Mouse Embryonic Hematopoietic Lineage Differentiation In Vitro
|
|
|
- Amberly Cox
- 6 years ago
- Views:
Transcription
1 Mouse Embryonic Hematopoietic Lineage Differentiation In Vitro by Chen Yao (Haydn) Liang A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy Graduate Department of Immunology University of Toronto Chen Yao (Haydn) Liang, 2014
2 ABSTRACT Hematopoietic differentiation from mouse embryonic stem cells (mesc) is the result of a complex and regulated process, dependent on a transcriptionally controlled network of signaling outcomes. Although several aspects of this differentiation process have been elucidated, critical components such as the identification of a hematopoietic-restricted progenitor have not been well characterized. In this study, mescs that lack the transcriptional regulator Ldb1 (Lim-domain-binding 1), which is required for the formation of a protein complex with Lmo2 (Lim-domain-only 2), Scl (stem cell leukemia protein; or Tal1, T cell acute lymphocytic leukemia 1), and GATA2 (GATA binding protein 2) were utilized. The loss of Ldb1 prevents the formation of this transcription regulatory protein complex, resulting in the lack of Runx1 (runt-related transcription factor 1) expression, which in turn leads to a loss of hematopoietic differentiation. This block in hematopoietic development allowed for the identification of a pre-hematopoietic lineage (phl, a developmental intermediate lacking hematopoietic lineage associated markers) progenitor that lacks CD41 (integrin 2 alpha) and CD45 (leukocyte common antigen) expression, but displays CD105 (endoglin), CD117 (ckit, stem cell factor receptor), CD202b (Tie2, angiopoietin receptor 2) expression. Through phenotypic and functional characterization of phl progenitor hematopoietic and endothelial differentiation potentials, as well as the gene expression profiling of Ldb1 deficient phl cells, it was observed that Ldb1-sufficient phl cells have the potential to give rise to CD45 + hematopoietic cells, which have the ability to subsequently develop into T-cells in vitro and thus fulfill a key aspect of definitive hematopoietic potential. It was also established that the loss of Ldb1 does not interfere with endothelial lineage differentiation. Another research focus of this thesis is the characterization of the effects of delivering HoxB4 (homeobox protein B4) protein to developing mescs in vitro by co-culture with OP9 stromal cells, ectopically expressing a secreted, soluble form of HoxB4. Previous findings showed that mesc-derived hematopoietic progenitors ectopically expressing HoxB4 via retroviral vectors could achieve long-term hematopoietic reconstitution after transfer into immune compromised mice. However, the use of retroviral vectors directly integrating into hematopoietic progenitors is potentially a major obstacle for clinical applications. Furthermore, the role HoxB4 plays in endothelial cell development is also poorly characterized. By engineering an HoxB4-OP9 stromal cell line that produces soluble HoxB4 proteins in co-culture with ESCs, coupled with the characterization of phl and CD201 + phl (phl expressing CD201, activated endothelial protein C receptor) progenitors, ESC/HoxB4-OP9 co-cultures were shown to yield comparable results of hematopoietic development as the retrovirally-induced ectopic HoxB4 expression Page ii
3 model. It is also shown that endothelial/hematopoietic bi-potent progenitors developed from HoxB4- OP9 co-cultures do not have significantly perturbed endothelial cell developmental potential. Together, the findings presented in this thesis support the use of pre-hematopoietic progenitors derived from embryonic stem cells as a viable approach to further characterize the potential to generate hematopoietic cells with in vivo reconstitution ability. Page iii
4 DEDICATION To my mother, Bei Min Qu, for your unconditional support. To Miho Tanaka, for preserving my sanity. To my colleagues, for giving me the utmost motivation. Page iv
5 ACKNOWLEDGEMENTS I am grateful for my supervisor Dr. Juan Carlos Zúñiga-Pflücker for giving me the opportunity to work on research projects ranging between T-cell development and function to ESC development, for the last 8 years. The many discussions we had taught me much more than simple scientific principles. Through this time I also had the privilege of being trained by amazing mentors, Dr. Lynn Rumfelt and Dr. Dzana Dervovic and their guidance and support made it possible for me to complete my PhD on time. I am thankful for my supervisory committee. Dr. Mira Puri had shown me that science is not just about cold facts. Dr. Norman Iscove for his excellent sense of observation, especially during committee meetings where his questions and comments always hit the mark and results in significant improvements for my project. Graduate school was much more endurable as a result of the many friendships I was fortunate enough to have made: Edwin Wang, Davie Lee, Ann McPherson, Julie Yu, Pak Hong Wong, and Courtney McIntosh. I thank them for their helpful discussions and the qualities that I admire in each of them. Finally, I would like to acknowledge my mother for her support which sometimes can be so subtle and quiet that I would not notice it until much time had passed. And to Miho Tanaka, we would not have met each other had we not chosen the road less travelled, together. Page v
6 TABLE OF CONTENTS ABSTRACT... ii DEDICATION... iv ACKNOWLEDGEMENTS... v TABLE OF CONTENTS... vi LIST OF FIGURES... ix Chapter I... ix Chapter II... ix Chapter III... ix LIST OF TABLES... xi Chapter II... xi Chapter III... xi LIST OF PUBLICATIONS... xii ABBREVIATIONS...xiii SELECTED CD NOMENCLATURE... xv ERUDITION... xvi CHAPTER I. INTRODUCTION... 1 Basic hematopoietic development concepts... 1 Human hematopoietic development in vivo... 2 In vitro hematopoietic development... 2 Assays used to study functional outcomes of hematopoietic development... 4 Molecular mechanisms of hematopoietic development... 5 Signaling pathways involved in hematopoietic development... 6 HSC Maintenance... 6 Current challenges and controversies THESIS FOCUS CHAPTER II Summary Introduction Results Phenotypical Characterization of Ldb1 -/- ESC/OP9 co-cultures Page vi
7 The generation of Sca1 + cells in Ldb1 -/- ESC/OP9 co-cultures Hematopoietic potential in the CD45 - fraction phl and phl+201 progenitors have T-cell potential Ldb1 -/- phl and phl+201 progenitors both have significantly decreased hematopoietic gene expression Ldb1 -/- ESCs can differentiate into endothelial cells in vitro Discussion Acknowledgements CHAPTER III Summary Introduction Results Secretable HoxB4 producing OP9 stromal cell line HoxB4 is secreted by the shoxb4-op9 stromal cells shoxb4-op9 increases hematopoietic development potential in vitro Role HoxB4 in the development of Sca1 + cells from ESCs in vitro Erythroid potential in progenitors derived from ESC/sHoxB4-OP9 co-cultures is more robust phl+201 progenitors produced from ESC/sHoxB4-OP9 co-cultures have lymphoid potential shoxb4-op9 does not play a role in endothelial cell development shoxb4-op9 induced gene expression changes in phl+201 cells are identical in hematopoietic progenitors ectopically expressing HoxB shoxb4 can induce gene expression changes in OP9 stromal cells Discussion Acknowledgements Chapter IV. Summary and Discussion Characterization of ESC hematopoiesis by time-lapse flow cytometry Defining the hematopoietic potential of phl and phl+201 cells Lessons learned from the gene expression microarray study Taking the gene expression microarray data one step further The role Ldb1 plays in endothelial cell development Engineering a soluble HoxB4 stromal cell line The effect of shoxb4 on ESC development in co-culture Defining the effect of shoxb4 on functional hematopoietic outcomes The effect of shoxb4 on gene expression in phl+201 progenitors The effect of shoxb4 on endothelial cell development Page vii
8 Assessing the potential changes in stromal cells Future directions CHAPTER V. MATERIALS AND METHODS Cell Lines ESC/OP9 co-culture, in vitro generation of T cells Flow cytometric analysis and cell sorting DNA Microarray QRT-PCR analysis Limiting dilution analysis Methyl-Cellulose Assay Matrigel culture REFERENCES Page viii
9 LIST OF FIGURES Chapter I Figure 1.1 Summaries of primitive and definitive hematopoietic development. Figure 1.2 Ldb1 stabilizes a complex of transcription factors (TF) involving Lmo2, Scl, and GATA2. Figure 1.3 Signaling factors involved in mammalian embryonic hematopoietic development. Figure 1.4 Purified LT-HSC from adult animals can reconstitute immune deficient hosts in vivo. Chapter II Figure 2.1 Time-lapse flow cytometry analysis of Ldb1 +/- and Ldb1 -/- ESC differentiation in coculture with OP9 stromal cells. Figure 2.2 Sca1 expression profile of Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures analyzed by flow cytometry between days 4 to 9 of co-cultures. Figure 2.3 Hematopoietic potential of CD45 - CD202b + and CD45 + cells was measured by methylcellulose assay. Figure 2.4 phl and phl+201 progenitors have T-cell potential. Figure 2.5 Limiting Dilution Assay was performed to determine the frequency of CD45 + cell generating capacity of phl and phl+201 progenitors. Figure 2.6 Gene array analysis of sorted phl progenitors from Ldb1 +/- and Ldb1 -/- ESC/OP9 cocultures. Figure 2.7 QRT-PCR was performed on sorted phl+201 progenitors. Figure 2.8 CD309 + cells were sorted on day 4 from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. Figure 2.9 Matrigel experiment was performed to assay for perturbations in endothelial cell development in CD309 + cells generated from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. Chapter III Figure 3.1 pmx-l-hoxb4-ires-hcd8 Construct. Figure 3.2 Western analysis of OP9 and HoxB4-OP9 stromal cells via anti-hoxb4 antibody shows the presence of the HoxB4 protein in the supernatant of HoxB4-OP9 cells, but not OP9 cells. Figure 3.3 Phenotypic characterization of ESC/HoxB4-OP9 co-cultures demonstrating dose dependent response. Page ix
10 Figure 3.4 The generation of Sca1 + cells are not significantly different in ESC/HoxB4-OP9 cocultures. Figure 3.5 Colony forming assay CD45 + and CD45 - CD202b + progenitors developed in ESC/OP9 or ESC/HoxB4-OP9 co-cultures. Figure 3.6 Both phl+201 progenitors derived from ESC/OP9 or ESC/HoxB4-OP9 co-cultures have T- cell potential. Figure 3.7 The role HoxB4-OP9 plays on endothelial development was assayed by the phenotypic characterization of the development of CD31 + cells. Figure 3.8 The role of HoxB4-OP9 in endothelial development was assayed by colony forming assays in MG cultures. Figure 3.9 QRT-PCR analysis of putative HoxB4 downstream genes in phl+201 progenitors derived from ESC/OP9 or ESC/HoxB4-OP9 co-cultures. Figure 3.10 Gene expression changes induced by HoxB4 in OP9 stromal cells were analyzed by QRT- PCR. Page x
11 LIST OF TABLES Chapter II Table 2.1 Surface markers used in the phenotypical characterization of ESC to HC development. Table 2.2 Average colony counts of hematopoietic colonies, broken down by cell type, which arose from CD45 + and CD45 - CD202b + progenitors at 8 days and 17 days in methyl-cellulose culture. Chapter III Table 3.1 The number of colonies observed in MC cultures from progenitors derived from ESC/OP9 or ESC/HoxB4-OP9 co-cultures. Page xi
12 LIST OF PUBLICATIONS Liang HC, Holmes R, and Zúñiga-Pflücker JC. Directed differentiation of embryonic stem cells to the T- lymphocyte lineage. Methods Mol. Biol : Dervovic DD, Liang HC, Cannons JL, Elford AR, Mohtashami M, Ohashi PS, Schwartzberg PL, Zúñiga- Pflücker JC. Cellular and Molecular Requirements for the Selection of In Vitro-Generated CD8 T Cells Reveal a Role for Notch. J. Immunol (4): Liang HC and Zúñiga-Pflücker JC. Stem Cells for an Immune System Restart. Stem Cell Research & Therapy. Submitted. Page xii
13 ABBREVIATIONS AGM Aorta-gonad-mesonephros region bfgf Basic fibroblast growth factor Bmp4 Bone morphogenic protein 4 CSF1 Colony stimulating factor 1 DAPI 4',6-diamidino-2-phenylindole EB Embryoid body FACS Fluorescence activated cell sorting GATA2 GATA binding protein 2 hang1 Human angiopoietin 1 HC Hematopoietic cell HoxB4 Homeobox protein B4 HSC Hematopoietic stem cell IL Interleukin KO Knockout Ldb1 Lim-domain binding protein 1 Lmo2 Lim-motif protein 2 LT Long-term mab Monoclonal antibody MACS Magnetic activated cell sorting mesc Murine/mouse embryonic stem cell mflt-3l Murine Flt-3 ligand MHC Major histocompatibility complex phl Pre-hematopoietic lineage phl+201 Pre-hematopoietic lineage expressing CD201 P-Sp Paraaortic splanchnopleura region Runx1 Runt related protein 1 Sca1 Ly-6A/E, lymphocyte antigen 6A/E SCF Stem cell factor Scl Stem cell leukemia protein Page xiii
14 Shh Sonic hedgehog Tal1 T cell acute lymphocytic leukemia 1 VEGF Vascular endothelial growth factor YS Yolk-sac Page xiv
15 SELECTED CD NOMENCLATURE CD4 L3T4, binds MHC class II CD8 Lyt-2/Lyt-3 (alpha/beta subunits), binds MHC class I CD31 Platelet endothelial cell adhesion molecule 1 CD34 Sialomucin, marker to enrich for early hematopoietic progenitors CD41 Integrin 2 alpha CD45 Ly-5, leukocyte common antigen CD105 Endoglin, accessory receptor for the TGF-beta superfamily CD117 c-kit, stem-cell growth factor receptor CD201 Activated endothelial protein C receptor CD202b Tie2, angiopoietin receptor 2 CD309 Flk1, KDR, VEGF receptor 2 Page xv
16 ERUDITION Whenever you feel like criticizing anyone just remember that all the people in this world haven t had the advantages that you ve had. - F. Scott Fitzgerald Page xvi
17 CHAPTER I. INTRODUCTION The hematopoietic developmental process has been studied in detail since the early 1900s. While E.R. Clark first reported the observation of vascular generation in tadpoles in 1909, and Stockard described blood-vessel development and coined the term angioblast in fish embryos in 1915, it was not until 1920 that F. Sabin described the formation of blood cells through the angioblast liquefying within the aorta [1-3]. Since then we have achieved several milestones in understanding this complex and highly conserved developmental process. Basic hematopoietic development concepts It was thought that mouse hematopoiesis develops initially in the yolk-sac (YS) where primitive hematopoietic development occurs [4]. This is also often referred to as fetal hematopoiesis, and was demonstrated by the production of embryonic erythrocytes that still possess a nucleus, express fetal hemoglobin, and an absence of any lymphoid potential [5]. This initial wave of fetal hematopoietic development appears to have short-term reconstituting function, as demonstrated by Dieterlen-Lievre using chicken-quail chimeras [6]. The translatability of these experiments across different biological systems such as in Rana sp. and Danio rerio, also demonstrates that this process is conserved across several species [2, 7-9]. Further studies in mice by Cumano and colleagues sought to address the issue of the anatomical origin of the long-term (LT) reconstituting hematopoietic stem cell found in adult animals [10]. As depicted in Figure 1.1a, early in murine embryonic development, hematopoietic progenitors can be found in the paraaortic splanchnopleura region (P-Sp) on embryonic day E7, but these progenitors are not LT reconstituting though they do possess lymphoid potential [11-13]. More advanced hematopoietic progenitors isolated from the aorta-gonad-mesonephros region (AGM) at E can give rise to lymphoid lineages and are LT reconstituting definitive hematopoietic stem cells (HSCs), but since this is post the establishment of the circulatory system in the embryo, their source was difficult to determine [13]. Cumano and colleagues approach was to culture explants containing hematopoietic progenitors isolated from YS and the embryo proper before the onset of circulation and determine their LT reconstitution potential [10]. Their results indicated that YS progenitors before the onset of circulation did not have LT reconstitution potential, while progenitors isolated from the AGM did, which indicates that primitive and definitive hematopoiesis may occur at distinct anatomical locations [10]. Overall, it was demonstrated that the definitive hematopoietic stem cell differs from the primitive hematopoietic stem cell by their lymphoid potential as well as their ability to achieve LT reconstitution Page 1
18 following transfer into adult recipients. More recently it has also been suggested that during fetal ontogeny, definitive hematopoietic development may also occur in subsets of endothelial cells in the heart [14]. Human hematopoietic development in vivo The study of in vivo hematopoietic development in humans is less understood relative to the mouse model due to limitations in experimental approaches. However, evidence resulting from studies in the human embryo has indicated that definitive HSCs emerge in the embryo proper from aortic endothelium similar to observations made in the mouse [15, 16]. It was also observed by morphological studies that HSC clusters can be found within the human aorta, and that these cells express CD34 (sialomucin) and CD45 (leukocyte common antigen), markers associated with the HSC fate [15]. Further similarities with the mouse model include the site of initial HSC formation within the aorta and subsequent hepatic colonization [15, 17, 18], and that definitive hematopoietic cells are derived from within the embryo and not the yolk-sac [15, 19]. In vitro hematopoietic development Given the observations made in animal models, several embryonic stem cell (ESC) based in vitro developmental models have been established in the last 10 years. The two most widely used models are the embryoid body (EB) system developed by the Keller group [20], and the OP9 co-culture system developed by the Honjo and Nakano groups [21, 22]. Embryoid bodies are generated by aggregating ESCs, and by doing so, cells expressing VEGFR2 (vascular endothelial growth factor receptor 2, or CD309), are induced to differentiate [20]. By isolating CD309 expressing cells and forming an aggregate a hemangioblast is produced that can give rise to hematopoietic cells that are multi-potent, which can give rise to erythroid, myeloid, and lymphoid lineages, but are not capable of LT reconstitution in vivo [20, 23]. On the other hand, by culturing ESCs with OP9 stromal cells, derived from the op/op mice that lack the CSF1 (colony stimulating factor 1) gene, it is also possible to partially recapitulate hematopoietic development [21]. The product of this developmental model is also multi-potent hematopoietic cells capable of erythroid, myeloid, and lymphoid development, but not capable of achieving LT reconstitution [21, 23]. Therefore, it is clear that the in vitro ESC hematopoietic developmental models lack fundamental components that can support the development of definitive HSCs in culture, and this prompted further research into the signaling pathways that form the basis of this developmental program. Page 2
19 Intra Embryonic Extra Embryonic A Primitive Hematopoiesis Definitive Hematopoiesis Yolk-Sac Paraaortic Splanchnopleura Region Aorta-gonadmesonephros region Fetal Liver Bone Marrow B Hemangioblast Primitive Hematopoietic Cell Definitive Hematopoietic Cell CD309 + CD202b + CD41 - CD202b + CD41 + CD202b - CD41 + CD202b - CD45 + Figure 1.1 Summaries of primitive and definitive hematopoietic development. A shows distinct anatomical regions of primitive and definitive hematopoietic development in both the mouse and human. B describes a subset of cell surface markers used in the characterization of this developmental process. CD309, vascular endothelial growth factor receptor 2. CD202b, Tie2 or angiopoietin receptor 2. CD41, integrin 2 alpha. CD45, leukocyte common antigen. Page 3
20 Assays used to study functional outcomes of hematopoietic development Many methods have been developed to assay the process of hematopoietic development, both in vitro and in vivo. While there are many protocols for assaying a myriad of molecular and functional outcomes, all of these methods can be categorized into two groups: functional assays and phenotypic/morphological assays. Phenotypic and morphological assays focus on the characterization of this developmental process by measuring distinct features that appear at specific time points. The oldest technique within this category is microscopy. Started over 100 years ago and still relevant today, hematopoietic cells are distinguished from its stromal environment by its non-adherent cell morphology [1, 24]. Time-lapse microscopy, in conjunction with advanced techniques of fluorescent labels has repeatedly demonstrated this process occurring with a market shifting of cell morphology, from adherent cell to non-adherent cell [7-9]. However, morphological observations have their limitations. The inability to efficiently isolate specific cells from a complex mixture based on appearance poses challenges to define developmental intermediates undergoing hematopoiesis. This shortcoming is addressed by the use of fluorescence activated cell sorting (FACS). FACS characterizes cell populations based on their protein expression. Applying this technique toward ESC to HSC development, several surrogate markers have been identified to be strongly associated with primitive hematopoietic progenitors and subsequent developmental intermediates. The most well-known of these is Flk1 (vascular endothelial growth factor receptor 2), which is found on hemangioblasts that are multipotent hematopoietic progenitors [20]. Other cell surface proteins used in the identification of HSCs include the LSK definition (Lineage - Sca1 + ckit + ) (Sca1, lymphocyte antigen 6A/E; ckit, or CD117 stem cell factor receptor) [25]. Cell surface markers used in this characterization process are shown in Figure 1.1b. Finally, as the hematopoietic developmental process is highly regulated and complex, the number of regulatory factors involved is also vast. The challenge of mapping out the regulatory network in an effort to characterize this process by means of distinct signal transduction and transcription regulation pathways is met with experimental techniques commonly found in molecular biology. From the simple assays such as PCR and western analysis to complex and high throughput gene arrays and ChIP (chromatin immune-precipitation) experiments, transcription factors such as Runx1 (or AML1, acute myeloid leukemia 1), Ldb1 (lim domain binding 1) have been identified to play crucial roles [26-28]. However, characterization of distinct features which appear during this process, whether it is a protein or gene being expressed or a cell shifting into a specific shape, functional outcomes cannot be determined. The definition of a hematopoietic progenitor is one based on function and not morphology Page 4
21 or phenotype. Therefore, a host of functional assays have been developed and used in conjunction with the methods mentioned above. The gold standard of all functional assays is of course, in vivo reconstitution. Successful serial reconstitution of immune deficient animals is the hallmark of a definitive hematopoietic stem cell [25]. Short of in vivo models, in vitro assays such as methyl cellulose and Matrigel have been used to demonstrate hematopoietic and endothelial potential at a clonal level. Coupled with in vitro culturing techniques that permit precursor-endpoint studies and limiting dilution assays, the tool-kit for functional analysis is thus complete [20, 29]. Molecular mechanisms of hematopoietic development At the core of the molecular mechanisms of this highly complex and regulated process is Runx1 (runt related protein 1, or AML1, acute myeloid leukemia 1 protein) [30]. In a Runx1 knockout (KO) mouse generated by Okuda et al., it was discovered that this genetic perturbation is embryonically lethal [30]. What was more interesting was that when examined more closely, there were signs of primitive hematopoiesis, but not definitive hematopoiesis [30]. Thus, Runx1 was established as a key transcription factor in the definitive hematopoietic development program. Other transcriptional regulators that act upstream of Runx1, include the Ldb1 (Lim domain binding 1)-Lmo2 (Lim motif 2)-Scl (stem cell leukemia protein)-gata2 (GATA box binding protein 2) protein complex that drives the expression of Runx1 [26, 31]. (Figure 1.2) Deficiency in Ldb1 abrogates the expression of Runx1 [26]. In the case of Scl, its absence leads to a redirection of the hematopoietic developmental program towards the cardiac smooth muscle lineage [27, 28]. Finally, GATA2 loss of function also results in embryonic lethality [32]. Downstream of Runx1 are transcription factors that play distinct roles in erythroid, myeloid, and lymphoid developmental programs such as GATA1 (GATA box binding protein 1), PU.1 (PU box binding protein 1, SPI1), and Runx1 itself, respectively [33-35]. Given that the main components of hematopoietic development depend heavily on multi-factorial transcriptional regulating protein complexes that can perform multiple functions depending on the context of their interactions, it is less likely that the hematopoietic developmental program is governed in a linear network connected by distinct signal transduction proteins or protein complexes where defined outcomes arise from a narrow set of inputs. Rather, it is likely that this process is regulated in a probabilistic manner similar to a quantum computer architecture where a defined input can lead to multiple outputs at varying probabilities based on the context of those inputs. The inputs in this case, would be a combination of cell-cell interactions as well as cytokine signals, and the output would be cell fate choices and the associated functional properties. Supporting this notion are studies of de-differentiation and the findings that indicate multiple unstable, Page 5
