Human Papillomaviruses Recruit Cellular DNA Repair and Homologous Recombination Factors to Viral Replication Centers
|
|
|
- Cassandra Wilcox
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi: /jvi Copyright 2012, American Society for Microbiology. All Rights Reserved Human Papillomaviruses Recruit Cellular DNA Repair and Homologous Recombination Factors to Viral Replication Centers Running title: Recruitment of repair factors to HPV replication foci Kenric A. Gillespie 1, Kavi P. Mehta 2, Laimonis A. Laimins 2, and Cary A. Moody 1* Lineberger Comprehensive Cancer Center and Department of Microbiology & Immunology, University of North Carolina at Chapel Hill, North Carolina , Department of Microbiology and Immunology, Northwestern University, Chicago, IL * Corresponding author: Mailing address: Lineberger Comprehensive Cancer Center, CB#7295, University of North Carolina, Chapel Hill, NC Phone: (919) Fax:
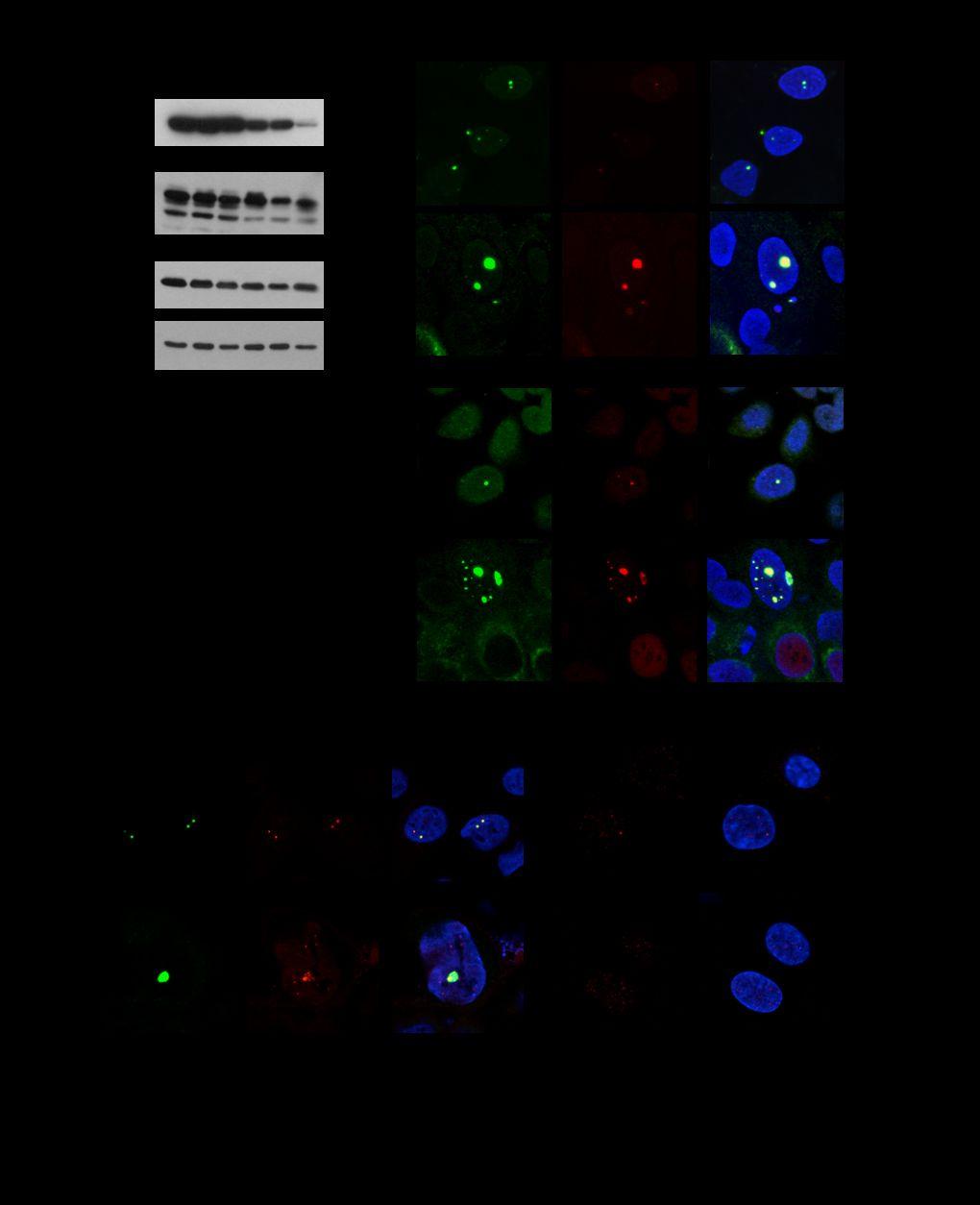
2 Abstract: Human papillomaviruses activate the ATM-dependent DNA damage response to induce viral genome amplification upon epithelial differentiation. Our studies show that along with members of the ATM pathway, HPV proteins also localize factors involved in homologous DNA recombination to distinct nuclear foci that contain HPV genomes and cellular replication factors. These studies indicate that HPV activates the ATM pathway to recruit repair factors to viral genomes and allow for efficient replication. Downloaded from on October 2, 2018 by guest 2
3 The life cycle of human papillomaviruses (HPV) is dependent on epithelial differentiation and is controlled through the action of viral as well as cellular factors (20). In undifferentiated cells, viral genomes are maintained as low copy episomes that replicate once per cell cycle along with cellular DNA. Upon differentiation, viral genomes are amplified to thousands of copies per cell along with induction of late gene expression and the assembly of progeny virions (20). While normal epithelial differentiation results in exit from the cell cycle, expression of the E6 and E7 proteins pushes a subset of differentiating cells into S or G2/M phases to induce genome amplification (26). E6 and E7 also activate the ATM DNA damage response that is necessary for the differentiation-dependent amplification of viral genomes (24). The DNA damage response (DDR) plays a crucial role in the maintenance of genomic stability by coordinating cell cycle progression with DNA repair. The DDR is regulated by two main kinases, ATM (Ataxia telangiectasia mutated) and ATR (ATM and Rad3-related) that belong to the phosphoinositide-3-kinase-related protein kinase family (PIKKs) (12). ATM responds primarily to double-strand breaks (DSBs), while ATR is activated in response to single stranded DNA at stalled replication forks. ATM and ATR phosphorylate multiple substrates in response to DNA damage, including proteins involved in cell cycle checkpoints, DNA repair and apoptosis (7, 21). The MRN complex, consisting of NBS1, Mre11 and Rad50, serves as the sensor for DSBs and recruits ATM to these sites, as well as promotes ATM activation through autophosphorylation (8, 17, 18). ATM activation leads to the phosphorylation of many substrates at sites of DNA damage, including Chk2, BRCA1 and NBS1, as well as the histone H2A variant H2AX (referred to as γh2ax) (5, 12). Previous studies demonstrated that high-risk HPV31 induces an ATM-dependent DNA damage response in both undifferentiated and differentiating cells, however, this activity is only required for genome amplification and not stable maintenance replication (24). In HPV positive cells, members of the ATM DNA damage pathway, including γh2ax, Chk2, BRCA1 and NBS1 are recruited in an ATM-dependent manner into distinct foci resembling those seen following DNA damage by 3
4 ionizing radiation. HPV genomes are also replicated at specific nuclear loci (24, 38), but it was unclear if these regions also contained activated members of the ATM pathway or were localized at distinctly separate locations To determine if DNA repair factors co-localize with HPV genomes we used immunofluorescence (IF) followed by Fluorescence In Situ Hybridization (FISH) to screen for HPV DNA. For these assays, we utilized the CIN 612 cell line, which stably maintains HPV31 genomes and was previously shown to activate the ATM-dependent DNA damage response (1, 24). CIN 612 cells were plated on glass coverslips and induced to differentiate in high calcium medium at approximately 90% confluency, or harvested as an undifferentiated sample (T0). For the localization assays, cells were first extracted prior to fixation using a cytoskeleton buffer containing Triton X-100, as described previously (15). This method removes proteins that are not bound tightly to chromatin and provides a clearer view of protein localization. Following extraction, cells were fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton X-100 in phosphate buffered saline and IF for DNA repair factors was performed. The samples were then cross-linked in methanol:acetic acid, and FISH for HPV DNA was performed using tyramide-enhanced fluorescence (Invitrogen), as described previously (15). IF/FISH was performed on detergent-extracted cells a minimum of two times for each DNA repair factor. The number of foci positive for both HPV DNA and each repair factor was quantified, with 25 to 40 FISH positive cells being counted for each experiment. Although all cells contain HPV genomes, only a subset of cells amplify viral DNA upon differentiation and are FISH positive (24). Consistent with previous reports (23, 24), incubation in high calcium media induces epithelial differentiation, as evidenced by the expression of differentiation-specific markers, such as involucrin and keratin 10 (K10) as well as viral genome amplification (data not shown). HPV DNA foci co-localize with markers of double strand DNA breaks. Previously, we found that HPV induces the formation of nuclear foci containing γh2ax, a target of ATM that is frequently used 101 as a marker of DNA breaks. p53 binding protein 1 (53BP1) is also an ATM target that is used to 4
5 identify sites of DNA damage. H2AX represents 2-20% of histone 2A and when phosphorylated marks the chromatin around DSBs (9, 14). 53BP1 is subsequently recruited to sites of DNA damage in response to histone ubiquitylation (2). Importantly, 53BP1 is necessary for the intra S and G2/M checkpoints (22, 35, 39). Western blot analysis of cell extracts indicated that both γh2ax and 53BP1 were present at increased levels in both undifferentiated and differentiated HPV positive cells compared to HPV negative HFKs (Figure 1A). To determine if γh2ax and 53BP1 localize to HPV DNA foci, CIN 612 cells were induced to differentiate in high calcium and examined by IF to detect γh2ax and 53BP1 and FISH to detect HPV DNA. As shown in Figure 1B, γh2ax, as well as 53BP1 (Figure 1C) were found localized to HPV DNA foci in both undifferentiated and differentiating cells. γh2ax was localized to % of cells containing HPV DNA foci in both undifferentiated and differentiated cells. Interestingly, 53BP1 was localized to 47-50% of undifferentiated cells that were FISH positive, and this increased to 88-93% upon differentiation. Consistent with viral genome amplification, the size of the foci containing γh2ax and 53BP1 increased in differentiating cells. Interestingly, upon differentiation we observed FISH positive cells containing either one or two large HPV DNA foci, or multiple smaller foci. As shown in Figure 1B, γh2ax localized to both foci types in detergent-extracted cells, and we observed similar results for 53BP1 as well as the other repair factors tested (data not shown). This increase in foci size coincides with increased replication of viral episomes as determined by Southern blot analyses (24). To ensure that episomal copies of the virus were not lost upon detergent extraction, resulting in only the detection of integrated HPV foci (15), we performed the IF/FISH assay without the pre-extraction step. In the absence of pre-extraction, we observed a similar distribution of cells containing large HPV DNA foci and multiple foci. In addition, the localization of γh2ax to HPV DNA foci was not affected (Figure 1B). These results indicate that DNA repair factors are recruited to episomal copies of HPV. It is possible that these large foci are the result of the coalescence of replication compartments, as has been shown for Herpes Simplex Virus (HSV) (6, 30, 31). In contrast to HPV positive cells, only faint focal staining of γh2ax and 53BP1 was observed in normal HFKs. Since both γh2ax and 53BP1 are important for the localization of cellular 5
