Cyclin B destruction triggers changes in kinetochore behavior essential for successful anaphase
|
|
|
- Brian Crawford
- 5 years ago
- Views:
Transcription
1 Parry, Hickson & O Farrell, 00, Curr. Biol. : Cyclin B destruction triggers changes in kinetochore behavior essential for successful anaphase Devin H. Parry, Gilles R. X. Hickson, and Patrick H. O Farrell* Department of Biochemistry and Biophysics, University of California San Francisco, San Francisco, CA , USA Abstract Successful mitosis requires that anaphase chromosomes sustain a commitment to move to their assigned spindle-poles. This requires stable spindle-attachment of anaphase kinetochores. Prior to anaphase, stable spindle-attachment depends on tension created by opposing forces on sisterkinetochores []. Since tension is lost when kinetochores disjoin, stable attachment in anaphase must have a different basis. After expression of non-degradable cyclin B (CYC- B S ) in Drosophila embryos, sister chromosomes disjoined normally but their anaphase behavior was abnormal []. Chromosomes exhibited cycles of reorientation from one pole to the other. Additionally, the unpaired kinetochores accumulated attachments to both poles (merotelic attachments), congressed (again) to a pseudo-metaphase plate, and reacquired associations with checkpoint proteins more characteristic of prometaphase kinetochores. Unpaired prometaphase kinetochores, which occurred in a mutant entering mitosis with unreplicated (unpaired) chromosomes, behaved just like the anaphase kinetochores at the CYC-B S arrest. Finally, the normal anaphase release of AuroraB/INCENP from kinetochores was blocked by CYC-B S expression, and reciprocally, was advanced in a CycB mutant. Given its established role in destabilizing kinetochore-microtubule interactions [], Aurora B dissociation is likely to be key to the change in kinetochore behavior. These findings show that, in addition to loss of sister chromosome cohesion, successful anaphase requires a transition in kinetochore behavior triggered by CYC-B destruction. Results and discussion CYC-B maintains AuroraB/INCENP localization at the kinetochore Stable cyclins have been shown to block mitotic exit in numerous systems [4-8], and detailed analyses of the cytological consequence of stabilization of each of the cognate mitotic cyclins of Drosophila have begun to reveal regulatory features that were not evident in other experimental systems [, 9]. A group of chromosomal passenger proteins that are localized between paired kinetochores at metaphase usually relocalize to the central spindle upon onset of anaphase [0]. Previous work showed that this relocalization is blocked upon expression of stable sea urchin cyclin B in mammalian cells [, ]. In agreement with this, expression of Drosophila CYC-B S in Drosophila embryos blocked relocalization of two interacting passenger proteins, INCENP (Figure A- B, and Figure SA) and Aurora B (Figure SE). Normal metaphase foci of INCENP split in two at anaphase, half segregating with each sister kinetochore without relocalization to the spindle (Figure B). Failure to release kinetochorelocalized AuroraB/INCENP and a slowing of anaphase A chromosome movements [] are the earliest perturbations of mitotic progression observed upon CYC-B S expression. The onset of these defects immediately follows or overlaps the time of destruction of normal CYC-B. Embryos expressing a different stabilized mitotic cyclin, CYC-B S, arrested with chromosomes at the spindle poles following normal anaphase movements [] and normal redistribution
2 Parry, Hickson & O Farrell, 00, Curr. Biol. : of AuroraB/INCENP from the kinetochore to the spindle midzone (Figure D and SB). Thus, CYC- B S and not CYC-B S maintains kinetochore localization of AuroraB/INCENP. As a result of partial redundancy among Drosophila cyclins, CycB null mutants undergo mitosis []. As in wild-type, AuroraB/INCENP is associated with kinetochores in metaphase cells lacking CYC-B, however its anaphase relocalization occurs prematurely (compare Figure A and C). Thus, the endogenous CYC-B in wild-type inhibits AuroraB/INCENP relocalization, and relocalization appears to await its destruction. Together, precocious relocalization in the CycB mutant, coincidence in onset of relocalization and time of CYC-B destruction, and the block to relocalization by persistent CYC-B, leads us to conclude that CYC-B destruction times AuroraB/INCENP relocalization. CYC-B degradation switches chromosomes from prometaphase to anaphase behavior. The dramatic transition in kinetochoreprotein interactions upon destruction of CYC-B might serve only to release the sequestered passenger proteins to play their important function at the spindle midzone in cytokinesis [4, 5]. However, elegant studies of the Aurora B kinase homolog of yeast, Ipl, suggest that Ipl can destabilize kinetochore interactions with the spindle [, 6, 7]. These studies, as well as supporting work in vertebrate cells [8, 9], suggest that loss of Aurora B function upon CYC-B destruction might alter kinetochore behavior. Indeed, our results suggest that CYC-B destruction does have an important influence on anaphase chromosome behavior. As we previously described, when Drosophila cells enter anaphase in the presence of CYC-B S, poleward movement of unpaired chromosomes is abortive and chromosome behavior is unusual []. We suggested that this chromosome behavior might represent extension of prometaphase/metaphase behavior, but influenced by the loss of kinetochore pairing at metaphase/anaphase. The behavior of unpaired prometaphase kinetochores was previously examined in a mutant in maize exhibiting premature loss of chromosome pairing, and following microsurgical production of single kinetochore chromosomes in mammalian cells [0, ]. In these experiments, single-kinetochore chromosomes behaved much as the chromosomes of Drosophila cells that progress to anaphase (to produce unpaired kinetochores) in the presence of CYC-B S []. To further test this parallel, we examined the Drosophila mutant, double parked, in which unpaired chromosomes exist in prometaphase. Double Parked is an essential replication protein that is also required for a checkpoint function that ordinarily prevents cells from entering mitosis with unreplicated DNA, and like analogous mutants in S. cerevisiae (e.g. cdc6), Drosophila cells lacking Double Parked enter mitosis with unreplicated DNA []. When a maternal supply of Double Parked is depleted, replication fails in double parked embryos and cells accumulate in mitosis (Figure A). The mitotic arrest occurs because unpaired chromosomes are incapable of normal bipolar alignment and consequently induce the spindle checkpoint [, ]. In fixed images of the double parked arrest, most chromosomes were scattered along the spindle with some clustered in a central pseudo-metaphase plate (Figure B,D), just as in CYC-B S arrested cells []. Real-time analysis showed that this is a dynamic situation with chromosomes making oscillatory movements between the poles (Figure C, supplementary data Movie ). For example, during the double parked arrest, a chromosome highlighted in green in Figure C lingered near the lower pole during four frames (5 sec), then moved abruptly towards the opposite pole over the next few frames (most movement within 5 sec, 69-04). This chromosome movement between the poles resembles that observed during the CYC-B S block and is consistent with reorientation of the kinetochore from one pole to the other as has been described for prometaphase chromosomes [4]. Despite the absence of prior replication, INCENP and Aurora B localized to the unpaired kinetochores in the double parked arrest (Figure D,D, S and data not shown), as in the CYC-B S arrest. Furthermore, despite the presence of only a single kinetochore, many of the chromosomes congress to a pseudo-metaphase plate in double parked and CYC-B S arrests (Figure and ). We conclude that, when CYC-B persists, unpaired chromosomes behave similarly before and after the metaphase/anaphase transition. While it was somewhat puzzling that some chromosomes congressed to a pseudo-metaphase plate in double parked embryos, a similar observation was made when single kinetochore chromosomes were present in prometaphase of
