INTERACTIONS WITH THE INSULIN RECEPTOR AND INHIBITS INSULIN ACTION
|
|
|
- Hester Burns
- 5 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on October 17, 2001 as Manuscript M Running Title: IRS1 Ser 307 Phosphorylation blocks IR IRS1 Interaction PHOSPHORYLATION OF SER 307 IN IRS-1 BLOCKS INTERACTIONS WITH THE INSULIN RECEPTOR AND INHIBITS INSULIN ACTION Vincent Aguirre, Eric D. Werner, Jodel Giraud, Yong Hee Lee Steve E. Shoelson, Morris F. White Howard Hughes Medical Institute Joslin Diabetes Center Harvard Medical School Boston, MA Address Correspondence to: Morris F. White, Ph.D. Howard Hughes Medical Institute Joslin Diabetes Center 1 Joslin Place Boston, MA TEL: (617) FAX: (617) morris.white@joslin.harvard.edu 1 Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
2 ABSTRACT Serine phosphorylation of IRS-1 inhibits insulin signal transduction in a variety of cell backgrounds, which might contribute to peripheral insulin resistance. However, owing to the large number of potential phosphorylation sites, the mechanism of inhibition has been difficult to determine. One serine residue located near the PTB-domain in IRS-1 (Ser 307 in rat IRS-1 or Ser 312 in human IRS-1) is phosphorylated via several mechanisms, including insulin-stimulated kinases or stress-activated kinases like the NH 2 -terminal Jun kinase. During a yeast tri-hybrid assay, phosphorylation of Ser 307 by Jnk-1 disrupted the interaction between the catalytic domain of the insulin receptor and the PTB-domain of IRS-1. In 32D myeloid progenitor cells, phosphorylation of Ser 307 inhibited insulin stimulation of the PI 3-kinase and MAP kinase cascades. These results suggest that inhibition of PTB-domain function in IRS-1 by phosphorylation at Ser 307 (Ser 312 in human IRS-1) might be a general mechanism to regulate insulin signaling. 2
3 INTRODUCTION The insulin signaling system plays an important role in many physiological processes including carbohydrate and fat metabolism, reproduction, cellular growth and survival (1). Acute insulin resistance is mediated, at least in part, by the action of proinflammatory cytokines that are produced during infection, physical trauma, or cancer (2-4). Chronic insulin resistance is an inevitable consequence of genetic variation which is exacerbated by aging and obesity, and contributes to multiple disorders including glucose intolerance, hyperlipidemia, hypertension and cardiovascular mortality, infertility and polycystic ovarian syndrome, and type 2 diabetes (5,6). Insulin resistance alone might not cause diabetes if pancreatic β-cells secrete enough insulin to compensate for reduced sensitivity; however, type 2 diabetes eventually develops, possibly because hyperinsulinemia itself exacerbates the pre-existing resistance until β-cells eventually fail to compensate (7). Understanding the molecular basis of insulin resistance will provide a rational basis for treatment of many related disorders. The insulin signaling system is complex and a common mechanism to explain the occurrence of acute and chronic insulin resistance is difficult to identify. Mutations in the insulin receptor are an obvious source of life-long insulin resistance, but they occur rarely and are not the common cause of type 2 diabetes (8-11). Generally, insulin resistance is a consequence of dysregulated insulin signaling that arises from various sources. Nonspecific or regulated degradation of elements in the insulin-signaling pathway might cause insulin resistance (12); elevated activity or expression of protein or lipid phosphatases, including PTP1B, SHIP2, or pten directly inhibit insulin signals(13,14). 3
4 Covalent modification of the IRS-proteins by serine phosphorylation is implicated in insulin resistance associated with obesity and trauma. Elevated circulating levels of several metabolites are known to promote serine phosphorylation of IRS-1, including free fatty acids, diacylglycerol, fatty acyl CoA s, ceramides and glucose (15). Moreover, adiposederived cytokines like TNFα also stimulate serine/threonine phosphorylation of IRS-1, which inhibits signaling (16). One of the branches of the TNFα signaling pathway involves activation of the c-jun N-terminal kinase (Jnk) (17-19). Jnk phosphorylates numerous cellular proteins, including IRS-1 and IRS-2, Shc and Gab-1(20). Previous work reveals that the major Jnk phosphorylation site in rat IRS-1 is located at Ser 307 (Ser 312 in human IRS-1) that is located on the COOH-terminal side of the phosphotyrosine-binding (PTB) domain (20). In this report, a yeast tri-hybrid assay reveals that Jnk-1 phosphorylation of Ser 307 inhibits the interaction between IRS-1 and the insulin receptor, which provides a rational mechanism to explain, at least in part, insulin resistance that occurs during trauma and obesity. 4
5 MATERIALS AND METHODS Antibodies and Reagents Phosphospecific MAPK, control MAPK, and phosphorospecific AKT antibodies were purchased from New England Biolabs. Control AKT and JNK1 antibodies were purchased from Santa Cruz. Phosphotyrosine antibodies were purchased from Transduction labs. Antibodies against IRS1, IRS2, and p85 have been described (21,22). Antibodies directed against phosphorylated serine 612 in IRS1 were purchased from Biosource International. Rabbit polyclonal serum directed against phosphorylated Ser 307 were generated using a synthetic peptide designed to contain phosphorylated Ser 307 and surrounding amino acids (Boston Biomolecules). Insulin was purchased from Boehringer Mannheim. IGF1 was a gift from Eli Lilly. TNF was purchased from R&D Systems. IRS1 tyrosine phosphorylation site mutants have been previously described (23,24). Point mutants for serine 307 and in the JNK binding domain of IRS1 were generated using appropriate oligonucleotides with the Stratagene Quikchange site-directed mutagenesis method. Constructs for JNK1 and GST-JIP-JBD have been described (25,26). Cell culture 32D cells were maintained in RPMI 1640 medium supplemented with 10% FBS, 5% WEHI conditioned medium (as a source of IL-3), and 5 mm histidinol and made quiescent by serum starvation for 4 hrs. 32D transfectants were generated by electroporation and selected in histidinol as previously described (27). HEK 293 cells were maintained in DMEM containing 10% FBS and made quiescent by serum starvation for 12 hrs. 5
6 Cell lysis, immunoprecipitation, and western analysis 32D cells were lysed in 50 mm Tris (ph 7.4), containing 130 mm NaCl, 5 mm EDTA, 1.0% NP-40, 100 mm NaF, 50 mm β-glycerophosphate, 100 µm NaVO 4, 1 mm PMSF, 5 µg/ml leupeptin, and 5 µg/ml aprotinin. Immunoprecipitations were performed for 2 hrs at 4 o C and collected on protein A Sepharose. Lysates and immunoprecipitates were resolved by SDS-PAGE, transferred to nitrocellulose, and proteins were detected by immunoblotting and either [ 125 I] protein A or enhanced chemiluminescence (Amersham-Pharmacia) and analysis by autoradiography or on a Molecular Dynamics Phosphorimager. HEK293 cells were lysed in 20 mm Tris (ph 7.4) containing 137 mm NaCl, 25 mm β-glycerophosphate, 2 mm sodium pyrophosphate, 2 mm EDTA, 1% Triton X-100, 10% glycerol, 1 mm PMSF, 5 µg/ml leupeptin, 5 µg/ml aprotinin, 2 mm benzamidine, and 0.5 mm DTT. Association of IRS-1 with JNK1 GST fusion proteins containing portions of IRS-1 were made by subcloning the indicated residues into pgex-2tk (Amersham-Pharmacia), expressed in E.coli (BL-21) and purified using glutathione-agarose (Amersham- Pharmacia). GST fusion proteins (111 pmole) were incubated with 293 cell lysates for 2 hrs at 4 o C. Proteins bound to the GST fusion proteins were fractionated by SDS-PAGE, transferred to nitrocellulose, and analyzed by western blot with antibodies against JNK1. In vitro kinase assay HEK 293 cells were transiently transfected with either pcdna3-flag-jnk1 or pcnda3 using Fugene-6. Transient transfectants were made quiescent by serum starvation for 12 hrs and assayed at 36 hrs. Following stimulation with 10 µg/ml anisomycin and lysis, flag-jnk1 was immunoprecipitated with M2 antibody (Sigma) for 2 hrs at 4 o C and immune complexes collected on anti-mouse agarose (Sigma). 6
