Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis
|
|
|
- Shanon Reed
- 6 years ago
- Views:
Transcription
1 The FASEB Journal article fj Published online June 5, 25. The FASEB Journal Research Communication Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis Nan Li, Dolores D. Mruk, Chris K. C. Wong, Will M. Lee, Daishu Han, and C. Yan Cheng, The Mary M. Wohlford Laboratory for Male Contraceptive Research, Center for Biomedical Research, Population Council, New York, New York, USA; Department of Biology, Hong Kong Baptist University, Hong Kong, China; School of Biological Sciences, University of Hong Kong, Hong Kong, China; and Institute of Basic Medical Sciences, Chinese Academy of Medical Sciences, School of Basic Medicine, Peking Union Medical College, Beijing, China ABSTRACT Ectoplasmic specialization (ES) is an actinrich adherens junction in the seminiferous epithelium of adult mammalian testes. ES is restricted to the Sertolispermatid (apical ES) interface, as well as the Sertoli cellcell (basal ES) interface at the blood-testis barrier (BTB). ES is typified by the presence of an array of bundles of actin microfilaments near the Sertoli cell plasma membrane. These actin microfilament bundles require rapid debundling to convert them from a bundled to branched/ unbundled configuration and vice versa to confer plasticity to support the transport of ) spermatids in the adluminal compartment and 2) preleptotene spermatocytes at the BTB while maintaining cell adhesion. is one of the plastin family members abundantly found in yeast, plant and animal cells that confers actin microfilaments their bundled configuration. Herein, plastin 3 was shown to be a component of the apical and basal ES in the rat testis, displaying spatiotemporal expression during the epithelial cycle. A knockdown (KD) of plastin 3 in Sertoli cells by RNA interference using an in vitro model to study BTB function showed that a transient loss of plastin 3 perturbed the Sertoli cell tight junction-permeability barrier, mediated by changes in the localization of basal ES proteins N- cadherin and b-catenin. More importantly, these changes were the result of an alteration of the actin microfilaments, converting from their bundled to branched configuration when examined microscopically, and validated by biochemical assays that quantified actin-bundling and polymerization activity. Moreover, these changes were confirmed by studies in vivo by plastin 3 KD in the testis in which mislocalization of N-cadherin and b-catenin was also detected at the BTB, concomitant with defects in the transport of spermatids and phagosomes and a disruption of cell adhesion most notably in elongated spermatids due to Abbreviations: Arp3, actin-related protein 3; BTB, blood-testis barrier; b.w., body weight; co-ip, coimmunoprecipitation; CTC, circulating tumor cell; Eps8, epidermal growth factor receptor pathway substrate 8; ES, ectoplasmic specialization; F2/DMEM, Ham s F2 Nutrient Mixture/DMEM (:, vol/ vol);, filamentous actin; FAK, focal adhesion kinase; (continued on next page) a loss of actin-bundling capability at the apical ES, which in turn affected localization of adhesion protein complexes at the site. In summary, plastin 3 is a regulator of actin microfilament bundles at the ES in which it dictates the configuration of the filamentous actin network by assuming either a bundled or unbundled/branched configuration via changes in its spatiotemporal expression during the epithelial cycle. Li, N., Mruk, D. D., Wong, C. K. C., Lee, W. M., Han, D., Cheng, C. Y. Actinbundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis. FASEB J. 29, (25). Key Words: actin microfilaments blood-testis barrier spermatid adhesion DURING SPERMATOGENESIS, EXTENSIVE REMODELING TAKES place at the Sertoli-germ cell interface to facilitate germ cell transport, in particular spermatids during spermiogenesis, across the seminiferous epithelium (). Thus, spermatids can be transported across the adluminal compartment while differentiating into spermatozoa that are then lined up at the edge of the tubule lumen at late-stage VIII of the epithelial cycle in rodents to prepare for spermiation (2 4). Similarly, restructuring of the Sertoli cell-cell interface also takes place at the blood-testis barrier (BTB) near the basement membrane to accommodate the transport of preleptotene spermatocytes across the barrier at stage VIII of the cycle (4 6). Thus, the timely occurrence of these events is essential for spermatogenesis. In the testis, there is a unique actin-based cell-cell anchoring junction known as ectoplasmic specialization (ES) at the Sertolispermatid and Sertoli cell-cell interface known as apical ES and basal ES, respectively (7 9). Unlike other anchoring junctions, ES is typified by the presence of an array of actin microfilament bundles that lie perpendicular to the Sertoli Correspondence: The Mary M. Wohlford Laboratory for Male Contraceptive Research, Center for Biomedical Research, Population Council, 23 York Ave., New York, NY 65, USA. y-cheng@popcbr.rockefeller.edu doi:.96/fj /5/29- FASEB
2 cell plasma membrane, sandwiched between the cisternae of endoplasmic reticulum and the apposing Sertolispermatid (apical ES) or Sertoli-Sertoli (basal ES) plasma membranes (8, ). Basal ES coexists with tight junction (TJ) and gap junction, and together with desmosome, they constitute the BTB (5, 8). The presence of these actin microfilament bundles at the ES also confers the ES its unusual adhesive strength (). Interestingly, the occurrence of germ cell transport as well as endocytic vesiclemediated trafficking events (e.g., endocytosis, transcytosis, and recycling) pertinent to spermatogenesis requires rapid reorganization of these microfilament bundles so that they can be efficiently converted from their bundled to unbundled/branched configuration and vice versa. Thus, it is envisioned that the dynamic changes of the actin microfilament bundles are conferred by the spatiotemporal expression of actin-bundling proteins, branched actin polymerization proteins, and depolymerization-inducing proteins during the epithelial cycle of spermatogenesis (3, 2). In order to better understand the molecular mechanism that regulates the dynamic nature of microfilament bundles at the ES, we sought to examine the role of an actinbundling protein, plastin 3, in the testis. Plastin, also known as fimbrin, is an actin-bundling proteinfamily,involvedintheformationoffilamentous actin () bundles in mammalian cells. This family consists of 3 known members that are evolutionarily conserved from yeast to plant and mammalian cells called plastin (I-plastin), plastin 2 (L-plastin), and plastin 3 (T-plastin), with a Ca 2 -binding site near the N terminus (3 5). There is ;75% sequence similarity among all 3 plastins in mouse and human (3, 5). Plastins are ;65 68 kda proteins in which plastin is predominantly found in small intestine, colon, and kidney. Plastin 2 is mostly expressed by hemopoietic cell lineages, such as leukocytes, and tumor cells. is highly expressed in normal cells ofsolidtissuessuchasneurons, fibroblasts, endothelial cells, epithelial cells, and melanocytes. Mutation defects of plastin 3 in humans are associated with osteoporosis and bone fracture (6), possibly due to defects in actin filament bundles in bone cells, thereby impeding the conversion of mechanical signals to intracellular biochemical signals in osteoblasts and osteoclasts (7), causing osteoporosis (8). The best-studied plastin is plastin 2 in immune cells (3, 4, 9, 2). For instance, plastin 2 is known to be involved in T- cell motility and activation (9), and plastin 2-deficient neutrophils failed to kill bacterial pathogens (2) because these cells lacked the ability to quickly rearrange their actin cytoskeleton in response to the pathogens. Studies have shown that phosphorylation of plastin 2 at Ser5 is involved in inducing adhesion in neutrophils (22), and the actinbundling activity of plastin 2 is an intracellular Ca 2 -dependent event (23). However, the function of plastins in the testis remains virtually unknown because there is no report in the literature on this subject except an earlier article with SDS-PAGE analysis supporting the presence of (continued from previous page) IB, immunoblotting; IF, immunofluorescence microscopy; IP, immunoprecipitation; KD, knockdown; qpcr, quantitative PCR; RNAi, RNA interference; sirna, small interfering RNA; SMA, spinal muscular atrophy; TER, transepithelial electrical resistance; TJ, tight junction; ZO-, zonula occludens plastin (fimbrin) in the rat testis (24) almost 26 years ago. Herein, we demonstrated that plastin 3 is restrictively expressed by Sertoli cells, but not germ cells, in the seminiferous epithelium. We also examined the likely involvement of plastin 3 in ES dynamics in light of its intrinsic actin-bundling activity in mammalian cells. MATERIALS AND METHODS Animals and antibodies Male Sprague-Dawley rats used for different experiments reported herein were purchased from Charles River Laboratories (Kingston, NY, USA). The use of animals was approved by the Rockefeller University Institutional Animal Care and Use Committee with Protocol Number Primary Sertoli cell cultures Sertoli cells were isolated from 2-d-old rat testes as earlier described (25). Sertoli cells were cultured in serum-free Ham s F2 Nutrient Mixture/DMEM (:, vol/vol) (F2/DMEM; Sigma-Aldrich, St. Louis, MO, USA) supplemented with growth factors and gentamicin at 35 C with 95% air-5% CO 2 (vol/vol) in a humidified atmosphere. Cells were seeded at a density of cells/cm 2 on 8 mm (diameter) round cover glasses (placed in 2-well dishes with 2 ml F2/DMEM per well; for immunofluorescence analysis), cells/cm 2 in 6-well dishes (with 5 ml F2/DMEM per well; for lysate preparation and RNA isolation), or cells/cm 2 on Millicell cell culture inserts (HA type, mixed cellulose esters, 2 mm diameter, effective surface area ;.6 cm 2,.45 mm pore size; EMD Millipore, Billerica, MA, USA) for transepithelial electrical resistance (TER) measurements (with.5 ml F2/DMEM in the apical and basal chamber) to monitor