Enterotoxin preconditioning restores calciumsensing receptor-mediated cytostasis in colon cancer cells
|
|
|
- Cory Francis
- 5 years ago
- Views:
Transcription

1 Thomas Jefferson University Jefferson Digital Commons Department of Pharmacology and Experimental Therapeutics Faculty Papers Department of Pharmacology and Experimental Therapeutics Enterotoxin preconditioning restores calciumsensing receptor-mediated cytostasis in colon cancer cells Giovanni Mario Pitari Thomas Jefferson University, Jieru E. Lin Thomas Jefferson University Fawad J. Shah Thomas Jefferson University Wilhelm J. Lubbe Thomas Jefferson University David Zuzga Thomas Jefferson University See next page for additional authors Let us know how access to this document benefits you Follow this and additional works at: Part of the Medical Pharmacology Commons Recommended Citation Pitari, Giovanni Mario; Lin, Jieru E.; Shah, Fawad J.; Lubbe, Wilhelm J.; Zuzga, David; Li, Peng; Schulz, Stephanie; and Waldman, Scott A, "Enterotoxin preconditioning restores calcium-sensing receptor-mediated cytostasis in colon cancer cells" (2008). Department of Pharmacology and Experimental Therapeutics Faculty Papers. Paper This Article is brought to you for free and open access by the Jefferson Digital Commons. The Jefferson Digital Commons is a service of Thomas Jefferson University's Center for Teaching and Learning (CTL). The Commons is a showcase for Jefferson books and journals, peer-reviewed scholarly publications, unique historical collections from the University archives, and teaching tools. The Jefferson Digital Commons allows researchers and interested readers anywhere in the world to learn about and keep up to date with Jefferson scholarship. This article has been accepted for inclusion in Department of Pharmacology and Experimental Therapeutics Faculty Papers by an authorized administrator of the Jefferson Digital Commons. For more information, please contact: JeffersonDigitalCommons@jefferson.edu.
2 Authors Giovanni Mario Pitari, Jieru E. Lin, Fawad J. Shah, Wilhelm J. Lubbe, David Zuzga, Peng Li, Stephanie Schulz, and Scott A Waldman This article is available at Jefferson Digital Commons:
3 1 Enterotoxin Preconditioning Restores Calcium-Sensing Receptor- Mediated Cytostasis in Colon Cancer Cells Giovanni M.Pitari, Jieru E.Lin, Fawad J.Shah, Wilhelm J.Lubbe, David Zuzga, Peng Li, Stephanie Schulz, and Scott A.Waldman Department of Pharmacology and Experimental Therapeutics, Thomas Jefferson University, Philadelphia, PA 19107, USA Corresponding Author: Giovanni M. Pitari, MD, PhD, Department of Pharmacology and Experimental Therapeutics, Thomas Jefferson University, 1100 Walnut St., MOB Suite 810, Philadelphia, PA 19107; Tel.: +(1) ; Fax: +(1) ; Running Title: Enterotoxins restore CaR-mediated cytostasis. Keywords: Colorectal cancer; Guanylyl cyclase C; Bacterial enterotoxins; Calciumsensing receptor. Word Counts: Abstract Words: 192; Total Words in Paper: 3,653. Abbreviations: GCC, guanylyl cyclase C; Ca 2+ o, extracellular Ca 2+ ; ETEC, enterotoxigenic E. coli infections; ST, heat-stable enterotoxin; CaR, calcium-sensing receptor; CNG channel, cyclic nucleotide gated channel; PKG, cgmp-dependent protein kinase; SOCs, store-operated Ca 2+ channels.
4 2 ABSTRACT Guanylyl cyclase C (GCC), the receptor for diarrheagenic bacterial enterotoxins (STs), inhibits colorectal cancer cell proliferation by co-opting Ca 2+ as the intracellular messenger. Similarly, extracellular Ca 2+ (Ca 2+ o) opposes proliferation and induces terminal differentiation in intestinal epithelial cells. In that context, human colon cancer cells develop a phenotype characterized by insensitivity to cytostasis imposed by Ca 2+ o. Here, preconditioning with ST, mediated by GCC signaling through cyclic nucleotide-gated channels, restored Ca 2+ o-dependent cytostasis, reflecting post-transcriptional regulation of Ca 2+ o-sensing receptors (CaRs). ST-induced GCC signaling deployed CaRs to the surface of human colon cancer cells, while elimination of GCC signaling in mice nearly abolished CaR expression in enterocytes. Moreover, ST-induced Ca 2+ o-dependent cytostasis was abrogated by CaR-specific antisense oligonucleotides. Importantly, following ST preconditioning, newly expressed CaRs at the cell surface represented tumor cell receptor targets for antiproliferative signaling by CaR agonists. Since expression of the endogenous paracrine hormones for GCC is uniformly lost early in carcinogenesis, these observations offer a mechanistic explanation for the Ca 2+ o-resistant phenotype of colon cancer cells. Restoration of antitumorigenic CaR signaling by GCC ligand replacement therapy represents a previously unrecognized paradigm for the prevention and treatment of human colorectal cancer employing dietary Ca 2+ supplementation.
5 3 INTRODUCTION Colorectal cancer is most prevalent in the developed world, representing the second leading cause of cancer-related mortality (1,2). Although the epidemiology of this disease is poorly understood, there is an unexplained inverse relationship between the worldwide incidence of colorectal cancer and enterotoxigenic E. coli (ETEC) infections (3,4). ETEC produce heat-stable enterotoxins (STs), exogenous ligands for the intestine-specific receptor guanylyl cyclase C (GCC) (5) and a principle cause of secretory diarrhea in endemic populations and animal herds (6-8). Regions of the world with the highest incidence of ETEC-associated diarrhea exhibit the lowest incidence of colon cancer (4). In that context, activation of GCC inhibits human colon carcinoma cell proliferation (4,9,10) and adenoma formation in mice (3). Reduced expression of the endogenous paracrine hormones for GCC, guanylin and uroguanylin, represents an early mutational event in colorectal carcinogenesis (11-13). GCC signaling through its second messenger cgmp promotes fluid and electrolyte secretion (5), opposes cell cycle progression and proliferation (4,9), and regulates migration, differentiation and apoptosis along the crypt-villus axis (14,15). Importantly, targeted GCC deletion (GCC -/- ) in mice increased intestinal tumorigenesis induced by the carcinogen azoxymethane or APC mutations by corrupting homeostatic crypt proliferation and genomic integrity (16). These observations suggest a model in which colorectal cancer is a disease of hormone insufficiency where dysregulation of GCC signaling, following loss of guanylin and uroguanylin, promotes tumorigenesis by disrupting mucosal homeostasis (16). However, beyond production of cgmp, molecular mechanisms by which GCC regulates processes underlying carcinogenesis remain undefined.
