Epoxyeicosatrienoic acids protect rat hearts against tumor necrosis
|
|
|
- Barbara Thomas
- 6 years ago
- Views:
Transcription
1 Epoxyeicosatrienoic acids protect rat hearts against tumor necrosis factor-α-induced injury Gang Zhao 1, Jianing Wang 1, Xizhen Xu 1, Yanyan Jing 1, Ling Tu 1,, Chen Chen 1, Katherine Cianflone 2, Peihua Wang 1, Ryan T. Dackor 3, Darryl C. Zeldin 3 and Dao Wen Wang 1 1 Department of Internal Medicine and Gene Therapy Center, Tongji Hospital, Tongji Medical College of Huazhong University of Science and Technology, Wuhan, , P.R. China; 2 Centre de Recherche Institut Universitaire de Cardiologie et Pneumologie de Quebec, Universite Laval, Y2186, 2725 Chemin Ste Foy, Ste Foy, Quebec, G1V 4G5, Canada; 3 Division of Intramural Research, National Institute of Environmental Health Sciences, NIH, Research Triangle Park, NC Corresponding Author: Dao Wen Wang, M.D., Ph.D. and Ling Tu, M.D., Ph.D. Department of Internal Medicine, Tongji Hospital, Tongji Medical College Huazhong University of Science and Technology 1095# Jiefang Ave. Wuhan , P. R. China Tel and Fax: (86-27) dwwang@tjh.tjmu.edu.cn or proftu@tom.com Running Title: EETs and cardiac protection 1
2 Document statistics: Number of Figures: 6. Number of References: 42. The number of words in the Abstract: 197 The number of words in the Introduction: 360 The number of words in the Discussion: 663. Abbreviations: EETs, Epoxyeicosatrienoic acids; AA, arachidonic acid; raav, recombinant adeno-associated viral vector; CYP2J2, cytochrome P450 2J2; GFP, green fluorescent protein; DMEM, Dulbecco s modified Eagle s medium; FBS, fetal bovine serum; DMSO, Dimethyl sulfoxide; TNF-α, tumor necrosis factor-alpha; PPOH, 2-(2- propynyloxy)-benzenehexanoic acid; PBMC, peripheral blood mononuclear cell; EGFR, epidermal growth factor receptor; PPARγ, peroxisome proliferator activated receptors γ; IL-6, Interleukin-6; IL-10, Interleukin-10; IL-1β, Interleukin-1β; svcam-1, soluble Vascuolar cell adhesion molecule-1; sicam-1, soluble intercellular adhesion molecule-1; VCAM-1,Vascular cell adhesion molecule-1; se-selectin, soluble endothelium-selectin; HUVECs, Human umbilical vein endothelial cells; GAPDH, Glyceraldehyde 3 phosphate dehydrogenase; enos, endothelial nitric-oxide synthase; PI3K, Phosphoinositide 3-kinase; AKT, protein kinase B; MAPK, mitogen-activated protein kinase; ERK, extracellular signal regulated kinase; PVDF, polyvinylidene difluoride; LVEDP, left ventricular end diastolic pressure; HEPES, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid N-2-hydroxyethylpiperazine-N[prime]-2-ethanesulfonic acid; AG1478, N-(3-Chlorophenyl)-6,7-dimethoxy-4-quinazolinamine; GW9662, 2-Chloro-5-nitro-N-phenylbenzamide; 2
3 Abstract Epoxyeicosatrienoic acids (EETs), the primary arachidonic acid metabolites of cytochrome P450 2J (CYP2J) epoxygenases, possess potent vasodilatory, anti-inflammatory, anti-apoptotic and mitogenic effects. To date, little is known about the role of CYP2J2 and EETs in TNF-α-induced cardiac injury. We therefore utilized cell culture and in vivo models to examine the effects of exogenously applied EETs or CYP2J2 overexpression on TNF-α-induced cardiac apoptosis and cardiac dysfunction. In neonatal rat cardiomyocytes, TNF-α-induced apoptosis was markedly attenuated by EETs or CYP2J2 overexpression, leading to significantly improved cell survival. Further studies showed that TNF-α decreased expression of the anti-apoptotic proteins Bcl-2 and Bcl-xL, decreased IκBα and PPARγ, and also inhibited PI3K-dependent Akt and EGFR signaling. Both EETs and CYP2J2 overexpression reversed the effects of TNF-α on these pathways. Furthermore, overexpression of CYP2J2 in rats prevented the decline in cardiac function that is normally observed in TNF-α-challenged animals. These results demonstrate that EETs or CYP2J2 overexpression can prevent TNF-α-induced cardiac cell injury and cardiac dysfunction by inhibiting apoptosis, reducing inflammation and enhancing PPARγ expression. Targeting the CYP2J2 epoxygenase pathway may represent a novel approach to mitigate cardiac injury in diseases such as heart failure where increased TNF-α levels are known to occur. 3
4 Introduction Cytochrome P450 epoxygenases of the 2J subfamily (CYP2J) are abundantly expressed in cardiac myocytes and metabolize arachidonic acid to four biologically-active epoxyeicosatrienoic acid (EET) isomers(1). CYP epoxygenase-derived EETs have anti-inflammatory properties (2), and have been reported to possess many potent biological effects in the renal and cardiovascular systems(3-5). For example, EETs inhibit cytokine-induced vascular cell adhesion molecule expression and leukocyte adhesion to the vascular wall(2), inhibit vascular smooth muscle cell migration(6), protect endothelial cells from apoptosis(7), upregulate enos(8) and promote endothelial cell proliferation and angiogenesis via activation of MAP kinases and PI3 kinase/akt signaling(8). 11,12- and 14,15-EETs have been characterized as effective mitogens(1, 9-14) which can enhance proliferation of renal epithelial cells(12, 15, 16). Interestingly, the role of 14,15-EET in mediating the mitogenic responses of EGF and HB-EGF has also been thoroughly documented in cultured LLCPk cells(9, 17). CYP2J2 is abundantly expressed in the human myocardium; however, little is known about its role in the heart, especially in the context of heart failure. Seubert et al. recently demonstrated a protective role of CYP2J2 overexpression or exogenously-administered EETs in ischemia-reperfusion injury in both rodents and dogs via a mechanism that involved opening of sarcolemmal and mitochondrial K + channels, as well as activation of p42/44 MAPK(18, 19). Importantly, the role of CYP2J2-derived EETs in non-ischemic cardiac injury and dysfunction remains unknown. In the present study, neonatal rat cardiomyocytes were used to evaluate the effects of CYP2J2 overexpression and exogenous EET supplementation on TNF-α-induced apoptosis and inflammation. An in vivo model of TNF-α-induced 4
5 cardiac dysfunction was also utilized to evaluate the hypothesis that CYP2J2 could improve cardiac function by inhibiting myocardial inflammation and apoptosis. 5
6 Materials and Methods Gene delivery vector construct Recombinant adeno-associated virus vectors containing GFP or CYP2J2 were prepared as previously described (20). The CYP2J2 or GFP cdnas subcloned into the pcdna3.1 plasmid were prepared as described (2, 21). pcdna3.1-gfp and pcdna3.1-cyp2j2 plasmids were purified using the QIAGEN Plasmid Maxi Kit (QIAGEN, Inc., Chatsworth, CA) according to the manufacturer's instructions. Primary culture and treatment of neonatal cardiomyocytes Neonatal cardiomyocytes were obtained from one-day-old rats as described previously (22). Experimental protocols were approved by the Institutional Animal Research Committee of Tongji Medical College and complied with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. The cells were incubated with TNF-α (10 ng/ml) in the presence/absence of exogenous 8,9-, 11,12- or 14,15-EETs (100 nm) or the selective CYP epoxygenase inhibitor PPOH (10 µm) for 24 hours. Alternatively, the cells were infected with raav-gfp or raav-cyp2j2 (MOI=500) for 48 hours prior to treatment with TNF-α. H9C2 cell culture and gene transfection H9C2 rat heart cells were obtained from American Type Culture Collection (ATCC, Rockville, Maryland, U.S.A) % confluent H9C2 cells were transfected with pcdna3.1/gfp or pcdna3.1/cyp2j2 plasmids (2μg DNA/9.4 cm 2 ) with Fugene 6 Transfection Reagent (Roche Molecular Biochemicals, Indianapolis, IN). Thirty-six hours after transfection, the percent transfection efficiency was determined by examining pcdna3.1/gfp transfected cells for GFP fluorescence under a fluorescence microscope (NIKON). Transfection efficiency was ~30% (see 6
7 supplementary material online, Figure S1A). The expression of CYP2J2 was confirmed by immunoblotting (see supplementary material online, Figure S1B) and was associated with increased production of stable EET metabolites, the DHETs (see supplementary material online, Figure S1C). H9C2 rat heart cell treatments To investigate the anti-apoptotic effect of CYP2J2 or EETs and possible signaling mechanisms involved, transfected H9C2 cells were treated for 45 min with/without the EGFR inhibitor AG1478 (100 nm) or the PPARγ inhibitor GW9662 (1 μm), followed by incubation with TNF-α (10ng/ml) for 24 hours. Alternatively, after pretreatment with/without MAPK inhibitor AG1478, or Apigenin or PPARγ inhibitor GW9662, cells were incubated with 11,12-EET (100 nm) for 30 min prior to TNF-α treatment. Next, cell viability was assessed using an MTT assay as described (23), and apoptosis was assessed by flow cytometry (FACS, Vantage, BD). In addition, caspase-3 activity was measured by colorimetric assay using DEVD-p-nitroanilide as a substrate as described previously (24). Flow cytometry Cells were harvested with trypsin/edta, resuspended in binding buffer [10 mm HEPES/NaOH (ph 7.4), 140 mm NaCl, and 2.5 mm CaCl 2 ], and incubated with FITC-conjugated Annexin V and 1 µg/ml propidium iodide (Annexin V-FITC kit; Bender MedSystems, San Bruno, CA) according to the manufacturer s protocol. Cells were then analyzed with a FACStar-Plus flow cytometer (Becton Dickinson, Franklin Lakes, NJ). The annexin V positive (FL1-H) cells were counted for early stages of apoptosis (the lower right quadrant) and late apoptotic or necrotic cells (the upper right quadrant). Animal treatment and gene delivery 7