6 factors involved in damage signaling and repair, these findings indicate that HPV promotes the recruitment of host repair factors to HPV genomes Phosphorylated ATM and Chk2 localize to HPV DNA foci. Since γh2ax and 53BP1 were localized to HPV DNA foci, we reasoned that ATM and Chk2 should also localize to these sites. patm and pchk2 are activated at higher levels in HPV positive cells compared to normal human foreskin keratinocytes (HFKs), and this is maintained upon differentiation (Figure 2A). To examine the possibility that these factors localize to HPV DNA foci, we performed IF/FISH on detergent-extracted CIN 612 cells. We observed that patm (55-60% of FISH positive cells) (Figure 2B) and Chk2 (83-90% of FISH positive cells) (Figure 2C) co-localized at sites containing HPV DNA in undifferentiated cells. Upon differentiation, the intensity of the signal and size of the foci increased, indicating that activated ATM and Chk2 are recruited to newly synthesized viral genomes. In differentiating cells, patm and Chk2 were localized to HPV DNA foci in 64-74% and 82-86% of FISH positive cells, respectively. Though we observed some Chk2 foci in the absence of HPV FISH foci (Figure 2C), we believe this reflects HPV s ability to induce a general DNA damage response as well as the limited sensitivity of the FISH assay (24). Normal HKFs exhibited only background staining of patm and Chk2 (data not shown), as demonstrated previously (24). HPV DNA foci represent compartments containing actively replicating genomes. To ensure that the HPV foci we observed by FISH represented active sites of viral replication, we next examined the levels and localization of the cellular replication proteins PCNA and RPA32 in relation to HPV DNA. As shown in Figure 3A, HPV positive cells exhibited higher levels of PCNA compared to normal HFKs, and this was maintained upon differentiation in high calcium, while levels of the RPA subunit 32 (RPA32) were similar between the two sets of cells. When examined by IF/FISH, we found that both PCNA and RPA32 also formed foci that co-localized with HPV genomes in undifferentiated cells, and these foci increased in size upon differentiation, indicating that these are indeed active viral replication 154 centers (Figure 3B). PCNA and RPA were localized to 81% and 77% of FISH positive 6
7 undifferentiated cells, respectively, and 80% and 85%, respectively of differentiating cells. Although we could detect PCNA and RPA at replication foci in undifferentiated cells, the intensity of the foci was very faint RPA consists of three subunits, RPA70, RPA32 and RPA14, and is the major single-stranded DNA (ssdna) binding protein in eukaryotic cells (3). RPA accumulates along stretches of ssdna generated by stalled replication forks or DNA damage. In response to DNA damage, the N terminus of RPA32 is phosphorylated at multiple residues by DNA damage kinases (ATM, ATR, DNA-PK) (27), which has been suggested to redirect the function of RPA from DNA replication to repair DNA synthesis (27, 44). To determine if HPV alters the phosphorylation state of RPA, we examined phosphorylation of the RPA32 Ser33 residue (prpa32 S33) by Western blot analysis and found that the levels of prpa32 S33 were not significantly increased in HPV positive cells upon differentiation as compared to uninfected HFKs (Figure 3A). However, using the IF/FISH assay, we detected prpa32 S33 at HPV replication foci in 30-60% of undifferentiated FISH positive cells (Figure 3C). Interestingly, the number of foci containing prpa that colocalized with HPV DNA increased to 83-85% in differentiated cells, with the size of the foci increasing in response to viral genome amplification. Since phosphorylated RPA localizes to nuclear foci where DNA repair is occurring following DNA damage (3, 45), HPV DNA synthesis may be dependent on repair replication. Cellular factors involved in homologous recombination repair are localized to viral replication compartments. Mammalian cells have two primary mechanisms by which they can repair double stranded breaks: homologous recombination (HR) and non-homologous end joining (NHEJ) (13, 28). HR is an accurate repair process that involves template-directed recombination and is restricted to the S and G2 phases of the cell cycle (25, 34). The MRN complex is the primary sensor that detects DSBs in the case of HR, and ssdna generated by resection of the DSB is protected by RPA containing phosphorylated RPA32. NHEJ is an error prone mechanism that can be carried out at any phase of the cell cycle, with the Ku70/Ku80 complex being the primary sensor of DSBs (19). Since we 7
8 previously observed formation of foci containing MRN components in HPV positive cells (24) and can also detect them at sites of viral replication (data not shown), we explored the potential linkage between HPV replication and cellular DNA repair through HR. We first examined the levels of the HR proteins Rad51 and BRCA1 by Western blot analysis. HPV positive cells have higher levels of both Rad51 and BRCA1 compared to undifferentiated HFKs (Figure 4A). Upon differentiation, levels of Rad51 remain high through 48hr post-exposure to calcium, whereas Brca1 is detected at higher levels throughout the time course (Figure 4A). To determine if the increase in Rad51 and BRCA1 coincided with the localization of these repair factors to HPV genomes, we performed IF for Rad51 and BRCA1 and FISH for HPV DNA on detergent extracted CIN 612 cells, as well as normal HFKs. In undifferentiated cells the amount of Rad51 co-localizing with HPV genomes was very faint and detected in 50-58% of FISH positive cells (Figure 4B). Upon differentiation, Rad51 colocalized with 60-68% of cells that were HPV positive by FISH, and the staining intensity increased upon viral genome amplification. Similar results were observed for BRCA1 (Figure 4C), with localization occurring at 60-78% of undifferentiated FISH positive cells, and 64-73% of differentiating FISH positive cells. In contrast to the localization of HR proteins to HPV DNA foci, we did not observe significant localization of the NHEJ factor DNA-PK (phosphorylated on Thr 2069) to sites of HPV replication (data not shown). The observation that HR proteins, including phosphorylated RPA, Rad51 and BRCA1 localize to HPV replication foci suggest that the amplification of viral genomes during the productive phase may require HR machinery to ensure high fidelity replication. γh2ax associates with viral genomes. The studies described above demonstrate that many components of the ATM DNA damage pathway co-localize with HPV genomes in foci that also contain cellular replication and homologous recombination repair factors. It is possible that these factors localize to these regions but are not bound to viral genomes. We therefore investigated if repair factors actually bind to HPV genomes. For our initial analyses we focused on γh2ax and examined binding to viral DNA by Chromatin Immunoprecipitation (ChIP) assays. We screened for binding of γh2ax to sequences around the HPV origin of replication in the upstream regulatory region (URR) in 8
9 undifferentiated as well as differentiated cells (Figure 5). An increase of approximately 7-fold in γh2ax binding to the origin was detected in undifferentiated cells over the IgG control and this increased upon differentiation to over 20-fold. This indicates that DNA repair enzymes are recruited to viral genomes during productive replication in differentiated cells Our studies demonstrate that members of the ATM DNA damage pathway are recruited to HPV replication foci in a manner resembling the effects seen following induction of DNA damage. We believe that the localization of the host cellular replication factors, PCNA and RPA, to sites of HPV DNA indicates that these centers contain viral genomes undergoing replication. The ATM DNA damage pathway can induce cell cycle arrest at either G1/S or G2/M depending on which arm of the pathway is activated. ATM signaling through p53 induces G1/S arrest while ATM signaling to Chk2 arrests cells in G2/M (7). HPV has been reported to amplify its DNA in cells that are in the G2 phase of the cell cycle, after cellular DNA has replicated (40). While HPV-induced ATM activation may contribute to productive replication via this mechanism, our studies suggest that the DDR factors also play an active role in HPV replication since they are recruited to HPV replication compartments. The DSB markers γh2ax and 53BP1 were found to co-localize with replicating HPV DNA. Our studies suggest the possibility that DSBs play a role in HPV genome amplification. DSBs could be introduced into amplifying HPV DNA if productive replication is mediated through a rolling circle, which has been suggested as the mechanism of differentiation-dependent genome amplification (10). In addition, the dramatic increase in viral replication taking place in differentiating cells could lead to replication stress, resulting in stalled replication forks and the formation of DSBs. Activation of ATM signaling and the recruitment of cellular HR repair factors would provide an efficient mechanism for HPV to repair and synthesize viral DNA. The finding that γh2ax localizes to viral replication centers and is bound to viral chromatin suggests that γh2ax may recruit host repair proteins necessary to carry out viral DNA replication. Within HPV virions, viral genomes are associated with histones (37, 9