6 4 Like GCC, extracellular Ca 2+ (Ca 2+ o) opposes proliferation and promotes differentiation of intestinal mucosa cells (17-20). Moreover, Ca 2+ supplementation abrogates intestinal hyperproliferation and tumor formation induced by a Western-style diet in APC Min/+ mice (21). In part, these antitumorigenic effects may reflect GCC-induced activation of cyclic nucleotide-gated (CNG) channels, inducing cytostasis through Ca 2+ o influx (4). Notably, Ca 2+ o in the colonic lumen may increase to 20 mm which activates calcium-sensing receptors (CaRs) (19,22), G protein-coupled receptors (23) expressed in apical membranes of colonocytes (24), an event resulting in reduced proliferation and tumorigenesis through inhibition of β-catenin/tcf-4 signaling, and increased cell maturation through p21 and p27 activities (25,26). Conversely, Ca 2+ o supports colorectal cancer cell proliferation by capacitative entry through store-operated Ca 2+ channels (SOCs) (27,28), which opposes antiproliferative GCC signaling through CNG and Ca 2+ o entry (28). Thus, opposing mechanisms regulating proliferative balance by Ca 2+ o comprise functional units reciprocally orchestrated in colon cancer cells (28). Here, the functional relationship between GCC and Ca 2+ o was explored in human colon carcinoma cells preconditioned with ST, in the context of a dynamic range of Ca 2+ o, and in GCC -/- mice. These studies revealed a previously unappreciated role for GCC in regulating CaR signaling, offering a novel paradigm for the prevention and treatment of human colorectal cancer employing hormone replacement therapy with GCC ligands in combination with oral Ca 2+ supplementation. MATERIALS AND METHODS Tumor Cell Proliferation. Proliferation of cancer cells was quantified in 96 well/plates by [methyl- 3 H]thymidine (0.2 µci/well) incorporation into DNA (9). Cells were pulse-labeled
7 5 (3 h) with 3 H-thymidine at the end of 24 h periods of proliferation induced by 10 mm L- glutamine. Following 3 H-thymidine labeling, media was aspirated, cells were incubated for 15 min with ice-cold 10% TCA and rinsed sequentially with 10% TCA and 100% methanol. The acid-insoluble material containing 3 H-labeled DNA was solubilized in 100 µl of 0.2 N NaOH, 80 µl aliquots were dissolved in 1 ml ScintiVerse and radioactivity quantified in a Packard β-scintillation spectrometer. All experiments were conducted on exponentially growing tumor cells. Tumor Cell Toxicity. Cytotoxicity, including occurrence of apoptosis or necrosis, was assessed by flow cytometry (9). Cancer cells (~10 6 cells per well in 6-well plates) were treated (24 h) with the indicated reagents. Then, cells were placed in suspension by trypsinization, pelleted by centrifugation, washed with PBS, and fixed in ice-cold 75% ethanol (30 min). After another wash with PBS, cells were resuspended in the staining solution (50 µg/ml propidium iodide, 100 µg/ml RNase A, 1 mm EDTA and 0.1% Triton X-100), and analyzed on a Coulter EPICS XL-MCL flow cytometer. Distribution in different phases of the cell cycle was analyzed using WinMDI software (version 2.8) provided by Joseph Trotter, Scripps Research Institute (La Jolla, CA). Twenty thousand cells, cleared from doublets, were analyzed from each sample. Cyclic GMP Assay. GCC-induced intracellular cgmp accumulation was assessed after treating cancer cells in triplicate with ST (15 min in 6 well/plates) employing Eagle s minimal essential medium (EMEM) supplemented with 2 mm L-glutamine. Reactions were terminated by adding ice-cold 100% ethanol, each well was washed twice with icecold 100% ethanol, and supernates separated from pellets by centrifugation (12,000 g, 15 min at 4 o C). Supernates containing cgmp were evaporated in a Savant SVC-100H
8 6 concentrator (Thermo Electron Corporation, Waltham, MA) and reconstituted with 50 mm sodium acetate (ph 4.0), and cgmp was quantified in each sample in triplicate by radioimmunoassay (4). Immunoblot analysis. Proteins from total cell lysates, cytosol or membrane extracts (28) prepared in SDS sample buffer were separated by electrophoresis on SDS-PAGE, transferred on nitrocellulose membranes, and then probed with rabbit polyclonal antibodies directed against CaR (dilution, 1:1,000) or human GAPDH (1:1,000), or with goat antivillin antibody [Villin(C-19); 1:1,000] in TBS-Tween (5% milk) overnight at 4 C. Then, membranes were probed with HRP-conjugated secondary antibody (dilution, 1:5,000) for 1 h at room temperature, and specific bands were visualized employing West Pico Chemiluminescent Substrate and subjected to densitometry. Immunoblots, performed under reducing conditions, exhibited only 1 specific band of ~125 kda corresponding to the monomer form of CaR. Immunostaining Analyses. For immunohistochemistry, tumor cells were washed 2 times with cold PBS and immediately fixed (30 min in 4% paraformaldehyde) at room temperature followed by quenching with 3% H 2 O 2. CaR or CD104 at the cell surface were visualized employing (overnight at 4 o C) rabbit anti-car (1:100) or mouse monoclonal anti-human CD104 (1:100), respectively, and the Histostain-plus kit. For immunofluorescence, GCC -/- mice, generated by neomycin-resistant gene insertion on an I- 129 background (29) and backcrossed with a C57BL/6 strain for 7 generations, were employed. All animals were treated in compliance with the National Institute of Health Guidelines for the Care and Use of Laboratory Animals and under a protocol approved by the Institutional Animal Care and Use Committee of Thomas Jefferson University. GCC +/+
9 7 and GCC -/- littermates (males, 2 months old; N, 3 per genotype) were sacrificed, the intestine collected, divided into anatomically comparable segments and processed for paraffin-based archiving. Paraffin embedded tissue sections (5 μm) were rehydrated, heated (2 times for 5 min in citrate buffer, ph 6) for antigen retrieval, incubated overnight (4 o C) with rabbit anti-car (1:100) or goat anti-villin (1:50) followed by incubation for 30 min (room temperature) with Alexa fluor 555 anti-rabbit IgG (for CaR) or Alexa fluor 488 anti-goat IgG (for villin). Digital images were acquired by computers attached to a light microscope or a confocal laser scanning microscope (LSM510, Carl Zeiss, Jena, Germany). Cell Transductions. A bp CaR fragment containing 8 bp of upstream DNA was cloned into MSCV-puro retroviral vectors in both sense and antisense orientations (30), confirmed by DNA sequencing. Antisense, sense, and empty vectors were transfected along with the packaging vector pcl Ampho into HEK 293 cells employing Fugene. Viral supernatants harvested at 48 and 72 h after transfection were used to transduce T84 cells (for 72 h). Then, tumor cells stably transduced with CaR antisense, CaR sense or empty vector were selected employing 5μg/ml puromycin in DMEM/F12. Statistical analysis. Data are expressed as the mean ± SEM of a representative of at least three experiments performed in triplicate. Data were analyzed employing the unpaired twotailed Student s t-test, and significance was assumed for p<0.05. RESULTS Colorectal Cancer Cells are Insensitive to Ca 2+ o-dependent Cytostasis. While intestinal epithelial cells along the crypt-villus axis transition from proliferation to differentiation at lumenal Ca 2+ o ~2-3 mm (22), neoplastic transformation progressively desensitizes these