3 Parry, Hickson & O Farrell, 00, Curr. Biol. : mammals [0]. These congressed single kinetochore chromosomes had attachments to both poles (merotelic attachment). We observed robust kinetochore fibers in double parked spindles, and in cases that were not confounded by the clustering of chromosomes in the middle, it was apparent that kinetochore fibers from both poles impinged on single kinetochores (Figure E and S). We interpret these observations as an indication of frequent merotelic attachment in the double parked arrest and have made similar findings in the CYC-B S arrested cells (see below). The finding that merotelic attachments accumulate in the double parked arrest suggests that kinetochore-pairing normally helps to prevent merotelic attachments under prometaphase conditions. We suggest that such an effect could be explained by an extension of the idea that trial and error processes contribute to bipolar attachment of paired kinetochores in prometaphase []. Because kinetochore-spindle interactions are unstable in prometaphase, all modes of attachment can be sampled, at least transiently, but the most stable mode ultimately predominates. Consequently, the most stable (correct bipolar attachment) precludes less stable and incorrect attachments (Figure 4A). Spindle tension stabilizes attachment, and it has been suggested [] that, upon bipolar arrangement, tension deforms the paired kinetochore, effectively pulling the attachment sites away from a centrally localized destabilizing activity (Figure 4). While tension also deforms a merotelically attached kinetochore [0,, 5], we suggest that the distortion is not as orderly as in bipolar attachment, and that the separation from the destabilizing activity is less effective. Consequently, when kinetochores are paired, bipolar attachments will accumulate as the most stable outcome, and hence exclude merotelic attachments. When kinetochores are unpaired the dynamics of formation and decay of merotelic attachments appears to favor their accumulation. Checkpoint reactivation in CYC-B S arrested cells Prior to the time at which CYC-B is usually degraded, we see no defects in mitotic progression in cells expressing CYC-B S. Sister chromatids separate from one another and other substrates of the APC/C are degraded [, 8]. The dissociation of BubR from kinetochores marks release of checkpoint control. CYC-B S -expressing cells having an anaphase configuration (prior to final arrest) had greatly decreased level of kineotchore staining (Figure F, SC and data not shown). However, at the final arrest point, BubR again localized to the kinetochores (Figure G, SC and data not shown). BubR staining did not completely disappear during anaphase and levels at final arrest did not match the highest levels at prometaphase (data not shown). Nevertheless, since a return of BubR to the kinetochore after sister chromatid separation was never observed in wild-type cells, there appears to be some reactivation of the checkpoint at the CYC-B S arrest. As additional reporters of checkpoint activity, we probed for ROD (Figure H-J and SD) and ZW0 (data not shown), components of a mitotic checkpoint that relocalize in a manner suggesting a role in sensing tension [6, 7]. In prometaphase cells, a ROD/ZW0 complex localizes tightly to kinetochores. As chromosomes develop bipolar attachment, the kinetochore staining for ROD/ZW0 is reduced and staining appears on kinetochore fibers. Upon disjunction of sister chromosomes the staining returns to the kinetochores. All these events appeared to occur normally in CYC-B S and CYC-B S expressing cells as they progressed toward an arrest (data not shown). However, during the course of arrest with CYC-B S, but not with CYC-B S, spindle microtubules once again stained for ROD/ZW0 (Figure I, versus J). The spindle localization of ROD/ZW0 in the CYC-B S arrest is another example of checkpoint components that have reverted to their characteristic pre-anaphase localization and further suggests that some aspects of the checkpoint have been reactivated, perhaps in response to defective chromosome-spindle interactions. CYC-B S arrested cells display merotelic spindle attachments Spindle staining of ROD/ZW0 during metaphase has been shown to require bipolar attachment and perhaps tension across the kinetochores [6, 7]. The spindle staining in cells at the CYC-B S arrest could imply some level of bipolar attachment. Initially, this seemed unlikely, as the anaphase chromosomes are unpaired and contain only one kinetochore when at the arrest point. However, staining for kinetochores and microtubules showed robust kinetochore fibers extending from both poles to the pseudo-metaphase
4 Parry, Hickson & O Farrell, 00, Curr. Biol. : plate, suggesting merotelic attachment (Figure A and S). Merotelic attachments were obvious when only one or a few chromosomes remained near the middle of the spindle (Figure B,C and S), as occurred frequently early after the transition to anaphase in the presence of CYC-B S (Figure B), and when the arrest was less complete (Figure C). At the level of CYC-B S expression in our experiments some cells are not fully arrested []. Live observations revealed occasional cells with slow mitotic progress but without the full complement of arrest behaviors. These prolonged mitoses show a high frequency of chromosome segregation anomalies. For example, Figure D shows frames from a movie in which chromosomes separated after a prolonged anaphase A as seen in cells destined for CYC-B S arrest []; however, unlike the complete arrest in which all the chromosomes lose their poleward orientation, most of the chromosomes remained at the poles and decondensed, while a single chromosome moved from one pole to the other. Kinetochores successfully retained at the pole following CYC-B S expression lacked INCENP and BubR, while chromosomes localized to the middle of the spindle displayed merotelic attachments, and stained strongly for INCENP and BubR (Figure C,C and S). The presence of two categories of kinetochore, one having and one lacking INCENP, suggests a switch-like event at individual kinetochores and that the cells with an incomplete arrest are near the threshold of the switch. Our findings show that CYC-B S promotes merotelic attachments, which accumulate following the initially successful chromosome disjunction at the transition to anaphase. Furthermore, our results are consistent with proposals that merotelic attachments underlie congression of chromosomes with a single kinetochore [0] and disrupt chromosome segregation [5]. We suggest that the accumulation of merotelic attachments at the CYC- B S arrest is the consequence of persistence of the dynamic phase of kinetochore spindle attachment beyond the time of sister kinetochore disjunction. Rather than preserving the established monopolar orientation of the anaphase kinetochores, persistence of dynamic exchange favors change toward the arrangements that are most stable for unpaired kinetochores, and one such arrangement is merotelic attachment (Figure 4). Conclusions Our results show that a change in kinetochore composition and behavior accompanies the metaphase/anaphase transition and that a change in kinetochore behavior is essential for the unerring commitment of chromosomes to their assigned poles. Since the success of mitosis depends on this change, we think of the transition as an integral part of the metaphase/anaphase transition. We show that destruction of CYC-B triggers and times the kinetochore transition at the onset of anaphase, and that a second mitotic cyclin, CYC- B, does not govern this kinetochore transition. The kinetochore transition is coordinated with the disjunction of sister chromosomes as a result of their common regulation by APC/C, which promotes the destruction of CYC-B as well as the regulators of sister cohesion, securin and cyclin A [, 8]. The change in kinetochore behavior can be understood as a change from dynamicallyexchanging tension-stabilized attachment to fixed stable attachment. The striking coupling of this change with release of Aurora B/INCENP from the kinetochore, and the identified role of Aurora B kinase in destabilizing kinetochore spindle attachments [] suggests a plausible mechanism in which the dissociation of Aurora B stabilizes spindle attachments. However, a stable derivative of the sea urchin cyclin B did not produce similar modifications of chromosome behavior in mammalian cells despite blocking release of GFP- Aurora B from the kinetochores []. Clearly, further work is required to elucidate the regulatory paths connecting kinetochore behavior with CYC-B destruction. We found that unpaired chromosomes developed merotelic attachments whenever AuroraB/INCENP is associated with unpaired kinetochores, whether this occurs in anaphase as a result of CYC-B S expression, or in prophase as result of failure in DNA replication (in the double parked arrest). We suggest that kinetochorepairing influences the outcome of dynamic reassortment of kinetochore attachments (Figure 4). Evidently, it is important to stabilize kinetochore-spindle attachments upon disjunction of sisters, otherwise attachments re-equilibrate to the most stable states available to unpaired kinetochores, including merotelic attachments.