7 Flag-Jnk1 was eluted with flag peptide [100 µg/ml] in kinase buffer [25 mm Hepes (ph 7.4), 25 mm β-glycerophosphate, 25 mm MgCl 2, 100 µm sodium orthovanadate, and 0.5 mm DTT] overnight at 4 o C. Kinase assays were initiated by addition of kinase and 50-µM γ[ 32 P]ATP to Baculovirus-expressed insulin receptor in a final volume of 50 µl kinase buffer. Reactions were terminated after 30-min at 22 C with ice-cold PBS and addition of SDS-sample buffer. After SDS-PAGE and transfer to nitrocellulose, [ 32 P] phosphorylation of substrate proteins was examined by autoradiography and Cerenkov counting. PI 3- kinase activity assays were performed on IRS-1 immunoprecipitates as previously described (28). Yeast Transformation and Interaction assay The yeast MATCHMAKER LexA two hybrid system reagents were purchased from CLONTECH. The yeast Saccharomyces cerevisiae strain EGY48 (Matα, trp1, his3, ura3, 6LexAop-LEU2, LYS2), plexa IR and pb42ad IRS1/2 constructs were a generous gift from Thomas A. Gustafson. EGY48 was sequentially transformed with plasmid constructs by the polyethylene glycol/lithium acetate method according to the CLONTECH protocol. Transformants were grown on the appropriate SD glucose agar plates for 3 days at 30 o C. Four independent colonies were streaked on SD glucose agar plates, grown overnight, replica plated onto SD galactose/raffinose agar plates, and regrown for 5 days at 30 o C to induce expression of B42 fusion proteins and determine interacting partners. Tri-Hybrid Disruption assay EGY48 (Matα, trp1, his3, ura3, 6LexAop-LEU2, LYS2) was sequentially transformed as above with plexa IR, various pb42ad IRS1 constructs and third plasmid (pdis) containing various cdnas. Transformants were grown 7
8 on the appropriate SD glucose agar plates for 3 days at 30 o C. Four independent colonies were streaked on SD glucose agar plates, incubated overnight, then replica plated onto SD galactose/raffinose agar plates. The plates were immediately replica cleaned, incubated overnight, replica cleaned, and incubated at 30 o C for 5 days to promote growth and induce the expression of B42 fusion proteins. 8
9 RESULTS Phosphorylation of Ser 307 in IRS-1 IRS-1 of rat or human origin contains many potential serine phosphorylation sites that are thought to play regulatory roles during insulin signaling. One of these sites, Ser 307 in rat IRS-1, was originally found to be phosphorylated specifically by the NH 2 -terminal Jun kinase (Jnk). Ser 307 was later found to be phosphorylated in IRS-1 isolated from cells and tissues stimulated with TNFα, insulin/igf-1, or anisomycin (20,29). Phosphorylation of Ser 307 is interesting because it inhibits insulin-stimulated tyrosine phosphorylation of rat IRS-1. To study the role of Ser 307 on insulin signaling, full-length rat IRS-1 or a mutant IRS-1 molecule containing a Ser 307 Ala substitution (A307 IRS1 ) were expressed stably in 32D IR cells. 32D cells are interleukin-3-dependent murine myeloid progenitor cells that express few insulin receptors and no IRS-proteins; over expression of IRS-1 and the insulin receptor reconstitutes many aspects of the insulin-signaling pathway in 32D cells (28,30). Immunoprecipitates of IRS-1 from 32D IR cells stimulated with insulin or anisomycin were analyzed by immunoblotting with phosphospecific antibodies against Ser 307 (αps 307 ), and for comparison Ser 612 (αps 612 ). Before stimulation, both antibodies reacted weakly with IRS-1, indicating that these phosphorylation sites were slightly phosphorylated under the basal conditions (Fig 1A). Anisomycin or insulin strongly stimulated phosphorylation of Ser 307 and Ser 612, whereas Ser 612 was phosphorylated only during anisomycin stimulation (Fig. 1A). As previously shown, TNFα stimulated Ser 307 phosphorylation more slowly than either insulin or anisomycin (Fig. 1B) (20). Thus, Ser 307 is a common phosphorylation 9
10 site for several signaling pathways that was detected specifically by immunoblotting with αps 307. Insulin-stimulated Ser 307 phosphorylation requires tyrosine phosphorylation of rat IRS-1 Insulin rapidly stimulated phosphorylation of Ser 307 in 32D IR cells (Fig. 2A). Ser 307 phosphorylation was more sensitive to insulin than IGF-1, owing presumably to the higher expression of recombinant insulin receptor in these cells. Previous results showed that PI 3-kinase inhibitors block insulin-stimulated Ser 307 phosphorylation (31). To determine whether insulin-stimulated Ser 307 phosphorylation requires tyrosine phosphorylation, IRS1 F18 was examined in 32D IR cells. IRS1 F18 lacks 18 potential tyrosine phosphorylation sites and fails to bind various SH2-proteins, including p85, Grb-2 and SHP2 (23). Interestingly, IRS1 F18 was not phosphorylated on Ser 307 during insulin stimulation, whereas it was phosphorylated normally during anisomycin and TNFα stimulation (Fig. 2B). Insulin-stimulated Ser 307 phosphorylation was restored by site-directed mutagenesis that replaced three PI 3-kinase binding sites in IRS1 F18, including Tyr 608, Tyr 628 and Tyr 658. By contrast, restoring the SHP2 binding sites in IRS1 F18 did not promote Ser 307 phosphorylation. Moreover, deletion of the Grb-2 binding motif in IRS-1 did not inhibit insulin-stimulated Ser 307 phosphorylation (Fig. 2C). Thus, several distinct signaling pathways, including insulin stimulation of the PI 3-kinase cascade, converge at Ser 307 to mediate negative feedback or heterologous inhibition of IRS-1 signaling to inhibit the insulin response. 10
11 Ser 307 phosphorylation inhibits IR IRS1 interaction During experiments in mammalian cells and yeast, the PTB-domain of IRS-1 binds to the phosphorylated NPEY-motif in the juxtamembrane region of the insulin receptor (32-35). In mammalian cells, this interaction promotes efficient phosphorylation of IRS-1 during insulin stimulation. Since Ser 307 is near the PTB-domain of IRS-1, phosphorylation of this residue during insulin or TNFα stimulation might disrupt the interaction between the insulin receptor and IRS-1. To determine if Jnk1-mediated phosphorylation of Ser 307 inhibits binding between the insulin receptor and IRS-1, a yeast tri-hybrid assay was developed to test the effect of Jnk-1 on the interaction between the insulin receptor (bait) and various IRS-1 constructs (prey). Prior work reveals that the PTB domain couples IR to IRS1 in yeast; however, to validate the tri-hybrid assay the specific interaction between Jnk-1 and IRS-1 in yeast was established. Human IRS-1 contains two putative JIP-homology motifs between residues and (residues and in the rat orthologs) that might specifically bind Jnk-1 (Fig. 3A) (20,36). The LxL sequence of this motif in JIP-1 is required for Jnk interaction (26). To establish which motif interacts with Jnk-1, various deletion constructs of IRS-1 (prey) were expressed with Jnk-1 (bait) in the yeast two-hybrid assay. Full length IRS-1 interacted with Jnk-1 as revealed by β-galactosidase activity in yeast growing on selective media, whereas an IRS-1 construct lacking both JIP-homology regions did not promote yeast growth (Fig. 3B). IRS-1 constructs retaining residues or , which contain the JIP homology region in the overlapping sequence, promoted growth and β-galactosidase activity (Fig 3B). These results suggest that the JIP- 11
12 homology motif in human IRS-1 between residues bound to Jnk1 in the yeast twohybrid system. In vitro binding experiments confirmed that the orthologous JIP-homology region (RPTRLSL 858 -motif) in rat IRS-1 binds Jnk-1. GST-fusion proteins containing a portion of rat IRS-1 with intact or mutant JIP-homology domains were incubated with 293 cell lysates containing Jnk-1. Jnk-1 associated with the immobilized fragments of wild-type IRS-1 (Fig. 3C). However, a Leu 852,858 Gly substitution, but not Leu 784,786 Gly sustitution abolished the ability of rat IRS-1 to pull-down Jnk-1 (Fig. 3C). Yeast expressing the human insulin receptor (bait) and human IRS-1 (prey) constructs grow efficiently on selective media; and growth was not inhibited when inactive Jnk-1 (Jnk1 APF ) or an empty vector were expressed in these yeast (Fig. 3D). By contrast, co-expression of active Jnk-1 prevented growth, suggesting that the IR IRS1 interaction was inhibited (Fig. 3D). Substitution of Ser 312 for alanine in human IRS-1 (orthologous to Ser 307 in rat IRS-1) blocked Jnk-1 mediated growth inhibition, suggesting that Jnk1 mediated phosphorylation of Ser 307 was required for the Jnk1-mediated disruption of the IR IRS1 interaction. Moreover, truncated human IRS-1 composed of residues , including Ser 312 but lacking the JIP-homology region, was insensitive to activate Jnk1 during growth on selective media (Fig. 3D). The insulin receptor did not interact with Jnk1 in a yeast two-hybrid assay (data not shown) and was not phosphorylated by Jnk1 during in vitro kinase assays using purified insulin receptor and Jnk1 (Fig. 3E). These results are consistent with the hypothesis that the interaction between the insulin receptor and IRS-1 is 12