10 8 cells to the cytostatic effects of Ca 2+ o (20), with ultimate loss of Ca 2+ o-induced cell cycle arrest (31). Accordingly, Ca 2+ o induced cytostasis (Figure 1A), but not cytotoxicity (Figure 1B), in moderately and well-differentiated colon cancer cells. However, Ca 2+ o failed to induce cell cycle arrest and a substantial proliferating cell population (~20-30%) persisted at 50 mm Ca 2+ o (Figure 1A), consistent with a phenotype of insensitivity to proliferative restriction by Ca 2+ o in human colon cancer cells. ST Potentiates Ca 2+ o-dependent Cytostasis by Inducing GCC Signaling Through CNG Channels. Acute (3 h) ST exposure, which restricts cell cycle progression of colon cancer cells by activating GCC signaling (4,9), induced cytostasis which was additive with Ca 2+ o (Figure 1C1). Indeed, cytostasis induced by acute ST exposure was not altered by Ca 2+ o over a range of 1-50 mm (Figure 1C2), suggesting that the antiproliferative effects of ST and Ca 2+ o are mediated by distinct, non-interfering mechanisms. In contrast, ST preconditioning (24 h), which induces desensitization to GCC-dependent cytostasis in colon cancer cells (10), potentiated the inhibition of proliferation by Ca 2+ o (Figure 1C1). Thus, ST synergistically increased the potency (IC 50 : Ca 2+ o alone, 9.91 ± 1.09 mm; ST plus Ca 2+ o, 6.49 ± 1.13 mm) and efficacy (mean % proliferation: Ca 2+ o alone, ± 1.84; ST plus Ca 2+ o, ± 2.14) of Ca 2+ o to inhibit colon cancer cell proliferation (p < 0.001; Figure 1C1). Importantly, while failing to inhibit cell proliferation, reflecting desensitization to cgmp signaling (10) (see conditions at 1 mm Ca 2+ o in Figure 1C1), ST preconditioning revealed a novel antiproliferative mechanism which required 3 mm Ca 2+ o (Figure 1C2). Potentiation of Ca 2+ o-induced cytostasis by ST preconditioning was mediated by GCC, since ST, but not an inactive analog, inhibited (IC 50, ± 3.21 nm; Figure 2A)
11 9 proliferation of human colon carcinoma cells expressing GCC, but not of cells lacking GCC (32) (Figure 2B). Similarly, the effects of ST preconditioning were greater in T84 cells, which exhibits the highest GCC expression (32), compared to Caco-2 cells (Figure 2B). Further, two membrane-permeant cgmp analogs mimicked (Figure 2B), whereas an inhibitor of cyclic nucleotide-hydrolyzing phosphodiesterases enhanced (Figure 2C), the effects of ST preconditioning on tumor cell proliferation. Conversely, two inhibitors of CNG channels and an intracellular Ca 2+ chelator, but not inhibitors of PKG or campdependent protein kinase, blocked ST-mediated potentiation of Ca 2+ o-dependent cytostasis (Figure 2C). Notably, ST did not induce cytotoxicity in colon cancer cells exposed to high Ca 2+ o (Figure 2D). Rather, ST delayed the progression of these tumor cells through the cell cycle (Figure 2D), consistent with the notion that GCC agonists are cytostatic agents for colon cancer (4,9,10). Together, these observations suggest that in colon cancer cells induction of Ca 2+ o-dependent cytostasis by ST preconditioning is mediated by cgmpdependent activation of CNG channels and Ca 2+ o entry, the same effector mechanism mediating GCC-induced cell cycle delay (4). GCC Regulates the Function of CaRs in Normal and Malignant Colonocytes. Increased antiproliferative effects of Ca 2+ o following ST preconditioning did not reflect enhanced GCC-dependent cytostasis by Ca 2+ o, because increased Ca 2+ o did not enhance cytostasis induced by GCC signaling (+ST for 3 h in Figure 1C2) and did not prevent GCC-mediated desensitization in cgmp signaling (10) (Figure 3A). Rather, ST preconditioning enhanced Ca 2+ o-dependent cytostasis by coupling increased Ca 2+ o with intracellular signaling mechanisms induced by cgmp signaling (Figure 2A-C). Ca 2+ o induces cytostasis in colonocytes, in part, by activating CaR signaling (22), a mechanism
12 10 typically lost during neoplastic transformation (26,33). ST preconditioning ( 3 h) augmented the complement of total cellular (Supplementary Figure 1A) and membranebound CaRs (Figure 3B) without significantly altering cytosolic CaRs (Figure 3B) or CaR mrna levels (data not shown). Further, examination of specific cell fractions (Supplementary Figure 1B) revealed that ST treatment (3 h) induces ~80 fold increase of CaR at tumor cell membranes (Supplementary Figure 1C), reflecting translocation of ~60% of the total complement of CaR protein in cancer cells from the cytosol to the membrane compartment (Supplementary Figure 1D). Importantly, GCC signal deprivation, either in ligand-free tumor cells (Figure 4, In Vitro panels on the left showing CaR surface staining of T84 cells) or in enterocytes of GCC -/- mice (Figure 4, In Vivo panels on the right), reduced CaRs in cell membranes. Induction of GCC signaling with bacterial enterotoxin in tumor cells or endogenous paracrine hormonal circuitry in enterocytes of GCC +/+ mice, in turn, increased the complement of CaRs at the cell surface (Figure 4). Effects of ST preconditioning on CaR staining at the surface of cancer cells were selective (Supplementary Figure 2), independent of elevated Ca 2+ o but dependent upon cgmp signaling through CNG (Figure 4). Moreover, compared to GCC +/+ mice, CaR expression in brush border membranes of GCC -/- enterocytes was uniformly attenuated along the entire rostrocaudal axis of the intestine in a GCC-specific fashion, since the expression of a marker for absorptive cells, villin, remained unchanged in the same anatomical locations (Figure 4, In Vivo panels showing representative sections from the jejunum). Thus, GCC signaling physiologically regulates post-transcriptional expression of CaRs in enterocytes, and ST preconditioning induces cell surface translocation of CaR receptors in human colon cancer cells.
13 11 ST Preconditioning, with Restoration of CaR Signaling, is a Novel Cytostatic Therapeutic Strategy in Colon Cancer Cells. To explore whether post-transcriptional regulation of CaRs mediates Ca 2+ o-dependent antiproliferation induced by ST preconditioning, CaR expression in colon cancer cells was eliminated employing specific antisense constructs (30) (Supplementary Figure 3; ~70% inhibition of CaR expression by densitometry, normalized to the respective GAPDH, in CaR antisense-transfected cells compared to CaR sense controls). While the response of tumor cells to Ca 2+ o was not altered by CaR antisense (Figure 5A), reflecting lack of effective CaR signaling in colon cancer cells, CaR antisense eliminated the response to Ca 2+ o of cancer cells preconditioned with ST (Figure 5B). Further, ST preconditioning of tumor cells produced a novel sensitivity of DNA synthesis to acute inhibition by 10 mm, but not 1 mm, Ca 2+ o (Supplementary Figure 4), consistent with membrane translocation of low-affinity CaRs which typically sense 3-10 mm Ca 2+ o (34). Moreover, beyond Ca 2+ o, other CaR agonists, including Mg 2+, Gd 3+ and spermine selectively inhibited proliferation of cancer cells preconditioned by ST (Figure 5C). Thus, ST preconditioning restores the sensitivity of human colon cancer cells to Ca 2+ o-dependent antiproliferation through CaRs, suggesting GCC-induced CaRs as previously unrecognized therapeutic targets for cytostatic strategies in patients with colorectal cancer. DISCUSSION Enterotoxins STs (18 amino-acid long) and endogenous paracrine hormones guanylin and uroguanylin (15/16 amino-acid long) are structurally homologous peptides (35,36), which specifically bind to and activate GCC at enterocyte brush-border membranes inducing PKG-dependent ion channel currents (36). In this way, endogenous hormones regulate
14 12 body fluid homeostasis, while STs, principle diarrheagenic agents maximally activating that pathway, permit enterotoxigenic bacteria to propagate in the environment (7,36-38). Of significance, GCC -/- mice exhibit increased proliferation, migration and apoptosis, and decreased differentiation along the crypt-villus axis (14), and are more susceptible to intestinal tumorigenesis induced by azoxymethane or APC mutations (16). Moreover, expression of guanylin and uroguanylin are uniformly lost early in intestinal carcinogenesis (11-13), loss of guanylin expression resulted in crypt hyperproliferation (15), and administration of GCC ligands inhibits intestinal tumor formation in APC min/+ mice (3) and cell cycle progression of colon cancer cells (9). Thus beyond fluid homeostasis, GCC controls proliferation and tumorigenesis in intestine. Mechanisms mediating antiproliferation by GCC appear diverse. GCC signaling through unidentified effectors engages colonic cell quality control mechanisms assuring genomic integrity and tumor suppression (16). Also, GCC- and cgmp-dependent Ca 2+ influx through CNGs induces colon cancer cell cytostasis by reducing nuclear rates of DNA synthesis (4,9). Here, a previously unappreciated mechanism contributing to tumor suppression by GCC is described (Figure 5D). ST preconditioning induces cgmpmediated CNG signaling, and synthesis and cell surface delivery of CaRs. Increased CaRs at the plasma membrane, in turn, provide new tumor-specific targets for antiproliferative signaling by CaR agonists, including Ca 2+ o, Mg 2+, Gd 3+ and spermine. Although mediated by the same proximal effector, Ca 2+ o influx through CNG, inhibition of DNA synthesis (4) and activation of CaR signaling (Figure 5D) represent two distinct, non-overlapping GCCdependent antiproliferative mechanisms, distinguished by acute (minutes) (9) and chronic ( 3 hr Figure 3B) kinetics, respectively. They may represent sequential temporal arms of
