology,
J Clin Pathol 1989;42:1173-1177 Type IV collagen and laminin staining patterns in benign and malignant cutaneous lesions RONA M MacKIE, D B CLELLAND, CHRISTINE J SKERROW From the Department ofdermatology,
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Histopathology: Cervical HPV and neoplasia
 Histopathology: Cervical HPV and neoplasia These presentations are to help you identify basic histopathological features. They do not contain the additional factual information that you need to learn about
Histopathology: Cervical HPV and neoplasia These presentations are to help you identify basic histopathological features. They do not contain the additional factual information that you need to learn about
(From Arnold Biological Laboratory, Brown University, ProVence) P~s 3 To 5. Materials and Methods
 P~s 3 To 5. Materials and Methods") Published Online: 25 January, 1955 Supp Info: http://doi.org/10.1083/jcb.1.1.13 Downloaded from jcb.rupress.org on June 21, 2018 HISTOLOGY AND CYTOCHEMISTRY OF HUMAN SKIN IX. THE Dm~mUT~ON Ot NoN-SP~.c~Ic
Published Online: 25 January, 1955 Supp Info: http://doi.org/10.1083/jcb.1.1.13 Downloaded from jcb.rupress.org on June 21, 2018 HISTOLOGY AND CYTOCHEMISTRY OF HUMAN SKIN IX. THE Dm~mUT~ON Ot NoN-SP~.c~Ic
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Benign and malignant epithelial lesions: Seborrheic keratosis: A common benign pigmented epidermal tumor occur in middle-aged or older persons more
 Benign and malignant epithelial lesions: Seborrheic keratosis: A common benign pigmented epidermal tumor occur in middle-aged or older persons more common on the trunk; but extremities, head and neck are
Benign and malignant epithelial lesions: Seborrheic keratosis: A common benign pigmented epidermal tumor occur in middle-aged or older persons more common on the trunk; but extremities, head and neck are
BIOCHEMISTRY OF SKIN AND CONNECTIVE TISSUES
 BIOCHEMISTRY OF SKIN AND CONNECTIVE TISSUES Sri Widia A Jusman Department of Biochemistry & Molecular Biology FMUI 1 2 SKIN Epidermis - horny layer (keratin-filled dead cells) - granular layer - spinous
BIOCHEMISTRY OF SKIN AND CONNECTIVE TISSUES Sri Widia A Jusman Department of Biochemistry & Molecular Biology FMUI 1 2 SKIN Epidermis - horny layer (keratin-filled dead cells) - granular layer - spinous
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:!
 LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:! Spinal cord and peripheral nerves! Eyes, Inner ear, nasal
LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:! Spinal cord and peripheral nerves! Eyes, Inner ear, nasal
Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells.
 SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells. A) Cirbp mrna expression levels in various mouse tissues collected around the clock
SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure S1. Expression of Cirbp mrna in mouse tissues and NIH3T3 cells. A) Cirbp mrna expression levels in various mouse tissues collected around the clock
Figure mouse globin mrna PRECURSOR RNA hybridized to cloned gene (genomic). mouse globin MATURE mrna hybridized to cloned gene (genomic).
. mouse globin MATURE mrna hybridized to cloned gene (genomic).") Splicing Figure 14.3 mouse globin mrna PRECURSOR RNA hybridized to cloned gene (genomic). mouse globin MATURE mrna hybridized to cloned gene (genomic). mrna Splicing rrna and trna are also sometimes spliced;
Splicing Figure 14.3 mouse globin mrna PRECURSOR RNA hybridized to cloned gene (genomic). mouse globin MATURE mrna hybridized to cloned gene (genomic). mrna Splicing rrna and trna are also sometimes spliced;
Loss of RhoA promotes skin tumor formation. Supplementary Figure 1. Loss of RhoA does not impair F-actin organization.