10 ), raising the possibility that H2AX is incorporated into newly replicated genomes and packaged into progeny viruses. This hypothesis will be investigated in more detail in future studies We previously found that HPV induces the activation of an ATM-dependent DNA damage response. While our current study indicates that an outcome of this response is the recruitment of repair factors to viral genomes, the mechanism by which HPV activates ATM signaling is unclear. One possibility is that expression of a viral protein elicits this response. In support of this, we, as well as others have shown that expression of E7 is sufficient to activate ATM and its downstream targets (24, 29, 32). We have also found that E7 interacts with ATM (24), as well as the MRN complex (Moody C.A., unpublished results). Recent studies indicate that the expression of the viral replication protein E1 can also activate ATM signaling (11, 15, 33). E7 and/or E1 may induce cellular DNA damage that consequently provides DNA repair/replication factors necessary for the synthesis of viral DNA. Alternatively, the interaction of HPV proteins with DNA repair factors could facilitate recruitment to viral DNA, which may be sufficient to nucleate a DNA damage response (36). It is also possible that viral replication results in the formation of intermediates that the cell recognizes as DNA damage. This may be sufficient to activate ATM signaling, resulting in the recruitment of repair factors necessary to allow viral genome amplification. Understanding how HPV triggers a DNA damage response will be the focus of future investigations. Our findings that cellular factors involved in HR, including Rad51, BRCA1, and prpa S33 are also recruited to HPV replication foci raises the intriguing possibility that homologous recombination is directly involved in HPV genome amplification. HSV, Epstein Barr Virus and SV40 have all been shown to recruit Rad51 to replication compartments (4, 16, 41). For EBV, as well as SV40, knockdown of Rad51 diminishes viral genome synthesis, suggesting that HR is necessary for efficient replication (4, 16). The localization of HR factors to HPV DNA foci may indicate that viral replication compartments may contain substrates, such as stalled replication forks or abnormal replication intermediates that require HR activity to repair to facilitate high fidelity viral DNA synthesis upon 10
11 differentiation. However, it is possible that HPV actively uses these factors to promote replication. In addition, if viral genome amplification does proceed through a rolling circle mechanism, HR may be required to circularize HPV genomes upon cleavage of viral concatemers to allow for packaging into virions. Overall, these studies show that members of the ATM DNA damage pathway along with factors involved in homologous recombination play active roles in the differentiation-dependent amplification of HPV genomes. Acknowledgements We thank Dale Ramsden for generously providing the pdna-pk Thr 2069 antibody. L.A.L. was supported by grants from the NCI, and C.A.M was supported by a Pathway to Independence Award from the NCI. References 1. Bedell, M. A., J. B. Hudson, T. R. Golub, M. E. Turyk, M. Hosken, G. D. Wilbanks, and L. A. Laimins Amplification of human papillomavirus genomes in vitro is dependent on epithelial differentiation. J Virol 65: Bekker-Jensen, S., and N. Mailand Assembly and function of DNA double-strand break repair foci in mammalian cells. DNA Repair (Amst) 9: Binz, S. K., A. M. Sheehan, and M. S. Wold Replication protein A phosphorylation and the cellular response to DNA damage. DNA Repair (Amst) 3: Boichuk, S., L. Hu, J. Hein, and O. V. Gjoerup Multiple DNA damage signaling and repair pathways deregulated by simian virus 40 large T antigen. J Virol 84: Ciccia, A., and S. J. Elledge The DNA damage response: making it safe to play with knives. Mol Cell 40: de Bruyn Kops, A., and D. M. Knipe Formation of DNA replication structures in herpes virusinfected cells requires a viral DNA binding protein. Cell 55: Derheimer, F. A., and M. B. Kastan Multiple roles of ATM in monitoring and maintaining DNA integrity. FEBS Lett 584:
12 Falck, J., J. Coates, and S. P. Jackson Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage. Nature 434: Fernandez-Capetillo, O., A. Lee, M. Nussenzweig, and A. Nussenzweig H2AX: the histone guardian of the genome. DNA Repair (Amst) 3: Flores, E. R., and P. F. Lambert Evidence for a switch in the mode of human papillomavirus type 16 DNA replication during the viral life cycle. J Virol 71: Fradet-Turcotte, A., F. Bergeron-Labrecque, C. A. Moody, M. Lehoux, L. A. Laimins, and J. Archambault Nuclear accumulation of the papillomavirus E1 helicase blocks S-phase progression and triggers an ATM-dependent DNA damage response. J Virol 85: Harper, J. W., and S. J. Elledge The DNA damage response: ten years after. Mol Cell 28: Hartlerode, A. J., and R. Scully Mechanisms of double-strand break repair in somatic mammalian cells. Biochem J 423: Huen, M. S., and J. Chen The DNA damage response pathways: at the crossroad of protein modifications. Cell Res 18: Kadaja, M., H. Isok-Paas, T. Laos, E. Ustav, and M. Ustav Mechanism of genomic instability in cells infected with the high-risk human papillomaviruses. PLoS Pathog 5:e Kudoh, A., S. Iwahori, Y. Sato, S. Nakayama, H. Isomura, T. Murata, and T. Tsurumi Homologous recombinational repair factors are recruited and loaded onto the viral DNA genome in Epstein-Barr virus replication compartments. J Virol 83: Lee, J. H., and T. T. Paull ATM activation by DNA double-strand breaks through the Mre11- Rad50-Nbs1 complex. Science 308: Lee, J. H., and T. T. Paull Direct activation of the ATM protein kinase by the Mre11/Rad50/Nbs1 complex. Science 304: Lieber, M. R The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu Rev Biochem 79: Longworth, M. S., and L. A. Laimins Pathogenesis of human papillomaviruses in differentiating epithelia. Microbiol Mol Biol Rev 68:
13 Matsuoka, S., B. A. Ballif, A. Smogorzewska, E. R. McDonald, 3rd, K. E. Hurov, J. Luo, C. E. Bakalarski, Z. Zhao, N. Solimini, Y. Lerenthal, Y. Shiloh, S. P. Gygi, and S. J. Elledge ATM and ATR substrate analysis reveals extensive protein networks responsive to DNA damage. Science 316: Mochan, T. A., M. Venere, R. A. DiTullio, Jr., and T. D. Halazonetis BP1, an activator of ATM in response to DNA damage. DNA Repair (Amst) 3: Moody, C. A., A. Fradet-Turcotte, J. Archambault, and L. A. Laimins Human papillomaviruses activate caspases upon epithelial differentiation to induce viral genome amplification. Proc Natl Acad Sci U S A 104: Moody, C. A., and L. A. Laimins Human papillomaviruses activate the ATM DNA damage pathway for viral genome amplification upon differentiation. PLoS Pathog 5:e Moynahan, M. E., and M. Jasin Mitotic homologous recombination maintains genomic stability and suppresses tumorigenesis. Nat Rev Mol Cell Biol 11: Munger, K., A. Baldwin, K. M. Edwards, H. Hayakawa, C. L. Nguyen, M. Owens, M. Grace, and K. Huh Mechanisms of human papillomavirus-induced oncogenesis. J Virol 78: Nuss, J. E., S. M. Patrick, G. G. Oakley, G. M. Alter, J. G. Robison, K. Dixon, and J. J. Turchi DNA damage induced hyperphosphorylation of replication protein A. 1. Identification of novel sites of phosphorylation in response to DNA damage. Biochemistry 44: Pardo, B., B. Gomez-Gonzalez, and A. Aguilera DNA repair in mammalian cells: DNA doublestrand break repair: how to fix a broken relationship. Cell Mol Life Sci 66: Pickering, M. T., and T. F. Kowalik Rb inactivation leads to E2F1-mediated DNA double-strand break accumulation. Oncogene 25: Quinlan, M. P., L. B. Chen, and D. M. Knipe The intranuclear location of a herpes simplex virus DNA-binding protein is determined by the status of viral DNA replication. Cell 36: Rixon, F. J., M. A. Atkinson, and J. Hay Intranuclear distribution of herpes simplex virus type 2 DNA synthesis: examination by light and electron microscopy. J Gen Virol 64 (Pt 9): Rogoff, H. A., M. T. Pickering, F. M. Frame, M. E. Debatis, Y. Sanchez, S. Jones, and T. F. Kowalik Apoptosis associated with deregulated E2F activity is dependent on E2F1 and Atm/Nbs1/Chk2. Mol Cell Biol 24:
14 Sakakibara, N., R. Mitra, and A. A. McBride The papillomavirus E1 helicase activates a cellular DNA damage response in viral replication foci. J Virol 85: San Filippo, J., P. Sung, and H. Klein Mechanism of eukaryotic homologous recombination. Annu Rev Biochem 77: Silverman, J., H. Takai, S. B. Buonomo, F. Eisenhaber, and T. de Lange Human Rif1, ortholog of a yeast telomeric protein, is regulated by ATM and 53BP1 and functions in the S-phase checkpoint. Genes Dev 18: Soutoglou, E., and T. Misteli Activation of the cellular DNA damage response in the absence of DNA lesions. Science 320: Stunkel, W., and H. U. Bernard The chromatin structure of the long control region of human papillomavirus type 16 represses viral oncoprotein expression. J Virol 73: Swindle, C. S., N. Zou, B. A. Van Tine, G. M. Shaw, J. A. Engler, and L. T. Chow Human papillomavirus DNA replication compartments in a transient DNA replication system. J Virol 73: Wang, B., S. Matsuoka, P. B. Carpenter, and S. J. Elledge BP1, a mediator of the DNA damage checkpoint. Science 298: Wang, H. K., A. A. Duffy, T. R. Broker, and L. T. Chow Robust production and passaging of infectious HPV in squamous epithelium of primary human keratinocytes. Genes Dev 23: Wilkinson, D. E., and S. K. Weller Recruitment of cellular recombination and repair proteins to sites of herpes simplex virus type 1 DNA replication is dependent on the composition of viral proteins within prereplicative sites and correlates with the induction of the DNA damage response. J Virol 78: Wong, P. P., A. Pickard, and D. J. McCance p300 alters keratinocyte cell growth and differentiation through regulation of p21(waf1/cip1). PLoS One 5:e Wooldridge, T. R., and L. A. Laimins Regulation of human papillomavirus type 31 gene expression during the differentiation-dependent life cycle through histone modifications and transcription factor binding. Virology 374:
15 Zernik-Kobak, M., K. Vasunia, M. Connelly, C. W. Anderson, and K. Dixon Sites of UVinduced phosphorylation of the p34 subunit of replication protein A from HeLa cells. J Biol Chem 272: Zou, Y., Y. Liu, X. Wu, and S. M. Shell Functions of human replication protein A (RPA): from DNA replication to DNA damage and stress responses. J Cell Physiol 208: Figure Legends Figure 1. Proteins associated with damaged DNA co-localize with HPV DNA foci. A) Whole cell extracts were harvested from CIN 612 cells or normal HFKs at T0 or after 48, 96 hours differentiation in high calcium medium, as described previously (23). Immunoblotting was performed using antibodies to γh2ax (Cell Signaling) and 53BP1 (Abcam). GAPDH served as a loading control. Ca=calcium. Protein levels were quantified by Image J, normalized to levels observed in uninfected keratinocytes. Shown is a representative experiment. B) IF for γh2ax (Millipore) was performed followed by FISH for HPV DNA on detergent extracted CIN 612 cells harvested at T0 or after 72 hours of calcium-induced differentiation. IF for γh2ax was performed on detergent-extracted normal HFKs at T0 or 72hr post-exposure to high calcium medium. Cellular DNA was counterstained with DAPI. Omission of the pre-extraction step did not alter the localization of γh2ax to HPV DNA foci (no preextraction). Images were collected on a Zeiss 710 confocal laser-scanning microscope and were processed using the Zeiss Zen software. C) IF for 53BP1 (Abcam) was performed on pre-extracted CIN 612 cells, followed by FISH for HPV DNA. IF for 53BP1 was also performed on detergentextracted normal HFKs at T0 or after 72 hours of differentiation in high calcium. Figure 2. patm and Chk2 localize to HPV DNA foci. (A) Whole cell extracts were harvested from CIN 612 cells and normal HFKs that were undifferentiated (T0) or induced to differentiate in high calcium medium for 48 and 96 hours as described previously (23). Western blot analysis was 15
16 performed using an antibody to ATM phosphorylated on Ser1981 (Epitomics), total ATM (Calbiochem), Chk2 phosphorylated on Thr68 (Cell Signaling), total Chk2 (Santa Cruz), followed by HRP-linked linked secondary antibodies (Cell Signaling). GAPDH served as a loading control. Protein levels were quantified by Image J, normalized to levels observed in uninfected keratinocytes. Ca=calcium. Shown is a representative experiment. (B,C) Immunofluorescence (IF) for patm Ser1981 (Epitomics) (B) and Chk2 (Abcam) (C) was followed by FISH for HPV DNA on detergentextracted CIN 612 cells that were undifferentiated (T0) or differentiated in high calcium medium for 72 hours. Cellular DNA was counterstained using DAPI. Figure 3. Cellular replication factors localize to HPV replication foci. (A) Immunoblotting was performed using antibodies to PCNA (Abcam), RPA32 (Bethyl Laboratories) and prpa32 S33 (Bethyl Laboratories) on whole cell lysates extracted from CIN 612 cells and HFKs that were undifferentiated or differentiated in high calcium medium for 48 and 96 hours. GAPDH served as a loading control. Protein levels were quantified by Image J, normalized to levels observed in uninfected keratinocytes. Shown is a representative experiment. (B,C) IF for PCNA (Abcam) (B), RPA32 (Bethyl Laboratories) (B) and prpa32 S33 (Bethyl Laboratories) (C) was performed followed by FISH for HPV DNA on detergent extracted CIN 612 cells that were undifferentiated or differentiated in high calcium medium for 72 hours. DAPI was used to counterstain cellular DNA. (C) IF was performed on detergent extracted HFKs at T0 or after 72 hours differentiation in high calcium medium using an antibody to prpa32 S33. DAPI was used to counterstain cellular DNA. Figure 4. HPV positive cells exhibit increased levels of homologous recombination factors, which are localized to HPV replication foci. (A) Whole cell extracts were harvested from CIN 612 cells and HFKs that were undifferentiated or differentiated in high calcium medium for the indicated times. Western blot analysis was performed using antibodies to Rad51 (Santa Cruz) and BRCA1 (Calbiochem). GAPDH served as a loading control. Protein levels were quantified by Image J, normalized to levels observed in uninfected keratinocytes. Shown is a representative experiment. (B, 16
17 C) IF for Rad51 (B) and BRCA1 (C) was performed followed by FISH for HPV DNA on detergent extracted CIN 612 cells harvested at T0 or after 72 hours differentiation in high calcium medium. IF was also performed on detergent extracted HFKs that were undifferentiated or differentiated in high calcium using antibodies to Rad51 (B) and BRCA1 (C). Cellular DNA was counterstained with DAPI Figure 5. γh2ax associates with HPV genomes. Chromatin Immunoprecipitation (ChIP) assays were performed on CIN 612 cells that were harvested at T0 or after 72 hours of differentiation in high calcium medium, as previously described by Wong et al (42). Immunoprecipitations were performed using an antibody to phospho-h2ax Ser139 (γh2ax) (Millipore). Quantitative PCR was done using a Roche Lightcycler 480 Sybr green Master Kit (Roche Applied Sciences). The primers used were specific to the Upstream Regulatory Region (URR) of HPV31. Forward: 5 AAC TGC CAA GGT TGT GTC ATG C 3, Reverse: 5 TGG CGT CTG TAG GTT TGC AC 3.The data is expressed as fold activation over the control mouse IgG (Santa Cruz), where the fold level was set at one to allow for comparisons. Ca=calcium. Shown is a representative of two experiments. 17


18


19

20


21


22
Human Papillomaviruses Activate the ATM DNA Damage Pathway for Viral Genome Amplification upon Differentiation
 Human Papillomaviruses Activate the ATM DNA Damage Pathway for Viral Genome Amplification upon Differentiation Cary A. Moody, Laimonis A. Laimins* Department of Microbiology and Immunology, Feinberg School
Human Papillomaviruses Activate the ATM DNA Damage Pathway for Viral Genome Amplification upon Differentiation Cary A. Moody, Laimonis A. Laimins* Department of Microbiology and Immunology, Feinberg School
Tumour growth environment modulates Chk1 signalling pathways and sensitivity to Chk1 inhibition
 Tumour growth environment modulates Chk1 signalling pathways and sensitivity to Chk1 inhibition Andrew J Massey Supplementary Information Supplementary Figure S1. Related to Fig. 1. (a) HT29 or U2OS cells
Tumour growth environment modulates Chk1 signalling pathways and sensitivity to Chk1 inhibition Andrew J Massey Supplementary Information Supplementary Figure S1. Related to Fig. 1. (a) HT29 or U2OS cells
Productive Replication of Human Papillomavirus 31 Requires the DNA Repair Factor Nbs1. Running Title: Nbs1 is necessary for productive HPV replication
 JVI Accepts, published online ahead of print on 21 May 2014 J. Virol. doi:10.1128/jvi.00517-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
JVI Accepts, published online ahead of print on 21 May 2014 J. Virol. doi:10.1128/jvi.00517-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Legends for Supplemental Figures.
