Kaposi s Sarcoma-Associated Herpes Virus (KSHV/HHV-8) Induces a Sustained ACCEPTED. NF-κB Activation During de novo Infection of Primary Human Dermal
|
|
|
- Gary Stanley
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on February 00 J. Virol. doi:./jvi.0-0 Copyright 00, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved JVI 0-0 REVISED MANUSCRIPT Kaposi s Sarcoma-Associated Herpes Virus (KSHV/HHV-) Induces a Sustained NF-κB Activation During de novo Infection of Primary Human Dermal Microvascular Endothelial Cells that is Essential for Viral Gene Expression. Sathish Sadagopan, Neelam Sharma-Walia, Mohanan Valiya Veettil, Hari Raghu, Ramu Sivakumar, Virginie Bottero and Bala Chandran* Department of Microbiology and Immunology, H.M. Bligh Cancer Research Laboratories, Chicago Medical School, Rosalind Franklin University of Medicine and Science, North Chicago, IL. Running Title: Sustained NF-κB activation by KSHV. *Corresponding author. Mailing Address: Department of Microbiology and Immunology, Chicago Medical School, Rosalind Franklin University of Medicine and Science, Green Bay Road, North Chicago, IL 00. Phone () -; Fax () -. bala.chandran@rosalindfranklin.edu Downloaded from on December 1, 01 by guest 1
2 ABSTRACT In vitro KSHV infection of primary human dermal microvascular endothelial (HMVEC-d) and foreskin fibroblast (HFF) cells is characterized by the induction of preexisting host signal cascades, sustained expression of latency-associated genes, transient expression of a limited number of lytic genes, and induction of several cytokines, growth and angiogenic factors. Since NF-κB is a key molecule involved in regulation of several of these factors, here, we examined NF-κB induction during de novo infection of HMVEC-d and HFF cells. Activation of NF-κB was observed as early as to 1 min post infection by KSHV and translocation of p-nf-κb into nuclei was detected by IFA, EMSA and p-elisa. IκB phosphorylation inhibitor (Bay-0) reduced this activation significantly. A sustained moderate level of NF-κB induction was observed during the observed h of in vitro KSHV latency. In contrast, high levels of ERK1/ activation during earlier time points and a moderate level of activation at later times were observed. The pmapkinase was activated only during later time points, and AKT was activated in a cyclic manner. Studies with UV-KSHV suggested a role for virus entry stages in NF-κB induction and requirement of KSHV viral gene expression for sustained induction. Inhibition of NF-κB did not affect target cell entry by KSHV but significantly reduced the expression of viral latent ORF and lytic genes. KSHV infection induced activation of several host transcription factors including AP-1 family as well as several cytokines growth factors and angiogenic factors, which were significantly affected by NF-κB inhibition. These results suggest that during de novo infection, KSHV induces the sustained levels of NF-κB to regulate viral and host cell genes, and thus possibly regulating the establishment of latent infection. Downloaded from on December 1, 01 by guest
3 INTRODUCTION Kaposi s sarcoma (KS) is a chronic inflammation associated malignancy characterized by spindle shaped endothelial cells, inflammatory cells, growth factors and cytokines. KS is an AIDS-defining vascular tumor and in the absence of HIV-1 infection, KS occurs in three distinct epidemiologic forms, the classic KS (CKS), endemic aggressive KS (EKS), and transplantation associated KS (TKS). Chang et al (1) reported the identification of a novel herpesvirus DNA sequence, Kaposi s sarcoma associated herpesvirus (KSHV/HHV-), in all four types of KS lesions, suggesting a potential common etiological factor for KS. KSHV is also etiologically associated with two lymphoproliferative disorders, namely body cavity-based B-cell lymphoma (BCBL) or primary effusion lymphoma (PEL) () and some forms of multicentric Castleman s disease (MCD). BCBL cell lines such as BCBL-1 and BC- carry KSHV in a latent form, and a lytic cycle can be induced by chemical agents (). KSHV DNA and transcripts have been detected in B cells from the peripheral blood, B cells in BCBL and MCD, flat endothelial cells lining the vascular spaces of KS lesions, typical KS spindle cells, CD+/CD+ monocytes in KS lesions, keratinocytes, and epithelial cells (1, 1, ). KSHV DNA is present in a latent form in the vascular endothelial and spindle cells of KS tissues and expression of latency associated LANA-1 (ORF ), v-cyclin D (ORF ), v-flip (K1) and Kaposin (K1) genes have been demonstrated in these cells (1, 1,,, ). Lytic infection is also detected in KS lesions with <1% of infiltrating inflammatory monocytic cells positive for lytic cycle proteins (1, 1). In addition, KSHV lytic cycle K gene expression has been also detected in the endothelial cells and spindle cells of KS tumors (0, ). Downloaded from on December 1, 01 by guest
4 KSHV infects a variety of in vitro target cells such as human B, endothelial, epithelial, and fibroblast cells (1, ). We have previously demonstrated that within min post infection (p.i.) of adherent target cells, KSHV induced the hosts cell pre-existing signal pathways such as FAK, Src, PI-K, RhoGTPases, PKCζ, MEK1/ and ERK1/ (,, ). In contrast to alpha and beta-herpesviruses, in vitro infection by KSHV does not result in a productive lytic cycle. Instead, KSHV infection of HMVEC-d and HFF cells is characterized by the sustained expression of latency associated ORFs, and K1 genes. A unique aspect of this in vitro infection is our demonstration of concurrent expression of a limited set of lytic cycle genes with anti-apoptotic and immune- modulation functions including lytic cycle switch ORF 0 or RTA gene (0). While the expression of latent ORF,, and K1 genes continues, nearly all lytic genes decline (, 0). Further examination revealed a steady quantitative increase in early lytic K, K and v-irf gene expression (). KSHV-K gene expression persists throughout the day period of observation (0), and down regulation of MHC class I A and C, ICAM-1, CD1/PECAM and B- molecules could be detected up to days in the infected HMVEC-d cells (1, 0, 0). Similar to our observation, very early ORF 0 expression and subsequent decline were also seen during primary KSHV infection of human cells (). Bechtel et al () showed that of the RNA transcripts detected in our system encoding ORFs such as K.1, K1, ORF / and ORF were present in the purified virion particles. However, other transcripts detected by us were absent suggesting de novo transcription of the remaining lytic genes during the initial hours of infection. The characteristic expression of limited lytic cycle genes could be a strategy which allows KSHV to evade the immune system, and to provide necessary factors and Downloaded from on December 1, 01 by guest
5 time to establish and/or maintain latency during the initial phases of infection. Establishment of latent infection by KSHV thus provides a good in vitro model for studying viral and host factors involved in the establishment and maintenance of latent infection. How KSHV achieves selective activation of RTA-responsive genes without initiating the full lytic cascade is a challenge to the understanding of KSHV latency. The newly synthesized ORF protein can potentially influence the functions of ORF 0 since LANA-1 has been shown to counter transactivation of certain lytic promoters by RTA. However, this may not be fully effective at early times during primary infection in the presence of abundant RTA (0). Our hypothesis is that KSHV induced signal cascades and host cell reprogramming induced during primary infection may be playing an important role in the establishment of latent infection and in suppression of the lytic cycle. As an initial step toward understanding how KSHV establishes in vitro latent infection, we have previously examined the modulation of host cell gene expression at and h p.i. of primary HMVEC-d and HFF cells using oligonucleotide arrays (). We observed the reprogramming of host transcription regulating apoptosis, cell cycle regulation, signaling, inflammatory response, and angiogenesis (). Notable among these is the strong induction of several pro-inflammatory cytokines and growth factors (). Since several of these factors could be induced by NF-κB, here, we examined the induction of NF-κB early during target cell infection and its role in KSHV infection. NF-κB belongs to a highly conserved family of transcription factors with an N- terminal Rel homology domain and a C-terminal transactivation domain, that includes c- rel, p0 (NF-κB 1 ), p (NF-κB ), p (rela) and relb (,, 1). Each of these Downloaded from on December 1, 01 by guest
6 polypeptides can form homodimers or dimerize with other Rel family members and the prototype NF-κB is composed of p0 and p. The function of NF-κB is regulated by a series of inhibitory molecules named IκBs. IκB molecules sequester NF-κB in the cytoplasm thus rendering them inactive. Post-translational modifications of IκBα, induced by various stimuli or viral infection that activate different signal transduction pathways, results in the activation of IκBα and subsequent proteolytic degradation. This causes the release of NF-κB that translocates to the nucleus and transcribes NF-κB dependent target genes. In lymphocytes, the IκB proteins are unstable, and high levels of NF-κB are constitutively present in the nucleus (). In B-lymphoma cells latently infected with human γ-herpesviruses like EBV and KSHV, NF-κB activity is further elevated by the expression of latent viral gene products that activate the NF-κB signaling pathway (1, 1, ). In EBV and KSHV infections, the latency associated proteins like LMP1 and vflip, respectively, have been shown to be responsible for the sustained activation of NF-κB (, ). Blocking NF-κB is known to disturb latency and down regulate NF-κB inducible cytokines resulting in apoptosis (, ). However, the role played by NF-κB during primary infection of endothelial cells has not been studied. In this study, we examined the induction of NF-κB during KSHV de novo infection of primary HMVEC-d and HFF cells and present a comprehensive view of the NF-κB and other factors induced during early and late stages of infection. We demonstrate that KSHV binding to target cells results in an early induction of NF-κB which is maintained at a sustained moderate level throughout the h time of observation, and activated NF-κB via the AP-1 family of transcription factors, plays a role in the regulation of viral and host genes such as cytokines and growth factors. Downloaded from on December 1, 01 by guest
7 MATERIALS AND METHODS Cells. Human foreskin fibroblast (HFF) cells (Clonetics, Walkersville, MD) were grown in DMEM (Gibco BRL, Grand Island, N.Y.) medium supplemented with % heat inactivated fetal bovine serum (FBS) (Hyclone, Logan, UT), mm L-glutamine and antibiotics. Human dermal microvascular endothelial cells (HMVEC-d; CC-, Clonetics) were maintained in endothelial basal media- (EBM-) in the presence of necessary growth factors (Clonetics). Cells were maintained in % CO at C. KSHV carrying BCBL-1 cells were cultured in RPMI (Gibco BRL, Grand Island, NY) medium with % heat inactivated FBS, L-Glutamine and antibiotics (). Recombinant green fluorescent protein-kshv (GFP-KSHV- KSHV.1)- carrying BCBL-1 cells (1) were cultured in RPMI (Gibco BRL) medium (). Antibodies, substrates and chemicals. Chemicals of the highest purity available were purchased. Rabbit antibodies detecting the phosphorylated forms of ERK1/ (Thr 0/Tyr 0 phospho-p/ MAP kinase), p MAP kinase (Thr/Tyr1 phospho-p MAPK), AKT, anti-mouse phospho p, total p, total AKT, total p antibodies and U01 (1,-Diamino-, -dicyano-1, -bis [-amino phenylthio] butadiene) were from Cell Signaling Technology, Beverly, Mass. Total ERK antibody was from Santa Cruz Biotechnology Inc, CA. LY00 (0(-morphodinyl)--phenyl-1 (H)-benzopyran--one), Bay-0 [(E)--(- Methylphenylsulfonyl) --propenenitrile], heparin, antibodies to β-tubulin and β-actin (CloneAC-0) antibodies were obtained from Sigma, St.Louis, MO. Anti-rabbit and anti-mouse- HRP or ALP linked antibodies were from Kirkegaard & Perry Laboratories, Inc., Gaithersberg, Md. Secondary antibodies for immunofluorescence were purchased from Molecular Probes- Invitrogen Corp., Carlsbad, CA. Downloaded from on December 1, 01 by guest
8 Virus. KSHV lytic cycle was induced in BCBL-1 cells and virus from the supernatants were purified as per procedures described previously (). Briefly, BCBL-1 cells were stimulated with 0ng/ml of TPA (Sigma) for days and virus in the spent culture medium was concentrated and DNase treated. UV inactivation of virus was done to prepare replication defective KSHV by inactivating KSHV with UV light (nm) for 0 min at a -cm distance followed by DNase treatment (). DNA was extracted from live and UV-KSHV, and viral DNA copy numbers were quantified by real-time DNA-PCR using primers amplifying the KSHV ORF gene (0). Cytotoxicity assay. Target cells were tested for their viability at various time points post serum starvation and in the presence of various concentrations of Bay-0 at C for different time points (, ). EBM- and DMEM containing different concentrations of various inhibitors were incubated with HMVEC-d cells for h. At different time points, supernatants were collected and assessed for the cellular toxicity using an LDH cytotoxicity assay kit (Promega, Madison, Wis). Percentage viability of HMVEC-d cells after h, h, h and h of serum starvation were %, 1%, % and 1%, respectively. Viability of cells after h incubation with µm, µm, µm, 1µM, 0µM, µm, 0µM, µm and 0µM concentrations of Bay-0 were %, %, %, %, %, 1%, %, % and %, respectively. Western blotting. Target cells grown to confluence in -cm flasks were serum starved and induced with KSHV ( MOI - Multiplicity of Infection or DNA copies/cell) at C. For inhibitor studies, cells were exposed to NF-κB inhibitor (Bay-0) for 1 h at C prior to KSHV infection. After treatment, cells were washed twice with phosphate buffered saline (PBS, ph.), and total protein was extracted. Total cell lysates (µg) were resolved on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels, transferred to nitrocellulose membranes, and immunoblotted with antibodies. Immunoreactive bands were Downloaded from on December 1, 01 by guest