15 13 an integrated antiproliferative strategy in which acute cytostasis, silenced by phosphodiesterase-dependent desensitization of cgmp signaling (10), is propagated by CaR-dependent signaling to ensure enduring colon tumor cytostasis imposed by GCC. Interestingly, CaR signaling also promotes phosphodiesterase-dependent hydrolysis of cgmp (39), a mechanism that may contribute to desensitize cgmp-mediated inhibition of DNA synthesis and represents a negative feedback loop for cgmp-induced CaR. First discovered in parathyroid cells where it senses blood Ca 2+ levels and regulates parathyroid hormone release (40), CaRs are expressed in many cell types, including osteoclasts, neurons and hematopoietic cells (23,41,42). In colonocytes, CaRs sense 1-10 mm Ca 2+ o in fecal water by interacting with the amino-terminal extracellular domain, inducing intracellular signaling through heterotrimeric G proteins (23,43). One emerging function of CaRs in colonocytes is the regulation of proliferation and differentiation and its putative role as a tumor suppressor (22,24,44). Although the CaR gene is not lost or mutated in colon cancer (33), CaR expression is inexplicably reduced during disease progression (25,26,33). The present finding that GCC regulates the post-transcriptional expression of CaRs in normal and malignant intestinal cells offers a mechanistic explanation for that observation, since GCC signaling is silenced early during colorectal tumorigenesis following loss of ligand expression (11-13). Accordingly, elimination of GCC signaling in mice abolished the expression of CaRs in enterocytes (Figure 4). Moreover, CaRs are primarily expressed in the apex and central regions of human crypts (26) where expression of endogenous GCC ligands and cgmp levels are greatest (45,46), suggesting that CaR signaling may be conditionally regulated along the crypt-villus axis by GCC and cgmp-dependent mechanisms. In this model, the common abilities of CaRs and
16 14 GCC to promote the transition from proliferation to differentiation and inhibit intestinal carcinogenesis may represent convergent, rather than parallel, signaling pathway(s). Finally, although they remain to be fully characterized, GCC effects on CaRs, including induction of translation, post-translational processing or trafficking to cell membranes, represent a previously unrecognized mechanism of transcriptionally-independent regulation of CaR surface expression and signaling (47). Beyond CaR, Ca 2+ o opposes tumorigenesis by forming benign insoluble complexes with toxic ionized fatty acids and bile acids (48), and promoting cis-dimerization of E-cadherin molecules underlying cell-cell adhesion (49) and growth suppression (50). This latter mechanism, which occurs at Ca 2+ o >0.5 mm (49) and induces the function of p27 (50), may explain, in part, the cytostatic effects of high Ca 2+ o in colon cancer cells not exposed to ST observed here. Indeed, these cells were not affected by CaR antisense delivery (Figure 5A) or application of CaR agonists (Figure 5C), indicating that in the absence of GCC activation CaRs is functionally silent in colon cancer cells. Loss of GCC (11-13) and CaR (25,26,33) signaling, in turn, may underlie the resistant phenotype of colon cancer cells to Ca 2+ o-induced cytostasis (20,31). These observations are significant since levels of 3-10 mm Ca 2+ o, required for significant CaR activation (34), are typically achieved in the fecal colonic water as a result of dietary Ca 2+ intake, absorption, and secretion (19,22). Moreover, allosteric CaR activators present in the intestinal lumen such as L-amino-acids (23) may further increase the CaR sensitivity for Ca 2+ o. Importantly, CaRs sense other polyvalent cations (i.e., Gd 3+, Mg 2+, Ni 2+, polylysine), including polyamines (spermine, spermidine and putrescine) endogenously produced by colonic bacteria (23), suggesting that GCC-induced CaRs may subserve diverse tumor suppressor pathways in intestine.
17 15 Strategies for cancer control include chemoprevention and chemotherapy. In contrast to its endogenous ligands, GCC is universally over-expressed in colorectal tumors (51,52), and oral administration of GCC ligands is a novel targeted approach to prevent tumor initiation and arrest disease progression in patients (3,4,9). Limitations to these therapies include adverse effects associated with the use of diarrheagenic bacterial enterotoxins (7,37,38). Conversely, dietary Ca 2+ supplementation has been proposed as a chemoprevention strategy against colon cancer (53). However, caution has been suggested based on the reduced efficacy of Ca 2+ o to inhibit intestinal cell proliferation following neoplastic transformation, likely reflecting reduced CaR expression (33). In that context, dietary Ca 2+ o could potentially promote growth of Ca 2+ o-insensitive tumor cells while suppressing proliferation of normal adjacent cells, which retain Ca 2+ o sensitivity (22). The present observations that GCC preconditioning restores antitumorigenic CaR signaling in human colon carcinoma cells may offer a solution to the limitations of Ca 2+ o supplementation strategies. Oral Ca 2+ o therapy, in turn, may prevent adverse effects of GCC-targeted strategies because activation of CaR opposes secretory diarrhea by bacterial enterotoxins (39). Taken together, these data suggest that combinatorial therapies including dietary Ca 2+ o supplementation and GCC ligand replacement represent a previously unrecognized paradigm for the prevention and treatment of human colorectal cancer. FUNDING National Institutes of Health (CA75123, CA95026) and Targeted Diagnostic and Therapeutics Inc. to S.A.W., the Pennsylvania Department of Health and Prevent Cancer Foundation to G.M.P., and a National Institutes of Health Institutional Training Award (5T32 CA09662) to W.J.L. The Pennsylvania Department of Health specifically disclaims
18 16 responsibility for any analyses, interpretations or conclusions. SUPPLEMENTARY MATERIAL Supplementary information is available at the Carcinogenesis s website. ACKNOWLEDGMENTS S.A.W. is the Samuel M.V. Hamilton Endowed Professor of Thomas Jefferson University.
19 17 REFERENCES 1. Jemal,A. et al. (2004) Cancer statistics, C.A. Cancer J. Clin., 54, Descriptive Epidemiology Group (2004) GLOBOCAN 2002 (International Agency for Research on Cancer, Lion). 3. Shailubhai,K. et al. (2000) Uroguanylin treatment suppresses polyp formation in the Apc(Min/+) mouse and induces apoptosis in human colon adenocarcinoma cells via cyclic GMP. Cancer Res., 60, Pitari,G.M. et al. (2003) Bacterial enterotoxins are associated with resistance to colon cancer. Proc. Natl Acad. Sci. U S A, 100, Schulz,S. et al. (1990) Guanylyl cyclase is a heat-stable enterotoxin receptor. Cell, 63, Hughes,J.M. et al. (1978) Role of cyclic GMP in the action of heat-stable enterotoxin of Escherichia coli. Nature, 271, Guarino,A. et al. (1987) Small and large intestinal guanylate cyclase activity in children: effect of age and stimulation by Escherichia coli heat-stable enterotoxin. Pediatr. Res., 21, Giannella,R.A. (1995) Escherichia coli heat-stable enterotoxins, guanylins, and their receptors: what are they and what do they do? J. Lab. Clin. Med., 125, Pitari,G.M. et al. (2001) Guanylyl cyclase C agonists regulate progression through the cell cycle of human colon carcinoma cells. Proc. Natl Acad. Sci. U S A, 98,