8 Forty eight male Sprague-Dawley rats weighing g were obtained from the Experimental Animal Center of Tongji Medical College. Experimental protocols were approved by the Institutional Animal Research Committee of Tongji Medical College and complied with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. Rats were randomized to 3 groups (16 animals per group) and received intravenous injection of either saline control, pcdna3.1-gfp control or pcdna3.1-cyp2j2 at a dose of 2.5mg/kg body weight(25)(2). Two weeks later, 8 rats from each group randomly received a single intravenous injection of TNF-α (10 µg/kg). Hemodynamic study At six and twenty four hours after TNF-α injection, rats were anesthetized by intraperitoneal administration of pentobarbital at 40mg/kg body weight. A polyethylene catheter(26) was inserted into the left ventricle (LV) through the right carotid artery for measurement of left ventricular end diastolic pressure (LVEDP) and maximum left ventricular pressure ascending speed (+dp/dt max)(26). 14, 15-DHET Determination in urine The 14,15-DHET ELISA kit (Detroit R&D Inc., Detroit, MI) was used to measure 14,15-DHET according to the manufacturer s instructions as previously described(8). EETs can be hydrolyzed to DHETs by acid treatment and thus DHET in acidified urine represents total DHETs. The difference between total 14,15-DHET and 14,15-DHET before acidification will be 14,15-EET levels. The concentration of 14,15-DHET and 14,15-EET were expressed as ng/ml urine or serum specimen. Determination of inflammatory mediators by ELISA The levels of svcam-1, sicam-1, se-selectin, IL-1β, IL-6 and IL-10 in serum 8
9 samples were determined using ELISA kits (R&D Systems, Inc., Minneapolis, USA) according to the manufacturer s instructions. Human umbilical vein endothelial cell culture and adhesion assays Human umbilical vein endothelial cells (HUVECs) were isolated from fresh human umbilical cords and cultured. The institutional review board of Tongji hospital approved this study and written informed consent was obtained from all participants. Experiments were conducted according to the principles expressed in the Declaration of Helsinki. HUVECs were infected with raav-cyp2j2 before incubating with TNF-α (10 ng/ml). Forty eight hours after infection, adhesion assays in PBMCs were performed as previously described (27, 28). Briefly, PBMCs from healthy volunteers (n=8) were isolated and labeled with Vybrant TM CFDA SE Cell Tracer Kit (Molecular Probes Inc., Oregon, USA) following the manufacturer s instructions. HUVECs were washed three times with PBS after incubation for 6 hours with TNFα (10 ng/ml). Labeled PBMCs were added to the medium ( /ml) and incubated for 30 min, followed by washing with PBS. A video record of adhesive events was made in at least eight video-microscopic fields on each slide after 5 min of washout. For monocytes, the number of adherent cells was counted, averaged per field and converted to cells/mm 2 using the calibrated field dimensions. Western blot analysis Protein concentrations were determined using the Bradford method. Western blots were performed according to methods described previously (29). Mainly, western blots were probed with antibodies against Bcl-2, Bcl-xl, and Bax (Santa Cruz Biotechnology, CA), PI3-kinase, phosphorylated Akt and Akt (Santa Cruz Biotechnology), phospho-erk1/2 and ERK1/2(Santa Cruz Biotechnology, CA), IkBα and PPARγ (Cell Signaling Technology, Beverly, MA), epidermal growth factor 9
10 receptor (EGFR) and phosphorylated EGFR (Cell Signaling Technology,Beverly, MA). And then, the membranes were treated with secondary antibody conjugated to horseradish peroxidase (Jackson Immunoresearch Laboratories, West Grove, PA) at room temperature for 2h. Antibody against β-actin was purchased from Sigma-Aldrich (Sigma-Aldrich, St. Louis, MO). Polyclonal antibodies against CYP2J2 were developed as described previously (30). The horseradish peroxidase-conjugated secondary antibodies (goat anti-rabbit and goat anti-mice) were bought from (Jackson Immunoresearch Laboratories, West Grove, PA). Statistics All data were expressed as means ± S.E.M. Comparisons between groups were performed by a one-way analysis of ANOVA with post hoc analyses performed using the Student-Newman-Keuls method. Statistical significance was defined as P <
11 Results Effects of EETs and CYP2J2 overexpression on apoptosis of primary rat cardiomyocytes The primary rat cardiomyocytes transfected with raav-mediated CYP2J2 epoxygenase had increased CYP2J2 expression and increased DHET levels relative to corresponding cells transfected with raav-gfp. But the DHET Levels was notably decreased by co-incubation with TNF-α or PPOH (Figure S1A-C). Flow cytometry analysis showed that 8,9-EET, 11,12-EET and 14,15-EET significantly decreased apoptosis of primary rat cardiomyocytes induced by TNF-α treatment, while PPOH, a selective CYP epoxygenase inhibitor, increased apoptosis compared to the corresponding cells without PPOH treatment (Figure 1A). Furthermore, raav-mediated CYP2J2 epoxygenase overexpression reduced spontaneous apoptosis and markedly reduced TNF-α-induced cardiomyocyte apoptosis, an effect that was blocked by PPOH (Figure 1B). Representative flow cytometry scatterplots from all treatment groups are shown in the supplementary data (Figure S2A and S2B). Western blot analysis showed that exogenous EET administration and raav-mediated CYP2J2 overexpression in primary rat cardiomyocytes significantly upregulated the anti-apoptotic proteins, Bcl-2 and Bcl-xL, and downregulated the pro-apoptotic protein, Bax (Figure 1C and 1D). Furthermore, PPOH upregulated Bax, providing further support for an anti-apoptotic effect of CYP epoxygenase-derived EETs. Effect of 11,12-EET and CYP2J2 overexpression on viability of rat heart H9C2 cells H9C2 cells transfected with pcdna3.1/cyp2j2 had increased CYP2J2 expression and increased DHET levels relative to H9C2 cells transfected with 11
12 pcdna3.1/gfp (Figure S3A-C). These results are similar to those previously reported for endothelial cells (3, 4). The results of MTT assays demonstrate that viability of H9C2 cells transfected with pcdna3.1/cyp2j2 was significantly greater than control cells transfected with pcdna3.1/gfp in both the presence and absence of TNF-α (Figure 2A). The pro-survival effects of CYP2J2 were significantly attenuated by both the EGFR inhibitor AG1478 (100 nm) and the PPARγ inhibitor GW9662 (1 μm). Likewise, addition of 11,12-EET to H9C2 cells had similar protective effects on cell viability in both the presence and absence of TNF-α, and these effects were attenuated by AG1478 and GW9662 (Figure 2B). These results show that the pro-survival effects of CYP2J2 overexpression and 11,12-EET treatment are mediated, at least in part, through activation of EGFR and PPARγ. Effect of 11,12-EET treatment and CYP2J2 overexpression on apoptosis of rat heart H9C2 cells We also evaluated the effects of EETs and CYP2J2 overexpression on apoptosis of H9C2 cells by flow cytometry. Our results show that cells overexpressing CYP2J2 had significantly reduced apoptosis 24 hours after TNF-α treatment compared to GFP-transfected cells treated with TNF-α (Figure 2C). Likewise, exogenous supplementation with 11,12-EET had similar protective effects (Figure 2D). Representative flow cytometry scatterplots from all treatment groups are shown in supplementary data (Figure S4A & S4B). The anti-apoptotic effects of CYP2J2 overexpression or exogenous 11,12-EET supplementation were attenuated by both AG1478 and GW9662 (Figure 2C and 2D). These results show that the anti-apoptotic effects of CYP2J2 overexpression and 11,12-EET treatment are mediated, at least in part, through activation of EGFR and PPARγ. Effect of 11,12-EET treatment and CYP2J2 overexpression on caspase-3 12
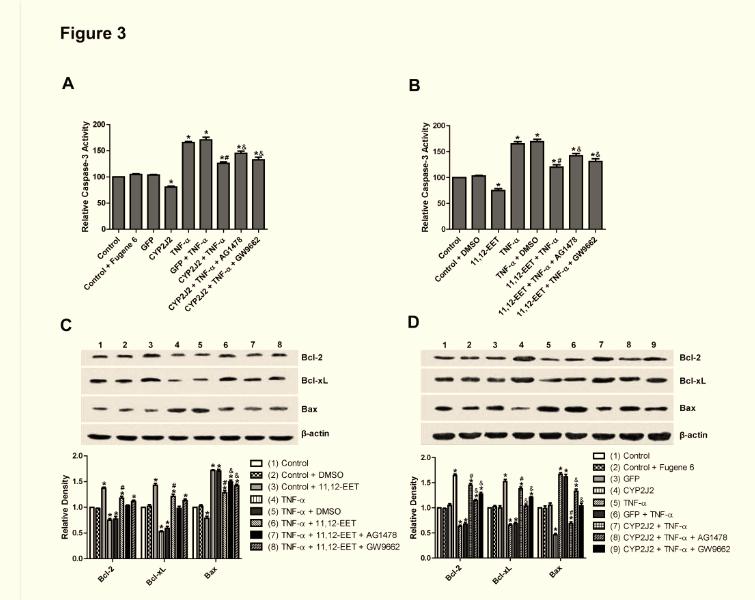
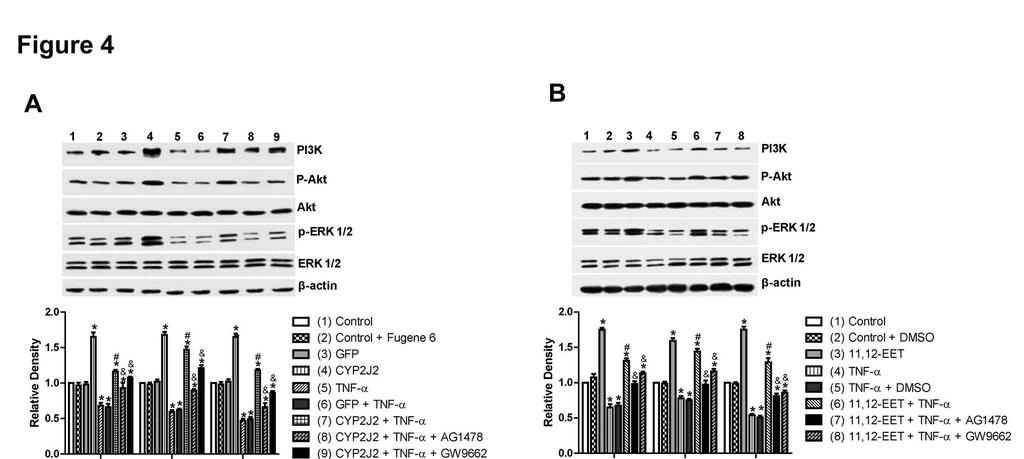
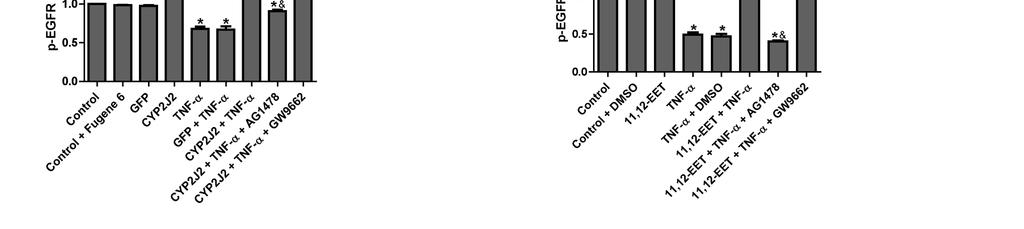
13 activity in rat heart H9C2 cells TNF-α treatment of H9C2 cells significantly increased caspase-3 activity. Transfection with pcdna3.1/cyp2j2 significantly attenuated the TNF-α-induced increase in caspase-3 compared to control and pcdna3.1/gfp transfected cells (Figure 3A). This effect was largely inhibited when cells were pre-treated with AG1478 or GW9662. Likewise, exogenous supplementation with 11,12-EET had similar protective effects on caspase-3 activity in TNF-α treated H9C2 cells, an effect that was also attenuated by AG1478 and GW9662 (Figure 3B). These results show that the effects of CYP2J2 overexpression and 11,12-EET treatment on caspase-3 activity are mediated, at least in part, through activation of EGFR and PPARγ. Effect of 11,12-EET treatment and CYP2J2 overexpression on pro- and anti-apoptotic protein expression in rat heart H9C2 cells Incubation of H9C2 cells with exogenous 11,12-EET significantly attenuated the down-regulation of anti-apoptotic proteins, Bcl-2 and Bcl-xL, and prevented the up-regulation of the pro-apoptotic protein, Bax, that is normally observed following TNF-α treatment (Figure 3C). Overexpression of CYP2J2 had similar effects on Bcl-2, Bcl-xL and Bax expression (Figure 3D). Furthermore, the effects of 11,12-EET treatment and CYP2J2 overexpression were attenuated by addition of either AG1478 or GW9662 (Figure 3C and 3D), supporting the roles of EGFR and PPARγ in the EET-mediated regulation of apoptosis signaling. Effect of 11,12-EET treatment and CYP2J2 overexpression on PI3K-dependent Akt phosphorylation and EGFR-ERK pathways in rat heart H9C2 cells TNF-α treatment inhibited expression of PI3K and PI3K-dependent Akt phosphorylation, while CYP2J2 overexpression induced PI3K protein expression and Akt phosphorylation in the absence or presence of TNF-α (Figure 4A). The effects of 13