5 Parry, Hickson & O Farrell, 00, Curr. Biol. : References. Nicklas RB: How cells get the right chromosomes. Science 997; 75: Parry DH, O'Farrell PH: The schedule of destruction of three mitotic cyclins can dictate the timing of events during exit from mitosis. Curr Biol 00; : Tanaka TU, Rachidi N, Janke C, Pereira G, Galova M, Schiebel E, Stark MJ, Nasmyth K: Evidence that the Ipl-Sli5 (Aurora kinase- INCENP) complex promotes chromosome biorientation by altering kinetochore-spindle pole connections. Cell 00; 08: Murray AW, Solomon MJ, Kirschner MW: The role of cyclin synthesis and degradation in the control of maturation promoting factor activity. Nature 989; 9: Ghiara JB, Richardson HE, Sugimoto K, Henze M, Lew DJ, Wittenberg C, Reed SI: A cyclin B homolog in S. cerevisiae: chronic activation of the Cdc8 protein kinase by cyclin prevents exit from mitosis. Cell 99; 65: Gallant P, Nigg EA: Cyclin B undergoes cell cycle-dependent nuclear translocation and, when expressed as a nondestructible mutant, causes mitotic arrest in HeLa cells. J Cell Biol 99; 7: Hinchcliffe EH, Cassels GO, Rieder CL, Sluder G: The coordination of centrosome reproduction with nuclear events of the cell cycle in the sea urchin zygote. J Cell Biol 998; 40: Sigrist S, Jacobs H, Stratmann R, Lehner CF: Exit from mitosis is regulated by Drosophila fizzy and the sequential destruction of cyclins A, B and B. Embo J 995; 4: Echard A, O'Farrell PH: The degradation of two mitotic cyclins contributes to the timing of cytokinesis. Curr Biol 00; in press. 0. Adams RR, Carmena M, Earnshaw WC: Chromosomal passengers and the (aurora) ABCs of mitosis. Trends Cell Biol 00; : Wheatley SP, Hinchcliffe EH, Glotzer M, Hyman AA, Sluder G, Wang Y: CDK inactivation regulates anaphase spindle dynamics and cytokinesis in vivo. J Cell Biol 997; 8: Murata-Hori M, Tatsuka M, Wang YL: Probing the dynamics and functions of aurora B kinase in living cells during mitosis and cytokinesis. Mol Biol Cell 00; : Jacobs HW, Knoblich JA, Lehner CF: Drosophila Cyclin B is required for female fertility and is dispensable for mitosis like Cyclin B. Genes Dev 998; : Kaitna S, Mendoza M, Jantsch-Plunger V, Glotzer M: Incenp and an aurora-like kinase form a complex essential for chromosome segregation and efficient completion of cytokinesis. Curr Biol 000; 0: Severson AF, Hamill DR, Carter JC, Schumacher J, Bowerman B: The aurora-related kinase AIR- recruits ZEN-4/CeMKLP to the mitotic spindle at metaphase and is required for cytokinesis. Curr Biol 000; 0: Biggins S, Severin FF, Bhalla N, Sassoon I, Hyman AA, Murray AW: The conserved protein kinase Ipl regulates microtubule binding to kinetochores in budding yeast. Genes Dev 999; : Kang J, Cheeseman IM, Kallstrom G, Velmurugan S, Barnes G, Chan CS: Functional cooperation of Dam, Ipl, and the inner centromere protein (INCENP)-related protein Sli5 during chromosome segregation. J Cell Biol 00; 55: Murata-Hori M, Wang YL: The kinase activity of aurora B is required for kinetochoremicrotubule interactions during mitosis. Curr Biol 00; : Kallio MJ, McCleland ML, Stukenberg PT, Gorbsky GJ: Inhibition of aurora B kinase blocks chromosome segregation, overrides the spindle checkpoint, and perturbs microtubule dynamics in mitosis. Curr Biol 00; : Khodjakov A, Cole RW, McEwen BF, Buttle KF, Rieder CL: Chromosome fragments possessing only one kinetochore can congress to the spindle equator. J Cell Biol 997; 6: Yu HG, Dawe RK: Functional redundancy in the maize meiotic kinetochore. J Cell Biol 000; 5:-4.. Whittaker AJ, Royzman I, Orr-Weaver TL: Drosophila double parked: a conserved, essential replication protein that colocalizes with the origin recognition complex and links DNA replication with mitosis and the downregulation of S phase transcripts. Genes Dev 000; 4: Garner M, van Kreeveld S, Su TT: mei- 4 and bub block mitosis at two distinct steps in response to incomplete DNA replication in Drosophila embryos. Curr Biol 00; : Nicklas RB: Recurrent pole-to-pole movements of the sex chromosome during prometaphase I in Melanoplus Differentialis spermatocytes. Chromosoma 96; :97-5.
6 Parry, Hickson & O Farrell, 00, Curr. Biol. : Cimini D, Howell B, Maddox P, Khodjakov A, Degrassi F, Salmon ED: Merotelic kinetochore orientation is a major mechanism of aneuploidy in mitotic mammalian tissue cells. J Cell Biol 00; 5: Williams BC, Gatti M, Goldberg ML: Bipolar spindle attachments affect redistributions of ZW0, a Drosophila centromere/kinetochore component required for accurate chromosome segregation. J Cell Biol 996; 4: Scaerou F, Aguilera I, Saunders R, Kane N, Blottiere L, Karess R: The rough deal protein is a new kinetochore component required for accurate chromosome segregation in Drosophila. J Cell Sci 999; : Nasmyth K, Peters JM, Uhlmann F: Splitting the chromosome: cutting the ties that bind sister chromatids. Science 000; 88: Cooke CA, Heck MM, Earnshaw WC: The inner centromere protein (INCENP) antigens: movement from inner centromere to midbody during mitosis. J Cell Biol 987; 05:
7 Parry, Hickson & O Farrell, 00, Curr. Biol. : Figures Figure : CYC-B destruction is required for normal anaphase distributions of INCENP, BubR and ROD UPPER PANELS: (A-D) INCENP localization in wild-type (A), CYC-B S expressing (B), CycB mutant (C), and CYC-B S expressing cells (D). INCENP staining is in red, Centromere Identifier (CID) staining in green and Hoechst staining of DNA in blue (see supplementary data Figure SA and SB for separated channels and tubulin staining). (A) INCENP, which is localized between paired CID staining foci in metaphase (cell #), disengages from kinetochores during anaphase (cells #,) to become strongly localized at the spindle midzone by telophase (cell #4) [9]. (B) INCENP foci remain associated with kinetochores in cells expressing CYC-B S. Cell # is late anaphase A (i.e. prior to final arrest). Cells # and # are at a later stage of arrest with kinetochores largely congregated at a pseudo-metaphase plate (note absence of pairing of CID staining foci as in the orderly arrangement seen in cell # of panel A). (C) INCENP, while kinetochore-localized during metaphase (cell #) in CycB null mutant cells, it relocalizes prematurely during anaphase (cells #,,). (D) Cells expressing another stable cyclin, CYC-B S, show normal anaphase movements and INCENP relocalization as they progress (cells #,) from metaphase cell # to an arrest with chromosomes at separated poles []. In all cases examined, Aurora B staining shows the same localization as that for INCENP (supplementary Figure SE and data not shown). MIDDLE PANELS: (E-G) BubR is lost from kinetochores during early anaphase in wildtype (E) and CYC-B S expressing (F) cells, but kinetochore localization is later restored in CYC-B S arrested cells (G). BubR staining is in green and Hoechst staining of the DNA in blue (see supplementary data Figure SC for separated channels and tubulin staining). (E) In wild-type, BubR localizes to the kinetochores in metaphase (cell # and #), but