22 dynamically regulated intermediate states during somatic to embryonic de-differentiation [36]. If in such a process, every environmental cue is kept under full control so that development follows a linear path, there would be no stochastic outcomes observed. However the observations of stochastic developmental results indicate a much more elegant, probabilistic regulatory network that requires far fewer nodes of control and regulation. Signaling pathways involved in hematopoietic development As a complex and highly regulated process, there are multiple signaling molecules both intrinsic as well as extrinsic that contribute to the development of an HSC. A well characterized developmental pathway is mediated by the Notch receptor. Disruption of Notch signaling during embryonic development leads to vascular and cardiac abnormalities, as it is required for the endothelial to mesenchymal differentiation that forms the stromal niche [37-39]. The formation of this stromal niche via Notch1 signaling is required for the embryonic development of HSCs, as well as for HSC specification by inducing the expression of Runx1 possibly through the Ldb1 complex [40, 41]. Upstream of Notch1, is signaling via VEGF by precursors of HSCs, and prior to this are signaling events mediated by Shh (sonic hedgehog), which are received by stromal cells and induced the production of VEGF [41]. Other factors that contribute to either the development or specification of the HSC lineage, or the development of the stromal niche that supports this process include the Nodal/Bmp4/Wnt3a pathways and signaling via FGF [41]. Figure 1.3 illustrates this process and the potential players involved in early hematopoietic development. HSC Maintenance As mentioned above, the in vitro hematopoietic developmental models designed with ESCs can only achieve a partial recapitulation of the hematopoietic process observed in vivo. The key deficiency with in vitro models is the lack of LT reconstitution potential in the multi-potent hematopoietic cells that are produced [23]. This has been an area of intense research, and recently it was shown by the Daley group that this is possible by activating signaling programs downstream of HoxB4 (homeobox protein B4) [42]. HoxB4, originally discovered in Drosophila [43], belong to a family of homeobox proteins that dictate the embryonic development of various tissues, ranging from neurologic to the hematopoietic [44, 45]. Recent studies of the role HoxB4 plays in hematopoietic development, by ectopically expressing HoxB4 in ESCs via retroviral vectors, conducted by the Daley group have successfully demonstrated that the resulting hematopoietic cells generated in vitro could achieve LT reconstitution, as well as Page 6
23 Ldb1 Ldb1 Lmo2 Scl Lmo2 Gata2 Runx1 Figure 1.2 Ldb1 stabilizes a complex of transcription factors (TF) involving Lmo2, Scl, and Gata2 (Nottingham el al., Blood 2007). During hematopoietic differentiation this complex induces the expression of Runx1, which is required for definitive hematopoiesis, as previously reported (Lancrin et al., Nature 2009). Surface marker analysis revealed that generation of CD41 + cell is impaired in the absence of Ldb1. Page 7
24 Establishing the Niche Environment Inducing the Hematopoietic Program Activating the Hematopoietic Program Nodal Shh Notch1 Bmp4 VEGFa Scl Ldb1 Wnt3a GATA2 Lmo2 Runx1 Formation of the Primitive Streak Stromal cells induced to secrete cytokines to support hematopoietic development VEGFa signaling induces the expression of Notch1 in developing hematopoietic precursors. Notch1 signaling on stromal elements enables the Ldb1 complex in hematopoietic progenitors to induce the transcription of Runx1. Figure 1.3 Signaling factors involved in mammalian embryonic hematopoietic development. Initially to form a stromal environment supportive of hematopoietic development, Nodal/Bmp4/Wnt3a is required for patterning of the primitive streak. Once the stromal environment is established, Shh secreted by the notochord induces the production of VEGF by stromal cells which signal to activate the transcription of Notch1 in hematopoietic precursors. Notch1 signaling then induces the transcription of members of the Ldb1 complex, which in turn induces the transcription of Runx1. Page 8
25 HSC Ectopic HoxB4 Figure 1.4 Purified LT-HSC from adult animals can reconstitute immune deficient hosts in vivo. However, hematopoietic cells generated in vitro cannot achieve the same functional outcome. The self-renewal capacity can be induced with the expression of HoxB4 ectopically in developing ESCs, indicating that the potential is present, though the trigger is unknown. Page 9
26 maintaining multi-potent property [42]. (Figure 1.4) This prompted research into the role of HoxB4 in hematopoietic development and HSC self-renewal [46-49]. Current challenges and controversies Despite the achievement of the above-mentioned milestones, there is still much that remains uncertain regarding the origin of the hematopoietic system. It was initially described by morphology and subsequently by fluorescence microscopy using tagged proteins, such as CD309 and Runx1 that hematopoietic development occurs in close relationship temporally and spatially with endothelial cell development [1, 7-9]. Currently an issue of contention in this field of research is in the connection between endothelial and hematopoietic development. It has been hypothesized that hematopoietic cells arise from a bi-potent progenitor that can give rise to endothelial cells as well as hematopoietic cells, termed as hemangioblast [1, 46]. However, there is also a school of thought that argues that hematopoietic cells arise from a unique subset of endothelial cells, called hemogenic endothelium [46-48]. Evidence from Huber et al. and Vogeli et al. indicated that a single cell can give rise to endothelial and hematopoietic lineage progeny, and these results served to support the hemangioblast hypothesis [24, 49, 50]. But in this approach there are caveats such as the misidentification of progeny that express surface markers commonly associated with mature endothelial cells and the possibility that prehematopoietic precursors may express the same surface proteins cannot be ruled out. On the other hand, evidence for the existence of a hemogenic endothelial cell can be demonstrated by the in vivo developmental morphology where a non-adherent hematopoietic cell can be seen budding off of adherent cells in a possibly unique type of cell division [7-9]. This too has uncertainties since morphology does not imply function and the definition of an endothelial cell is mostly by their function as structural components of the vascular system [51]. In vitro studies that utilized genetic modifications that result in a block in hematopoietic development, performed by Okuda et al., Chen et al., and our group, have shown that it is possible to achieve endothelial cell development in the absence of hematopoietic development [30, 52, 53]. It is also noted that the block in hematopoietic development did not seem to allow a redirection towards the endothelial cell lineage even when the genetic modification occurs upstream of Runx1 expression [53]. Therefore it may be more likely that endothelial and hematopoietic cell development are regulated under distinct but related programs and that they may not occur concurrently as the hemangioblast hypothesis would predict. Other areas of intense debate are found in ESC-derived hematopoietic developmental models, and generally revolve around the common theme of the inability of ESCs to give rise to in vivo- Page 10
27 reconstituting HSCs. The first issue involves the characterization of this process using cell surface markers. While it is well understood that cell surface markers do not dictate the function of the cell expressing them, though occasionally the function of those markers may allow us to infer function; it is nonetheless very useful to identify distinct cell populations based on a defined set of cell surface markers that are expressed or absent. Successful characterization using this approach allows this process to be studied in similar ways as other developmental pathways are studied, by taking full advantage of powerful techniques such as fluorescence activated cell sorting (FACS) to isolate distinct subsets of complex population mixtures to test for function both in vitro as well as in vivo. Over the last several decades, this has not been achieved in a systematic and thorough manner. Initially, CD309 was shown to be expressed by endothelial cells at the site of hematopoietic differentiation in vivo [54]. Developing ESCs were then characterized by CD105 expression [55]. And in mature HSCs, CD117 and Sca1 expression was used for their identification as LSK (Lineage-negative Sca1 + c-kit + ) cells [25]. In all of the above mentioned reports, there may be many markers characterized in total, but in each report only a limited number of markers were analyzed in sparse temporal windows of ESC hematopoietic development. It would be insightful if the in vitro ESC hematopoietic developmental process was characterized using all of the above-mentioned markers and compared with corresponding in vivo subsets. When discussing controversies regarding in vitro ESC hematopoietic development, of course one must also consider the novel questions posed to us by the findings using ectopic expression of HoxB4. While its functional outcome is clear as demonstrated by the Daley group, its mechanism is not well understood [42]. Attempts at dissecting the transcriptional regulatory programs involved following the ectopic expression of HoxB4 in hematopoietic cells derived from ESCs in vitro were performed by Oshima et al [56]. By studying the changes in gene expression in these cells it was possible to elucidate the pathways that are activated as a result, however since HoxB4 was ectopically expressed in ESCs before the onset of differentiation, it is not clear whether the changes in gene expression are the result of HoxB4 expression or a combination of cross-talk between HoxB4 induced programs and feedbacks from the developmental environment. Furthermore, it was also noted by Lee et al. that ectopic expression of HOXB4 in human ESCs did not yield the same effect as was observed in murine ESCs [57]. Finding the gap between these two systems will be critical in the translation of the results from studies in murine ESC hematopoietic development toward the human system and eventually leading to clinical applications. Page 11
28 THESIS FOCUS This thesis focuses on the role of Ldb1 in mesc to HSC development, the characterization of a novel pre-hematopoietic lineage progenitor population, and the role of HoxB4 in mesc to HSC development. Ldb1 deficient ESCs displayed with a complete block in hematopoietic development. This phenotype was used to identify precursors to the definitive hematopoietic lineage cells and characterize this population based on surface marker expression, gene expression profile, as well as their hematopoietic and endothelial potential. Further analysis and comparison between Ldb1 deficient and wild-type precursor cells was conducted to determine the role that the Ldb1-depedent transcriptional network plays in murine embryonic hematopoietic development. Subsequent to the identification of a pre-hematopoietic lineage cell population, the role of HoxB4 was explored by introducing soluble HoxB4 proteins into the ESC/OP9 co-culture system. Previous findings by others had indicated that ectopic HoxB4 expression bestows upon in vitro-derived hematopoietic cells the ability to self-renew and in turn become effective in reconstitution experiments in vivo. By ectopically expressing a soluble HoxB4 protein in OP9 stromal cells, the downstream differentiation program in developing ESCs is expected to be affected by extrinsic HoxB4 expression without perturbing their genome. Exposure to HoxB4 in culture have shown by phenotypic characterization as well as gene expression profiling, to lead to an increase in the efficiency of hematopoietic cell differentiation from mescs. These findings, together with the characterization of a pre-hematopoietic lineage progenitor, serve as important steps towards the development of new and improved methods for the production of hematopoietic cells from ESCs, which may help shed light on the molecular mechanism that underpins the initiation of hematopoiesis. Page 12
29 CHAPTER II Block in hematopoiesis from Ldb1-deficient mouse embryonic stem cells reveals a pre-hematopoietic stage within hemogenic endothelial cells Haydn C.-Y. Liang*, Paul Love, and Juan Carlos Zúñiga-Pflücker* *Department of Immunology, University of Toronto, Toronto, Ontario M5S 1A8, Canada. National Institute for Child Health and Development, National Institutes of Health, Bethesda, MD Ldb1 +/- and Ldb1 -/- ESCs were provided by Dr. Love. Gene expression microarray analysis was performed by Dr. Love. Samples for gene array and all data shown were prepared and performed by H.C.Y. Liang. Manuscript in preparation. Page 13
30 Summary Though embryonic hematopoietic development in the mouse has been studied in detail, the identification of a hematopoietic-restricted progenitor remains elusive. During embryonic hematopoietic development, Runx1 (runt related protein 1, AML1) expression is driven by a transcriptional regulatory complex composed of Lmo2 (Lim motif 2), Scl (stem cell leukemia protein), GATA2 (GATA box binding protein 2), and Ldb1 (Lim-domain-binding 1). The phenotypic and functional effects of Ldb1 -/- embryonic stem cells (ESCs) within the context of hematopoietic development are also not well known. By utilizing ESCs deficient in the expression of Ldb1 in conjunction with the ESC/OP9 stromal cell co-culture method, the phenotypic and functional outcomes of this deficiency within this context were assessed in this study. Here, we have demonstrated that Ldb1 -/- ESCs have a developmental block before the CD41 + (integrin 2 alpha) CD202b + (angiopoietin receptor 2, Tie2) stage, but do not have any significant impairment on the generation of Sca1 + (lymphocyte antigen 6A/E) or CD31 + (platelet endothelial cell adhesion molecule 1) cells. Functionally, Ldb1 -/- ESCs are unable to give rise to hematopoietic cells in co-culture with OP9 stromal cells, but are able to successfully achieve endothelial cell differentiation. Moreover, through gene array and QRT-PCR analysis on purified prehematopoietic lineage progenitor populations, several hematopoietic genes that have decreased expression profiles when Ldb1 is absent were identified. Taking advantage of the block in hematopoietic development, two novel pre-hematopoietic lineage (phl) progenitors were isolated before the onset of CD41 + or CD45 + (leukocyte common antigen) cells, which were shown to have lymphopoietic potential and significant hematopoietic potential. Overall, the elucidation of the role Ldb1 plays in hematopoietic development and the subsequent identification and characterization of phl progenitors have provided the foundation to further study phl progenitors for in vivo reconstitution applications. Page 14
31 Introduction Hematopoietic differentiation from mouse ESCs is the result of a complex and well-regulated process, dependent on a transcriptionally regulated network of signaling outcomes [52, 58-60]. In vivo, definitive hematopoietic stem cells (HSCs) arise in the AGM (aorta-gonad-mesonephros) at embryonic day E9.5 [13]. In vitro, this process is partially recapitulated by the ESC/OP9 co-culture method with initial development of the CD309 (vascular endothelial growth factor receptor 2) expressing hemangioblast, followed by the appearance of CD45 + hematopoietic cells capable of giving rise to erythroid, myeloid, and lymphoid progeny [21, 23]. Orchestrating this developmental process is Runx1 or AML1 (acute myeloid leukemia 1 protein), where it plays a pivotal role in allowing the differentiation to proceed from primitive to definitive hematopoietic stem cells, as demonstrated by the embryonic lethality of knockout animals [30]. Upstream of Runx1 lies a transcriptional regulatory complex involving four factors: Ldb1 (Lim domain binding 1)-Lmo2 (Lim motif 2)-Scl (stem cell leukemia protein)-gata2 (GATA box binding protein 2) [26, 31]. Within this complex, Ldb1 functions as an integrator, the lack of which results in the abrogation of Runx1 induction during embryonic hematopoietic development. Data from chimera studies conducted by the Love group using Ldb1 -/- and Ldb1 +/+ ESCs in vivo have also demonstrated that Ldb1 -/- ESCs are unable to give rise to long-term reconstituting HSCs as a result of a significant disruption in both HSC specification as well as maintenance [61]. While much about this developmental pathway has been described in detail, a prehematopoietic progenitor cell remains to be identified and characterized. In this study, ESCs that lack the transcriptional regulator Ldb1 were utilized. To study the role of Ldb1 in this developmental process and taking advantage of the developmental block observed in Ldb1 -/- ESCs; time-lapse flow cytometry was employed to analyze changes in cell surface marker expressions as well as to characterize progenitor populations before the onset of specification. Through this analysis, it was confirmed that Ldb1 -/- ESCs have a definite block in hematopoietic cell differentiation as previous reported [61], and that endothelial cell development appeared to be unaffected in the absence of Ldb1. The endothelial developmental potential of Ldb1 -/- ESCs was further demonstrated by functional assays which showed that the loss of Ldb1 did not perturb this process. Following this analysis, specifically defined prehematopoietic lineage (phl) progenitors, were purified by flow cytometry. The potential to give rise to CD45 + hematopoietic cells as well as the ability to develop into T-cells in vitro from phl (prehematopoietic lineage cells lacking the expression of CD41 and CD45) and phl+201 (pre-hematopoietic lineage cells lacking the expression of CD41 and CD45, but express CD201 (activated endothelial protein Page 15
32 C receptor) progenitors were demonstrated, and thus fulfill a key aspect of definitive hematopoietic potential. Further gene expression profiling of Ldb1 -/- phl progenitors indicated that several functional gene clusters have decreased expression. Genes known to be involved in the hematopoietic developmental process such as genes within the Ldb1 complex were the most significantly reduced. These findings support the use of pre-hematopoietic progenitors derived from ESCs to further characterize the potential to generate hematopoietic cells with clinically relevant outcomes. Results Phenotypical Characterization of Ldb1 -/- ESC/OP9 co-cultures Ldb1 participates within a transcriptional regulatory complex that drives the expression of Runx1. Studies of hematopoietic development employing various fluorescent and/or time-lapse microscopy techniques have yielded valuable insights regarding this differentiation process [1, 7-9], we made use of a time-lapse flow-cytometry experiment to fully characterize the differentiation of ESCs in vitro on a population level by characterizing cell surface marker expression. The experimental design is detailed in Chapter V (Materials and Methods). Briefly, ESCs were co-cultured with OP9 stromal cells where hematopoietic development is supported. The study of this process by phenotypic surface marker characterization, functional assays of differentiation potential, and gene expression profiling took place within the period of 20 days of co-culture. For the phenotypic surface marker expression characterization, markers were chosen based on previous findings that associated their expression profile toward endothelial, hematopoietic, or shared fates. The markers described in this study are listed in Table 2.1, and are in concordance with previous findings that support the notion that hematopoietic and endothelial cells arise from a common hemogenic-endothelial progenitor [7-9]. Our results show that when starting equally with 5000 undifferentiated ESCs per co-culture, at early time-points CD309 + cells emerge on day 3-4 in both Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures (Figure 2.1a). To visualize a developmental pattern, the relative frequencies of cells that are positive for CD309 and/or CD201 were plotted as a function of time (Figure 2.1e). As shown in Figure 2.1e, the pattern of CD309 + cell development between both co-cultures follow the same profiles with respect to time, however there is a marked difference in the development of CD201 + cells, in which Ldb1 -/- ESCs appear to have less robust capacity to give rise to CD201 + cells. I next examined the development of primitive and definitive hematopoietic cells based on CD41 and CD45 expression. Figure 2.1b shows that CD41 + cells can be observed as early as day 4 and CD45 + cells can be observed as early as day 5 in Ldb1 +/- co-cultures; whereas Ldb1 -/- co-cultures did not produce Page 16
33 any CD41 + or CD45 + cells. There appears to be a step-wise order to the observed developmental process based on the expression of these markers, in which initially CD41 + CD45 - cells emerge, followed by CD41 + CD45 + cells and then finally by a gradual shift towards the CD41 - CD45 + phenotype. Figure 2.1f shows the compilation of this time-lapse flow-cytometry data, and further characterizes the profile of this developmental process. Since it has been proposed that hematopoietic cells (HCs) derive from the same hemogenicendothelium precursor as endothelial cells [7-9], an analysis involving CD202b and CD41 was performed. Figure 2.1c shows again a step-wise developmental pathway in which CD202b + CD41 - cells emerge initially at day 4-5 in both Ldb1 +/- and Ldb1 -/- co-cultures, followed by the rapid emergence of CD202b + CD41 + cells, and finally a shift toward the CD41 + CD202b - phenotype. This latter shift in phenotype occurs within a 16 hour time period. Figure 2.1g shows the compilation of this process and demonstrates that in Ldb1 -/- co-cultures the emergence of CD202b + cells are identical in timing as those observed in Ldb1 +/- co-cultures. However, as noted above, no CD41 + cells were present in Ldb1 -/- cocultures. When considering the emergence of HCs, a key marker of interest is CD117 (c-kit, stem cell factor receptor). Lin - Sca1 + c-kit + cells found in the adult bone marrow (LSKs) have long been recognized as containing hematopoietic stem cells [25]. In keeping with the shared hemogenic-endothelium precursor hypothesis mentioned above [7-9], CD105 (endoglin) was also included along with CD117 in a comparison between Ldb1 +/- and Ldb1 -/- co-cultures. Figure 2.1d shows that similar to the CD309 expression profile, there was no clear difference in the development of populations expressing CD117 and/or CD105 with respect to time. There were no distinct populations found to be present or absent exclusively in Ldb1 -/- co-cultures. Figure 2.1h shows the compilation of this analysis, in which the profile of CD117 + and/or CD105 + cells emerging in these co-cultures did not differ significantly. This is the first temporal comprehensive phenotypic characterization of this developmental process known to our group, and through the analysis of marker pairs and dissecting developmental patterns based on the expression of hematopoietic lineage associated markers (CD41, CD45, CD309, CD117) and endothelial lineage associated markers (CD202b, CD105, CD201); it is clear that Day 5 is when the Ldb1 sufficient and deficient ESC/OP9 co-cultures diverged in differentiation outcomes and the hematopoietic developmental block becomes apparent. Page 17
34 Table 2.1. Surface markers used in the phenotypical characterization of ESC to HC development. Marker Alternative nomenclature Rationale CD31 Pecam1 Found on mature endothelial cells CD41 Integrin 2 alpha Found on primitive hematopoietic cells CD45 Ptprc, pan-hematopoietic marker Found on definitive hematopoietic cells CD105 Endoglin Associated with endothelial cells, involved in endocardial cushion formation during fetal ontogeny [62]. CD117 ckit Used in the isolation of LSK cells CD201 Activated Endothelial Protein C Receptor Found of mature endothelial cells and bone marrow HSCs CD202b Tie2, Angiopoietin 2 receptor Found on mature endothelial cells. Required also for early fetal hematopoietic development via the formation of the endothelial niche [63]. CD309 Flk1, Kdr, Vegfr2 Required for the development of hematopoietic and endothelial cells H-2K k MHC-class I k haplotype Found on OP9 stromal cells and not ES-R1 cells. Sca1 Ly-6A/E Used in the isolation of LSK cells Page 18