the TJ-permeability barrier function. About 36 h thereafter, cultures were subjected to a brief hypotonic treatment using 2 mm Tris (ph 7.4) at 22 C for 2.5 min as described (26) to lyse residual germ cells, and Sertoli cells were rinsed twice with F2/DMEM to remove cellular debris. These cultures were ;98% pure, with negligible contaminations of Leydig cells, peritubular myoid cells, or germ cells using specific markers for these cells by either immunoblotting (IB) or RT-PCR as described (27). All dishes or cover glasses were coated with Matrigel (BD Biosciences, San Jose, CA, USA) diluted with F2/ DMEM at :7 (for bicameral inserts, a dilution of :5 was used) (25). After Sertoli cells were cultured for ;2 d, a functional TJpermeability barrier was established, and ultrastructures of TJ, basal ES, gap junction, and desmosome were detected by electron microscopy that mimic the Sertoli cell BTB in vivo (28 3). In fact, this in vitro system was widely used by investigators in the field to study Sertoli cell BTB function (3 35), and these results were subsequently reproduced in studies in vivo (34, 36, 37), illustrating the physiologic relevancy of this in vitro system. Knockdown of plastin 3 in primary Sertoli cells in vitro by RNA interference Sertoli cells were cultured in vitro for 3 d to allow the establishment of a functional TJ-permeability barrier that mimic the Sertoli cell BTB in vivo, containing ultrastructures of TJ, basal ES, gap junction, and desmosome as earlier described (25, 29, 3). Thereafter, cells were transfected with plastin 3-specific small interfering RNA (sirna) duplexes vs. nontargeting negative control sirna duplexes at nm [for immunofluorescence microscopy (IF)andIB]or5nM(forTJ-barrierfunctionassessment)using 2 Vol. 29 September 25 The FASEB Journal x LI ET AL.
3 Lipofectamine RNAiMAX Reagent (Life Technologies, Norwalk, CT, USA) as transfection medium. The desired concentrations of sirna duplexes for different experiments were selected based on results of pilot experiments that yielded detectable phenotypes without detectable cytotoxicity based on an XTT (sodium 39-[- (phenylaminocarbonyl)-3,4-tetrazolium]-bis (4-methoxy-6-nitro) benzene sulfonic acid hydrate) assay (Cell Proliferation Kit II, Roche Life Sciences, Branford, CT, USA) as described (29). sirna duplexes that specifically targeted plastin 3 were obtained from Ambion (Austin, TX, USA): sense, 59-GCCUAUUUCCAU- CUACUCAtt-39, antisense,59-ugaguagauggaaauaggctt-39 (s3565); sense, 59-CACCCUUCAUCAUUCAGGAtt-39,antisense, 59-UCCUGAAUGAUGAAGGGUGta-39 (s35652); and sense, 59- CCUCUUUAAUAAAUAUCCAtt-39, antisense,59-uggauauuu- AUUAAAGAGGtt-39 (s35653). However, only s3565 sirna duplexes were used in all subsequent experiments because pilot experiments had demonstrated that the efficacy of s35652 and s35653 in silencing plastin 3 in Sertoli cells was 5% vs. 7% obtained with s3565 sirna duplexes. Nontargeting sirna duplex (Silencer Select Negative Control # sirna; Ambion) that served as a negative control was included in all experiments, which was comprised of a 9 bp nontargeting sequence with 3 dt overhangs, bearing no significant homology to any known human, mouse, or rat gene sequences as indicated by the manufacturer, which also failed to induce gross changes in gene expression in transfected Sertoli cells as noted in our studies herein. After transfection for 24 h, cells were washed twice and cultured with fresh F2/DMEM including supplements for an additional 24 h before termination to be used for IB, IF, and biochemical assays. For fluorescence microscopy, cells were cotransfected with nm siglo Red Transfection Indicator (Dharmacon, GE Healthcare Life Sciences, Lafayette, CO, USA) to track successful transfection. Assessment of TJ-permeability barrier in vitro Sertoli cell TJ-permeability barrier that assessed the barrier integrity was monitored by quantifying TER across the cell epithelium on the bicameral unit using a Millicell electrical resistance system (EMD Millipore) as described (25). Each time point had quadruplicate culture inserts, and each experiment was repeated 3 times using different batches of Sertoli cells. Actin-bundling assay Actin-bundling assay was prepared as earlier described (38) with some modifications. was first obtained using an Actin Binding Protein Biochem Kit (catalog #BK3; Cytoskeleton, Denver, CO, USA) as follows. In brief, purified nonmuscle actin was reconstituted to mg/ml at 4 C with sterile water. A total of 25 ml of this actin solution was diluted to mg/ml with 225 ml general actin buffer [5 mm Tris-HCl,.2 mm CaCl 2, and.2 mm ATP (ph 8.)]. The actin solution was then placed on ice for 3 min to obtain G-actin, then supplemented with 25 ml actin polymerization buffer [5 mm KCl, 2 mm MgCl 2, mm Tris, and mm ATP (ph 7.5)] and incubated at room temperature (22 C) for h to allow the assembly of from G-actin, which wasthenusedasthesubstratetoassesstheabilityofsertolicell lysate to induce actin microfilament bundling. Sertoli cell lysates were first obtained from cells previously transfected with plastin 3-specific sirna duplexes vs. nontargeting negative control duplexes using a Tris lysis buffer [2 mm Tris, 2 mm NaCl, and.5% Triton X- (vol/vol) (ph 7.5) at 22 C, freshly supplemented with protease and phosphatase inhibitor cocktails (Sigma- Aldrich) at a : dilution (vol/vol)], and cellular debris was removed by centrifugation at 2,8 g at 4 C for h. A total of ml Sertoli cell lysates (;4 5 mg protein) from both groups (clear supernatant from the above step) containing equal amounts of proteins (based on protein estimation using a Bio-Rad DC Protein Assay kit; Hercules, CA, USA) vs. ml Tris lysis buffer (served as a negative control) was then added into 4 ml freshly prepared obtained above. This mixture was incubated for h at room temperature to allow actin bundling, centrifuged at 4, g for 5 min at 24 C to sediment bundled in the pellet, whereas the linear and unbundled actin microfilaments remain in the supernatant. The whole pellet and 5 ml supernatant of each sample including both groups vs. negative control were analyzed by immunoblot using a specific b-actin antibody. A total of 4 mg of the same cell lysate from each sample was also analyzed by immunoblot to monitor the efficiency of plastin 3 knockdown (KD) and to serve as a protein loading control. This experiment was repeated 5 times using different cell preparations. Actin polymerization assay Actin polymerization assay that also assessed the kinetics of actin polymerization was performed as earlier described (39, 4). Effects of plastin 3 KD vs. nontargeting control on actin polymerization were assessed through the initial rate of fluorescence increase that occurs during pyrene-conjugated G-actin incorporation into existing actin microfilaments using Actin Polymerization Biochem Kits from Cytoskeleton. In brief, on d 5 (i.e., 2 d after transfection), Sertoli cells were lysed in Tris lysis buffer in which cellular debris was removed by centrifugation at 2,8 g at 4 C for h. The clear supernatant that contained lysates from Sertoli cells following plastin 3 KD vs. control was immediately subjected to actin polymerization assay as described (39, 4). Cell lysates (;3 ml) from control and plastin 3-silenced groups with equal amounts of protein (;2 5 mg) were added to the final reaction mix( ml) containing 6 ml pyrene-conjugated G-actin stock and ml of 3 actin polymerization buffer. The kinetics of fluorescence increase were monitored in a Corning 96 well black flat bottom polystyrene microplate (via top reading; Corning, Lowell, MA, USA) by enhanced fluorescence emission at nm as described (39) using a FilterMax F5 Multi-Mode Microplate Reader and the Multi-Mode Analysis Software (Molecular Devices, Sunnyvale, CA, USA). A kinetics reading was taken at room temperature every 3 s for cycles with an excitation filter/emission filter at 36/43 nm, and 5 ms integration time was used. The initial rate of actin polymerization ( 2 min) assessed by the rate of increase in fluorescence intensity was obtained by linear regression analysis using Microsoft Excel (Redmond, WA, USA). Each sample was run in duplicate, and this experiment was repeated 3 times, excluding pilot experiments that yielded similar results. silencing in adult rat testis in vivo To knock down plastin 3 in vivo, rats[; g body weight (b.w.)] were transfected with sirna duplexes via intratesticular injection using a 28 gauge needle of an insulin syringe on d, to be followed by a second transfection on d as earlier described (4, 42). One testis of a rat received nontargeting negative control sirna duplexes, whereas the other testis received plastin 3- specific sirna duplexes shown to perturb the Sertoli cell TJ barrier in vitro. sirna duplexes at desired concentration ( nm) were constituted in a transfection mix containing 3 ml Lipofectamine RNAiMAX sirna transfection reagent and 93 ml Opti- MEM Reduced Serum Medium (Invitrogen, Life Technologies) to a final volume of 2 ml. Thus, each testis (;.6 g, with a volume of ;.6 ml) of an adult rat received 2 ml ofthistransfection solution.inshort,the28gauge (3 mm long) needle attached to a ml insulin syringe containing the transfection mix was inserted from the apical to near the basal end of the testis vertically. As the needle was withdrawn apically, transfection mixture was released gently and gradually from the syringe so that the entire testis was slowly filled with the sirna mixture that spread throughout the PLASTIN 3 REGULATES ES DYNAMICS 3