 Supplementary Figure Legends Supplementary Figure 1. Loss of RhoA does not impair F-actin organization. a. Representative IF images of F-actin staining of big and small control (left) and RhoA ko tumors
Supplementary Figure Legends Supplementary Figure 1. Loss of RhoA does not impair F-actin organization. a. Representative IF images of F-actin staining of big and small control (left) and RhoA ko tumors
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: van Seters M, van Beurden M, ten Kate FJW, et al. Treatment
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: van Seters M, van Beurden M, ten Kate FJW, et al. Treatment
Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene
 Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene YUELIN ZHANG, WEIHUA FAN, MARK KINKEMA, XIN LI, AND
Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene YUELIN ZHANG, WEIHUA FAN, MARK KINKEMA, XIN LI, AND
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Proteins. Amino acids, structure and function. The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka
 Proteins Amino acids, structure and function The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka O O HO N N HN OH Ser65-Tyr66-Gly67 The Nobel prize in chemistry 2008 Osamu Shimomura,
Proteins Amino acids, structure and function The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz Brian K. Kobilka O O HO N N HN OH Ser65-Tyr66-Gly67 The Nobel prize in chemistry 2008 Osamu Shimomura,
Histology: Epithelial tissue
 Histology: Epithelial tissue Epithelial Tissue is presented in two forms: 1. Covering Epithelia: 2. Glandular Epithelia: 1. Simple Epithelium: contain only one layer of cells. 2. Stratified Epithelium:
Histology: Epithelial tissue Epithelial Tissue is presented in two forms: 1. Covering Epithelia: 2. Glandular Epithelia: 1. Simple Epithelium: contain only one layer of cells. 2. Stratified Epithelium:
Integumentary System. Integumentary System
 1. General aspects a. The integumentary system consists of several organs major organ of the system is the skin other organs are relatively small and they can be considered as specialized structures of
1. General aspects a. The integumentary system consists of several organs major organ of the system is the skin other organs are relatively small and they can be considered as specialized structures of
BCH Graduate Survey of Biochemistry
 BCH 5045 Graduate Survey of Biochemistry Instructor: Charles Guy Producer: Ron Thomas Director: Glen Graham Lecture 7 Slide sets available at: http://hort.ifas.ufl.edu/teach/guyweb/bch5045/index.html David
BCH 5045 Graduate Survey of Biochemistry Instructor: Charles Guy Producer: Ron Thomas Director: Glen Graham Lecture 7 Slide sets available at: http://hort.ifas.ufl.edu/teach/guyweb/bch5045/index.html David
Problem-solving Test: The Mechanism of Protein Synthesis
 Q 2009 by The International Union of Biochemistry and Molecular Biology BIOCHEMISTRY AND MOLECULAR BIOLOGY EDUCATION Vol. 37, No. 1, pp. 58 62, 2009 Problem-based Learning Problem-solving Test: The Mechanism
Q 2009 by The International Union of Biochemistry and Molecular Biology BIOCHEMISTRY AND MOLECULAR BIOLOGY EDUCATION Vol. 37, No. 1, pp. 58 62, 2009 Problem-based Learning Problem-solving Test: The Mechanism
Anti-PD-L1 antibody [28-8] ab205921
![Anti-PD-L1 antibody [28-8] ab205921](/thumbs/84/91196262.jpg "Anti-PD-L1 antibody [28-8] ab205921") Anti-PD-L1 antibody [28-8] ab205921 2 Abreviews 16 References 15 Images Overview Product name Anti-PD-L1 antibody [28-8] Description Tested applications Species reactivity Immunogen Rabbit monoclonal [28-8]
Anti-PD-L1 antibody [28-8] ab205921 2 Abreviews 16 References 15 Images Overview Product name Anti-PD-L1 antibody [28-8] Description Tested applications Species reactivity Immunogen Rabbit monoclonal [28-8]
Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR
 Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR Pages with reference to book, From 305 To 307 Irshad N. Soomro,Samina Noorali,Syed Abdul Aziz,Suhail Muzaffar,Shahid
Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR Pages with reference to book, From 305 To 307 Irshad N. Soomro,Samina Noorali,Syed Abdul Aziz,Suhail Muzaffar,Shahid
Spherical Nucleic Acids For Advanced Wound Healing Applications Chad A. Mirkin
 Spherical Nucleic Acids For Advanced Wound Healing Applications Chad A. Mirkin Departments of Chemistry, Infectious Disease, Materials Science & Engineering, Chemical & Biological Engineering, and Biomedical
Spherical Nucleic Acids For Advanced Wound Healing Applications Chad A. Mirkin Departments of Chemistry, Infectious Disease, Materials Science & Engineering, Chemical & Biological Engineering, and Biomedical
Supporting Information
 Supporting Information Plikus et al. 10.1073/pnas.1215935110 SI Text Movies S1, S2, S3, and S4 are time-lapse recordings from individually cultured Period2 Luc vibrissa follicles show that circadian cycles
Supporting Information Plikus et al. 10.1073/pnas.1215935110 SI Text Movies S1, S2, S3, and S4 are time-lapse recordings from individually cultured Period2 Luc vibrissa follicles show that circadian cycles
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Desmoplastic Melanoma R/O BCC. Clinical Information. 74 y.o. man with lesion on left side of neck r/o BCC
 R/O BCC Sabine Kohler, M.D. Professor of Pathology and Dermatology Dermatopathology Service Stanford University School of Medicine Clinical Information 74 y.o. man with lesion on left side of neck r/o
R/O BCC Sabine Kohler, M.D. Professor of Pathology and Dermatology Dermatopathology Service Stanford University School of Medicine Clinical Information 74 y.o. man with lesion on left side of neck r/o
Anti-Lamin B1/LMNB1 Picoband Antibody
 Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
Expression and clinical significance of ADAM17 protein in esophageal squamous cell carcinoma
 Expression and clinical significance of ADAM17 protein in esophageal squamous cell carcinoma H.B. Liu, Y. Zhu, Q.C. Yang, Y. Shen, X.J. Zhang and H. Chen Department of Pathology First People s Hospital
Expression and clinical significance of ADAM17 protein in esophageal squamous cell carcinoma H.B. Liu, Y. Zhu, Q.C. Yang, Y. Shen, X.J. Zhang and H. Chen Department of Pathology First People s Hospital
DNA codes for RNA, which guides protein synthesis.
 Section 3: DNA codes for RNA, which guides protein synthesis. K What I Know W What I Want to Find Out L What I Learned Vocabulary Review synthesis New RNA messenger RNA ribosomal RNA transfer RNA transcription
Section 3: DNA codes for RNA, which guides protein synthesis. K What I Know W What I Want to Find Out L What I Learned Vocabulary Review synthesis New RNA messenger RNA ribosomal RNA transfer RNA transcription
Pepsin Solution ready-to-use
 SIE HABEN DIE VISION, WIR HABEN DIE SUBSTANZ. Pepsin Solution Single component Pepsin Solution: only one component refrigerator stable Pepsin is a commonly used digestive enzyme for immunohistochemical
SIE HABEN DIE VISION, WIR HABEN DIE SUBSTANZ. Pepsin Solution Single component Pepsin Solution: only one component refrigerator stable Pepsin is a commonly used digestive enzyme for immunohistochemical
supplementary information
 DOI: 10.1038/ncb2133 Figure S1 Actomyosin organisation in human squamous cell carcinoma. (a) Three examples of actomyosin organisation around the edges of squamous cell carcinoma biopsies are shown. Myosin
DOI: 10.1038/ncb2133 Figure S1 Actomyosin organisation in human squamous cell carcinoma. (a) Three examples of actomyosin organisation around the edges of squamous cell carcinoma biopsies are shown. Myosin
Page 1 of 5 Biochemistry I Fall 2017 Practice for Exam2 Dr. Stone Name
 Page 1 of 5 Biochemistry I Fall 2017 Practice for Exam2 Dr. Stone ame o answers will be provided. ere are some constants and equations that may be useful: K a = [+][A-]/[A] p = pka + log [A-]/[A] K a for
Page 1 of 5 Biochemistry I Fall 2017 Practice for Exam2 Dr. Stone ame o answers will be provided. ere are some constants and equations that may be useful: K a = [+][A-]/[A] p = pka + log [A-]/[A] K a for