35 CD309 A. Ldb1 +/- Ldb1 -/- 0 hour 8 hour 16 hour 0 hour 8 hour 16 hour Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8 CD201 Page 19












36 CD41 B. Ldb1 +/- Ldb1 -/- 0 hour 8 hour 16 hour 0 hour 8 hour 16 hour Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8 CD45 Page 20






37 CD41 C. Ldb1 +/- Ldb1 -/- 0 hour 8 hour 16 hour 0 hour 8 hour 16 hour Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8 CD202b Page 21



38 CD117 D. Ldb1 +/- Ldb1 -/- 0 hour 8 hour 16 hour 0 hour 8 hour 16 hour Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8 CD105 Page 22


39 % cells positive % cells positive % cells positive % cells positive % cells positive % cells positive E. Ldb1 +/- Ldb1 -/- F. G. Page 23

40 % cells positive % cells positive H. Ldb1 +/- Ldb1 -/- Figure 2.1 Time-lapse flow cytometry analysis of Ldb1 +/- and Ldb1 -/- ESC differentiation in co-culture with OP9 stromal cells. Starting with 5000 undifferentiated ESCs, cells were isolated from ESC/OP9 cocultures and analyzed by flow cytometry at every 8 hours between days 1 to 10 after the initiation of cocultures. A, B show flow cytometry plots for both co-cultures analyzed by CD41, CD45, CD201, and CD309 expression between days 1 to 8. C, D show flow cytometry plots for both co-cultures analyzed by CD41, CD202b (Tie2), CD117, and CD105 (Endoglin) expression. E-H show the bar graph representations of A-D; analyzed across 4 replicates. Error bars denote SEM of the top population as a representation of reproducibility. SEM Standard Error of the Mean. Page 24
41 The generation of Sca1 + cells in Ldb1 -/- ESC/OP9 co-cultures Sca1 or Ly-6A/E is expressed on HSCs found in the adult bone marrow [25], and in emerging hematopoietic lineage cells within the dorsal aorta [8]. Previously published findings by the Stanford group indicated that Sca1 deficient HSCs are less competitive when used in reconstitution with irradiated animals and it was also shown that these same cells possess significantly reduced capacity to secondarily reconstitute irradiated animals [64]. In our study, I examined the capacity for Ldb1 -/- ESCs to generate Sca1 + cells within the context of early hematopoiesis in association with the expression of CD202b, as well as within the context of late hematopoiesis, in association with the expression of CD117 (Figure 2.2a). Our group previously reported that at day 8 of ESC/OP9 co-cultures, CD45 + cells also expressed CD117 and Sca1 [55]. In the current study, I expanded this analysis to study this process at earlier and later time points focusing on the emergence of Sca1 + cells in relation to other sub-populations. Figure 2.2a shows that at day 4 of co-cultures, there is a significant CD202b + Sca1 - population and a CD202b - Sca1 + population, but not a significant double positive population. This is similarly observed in both Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. At day 5, a significant double positive population emerges and persists in both co-cultures until day 9. Starting at day 7, the population distribution of CD202b + Sca1 + double positive population in Ldb1 -/- co-cultures appeared to contain a different distribution relative to Ldb1 +/- co-cultures, but this was observed at later time points. With respect to Sca1 and CD117 expressing cells, Figure 2.2a also shows that in both Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures, Sca1 + and CD117 + cells are present, but since there are no CD45 + cells in Ldb1 -/- co-cultures the CD45 + Sca1 + CD117 + cells previously observed in such co-cultures were absent under these conditions. Figure 2.2b shows a bar graph representation of the flow cytometry plots shown in Figure 2.2a. The profiles of cells expressing Sca1 and/or CD202b or CD117 display the same trend in both Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. These results suggest that the initial steps of pre-hematopoietic differentiation appear to be supported in the absence of Ldb1. Hematopoietic potential in the CD45 - fraction Following the observations made above, I hypothesized that there is a CD202b + CD41 - CD45 - prehematopoietic progenitor that may have the potential to become hematopoietic under permissive Page 25