9 developed by enhanced chemiluminescence reaction (NEN Life Sciences Products, Boston, Mass.) and quantified following standard protocols (). Immunofluorescence assay. HMVEC-d cells and HFF cells grown in eight-well chamber slides (% confluence) (Nalge Nunc International, Naperville, Il.) were serum starved, Bay-0 pretreated or left untreated and incubated with KSHV for 0 min and min, respectively. For colocalization studies HMVEC-d cells were infected with KSHV for h in serum free EBM- media, followed by the addition of EBM- with serum and growth factors and incubated for an additional h. GFP-KSHV infection was done as per procedures described previously (). At respective time points, cells were washed with PBS, fixed with.% paraformaldehyde for min at RT, permeabilised for min with 0.1% Triton X-0, and blocked for min with % BSA in PBS. The cells were incubated with a 1:00 dilution of rabbit anti-p antibody or anti LANA antibody for 1 h at room temperature followed by incubation with goat anti-rabbit antibody labeled with Alexa fluor (Molecular Probes-Invitrogen Corp). After washing with PBS, the cells were mounted with anti-fade reagent containing DAPI (, - diamidino-- phenylindole) and observed under a fluorescent microscope equipped with the Nikon Metamorph digital imaging system. Nuclear extracts preparation. HMVEC-d cells and HFF cells were left untreated or pretreated with Bay-0 at C for 1 h and infected with KSHV ( DNA copies/cell) for 1 min, 0 min and 0 min. Nuclear extracts were prepared using Nuclear Extract Kit (Active Motif Corp, Carlsbad, CA) as per manufacturer s instructions. After measuring protein concentrations by BCA protein assay reagent (Pierce Biotechnology, Rockford, IL), extracts were stored at 0 C. Purity of the nuclear extracts was assessed by immunoblotting using anti-lamin B antibodies, and cytoskeletal contamination was checked for by using anti-β-actin and anti-β-tubulin antibodies. Downloaded from on December 1, 01 by guest
10 NF-κB DNA binding assay: µg of Bay-0 pretreated and untreated nuclear extracts were assayed for activated NF-κB by an ELISA based assay kit (Active Motif). This assay, which has been reported to be more sensitive than the gel shift assay, uses well plates coated with oligonucleotides containing the NF-κB consensus sequence ( -GGGACTTTCC- ). Excess (0 pmol) mutant probe ( AGTTGAGGCCATTTC CCAGGC- ) and wild type probe ( - AGTTGAGGGGACTTTCCCAGGC- ) were added to reactions in the competition experiments. Plates were washed three times in wash buffer (PBS, 0.1% Tween 0), incubated with an HRP conjugated anti-rabbit immunoglobulin G (IgG) antibody for 1 h, washed times, incubated with 0µl of developing solution for - min, followed by addition of 0µl stop solution as per the manufacturer s instructions. Plates were read with an ELISA plate reader at 0nm with a reference wavelength of nm. Electrophoretic mobility shift assay (EMSA). The κb, Oct1 consensus oligonucleotides were obtained from Santa Cruz Biotechnology, Inc. The double stranded oligonucleotides were labeled at the ' end with [γ P]-ATP (Perkin Elmer) using T polynucleotide kinase (Gibco BRL). Binding reactions were incubated on ice for 0 min and were performed in 0 µl reaction volume containing 0 mmol/l NaCl, mmol/l Tris-HCl ph., 1 mmol/l MgCl, 0. mmol/l EDTA, 0. mmol/l DTT, % (v/v) glycerol, 1 µg poly (di-dc), µg nuclear extract and labeled probe (,000 cpm). The resulting DNA-protein complexes were then size fractionated from the free DNA probe by electrophoresis at 00 V on a % native polyacrylamide gel. The gel was Downloaded from on December 1, 01 by guest 0 dried at 0 C and autoradiographed. Competition EMSA was performed by adding a 0X 1 molar excess of unlabeled double-stranded, κb oligonucleotide probe. Nucleotide sequences of the annealed DNA probes for κb consensus and Oct1 consensus used were; κb consensus: ( -
11 AGTTGAGGGGACTTTCCCAGGC- ) and Oct-1 consensus ( - TGTGGAATGCAAATCACTAGAA- ) Measurement of KSHV internalization by real-time DNA PCR. Target cells that were left untreated or treated with inhibitor were infected with KSHV at DNA copies/cell. After h incubation, cells were washed twice with PBS to remove the unbound virus, treated with trypsin- EDTA for min at C to remove the bound but non-internalized virus, washed, and total DNA was isolated using a DNeasy kit (Qiagen Valencia, CA). A total of 0 ng of DNA samples, KSHV-ORF gene TaqMan probe (0), gene specific primers and Quantitect PCR mix were used. The KSHV ORF gene, cloned in the pgem-t vector (Promega), was used for the external standard. Known amounts of ORF plasmid were used in the amplification reactions along with the test samples. The lower limit of ORF gene detection was to 0 copies, and the most accurate detection was from 0 to copies. The cycle threshold (C t ) values were used to plot the standard graph and to calculate the relative copy numbers of viral DNA in the samples. Real-time RT-PCR. KSHV infected cells, untreated or pretreated with Bay-0 prior to infection were washed twice with 1X PBS to remove the unbound virus, lysed with RLT buffer (Qiagen), and the monolayer was scraped to collect the lysate. Total RNA was isolated from the lysate (1 min, 0 min, 0 min, 0 min, h, h and h p.i.) using RNeasy kits (Qiagen) according to the manufacturer s protocols, quantified spectrophotometrically and stored at 0 C. The ORF 0, ORF, K, K, and v-irf transcripts were detected by real-time RT-PCR using specific real-time primers and specific TaqMan probes as described previously (). The expression levels of the viral transcripts were normalized to GAPDH gene expression. Downloaded from on December 1, 01 by guest
12 Transcription factor activation assay. µg of each nuclear extract was used for this assay. Transcription factors FosB, cfos, Fra1, Fra, phospho-c-jun, JunB and JunD in the nuclear extracts were measured using the ELISA-based TransAM AP-1 Family kit (Active Motif Corp.) as per manufacturer s instructions. In this assay, transcription factors bind to the immobilized oligonucleotide containing the consensus sequences specific for the particular transcription factor, which is then detected by a sandwich ELISA. The active form of the transcription factor contained in the nuclear extract specifically binds to this oligonucleotide mixture. The primary antibodies used to detect each of the AP-1 transcription factors recognize an epitope in the phosphorylated-c-jun, JunB, JunD, cfos, FosB, Fra1 and Fra that is accessible only when these transcription factors are activated and bound to their target DNA. The detection limit for the TransAM AP-1 Family kit is <0. µg nuclear extract/well. Competition assay was done by premixing nuclear extracts for 0 min at C with wild-type (wt) consensus and mutated (mut) consensus oligonucleotides provided in the kit before adding it to the probe immobilized on the plate. Cytokine array: Conditioned media obtained by centrifuging serum starved, untreated or NF-κB inhibitor (Bay-0) pretreated HMVEC-d cells infected with KSHV (0 DNA copies/cell) were used to study the cytokine profile using a human protein cytokine-array kit from Ray Biotech (Norcross, GA). Uninfected HMVEC-d cells were used as control. The cytokine detection membranes were blocked with a blocking buffer for 1 h at room temperature then incubated with conditioned media at C overnight. The membranes were washed, incubated with 1ml of primary biotin-conjugated antibody at room temperature for h, subsequently washed, incubated with ml of horseradish peroxidase-conjugated streptavidin at room temperature for 0 min, developed by using enhanced chemiluminescence- type solution, Downloaded from on December 1, 01 by guest 1
13 exposed to film and processed by autoradiography. Signal intensities were quantitated using an Alpha Inotech Image analysis system. All the arrays were normalized to the same background levels with positive and negative substrate controls using the software Ray Bio Human Antibody Array.1 Analysis Tool. Downloaded from on December 1, 01 by guest 1
14 RESULTS KSHV induces the activation of NF-κB early during infection of HMVEC-d and HFF cells. In a normal resting cell, NF-κB is sequestered in the cytoplasm due to its association with a family of inhibitory proteins, called IκB. A variety of external stimuli like viral infections, growth factors and cytokines are known to phosphorylate IκB through the IKK complex leading to the activation of NF-κB. Treatment of HMVEC-d and HFF cells with 0ng/ml TNFα, a known stimulator of NF-κB pathway, for 0 min showed about -fold increase in the phosphorylation levels of p and IκBα (Fig. 1A, 1B and 1C lane ). When target cells were infected with KSHV ( DNA copies/cell), we observed rapid NF-κB activation as detected by NF-κB-p phosphorylation as early as 1 min p.i. of HMVEC- d cells (Fig. 1A, top panel lanes 1-) or at min p.i. of HFF cells (Fig. 1B, top panel lanes -). The NF-κB activation observed in both the cell types was sustained till min of our observation. When phospho-iκb antibodies were used to determine whether p activation was due to IκB phosphorylation, we observed the phosphorylation of IκBα in infected HFF cells as early as min p.i. (Fig. 1C, top panel lanes 1-). NF-κB-p phosphorylation observed at nearly the same time points suggested that KSHV infection results in IκBα phosphorylation which in turn could be responsible for p activation. Similar IκBα phosphorylation was seen in HMVEC-d cells (data not shown). Equal loading of total lysates between different treatments was confirmed by the detection of similar β-actin protein levels in all samples (Fig. 1A, 1B and 1C, lower panels). Infection did not affect the total p levels in both HMVEC-d (Fig. 1A, middle panel) and HFF cells (Fig. 1B, middle panel) or total IκB levels in HFF cells (Fig. 1C, middle panel). These results demonstrated that KSHV activates NF-κB early during infection of adherent HMVEC-d and HFF cells. Downloaded from on December 1, 01 by guest 1
15 Specificity of KSHV induced NF-κB activation in HMVEC-d and HFF cells. Bay-0 is an inhibitor of IκB phosphorylation and is known to inhibit NF-κB activation (). To determine whether abrogation of IκB phosphorylation could inhibit KSHV induced NF-κB activation, cells pretreated with varying concentrations of Bay-0 were infected with KSHV for 1 min and then analyzed for NF-κB activation. We observed a dose dependent inhibition of KSHV induced p activation by Bay-0 in infected HMVEC-d and HFF cells (Fig. 1D and E, top panel, lanes -). KSHV binds to the adherent target cell surface heparan sulphate via its envelope glycoproteins gb and gpk.1a (1, ). Blocking this interaction by heparin, an analogue of heparan sulphate, prevents KSHV binding to the target cells and infection (, ). To demonstrate if NF-κB activation was due to KSHV binding and entry into the target cell and not due to contaminating materials or LPS, cells were infected for 0 min with KSHV pre-incubated with heparin and lysates were analyzed for NF-κB-p phosphorylation. Heparin treatment blocked the KSHV induced NF-κB activation by about 1% and % in HMVEC-d and HFF cells respectively (Fig.1D and 1E, upper panel, lane ), indicating that NF-κB activation was indeed due to KSHV infection. We have previously shown that KSHV infection induces a rapid transient MEK1/ and ERK1/ phosphorylation in HMVEC-d and HFF cells (). When lysates from Bay-0 pretreated cells were tested with phospho ERK1/ antibodies, Bay-0 pretreatment had no effect on KSHV induced ERK1/ phosphorylation (Fig. 1F, upper panel, lanes -). In contrast, pretreatment of cells with µm U01, a MEK1/ specific inhibitor, resulted in about % inhibition of KSHV induced ERK1/ phosphorylation (Fig. 1F, upper panel, lane ). There was no change in the total ERK levels (Fig. 1F, middle panel, lanes 1-). Equal loading was Downloaded from on December 1, 01 by guest 1
16 confirmed using anti β-actin antibodies (Fig. 1F, bottom panel, lanes 1-). These results demonstrated the specificity of inhibition by Bay-0 pretreatment as well as specificity of KSHV induced NF-κB activity. KSHV triggers the rapid nuclear translocation of activated NF-κB-p. Once activated in a stimulus specific manner, NF-κB rapidly translocates into the nucleus and induces the transcription of various cellular genes (). Since KSHV induced the NF-κB early during infection, we examined the uninfected and infected cells by IFA using polyclonal antibody against NF-κB-p. Rapid nuclear translocation of p in > 0% of KSHV infected HMVEC-d (Fig., panels C and D) and HFF (Fig., panels I and J) cells was observed after 0 min and min p.i., respectively. In contrast NF-κB-p was predominantly localized in the cytoplasm of uninfected cells (Fig., panels A, B, G and H). Pretreatment of Bay-0 significantly inhibited nuclear translocation in both HMVEC-d (Fig., panels E and F) and HFF cells (Fig., panels K and L). These results confirmed the specificity of NF-κB induction and further supported our observation that KSHV induces NF-κB early during infection of target cells. When infected cells were examined at h p.i., >0% of cells displayed p nuclear translocation (Fig., panels A and B). Infection of HMVEC-d cells with MOI of GFP-KSHV revealed that about 0% of cells were infected under these conditions (Fig., panels C and D). Uninfected cells were negative for ORF staining and showed cytoplasmic p staining (Fig., panels E, F, G and H). In contrast, cells infected with KSHV displayed characteristic punctuate nuclear staining of ORF protein and greater than 0% of cells were positive for both ORF and nuclear p at h p.i. (Fig., panels I, J, K and L). Some of the uninfected cells also displayed p nuclear translocation, which could be due to the action of cytokines and Downloaded from on December 1, 01 by guest 1