4 TABLE. Antibodies used for different experiments in this report Antibody Host species Vendor Catalog number Working dilution IB IF Rabbit Abcam ab37585 :5 Mouse Santa Cruz Biotechnology sc-6628 : Arp3 Mouse Sigma-Aldrich A5979 :3 :5 Eps8 Mouse BD Biosciences 643 :5 :5 N-cadherin Rabbit Santa Cruz Biotechnology sc-7939 :2 N-cadherin Mouse Invitrogen, Life Technologies : Actin Goat Santa Cruz Biotechnology sc-66 :3 a-catenin Rabbit Santa Cruz Biotechnology sc-7894 :2 a-catenin Mouse Invitrogen, Life Technologies 3-97 : b-catenin Rabbit Invitrogen, Life Technologies 7-27 :25 : Claudin- Rabbit Invitrogen, Life Technologies :5 JAM-A Rabbit Invitrogen, Life Technologies 36-7 :25 Laminin-g3 Rabbit C.Y.C. lab Reference (43) :5 Occludin Rabbit Invitrogen, Life Technologies 7-5 :25 ZO- Rabbit Invitrogen, Life Technologies 6-73 :25 : Vimentin Mouse Santa Cruz Biotechnology sc-626 :3 : Vimentin Rabbit Cell Signaling Technology 3932 :5 Nectin-3 Rabbit Santa Cruz Biotechnology sc :5 Nectin-3 Goat Santa Cruz Biotechnology sc-486 :2 b-integrin Rabbit Santa Cruz Biotechnology sc-8978 :2 :5 Formin Mouse Abcam ab6858 :5 Palladin Rabbit Proteintech 853--AP : :5 Filamin A Mouse Abcam ab8837 :5 CAR Rabbit Santa Cruz Biotechnology sc-545 :2 : p-fak-tyr 397 Rabbit Invitrogen, Life Technologies G :5 GAPDH Mouse Abcam ab824 : Abcam, Cambridge, MA, USA; Santa Cruz Biotechnology, Santa Cruz, CA, USA; Cell Signaling Technology, Danvers, MA, USA; Proteintech, Chicago, IL, USA. CAR, coxsackievirus and adenovirus receptor; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. testis to avoid an acute rise in intratesticular hydrostatic pressure. Rats were euthanized on d 3 (i.e., 2 d after the second transfection) by slow (2 3%/min) displacement of chamber air with compressed CO 2 via a CO 2 tank with a desired regulator. Pilot experiments showed that the phenotypes were similar when rats were terminated on d 3 vs. d 4; thus, animals euthanized on d 3 andd4werepooledforanalysiswith6rats.forrnaextraction (for RT-PCR), lysate preparation (for IB), and fluorescence analysis, testes were snap frozen in liquid nitrogen and stored at 28 C until used. Some testes (n =3)werefixed in Bouin s fixative to obtain paraffin sections for histologic analysis using hematoxylin and eosin staining. IB, coimmunoprecipitation, RT-PCR, and quantitative PCR Lysates of Sertoli cells, germ cells, and testes were obtained by using immunoprecipitation (IP) lysis buffer [ mm Tris,.5 M NaCl, % NP-4 (vol/vol), and % glycerol (vol/vol) (ph 7.4) at 22 C] freshly supplemented with protease and phosphatase inhibitor cocktails at a : dilution (vol/vol) as described (39, 42). Immunoblot analysis was performed as earlier described (39) using corresponding specific antibodies (Table ). Chemiluminescence was performed using a homemade kit in our laboratory as described (44). Coimmunoprecipitation (co-ip) was performed using lysates (;6 mg protein) from testes as described (39, 42) to assess the binding partners of plastin 3. RNA extraction, PCR, and quantitative PCR (qpcr) using specific primer pairs (Table 2) were performed as described (45). The authenticity of PCR products was verified by direct DNA sequencing performed at Genewiz (South Plainfield, NJ, USA). qpcr was performed at The Rockefeller University Genomics Resource Center (New York, NY, USA). Dual-labeled immunofluorescence analysis Dual-labeled immunofluorescence analysis was performed using frozen cross sections of testes and specific primary antibodies and corresponding secondary antibodies (Table ). Negative controls were performed using normal IgG of either mouse or rabbit (depending on the source of primary antibody) or the omission of secondary antibody, which yielded no detectable staining, illustrating the specificity of the antibody. was stained by rhodamine-conjugated phalloidin as described (46). Cell nuclei were visualized by DAPI (Invitrogen, Life Technologies). Fluorescence images were obtained in an Olympus BX6 fluorescence microscope with images captured using the built-in Olympus DP- 7 digital camera (Tokyo, Japan). Image files were analyzed using Photoshop in Adobe Creative Suite (version 3.; San Jose, CA, USA) such as for image overlay to assess protein colocalization. Confocal microscopy Confocal microscopy was performed at the Rockefeller University Bio-Imaging Resource Center (New York, NY, USA) as earlier described (47, 48). It was used to examine changes in the distribution of basal ES protein N-cadherin vs. TJ protein zonula occludens (ZO-) at the Sertoli cell-cell interface following plastin 3 KD by RNA interference (RNAi). Sertoli cells were cultured at cells/cm 2 on Matrigel-coated Transwell Permeable Supports (Costar polyester membrane inserts, 24 mm diameter,.4 mm pore size; Corning) for 3 d and transfected with plasmin 3-specific sirna duplexes vs. nontargeting control duplexes for 24 h, thereafter, cells were washed twice and allowed to culture for an additional day before being fixed in paraformaldehyde and processed for confocal microscopy including 4 Vol. 29 September 25 The FASEB Journal x LI ET AL.
5 TABLE 2. Primers used for different RT-PCR experiments in this report Gene GenBank accession number Primer sequence (59 39) Nucleotide position Expected size (bp) Plastin NM_878.2 Sense: TGAACCAGACACGATTGACG Antisense: TGATGATCTGCCAGAGAAGC Plastin 2 NM_244. Sense: GTCACAGCCACTGATGTTGT Antisense: GTCGGATAAGTCGCTGTACA NM_384. Sense: CTGAATTCTGCTTCTGCC Antisense: CAGTGCTTCATTCCTGCT S-6 NM_6946. Sense: TCCGCTGCAGTCCGTTCAAGTCTT Antisense: GCCAAACTTCTTGGATTCGCAGCG staining with primary and secondary antibodies as described (47). Images were obtained using an inverted Zeiss LSM 78 laserscanning confocal microscope (Carl Zeiss MicroImaging, Thornwood, NY, USA) equipped with multiple laser lines (diode, 45 nm; argon/2, 458, 477, 488, and 54 nm; HeNe, 543 nm; and HeNe2, 633 nm) and the Zeiss LSM 78 software package. Optical sections,.8 mm were collected at.4 mm intervals along the z axis to obtain a series of images (Z-stack). Image deconvolution of Z-stacks was performed using 3D Huygens Deconvolution Software (Scientific Volume Imaging, Hilvesum, The Netherlands) as described (48, 49). Data reported herein were representative findings of an experiment that we repeated 3 times that yielded similar results. Image analysis To assess changes in protein localization in Sertoli cells following in vitro plastin 3 KD, at least 2 cells were randomly selected and examined in control vs. experimental groups with 3 experiments. Fluorescence intensity of a target protein in Sertoli cells or in the seminiferous epithelium of testes following in vivo silencing was quantifiedusingimagej.45(nih,bethesda,md,usa; nih.gov/ij) as earlier described (4). At least 2 randomly selected stage VII vs. stage VIII tubules from cross sections of a rat testis were examined with 3 rats (a total of ;6 tubules). Analysis was focused on these 2 stages because plastin 3 restrictively expressed at the apical ES in stage VII tubules, and defects in spermatid adhesion and transport that rendered stage VII to become VIII tubules that compromised spermiation were readily detected in stage VIII tubules. Statistical analysis For studies using Sertoli cell cultures, triplicate coverslips, dishes, or bicameral units were used. Each data point or bar graph is amean6 SD of 3 5 experiments (or n = 5 rats). Statistical analysis was performed using the GB-STAT software package (version 7.; Dynamic Microsystems, Silver Spring, MD, USA) by -way ANOVA, followed by Dunnett s test. In selected experiments, Student s t test was used for paired comparisons. RESULTS is expressed stage specifically by Sertoli cells in the seminiferous epithelium of adult rat testes during the epithelial cycle Using primer pairs specific to plastin, 2, and 3 (Table 2) for RT-PCR, it was shown that plastin and 2 were expressed by both Sertoli and germ cells, but plastin 3 was expressed only by Sertoli cells, but not germ cells, in the seminiferous epithelium (Fig. A). A specific anti-plastin 3 antibody (Table ) illustrated by immunoblot analysis using lysate of Sertoli cells (Fig. A) was then used for our studies. Using lysates of testes and germ cells vs. Sertoli cells and a specific plastin 3 antibody (Fig. B and Table ) for immunoblot analysis, plastin 3 was found to be expressed by Sertoli cells, but not germ cells (Fig. C, upperpanel), consistent with RT-PCR data shown in Fig. A and qpcr analysis shown in Fig. C (lower panel). There was a timedependent expression of plastin 3 by Sertoli cells when these cells were cultured in vitro during the assembly of a functional TJ-permeability barrier (Fig. D). When Sertoli cells were cultured alone for 4 d, plastin 3 was found to partially colocalize with actin microfilaments (Fig. E). The pattern of plastin 3 distribution in Sertoli cells was not uniform across all cells as shown in Fig. E, plausibly the resultofthecyclicnatureofthesecellsbecausetheywere isolated from tubules of different stages. Besides Sertoli cells, Leydig cells also expressed plastin 3 in the interstitial space in the seminiferous epithelium (Fig. F). The expression of plastin 3 by Sertoli cells is most predominant at the -rich ultrastructure the basal ES/BTB near the basement membrane and the apical ES at the Sertolispermatid interface (Fig. G). Although plastin 3 was detected at the basal ES/BTB (annotated by yellow arrowheads in Fig. G) in virtually all stages of the epithelial cycle with similar intensity such as at stages V VII, its expression considerably diminished at stage VIII. Interestingly, its expression at the apical ES was highly stage specific and limited to VII tubules since it was weakly expressed at the apical ES in V VI tubules and not detectable at any other stages (Fig. G). also partially colocalized with at both apical and basal ES (Fig. G). is associated with ES in adult rat testes At the basal ES/BTB near the basement membrane, plastin 3 was found to colocalize, at least in part, with integral membrane proteins of TJ (e.g., claudin-) or basal ES (e.g., N-cadherin), as well as TJ adaptor (e.g., ZO-) and also vimentin and in stage VII tubules (Fig. 2A). also partially colocalized with integral membrane protein and nectin-3 (but not b-integrin), as well as at the apical ES (Fig. 2B). Although plastin 3 colocalized in part with basal ES (Fig. 2A) and apical ES (Fig. 2B) proteins, a study by co-ip illustrated that plastin 3 only structurally interacts with actin and vimentin, but not constituent proteins nor actin regulatory proteins at the apical and/or basal ES (Fig. 2C), such as those shown in Fig. 2A, B. This is not entirely unexpected because basal ES protein PLASTIN 3 REGULATES ES DYNAMICS 5