42 Sca1 A. Ldb1 +/- Ldb1 -/- Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD202b CD117 CD202b CD117 Page 26

43 % cells positive % cells positive % cells positive % cells positive B. Ldb1 +/- Ldb1 -/- C. Figure 2.2 Sca1 expression profile of Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures, initiated equally with 5000 undifferentiated ESCs per co-culture and analyzed by flow cytometry between days 4 to 9 of co-cultures. A shows the flow cytometry analysis of Sca1 expression in co-culture with respect to CD202b (Tie2) and CD117. B and C shows the bar graph representation of A, analyzed across 3 replicates. Error bars represent SEM for top population as a representation of reproducibility. Page 27
44 Table 2.2 Average colony counts of hematopoietic colonies, broken down by cell type, that arose from CD45 + and CD45 - CD202b + progenitors at 8 days and 17 days in methyl-cellulose culture. (n=3) Day 8+8 CD45 - CD202b + CD45 + Bone Marrow Erythrocyte Macrophage Granulocyte GM Pre-B Day 8+17 CD45 - CD202b + CD45 + Bone Marrow Erythrocyte Macrophage Granulocyte GM Pre-B Page 28
45 conditions. To test this hypothesis, CD45 - CD202b + cells from day 8 of ESC/OP9 co-cultures, starting with 10,000 ESCs per co-culture, were isolated by fluorescent activated cell sorting (FACS) and cultured in methyl-cellulose (MC) media supplemented with hematopoietic cytokines in parallel with CD45 + cells. Colonies were examined and counted 8 and 17 days after the initiation of MC cultures. Figure 2.3a shows that as expected, the CD45 - CD202b + fraction has erythroid potential as demonstrated by the successful generation of erythroid colonies at day 8 of MC culture. Whole bone marrow cells were used as positive controls. Figure 2.3b shows that at day 8 both the CD45 - CD202b + and CD45 + progenitors produced the same number of erythroid colonies. However at day 17 of MC cultures CD45 + progenitors had significantly more erythroid colonies. There was also a significant increase in the number of erythroid colonies from day 8 to day 17 of MC cultures using CD45 + progenitors. Despite the increase in the number of colonies in the CD45 + MC cultures, the pigmentation, as well as colony size, seen in the CD45 - CD202b + MC cultures were markedly more accentuated. Table 2.2 shows the colony counts for bone marrow control, CD45 + and CD45 - CD202b + samples. These results validate the initial hypothesis by demonstrating that robust hematopoietic potential is contained within the pre-hematopoietic CD45 - CD202b + fraction. phl and phl+201 progenitors have T-cell potential To more precisely characterize the pre-hematopoietic progenitors within the CD45 - CD202b + fraction of ESC/OP9 co-cultures, two specific subsets were further characterized for hematopoietic potential. The first was defined as CD41 - CD45 - CD201 - CD309 - CD105 + CD117 + CD202b +, and termed prehematopoietic lineage (phl) progenitors. The second subset is related, as it shares the same makers, but arises right after phl progenitors are first detected, and differs by only the expression of CD201, and thus termed phl+201 progenitors. Figure 2.4a shows the gating strategy used for the isolation of phl and phl+201 progenitors by FACS. Since lymphopoietic potential is a hallmark of hematopoietic stem cells, T-cell development potential was assessed using the OP9-DL1 system [65]. Previously developed by our group, it was demonstrated that when OP9 stromal cells were made to ectopically express the Notch1 ligand Deltalike 1, T-cell development from fetal and adult hematopoietic progenitors were supported at the expense of B-cell development [66]. Recent characterizations of the resulting T-cells derived from this system were shown to be both phenotypical and functionally identical to ex-vivo derived CD8 T-cells [67]. Both phl and phl+201 progenitors gave rise to T-lineage cells, as shown in Figure 2.4b, with progression through the double negative stages as well as the double positive and single positive stages of T-cell Page 29



.")

46 # of colonies A. Bone Marrow CD45 - CD202b + CD45 + Ery Ery M Ery G Ery B. * ** P= P= CD45 - CD202b + CD45 + Figure 2.3 Hematopoietic potential of CD45 - CD202b + and CD45 + cells was measured by methyl-cellulose assay. A shows images of erythoid colonies that have formed after 8 days in methyl-cellulose media. B shows the frequency of erythoid colonies that have formed after 8 and 17 days in methyl-cellulose media. Error bars denote SEM (n=3). In Day 17 co-cultures, CD45 + progenitors were able to produce significantly more erythroid colonies than CD45 - CD202b + progenitors. (P=0.0003) In CD45 + progenitors, there was also a significant increase of the number of erythroid colonies formed from day 8 to day 17 of culture in methyl-cellulose media. All images were taken at 10x magnification. Page 30
 CD202b CD202b")
47 CD105 CD105 CD117 CD117 SS DAPI # of cells A. FSC FSC CD41, CD45, CD309, H-2K k, (CD201) CD202b CD202b CD201 CD201 Page 31



48 CD4 CD4 B. phl Ldb1 +/- phl+201 ES-R1 phl Ldb1 -/- CD8 CD25 Figure 2.4 phl and phl+201 progenitors have T-cell potential. A shows the definition of phl and phl+201 cell as FACS strategies for phl (left) and phl+201 (right). Experiments requiring the isolation of phl and other pre-hematopoietic progenitors were initiated with 10,000 ESCs per co-culture to maximize yield. CD201 is included in the exclusion panel of markers when isolating phl progenitors. B shows the successful differentiation of phl and phl+201 progenitors into T lineage cells in vitro as shown by T-cell associated surface marker expression. Page 32
49 differentiation. The phl progenitors were also sorted from Ldb1 -/- ESC/OP9 co-cultures and as expected did not show any T-cell potential under permissive conditions. (Figure 2.4b) To determine the frequency of precursors that can undergo definitive hematopoiesis within the phl and phl+201 progenitor populations, a limiting dilution assay was performed. Figure 2.5 shows the precursor frequency of phl and phl+201 progenitors that can generate CD45 + cells in OP9 co-cultures after isolation by FACS. A frequency of 1 in 388 phl cells can give rise to CD45 + cells, while 1 in 546 phl+201 cells yield CD45 + cells. Figure 2.5 also shows the 95% confidence interval limits from this frequency to be between 1 in cells for phl progenitors and 1 in cells for phl+201 progenitors. Given the overlap with the obtained frequency for phl+201 cell and the 95% confidence intervals for phl cells, there is no significant difference between these two subsets in their ability to give rise to definitive hematopoietic progenitors. Ldb1 -/- phl and phl+201 progenitors both have significantly decreased hematopoietic gene expression To further elucidate the role that Ldb1 plays in the development of ESC to HC in vitro, a gene expression microarray analysis was performed in collaboration with Dr. P. Love, with purified phl progenitors, since we showed that this subset contained strong hematopoietic potential. Figure 2.6a shows functional genetic clusters that have increased or decreased expression when Ldb1 is absent in FACS-purified phl progenitors. Of note, gene clusters with significantly decreased gene expression profiles without Ldb1 were those that are involved in hematopoietic development, the homeobox domain protein regulatory network, Myc (myelocytomatosis oncogene) targets, and genes expressed in CD31 + cells, presumably endothelial cells due to the prevalence of CD31 expression in that specific cell type. Gene clusters with significantly increased gene expression profiles in the absence of Ldb1 were relatively few. The ones that were most significant were also genes that are involved in the homeobox domain protein regulatory network and epithelial cell development. Figure 2.6b shows a specific list of genes of interest that showed significant decreased in expression in the absence of Ldb1. Among this list, we noted that several Ldb1 complex members showed reduced expression in the absence of Ldb1, such as Scl/Tal1 and Lmo2. As expected, the downstream effector Runx1 showed a significantly decreased expression in the absence of Ldb1, therefore increasing the confidence in our findings. To confirm the observations made in the gene array analysis, a quantitative real-time PCR reaction (QRT-PCR) was performed using phl+201 progenitors. In this analysis, phl+201 progenitors Page 33
50 Number of CD45 - co-cultures (1/546) (1/388) 37% Number of progenitor cells seeded Figure 2.5 Limiting Dilution Assay was performed to determine the frequency of CD45 + cell generating capacity of phl and phl+201 progenitors. Progenitors were sorted and seeded into OP9 co-cultures for 5 days and assayed for CD45+ cells. Horizontal bars indicate the 95% confidence intervals; progenitor frequencies were: 1/388 and 1/546 for phl and phl+201, respectively. Page 34
51 A. B. Fold Change in Ldb-1 -/- phl Figure 2.6 Gene array analysis of sorted phl progenitors from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. Cells were sorted from day 6 co-cultures. A demonstrates functional gene clusters that have increased or decreased expression when Ldb1 is absent in phl progenitors. B shows key genes involved in hematopoietic differentiation that have increased or decreased expression when Ldb1 is absent in phl progenitors. Page 35
52 were utilized with the expectation that since they are phenotypically related to phl progenitors, these cells should share similar, if not nearly identical, gene expression patterns. Figure 2.7 shows the results of this analysis and corroborating with the results of the gene array analysis, Runx1, Oct4, Tie2, Myb, and Flk1 all showed statistically significantly decreased gene expression in phl+201 progenitors. Other genes assayed in this analysis included PU.1, a downstream effector of Runx1 [33], Flk1, CD41, and CD45. As expected, they also showed statistically significantly decreased gene expression in Ldb1 -/- phl+201 progenitors. However, unlike the results observed with phl progenitors, phl+201 progenitors did not show significantly decreased Scl/Tal1 expression. Ldb1 -/- ESCs can differentiate into endothelial cells in vitro To determine whether Ldb1 plays a role in the development of endothelial cells, co-cultures of Ldb1 +/- and Ldb1 -/- ESC/OP9 cells were performed and then assayed for the presence of endothelial lineage cells. Figure 2.8a shows flow-cytometric analysis for the expression of CD31 and CD45 on coculture-derived cells. We observed that CD31 + cell could be generated from Ldb1 -/- ESC/OP9 cocultures, and similar frequency was observed from Ldb1 +/- ESCs, including time of appearance. When multiple replicates were performed, the compiled data shown in Figure 2.8b, it revealed that CD31 + CD45- cells were generated in a similar manner from both Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. It is interesting to note that though it was not the intention of this experiment, a CD31 + CD45 + subpopulation was nonetheless observed in Ldb1 +/- but not Ldb1 -/- ESC/OP9 co-cultures. To functionally test for the potential to give rise to endothelial cells from Ldb1 -/- progenitors, a subsequent experiment using Matrigel (MG) was performed. Matrigel is an extra-cellular membrane protein mixture derived from the Engelbreth-Holm-Swarm mouse sarcoma [68]. It has been used since the 1980s for the culturing of endothelial cells and can be used to functionally assay for endothelial potential in a similar methyl-cellulose culture is used to assess hematopoietic potential [69]. When CD309 + progenitors were isolated from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures by magnetically activated cell sorting (MACS) and then cultured in Matrigel, endothelial cell colonies were observed arising from either progenitors at day 2 of MG culture (Figure 2.9a). On day 5 of MG cultures the colonies were more developed in both MG cultures. Figure 2.9b shows the number of colonies counted on day 5 of MG culture within a field of view that had the most number of endothelial cell colonies per sample, averaged over 3 samples each. We did not observe a statistically significant different number of EC colonies arising in MG cultures using Ldb1 +/- and Ldb1 -/- progenitors. Page 36
53 * * * * * * * Figure 2.7 QRT-PCR was performed on sorted phl+201 progenitors. Key genes involved in hematopoietic development that was previously observed to have decreased genetic expression in Ldb1 - /- phl progenitors were included in this analysis. Bars denote the fold different in gene expression between Ldb1 -/- and Ldb1 +/- phl+201 progenitors and error bars denote SEM (n=3). Asterisks indicated genes that have significantly decreased expression (p<0.05). Page 37



54 Ldb1 -/- CD31 Ldb1 +/- CD31 A. Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD45 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD45 Page 38
55 % cells positive % cells positive B. Ldb1 +/- Ldb1 -/- Figure 2.8 CD309 + cells were sorted on day 4 from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. Cells were them plated in Matrigel media which supports the differentiation of endothelial cells. Ldb1 deficiency does not have an impact on endothelial cell differentiation. A shows flow cytometry analysis of the generation of CD31 + and CD45 + cells in Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. B shows the bar graph representation of replicates of A (n=3). Page 39