20 Pitari,G.M. et al. (2005) Interruption of homologous desensitization in cyclic guanosine 3',5'-monophosphate signaling restores colon cancer cytostasis by bacterial enterotoxins. Cancer Res., 65, Cohen,M.B. et al. (1998) Guanylin mrna expression in human intestine and colorectal adenocarcinoma. Lab. Invest., 78, Notterman,D.A. et al. (2001) Transcriptional gene expression profiles of colorectal adenoma, adenocarcinoma, and normal tissue examined by oligonucleotide arrays. Cancer Res., 61, Birkenkamp-Demtroder,K. et al. (2002) Gene expression in colorectal cancer. Cancer Res., 62, Li,P. et al. (2007) Homeostatic regulation of the crypt-to-villus axis by the bacterial enterotoxin receptor guanylyl cyclase C restricts the proliferating compartment in intestine. Am. J. Pathol., 171, Steinbrecher,K.A. et al. (2002) Targeted inactivation of the mouse guanylin gene results in altered dynamics of colonic epithelial proliferation. Am. J. Pathol., 161, Li,P. et al. (2007) Guanylyl cyclase C suppresses intestinal tumorigenesis by restricting proliferation and maintaining genomic integrity. Gastroenterology, 133, Lipkin,M. et al. (1995) Calcium and the prevention of colon cancer. J. Cell. Biochem. Suppl., 22, Lipkin,M. (1999) Preclinical and early human studies of calcium and colon cancer prevention. Ann. N.Y. Acad. Sci., 889,
21 Whitfield,J.F. (1992) Calcium signals and cancer. Crit. Rev. Oncog., 3, Buras,R.R. et al. (1995) The effect of extracellular calcium on colonocytes: evidence for differential responsiveness based upon degree of cell differentiation. Cell Prolif., 28, Newmark,H.L. et al. (2001) A Western-style diet induces benign and malignant neoplasms in the colon of normal C57Bl/6 mice. Carcinogenesis, 22, Whitfield,J.F. et al. (1995) Calcium-cell cycle regulator, differentiator, killer, chemopreventor, and maybe, tumor promoter. J. Cell. Biochem. Suppl., 22, Hofer,A.M. et al. (2003) Extracellular calcium sensing and signalling. Nat. Rev. Mol. Cell Biol., 4, Kallay,E. et al. (2000) Dietary calcium and growth modulation of human colon cancer cells: role of the extracellular calcium-sensing receptor. Cancer Detect. Prev., 24, Chakrabarty,S. et al. (2003) Extracellular calcium and calcium sensing receptor function in human colon carcinomas: promotion of E-cadherin expression and suppression of beta-catenin/tcf activation. Cancer Res., 63, Chakrabarty,S. et al. (2005) Calcium sensing receptor in human colon carcinoma: interaction with Ca(2+) and 1,25-dihydroxyvitamin D(3). Cancer Res., 65, Weiss,H. et al. (2001) Inhibition of store-operated calcium entry contributes to the anti-proliferative effect of non-steroidal anti-inflammatory drugs in human colon cancer cells. Int. J. Cancer, 92,
22 Kazerounian,S. et al. (2005) Proliferative Signaling by Store-Operated Calcium Channels Opposes Colon Cancer Cell Cytostasis Induced By Bacterial Enterotoxins. J. Pharmacol. Exp. Ther., 314, Schulz,S. et al. (1997) Disruption of the guanylyl cyclase-c gene leads to a paradoxical phenotype of viable but heat-stable enterotoxin-resistant mice. J. Clin. Invest., 100, Yamauchi,M. et al. (2005) Involvement of calcium-sensing receptor in osteoblastic differentiation of mouse MC3T3-E1 cells. Am. J. Physiol. Endocrinol. Metab., 288, E Buset,M. et al. (1986) Inhibition of human colonic epithelial cell proliferation in vivo and in vitro by calcium. Cancer Res., 46, Waldman,S.A. et al. (1998) Heterogeneity of guanylyl cyclase C expressed by human colorectal cancer cell lines in vitro. Cancer Epidemiol. Biomarkers Prev., 7, Kallay,E. et al. (2003) Molecular and functional characterization of the extracellular calcium-sensing receptor in human colon cancer cells. Oncol. Res., 13, Garrett,J.E. et al. (1995) Molecular cloning and functional expression of human parathyroid calcium receptor cdnas. J. Biol. Chem., 270, Forte,L.R. (1999) Guanylin regulatory peptides: structures, biological activities mediated by cyclic GMP and pathobiology. Regul. Pept., 81, Lucas,K.A. et al. (2000) Guanylyl cyclases and signaling by cyclic GMP. Pharmacol. Rev., 52,
23 Acosta-Martinez,F. et al. (1980) Escherichia coli heat-stable enterotoxin in feces and intestines of calves with diarrhea. Am. J. Vet. Res., 41, Haberberger,R.L., Jr. et al. (1991) Enteritis due to multiresistant enteroadherent Escherichia coli. Lancet, 337, Geibel,J. et al. (2006) Calcium-sensing receptor abrogates secretagogue- induced increases in intestinal net fluid secretion by enhancing cyclic nucleotide destruction. Proc. Natl Acad. Sci. U S A, 103, Brown,E.M. et al. (1993) Cloning and characterization of an extracellular Ca(2+)- sensing receptor from bovine parathyroid. Nature, 366, House,M.G. et al. (1997) Expression of an extracellular calcium-sensing receptor in human and mouse bone marrow cells. J. Bone Miner. Res., 12, Chattopadhyay,N. et al. (2000) Cellular "sensing" of extracellular calcium (Ca(2+)(o)): emerging roles in regulating diverse physiological functions. Cell Signal., 12, Gama,L. et al. (1997) Ca2+-sensing receptors in intestinal epithelium. Am. J. Physiol., 273, C Lamprecht,S.A. et al. (2003) Chemoprevention of colon cancer by calcium, vitamin D and folate: molecular mechanisms. Nat. Rev. Cancer, 3, Cohen,M.B. et al. (1995) Immunohistochemical localization of guanylin in the rat small intestine and colon. Biochem. Biophys. Res. Commun., 209, Whitaker,T.L. et al. (1997) Uroguanylin and guanylin: distinct but overlapping patterns of messenger RNA expression in mouse intestine. Gastroenterology, 113,
24 Bouschet,T. et al. (2005) Receptor-activity-modifying proteins are required for forward trafficking of the calcium-sensing receptor to the plasma membrane. J. Cell. Sci., 118, Newmark,H.L. et al. (1984) Colon cancer and dietary fat, phosphate, and calcium: a hypothesis. J. Natl Cancer Inst., 72, Pertz,O. et al. (1999) A new crystal structure, Ca2+ dependence and mutational analysis reveal molecular details of E-cadherin homoassociation. Embo J., 18, St Croix,B. et al. (1998) E-Cadherin-dependent growth suppression is mediated by the cyclin-dependent kinase inhibitor p27(kip1). J. Cell Biol., 142, Witek,M.E. et al. (2005) The putative tumor suppressor Cdx2 is overexpressed by human colorectal adenocarcinomas. Clin. Cancer Res., 11, Schulz,S. et al. (2006) A validated quantitative assay to detect occult micrometastases by reverse transcriptase-polymerase chain reaction of guanylyl cyclase C in patients with colorectal cancer. Clin. Cancer Res., 12, Cho,E. et al. (2004) Dairy foods, calcium, and colorectal cancer: a pooled analysis of 10 cohort studies. J. Natl Cancer Inst., 96,
25 23 LEGENDS TO FIGURES Fig. 1. Bacterial enterotoxin ST potentiates Ca 2+ o mediated cytostasis in colorectal cancer cells. (A) Dose responses of cytostasis by Ca 2+ o in various human colon carcinoma cells., P < 0.01, versus control for Caco-2 cells; #, P < 0.05 and ##, P < 0.01, versus respective controls in each of the three cell lines. (B) Absence of cytotoxicity by Ca 2+ o in human colon cancer cells. Cytotoxicity was assessed as the proportion of tumor cells identified by flow cytometry in the sub-g 1 fractions, which correspond to cells undergoing apoptosis or necrosis, of the cell cycle. Data are mean ± SEM from 3 independent experiments. (C1) Concentration dependence of T84 cell proliferation on Ca 2+ o in the presence or absence of 1 μm ST. Results in (A) and (C1) are expressed as the percentage of respective control incubations treated with 1 mm Ca 2+ o. *, p<0.05 and **, P < 0.01, comparing respective conditions treated with Ca 2+ o alone and Ca 2+ o plus ST for 24 h. In (C2), data obtained in (C1) are expressed as {100-[(condition treated with ST plus Ca 2+ o)/(respective control condition treated with Ca 2+ o alone) x 100]}. Fig. 2. ST potentiates Ca 2+ o mediated cytostasis by inducing GCC signaling through CNG. (A) Dose response of ST-induced potentiation of antiproliferation by Ca 2+ o in T84 cells. (B) ST (1 μm) actions on Ca 2+ o are mediated by GCC and cgmp in colorectal cancer cells. TJU, 1 μm of the inactive ST analog TJU 1-103; 8-br-cGMP (5 mm) and 8-pCPTcGMP (0.5 mm), membrane-permeant cgmp analogues. (C) CNG channel and Ca 2+ o entry are the proximal molecular effectors of enhanced Ca 2+ o cytostasis by ST (1 μm) in T84 cells. IBMX (50 μm), the general phosphodiesterase inhibitor 3-isobutyl-1- methylxanthine; L-DLT (L-cis-diltiazem, 10 μm) and PET (8-br-PET-cGMPS, 50 μm), two inhibitors of CNG channels; BAPTA (2 μm), the intracellular Ca 2+ chelator BAPTA-
