Morphology and Dynamics of Clathrin/ GGA1-Coated Carriers. Budding from the Trans-Golgi Network
|
|
|
- Wilfred Jennings
- 6 years ago
- Views:
Transcription
1 MBC in Press, published on January 26, 2003 as /mbc Morphology and Dynamics of Clathrin/ GGA1-Coated Carriers Budding from the Trans-Golgi Network Rosa Puertollano*, Nicole N. van der Wel, Lois E. Greene, Evan Eisenberg, Peter J. Peters and Juan S. Bonifacino* *Cell Biology and Metabolism Branch National Institute of Child Health and Human Development Laboratory of Cell Biology National Heart, Lung, and Blood Institute National Institutes of Health Bethesda, MD 20892, USA Division of Tumor Biology The Netherlands Cancer Institute Amsterdam, The Netherlands Corresponding author: Juan S. Bonifacino, Cell Biology and Metabolism Branch, National Institute of Child Health and Human Development, Building 18T/Room 101, National Institutes of Health, Bethesda, MD 20892, USA. Tel: Fax:
2 2
3 ABSTRACT Sorting of transmembrane proteins and their ligands at various compartments of the endocytic and secretory pathways is mediated by selective incorporation into clathrin-coated intermediates. Previous morphological and biochemical studies have shown that these clathrin-coated intermediates consist of spherical vesicles with a diameter of nm. Here we report the use of fluorescent imaging of live cells to demonstrate the existence of a different type of transport intermediate containing associated clathrin coats. Clathrin and the adaptors GGA1 and AP-1, labeled with different spectral variants of the green fluorescent protein, are shown to co-localize to the trans-golgi network and to a population of vesicles and tubules budding from it. These intermediates are highly pleiomorphic and move towards to peripheral cytoplasm for distances of up to 10 µm with average speeds of ~1 µm/s. The labeled clathrin and GGA1 cycle on and off membranes with half-times of s, independently of vesicle budding. Our observations suggest the existence of a novel type of TGN-derived carrier containing associated clathrin, GGA1 and AP-1 that are larger than conventional clathrin-coated vesicles, and that undergo long-range translocation in the cytoplasm before losing their coats. 3
4 INTRODUCTION Clathrin coats associated with the cytosolic face of membranes mediate the sorting of transmembrane proteins and their bound ligands at the plasma membrane, the trans-golgi network (TGN) and endosomes (Kirchhausen, 2000; Brodsky et al., 2001). The major building blocks of clathrin coats are clathrin triskelia composed of three heavy chains and three light chains (a or b isoforms), which assemble into polyhedral lattices. At each intracellular location, clathrin coats contain a characteristic set of heterotetrameric adaptor protein (AP) complexes that mediate both attachment of the clathrin lattices to membranes and concentration of specific transmembrane proteins. The AP-2 complex plays such roles at the plasma membrane, while the AP-1 complex does so at the TGN and/or endosomes (Kirchhausen, 2000; Brodsky et al., 2001; Robinson and Bonifacino, 2001). Recently, a family of monomeric proteins termed GGAs (i.e., GGA1, GGA2 and GGA3 in humans) has also been shown to promote recruitment of clathrin and sorting of transmembrane proteins at the TGN (Robinson and Bonifacino, 2001; Boman, 2001). In particular, the GGAs have been implicated in the sorting the cation-dependent and cation-independent mannose 6-phosphate receptors (CD- and CI-MPRs, respectively) from the TGN to the endosomal-lysosomal system (Puertollano et al., 2001a; Zhu et al., 2001; Takatsu et al., 2001). This sorting is mediated by interaction of the amino-terminal VHS domain of the GGAs and acidic-cluster dileucine sorting signals present in the 4
5 cytoplasmic domains of the MPRs (Puertollano et al., 2001a; Zhu et al., 2001; Takatsu et al., 2001). The VHS domain of the GGAs has also been shown to interact with acidic-cluster dileucine signals in the cytoplasmic domains of other transmembrane proteins such as sortilin (Nielsen et al., 2001), the LDL receptor-related protein 3 (LRP3) (Takatsu et al., 2001), and β-secretase (or BACE) (He et al., 2002), all of which may also cycle between the TGN and endosomes. Despite recent advances in the elucidation of the role of the GGAs in protein sorting, many outstanding issues remain concerning (a) whether the GGAs co-localize with clathrin and AP-1 on transport intermediates budding from the TGN, (b) the morphology of these GGA-containing intermediates, (c) the intracellular destination of these carriers, and (d) the dynamics of GGA cycling on and off TGN membranes in vivo. These issues have recently become amenable to analysis through the use of fluorescence imaging technologies on live cells. The dynamics of plasma membrane clathrin-coated pits, for example, have been examined using clathrin constructs tagged with variants of the green fluorescent protein (GFP) (Damer and O'Halloran, 2000; Gaidarov et al., 1999; Wu et al., 2001). This approach revealed that clathrin-coated vesicles (CCVs) emanate repeatedly from the same sites, suggesting the existence of hot-spots for CCV formation from the plasma membrane (Gaidarov et al., 1999). The same approach was used to demonstrate a rapid (t 1/2 ~16s), ATP-dependent exchange of clathrin triskelia on plasma membrane coated pits, which may allow remodeling of the coat as the membrane invaginates (Wu et al., 2001). Strikingly, clathrin exchange 5
6 on the pits occurs even when endocytosis is blocked (Wu et al., 2001), indicating that the coats are not static assemblies and that uncoating does not require vesicle detachment from the membrane. To date, the characteristics of protein coats associated with the TGN in living cells remain to be similarly examined. Here we report the results of a study on the morphology and dynamics of GGA1-, AP-1- and clathrin-coated structures at the TGN in vivo. Using GGA1, the γ1 subunit of AP-1, and clathrin light chain b isoform labeled with different spectral variants of GFP, we show that all of these coat proteins co-localize to the TGN and to a population of vesicular-tubular carriers budding from the TGN. Surprisingly, these carriers are pleiomorphic and apparently larger than conventional CCVs. They move centrifugally for distances of up to 10 µm and with average speeds of 1 µm/s. The fluorescently-labeled clathrin and GGA1 constructs cycle on and off membranes with t 1/2 of 10-20s. This cycling is not dependent on budding from the TGN, suggesting that assembly and disassembly of the coats can be functionally uncoupled from vesicle budding. These results suggest the existence of a novel type of TGN-derived carriers containing associated clathrin that are larger and more dynamic than conventional CCVs, and that undergo long-range translocation in the cytoplasm before losing their coat. 6
7 MATERIALS AND METHODS Recombinant DNA procedures and antibodies A cdna encoding full-length human GGA1 was cloned into the SalI and BamHI sites of the pegfp-c1, peyfp-c1, and pecfp-c1 vectors (Clontech). Clathrin light chain b was amplified from human placenta Quick-Clone cdna (Clontech) and cloned into the peyfp-c2 vector via EcoRI and SalI sites. Plasmids encoding CFP-VSV-G (ts045 mutant) and CFP-galactosyl transferase were the kind gift of Jennifer Lippincott-Schwartz (NICHD). The cloning of the Myc-epitope-tagged GGA1 and CD-MPR-CFP was described previously (Dell'Angelica et al., 2000; Puertollano et al., 2001a). The following commercial antibodies were used: mouse monoclonal and rabbit polyclonal anti-myc antibodies (9E10, Covance, Princeton, NJ); mouse monoclonal antibody to γ1-adaptin (100/3, Sigma Chemical Co., St. Louis, MO); mouse monoclonal antibody to clathrin (CHC, Transduction Laboratories, Lexington, KY). The preparation of rabbit polyclonal antibody to AP-3 (β3c1) has been described in a previous report (Dell'Angelica et al., 2000). A polyclonal antibody to GGA1 was the kind gift of Margaret S. Robinson (University of Cambridge, UK). 7
8 Immunofluorescence microscopy Non-polarized MDCK cells were grown on coverslips and fixed in methanol/acetone (1:1, v/v) for 10 min at -20 C and subsequently air-dried. Incubation with primary antibodies diluted in PBS, 0.1% (w/v) saponin, 0.1% BSA, was carried out for 1 h at room temperature. Unbound antibodies were removed by rinsing with PBS for 5 min, and cells were subsequently incubated with secondary antibodies (Cy3-conjugated donkey anti-rabbit Ig and Alexa 448- conjugated donkey anti-mouse Ig) diluted in PBS, 0.1% (w/v) saponin, 0.1% BSA, for min at room temperature. After a final rinse with PBS, coverslips were mounted onto glass slides with Fluoromount G (Southern Biotechnology Associates, Birmingham, AL). Fluorescence images were acquired on a Zeiss LSM 410 or LSM 510 confocal microscope (Carl Zeiss Inc., Thornwood, NY). Immunoelectron Microscopy Cryogold immunoelectron microscopy of monocyte-derived dendritic cells and stably transfected MDCK cells was performed as previously described (Porcelli et al., 1992; Peters, 1998). Fluorescent imaging of living cells MDCK cells were grown on LabTek chambers (Nalge Nunc, Naperville, IL), transfected with different constructs tagged with GFP spectral variants by using FuGENE 6 (Roche Molecular Biochemicals), and transferred into culture medium 8