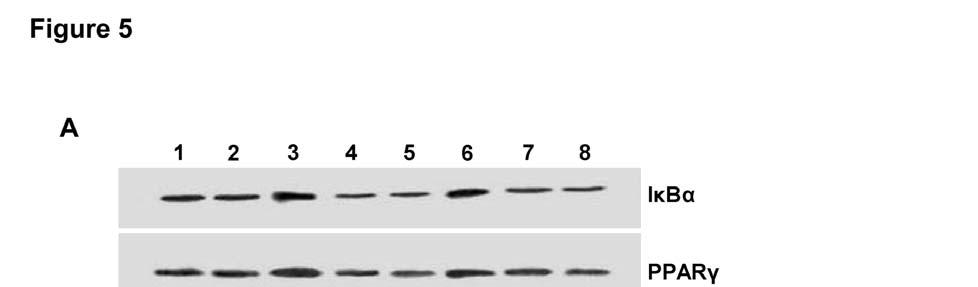
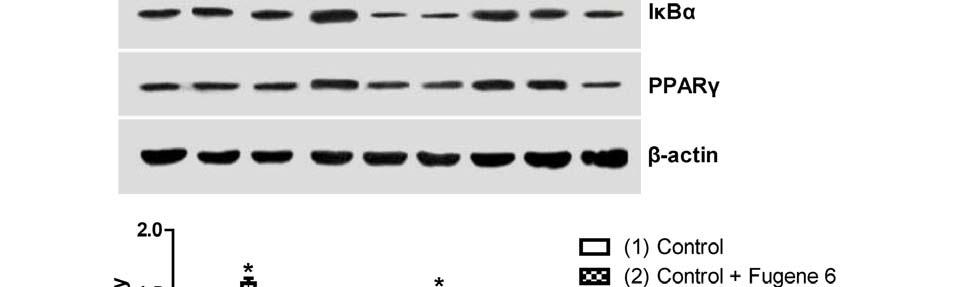
14 CYP2J2 overexpression were apparently inhibited when cells were pretreated with AG1478 or GW9662. Similarly, exogenous treatment with 11,12-EET also increased Akt phosphorylation and enhanced PI3K expression, an effect that was blocked by both AG1478 and GW9662 (Figure 4B). Treatment of cells with TNF-α also led to significantly reduced phosphorylation of extracellular signal-regulated kinases (ERK) 1/2 and EGFR in control cells. CYP2J2 overexpression or exogenous 11,12-EET significantly attenuated the TNF-α-induced effects on ERK1/2 and EGFR phosphorylation (Figure 4A-D), and in addition, ERK1/2 inhibitor apigenin (25 um) significantly and partially abolished the protective effects of EETs on TNF-α induced H9C2 cells apoptosis (Figure 4E). Furthermore, the effects of CYP2J2 overexpression and 11,12-EET treatment on EGFR-activation were inhibited by addition of AG1478. GW9662 also attenuated EGFR activation, although the effects were relatively small (Figure 4C and D). Collectively, these findings suggest that CYP2J2 overexpression or exogenous 11,12-EET activates the EGFR-ERK1/2 signaling pathway, a process that may be involved in the anti-apoptotic effects observed in cardiac myocytes. Effects of 11,12-EET treatment and CYP2J2 overexpression on PPARγ and IκBα expression in neonatal rat cardiomyocytes CYP epoxygenase-derived eicosanoids are known to possess potent anti-inflammatory properties, some of which may be mediated by activation of PPARγ and/or inhibition of IκBα degradation (2, 31). Interestingly, CYP2J2 overexpression or addition of exogenous 11,12-EET significantly upregulated PPARγ and IκBα, both in the presence and absence of TNF-α (Figure 5A and 5B). These effects were significantly attenuated by addition of the GW9662 or AG1478 prior to TNF-α treatment (Figure 5A and 5B). Therefore, the EET/PPARγ/IκBα pathway may 14
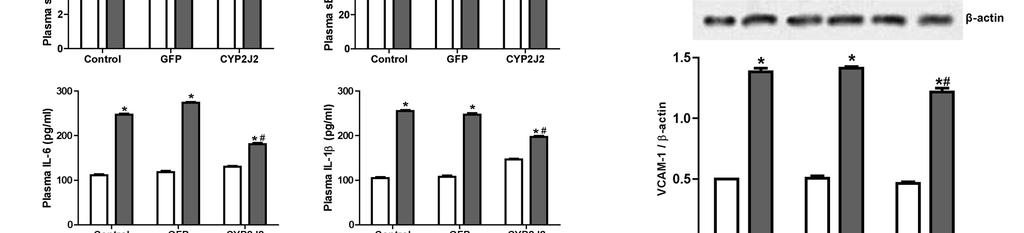
15 represent a novel protective pathway for inhibition of TNF-α-induced effects in cardiomyocytes. Improvement of hemodynamics and systemic inflammation in TNF-α treated rats after CYP2J2 gene delivery Western blot for expression of CYP epoxygenases indicated that CYP2J2 gene delivery induced significant expression of CYP2J2 in vivo in the heart and aorta (Figure S5A). Overexpression of CYP epoxygenases was associated with a significant increase in urinary 14,15-DHET and serum 14,15-EET levels 2 weeks after gene delivery compared to levels in rats injected with saline or with pcdna3.1/gfp (p<0.05; Figure S5B and S5C). These results indicate that CYP2J2 gene delivery induced significant epoxygenase expression and increases in and enzyme activity in vivo. Hemodynamic assessments showed that TNF-α treatment of rats induced a marked deterioration of cardiac function compared with control rats (Table 1). Treatment with pcdna3.1/cyp2j2 resulted in a significant decrease in LVEDP ( # p<0.05) and a notable increase in +dp/dt max ( # p<0.05) compared to saline and pcdna3.1/gfp-treated rats. Together, these results demonstrate an improvement in systolic and diastolic function after CYP2J2 gene delivery. TNF-α injection significantly increased serum levels of pro-inflammatory cytokines including svcam-1, se-selectin, IL-1β and IL-6, but decreased serum levels of IL-10, an anti-inflammatory cytokine (Figure 6A). Interestingly, the pro-inflammatory effects of TNF-α were markedly attenuated by CYP2J2 gene delivery (Figure 6A). Effect of CYP2J2 overexpression on TNF-α-induced adhesion of PBMC to 15
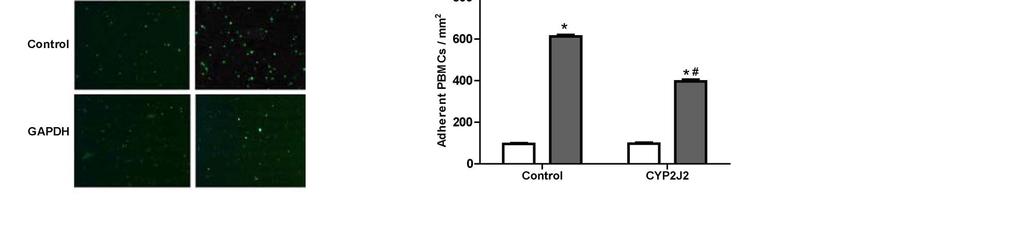
16 endothelial cells CYP2J2 overexpression inhibited TNF-α-induced VCAM-1 expression in cultured HUVECs (Figure 6B). Likewise, TNF-α greatly increased the number of PBMCs which adhered to HUVECs. This effect was significantly attenuated by CYP2J2 overexpression (Figure 6C and 6D). 16
17 Discussion In the present study, we show that CYP2J2 transfection and exogenous addition of 11,12-EET can each significantly increase cell viability and protect against TNF-α-induced apoptosis in both primary neonatal cardiomyocytes and in the H9C2 rat cardiac cell line. Furthermore, CYP2J2 overexpression and exogenous addition of 11,12-EET significantly attenuate the TNF-α-induced increase in caspase-3 activity and decrease in expression of pro-survival proteins Bcl-2 and Bcl-xL, a process which involves activation of PI3K/Akt, EGF/ERK and PPARγ signaling pathways. Finally, in vivo overexpression of CYP2J2 significantly inhibits inflammation and ameliorates the cardiac dysfunction induced by TNF-α. Cardiomyocyte apoptosis and inflammation induced by cytokines such as TNF-α are important in the pathogenesis of acute and chronic myocardial injury and heart failure(32-35). Apoptosis plays an important role in the basic pathogenic mechanisms that underlie a variety of heart diseases. Owing to the chronic nature of heart failure, a very low level of cellular apoptosis over a period of months-years can be detrimental. Importantly, cardiac myocytes are terminally differentiated and have little potential for division as a rescue mechanism. As a result, it is appealing to develop therapeutic strategies for heart failure that enhance cell viability and inhibit apoptosis. The cardioprotective effects of 11,12-EET and CYP2J2 overexpression that we observed in this study are consistent with the observation by Seubert et al. that CYP2J2 overexpression in transgenic mice protects against post-ischemic myocardial dysfunction(18). EETs and CYP2J2 overexpression have also been shown to attenuate cytokine-induced endothelial cell adhesion molecule expression, and endogenous EETs prevented leukocyte adhesion to the vascular wall by a mechanism involving transcription factor NF-κB and IκB kinase(2). The current study 17
18 also demonstrated inhibitory effects of CYP2J2 overexpression on cytokine-induced leukocyte adhesion to endothelial cells. In vivo experiments showed that CYP2J2 overexpression attenuates production of VCAM, E-selectin, IL-1β and IL-6 while enhancing production of the anti-inflammatory cytokine, IL-10. In addition, CYP2J2 gene delivery via intravenous injection significantly increased CYP2J2 expression in vivo in most organs, mainly aorta, heart, liver and kidney (36-38)(3, 5, 6), which may also explain some effects of heart protection. These results support the view that CYP2J2-derived EETs protect the heart via their anti-inflammatory, anti-apoptotic and pro-survival effects. Additional experiments demonstrate an important role for PPARγ and EGFR signaling in mediating the cardioprotective effects of EETs and CYP2J2 overexpression. Treatment of H9C2 cells with GW9662 (a PPARγ inhibitor) or AG1478 (an EGFR inhibitor) significantly inhibited the anti-apoptotic and pro-survival effects of 11,12-EET treatment and CYP2J2 overexpression, with EGFR inhibition being especially detrimental. These results demonstrate that EET-induced EGFR transactivation and PPARγ signaling is an essential event in preventing cardiac myocyte apoptosis and enhancing survival. These results are supported by previous studies in other cell types showing EGFR transactivation in response to EET treatment (9, 39). PPARγ agonists have consistently been shown to decrease both chemokine secretion and matrix metalloproteinase (MMP) expression by inhibiting NF-κB activation(40-42). PPARγ activation is also known to attenuate inflammation and apoptosis during cardiac ischemia and injury (43). A study by Liu et al. showed that PPARγ inhibition abolished the EET/AUDA-mediated anti-inflammatory effect in endothelial cells subjected to laminar flow (31). In addition, PPARγ is an effector of 18
19 EETs via Competition and direct binding assays and PPARγ mediated anti-inflammation effects of EETs in endothelium(31); PPARγ agonist reduced LPS induced NF-κB activation by antagonizing NF-κB DNA activation(44). In addition, PPARγ agonists similarly inhibit cardiomyocyte inflammatory responses in culture (45, 46). Consistent with these findings, we found that 11,12-EET and CYP2J2 overexpression partially, but significantly prevented the loss of PPARγ expression that is normally induced by TNF-α. Furthermore, we also found that the PPARγ inhibitor GW9662 hindered the 11,12-EET and CYP2J2-mediated downregulation of caspase-3 activity and Akt/ERK activation, which may be linked to increase in heparin-binding epithelial growth factor (HB-EGF) release(39, 47). Together, these findings suggest that PPARγ activation plays some role in EET-mediated cardioprotection (Scheme 1). In our study, CYP2J2 overexpression or EET activates MAPK1/2, but PPARγ inhibitor GW9662 blocked this effect; However, PPARγ inhibitor GW9662 has much less inhibitory effect on EET-mediated EGFR activation. These results suggest that PPARγ in heart may have crosstalk with EGFR downstream pathway. In previous studies, we demonstrated that overexpression of epoxygenases can reduce systolic blood pressure in hypertensive rats via enhancing production in cardiomyocytes(36), attenuate pulmonary artery hypertension with significantly decreased inflammatory cytokines in macrataline induced pulmonary artery hypertension model; EETs protect endothelial cells from apoptosis and increased enos activity and expression level(48). Dr. Liao demonastrated that EETs inhibited leukocyte adheresion to endothelial cells and reduced IL-1 via inhibition of NF-κB in endothelial cells(2). These data indicated that EETs really have systemic effects and these cardiovascular protective effects shold be able to exert benefit effects on heart, 19