26 24 AM; RP8pCPT (50 μm), the PKG inhibitor Rp-8-pCPT-cGMP; Rp-cAMPS (50 μm), an inhibitor of camp-dependent protein kinase. Inhibitors were employed at concentrations that completely inhibit their target enzymes (4). Results in (A) are expressed as the percentage of respective control incubations treated with 10 mm Ca 2+ o, while in (B) and (C) are expressed as in Figure 1c2. *, p<0.05 and **, P < 0.01, versus respective parallel control conditions treated with 10 mm Ca 2+ o. (D) ST (1 μm) induces cytostasis, but not cytotoxicity, in synchronized T84 cells exposed to 10 mm Ca 2+ o. The percentage of tumor cells in each phase of the cell cycle, including the sub-g 1 fraction of apoptotic/necrotic cells, was quantified by flow cytometry. Data are from a representative experiment. Fig. 3. ST induces CaR translocation in colon cancer cells. (A) Intracellular cgmp accumulation induced by ST (1 μm for 15 min) in T84 cells pre-treated with ST (1 μm for 24 h) or the vehicle control in the presence of the indicated Ca 2+ o concentration. *, p<0.05, versus respective control conditions not pre-treated with ST. (B) Time-course of ST (1 μm) effects on cytosol (upper panel) or membrane (lower panel) CaR expression in T84 cells. PBS, the vehicle control; GAPDH and villin, the loading controls. Fig. 4. Ligand-dependent GCC signaling upregulates surface CaR receptors in normal and transformed intestinal epithelial cells. Membranes of non-permeabilized T84 cells (In Vitro) were stained (brown) with specific rabbit polyclonal anti-car in the presence of low (0.5 mm; left column) or high (5 mm; right three columns) Ca 2+ o (magnification, 40X). In blue, counterstaining of nuclei with hematoxylin. Tumor cells were treated for 24 h with PBS (vehicle control; upper row), ST (1 μm; middle row) or 8-br-cGMP (5 mm; lower row), in the absence (left two columns) or presence (right two columns) of the indicated CNG inhibitors (see Figure 1 legend for keys). Also, epithelial cells in villus tips from the
27 25 jejunum of GCC +/+ and GCC -/- mice were stained with DAPI (blue, nuclei) and specific antibodies against villin (green, enterocyte brush-border membranes) and CaR (red), and subjected to confocal microscopy (magnification, 63X). Fig. 5. CaR induced by ST is a novel therapeutic target in human colon cancer cells. (A and B) The proliferation of T84 cells stably expressing the empty vector (Vector, control), the CaR antisense vector (AS-CaR) or the CaR sense vector (S-CaR) was examined. Cells were treated for 24 h with Ca 2+ o (5 mm) alone (A) or ST (1 μm) plus Ca 2+ o (5 mm) (B). Results are expressed as in Figure 1c2; the controls were low (0.5 mm; A) or high (5 mm; B) Ca 2+ o. *, p<0.05, versus parallel control conditions. (C) T84 cell proliferation exposed to CaR agonists was measured as the percentage of respective control incubations treated with 0.5 mm Ca 2+ o (first two columns on the left). CaR agonists were added to tumor incubations for 24 h in the presence of the vehicle control (PBS) or ST (1 μm), and include: Ca 2+, 10 mm; Mg 2+, 20 mm; Gd 3+, 100 μm, and spermine, 100 μm. CaR agonists are used at concentrations that maximally induce CaR signaling (34,40). *, p<0.05 and **, P < 0.01, versus respective control (PBS) conditions. (D) Proposed molecular mechanism for restoration of CaR signaling by GCC associated with colorectal cancer suppression (see Discussion for description).
28 Pitari_Figure 1 A Proliferation, % C1 Proliferation, % T84 Caco-2 SW PBS ST, 24 h ** # ## [Ca 2+ ] o, mm +ST, 3 h ** * B Cytotoxicity, % C2 % Inhibition of Proliferation Apoptosis [Ca 2+ ] o, 1 mm [Ca 2+ ] o, 10 mm [Ca 2+ ] o, 30 mm T84 Necrosis ST, 24 h ST, 3 h Apoptosis Caco-2 Necrosis [Ca 2+ ] o, mm [Ca 2+ ] o, mm
29 A ** Proliferation, % ST, nm * * B 75 T84 Caco-2 SW ** * * * * * * * % Inhibition of Proliferation 8-br-cGMP ST 8-br-cGMP ST 8-br-cGMP TJU ST 8-pCPT-cGMP 8-pCPT-cGMP 8-pCPT-cGMP C % Inhibition of Proliferation IBMX L-DLT PET BAPTA RP8pCPT RP-cAMPS 0 * * ST * * PBS Pitari_Figure 2 D G 2 /M 21% S 37% PBS sub-g 1 11% G 1 31% S 19% G 2 /M 16% ST sub-g 1 9% G 1 56%
30 Pitari_Figure 3 A ST-Induced cgmp, pmol/mg cell protein ST Pretreatment +ST Pretreatment * * 1 10 [Ca 2+ ] o, mm B CaR GAPDH CaR Villin 0 h 3 h 24 h PBS ST PBS ST PBS ST 0 h 3 h 24 h PBS ST PBS ST PBS ST Cytosol Membrane








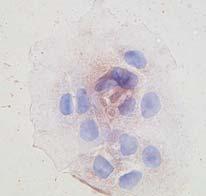




31 Pitari_Figure 4 In Vitro In Vivo GCC -/- Ca 2+ o 0.5 mm Ca 2+ o 5 mm L-DLT PET (-) GCC Signaling PBS DAPI Villin CaR Merge GCC +/+ (+) GCC Signaling ST 8-brcGMP DAPI Villin CaR Merge
32 Pitari_Figure 5 A C % Inhibition of Proliferation Ca 2+ * * * Vector AS-CaR S-CaR Proliferation, % Control PBS ST * * * ** Ca 2+ Mg 2+ Gd 3+ Spermine B 75 ST + Ca 2+ * D ST Uroguanylin Guanylin Spermine Ca 2+ Mg 2+ % Inhibition of Proliferation * Vector AS-CaR S-CaR Membrane GTP GCC cgmp CNG Ca 2+ CaR Tumor Suppression
Targeting the cgmp Pathway to Treat Colorectal Cancer
 Thomas Jefferson University Jefferson Digital Commons Department of Pharmacology and Experimental Therapeutics Faculty Papers Department of Pharmacology and Experimental Therapeutics 29 Targeting the cgmp
Thomas Jefferson University Jefferson Digital Commons Department of Pharmacology and Experimental Therapeutics Faculty Papers Department of Pharmacology and Experimental Therapeutics 29 Targeting the cgmp
Receptors Families. Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia
 Receptors Families Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia Receptor Families 1. Ligand-gated ion channels 2. G protein coupled receptors 3. Enzyme-linked
Receptors Families Assistant Prof. Dr. Najlaa Saadi PhD Pharmacology Faculty of Pharmacy University of Philadelphia Receptor Families 1. Ligand-gated ion channels 2. G protein coupled receptors 3. Enzyme-linked
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
(A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a
 RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a") Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
TITLE: Autocrine and Paracrine Control of Breast Cancer Growth by Sex Hormone-Binding Globulin
 AD Award Number: DAMD17-02-1-0572 TITLE: Autocrine and Paracrine Control of Breast Cancer Growth by Sex Hormone-Binding Globulin PRINCIPAL INVESTIGATOR: William Rosner, M.D. Scott M. Kahn, Ph.D. CONTRACTING
AD Award Number: DAMD17-02-1-0572 TITLE: Autocrine and Paracrine Control of Breast Cancer Growth by Sex Hormone-Binding Globulin PRINCIPAL INVESTIGATOR: William Rosner, M.D. Scott M. Kahn, Ph.D. CONTRACTING
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Receptors Functions and Signal Transduction- L4- L5
 Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice
 Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice Jimmy A. Rotolo 1, Branka Stancevic 1, Jianjun Zhang 1, Guoqiang Hua 1, John Fuller 1, Xianglei Yin 1, Adriana Haimovitz-Friedman 2, Kisu
Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice Jimmy A. Rotolo 1, Branka Stancevic 1, Jianjun Zhang 1, Guoqiang Hua 1, John Fuller 1, Xianglei Yin 1, Adriana Haimovitz-Friedman 2, Kisu
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
AMPK Phosphorylation Assay Kit
 AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.