9 buffered with 25 mm Hepes/KOH ph 7.4. Experiments were performed using an inverted confocal laser scanning microscope (Carl Zeiss LSM 510) equipped with a stage heated to 37 C, Argon, HeNe, and Krypton lasers, and a 63X 1.4 NA objective. YFP and CFP fluorescence were visualized using excitation filters at 514 and 413 nm and emission filters at 530 and nm, respectively, while GFP was excited with a 488 nm filter and imaged through a nm emission filter. When indicated, endosomes were loaded with rhodamine-conjugated human serum albumin or rhodamine-conjugated transferrin (Molecular Probes, Eugene, OR), which were imaged using a 543 nm laser. Cells expressing CFP- VSV-G (ts045 mutant) were cultured at 40 C for 24 h, shifted to 20 C for 1 h and then to 32 C prior to visualization. Images were recorded every 3 or 6 s. To estimate the size of GGA1-coated intermediaries, MDCK cells expressing YFP- GGA1 were incubated with medium containing yellow-green fluorescent beads (Molecular Probes, FluoSpheres carboxylate-modified microspheres 2.0 µm) and analyzed by time-lapse confocal microscopy as previously described. Data were processed to QuickTime format in Adobe Premiere 5.0. Photobleaching was performed at high laser power (100% power, 100% transmission). Recovery was followed by scanning the whole cell at lower power (40%) and attenuated transmission (3%) at intervals from 5 s to 5 min. A nearby background region of equal area was similarly measured and subtracted. Background pixel values were <20% of the average intensity of these structures. 9
10 Fluorescence intensity was normalized to the first time point. 10
11 RESULTS Distribution of coat proteins associated with the TGN in MDCK cells. We chose to perform our imaging studies on non-polarized MDCK epithelial cells because they (i) are large and flat, (ii) have an extensive TGN, and (iii) yield low to moderate expression of constructs driven from the human cytomegalovirus promoter, as is the case for the plasmid vectors used here. The use of MDCK cells, however, necessitated a re-examination of the distribution of various coat proteins associated with the TGN, as previously established for other cell lines. We started by performing immunofluorescence microscopy of MDCK cells stably transfected with a Myc-tagged GGA1 construct (Myc-GGA1). Immunoblot analyses with an antibody to GGA1 revealed that the expression levels of Myc-GGA1 were 4-5-fold higher than those of endogenous GGA1 (data not shown). Consistent with previous findings in other cell lines (Puertollano et al., 2001b; Hirst et al., 2001), we observed extensive co-localization of Myc-GGA1 with clathrin in the area of the TGN (Fig. 1, A-C). We also observed considerable overlap of Myc-GGA1 and AP-1 staining at the TGN (Fig. 1, D-F). Outside the TGN, Myc-GGA1, AP-1, and clathrin displayed various degrees of colocalization to cytoplasmic structures having apparent diameters of ~400 nm (Fig. 1, insets). Quantification of these results showed that almost 100% of all resolvable GGA1-containing foci co-localized with clathrin (148 foci counted), while 40% co-localized with AP-1 (110 foci counted). Another AP complex, AP-3, 11
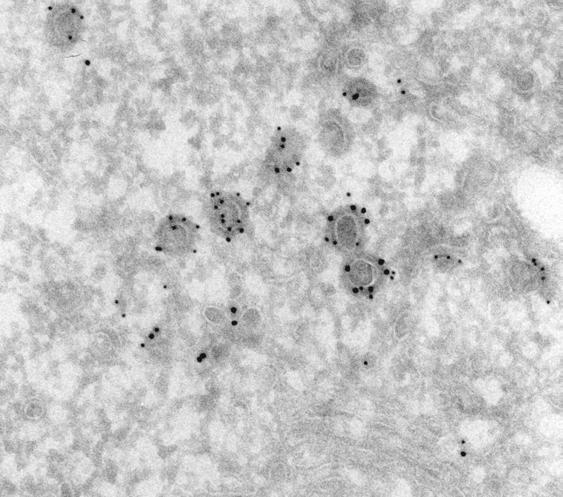
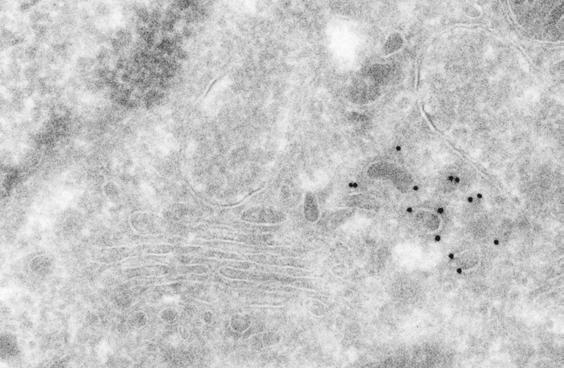
12 was more peripherally distributed than either GGA1 or AP-1 and therefore exhibited substantially less co-localization with Myc-GGA1 in the area of the TGN (Fig. 1, G-I). Some peripheral foci, however, appeared to contain both AP-3 and Myc-GGA1 (Fig. 1, G-I, inset). Immunoelectron microscopy of monocyte-derived dendritic cells, in which endogenous GGA1 can be more easily detected, revealed localization of endogenous GGA1 to coated buds with diameters of nm in the area of the TGN (Fig. 2A), as well as in the peripheral cytoplasm (data not shown). A similar localization was observed for Myc-GGA1 in stably transfected MDCK cells (Fig. 2, B-D). Many coated buds were found to contain both Myc-GGA1 and clathrin (Fig. 2B, open arrowheads). We also observed partial co-localization of Myc- GGA1 and AP-1 on coated buds with diameters of nm at both the TGN (Fig. 2, C-D, open arrowheads) and the cell periphery (Fig. 2D, open arrowheads). However, some individual buds contained gold particles labeling only Myc-GGA1 or AP-1 (Fig. 2, C-D). We detected some gold labeling for all of these coat proteins in the cytosol, consistent with the fact that they cycle between membranes and the cytosol. Buds containing these coat proteins often occurred in mixed clusters, which might correspond to the larger peripheral structures observed by fluorescence microscopy. Characteristics of coated carriers budding from the TGN. 12
13 To examine the morphology and dynamics of coated carrier vesicles budding from the TGN, we constructed plasmids encoding various spectral variants of GFP attached to clathrin light chain b isoform, GGA1 and the γ1-adaptin subunit of AP-1. These proteins were expressed by transfection into MDCK cells. At low to moderate levels of expression, all three GFP-tagged proteins localized to both juxtanuclear and peripheral structures (Fig. 3, A-C), similar to those shown in Fig. 1. Time-lapse confocal microscopy of live cells expressing each of these GFPtagged constructs revealed that some of the peripheral foci were relatively static (arrowheads) while others moved rapidly from the TGN towards the peripheral cytoplasm (arrows) (Fig. 3, A-C). The structures emanating from the TGN were relatively large ( nm average diameters, Table 1) in comparison to plasma membrane clathrin-coated pits (Gaidarov et al., 1999; Wu et al., 2001). This difference could be readily appreciated in fields where the larger TGN-derived, clathrin-coated intermediates moved in relation to smaller, less mobile clathrincoated pits at the cell edges (Fig. 3D, size comparison shown in the insets). In addition, the intermediates appeared larger than 200 nm-diameter fluorescent beads of similar brightness added to the cell cultures before imaging (Fig. 4A, compare fluorescent beads in the insets with moving intermediate indicated by arrow). The TGN-derived intermediates decorated with any of the three GFPfusion proteins were quite pleiomorphic. Most appeared spheroidal or ellipsoidal. Some detached from the TGN as single vesicles whereas others 13
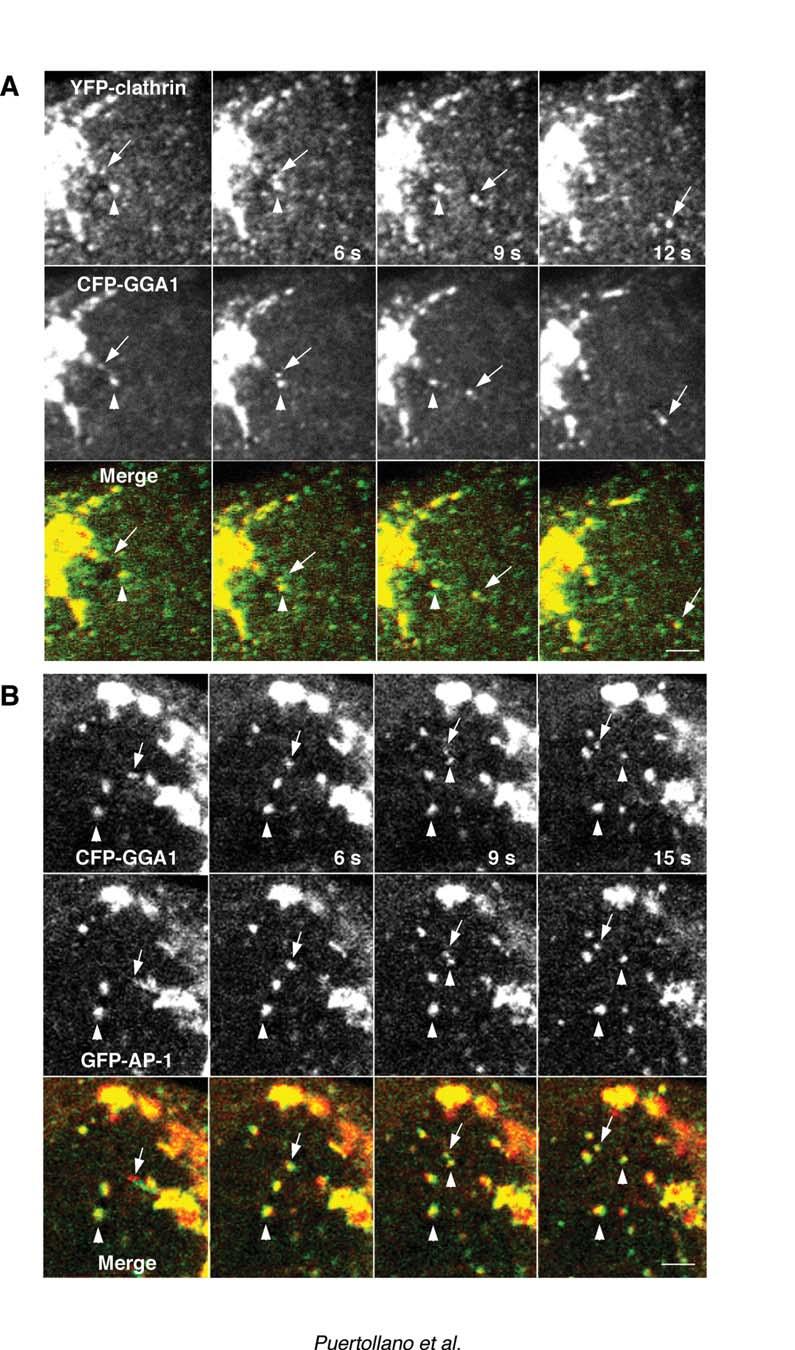
14 pulled off in groups resembling beads on a string (e.g., Figs. 3C, 6A, 7A and 7B). They often changed shapes and some even divided during transport (e.g., Fig. 5B, arrow between 6 and 9 s). Others joined or fused during transport, generating structures that were bigger and brighter than the original vesicles (Fig. 4B). The TGN-derived intermediates moved centrifugally with average speeds of ~1 µm/s. Their trajectories were quasi-linear, as if they were tethered to a system of radial tracks. Treatment with the microtubule-depolymerizing agent, nocodazole, abolished the long-range translocation of the TGN-derived intermediates (data not shown) suggesting that movement occurred along microtubules. Co-expression of combinations of coat proteins, each tagged with a different GFP variant, allowed us to analyze for their presence on the same coated intermediates. We observed that most TGN-derived intermediates contained both clathrin and GGA1 (an example is shown in Fig. 5A). We could also observe budding of intermediates that contained both GGA1 and AP-1 (Fig. 5B. As previously shown (Puertollano et al., 2001a), tubules and vesicles carrying the CD-MPR from the TGN almost always exhibited associated GGA1. In the example shown in Fig. 6A, a string of GGA1-containing structures can be seen decorating in discontinuous fashion a CD-MPR-carrying tubule. We occasionally saw endosomal recycling tubules labeled with internalized rhodamine-transferrin pulling off the juxtanuclear area of the cell, but these 14
15 almost never contained associated GGA1 (Fig. 6B). While GGA1- and AP-1- containing intermediates detached and moved away from the TGN, the Golgi resident enzyme galactosyl transferase stayed behind in the Golgi complex (Fig. 7, A and B). GGA1-containing intermediates were distinct from post-golgi carriers (i.e., PGCs; Hirschberg et al., 1998; Polishchuk et al., 2000) that transport the vesicular stomatitis virus G (VSV-G) protein to the plasma membrane (Fig. 7C). These observations illustrate the occurrence of protein sorting at the TGN by generation of distinct types of vesicular carriers. Interaction of GGA1-containing carriers with endosomes. Upon arrival at the cell periphery, the fluorescent signal of most TGN-derived, GGA1-containing carriers was lost perhaps due to uncoating or movement outside the plane of focus. This prevented us from establishing the final destination of these carriers. Some GGA1-containing structures, however, were found to interact with endosomes labeled by internalization of rhodaminealbumin for 15 minutes. Fig. 8 shows an example of such interactions. In this example, a GFP-GGA1-coated carrier (green) and a rhodamine-albumincontaining endosome (red) converge to form a single hybrid organelle (yellow). The two markers remain associated with the same structure and move together for 51 seconds, although at times they appear segregated to different domains of the hybrid organelle. This structure eventually undergoes uneven fission into a vesicle that contains both GGA1 and albumin and another vesicle that contains 15