Molecular Biology (BIOL 4320) Exam #2 May 3, 2004
 Exam #2 May 3, 2004") Molecular Biology (BIOL 4320) Exam #2 May 3, 2004 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number. Good
Molecular Biology (BIOL 4320) Exam #2 May 3, 2004 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number. Good
****************************************************************************************************** INTEGUMENTARY SYSTEM
 BIOLOGY 211: HUMAN ANATOMY & PHYSIOLOGY ****************************************************************************************************** INTEGUMENTARY SYSTEM ******************************************************************************************************
BIOLOGY 211: HUMAN ANATOMY & PHYSIOLOGY ****************************************************************************************************** INTEGUMENTARY SYSTEM ******************************************************************************************************
Ch. 4: Skin and Body Membranes
 Ch. 4: Skin and Body Membranes I. Body Membranes A. Function of body membranes 1. Cover body surfaces 2. Line body cavities 3. Form protective sheets around organs II. Classification of Body Membranes
Ch. 4: Skin and Body Membranes I. Body Membranes A. Function of body membranes 1. Cover body surfaces 2. Line body cavities 3. Form protective sheets around organs II. Classification of Body Membranes
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide gel electrophoresis/genetics)
") Proc. Natl. Acad. Sci. USA Vol. 73, No. 6, pp. 242-246, June 976 Microbiology Mapping of the influenza virus genome: Identification of the hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide
Proc. Natl. Acad. Sci. USA Vol. 73, No. 6, pp. 242-246, June 976 Microbiology Mapping of the influenza virus genome: Identification of the hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Review of Biochemistry
 Review of Biochemistry Chemical bond Functional Groups Amino Acid Protein Structure and Function Proteins are polymers of amino acids. Each amino acids in a protein contains a amino group, - NH 2,
Review of Biochemistry Chemical bond Functional Groups Amino Acid Protein Structure and Function Proteins are polymers of amino acids. Each amino acids in a protein contains a amino group, - NH 2,
Increased DNA synthesis of uninvolved psoriatic epidermis is maintained in vitro
 British Journal of Dermatology (1985) 112, 263-270. Increased DNA synthesis of uninvolved psoriatic epidermis is maintained in vitro K.KRAGBALLE, L.DESJARLAIS AND C.L.MARCELO University of Michigan Medical
British Journal of Dermatology (1985) 112, 263-270. Increased DNA synthesis of uninvolved psoriatic epidermis is maintained in vitro K.KRAGBALLE, L.DESJARLAIS AND C.L.MARCELO University of Michigan Medical
Classification of Epidermal Keratins according to Their Immunoreactivity, Isoelectric Point, and Mode of Expression
 Classification of Epidermal Keratins according to Their Immunoreactivity, Isoelectric Point, and Mode of Expression RIVA EICHNER,*$ PAULA BONITZ,* and TUNG-TIEN SUN* #41l Departments of *Cell Biology and
Classification of Epidermal Keratins according to Their Immunoreactivity, Isoelectric Point, and Mode of Expression RIVA EICHNER,*$ PAULA BONITZ,* and TUNG-TIEN SUN* #41l Departments of *Cell Biology and
Skin and Body Membranes Body Membranes Function of body membranes Cover body surfaces Line body cavities Form protective sheets around organs
 Skin and Body Membranes Body Membranes Function of body membranes Cover body surfaces Line body cavities Form protective sheets around organs Classification of Body Membranes Epithelial membranes Cutaneous
Skin and Body Membranes Body Membranes Function of body membranes Cover body surfaces Line body cavities Form protective sheets around organs Classification of Body Membranes Epithelial membranes Cutaneous
ab SREBP-2 Translocation Assay Kit (Cell-Based)
") ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
Papillary Lesions of the Breast A Practical Approach to Diagnosis. (Arch Pathol Lab Med. 2016;140: ; doi: /arpa.