56 # of colonies A. Ldb1 +/- Ldb1 -/- Day 4+2 Day 4+5 B. P= Ldb-1 +/- Ldb-1 -/- Figure 2.9 Matrigel experiment was performed to assay for perturbations in endothelial cell development in CD309 + cells generated from Ldb1 +/- and Ldb1 -/- ESC/OP9 co-cultures. A shows images of endothelial cell colonies differentiated in Matrigel media. All images were taken at 5x magnification. B shows the average counts of endothelial cell colonies on day 5. Error bars denote SEM. (n=3) SEM Standard Error of the Mean. Page 40
57 Discussion The main aim of this study was to identify and characterize an in vitro ESC derived prehematopoietic progenitor that is restricted to the hematopoietic lineage, by taking advantage of a developmental block caused by the loss of Ldb1. In pursuing this objective, the role Ldb1 plays within this process was also characterized and studied. Novel progenitor populations were identified by cell surface marker expression via systematic phenotypic characterization of the hematopoietic development of Ldb1 -/- ESCs in vitro. Through this approach and subsequent functional characterizations, two pre-hematopoietic progenitors that displayed hematopoietic and lymphoid potential in vitro, phl and phl+201, were identified and isolated.. Gene expression profiles of Ldb1 -/- phl and phl+201 progenitors were analyzed and it was found that Ldb1 plays a major role in hematopoietic development. However the loss of Ldb1 does not appear to have a significant effect in the expression of regulatory factors involved in other developmental pathways. Finally, the specificity of Ldb1 s involvement in the hematopoietic developmental pathway was confirmed by functional characterization of Ldb1 -/- progenitors where the lack of Ldb1 was found to not have a significant impact on endothelial cell development. CD309 + and CD202b + cells were observed to emerge from ESC/OP9 co-cultures at day 3 and day 4, respectively; confirming previous findings by other groups [24, 53]. Ldb1 -/- ESCs were unable to give rise to hematopoietic lineage cells since there was an absence of CD41 + and CD45 + cells as expected [61]. Further characterization of cell populations expressing CD201, CD202b, CD105, and CD117 indicated that a lack of Ldb1 did not impact significantly the generation of populations of cells other than those with a CD41 + and CD45 + phenotype; suggesting that Ldb1 -/- ESCs have a specific defect in hematopoietic lineage development. Sca1 has been more extensively studied in the context of T-cell development and its regulation in that role is regulated via the Src kinase pathway [70-72]. Its human homolog has not been identified, though in the mouse model it is commonly relied upon in the isolation of adult and fetal HSCs as part of the Lin - Sca1 + ckit + (LSK) population [25]. Generation of cells expressing Sca1 from Ldb1 -/- ESCs showed that the lack of Ldb1 did not impact the generation of Sca1 + cells, supports the notion that Ldb1 -/- ESCs do not have defects outside of the hematopoietic lineage and that the development of Sca1 + cells may be orchestrated apart from the main hematopoietic developmental program. Since Sca1 + CD117 + cells were observed in Ldb1 -/- ESC/OP9 co-cultures, we hypothesized that these may be a CD41 - CD45 - prehematopoietic progenitor that may be precursors to the hematopoietic lineage. Methyl-cellulose Page 41
58 cultures with CD45 - CD202b + progenitors supported this hypothesis, as these cells were able to give rise to erythroid colonies. The CD45 - CD202b + population was then further fractionated, and 2 subsets that possessed hematopoietic capacity, phl and phl+201, were identified. Lymphoid potential within phl and phl+201 was demonstrated by in vitro assays, and thus demonstrating the presence of cells capable of definitive hematopoiesis [10, 20]. The frequency of phl and phl+201 cells to give rise to CD45 + cells was determined to be within the 1 in 300 to 500 range, suggesting that further purification is needed, or that the conditions to induce hematopoietic lineage conversion require further optimization. Together with the observation that Ldb1 -/- phl cells do not survive in co-culture with OP9-DL1 cells (data not shown), these findings support the notion that phl and phl+201 progenitors are restricted to the hematopoietic lineage. Following the cell surface characterization of Ldb1 -/- ESCs, gene expression microarray analysis of Ldb1 -/- and wild-type phl cells was performed. The hypothesis that without Ldb1, many genes related to hematopoietic development would be affected and would have significantly decreased expression was supported by our findings. Of note, genes that are associated with the endothelial cell lineage also showed decreased expression, supporting the view that hematopoietic cells are derived from a common hemogenic endothelial progenitor [7-9]. To confirm and further characterize the related phl+201 progenitors, QRT-PCR experiments were performed, comparing between Ldb1 +/- and wild-type phl+201 progenitors. As phl+201 and phl are very similar in phenotype and show coordinated appearance in culture, it was expected that these subsets would share similar gene expression profiles. The hematopoietic genes chosen for analysis indicated that this was indeed the case. Key hematopoietic genes such as Runx1 and Oct4 showed significantly decreased expression as expected, although Scl did not show a similar trend. To address whether Ldb1 function affects the development of endothelial cells and therefore disrupts pre-hematopoietic development, flow cytometry analysis was performed to look for the production of CD31 + cells, which would inform us of their endothelial cell developmental potential [51, 73]. However, CD31 is not exclusively expressed on endothelial cells [74] and therefore Matrigel cultures were performed to assess whether endothelial cell development was impaired. Ldb1 -/- progenitors demonstrated robust endothelial cell developmental potential, and therefore we can conclude that the absence of Ldb1 appears to specifically impact hematopoietic development and does not affect the earlier stages of development that include progenitors with shared hematopoietic and endothelial cell differentiation potential. Page 42
59 Overall, the examination of Ldb1 deficient ESC hematopoietic development revealed that this loss of function mutation ablates hematopoietic development but not endothelial cell development. Additionally, the clear block in hematopoietic differentiation found in Ldb1 -/- ESCs enabled characterization of a pre-hematopoietic stage, which was shown to be CD41 - CD45 - CD105 + CD117 + CD202b + and capable of definitive hematopoietic development. Further characterization of this subset is still required to determine how its appearance is regulated in vitro, and to also determine whether these cells are capable of in vivo hematopoietic reconstitution. Acknowledgements We thank Roxanne Holmes for assisting with ESC/OP9 co-cultures. Sumugan Shamuganathan for helping to design QRT-PCR primers. This work supported by the Canadian Cancer Society, and the Canadian Institutes of Health. Page 43
60 CHAPTER III Soluble HoxB4 enhances hematopoietic and endothelial cell differentiation of mouse embryonic stem cells in vitro Haydn C.-Y. Liang*, Sumugan Shamuganathan*, Bilal Ahmadi*, and Juan Carlos Zúñiga-Pflücker* *Department of Immunology, University of Toronto, Toronto, Ontario M5S 1A8, Canada. All data shown were prepared and performed by H.C.-Y. Liang. Manuscript in preparation. Page 44
61 Summary Hematopoietic development has been studied in detail and the results of decades of research have enabled us to model this developmental process in vitro with embryonic stem cells (ESCs). However, the differentiation of a functional hematopoietic stem cell (HSC) remains an elusive goal. In both human and mouse models, it has been shown that pluripotent hematopoietic cells capable of lymphoid development can be generated in vitro. However the resulting progenitors consistently fail to achieve in vivo reconstitution of immune deficient hosts. Recent studies involving HoxB4 (homeobox protein B4) has indicated that its expression in developing ESCs has a significant impact on the selfrenewal capacity of downstream hematopoietic progeny and can bestow upon the resulting hematopoietic cells the potential for long term reconstitution. However, the potential for HoxB4 to be applied practically in a clinical setting remain unaddressed. With the use of retroviral expression vectors being unfeasible, the challenge is compounded by the short half-life of purified HoxB4 protein. Here we demonstrate that soluble Hoxb4 delivered by OP9 stromal cells in co-culture with ESCs can effectively emulate effects of ectopic HoxB4 expression by retroviral vectors. By taking advantage of the homeobox domain and allowing HoxB4 to be constantly produced in proximity to developing ESCs, we successfully demonstrated increased expression of downstream HoxB4 effectors in in vitro developed hematopoietic progenitors comparable to previous studies involving similar progenitors expressing HoxB4 ectopically. When co-cultured with HoxB4 producing OP9 cells, developing ESCs also demonstrated increased hematopoietic potential at earlier time points, as well as with increased frequency. Functional analyses have indicated that erythroid development is significantly improved while endothelial cell development remains unaffected. Overall, the results of this study support the use of soluble HoxB4 in clinical applications and demonstrate the effectiveness of a novel cell conditioning system that can be utilized in a wide variety of applications. Page 45
62 Introduction Being able to self-renew and differentiate into all blood cell lineages, hematopoietic stem cells (HSCs) are the basis of the circulatory and immune system. In vivo, their ontogeny is thought to initiate from the para-aortic splanchnopleura (P-Sp) as primitive HSCs incapable of long term (LT) reconstitution while able to give rise to lymphoid progeny [11-13]. This is followed by a second wave of definitive HSC development found in the aorta-gonad-mesonephros (AGM), which does possess the potential for LT reconstitution [13]. In vitro hematopoietic development utilizes the totipotency of embryonic stem cells (ESCs) either alone as embryoid body (EB) cultures, or in co-culture with stromal cells such as OP9 cells [20, 21]. Regardless of method, the resulting hematopoietic cells (HCs), while capable of giving rise to lymphoid progeny under permissive conditions, fail to successfully demonstrate the ability to long term reconstitute immune deficient hosts [20, 23]. This fundamental difference between in vitro and in vivo derived hematopoietic progenitors has been a focus of investigation for decades. One gene is of particular interest in this context. HoxB4 (homeobox protein B4), originally discovered in Drosophila, belongs to the family of homeobox proteins that play crucial roles in embryonic development and segmentation [43]. While HoxB4 has been implicated in other developmental processes aside from hematopoiesis, in this context the most prominent discovery pertains to its potential role in HSC maintenance. When HoxB4 is ectopically expressed by mouse ESCs, it is able to bestow the capacity for self-renewal upon in vitro derived HCs and enable their long term reconstitution of immune deficient hosts [42, 75]. However, previous studies employed methodologies for inducing the downstream effects of HoxB4 expression that are unfeasible for translation toward clinical applications. The use of retroviral expression vectors opens up the possibility for malignant transformation, while the purified HoxB4 protein have been reported to have a short half-life on the order of hours in vitro [75, 76]. By taking advantage of HoxB4 s innate ability to be internalized [77], OP9 stromal cells ectopically expressing sufficient levels of soluble HoxB4 (shoxb4) were developed to affect a measurable impact on the hematopoietic development of ESCs in co-culture. Phenotypic and functional characterizations of the resulting hematopoietic progenitors indicate that the effect of shoxb4 on developing ESCs delivered in this manner is comparable to previous observations involving ESCs ectopically expressing HoxB4. As a secondary objective, the effects of HoxB4 on various stages of hematopoiesis, as well as endothelial development during ESC to HC development in vitro were also investigated. Since HoxB4 is Page 46
63 not ectopically expressed in the developing ESCs as was in previous studies [42, 75], the outcomes therefore may differ. Furthermore, when an external force that affects hematopoiesis is introduced, there is always the possibility that the closely related endothelial developmental pathway may also be perturbed [7-9]. The shoxb4 producing OP9 stromal cells provided the optimal condition to examine these questions more closely. Through phenotypic characterization and functional assays using methylcellulose and Matrigel cultures, our results have indicated that shoxb4 increase both the progression of hematopoiesis as well as the number of resulting HCs; while showing no observable effects on endothelial cell development. Page 47
64 Results Secretable HoxB4 producing OP9 stromal cell line To enable the secretion of HoxB4 by OP9 stromal cells, a retroviral bi-cistronic vector was used. The pmx vector [78] used included the gene encoding for the human CD8 alpha chain (hcd8a) preceded by an intra-ribosomal-entry-site (IRES). The HoxB4 gene fused to a 5 IgG secretion signal peptide was cloned upstream of the IRES within the pmx vector (Figure 3.1). OP9 cells were transduced with the resulting retroviral construct, and shoxb4-expressing OP9 cells (shoxb4-op9) were isolated using FACS (fluorescence activated cell sorting) with the hcd8a as a surrogate marker to identify shoxb4 transduced cells. HoxB4 is secreted by the shoxb4-op9 stromal cells To confirm that HoxB4 is secreted in culture, the supernatant of shoxb4-op9 cells were assayed for the presence of the HoxB4 protein. Western blot analysis of supernatants isolated from OP9 and shoxb4-op9 stromal cells indicated that HoxB4 is indeed secreted by OP9 cells in culture. (Figure 3.2) The anti-hoxb4 antibody used did not bind BSA (bovine serum albumin) or supernatants from control OP9 stromal cells, and specific binding to HoxB4 from shoxb4-op9 cell supernatant was detected, and the product had the expected molecular size. shoxb4-op9 increases hematopoietic development potential in vitro Two distinct shoxb4-op9 cell lines were made that expressed shoxb4 at different levels, as characterized indirectly by flow cytometry detecting the hcd8a protein (Figure 3.3a). This provided us with a unique opportunity to determine the relative effect on ESC-derived hematopoietic development in vitro with respect to varying concentrations of HoxB4 in the cultures. Figure 3.3b shows the analysis of ESC/OP9 and ESC/sHoxB4-OP9 co-cultures from 2 different HoxB4 expression levels with respect to the development of CD41 + (integrin 2 alpha) and CD45 + (leukocyte common antigen) cells. The results showed a dose dependent response to HoxB4 in the appearance of both CD41 + and CD45 + cells. In ESC/sHoxB4-OP9 (low) co-cultures CD41 + cells appeared to emerge at day 5 with 5 times higher frequency relative to cells in ESC/OP9 co-cultures. In ESC/sHoxB4- OP9 co-cultures CD41 + cells emerged 1 day earlier, at day 4, and by day 5 the frequencies were 25 times higher than OP9 co-cultures. This difference was repeated throughout the course of ESC/OP9 co-culture development. By day 9 the difference in the generation of CD41 + and CD45 + cells relative to the level of Page 48
65 IgG Secretion Signal Peptide IRES 5 3 HoxB4 hcd8 Figure 3.1 pmx-l-hoxb4-ires-hcd8 Construct. A human CD8 alpha (hcd8a) gene is placed downstream of an intra-ribosomal entry site (IRES), which is also downstream of the HoxB4 gene. Fused to the 5- prime of the HoxB4 gene is the excretion signal peptide taken from the IgG kappa gene. Page 49
66 BSA shoxb4-op9 OP9 shoxb4 GP+E kda 64 kda 26 kda 19 kda 6 kda Figure 3.2 Western analysis of OP9 and shoxb4-op9 stromal cells via anti-hoxb4 antibody shows the presence of the HoxB4 protein in the supernatant of shoxb4-op9 cells, but not OP9 cells. Bovine serum albumin (BSA) is used to detect non-specific binding and supernatant from GP+E.86 cells transiently transduced with the shoxb4 plasmid is used to confirm the specificity of the anti-hoxb4 antibody. BSA, bovine serum albumin. Page 50
67 A. hcd8 Page 51



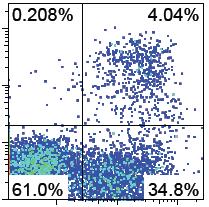








68 CD45 B. OP9 shoxb4-op9 (low) shoxb4-op9 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD41 Page 52













69 CD41 C. OP9 shoxb4-op9 (low) shoxb4-op9 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD202b Page 53
70 % cells positive % cells positive D. OP9 shoxb4-op9 (low) shoxb4-op9 E. OP9 shoxb4-op9 (low) shoxb4-op9 Figure 3.3 Phenotypic characterization of ESC/sHoxB4-OP9 co-cultures demonstrating dose dependent response. Cell surface marker expression was assayed by flow cytometry. A. Various levels of shoxb4 expressing OP9 stromal cells were isolated based on their expression of the hcd8a surrogate marker via FACS. B. Phenotypic characterization of ESC/OP9 co-cultures with varying levels of shoxb4 expression by OP9 stromal cells with respect to CD41 + and CD45 + cells. C. Phenotypic characterization of ESC/OP9 cocultures with varying levels of shoxb4 expression by OP9 stromal cells with respect to CD202b + and CD41 + cells. D. Compilation of triplicates of phenotypic characterizations shown in B. E. Compilation of triplicates of phenotypic characterizations shown in C. hcd8, human CD8 alpha chain. CD41, integrin 2 alpha. CD45, leukocyte common antigen. CD202b, Tie2 or angiopoietin receptor 2. Page 54
71 shoxb4 expression in co-cultures became less clear, but relative to OP9 co-cultures without shoxb4 expression there were significantly more CD41 + and CD45 + cells produced in the ESC/sHoxB4-OP9 cocultures. Figure 3.3c shows the analysis of the same co-cultures, but focusing instead on CD202b + (Tie2, angiopoietin receptor 2) and CD41 + cells. At day 4, co-cultures exposed to HoxB4 demonstrate a clear evidence of CD202b + cell development. As seen in Figure 3b, there was a strong dose dependent effect with respect to the levels of shoxb4 expressed by the OP9 cells. A similar pattern was repeated throughout the developmental process, and by day 9 the frequency of the CD41 + single positive population in ESC/sHoxB4-OP9 (low) co-cultures increased by 10 fold with respect to control ESC/OP9 co-cultures; and showed a 20 fold increase in ESC/sHoxB4-OP9 co-cultures. Figure 3.3d shows a compilation of data describing the development of CD41 + and CD45 + cells between days 4 to 9 of co-cultures. As seen in Figure 3.3b, there was a positive correlation between the levels of HoxB4 expression by OP9 cells and the frequency of CD41 + and CD45 + cells generated in their respective co-cultures. Figure 3.3c shows a compilation of data describing the development of CD202b + and CD41 + cells between days 4 to 9 of ESC/OP9 co-cultures. As described in Figure 3.3c, there is also a positive correlation similar to the one observed in Figure 3.3d. Role HoxB4 in the development of Sca1 + cells from ESCs in vitro Sca1 or Ly-6A/E is expressed on HSCs found in the adult bone marrow. [25] Previous studies indicate that Sca1 deficient HSCs are less competitive when used in reconstitution assays of irradiated animals [64]. In this study, the role HoxB4 plays in the development of Sca1 + cells within the context of early ESC-derived hematopoiesis was examined in association with the expression of CD202b; as well as within the context of late hematopoiesis in association with the expression of CD117 (ckit, stem cell factor receptor) (Figure 3.4a). Our group previously showed that at day 8 of ESC/OP9 co-cultures CD45 + cells also expressed CD117 and Sca1. [55] In the current study, this analysis was expanded further to study this process at earlier and later time points focusing on the emergence of Sca1 + cells in relation to other subpopulations. Figure 3.4a shows that at day 4 of co-culture, there is a significant CD202b + Sca1 - population and a CD202b - Sca1 + population, but not a significant double positive population. This was similarly observed in both ESC/OP9 and ESC/sHoxB4-OP9 co-cultures. At day 5 a significant double positive population emerges and persists in both co-cultures until day 9. Page 55





72 Sca1 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 A. OP9 shoxb4-op9 CD202b CD117 CD202b CD117 Page 56

73 % cells positive % cells positive B. OP9 shoxb4-op9 C. Figure 3.4 The generation of Sca1 + cells is not significantly different in ESC/sHoxB4-OP9 co-cultures. A. ESC/OP9 and ESC/sHoxB4-OP9 co-cultures were phenotypically characterized by flow cytometry from day 4 to day 9 focusing on cells that express Sca1, CD202b, and/or CD117. B and C shows the aggregate data for this characterization of the development of CD202b and Sca1 expressing cells as well as CD117 and Sca1 expressing cells respectively. Sca1, lymphocyte antigen 6A/E. CD202b, Tie2 or angiopoietin receptor 2. CD117, ckit or stem cell factor receptor. Page 57
74 With respect to Sca1 and CD117 expressing cells, Figure 3.4a also shows that in both ESC/OP9 and ESC/sHoxB4-OP9 co-cultures, Sca1 + and CD117 + cells are observed. Overall, there was a marginal reduction in the production of Sca1 + cells in ESC/sHoxB4-OP9 co-cultures. Figure 3.4b provides a bar graph representation of the flow cytometry results shown in Figure 3.3a. The profile of cells expressing Sca1 and/or CD202b or CD117 shows the same trend in both ESC/OP9 and ESC/sHoxB4-OP9 co-cultures. Erythroid potential in progenitors derived from ESC/sHoxB4-OP9 co-cultures is more robust Following the observations made above, I hypothesized that shoxb4-op9 may positively impact ESC-derived hematopoiesis by accelerating the developmental process as well as by inducing more robust proliferation. To test this notion, functional properties of CD45 - CD202b + and CD45 + cells obtained from day 8 ESC/OP9 and ESC/sHoxB4-OP9 co-cultures were tested. Following fluorescent activated cell sorting (FACS), co-culture-derived progenitors were placed in methyl-cellulose (MC) media, and colonies were examined and counted 8 and 17 days after the initiation of MC culture to assay for hematopoietic colony formation. Figure 3.5a shows that progenitors derived from ESC/sHoxB4-OP9 co-cultures produced more robust erythroid colonies in MC cultures. Figure 3.5b shows that CD45 - CD202b + progenitors produced the same number of erythroid colonies regardless of the co-culture they were derived from; while in contrast CD45 + progenitors derived from ESC/sHoxB4-OP9 co-cultures produced significantly more erythroid colonies relative to their ESC/OP9 co-culture derived counterparts (p=0.0003). At day 17, MC cultures of both CD45 - CD202b + and CD45 + progenitors derived from ESC/sHoxB4-OP9 colonies produced significantly more erythroid colonies than progenitors derived from ESC/OP9 co-cultures (p=0.0071, p<0.0001, respectively). Table 3.1 shows the number of colonies observed from bone marrow cells as well as CD45 + and CD45 - CD202b + progenitors produced from ESC/OP9 and ESC/sHoxB4-OP9 co-cultures. phl+201 progenitors produced from ESC/sHoxB4-OP9 co-cultures have lymphoid potential While HoxB4 plays a role in the development of hematopoietic cells in ESC/OP9 co-cultures as demonstrated by assays of the development of CD41 + and CD45 + cells, as well as the erythropoietic potential of CD45 + or CD45 - CD202b + progenitors on MC media; the question of lymphopoietic development potential of phl+201 progenitors remain unanswered. phl+201 progenitors (prehematopoietic lineage progenitors previously characterized by the following phenotype: CD202b + CD105 + (endoglin) CD117 + CD309 - (Flk1, vascular endothelial growth factor receptor 2) CD41 - CD45 - Page 58