16 only GGA1. The latter finally disappears likely due to uncoating. We have observed variations on this phenomenon. Sometimes the GFP-GGA1 fluorescence is quickly lost after the two vesicles merge. Other times the GGA1- and albumin-containing vesicles interact transiently and then part ways, reminiscent of kiss-and-run interactions (Storrie and Desjardins, 1996). We interpret that these events represent the transfer of cargo from the TGN-derived carriers to the endosomal system. Dynamics of clathrin and GGA1 coats. The ability to visualize fluorescently-tagged clathrin and GGA1 in association with the TGN allowed us to examine the rates of exchange of these proteins between membranes and the cytosol using the technique of fluorescence recovery after photobleaching (Jacobson et al., 1976). This was performed by photobleaching a region of the TGN and quantifying the time course of fluorescence recovery in that region. We found that both GFP-clathrin (Fig. 9A) and GFP-GGA1 fluorescence (Fig. 9B) recovered with exponential kinetics and t 1/2 of ~20 s and 10 s, respectively (Fig. 9D). These rates were within the range previously reported for clathrin at the plasma membrane (Wu et al., 2001) and COPI on the Golgi complex (Presley et al., 2002). Hence, the association of all of these proteins coats with membranes is a highly dynamic process. The recovery appeared to occur on the same structures that were photobleached (Fig. 9C, arrows). 16
17 To assess whether membrane binding and release of GFP-clathrin and GFP-GGA1 were coupled to vesicle budding from the TGN, we examined the effect of lowering the temperature to 20 o C, a manipulation that blocks protein export from the TGN to various compartments (Griffiths et al., 1985; Xu and Shields, 1993; Wacker et al., 1997), including those of the endosomal-lysosomal system (Nishimura et al., 1990). We observed that budding of clathrin- and GGA1-containing intermediates completely ceased at this temperature (data not shown). Strikingly, the majority of GGA1 and clathrin fluorescence on the TGN still recovered at 20 o C, albeit more slowly (Fig. 9D). These results indicate that GGA1 and clathrin continue to undergo cycling on and off the TGN membrane even when vesicle budding from the TGN is inhibited. Thus, the association and dissociation of these TGN coat proteins with membranes is not obligatorily coupled to vesicle formation and budding. 17
18 DISCUSSION The results presented here demonstrate that clathrin, GGA1 and AP-1 are associated with a population of vesicular-tubular carriers budding from the TGN. These carriers appear larger and more pleiomorphic than conventional CCVs. After detaching from the TGN, the carriers move along microtubules for distances of up to ~10 µm towards the peripheral cytoplasm. The signal of the fluorescently-labeled coat proteins associated with the carriers disappears at different times after reaching the cell periphery, probably due to uncoating. Some coated carriers, however, persist and can be seen engaging in fusion or kiss-and-run interactions with peripheral endosomes before they lose their coat. Thus, it is likely that a function of these coated carriers is to move cargo between the TGN and peripheral endosomes. A cargo molecule transported by these intermediates is the CD-MPR, which interacts via acidic cluster-dileucine signals with the VHS domain of the GGAs (Puertollano et al., 2001a; Takatsu et al., 2001; Doray et al., 2002a). Indeed, we observed that the CD-MPR leaves the TGN on vesicular-tubular structures decorated with GGA1. Other transmembrane proteins such as the CI-MPR (Puertollano et al., 2001a; Zhu et al., 2001; Takatsu et al., 2001), sortilin (Nielsen et al., 2001), the low density lipoprotein related protein 3 (Takatsu et al., 2001), and β-secretase or BACE (He et al., 2002), also have acidic-cluster dileucine signals 18
19 that interact with the GGAs and might therefore be exported from the TGN on the same carriers. In contrast, recycling transferrin receptors and the VSV-G protein traffic in distinct sets of tubules and vesicles devoid of GGA1. The presence of both clathrin and GGA1 on the same carriers adds to the evidence that the GGAs function in association with clathrin (Puertollano et al., 2001b; Costaguta et al., 2001; Mullins and Bonifacino, 2001). The occurrence of AP-1 on these TGN-derived carriers, however, is intriguing because the exact role of AP-1 in sorting is currently a matter of debate. Our observations with GFP-γ1-adaptin agree with those of Huang et al. (Huang et al., 2001) obtained using YFP-µ1 to label the AP-1 complex. These authors reported that most YFP- µ1a-labeled vesicles also move from the TGN to the periphery of the cells. This sense of transport contrasts with the proposed role of AP-1 in recycling MPRs from endosomes to the TGN, inferred from the accumulation of MPRs in peripheral endosomes of µ1a-deficient fibroblasts (Meyer et al., 2000). A possible explanation for these seemingly contradictory observations could be that AP-1 actually mediates removal of cargo from the intermediates as they move towards the cell periphery or upon their merge with peripheral endosomes. AP-1 has in fact been shown to function in the removal of membrane proteins from immature secretory granules after their budding from the TGN (Klumperman et al., 1998). This would be analogous to the behavior of COPI, which is present on VTCs moving from ER exit sites to the Golgi complex (Presley et al., 1997), even though 19
20 it plays a role in protein recycling from the Golgi complex to the ER (Letourneur et al., 1994). Another possibility is that AP-1 plays a role in sorting at the TGN, as previously assumed. Recent work suggests that the GGAs and AP-1 do cooperate to package MPRs into TGN-derived intermediates (Doray et al., 2002b). The TGN-derived coated intermediates described here appear to belong to a growing family of large intracellular transport carriers including VTCs (vesicular tubular clusters) that mediate transport from the endoplasmic reticulum (ER) to the Golgi complex (Aridor et al., 1995; Presley et al., 1997) and PGCs (post-golgi carriers) involved in transport of VSV-G protein from the TGN to the plasma membrane (Hirschberg et al., 1998; Polishchuk et al., 2000). The large carriers described here are the first ones shown to contain associated clathrin and GGA1. The decoration of tubules carrying the CD-MPR with GGA1 appears to occur in discontinuous fashion, as if defining specific domains on the tubules. The CD-MPR was often more concentrated in the tubule domains containing associated GGA1, suggesting that the segregation of CD-MPR from other cargo molecules may persist after budding from the TGN. The vesicular-tubular carriers containing CD-MPR and even the individual foci labeled for clathrin, GGA1 and AP-1 were apparently larger than plasma membrane coated pits and conventional CCVs. This difference could be easily appreciated in microscopic fields where both types of clathrin-coated structures were visible (e.g., Fig. 3D). Although the intensity of the fluorescence signal can impinge upon estimations of size by optical microscopy, in the case of 20
21 clathrin it is reasonable to assume that the probability of GFP-labeled clathrin to be incorporated into clathrin lattices is the same in different parts of the cell. Thus, brighter clathrin-coated structures are also larger. Corroboration of the larger size of the TGN-derived intermediates was obtained by comparison to fluorescent beads of known size and similar brightness (Fig. 4). The larger size of the TGN-intermediates was not due to streaking since the speed of scanning (typically µm/s) was much higher than the speed of the intermediates (~1 µm/s). The fine structure of the TGN-derived carriers could not be resolved by fluorescence microscopy because of limits on the resolution by this technique (~200 nm under the conditions of our experiments, Inoue, 1989). We envision that they consist of tubular or irregularly-shaped membrane-bound organelles (akin to VTCs, Aridor et al., 1995 or PGCs, Polishchuk et al., 2000) with nm coated buds that define specific domains within these organelles. The apparently larger size of the coated foci relative to CCVs could be due to the presence of several nm coated buds on the carriers. It is also possible that the larger foci represent clusters of CCVs that are somehow tethered together. An important property of the TGN-associated coats studied here is that they are constantly cycling between membranes and the cytosol. Even when vesicle budding from the TGN is inhibited by incubation at 20 o C, the coats continue to exchange. Therefore, dissociation of the coats does not require 21
22 formation of vesicular intermediates. In this regard, the clathrin- GGA- and AP- 1-containing TGN coats behave like plasma membrane clathrin coats (Wu et al., 2001) and COPI (Presley et al., 2002), which also cycle on and off membranes when vesicle budding is inhibited. We were unable to examine the dynamics of coats on the TGN-derived intermediates themselves because of their mobility. In any event, whether dynamically or statically, the intermediates do retain their coats until they reach the periphery. The coats could thus be directly responsible for the recruitment of motor molecules that effect tracking along microtubules. The kinesin superfamily protein KIF13A, for example, is a plus-end microtubule motor that interacts with the β1-adaptin subunit of AP-1 (Nakagawa et al., 2000). The coats could also bind tethering proteins that allow interactions or fusion of the intermediates with endosomes. For instance, rabaptin-5 is an effector of the early endosomal rab4 and rab5 GTP-binding proteins (Stenmark et al., 1995; Vitale et al., 1998) that also interacts with the γ1-adaptin subunit of AP-1 (Shiba et al., 2002). The persistence of the coats on the intermediates may thus enable their involvement in organelle targeting events, in addition to their roles in vesicle formation and cargo selection. Of course, our observations do not rule the existence of small CCVs that could mediate short-range transfer of cargo to the larger intermediates or to endosomes located in the vicinity of the TGN. In fact, the endosomal recycling compartment (Yamashiro et al., 1984) and late endosomes (Rabinowitz et al., 22