is absent from the kinetochores during anaphase (cell #). (F) In cells expressing CYC-B S, BubR localizes normally in metaphase (cell #) and is absent from the kinetochores during the early anaphase A movements (cell #). Note that we documented many CYC-B S expressing cells in early anaphase, and reduction in kinetochore staining by Bub-R antibody at this stage is a general finding. (G) BubR again localizes to the kinetochores in cells arrested with CYC-B S. LOWER PANELS: (H-J) ROD localization in wild-type (H), CYC-B S expressing (I) and CYC-B S expressing (J) cells. ROD staining is in red, Hoechst staining of the DNA in blue and CID staining in green (see supplementary data Figure SD for separated channels and tubulin staining). (H) ROD staining is on the kinetochores and along the spindle during metaphase (cell #), but only on the kinetochores during anaphase (cell #) in wildtype cells. (I) ROD staining in cells arrested with CYC-B S, localized to some of the kinetochores with some dispersion along the spindle. (J) ROD staining can be seen only at the kinetochores in cells arrested with CYC-B S. In all cases examined, ZW0 staining shows the same localization as that for ROD (data not shown) Heat shock and antibody staining protocols have been previously described []. Antibodies were generously provided by Gary Karpen (chicken α-cid), Bill Earnshaw (rabbit α- INCENP), Claudio Sunkel (rabbit α-bubr), Roger Karess (rabbit α-rod) and David Glover (rabbit α- Aurora B). Homozygous cyclin B null mutants were identified by the absence of a balancer chromosome expressing lacz under the control of the ftz promoter, detected by β-galactosidase staining in a striped pattern. Scale bars indicate 5µm. Figure : Unreplicated chromosomes in double parked mitosis behave like disjoined chromosomes at CYC-B S arrest. The double parked mutant embryo fails to replicate DNA in S phase of cycle 6 and arrests in the subsequent mitosis []. (A) A double parked mutant embryo (left) and a control embryo (right) are stained for phospho-histone H, which specifically labels mitotic chromosomes, in green and α-tubulin in red. The mutant embryo was identified by the absence of a balancer chromosome expressing lacz under the control of the ftz promoter, detected by β-galactosidase staining (also red) in a striped pattern (see control embryo). (B) A region (boxed) of the double parked mutant embryo from (A) is enlarged to show deranged distribution of chromosomes along the spindle. The arrow indicates clustering of chromosomes at a pseudo-metaphase plate in some cells. (C) Frames from a real-time movie (supplementary data: Movie
8 Parry, Hickson & O Farrell, 00, Curr. Biol. : ) showing dynamic chromosome movements in the double parked mutant mitotic arrest. The approximate spindle orientation is given with the dotted red line. The highlighted chromosome (artificially green), initially oriented towards the lower pole in the first 4 frames, is seen to undergo spindle reorientation and rapid movement towards the upper pole in the final 4 frames. Time indicated is seconds elapsed from the start of the series. Standard crossing schemes were used to create a line of flies carrying the dup a mutation and the histone-gfp transgene for live analysis. Scale bar indicates µm. (D) INCENP staining (red) in a double parked mutant cell showing association of INCENP foci with CID staining (green) of kinetochores (DNA stained blue). Image is a projection of multiple focal planes. Scale bar indicates 5µm. (D ) Same image as (D), but with tubulin staining in green showing merotelic attachments of kinetochores to the spindle. (E) Four focal planes of a cell from D (circled) showing that the kinetochore marked by the arrowhead is associated with kinetochore fibers from both poles. See supplementary data Figure S for the separated channels. Figure : CYC-B S promotes attachment of unpaired anaphase kinetochores to both spindle poles and increases anomalous segregation. In the CYC-B S arrest, kinetochores frequently make merotelic attachments to spindle microtubules and these kinetochores stain strongly for INCENP. Spindle staining is shown in green (A, B, C), INCENP staining in red (A, B, C, D) and DNA staining in blue (A, B, and C ) or CID kinetochore staining in green (C ) or in blue (C). (A) A cell arrested with CYC-B S with disjoined chromosomes in pseudo-metaphase plate and a spindle exhibiting robust kinetochore fibers impinging on the unpaired kinetochores. (B) A CYC-B S expressing cell early in the progress toward arrest with one kinetochore in the spindle midzone with merotelic attachments (arrow). Presumably, this is one of the first of the disjoined chromosomes to return to the midzone after earlier anaphase movements []. (C,C ) This cell appears to be disrupted (and probably retarded) but not arrested by CYC-B S expression, as suggested by the presence of some decondensing chromosomes at the poles. A few kinetochores remain localized in the middle of the spindle and only these stain for INCENP. (F) Frames from a movie of a cell proceeding through mitosis despite expression of CYC-B S. The times are approximate times after metaphase, and indicate greatly retarded anaphase (about times normal). Near the end of anaphase movements, a chromosome (arrowheads) can be seen to make a movement from the lower pole towards the upper pole. We believe this phenomenon to be analogous to the fixed image shown in (C) and to represent CYC-B S stimulated re-orientation of a single chromosome instead of the entire complement. Scale bars on the fixed images indicate µm. See supplementary Figure S for separated channels and an additional example. Figure 4: Model of kinetochore-spindle interactions A model illustrating the different states of kinetochore-spindle interactions in prometaphase and anaphase. Chromosomes are shown in blue, microtubules in black, kinetochores in yellow and the AuroraB/INCENP complex in red. (A) In prometaphase, destabilization of inappropriate attachments by the AuroraB/INCENP complex allows interconversion of the different attachment states until bipolar attachment is achieved. At that point tension pulls the sister kinetochores away from the AuroraB/INCENP complex, more microtubules are recruited and the bipolar state is stabilized (after []). (B) After the metaphase/anaphase transition in wild-type, the AuroraB/INCENP complex dissociates from the kinetochores, ensuring stable attachment during anaphase. (C) In the presence of CYC-B S, the AuroraB/INCENP complex does not dissociate from the kinetochores and the instability that characterizes prometaphase kinetochore-spindle interaction now persists. Since the disjoined kinetochores are unpaired, attachments interconvert without the option of normal bipolar attachment. As is the case with unreplicated prometaphase chromosomes in the double parked mutant, this interconversion (gray double headed arrows at the right) leads to disorderly chromosome movements and merotelic attachments.