75 A. OP9 shoxb4-op9 CD45 - CD202b + CD45 + CD45 - CD202b + CD45 + Page 59


76 # of erythroid colonies B. OP9 shoxb4-op9 Day 8+8 Day 8+17 P= * P= * P< * CD45 - CD202b + CD45 + CD45 - CD202b + CD45 + Figure 3.5 Colony forming assay CD45 + and CD45 - CD202b + progenitors developed in ESC/OP9 or ESC/sHoxB4-OP9 co-cultures. Progenitors were isolated by FACS at day 8 of co-culture and plated on MC culture. Analyses of MC cultures were performed on day 8 and 17 post MC culture initiation. A. Shows the development of erythroid colonies in MC culture from progenitors derived in either ESC/OP9 or ESC/sHoxB4-OP9 co-cultures. Up to three representative fields are shown. B. Shows the number of erythroid colonies observed on day 8 and 17 of MC cultures. MC, methyl-cellulose. CD45, leukocyte common antigen. CD202b, Tie2 or angiopoietin receptor 2. Page 60
77 Table 3.1 The number of colonies observed in MC cultures from progenitors derived from ESC/OP9 or ESC/sHoxB4-OP9 co-cultures. GM, granulocyte-macrophage colonies. ESC/OP9 ESC/sHoxB4-OP9 Day 8+8 CD45 - CD202b + CD45 + CD45 - CD202b + CD45 + Bone Marrow Erythrocyte Macrophage Granulocyte GM Pre-B ESC/OP9 ESC/sHoxB4-OP9 Day 8+17 CD45 - CD202b + CD45 + CD45 - CD202b + CD45 + Bone Marrow Erythrocyte Macrophage Granulocyte GM Pre-B Page 61
78 CD201 + (activated endothelial cell protein c receptor)) were characterized functionally to be progenitors that are phenotypically before hematopoietic lineage commitment and possessing lymphoid potential. As phl+201 progenitors differentiated in co-cultures with shoxb4-op9 cells and could be induced to give rise to T-cells by co-culture with OP9-DL1 (OP9 stromal cells expressing the notch ligand delta-like 1) cells in the absence of HoxB4, the hypothesis was that the initial exposure to HoxB4 would not have any deleterious effects on subsequent T-cell development. The expectation was that once removed from this stromal environment, the short half-life of HoxB4 would ensure that no further influence was exerted on developing cells. [75] In Figure 3.6, phl+201 progenitors were isolated by FACS and subsequently cultured on OP9-DL1 cells, to induce T-cell development in vitro, which was assessed by examining for the cell surface expression of CD4 and CD8. Surprisingly, phl+201 progenitors derived from ESC/sHoxB4 co-cultures demonstrated less robust T-cell development in this developmental model. shoxb4-op9 does not play a role in endothelial cell development Based on previous studies that have indicated that hematopoietic cells arise from a hematopoietic/endothelial bi-potent progenitor [7-9], it was hypothesized that this observed effect on hematopoietic development could be due to the effects exerted on the hematopoietic lineage and that shoxb4 may also play a role in the endothelial cell developmental process. To test this hypothesis, the development of CD31 + (platelet endothelial cell adhesion molecule 1) cells were phenotypically characterized. Figure 3.7a shows that the development of CD31 + cells with respect to CD45 + cells was not significantly different between ESC/OP9 and ESC/sHoxB4-OP9 co-cultures. Compilation of this data across triplicates in Figure 3.7b also confirmed this finding. The development of CD31 + cells followed the same pattern in either co-cultures. Functionally, CD309 + cells isolated from ESC/OP9 and ESC/sHoxB4-OP9 co-cultures were subsequently cultured on Matrigel media (MG) that was permissive for endothelial cell development; and both demonstrated robust endothelial cell colony formation (Figure 3.8a). The number of endothelial cell colonies formed from CD309 + progenitors from either co-culture conditions indicate that there is no statistical significance between progenitors derived from either co-culture conditions (P=0.0615) (Figure 3.8b). Taken altogether, the effects of shoxb4 on hematopoietic development seem to act exclusively on the hematopoietic developmental pathway, independent of endothelial differentiation. Page 62
79 CD105 CD117 SS DAPI # of cells A. FSC FSC CD41, CD45, CD309, H-2K k CD202b CD201 Page 63

80 CD4 B. OP9 shoxb4-op9 CD8 Figure 3.6 Both phl+201 progenitors derived from ESC/OP9 and ESC/sHoxB4-OP9 co-cultures have T-cell potential. A. Shows the definition of a phl+201 cell as FACS strategies for phl+201. B. Shows the successful differentiation of phl+201 progenitors into T lineage cells in vitro as shown by T-cell associated surface marker expression of typical T-cell associated markers CD4 and CD8. phl+201, prehematopoietic lineage commitment progenitor expressing CD201. Page 64







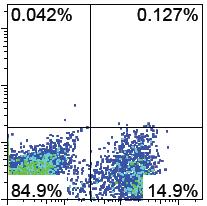
81 CD31 OP9 shoxb4-op9 A. Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 CD45 Page 65
82 % CD31 + cells B. OP9 shoxb4-op9 Figure 3.7 The role shoxb4-op9 plays on endothelial development was assayed by the phenotypic characterization of the development of CD31 + cells. A. Shows the development of CD31 + cells with respect to CD45 + cells in either ESC/OP9 or ESC/sHoxB4-OP9 co-cultures. B. Shows the aggregate data of triplicated experiments. Page 66