20 especially decrease in inflammatory cytokines. In the present study, artery blood pressure is not high and therefore overexpression of epoxygenase has no hypotensive effect. This study has some limitations. Further studiess are needed to figure out whether heart protective effects of EETs in antiinflammation and anti-apoptosis involve activation of G-protein coupled receptors and their hyperpolarization role via activating calcium-activated potassium channels, as well as more detailed molecular mechanisms and targets of EETs. In summary, the present study provides insight on the protective role of EETs against cardiac dysfunction and myocardial injuries. The potency of EET-mediated cardiac protection suggests that abundant CYP2J2 expression and EET production in the myocardium are important protective factors whose mechanisms are likely mediated via EGFR and PPARγ activation. Our data support the development of tools to enhance endogenous EETs and/or to deliver exogenous EETs as a novel approach to the treatment of cardiovascular complications including heart failure. 20
21 Acknowledgements: This research was supported by grant from the National Natural Science Foundation of China (No , and ) and National Basic Research Program of China (973 Program) (No. 2007CB512004). This research was also supported in part by the Intramural Research Program of the National Institutes of Health, National Institute of Environmental Health Sciences. Gang Zhao, Jianing Wang and Xizhen Xu equally contributed to this work. 21
22 References: 1. Capdevila, J. H., J. R. Falck, and R. C. Harris Cytochrome P450 and arachidonic acid bioactivation. Molecular and functional properties of the arachidonate monooxygenase. J Lipid Res 41: Node, K., Y. Huo, X. Ruan, B. Yang, M. Spiecker, K. Ley, D. C. Zeldin, and J. K. Liao Anti-inflammatory properties of cytochrome P450 epoxygenase-derived eicosanoids. Science 285: Miura, H., and D. D. Gutterman Human coronary arteriolar dilation to arachidonic acid depends on cytochrome P-450 monooxygenase and Ca2+-activated K+ channels. Circ Res 83: Fisslthaler, B., R. Popp, L. Kiss, M. Potente, D. R. Harder, I. Fleming, and R. Busse Cytochrome P450 2C is an EDHF synthase in coronary arteries. Nature 401: Casolari, C., T. Rossi, G. Baggio, A. Coppi, G. Zandomeneghi, A. I. Ruberto, C. Farina, G. Fabio, A. Zanca, and M. Castelli Interaction between saquinavir and antimycotic drugs on C. albicans and C. neoformans strains. Pharmacol Res 50: Sun, J., X. Sui, J. A. Bradbury, D. C. Zeldin, M. S. Conte, and J. K. Liao Inhibition of vascular smooth muscle cell migration by cytochrome p450 epoxygenase-derived eicosanoids. Circ Res 90: Yang, S., L. Lin, J. X. Chen, C. R. Lee, J. M. Seubert, Y. Wang, H. Wang, Z. R. Chao, D. D. Tao, J. P. Gong, Z. Y. Lu, D. W. Wang, and D. C. Zeldin Cytochrome P-450 epoxygenases protect endothelial cells from apoptosis induced by tumor necrosis factor-alpha via MAPK and PI3K/Akt signaling pathways. Am J Physiol Heart Circ Physiol 293: H142-H Jiang, J. G., R. J. Chen, B. Xiao, S. Yang, J. N. Wang, Y. Wang, L. A. Cowart, X. Xiao, D. W. Wang, and Y. Xia Regulation of endothelial nitric-oxide synthase activity through phosphorylation in response to epoxyeicosatrienoic acids. Prostaglandins Other Lipid Mediat 82: Chen, J. K., J. Capdevila, and R. C. Harris Heparin-binding EGF-like growth factor mediates the biological effects of P450 arachidonate epoxygenase metabolites in epithelial cells. Proc Natl Acad Sci U S A 99: Zhang, C., and D. R. Harder Cerebral capillary endothelial cell mitogenesis and morphogenesis induced by astrocytic epoxyeicosatrienoic Acid. Stroke 33: Medhora, M., J. Daniels, K. Mundey, B. Fisslthaler, R. Busse, E. R. Jacobs, and D. R. Harder Epoxygenase-driven angiogenesis in human lung microvascular endothelial cells. Am J Physiol Heart Circ Physiol 284: H215-H Michaelis, U. R., B. Fisslthaler, M. Medhora, D. Harder, I. Fleming, and R. Busse Cytochrome P450 2C9-derived epoxyeicosatrienoic acids induce angiogenesis via cross-talk with the epidermal growth factor receptor (EGFR). Faseb J 17: Pratt, P. F., M. Medhora, and D. R. Harder Mechanisms regulating cerebral blood flow as therapeutic targets. Curr Opin Investig Drugs 5: Michaelis, U. R., J. R. Falck, R. Schmidt, R. Busse, and I. Fleming Cytochrome P4502C9-derived epoxyeicosatrienoic acids induce the expression of cyclooxygenase-2 in endothelial cells. Arterioscler Thromb Vasc Biol 25: Graber, M. N., A. Alfonso, and D. L. Gill Recovery of Ca2+ pools and growth in Ca2+ pool-depleted cells is mediated by specific epoxyeicosatrienoic acids derived from arachidonic acid. J Biol Chem 272: Chen, J. K., J. Capdevila, and R. C. Harris Overexpression of C-terminal Src kinase blocks 14, 15-epoxyeicosatrienoic acid-induced tyrosine phosphorylation and mitogenesis. J Biol Chem 275: Chen, J. K., D. W. Wang, J. R. Falck, J. Capdevila, and R. C. Harris Transfection of an active cytochrome P450 arachidonic acid epoxygenase indicates that 14,15-epoxyeicosatrienoic acid functions as an intracellular second messenger in response to epidermal growth factor. J Biol Chem 274: Seubert, J., B. Yang, J. A. Bradbury, J. Graves, L. M. Degraff, S. Gabel, R. Gooch, J. Foley, J. Newman, L. Mao, H. A. Rockman, B. D. Hammock, E. Murphy, and D. C. Zeldin Enhanced postischemic functional recovery in CYP2J2 transgenic hearts involves mitochondrial ATP-sensitive K+ channels and p42/p44 MAPK pathway. Circ Res 95: Gross, G. J., J. R. Falck, E. R. Gross, M. Isbell, J. Moore, and K. Nithipatikom Cytochrome P450 and arachidonic acid metabolites: role in myocardial ischemia/reperfusion injury revisited. Cardiovasc Res 68: Jiang, J. G., C. L. Chen, J. W. Card, S. Yang, J. X. Chen, X. N. Fu, Y. G. Ning, X. Xiao, D. C. Zeldin, and D. W. Wang Cytochrome P450 2J2 promotes the neoplastic phenotype of carcinoma cells and is up-regulated in human tumors. Cancer Res 65: Yang, B., L. Graham, S. Dikalov, R. P. Mason, J. R. Falck, J. K. Liao, and D. C. Zeldin Overexpression of cytochrome P450 CYP2J2 protects against hypoxia-reoxygenation injury in cultured bovine aortic endothelial cells. Mol Pharmacol 60:
23 22. Prasad, A. M., H. Ma, C. Sumbilla, D. I. Lee, M. G. Klein, and G. Inesi Phenylephrine hypertrophy, Ca2+-ATPase (SERCA2), and Ca2+ signaling in neonatal rat cardiac myocytes. Am J Physiol Cell Physiol 292: C2269-C Wang, Y., X. Wei, X. Xiao, R. Hui, J. W. Card, M. A. Carey, D. W. Wang, and D. C. Zeldin Arachidonic acid epoxygenase metabolites stimulate endothelial cell growth and angiogenesis via mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt signaling pathways. J Pharmacol Exp Ther 314: Kotamraju, S., E. A. Konorev, J. Joseph, and B. Kalyanaraman Doxorubicin-induced apoptosis in endothelial cells and cardiomyocytes is ameliorated by nitrone spin traps and ebselen. Role of reactive oxygen and nitrogen species. J Biol Chem 275: Zhao, C., P. Wang, X. Xiao, J. Chao, L. Chao, D. W. Wang, and D. C. Zeldin Gene therapy with human tissue kallikrein reduces hypertension and hyperinsulinemia in fructose-induced hypertensive rats. Hypertension 42: Xu, X., W. Wan, L. Ji, S. Lao, A. S. Powers, W. Zhao, J. M. Erikson, and J. Q. Zhang Exercise training combined with angiotensin II receptor blockade limits post-infarct ventricular remodelling in rats. Cardiovasc Res 78: Luu, N. T., G. E. Rainger, and G. B. Nash Kinetics of the different steps during neutrophil migration through cultured endothelial monolayers treated with tumour necrosis factor-alpha. J Vasc Res 36: Tsouknos, A., G. B. Nash, and G. E. Rainger Monocytes initiate a cycle of leukocyte recruitment when cocultured with endothelial cells. Atherosclerosis 170: Wang, H., L. Lin, J. Jiang, Y. Wang, Z. Y. Lu, J. A. Bradbury, F. B. Lih, D. W. Wang, and D. C. Zeldin Up-regulation of endothelial nitric-oxide synthase by endothelium-derived hyperpolarizing factor involves mitogen-activated protein kinase and protein kinase C signaling pathways. J Pharmacol Exp Ther 307: Wu, S., C. R. Moomaw, K. B. Tomer, J. R. Falck, and D. C. Zeldin Molecular cloning and expression of CYP2J2, a human cytochrome P450 arachidonic acid epoxygenase highly expressed in heart. J Biol Chem 271: Liu, Y., Y. Zhang, K. Schmelzer, T. S. Lee, X. Fang, Y. Zhu, A. A. Spector, S. Gill, C. Morisseau, B. D. Hammock, and J. Y. Shyy The antiinflammatory effect of laminar flow: the role of PPARgamma, epoxyeicosatrienoic acids, and soluble epoxide hydrolase. Proc Natl Acad Sci U S A 102: Gottlieb, R. A., K. O. Burleson, R. A. Kloner, B. M. Babior, and R. L. Engler Reperfusion injury induces apoptosis in rabbit cardiomyocytes. J Clin Invest 94: Krown, K. A., M. T. Page, C. Nguyen, D. Zechner, V. Gutierrez, K. L. Comstock, C. C. Glembotski, P. J. Quintana, and R. A. Sabbadini Tumor necrosis factor alpha-induced apoptosis in