23 1992) are mostly concentrated in the juxtanuclear area of the cell. It would be virtually impossible to observe direct transfer of materials from the TGN to these compartments. Small CCVs could also pinch off from the large intermediates as they move towards the peripheral cytoplasm. Finally, small CCVs could be too weakly labeled or transient for detection. Nevertheless, our findings do reveal the existence of a previously unrecognized type of intermediate containing associated clathrin coats, which could be involved in long-range delivery of biosynthetic cargo to peripheral cellular locations. 23
24 ACKNOWLEDGEMENTS We thank Jennifer Lippincott-Schwartz for helpful discussions and Margaret S. Robinson for kind gifts of reagents. 24
25 REFERENCES Aridor, M., Bannykh, S. I., Rowe, T., and Balch, W. E. (1995). Sequential coupling between copii and copi vesicle coats in endoplasmic reticulum to Golgi transport. J. Cell Biol. 131, Boman, A. L. (2001). GGA proteins: New players in the sorting game. J. Cell Sci. 114, Brodsky, F. M., Chen, C. Y., Knuehl, C., Towler, M. C., and Wakeham, D. E. (2001). Biological basket weaving: Formation and function of clathrin-coated vesicles. Annu. Rev. Cell Dev. Biol. 17, Costaguta, G., Stefan, C. J., Bensen, E. S., Emr, S. D., and Payne, G. S. (2001). Yeast gga coat proteins function with clathrin in Golgi to endosome transport. Mol Biol Cell 12, Damer, C. K., and O'Halloran, T. J. (2000). Spatially regulated recruitment of clathrin to the plasma membrane during capping and cell translocation. Mol. Biol. Cell 11, Dell'Angelica, E. C., Puertollano, R., Mullins, C., Aguilar, R. C., Vargas, J. D., Hartnell, L. M., and Bonifacino, J. S. (2000). Ggas: A family of ADP ribosylation 25
26 factor-binding proteins related to adaptors and associated with the Golgi complex. J. Cell Biol. 149, Doray, B., Bruns, K., Ghosh, P., and Kornfeld, S. (2002a). Interaction of the cationdependent mannose 6-phosphate receptor with GGA proteins. J Biol Chem 277, Doray, B., Ghosh, P., Griffith, J., Geuze, H. J., and Kornfeld, S. (2002b). Cooperation of ggas and ap-1 in packaging man-6-p receptors at the trans-golgi network. Science in press. Gaidarov, I., Santini, F., Warren, R. A., and Keen, J. H. (1999). Spatial control of coated-pit dynamics in living cells. Nat. Cell Biol. 1, 1-7. Griffiths, G., Pfeiffer, S., Simons, K., and Matlin, K. (1985). Exit of newly synthesized membrane proteins from the trans cisternae of the Golgi complex to the plasma membrane. J. Cell Biol. 101, He, X., Chang, W. P., Koelsch, G., and Tang, J. (2002). Memapsin 2 (betasecretase) cytosolic domain binds to the VHS domains of GGA1 and GGA2: Implications on the endocytosis mechanism of memapsin 2. FEBS Lett 524, Hirschberg, K., Miller, C. M., Ellenberg, J., Presley, J. F., Siggia, E. D., Phair, R. D., and Lippincott-Schwartz, J. (1998). Kinetic analysis of secretory protein traffic 26
27 and characterization of golgi to plasma membrane transport intermediates in living cells. J. Cell Biol. 143, Hirst, J., Lindsay, M. R., and Robinson, M. S. (2001). Ggas: Roles of the different domains and comparison with ap-1 and clathrin. Mol. Biol. Cell 12, Huang, F., Nesterov, A., Carter, R. E., and Sorkin, A. (2001). Trafficking of yellow-fluorescent-protein-tagged mu1 subunit of clathrin adaptor ap-1 complex in living cells. Traffic 2, Inoue, S. (1989). Imaging of unresolved objects, superresolution, and precision of distance measurement with video microscopy. Methods Cell Biol. 30, Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., and Poste, G. (1976). Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5, 428. Kirchhausen, T. (2000). Clathrin. Annu. Rev. Biochem. 69, Klumperman, J., Kuliawat, R., Griffith, J. M., Geuze, H. J., and Arvan, P. (1998). Mannose 6-phosphate receptors are sorted from immature secretory granules via adaptor protein ap-1, clathrin, and syntaxin 6-positive vesicles. J. Cell Biol. 141,
28 Letourneur, F., Gaynor, E. C., Hennecke, S., Demolliere, C., Duden, R., Emr, S. D., Riezman, H., and Cosson, P. (1994). Coatomer is essential for retrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell 79, Meyer, C., Zizioli, D., Lausmann, S., Eskelinen, E. L., Hamann, J., Saftig, P., von Figura, K., and Schu, P. (2000). Mu1a-adaptin-deficient mice: Lethality, loss of ap-1 binding and rerouting of mannose 6-phosphate receptors. EMBO J. 19, Mullins, C., and Bonifacino, J. S. (2001). Structural requirements for function of yeast ggas in vacuolar protein sorting, alpha-factor maturation, and interactions with clathrin. Mol. Cell. Biol. 21, Nakagawa, T., Setou, M., Seog, D., Ogasawara, K., Dohmae, N., Takio, K., and Hirokawa, N. (2000). A novel motor, kif13a, transports mannose-6-phosphate receptor to plasma membrane through direct interaction with ap-1 complex. Cell 103, Nielsen, M. S., Madsen, P., Christensen, E. I., Nykjaer, A., Gliemann, J., Kasper, D., Pohlmann, R., and Petersen, C. M. (2001). The sortilin cytoplasmic tail conveys Golgi-endosome transport and binds the VHS domain of the GGA2 sorting protein. EMBO J. 20, Nishimura, Y., Kawabata, T., Yano, S., and Kato, K. (1990). Inhibition of intracellular sorting and processing of lysosomal cathepsins h and l at reduced 28
29 temperature in primary cultures of rat hepatocytes. Arch. Biochem. Biophys. 283, Peters, P. J. (1998). Cryo-immunogold electron microscopy. In Current protocols in cell biology, J. S. Bonifacino, M. Dasso, J. B. Harford, J. Lippincott-Schwartz, and K. Yamada, eds. (New York, John Wiley & Sons), pp Polishchuk, R. S., Polishchuk, E. V., Marra, P., Alberti, S., Buccione, R., Luini, A., and Mironov, A. A. (2000). Correlative light-electron microscopy reveals the tubular-saccular ultrastructure of carriers operating between Golgi apparatus and plasma membrane. J Cell Biol 148, Porcelli, S., Morita, C. T., and Brenner, M. B. (1992). Cd1b restricts the response of human CD4-8- t lymphocytes to a microbial antigen. Nature 360, Presley, J. F., Cole, N. B., Schroer, T. A., Hirschberg, K., Zaal, K. J., and Lippincott-Schwartz, J. (1997). Er-to-Golgi transport visualized in living cells. Nature 389, Presley, J. F., Ward, T. H., Pfeiffer, A., Siggia, E. D., Phair, R. D., and Lippincott- Schwartz, J. (2002). Dissection of copi and arf1 dynamics in vivo and role in Golgi membrane transport. Nature 417,
30 Puertollano, R., Aguilar, R. C., Gorshkova, I., Crouch, R. J., and Bonifacino, J. S. (2001a). Sorting of mannose 6-phosphate receptors mediated by the ggas. Science 292, Puertollano, R., Randazzo, P., Hartnell, L. M., Presley, J., and Bonifacino, J. S. (2001b). The ggas promote ARF-dependent recruitment of clathrin to the TGN. Cell 105, Rabinowitz, S., Horstmann, H., Gordon, S., and Griffiths, G. (1992). Immunocytochemical characterization of the endocytic and phagolysosomal compartments in peritoneal macrophages. J. Cell Biol. 116, Robinson, M. S., and Bonifacino, J. S. (2001). Adaptor-related proteins. Curr. Opin. Cell Biol. 13, Shiba, Y., Takatsu, H., Shin, H. W., and Nakayama, K. (2002). Gamma-adaptin interacts directly with rabaptin-5 through its ear domain. J. Biochem. (Tokyo) 131, Stenmark, H., Vitale, G., Ullrich, O., and Zerial, M. (1995). Rabaptin-5 is a direct effector of the small gtpase rab5 in endocytic membrane fusion. Cell 83, Storrie, B., and Desjardins, M. (1996). The biogenesis of lysosomes: Is it a kiss and run, continuous fusion and fission process? Bioessays 18,
31 Takatsu, H., Katoh, Y., Shiba, Y., and Nakayama, K. (2001). Golgi-localizing, gamma-adaptin ear homology domain, ADP-ribosylation factor-binding (GGA) proteins interact with acidic dileucine sequences within the cytoplasmic domains of sorting receptors through their vps27p/hrs/stam (VHS) domains. J. Biol. Chem. 276, Vitale, G., Rybin, V., Christoforidis, S., Thornqvist, P., McCaffrey, M., Stenmark, H., and Zerial, M. (1998). Distinct rab-binding domains mediate the interaction of rabaptin-5 with GTP-bound rab4 and rab5. EMBO J. 17, Wacker, I., Kaether, C., Kromer, A., Migala, A., Almers, W., and Gerdes, H. H. (1997). Microtubule-dependent transport of secretory vesicles visualized in real time with a GFP-tagged secretory protein. J Cell Sci 110, Wu, X., Zhao, X., Baylor, L., Kaushal, S., Eisenberg, E., and Greene, L. E. (2001). Clathrin exchange during clathrin-mediated endocytosis. J. Cell Biol. 155, Xu, H., and Shields, D. (1993). Prohormone processing in the trans-golgi network: Endoproteolytic cleavage of prosomatostatin and formation of nascent secretory vesicles in permeabilized cells. J. Cell Biol. 122, Yamashiro, D. J., Tycko, B., Fluss, S. R., and Maxfield, F. R. (1984). Segregation of transferrin to a mildly acidic (ph 6.5) para-golgi compartment in the recycling pathway. Cell 37,