17 growth factors released from the HMVEC-d cells as shown in Fig.. These results suggested that NF-κB stimulation also occurs at a later time p.i., a time point where only the expression of latent KSHV genes were observed (, 0). KSHV infection increases the NF-κB DNA binding activity in HMVEC-d and HFF cells. Activated NF-κB after translocating into the nucleus binds to κb site sequences in the various target genes and initiates transcription. To determine whether KSHV infection induces DNA binding activity of NF-κB, we analyzed the nuclear extracts prepared from infected untreated or Bay-0 pretreated cells by an ELISA-based DNA binding assay. Purity of the nuclear extracts was confirmed using anti-lamin B and anti-tubulin antibodies (data not shown) (). When infected cell values were normalized to the uninfected cell values, nuclear extracts from HMVEC-d and HFF cells infected for 0 min displayed enhanced NF-κB DNA binding activity (Fig. A). The specificity of increase in NF-κB DNA binding activity was demonstrated by inhibition using excess wild-type oligonucleotides, but not with excess mutant consensus site oligonucleotides (Fig. A). The specificity was also demonstrated by the dose-dependent decrease in NF-κB DNA binding activity in Bay-0 pretreated HMVEC-d and HFF cells (Fig. B). Time course analysis of DNA binding activity with µm Bay-0 pretreated cells showed an inhibition of about %, % and % at 1 min, 0 min and 0 min p.i., respectively, in HMVEC-d cells, and about %, % and % at 1 min, 0 min and 0 min p.i., respectively, in HFF cells (Fig. C). These results correlated well with the Western blot data. For all further studies µm and µm concentrations of Bay-0 were used and only the concentration showing optimal activity is presented in all the figures. Electrophoretic mobility shift assay (EMSA) detects the increase in NF-κB DNA binding activity in infected HMVEC-d and HFF cells. Downloaded from on December 1, 01 by guest 1
18 To confirm the above finding, EMSA was performed to demonstrate NF-κB protein binding to the NF-κB sites. Retardation in migration of radio labeled NF-κB consensus site oligonucleotides was observed with infected cell nuclear extracts (Fig. A and B, lanes -), which was reduced by µm Bay-0 pretreatment (Fig. A and B, lanes -). The specificity of this reaction was demonstrated by the absence of NF-κB binding to the target DNA in the competition assays using 0 times molar excess of cold double stranded κb oligonucleotide probe (Fig. A and B, lane ), while the binding was not affected with normal probe (Fig. A and B, lane ). Binding of Oct1 protein to its specific probe remained unchanged (Fig. A and B, lower panel, lanes 1-), which also demonstrated the specificity of NF-κB inhibition by Bay-0. These results demonstrated that KSHV infection activated NF- κb translocates to the nucleus and recognized the NF-κB specific sites thus suggesting possible transcription of NF-κB dependent genes. Early induction of NF-κB by KSHV indicated a role for virus binding and entry stages. To determine whether NF-κB induction requires a KSHV-induced signal cascade and/or viral gene expression, we examined the NF-κB levels in HMVEC-d cells infected with MOI of either live or UV-inactivated KSHV. Live-KSHV induced NF-κB to a greater extent than UV- KSHV, with about.1,, and.-fold increase in NF-κB activation with live-kshv (Fig. C), compared to.1,., and.-fold with UV-KSHV (Fig. D) at h, h and h p.i., respectively in HMVEC-d cells. Oct-1 levels remained unaltered with live-kshv and UV- KSHV infection at all time points. Although NF-κB induction with UV-KSHV was significantly higher than uninfected cells and sustained, this induction was comparatively lower than the induction observed with live-kshv at all parallel time points. This suggested that early Downloaded from on December 1, 01 by guest 1
19 induction of NF-κB by KSHV must be mediated by virus binding and entry stages, and KSHV viral gene expression appears to be required for the augmented continued induction of NF-κB. KSHV induces a sustained level of NF-κB induction during de novo infection of HMVEC-d and HFF cells. Early during infection of adherent target cells, KSHV induced the FAK, Src, PI-K, Rho- GTPases, PKC-ζ and ERK1/ signal pathways, which were characterized by a rapid early response followed by decline to the resting state within - h of observation (1,,, ). Since we observed NF-κB activation up to min p.i. (Fig. 1), to determine the extent of NF- κb induction and compare them to other KSHV induced signal cascades, infected HMVEC-d and HFF cells collected at different time point p.i. were analyzed for the phosphorylated forms of NF-κB-p, ERK1/, AKT and pmapk. We observed a modest level of NF-κB-p activation in the infected HMVEC-d cells at all time points tested (Fig. A). Phosphorylated p was detected as early as min p.i. and sustained activation of about >-fold increase over the uninfected cells was observed until h p.i. (Fig. A, lanes 1-1), as well as at h p.i. (data not shown). In contrast, there was a rapid increase in ERK1/ phosphorylation, peaking at 0 min p.i. with about 1.-fold activation over the uninfected cells, decreased by h p.i., followed by a second cycle of activation, which peaked at h p.i. and returning to the basal level at h p.i. (Fig. A). KSHV induces FAK, Src, and PI-K pathways and NF-κB could be activated through the PIK-AKT pathway (). When we examined the activation of AKT, a -fold increase in phospho-akt level was observed by min p.i., followed by a reduction till 0 min p.i., a second cycle of increase at 1 h (.- fold) which peaked to.-fold at h p.i. Although the p-akt levels decreased between - h p.i., it showed a higher level of activation over the uninfected Downloaded from on December 1, 01 by guest 1
20 cells. A third intense -fold activation was seen at h p.i. which decreased to -fold at h p.i. (Fig. A). Previously, we did not observe the induction of LPS/stress-activated p-mapk during 0 min of KSHV infection in HMVEC-d, HFF and cells (). A recent study reported that pmapk activation is essential for KSHV infection in human umbilical vein endothelial cells (). Furthermore, KSHV latency associated gene product Kaposin B (K1-B) has been shown to enhance the release of pro-inflammatory cytokines by activating the pmapk-mitogen- activated protein kinase (MAPK)-associated protein kinase (MK) pathway and blocking cytokine mrna decay by binding to MK (0). Cytokines that are secreted upon KSHV infection can act in an autocrine or paracrine fashion to activate pmapk. Activation of many of these cytokines are known to be controlled by NF-κB and pmapk-mk-kaposin B may play an important role in stabilizing the cytokine expression that is activated by NF-κB. Hence, we examined the kinetics of pmapk activation by KSHV. In concordance with our earlier results (), there was only minimal activation of pmapk at earlier time points (Fig. A, lanes -). However, an appreciable level of pmapk activation was observed at h p.i. with about.-fold activation over uninfected cells, peaking at h p.i. with.-fold induction and returning to basal level at h p.i. (Fig. A, lanes -1). Equal loading of samples was demonstrated by detection of similar levels of total p, AKT, p, ERK and β-actin throughout the observation period (Fig. A), which also clearly suggested that KSHV infection induces the phosphorylation of these proteins without affecting the total protein synthesis levels. KSHV infected HFF cells also displayed nearly similar p, ERK1/, AKT and pmapk activation kinetics as seen in HMVEC-d cells (Fig. B). There was modest steady state activation of NF-ΚB- p at all time points, which peaked at h p.i. (.- fold activation), Downloaded from on December 1, 01 by guest 0
21 which reached the basal level at h p.i. We observed about -fold ERK1/ phosphorylation as early as min p.i., peaking at min to.-fold, which returned to basal level between - h p.i. (Fig. B). A second cycle of moderate ERK1/ activation was seen at - h p.i. (Fig. B lanes -1). The induction kinetics of phospho AKT was similar to that of HMVEC-d cells but with cycles of activation, the first cycle starting at min p.i., peaking at h p.i., returning to basal level between -1 h p.i., with a second cycle of AKT activation seen at h p.i. (.- fold activation), which was sustained at a moderate level until h p.i. (Fig. B, lanes -1). Similar to HMVEC-d cells, minimal activation of pmapk at earlier time points was observed in HFF cells (Fig. B, lanes -), started to increase at h p.i., reached a maximum at 1 h p.i. and returned to the basal levels at h p.i. (Fig. B lanes -1). Taken together, these results demonstrated that KSHV infection induces a sustained level of NF-κB during the h period of observation which is in contrast to the biphasic ERK1/ and AKT activation, and activation of pmapkinase during the later time points. These results also suggested that during primary infection of adherent target cells, KSHV must have evolved to differentially induce these signal molecules. KSHV induced NF-κB does not play a role in entry of virus into the target cells. KSHV entry overlaps with the induction of host cell pre-existing signal pathways such as FAK, Src, PI-K, Rho-GTPases, PKC-ζ and ERK1/ (,, ). We have demonstrated the role of FAK and PI-K in virus entry, role of RhoA-GTPase in microtubule modulation, transport of capsid in the cytoplasm and nuclear delivery and a role for ERK1/ in viral gene expression (1,,, ). Target cells pre-incubated with the PI-K inhibitor LY00 inhibited the internalization of KSHV DNA (), whereas there was no significant reduction when cells were pretreated with the MEK/ERK inhibitor, U01 (). To determine whether Downloaded from on December 1, 01 by guest 1
22 KSHV induced NF-κB plays a role in viral infection, we examined virus DNA internalization by quantitative real-time DNA PCR for KSHV ORF gene. Treatment of cells with three different concentrations of Bay-0 did not have any effect on KSHV DNA entry in both HMVEC-d and HFF cells (Fig. A). In contrast, preincubation of cells with 0µM LY00 reduced internalization by about 0% and % in HMVEC-d and HFF cells, respectively (Fig. A). These results demonstrated that KSHV induced NF-κB does not have a significant role in viral DNA internalization. Inhibition of NF-κB blocks the expression of KSHV latent and lytic gene expression. To analyze the potential role of NF-κB activation in KSHV gene expression, HMVEC-d and HFF cells were pre-incubated with different concentrations of Bay-0 at C for 1 h, infected with KSHV for h, and viral messages, collected at different time points p.i., were quantitated by real-time RT PCR. We have previously shown that de novo KSHV infection of HMVEC-d and HFF cells is characterized by the sustained expression of latency-associated ORF (LANA-1), ORF and K1 genes, transient expression of a limited number of lytic genes including ORF 0 (RTA), virf, K and K genes (0, ). In this present study, we also observed a steady increase in ORF gene expression from as early as 1 min p.i. peaking at h p.i., while ORF 0 gene expression was fold higher at initial time points, peaked at h p.i. and gradually reduced to basal levels at h p.i. in HMVEC-d cells (Fig. B and C). As the gene expression for ORF 0 and ORF was high at h and h respectively, expression of other lytic genes like K, K and virf were studied and compared at h, h and h p.i. Likewise, in HFF cells there was a stable increase in ORF gene expression during the observed h time period (Fig. D). In contrast, very high levels of lytic gene expression were seen at earlier time points, which peaked between - h p.i. and slowly declined thereafter in Downloaded from on December 1, 01 by guest
23 HMVEC-d and HFF cells (Fig. C and D). As we have previously demonstrated (), no viral gene expression was seen when target cells were infected with UV-treated KSHV (Fig. E and F). In contrast, treatment of cells with µm Bay-0 for 1 h reduced the expression of both latent and lytic KSHV gene expression significantly (Fig. E and F). The expression of ORF gene in HMVEC-d cells was reduced by about %, % and % at h, h and h p.i., respectively (Fig. E). Similarly, expression of ORF gene in HFF cells was reduced by about %, % and 0% at h, h and h p.i., respectively (Fig. F). About 0% to % reduction in the lytic genes was observed in Bay-0 treated HMVEC-d cells (Fig. E), and % to % inhibition was seen in HFF cells (Fig. F). These results demonstrated that NF-κB induced by KSHV early during target cell infection play important roles in viral latent and lytic gene expression, thus contributing to KSHV infection and pathogenesis. KSHV induced NF-κB plays a major role in the activation of AP-1 family transcription factors. The role-played by NF-κB and AP-1 transcription factors independently in modulating KSHV latent and lytic gene expression in PEL cells is well documented (, ). However there are no reports on the effect of NF-κB inhibition on AP-1 transcription factors during de novo KSHV infection. Our studies suggested that NF-κB activation is required for transcription initiation of both latent and lytic genes in primary adherent target cells. To determine whether this is due to the ability of NF-κB to modulate a variety of host transcription factors, we next examined the ability of KSHV infection to induce AP-1 transcription factors which is known to be involved in KSHV latent and lytic gene expression (). Nuclear extracts from uninfected and infected HMVEC-d cells were assessed in an ELISA based assay for the ability of the AP-1 transcription factors to bind to their respective Downloaded from on December 1, 01 by guest