83 Day 4+5 Day 4+2 A. OP9 shoxb4-op9 Page 67
 and subsequently cultured in Matrigel")
84 # of endothelial cell colonies B. P= OP9 shoxb4-op9 Figure 3.8 The role of shoxb4-op9 in endothelial development was assayed by colony forming assays in MG cultures. CD309 + progenitors derived from either ESC/OP9 or ESC/sHoxB4-OP9 co-cultures were isolated at day 4 of co-culture by magnetic activated cell sorting (MACS) and subsequently cultured in Matrigel media. A. Shows the development of endothelial cell colonies on MG culture at day 2 and 5 of culture. B. Shows the number of endothelial cell colonies observed. There was no statistical significance between progenitors derived from co-cultures with or without shoxb4. Page 68
Modeling Developmental Hematopoiesis Using Pluripotent Stem Cells
 Modeling Developmental Hematopoiesis Using Pluripotent Stem Cells Christopher Sturgeon February 14, 2017 Pluripotent Stem Cells self-renewal hpsc Mesoderm blood cardiovascular muscle Endoderm lung liver
Modeling Developmental Hematopoiesis Using Pluripotent Stem Cells Christopher Sturgeon February 14, 2017 Pluripotent Stem Cells self-renewal hpsc Mesoderm blood cardiovascular muscle Endoderm lung liver
Haematopoietic stem cells
 Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Meeting Report. From December 8 to 11, 2012 at Atlanta, GA, U.S.A
 Meeting Report Affiliation Department of Transfusion Medicine and Cell Therapy Name Hisayuki Yao Name of the meeting Period and venue Type of your presentation Title of your presentation The 54 th Annual
Meeting Report Affiliation Department of Transfusion Medicine and Cell Therapy Name Hisayuki Yao Name of the meeting Period and venue Type of your presentation Title of your presentation The 54 th Annual
Hematopoiesis. - Process of generation of mature blood cells. - Daily turnover of blood cells (70 kg human)
") Hematopoiesis - Process of generation of mature blood cells - Daily turnover of blood cells (70 kg human) 1,000,000,000,000 total cells 200,000,000,000 red blood cells 70,000,000,000 neutrophils Hematopoiesis
Hematopoiesis - Process of generation of mature blood cells - Daily turnover of blood cells (70 kg human) 1,000,000,000,000 total cells 200,000,000,000 red blood cells 70,000,000,000 neutrophils Hematopoiesis
Stem cells: units of development and regeneration. Fernando D. Camargo Ph.D. Whitehead Fellow Whitehead Institute for Biomedical Research.
 Stem cells: units of development and regeneration Fernando D. Camargo Ph.D. Whitehead Fellow Whitehead Institute for Biomedical Research Concepts 1. Embryonic vs. adult stem cells 2. Hematopoietic stem
Stem cells: units of development and regeneration Fernando D. Camargo Ph.D. Whitehead Fellow Whitehead Institute for Biomedical Research Concepts 1. Embryonic vs. adult stem cells 2. Hematopoietic stem
The first wave of B lymphopoiesis develops independent of stem cells in the murine
 The first wave of B lymphopoiesis develops independent of stem cells in the murine embryo Short title: HSC-independent B-1 cell lineage Momoko Yoshimoto., MD., PhD Wells Center for Pediatric Research,
The first wave of B lymphopoiesis develops independent of stem cells in the murine embryo Short title: HSC-independent B-1 cell lineage Momoko Yoshimoto., MD., PhD Wells Center for Pediatric Research,
PhD THESIS Epigenetic mechanisms involved in stem cell differentiation
 Romanian Academy Institute of Cellular Biology and Pathology "Nicolae Simionescu" PhD THESIS Epigenetic mechanisms involved in stem cell differentiation Coordinator: Acad. Maya Simionescu PhD Student:
Romanian Academy Institute of Cellular Biology and Pathology "Nicolae Simionescu" PhD THESIS Epigenetic mechanisms involved in stem cell differentiation Coordinator: Acad. Maya Simionescu PhD Student:
Notch signalling pathway in murine embryonic stem cell derived haematopoiesis CAOXIN HUANG
 This thesis has been submitted in fulfilment of the requirements for a postgraduate degree (e.g. PhD, MPhil, DClinPsychol) at the University of Edinburgh. Please note the following terms and conditions
This thesis has been submitted in fulfilment of the requirements for a postgraduate degree (e.g. PhD, MPhil, DClinPsychol) at the University of Edinburgh. Please note the following terms and conditions
Bone Marrow Stroma in Myelodysplastic Syndromes
 Bone Marrow Stroma in Myelodysplastic Syndromes Universidad de Salamanca Prof Mª M Consuelo del Cañizo Hematology Dept. University Hospital, Salamanca SPAIN Bone marrow stroma in MDS Introduction Mesenchymal
Bone Marrow Stroma in Myelodysplastic Syndromes Universidad de Salamanca Prof Mª M Consuelo del Cañizo Hematology Dept. University Hospital, Salamanca SPAIN Bone marrow stroma in MDS Introduction Mesenchymal
Hematopoiesis. BHS Liège 27/1/2012. Dr Sonet Anne UCL Mont-Godinne
 Hematopoiesis BHS Liège 27/1/2012 Dr Sonet Anne UCL Mont-Godinne Hematopoiesis: definition = all the phenomenons to produce blood cells Leukocytes = White Blood Cells Polynuclear = Granulocytes Platelet
Hematopoiesis BHS Liège 27/1/2012 Dr Sonet Anne UCL Mont-Godinne Hematopoiesis: definition = all the phenomenons to produce blood cells Leukocytes = White Blood Cells Polynuclear = Granulocytes Platelet
Nature Immunology: doi: /ni Supplementary Figure 1. Huwe1 has high expression in HSCs and is necessary for quiescence.
 Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
Supplementary Table S1. Primers used for quantitative real-time polymerase chain reaction. Marker Sequence (5 3 ) Accession No.
 Accession No.") Supplementary Tables Supplementary Table S1. Primers used for quantitative real-time polymerase chain reaction Marker Sequence (5 3 ) Accession No. Angiopoietin 1, ANGPT1 A CCCTCCGGTGAATATTGGCTGG NM_001146.3
Supplementary Tables Supplementary Table S1. Primers used for quantitative real-time polymerase chain reaction Marker Sequence (5 3 ) Accession No. Angiopoietin 1, ANGPT1 A CCCTCCGGTGAATATTGGCTGG NM_001146.3
Nature Immunology: doi: /ni.3412
 Supplementary Figure 1 Gata1 expression in heamatopoietic stem and progenitor populations. (a) Unsupervised clustering according to 100 top variable genes across single pre-gm cells. The two main cell
Supplementary Figure 1 Gata1 expression in heamatopoietic stem and progenitor populations. (a) Unsupervised clustering according to 100 top variable genes across single pre-gm cells. The two main cell
Cytokines, adhesion molecules and apoptosis markers. A comprehensive product line for human and veterinary ELISAs
 Cytokines, adhesion molecules and apoptosis markers A comprehensive product line for human and veterinary ELISAs IBL International s cytokine product line... is extremely comprehensive. The assays are
Cytokines, adhesion molecules and apoptosis markers A comprehensive product line for human and veterinary ELISAs IBL International s cytokine product line... is extremely comprehensive. The assays are
BCR-ABL - LSK BCR-ABL + LKS - (%)
") Marker Clone BCR-ABL + LSK (%) BCR-ABL + LKS - (%) BCR-ABL - LSK (%) P value vs. BCR-ABL + LKS - P value vs. BCR-ABL - LSK CD2 RM2-5 12.9 ± 3.6 36.7 ± 6.5 19.3 ± 2.4 0.01 0.10 CD5 53-7.3 13.9 ± 3.2 20.8
Marker Clone BCR-ABL + LSK (%) BCR-ABL + LKS - (%) BCR-ABL - LSK (%) P value vs. BCR-ABL + LKS - P value vs. BCR-ABL - LSK CD2 RM2-5 12.9 ± 3.6 36.7 ± 6.5 19.3 ± 2.4 0.01 0.10 CD5 53-7.3 13.9 ± 3.2 20.8
CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow
 White Paper September 2016 CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow Lily C. Trajman, PhD Introduction: Hematopoietic Stem Cells (HSCs)
White Paper September 2016 CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow Lily C. Trajman, PhD Introduction: Hematopoietic Stem Cells (HSCs)
Hematopoiesis/Hematopoiesis Physiology
 Hematopoiesis/Hematopoiesis Physiology Definitions Hematopoiesis is the process of continuous generation of mature blood cells in the bone marrow (Figure 1). Blood cells represent different kinds of mature
Hematopoiesis/Hematopoiesis Physiology Definitions Hematopoiesis is the process of continuous generation of mature blood cells in the bone marrow (Figure 1). Blood cells represent different kinds of mature
Getting to the root of Cancer
 Cancer Stem Cells: Getting to the root of Cancer Dominique Bonnet, Ph.D Senior Group Leader, Haematopoietic Stem Cell Laboratory Cancer Research UK, London Research Institute Venice, Sept 2009 Overview
Cancer Stem Cells: Getting to the root of Cancer Dominique Bonnet, Ph.D Senior Group Leader, Haematopoietic Stem Cell Laboratory Cancer Research UK, London Research Institute Venice, Sept 2009 Overview
Scientific report: Delineating cellular stages and regulation of human NK cell development to improve NK cell-based therapy for cancer (Dnr )
") Scientific report: Delineating cellular stages and regulation of human NK cell development to improve NK cell-based therapy for cancer (Dnr 130259) The main goal of this project focuses on establishing
Scientific report: Delineating cellular stages and regulation of human NK cell development to improve NK cell-based therapy for cancer (Dnr 130259) The main goal of this project focuses on establishing
CD90 + Human Dermal Stromal Cells Are Potent Inducers of FoxP3 + Regulatory T Cells
 CD90 + Human Dermal Stromal Cells Are Potent Inducers of FoxP3 + Regulatory T Cells Karin Pfisterer, Karoline M Lipnik, Erhard Hofer and Adelheid Elbe-Bürger Journal of Investigative Dermatology (2015)
CD90 + Human Dermal Stromal Cells Are Potent Inducers of FoxP3 + Regulatory T Cells Karin Pfisterer, Karoline M Lipnik, Erhard Hofer and Adelheid Elbe-Bürger Journal of Investigative Dermatology (2015)
sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5
 sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
CRISPR-mediated Editing of Hematopoietic Stem Cells for the Treatment of β-hemoglobinopathies
 CRISPR-mediated Editing of Hematopoietic Stem Cells for the Treatment of β-hemoglobinopathies Jennifer Gori American Society of Gene & Cell Therapy May 11, 2017 editasmedicine.com 1 Highlights Developed
CRISPR-mediated Editing of Hematopoietic Stem Cells for the Treatment of β-hemoglobinopathies Jennifer Gori American Society of Gene & Cell Therapy May 11, 2017 editasmedicine.com 1 Highlights Developed
DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS
 DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS James Clinton, Ph.D. Scientist, ATCC February 19, 2015 About ATCC Founded in 1925, ATCC is a non-profit
DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS James Clinton, Ph.D. Scientist, ATCC February 19, 2015 About ATCC Founded in 1925, ATCC is a non-profit
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Stem cells are undifferentiated cells which are maintained within a specific niche. A stem cell
 Abstract Stem cells are undifferentiated cells which are maintained within a specific niche. A stem cell niche is a microenvironment of cells that maintain stem cell functionality, and one example is the
Abstract Stem cells are undifferentiated cells which are maintained within a specific niche. A stem cell niche is a microenvironment of cells that maintain stem cell functionality, and one example is the
The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D.
 The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D. OBJECTIVES 1. To understand how ordered Ig gene rearrangements lead to the development
The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D. OBJECTIVES 1. To understand how ordered Ig gene rearrangements lead to the development
Recommended reading: Abbas et al. 5th edition, chapters 7 and 10; Janeway and Travers, 5th edition, chapter 7.
 Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai 10/05/05; 11 AM Shiv Pillai T Lymphocyte Development Recommended reading:
Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai 10/05/05; 11 AM Shiv Pillai T Lymphocyte Development Recommended reading:
SUPPLEMENTARY INFORMATION doi: /nature12026
 doi:1.138/nature1226 a 4 35 3 MCSF level (pg/ml) 25 2 15 1 5 1h3 3h 5h 7h 15h 24h b MPP (CD135 KSL) HSC (CD34 CD15 KSLF) c % 4 ** LPS 3 GFP pos cells 2 PU.1 GFP LPS 1 FSCA Ctl NI 24h LPS Sup.Fig.1 Effect
doi:1.138/nature1226 a 4 35 3 MCSF level (pg/ml) 25 2 15 1 5 1h3 3h 5h 7h 15h 24h b MPP (CD135 KSL) HSC (CD34 CD15 KSLF) c % 4 ** LPS 3 GFP pos cells 2 PU.1 GFP LPS 1 FSCA Ctl NI 24h LPS Sup.Fig.1 Effect
Chronic variable stress activates hematopoietic stem cells
 SUPPLEMENTARY INFORMATION Chronic variable stress activates hematopoietic stem cells Timo Heidt *, Hendrik B. Sager *, Gabriel Courties, Partha Dutta, Yoshiko Iwamoto, Alex Zaltsman, Constantin von zur
SUPPLEMENTARY INFORMATION Chronic variable stress activates hematopoietic stem cells Timo Heidt *, Hendrik B. Sager *, Gabriel Courties, Partha Dutta, Yoshiko Iwamoto, Alex Zaltsman, Constantin von zur
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
Hepatogenesis I Liver development
 Hepatogenesis I Liver development HB 308 George Yeoh Room 2.59 MCS Building yeoh@cyllene.uwa.edu.au Topics Early liver development Tissue interaction - role of morphogens and cytokines Liver enriched transcription
Hepatogenesis I Liver development HB 308 George Yeoh Room 2.59 MCS Building yeoh@cyllene.uwa.edu.au Topics Early liver development Tissue interaction - role of morphogens and cytokines Liver enriched transcription
The Role of the Embryonic Microenvironment in Hematopoietic Cell Development
 The Role of the Embryonic Microenvironment in Hematopoietic Cell Development The Role of the Embryonic Microenvironment in Hematopoietic Cell Development De rol van de embryonale micro-omgeving op de
The Role of the Embryonic Microenvironment in Hematopoietic Cell Development The Role of the Embryonic Microenvironment in Hematopoietic Cell Development De rol van de embryonale micro-omgeving op de
Chapter 7 Conclusions
 VII-1 Chapter 7 Conclusions VII-2 The development of cell-based therapies ranging from well-established practices such as bone marrow transplant to next-generation strategies such as adoptive T-cell therapy
VII-1 Chapter 7 Conclusions VII-2 The development of cell-based therapies ranging from well-established practices such as bone marrow transplant to next-generation strategies such as adoptive T-cell therapy
Development of the hematopoietic system in the mouse
 Experimental Hematology 27 (1999) 777 787 Development of the hematopoietic system in the mouse Gordon Keller a,b, Georges Lacaud a, and Scott Robertson a a National Jewish Medical and Research Center,
Experimental Hematology 27 (1999) 777 787 Development of the hematopoietic system in the mouse Gordon Keller a,b, Georges Lacaud a, and Scott Robertson a a National Jewish Medical and Research Center,
EML Erythroid and Neutrophil Differentiation Protocols Cristina Pina 1*, Cristina Fugazza 2 and Tariq Enver 3
 EML Erythroid and Neutrophil Differentiation Protocols Cristina Pina 1*, Cristina Fugazza 2 and Tariq Enver 3 1 Department of Haematology, University of Cambridge, Cambridge, UK; 2 Dipartimento de Biotecnologie
EML Erythroid and Neutrophil Differentiation Protocols Cristina Pina 1*, Cristina Fugazza 2 and Tariq Enver 3 1 Department of Haematology, University of Cambridge, Cambridge, UK; 2 Dipartimento de Biotecnologie
Early cell death (FGF) B No RunX transcription factor produced Yes No differentiation
 B No RunX transcription factor produced Yes No differentiation") Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11095 Supplementary Table 1. Summary of the binding between Angptls and various Igdomain containing receptors as determined by flow cytometry analysis. The results were summarized from
doi:10.1038/nature11095 Supplementary Table 1. Summary of the binding between Angptls and various Igdomain containing receptors as determined by flow cytometry analysis. The results were summarized from
Transcript-indexed ATAC-seq for immune profiling
 Transcript-indexed ATAC-seq for immune profiling Technical Journal Club 22 nd of May 2018 Christina Müller Nature Methods, Vol.10 No.12, 2013 Nature Biotechnology, Vol.32 No.7, 2014 Nature Medicine, Vol.24,
Transcript-indexed ATAC-seq for immune profiling Technical Journal Club 22 nd of May 2018 Christina Müller Nature Methods, Vol.10 No.12, 2013 Nature Biotechnology, Vol.32 No.7, 2014 Nature Medicine, Vol.24,
SUPPLEMENTARY INFORMATION
 a. Smo+/+ b. Smo+/+ 5.63 5.48 c. Lin- d. e. 6 5 4 3 Ter119 Mac B T Sca1 Smo+/+ 25 15 2 o BMT 2 1 5 * Supplementary Figure 1: Deletion of Smoothened does not alter the frequency of hematopoietic lineages
a. Smo+/+ b. Smo+/+ 5.63 5.48 c. Lin- d. e. 6 5 4 3 Ter119 Mac B T Sca1 Smo+/+ 25 15 2 o BMT 2 1 5 * Supplementary Figure 1: Deletion of Smoothened does not alter the frequency of hematopoietic lineages
Supplementary Material
 Supplementary Material Taghon, Yui, and Rothenberg: Mast cell diversion of T-lineage precursor cells by the essential T-lineage transcription factor GATA Supplemental Table Supplemental Figures 1-6 Supplemental
Supplementary Material Taghon, Yui, and Rothenberg: Mast cell diversion of T-lineage precursor cells by the essential T-lineage transcription factor GATA Supplemental Table Supplemental Figures 1-6 Supplemental
International Graduate Research Programme in Cardiovascular Science
 1 International Graduate Research Programme in Cardiovascular Science This work has been supported by the European Community s Sixth Framework Programme under grant agreement n LSHM-CT-2005-01883 EUGeneHeart.
1 International Graduate Research Programme in Cardiovascular Science This work has been supported by the European Community s Sixth Framework Programme under grant agreement n LSHM-CT-2005-01883 EUGeneHeart.
Hematopoiesis. Hematopoiesis. Hematopoiesis
 Chapter. Cells and Organs of the Immune System Hematopoiesis Hematopoiesis- formation and development of WBC and RBC bone marrow. Hematopoietic stem cell- give rise to any blood cells (constant number,
Chapter. Cells and Organs of the Immune System Hematopoiesis Hematopoiesis- formation and development of WBC and RBC bone marrow. Hematopoietic stem cell- give rise to any blood cells (constant number,
TITLE: Effects of Hematopoietic Stem Cell Age on CML Disease Progression
 AD Award Number: W81XWH-04-1-0795 TITLE: Effects of Hematopoietic Stem Cell Age on CML Disease Progression PRINCIPAL INVESTIGATOR: Kenneth Dorshkind, Ph.D. CONTRACTING ORGANIZATION: University of California,
AD Award Number: W81XWH-04-1-0795 TITLE: Effects of Hematopoietic Stem Cell Age on CML Disease Progression PRINCIPAL INVESTIGATOR: Kenneth Dorshkind, Ph.D. CONTRACTING ORGANIZATION: University of California,
SEVENTH EDITION CHAPTER
 Judy Owen Jenni Punt Sharon Stranford Kuby Immunology SEVENTH EDITION CHAPTER 16 Tolerance, Autoimmunity, and Transplantation Copyright 2013 by W. H. Freeman and Company Immune tolerance: history * Some
Judy Owen Jenni Punt Sharon Stranford Kuby Immunology SEVENTH EDITION CHAPTER 16 Tolerance, Autoimmunity, and Transplantation Copyright 2013 by W. H. Freeman and Company Immune tolerance: history * Some
Lymphoid architecture & Leukocyte recirculation. Thursday Jan 26th, 2017
 Lymphoid architecture & Leukocyte recirculation Thursday Jan 26th, 2017 Topics The life of immune cells Where are they born? Where are they educated? Where do they function? How do they get there? The
Lymphoid architecture & Leukocyte recirculation Thursday Jan 26th, 2017 Topics The life of immune cells Where are they born? Where are they educated? Where do they function? How do they get there? The
T Cell Development. Xuefang Cao, MD, PhD. November 3, 2015
 T Cell Development Xuefang Cao, MD, PhD November 3, 2015 Thymocytes in the cortex of the thymus Early thymocytes development Positive and negative selection Lineage commitment Exit from the thymus and
T Cell Development Xuefang Cao, MD, PhD November 3, 2015 Thymocytes in the cortex of the thymus Early thymocytes development Positive and negative selection Lineage commitment Exit from the thymus and
Immunology - Lecture 2 Adaptive Immune System 1
 Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nture07679 Emryonic Stem (ES) cell Hemngiolst Flk1 + Blst Colony 3 to 3.5 Dys 3-4 Dys ES differentition Sort of Flk1 + cells Supplementry Figure 1. Chrcteristion of lst colony development.
doi: 10.1038/nture07679 Emryonic Stem (ES) cell Hemngiolst Flk1 + Blst Colony 3 to 3.5 Dys 3-4 Dys ES differentition Sort of Flk1 + cells Supplementry Figure 1. Chrcteristion of lst colony development.
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products
 stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
stem cell products Basement Membrane Matrix Products Rat Mesenchymal Stem Cell Growth and Differentiation Products Stem Cell Qualified Extracellular Matrix Proteins Stem cell research requires the finest
Supplementary Figure 1. Successful excision of genes from WBM lysates and