cardiac myocytes. Involvement of the sphingolipid signaling cascade in cardiac cell death. J Clin Invest 98: Kubota, T., C. F. McTiernan, C. S. Frye, S. E. Slawson, B. H. Lemster, A. P. Koretsky, A. J. Demetris, and A. M. Feldman Dilated cardiomyopathy in transgenic mice with cardiac-specific overexpression of tumor necrosis factor-alpha. Circ Res 81: Horton, J. W., D. L. Maass, and C. Ballard-Croft Rho-associated kinase modulates myocardial inflammatory cytokine responses. Shock 24: Xiao, B., X. Li, J. Yan, X. Yu, G. Yang, X. Xiao, J. W. Voltz, D. C. Zeldin, and D. W. Wang Overexpression of cytochrome P450 epoxygenases prevents development of hypertension in spontaneously hypertensive rats by enhancing atrial natriuretic peptide. J Pharmacol Exp Ther 334: Wang, Y., X. Wei, X. Xiao, R. Hui, J. W. Card, M. A. Carey, D. W. Wang, and D. C. Zeldin Arachidonic acid epoxygenase metabolites stimulate endothelial cell growth and angiogenesis via mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt signaling pathways. J Pharmacol Exp Ther 314: Zheng, C., L. Wang, R. Li, B. Ma, L. Tu, X. Xu, R. T. Dackor, D. C. Zeldin, and D. W. Wang Gene delivery of cytochrome p450 epoxygenase ameliorates monocrotaline-induced pulmonary artery hypertension in rats. Am J Respir Cell Mol Biol 43: Cheng, L. M., J. G. Jiang, Z. Y. Sun, C. Chen, R. T. Dackor, D. C. Zeldin, and D. W. Wang The epoxyeicosatrienoic acid-stimulated phosphorylation of EGF-R involves the activation of metalloproteinases and the release of HB-EGF in cancer cells. Acta Pharmacol Sin 31: Ricote, M., A. C. Li, T. M. Willson, C. J. Kelly, and C. K. Glass The peroxisome proliferator-activated receptor-gamma is a negative regulator of macrophage activation. Nature 391: Chinetti, G., J. C. Fruchart, and B. Staels Peroxisome proliferator-activated receptors (PPARs): nuclear receptors with functions in the vascular wall. Z Kardiol 90 Suppl 3: Moraes, L. A., L. Piqueras, and D. Bishop-Bailey Peroxisome proliferator-activated receptors and inflammation. Pharmacol Ther 110: Liu, H. R., L. Tao, E. Gao, B. L. Lopez, T. A. Christopher, R. N. Willette, E. H. Ohlstein, T. L. Yue, and X. L. Ma Anti-apoptotic effects of rosiglitazone in hypercholesterolemic rabbits subjected to myocardial 23
24 ischemia and reperfusion. Cardiovasc Res 62: Takano, H., T. Nagai, M. Asakawa, T. Toyozaki, T. Oka, I. Komuro, T. Saito, and Y. Masuda Peroxisome proliferator-activated receptor activators inhibit lipopolysaccharide-induced tumor necrosis factor-alpha expression in neonatal rat cardiac myocytes. Circ Res 87: Ding, G., L. Cheng, Q. Qin, S. Frontin, and Q. Yang PPARdelta modulates lipopolysaccharide-induced TNFalpha inflammation signaling in cultured cardiomyocytes. J Mol Cell Cardiol 40: Ye, P., H. Fang, X. Zhou, Y. L. He, and Y. X. Liu Effect of peroxisome proliferator-activated receptor activators on tumor necrosis factor-alpha expression in neonatal rat cardiac myocytes. Chin Med Sci J 19: Chen, J. K., J. Capdevila, and R. C. Harris Heparin-binding EGF-like growth factor mediates the biological effects of P450 arachidonate epoxygenase metabolites in epithelial cells. Proc Natl Acad Sci U S A 99: Xu, X., X. A. Zhang, and D. W. Wang The roles of CYP450 epoxygenases and metabolites, epoxyeicosatrienoic acids, in cardiovascular and malignant diseases. Adv Drug Deliv Rev 63:
25 Figure legends Figure 1. Effect of epoxygenase overexpression and exogenous EETs on apoptosis of primary rat cardiomyocytes. A) Exogenous administration of EETs decreases apoptosis in primary rat cardiomyocytes induced by TNF-α treatment, while the apoptosis is increased by treatment with PPOH compared to the corresponding cells (PPOH, a selective inhibitor of CYP epoxygenases) (*p<0.05 vs. control, # p<0.05 vs. the corresponding group without TNFα treatment). B) Apoptosis of primary rat cardiomyocytes induced by TNF-α is decreased by raav-cyp2j2 transfection and increased by PPOH treatment (*p<0.05 vs. control, # p<0.05 vs. raav-gfp + TNFα, and & p<0.05 vs. raav-2j2 + TNFα). C and D) Representative western blots and densitometry results showing altered levels of Bcl-2, Bcl-xL and Bax following exogenous administration of EETs or raav-cyp2j2 transfection (*p<0.05 vs. control, # p<0.05 vs. raav-gfp + TNFα). Figure 2. Effect of CYP epoxygenase overexpression and exogenous EET treatment on viability and apoptosis of H9C2 rat heart cells. A) CYP2J2 overexpression increases cell viability and inhibits the detrimental effects of TNF-α. The CYP2J2-mediated increase in cell viability is notably attenuated by co-incubation with AG1478 or GW9662 ( * p<0.05 vs. the control; # p<0.05 compared to the GFP + TNF-α; & p<0.05 compared to CYP2J2 + TNF-α. B) Viability of H9C2 cells is increased after treatment with 11,12-EET. The EET-mediated increase in cell viability is attenuated by AG1478 and GW9662 (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 compared to 11,12-EET + TNF-α). C) TNF-α-induced apoptosis of H9C2 cells is inhibited by CYP2J2 overexpression, an effect that is attenuated by co-incubation with AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α; & p<0.05 vs. CYP2J2 + TNF-α. D) TNF-α-induced apoptosis of H9C2 cells is 25
26 inhibited by 11,12-EET treatment, an effect that is attenuated by AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 vs. 11,12-EET + TNF-α). Figure 3. Effect of CYP2J2 overexpression and exogenous EET treatment on caspase-3 activity and apoptosis-regulating proteins in H9C2 rat heart cells. A) CYP2J2 overexpression attenuates TNF-α-induced caspase-3 activity, an effect that is attenuated by either AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α; & p<0.05 vs. CYP2J2 + TNF-α). B) 11,12-EET treatment attenuates TNF-α-induced caspase-3 activity. The EET-mediated effect is attenuated b y AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 vs. 11,12-EET+ TNF-α). C) and D) Representative western blots and densitometry results showing altered levels of Bcl-2, Bcl-xL and Bax following exogenous administration of EETs or overexpression of CYP2J2, and effects of AG1478 and GW9662 (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α or vs. GFP + TNF-α; & p<0.05 vs. 11,12-EET + TNF-α or vs. CYP2J2 + TNF-α). Figure 4. Effect of CYP2J2 overexpression and 11,12-EET on activity of PI3K/Akt, ERK1/2 and EGFR in cardiac myocytes. A) Representative western blots and densitometry results of PI3K/β-actin, phospho-akt /Akt and phospho-erk1/2/erk1/2 upon CYP2J2 transfection with or without AG1478 or GW9662. (*p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α; & p<0.05 vs. CYP2J2 + TNF-α). B) Representative western blots and densitometry results of PI3K/β-actin, phospho-akt/akt and phospho-erk1/2/erk1/2 upon 11,12-EET incubation with or without AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 vs. 11,12-EET + TNF-α). C) Representative western blots and densitometry results of phospho-egfr upon CYP2J2 transfection with or without AG1478 or GW9662 (*p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α; & p<0.05 vs. CYP2J2 + TNF-α. D) 26
27 Representative western blots and densitometry results of phospho-egfr upon 11,12-EET incubation with or without AG1478 or GW9662; E) Effect of MAPK1/2 inhibitor apigenin (25 um) on TNFα-induced cardiomyocyte apoptosis (*p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 vs. 11,12-EET + TNF-α). Figure 5. Effect of CYP2J2 overexpression and 11,12-EET treatment on PPARγ and IκBα expression in neonatal rat cardiac myocytes. A) Representative western blots and densitometry results showing effects of CYP2J2 overexpression on PPARγ and IκBα expression in TNF-α treated neonatal rat cardiomyocytes (*p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α; & p<0.05 vs. CYP2J2 + TNF-α). B) Representative western blots and densitometry results showing effects of 11,12-EET treatment on PPARγ and IκBα expression in TNF-α treated neonatal rat cardiomyocytes ( * p<0.05 vs. control; # p<0.05 vs. DMSO + TNF-α; & p<0.05 vs. 11,12-EET + TNF-α). Figure 6. Effect of CYP2J2 gene delivery on in vivo inflammatory cytokine production and in vitro adhesion of PBMCs to endothelial cells. A) Inflammatory cytokine levels in serum of rats treated with TNF-α (n=8 for each group; *p<0.05 vs. control without TNF-α; # p<0.05 vs. GFP with TNF-α. B) Representative western blots and densitometry results of VCAM-1 in HUVECs treated with TNF-α (n=8 for each group; *p<0.05 vs. control; # p<0.05 vs. GFP + TNF-α). C) and D) Representative image (x200) and numbers of PBMCs adhering to TNF-α treated HUVECs (n=8 for each group; *p<0.05 vs. control; # p<0.05 vs. TNF-α). Scheme 1. Signaling pathways of heart protective effects of EETs. indicates inhibitory or blocking effects, and indicates increasing or enhancing effcts. 27
28 Table 1. Effects of CYP2J2 gene delivery on hemodynamic parameters in rats LVEDP(mmHg) +dp/dt max (mmhg/s) Control 1.31 ± ± TNF-α 3.66 ± 0.26* ± 391.4* GFP 1.22 ± ± GFP + TNF-α 3.45 ± 0.84* ± 496.2* CYP2J ± 0.39* ± 57.7* CYP2J2 + TNF-α 2.55 ± 0.88* # ± 79.9* # Values are expressed as mean ± SEM (n=8 per group). *p<0.05 vs. control, # p<0.05 vs. GFP+TNF-α. 28
29 29
30 30
31 31
32 32
33 E 33


34 34
35 35
36 CYP Epoxygenases EETs TNF-alpha EGFR PPARγ PI3K/AKT EGFR NF-KappaB ERK PI3K/AKT ERK PPARγ Mitochondrial dysfunction Caspase-3 Cardiomyocyte Apoptosis Inflammation Cardiomyocytes Injury and Cardiac dysfunction 36
Promotive effects of epoxyeicosatrienoic acids (EETs) on the proliferation of tumor cells