 Light microscopic pictures show mouse embryos at developmental stage E12.") Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Plasma membranes. Plasmodesmata between plant cells. Gap junctions between animal cells Cell junctions. Cell-cell recognition
 Cell Communication Cell Signaling Cell-to-cell communication is essential for multicellular organisms Communicate by chemical messengers Animal and plant cells have cell junctions that directly connect
Cell Communication Cell Signaling Cell-to-cell communication is essential for multicellular organisms Communicate by chemical messengers Animal and plant cells have cell junctions that directly connect
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
2013 W. H. Freeman and Company. 12 Signal Transduction
 2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
(Stratagene, La Jolla, CA) (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment
 (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment") SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Product Manual. Human LDLR ELISA Kit. Catalog Number. FOR RESEARCH USE ONLY Not for use in diagnostic procedures
 Product Manual Human LDLR ELISA Kit Catalog Number STA-386 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Cholesterol is an essential component of cellular membranes,
Product Manual Human LDLR ELISA Kit Catalog Number STA-386 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Cholesterol is an essential component of cellular membranes,
Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/-
 Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Summary and Concluding Remarks
 Summary and Concluding Remarks Chapter 6 The intestinal epithelium provides an excellent model system for investigating molecular mechanisms regulating cell lineage establishment, stem cell proliferation,
Summary and Concluding Remarks Chapter 6 The intestinal epithelium provides an excellent model system for investigating molecular mechanisms regulating cell lineage establishment, stem cell proliferation,
Supplemental Information
 Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
supplementary information
 DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
Cell Signaling part 2
 15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
Doctor of Philosophy
 Regulation of Gene Expression of the 25-Hydroxyvitamin D la-hydroxylase (CYP27BI) Promoter: Study of A Transgenic Mouse Model Ivanka Hendrix School of Molecular and Biomedical Science The University of
Regulation of Gene Expression of the 25-Hydroxyvitamin D la-hydroxylase (CYP27BI) Promoter: Study of A Transgenic Mouse Model Ivanka Hendrix School of Molecular and Biomedical Science The University of
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
Ayman Mesleh & Leen Alnemrawi. Bayan Abusheikha. Faisal
 24 Ayman Mesleh & Leen Alnemrawi Bayan Abusheikha Faisal We were talking last time about receptors for lipid soluble hormones.the general mechanism of receptors for lipid soluble hormones: 1. Receptors
24 Ayman Mesleh & Leen Alnemrawi Bayan Abusheikha Faisal We were talking last time about receptors for lipid soluble hormones.the general mechanism of receptors for lipid soluble hormones: 1. Receptors
Supplemental Data Figure S1 Effect of TS2/4 and R6.5 antibodies on the kinetics of CD16.NK-92-mediated specific lysis of SKBR-3 target cells.
 Supplemental Data Figure S1. Effect of TS2/4 and R6.5 antibodies on the kinetics of CD16.NK-92-mediated specific lysis of SKBR-3 target cells. (A) Specific lysis of IFN-γ-treated SKBR-3 cells in the absence
Supplemental Data Figure S1. Effect of TS2/4 and R6.5 antibodies on the kinetics of CD16.NK-92-mediated specific lysis of SKBR-3 target cells. (A) Specific lysis of IFN-γ-treated SKBR-3 cells in the absence
To determine the effect of over-expression and/or ligand activation of. PPAR / on cell cycle, cell lines were cultured as described above until ~80%
 Supplementary Materials and Methods Cell cycle analysis To determine the effect of over-expression and/or ligand activation of PPAR / on cell cycle, cell lines were cultured as described above until ~80%
Supplementary Materials and Methods Cell cycle analysis To determine the effect of over-expression and/or ligand activation of PPAR / on cell cycle, cell lines were cultured as described above until ~80%
Supplementary Figures
 Supplementary Figures Supplementary Figure 1 Increased ABHD5 expression in human colon cancer associated macrophages. (a) Murine peritoneal macrophages were treated with regular culture medium (Ctrl) or
Supplementary Figures Supplementary Figure 1 Increased ABHD5 expression in human colon cancer associated macrophages. (a) Murine peritoneal macrophages were treated with regular culture medium (Ctrl) or
Ex vivo Human Antigen-specific T Cell Proliferation and Degranulation Willemijn Hobo 1, Wieger Norde 1 and Harry Dolstra 2*
 Ex vivo Human Antigen-specific T Cell Proliferation and Degranulation Willemijn Hobo 1, Wieger Norde 1 and Harry Dolstra 2* 1 Department of Laboratory Medicine - Laboratory of Hematology, Radboud University
Ex vivo Human Antigen-specific T Cell Proliferation and Degranulation Willemijn Hobo 1, Wieger Norde 1 and Harry Dolstra 2* 1 Department of Laboratory Medicine - Laboratory of Hematology, Radboud University
Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival
 Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
TRACP & ALP Assay Kit
 Cat. # MK301 For Research Use TRACP & ALP Assay Kit Product Manual Table of Contents I. Description...3 II. III. IV. Introduction...3 Components...4 Materials Required but not Provided...4 V. Storage...4
Cat. # MK301 For Research Use TRACP & ALP Assay Kit Product Manual Table of Contents I. Description...3 II. III. IV. Introduction...3 Components...4 Materials Required but not Provided...4 V. Storage...4
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION
 Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
Supporting Information
 Supporting Information Fujishita et al. 10.1073/pnas.0800041105 SI Text Polyp Scoring. Intestinal polyps were counted as described (1). Briefly, the small and large intestines were excised, washed with
Supporting Information Fujishita et al. 10.1073/pnas.0800041105 SI Text Polyp Scoring. Intestinal polyps were counted as described (1). Briefly, the small and large intestines were excised, washed with
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Dr Rodney Itaki Lecturer Anatomical Pathology Discipline. University of Papua New Guinea School of Medicine & Health Sciences Division of Pathology
 Neoplasia Dr Rodney Itaki Lecturer Anatomical Pathology Discipline University of Papua New Guinea School of Medicine & Health Sciences Division of Pathology General Considerations Overview: Neoplasia uncontrolled,
Neoplasia Dr Rodney Itaki Lecturer Anatomical Pathology Discipline University of Papua New Guinea School of Medicine & Health Sciences Division of Pathology General Considerations Overview: Neoplasia uncontrolled,
Propagation of the Signal
 OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
ab SREBP-2 Translocation Assay Kit (Cell-Based)
") ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
Neoplasia 18 lecture 6. Dr Heyam Awad MD, FRCPath
 Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Neoplasia 18 lecture 6 Dr Heyam Awad MD, FRCPath ILOS 1. understand the role of TGF beta, contact inhibition and APC in tumorigenesis. 2. implement the above knowledge in understanding histopathology reports.
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1
 GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
LDL Uptake Cell-Based Assay Kit
 LDL Uptake Cell-Based Assay Kit Catalog Number KA1327 100 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
LDL Uptake Cell-Based Assay Kit Catalog Number KA1327 100 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
RNA extraction, RT-PCR and real-time PCR. Total RNA were extracted using
 Supplementary Information Materials and Methods RNA extraction, RT-PCR and real-time PCR. Total RNA were extracted using Trizol reagent (Invitrogen,Carlsbad, CA) according to the manufacturer's instructions.