13 inhibited by phosphorylation of Ser 307 during association of Jnk1 with the JIP-homology region of IRS-1. Phosphorylation of Ser 307 inhibits insulin signaling Anisomycin was used to promote phosphorylation of Ser 307 in rat IRS-1 and A307 IRS1 to establish the effect on insulin-stimulated PI 3-kinase and MAP kinase cascades. Anisomycin rapidly stimulated Ser 307 phosphorylation of IRS-1 with a half maximal effect below 100 ng/ml (Fig. 4A and B). By contrast, αps 307 weakly immunoblotted A307 IRS1 before and after anisomycin treatment, confirming that Ser 307 was removed from the mutant molecule (Fig. 4A and B). The residual immunoblotting of A307 IRS1 by αps 307 might reflect cross reactivity with other phosphorylation sites in IRS-1, such as Ser 612. Tyrosine phosphorylation of IRS-1 was detected by immunoblotting with αpy, as previously described (37). Treatment of 32D IR /IRS1 cells for 30 min with 0.01 or 0.1- µg/ml anisomycin inhibited insulin-stimulated tyrosine phosphorylation by 35%; and inhibition reached 45% with 1.0 µg/ml anisomycin (Fig. 5A). Low concentrations of anisomycin had no inhibitory effect on the tyrosine phosphorylation of A307 IRS1, and inhibition barely reached 15% at 1.0 µg/ml anisomycin (Fig 5A). The inhibitory effect of anisomycin did not occur through degradation of IRS-1 or inhibition of insulin receptor autophosphorylation (data not shown). However, anisomycin-mediated inhibition of IRS1 tyrosine phosphorylation required a functional JIP-homology domain. Mutation of the LxL sequence of the Jnk binding domain of JIP-1 to GxG abrogates Jnk interaction (personal communication, Roger Davis and 26). Inactivation of the JIP-homology region by point mutations of the L 856 SL-motif to a G 856 SG-motif completely eliminated the inhibitory 13
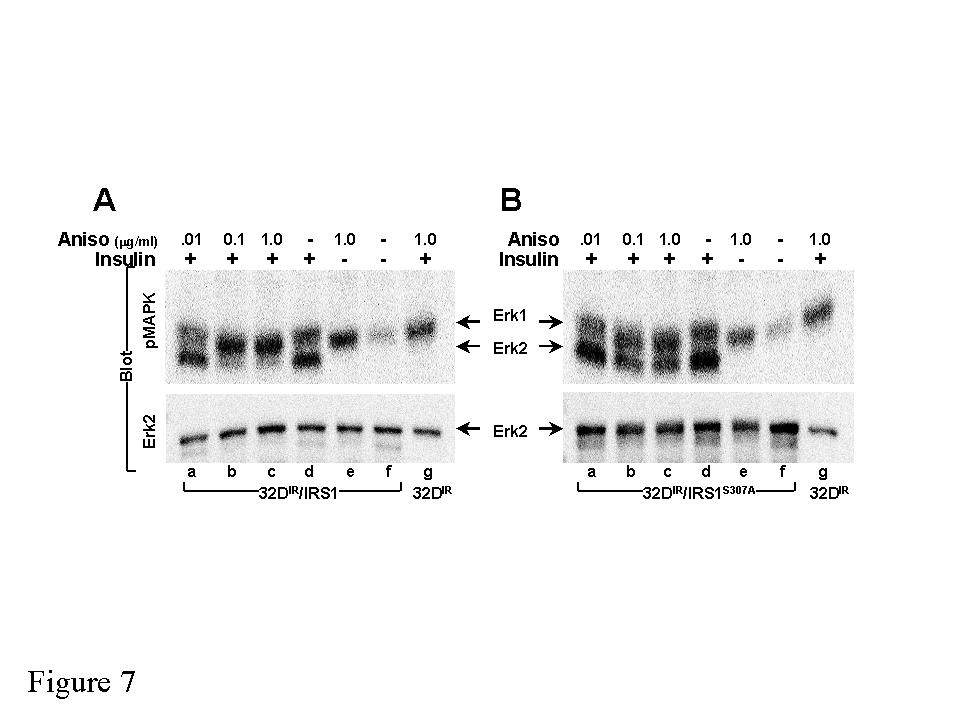
14 effect of anisomycin (Fig. 6). These results confirm that the intact JIP-homology region mediates inhibition of insulin-stimulated tyrosine phosphorylation, most likely through phosphorylation of Ser 307. Many insulin signals are mediated through the binding of tyrosine phosphorylated motifs in IRS-1 to the src-homology 2 domains in various signaling proteins (SH2- proteins), including the PI 3-kinase and Grb2 (38). Consistent with the inhibition of insulin-stimulated tyrosine phosphorylation of IRS-1, anisomycin inhibited the binding of p85 to IRS-1 in 32D IR cells; however, the binding of A307 IRS1 to p85 during insulin stimulation was not inhibited by anisomycin (Fig. 5B). In the same dose-dependent manner, anisomycin inhibited insulin-stimulated PI 3-kinase activity associated with IRS-1, but had no effect on PI 3-kinase activity associated with A307 IRS1 during insulin stimulation (Fig. 5C). Insulin promotes the association of Grb-2 with IRS-1 or Shc, which stimulates the phosphorylation of Erk1 and Erk2 that is detected by immunoblotting with α- phosphomapk. In 32D IR cells, analysis of the inhibitory effect of anisomycin on Erk-1 phosphorylation was confounded by stimulation of Erk1 phosphorylation by anisomycin and insulin through the Shc pathway in the absence of IRS1 or A307 IRS1 expression (Fig. 7A). By contrast, Erk2 was only phosphorylated during insulin stimulation of 32D IR cells expressing either IRS-1 or A307 IRS1 (Fig. 7A and B). Anisomycin completely inhibited insulin-stimulation of Erk2 phosphorylation (60% inhibition at 0.1 µg/ml) in 32D IR /IRS1 cells. By contrast, Erk2 phosphorylation was barely inhibited 20% at the highest anisomycin concentration in 32D/A307 IRS1 cells (Fig 7B). These data reveal that phosphorylation of Ser 307 inhibits IRS1-mediated Erk2 phosphorylation. 14
15 15
16 DISCUSSION Our results reveal a general mechanism for the negative feedback and heterologous regulation of the IRS-1 branch of the insulin-signaling pathway through inhibition of PTBdomain function by phosphorylation of Ser 307. Previous work established that the interaction in yeast between the insulin receptor catalytic domain and IRS-1 is mediated entirely through the binding of the phosphorylated NPEY-motif in the insulin receptor to the PTB-domain in IRS-1(32,33,39). Based on this prior information, we conclude that disruption of the binding between the insulin receptor and IRS-1 in yeast expressing Jnk-1 occurs because phosphorylation of Ser 307 disrupts PTB-domain function (35, 34) All of the control experiments confirm this conclusion, including association of the insulin receptor and IRS-1 in yeast expressing a kinase dead Jnk-1 construct; association of the insulin receptor and human IRS-1 mutant with a Ser 312 Ala in yeast expressing a functional Jnk-1 construct; and association of the insulin receptor and an IRS-1 construct lacking the JIPhomology region in yeast expressing Jnk1. Although the yeast 3-hybrid assay reveals that Ser 307 phosphorylation completely abrogates insulin receptor IRS1 interaction, the 32D IR cell-based experiments suggest that it only inhibits IRS-1 tyrosine phosphorylation by 50% at best. Previous work reveals that efficient phosphorylation of IRS-1 depends on two NH 2 -terminal domains, the pleckstrin homology (PH) domain and the adjacent phosphotyrosine-binding (PTB) domain (35,40). Deletion of both the PH and PTB domains completely inhibits phosphorylation during insulin stimulation of 32D IR cells, whereas deletion of either the PH or PTB domain partially reduces tyrosine phosphorylation. Since Ser 307 phosphorylation inhibits PTB 16
17 domain function, persistent coupling mediated through the PH domain might be responsible for incomplete inhibition of IRS-1 tyrosine phosphorylation. Efficient coupling of IRS-1 to low levels of insulin receptors requires both domains, whereas either the PH or PTB domain is sufficient in cells expressing high levels of insulin receptor. Therefore, in cells with a low number of receptors, Ser 307 phosphorylation might play a major regulatory role, whereas Ser 307 phosphorylation might be inefficient in cells with a high number of receptors (35). Under the later condition, more drastic regulatory mechanisms might be required, including degradation of IRS-1. Considerable evidence is largely consistent with the hypothesis that serine phosphorylation of the insulin receptor or the IRS-proteins inhibits signal transduction. Despite the potential importance of this regulatory pathway, the sites of phosphorylation and the inhibitory mechanisms involved have been difficult to identify. Increased serine phosphorylation of IRS-1 is a common finding during insulin resistance and type 2 diabetes (41). Serine phosphorylated IRS-1 inhibits insulin-stimulated autophosphorylation of the insulin receptor, PI 3-kinase activation, glucose uptake, and other insulin-stimulated biological responses (3,3,42-50). Besides the Jnk phosphorylation site at Ser 307, IRS-1 contains serine/threonine residues in consensus sequences for many other protein kinases, including casein kinase II, camp-dependent protein kinase, protein kinase C, cdc2 kinase, MAP kinase, and PKB/akt (21,42,44,51-53). Recent reports suggest that serine phosphorylation of IRS-1 inhibits its ability to associate with the insulin receptor and to serve as a substrate for tyrosine phosphorylation (3,42,45,50,53,54). Thus, the identification of serine/threonine phosphorylation-based mechanisms of signal inhibition 17