 1 Legends for Supplemental Figures. Supplemental Figure 1. Proteasome Inhibition does not abrogate the expression of FA core complex proteins or formation of the FA core complex. (Full-length blots are
1 Legends for Supplemental Figures. Supplemental Figure 1. Proteasome Inhibition does not abrogate the expression of FA core complex proteins or formation of the FA core complex. (Full-length blots are
DSB. Double-Strand Breaks causate da radiazioni stress ossidativo farmaci
 DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci METODI DDR foci formation in irradiated (2 Gy) cells fixed 2 h later IRIF IRradiation Induced Focus Laser micro-irradiation DDR
DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci METODI DDR foci formation in irradiated (2 Gy) cells fixed 2 h later IRIF IRradiation Induced Focus Laser micro-irradiation DDR
Mechanism of Genomic Instability in Cells Infected with the High-Risk Human Papillomaviruses
 Mechanism of Genomic Instability in Cells Infected with the High-Risk Human Papillomaviruses Meelis Kadaja 1,2, Helen Isok-Paas 3, Triin Laos 1, Ene Ustav 2,3, Mart Ustav 3 * 1 Institute of Molecular and
Mechanism of Genomic Instability in Cells Infected with the High-Risk Human Papillomaviruses Meelis Kadaja 1,2, Helen Isok-Paas 3, Triin Laos 1, Ene Ustav 2,3, Mart Ustav 3 * 1 Institute of Molecular and
Targeting the ATR Kinase in Cancer Therapy
 Targeting the ATR Kinase in Cancer Therapy 2017 Chabner Colloquium October 30, 2017 Lee Zou MGH Cancer Center Harvard Medical School Disclosure Consultant/advisory role: Loxo Oncology DNA Damage and Replication
Targeting the ATR Kinase in Cancer Therapy 2017 Chabner Colloquium October 30, 2017 Lee Zou MGH Cancer Center Harvard Medical School Disclosure Consultant/advisory role: Loxo Oncology DNA Damage and Replication
Brian T Burgess, DO, PhD, GYN Oncology Fellow Rachel W. Miller, MD, GYN Oncology
 Brian T Burgess, DO, PhD, GYN Oncology Fellow Rachel W. Miller, MD, GYN Oncology Epithelial Ovarian Cancer - Standard Current Treatment: Surgery with De-bulking + Platinum-Taxane based Chemotherapy - No
Brian T Burgess, DO, PhD, GYN Oncology Fellow Rachel W. Miller, MD, GYN Oncology Epithelial Ovarian Cancer - Standard Current Treatment: Surgery with De-bulking + Platinum-Taxane based Chemotherapy - No
Removal of Shelterin Reveals the Telomere End-Protection Problem
 Removal of Shelterin Reveals the Telomere End-Protection Problem DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to
Removal of Shelterin Reveals the Telomere End-Protection Problem DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to
Received 2 August 2006/Accepted 21 November 2006
 JOURNAL OF VIROLOGY, Feb. 2007, p. 1934 1950 Vol. 81, No. 4 0022-538X/07/$08.00 0 doi:10.1128/jvi.01670-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Human Cytomegalovirus
JOURNAL OF VIROLOGY, Feb. 2007, p. 1934 1950 Vol. 81, No. 4 0022-538X/07/$08.00 0 doi:10.1128/jvi.01670-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Human Cytomegalovirus
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
HCMV disrupts both ATM and ATR-mediated. DNA damage responses during lytic infection ACCEPTED
 JVI Accepts, published online ahead of print on 6 December 2006 J. Virol. doi:10.1128/jvi.01670-06 Copyright 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
JVI Accepts, published online ahead of print on 6 December 2006 J. Virol. doi:10.1128/jvi.01670-06 Copyright 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
SV40 Utilizes ATM Kinase Activity to Prevent Non-homologous End Joining of Broken Viral DNA Replication Products
 SV40 Utilizes ATM Kinase Activity to Prevent Non-homologous End Joining of Broken Viral DNA Replication Products Gregory A. Sowd 1 *, Dviti Mody 1, Joshua Eggold 1, David Cortez 2, Katherine L. Friedman
SV40 Utilizes ATM Kinase Activity to Prevent Non-homologous End Joining of Broken Viral DNA Replication Products Gregory A. Sowd 1 *, Dviti Mody 1, Joshua Eggold 1, David Cortez 2, Katherine L. Friedman
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
SUPPLEMENTARY INFORMATION
 Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Chapter 2. Aims & Objectives
 2.1. Statement of the problem: Earlier reports have shown ambiguous alteration of histone marks in response to DNA damage in asynchronized population of cells. These histone marks not only undergo dynamic
2.1. Statement of the problem: Earlier reports have shown ambiguous alteration of histone marks in response to DNA damage in asynchronized population of cells. These histone marks not only undergo dynamic
Tumor cell reassortment within the cell cycle (including checkpoints and cell-cycle arrest)
") Tumor cell reassortment within the cell cycle (including checkpoints and cell-cycle arrest) Carsten Herskind Dept. of Radiation Oncology. Universitätsmedizin Mannheim Medical Faculty Mannheim, Heidelberg
Tumor cell reassortment within the cell cycle (including checkpoints and cell-cycle arrest) Carsten Herskind Dept. of Radiation Oncology. Universitätsmedizin Mannheim Medical Faculty Mannheim, Heidelberg
Removal of Shelterin Reveals the Telomere End-Protection Problem
 Removal of Shelterin Reveals the Telomere End-Protection Problem DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to
Removal of Shelterin Reveals the Telomere End-Protection Problem DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to
THE ACTIVATION AND CONSEQUENCES OF THE ATM MEDIATED DNA DAMAGE RESPONSE IN HPV INFECTED CELLS. Bryan Allen Johnson
 THE ACTIVATION AND CONSEQUENCES OF THE ATM MEDIATED DNA DAMAGE RESPONSE IN HPV INFECTED CELLS Bryan Allen Johnson A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill
THE ACTIVATION AND CONSEQUENCES OF THE ATM MEDIATED DNA DAMAGE RESPONSE IN HPV INFECTED CELLS Bryan Allen Johnson A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 AD Award Number: W81XWH-05-1-0494 TITLE: Tumor Suppression by BRCA-1: A Critical Role at DNA Replication Forks PRINCIPAL INVESTIGATOR: Jean Gautier, Ph.D. CONTRACTING ORGANIZATION: Columbia University
AD Award Number: W81XWH-05-1-0494 TITLE: Tumor Suppression by BRCA-1: A Critical Role at DNA Replication Forks PRINCIPAL INVESTIGATOR: Jean Gautier, Ph.D. CONTRACTING ORGANIZATION: Columbia University
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
DSB. Double-Strand Breaks causate da radiazioni stress ossidativo farmaci
 DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to the detection and repair of DNA damage DSBs induce a local decrease
DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA Higher-order chromatin packaging is a barrier to the detection and repair of DNA damage DSBs induce a local decrease
Review Human Papillomavirus and the DNA Damage Response: Exploiting Host Repair Pathways for Viral Replication
 Review Human Papillomavirus and the DNA Damage Response: Exploiting Host Repair Pathways for Viral Replication Chelsey C. Spriggs and Laimonis A. Laimins * Department of Microbiology-Immunology, Northwestern
Review Human Papillomavirus and the DNA Damage Response: Exploiting Host Repair Pathways for Viral Replication Chelsey C. Spriggs and Laimonis A. Laimins * Department of Microbiology-Immunology, Northwestern
Chapt 15: Molecular Genetics of Cell Cycle and Cancer
 Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
Chapt 15: Molecular Genetics of Cell Cycle and Cancer Student Learning Outcomes: Describe the cell cycle: steps taken by a cell to duplicate itself = cell division; Interphase (G1, S and G2), Mitosis.