 Papillary Lesions of the Breast A Practical Approach to Diagnosis (Arch Pathol Lab Med. 2016;140:1052 1059; doi: 10.5858/arpa.2016-0219-RA) Papillary lesions of the breast Span the spectrum of benign,
Papillary Lesions of the Breast A Practical Approach to Diagnosis (Arch Pathol Lab Med. 2016;140:1052 1059; doi: 10.5858/arpa.2016-0219-RA) Papillary lesions of the breast Span the spectrum of benign,
Supplementary Materials and Methods
 Supplementary Materials and Methods Reagents and antibodies was purchased from iaffin GmbH & Co KG. Cisplatin (ristol-myers Squibb Co.) and etoposide (Sandoz Pharma Ltd.) were used. Antibodies recognizing
Supplementary Materials and Methods Reagents and antibodies was purchased from iaffin GmbH & Co KG. Cisplatin (ristol-myers Squibb Co.) and etoposide (Sandoz Pharma Ltd.) were used. Antibodies recognizing
Supplemental Figure 1
 Supplemental Figure 1 A S100A4: SFLGKRTDEAAFQKLMSNLDSNRDNEVDFQEYCVFLSCIAMMCNEFFEGFPDK Overlap: SF G DE KLM LD N D VDFQEY VFL I M N FF G PD S100A2: SFVGEKVDEEGLKKLMGSLDENSDQQVDFQEYAVFLALITVMCNDFFQGCPDR
Supplemental Figure 1 A S100A4: SFLGKRTDEAAFQKLMSNLDSNRDNEVDFQEYCVFLSCIAMMCNEFFEGFPDK Overlap: SF G DE KLM LD N D VDFQEY VFL I M N FF G PD S100A2: SFVGEKVDEEGLKKLMGSLDENSDQQVDFQEYAVFLALITVMCNDFFQGCPDR
Life Sciences 1A Midterm Exam 2. November 13, 2006
 Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Phosphoserine Detection Kit
 Kit 0701/PSER-KIT 02/080507 Background and Specificity extracellular signals to the nucleus. Phosphorylated epitopes may serve as docking sites for the assembley of protein complexes or may alter the 3-dimensional
Kit 0701/PSER-KIT 02/080507 Background and Specificity extracellular signals to the nucleus. Phosphorylated epitopes may serve as docking sites for the assembley of protein complexes or may alter the 3-dimensional
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer. Application Note
 Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
Proteins. AP Biology. Proteins. Proteins. Proteins. Effect of different R groups: Nonpolar amino acids. Amino acids H C OH H R. Structure.
 2008-2009 Most structurally & functionally diverse group : involved in almost everything (pepsin, DNA polymerase) (keratin, collagen) (hemoglobin, aquaporin) (insulin & other hormones) (antibodies) (actin
2008-2009 Most structurally & functionally diverse group : involved in almost everything (pepsin, DNA polymerase) (keratin, collagen) (hemoglobin, aquaporin) (insulin & other hormones) (antibodies) (actin
EXPERIMENTAL THERMAL BURNS I. A study of the immediate and delayed histopathological changes of the skin.
 EXPERIMENTAL THERMAL BURNS I A study of the immediate and delayed histopathological changes of the skin. RJ Brennan, M.D. and B. Rovatti M.D. The purpose of this study was to determine the progressive
EXPERIMENTAL THERMAL BURNS I A study of the immediate and delayed histopathological changes of the skin. RJ Brennan, M.D. and B. Rovatti M.D. The purpose of this study was to determine the progressive
C ORNEAL epithelium is a four- to five-layered, stratified
 Published Online: 1 July, 1986 Supp Info: http://doi.org/10.1083/jcb.103.1.49 Downloaded from jcb.rupress.org on December 26, 2018 Differentiation-related Expression of a Major 64K Corneal Keratin In Vivo
Published Online: 1 July, 1986 Supp Info: http://doi.org/10.1083/jcb.103.1.49 Downloaded from jcb.rupress.org on December 26, 2018 Differentiation-related Expression of a Major 64K Corneal Keratin In Vivo
Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC-
 Supplemental material and methods Reagents. Hydralazine was purchased from Sigma-Aldrich. Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC- 133, human thyroid medullary
Supplemental material and methods Reagents. Hydralazine was purchased from Sigma-Aldrich. Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC- 133, human thyroid medullary
Histopathology of Melanoma
 THE YALE JOURNAL OF BIOLOGY AND MEDICINE 48, 409-416 (1975) Histopathology of Melanoma G. J. WALKER SMITH Department ofpathology, Yale University School ofmedicine, 333 Cedar Street, New Haven, Connecticut