9 Parry, Hickson & O Farrell, 00, Curr. Biol. : Supplementary data the progressive relocalization of INCENP during anaphase. In all cases examined, Aurora B staining shows the same localization as that for INCENP (data not shown). Figure SA: INCENP localization in wildtype, CYC-B S expressing and CycB mutant cells. This figure parallels the panels A-C of text Figure. It shows the separated channels for Hoechst, CID, and INCENP as well as staining for α-tubulin. During anaphase in wild-type cells (lefthand panels), INCENP disengages from the kinetochores and moves onto the microtubules of the central spindle during anaphase. Cell # is in metaphase with INCENP foci localized between paired CID staining foci. Cells # and # are anaphase cells that illustrate the progressive decline of INCENP associated with the kinetochores and the accumulation on the spindle. By telophase, cell #4, INCENP is concentrated in the spindle midzone. In CYC-B S -expressing cells (center panels), INCENP foci remain associated with kinetochores in cells expressing CYC-B S. Cell # is late anaphase A (i.e. prior to final arrest). Cells # and # are at a later stage of arrest with kinetochores having lost their orientation towards their designated poles. In CycB mutant cells (righthand panels), INCENP moves off of the kinetochores prematurely during anaphase. Cell # (right subpanels) is in metaphase and cells #-4 are in anaphase and are earlier (#) or roughly comparable (#,4) to the wild-type anaphase of cell # in the left-hand panels. In all cases examined, Aurora B staining shows the same localization as that for INCENP (data not shown). Figure SB: INCENP localization in CYC- B S expressing cells. Shown are the separated channels from Fig. D (Hoechst, CID, INCENP). Unlike CYC-B S, CYC-B S does not block relocalization of INCENP. Cells expressing CYC-B S show normal anaphase movements as they progress to an arrest with chromosomes at separated poles [] and INCENP relocalizes normally during this anaphase. Cell# (B S meta, left) is close to metaphase and cells # (B S -early, center) and # (B S -late, right) show Figure SC: BubR localization in CYC-B S expressing cells. Shown are the separated channels from Fig. E-G with additional α-tubulin staining of the same cells and merged images of DNA (blue), BubR (green) and tubulin (red). BubR disengages from kinetochores at anaphase in both wild-type (lefthand panels) and CYC-B S expressing cells (center panels), but kinetochore localization is later restored in the latter upon arrest (right-hand panels). In wild-type (left-hand panels), BubR localizes to the kinetochores in metaphase (cell # and #), but is absent from the kinetochores during anaphase (cell #). In cells expressing CYC-B S, BubR localizes normally in metaphase (center panels, cell #) and is absent from the kinetochores during the early anaphase A movements (center panels, cell #). However, BubR again localizes to the kinetochores in cells arrested with CYC-B S (right-hand panels). Figure SD: ROD localization in CYC-B S expressing cells. Shown are the separated channels from Fig. H-J with additional α-tubulin staining of the same cells and merged images of DNA (blue), CID (green) and ROD (red). ROD localization (H), CYC-B S expressing (I) and CYC-B S expressing (J) cells. In wild-type cells (left-hand panels), ROD staining can be seen on the kinetochores and along the spindle in metaphase (cell #), while it localizes exclusively to the kinetochores in anaphase (cell #). In cells arrested with CYC-B S (center panels), ROD staining can be seen spread along the spindle, as well as on some of the kinetochores. In cells arrested with CYC-B S (right-hand panels), ROD staining can be seen only at the kinetochores. In all cases examined, ZW0 staining shows the same localization as that for ROD (data not shown).
10 Parry, Hickson & O Farrell, 00, Curr. Biol. : Figure SE: Aurora B localization in CYC- B S expressing cells. Aurora B behaves the same as INCENP (see Figures and SA) and fails to disengage from the kinetochores in a CYC-B S arrest. Two cells are shown stained for hoechst (left panel, blue in the merged image), Aurora B (center-left, red in the merged image), and tubulin (center-right, green in the merged image). Heat shock and antibody staining protocols have been previously described (Parry & O Farrell, 00). The rabbit a-aurora B antibody was a gift from David Glover and was used at a dilution of :00. Figure S: Unreplicated chromosomes in double parked mitosis behave like disjoined chromosomes at CYC-B S arrest. Shown are the separated channels from Fig. D-E with alternative merged images. double parked mutant embryos fail to replicate DNA in S phase of cycle 6 and arrest in the subsequent mitosis. Three cells at such an arrest are shown in the upper panels; as in the CYC-B S arrest, the INCENP staining pattern closely resembles that of CID at kinetochores. The images in the upper panels are projections of multiple focal planes. The lower panels show four consecutive focal planes from the cell circled in the merged image in the upper right-hand panel. It can be seen that the kinetochore marked by the arrowhead is merotelically attached (having kinetochore fibers extending from both poles). Figure S: CYC-B S promotes attachment of unpaired anaphase kinetochores to both spindle poles and increases anomalous segregation. kinetochore fibers impinging on the unpaired kinetochores. Row B shows a CYC-B S expressing cell early in the progress toward arrest with one kinetochore in the spindle midzone exhibiting merotelic attachment. Presumably, this is one of the first of the disjoined chromosomes to return to the midzone after earlier anaphase movements []. The cell in row C appears to be disrupted (and probably retarded) but not arrested by CYC-B S expression, as suggested by the presence of some decondensing chromosomes at the poles. A few kinetochores remain localized in the middle of the spindle and only these stain for INCENP. The cell in row D is another example of a cell that has not been arrested by CYC-B S expression, but is exhibiting a merotelically oriented chromosome whose kinetochore stains for INCENP. The cell in row E is a similar example showing that BubR also associates with kinetochores of merotelically attached chromosomes induced by CYC-B S expression. In the merged images, spindles are in green (rows A, B, D, E), INCENP (rows A, B, C, D) and BubR (row E) are in red, DNA is blue (rows A, C, E) and CID is in green (row C) or blue (rows B, D). Movies Movie : double parked mutant. 0X 7.5 min. Real-time record of double parked mutant embryos carrying the histone-gfp transgene. A QuickTime movie was made using time lapse imaging with the DeltaVision microscopy system. Clock in lower right corner rotates once per minute of elapsed time. In the CYC-B S arrest, kinetochores frequently make merotelic attachments to spindle microtubules and these kinetochores stain strongly for INCENP. Rows A-C show the separated channels from Fig. A-C, while rows D and E are additional images not present in Fig.. Row A shows a single focal plane of a cell arrested with CYC-B S with disjoined chromosomes at a pseudometaphase plate and a spindle exhibiting robust










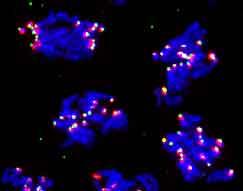
11 Fig. A 4 B C INCENP CID DNA wt D B s B mut 4 B s ~meta E B s -early F B s -late G BubR DNA wt B s -early B s -late ROD CID DNA H wt I B s J B s












12 Fig. A C 0s 7s 4s 5s D 69s 86s D' 04s s B E






13 Fig. A D 68s B 0s C 99s 68s C' 506s
14 Fig. 4 A paired chromosomes in prometaphase state Interconversion of attachment states Bipolar attachments stabilized B wild-type metaphase/anaphase transition Key: AurB/INCENP AurB/INCENP dissociates Attachments stable microtubules kinetochores C CYC-B s metaphase/anaphase transition AurB/INCENP persists Attachments unstable Interconversion of attachment states







15 wild-type CYC-B s B mutant Hoechst CID INCENP α-tubulin Fig. SA







16 Fig. SB B s ~meta B s -early B s -late Hoechst CID INCENP








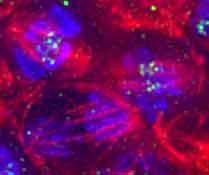