 M 653 57 453 394 298 234")

 Relative expression of plastin, plastin 2,")
, and germ cells (GC) vs.")
, spleen (SP), and liver (L) that")


 Specificity of the anti-plastin 3 antibody")
 vs.")

 Steady-state protein level of plastin 3 in")
 was examined by IB with b-actin")
6 T SC GC Plastin, 88 bp bp M T SC GC C -Actin, 42 kda 3 - S6, 385 bp Plastin 2, 29 bp M SC GC L -, 68 kda S6, 385 bp D 34 -, 53 bp qpcr Analysis 3 2 T SC GC , 68 kda - Actin, 42 kda F /DAPI 4 Time of Sertoli cells in culture (day) Actin, 42 kda E /DAPI GC -, 68 kda 95 - T SC 7 - SP T SC G C 3T 3 S6, 385 bp bp B SI Relative plastin 3 mrna level (Arbitrary unit) M Mr x -3 bp A /DAPI Late VII IX VII G VII VI V VIII Figure. Expression of plastins in the rat testis. A) Relative expression of plastin, plastin 2, and plastin 3 in adult rat testes (T), Sertoli cells (SC), and germ cells (GC) vs. the corresponding positive controls of small intestine (SI), spleen (SP), and liver (L) that are known to express these plastins was analyzed by RT-PCR using specific primer pairs (Table 2). S-6 served as a loading and PCR control. M, DNA markers in base pair (bp). B) Specificity of the anti-plastin 3 antibody (Table ) was assessed by IB using lysates of Sertoli cells (SC) vs. germ cells (GC) and NIH 3T3 cells (mouse embryonic fibroblast cell line, positive control). C ) Steady-state protein level of plastin 3 in adult rat testes (T), Sertoli cells (SC), and germ cells (GC) was examined by IB with b-actin serving as a protein loading control (upper panel) vs. steady-state mrna level by qpcr (lower panel). Each bar in the (continued on next page) 6 Vol. 29 September 25 The FASEB Journal x LI ET AL.
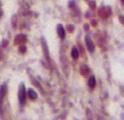
7 N-cadherin (a basal ES protein), whereas it colocalizes with occludin (a TJ protein) at the BTB, does not display any structural interaction with occludin by co-ip (5). KD in Sertoli cells with an established TJ barrier by RNAi perturbs the TJ-permeability barrier via mis-localization of basal ES proteins Using sirna duplexes specific to plastin 3 vs. nontargeting negative control duplexes to knock down plastin 3 in Sertoli cells, plastin 3 was selectively silenced without affecting the expression of either plastin or plastin 2 when RNAs extracted from these cells were used for RT-PCR to examine the steady-state mrna levels of all 3 plastins (Fig. 3A). These findings were confirmed by immunoblot analysis when the steady-state protein level of plastin 3 was quantified in plastin 3 KD cells vs. nontargeting negative control cells, illustrating an ;7% KD (Fig. 3B). Furthermore, the expression of several integral membrane proteins, adaptors, actin regulatory proteins, and cytoskeletal proteins at the BTB was also not affected, illustrating that there was no apparent off-target effects (Fig. 3B). When plastin 3 was knocked down by ;7%, the Sertoli TJpermeability barrier was found to be perturbed (Fig. 3C), likely the result of a loss of actin filament-bundling ability conferred by plastin 3 at the Sertoli cell barrier. It was noted that a KD of plastin 3 by ;7% based on IF led to changes in the distribution of basal ES proteins (e.g., N-cadherin and b-catenin) at the Sertoli cell-cell interface, but not TJ proteins (claudin- and ZO-). Both N-cadherin and b-catenin were no longer tightly restricted to the Sertoli cell-cell interface; instead, they were diffusely localized at the site in plastin 3 KD cells vs. control cells (Fig. 3D, see white brackets), likely due to an increase in protein endocytosis. These changes in localization were summarized semiquantitatively in histograms shown in Fig. 3D (see right panel), suggesting that the plastin 3 KDinduced TJ-barrier disruption is mediated by changes in localization of basal ES proteins at the BTB. These observations were further supported by a study using confocal microscopy that assessed changes in the localization of basal ES protein N-cadherin vs. TJ protein ZO- in both x-y (horizontal view) and x-z (vertical view) planes (Fig. 4A). Changes in N-cadherin localization following plastin 3 KD were semiquantitatively assessed by averaging the width of green fluorescence (N-cadherin) on 4 sides of a Sertoli cell in plastin 3-silenced cells at the x-z plane vs. control cells from 3 independent experiments (Fig. 4B), supporting findings shown in Fig. 3D. KD perturbs organization at the Sertoli cell BTB mediated by a disruption of intrinsic actin-bundling and actin polymerization activity Because basal ES is an actin microfilament-rich adherens junction (8, ) and plastin 3 is an actin-bundling protein, we next investigated changes in organization in Sertoli cells following plastin 3 KD in Sertoli cells. As noted in control Sertoli cells, actin microfilaments appeared as a properly organized network, analogous to bundles of (Fig. 5A). Following plastin 3 KD, however, extensively branched actin microfilaments were detected, but vimentin-based intermediate filaments were not affected (Fig. 5A). It was also noted that whereas the level of epidermal growth factor receptor pathway substrate 8 (Eps8; an actin barbed end capping and bundling protein) and actin-related protein (Arp; a branched actin nucleation protein, effectively converting bundled actin microfilaments to an unbundled/branched configuration) 3 was unaffected following plastin 3 KD (Fig. 3B), the localization of both Eps8 and Arp3 was perturbed in which these actin regulatory proteins were relocated to the cell cytosol, no longer found conspicuously at or near the Sertoli cell-cell interface (Fig. 5A) to support the proper organization of F- actin at or near the TJ barrier. Based on a biochemical actin-bundling assay, the KD of plastin 3 indeed perturbed actin-bundling activity in Sertoli cells vs. control cells (Fig. 5B). Furthermore, the actin polymerization kinetics was also perturbed (Fig. 5C). Collectively, these data illustrate that plastin 3 is a crucial regulator of actin microfilament bundles at the basal ES/BTB by conferring actin bundling and polymerization of microfilaments. KD in the testis in vivo perturbs spermatogenesis To better understand the role of plastin 3 in the testis during spermatogenesis, plastin 3 was silenced by transfecting adult rat testes with plastin 3-specific sirna duplexes to knock down plastin 3 by ;55% based on fluorescence analysis (Fig. 6A), which is consistent with results of qpcr (Fig. 6B). Testes from these rats were examined for any changes in the status of spermatogenesis. Because plastin 3 was highly expressed at the apical ES in stage VII tubules, and in vitro studies illustrate its involvement in bundled actin microfilaments at the ES (Fig. 5), we assessed if its KD would impede the transport of elongated spermatids in the adluminal compartment, accelerating the progression of stage VII to VIII tubules, histogram shown in the lower panel is a mean 6 SD of 3 qpcr experiments. D) The relative steady-state protein level of plastin 3 expressed by Sertoli cells that were cultured in vitro for 4 d during the establishment of a functional TJ-permeability barrier with actin serving as a protein loading control. E) Staining of Sertoli cells with anti-plastin 3 antibody (green) and rhodamine phalloidin (red) to visualize plastin 3 and, respectively, illustrating partial colocalization of plastin 3 and in Sertoli cell cytosol. Sertoli cell nuclei were visualized by DAPI (blue). Scale bar, 4 mm. F) Localization of plastin 3 (green) in the seminiferous epithelium and interstitium of adult rat testis; cell nuclei were visualized by DAPI (blue). was expressed by Sertoli cells and interstitial cells, most notably Leydig cells, endothelial cells of microvessels, and resident macrophages. Scale bar, mm. G) Stage-specific expression of plastin 3 (green) and its colocalization with (red) in the seminiferous epithelium of adult rat testes; cell nuclei were visualized by DAPI (blue). (yellow arrowheads) colocalized with at the BTB near the basement membrane (annotated by dashed white line) in virtually all stages of the epithelial cycle such as V VII but considerably diminished in stage VIII; however, its expression at the apical ES was limited mostly to stage VII. Data shown herein are representative findings of an experiment, but each experiment was repeated 3 4 times using different samples or cell preparations excluding pilot experiments. Scale bar, 7 mm. PLASTIN 3 REGULATES ES DYNAMICS 7

 - ZO-, 2 kda (-) - N-cadherin, 3 kda (-) -")
 - Vimentin, 55 kda () - Actin, 42 kda () - IgG H, 5")


 with TJ (claudin- and ZO-) (red) and")
 (red) and microfilament () (red), at")

 side of spermatid")

, which predominantly localized to the convex side of spermatid heads")

.")


8 A Claudin C Co-IP: ZO- N-cadherin Vimentin Testis lysate Anti-claudin IgG Rabbit IgG - - IB: -, 68 kda - IgG H, 5 kda - IgG L, 25 kda Co-IP: B β-integrin Nectin-3 Testis lysate Anti-plastin 3 IgG Rabbit IgG IB: - Occludin, 65 kda (-) - ZO-, 2 kda (-) - N-cadherin, 3 kda (-) - β-integrin, 4 kda (-) - Nectin 3, 83 kda (-) - Palladin, 95 kda (-) - Filamin A, 28 kda (-) - Vimentin, 55 kda () - Actin, 42 kda () - IgG H, 5 kda - IgG L, 25 kda Co-IP: Testis lysate Anti-Arp3 IgG Anti-Eps8 IgG Mouse IgG IB: -, 68 kda IgG H, 5 kda - IgG L, 25 kda Figure 2. is a component of the basal ES at the BTB and the apical ES at the Sertoli-Sertoli and Sertoli-spermatid interface in adult rat testes. A) Dual-labeled immunofluorescence analysis confirmed partial colocalization of plastin 3 (green) with TJ (claudin- and ZO-) (red) and basal ES (N-cadherin) (red) proteins, as well as cytoskeletal proteins of the intermediate filament (vimentin) (red) and microfilament () (red), at the BTB in stage VII tubules. Cell nuclei were visualized by DAPI (blue). Scale bar, 7 mm. B) (green) was not colocalized with apical ES protein b-integrin (red) because b-integrin was restricted to the convex (dorsal) side of spermatid heads, but plastin 3 was localized mostly to the tip of the spermatid head in stage VII tubules. However, plastin 3 (green) colocalized, at least in part, with nectin-3 (red), which predominantly localized to the convex side of spermatid heads but spread to the tip of spermatid heads where plastin 3 was found. also colocalized with, at least in part, at the apical ES. Boxed areas in micrographs were enlarged and shown as insets below. Scale bars, 7 and 35 mm (insets). C) A study by co-ip to assess protein-protein interactions between plastin 3 and selected component proteins of the BTB and the apical ES including cytoskeletal elements using corresponding antibodies as immunoprecipitating antibodies. Rabbit or mouse IgG served as a negative control, and testis lysate without subjection to co-ip served as a positive control. Data shown herein are representative findings of an experiment that was repeated 3 4 times and yielded similar results. 8 Vol. 29 September 25 The FASEB Journal x LI ET AL.