18 might reveal a molecular basis for insulin resistance that promotes the pathogenesis of type 2 diabetes. Ser 307 phosphorylation promotes general inhibition of IRS-1 signaling as revealed by reduced activation of both the PI 3-kinase and MAP kinase cascades. This effect does not occur through inhibition of insulin receptor autophosphorylation, but is consistent with reduced coupling between the insulin receptor and IRS-1. Association of p85 or Grb-2 with IRS-1 depends on distinct sets of tyrosine phosphorylation motifs that are separated by up to 300 amino acids in the primary sequence. These results are consistent with general inhibition of tyrosine phosphorylation expected during inhibition of PTB-domain function by Ser 307 phosphorylation. This general inhibition is in contrast to the specific inhibition of p85 association at tyrosine phosphorylation motifs directly adjacent to previously identified inhibitory serine residues (45,55,56). At least three kinases apparently mediate phosphorylation of Ser 307, including a TNFα/anisomycin-stimulated kinase other than Jnk, and an insulin/igf-1 stimulated kinase that is inhibited by wortmannin/ly and requires PI 3-kinase activity. We originally thought that Jnk might be the common final step that mediates Ser 307 phosphorylation downstream of various cytokines, an especially attractive hypothesis since Jnk1 binds to IRS-1(20) and the Jnk-binding region of IRS-1 is required for the inhibition of insulinstimulated tyrosine phosphorylation of IRS-1 by anisomycin. However, some experiments with potential physiologic mediators of insulin resistance do not support this hypothesis. Whereas anisomycin and TNFα stimulate Jnk and Ser 307 phosphorylation, the MEK kinase inhibitor PD98059 completely inhibits Ser 307 phosphorylation with no effect on Jnk activity (31). Although insulin activates Jnk in certain cells, this pathway is not inhibited by 18
19 wortmannin/ly294002, suggesting that a distinct cascade is involved. Therefore, in addition to Jnk at least two other kinases apparently mediate phosphorylation of Ser 307. These kinases might possess the common ability to bind to the Jnk binding domain in IRS1, although other mechanisms could be involved. In summary, potential mediators of chronic insulin resistance, such as TNFα and hyperinsulinemia, lead to progressive accumulation of IRS-1 molecules that are phosphorylated on Ser 307 and couple less efficiently to the insulin receptor. Chronic Ser 307 phosphorylation might also target IRS-1 for degradation or to subcellular compartments inaccessible to the activated insulin receptor. Other IRS-proteins, especially IRS-2, might be similarly sensitive to serine phosphorylation. IRS-2 contains a JIP-homology region, although a residue analogous to Ser 307 does not exist in IRS-2 (20). Nevertheless IRS2 is serine phosphorylated during TNFα or anisomycin stimulation, which inhibits insulinstimulated tyrosine phosphorylation. Since IRS-1 is essential to sustain compensatory insulin secretion in mice, serine phosphorylation mediated inhibition might promote both peripheral insulin resistance and β-cell failure. Identification of the phosphorylation sites in IRS-2 that inhibit insulin-stimulated tyrosine phosphorylation and the kinase specific to those sites are important targets for future mechanism-based drug discovery. 19
20 FIGURE LEGENDS Figure 1. Anisomycin and TNFα induce phosphorylation of Ser 307 in IRS1. (A) IRS1 immunoprecipitates from 32D IR /IRS1 cells treated with 10 nm insulin for the indicated times (Ins) or 1.0 µg/ml anisomycin were analyzed by immunoblotting with antibodies against phosphorylated Ser 307 (αps 307 ) or Figure 2. Ser 612 (αps 612 ), and IRS1. (B) IRS1 immunoprecipitates from 32D IR /IRS1 cells treated with 25 ng/ml TNFα for the indicated times were analyzed by immunoblotting with αp S307 or IRS1. Insulin induces Serine 307 phosphorylation in a PI3K-dependent manner. (A) IRS1 immunoprecipitates from 32D/IR/IRS1 cells treated with 10 nm insulin (Ins) or 10 nm IGF1 for the indicated times were analyzed by immunoblotting with antibodies against Serine 307 (ps307). IRS1 immunoprecipitates from 32D/IR/IRS1 treated with the indicated doses of insulin and IGF1 for 30 min were analyzed by immunoblotting with antibodies against pser 307 and IRS1. (B) IRS1 immunoprecipitates from 32D/IR/IRS1 F18 cells treated with 5 µg/ml anisomycin (A), 10 nm insulin (I), or 25 ng/ml TNF (T) were analyzed by immunoblotting with antibodies against pser307 and IRS1. (C) Schematic of various IRS1 tyrosine phosphorylation site mutants. IRS1 immunoprecipitates from 32D/IR/IRS1 and 32D/IR cells stably expressing various IRS1 tyrosine phosphorylation site mutants treated with 10 nm insulin (Ins) for 30 min were analyzed by immunoblotting with antibodies against pser307 and IRS1. 20
21 Figure 3. Ser 307 phosphorylation inhibits insulin action through disruption of the IR/IRS1 interaction. (A) Schematic comparing JBD at 856/858 in the rat IRS1 sequence and 861/863 in the human IRS1 sequence. (B) Schematic representation of yeast interaction analysis between human JNK1 (bait) and various truncations of human IRS1 (prey). Symbols are representative of the presence (+) or lack (-) of interaction between given partners, as determined by growth on selective media (using two independent reporters, LEU2 and LacZ). (C) The Jnk-binding domain of JIP1 (JIP JBD), the Y2 region (amino acids ) of IRS1, the Y2 region of IRS-1 in which the pair of leucine in the potential Jnk-binding domain at residues 856 and 858, were mutated to glycine, and the Y2 region of IRS-1 in which the pair of leucine in the potential Jnk-binding domain at residues 784 and 786 were mutated to glycine, were expressed as GST fusion proteins (111 pmole) and incubated with HEK293 whole cell lysates (WCL). WCL and GST-pull-downs were analyzed by immunoblotting with antibodies against JNK1. (D) Yeast tri-hybrid disruption analysis of the interaction between the human insulin receptor (bait), human IRS1 (prey), and human IRS1 S312A (prey) by human JNK1 (disruptant). The IR IRS1 (bait-prey) interaction is scored by growth on selective medium. Disruption of the IR IRS1 interaction by JNK1, catalytically inactive JNK1 (JNK1 APF ), or empty vector control (EV) is assayed as a lack of growth on selective medium. (E) IRS1 and baculovirus expressed insulin receptor (IR-β) were subjected to in vitro kinase assay with 21
22 JNK1 expressed and activated with 5.0 µg/ml anisomycin in HEK293 cells. HEK293 cells transfected with empty vector (EV) were assayed in parallel. Figure 4. Figure 5. Anisomycin induces phosphorylation of Ser 307 in IRS1 in a dose and timedependent manner. (A) IRS1 immunoprecipitates from 32D IR, 32D IR /IRS1 and 32D IR /A307 IRS1 cells treated with 1.0 µg/ml anisomycin for the indicated times were analyzed by immunoblotting with antibodies against Serine 307 (ps307) and IRS1. (B) IRS1 immunoprecipitates from 32D IR, 32D IR /IRS1 and 32D IR /A307 IRS1 cells treated with the indicated doses of anisomycin for 30 min were analyzed by immunoblotting with antibodies against phosphorylated Ser 307 and IRS1. Serine-307 of IRS1 is required for the inhibition of insulin signaling by anisomycin. (A) Proteins in whole cell lysates (WCL) from 32D IR /IRS1 and 32D IR /A307 IRS1 cells treated with the indicated doses of anisomycin for 30 min prior to stimulation with 10-nM insulin (Ins) for 5-min were analyzed with antibodies against phosphotyrosine (py). (B) Immunoprecipitates of the p85 regulatory subunit of the PI 3-kinase from 32D IR /IRS1 and 32D IR /A307 IRS1 cells treated with the indicated doses of anisomycin for 30 min prior to stimulation with 10-nM insulin (Ins) for 5-min were analyzed with antibodies against IRS1. (C) IRS1 immunoprecipitates from 32D IR /IRS1 and 32DIR/A307 IRS1 cells treated with the indicated doses of anisomycin for 30 min prior to stimulation with 10-nM insulin for 5-min were analyzed for associated PI 3-kinase activity. Phosphorylated inositol was resolved by chromatography and visualized by phosphorimaging. 22