24 wild type DNA sequences. Since we observed NF-κB activation very early during infection, nuclear extracts from HMVEC-d cells infected with KSHV for 1 min, 0 min and 0 min were assayed for the AP-1 family of transcription factors. Infection of HMVEC-d cells with KSHV mediated a differential activation of AP-1 family transcription factors (Fig. ). As shown in Fig. A, compared to uninfected cells, KSHV infection increased the activated forms of both Fos and the Jun family of transcription factors. A higher level of activation was observed for p-c-jun, JunB, JunD and cfos, while FosB, Fra1 and Fra transcription factors were moderately activated (Fig. A). Specificity experiments carried out with wild type (wt) and mutated (mut) oligos demonstrated a significant reduction in the ability of transcription factors to bind their respective target sequences by pre-incubation with wt oligonucleotides (data not shown). To analyze the effect of NF-κB inhibitor Bay-0 in KSHV mediated induction of AP-1 transcription factors, HMVEC-d cells were pretreated with the drug, infected with KSHV for 1 min, 0 min and 0 min, and activities of various transcription factors in the nuclear extracts of infected cells were measured. Only the optimum time point values are represented in the graph (Fig. B and C). In concordance with previous results (Fig. A), neither KSHV infection nor inhibitors of NF-κB pathway had any effect on the activation of Fra1 and Fra, whereas there was a very moderate effect of Bay-0 on JunB with about < 0% inhibition (Fig. B). In contrast, Bay-0 displayed differential inhibitory effect on the activation of other AP-1 components (Fig. B). About 0%-0% FosB and JunD inhibition was observed. The highest inhibition of about >0-0% was observed for cfos with Bay-0. In contrast, p-c- Jun activation increased by about % and 0% with µm and 0µM Bay-0, respectively, over untreated cells infected with KSHV (Fig. C). Our previous studies have demonstrated that MEK1/ inhibitor U01 prevented the activation of p-c-jun by Downloaded from on December 1, 01 by guest
25 approximately 0% and cfos by % in HFF cells (). Similarly, U01 when used as a specificity control in this study, inhibited p-c-jun, cfos, FosB, JunB and JunD activities by about %, 0%, 1%, % and %, respectively, and did not have any effect on Fra1 and Fra (Fig. B and C). These results indicate that NF-κB has a differential impact on the activation of AP-1 family of transcription factors in KSHV infected adherent target cells. KSHV infection leads to NF-κB mediated up regulation of cytokines. KS lesion is an inflammatory angioproliferative lesion characterized by the presence of a variety of inflammatory cells, pro-inflammatory cytokines and angiogenic factors in the lesions (1). Cultured KS lesion spindle cells require cytokines for their survival and proliferation (1) suggesting that cytokines probably act in both an autocrine and paracrine fashion. In our oligonucleotide array analysis of KSHV infected HMVEC-d and HFF cells at h and h p.i., we observed the reprogramming of host transcriptional machinery regulating a variety of cellular processes including apoptosis, cell cycle regulation, signaling, inflammatory response, and angiogenesis (). Since NFκB is known to regulate the majority of these factors, we next analyzed the role of KSHV induced NFκB in the regulation of these factors. Conditioned media collected from KSHV infected HMVEC-d cells at various time points post-infection were used to study the cytokine profile. Compared to the uninfected HMVEC-d cells, KSHV infection induced an increase in secretion of the following category of factors: a) pro-inflammatory cytokines such as IL-, IL-, IL-, IL-, IL-1, GRO, GROα and IFNγ (Fig. A, Table 1); b) anti-inflammatory cytokines such as IL-, IL- and IL-1 (Table 1); c) growth factors such as PDGF, Leptin, TGFβ1, TGFβ, IGF-1, GM-CSF, G-CSF, M-CSF and EGF (Fig. B, Table 1); d) angiogenic factors like SDF-1, angiogenin and SCF (Fig. C, Table 1); e) chemokines such as MCP-, MCP-, TARC, CKβ -1, eotaxin, eotaxin-, GCP-, MIF, MIG, Downloaded from on December 1, 01 by guest
26 MDC, ENA- and BLC (Fig. D, Table 1); f) Tissue inhibitors of matrix metalloproteinases such as TIMP-1 and TIMP- (Table 1). Further analyzes of the activation kinetics of these factors revealed three distinct patterns of induction (Fig., Table 1). In pattern one, factors such as IL-, IL-1, IL-, IL-, IL-1, G- CSF, SDF-1, TARC, ENA- and leptin were induced at a significant level at h p.i., reached a maximum induction at h p.i. and lowered to the h level or basal level at h p.i. In pattern two, several of the factors like IL-, IL-, LIGHT, GRO, IL-, GM-CSF, EGF, TGFβ, angiogenin and eotaxin- were induced at a significant level only at h p.i. and continued to be induced even at h p.i. Cytokines such as IL-, IFNγ, GROα, TNFβ, PDGF-BB, TGFβ1, IGF- 1, M-CSF, MCP-, CKβ -1, eotaxin, GCP-, MIF, BLC, MCP-, MDC and MIG were secreted at all time points tested, which could probably be playing a potential role in the constitutive activation of NF-κB and KSHV biology. Many of the KSHV infection induced cytokines, growth factors and angiogenic factors were inhibited by µm Bay-0 pretreatment (Table 1). We observed a -fold and -fold reduction in IL- induction at h and h p.i., respectively. IL-, IL-, GROα and IFNγ showed greater than -fold reduction after Bay-0 pretreatment. Similarly, the observed remarkable increase in IGF-1, PDGF-BB, leptin, TGFβ-1, M-CSF, GM-CSF and G-CSF growth factors after KSHV infection was also reduced by more than -fold with Bay-0. Among the chemokines, MCPs, MIG, MDC, MIPα, TARC, CKβ -1, eotaxins, MIF, PARC, GCP- and BLC showed more than a -fold increase and most of these chemokines were significantly reduced by NF-κB inhibition. Appreciable changes were not detected in the growth factor binding protein and tissue inhibitors of matrix metalloproteinase induction by Bay-0 pretreatment, whereas anti-inflammatory cytokines like IL-, IL-, IL- and IL-1 showed Downloaded from on December 1, 01 by guest
27 more than -fold reduction with µm Bay-0 pretreatment, in comparison to the supernatant from untreated cells infected with KSHV. We also observed the up regulation of a variety of angiogenic factors such as angiogenin, SCF, SDF-1 and VEGF, and these were also inhibited by Bay-0 pretreatment. Since the genes encoding these wide ranges of cytokines secreted upon KSHV infection possesses NF-κB binding site in their promoter region, their inhibition clearly demonstrated the role of KSHV induced NF-κB in the regulation of these factors. Downloaded from on December 1, 01 by guest
28 DISCUSSION During infection of target cells leading to a productive lytic replicative cycle or to the establishment of latency in certain target cells, herpes viruses need to overcome several obstacles like apoptosis, host intrinsic, innate and adaptive immune responses and transcriptional restrictions. These obstacles need to be counteracted not only during the early time of infection but also during the entire time of latent infection. Establishment of latent infection during in vitro infection of primary human endothelial or fibroblast cells by KSHV provides an opportunity to analyze the various complex interactions between viral and host factors and the potential mechanism of establishment and maintenance of latent infection. Our previous studies have revealed that to over come the obstacles early during infection even before de novo viral gene transcription and expression, KSHV has adopted a best strategy of manipulating the host cells pre-existing signal pathways via the interactions with cell surface receptors (Fig. ). KSHV binds to the adherent target cell surface heparan sulfate molecule, to integrins, to the transporter CD/xCT complex, and possibly to other molecule(s). This is followed by virus entry overlapping with the induction of host cell pre-existing signal pathways such as FAK, Src, PI- K, Rho-GTPases, PKC-ζ and ERK1/. In this report, we provide multiple comprehensive evidences to suggest that in addition to the signal cascades, and in contrast to the differential induction of ERK1/ and pmapk molecules, KSHV infection also induces NF-κB very early during infection which is sustained throughout the period of observation. Our studies give a snapshot of the complex events occurring early during infection of adherent target cells (Fig. ). For clarity, we have summarized below these events and their potential implications on KSHV biology and pathogenesis. Downloaded from on December 1, 01 by guest
29 Role of NF-κB in KSHV gene expression during endothelial cell infection. Several inhibitors are shown to inhibit NF-κB activation at different levels such as the prevention of IκB phosphorylation by Bay-0, blocking IκB degradation by protease inhibitor like MG1, or by preventing the nuclear translocation of NF-κB by CAPE or SN0. We used Bay-0 and not the protease inhibitors as they might affect the Notch signaling pathway involved in KSHV pathogenesis (). KSHV induced NF-κB was blocked by Bay-0 and dose response studies indicate that both HMVEC-d and HFF cell have varying sensitivity to the inhibitor. Similar variation with Bay-0 pretreatment was observed between HEK cells and murine pre B cells upon TNFα treatment (, ). We have previously demonstrated that KSHV induced ERK1/ play roles in the regulation of ORF 0 and ORF gene expression, probably in the initiation of their expression. KSHV induced NF-κB also appears to influence viral gene expression, which could be due to direct interactions with the viral gene transcription initiation region or by indirect methods such as the activation of host transcription factors and/or host genes which in turn play roles in viral gene expression. Examination of ORF 0 and ORF genes promoter regions show that only the ORF 0 gene but not the ORF gene possess NF-κB binding sites in its promoter region (), and thus suggesting that NF-κB could directly influence the transcriptional activation of the ORF 0 gene. KSHV latency associated vflip has been shown to persistently activate NF-κB by interacting with the IKKα, IKKβ, IKKγ complex and this has been taken as evidence for NFκB s role in the maintenance of KSHV latency in the PEL cells (1). However, how NF-κB Downloaded from on December 1, 01 by guest 1 regulates the latent genes is not known. ORF 0 (RTA) is believed to contribute in the establishment of latency through activation of LANA-1 expression in the early stages of infection (). LANA-1 has been shown to physically interact with RBPJk, bind to the RTA
30 promoter and block the activation of RTA (). Thus there exists a feed back loop through which LANA-1 and RTA possibly regulates each other. Even though there is no NF-κB binding site in the ORF promoter, since the impact of blocking RTA could be manifested at various levels, the inhibition of ORF gene expression by NF-κB inhibition could also be due to the block in RTA expression by Bay-0 pretreatment. KSHV virf and K are expressed early during infection of HMVEC-d cells and Bay-0 pretreatment inhibited the expression of these genes. Since RTA (ORF 0) protein is known to control the transcription of both K and virf by binding to the RRE (RTA response element) present in the promoter region of these lytic genes, K and virf inhibition upon NF-κB blockade could be attributed directly to RTA inhibition. The regulation of lytic K gene is known to be independent of RTA () and was not influenced by ERK1/ early during infection (, ). Since the K gene also does not have an NF-κB binding site, inhibition of K gene observed after Bay-0 pretreatment could also be an indirect effect of various transcription factors under the control of NF-κB. Our results are in concordance with the studies by Keller et al () who did not observe any increase in lytic gene activation in PEL cells after Bay-0 treatment. KSHV induced NF-κB and AP-1 activation. Activation of any viral or cellular gene is not controlled by a single transcription factor but by interplay between various transcription factors, and one transcription factor could control the expression of other. It is interesting to note that LANA-1 and K genes possess several transcription factor binding motifs which include AP-1, SP1, cmyc and c-jun. Previous reports demonstrated that AP-1 activity may be critical for very early activation of the RTA and K promoters during lytic cycle (), and our studies have shown that ERK1/ via the activation of AP-1 and other MAPK related transcription factors Downloaded from on December 1, 01 by guest 0
31 plays important roles in the activation of LANA-1 and RTA genes (). Inhibition of ERK1/ using MEK inhibitor U01 blocked RTA, LANA-1, K and virf gene expression but had minimal effect on K () whereas Bay-0 pretreatment inhibited all of the genes. These studies demonstrate that KSHV gene expression is controlled by the regulation of multiple transcription factors, and inhibiting ERK1/ probably inhibited only the factors downstream of ERK1/. In contrast, Bay-0 pretreatment leads to the inhibition of both NF-κB and the AP-1 family of transcription factors resulting in the blockade of all the viral genes tested. By inducing NF-κB and subsequent transcription factors like AP-1, KSHV must be overcoming the transcription block in quiescent cells. A rapid NF-κB activation is evident in KSHV infected target cells. At nearly the same time period there is activation of the AP-1 family of transcription factors accompanied by KSHV lytic gene expression. NF-κB could be influencing AP-1 transcription factors, which has κb binding sites in their promoter. We observed an appreciable increase in cfos, FosB, c-jun, JunB and JunD transcription factors after KSHV infection and this activation was inhibited significantly by NF-κB inhibition (Fig. B). Down stream phosphorylation of AP-1 transcription factors were probably inhibited by NF-κB inhibition and thus clearly demonstrating that the regulation of AP-1 transcription factors could also be due to NF-κB. The effect of NF-κB inhibition on AP-1 transcription factors could be at multiple levels a) it might affect the gene transcription, as the AP-1 family of transcription factors are known to have a κb binding site in their promoter, b) it might affect the phosphorylation of transcription factors and c) DNA binding activity of transcription factors could also be inhibited. Further studies are needed to identify the mechanism of AP-1 down regulation by NF-κB inhibition. Downloaded from on December 1, 01 by guest 1