17 Fig. SC wild-type B s -early B s -late Hoechst BubR α-tubulin α-tubulin BubR DNA














18 Fig. SD wild-type B s B s Hoechst CID ROD α-tubulin ROD α-tubulin DNA





19 Fig. S Hoechst CID INCENP α-tubulin merged INCENP α-tubulin CID Hoechst CID INCENP α-tubulin merged INCENP α-tubulin DNA









20 Fig. S Hoechst CID INCENP α-tubulin merged A B C D Hoechst CID BubR α-tubulin merged E
Cyclin B Destruction Triggers Changes in Kinetochore Behavior Essential for Successful Anaphase
 Current Biology, Vol. 13, 647 653, April 15, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/S0960-9822(03)00242-2 Cyclin B Destruction Triggers Changes in Kinetochore Behavior Essential
Current Biology, Vol. 13, 647 653, April 15, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/S0960-9822(03)00242-2 Cyclin B Destruction Triggers Changes in Kinetochore Behavior Essential
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
Bacterial cell. Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Mitosis and the Cell Cycle
 Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Chromosomes Can Congress To The Metaphase Plate Prior. To Bi-Orientation
 Chromosomes Can Congress To The Metaphase Plate Prior To Bi-Orientation Tarun M. Kapoor 1,2, Michael Lampson 1, Polla Hergert 3, Lisa Cameron 2,4, Daniela Cimini 4, E.D. Salmon 2,4, Bruce F. McEwen 3,
Chromosomes Can Congress To The Metaphase Plate Prior To Bi-Orientation Tarun M. Kapoor 1,2, Michael Lampson 1, Polla Hergert 3, Lisa Cameron 2,4, Daniela Cimini 4, E.D. Salmon 2,4, Bruce F. McEwen 3,
Cell Cycle, Mitosis, and Microtubules. LS1A Final Exam Review Friday 1/12/07. Processes occurring during cell cycle
 Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Chapter 8: Cellular Reproduction
 Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Cellular Reproduction, Part 2: Meiosis Lecture 10 Fall 2008
 Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
-The cell s hereditary endowment of DNA -Usually packaged into chromosomes for manageability
 Binary Fission-Bacterial Cell Division -Asexual reproduction of prokaryotes -No mitosis -Circular DNA and organelles replicate, the copies migrate to opposite sides of the elongating cell, and the cell
Binary Fission-Bacterial Cell Division -Asexual reproduction of prokaryotes -No mitosis -Circular DNA and organelles replicate, the copies migrate to opposite sides of the elongating cell, and the cell
The Cell Cycle. Packet #9. Thursday, August 20, 2015
 1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
Lecture 10. G1/S Regulation and Cell Cycle Checkpoints. G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint
 Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
(a) Reproduction. (b) Growth and development. (c) Tissue renewal
 Reproduction. (b) Growth and development. (c) Tissue renewal") 100 µm 200 µm 20 µm (a) Reproduction (b) Growth and development (c) Tissue renewal 1 20 µm 2 0.5 µm Chromosomes DNA molecules Chromosome arm Centromere Chromosome duplication (including DNA synthesis)
100 µm 200 µm 20 µm (a) Reproduction (b) Growth and development (c) Tissue renewal 1 20 µm 2 0.5 µm Chromosomes DNA molecules Chromosome arm Centromere Chromosome duplication (including DNA synthesis)
The Cell Cycle. Dr. SARRAY Sameh, Ph.D
 The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
Biology is the only subject in which multiplication is the same thing as division
 The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
Cell Division and Mitosis
 Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
Cell division functions in 1. reproduction, 2. growth, and 3. repair
 Cell division functions in 1. reproduction, 2. growth, and 3. repair What do you think you are looking at here??? Can something like you or I do this??? Fig. 12.1 How did you start out? How did you grow?
Cell division functions in 1. reproduction, 2. growth, and 3. repair What do you think you are looking at here??? Can something like you or I do this??? Fig. 12.1 How did you start out? How did you grow?
Mitosis THE CELL CYCLE. In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for..
 Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
Chapter 2. Mitosis and Meiosis
 Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach
![klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach](/thumbs/91/104639484.jpg "klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach") DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
How Cells Divide. Chapter 10
 How Cells Divide Chapter 10 Bacterial Cell Division Bacteria divide by binary fission. -the single, circular bacterial chromosome is replicated -replication begins at the origin of replication and proceeds
How Cells Divide Chapter 10 Bacterial Cell Division Bacteria divide by binary fission. -the single, circular bacterial chromosome is replicated -replication begins at the origin of replication and proceeds
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Life Cycles Sexual reproduction mitosis
 meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division
 Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
Creating Identical Body Cells
 Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
Prof. R. V. Skibbens. BIOS 10 and BIOS 90: BioScience in the 21 st Century. Cell Cycle, Cell Division and intro to Cancer.
 Prof. R. V. Skibbens August 31, 2015 BIOS 10 and BIOS 90: BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell Cycle Why a cell cycle? What is the goal? trauma growth development
Prof. R. V. Skibbens August 31, 2015 BIOS 10 and BIOS 90: BioScience in the 21 st Century Cell Cycle, Cell Division and intro to Cancer Cell Cycle Why a cell cycle? What is the goal? trauma growth development
General Biology. Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell division
 General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!!
 Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!! Chromosomes Are made of chromatin: a mass of genetic material composed
Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!! Chromosomes Are made of chromatin: a mass of genetic material composed
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle. 9.1 Multiple-Choice Questions
 Chapter 9 The Cell Cycle. 9.1 Multiple-Choice Questions") Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle 9.1 Multiple-Choice Questions 1) Starting with a fertilized egg (zygote), a series of five cell divisions would produce an early embryo with how
Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle 9.1 Multiple-Choice Questions 1) Starting with a fertilized egg (zygote), a series of five cell divisions would produce an early embryo with how
The Cell Cycle CHAPTER 12
 The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
General Biology. Overview: The Key Roles of Cell Division. Unicellular organisms
 General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
Chapter 10 Cell Growth and Division
 Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Ploidy and Human Cell Types. Cell Cycle and Mitosis. DNA and Chromosomes. Where It All Began 11/19/2014. Chapter 12 Pg
 Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Name. A.P. Biology Chapter 12 The Cell Cycle
 A.P. Biology Chapter 12 The Cell Cycle Name Living species MUST possess the ability to r if they are to flourish. The Cell Cycle follows the life of a cell from its o until its d. The Key Roles Of Cell
A.P. Biology Chapter 12 The Cell Cycle Name Living species MUST possess the ability to r if they are to flourish. The Cell Cycle follows the life of a cell from its o until its d. The Key Roles Of Cell
DAPI ASY1 DAPI/ASY1 DAPI RAD51 DAPI/RAD51. Supplementary Figure 1. Additional information on meiosis in R. pubera. a) The
 The") a % 10 Number of crossover per bivalent b 0 1 c DAPI/telomere 80 1 60 40 1 2 20 d 0 0 1 2 >=3 DAPI ASY1 DAPI/ASY1 e DAPI RAD51 DAPI/RAD51 Supplementary Figure 1. Additional information on meiosis in R.
a % 10 Number of crossover per bivalent b 0 1 c DAPI/telomere 80 1 60 40 1 2 20 d 0 0 1 2 >=3 DAPI ASY1 DAPI/ASY1 e DAPI RAD51 DAPI/RAD51 Supplementary Figure 1. Additional information on meiosis in R.