32 Zhu, Y., Doray, B., Poussu, A., Lehto, V. P., and Kornfeld, S. (2001). Binding of GGA2 to the lysosomal enzyme sorting motif of the mannose 6- phosphate receptor. Science 292,
33 LEGENDS TO FIGURES Figure 1. Intracellular distribution of Myc-GGA1 in MDCK cells analyzed by immunofluorescence microscopy. MDCK cells stably expressing Myc-GGA1 were fixed, permeabilized, and double-labeled by incubation with antibodies to GGA1 (A, red channel), the Myc epitope (D, red channel; G, green channel), clathrin (B, green channel), the γ1-adaptin subunit of AP-1 (E, green channel), and the β3a subunit of the AP-3 complex (H, red channel), as indicated in the figure, followed by the corresponding fluorescently-conjugated secondary antibodies. Stained cells were examined by confocal fluorescence microscopy. Merging the images in the red and green channels generated the third picture on each row (C, F, and I); yellow indicates overlapping localization. Insets show two-fold-magnified views of peripheral cytoplasmic regions. Bar, 10 µm. Figure 2. Immunoelectron microscopy localization of GGA1, Myc-GGA1, clathrin and AP-1. (A) The localization of endogenous GGA1 in monocyte-derived dendritic cells was analyzed by cryo-immunogold electron microscopy using antibodies to GGA1 (10 nm gold). (B-D) MDCK cells stably expressing Myc- GGA1 were analyzed by cryo-immunogold electron microscopy using antibodies to: (B) the Myc epitope (15 nm gold) and clathrin (10 nm gold), (C, D) GGA1 (15 nm gold) and the γ1 subunit of AP-1 (10 nm gold). Open arrowheads in B 33
34 indicate coated buds containing both Myc-GGA1 and clathrin, whereas closed arrowheads point to 15 nm gold particles marking the location of Myc-GGA1. Open arrowheads in C-D point to coated buds containing both Myc-GGA1 (or GGA1) and AP-1, whereas closed arrowheads point to 15 nm gold particles marking the location of Myc-GGA1 (or GGA1). G, Golgi complex; n, nucleus; p, plasma membrane; m, mitochondrion. Bars, 200 nm. Figure 3. Characterization of TGN-derived carriers containing clathrin, GGA1, and AP-1. MDCK cells were grown on LabTek chambers and transiently transfected with constructs encoding GFP-clathrin light chain b (A, D), GFP- GGA1 (B), or GFP-γ1-adaptin (GFP-AP-1, C). At 15 h after transfection, cells expressing moderate levels of the fluorescent proteins were analyzed by timelapse microscopy and images acquired at the indicated intervals. Insets in panel D show two-fold-magnified comparisons of clathrin-containing carriers budding from the TGN with clathrin-coated pits at the plasma membrane. Arrows point to pleiomorphic carriers budding from the TGN and moving towards the cell periphery while arrowheads indicate immobile structures. N: Nucleus. Bars: 5 µm (panels A, B, C); 4 µm (panel D); 1 µm (panel D insets). Quicktime videos of the experiments shown in panels A, B, and C can be seen in Supplemental Materials (videos 1, 2 and 3, respectively). 34
35 Figure 4. Comparison of the size of GGA1-coated intermediaries to that of fluorescent beads. MDCK cells expressing YFP-GGA1 were incubated with 0.2 µm diameter fluorescent beads prior to analysis by time-lapse confocal microscopy. Arrow in panel A points to a YFP-GGA1-containing intermediate moving from the TGN toward the cell periphery. Notice that this carrier appears larger (~0.35 µm) than the beads shown in the inset (~0.2 µm) even though the three structures have similar fluorescent intensities (37.7 and 38.6 arbitrary units respectively). In some cases, large intermediates appear to arise from clustering of smaller vesicles, as shown in panel B (fluorescent bead is shown in the inset). Bars, 0.5 µm (A), 1 µm (B). Figure 5. Presence of clathrin and AP-1 on GGA1-containing carriers. MDCK cells were simultaneously transfected with constructs encoding YFP-clathrin and CFP-GGA1 (A) or GFP-γ1 (GFP-AP-1) and CFP-GGA1 (B). Confocal microscopy images were acquired every 3 s. Arrows indicate vesicular carriers budding from the TGN while arrowheads point to immobile structures. The yellow color in the merged images indicates co-localization between clathrin and GGA1 (panel A) or GGA1 and AP-1 (panel B). Bars, 2 µm. Figure 6. Cargo specificity of GGA1-containing intermediates. (A). Confocal microscopy imaging of live MDCK cells co-expressing CD-MPR-CFP and YFP- 35
36 GGA1. Images were collected at 6-sec intervals. Arrows indicate the formation and detachment of long tubular structures containing CD-MPR-CFP from the TGN. Notice that YFP-GGA1 associates with those tubular structures in a discontinuous pattern. (B) MDCK cells expressing GFP-GGA1 were incubated with rhodamine-transferrin for 20 minutes to load recycling endosomes and then examined by video microscopy. Images show that transferrin-containing intermediates in transit from the juxtanuclear area of the cell are devoid of GFP- GGA1. Bars, 3 µm. Figure 7. Exclusion of galactosyl transferase and VSV-G protein from GGA1 carriers budding from the TGN. (A, B). The Golgi-resident protein, galactosyl transferase (GT), is not incorporated into vesicular carriers budding from the TGN. MDCK cells were doubly-transfected with a trans-golgi resident protein (CFP-GT) and with YFP-GGA1 (A) or GFP-γ1 (GFP-AP-1, B). Time-lapse microscopy showed the absence of CFP-GT (red channel) from rows of coated intermediates budding from the TGN (green channel, arrows). (C). VSV-G and GGA-1 localize to different transport intermediaries. MDCK cells, transfected with YFP-GGA1 and CFP-VSV-G (ts045 mutant) were incubated for 24 h at 40 C, shifted to 20 C for 1 h to accumulate cargo in the TGN, and then warmed to 32 C prior to visualization. Consecutive video images from time-lapse series show a VSV-G-enriched vesicle budding from the TGN (arrow, red channel). Note the 36
37 absence of GGA1 (green channel) in this structure. The arrowhead points to a row of YFP-GGA1-containing vesicles rapidly budding from the TGN. N: nucleus. Bars, 10 µm (A and B), 5 µm (C). Figure 8. Interaction of GGA1-containing carriers with endosomes. MDCK cells were transfected with GFP-GGA1. At 15 h after transfection, the cells were loaded with rhodamine-albumin for 15 min prior to analysis by time-lapse confocal microscopy. Merged images for GFP-GGA1 (green) and rhodaminealbumin (red) are shown. Arrows point to vesicular structures containing rhodamine-albumin and GFP-GGA1. The insets show four-fold magnification of the structures pointed by the arrows. Bars, 2 µm and 0.5 µm (inset). Figure 9. Dynamics of clathrin and GGA1 analyzed by fluorescence recovery after photobleaching. (A-C) MDCK cells were transiently transfected with constructs encoding GFP-clathrin (A) or GFP-GGA1 (B, C). Fluorescence associated with the Golgi complex was photobleached with high-intensity laser light and subsequent recovery of fluorescence was monitored by scanning the whole cell at low laser power at 5 s intervals for 5 min. The arrows in panel C indicate that the GFP-GGA1 recovers on the same TGN structures where it was located prior to photobleaching. N: nucleus. Bars, 10 µm (A and B), 5 µm (C). (D) Kinetics of GFP-clathrin and GFP-GGA1 recovery after photobleaching at 37 C 37
38 and 20 C. MDCK cells expressing GFP-clathrin or GFP-GGA1 were photobleached and then scanned at low laser power for five minutes. The graph shows the combined results of 10 determinations per condition. Quicktime videos of the experiments shown in panes A, B, and C can be seen in Supplemental Materials (videos A, B, and C). 38
39 Table 1. Quantification of the size of foci containing fluorescently-labeled clathrin and GGA1. MDCK cells expressing GFP-fusion proteins were analyzed for the size of moving foci emanating from the TGN (GFP-clathrin and GFP- GGA1) and at plasma membrane (GFP-clathrin). Sizes were estimated from fluorescence microscopy images. Size values are the mean ± SD of the number of determinations (n) indicated in the Table. GFP Fusion Proteins Size (nm) Number GFP-clathrin 360 ± 60 n= 30 (moving from the TGN area) GFP-clathrin 180 ± 40 n= 30 (plasma membrane) GFP-GGA1 380 ± 50 n= 24 (moving from the TGN area) 39

40


41 A G D p n B G m G C G G

42

43
44

45

46

47

48
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic
 Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
Molecular Cell Biology - Problem Drill 17: Intracellular Vesicular Traffic Question No. 1 of 10 1. Which of the following statements about clathrin-coated vesicles is correct? Question #1 (A) There are
Vesicle Transport. Vesicle pathway: many compartments, interconnected by trafficking routes 3/17/14
 Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
Vesicle Transport Vesicle Formation Curvature (Self Assembly of Coat complex) Sorting (Sorting Complex formation) Regulation (Sar1/Arf1 GTPases) Fission () Membrane Fusion SNARE combinations Tethers Regulation
endomembrane system internal membranes origins transport of proteins chapter 15 endomembrane system
 endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
endo system chapter 15 internal s endo system functions as a coordinated unit divide cytoplasm into distinct compartments controls exocytosis and endocytosis movement of molecules which cannot pass through
Homework Hanson section MCB Course, Fall 2014
 Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
Homework Hanson section MCB Course, Fall 2014 (1) Antitrypsin, which inhibits certain proteases, is normally secreted into the bloodstream by liver cells. Antitrypsin is absent from the bloodstream of
Intracellular Vesicular Traffic Chapter 13, Alberts et al.