32 Inhibition of KSHV induced NF-κB and c-jun activation. One of our surprising observations is that in cells pretreated with Bay-0 before KSHV infection, c-jun was further up regulated, and there was a dose dependent increase in c-jun DNA binding activity. Generally AP-1 complexes function as positive regulators of cell proliferation by regulating the expression of essential cell cycle proteins such as D1, p, p1, p1 and p1 and differential effects are also present among different members (0). Constitutive activation of c-jun is known to induce apoptosis (). In c-jun knockout cells, there was an up regulation of NF-κB, and when c-jun was ectopically expressed, NF-κB levels were reduced (). The absence of NF-κB mediated inhibition of JNK activation was known to contribute to TNFα induced apoptosis (). These evidences clearly suggest that there exists a negative regulation between c-jun and NF-κB. It is reasonable to speculate that the increase in c-jun activation upon NF-κB inhibition could lead to an increase in latent and lytic gene activation. However, except c-jun, all other AP-1 family of transcription factors was inhibited with NF-κB inhibition. Hence, it is evident that viral gene regulation is not controlled by a single transcription factor instead by the involvement of a variety of transcription factors. It is likely that KSHV activates c-jun for the production of cytokines and lytic gene expression. Since hyper activation of c-jun may be deleterious for the cell as it is pro-apoptotic, KSHV probably activates NF-κB to regulate c-jun phosphorylation so as to allow the infected cells to survive. Implications for the sustained NF-κB induction by KSHV. The sustained induction of NF-κB during infection of endothelial cells in vivo is probably required to overcome apoptosis, host intrinsic, innate and adaptive immune responses and transcriptional restriction not only during the early stages of infection but also during the establishment and subsequent maintenance of latency. Activated NF-κB is known to be critical to escape immune system surveillance and to Downloaded from on December 1, 01 by guest
33 block apoptosis by directly binding to p and thus preventing p s transcriptional activity or through competition for transcriptional co-activators like CBP/p00 () or by activating various anti-apoptotic molecules or by inducing the secretion of various cytokines. KSHV infection induced sustained NF-κB activation probably performs all these functions. Activation of transcription factors requires the phosphorylation of multiple upstream signaling molecules. Our previous studies have shown that KSHV induces multiple signaling pathways, which probably aids in the successful establishment of infection and to evade surveillance by the immune system (1,, ). The initial binding event appears to be enough for the activation of NF-κB, and this early phase of activation (Fig. ) could be due to both virus internalization and the expression of viral lytic genes, as many of the transiently expressed early viral lytic genes have roles in inducing NF-κB. During this phase, NF-κB probably induces proteins necessary for its sustained activation, and the later time point of activation could be due to the effect of anti-apoptotic molecules and the secretion of cytokines, as many of these NF-κB regulated cytokines are known to activate NF-κB (Fig. ). NF-κB activation possibly leads to the induction of various anti-apoptotic molecules, and NF-κB is in turn probably activated by the anti-apoptotic KSHV latency associated gene like vflip shown to be responsible for the constitutive activation of NF-κB in PEL cells (1). In addition to up regulating anti-apoptotic proteins, NF-κB is known to induce signaling factors involved in NF-κB activation pathway, by doing so, NF-κB probably assures the constitutive expression of proteins necessary for its persistent activation in KSHV infected endothelial cells. Alternatively, Notch could also be responsible for the persistent activation of NF-κB, as Notch is known to augment NF-κB activity by retaining NF-κB in the nucleus (1). Activation of the Notch signal pathway by KSHV is known to be involved in the regulation of lytic gene Downloaded from on December 1, 01 by guest
34 expression (). RBPJκ was shown to bind with RTA and recruit it to its cognate recognition site and thus relieving the RBPJκ mediated repression and up regulation of target gene expression. The upstream events leading to the activation of NF-κB, viral envelope glycoprotein and the interacting receptor(s) involved in the early induction are not known at present and are under study. Activation of NF-κB by UV-KSHV demonstrated that virus binding and entry is enough to induce the activation of NF-κB at the early phase, and activation during the late phase could be due to a combination of vflip action and by the variety of cytokines and growth factors secreted from the infected cells. A recent report by Caselli et al. () showed that UV-virus could not activate NF-κB. This discrepancy may be due to technical reasons such as the difference in virus titers. UV treatment of KSHV results in a reduction in viral copy numbers (presumably due to virus adhering to the plastic). Hence it is necessary to DNase treat the virus and estimate the copy numbers after UV treatment, and utilize similar copy numbers as that of live KSHV. ERK1/, pmapk and AKT induction by KSHV. ERK1/ phosphorylation was critical for the initiation of KSHV latent and lytic gene expression (). Our long-term activation study demonstrates biphasic activation kinetics of ERK1/. The high level of early phase ERK1/ activation coincided with NF-κB activation, which could be attributed to virus binding and entry. The late phase of ERK1/ activation was seen at h p.i., which coincided with LANA-1 expression indicating that the second phase could be due to the establishment of latency in these cells as blocking ERK was known to inhibit LANA-1 expression (). LANA-1 up regulates vil- expression by inducing AP-1 transcription factors, which is known to be activated by the MAPK pathway (, ). Biphasic MAPK activation is also seen in other viruses, with the second phase coinciding with viral genome synthesis (,,, 1). We have previously demonstrated Downloaded from on December 1, 01 by guest
35 that AP-1 and MAPK family of transcription factors activation via ERK1/ is critical for the latent and lytic gene expression (). Hence it is possible that the c-jun phosphorylation we observed after NF-κB inhibition could also be due to ERK phosphorylation, which remains unaffected with Bay-0 pretreatment, whereas the requirement of multiple transcription factors for viral gene expression cannot be ruled out. Besides the sustained NF-κB activation and biphasic ERK1/ activation, we observed a late time point activation of pmapk, which was in concordance with the result observed in our previous study (). Kaposin B is known to induce p/mk pathway and stabilize cytokine mrnas (0). Unlike ERK1/ and NF-κB, pmapk is not activated by KSHV binding to target cells; instead the activation was seen only after h p.i. The pmapk pathway is normally activated by stress, growth factors and cytokines resulting in proliferation, differentiation, development and inflammation. Hence KSHV induced activation of NF-κB early during infection is probably necessary for the cytokine release and it is probably sustained by the activation of pmapk during the later time period of latent infection (Fig. ). Combined activation of both the MAPK pathway and NF-κB pathway has been shown to be necessary for COX- induction and prostacyclin release in endothelial cells () and we have observed a rapid sustained induction of COX- in KSHV infected endothelial cells (). Taken together, this suggests that the signal pathways may co-operate and induce the secretion of the cytokines, chemokines and growth factors (Fig ). COX- expression is known to be mediated through AKT via the NF-κB pathway (). AKT is a survival signal molecule that is activated during many viral infections (, 0, ). We observed a triphasic activation of AKT in both target cells. The initial activation could have been due to virus binding and entry and the second phase could be due to viral gene expression. Downloaded from on December 1, 01 by guest
36 KSHV interaction with target cells induced PI-K during the early stages of infection (). AKT is the immediate downstream signaling molecule of PI-K; hence virus binding to integrin could have initiated the AKT phosphorylation. Induction of lytic genes could have contributed to the second phase of activation. The variety of growth factors and cytokines induced during the later time points could act both in paracrine and autocrine fashions to stimulate the third phase of PIK/AKT pathway activation. In summary, these results suggest that in adherent target cells, KSHV induces the differential activation of various signaling molecules but sustains the activation of NF-κB to modulate various transcription factors responsible for latent and lytic gene regulation. Implications of KSHV induced NF-κB and induction of cytokines. The role played by NF-κB in regulating cytokines like IL-, IL-, GROα, IL-1β, IFN-γ, and VEGF in PEL cells is well documented (,,, ). Our studies clearly demonstrated that KSHV infection of primary endothelial cells leads to the increased secretion of human cytokines, chemokines and growth factors via the activation of NF-κB. Among the strongest up regulated host molecules on the array were cytokines and chemokines like GROα, IL-, IL-, IL-, IL-, IL-, IFN- γ, GM-CSF, PDGF, IGF-1, eotaxin, MCPs, MIF and angiogenin. Among these GRO, IL-, IL-, IL-, IFN-γ, GM-CSF, PDGF, IGF and MCPs are well established target genes of NF-κB (). Except a few cytokines, growth factors, chemokines and angiogenic factors that were modulated by KSHV infection at all three time points of our observation, there were many cytokines that were released only at one or two time points, thus suggesting KSHV may be selectively regulating these factors for its advantage. Further studies are essential to define the variations in KSHV induced cytokines. Downloaded from on December 1, 01 by guest
37 Several lines of evidence demonstrate that KSHV is etiologically associated in KS pathogenesis (1). Expression of limited KSHV latent proteins such as LANA-1, vflip, vcyclind, Kaposins and the lytic protein K are detected in the KS lesion endothelial cells and lytic cycle proteins are detected in the limited percentage of KS lesion associated inflammatory cells (0). KS tumorigenesis appears to require an ongoing lytic infection since interruption of lytic replication by drugs such as ganciclovir appears to prevent KS development (). KSHV latent gene products such as vflip acting on NF-κB in latently infected endothelial cells and lytic infection in inflammatory cells expressing vgpcr, vil-, vmips, etc could collectively contribute to the initiation and maintenance of the KS lesion associated inflammatory microenvironment. Our observation of a robust induction of cytokines, growth factors and angiogenic factors by KSHV at h, h and h p.i. of endothelial cells () together with our demonstration of sustained NF-κB activation, a key inflammatory induction molecule, suggest that primary infection of endothelial cells could also create the microenvironment observed in the KS lesions. The persistent NF-κB activation by KSHV could be mediated by a combination of viral latent genes like vflip expression in the endothelial cells and by the cytokines and growth factors secreted in the infected cells supernatant (0). The model that emerges from our current and previous studies is that, primary infection of endothelial cells by KSHV initiates the host cell cytokine and growth factor cascades which are probably subsequently maintained by the interplay between viral and host genes, and KSHV utilizes the cytokines and growth factors for its own advantage such as for the maintenance of latent infection and immune evasion (Fig. ). The variety of cytokines and growth factors seen during KSHV primary infection of endothelial cells in our studies are strikingly similar to the cytokines and growth factors detected in the KS lesions. Although KSHV codes for several Downloaded from on December 1, 01 by guest
38 cytokines and chemokines that are known to activate NF-κB, none of them are shown to be expressed in the latently infected KS lesion endothelial cells (). It is possible that NF-κB, COX-, PGE and other cytokines induced during in vivo infection of endothelial cells could be responsible for the growth factors and cytokines seen in the microenvironment of KS lesions. A recent study by Grossmann et al. (1) showed the activation of NF-κB by vflip is required for the spindle shape of virus infected endothelial cells, which contributes to their cytokine release. Activation of several cytokines and growth factors in our study could be attributed to multiple viral proteins apart from vflip. Establishment of latency by KSHV is a very complex process and no single viral or host gene, transcription factor, signal molecule or cytokine activation could independently be responsible for this. Instead, it is probably mediated by a combination of all these factors selected over the time of evolution of KSHV along with the host. Hence, the outcome of in vitro KSHV infection of HMVEC-d cells and by analogy the in vivo infection of endothelial cells probably represents a complex interplay between host cell signal molecules, cytokines, growth factors, transcription factors and viral latent gene products resulting in an equilibrium state in which virus maintains its latency, blocks apoptosis, blocks host cell intrinsic and innate responses, and escapes from the host adaptive immune responses (Fig. ). KSHV probably utilizes NF-κB, COX- and other host cell factors including the inflammatory factors for its advantage such as the establishment of latent infection and immune-modulation. However, the combination of factors such as the absence of immune regulation, unchecked KSHV lytic cycle and increased virus load resulting in widespread KSHV infection of endothelial cells leading to induction of inflammatory cytokines and growth factors, and inability of the host to modulate this inflammation may be contributing to KSHV induced KS lesions. Thus, it is possible that Downloaded from on December 1, 01 by guest