 Washing sirna intensity (Arbitrary unit) Distance from the cell-cell interface (Arbitrary unit) 8..5 2 2..5..5 N-Cadherin -Catenin Claudin Figure 3.")

9 A B bp TJ protein Basal ES protein Actin regulatory protein Hemidesmosome protein Cytoskeletal protein Relative plastin 3 protein level (Arbitrary unit).5. Marker sirna sirna, 53 bp Plastin, 88 bp Plastin 2, 29 bp S6, 385 bp -, 68 kda - CAR, 46 kda - JAM-A, 36 kda - Occludin, 65 kda - ZO-, 2 kda - N-cadherin, 27 kda - -catenin, 2 kda - β-catenin, 92 kda - Formin, 8 kda - Eps8, 97 kda - Arp3, 45 kda - Palladin, 95 kda - β-integrin, 4 kda - Vimentin, 55 kda - Actin, 42 kda - GAPDH, 35 kda C D /DAPI N-cadherin/DAPI Claudin /DAPI -catenin/dapi ZO-/DAPI TER (Ohm cm 2 ) Transfection Time of Sertoli cells in culture (Day) Washing sirna intensity (Arbitrary unit) Distance from the cell-cell interface (Arbitrary unit) N-Cadherin -Catenin Claudin Figure 3. A KD of plastin 3 by RNAi perturbs the Sertoli cell TJ-permeability barrier function via changes in the localization of basal ES proteins at the cell-cell interface. Sertoli cells were cultured for 3 d alone to form a functional TJ-permeability barrier. Thereafter, cells were transfected with rat plastin 3-specific sirna duplexes vs. nontargeting negative control duplexes (Ctrl sirna) for 24 h. Subsequently, cells were washed twice and cultured for an additional 24 h and terminated for RNA extraction, lysate preparation, or IF analysis. A) A study by RT-PCR confirmed the specific KD of plastin 3 by RNAi because the expression of either plastin or plastin 2 was unaffected. B) A study by IB also illustrated the specific KD of plastin 3 by up to ;7%, and several BTB-associated proteins including actin-regulating proteins were unaffected without any apparent off-target effect. Each bar in the histogram is a mean 6 SD of 4 experiments. P,.. C) A KD of plastin 3 was found to perturb the Sertoli cell TJ permeability. Each data point is a mean 6 SD of triplicates of a representative experiment, which was repeated 3 times and yielded similar results. P,.5. D) A study by IF to confirm the silencing of plastin 3 by ;7%. The KD of plastin 3 also affected the localization of basal ES proteins N-cadherin and b-catenin, but not TJ proteins claudin- and ZO-, at the Sertoli cell-cell interface in which these proteins moved away from the cell junction site (see white brackets). Scale bar, 4 mm. ZO- PLASTIN 3 REGULATES ES DYNAMICS 9
 3 2 Plastin3 sirna Figure 4. Confocal microscopy analysis of TJ vs.")
 or basal ES protein N-cadherin (green fluorescence) are shown in the top 2 panels, which are")
 on the right.")
 on the right.")

 About 8 Ser")
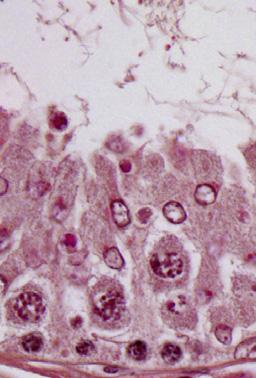
10 A sirna sirna ZO-/DAPI N-cadherin/DAPI z y ZO- N-cadherin x-y section x z y x-z section x B Localization of ZO- at the cell-cell interface (Arbitrary unit) 3 2 Plastin3 sirna Localization of N-cadherin at the cell-cell interface (arbitrary unit) 3 2 Plastin3 sirna Figure 4. Confocal microscopy analysis of TJ vs. basal ES proteins at the Sertoli cell-cell interface following plastin 3 KD by RNAi. A) Sertoli cells transfected with nontargeting negative control sirna duplexes vs. plastin 3-specific duplexes for plastin 3 KD and stained for either TJ protein ZO- (green fluorescence) or basal ES protein N-cadherin (green fluorescence) are shown in the top 2 panels, which are the horizontal views of the Sertoli cell epithelium, showing the optical slice from the x-y plane (i.e., parallel to the plane of Sertoli cell attachment). See x-y section orientation annotated in the graph (in red) on the right. The bottom panel in (A) illustrates the vertical view of the Sertoli cell epithelium, showing the reconstructed optical slide from the x-z plane (i.e., perpendicular to the plane of Sertoli cell attachment). See x-z section orientation annotated in the graph (in blue) on the right. Corresponding optical slice positions on the x-y plane are annotated by white brackets marked by dotted lines (middle panel) and shown in the bottom x-z plane and annotated by black brackets. These findings illustrate that basal ES protein N-cadherin considerably endocytosed following plastin 3 KD, thereby destabilizing the Sertoli cell-permeability barrier as shown in Fig. 3C. Scale bar, 8 mm. B) About 8 Sertoli cells in each experimental group of 3 independent experiments (i.e., 24 cells total) were randomly selected and analyzed as shown in (A) and summarized in the histogram in which the localization of N-cadherin (green fluorescence) or ZO- (green fluorescence) at the cell-cell interface was assessed in plastin 3 KD cells vs. control cells. Localization refers to the mean of the width of green fluorescence on 4 sides of a Sertoli cell of 24 randomly selected cells from 3 experiments, and the width of N-cadherin signal in control cells was arbitrarily set at. Each bar is a mean 6 SD of 3 experiments. P,.. because one of the primary functions of apical ES is to confer spermatid transport besides adhesion. It was noted that in Sprague-Dawley rats transfected with nontargeting negative control sirna duplexes, the frequencies of stage VII and VIII tubules were ;2 and ;8%, respectively, among the 6 tubules randomly scored from 3 rats, consistent with findings of an earlier report (5). Indeed, the KD of plastin 3 led to a considerable reduction in the number of VII and a concomitant increase in VIII tubules (Fig. 6C), supporting the concept that a disruption of actin microfilament bundles at the apical ES perturbed spermatid transport and adhesion, such that stage VII tubules underwent spermiation, making them analogous to stage VIII tubules morphologically (Fig. 6D). Histologic analysis of the testis following plastin 3 KD vs. the nontargeting control testis indeed supports this concept because multiple elongated spermatids were found to be embedded deep inside the seminiferous epithelium in an apparent stage VIII tubule (top panel in Fig. 6D),whichinfactwas a stage VII tubule. Also, residual elongated spermatids were found to be embedded deep inside the seminiferous epithelium in stage X tubules (lower panel in Fig. 6D). Furthermore, defects in the transport of phagocytes were also noted. For instance, phagosomes that were found near the basement membrane a typical feature of stage IX tubules in control testes (left panel in Fig. 6E) remained near the tubule lumen in plastin 3 KD testes in stage IX tubules including a late-stage IX tubule as shown herein (Fig. 6E), Vol. 29 September 25 The FASEB Journal x LI ET AL.
 2.5 2 2 3 Time (min) Relative rate of actin polymerization (Arbitrary unit).5 Figure 5.")
 Following the KD of plastin 3, the F- actin (green or gray) network in Sertoli cells became branched, truncated, and intersected with each other, illustrating a reduced bundling capability, and")

 A study by biochemical assay to assess actin-bundling activity illustrated that the KD of plastin 3 led to a reduced actin-bundling capability (left panel).")
11 A sirna sirna F-Actin/DAPI Eps8/DAPI F-Actin Vimentin/DAPI B Pellet S/N alone sirna - Actin, 42 kda - Actin, 42 kda -, 68 kda - Actin, 42 kda Palladin/DAPI Arp3/DAPI Relative bundled (Arbitrary unit).5 C Fluorescence (Arbitrary unit) Time (min) Relative rate of actin polymerization (Arbitrary unit).5 Figure 5. KD of plastin 3 leads to disorganization of actin microfilaments in Sertoli cells. A) Following the KD of plastin 3, the F- actin (green or gray) network in Sertoli cells became branched, truncated, and intersected with each other, illustrating a reduced bundling capability, and the vimentin-based intermediate filaments were mostly unaffected. These changes in actin microfilament organization following plastin 3 KD also associated with a mis-localization of actin barbed end capping and bundling protein Eps8, branched actin polymerization protein Arp3, and actin-bundling protein palladin. For instance, both Eps8 and Arp3 were not found at the Sertoli cell-cell interface (see yellow arrowheads). B) A study by biochemical assay to assess actin-bundling activity illustrated that the KD of plastin 3 led to a reduced actin-bundling capability (left panel). Pellet contained bundled, whereas supernatant (S/N) contained single unbundled actin microfilaments (see Materials and Methods for details). Each bar in the histogram (right panel) is a mean 6 SD of 4 experiments. P,.5. C) A KD of plastin 3 by RNAi also perturbed the kinetics of actin polymerization (left panel) in Sertoli cells. In brief, Sertoli cells ( cells/cm 2 ) cultured for 3 d were transfected with corresponding sirna duplexes for 24 h. Thereafter, sirna duplexes were removed, and Sertoli cells were cultured alone in fresh F2/DMEM for another 24 h and harvested on d 5 to obtain lysates and subjected to a fluorometric-based actin polymerization assay, in which the polymerization of pyrene-labeled actin was monitored by enhanced fluorescence emission at nm over a 35-min period. The relative rate of actin polymerization (right panel) (i.e., increase in fluorescence intensity over time) during the initial linear phase in the first 2 min based on the kinetics data shown on the left panel was estimated by linear regression. The rate of actin polymerization of control cells (transfected with nontargeting negative control duplexes) was arbitrarily set as. This experiment was repeated 4 times and yielded similar results. Each bar in the histogram is a mean 6 SD of 4 experiments. P,.5. PLASTIN 3 REGULATES ES DYNAMICS

 D.")