23 Figure 6. Proteins in whole cell lysates from 32D IR /IRS1 and 32D IR /GSG IRS1 cells treated with the indicated doses of anisomycin for 30 min prior to stimulation with 10-nM insulin (Ins) for 5-min were analyzed with antibodies against phosphotyrosine (py). Figure 7. Serine-307 is required for the inhibition of IRS1-dependent insulin-stimulated Erk2 activity by anisomycin. Proteins in whole cell lysates from 32D IR, 32D IR /IRS1 (A) and 32D IR /A307 IRS1 (B) cells treated with the indicated doses of anisomycin for 30 min prior to stimulation with 10-nM insulin (Ins) for 5 min were analyzed with antibodies against phospho-mapk (pmapk) and Erk2. 23
24 Acknowledgements YHL was supported in part by Korean Science and Engineering Foundation. The authors would like to thank Lauren Kelly for excellent secretarial assistance, Liangyou Rui for technical assistance and critical review, and Jeff Thomas for technical assistance. 24
25 References 1. White, M. F. and Myers, M. G. (2001) The Molecular Basis of Insulin Action. In DeGroot, L. J. and Jameson, J. L., editors. Endocrinology, 2. Loddick, S. A. and Rothwell, N. J. (1999) Proc.Natl.Acad.Sci.U.S.A 96, Hotamisligil, G. S., Peraldi, P., Budvari, A., Ellis, R. W., White, M. F., and Spiegelman, B. M. (1996) Science 271, Yoshikawa, T, Noguchi, Y, and Satoh, S. (1999) Biochem Biophys Res Commun 256, Maezono, K., Osman, A., Patti, M. E., Cusi, K., Pendergrass, M., DeFronzo, R. A., and Mandarino, L. (1998) Diabetes 47, A Kahn, B. B. and Flier, J. S. (2000) J Clin Invest 106, Kahn, B. B. (1998) Cell 92, Hani, E. H., Suaud, L., Boutin, P., Chevre, J. C., Durand, E., Philippi, A., Demenais, F., Vionnet, N., Furuta, H., Velho, G., Bell, G. I., Laine, B., and Froguel, P. (1998) J Clin.Invest. 101,
26 9. Carboni, J. M., Yan, N., Cox, A. D., Bustelo, X., Graham, S. M., Lynch, M. J., Weinmann, R., Seizinger, B. R., Der, C. J., Barbacid, M., and et al. (1995) Oncogene 10, Vaxillaire, M., Rouard, M., Yamagata, K., Oda, N., Kaisaki, P. J., Boriraj, V. V., Chevre, J. C., Boccio, V., Cox, R. D., Lathrop, G. M., Dussoix, P., Philippe, J., Timsit, J., Charpentier, G., Velho, G., Bell, G. I., and Froguel, P. (1997) Hum.Mol Genet 6, Comb, D. G. and Roseman, S. (1958) J.Biol.Chem. 232, Sun, X. J., Goldberg, J. L., Qiao, L. Y., and Mitchell, J. J. (1999) Diabetes 48, Ishihara, H., Sasaoka, T., Hori, H., Wada, T., Hirai, H., Haruta, T., Langlois, W. J., and Kobayashi, M. (1999) Biochem.Biophys.Res.Commun. 260, Clement, S., Krause, U., Desmedt, F., Tanti, J.-F., Behrends, J., Pesesse, X., Sasaki, T., Penninger, J., Doherty, M., Malaisse, W., Dumont, J. E., Le Maechand-Brustel, Y., Erneux, C., Hue, L., and Schurmans, S. (2001) Nature 409, Shulman, G. I. (2000) J Clin Invest 106, Peraldi, P., Hotamisligil, G. S., Buurman, W. A., White, M. F., and Spiegelman, B. M. (1996) J.Biol.Chem. 271,
27 17. Yuasa, T., Ohno, S., Kehrl, J. H., and Kyriakis, J. M. (1998) J.Biol Chem. 273, Kuan, C. Y., Yang, D. D., Samanta Roy, D. R., Davis, R. J., Rakic, P., and Flavell, R. A. (1999) Neuron 22, Rincon, M., Whitmarsh, A., Yang, D. D., Weiss, L., Derijard, B., Jayaraj, P., Davis, R. J., and Flavell, R. A. (1998) J.Exp.Med. 188, Aguirre, V., Uchida, T., Yenush, L., Davis, R. J., and White, M. F. (2000) J.Biol Chem. 275, Sun, X. J., Wang, L. M., Zhang, Y., Yenush, L., Myers, M. G., Jr., Glasheen, E. M., Lane, W. S., Pierce, J. H., and White, M. F. (1995) Nature 377, Pons, S., Asano, T., Glasheen, E. M., Miralpeix, M., Zhang, Y., Fisher, T. L., Myers, M. G., Jr., Sun, X. J., and White, M. F. (1995) Mol.Cell Biol. 15, Myers, M. G., Jr., Zhang, Y., Aldaz, G. A. I., Grammer, T. C., Glasheen, E. M., Yenush, L., Wang, L. M., Sun, X. J., Blenis, J., Pierce, J. H., and White, M. F. (1996) Mol.Cell Biol. 16, Myers, M. G., Jr., Wang, L. M., Sun, X. J., Zhang, Y., Yenush, L., Schlessinger, J., Pierce, J. H., and White, M. F. (1994) Mol.Cell Biol. 14, Derijard, B., Hibi, M., Wu, I. H., Barrett, T., Su, B., Deng, T., Karin, M., and Davis, R. J. (1994) Cell 76,
28 26. Dickens, M., Rogers, J. S., Cavanagh, J., Raitano, A., Xia, Z., Halpern, J. R., Greenberg, M. E., Sawyers, C. L., and Davis, R. J. (1997) Science 277, Yenush, L., Zanella, C., Uchida, T., Bernal, D., and White, M. F. (1998) Mol.Cell Biol. 18, Uchida, T., Myers, M. G., Jr., and White, M. F. (2000) Mol Cell Biol 20, Rui, L., Aguirre, V., Kim, J. K., Shulman G.I., Lee, A., orbould, A., Dunaif, A., and White, M. F. (2001) J Clin Invest 107, Wang, L. M., Myers, M. G., Jr., Sun, X. J., Aaronson, S. A., White, M. F., and Pierce, J. H. (1993) Science 261, Rui, L., Aguirre, V., Kim, J. K., Shulman G.I., Lee, A., Corbould, A., Dunaif, A., and White, M. F. (2001) Journal of Clinical Investigation 107, Craparo, A., O'Neill, T. J., and Gustafson, T. A. (1995) J.Biol.Chem. 270, O'Neill, T. J., Craparo, A., and Gustafson, T. A. (1994) Mol.Cell Biol. 14, Wolf, G., Trub, T., Ottinger, E., Groninga, L., Lynch, A., White, M. F., Miyazaki, M., Lee, J., and Shoelson, S. E. (1995) J.Biol.Chem. 270, Yenush, L., Makati, K. J., Smith-Hall, J., Ishibashi, O., Myers, M. G., Jr., and White, M. F. (1996) J.Biol.Chem. 271,
29 36. Yasuda, J., Whitmarsh, A. J., Cavanagh, J., Sharma, M., and Davis, R. J. (1999) Mol.Cell Biol. 19, Myers, M. G., Jr., Zhang, Y., Yenush, L., Glasheen, E. M., Grammer, T. C., Wang, L. M., Blenis, J., Sun, X. J., Pierce, J. H., and White, M. F. (1995) Diabetes 44, 49A 38. Yenush, L. and White, M. F. (1997) Bio.Essays 19, Gustafson, T. A., He, W., Craparo, A., Schaub, C. D., and O'Neill, T. J. (1995) Mol.Cell Biol. 15, Burks, D. J., Pons, S., Towery, H., Smith-Hall, J., Myers, M. G., Jr., Yenush, L., and White, M. F. (1997) J.Biol.Chem. 272, Virkamaki, A., Ueki, K., and Kahn, C. R. (1999) J.Clin.Invest. 103, De Fea, K. and Roth, R. A. (1997) J.Biol.Chem. 272, Chin, J. E., Liu, F., and Roth, R. A. (1994) Mol.Endocrinol. 8(1), Li, J., De Fea, K., and Roth, R. A. (1999) J.Biol.Chem. 274, Mothe, I. and Van Obberghen, E. (1996) J.Biol.Chem. 271, Tanti, J. F., Gremeaux, T., Van Obberghen, E., and Le Marchand-Brustel, Y. (1994) J.Biol.Chem. 269(8), De Meyts, P. (1995) Diabetologia 37, s135-s148 29
30 48. Richardson, J. M. and Pessin, J. E. (1993) J.Biol.Chem. 268, Kroder, G., Beossenmaier, B., Kellerer, M., Capp, E., Stoyanov, B., Muhlhofer, A., Berti, L., Horikoshi, H., Ullrich, A., and Haring, H. (1996) J.Clin.Invest. 97, Paz, K., Hemi, R., LeRoith, D., Karasik, A., Elhanany, E., Kanety, H., and Zick, Y. (1997) J.Biol.Chem. 272, Sun, X. J., Rothenberg, P. L., Kahn, C. R., Backer, J. M., Araki, E., Wilden, P. A., Cahill, D. A., Goldstein, B. J., and White, M. F. (1991) Nature 352, Barthel, A., Nakatani, K., Dandekar, A. A., and Roth, R. A. (1998) Biochem Biophys Res Commun 243, De Fea, K. and Roth, R. A. (1997) Biochemistry 36, Kanety, H., Feinstein, R., Papa, M. Z., Hemi, R., and Karasik, A. (1995) J.Biol.Chem. 270, Delahaye, L., Mothe-Satney, I., Myers, M. G., Jr., White, M. F., and Van Obberghen, E. (1998) Endocrinology 139, De Fea, K. and Roth, R. A. (1997) Biochemistry 36,
31

32
33
34
35
36
37
38 Phosphorylation of SER307 in IRS-1 blocks interactions with the insulin receptor and inhibits insulin action Vincent Aguirre, Eric D. Werner, Jodel Giraud, Yong Hee Lee, Steve E. Shoelson and Morris F. White J. Biol. Chem. published online October 17, 2001 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts
Phosphorylation of Ser 307 in Insulin Receptor Substrate-1 Blocks Interactions with the Insulin Receptor and Inhibits Insulin Action*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 2, Issue of January 11, pp. 1531 1537, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Phosphorylation
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 2, Issue of January 11, pp. 1531 1537, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Phosphorylation
The c-jun NH 2 -terminal Kinase Promotes Insulin Resistance during Association with Insulin Receptor Substrate-1 and Phosphorylation of Ser 307 *
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 275, No. 12, Issue of March 24, pp. 9047 9054, 2000 2000 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The c-jun NH 2 -terminal
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 275, No. 12, Issue of March 24, pp. 9047 9054, 2000 2000 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The c-jun NH 2 -terminal
Insulin/IGF-1 and TNF-α stimulate phosphorylation of IRS-1 at inhibitory Ser 307 via distinct pathways