39 effective inhibition of inflammatory responses including NF-κB, COX- and PGE, could lead to reduced latent KSHV infection of endothelial cells, which may in turn lead to a reduction in the accompanying inflammation and KS lesions. ACKNOWLEDGMENTS This study was supported in part by Public Health Service Grant CA 0 and Rosalind Franklin University of Medicine and Science - H.M. Bligh Cancer Research Fund to BC. We thank Keith Philibert for critically reading this manuscript. Downloaded from on December 1, 01 by guest
40 References: 1. Akula, S. M., N. P. Pramod, F. Z. Wang, and B. Chandran Human herpesvirus envelope-associated glycoprotein B interacts with heparan sulfate-like moieties. Virology :-.. Akula, S. M., F. Z. Wang, J. Vieira, and B. Chandran Human herpesvirus interaction with target cells involves heparan sulfate. Virology :-.. An, J., A. K. Lichtenstein, G. Brent, and M. B. Rettig. 00. The Kaposi sarcomaassociated herpesvirus (KSHV) induces cellular interleukin expression: role of the KSHV latency-associated nuclear antigen and the AP1 response element. Blood :-.. An, J., Y. Sun, R. Sun, and M. B. Rettig. 00. Kaposi's sarcoma-associated herpesvirus encoded vflip induces cellular IL- expression: the role of the NF-kappaB and JNK/AP1 pathways. Oncogene :1-.. Baeuerle, P. A., and D. Baltimore. 1. NF-kappa B: ten years after. Cell :1-0.. Baldwin, A. S., Jr. 1. The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu. Rev. Immunol. 1:-.. Bechtel, J. T., R. C. Winant, and D. Ganem. 00. Host and viral proteins in the virion of Kaposi's sarcoma-associated herpesvirus. J. Virol. :-.. Cahir-McFarland, E. D., K. Carter, A. Rosenwald, J. M. Giltnane, S. E. Henrickson, L. M. Staudt, and E. Kieff. 00. Role of NF-kappa B in cell survival and transcription of latent membrane protein 1-expressing or Epstein-Barr virus latency III-infected cells. J. Virol. :-.. Caselli, E., S. Fiorentini, C. Amici, D. Di Luca, A. Caruso, and M. G. Santoro. 00. Human herpesvirus acute infection of endothelial cells induces monocyte chemoattractant protein 1-dependent capillary-like structure formation: role of the IKK/NF-{kappa}B pathway. Blood doi:./ blood Casper, C., W. G. Nichols, M. L. Huang, L. Corey, and A. Wald. 00. Remission of HHV- and HIV-associated multicentric Castleman disease with ganciclovir treatment. Blood :1-1.. Cesarman, E., Y. Chang, P. S. Moore, J. W. Said, and D. M. Knowles. 1. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N. Engl. J. Med. : Chang, Y., E. Cesarman, M. S. Pessin, F. Lee, J. Culpepper, D. M. Knowles, and P. S. Moore. 1. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science : Chaudhary, P. M., A. Jasmin, M. T. Eby, and L. Hood. 1. Modulation of the NFkappa B pathway by virally encoded death effector domains-containing proteins. Oncogene 1:-. 1. Coscoy, L., and D. Ganem A viral protein that selectively downregulates ICAM- 1 and B- and modulates T cell costimulation. J. Clin. Invest. : Dourmishev, L. A., A. L. Dourmishev, D. Palmeri, R. A. Schwartz, and D. M. Lukac. 00. Molecular genetics of Kaposi's sarcoma-associated herpesvirus (human herpesvirus-) epidemiology and pathogenesis. Microbiol. Mol. Biol. Rev. : Ensoli, B., and R. C. Gallo. 1. AIDS-associated Kaposi's sarcoma: a new perspective of its pathogenesis and treatment. Proc. Assoc. Am. Physicians. :-1. Downloaded from on December 1, 01 by guest 0
41 Ganem, D. 1. KSHV and Kaposi's sarcoma: the end of the beginning? Cell 1: Grossmann, C., S. Podgrabinska, M. Skobe, and D. Ganem. 00. Activation of NFkappaB by the latent vflip gene of Kaposi's sarcoma-associated herpesvirus is required for the spindle shape of virus-infected endothelial cells and contributes to their proinflammatory phenotype. J. Virol. 0: Hammarskjold, M. L., and M. C. Simurda. 1. Epstein-Barr virus latent membrane protein transactivates the human immunodeficiency virus type 1 long terminal repeat through induction of NF-kappa B activity. J. Virol. : Haque, M., K. Ueda, K. Nakano, Y. Hirata, C. Parravicini, M. Corbellino, and K. Yamanishi Major histocompatibility complex class I molecules are downregulated at the cell surface by the K protein encoded by Kaposi's sarcoma-associated herpesvirus/human herpesvirus-. J. Gen. Virol. : Hayden, M. S., and S. Ghosh. 00. Signaling to NF-kappaB. Genes Dev. 1:1-.. Huang, T. T., S. L. Feinberg, S. Suryanarayanan, and S. Miyamoto. 00. The zinc finger domain of NEMO is selectively required for NF-kappa B activation by UV radiation and topoisomerase inhibitors. Mol. Cell. Biol. :1-.. Hung, J. H., I. J. Su, H. Y. Lei, H. C. Wang, W. C. Lin, W. T. Chang, W. Huang, W. C. Chang, Y. S. Chang, C. C. Chen, and M. D. Lai. 00. Endoplasmic reticulum stress stimulates the expression of cyclooxygenase- through activation of NF-kappaB and pp mitogen-activated protein kinase. J. Biol. Chem. :-.. Ikeda, A., X. Sun, Y. Li, Y. Zhang, R. Eckner, T. S. Doi, T. Takahashi, Y. Obata, K. Yoshioka, and K. Yamamoto p00/cbp-dependent and -independent transcriptional interference between NF-kappaB RelA and p. Biochem. Biophys. Res. Commun. :-.. Jeong, S. J., C. A. Pise-Masison, M. F. Radonovich, H. U. Park, and J. N. Brady. 00. Activated AKT regulates NF-kappaB activation, p inhibition and cell survival in HTLV-1-transformed cells. Oncogene :1-.. Jiang, S. J., T. M. Lin, G. Y. Shi, H. L. Eng, H. Y. Chen, and H. L. Wu. 00. Inhibition of bovine herpesvirus- replication by arsenite through downregulation of the extracellular signal-regulated kinase signaling pathway. J. Biomed. Sci. :00-.. Keller, S. A., D. Hernandez-Hopkins, J. Vider, V. Ponomarev, E. Hyjek, E. J. Schattner, and E. Cesarman. 00. NF-kappaB is essential for the progression of KSHV- and EBV-infected lymphomas in vivo. Blood :-0.. Keller, S. A., E. J. Schattner, and E. Cesarman Inhibition of NF-kappaB induces apoptosis of KSHV-infected primary effusion lymphoma cells. Blood :-.. Kim, S. M., E. J. Kim, S. I. Park, and J. H. Nam. 00. The role of ERK1/ activation in the infection of HeLa cells with Human coxsackievirus B. Acta Virol. : Krishnan, H. H., P. P. Naranatt, M. S. Smith, L. Zeng, C. Bloomer, and B. Chandran. 00. Concurrent expression of latent and a limited number of lytic genes with immune modulation and antiapoptotic function by Kaposi's sarcoma-associated herpesvirus early during infection of primary endothelial and fibroblast cells and subsequent decline of lytic gene expression. J. Virol. :01-0. Downloaded from on December 1, 01 by guest 1
42 Krishnan, H. H., N. Sharma-Walia, D. N. Streblow, P. P. Naranatt, and B. Chandran. 00. Focal adhesion kinase is critical for entry of Kaposi's sarcomaassociated herpesvirus into target cells. J. Virol. 0:-1.. Laherty, C. D., H. M. Hu, A. W. Opipari, F. Wang, and V. M. Dixit. 1. The Epstein-Barr virus LMP1 gene product induces A0 zinc finger protein expression by activating nuclear factor kappa B. J. Biol. Chem. :1-.. Lan, K., T. Choudhuri, M. Murakami, D. A. Kuppers, and E. S. Robertson. 00. Intracellular activated Notch1 is critical for proliferation of Kaposi's sarcoma-associated herpesvirus-associated B-lymphoma cell lines in vitro. J. Virol. 0:-1.. Lan, K., D. A. Kuppers, and E. S. Robertson. 00. Kaposi's sarcoma-associated herpesvirus reactivation is regulated by interaction of latency-associated nuclear antigen with recombination signal sequence-binding protein Jkappa, the major downstream effector of the Notch signaling pathway. J. Virol. :-.. Lan, K., D. A. Kuppers, S. C. Verma, and E. S. Robertson. 00. Kaposi's sarcomaassociated herpesvirus-encoded latency-associated nuclear antigen inhibits lytic replication by targeting Rta: a potential mechanism for virus-mediated control of latency. J. Virol. :-.. Lan, K., D. A. Kuppers, S. C. Verma, N. Sharma, M. Murakami, and E. S. Robertson. 00. Induction of Kaposi's sarcoma-associated herpesvirus latencyassociated nuclear antigen by the lytic transactivator RTA: a novel mechanism for establishment of latency. J. Virol. :-.. Liang, Y., and D. Ganem. 00. Lytic but not latent infection by Kaposi's sarcomaassociated herpesvirus requires host CSL protein, the mediator of Notch signaling. Proc. Natl. Acad. Sci. U. S. A. 0:0-.. Luo, H., B. Yanagawa, J. Zhang, Z. Luo, M. Zhang, M. Esfandiarei, C. Carthy, J. E. Wilson, D. Yang, and B. M. McManus. 00. Coxsackievirus B replication is reduced by inhibition of the extracellular signal-regulated kinase (ERK) signaling pathway. J. Virol. :-.. Matta, H., R. M. Surabhi, J. Zhao, V. Punj, Q. Sun, S. Schamus, L. Mazzacurati, and P. M. Chaudhary. 00. Induction of spindle cell morphology in human vascular endothelial cells by human herpesvirus -encoded viral FLICE inhibitory protein K1. Oncogene doi:./sj.onc McCormick, C., and D. Ganem. 00. The kaposin B protein of KSHV activates the p/mk pathway and stabilizes cytokine mrnas. Science 0: Miles, S. A., A. R. Rezai, J. F. Salazar-Gonzalez, M. Vander Meyden, R. H. Stevens, D. M. Logan, R. T. Mitsuyasu, T. Taga, T. Hirano, T. Kishimoto, and et al. 10. AIDS Kaposi sarcoma-derived cells produce and respond to interleukin. Proc. Natl. Acad. Sci. U. S. A. :0-0.. Miyamoto, S., P. J. Chiao, and I. M. Verma. 1. Enhanced I kappa B alpha degradation is responsible for constitutive NF-kappa B activity in mature murine B-cell lines. Mol. Cell. Biol. 1:-.. Mocarski, E. S., Jr. 1. Propagating Kaposi's sarcoma-associated herpesvirus. N. Engl. J. Med. :1-1.. Naranatt, P. P., S. M. Akula, C. A. Zien, H. H. Krishnan, and B. Chandran. 00. Kaposi's sarcoma-associated herpesvirus induces the phosphatidylinositol -kinase-pkc- Downloaded from on December 1, 01 by guest
43 zeta-mek-erk signaling pathway in target cells early during infection: implications for infectivity. J. Virol. :1-1.. Naranatt, P. P., H. H. Krishnan, M. S. Smith, and B. Chandran. 00. Kaposi's sarcoma-associated herpesvirus modulates microtubule dynamics via RhoA-GTPdiaphanous signaling and utilizes the dynein motors to deliver its DNA to the nucleus. J. Virol. :1-.. Naranatt, P. P., H. H. Krishnan, S. R. Svojanovsky, C. Bloomer, S. Mathur, and B. Chandran. 00. Host gene induction and transcriptional reprogramming in Kaposi's sarcoma-associated herpesvirus (KSHV/HHV-)-infected endothelial, fibroblast, and B cells: insights into modulation events early during infection. Cancer Res. :-.. Okuno, T., Y. B. Jiang, K. Ueda, K. Nishimura, T. Tamura, and K. Yamanishi. 00. Activation of human herpesvirus open reading frame K independent of ORF0 expression. Virus Res. 0:-.. Pahl, H. L. 1. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 1:-.. Pan, H., J. Xie, F. Ye, and S. J. Gao. 00. Modulation of Kaposi's sarcoma-associated herpesvirus infection and replication by MEK/ERK, JNK, and p multiple mitogenactivated protein kinase pathways during primary infection. J. Virol. 0: Pati, S., M. Cavrois, H. G. Guo, J. S. Foulke, Jr., J. Kim, R. A. Feldman, and M. Reitz Activation of NF-kappaB by the human herpesvirus chemokine receptor ORF: evidence for a paracrine model of Kaposi's sarcoma pathogenesis. J. Virol. : Pleschka, S., T. Wolff, C. Ehrhardt, G. Hobom, O. Planz, U. R. Rapp, and S. Ludwig Influenza virus propagation is impaired by inhibition of the Raf/MEK/ERK signalling cascade. Nat. Cell Biol. : Polson, A. G., D. Wang, J. DeRisi, and D. Ganem. 00. Modulation of host gene expression by the constitutively active G protein-coupled receptor of Kaposi's sarcomaassociated herpesvirus. Cancer Res. :-0.. Sanchez-Perez, I., S. A. Benitah, M. Martinez-Gomariz, J. C. Lacal, and R. Perona. 00. Cell stress and MEKK1-mediated c-jun activation modulate NFkappaB activity and cell viability. Mol. Biol. Cell 1:-.. Sanchez-Perez, I., and R. Perona. 1. Lack of c-jun activity increases survival to cisplatin. FEBS Lett. :-1.. Schulz, T. F. 1. Kaposi's sarcoma-associated herpesvirus (human herpesvirus-). J. Gen. Virol. : Schulz, T. F., J. Sheldon, and J. Greensill. 00. Kaposi's sarcoma associated herpesvirus (KSHV) or human herpesvirus (HHV). Virus Res. :-1.. Sharma-Walia, N., H. H. Krishnan, P. P. Naranatt, L. Zeng, M. S. Smith, and B. Chandran. 00. ERK1/ and MEK1/ induced by Kaposi's sarcoma-associated herpesvirus (human herpesvirus ) early during infection of target cells are essential for expression of viral genes and for establishment of infection. J. Virol. :0-.. Sharma-Walia, N., P. P. Naranatt, H. H. Krishnan, L. Zeng, and B. Chandran. 00. Kaposi's sarcoma-associated herpesvirus/human herpesvirus envelope glycoprotein gb induces the integrin-dependent focal adhesion kinase-src-phosphatidylinositol -kinaserho GTPase signal pathways and cytoskeletal rearrangements. J. Virol. :0-. Downloaded from on December 1, 01 by guest
44 Sharma-Walia, N., H. Raghu, S. Sadagopan, R. Sivakumar, M. V. Veettil, P. P. Naranatt, M. M. Smith, and B. Chandran. 00. Cyclooxygenase induced by Kaposi's sarcoma-associated herpesvirus early during in vitro infection of target cells plays a role in the maintenance of latent viral gene expression. J. Virol. 0:-. 0. Shaulian, E., and M. Karin. 00. AP-1 as a regulator of cell life and death. Nat. Cell Biol. :E Shin, H. M., L. M. Minter, O. H. Cho, S. Gottipati, A. H. Fauq, T. E. Golde, G. E. Sonenshein, and B. A. Osborne. 00. Notch1 augments NF-kappaB activity by facilitating its nuclear retention. EMBO J. :1-1.. St-Germain, M. E., V. Gagnon, S. Parent, and E. Asselin. 00. Regulation of COX- protein expression by Akt in endometrial cancer cells is mediated through NFkappaB/IkappaB pathway. Mol. Cancer. :.. Staskus, K. A., W. Zhong, K. Gebhard, B. Herndier, H. Wang, R. Renne, J. Beneke, J. Pudney, D. J. Anderson, D. Ganem, and A. T. Haase. 1. Kaposi's sarcomaassociated herpesvirus gene expression in endothelial (spindle) tumor cells. J. Virol. 1:1-1.. Sun, Q., H. Matta, G. Lu, and P. M. Chaudhary. 00. Induction of IL- expression by human herpesvirus encoded vflip K1 via NF-kappaB activation. Oncogene :1-.. Sun, R., S. F. Lin, K. Staskus, L. Gradoville, E. Grogan, A. Haase, and G. Miller. 1. Kinetics of Kaposi's sarcoma-associated herpesvirus gene expression. J. Virol. :-.. Syeda, F., J. Grosjean, R. A. Houliston, R. J. Keogh, T. D. Carter, E. Paleolog, and C. P. Wheeler-Jones. 00. Cyclooxygenase- induction and prostacyclin release by protease-activated receptors in endothelial cells require cooperation between mitogenactivated protein kinase and NF-kappaB pathways. J. Biol. Chem. 1:-1.. Tang, F., G. Tang, J. Xiang, Q. Dai, M. R. Rosner, and A. Lin. 00. The absence of NF-kappaB-mediated inhibition of c-jun N-terminal kinase activation contributes to tumor necrosis factor alpha-induced apoptosis. Mol. Cell Biol. :1-.. Thomas, K. W., M. M. Monick, J. M. Staber, T. Yarovinsky, A. B. Carter, and G. W. Hunninghake. 00. Respiratory syncytial virus inhibits apoptosis and induces NFkappa B activity through a phosphatidylinositol -kinase-dependent pathway. J. Biol. Chem. :-01.. Thornburg, N. J., W. Kulwichit, R. H. Edwards, K. H. Shair, K. M. Bendt, and N. Raab-Traub. 00. LMP1 signaling and activation of NF-kappaB in LMP1 transgenic mice. Oncogene :-. 0. Tomescu, C., W. K. Law, and D. H. Kedes. 00. Surface downregulation of major histocompatibility complex class I, PE-CAM, and ICAM-1 following de novo infection of endothelial cells with Kaposi's sarcoma-associated herpesvirus. J. Virol. :-. 1. Vieira, J., P. O'Hearn, L. Kimball, B. Chandran, and L. Corey Activation of Kaposi's sarcoma-associated herpesvirus (human herpesvirus ) lytic replication by human cytomegalovirus. J. Virol. :1-1.. Wang, F. Z., S. M. Akula, N. P. Pramod, L. Zeng, and B. Chandran Human herpesvirus envelope glycoprotein K.1A interaction with the target cells involves heparan sulfate. J. Virol. :1-. Downloaded from on December 1, 01 by guest