 B")














12 A sirna /DAPI /DAPI Relative fluorescence intensity (Arbitrary unit) D.5 Stage VIII sirna Stage VIII E Stage IX Plastin3 sirna C Plastin3 sirna Percentage of stage VII and VIII tubules Relative plastin 3 mrna level (Arbitrary unit) B Plastin3 sirna VII VIII sirna Stage IX Stage IX Stage X Stage X Figure 6. A KD of plastin 3 in adult rat testes in vivo impairs spermatid adhesion and spermatid transport. Testes of adult rats (;7 d postpartum; ; g b.w.) were transfected with plastin 3-specific sirna duplexes vs. nontargeting negative controls. About 2 3 d thereafter, testes were collected either to obtain frozen sections or fixed in Bouin s fixative to obtain paraffin sections for histologic analysis. A) Frozen sections of testes were stained for plastin 3 (green); cell nuclei were visualized with DAPI (blue). Due to the stage-specific expression of plastin 3 at the apical ES in stage VII tubules, we focused our analysis on this stage. At least 3 stage VII tubules of each testis from the plastin 3 RNAi group (n = 4 rats) and control group (n = 4 rats) were analyzed. fluorescence staining intensity was considerably diminished following the KD of plastin 3 in vivo. Each bar in (continued on next page) 2 Vol. 29 September 25 The FASEB Journal x LI ET AL.
13 illustrating a failure of phagosome transport that is known to be an actin-based cytoskeleton-dependent event (52). In short, these data illustrate that the ES function was compromised due to plastin 3 KD in the testis. Changes in the basal ES protein distribution at the BTB following plastin 3 in vivo KD in the testis KD of plastin 3 in the testis in vivo ledtoareducedexpression of plastin 3 at the BTB; however, plastin 3 KD also led to reduced and Eps8 at the BTB (Fig. 7A, B). Consistent with findings in vitro shown in Fig. 3D, the localization of basal ES proteins N-cadherin and b-catenin, but not TJ proteins claudin- and ZO-, was no longer tightly restricted to the BTB and diffused away from the site (Fig. 7A, C). Changes in the apical ES protein distribution in vivo following plastin 3 KD in the testis Because the expression of plastin 3 is limited to stage VII tubules, the following analysis was performed using stage VII tubules. KD of plastin 3 in the testis in vivo that led to a reduced expression of plastin 3 at the apical ES also perturbed the organization and/or localization of at the apical ES (Fig. 8A). For instance, is no longer confined predominantly to the concave (ventral) side of spermatid heads as in control testes but mostly to the tip (yellow arrowheads in Fig. 8A) or on the convex (dorsal) side (white arrowheads) of spermatid heads. These findings are consistent with data obtained in vitro (Fig. 5). Furthermore, the expression of Arp3 and Eps8 at the apical ES near the concave side of spermatid heads was also considerably down-regulated (Fig. 8A). These changes in actin regulatory protein localization and/or expression that impeded organization following plastin 3 KD thus perturbed the localization of apical ES proteins nectin-3, b-integrin, and laminin-g3, as well as an apical ES regulatory protein kinase p-focal adhesion kinase (FAK)-Tyr 397, at the Sertoli-spermatid interface (Fig. 8B). For instance, nectin-3 no longer tightly associated with the convex (dorsal) side of spermatid heads as found in control testes. Instead, it became disengaged and moved away from the site in many spermatids (green arrowheads in Fig. 8B), whereas b-integrin that was found covering the convex side of spermatid heads in control testes became limited to the tip of spermatid heads in some spermatids (red arrowheads), and laminin-g3 chain was generally down-regulated in plastin 3-silenced testes (Fig. 8B). p- FAK-Tyr 397 was no longer tightly associated with the convex side of spermatid heads but diffusely localized at the apical ES (Fig. 8B). These changes thus support findings shown in Fig. 6C E regarding defects in spermatid adhesion and transport of spermatids and phagosomes following plastin 3 KD in the testis. DISCUSSION Mammalian cells usually express just one form of plastins, such as plastin in small intestine, colon, and kidney, plastin 2 in leukocytes and tumor cells, and plastin 3 in normal cells of solid tissues (e.g., neurons, fibroblasts, endothelial cells, epithelial cells, and melanocytes) (3 5, 2). Sertoli cells, however, express all 3 plastins with plastin 3 being the most predominant form. Germ cells also express plastin and 2, but not plastin 3, in the seminiferous epithelium as reported herein. Recent studies in humans have illustrated the physiologic and pathophysiologic significance of plastin 3. For instance, plastin 3 is a reliable marker for Sézary syndrome (a type of cutaneous lymphoma) diagnosis, in which the expression of plastin 3 by CD4 T cells is several orders of magnitude vs. CD4 Tcells in normal subjects (53). is also identified as a protective modifier of spinal muscular atrophy (SMA) because higher expression of plastin 3 was detected in peripheral blood of patients who had lesser clinical severity of SMA, such as the capability of walking unaided in some female patients (54). is also becoming a novel the histogram shown on the right is a mean 6 SD of n = 4 rats, and the fluorescence intensity of control rats was arbitrarily set as. P,.5. B) The steady-state mrna level of plastin 3 in the testis of the plastin 3 KD group vs. nontargeting control group was examined by qpcr using glyceraldehyde 3-phosphate dehydrogenase as a loading control and reference gene. The mrna level of plastin 3 was reduced by 4% with 4 rats in each group when testes were transfected by sirna duplexes in vivo. P,.5. C) The number of stage VII tubules expressed as percentage of all tubules examined was found to be reduced concomitant with an increase in stage VIII tubules in the testis after plastin 3 KD due to defects in spermatid adhesion following a loss of actinbundling protein plastin 3 at the apical ES, in which spermiation took place in stage VII tubules, rendering them to appear as stage VIII tubules. About 2 tubules from each testis with 3 rats from each group were randomly counted to score stage VII and VIII tubules. P,.5. D) In the control testis that was transfected with nontargeting negative control sirna duplexes, elongated spermatids line up near the tubule lumen edge as shown in this stage VIII tubule to prepare for spermiation (left top panel), and phagosome (green arrowhead) was also detected near the tubule lumen that was derived from residual bodies phagocytosed by the Sertoli cell to be transported to the base of the seminiferous epithelium at stage IX (see panel in E). However, after plastin 3 KD, in this apparent stage VIII tubule (right top panel), elongated spermatids remained entrapped in the seminiferous epithelium (yellow arrowheads), which appeared to be spermatids from a putative stage VII tubule that failed to be transported to the tubule lumen following plastin 3 KD. But spermatids near the tubule lumen were induced to undergo spermiation prematurely due to defects of actin microfilament organization at the apical ES as a result of plastin 3 KD. In the bottom panel in (D), an elongated spermatid also remained entrapped deep inside the epithelium (yellow arrowhead) in this stage X tubule (right panel) vs. the control (left panel). Scale bars, 7 and 4 mm (insets). E) In the control testis, phagosomes were transported to the base of the epithelium in stage IX (left panel); however, a KD of plastin 3 in the testis caused defects in actin microfilaments, impeding phagosome transport so that many phagosomes remained near the tubule lumen (annotated by green arrowheads), and this failure in phagosome transport persisted in late-stage IX (bottom-right panel). In short, these defects were widespread in many of the apparent stage VIII/IX tubules following plastin 3 KD resulting from defects in spermatid adhesion/transport due to disorganization of actin microfilaments at the ES. Insets are magnified images of the corresponding boxed areas in micrographs. Scale bars, 7 and 4 mm (insets). PLASTIN 3 REGULATES ES DYNAMICS 3
.5 Eps8 sirna C Relative distance from the BTB (Arbitrary unit) 2 N-cadherin -catenin sirna Figure 7.")

14 A sirna /DAPI /DAPI /DAPI /DAPI Eps8 Eps8/DAPI Eps8 Eps8/DAPI N-cadherin N-cadherin/DAPI N-cadherin N-cadherin/DAPI -catenin -catenin/dapi -catenin -catenin/dapi Claudin Claudin /DAPI Claudin Claudin /DAPI ZO ZO/DAPI ZO ZO/DAPI B Relative fluorescence intensity (Arbitrary unit).5 Eps8 sirna C Relative distance from the BTB (Arbitrary unit) 2 N-cadherin -catenin sirna Figure 7. Changes in the distribution of, actin regulatory protein Eps8, and basal ES proteins following a KD of plastin 3 in adult rat testes in vivo. Frozen sections of rat testes transfected with sirna duplexes to knock down plastin 3 vs. nontargeting negative control duplexes were stained for plastin 3,, actin barbed end capping and bundling protein Eps8, as well as basal ES proteins N-cadherin and b-catenin vs. TJ proteins claudin- and ZO-. Cell nuclei were visualized with DAPI (blue). Stage VII/VIII/IX tubules were randomly selected for analysis. A) KD of plastin 3 was verified in these tubules in which plastin 3 fluorescence staining was considerably diminished. The fluorescence intensity of both and Eps8 at the BTB was also weakened after plastin 3 KD vs. control, supporting the notion that there were less and microfilament bundles in plastin 3 KD testes. Consistent with the findings in vitro, the basal ES proteins N-cadherin and b-catenin were no longer restrictively localized to the BTB near the basement membrane (annotated by dashed white line); instead, they displayed a considerably diffused pattern (see white brackets in plastin 3 KD testes vs. control testes). Interestingly, the TJ proteins claudin- and ZO- were not affected by the plastin 3 KD. Scale bar, 4 mm. B and C) Semiquantitative analysis of fluorescence image data shown in (A) including fluorescence intensity (B) or the mis-localization of basal ES proteins by diffusing away from the BTB (C). Data in the control group were arbitrarily set at. Each bar is a mean 6 SD of 3 rats. P,.. marker for circulating tumor cells (CTCs) in blood, possessing important prognostic value for metastasis because it is highly expressed in metastatic CTCs (55). These findings also support the notion that plastin 3 is crucial to confer actin-based cytoskeletal dynamics to mammalian cells, such as metastatic cells for locomotion. Although the Sertoli cell in vivo is not a motile cell type per se, unlike fibroblasts, macrophages, or neutrophils, Sertoli cells in the 4 Vol. 29 September 25 The FASEB Journal x LI ET AL.