 Intracellular Vesicular Traffic Chapter 13, Alberts et al. The endocytic and biosynthetic-secretory pathways The intracellular compartments of the eucaryotic ell involved in the biosynthetic-secretory
Intracellular Vesicular Traffic Chapter 13, Alberts et al. The endocytic and biosynthetic-secretory pathways The intracellular compartments of the eucaryotic ell involved in the biosynthetic-secretory
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson
 Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
Lysosomes and endocytic pathways 9/27/2012 Phyllis Hanson General principles Properties of lysosomes Delivery of enzymes to lysosomes Endocytic uptake clathrin, others Endocytic pathways recycling vs.
04_polarity. The formation of synaptic vesicles
 Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Mechanism of Vesicular Transport
 Mechanism of Vesicular Transport Transport vesicles play a central role in the traffic of molecules between different membrane-enclosed enclosed compartments. The selectivity of such transport is therefore
Mechanism of Vesicular Transport Transport vesicles play a central role in the traffic of molecules between different membrane-enclosed enclosed compartments. The selectivity of such transport is therefore
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation
 Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 2/21/17 glycosylation is a non-template derived phenomenon - the presence of
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 2/21/17 glycosylation is a non-template derived phenomenon - the presence of
Protein Trafficking in the Secretory and Endocytic Pathways
 Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Chapter 1: Vesicular traffic. Biochimica cellulare parte B 2017/18
 Chapter 1: Vesicular traffic Biochimica cellulare parte B 2017/18 Major Protein-sorting pathways in eukaryotic cells Secretory and endocytic pathways Unifying principle governs all protein trafficking
Chapter 1: Vesicular traffic Biochimica cellulare parte B 2017/18 Major Protein-sorting pathways in eukaryotic cells Secretory and endocytic pathways Unifying principle governs all protein trafficking
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation
 Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 3/8/2011 glycosylation is a non-template derived phenomenon - the presence of
Localization and Retention of Glycosyltransferases And the Role of Vesicle Trafficking in Glycosylation Richard Steet, Ph.D. 3/8/2011 glycosylation is a non-template derived phenomenon - the presence of
Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Maintenance of Golgi structure and function depends on the integrity of ER export
 JCB Article Published November 12, 2001 Maintenance of Golgi structure and function depends on the integrity of ER export Theresa H. Ward, Roman S. Polishchuk, Steve Caplan, Koret Hirschberg, and Jennifer
JCB Article Published November 12, 2001 Maintenance of Golgi structure and function depends on the integrity of ER export Theresa H. Ward, Roman S. Polishchuk, Steve Caplan, Koret Hirschberg, and Jennifer
Lecture 6 - Intracellular compartments and transport I
 01.25.10 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
01.25.10 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
1. endoplasmic reticulum This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Intracellular vesicular traffic. B. Balen
 Intracellular vesicular traffic B. Balen Three types of transport in eukaryotic cells Figure 12-6 Molecular Biology of the Cell ( Garland Science 2008) Endoplasmic reticulum in all eucaryotic cells Endoplasmic
Intracellular vesicular traffic B. Balen Three types of transport in eukaryotic cells Figure 12-6 Molecular Biology of the Cell ( Garland Science 2008) Endoplasmic reticulum in all eucaryotic cells Endoplasmic
Molecular Trafficking
 SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
SCBM 251 Molecular Trafficking Assoc. Prof. Rutaiwan Tohtong Department of Biochemistry Faculty of Science rutaiwan.toh@mahidol.ac.th Lecture outline 1. What is molecular trafficking? Why is it important?
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human
 and HeLa (bottom) cells expressing for 24 h human") Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
1. This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Appendix: Subcellular localization of MIG-14::GFP in C. elegans body wall muscle cells. Pei-Tzu Yang and Hendrik C. Korswagen
 Appendix: Subcellular localization of MIG-14::GFP in C. elegans body wall muscle cells Pei-Tzu Yang and Hendrik C. Korswagen 129 SUBCELLULAR LOCALIZATION OF MIG-14 Abstract MIG-14/Wls is a Wnt binding
Appendix: Subcellular localization of MIG-14::GFP in C. elegans body wall muscle cells Pei-Tzu Yang and Hendrik C. Korswagen 129 SUBCELLULAR LOCALIZATION OF MIG-14 Abstract MIG-14/Wls is a Wnt binding
Summary of Endomembrane-system
 Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Summary of Endomembrane-system 1. Endomembrane System: The structural and functional relationship organelles including ER,Golgi complex, lysosome, endosomes, secretory vesicles. 2. Membrane-bound structures
Lab module 6a Receptor-mediated endocytosis
 Goal for the module Lab module 6a Receptor-mediated endocytosis To follow cell surface receptors as they are internalized. Pre-lab homework Read about receptor-mediated endocytosis and other forms of internalization
Goal for the module Lab module 6a Receptor-mediated endocytosis To follow cell surface receptors as they are internalized. Pre-lab homework Read about receptor-mediated endocytosis and other forms of internalization
Practice Exam 2 MCBII
 1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
1. Which feature is true for signal sequences and for stop transfer transmembrane domains (4 pts)? A. They are both 20 hydrophobic amino acids long. B. They are both found at the N-terminus of the protein.
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/7/334/rs4/dc1 Supplementary Materials for Rapidly rendering cells phagocytic through a cell surface display technique and concurrent Rac activation Hiroki Onuma,
www.sciencesignaling.org/cgi/content/full/7/334/rs4/dc1 Supplementary Materials for Rapidly rendering cells phagocytic through a cell surface display technique and concurrent Rac activation Hiroki Onuma,
Endocytosis and Intracellular Trafficking of Notch and Its Ligands
 CHA P T E R F IVE Endocytosis and Intracellular Trafficking of Notch and Its Ligands Shinya Yamamoto, *,1 Wu-Lin Charng, *,1 and Hugo J. Bellen *,,, Contents 1. Notch Signaling and its Regulation by Endocytosis
CHA P T E R F IVE Endocytosis and Intracellular Trafficking of Notch and Its Ligands Shinya Yamamoto, *,1 Wu-Lin Charng, *,1 and Hugo J. Bellen *,,, Contents 1. Notch Signaling and its Regulation by Endocytosis
In the previous chapter we explored how proteins are targeted
 17 VESICULAR TRAFFIC, SECRETION, AND ENDOCYTOSIS Electron micrograph of clathrin cages, like those that surround clathrin-coated transport vesicles, formed by the in vitro polymerization of clathrin heavy
17 VESICULAR TRAFFIC, SECRETION, AND ENDOCYTOSIS Electron micrograph of clathrin cages, like those that surround clathrin-coated transport vesicles, formed by the in vitro polymerization of clathrin heavy
Visualization of TGN to Endosome Trafficking through Fluorescently Labeled MPR and AP-1 in Living Cells V
 Molecular Biology of the Cell Vol. 14, 142 155, January 2003 Visualization of TGN to Endosome Trafficking through Fluorescently Labeled MPR and AP-1 in Living Cells V Satoshi Waguri,* Frédérique Dewitte,*
Molecular Biology of the Cell Vol. 14, 142 155, January 2003 Visualization of TGN to Endosome Trafficking through Fluorescently Labeled MPR and AP-1 in Living Cells V Satoshi Waguri,* Frédérique Dewitte,*
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Beck et al., http://www.jcb.org/cgi/content/full/jcb.201011027/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Membrane binding of His-tagged proteins to Ni-liposomes.
Supplemental material Beck et al., http://www.jcb.org/cgi/content/full/jcb.201011027/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Membrane binding of His-tagged proteins to Ni-liposomes.
Chapter 3. Expression of α5-megfp in Mouse Cortical Neurons. on the β subunit. Signal sequences in the M3-M4 loop of β nachrs bind protein factors to
 22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS
 Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
ab CytoPainter Golgi/ER Staining Kit
 ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering
 nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering Toru Komatsu, Igor Kukelyansky, J Michael McCaffery, Tasuku Ueno, Lidenys C Varela & Takanari Inoue Supplementary
nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering Toru Komatsu, Igor Kukelyansky, J Michael McCaffery, Tasuku Ueno, Lidenys C Varela & Takanari Inoue Supplementary
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
Module 3 Lecture 7 Endocytosis and Exocytosis
 Module 3 Lecture 7 Endocytosis and Exocytosis Endocytosis: Endocytosis is the process by which cells absorb larger molecules and particles from the surrounding by engulfing them. It is used by most of
Module 3 Lecture 7 Endocytosis and Exocytosis Endocytosis: Endocytosis is the process by which cells absorb larger molecules and particles from the surrounding by engulfing them. It is used by most of
Advanced Cell Biology. Lecture 33
 Advanced Cell Biology. Lecture 33 Alexey Shipunov Minot State University April 22, 2013 Shipunov (MSU) Advanced Cell Biology. Lecture 33 April 22, 2013 1 / 38 Outline Questions and answers Intracellular
Advanced Cell Biology. Lecture 33 Alexey Shipunov Minot State University April 22, 2013 Shipunov (MSU) Advanced Cell Biology. Lecture 33 April 22, 2013 1 / 38 Outline Questions and answers Intracellular
Legionella pneumophila: an intracellular pathogen of phagocytes Prof. Craig Roy
 an intracellular pathogen of phagocytes Section of Microbial Pathogenesis, Yale University School of Medicine 1 Legionella pneumophila Gram-negative bacterium Facultative intracellular pathogen Protozoa
an intracellular pathogen of phagocytes Section of Microbial Pathogenesis, Yale University School of Medicine 1 Legionella pneumophila Gram-negative bacterium Facultative intracellular pathogen Protozoa
Adaptor and Clathrin Exchange at the Plasma Membrane and trans-golgi Network
 Molecular Biology of the Cell Vol. 14, 516 528, February 2003 Adaptor and Clathrin Exchange at the Plasma Membrane and trans-golgi Network Xufeng Wu,* Xiaohong Zhao,* Rosa Puertollano, Juan S. Bonifacino,
Molecular Biology of the Cell Vol. 14, 516 528, February 2003 Adaptor and Clathrin Exchange at the Plasma Membrane and trans-golgi Network Xufeng Wu,* Xiaohong Zhao,* Rosa Puertollano, Juan S. Bonifacino,
/07/$15.00/0 Molecular Endocrinology 21(12): Printed in U.S.A.