Chapter 10. Cell Growth and Division
 Chapter 10 Cell Growth and Division Cell Growth A. Limits to Cell Growth 1. Two main reasons why cells divide: a. Demands on DNA as the cell get too large Cell Growth b. Moving nutrients and waste across
Chapter 10 Cell Growth and Division Cell Growth A. Limits to Cell Growth 1. Two main reasons why cells divide: a. Demands on DNA as the cell get too large Cell Growth b. Moving nutrients and waste across
基醫所. The Cell Cycle. Chi-Wu Chiang, Ph.D. IMM, NCKU
 基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
Monday, October 6 Put these items into the appropriate category:
 Monday, October 6 Put these items into the appropriate category: Active Transport Facilitated Diffusion Osmosis Simple Diffusion The smell of rotten eggs spreading through the room Requires ATP expenditure
Monday, October 6 Put these items into the appropriate category: Active Transport Facilitated Diffusion Osmosis Simple Diffusion The smell of rotten eggs spreading through the room Requires ATP expenditure
Genetics. Instructor: Dr. Jihad Abdallah Lecture 2 The cell cycle and Cell Division
 Genetics Instructor: Dr. Jihad Abdallah Lecture 2 The cell cycle and Cell Division 1 The cell cycle Living cells go through a series of stages known as the cell cycle. They undergo a continuous alternation
Genetics Instructor: Dr. Jihad Abdallah Lecture 2 The cell cycle and Cell Division 1 The cell cycle Living cells go through a series of stages known as the cell cycle. They undergo a continuous alternation
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Genetics and Cellular Function
 Genetics and Cellular Function DNA replication and the cell cycle Mitosis Mitosis Mitosis: division of cells that results in daughter cells with the same the genetic information that the original cell
Genetics and Cellular Function DNA replication and the cell cycle Mitosis Mitosis Mitosis: division of cells that results in daughter cells with the same the genetic information that the original cell
BIOLOGY 4/6/2015. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
Name: Date: Block: 10-2 Cell Division Worksheet
 10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
8.4 The cell cycle multiplies cells. 8.4 The cell cycle multiplies cells
 8.4 The cell cycle multiplies cells! Cell division is a highly orchestrated process! The cell cycle is an ordered sequence of events that extends from the time a cell is first formed from a dividing parent
8.4 The cell cycle multiplies cells! Cell division is a highly orchestrated process! The cell cycle is an ordered sequence of events that extends from the time a cell is first formed from a dividing parent
Cell Division. During interphase, a cell s DNA is in a loose form called. It condenses into tightly coiled structures called chromosomes during.
 Cell Division The is a cell s total DNA. Prokaryotes DNA is found mostly in a single called the and also in small circles called. Eukaryotes have several DNA double helices packaged into. During interphase,
Cell Division The is a cell s total DNA. Prokaryotes DNA is found mostly in a single called the and also in small circles called. Eukaryotes have several DNA double helices packaged into. During interphase,
How deregulated cell cycles (might) trigger cancer. Dan Fisher. Friday 27th October Lucie Fisher, 3 Eddie Fisher, 6.
 trigger cancer. Dan Fisher. Friday 27th October Lucie Fisher, 3 Eddie Fisher, 6.") How deregulated cell cycles (might) trigger cancer http://www.igmm.cnrs.fr Dan Fisher Friday 27th October 2017 Lucie Fisher, 3 Eddie Fisher, 6 1 transformed cells transformed cells Structure of this lecture
How deregulated cell cycles (might) trigger cancer http://www.igmm.cnrs.fr Dan Fisher Friday 27th October 2017 Lucie Fisher, 3 Eddie Fisher, 6 1 transformed cells transformed cells Structure of this lecture
Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division
 The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
BIOLOGY. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
Mitosis in Onion Root Tip Cells
 Mitosis in Onion Root Tip Cells A quick overview of cell division The genetic information of plants, animals and other eukaryotic organisms resides in several (or many) individual DNA molecules, or chromosomes.
Mitosis in Onion Root Tip Cells A quick overview of cell division The genetic information of plants, animals and other eukaryotic organisms resides in several (or many) individual DNA molecules, or chromosomes.
10.2 The Cell Cycle *
 OpenStax-CNX module: m52672 1 10.2 The Cell Cycle * Shannon McDermott Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
OpenStax-CNX module: m52672 1 10.2 The Cell Cycle * Shannon McDermott Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
BIOLOGY - CLUTCH CH.12 - CELL DIVISION.
 !! www.clutchprep.com CONCEPT: CELL DIVISION Cell division is the process by which one cell splits into two or more daughter cells. Cell division generally requires that cells produce enough materials,
!! www.clutchprep.com CONCEPT: CELL DIVISION Cell division is the process by which one cell splits into two or more daughter cells. Cell division generally requires that cells produce enough materials,
The Cell Cycle. Chapter 12. PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
Unduplicated. Chromosomes. Telophase
 10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
Chapter 8 The Cell Cycle
 What molecule stores your genetic information or determines everything about you? DNA a nucleic acid How are DNA molecules arranged in the nucleus? As you can see DNA is: Chapter 8 The Cell Cycle 1. Arranged
What molecule stores your genetic information or determines everything about you? DNA a nucleic acid How are DNA molecules arranged in the nucleus? As you can see DNA is: Chapter 8 The Cell Cycle 1. Arranged
10-2 Cell Division. Slide 1 of 38. End Show. Copyright Pearson Prentice Hall
 1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
Histones- protein molecules that are used to fold and package DNA into chromosomes.
 Chromosome- a portion of the DNA in a cell, a chromosome is created when the DNA segment coils around histones then twists further to create a long twisted mass. Histones- protein molecules that are used
Chromosome- a portion of the DNA in a cell, a chromosome is created when the DNA segment coils around histones then twists further to create a long twisted mass. Histones- protein molecules that are used
UNC-Duke Biology Course for Residents Fall Cell Cycle Effects of Radiation
 UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
The Cell Cycle 4/10/12. Chapter 12. Overview: The Key Roles of Cell Division
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Chapter 12. living /non-living? growth repair renew. Reproduction. Reproduction. living /non-living. fertilized egg (zygote) next chapter
 next chapter") Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Cell Cycle and Mitosis
 Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
Why do cells reproduce?
 Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis Examples of Cell
Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis Examples of Cell
The faithful separation of chromosomes prior to cell division at
 Modeling the temporal evolution of the spindle assembly checkpoint and role of Aurora B kinase Hitesh B. Mistry a, David E. MacCallum b, Robert C. Jackson b, Mark A. J. Chaplain a, and Fordyce A. Davidson
Modeling the temporal evolution of the spindle assembly checkpoint and role of Aurora B kinase Hitesh B. Mistry a, David E. MacCallum b, Robert C. Jackson b, Mark A. J. Chaplain a, and Fordyce A. Davidson
NOTHING TO DECLARE. Eugene Pergament, MD, PhD. FACMG. Northwestern Reproductive Genetics, Inc ORIGIN OF ANEUPLOIDY
 NOTHING TO DECLARE Eugene Pergament, MD, PhD. FACMG Northwestern Reproductive Genetics, Inc CHROMOSOME SEGREGATION ERRORS 1-2% 8-9% ~90% Origin of Aneuploidy 1. Cohesin complex 2.Kinetochore-Microtubule
NOTHING TO DECLARE Eugene Pergament, MD, PhD. FACMG Northwestern Reproductive Genetics, Inc CHROMOSOME SEGREGATION ERRORS 1-2% 8-9% ~90% Origin of Aneuploidy 1. Cohesin complex 2.Kinetochore-Microtubule
Regulation of Cell Division
 Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Regulation of Cell Division Two HeLa cancer cells are just completing cytokinesis. Explain how the cell division of cancer cells like these is misregulated. Identify genetic and other changes that might
Biology is the only subject in which multiplication is the same thing as division. AP Biology
 Biology is the only subject in which multiplication is the same thing as division Chapter 12. The Cell Cycle: Cell Growth, Cell Division Where it all began You started as a cell smaller than a period at
Biology is the only subject in which multiplication is the same thing as division Chapter 12. The Cell Cycle: Cell Growth, Cell Division Where it all began You started as a cell smaller than a period at
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Chapter 12. The Cell Cycle
 Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Mitosis vs. Meiosis. The Somatic Cell Cycle (Mitosis) The somatic cell cycle consists of 3 phases: interphase, m phase, and cytokinesis.