 on the proliferation of tumor cells") [Chinese Journal of Cancer 27:11, 390-395; November Promotive 2008]; effects 2008 of epoxyeicosatrienoic Sun Yat-sen University acids Cancer (EETs) on Center the proliferation of tumor cells Basic Research
[Chinese Journal of Cancer 27:11, 390-395; November Promotive 2008]; effects 2008 of epoxyeicosatrienoic Sun Yat-sen University acids Cancer (EETs) on Center the proliferation of tumor cells Basic Research
Impact factor: Reporter:4A1H0019 Chen Zi Hao 4A1H0023 Huang Wan ting 4A1H0039 Sue Yi Zhu 4A1H0070 Lin Guan cheng 4A1H0077 Chen Bo xuan
 Curcumin Protects Neonatal Rat Cardiomyocytes against High Glucose-Induced Apoptosis via PI3K/Akt Signalling Pathway Wei Yu,1,2 Wenliang Zha,1 Zhiqiang Ke,1 Qing Min,2 Cairong Li,1 Huirong Sun,3 and Chao
Curcumin Protects Neonatal Rat Cardiomyocytes against High Glucose-Induced Apoptosis via PI3K/Akt Signalling Pathway Wei Yu,1,2 Wenliang Zha,1 Zhiqiang Ke,1 Qing Min,2 Cairong Li,1 Huirong Sun,3 and Chao
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Protection against doxorubicin-induced myocardial dysfunction in mice by cardiac-specific expression of carboxyl terminus of hsp70-interacting protein
 Protection against doxorubicin-induced myocardial dysfunction in mice by cardiac-specific expression of carboxyl terminus of hsp70-interacting protein Lei Wang 1, Tian-Peng Zhang 1, Yuan Zhang 2, Hai-Lian
Protection against doxorubicin-induced myocardial dysfunction in mice by cardiac-specific expression of carboxyl terminus of hsp70-interacting protein Lei Wang 1, Tian-Peng Zhang 1, Yuan Zhang 2, Hai-Lian
14,15 DHET Human Urine Test Kit
 14,15 DHET Human Urine Test Kit For measuring free and glucuronidated 14,15 DHET Catalog Number: DH3, DH13, DH23, DH103 V. 10232012 Introduction This competitive ELISA kit is for determination of free
14,15 DHET Human Urine Test Kit For measuring free and glucuronidated 14,15 DHET Catalog Number: DH3, DH13, DH23, DH103 V. 10232012 Introduction This competitive ELISA kit is for determination of free
- 1 - Cell types Monocytes THP-1 cells Macrophages. LPS Treatment time (Hour) IL-6 level (pg/ml)
 IL-6 level (pg/ml)") Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Effects of the angiotensin II type-1 receptor antagonist telmisartan on endothelial activation induced by advanced glycation endproducts
 Effects of the angiotensin II type-1 receptor antagonist telmisartan on endothelial activation induced by advanced glycation endproducts Serena Del Turco, Teresa Navarra, Giuseppina Basta, Raffaele De
Effects of the angiotensin II type-1 receptor antagonist telmisartan on endothelial activation induced by advanced glycation endproducts Serena Del Turco, Teresa Navarra, Giuseppina Basta, Raffaele De
Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108
 37:108") Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108 https://doi.org/10.1186/s13046-018-0774-7 CORRECTION Correction to: Novel smac mimetic APG- 1387 elicits ovarian cancer cell killing
Li et al. Journal of Experimental & Clinical Cancer Research (2018) 37:108 https://doi.org/10.1186/s13046-018-0774-7 CORRECTION Correction to: Novel smac mimetic APG- 1387 elicits ovarian cancer cell killing
renoprotection therapy goals 208, 209
 Subject Index Aldosterone, plasminogen activator inhibitor-1 induction 163, 164, 168 Aminopeptidases angiotensin II processing 64 66, 214 diabetic expression 214, 215 Angiotensin I intrarenal compartmentalization
Subject Index Aldosterone, plasminogen activator inhibitor-1 induction 163, 164, 168 Aminopeptidases angiotensin II processing 64 66, 214 diabetic expression 214, 215 Angiotensin I intrarenal compartmentalization
Supplementary Figure 1:
 Supplementary Figure 1: (A) Whole aortic cross-sections stained with Hematoxylin and Eosin (H&E), 7 days after porcine-pancreatic-elastase (PPE)-induced AAA compared to untreated, healthy control aortas
Supplementary Figure 1: (A) Whole aortic cross-sections stained with Hematoxylin and Eosin (H&E), 7 days after porcine-pancreatic-elastase (PPE)-induced AAA compared to untreated, healthy control aortas
Ginkgo biloba extract postconditioning reduces myocardial ischemia reperfusion injury
 Ginkgo biloba extract postconditioning reduces myocardial ischemia reperfusion injury K. Ran 1, D.-L. Yang 1, Y.-T. Chang 1, K.-M. Duan 2, Y.-W. Ou 2, H.-P. Wang 3 and Z.-J. Li 1 1 Department of Anesthesiology,
Ginkgo biloba extract postconditioning reduces myocardial ischemia reperfusion injury K. Ran 1, D.-L. Yang 1, Y.-T. Chang 1, K.-M. Duan 2, Y.-W. Ou 2, H.-P. Wang 3 and Z.-J. Li 1 1 Department of Anesthesiology,
Adipose Tissue Dysfunction and Diabetic Cardiovascular Disease
 Adipose Tissue Dysfunction and Diabetic Cardiovascular Disease Xin-Liang Ma, MD, PhD Thomas Jefferson University Philadelphia, PA USA CVD As The #1 Causes of Death In USA Male Female Heart Disease and
Adipose Tissue Dysfunction and Diabetic Cardiovascular Disease Xin-Liang Ma, MD, PhD Thomas Jefferson University Philadelphia, PA USA CVD As The #1 Causes of Death In USA Male Female Heart Disease and
CYP2J3 Gene Delivery Reduces Insulin Resistance via Upregulation of enos in Fructose-treated Rats
 CARDIO VASCULAR DIABETOLOGY ORIGINAL INVESTIGATION Open Access CYP2J3 Gene Delivery Reduces Insulin Resistance via Upregulation of enos in Fructose-treated Rats Xizhen Xu 1, Ling Tu 1, Luyun Wang 2, Xiaosai
CARDIO VASCULAR DIABETOLOGY ORIGINAL INVESTIGATION Open Access CYP2J3 Gene Delivery Reduces Insulin Resistance via Upregulation of enos in Fructose-treated Rats Xizhen Xu 1, Ling Tu 1, Luyun Wang 2, Xiaosai
EETs mediate cardioprotection of salvianolic acids through MAPK signaling pathway
 Acta Pharmaceutica Sinica B 2013;3(1):25 31 Institute of Materia Medica, Chinese Academy of Medical Sciences Chinese Pharmaceutical Association Acta Pharmaceutica Sinica B www.elsevier.com/locate/apsb
Acta Pharmaceutica Sinica B 2013;3(1):25 31 Institute of Materia Medica, Chinese Academy of Medical Sciences Chinese Pharmaceutical Association Acta Pharmaceutica Sinica B www.elsevier.com/locate/apsb
B-cell. Astrocyte SCI SCI. T-cell
 RF #2015 P-01 PI: Azizul Haque, PhD Grant Title: Targeting Enolase in Spinal Cord Injury 12-month Technical Progress Report Progress Report (First Six Months): Enolase is one of the most abundantly expressed
RF #2015 P-01 PI: Azizul Haque, PhD Grant Title: Targeting Enolase in Spinal Cord Injury 12-month Technical Progress Report Progress Report (First Six Months): Enolase is one of the most abundantly expressed
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary Figure 1. Confocal immunofluorescence showing mitochondrial translocation of Drp1. Cardiomyocytes treated with H 2 O 2 were prestained
 Supplementary Figure 1. Confocal immunofluorescence showing mitochondrial translocation of Drp1. Cardiomyocytes treated with H 2 O 2 were prestained with MitoTracker (red), then were immunostained with
Supplementary Figure 1. Confocal immunofluorescence showing mitochondrial translocation of Drp1. Cardiomyocytes treated with H 2 O 2 were prestained with MitoTracker (red), then were immunostained with
A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified
 Cell culture and animal model A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum at 37 C in humidified atmosphere containing
Cell culture and animal model A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum at 37 C in humidified atmosphere containing
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Supplementary Materials
 Supplementary Materials Figure S1. MTT Cell viability assay. To measure the cytotoxic potential of the oxidative treatment, the MTT [3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyl tetrazolium bromide] assay
Supplementary Materials Figure S1. MTT Cell viability assay. To measure the cytotoxic potential of the oxidative treatment, the MTT [3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyl tetrazolium bromide] assay
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
PUBLICATIONS. cells. J. Physiol. (London) 517P:91P (Manchester, England, UK).
 517P:91P (Manchester, England, UK).") 277 PUBLICATIONS Abstracts Haddad JJ, Land SC (1999). Differential activation of oxygen-responsive transcription factors over fetal-to-neonatal alveolar oxygen tensions in rat fetal distal lung epithelial
277 PUBLICATIONS Abstracts Haddad JJ, Land SC (1999). Differential activation of oxygen-responsive transcription factors over fetal-to-neonatal alveolar oxygen tensions in rat fetal distal lung epithelial
Interaction between CYP 2C19*3 polymorphism and smoking in relation to laryngeal carcinoma in the Chinese Han population
 Interaction between CYP 2C19*3 polymorphism and smoking in relation to laryngeal carcinoma in the Chinese Han population J. Feng 1,4 *, L. Li 2 *, Y.-S. Zhao 3, S.-Q. Tang 4, H.-B. Yang 4 and S.-X. Liu
Interaction between CYP 2C19*3 polymorphism and smoking in relation to laryngeal carcinoma in the Chinese Han population J. Feng 1,4 *, L. Li 2 *, Y.-S. Zhao 3, S.-Q. Tang 4, H.-B. Yang 4 and S.-X. Liu
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Intracellular MHC class II molecules promote TLR-triggered innate. immune responses by maintaining Btk activation
 Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining Btk activation Xingguang Liu, Zhenzhen Zhan, Dong Li, Li Xu, Feng Ma, Peng Zhang, Hangping Yao and Xuetao
Reperfusion Injury: How Can We Reduce It?