Supplementary Information Materials and Methods RNA extraction, RT-PCR and real-time PCR. Total RNA were extracted using Trizol reagent (Invitrogen,Carlsbad, CA) according to the manufacturer's instructions.
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC-
 Supplemental material and methods Reagents. Hydralazine was purchased from Sigma-Aldrich. Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC- 133, human thyroid medullary
Supplemental material and methods Reagents. Hydralazine was purchased from Sigma-Aldrich. Cell Culture. The human thyroid follicular carcinoma cell lines FTC-238, FTC-236 and FTC- 133, human thyroid medullary
Regulation of cell function by intracellular signaling
 Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
Regulation of cell function by intracellular signaling Objectives: Regulation principle Allosteric and covalent mechanisms, Popular second messengers, Protein kinases, Kinase cascade and interaction. regulation
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
LDL Uptake Cell-Based Assay Kit
 LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
Human Leptin ELISA Kit
 Product Manual Human Leptin ELISA Kit Catalog Numbers MET-5057 MET-5057-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Leptin is a polypeptide hormone
Product Manual Human Leptin ELISA Kit Catalog Numbers MET-5057 MET-5057-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Leptin is a polypeptide hormone
General Principles of Endocrine Physiology
 General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
ab Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions.
 ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway
 Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
By the name of Allah
 By the name of Allah Receptors function and signal transduction ( Hormones and receptors Types) We were talking about receptors of the neurotransmitters; we have 2 types of receptors: 1- Ionotropic receptors
By the name of Allah Receptors function and signal transduction ( Hormones and receptors Types) We were talking about receptors of the neurotransmitters; we have 2 types of receptors: 1- Ionotropic receptors
In vitro human regulatory T cell suppression assay
 Human CD4 + CD25 + regulatory T cell isolation, in vitro suppression assay and analysis In vitro human regulatory T cell suppression assay Introduction Regulatory T (Treg) cells are a subpopulation of
Human CD4 + CD25 + regulatory T cell isolation, in vitro suppression assay and analysis In vitro human regulatory T cell suppression assay Introduction Regulatory T (Treg) cells are a subpopulation of
NF-κB p65 (Phospho-Thr254)
") Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
INTERACTION DRUG BODY
 INTERACTION DRUG BODY What the drug does to the body What the body does to the drug Receptors - intracellular receptors - membrane receptors - Channel receptors - G protein-coupled receptors - Tyrosine-kinase
INTERACTION DRUG BODY What the drug does to the body What the body does to the drug Receptors - intracellular receptors - membrane receptors - Channel receptors - G protein-coupled receptors - Tyrosine-kinase
Chapter 11. Cell Communication. Signal Transduction Pathways
 Chapter 11 Cell Communication Signal Transduction Pathways Signal-Transduction Pathway Signal on a cell s surface is converted into a specific cellular response Local signaling (short distance) - Paracrine
Chapter 11 Cell Communication Signal Transduction Pathways Signal-Transduction Pathway Signal on a cell s surface is converted into a specific cellular response Local signaling (short distance) - Paracrine
Cell Signaling (part 1)
") 15 Cell Signaling (part 1) Introduction Bacteria and unicellular eukaryotes respond to environmental signals and to signaling molecules secreted by other cells for mating and other communication. In multicellular
15 Cell Signaling (part 1) Introduction Bacteria and unicellular eukaryotes respond to environmental signals and to signaling molecules secreted by other cells for mating and other communication. In multicellular
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Fang et al. NMuMG. PyVmT unstained Anti-CCR2-PE MDA-MB MCF MCF10A
 A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
ab Lysosome/Cytotoxicity Dual Staining Kit
 ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
Immature organoids appear after hours.
 THE ESSENTIALS OF LIFE SCIENCE RESEARCH GLOBALLY DELIVERED Allison Ruchinskas, B.S., and James Clinton, Ph.D. ATCC Cell Systems, Gaithersburg, MD INTRODUCTION Figure 1. Mouse small intestinal organoid
THE ESSENTIALS OF LIFE SCIENCE RESEARCH GLOBALLY DELIVERED Allison Ruchinskas, B.S., and James Clinton, Ph.D. ATCC Cell Systems, Gaithersburg, MD INTRODUCTION Figure 1. Mouse small intestinal organoid
Cover Page. The handle holds various files of this Leiden University dissertation.
 Cover Page The handle http://hdl.handle.net/1887/22278 holds various files of this Leiden University dissertation. Author: Cunha Carvalho de Miranda, Noel Filipe da Title: Mismatch repair and MUTYH deficient
Cover Page The handle http://hdl.handle.net/1887/22278 holds various files of this Leiden University dissertation. Author: Cunha Carvalho de Miranda, Noel Filipe da Title: Mismatch repair and MUTYH deficient
A protocol for enhancement of the AAV-mediated expression of transgenes
 A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Focus Application. Compound-Induced Cytotoxicity
 xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
- 1 - Cell types Monocytes THP-1 cells Macrophages. LPS Treatment time (Hour) IL-6 level (pg/ml)
 IL-6 level (pg/ml)") Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11429 S1a 6 7 8 9 Nlrc4 allele S1b Nlrc4 +/+ Nlrc4 +/F Nlrc4 F/F 9 Targeting construct 422 bp 273 bp FRT-neo-gb-PGK-FRT 3x.STOP S1c Nlrc4 +/+ Nlrc4 F/F casp1
SUPPLEMENTARY INFORMATION doi:10.1038/nature11429 S1a 6 7 8 9 Nlrc4 allele S1b Nlrc4 +/+ Nlrc4 +/F Nlrc4 F/F 9 Targeting construct 422 bp 273 bp FRT-neo-gb-PGK-FRT 3x.STOP S1c Nlrc4 +/+ Nlrc4 F/F casp1
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Note: During 30 minute incubation; proceed thru appropriate sections below (e.g. sections II, III and V).
.") LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
LEGEND MAX β Amyloid x 40 LEGEND MAX β Amyloid x 40 ELISA Kit Components and Protocol Kit Components Capture Antibody Coated Plate 1 stripwell plate 1 40 Standard (2) 20μg vial 5X Wash Buffer 125mL Standard
Mouse GLP-2 EIA FOR LABORATORY USE ONLY
 YK142 Mouse GLP-2 EIA FOR LABORATORY USE ONLY Kasumigaseki place, 3-6-7, Kasumigaseki, Chiyoda-ku, Tokyo 100-0013 Japan http://www.sceti.co.jp/english/export e-mail exp-pet@sceti.co.jp
YK142 Mouse GLP-2 EIA FOR LABORATORY USE ONLY Kasumigaseki place, 3-6-7, Kasumigaseki, Chiyoda-ku, Tokyo 100-0013 Japan http://www.sceti.co.jp/english/export e-mail exp-pet@sceti.co.jp
Anti-Lamin B1/LMNB1 Picoband Antibody
 Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
The effect of insulin on chemotherapeutic drug sensitivity in human esophageal and lung cancer cells
 The effect of insulin on chemotherapeutic drug sensitivity in human esophageal and lung cancer cells Published in: Natl Med J China, February 10, 2003; Vol 83, No 3, Page 195-197. Authors: JIAO Shun-Chang,
The effect of insulin on chemotherapeutic drug sensitivity in human esophageal and lung cancer cells Published in: Natl Med J China, February 10, 2003; Vol 83, No 3, Page 195-197. Authors: JIAO Shun-Chang,
a b G75 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server.
 a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS
 Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Physiology Unit 1 CELL SIGNALING: CHEMICAL MESSENGERS AND SIGNAL TRANSDUCTION PATHWAYS In Physiology Today Cell Communication Homeostatic mechanisms maintain a normal balance of the body s internal environment
Supplementary Figure 1. MAT IIα is Acetylated at Lysine 81.
 IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
ulcer healing role 118 Bicarbonate, prostaglandins in duodenal cytoprotection 235, 236
 Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein
 Representative blots (upper and lower panels) showing the AdipoR1 protein") Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein content relative to GAPDH in two independent experiments.
Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein content relative to GAPDH in two independent experiments.