 The somatic cell cycle consists of 3 phases: interphase, m phase, and cytokinesis.") Mitosis vs. Meiosis In order for organisms to continue growing and/or replace cells that are dead or beyond repair, cells must replicate, or make identical copies of themselves. In order to do this and
Mitosis vs. Meiosis In order for organisms to continue growing and/or replace cells that are dead or beyond repair, cells must replicate, or make identical copies of themselves. In order to do this and
The Cell Cycle and How Cells Divide
 The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
10-2 Cell Division. Chromosomes
 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
Regulation of Cell Division. AP Biology
 Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
Regulation of Cell Division 2006-2007 Coordination of cell division A multicellular organism needs to coordinate cell division across different tissues & organs critical for normal growth, development
Bis2A 15.0 The Cell Cycle *
 OpenStax-CNX module: m56084 1 Bis2A 15.0 The Cell Cycle * Mitch Singer Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
OpenStax-CNX module: m56084 1 Bis2A 15.0 The Cell Cycle * Mitch Singer Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis
 Mitosis") Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis Chromosomes Chromosomes were first observed by the German embryologist Walther Fleming in 1882. Chromosome number varies among organisms most
Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis Chromosomes Chromosomes were first observed by the German embryologist Walther Fleming in 1882. Chromosome number varies among organisms most
BIOLOGY. The Cell Cycle CAMPBELL. Reece Urry Cain Wasserman Minorsky Jackson. Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick
 CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 12 The Cell Cycle Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick The Key Roles of Cell Division The ability
CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 12 The Cell Cycle Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick The Key Roles of Cell Division The ability
Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis
 Mitosis") Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis Prokaryotes Have a Simpler Cell Cycle Cell division in prokaryotes takes place in two stages, which together make up a simple cell cycle 1. Copy
Breaking Up is Hard to Do (At Least in Eukaryotes) Mitosis Prokaryotes Have a Simpler Cell Cycle Cell division in prokaryotes takes place in two stages, which together make up a simple cell cycle 1. Copy
Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa
 HeLa") SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney
 2013 Course University of Sydney") BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
BCHM3972 Human Molecular Cell Biology (Advanced) 2013 Course University of Sydney Page 2: Immune Mechanisms & Molecular Biology of Host Defence (Prof Campbell) Page 45: Infection and Implications for Cell
Cell Division (Mitosis)
") Cell Division (Mitosis) Chromosomes The essential part of a chromosome is a single very long strand of DNA. This DNA contains all the genetic information for creating and running the organism. Each chromosome
Cell Division (Mitosis) Chromosomes The essential part of a chromosome is a single very long strand of DNA. This DNA contains all the genetic information for creating and running the organism. Each chromosome
Why do cells divide? The Cell Cycle: Cell Growth, Cell Division. Making new cells. Getting the right stuff. Overview of mitosis 1/5/2015
 Why do cells divide? The Cell Cycle: Cell Growth, Cell Division For reproduction asexual reproduction one-celled organisms For growth from fertilized egg to multi-celled organism For repair & renewal replace
Why do cells divide? The Cell Cycle: Cell Growth, Cell Division For reproduction asexual reproduction one-celled organisms For growth from fertilized egg to multi-celled organism For repair & renewal replace
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
General Embryology. School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan
 General Embryology 2019 School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan https://www.facebook.com/dramjad-shatarat What is embryology? Is the science that
General Embryology 2019 School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan https://www.facebook.com/dramjad-shatarat What is embryology? Is the science that
Mitosis. AND Cell DiVISION
 Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
Mitosis/Meiosis Simulation Activities
 Mitosis/Meiosis Simulation Activities In this simulation, you will demonstrate an understanding of mitosis, meiosis, segregation, independent assortment, and crossing over, all processes involved with
Mitosis/Meiosis Simulation Activities In this simulation, you will demonstrate an understanding of mitosis, meiosis, segregation, independent assortment, and crossing over, all processes involved with
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Courtheoux et al., http://www.jcb.org/cgi/content/full/jcb.200902093/dc1 S1 Figure S2. Visualization of multiple merotelic attachments.
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Courtheoux et al., http://www.jcb.org/cgi/content/full/jcb.200902093/dc1 S1 Figure S2. Visualization of multiple merotelic attachments.
Mitosis Notes AP Biology Mrs. Laux
 I. Cell Cycle-includes interphase and mitosis (IPPMAT) A. Interphase 1. accounts for 90% of the cycle 2. cell grows and copies its chromosomes in preparation for cell division 3. produces proteins and
I. Cell Cycle-includes interphase and mitosis (IPPMAT) A. Interphase 1. accounts for 90% of the cycle 2. cell grows and copies its chromosomes in preparation for cell division 3. produces proteins and
Chapter 4 The Chromosome Theory of Inheritance
 Chapter 4 The Chromosome Theory of Inheritance 4-1 Sections to study 4.1 Chromosomes: The carriers of genes 4.2 Mitosis: Cell division that preserves chromosome number 4.3 Meiosis: Cell division that halve
Chapter 4 The Chromosome Theory of Inheritance 4-1 Sections to study 4.1 Chromosomes: The carriers of genes 4.2 Mitosis: Cell division that preserves chromosome number 4.3 Meiosis: Cell division that halve
2014 Pearson Education, Inc.
 2 The Cell Cycle CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson The Key Roles of Cell Division The ability of organisms to produce more of their own kind best distinguishes living
2 The Cell Cycle CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson The Key Roles of Cell Division The ability of organisms to produce more of their own kind best distinguishes living
Chapter 8. The Cellular Basis of Reproduction and Inheritance. Lecture by Mary C. Colavito
 Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for Biology: Concepts & Connections, Sixth Edition Campbell, Reece, Taylor, Simon, and Dickey Copyright 2009 Pearson Education,
Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for Biology: Concepts & Connections, Sixth Edition Campbell, Reece, Taylor, Simon, and Dickey Copyright 2009 Pearson Education,
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
Cellular Reproduction
 9 Cellular Reproduction section 1 Cellular Growth Before You Read Think about the life cycle of a human. On the lines below, write some of the stages that occur in the life cycle of a human. In this section,
9 Cellular Reproduction section 1 Cellular Growth Before You Read Think about the life cycle of a human. On the lines below, write some of the stages that occur in the life cycle of a human. In this section,
Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide)
") Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide) Meiosis: cell division that forms daughter cells with half the number
Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide) Meiosis: cell division that forms daughter cells with half the number
Essential Questions. Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase?
 Essential Questions Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase? Cellular Growth Vocabulary Review selective permeability New cell cycle
Essential Questions Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase? Cellular Growth Vocabulary Review selective permeability New cell cycle
10-2 Cell Division mitosis. cytokinesis. Chromosomes chromosomes Slide 1 of 38
 In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. The cell cycle has 4 main stages. The cell cycle is a regular
 Chapter 10 Chapter 10 KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. The cell cycle has 4 main stages. The cell cycle is a regular pattern of growth, DNA replication,
Chapter 10 Chapter 10 KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. The cell cycle has 4 main stages. The cell cycle is a regular pattern of growth, DNA replication,
Chapter 12 The Cell Cycle: Cell Growth, Cell Division
 Chapter 12 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a period at the end of a sentence And now look at you How did you get from there to
Chapter 12 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a period at the end of a sentence And now look at you How did you get from there to