HPV31 utilizes the ATR-Chk1 pathway to maintain elevated RRM2 levels and a replication-competent environment in differentiating Keratinocytes
 HPV31 utilizes the ATR-Chk1 pathway to maintain elevated RRM2 levels and a replication-competent environment in differentiating Keratinocytes Daniel C. Anacker, University of North Carolina Heather L.
HPV31 utilizes the ATR-Chk1 pathway to maintain elevated RRM2 levels and a replication-competent environment in differentiating Keratinocytes Daniel C. Anacker, University of North Carolina Heather L.
Supplementary Figure 1
 Supplementary Figure 1 a γ-h2ax MDC1 RNF8 FK2 BRCA1 U2OS Cells sgrna-1 ** 60 sgrna 40 20 0 % positive Cells (>5 foci per cell) b ** 80 sgrna sgrna γ-h2ax MDC1 γ-h2ax RNF8 FK2 MDC1 BRCA1 RNF8 FK2 BRCA1
Supplementary Figure 1 a γ-h2ax MDC1 RNF8 FK2 BRCA1 U2OS Cells sgrna-1 ** 60 sgrna 40 20 0 % positive Cells (>5 foci per cell) b ** 80 sgrna sgrna γ-h2ax MDC1 γ-h2ax RNF8 FK2 MDC1 BRCA1 RNF8 FK2 BRCA1
An E2F1-mediated DNA damage response contributes to the replication of human cytomegalovirus
 University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 5-12-2011 An E2F1-mediated DNA damage response contributes to the replication
University of Massachusetts Medical School escholarship@umms Open Access Articles Open Access Publications by UMMS Authors 5-12-2011 An E2F1-mediated DNA damage response contributes to the replication
Lecture 10. G1/S Regulation and Cell Cycle Checkpoints. G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint
 Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS
 CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS Summary of the regulation of cyclin/cdk complexes during celll cycle Cell cycle phase Cyclin-cdk complex inhibitor activation Substrate(s) G1 Cyclin D/cdk 4,6
CELL CYCLE MOLECULAR BASIS OF ONCOGENESIS Summary of the regulation of cyclin/cdk complexes during celll cycle Cell cycle phase Cyclin-cdk complex inhibitor activation Substrate(s) G1 Cyclin D/cdk 4,6
Cell Cycle. Trends in Cell Biology
 Cell Cycle Trends in Cell Biology Cell Cycle The orderly sequence of events by which a cell duplicates its contents and divides into two Daughter Cells Activities of a cell from one cell division to the
Cell Cycle Trends in Cell Biology Cell Cycle The orderly sequence of events by which a cell duplicates its contents and divides into two Daughter Cells Activities of a cell from one cell division to the
MOLECULAR MECHANISMS OF MAMMALIAN DNA REPAIR AND THE DNA DAMAGE CHECKPOINTS
 Annu. Rev. Biochem. 2004. 73:39 85 doi: 10.1146/annurev.biochem.73.011303.073723 Copyright 2004 by Annual Reviews. All rights reserved First published online as a Review in Advance on January 20, 2004
Annu. Rev. Biochem. 2004. 73:39 85 doi: 10.1146/annurev.biochem.73.011303.073723 Copyright 2004 by Annual Reviews. All rights reserved First published online as a Review in Advance on January 20, 2004
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
CELL CYCLE REGULATION AND CANCER. Cellular Reproduction II
 CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
Multistep nature of cancer development. Cancer genes
 Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
Multistep nature of cancer development Phenotypic progression loss of control over cell growth/death (neoplasm) invasiveness (carcinoma) distal spread (metastatic tumor) Genetic progression multiple genetic
Cancer Biology How a cell responds to DNA Damage
 1 Cancer Biology How a cell responds to DNA Damage Jann Sarkaria Department of Oncology Mayo Clinic 2 EDUCATIONAL GOALS How proteins can transmit signals to each other. The definition of a tumor suppressor
1 Cancer Biology How a cell responds to DNA Damage Jann Sarkaria Department of Oncology Mayo Clinic 2 EDUCATIONAL GOALS How proteins can transmit signals to each other. The definition of a tumor suppressor
ABSTRACT. by Jason Patrick Clark
 ABSTRACT INVESTIGATING CELLULAR DNA DAMAGE RESPONSES INDUCED DURING ADENOVIRUS EARLY REGION 4 MUTANT INFECTION AND THEIR IMPACT ON VIRAL DNA REPLICATION by Jason Patrick Clark DNA double-strand breaks
ABSTRACT INVESTIGATING CELLULAR DNA DAMAGE RESPONSES INDUCED DURING ADENOVIRUS EARLY REGION 4 MUTANT INFECTION AND THEIR IMPACT ON VIRAL DNA REPLICATION by Jason Patrick Clark DNA double-strand breaks
Prof. R. V. Skibbens. Cell Cycle, Cell Division and Cancer (Part 2)
") Prof. R. V. Skibbens November 22, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality - clocks go in only one direction G1 doesn t have replication-inducing
Prof. R. V. Skibbens November 22, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality - clocks go in only one direction G1 doesn t have replication-inducing
Transcription and RNA processing
 Transcription and RNA processing Lecture 7 Biology 3310/4310 Virology Spring 2018 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at the
Transcription and RNA processing Lecture 7 Biology 3310/4310 Virology Spring 2018 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at the
Prof. R. V. Skibbens
 Prof. R. V. Skibbens December 2, 2011 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality The Cell Cycle clock goes in only one direction S-phase cells
Prof. R. V. Skibbens December 2, 2011 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer (Part 2) Directionality The Cell Cycle clock goes in only one direction S-phase cells
Breast Cancer Susceptibility and HLA Complex: Source of Missing Heritability?