: Printed in U.S.A.") 0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Chapter 13: Vesicular Traffic
 Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
Chapter 13: Vesicular Traffic Know the terminology: ER, Golgi, vesicle, clathrin, COP-I, COP-II, BiP, glycosylation, KDEL, microtubule, SNAREs, dynamin, mannose-6-phosphate, M6P receptor, endocytosis,
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001
 BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
BIOL 4374/BCHS 4313 Cell Biology Exam #2 March 22, 2001 SS# Name This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses. Good luck! 1. (2) In the
Intracellular Compartments and Protein Sorting
 Intracellular Compartments and Protein Sorting Intracellular Compartments A eukaryotic cell is elaborately subdivided into functionally distinct, membrane-enclosed compartments. Each compartment, or organelle,
Intracellular Compartments and Protein Sorting Intracellular Compartments A eukaryotic cell is elaborately subdivided into functionally distinct, membrane-enclosed compartments. Each compartment, or organelle,
Downloaded from:
 Nichols, BJ; Kenworthy, AK; Polishchuk, RS; Lodge, R; Roberts, TH; Hirschberg, K; Phair, RD; Lippincott-Schwartz, J (2001) Rapid cycling of lipid raft markers between the cell surface and Golgi complex.
Nichols, BJ; Kenworthy, AK; Polishchuk, RS; Lodge, R; Roberts, TH; Hirschberg, K; Phair, RD; Lippincott-Schwartz, J (2001) Rapid cycling of lipid raft markers between the cell surface and Golgi complex.
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
Overview of clathrin-mediated endocytosis
 Overview of clathrin-mediated endocytosis Accessory and adaptor proteins promote clathrin nucleation on the plasma membrane and some help deform membrane. Clathrin assembly into lattices stabilize the
Overview of clathrin-mediated endocytosis Accessory and adaptor proteins promote clathrin nucleation on the plasma membrane and some help deform membrane. Clathrin assembly into lattices stabilize the
Summary and Discussion antigen presentation
 Summary and Discussion antigen presentation 247 248 Summary & Discussion Summary and discussion: antigen presentation For a cell to communicate information about its internal health and status to the immune
Summary and Discussion antigen presentation 247 248 Summary & Discussion Summary and discussion: antigen presentation For a cell to communicate information about its internal health and status to the immune
Supplemental information contains 7 movies and 4 supplemental Figures
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
ArfGAP1 dynamics and its role in COPI coat assembly on Golgi membranes of living cells
 JCB: ARTICLE ArfGAP1 dynamics and its role in COPI coat assembly on Golgi membranes of living cells Wei Liu, 1 Rainer Duden, 3 Robert D. Phair, 2 and Jennifer Lippincott-Schwartz 1 1 Cell Biology and Metabolism
JCB: ARTICLE ArfGAP1 dynamics and its role in COPI coat assembly on Golgi membranes of living cells Wei Liu, 1 Rainer Duden, 3 Robert D. Phair, 2 and Jennifer Lippincott-Schwartz 1 1 Cell Biology and Metabolism
CELL BIOLOGY - CLUTCH CH INTRACELLULAR PROTEIN TRANSPORT.
 !! www.clutchprep.com CONCEPT: MEMBRANE ENCLOSED ORGANELLES Table of eukaryotic organelles and their functions Organelle Function % volume of cell Cytosol Aqueous fluid where metabolic pathways and chemical
!! www.clutchprep.com CONCEPT: MEMBRANE ENCLOSED ORGANELLES Table of eukaryotic organelles and their functions Organelle Function % volume of cell Cytosol Aqueous fluid where metabolic pathways and chemical
17/01/2017. Protein trafficking between cell compartments. Lecture 3: The cytosol. The mitochondrion - the power plant of the cell
 ell biology 2017 version 13/1 2017 ote endosome vs lysosome handout Lecture 3: Text book Alberts et al.: hapter 12-14 (Topics covered by the lecture) A lot of reading! Focus on principles ell Biology interactive
ell biology 2017 version 13/1 2017 ote endosome vs lysosome handout Lecture 3: Text book Alberts et al.: hapter 12-14 (Topics covered by the lecture) A lot of reading! Focus on principles ell Biology interactive
GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
 MBC in Press, published on October 31, 2003 as 10.1091/mbc.E03-07-0517 GLUT4 retention by intracellular cycling GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
MBC in Press, published on October 31, 2003 as 10.1091/mbc.E03-07-0517 GLUT4 retention by intracellular cycling GLUT4 is retained by an intracellular cycle of vesicle formation and fusion with endosomes.
Dynamic Interaction of HIV-1 Nef with the Clathrin-Mediated Endocytic Pathway at the Plasma Membrane
 Traffic 2007; 8: 61 76 Blackwell Munksgaard Dynamic Interaction of HIV-1 Nef with the Clathrin-Mediated Endocytic Pathway at the Plasma Membrane # 2006 The Authors Journal compilation # 2006 Blackwell
Traffic 2007; 8: 61 76 Blackwell Munksgaard Dynamic Interaction of HIV-1 Nef with the Clathrin-Mediated Endocytic Pathway at the Plasma Membrane # 2006 The Authors Journal compilation # 2006 Blackwell
PROTEIN TRAFFICKING. Dr. SARRAY Sameh, Ph.D
 PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
PROTEIN TRAFFICKING Dr. SARRAY Sameh, Ph.D Overview Proteins are synthesized either on free ribosomes or on ribosomes bound to endoplasmic reticulum (RER). The synthesis of nuclear, mitochondrial and peroxisomal
LDLR-related protein 10 (LRP10) regulates amyloid precursor protein (APP) trafficking and processing: evidence for a role in Alzheimer s disease
 regulates amyloid precursor protein (APP) trafficking and processing: evidence for a role in Alzheimer s disease") Brodeur et al. Molecular Neurodegeneration 2012, 7:31 RESEARCH ARTICLE Open Access LDLR-related protein 10 (LRP10) regulates amyloid precursor protein (APP) trafficking and processing: evidence for a role
Brodeur et al. Molecular Neurodegeneration 2012, 7:31 RESEARCH ARTICLE Open Access LDLR-related protein 10 (LRP10) regulates amyloid precursor protein (APP) trafficking and processing: evidence for a role
Molecular Cell Biology 5068 In class Exam 1 October 2, Please print your name: Instructions:
 Molecular Cell Biology 5068 In class Exam 1 October 2, 2012 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your number
Molecular Cell Biology 5068 In class Exam 1 October 2, 2012 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your number
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest.
 Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Thursday, October 16 th
 Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Subcellular biochemistry
 Department of Medical Biochemistry Semmelweis University Subcellular biochemistry February-March 2017 Subcellular biochemistry (biochemical aspects of cell biology) Miklós Csala Semmelweis University Dept.
Department of Medical Biochemistry Semmelweis University Subcellular biochemistry February-March 2017 Subcellular biochemistry (biochemical aspects of cell biology) Miklós Csala Semmelweis University Dept.
Immunocytochemical localization of β-cop to the ER-Golgi boundary and the TGN
 Journal of Cell Science 108, 2839-2856 (1995) Printed in Great Britain The Company of Biologists Limited 1995 2839 Immunocytochemical localization of β-cop to the ER-Golgi boundary and the TGN Gareth Griffiths
Journal of Cell Science 108, 2839-2856 (1995) Printed in Great Britain The Company of Biologists Limited 1995 2839 Immunocytochemical localization of β-cop to the ER-Golgi boundary and the TGN Gareth Griffiths
Conformational changes of coat proteins during vesicle formation
 FEBS Letters 581 (2007) 2083 2088 Minireview Conformational changes of coat proteins during vesicle formation Julian D. Langer, Emily H. Stoops, Julien Béthune, Felix T. Wieland * Biochemie Zentrum der
FEBS Letters 581 (2007) 2083 2088 Minireview Conformational changes of coat proteins during vesicle formation Julian D. Langer, Emily H. Stoops, Julien Béthune, Felix T. Wieland * Biochemie Zentrum der
Lipidne mikrodomene. funkcija
 Lipidne mikrodomene funkcija 1 Cellular processes involving lipid rafts - Signal transduction - Protein and lipid trafficking and sorting - Endosome(clathrin)-independent endocytosis: - potocytosis and
Lipidne mikrodomene funkcija 1 Cellular processes involving lipid rafts - Signal transduction - Protein and lipid trafficking and sorting - Endosome(clathrin)-independent endocytosis: - potocytosis and
MCB130 Midterm. GSI s Name:
 1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
Zool 3200: Cell Biology Exam 4 Part I 2/3/15
 Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Key Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
JCB. Evidence that the entire Golgi apparatus cycles in interphase HeLa cells: sensitivity of Golgi matrix proteins to an ER exit block.