 MI/CAD: Practical Question in Management of AMI Patients Reperfusion Injury: How Can We Reduce It? Hyun-Jai Cho, M.D., Ph.D Cardiovascular Center & Department of Internal Medicine Seoul National University
MI/CAD: Practical Question in Management of AMI Patients Reperfusion Injury: How Can We Reduce It? Hyun-Jai Cho, M.D., Ph.D Cardiovascular Center & Department of Internal Medicine Seoul National University
Full Record.
 第 1 頁, 共 2 頁 Full Record Record 1 of 6 (Set #2) Title: Insulin-like growth factor-1 mediates stretch-induced upregulation of myostatin expression in neonatal rat cardiomyocytes Author(s): Shyu KG, Ko WH,
第 1 頁, 共 2 頁 Full Record Record 1 of 6 (Set #2) Title: Insulin-like growth factor-1 mediates stretch-induced upregulation of myostatin expression in neonatal rat cardiomyocytes Author(s): Shyu KG, Ko WH,
E. Cervio, P. Danieli, C. Ciuffreda, F. Pisano, M. Roccio, M. Gnecchi. The authors have no financial disclosures to declare
 16 th ISCT Annual Meeting SOLUBLE FACTORS RELEASED BY HUMAN MESENCHYMAL STEM CELLS OF FETAL ORIGIN LEAD TO CARDIOMYOCYTE PROTECTION THROUGH THE INHIBITION OF PRO-APOPTOTIC SIGNALING E. Cervio, P. Danieli,
16 th ISCT Annual Meeting SOLUBLE FACTORS RELEASED BY HUMAN MESENCHYMAL STEM CELLS OF FETAL ORIGIN LEAD TO CARDIOMYOCYTE PROTECTION THROUGH THE INHIBITION OF PRO-APOPTOTIC SIGNALING E. Cervio, P. Danieli,
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Effects and mechanisms of Fenofibrate on the secretion of vascular endothelial contraction factors in hypertensive rats
 Effects and mechanisms of Fenofibrate on the secretion of vascular endothelial contraction factors in hypertensive rats Y. Zhu 1, H.-S. Wang 1, X.-M. Li 1 and C. Qu 2 1 Department of Cardiac Surgery, General
Effects and mechanisms of Fenofibrate on the secretion of vascular endothelial contraction factors in hypertensive rats Y. Zhu 1, H.-S. Wang 1, X.-M. Li 1 and C. Qu 2 1 Department of Cardiac Surgery, General
Research article. Department of Pharmacology and Toxicology, Medical College of Georgia, Georgia Health Sciences University, Augusta, Georgia, USA.
 Research article Calpain mediates pulmonary vascular remodeling in rodent models of pulmonary hypertension, and its inhibition attenuates pathologic features of disease Wanli Ma, 1 Weihong Han, 1 Peter
Research article Calpain mediates pulmonary vascular remodeling in rodent models of pulmonary hypertension, and its inhibition attenuates pathologic features of disease Wanli Ma, 1 Weihong Han, 1 Peter
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Figure S I: Effects of D4F on body weight and serum lipids in apoe -/- mice.
 Supplementary Figures: Supplementary Figure S I: Effects of D4F on body weight and serum lipids in apoe -/- mice. Male apoe -/- mice were fed a high-fat diet for 8 weeks, and given PBS (model group) or
Supplementary Figures: Supplementary Figure S I: Effects of D4F on body weight and serum lipids in apoe -/- mice. Male apoe -/- mice were fed a high-fat diet for 8 weeks, and given PBS (model group) or
ulcer healing role 118 Bicarbonate, prostaglandins in duodenal cytoprotection 235, 236
 Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
Subject Index Actin cellular forms 48, 49 epidermal growth factor, cytoskeletal change induction in mucosal repair 22, 23 wound repair 64, 65 polyamine effects on cytoskeleton 49 51 S-Adenosylmethionine
A novel bfgf antagonist peptide inhibits breast cancer cell growth
 210 A novel bfgf antagonist peptide inhibits breast cancer cell growth QUCHOU LI 1, SUSU GAO 1, YONGLIN YU 1, WENHUI WANG 1, XILEI CHEN 1, RUIXUE WANG 1, TAO LI 1, CONG WANG 1, XIAOKUN LI 1,2 and XIAOPING
210 A novel bfgf antagonist peptide inhibits breast cancer cell growth QUCHOU LI 1, SUSU GAO 1, YONGLIN YU 1, WENHUI WANG 1, XILEI CHEN 1, RUIXUE WANG 1, TAO LI 1, CONG WANG 1, XIAOKUN LI 1,2 and XIAOPING
PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells
 PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells C.-G. Sun 1 *, J. Zhuang 1 *, W.-J. Teng 1, Z. Wang 2 and S.-S. Du 3 1 Department of Oncology,
PUMA gene transfection can enhance the sensitivity of epirubicin-induced apoptosis of MCF-7 breast cancer cells C.-G. Sun 1 *, J. Zhuang 1 *, W.-J. Teng 1, Z. Wang 2 and S.-S. Du 3 1 Department of Oncology,
Supporting Information
 Copyright WILEY-VCH Verlag GmbH & Co. KGaA, 69469 Weinheim, Germany, 212. Supporting Information for Adv. Funct. Mater., DOI:.2/adfm.2122233 MnO Nanocrystals: A Platform for Integration of MRI and Genuine
Copyright WILEY-VCH Verlag GmbH & Co. KGaA, 69469 Weinheim, Germany, 212. Supporting Information for Adv. Funct. Mater., DOI:.2/adfm.2122233 MnO Nanocrystals: A Platform for Integration of MRI and Genuine
Introduction. Acute sodium overload produces renal tubulointerstitial inflammation in normal rats
 Acute sodium overload produces renal tubulointerstitial inflammation in normal rats MI Roson, et al. Kidney International (2006) Introduction Present by Kanya Bunnan and Wiraporn paebua Tubular sodium
Acute sodium overload produces renal tubulointerstitial inflammation in normal rats MI Roson, et al. Kidney International (2006) Introduction Present by Kanya Bunnan and Wiraporn paebua Tubular sodium
Dr. Hany M. Abo-Haded
 BY Dr. Hany M. Abo-Haded Pediatric Cardiology Unit, Mansoura Universty Children Hospital, Mansoura, Egypt Co-authors: Dina S El-Agamy and Mohamed A Elkablawy Background Doxorubicin (DOX) is an anthracycline
BY Dr. Hany M. Abo-Haded Pediatric Cardiology Unit, Mansoura Universty Children Hospital, Mansoura, Egypt Co-authors: Dina S El-Agamy and Mohamed A Elkablawy Background Doxorubicin (DOX) is an anthracycline
Supporting Information
 Supporting Information Natural small molecule FMHM inhibits lipopolysaccharide-induced inflammatory response by promoting TRAF6 degradation via K48-linked polyubiquitination Ke-Wu Zeng 1, Li-Xi Liao 1,
Supporting Information Natural small molecule FMHM inhibits lipopolysaccharide-induced inflammatory response by promoting TRAF6 degradation via K48-linked polyubiquitination Ke-Wu Zeng 1, Li-Xi Liao 1,
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
alveolar macrophages (AMs) after 24 hours of in vitro culture in complete medium
 after 24 hours of in vitro culture in complete medium") Online Supplement for: NF-κB ACTIVATION IN ALVEOLAR MACROPHAGES REQUIRES IκB KINASE-β, BUT NOT NF-κB INDUCING KINASE Supershift and Competition Assays for NF-κB Competition and supershift assays were performed
Online Supplement for: NF-κB ACTIVATION IN ALVEOLAR MACROPHAGES REQUIRES IκB KINASE-β, BUT NOT NF-κB INDUCING KINASE Supershift and Competition Assays for NF-κB Competition and supershift assays were performed
a. b. c. d. e. f. g. h. i. j. k. l. m. n. o. p.
 a. b. c. d. e. f. g. h. i. j. k. l. 2.5 2 1.5 1.5 IL-1β 12 8 6 4 2 IL-1β 9 8 7 6 4 3 3 2.9 IL-1β m. n. o. p. 1.8 1.6 1.4 1.2 1.8.6.4.2 6h LPS 2 15 1 5 6h LPS 2 6h LPS 6 4 3 6h LPS Supplementary Figure
a. b. c. d. e. f. g. h. i. j. k. l. 2.5 2 1.5 1.5 IL-1β 12 8 6 4 2 IL-1β 9 8 7 6 4 3 3 2.9 IL-1β m. n. o. p. 1.8 1.6 1.4 1.2 1.8.6.4.2 6h LPS 2 15 1 5 6h LPS 2 6h LPS 6 4 3 6h LPS Supplementary Figure
c Ischemia (30 min) Reperfusion (8 w) Supplementary Figure bp 300 bp Ischemia (30 min) Reperfusion (4 h) Dox 20 mg/kg i.p.
 Reperfusion (8 w) Supplementary Figure bp 300 bp Ischemia (30 min) Reperfusion (4 h) Dox 20 mg/kg i.p.") a Marker Ripk3 +/ 5 bp 3 bp b Ischemia (3 min) Reperfusion (4 h) d 2 mg/kg i.p. 1 w 5 w Sacrifice for IF size A subset for echocardiography and morphological analysis c Ischemia (3 min) Reperfusion (8
a Marker Ripk3 +/ 5 bp 3 bp b Ischemia (3 min) Reperfusion (4 h) d 2 mg/kg i.p. 1 w 5 w Sacrifice for IF size A subset for echocardiography and morphological analysis c Ischemia (3 min) Reperfusion (8
Molecular Basis of Cardioprotection by Erythropoietin
 56 Current Molecular Pharmacology, 2009, 2, 56-69 Molecular Basis of Cardioprotection by Erythropoietin D. Burger 1, A. Xenocostas 2,3 and Q.P. Feng*,1,2,3 1 Department of Physiology and Pharmacology,
56 Current Molecular Pharmacology, 2009, 2, 56-69 Molecular Basis of Cardioprotection by Erythropoietin D. Burger 1, A. Xenocostas 2,3 and Q.P. Feng*,1,2,3 1 Department of Physiology and Pharmacology,
The biological bases of treatment related cardiac and lung toxicity
 The biological bases of treatment related cardiac and lung toxicity Monica Mangoni Cardiopulmonary toxicity of anticancer treatments Chemotherapy TKIs MoAB CARDIAC TOXICITY Radiation effect -------------------------------Early
The biological bases of treatment related cardiac and lung toxicity Monica Mangoni Cardiopulmonary toxicity of anticancer treatments Chemotherapy TKIs MoAB CARDIAC TOXICITY Radiation effect -------------------------------Early
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Supporting Information. FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce. apoptosis
 1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
J Jpn Coll Angiol, 2009, 49:
 Online publication October 6, 2009 48 2 20 J Jpn Coll Angiol, 2009, 49: 293 297 atrial natriuretic peptide, brain natriuretic peptide, guanylyl cyclase-a receptor, cardiac remodeling, cardiac hypertrophy
Online publication October 6, 2009 48 2 20 J Jpn Coll Angiol, 2009, 49: 293 297 atrial natriuretic peptide, brain natriuretic peptide, guanylyl cyclase-a receptor, cardiac remodeling, cardiac hypertrophy
Equivalent Cardioprotection Induced by Ischemic and Hypoxic Preconditioning
 Original Cardiovascular 229 Equivalent Cardioprotection Induced by Ischemic and Hypoxic Preconditioning Xujin Xiang 1 Haixia Lin 2 Jin Liu 1 Zeyan Duan 1 1 Department of Anesthesiology, West China Hospital,
Original Cardiovascular 229 Equivalent Cardioprotection Induced by Ischemic and Hypoxic Preconditioning Xujin Xiang 1 Haixia Lin 2 Jin Liu 1 Zeyan Duan 1 1 Department of Anesthesiology, West China Hospital,
Supplementary Materials and Methods
 Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
SCIRF Award #2016 I-03 PI: Azizul Haque, PhD Grant Title: Neuron-specific Enolase and SCI
 SCIRF Award #2016 I-03 PI: Azizul Haque, PhD Grant Title: Neuron-specific Enolase and SCI 10-month Technical Progress Report Enolase is a multifunctional glycolytic enzyme involved in growth control, hypoxia,
SCIRF Award #2016 I-03 PI: Azizul Haque, PhD Grant Title: Neuron-specific Enolase and SCI 10-month Technical Progress Report Enolase is a multifunctional glycolytic enzyme involved in growth control, hypoxia,
Integrin CD11b negatively regulates TLR-triggered inflammatory responses by. activating Syk and promoting MyD88 and TRIF degradation via cbl-b
 Integrin CD11b negatively regulates TLR-triggered inflammatory responses by activating Syk and promoting MyD88 and TRIF degradation via cbl-b Chaofeng Han, Jing Jin, Sheng Xu, Haibo Liu, Nan Li, and Xuetao
Integrin CD11b negatively regulates TLR-triggered inflammatory responses by activating Syk and promoting MyD88 and TRIF degradation via cbl-b Chaofeng Han, Jing Jin, Sheng Xu, Haibo Liu, Nan Li, and Xuetao
Correlations among copeptin, ischemia-modified albumin, and the extent of myocardial injury in patients with acute carbon monoxide poisoning
 Correlations among copeptin, ischemia-modified albumin, and the extent of myocardial injury in patients with acute carbon monoxide poisoning J. Li, J.S. Wang, Z.X. Xie, W.Z. Wang, L. Wang, G.Y. Ma, Y.Q.