 Supplementary Information: Supplementary Figure 1. Successful excision of genes from WBM lysates and survival of mice with different genotypes. (a) The proper excision of Pten, p110α, p110α and p110δ was
Supplementary Information: Supplementary Figure 1. Successful excision of genes from WBM lysates and survival of mice with different genotypes. (a) The proper excision of Pten, p110α, p110α and p110δ was
Effector T Cells and
 1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
Highly Efficient CRISPR/Cas9 Gene Editing and Long-Term Engraftment of Human Hematopoietic Stem and Progenitor Cells
 Highly Efficient CRISPR/Cas9 Gene Editing and Long-Term Engraftment of Human Hematopoietic Stem and Progenitor Cells J. M. Heath, A. Chalishazar, C.S. Lee, W. Selleck, C. Cotta-Ramusino, D. Bumcrot, J.L.
Highly Efficient CRISPR/Cas9 Gene Editing and Long-Term Engraftment of Human Hematopoietic Stem and Progenitor Cells J. M. Heath, A. Chalishazar, C.S. Lee, W. Selleck, C. Cotta-Ramusino, D. Bumcrot, J.L.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
X P. Supplementary Figure 1. Nature Medicine: doi: /nm Nilotinib LSK LT-HSC. Cytoplasm. Cytoplasm. Nucleus. Nucleus
 a b c Supplementary Figure 1 c-kit-apc-eflu780 Lin-FITC Flt3-Linc-Kit-APC-eflu780 LSK Sca-1-PE-Cy7 d e f CD48-APC LT-HSC CD150-PerCP-cy5.5 g h i j Cytoplasm RCC1 X Exp 5 mir 126 SPRED1 SPRED1 RAN P SPRED1
a b c Supplementary Figure 1 c-kit-apc-eflu780 Lin-FITC Flt3-Linc-Kit-APC-eflu780 LSK Sca-1-PE-Cy7 d e f CD48-APC LT-HSC CD150-PerCP-cy5.5 g h i j Cytoplasm RCC1 X Exp 5 mir 126 SPRED1 SPRED1 RAN P SPRED1
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti.
 Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti. Chapter 1: Overview of Immunology Prepared by David Scott, MD, Scripps
Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti. Chapter 1: Overview of Immunology Prepared by David Scott, MD, Scripps
Edinburgh Research Explorer
 Edinburgh Research Explorer Blood-forming endothelium in human ontogeny: Lessons from in utero development and embryonic stem cell culture Citation for published version: Zarnbidis, ET, Oberlin, E, Tavian,
Edinburgh Research Explorer Blood-forming endothelium in human ontogeny: Lessons from in utero development and embryonic stem cell culture Citation for published version: Zarnbidis, ET, Oberlin, E, Tavian,
Supplement Material. Spleen weight (mg) LN cells (X106) Acat1-/- Acat1-/- Mouse weight (g)
 LN cells (X106) Acat1-/- Acat1-/- Mouse weight (g)") Supplement Material A Spleen weight (mg) C Mouse weight (g) 1 5 1 2 9 6 3 2 5 2 1 5 Male LN cells (X16) 4 ** ** Female B 3 2 1 Supplemental Figure I. Spleen weight (A), Inguinal lymph node (LN) cell number
Supplement Material A Spleen weight (mg) C Mouse weight (g) 1 5 1 2 9 6 3 2 5 2 1 5 Male LN cells (X16) 4 ** ** Female B 3 2 1 Supplemental Figure I. Spleen weight (A), Inguinal lymph node (LN) cell number
NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression
 Manuscript EMBO-2010-75657 NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression Masaki Kashiwada, Suzanne L. Cassel, John D. Colgan and Paul B. Rothman Corresponding author: Paul Rothman, University
Manuscript EMBO-2010-75657 NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression Masaki Kashiwada, Suzanne L. Cassel, John D. Colgan and Paul B. Rothman Corresponding author: Paul Rothman, University
Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features
 Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features Loretta Gammaitoni, Lidia Giraudo, Valeria Leuci, et al. Clin Cancer Res
Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features Loretta Gammaitoni, Lidia Giraudo, Valeria Leuci, et al. Clin Cancer Res
T. R. Golub, D. K. Slonim & Others 1999
 T. R. Golub, D. K. Slonim & Others 1999 Big Picture in 1999 The Need for Cancer Classification Cancer classification very important for advances in cancer treatment. Cancers of Identical grade can have
T. R. Golub, D. K. Slonim & Others 1999 Big Picture in 1999 The Need for Cancer Classification Cancer classification very important for advances in cancer treatment. Cancers of Identical grade can have
Of hemangioblast, hemogenic endothelium and primitive versus definitive hematopoiesis
 Of hemangioblast, hemogenic endothelium and primitive versus definitive hematopoiesis Georges Lacaud 1* and Valerie Kouskoff 2* 1 Stem Cell Biology Group, Cancer Research UK Manchester Institute, The University
Of hemangioblast, hemogenic endothelium and primitive versus definitive hematopoiesis Georges Lacaud 1* and Valerie Kouskoff 2* 1 Stem Cell Biology Group, Cancer Research UK Manchester Institute, The University
ChipSeq. Technique and science. The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation
 Center for Biomics ChipSeq Technique and science The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation Wilfred van IJcken Sequencing Seminar Illumina September 8 Scheme
Center for Biomics ChipSeq Technique and science The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation Wilfred van IJcken Sequencing Seminar Illumina September 8 Scheme
Chapter 2 (pages 22 33): Cells and Tissues of the Immune System. Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Cells and Tissues of the Immune System. Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") Allergy and Immunology Review Corner: Cellular and Molecular Immunology, 8th Edition By Abul K. Abbas, MBBS; Andrew H. H. Lichtman, MD, PhD; and Shiv Pillai, MBBS, PhD. Chapter 2 (pages 22 33): Cells and
Allergy and Immunology Review Corner: Cellular and Molecular Immunology, 8th Edition By Abul K. Abbas, MBBS; Andrew H. H. Lichtman, MD, PhD; and Shiv Pillai, MBBS, PhD. Chapter 2 (pages 22 33): Cells and
12 Dynamic Interactions between Hematopoietic Stem and Progenitor Cells and the Bone Marrow: Current Biology of Stem Cell Homing and Mobilization
 Table of Contents: PART I: Molecular and Cellular Basis of Hematology 1 Anatomy and Pathophysiology of the Gene 2 Genomic Approaches to Hematology 3 Regulation of Gene Expression, Transcription, Splicing,
Table of Contents: PART I: Molecular and Cellular Basis of Hematology 1 Anatomy and Pathophysiology of the Gene 2 Genomic Approaches to Hematology 3 Regulation of Gene Expression, Transcription, Splicing,
RUNX1 and FPD/AML Translational Research. The Leukemia and Lymphoma Society / Babich Family Foundation Partnership. September 2016
 www.lls.org www.runx1.com RUNX1 and FPD/AML Translational Research The Leukemia and Lymphoma Society / Babich Family Foundation Partnership September 2016 Prepared by L. Greenberger, PhD Chief Scientific
www.lls.org www.runx1.com RUNX1 and FPD/AML Translational Research The Leukemia and Lymphoma Society / Babich Family Foundation Partnership September 2016 Prepared by L. Greenberger, PhD Chief Scientific
Regenerative Medicine for Cardiomyocytes
 Regenerative Medicine Regenerative Medicine for JMAJ 47(7): 328 332, 2004 Keiichi FUKUDA Assistant Professor, Institute for Advanced Cardiac Therapeutics, Keio University School of Medicine Abstract: Heart
Regenerative Medicine Regenerative Medicine for JMAJ 47(7): 328 332, 2004 Keiichi FUKUDA Assistant Professor, Institute for Advanced Cardiac Therapeutics, Keio University School of Medicine Abstract: Heart
MACROPHAGE "MONOCYTES" SURFACE RECEPTORS
 LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
4. TEXTBOOK: ABUL K. ABBAS. ANDREW H. LICHTMAN. CELLULAR AND MOLECULAR IMMUNOLOGY. 5 TH EDITION. Chapter 2. pg
 LECTURE: 03 Title: CELLS INVOLVED IN THE IMMUNE RESPONSE LEARNING OBJECTIVES: The student should be able to: Identify the organs where the process of the blood formation occurs. Identify the main cell
LECTURE: 03 Title: CELLS INVOLVED IN THE IMMUNE RESPONSE LEARNING OBJECTIVES: The student should be able to: Identify the organs where the process of the blood formation occurs. Identify the main cell
The Role of Rac Signaling in The Perivascular Niche
 The Role of Rac Signaling in The Perivascular Niche Felicia Ciuculescu Diaspora and Higher Education and Research Perspectives in Personalized Medicine- from Concept to Clinical Application Center for
The Role of Rac Signaling in The Perivascular Niche Felicia Ciuculescu Diaspora and Higher Education and Research Perspectives in Personalized Medicine- from Concept to Clinical Application Center for
Nature Immunology: doi: /ni Supplementary Figure 1. Characteristics of SEs in T reg and T conv cells.
 Supplementary Figure 1 Characteristics of SEs in T reg and T conv cells. (a) Patterns of indicated transcription factor-binding at SEs and surrounding regions in T reg and T conv cells. Average normalized
Supplementary Figure 1 Characteristics of SEs in T reg and T conv cells. (a) Patterns of indicated transcription factor-binding at SEs and surrounding regions in T reg and T conv cells. Average normalized
Frontiers of Science Foundation Scholar. Toby Bothwell. Freshman, University of Arkansas. final research manuscripts
 Frontiers of Science Foundation Scholar Toby Bothwell Oklahoma city, OKlahoma Freshman, University of Arkansas final research manuscripts The Effect of Estradiol and Estrogen Receptors α and β on Bone
Frontiers of Science Foundation Scholar Toby Bothwell Oklahoma city, OKlahoma Freshman, University of Arkansas final research manuscripts The Effect of Estradiol and Estrogen Receptors α and β on Bone
September 20, Submitted electronically to: Cc: To Whom It May Concern:
 History Study (NOT-HL-12-147), p. 1 September 20, 2012 Re: Request for Information (RFI): Building a National Resource to Study Myelodysplastic Syndromes (MDS) The MDS Cohort Natural History Study (NOT-HL-12-147).
History Study (NOT-HL-12-147), p. 1 September 20, 2012 Re: Request for Information (RFI): Building a National Resource to Study Myelodysplastic Syndromes (MDS) The MDS Cohort Natural History Study (NOT-HL-12-147).
Mariusz Z. Ratajczak M.D., Ph.D., d.hc. Stem Cell Institute at the James Graham Brown Cancer Center, University of Louisville.
 Umbilical cord blood-derived CD45 - /SSEA-4 + /OCT-4 + /CD133 + /CXCR4 + /Lin - very small embryonic/epiblast like stem cells (VSELs) Potential Clinical Applications Mariusz Z. Ratajczak M.D., Ph.D., d.hc.
Umbilical cord blood-derived CD45 - /SSEA-4 + /OCT-4 + /CD133 + /CXCR4 + /Lin - very small embryonic/epiblast like stem cells (VSELs) Potential Clinical Applications Mariusz Z. Ratajczak M.D., Ph.D., d.hc.
Can we classify cancer using cell signaling?
 Can we classify cancer using cell signaling? Central hypotheses (big ideas) Alterations to signaling genes would cause leukemic cells to react in an inappropriate or sensitized manner to environmental
Can we classify cancer using cell signaling? Central hypotheses (big ideas) Alterations to signaling genes would cause leukemic cells to react in an inappropriate or sensitized manner to environmental
A dissertation by. Clare Rachel Watsford
 Young People s Expectations, Preferences and Experiences of Seeking Help from a Youth Mental Health Service and the Effects on Clinical Outcome, Service Use and Future Help-Seeking Intentions A dissertation
Young People s Expectations, Preferences and Experiences of Seeking Help from a Youth Mental Health Service and the Effects on Clinical Outcome, Service Use and Future Help-Seeking Intentions A dissertation
Accelerate Your Research with Conversant Bio
 Accelerate Your Research with Conversant Bio 400+ Participating MDs 50+ Partner sites for tissue procurement Continuous expansion of sourcing capabilities Closely monitored chain of custody Full regulatory
Accelerate Your Research with Conversant Bio 400+ Participating MDs 50+ Partner sites for tissue procurement Continuous expansion of sourcing capabilities Closely monitored chain of custody Full regulatory
Cell signalling pathways in the HSC niche. Dr. Abdullah Aljedai
 Cell signalling pathways in the HSC niche Dr. Abdullah Aljedai 31-10-2009 Learning objectives & resources 1- To introduce the concept of cell signaling process in the haemopoietic system. 2- To outline
Cell signalling pathways in the HSC niche Dr. Abdullah Aljedai 31-10-2009 Learning objectives & resources 1- To introduce the concept of cell signaling process in the haemopoietic system. 2- To outline
Stem cells and Cancer. John Glod. December 2, 2009
 Stem cells and Cancer John Glod Lehigh University Lehigh University December 2, 2009 The Tumor Microenvironment Littlepage et al Cancer Cell 2005 Cancer Stem Cells A small group of cells within the larger
Stem cells and Cancer John Glod Lehigh University Lehigh University December 2, 2009 The Tumor Microenvironment Littlepage et al Cancer Cell 2005 Cancer Stem Cells A small group of cells within the larger
TITLE: Properties of Leukemia Stem Cells in a Novel Model of CML Progression to Lymphoid Blast Crisis
 AD Award Number: W81XWH-05-1-0608 TITLE: Properties of Leukemia Stem Cells in a Novel Model of CML Progression to Lymphoid Blast Crisis PRINCIPAL INVESTIGATOR: Craig T. Jordan, Ph.D. CONTRACTING ORGANIZATION:
AD Award Number: W81XWH-05-1-0608 TITLE: Properties of Leukemia Stem Cells in a Novel Model of CML Progression to Lymphoid Blast Crisis PRINCIPAL INVESTIGATOR: Craig T. Jordan, Ph.D. CONTRACTING ORGANIZATION:
Stem Cells. Induced Stem Cells
 Induced Stem Cells Stem Cells Mouse and human somatic cells can either be reprogrammed to a pluripotent state or converted to another lineage with a combination of transcription factors suggesting that
Induced Stem Cells Stem Cells Mouse and human somatic cells can either be reprogrammed to a pluripotent state or converted to another lineage with a combination of transcription factors suggesting that
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Heart 3: Organogenesis, CHD, prenatal circulation
 Heart 3: Organogenesis, CHD, prenatal circulation Heart development Development of vasculature Pathogenesis of CHD Prenatal circulation and its remodeling after birth David Sedmera Charles University First
Heart 3: Organogenesis, CHD, prenatal circulation Heart development Development of vasculature Pathogenesis of CHD Prenatal circulation and its remodeling after birth David Sedmera Charles University First
IMMUNE CELL SURFACE RECEPTORS AND THEIR FUNCTIONS
 LECTURE: 07 Title: IMMUNE CELL SURFACE RECEPTORS AND THEIR FUNCTIONS LEARNING OBJECTIVES: The student should be able to: The chemical nature of the cellular surface receptors. Define the location of the
LECTURE: 07 Title: IMMUNE CELL SURFACE RECEPTORS AND THEIR FUNCTIONS LEARNING OBJECTIVES: The student should be able to: The chemical nature of the cellular surface receptors. Define the location of the
THE HYPOXIC HEMATOPOIETIC STEM CELL NICHE Consequences of Hypoxiainduced Transcription on Stem Cell Fate
 THE HYPOXIC HEMATOPOIETIC STEM CELL NICHE Consequences of Hypoxiainduced Transcription on Stem Cell Fate Rehn, Matilda Published: 2011-01-01 Link to publication Citation for published version (APA): Rehn,
THE HYPOXIC HEMATOPOIETIC STEM CELL NICHE Consequences of Hypoxiainduced Transcription on Stem Cell Fate Rehn, Matilda Published: 2011-01-01 Link to publication Citation for published version (APA): Rehn,
DISCLOSURE. I have the following financial relationships:
 DISCLOSURE I have the following financial relationships: Consultant for: Fate Therapeutics, GlaxoSmithKline, Bone Therapeutics, G1 Therapeutics Contracted Research for: GlaxoSmithKline Royalties from:
DISCLOSURE I have the following financial relationships: Consultant for: Fate Therapeutics, GlaxoSmithKline, Bone Therapeutics, G1 Therapeutics Contracted Research for: GlaxoSmithKline Royalties from:
MHC class I MHC class II Structure of MHC antigens:
 MHC class I MHC class II Structure of MHC antigens: MHC class I antigens consist of a transmembrane heavy chain (α chain) that is non-covalently associated with β2- microglobulin. Membrane proximal domain
MHC class I MHC class II Structure of MHC antigens: MHC class I antigens consist of a transmembrane heavy chain (α chain) that is non-covalently associated with β2- microglobulin. Membrane proximal domain
The role of DNMT3A and HOXA9 hypomethylation in acute myeloid leukemia (AML)
") Campbell Drohan BIOL 463 December 2016 The role of DNMT3A and HOXA9 hypomethylation in acute myeloid leukemia (AML) Introduction Epigenetic modifications of DNA and histones are key to gene regulation
Campbell Drohan BIOL 463 December 2016 The role of DNMT3A and HOXA9 hypomethylation in acute myeloid leukemia (AML) Introduction Epigenetic modifications of DNA and histones are key to gene regulation
Understanding the regulation of vertebrate hematopoiesis and blood disorders big lessons from a small fish
 REVIEW ARTICLE Understanding the regulation of vertebrate hematopoiesis and blood disorders big lessons from a small fish Anne L. Robertson 1, Serine Avagyan 2, John M. Gansner 3 and Leonard I. Zon 4 1
REVIEW ARTICLE Understanding the regulation of vertebrate hematopoiesis and blood disorders big lessons from a small fish Anne L. Robertson 1, Serine Avagyan 2, John M. Gansner 3 and Leonard I. Zon 4 1
The Adaptive Immune Response. T-cells
 The Adaptive Immune Response T-cells T Lymphocytes T lymphocytes develop from precursors in the thymus. Mature T cells are found in the blood, where they constitute 60% to 70% of lymphocytes, and in T-cell
The Adaptive Immune Response T-cells T Lymphocytes T lymphocytes develop from precursors in the thymus. Mature T cells are found in the blood, where they constitute 60% to 70% of lymphocytes, and in T-cell
ChIPSeq. Technique and science. The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation
 Center for Biomics ChIPSeq Technique and science The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation Wilfred van IJcken EU Sequencing Seminar Illumina June 16 Genomics
Center for Biomics ChIPSeq Technique and science The genome wide dynamics of the binding of ldb1 complexes during erythroid differentiation Wilfred van IJcken EU Sequencing Seminar Illumina June 16 Genomics
Role of oocyte-secreted factors in prevention of cumulus cell apoptosis and enhancement of oocyte developmental competence
 Role of oocyte-secreted factors in prevention of cumulus cell apoptosis and enhancement of oocyte developmental competence Tamer Hussein, MScMed Research Centre for Reproductive Health, Discipline of Obstetrics
Role of oocyte-secreted factors in prevention of cumulus cell apoptosis and enhancement of oocyte developmental competence Tamer Hussein, MScMed Research Centre for Reproductive Health, Discipline of Obstetrics
Chapter 6. Villous Growth
 Core Curriculum in Perinatal Pathology Chapter 6 Villous Growth Overview of vasculogenesis and angiogenesis Vasculogenesis Extraembryonic Vasculogenesis Angiogenesis Branching angiogenesis Sprouting angiogenesis
Core Curriculum in Perinatal Pathology Chapter 6 Villous Growth Overview of vasculogenesis and angiogenesis Vasculogenesis Extraembryonic Vasculogenesis Angiogenesis Branching angiogenesis Sprouting angiogenesis
Lost in translation: pluripotent stem cell-derived hematopoiesis
 Review Lost in translation: pluripotent stem cell-derived hematopoiesis Mania Ackermann 1,2,, Steffi Liebhaber 1,2,, Jan-Henning Klusmann 3 & Nico Lachmann 1,2,4,* Abstract Pluripotent stem cells (PSCs)
Review Lost in translation: pluripotent stem cell-derived hematopoiesis Mania Ackermann 1,2,, Steffi Liebhaber 1,2,, Jan-Henning Klusmann 3 & Nico Lachmann 1,2,4,* Abstract Pluripotent stem cells (PSCs)
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 AD Award Number: W81XWH-13-1-0057 TITLE: Role of TIRAP in Myelodysplastic Syndromes PRINCIPAL INVESTIGATOR: Linda Ya-ting Chang CONTRACTING ORGANIZATION: British Columbia Cancer Agency Branch Vancouver,
AD Award Number: W81XWH-13-1-0057 TITLE: Role of TIRAP in Myelodysplastic Syndromes PRINCIPAL INVESTIGATOR: Linda Ya-ting Chang CONTRACTING ORGANIZATION: British Columbia Cancer Agency Branch Vancouver,
Characterization of human myeloid progenitors and their differentiation
 Characterization of human myeloid progenitors and their differentiation Edvardsson, Louise 2006 Link to publication Citation for published version (APA): Edvardsson, L. (2006). Characterization of human
Characterization of human myeloid progenitors and their differentiation Edvardsson, Louise 2006 Link to publication Citation for published version (APA): Edvardsson, L. (2006). Characterization of human
ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1
 ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
Topics of this lecture : RBC. Structural characteristics Hemoglobin Erythropoiesis Erythrocytes destruction
 Topics of this lecture : RBC Structural characteristics Hemoglobin Erythropoiesis Erythrocytes destruction Structural characteristics Its small size and biconcave shape provides more surface area than
Topics of this lecture : RBC Structural characteristics Hemoglobin Erythropoiesis Erythrocytes destruction Structural characteristics Its small size and biconcave shape provides more surface area than