 JCB Published Online: 5 November, 2001 Supp Info: http://doi.org/10.1083/jcb.200103104 Downloaded from jcb.rupress.org on October 8, 2018 Article Evidence that the entire Golgi apparatus cycles in interphase
JCB Published Online: 5 November, 2001 Supp Info: http://doi.org/10.1083/jcb.200103104 Downloaded from jcb.rupress.org on October 8, 2018 Article Evidence that the entire Golgi apparatus cycles in interphase
Renáta Schipp Gergely Berta Department of Medical Biology
 The cell III. Renáta Schipp Gergely Berta Department of Medical Biology Size and Biology Biology is a visually rich subject many of the biological events and structures are smaller than the unaided human
The cell III. Renáta Schipp Gergely Berta Department of Medical Biology Size and Biology Biology is a visually rich subject many of the biological events and structures are smaller than the unaided human
AP Biology
 Tour of the Cell (1) 2007-2008 Types of cells Prokaryote bacteria cells - no organelles - organelles Eukaryote animal cells Eukaryote plant cells Cell Size Why organelles? Specialized structures - specialized
Tour of the Cell (1) 2007-2008 Types of cells Prokaryote bacteria cells - no organelles - organelles Eukaryote animal cells Eukaryote plant cells Cell Size Why organelles? Specialized structures - specialized
Supplemental Materials Molecular Biology of the Cell
 Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
Supplemental Materials Molecular Biology of the Cell Gilberti et al. SUPPLEMENTAL FIGURE LEGENDS: Figure S1: The effect of pharmacological inhibitors on particle uptake. The data presented in Figure 1
SUPPLEMENTARY FIGURE LEGENDS
 SUPPLEMENTARY FIGURE LEGENDS Supplemental FIG. 1. Localization of myosin Vb in cultured neurons varies with maturation stage. A and B, localization of myosin Vb in cultured hippocampal neurons. A, in DIV
SUPPLEMENTARY FIGURE LEGENDS Supplemental FIG. 1. Localization of myosin Vb in cultured neurons varies with maturation stage. A and B, localization of myosin Vb in cultured hippocampal neurons. A, in DIV
The clathrin adaptor Numb regulates intestinal cholesterol. absorption through dynamic interaction with NPC1L1
 The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Lecture 6 9/17 Dr. Hirsh Organization of Cells, continued
 Cell structure of Eukaryotic cells Lecture 6 9/17 Dr. Hirsh Organization of Cells, continued Lots of double-membraned organelles Existence of an Endo-membrane system separation of areas of cell, transport
Cell structure of Eukaryotic cells Lecture 6 9/17 Dr. Hirsh Organization of Cells, continued Lots of double-membraned organelles Existence of an Endo-membrane system separation of areas of cell, transport
7.06 Spring of PROBLEM SET #6
 7.6 Spring 23 1 of 6 7.6 PROBLEM SET #6 1. You are studying a mouse model of hypercholesterolemia, a disease characterized by high levels of cholesterol in the blood. In normal cells, LDL particles in
7.6 Spring 23 1 of 6 7.6 PROBLEM SET #6 1. You are studying a mouse model of hypercholesterolemia, a disease characterized by high levels of cholesterol in the blood. In normal cells, LDL particles in
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting
 Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
Nature Biotechnology: doi: /nbt.3828
 Supplementary Figure 1 Development of a FRET-based MCS. (a) Linker and MA2 modification are indicated by single letter amino acid code. indicates deletion of amino acids and N or C indicate the terminus
Supplementary Figure 1 Development of a FRET-based MCS. (a) Linker and MA2 modification are indicated by single letter amino acid code. indicates deletion of amino acids and N or C indicate the terminus
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Significance of the MHC
 CHAPTER 8 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
CHAPTER 8 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
Zool 3200: Cell Biology Exam 4 Part II 2/3/15
 Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Organization of ATPases
 The Primary Active Transporter II: The ATPase Objectives: Organization P type with NPA domains Proton pumps of the rotary V type ATPase 1 Organization of P type, solute transport, found in plasma membranes
The Primary Active Transporter II: The ATPase Objectives: Organization P type with NPA domains Proton pumps of the rotary V type ATPase 1 Organization of P type, solute transport, found in plasma membranes
The Cell Organelles. Eukaryotic cell. The plasma membrane separates the cell from the environment. Plasma membrane: a cell s boundary
 Eukaryotic cell The Cell Organelles Enclosed by plasma membrane Subdivided into membrane bound compartments - organelles One of the organelles is membrane bound nucleus Cytoplasm contains supporting matrix
Eukaryotic cell The Cell Organelles Enclosed by plasma membrane Subdivided into membrane bound compartments - organelles One of the organelles is membrane bound nucleus Cytoplasm contains supporting matrix
Zool 3200: Cell Biology Exam 4 Part I 2/3/15
 Name: Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 4 Part I 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Supplementary table and figures
 3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
Chapter 17: Vesicular traffic, secretion, and endocytosis
 Chapter 17: Vesicular traffic, secretion, and endocytosis SEM of the formation of clathrin-coated vesicles on the cytosolic face of the plasma membrane Outline: 1. Techniques for studying the secretory
Chapter 17: Vesicular traffic, secretion, and endocytosis SEM of the formation of clathrin-coated vesicles on the cytosolic face of the plasma membrane Outline: 1. Techniques for studying the secretory
Antigen presenting cells
 Antigen recognition by T and B cells - T and B cells exhibit fundamental differences in antigen recognition - B cells recognize antigen free in solution (native antigen). - T cells recognize antigen after
Antigen recognition by T and B cells - T and B cells exhibit fundamental differences in antigen recognition - B cells recognize antigen free in solution (native antigen). - T cells recognize antigen after
Chapter 2: Exocytosis and endocytosis. Biochimica cellulare parte B 2016/17
 Chapter 2: Exocytosis and endocytosis Biochimica cellulare parte B 2016/17 Exocytosis and endocytosis Transport from the trans-golgi network to the cell exterior: exocytosis. All eukaryotic cells continuously
Chapter 2: Exocytosis and endocytosis Biochimica cellulare parte B 2016/17 Exocytosis and endocytosis Transport from the trans-golgi network to the cell exterior: exocytosis. All eukaryotic cells continuously
I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins
 Lecture 6: Membranes and Cell Transport Biological Membranes I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins 1. Characteristics a. Phospholipids form bilayers
Lecture 6: Membranes and Cell Transport Biological Membranes I. Fluid Mosaic Model A. Biological membranes are lipid bilayers with associated proteins 1. Characteristics a. Phospholipids form bilayers
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Molecular Cell Biology Bulk transport: Receptor mediated endocytosis
 Paper : 15 Module :07 Development Team Principal Investigator : Prof. Neeta Sehgal Department of Zoology, University of Delhi Co-Principal Investigator : Prof. D.K. Singh Department of Zoology, University
Paper : 15 Module :07 Development Team Principal Investigator : Prof. Neeta Sehgal Department of Zoology, University of Delhi Co-Principal Investigator : Prof. D.K. Singh Department of Zoology, University
Eukaryotic cells use internal membrane-bound compartments
 Exo1: A new chemical inhibitor of the exocytic pathway Yan Feng*, Sidney Yu, Troy K. R. Lasell, Ashutosh P. Jadhav*, Eric Macia, Pierre Chardin, Paul Melancon, Michael Roth, Timothy Mitchison*, and Tomas
Exo1: A new chemical inhibitor of the exocytic pathway Yan Feng*, Sidney Yu, Troy K. R. Lasell, Ashutosh P. Jadhav*, Eric Macia, Pierre Chardin, Paul Melancon, Michael Roth, Timothy Mitchison*, and Tomas
Molecular Cell Biology 5068 In Class Exam 1 September 29, Please print your name:
 Molecular Cell Biology 5068 In Class Exam 1 September 29, 2015 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 1 September 29, 2015 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Structures in Cells. Cytoplasm. Lecture 5, EH1008: Biology for Public Health, Biomolecules
 Structures in Cells Lecture 5, EH1008: Biology for Public Health, Biomolecules Limian.zheng@ucc.ie 1 Cytoplasm Nucleus Centrioles Cytoskeleton Cilia Microvilli 2 Cytoplasm Cellular material outside nucleus
Structures in Cells Lecture 5, EH1008: Biology for Public Health, Biomolecules Limian.zheng@ucc.ie 1 Cytoplasm Nucleus Centrioles Cytoskeleton Cilia Microvilli 2 Cytoplasm Cellular material outside nucleus
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/3/6/e1700338/dc1 Supplementary Materials for HIV virions sense plasma membrane heterogeneity for cell entry Sung-Tae Yang, Alex J. B. Kreutzberger, Volker Kiessling,
advances.sciencemag.org/cgi/content/full/3/6/e1700338/dc1 Supplementary Materials for HIV virions sense plasma membrane heterogeneity for cell entry Sung-Tae Yang, Alex J. B. Kreutzberger, Volker Kiessling,
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
BIOSC 041. v Today s lecture. v Today s lab. v Note- Monday is a holiday good time to do some reading!
 BIOSC 041 v Today s lecture Review questions Chapter 6, Cells More review questions v Today s lab Quick review of lab safety The Scientific Method start thinking about which environments you might want
BIOSC 041 v Today s lecture Review questions Chapter 6, Cells More review questions v Today s lab Quick review of lab safety The Scientific Method start thinking about which environments you might want
A positive signal prevents secretory membrane cargo from recycling between the Golgi and the ER
 Article A positive signal prevents secretory membrane cargo from recycling between the Golgi and the ER Matteo Fossati 1,, Sara F Colombo 1 & Nica Borgese 1,2,* Abstract The Golgi complex and ER are dynamically
Article A positive signal prevents secretory membrane cargo from recycling between the Golgi and the ER Matteo Fossati 1,, Sara F Colombo 1 & Nica Borgese 1,2,* Abstract The Golgi complex and ER are dynamically
Structures in Cells. Lecture 5, EH1008: Biology for Public Health, Biomolecules.
 Structures in Cells Lecture 5, EH1008: Biology for Public Health, Biomolecules Limian.zheng@ucc.ie 1 Cytoplasm Nucleus Centrioles Cytoskeleton Cilia Microvilli 2 Cytoplasm Cellular material outside nucleus
Structures in Cells Lecture 5, EH1008: Biology for Public Health, Biomolecules Limian.zheng@ucc.ie 1 Cytoplasm Nucleus Centrioles Cytoskeleton Cilia Microvilli 2 Cytoplasm Cellular material outside nucleus
Dynamics of proteins in Golgi membranes: comparisons between mammalian and plant cells highlighted by photobleaching techniques
 CMLS, Cell. Mol. Life Sci. 61 (2004) 172 185 1420-682X/04/020172-14 DOI 10.1007/s00018-003-3355-6 Birkhäuser Verlag, Basel, 2004 CMLS Cellular and Molecular Life Sciences Dynamics of proteins in Golgi
CMLS, Cell. Mol. Life Sci. 61 (2004) 172 185 1420-682X/04/020172-14 DOI 10.1007/s00018-003-3355-6 Birkhäuser Verlag, Basel, 2004 CMLS Cellular and Molecular Life Sciences Dynamics of proteins in Golgi
A Tour of the Cell. reference: Chapter 6. Reference: Chapter 2
 A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
Cell Structure and Function. Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages and 68-69
 Cell Structure and Function Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages 45 59 and 68-69 Assignments for this Unit Pick up the notes/worksheet for this unit and the project There
Cell Structure and Function Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages 45 59 and 68-69 Assignments for this Unit Pick up the notes/worksheet for this unit and the project There