THE YALE JOURNAL OF BIOLOGY AND MEDICINE 48, 409-416 (1975) Histopathology of Melanoma G. J. WALKER SMITH Department ofpathology, Yale University School ofmedicine, 333 Cedar Street, New Haven, Connecticut
Overview of the Expressway Cell-Free Expression Systems. Expressway Mini Cell-Free Expression System
 Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
HiPer Western Blotting Teaching Kit
 HiPer Western Blotting Teaching Kit Product Code: HTI009 Number of experiments that can be performed: 5/20 Duration of Experiment: ~ 2 days Day 1: 6-8 hours (SDS- PAGE and Electroblotting) Day 2: 3 hours
HiPer Western Blotting Teaching Kit Product Code: HTI009 Number of experiments that can be performed: 5/20 Duration of Experiment: ~ 2 days Day 1: 6-8 hours (SDS- PAGE and Electroblotting) Day 2: 3 hours
PowerPoint Lecture Slide Presentation by Patty Bostwick-Taylor, Florence-Darlington Technical College Skin and Body Membranes
 PowerPoint Lecture Slide Presentation by Patty Bostwick-Taylor, Florence-Darlington Technical College Skin and Body Membranes 4 Body Membranes Function of body membranes Cover body surfaces Line body cavities
PowerPoint Lecture Slide Presentation by Patty Bostwick-Taylor, Florence-Darlington Technical College Skin and Body Membranes 4 Body Membranes Function of body membranes Cover body surfaces Line body cavities
Received 26 January 1996/Returned for modification 28 February 1996/Accepted 15 March 1996
 MOLECULAR AND CELLULAR BIOLOGY, June 1996, p. 3012 3022 Vol. 16, No. 6 0270-7306/96/$04.00 0 Copyright 1996, American Society for Microbiology Base Pairing at the 5 Splice Site with U1 Small Nuclear RNA
MOLECULAR AND CELLULAR BIOLOGY, June 1996, p. 3012 3022 Vol. 16, No. 6 0270-7306/96/$04.00 0 Copyright 1996, American Society for Microbiology Base Pairing at the 5 Splice Site with U1 Small Nuclear RNA
Each Other. EDTA), quickly cooled in an ice slurry, and made 3 M KCl. before being bound to the column. Sindbis virus RNAs (49S
, quickly cooled in an ice slurry, and made 3 M KCl. before being bound to the column. Sindbis virus RNAs (49S") JOURNAL OF VIROLOGY, Mar. 1986, p. 917-921 0022-538X/86/030917-05$02.00/0 Vol. 57, No. 3 RNA Virus Genomes Hybridize to Cellular rrnas and to Each Other MARCELLA A. McCLURElt* AND JACQUES PERRAULT'2: Department
JOURNAL OF VIROLOGY, Mar. 1986, p. 917-921 0022-538X/86/030917-05$02.00/0 Vol. 57, No. 3 RNA Virus Genomes Hybridize to Cellular rrnas and to Each Other MARCELLA A. McCLURElt* AND JACQUES PERRAULT'2: Department
Skin and Body Membranes
 4 Skin and Body Membranes PowerPoint Lecture Slide Presentation by Jerry L. Cook, Sam Houston University ESSENTIALS OF HUMAN ANATOMY & PHYSIOLOGY EIGHTH EDITION ELAINE N. MARIEB Skin and Body Membranes
4 Skin and Body Membranes PowerPoint Lecture Slide Presentation by Jerry L. Cook, Sam Houston University ESSENTIALS OF HUMAN ANATOMY & PHYSIOLOGY EIGHTH EDITION ELAINE N. MARIEB Skin and Body Membranes
microrna Presented for: Presented by: Date:
 microrna Presented for: Presented by: Date: 2 micrornas Non protein coding, endogenous RNAs of 21-22nt length Evolutionarily conserved Regulate gene expression by binding complementary regions at 3 regions
microrna Presented for: Presented by: Date: 2 micrornas Non protein coding, endogenous RNAs of 21-22nt length Evolutionarily conserved Regulate gene expression by binding complementary regions at 3 regions
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Yatsenko AN, Georgiadis AP, Röpke A, et al. X-linked TEX11
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Yatsenko AN, Georgiadis AP, Röpke A, et al. X-linked TEX11
Unit 4 - The Skin and Body Membranes 1
 Unit 4 - The Skin and Body Membranes 1 I. Unit 4: Skin and Body Membranes A. Body Membranes 1. Function of body membranes a) Cover body surfaces b) Line body cavities c) Form protective sheets around organs
Unit 4 - The Skin and Body Membranes 1 I. Unit 4: Skin and Body Membranes A. Body Membranes 1. Function of body membranes a) Cover body surfaces b) Line body cavities c) Form protective sheets around organs
The Integumentary System. Mosby items and derived items 2010, 2006, 2002, 1997, 1992 by Mosby, Inc., an affiliate of Elsevier Inc.