 Insulin/IGF-1 and TNF-α stimulate phosphorylation of IRS-1 at inhibitory Ser 307 via distinct pathways Liangyou Rui, 1 Vincent Aguirre, 1 Jason K. Kim, 2 Gerald I. Shulman, 2 Anna Lee, 3 Anne Corbould,
Insulin/IGF-1 and TNF-α stimulate phosphorylation of IRS-1 at inhibitory Ser 307 via distinct pathways Liangyou Rui, 1 Vincent Aguirre, 1 Jason K. Kim, 2 Gerald I. Shulman, 2 Anna Lee, 3 Anne Corbould,
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
William C. Comb, Jessica E. Hutti, Patricia Cogswell, Lewis C. Cantley, and Albert S. Baldwin
 Molecular Cell, Volume 45 Supplemental Information p85 SH2 Domain Phosphorylation by IKK Promotes Feedback Inhibition of PI3K and Akt in Response to Cellular Starvation William C. Comb, Jessica E. Hutti,
Molecular Cell, Volume 45 Supplemental Information p85 SH2 Domain Phosphorylation by IKK Promotes Feedback Inhibition of PI3K and Akt in Response to Cellular Starvation William C. Comb, Jessica E. Hutti,
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
G-Protein Signaling. Introduction to intracellular signaling. Dr. SARRAY Sameh, Ph.D
 G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
KEY CONCEPT QUESTIONS IN SIGNAL TRANSDUCTION
 Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Cell Signaling part 2
 15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
Insulin Resistance Due to Phosphorylation of. IRS-1 at Serine 302
 JBC Papers in Press. Published on June 14, 2004 as Manuscript M405203200 M3:11187 Insulin Resistance Due to Phosphorylation of IRS-1 at Serine 302 Running Title: Insulin Resistance Due to IRS-1 Phosphorylation
JBC Papers in Press. Published on June 14, 2004 as Manuscript M405203200 M3:11187 Insulin Resistance Due to Phosphorylation of IRS-1 at Serine 302 Running Title: Insulin Resistance Due to IRS-1 Phosphorylation
Insulin Receptor Substrate 3 (IRS-3) and IRS-4 Impair IRS-1- and IRS-2-Mediated Signaling
 and IRS-4 Impair IRS-1- and IRS-2-Mediated Signaling") MOLECULAR AND CELLULAR BIOLOGY, Jan. 2001, p. 26 38 Vol. 21, No. 1 0270-7306/01/$04.00 0 DOI: 10.1128/MCB.21.1.26 38.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Insulin
MOLECULAR AND CELLULAR BIOLOGY, Jan. 2001, p. 26 38 Vol. 21, No. 1 0270-7306/01/$04.00 0 DOI: 10.1128/MCB.21.1.26 38.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Insulin
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression. through inhibition of Hsp90
 Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
The elements of G protein-coupled receptor systems
 The elements of G protein-coupled receptor systems Prostaglandines Sphingosine 1-phosphate a receptor that contains 7 membrane-spanning domains a coupled trimeric G protein which functions as a switch
The elements of G protein-coupled receptor systems Prostaglandines Sphingosine 1-phosphate a receptor that contains 7 membrane-spanning domains a coupled trimeric G protein which functions as a switch
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Insulin Resistance Due to Phosphorylation of Insulin Receptor Substrate-1 at Serine 302*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 34, Issue of August 20, pp. 35298 35305, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Insulin Resistance
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 34, Issue of August 20, pp. 35298 35305, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Insulin Resistance
Insulin Resistance. Biol 405 Molecular Medicine
 Insulin Resistance Biol 405 Molecular Medicine Insulin resistance: a subnormal biological response to insulin. Defects of either insulin secretion or insulin action can cause diabetes mellitus. Insulin-dependent
Insulin Resistance Biol 405 Molecular Medicine Insulin resistance: a subnormal biological response to insulin. Defects of either insulin secretion or insulin action can cause diabetes mellitus. Insulin-dependent
The rabbit femoral artery was prepared and each arterial ring was permeabilized
 Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
The Pleckstrin Homology and Phosphotyrosine Binding Domains of Insulin Receptor Substrate 1 Mediate Inhibition of Apoptosis by Insulin
 MOLECULAR AND CELLULAR BIOLOGY, Nov. 1998, p. 6784 6794 Vol. 18, No. 11 0270-7306/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. The Pleckstrin Homology and Phosphotyrosine
MOLECULAR AND CELLULAR BIOLOGY, Nov. 1998, p. 6784 6794 Vol. 18, No. 11 0270-7306/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. The Pleckstrin Homology and Phosphotyrosine
# This work was supported by grant DK27626 from the National Institutes of. *To whom correspondence should be addressed: Dept. of Molecular & Cellular
 JBC Papers in Press. Published on May 22, 2002 as Manuscript M200367200 3:33 PM 5/14/2002 Construction and Characterization of a Monomeric Insulin Receptor# James Jianping Wu and Guido Guidotti* Department
JBC Papers in Press. Published on May 22, 2002 as Manuscript M200367200 3:33 PM 5/14/2002 Construction and Characterization of a Monomeric Insulin Receptor# James Jianping Wu and Guido Guidotti* Department
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
Diabetes Mellitus and Breast Cancer
 Masur K, Thévenod F, Zänker KS (eds): Diabetes and Cancer. Epidemiological Evidence and Molecular Links. Front Diabetes. Basel, Karger, 2008, vol 19, pp 97 113 Diabetes Mellitus and Breast Cancer Ido Wolf
Masur K, Thévenod F, Zänker KS (eds): Diabetes and Cancer. Epidemiological Evidence and Molecular Links. Front Diabetes. Basel, Karger, 2008, vol 19, pp 97 113 Diabetes Mellitus and Breast Cancer Ido Wolf
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Supplementary Material for
 Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
On Line Data Supplement
 On Line Data Supplement Chemicals and Other Materials [γ- 32 P]ATP, L-[ 35 S]methionine, L-[ 3 H]leucine, m 7 GTP-Sepharose, glutathione- Sepharose 4B and ECL reagents were purchased from Amersham Pharmacia
On Line Data Supplement Chemicals and Other Materials [γ- 32 P]ATP, L-[ 35 S]methionine, L-[ 3 H]leucine, m 7 GTP-Sepharose, glutathione- Sepharose 4B and ECL reagents were purchased from Amersham Pharmacia
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Phosphoserine Detection Kit
 Kit 0701/PSER-KIT 02/080507 Background and Specificity extracellular signals to the nucleus. Phosphorylated epitopes may serve as docking sites for the assembley of protein complexes or may alter the 3-dimensional
Kit 0701/PSER-KIT 02/080507 Background and Specificity extracellular signals to the nucleus. Phosphorylated epitopes may serve as docking sites for the assembley of protein complexes or may alter the 3-dimensional
A high-fructose diet induces changes in pp185 phosphorylation in muscle and liver of rats
 Fructose Brazilian diet Journal induces of Medical changes and in Biological rat pp185 Research () 33: 1421-1427 ISSN -879X Short Communication 1421 A high-fructose diet induces changes in pp185 phosphorylation
Fructose Brazilian diet Journal induces of Medical changes and in Biological rat pp185 Research () 33: 1421-1427 ISSN -879X Short Communication 1421 A high-fructose diet induces changes in pp185 phosphorylation
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Validation & Assay Performance Summary
 Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Translocation of IRS-1 to Cytosol Attenuates Insulin-Stimulated Glucose Transport without Affecting PI3-Kinase Activity
 4 The Open Diabetes Journal, 2, 3, 4-2 Open Access Translocation of IRS- to Cytosol Attenuates Insulin-Stimulated Glucose Transport without Affecting PI3-Kinase Activity Junko Kawahara, Isao Usui *, Tetsuro
4 The Open Diabetes Journal, 2, 3, 4-2 Open Access Translocation of IRS- to Cytosol Attenuates Insulin-Stimulated Glucose Transport without Affecting PI3-Kinase Activity Junko Kawahara, Isao Usui *, Tetsuro
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3076 Supplementary Figure 1 btrcp targets Cep68 for degradation during mitosis. a) Cep68 immunofluorescence in interphase and metaphase. U-2OS cells were transfected with control sirna
DOI: 10.1038/ncb3076 Supplementary Figure 1 btrcp targets Cep68 for degradation during mitosis. a) Cep68 immunofluorescence in interphase and metaphase. U-2OS cells were transfected with control sirna