45 Wang, F. Z., S. M. Akula, N. Sharma-Walia, L. Zeng, and B. Chandran. 00. Human herpesvirus envelope glycoprotein B mediates cell adhesion via its RGD sequence. J. Virol. :-1.. Wang, H. W., M. W. Trotter, D. Lagos, D. Bourboulia, S. Henderson, T. Makinen, S. Elliman, A. M. Flanagan, K. Alitalo, and C. Boshoff. 00. Kaposi sarcoma herpesvirus-induced cellular reprogramming contributes to the lymphatic endothelial gene expression in Kaposi sarcoma. Nat. Genet. :-.. Wang, S. E., F. Y. Wu, H. Chen, M. Shamay, Q. Zheng, and G. S. Hayward. 00. Early activation of the Kaposi's sarcoma-associated herpesvirus RTA, RAP, and MTA promoters by the tetradecanoyl phorbol acetate-induced AP1 pathway. J. Virol. :-.. Wang, X. T., P. Y. Liu, and J. B. Tang. 00. PDGF gene therapy enhances expression of VEGF and bfgf genes and activates the NF-kappaB gene in signal pathways in ischemic flaps. Plast. Reconstr. Surg. :1-1.. Xie, J., H. Pan, S. Yoo, and S. J. Gao. 00. Kaposi's sarcoma-associated herpesvirus induction of AP-1 and interleukin during primary infection mediated by multiple mitogen-activated protein kinase pathways. J. Virol. :-.. Zhong, W., H. Wang, B. Herndier, and D. Ganem. 1. Restricted expression of Kaposi sarcoma-associated herpesvirus (human herpesvirus ) genes in Kaposi sarcoma. Proc. Natl. Acad. Sci. U. S. A. :1-. Downloaded from on December 1, 01 by guest
46 1 Table 1. Cytokines up regulated during KSHV infection of HMVEC-d cells. Conditioned media from h, h and h KSHV infected cells, and cells pretreated with Bay-0 followed by infection with KSHV were assayed using Ray Bio human cytokine antibody array V according to manufacturer s instructions. The densitometric values were substituted in the Ray Bio Human Cytokine antibody array V analysis tool and the entire array values were normalized to the same background level. The cytokines were categorized into pro-inflammatory cytokines, anti-inflammatory cytokines, growth factors, angiogenic factors, chemokines, tissue inhibitors of matrix metalloproteinases and growth factor binding proteins. The fold increase in the cytokine levels were calculated by dividing the respective values obtained from infected cells supernatant, both untreated and Bay-0 treated, with the values obtained from uninfected cell supernatant. Downloaded from on December 1, 01 by guest
47 Table 1. Cytokines up regulated during KSHV infection of HMVEC-d cells Fold Activation Cytokines Bay + Bay + Bay + KSHV h KSHV h KSHV h KSHV h KSHV h KSHV h Pro- inflammatory cytokines IL IL IL IL IL IL-1beta IL1-p IL-1alpha IL IFN-gamma LIGHT TNF-alpha GRO GRO-alpha TNF-beta Anti- inflammatory cytokines IL IL IL IL IL LIF Growth Factor PDGF-BB Leptin TGF-beta IGF GM-CSF TGF-beta G-CSF BDNF FGF FGF FGF FGF NT EGF TGF-beta PlGF M-CSF GDNF HGF NT Osteoprotegerin Downloaded from on December 1, 01 by guest
48 Table 1. Continued Fold Activation Cytokines Bay + Bay + Bay + KSHV h KSHV h KSHV h KSHV h KSHV h KSHV h Angiogenic factors SDF Angiogenin SCF Oncostatin M TPO VEGF Flt- Ligand Chemokines MCP TARC CK beta Eotaxin GCP MIF BLC MCP MDC MIG Eotaxin ENA PARC Eotaxin IP MIP--alpha I MCP RANTES MIP-1 beta MIP-1-delta Fractalkine NAP Growth factor binding proteins IGFBP IGFBP IGFBP IGFBP Tissue inhibitors of matrix metalloproteinases TIMP TIMP Fold Activation Downloaded from on December 1, 01 by guest
49 FIGURE LEGENDS Figure 1. (A), (B) and (C): Detection of activated NF-κB in KSHV infected HMVEC-d and HFF cells, HMVEC-d (A) and HFF (B) cells grown to 0% confluence were serum starved, infected with KSHV ( DNA copies/cell) and p protein phosphorylation monitored at the indicated time points. Cells were washed, lysed with RIPA lysis buffer, lysates adjusted to equal amounts of protein, resolved on SDS-% PAGE, and transferred to nitrocellulose membranes. Membranes were immunoblotted with monoclonal antibodies against phospho-p protein (top panel) or total p protein (middle panel) or β-actin (bottom panel). (C) HFF cells that were either uninfected or infected with KSHV ( DNA copies/cell) at various time points were Western blotted using phospho-iκb (top panel), total IκB (middle panel) and β-actin antibodies (bottom panel). Phosphorylated p level in uninfected cells was considered as 1 for comparison. (D), (E) and (F): Specificity of NF-κB induction by KSHV and inhibition by Bay-0. Serum-starved HMVEC-d cells (D) and HFF cells (E and F) untreated or pretreated with, or 0µM Bay-0 (lanes, and, respectively) were either uninfected (lane 1) or infected with DNA copies/cell of KSHV for 1 min. For control, serum starved cells were infected for 0 min with virus pre-incubated with 0 µg/ml of heparin for 0 min at C (lane ). Cell lysates were reacted in Western blot reactions with anti-phospho p antibodies (top panel). Membranes were stripped and reprobed with anti-p antibodies (middle panel) and β-actin antibodies (bottom panel). NF-κB induction with virus alone was considered 0%, and data are presented as the percentage inhibition of p phosphorylation. (F). Bay-0 pretreated HFF cell lysates were immunoblotted with phospho ERK1/ antibodies (top panel, lanes 1-). ERK1/ phosphorylation in virus infected cells was measured in the presence of MAPK inhibitor U01 (top panel, lane ). Blots were stripped and reprobed for total ERK (middle panel) and Downloaded from on December 1, 01 by guest
50 β-actin (bottom panel) levels. Each blot is representative of at least three independent experiments and percentage inhibition was calculated with respect to the phosphorylated levels of p in KSHV infected HMVEC-d cells without Bay-0 pretreatment. Figure. Nuclear translocation of NF-κB-p in KSHV infected cells. Serum starved HMVEC-d and HFF cells grown in eight well chamber slides were infected with KSHV ( DNA copies/cell) for 0 min and min respectively, washed, fixed, permeabilised and stained with anti-p polyclonal antibody. HMVEC-d and HFF cells were either uninfected (A, B, G and H) or infected with KSHV ( DNA copies/cell) (C, D, I and J) or incubated with µm Bay- 0 followed by infection with KSHV (E, F, K and L) and stained for NF-κB-p. DAPI was used as a nuclear stain and merged with p staining. Figure. Colocalization of NF-κB-p and ORF- (LANA-1) in KSHV infected HMVEC- d cells. (A) Serum starved HMVEC-d cells grown in eight well chamber slides were infected with KSHV ( DNA copies/cell) for h, washed, fixed, permeabilised and stained with anti- p polyclonal antibody. (B) Merged image of DAPI stained nuclei with p staining. (C) HMVEC-d cells infected with GFP-KSHV for h. (D) Merged image of DAPI stained nuclei with GFP staining. (E-L) show the colocalization of LANA-1 and p. LANA (green; E and I), p (red; F and J) in uninfected (E-H) and infected (I-L) HMVEC-d cells. (G and K) DAPI staining merged with LANA-1 and p (H and L). Arrows indicate p nuclear staining and arrow heads indicate cells positive for LANA-1 and p nuclear staining. Figure. Detection of KSHV induced nuclear translocation of NF-κB-p by ELISA. (A) Downloaded from on December 1, 01 by guest 0
51 Nuclear extracts from HMVEC-d and HFF cells infected with KSHV ( DNA copies/cell) for 0 min were prepared and assayed for NF-κB DNA binding activity by ELISA. Plates immobilized with oligonucleotide specific for the κb site were incubated with nuclear extracts (µg/well), followed by ELISA with anti-p antibody. The competition experiment was done in a similar fashion but using plates coated with excess (0pmol) NF-κB consensus site mutant κb or wt type oligonucleotides. Data represent the average ± standard deviations of three experiments. (B) HMVEC-d and HFF cells untreated or pretreated with varying concentrations of Bay-0 for 1 h were infected with KSHV ( DNA copies/cell) for 0 min, nuclear extracts prepared and assayed for NF-κB DNA binding activity. Percentage nuclear translocation of NF-κB-p inhibition by Bay-0 pretreatment was calculated with respect to the DNA binding activities in untreated KSHV infected cells. (C) Histograms depicting the kinetics of percentage inhibition of DNA binding activity in nuclear extracts from HMVEC-d and HFF cells pretreated with µm Bay-0 for 1 h and then infected with KSHV ( DNA copies/cell) for different time points. Data represent the average ± standard deviations of three experiments. Figure. EMSA detection of KSHV induced NF-κB DNA binding activity. Top panel, Lanes 1-: Untreated or µm Bay-0 pretreated (1 h) HMVEC-d (A) and HFF (B) cells were uninfected or infected with KSHV ( DNA copies/cell) for the indicated times. Nuclear extracts prepared from these cells were incubated with κb specific probe. The specificity of DNA-protein interaction was assessed by induction with TNFα (0ng/ml) (lane ), free probe (lane ), normal probe (lane ) or competitive EMSA using a 0X molar excess of unlabeled double-stranded oligonucleotide κb probe (cold probe lane ). Nuclear extracts from HMVECd cells infected with DNA copies/cell of live KSHV (C) or UV-KSHV (D) for h, h and Downloaded from on December 1, 01 by guest 1
52 h were analyzed by EMSA. Each EMSA is a representative of at least three independent experiments. Bottom panels (A and B) Lanes 1- and bottom panels (C and D) Lanes 1-: Oct1 probe used as loading control and specificity control to demonstrate the binding of NF-κB to the specific probe. Arrows denote the position of the induced NF-κB complexes. Figure. Sustained activation of NF-κB during de novo infection of target cells by KSHV. Proteins prepared from HMVEC-d (A) and HFF cells (B) that were uninfected (UI) or infected with KSHV ( DNA copies/cell) for min, min, 0 min, 1 h, h, h, h, 1 h, h, h h and h were resolved by SDS-PAGE and transferred onto nitrocellulose membranes. Phosphorylated and total p, ERK 1/, AKT and pmapk proteins were detected with the respective antibodies. Each blot is representative of at least three independent experiments. The phosphorylated levels in the uninfected cells were considered as 1 for comparison. For control, cells were induced with TNFα for 0 min. Figure. Effect of Bay-0 on KSHV entry and KSHV ORF, ORF 0, K, K and virf gene expression. (A) HMVEC-d and HFF cells untreated or incubated with different concentrations of inhibitors for 1 h at C were infected with KSHV at DNA copies/cell. For control, cells were pre-incubated with 0µM LY00 (LY) for 1 h at C before adding virus. After h incubation, cells were washed twice with PBS to remove the unbound virus, treated with trypsin-edta for min at C to remove the bound but non-internalized virus, washed, and total DNA was isolated. This was normalized, and the numbers of KSHV genome copies were estimated by real-time DNA PCR for ORF. The Ct values were used to plot the standard graph and to calculate the relative copy number of viral DNA in the samples. Data are Downloaded from on December 1, 01 by guest
53 presented as the percentage entry of KSHV DNA internalization obtained when the cells were incubated with virus alone. Each reaction was done in duplicate, and each bar represents the mean standard deviation of three experiments. (B-F) Untreated HMVEC-d and HFF cells or cells pretreated with µm Bay-0 for 1 h were infected with KSHV ( DNA copies/cell) for h, h and h, RNA was isolated and treated with DNase I for 1 h. A total of 0ng of DNasetreated RNA was subjected to real-time RT-PCR with ORF, ORF 0, K, K and virf gene-specific primers and TaqMan probes. Standard graphs generated using known concentrations of DNase-treated in vitro transcribed ORF, ORF 0, K K and virf transcripts were used to calculate the relative copy numbers of viral transcripts and were normalized with GAPDH. Each reaction was done in duplicate, and each point represents the average + standard deviation of three independent experiments. (B) Kinetics of ORF and 0 gene expression in HMVEC-d cells. (C and D) Comparative kinetics of ORFs, 0, K, K and virf in HMVEC-d and HFF cells respectively. (E and F) Histogram depicting the percentage inhibition of KSHV ORFs, 0, K, K and virf expression in the presence of Bay-0 in HMEVC-d and HFF cells respectively. Figure. Effect of NF-κB inhibition on AP-1 transcription factors activation. (A) Nuclear extracts prepared from uninfected HMVEC-d cells or HMVEC-d cells infected with KSHV for 1 min, 0 min and 0 min were tested for the activation of AP-1 regulated transcription factors by incubating the nuclear extracts with the plate-immobilized oligonucleotides containing the AP-1 transcription factor-specific site, followed by ELISA with antibodies to the respective transcription factors. The histogram represents the activation levels of phospho-c-jun, JunB, JunD, Fra1, Fra, Fos-B and c-fos in the nuclear extracts from KSHV-infected HMVEC-d cells. Downloaded from on December 1, 01 by guest