 Breast Cancer Susceptibility and HLA Complex: Source of Missing Heritability? M. Tevfik DORAK Environmental & Occupational Health College of Public Health Florida International University Miami, USA http://www.dorak.info
Breast Cancer Susceptibility and HLA Complex: Source of Missing Heritability? M. Tevfik DORAK Environmental & Occupational Health College of Public Health Florida International University Miami, USA http://www.dorak.info
HLA Complex Genetics & Biology
 HLA Complex Genetics & Biology M. Tevfik DORAK Environmental & Occupational Health College of Public Health Florida International University Miami, USA http://www.dorak.info Mount Sinai Medical Center
HLA Complex Genetics & Biology M. Tevfik DORAK Environmental & Occupational Health College of Public Health Florida International University Miami, USA http://www.dorak.info Mount Sinai Medical Center
ATM and ATR Activities Maintain Replication Fork Integrity during SV40 Chromatin Replication
 ATM and ATR Activities Maintain Replication Fork Integrity during SV40 Chromatin Replication Gregory A. Sowd, Nancy Yan Li, Ellen Fanning* Department of Biological Sciences, Vanderbilt University, Vanderbilt
ATM and ATR Activities Maintain Replication Fork Integrity during SV40 Chromatin Replication Gregory A. Sowd, Nancy Yan Li, Ellen Fanning* Department of Biological Sciences, Vanderbilt University, Vanderbilt
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney
 2013 Course University of Sydney") BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Mitosis and the Cell Cycle
 Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts
 Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts (TIG-3 cells) were rendered senescent by either serial passage
Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts (TIG-3 cells) were rendered senescent by either serial passage
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Control of Cell Cycle. Unit 2 Part f III
 Control of Cell Cycle Unit 2 Part f III How often do cells divide and why? The timing and rate of cell division in different parts of the plant or animals are crucial to normal growth, development and
Control of Cell Cycle Unit 2 Part f III How often do cells divide and why? The timing and rate of cell division in different parts of the plant or animals are crucial to normal growth, development and
Repair of Broken Chromosomes and Maintenance of Chromosome Stability
 Repair of Broken Chromosomes and Maintenance of Chromosome Stability Jim Haber Brandeis University Genome instability in tumor cells Truncations Translocations Inversions Duplications Amplifications Deletions
Repair of Broken Chromosomes and Maintenance of Chromosome Stability Jim Haber Brandeis University Genome instability in tumor cells Truncations Translocations Inversions Duplications Amplifications Deletions
Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis
 Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis MUDr. Jiří Vachtenheim, CSc. CELL CYCLE - SUMMARY Basic terminology: Cyclins conserved proteins with homologous regions; their cellular
Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis MUDr. Jiří Vachtenheim, CSc. CELL CYCLE - SUMMARY Basic terminology: Cyclins conserved proteins with homologous regions; their cellular
Basics of Radiation Biology
 Basics of Radiation Biology Sally A. Amundson Columbia University Center for Radiological Research http://www.cmcr.columbia.edu/ Overview Radiation damage to cells DNA Effects of radiation damage on cells
Basics of Radiation Biology Sally A. Amundson Columbia University Center for Radiological Research http://www.cmcr.columbia.edu/ Overview Radiation damage to cells DNA Effects of radiation damage on cells
Basics of Radiation Biology
 Basics of Radiation Biology Sally A. Amundson Columbia University Center for Radiological Research http://www.cmcr.columbia.edu/ Overview Radiation damage to cells DNA Effects of radiation damage on cells
Basics of Radiation Biology Sally A. Amundson Columbia University Center for Radiological Research http://www.cmcr.columbia.edu/ Overview Radiation damage to cells DNA Effects of radiation damage on cells
Shuttling towards a predictive assay for radiotherapy
 Editorial Shuttling towards a predictive assay for radiotherapy Faissal Ouenzar, Michael J. Hendzel, Michael Weinfeld Department of Oncology, University of Alberta, Edmonton, Canada Correspondence to:
Editorial Shuttling towards a predictive assay for radiotherapy Faissal Ouenzar, Michael J. Hendzel, Michael Weinfeld Department of Oncology, University of Alberta, Edmonton, Canada Correspondence to:
Cell cycle control (mammalian)
") Apr. 21, 2005 Cell cycle control (mammalian) Basic mechanisms & protein components Checkpoints Chap. 21, by Lodish et al., 5 th ed. 2004 Chap. 17, by Alberts et al., 4 th ed. 2002 鍾明怡 mychung@vghtpe.gov.tw
Apr. 21, 2005 Cell cycle control (mammalian) Basic mechanisms & protein components Checkpoints Chap. 21, by Lodish et al., 5 th ed. 2004 Chap. 17, by Alberts et al., 4 th ed. 2002 鍾明怡 mychung@vghtpe.gov.tw
Oncolytic virus strategy
 Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
Post-Translational Regulation of the RSF1 Chromatin Remodeler under DNA Damage
 Molecules and Cells Minireview Post-Translational Regulation of the RSF1 Chromatin Remodeler under DNA Damage Sunwoo Min 1,2,3,5, Yong Won Choi 1,4,5, Hansol Yun 1,2,3, Sujin Jo 3, Jae-Hoon Ji 3, and Hyeseong
Molecules and Cells Minireview Post-Translational Regulation of the RSF1 Chromatin Remodeler under DNA Damage Sunwoo Min 1,2,3,5, Yong Won Choi 1,4,5, Hansol Yun 1,2,3, Sujin Jo 3, Jae-Hoon Ji 3, and Hyeseong
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Inactivation of p53 Rescues the Maintenance of High Risk HPV DNA Genomes Deficient in Expression of E6
 Inactivation of p53 Rescues the Maintenance of High Risk HPV DNA Genomes Deficient in Expression of E6 Laurel D. Lorenz, Jessenia Rivera Cardona, Paul F. Lambert* McArdle Laboratory for Cancer Research,
Inactivation of p53 Rescues the Maintenance of High Risk HPV DNA Genomes Deficient in Expression of E6 Laurel D. Lorenz, Jessenia Rivera Cardona, Paul F. Lambert* McArdle Laboratory for Cancer Research,
Roles of ATM and ATR-Mediated DNA Damage Responses during Lytic BK Polyomavirus Infection
 Roles of ATM and ATR-Mediated DNA Damage Responses during Lytic BK Polyomavirus Infection Mengxi Jiang 1, Linbo Zhao 2, Monica Gamez 1, Michael J. Imperiale 1,2 * 1 Department of Microbiology and Immunology
Roles of ATM and ATR-Mediated DNA Damage Responses during Lytic BK Polyomavirus Infection Mengxi Jiang 1, Linbo Zhao 2, Monica Gamez 1, Michael J. Imperiale 1,2 * 1 Department of Microbiology and Immunology
Factors determining DNA double strand break repair pathway choice in G2 phase.
 Supplementary Data Factors determining DN double strand break repair pathway choice in G2 phase. tsushi Shibata 1, Sandro Conrad 2, Julie irraux 1, Verena Geuting 2, Olivia arton 2, mani Ismail 1, ndreas
Supplementary Data Factors determining DN double strand break repair pathway choice in G2 phase. tsushi Shibata 1, Sandro Conrad 2, Julie irraux 1, Verena Geuting 2, Olivia arton 2, mani Ismail 1, ndreas
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
Regulation of cell cycle. Dr. SARRAY Sameh, Ph.D
 Regulation of cell cycle Dr. SARRAY Sameh, Ph.D Control of cell cycle: Checkpoints Are the cell cycle controls mechanisms in eukaryotic cells. These checkpoints verify whether the processes at each phase
Regulation of cell cycle Dr. SARRAY Sameh, Ph.D Control of cell cycle: Checkpoints Are the cell cycle controls mechanisms in eukaryotic cells. These checkpoints verify whether the processes at each phase
Genomic instability. Amin Mahpour
 Genomic instability Amin Mahpour 1 Some questions to ponder What is Genomic instability? What factors contribute to the genomic integrity? How we identify these aberrations? 2 PART I: MOLECULAR BIOLOGY
Genomic instability Amin Mahpour 1 Some questions to ponder What is Genomic instability? What factors contribute to the genomic integrity? How we identify these aberrations? 2 PART I: MOLECULAR BIOLOGY
ATM in focus: A damage sensor and cancer target
 BioDiscovery MONOGRAPH ATM in focus: A damage sensor and cancer target Hilal S Khalil, Hemanth Tummala, Nikolai Zhelev * School of Contemporary Sciences, University of Abertay Dundee, Dundee DD1 1HG, Scotland,
BioDiscovery MONOGRAPH ATM in focus: A damage sensor and cancer target Hilal S Khalil, Hemanth Tummala, Nikolai Zhelev * School of Contemporary Sciences, University of Abertay Dundee, Dundee DD1 1HG, Scotland,
Chapter 12. living /non-living? growth repair renew. Reproduction. Reproduction. living /non-living. fertilized egg (zygote) next chapter
 next chapter") Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
SUPPLEMENTAL FIGURE LEGENDS
 SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
Epstein-Barr Virus Polypeptides: Identification of Early Proteins and Their Synthesis and Glycosylation
 JOURNAL OF VIROLOGY, Aug. 1981, p. 651-655 0022-538X/81/080651-05$02.00/0 Vol. 39, No. 2 Epstein-Barr Virus Polypeptides: Identification of Early Proteins and Their Synthesis and Glycosylation ROBERT J.
JOURNAL OF VIROLOGY, Aug. 1981, p. 651-655 0022-538X/81/080651-05$02.00/0 Vol. 39, No. 2 Epstein-Barr Virus Polypeptides: Identification of Early Proteins and Their Synthesis and Glycosylation ROBERT J.
Kaposi's Sarcoma-Associated Herpesvirus Induces ATM and H2AX. DNA Damage Response Early During de novo Infection of Primary
 JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists. International authors and editors
 We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,900 116,000 120M Open access books available International authors and editors Downloads Our
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,900 116,000 120M Open access books available International authors and editors Downloads Our
Role of genetic testing in familial breast cancer outside of BRCA1 and BRCA2
 Role of genetic testing in familial breast cancer outside of BRCA1 and BRCA2 Introduction Most commonly diagnosed cancer in South African women and the second most commonly diagnosed cancer in Black women
Role of genetic testing in familial breast cancer outside of BRCA1 and BRCA2 Introduction Most commonly diagnosed cancer in South African women and the second most commonly diagnosed cancer in Black women
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Regulation of Cell Division
 Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Open Access Book Chapter
 ERC-OAPEN-2015, Deposit cover page Open Access Book Chapter Published version Chapter title Signalling DNA Damage Title in English Author(s) Andres Joaquin Lopez-Contreras and Oscar Fernandez-Capetillo
ERC-OAPEN-2015, Deposit cover page Open Access Book Chapter Published version Chapter title Signalling DNA Damage Title in English Author(s) Andres Joaquin Lopez-Contreras and Oscar Fernandez-Capetillo
Rabbit Polyclonal antibody to NFkB p65 (v-rel reticuloendotheliosis viral oncogene homolog A (avian))
)") Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Transcription and RNA processing
 Transcription and RNA processing Lecture 7 Biology W3310/4310 Virology Spring 2016 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at
Transcription and RNA processing Lecture 7 Biology W3310/4310 Virology Spring 2016 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Dual role of Nbs1 in the ataxia telangiectasia mutateddependent
 MINIREVIEW Dual role of Nbs1 in the ataxia telangiectasia mutateddependent DNA damage response Joo-Hyeon Lee and Dae-Sik Lim Department of Biological Sciences, Korea Advanced Institute of Science and Technology,
MINIREVIEW Dual role of Nbs1 in the ataxia telangiectasia mutateddependent DNA damage response Joo-Hyeon Lee and Dae-Sik Lim Department of Biological Sciences, Korea Advanced Institute of Science and Technology,
Molecular Cell Biology (Bio 5068) Cell Cycle I. Ron Bose, MD PhD November 14, 2017
 Cell Cycle I. Ron Bose, MD PhD November 14, 2017") Molecular Cell Biology (Bio 5068) Cell Cycle I Ron Bose, MD PhD November 14, 2017 CELL DIVISION CYCLE M G2 S G1 DISCOVERY AND NAMING OF CYCLINS A protein (called cyclin ) was observed to increase as cells
Molecular Cell Biology (Bio 5068) Cell Cycle I Ron Bose, MD PhD November 14, 2017 CELL DIVISION CYCLE M G2 S G1 DISCOVERY AND NAMING OF CYCLINS A protein (called cyclin ) was observed to increase as cells
The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters.