Enzyme-coupled Receptors. Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors
 Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Perspective: The Insulin Signaling System A Common Link in the Pathogenesis of Type 2 Diabetes
 Perspective: The Insulin Signaling System A Common Link in the Pathogenesis of Type 2 Diabetes Dominic J. Withers and Morris White Endocrinology 2000 141: 1917-1921, doi: 10.1210/en.141.6.1917 To subscribe
Perspective: The Insulin Signaling System A Common Link in the Pathogenesis of Type 2 Diabetes Dominic J. Withers and Morris White Endocrinology 2000 141: 1917-1921, doi: 10.1210/en.141.6.1917 To subscribe
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/10/471/eaah5085/dc1 Supplementary Materials for Phosphorylation of the exocyst protein Exo84 by TBK1 promotes insulin-stimulated GLUT4 trafficking Maeran Uhm,
www.sciencesignaling.org/cgi/content/full/10/471/eaah5085/dc1 Supplementary Materials for Phosphorylation of the exocyst protein Exo84 by TBK1 promotes insulin-stimulated GLUT4 trafficking Maeran Uhm,
Chapter 15: Signal transduction
 Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
FEBS FEBS Letters 410 (1997)
") FEBS 18800 FEBS Letters 410 (1997) 361-367 Inhibition of protein tyrosine phosphatases causes phosphorylation of tyrosine-331 in the p60 TNF receptor and inactivates the receptor-associated kinase Bryant
FEBS 18800 FEBS Letters 410 (1997) 361-367 Inhibition of protein tyrosine phosphatases causes phosphorylation of tyrosine-331 in the p60 TNF receptor and inactivates the receptor-associated kinase Bryant
The Adapter Protein ZIP Binds Grb14 and Regulates Its Inhibitory Action on Insulin Signaling by Recruiting Protein Kinase C
 MOLECULAR AND CELLULAR BIOLOGY, Oct. 2002, p. 6959 6970 Vol. 22, No. 20 0270-7306/02/$04.00 0 DOI: 10.1128/MCB.22.20.6959 6970.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
MOLECULAR AND CELLULAR BIOLOGY, Oct. 2002, p. 6959 6970 Vol. 22, No. 20 0270-7306/02/$04.00 0 DOI: 10.1128/MCB.22.20.6959 6970.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh-
 or GSK3b-specific shrna (sh-") 1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
Cellular Signaling Pathways. Signaling Overview
 Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
A Reciprocal Interdependence between Nck and PI(4,5)P 2 Promotes Localized N-WASp-Mediated Actin Polymerization in Living Cells
P 2 Promotes Localized N-WASp-Mediated Actin Polymerization in Living Cells") Molecular Cell, Volume 36 Supplemental Data A Reciprocal Interdependence between Nck and PI(4,5)P 2 Promotes Localized N-WASp-Mediated Actin Polymerization in Living Cells Gonzalo M. Rivera, Dan Vasilescu,
Molecular Cell, Volume 36 Supplemental Data A Reciprocal Interdependence between Nck and PI(4,5)P 2 Promotes Localized N-WASp-Mediated Actin Polymerization in Living Cells Gonzalo M. Rivera, Dan Vasilescu,
Previous Class. Today. Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism. Protein Kinase Catalytic Properties
 Previous Class Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism Today Protein Kinase Catalytic Properties Protein Phosphorylation Phosphorylation: key protein modification
Previous Class Detection of enzymatic intermediates: Protein tyrosine phosphatase mechanism Today Protein Kinase Catalytic Properties Protein Phosphorylation Phosphorylation: key protein modification
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
JBC Papers in Press. Published on November 10, 2005 as Manuscript M
 JBC Papers in Press. Published on November 10, 2005 as Manuscript M505546200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m505546200 Evidence that Ser 87 of Bim EL is phosphorylated
JBC Papers in Press. Published on November 10, 2005 as Manuscript M505546200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m505546200 Evidence that Ser 87 of Bim EL is phosphorylated
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
The signaling lifetime of protein kinase C (PKC) 4 is under the control of multiple mechanisms. Phosphorylation controls the
 4 is under the control of multiple mechanisms. Phosphorylation controls the") THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 46, pp. 33776 33787, November 16, 2007 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Amplitude Control
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 46, pp. 33776 33787, November 16, 2007 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Amplitude Control
Table S1. Sequence of human and mouse primers used for RT-qPCR measurements.
 Table S1. Sequence of human and mouse primers used for RT-qPCR measurements. Ca9, carbonic anhydrase IX; Ndrg1, N-myc downstream regulated gene 1; L28, ribosomal protein L28; Hif1a, hypoxia inducible factor
Table S1. Sequence of human and mouse primers used for RT-qPCR measurements. Ca9, carbonic anhydrase IX; Ndrg1, N-myc downstream regulated gene 1; L28, ribosomal protein L28; Hif1a, hypoxia inducible factor
Biol403 MAP kinase signalling
 Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Signaling. Dr. Sujata Persad Katz Group Centre for Pharmacy & Health research
 Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis
 Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis") Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Effec<ve Use of PI3K and MEK Inhibitors to Treat Mutant K Ras G12D and PIK3CA H1047R Murine Lung Cancers
 Effec
Effec
Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers
 Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers Amy Grunbeck, Thomas Huber, Pallavi Sachdev, Thomas P. Sakmar Laboratory of Molecular Biology and Biochemistry, The
Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers Amy Grunbeck, Thomas Huber, Pallavi Sachdev, Thomas P. Sakmar Laboratory of Molecular Biology and Biochemistry, The
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10962 Supplementary Figure 1. Expression of AvrAC-FLAG in protoplasts. Total protein extracted from protoplasts described in Fig. 1a was subjected to anti-flag immunoblot to detect AvrAC-FLAG
doi:10.1038/nature10962 Supplementary Figure 1. Expression of AvrAC-FLAG in protoplasts. Total protein extracted from protoplasts described in Fig. 1a was subjected to anti-flag immunoblot to detect AvrAC-FLAG
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Communication. Identification of Methionine N -Acetyltransferase from Saccharomyces cerevisiae
 Communication THE JOURNAL OP BIOLOGICAL CHEMISTRY Vol. 265, No. 7, Issue of March 5, pp. 3603-3606,lSSO 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A. Identification
Communication THE JOURNAL OP BIOLOGICAL CHEMISTRY Vol. 265, No. 7, Issue of March 5, pp. 3603-3606,lSSO 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A. Identification
Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways
![Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways](/thumbs/96/128474991.jpg "Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways") Electronic Supplementary Material (ESI) for MedChemComm. This journal is The Royal Society of Chemistry 2016 Supplementary Data Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic
Electronic Supplementary Material (ESI) for MedChemComm. This journal is The Royal Society of Chemistry 2016 Supplementary Data Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Signal Transduction Pathways Involved in Phosphorylation and
 JBC Papers in Press. Published on March 28, 2001 as Manuscript M009047200 Signal Transduction Pathways Involved in Phosphorylation and Activation of p70 S6K Following Exposure to UVA Irradiation* Yiguo
JBC Papers in Press. Published on March 28, 2001 as Manuscript M009047200 Signal Transduction Pathways Involved in Phosphorylation and Activation of p70 S6K Following Exposure to UVA Irradiation* Yiguo
The clathrin adaptor Numb regulates intestinal cholesterol. absorption through dynamic interaction with NPC1L1
 The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
The clathrin adaptor Numb regulates intestinal cholesterol absorption through dynamic interaction with NPC1L1 Pei-Shan Li 1, Zhen-Yan Fu 1,2, Ying-Yu Zhang 1, Jin-Hui Zhang 1, Chen-Qi Xu 1, Yi-Tong Ma
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
a b G75 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server.