 The Integumentary System The Skin Structure two primary layers called epidermis and dermis Epidermis Outermost and thinnest primary layer of skin Composed of several layers of stratified squamous epithelium
The Integumentary System The Skin Structure two primary layers called epidermis and dermis Epidermis Outermost and thinnest primary layer of skin Composed of several layers of stratified squamous epithelium
Introduction retroposon
 17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
For example, monosaccharides such as glucose are polar and soluble in water, whereas lipids are nonpolar and insoluble in water.
 Biology 4A Laboratory Biologically Important Molecules Objectives Perform tests to detect the presence of carbohydrates, lipids, proteins, and nucleic acids Recognize the importance of a control in a biochemical
Biology 4A Laboratory Biologically Important Molecules Objectives Perform tests to detect the presence of carbohydrates, lipids, proteins, and nucleic acids Recognize the importance of a control in a biochemical
Protein Synthesis
 Protein Synthesis 10.6-10.16 Objectives - To explain the central dogma - To understand the steps of transcription and translation in order to explain how our genes create proteins necessary for survival.
Protein Synthesis 10.6-10.16 Objectives - To explain the central dogma - To understand the steps of transcription and translation in order to explain how our genes create proteins necessary for survival.
INTEGUMENTARY 1-Epidermis, 2-Dermis, Structure of thick and thin skin I- Epidermis . Stratum basale
 INTEGUMENTARY The skin (integument, cutis ) and its derivatives constitute the integumentary system. It form the external covering of the body and is the largest organ of the body. The skin consists of
INTEGUMENTARY The skin (integument, cutis ) and its derivatives constitute the integumentary system. It form the external covering of the body and is the largest organ of the body. The skin consists of
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Introduction. Skin and Body Membranes. Cutaneous Membranes Skin 9/14/2017. Classification of Body Membranes. Classification of Body Membranes
 Introduction Skin and Body Membranes Body membranes Cover surfaces Line body cavities Form protective and lubricating sheets around organs Classified in 5 categories Epithelial membranes 3 types- cutaneous,
Introduction Skin and Body Membranes Body membranes Cover surfaces Line body cavities Form protective and lubricating sheets around organs Classified in 5 categories Epithelial membranes 3 types- cutaneous,
Integumentary System (Script) Slide 1: Integumentary System. Slide 2: An overview of the integumentary system
 Slide 1: Integumentary System. Slide 2: An overview of the integumentary system") Integumentary System (Script) Slide 1: Integumentary System Slide 2: An overview of the integumentary system Skin is the body s largest and heaviest organ making up 15% of body weight. Most skin is 1 to
Integumentary System (Script) Slide 1: Integumentary System Slide 2: An overview of the integumentary system Skin is the body s largest and heaviest organ making up 15% of body weight. Most skin is 1 to
Chemical Biology, Option II Mechanism Based Proteomic Tagging Case History CH1
 Proteome Wide Screening of Serine Protease Activity Proc Natl Acad Sci 1999, 97, 14694; Proteomics 2001, 1, 1067; Proc Natl Acad Sci 2002, 99, 10335; Biochemistry 2001, 40, 4005; J. Am. Chem. Soc., 2005,
Proteome Wide Screening of Serine Protease Activity Proc Natl Acad Sci 1999, 97, 14694; Proteomics 2001, 1, 1067; Proc Natl Acad Sci 2002, 99, 10335; Biochemistry 2001, 40, 4005; J. Am. Chem. Soc., 2005,
University of Groningen. Acantholysis in pemphigus van der Wier, Gerda
 University of Groningen Acantholysis in pemphigus van der Wier, Gerda IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the
University of Groningen Acantholysis in pemphigus van der Wier, Gerda IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the