54 Data represent the average standard deviation of three experiments, and values shown here are after normalization with uninfected cells. (B) Histogram depicting the percentage inhibition of DNA binding of AP-1 transcription factors in nuclear extracts from HMVEC-d pretreated with different concentrations of Bay-0 and µm U01 followed by infection with KSHV. (C) Histogram depicting the percentage activation of DNA binding of the p-c-jun transcription factor in HMVEC-d nuclear extracts. Percentage inhibition and percentage activation were calculated with respect to the DNA binding activities in KSHV-infected HMVEC-d cells without Bay- 0 pretreatment. Data represent the average + standard deviations of three experiments. Figure. Upregulation of pro-inflammatory cytokines, growth factors, angiogenic factors and chemokines in HMVEC-d cells by KSHV. Densitometric analysis of cytokine array blots was carried out to determine the difference in the release of human cytokines from serum starved, untreated HMVEC-d cells and KSHV infected cells at three different time points. The values were normalised to identical background levels using the Ray Bio Human Cytokine antibody array V analysis tool. The fold increases in the cytokine levels were calculated by dividing the respective values obtained from infected cells supernatant, with the values obtained from uninfected cells supernatant and cytokines showing significant change are represented in a line graph format. (A) Pro-inflammatory cytokines; (B) Growth factors; (C) Angiogenic factors, and (D) Chemokines that showed significant changes with respect to uninfected are represented in the graph. Figure. Schematic representation depicting the early and late induction phases of NF-κB during in vitro KSHV infection of HMVEC-d cells and their potential roles in transcription Downloaded from on December 1, 01 by guest
55 1 1 1 factor regulation, establishment and maintenance of KSHV infection and cytokine secretion. In the early phase of NF-κB induction (blue arrows), virus binding and entry leads to signal pathway induction such as FAK, Src, PI-K, AKT, PKC-ζ, MAPK-ERK1/ and NF-κB signal molecules. Activated NF-κB translocates into the nucleus, which coincides with viral DNA entry into the infected cell nuclei, concurrent transient expression of limited viral lytic genes, and persistent latent gene expression. Overlapping with these events, a limited number of cytokine and growth factors are induced which is initiated by transcription factors like AP-1 (induced by ERK1/ and NF-κB). Early activation of NF-κB and ERK1/ also leads to the activation and release of NFκB inducible host factors, which act in an autocrine and paracrine fashion on the infected as well as neighboring cells. Autocrine action of these factors along with viral gene expression probably contributes to the second or late phase of signal pathway activation (red arrows) including sustained NF-κB activation, phosphorylation of pmapk, ERK1/ and AKT required for the maintenance of latency. Blue and red arrows together indicate pathways induced both during early and late phases of KSHV infection. Downloaded from on December 1, 01 by guest

56

57

58

59

60

61

62

63
64

65
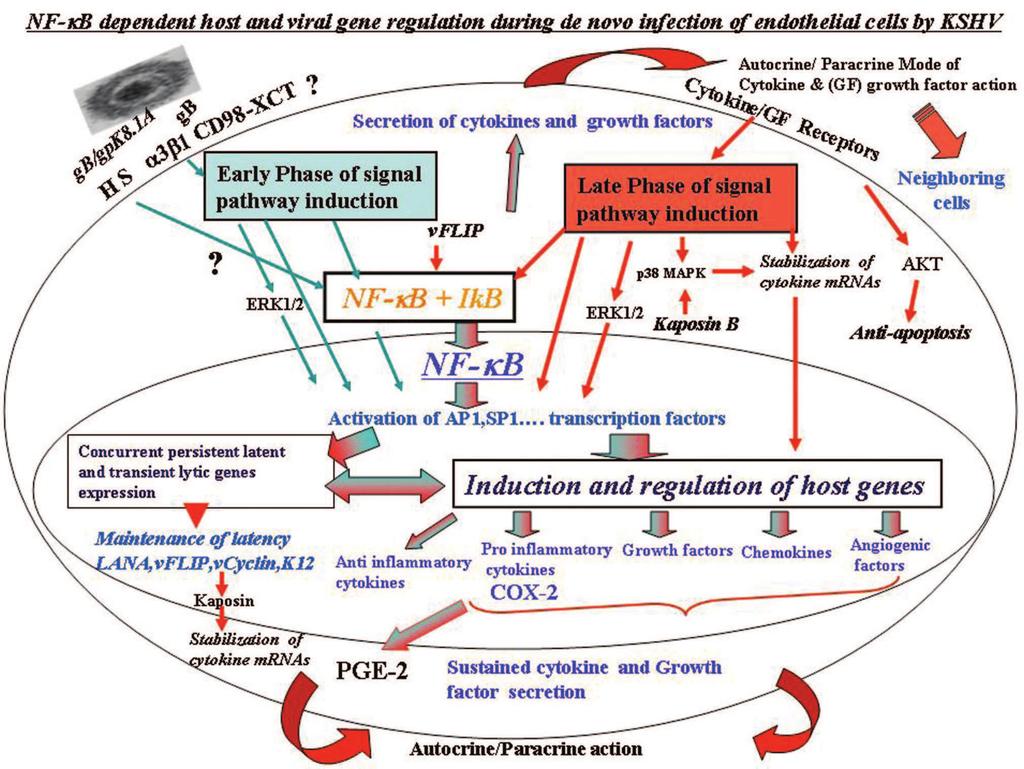
66
Sathish Sadagopan, Neelam Sharma-Walia, Mohanan Valiya Veettil, Hari Raghu, Ramu Sivakumar, Virginie Bottero, and Bala Chandran*
 JOURNAL OF VIROLOGY, Apr. 2007, p. 3949 3968 Vol. 81, No. 8 0022-538X/07/$08.00 0 doi:10.1128/jvi.02333-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
JOURNAL OF VIROLOGY, Apr. 2007, p. 3949 3968 Vol. 81, No. 8 0022-538X/07/$08.00 0 doi:10.1128/jvi.02333-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
Received 1 February 2006/Accepted 5 April 2006
 JOURNAL OF VIROLOGY, July 2006, p. 6534 6552 Vol. 80, No. 13 0022-538X/06/$08.00 0 doi:10.1128/jvi.00231-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Cyclooxygenase 2 Induced
JOURNAL OF VIROLOGY, July 2006, p. 6534 6552 Vol. 80, No. 13 0022-538X/06/$08.00 0 doi:10.1128/jvi.00231-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Cyclooxygenase 2 Induced
Received 24 April 2007/Accepted 20 November 2007
 JOURNAL OF VIROLOGY, Feb. 2008, p. 1759 1776 Vol. 82, No. 4 0022-538X/08/$08.00 0 doi:10.1128/jvi.00873-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
JOURNAL OF VIROLOGY, Feb. 2008, p. 1759 1776 Vol. 82, No. 4 0022-538X/08/$08.00 0 doi:10.1128/jvi.00873-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
Updated information and services can be found at:
 REFERENCES CONTENT ALERTS Lipid Rafts of Primary Endothelial Cells Are Essential for Kaposi's Sarcoma-Associated Herpesvirus/Human Herpesvirus 8-Induced Phosphatidylinositol 3-Kinase and RhoA-GTPases Critical
REFERENCES CONTENT ALERTS Lipid Rafts of Primary Endothelial Cells Are Essential for Kaposi's Sarcoma-Associated Herpesvirus/Human Herpesvirus 8-Induced Phosphatidylinositol 3-Kinase and RhoA-GTPases Critical
Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators
 /, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
/, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
NFAT and CREB Regulate Kaposi s Sarcoma-Associated Herpesvirus-Induced Cyclooxygenase 2 (COX-2)
") JOURNAL OF VIROLOGY, Dec. 2010, p. 12733 12753 Vol. 84, No. 24 0022-538X/10/$12.00 doi:10.1128/jvi.01065-10 Copyright 2010, American Society for Microbiology. All Rights Reserved. NFAT and CREB Regulate
JOURNAL OF VIROLOGY, Dec. 2010, p. 12733 12753 Vol. 84, No. 24 0022-538X/10/$12.00 doi:10.1128/jvi.01065-10 Copyright 2010, American Society for Microbiology. All Rights Reserved. NFAT and CREB Regulate
Harinivas H. Krishnan, 1 Pramod P. Naranatt, 1 Marilyn S. Smith, 1 Ling Zeng, 1 Clark Bloomer, 2 and Bala Chandran 1 *
 JOURNAL OF VIROLOGY, Apr. 2004, p. 3601 3620 Vol. 78, No. 7 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.7.3601 3620.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Concurrent
JOURNAL OF VIROLOGY, Apr. 2004, p. 3601 3620 Vol. 78, No. 7 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.7.3601 3620.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Concurrent
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Kaposi s sarcoma (KS) is an angioproliferative and highly vascularized
 is an angioproliferative and highly vascularized") Kaposi s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen Interacts with Multifunctional Angiogenin To Utilize Its Antiapoptotic Functions Nitika Paudel, a Sathish Sadagopan, a Sayan Chakraborty,
Kaposi s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen Interacts with Multifunctional Angiogenin To Utilize Its Antiapoptotic Functions Nitika Paudel, a Sathish Sadagopan, a Sayan Chakraborty,
Kaposi's Sarcoma-Associated Herpesvirus Induces ATM and H2AX. DNA Damage Response Early During de novo Infection of Primary
 JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Overview of primary HHV-8 infection
 Overview of primary HHV-8 infection HHV-8, also known as Kaposi sarcoma-associated herpesvirus (KSHV), is a gamma herpesvirus primarily transmitted through saliva. The virus initially replicates in epithelial
Overview of primary HHV-8 infection HHV-8, also known as Kaposi sarcoma-associated herpesvirus (KSHV), is a gamma herpesvirus primarily transmitted through saliva. The virus initially replicates in epithelial
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
RayBio Human Thyroglobulin ELISA Kit
 RayBio Human Thyroglobulin ELISA Kit Catalog #: ELH-Thyroglobulin User Manual Last revised July 6, 2017 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
RayBio Human Thyroglobulin ELISA Kit Catalog #: ELH-Thyroglobulin User Manual Last revised July 6, 2017 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
MINIREVIEW. Early Events in Kaposi s Sarcoma-Associated Herpesvirus Infection of Target Cells. Bala Chandran*
 JOURNAL OF VIROLOGY, Mar. 2010, p. 2188 2199 Vol. 84, No. 5 0022-538X/10/$12.00 doi:10.1128/jvi.01334-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. MINIREVIEW Early Events
JOURNAL OF VIROLOGY, Mar. 2010, p. 2188 2199 Vol. 84, No. 5 0022-538X/10/$12.00 doi:10.1128/jvi.01334-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. MINIREVIEW Early Events
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
Pre-made Reporter Lentivirus for NF-κB Signal Pathway
 Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB
 Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB Kenji Kawamura, Yoshio Kano. Kibi International University, Takahashi-city, Japan. Disclosures: K.
Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB Kenji Kawamura, Yoshio Kano. Kibi International University, Takahashi-city, Japan. Disclosures: K.
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Supplementary Materials and Methods
 Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Erzsebet Kokovay, Susan Goderie, Yue Wang, Steve Lotz, Gang Lin, Yu Sun, Badrinath Roysam, Qin Shen,
 Cell Stem Cell, Volume 7 Supplemental Information Adult SVZ Lineage Cells Home to and Leave the Vascular Niche via Differential Responses to SDF1/CXCR4 Signaling Erzsebet Kokovay, Susan Goderie, Yue Wang,
Cell Stem Cell, Volume 7 Supplemental Information Adult SVZ Lineage Cells Home to and Leave the Vascular Niche via Differential Responses to SDF1/CXCR4 Signaling Erzsebet Kokovay, Susan Goderie, Yue Wang,
alveolar macrophages (AMs) after 24 hours of in vitro culture in complete medium
 after 24 hours of in vitro culture in complete medium") Online Supplement for: NF-κB ACTIVATION IN ALVEOLAR MACROPHAGES REQUIRES IκB KINASE-β, BUT NOT NF-κB INDUCING KINASE Supershift and Competition Assays for NF-κB Competition and supershift assays were performed
Online Supplement for: NF-κB ACTIVATION IN ALVEOLAR MACROPHAGES REQUIRES IκB KINASE-β, BUT NOT NF-κB INDUCING KINASE Supershift and Competition Assays for NF-κB Competition and supershift assays were performed
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
RayBio Human ENA-78 ELISA Kit
 RayBio Human ENA-78 ELISA Kit Catalog #: ELH-ENA78 User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite 100 Norcross,
RayBio Human ENA-78 ELISA Kit Catalog #: ELH-ENA78 User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite 100 Norcross,
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Biol403 MAP kinase signalling
 Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
NF-κB p65 (Phospho-Thr254)
") Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
Kaposi s Sarcoma-Associated Herpesvirus Induces Nrf2 during De Novo Infection of Endothelial Cells to Create a Microenvironment Conducive to Infection
 Kaposi s Sarcoma-Associated Herpesvirus Induces Nrf2 during De Novo Infection of Endothelial Cells to Create a Microenvironment Conducive to Infection Olsi Gjyshi, Virginie Bottero, Mohanan Valliya Veettil,
Kaposi s Sarcoma-Associated Herpesvirus Induces Nrf2 during De Novo Infection of Endothelial Cells to Create a Microenvironment Conducive to Infection Olsi Gjyshi, Virginie Bottero, Mohanan Valliya Veettil,
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Intrinsic cellular defenses against virus infection
 Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
- 1 - Cell types Monocytes THP-1 cells Macrophages. LPS Treatment time (Hour) IL-6 level (pg/ml)
 IL-6 level (pg/ml)") Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Signaling. Dr. Sujata Persad Katz Group Centre for Pharmacy & Health research
 Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Genome of Hepatitis B Virus. VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department
 Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set
 p24 / Capsid Protein p24 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
Basis of Immunology and
 Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation
 MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation Tao Li 1 *, Michael J. Morgan 1 *, Swati Choksi 1, Yan Zhang 1, You-Sun Kim 2#, Zheng-gang
MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation Tao Li 1 *, Michael J. Morgan 1 *, Swati Choksi 1, Yan Zhang 1, You-Sun Kim 2#, Zheng-gang
Supplementary Material
 Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Rabbit Polyclonal antibody to NFkB p65 (v-rel reticuloendotheliosis viral oncogene homolog A (avian))
)") Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Received 2 November 2007/Accepted 19 February 2008
 JOURNAL OF VIROLOGY, May 2008, p. 4235 4249 Vol. 82, No. 9 0022-538X/08/$08.00 0 doi:10.1128/jvi.02370-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
JOURNAL OF VIROLOGY, May 2008, p. 4235 4249 Vol. 82, No. 9 0022-538X/08/$08.00 0 doi:10.1128/jvi.02370-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Absence of Kaposi s Sarcoma-Associated Herpesvirus in Patients with Pulmonary
 Online Data Supplement Absence of Kaposi s Sarcoma-Associated Herpesvirus in Patients with Pulmonary Arterial Hypertension Cornelia Henke-Gendo, Michael Mengel, Marius M. Hoeper, Khaled Alkharsah, Thomas
Online Data Supplement Absence of Kaposi s Sarcoma-Associated Herpesvirus in Patients with Pulmonary Arterial Hypertension Cornelia Henke-Gendo, Michael Mengel, Marius M. Hoeper, Khaled Alkharsah, Thomas
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Sarah A. Kopecky and Douglas S. Lyles*
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Supplementary Information
 Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic
 Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
Chemical Chaperones Mitigate Experimental Asthma By Attenuating Endoplasmic Reticulum Stress Lokesh Makhija, BE, Veda Krishnan, MSc, Rakhshinda Rehman, MTech, Samarpana Chakraborty, MSc, Shuvadeep Maity,
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Supplemental Figure 1. Western blot analysis indicated that MIF was detected in the fractions of
 Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
well for 2 h at rt. Each dot represents an individual mouse and bar is the mean ±
 Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Online Data Supplement. Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2
 Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Anti-Lamin B1/LMNB1 Picoband Antibody
 Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
Anti-Lamin B1/LMNB1 Picoband Antibody Catalog Number:PB9611 About LMNB1 Lamin-B1 is a protein that in humans is encoded by the LMNB1 gene. The nuclear lamina consists of a two-dimensional matrix of proteins
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit
 RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,