 O[superscript 6]-Methylguanine DNA lesions induce an intra-s-phase arrest from which cells exit into apoptosis governed by early and late multi-pathway signaling The MIT Faculty has made this article openly
O[superscript 6]-Methylguanine DNA lesions induce an intra-s-phase arrest from which cells exit into apoptosis governed by early and late multi-pathway signaling The MIT Faculty has made this article openly
Rishabh Kala 1, Harsh N. Shah 1, Samantha L. Martin 1 and Trygve O. Tollefsbol 1,2,3,4,5*
 Kala et al. BMC Cancer (2015) 15:672 DOI 10.1186/s12885-015-1693-z RESEARCH ARTICLE Open Access Epigenetic-based combinatorial resveratrol and pterostilbene alters DNA damage response by affecting SIRT1
Kala et al. BMC Cancer (2015) 15:672 DOI 10.1186/s12885-015-1693-z RESEARCH ARTICLE Open Access Epigenetic-based combinatorial resveratrol and pterostilbene alters DNA damage response by affecting SIRT1
Ch. 18 Regulation of Gene Expression
 Ch. 18 Regulation of Gene Expression 1 Human genome has around 23,688 genes (Scientific American 2/2006) Essential Questions: How is transcription regulated? How are genes expressed? 2 Bacteria regulate
Ch. 18 Regulation of Gene Expression 1 Human genome has around 23,688 genes (Scientific American 2/2006) Essential Questions: How is transcription regulated? How are genes expressed? 2 Bacteria regulate
Regulation of Cell Division. AP Biology
 Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Eukaryotic Gene Regulation
 Eukaryotic Gene Regulation Chapter 19: Control of Eukaryotic Genome The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different,
Eukaryotic Gene Regulation Chapter 19: Control of Eukaryotic Genome The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different,
DNA Repair 9 (2010) Contents lists available at ScienceDirect. DNA Repair. journal homepage:
 Contents lists available at ScienceDirect. DNA Repair. journal homepage:") DNA Repair 9 (2010) 1219 1228 Contents lists available at ScienceDirect DNA Repair journal homepage: www.elsevier.com/locate/dnarepair Mini-review Assembly and function of DNA double-strand break repair
DNA Repair 9 (2010) 1219 1228 Contents lists available at ScienceDirect DNA Repair journal homepage: www.elsevier.com/locate/dnarepair Mini-review Assembly and function of DNA double-strand break repair
Mutant p53 establishes targetable tumor dependency by promoting unscheduled replication
 Mutant p53 establishes targetable tumor dependency by promoting unscheduled replication Shilpa Singh, 1 Catherine A. Vaughan, 1 Rebecca A. Frum, 2 Steven R. Grossman, 2 Sumitra Deb, 1 and Swati Palit Deb
Mutant p53 establishes targetable tumor dependency by promoting unscheduled replication Shilpa Singh, 1 Catherine A. Vaughan, 1 Rebecca A. Frum, 2 Steven R. Grossman, 2 Sumitra Deb, 1 and Swati Palit Deb
Accumulation of DNA double strand breaks. in normal tissues after fractionated low dose irradiation
 Aus der Klinik für Strahlentherapie und Radioonkologie Direktor: Prof. Dr. Ch. Rübe UniversitätskliniKum des Saarlandes Accumulation of DNA double strand breaks in normal tissues after fractionated low
Aus der Klinik für Strahlentherapie und Radioonkologie Direktor: Prof. Dr. Ch. Rübe UniversitätskliniKum des Saarlandes Accumulation of DNA double strand breaks in normal tissues after fractionated low
Viruses Tomasz Kordula, Ph.D.
 Viruses Tomasz Kordula, Ph.D. Resources: Alberts et al., Molecular Biology of the Cell, pp. 295, 1330, 1431 1433; Lehninger CD Movie A0002201. Learning Objectives: 1. Understand parasitic life cycle of
Viruses Tomasz Kordula, Ph.D. Resources: Alberts et al., Molecular Biology of the Cell, pp. 295, 1330, 1431 1433; Lehninger CD Movie A0002201. Learning Objectives: 1. Understand parasitic life cycle of
Cell Cycle, Mitosis, and Microtubules. LS1A Final Exam Review Friday 1/12/07. Processes occurring during cell cycle
 Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:
 into Metastatic Breast Cancer Cells: Introduction - The Broad Picture:") Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Transformation of Normal HMECs (Human Mammary Epithelial Cells) into Metastatic Breast Cancer Cells: Introduction - The Broad Picture: Spandana Baruah December, 2016 Cancer is defined as: «A disease caused
Cell Damage. Standardized and automatic determination of DNA double strand breaks using immunofluorescence
 Standardized and automatic determination of DNA double strand breaks using immunofluorescence Patents: CN 2702421, EP 2208068, TR 201308237, 200880115751 The Platform Motorized, inverse fluorescence microscope
Standardized and automatic determination of DNA double strand breaks using immunofluorescence Patents: CN 2702421, EP 2208068, TR 201308237, 200880115751 The Platform Motorized, inverse fluorescence microscope
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Importance of ATM in Radiotherapy
 Importance of ATM in Radiotherapy Radiology and Physical Medicine María Trinidad Rueda Cáceres Radiotherapy Importance of ATM in Radiotherapy One of the standard treatment options for various malignant
Importance of ATM in Radiotherapy Radiology and Physical Medicine María Trinidad Rueda Cáceres Radiotherapy Importance of ATM in Radiotherapy One of the standard treatment options for various malignant
The interaction between Rad9 and Tousled-like kinase 1 in the cell cycle and the DNA damage response
 The interaction between Rad9 and Tousled-like kinase 1 in the cell cycle and the DNA damage response By Ryan Patrick Kelly A thesis submitted to the Graduate Program in Biochemistry in conformity with
The interaction between Rad9 and Tousled-like kinase 1 in the cell cycle and the DNA damage response By Ryan Patrick Kelly A thesis submitted to the Graduate Program in Biochemistry in conformity with
Regulation of Cell Division (Ch. 12)
") Regulation of Cell Division (Ch. 12) Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development &
Regulation of Cell Division (Ch. 12) Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development &
Chapter 12. Regulation of Cell Division. AP Biology
 Chapter 12. Regulation of Cell Division Coordination of cell division! Multicellular organism " need to coordinate across different parts of organism! timing of cell division! rates of cell division "
Chapter 12. Regulation of Cell Division Coordination of cell division! Multicellular organism " need to coordinate across different parts of organism! timing of cell division! rates of cell division "
Karyotype analysis reveals transloction of chromosome 22 to 9 in CML chronic myelogenous leukemia has fusion protein Bcr-Abl
 Chapt. 18 Cancer Molecular Biology of Cancer Student Learning Outcomes: Describe cancer diseases in which cells no longer respond Describe how cancers come from genomic mutations (inherited or somatic)
Chapt. 18 Cancer Molecular Biology of Cancer Student Learning Outcomes: Describe cancer diseases in which cells no longer respond Describe how cancers come from genomic mutations (inherited or somatic)
DNA double strand break repair: a radiation perspective
 DNA double strand break repair: a radiation perspective Kavanagh, J. N., Redmond, K. M., Schettino, G., & Prise, K. M. (2013). DNA double strand break repair: a radiation perspective. Antioxidants & Redox
DNA double strand break repair: a radiation perspective Kavanagh, J. N., Redmond, K. M., Schettino, G., & Prise, K. M. (2013). DNA double strand break repair: a radiation perspective. Antioxidants & Redox