.")



 side of")
 of spermatid heads,")


 and b-integrin (an integral membrane in Sertoli")

.")
15 A /DAPI sirna /DAPI Plastin3 sirna..5 /DAPI /DAPI..5 Arp3 Arp3/DAPI Arp3 Arp3/DAPI Arp3..5 Eps8 Eps8/DAPI Eps8 Eps8/DAPI Eps8..5 B sirna Nectin 3 Nectin 3/DAPI Nectin 3 Nectin 3/DAPI -integrin -integrin/dapi -integrin -integrin/dapi Laminin- 3 Laminin- 3/DAPI Laminin- 3 Laminin- 3/DAPI p-fak-tyr 397 p-fak-tyr 397 /DAPI p-fak-tyr 397 p-fak-tyr 397 /DAPI Figure 8. Changes in the distribution of, actin regulatory proteins, and apical ES proteins following a KD of plastin 3 in adult rat testes in vivo. Frozen sections of testes transfected with sirna duplexes to knock down plastin 3 vs. nontargeting negative control duplexes were stained for plastin 3,, branched actin polymerization protein Arp3, and actin barbed end capping and bundling protein Eps8, as well as apical ES constituent proteins nectin-3, b-integrin, and laminin-g3, and apical ES regulatory protein p-fak-tyr 397. Nuclei were stained with DAPI (blue). A) In stage VII tubules, plastin 3 staining was considerably diminished at the apical ES following plastin 3 KD in the testis in vivo vs. control. Actin microfilaments prominently localized to the concave (ventral) side of spermatid heads as shown in the seminiferous epithelium of control testes were found to become diffusively localized at the site and considerably diminished; and in some cases, was mostly expressed on the convex (dorsal) side of spermatid heads but near the base (white arrowheads) and also near the tip (yellow arrowheads) of spermatid heads, considerably different from the control testes. Furthermore, both Arp8 and Eps8 were considerably diminished at the apical ES following plastin 3 KD in the testis. Right panel illustrates semiquantitative analysis of the IF data shown on the left panel. P,.5, P,.. B) Apical ES proteins nectin-3 (an integral membrane protein in spermatids) and b-integrin (an integral membrane in Sertoli cells) were also found to be mis-localized in which nectin-3 appeared to be detached from the convex side of spermatid heads (green arrowheads), whereas b-integrin shifted its expression mostly to the tip of spermatids in some cases (red arrowheads). For laminin-g3 chain (a spermatid-specific apical ES protein), its expression was considerably down-regulated, whereas apical ES regulatory protein p-fak-tyr 397 no longer tightly localized to the convex side of spermatid heads; instead, it was diffusely localized. Scale bars, 4 mm. PLASTIN 3 REGULATES ES DYNAMICS 5
Wang, CQF; Mruk, DD; Lee, WWM; Cheng, CY. Citation Experimental Cell Research, 2007, v. 313 n. 7, p
 Title Coxsackie and adenovirus receptor (CAR) is a product of Sertoli and germ cells in rat testes which is localized at the Sertoli- Sertoli and Sertoli-germ cell interface Author(s) Wang, CQF; Mruk,
Title Coxsackie and adenovirus receptor (CAR) is a product of Sertoli and germ cells in rat testes which is localized at the Sertoli- Sertoli and Sertoli-germ cell interface Author(s) Wang, CQF; Mruk,
REVIEWS. Germ Cell Transport Across the Seminiferous Epithelium During Spermatogenesis
 Germ Cell Transport Across the Seminiferous Epithelium During Spermatogenesis Transport of germ cells across the seminiferous epithelium is crucial to spermatogenesis. Its disruption causes infertility.
Germ Cell Transport Across the Seminiferous Epithelium During Spermatogenesis Transport of germ cells across the seminiferous epithelium is crucial to spermatogenesis. Its disruption causes infertility.
N-WASP Is Required for Structural Integrity of the Blood-Testis Barrier
 N-WASP Is Required for Structural Integrity of the Blood-Testis Barrier The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
N-WASP Is Required for Structural Integrity of the Blood-Testis Barrier The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Differential interactions between transforming growth factor-β3/ TβR1, TAB1, and CD2AP disrupt blood-testis barrier and sertoligerm cell adhesion
 Title Differential interactions between transforming growth factor-β3/ TβR1, TAB1, and CD2AP disrupt blood-testis barrier and sertoligerm cell adhesion Author(s) Xia, W; Mruk, DD; Lee, WM; Cheng, CY Citation
Title Differential interactions between transforming growth factor-β3/ TβR1, TAB1, and CD2AP disrupt blood-testis barrier and sertoligerm cell adhesion Author(s) Xia, W; Mruk, DD; Lee, WM; Cheng, CY Citation
TGF-h3 regulates anchoring junction dynamics in the seminiferous epithelium of the rat testis via the Ras/ERK signaling pathway: An in vivo study
 Developmental Biology 280 (2005) 321 343 www.elsevier.com/locate/ydbio TGF-h3 regulates anchoring junction dynamics in the seminiferous epithelium of the rat testis via the Ras/ERK signaling pathway: An
Developmental Biology 280 (2005) 321 343 www.elsevier.com/locate/ydbio TGF-h3 regulates anchoring junction dynamics in the seminiferous epithelium of the rat testis via the Ras/ERK signaling pathway: An
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Human Apolipoprotein A1 EIA Kit
 A helping hand for your research Product Manual Human Apolipoprotein A1 EIA Kit Catalog Number: 83901 96 assays 1 Table of Content Product Description 3 Assay Principle 3 Kit Components 3 Storage 4 Reagent
A helping hand for your research Product Manual Human Apolipoprotein A1 EIA Kit Catalog Number: 83901 96 assays 1 Table of Content Product Description 3 Assay Principle 3 Kit Components 3 Storage 4 Reagent
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Supplemental Information. Autophagy in Oncogenic K-Ras. Promotes Basal Extrusion. of Epithelial Cells by Degrading S1P. Current Biology, Volume 24
 Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
ab CytoPainter Golgi/ER Staining Kit
 ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
Toxicants target cell junctions in the testis: Insights from the indazole-carboxylic acid model
 Spermatogenesis ISSN: (Print) 2156-5562 (Online) Journal homepage: https://www.tandfonline.com/loi/kspe20 Toxicants target cell junctions in the testis: Insights from the indazole-carboxylic acid model
Spermatogenesis ISSN: (Print) 2156-5562 (Online) Journal homepage: https://www.tandfonline.com/loi/kspe20 Toxicants target cell junctions in the testis: Insights from the indazole-carboxylic acid model
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses
 The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses") Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses using an anti-cre antibody; testes at 1 week (left panel),
Supplemental Figure 1. (A) The localization of Cre DNA recombinase in the testis of Cyp19a1-Cre mice was detected by immunohistchemical analyses using an anti-cre antibody; testes at 1 week (left panel),
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Regulation of blood-testis barrier dynamics: an in vivo study
 Research Article 783 Regulation of blood-testis barrier dynamics: an in vivo study Ching-hang Wong, Dolores D. Mruk, Wing-yee Lui and C. Yan Cheng* Population Council, Center for Biomedical Research, 1230
Research Article 783 Regulation of blood-testis barrier dynamics: an in vivo study Ching-hang Wong, Dolores D. Mruk, Wing-yee Lui and C. Yan Cheng* Population Council, Center for Biomedical Research, 1230
Supplementary table and figures
 3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
3D single molecule tracking with multifocal plane microscopy reveals rapid intercellular transferrin transport at epithelial cell barriers Sripad Ram, Dongyoung Kim, Raimund J. Ober and E. Sally Ward Supplementary
Supplemental Information. Otic Mesenchyme Cells Regulate. Spiral Ganglion Axon Fasciculation. through a Pou3f4/EphA4 Signaling Pathway
 Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Fluorescence Microscopy
 Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Adapted from Preg. & Part., Senger
 MALE ENDOCRINOLOGY AND SPERMATOGENESIS (Chapter 10) AVS 222 (Instructor: Dr. Amin Ahmadzadeh) I. MALE ENDOCRINOLOGY (Figure10-1 to 10-3) A. Glands and their respective hormones 1) Hypothalamic hormone:
MALE ENDOCRINOLOGY AND SPERMATOGENESIS (Chapter 10) AVS 222 (Instructor: Dr. Amin Ahmadzadeh) I. MALE ENDOCRINOLOGY (Figure10-1 to 10-3) A. Glands and their respective hormones 1) Hypothalamic hormone:
human Total Cathepsin B Catalog Number: DY2176
 human Total Cathepsin B Catalog Number: DY2176 This DuoSet ELISA Development kit contains the basic components required for the development of sandwich ELISAs to measure natural and recombinant human Total
human Total Cathepsin B Catalog Number: DY2176 This DuoSet ELISA Development kit contains the basic components required for the development of sandwich ELISAs to measure natural and recombinant human Total
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
") Supplementary Figure 1 (Mu) SBP (mmhg) 2 18 16 p
Supplementary Figure 1 (Mu) SBP (mmhg) 2 18 16 p 04_polarity. The formation of synaptic vesicles
 Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Focus Application. Compound-Induced Cytotoxicity
 xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity For life science research only. Not for use in diagnostic procedures. Featured Study: Using the Time Resolving
xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity For life science research only. Not for use in diagnostic procedures. Featured Study: Using the Time Resolving
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
SUPPORTING MATREALS. Methods and Materials
 SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
were isolated from the freshly drawn blood of healthy donors and ACS patients using the
 Supplemental Figure 1. Quality control of CD4 + T-cell purification. CD4 + T cells were isolated from the freshly drawn blood of healthy donors and ACS patients using the RosetteSep CD4 + T Cell Enrichment
Supplemental Figure 1. Quality control of CD4 + T-cell purification. CD4 + T cells were isolated from the freshly drawn blood of healthy donors and ACS patients using the RosetteSep CD4 + T Cell Enrichment
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Supplemental Experimental Procedures
 Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated,
 Mammary epithelial cells were freshly isolated,") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated, embedded in matrigel and exposed
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated, embedded in matrigel and exposed
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Catalog Number KA1538 48 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use...
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
") EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric) Base Catalog # PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Total Histone H3 Acetylation Detection Fast Kit (Colorimetric)
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
Exam Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) All of the following are synthesized along various sites of the endoplasmic reticulum
Electron micrograph of phosphotungstanic acid-stained exosomes derived from murine
 1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS
 TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
STAT3 (py705) (Human/Mouse/Rat) ELISA Kit
 (Human/Mouse/Rat) ELISA Kit") STAT3 (py705) (Human/Mouse/Rat) ELISA Kit Catalog Number KA2175 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT3 (py705) (Human/Mouse/Rat) ELISA (Enzyme-Linked
STAT3 (py705) (Human/Mouse/Rat) ELISA Kit Catalog Number KA2175 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT3 (py705) (Human/Mouse/Rat) ELISA (Enzyme-Linked
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental information contains 7 movies and 4 supplemental Figures
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.
 Light microscopic pictures show mouse embryos at developmental stage E12.") Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
NIH Public Access Author Manuscript Crit Rev Biochem Mol Biol. Author manuscript; available in PMC 2010 September 1.
 NIH Public Access Author Manuscript Published in final edited form as: Crit Rev Biochem Mol Biol. 2009 ; 44(5): 245 263. doi:10.1080/10409230903061207. An intracellular trafficking pathway in the seminiferous
NIH Public Access Author Manuscript Published in final edited form as: Crit Rev Biochem Mol Biol. 2009 ; 44(5): 245 263. doi:10.1080/10409230903061207. An intracellular trafficking pathway in the seminiferous
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
RayBio Human Phospho-DDR2 (Tyr740) and Total DDR2 ELISA Kit
 and Total DDR2 ELISA Kit") RayBio Human Phospho-DDR2 (Tyr740) and Total DDR2 ELISA Kit Catalog #: PEL-DDR2-Y740-T User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607
RayBio Human Phospho-DDR2 (Tyr740) and Total DDR2 ELISA Kit Catalog #: PEL-DDR2-Y740-T User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607
Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit
 Protein Quantity Microplate Assay Kit") PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
Evaluation of directed and random motility in microslides Assessment of leukocyte adhesion in flow chambers
 Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
RayBio Human Phospho-DDR1 (Tyr792) ELISA Kit
 ELISA Kit") RayBio Human Phospho-DDR1 (Tyr792) ELISA Kit Catalog #: PEL-DDR1-Y792 User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane,
RayBio Human Phospho-DDR1 (Tyr792) ELISA Kit Catalog #: PEL-DDR1-Y792 User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane,
RayBio Human, Mouse and Rat Phospho-NF-kB P65 (Ser536) and Total NF-kB P65 ELISA Kit
 and Total NF-kB P65 ELISA Kit") RayBio Human, Mouse and Rat Phospho-NF-kB P65 (Ser536) and Total NF-kB P65 ELISA Kit Catalog #: PEL-NFKBP65-S536-T User Manual Last revised October 10, 2017 Caution: Extraordinarily useful information
RayBio Human, Mouse and Rat Phospho-NF-kB P65 (Ser536) and Total NF-kB P65 ELISA Kit Catalog #: PEL-NFKBP65-S536-T User Manual Last revised October 10, 2017 Caution: Extraordinarily useful information
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supplemental Materials. STK16 regulates actin dynamics to control Golgi organization and cell cycle
 Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
RayBio Human Phospho-DDR1 (Tyr792) and Total DDR1 ELISA Kit
 and Total DDR1 ELISA Kit") RayBio Human Phospho-DDR1 (Tyr792) and Total DDR1 ELISA Kit Catalog #: PEL-DDR1-Y792-T User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607
RayBio Human Phospho-DDR1 (Tyr792) and Total DDR1 ELISA Kit Catalog #: PEL-DDR1-Y792-T User Manual Last revised March 22, 2018 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607
EPIGENTEK. EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE
 Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE") EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric)
EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric) Base Catalog # P-4059 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Global Acetyl Histone H3K27 Quantification Kit (Colorimetric)
Focal adhesion kinase is a regulator of F-actin dynamics New insights from studies in the testis
 REVIEW Spermatogenesis 3:3, e25385; July/August/September 2013; 2013 Landes Bioscience REVIEW Focal adhesion kinase is a regulator of F-actin dynamics New insights from studies in the testis Stephen YT
REVIEW Spermatogenesis 3:3, e25385; July/August/September 2013; 2013 Landes Bioscience REVIEW Focal adhesion kinase is a regulator of F-actin dynamics New insights from studies in the testis Stephen YT
Animal Tissue Culture SQG 3242 Biology of Cultured Cells. Dr. Siti Pauliena Mohd Bohari
 Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
Animal Tissue Culture SQG 3242 Biology of Cultured Cells Dr. Siti Pauliena Mohd Bohari The Culture Environment Changes of Cell s microenvironment needed that favor the spreading, migration, and proliferation
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Focus Application. Compound-Induced Cytotoxicity
 xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
GFP-LC3 +/+ CLU -/- kda CLU GFP. Actin. GFP-LC3 +/+ CLU -/- kda CLU GFP. Actin
 Supplementary Fig. 1 a CQ treatment ScrB OGX11 MG132 I II AZD5363 I II b GFP / / GFP / / GFP / / GFP / / GFP GFP Actin Actin ctrl CQ GFP / / GFP / / GFP / / GFP / / GFP GFP Actin Actin rapamycin rapamycincq
Supplementary Fig. 1 a CQ treatment ScrB OGX11 MG132 I II AZD5363 I II b GFP / / GFP / / GFP / / GFP / / GFP GFP Actin Actin ctrl CQ GFP / / GFP / / GFP / / GFP / / GFP GFP Actin Actin rapamycin rapamycincq
Supplemental information
 Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental
Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental
SUPPLEMENTARY INFORMATION
 DOI: 1.138/ncb222 / b. WB anti- WB anti- ulin Mitotic index (%) 14 1 6 2 T (h) 32 48-1 1 2 3 4 6-1 4 16 22 28 3 33 e. 6 4 2 Time (min) 1-6- 11-1 > 1 % cells Figure S1 depletion leads to mitotic defects
DOI: 1.138/ncb222 / b. WB anti- WB anti- ulin Mitotic index (%) 14 1 6 2 T (h) 32 48-1 1 2 3 4 6-1 4 16 22 28 3 33 e. 6 4 2 Time (min) 1-6- 11-1 > 1 % cells Figure S1 depletion leads to mitotic defects
Supplementary Figure (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were
 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were") Supplementary Figure 1. Gd@C 82 (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were treated with PBS, Gd@C 82 (OH) 22, C 60 (OH) 22 or GdCl
Supplementary Figure 1. Gd@C 82 (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were treated with PBS, Gd@C 82 (OH) 22, C 60 (OH) 22 or GdCl
Supplementary Information
 Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
STAT1 (ps727) (Human/Mouse) ELISA Kit
 (Human/Mouse) ELISA Kit") STAT1 (ps727) (Human/Mouse) ELISA Kit Catalog Number KA2171 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT1 (ps727) (Human/Mouse) ELISA (Enzyme-Linked Immunosorbent
STAT1 (ps727) (Human/Mouse) ELISA Kit Catalog Number KA2171 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT1 (ps727) (Human/Mouse) ELISA (Enzyme-Linked Immunosorbent
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
SUPPLEMENTARY INFORMATION
 b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set
 gp120 / Glycoprotein 120 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) gp120 / Glycoprotein 120 ELISA Pair Set Catalog Number : SEK11233 To achieve the best assay results, this manual must be read carefully before using this product
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the
 Schematic of Crb3 isoforms, ERLI and CLPI, indicating the") Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
TECHNICAL BULLETIN. Catalog Number RAB0447 Storage Temperature 20 C
 Phospho-Stat3 (ptyr 705 ) and pan-stat3 ELISA Kit for detection of human, mouse, or rat phospho-stat3 (ptyr 705 ) and pan-stat3 in cell and tissue lysates Catalog Number RAB0447 Storage Temperature 20
Phospho-Stat3 (ptyr 705 ) and pan-stat3 ELISA Kit for detection of human, mouse, or rat phospho-stat3 (ptyr 705 ) and pan-stat3 in cell and tissue lysates Catalog Number RAB0447 Storage Temperature 20
Supplemental Information
 Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway
 Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Human Urokinase / PLAU / UPA ELISA Pair Set
 Human Urokinase / PLAU / UPA ELISA Pair Set Catalog Number : SEK10815 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Human Urokinase / PLAU / UPA ELISA Pair Set Catalog Number : SEK10815 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
Supporting Information File S2
 Pulli et al. Measuring Myeloperoxidase Activity in Biological Samples Page 1 of 6 Supporting Information File S2 Step-by-Step Protocol Reagents: Sucrose (Sigma, S3089) CaCl 2 (Sigma, C5770) Heparin Sodium
Pulli et al. Measuring Myeloperoxidase Activity in Biological Samples Page 1 of 6 Supporting Information File S2 Step-by-Step Protocol Reagents: Sucrose (Sigma, S3089) CaCl 2 (Sigma, C5770) Heparin Sodium
Ch 2: The Cell. Goals: Anatomy of a typical cell Cell Membrane Discussion of internal structure of a cell with emphasis on the various organelles
 Ch 2: The Cell Goals: Anatomy of a typical cell Cell Membrane Discussion of internal structure of a cell with emphasis on the various organelles Developed by John Gallagher, MS, DVM Some Terminology: 1.
Ch 2: The Cell Goals: Anatomy of a typical cell Cell Membrane Discussion of internal structure of a cell with emphasis on the various organelles Developed by John Gallagher, MS, DVM Some Terminology: 1.
Dental Research Institute, Faculty of Dentistry, University of Toronto, Toronto, Canada *For correspondence:
 Zymogram Assay for the Detection of Peptidoglycan Hydrolases in Streptococcus mutans Delphine Dufour and Céline M. Lévesque * Dental Research Institute, Faculty of Dentistry, University of Toronto, Toronto,
Zymogram Assay for the Detection of Peptidoglycan Hydrolases in Streptococcus mutans Delphine Dufour and Céline M. Lévesque * Dental Research Institute, Faculty of Dentistry, University of Toronto, Toronto,
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small cell lung cancer
, NCI-H460 (human non-small cell lung cancer") Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Supplementary Figures
 Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit
/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit") STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit Catalog Number KA2176 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay...
STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit Catalog Number KA2176 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay...
LDL Uptake Cell-Based Assay Kit
 LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1%
 FIGURE LEGENDS Supplementary Fig 1 (A) sumoylation pattern detected under denaturing condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1% SDS in the presence and absence
FIGURE LEGENDS Supplementary Fig 1 (A) sumoylation pattern detected under denaturing condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1% SDS in the presence and absence
Procaspase-3. Cleaved caspase-3. actin. Cytochrome C (10 M) Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk
 Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk") A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.