 a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene
 Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene YUELIN ZHANG, WEIHUA FAN, MARK KINKEMA, XIN LI, AND
Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene YUELIN ZHANG, WEIHUA FAN, MARK KINKEMA, XIN LI, AND
Aleksandar Sekulić, 2 Christine C. Hudson, 2 James L. Homme, Peng Yin, Diane M. Otterness, Larry M. Karnitz, and Robert T. Abraham 3 ABSTRACT
 [CANCER RESEARCH 60, 3504 3513, July 1, 2000] A Direct Linkage between the Phosphoinositide 3-Kinase-AKT Signaling Pathway and the Mammalian Target of Rapamycin in Mitogen-stimulated and Transformed Cells
[CANCER RESEARCH 60, 3504 3513, July 1, 2000] A Direct Linkage between the Phosphoinositide 3-Kinase-AKT Signaling Pathway and the Mammalian Target of Rapamycin in Mitogen-stimulated and Transformed Cells
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
MCB*4010 Midterm Exam / Winter 2008
 MCB*4010 Midterm Exam / Winter 2008 Name: ID: Instructions: Answer all 4 questions. The number of marks for each question indicates how many points you need to provide. Write your answers in point form,
MCB*4010 Midterm Exam / Winter 2008 Name: ID: Instructions: Answer all 4 questions. The number of marks for each question indicates how many points you need to provide. Write your answers in point form,
Chapter 11: Enzyme Catalysis
 Chapter 11: Enzyme Catalysis Matching A) high B) deprotonated C) protonated D) least resistance E) motion F) rate-determining G) leaving group H) short peptides I) amino acid J) low K) coenzymes L) concerted
Chapter 11: Enzyme Catalysis Matching A) high B) deprotonated C) protonated D) least resistance E) motion F) rate-determining G) leaving group H) short peptides I) amino acid J) low K) coenzymes L) concerted
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Effects of Second Messengers
 Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
EXPERIMENTAL PROCEDURES
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 11, pp. 8207 8217, March 12, 2010 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Mutual Regulation of
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 11, pp. 8207 8217, March 12, 2010 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Mutual Regulation of
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
INTRODUCTION. Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells
 Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells") Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
phospho-insulin Receptor b Subunit (ptyr 1158 ) ELISA Catalog Number PI0100 Storage Temperature 2-8 C Technical Bulletin
 ELISA Catalog Number PI0100 Storage Temperature 2-8 C Technical Bulletin") phospho-insulin Receptor b Subunit (ptyr 1158 ) ELISA Catalog Number PI0100 Storage Temperature 2-8 C Technical Bulletin Product Description phospho-insulin Receptor β Subunit (ptyr 1158 ) ELISA is a solid
phospho-insulin Receptor b Subunit (ptyr 1158 ) ELISA Catalog Number PI0100 Storage Temperature 2-8 C Technical Bulletin Product Description phospho-insulin Receptor β Subunit (ptyr 1158 ) ELISA is a solid
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
CRISTINA M. RONDINONE*, LING-MEI WANG, PETER LONNROTH*, CHRISTIAN WESSLAU*, JACALYN H. PIERCE, AND ULF SMITH* MATERIALS AND METHODS
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 4171 4175, April 1997 Pharmacology Insulin receptor substrate (IRS) 1 is reduced and IRS-2 is the main docking protein for phosphatidylinositol 3-kinase in adipocytes
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 4171 4175, April 1997 Pharmacology Insulin receptor substrate (IRS) 1 is reduced and IRS-2 is the main docking protein for phosphatidylinositol 3-kinase in adipocytes
Role of Phosphoinositide 3-Kinase in Activation of Ras and Mitogen-Activated Protein Kinase by Epidermal Growth Factor
 Role of Phosphoinositide 3-Kinase in Activation of Ras and Mitogen-Activated Protein Kinase by Epidermal Growth Factor Stefan Wennström and Julian Downward Mol. Cell. Biol. 1999, 19(6):4279. REFERENCES
Role of Phosphoinositide 3-Kinase in Activation of Ras and Mitogen-Activated Protein Kinase by Epidermal Growth Factor Stefan Wennström and Julian Downward Mol. Cell. Biol. 1999, 19(6):4279. REFERENCES
Lecture #27 Lecturer A. N. Koval
 Lecture #27 Lecturer A. N. Koval Hormones Transduce Signals to Affect Homeostatic Mechanisms Koval A. (C), 2011 2 Lipophilic hormones Classifying hormones into hydrophilic and lipophilic molecules indicates
Lecture #27 Lecturer A. N. Koval Hormones Transduce Signals to Affect Homeostatic Mechanisms Koval A. (C), 2011 2 Lipophilic hormones Classifying hormones into hydrophilic and lipophilic molecules indicates
MAPK Pathway
 MAPK Pathway Mitogen-activated protein kinases (MAPK) are proteins that are serine/ threonine specific kinases which are activated by a wide range of stimuli including proinflammatory cytokines, growth
MAPK Pathway Mitogen-activated protein kinases (MAPK) are proteins that are serine/ threonine specific kinases which are activated by a wide range of stimuli including proinflammatory cytokines, growth
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Final Review Sessions. 3/16 (FRI) 126 Wellman (4-6 6 pm) 3/19 (MON) 1309 Surge 3 (4-6 6 pm) Office Hours
 126 Wellman (4-6 6 pm) 3/19 (MON) 1309 Surge 3 (4-6 6 pm) Office Hours") Final Review Sessions 3/16 (FRI) 126 Wellman (4-6 6 pm) 3/19 (MON) 1309 Surge 3 (4-6 6 pm) Office ours 3/14 (WED) 9:30 11:30 am (Rebecca) 3/16 (FRI) 9-11 am (Abel) Final ESSENTIALS Posted Lecture 20 ormonal
Final Review Sessions 3/16 (FRI) 126 Wellman (4-6 6 pm) 3/19 (MON) 1309 Surge 3 (4-6 6 pm) Office ours 3/14 (WED) 9:30 11:30 am (Rebecca) 3/16 (FRI) 9-11 am (Abel) Final ESSENTIALS Posted Lecture 20 ormonal
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Role for the Adaptor Protein Grb10 in the Activation of Akt
 MOLECULAR AND CELLULAR BIOLOGY, Feb. 2002, p. 979 991 Vol. 22, No. 4 0270-7306/02/$04.00 0 DOI: 10.1128/MCB.22.4.979 991.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Role
MOLECULAR AND CELLULAR BIOLOGY, Feb. 2002, p. 979 991 Vol. 22, No. 4 0270-7306/02/$04.00 0 DOI: 10.1128/MCB.22.4.979 991.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Role
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding
 Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding Hong-mei Gu, 1 Ayinuer Adijiang, 1 Matthew Mah, and Da-wei Zhang 2 Departments of Pediatrics and Biochemistry,
Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding Hong-mei Gu, 1 Ayinuer Adijiang, 1 Matthew Mah, and Da-wei Zhang 2 Departments of Pediatrics and Biochemistry,
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
Revisiting post-translational regulation of cell cycle CDKs
 Revisiting post-translational regulation of cell cycle CDKs Cyclin-CDK complexes during cell cycle Cyclin B-CDK1 G0 E2F prb Cyclin A-CDK1 G2 M Cyclin D-CDK4 CDK6 + prb Cyclin A-CDK2 S R G1 Cyclin E-CDK2
Revisiting post-translational regulation of cell cycle CDKs Cyclin-CDK complexes during cell cycle Cyclin B-CDK1 G0 E2F prb Cyclin A-CDK1 G2 M Cyclin D-CDK4 CDK6 + prb Cyclin A-CDK2 S R G1 Cyclin E-CDK2