Correlations among copeptin, ischemia-modified albumin, and the extent of myocardial injury in patients with acute carbon monoxide poisoning J. Li, J.S. Wang, Z.X. Xie, W.Z. Wang, L. Wang, G.Y. Ma, Y.Q.
C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is
 is a n«sjfc&c- waefc-jduble phycobiliprotein. pigment isolated from Spirulina platensis. This water- soluble protein pigment is") ' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
' ^Summary C-Phycocyanin (C-PC) is a n«sjfc&c- waefc-jduble phycobiliprotein pigment isolated from Spirulina platensis. This water- soluble protein pigment is of greater importance because of its various
The Study of Endothelial Function in CKD and ESRD
 The Study of Endothelial Function in CKD and ESRD Endothelial Diversity in the Human Body Aird WC. Circ Res 2007 Endothelial Diversity in the Human Body The endothelium should be viewed for what it is:
The Study of Endothelial Function in CKD and ESRD Endothelial Diversity in the Human Body Aird WC. Circ Res 2007 Endothelial Diversity in the Human Body The endothelium should be viewed for what it is:
Arachidonic acid cytochrome P450 epoxygenase pathway
 Arachidonic acid cytochrome P450 epoxygenase pathway Arthur A. Spector 1 Department of Biochemistry, Carver College of Medicine, University of Iowa, Iowa City, IA 52242 Abstract Cytochrome P450 (CYP) epoxygenases
Arachidonic acid cytochrome P450 epoxygenase pathway Arthur A. Spector 1 Department of Biochemistry, Carver College of Medicine, University of Iowa, Iowa City, IA 52242 Abstract Cytochrome P450 (CYP) epoxygenases
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Supplementary Information
 Supplementary Information Figure S1. Int6 gene silencing efficiency. (A) Western Blot analysis of Int6 expression at different times after sirna transfection. Int6 expression is strongly silenced in Int6
Supplementary Information Figure S1. Int6 gene silencing efficiency. (A) Western Blot analysis of Int6 expression at different times after sirna transfection. Int6 expression is strongly silenced in Int6
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Arachidonic Acid Epoxygenase Metabolites Stimulate Endothelial Cell Growth. and Angiogenesis via MAP kinase and PI3 Kinase/Akt Signaling Pathways
 JPET This Fast article Forward. has not been Published copyedited and on formatted. April 19, The 2005 final version as DOI:10.1124/jpet.105.083477 may differ from this version. JPET #83477 Arachidonic
JPET This Fast article Forward. has not been Published copyedited and on formatted. April 19, The 2005 final version as DOI:10.1124/jpet.105.083477 may differ from this version. JPET #83477 Arachidonic
Reactive oxygen species: Importance for ischemia/reperfusion (injury)
") Physiologisches Institut Reactive oxygen species: Importance for ischemia/reperfusion (injury) Prof. Dr. Rainer Schulz Reactive oxygen species (ROS) in ischemia/reperfusion injury (IRI) ROS GOOD: Endogenous
Physiologisches Institut Reactive oxygen species: Importance for ischemia/reperfusion (injury) Prof. Dr. Rainer Schulz Reactive oxygen species (ROS) in ischemia/reperfusion injury (IRI) ROS GOOD: Endogenous
About OMICS Group Conferences
 About MICS Group MICS Group International is an amalgamation of pen Access publications and worldwide international science conferences and events. Established in the year 2007 with the sole aim of making
About MICS Group MICS Group International is an amalgamation of pen Access publications and worldwide international science conferences and events. Established in the year 2007 with the sole aim of making
The effect of elemene reversing the multidurg resistance of A549/DDP lung cancer cells
 213 12 33 12 TUMOR Vol. 33, December 213 www.tumorsci.org 161 Basic Research DOI: 1.3781/j.issn.1-7431.213.12. 5 Copyright 213 by TUMOR A549/DDP 1, 2 2 1 3 4 1 1 1. 3619 2. 355 3. 3611 4. 361 elemene cisplatin
213 12 33 12 TUMOR Vol. 33, December 213 www.tumorsci.org 161 Basic Research DOI: 1.3781/j.issn.1-7431.213.12. 5 Copyright 213 by TUMOR A549/DDP 1, 2 2 1 3 4 1 1 1. 3619 2. 355 3. 3611 4. 361 elemene cisplatin
Supplementary Material and Methods
 Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
IL-24 AND ITS ROLE IN WOUND HEALING
 IL-24 AND ITS ROLE IN WOUND HEALING Nancy J. Poindexter, Ryan Williams, Garth Powis, Sunil Chada, and Elizabeth A. Grimm & Introgen Therapeutics, Inc., Houston, TX IL-24/MDA 24/MDA-77 is a Tumor Suppressor
IL-24 AND ITS ROLE IN WOUND HEALING Nancy J. Poindexter, Ryan Williams, Garth Powis, Sunil Chada, and Elizabeth A. Grimm & Introgen Therapeutics, Inc., Houston, TX IL-24/MDA 24/MDA-77 is a Tumor Suppressor
Cardiovascular Protection and the RAS
 Cardiovascular Protection and the RAS Katalin Kauser, MD, PhD, DSc Senior Associate Director, Boehringer Ingelheim Pharmaceutical Inc. Micardis Product Pipeline Scientific Support Ridgefield, CT, USA Cardiovascular
Cardiovascular Protection and the RAS Katalin Kauser, MD, PhD, DSc Senior Associate Director, Boehringer Ingelheim Pharmaceutical Inc. Micardis Product Pipeline Scientific Support Ridgefield, CT, USA Cardiovascular
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Oxidative Stress in PGE2 Treated H9c2 Cardiomyocytes. Rose Picklo
 Oxidative Stress in PGE2 Treated H9c2 Cardiomyocytes Rose Picklo Abstract Heart disease is the leading cause of adult death in the United States. Chronic ischemia resulting from obstruction of coronary
Oxidative Stress in PGE2 Treated H9c2 Cardiomyocytes Rose Picklo Abstract Heart disease is the leading cause of adult death in the United States. Chronic ischemia resulting from obstruction of coronary
E and the heart: Possible role as antioxidant. Acta Vitaminol. Enzymol. 5: 11-22, ) Jolly, S. R., Kane, W. J., Bailie, M. B. et al.
 Jolly, S. R., Kane, W. J., Bailie, M. B. et al.") 1) Ferrari, R., Visoli, O., Guarnieri, C. et al.: Vitamin E and the heart: Possible role as antioxidant. Acta Vitaminol. Enzymol. 5: 11-22, 1983. 2) Jolly, S. R., Kane, W. J., Bailie, M. B. et al.: Canine
1) Ferrari, R., Visoli, O., Guarnieri, C. et al.: Vitamin E and the heart: Possible role as antioxidant. Acta Vitaminol. Enzymol. 5: 11-22, 1983. 2) Jolly, S. R., Kane, W. J., Bailie, M. B. et al.: Canine
Reduction of metastatic and angiogenic potency of malignant cancer by Eupatorium. fortunei via suppression of MMP-9 activity and VEGF production
 Supplementary Information Reduction of metastatic and angiogenic potency of malignant cancer by Eupatorium fortunei via suppression of MMP-9 activity and VEGF production Aeyung Kim, Minju Im, Nam-Hui Yim
Supplementary Information Reduction of metastatic and angiogenic potency of malignant cancer by Eupatorium fortunei via suppression of MMP-9 activity and VEGF production Aeyung Kim, Minju Im, Nam-Hui Yim
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
LncRNA LET function as a tumor suppressor in breast cancer development
 European Review for Medical and Pharmacological Sciences 2018; 22: 6002-6007 LncRNA LET function as a tumor suppressor in breast cancer development C.-X. ZHOU, X. WANG, N. YANG, S.-K. XUE, W.-C. LI, P.-P.
European Review for Medical and Pharmacological Sciences 2018; 22: 6002-6007 LncRNA LET function as a tumor suppressor in breast cancer development C.-X. ZHOU, X. WANG, N. YANG, S.-K. XUE, W.-C. LI, P.-P.
Nature Immunology: doi: /ni Supplementary Figure 1. IC261 inhibits a virus-induced type I interferon response.
 Supplementary Figure 1 IC261 inhibits a virus-induced type I interferon response. (a) HEK293T cells were cultured in 384 wells and transiently transfected with 50 ng of the IFN-β promoter-luc construct
Supplementary Figure 1 IC261 inhibits a virus-induced type I interferon response. (a) HEK293T cells were cultured in 384 wells and transiently transfected with 50 ng of the IFN-β promoter-luc construct
ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1
 ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
Original Article Bufalin attenuates the proliferation of breast cancer MCF-7 cells in vitro and in vivo by inhibiting the PI3K/Akt pathway
 Int J Clin Exp Med 2016;9(6):10297-10303 www.ijcem.com /ISSN:1940-5901/IJCEM0022878 Original Article Bufalin attenuates the proliferation of breast cancer MCF-7 cells in vitro and in vivo by inhibiting
Int J Clin Exp Med 2016;9(6):10297-10303 www.ijcem.com /ISSN:1940-5901/IJCEM0022878 Original Article Bufalin attenuates the proliferation of breast cancer MCF-7 cells in vitro and in vivo by inhibiting
C57BL/6 Mice are More Appropriate. than BALB/C Mice in Inducing Dilated Cardiomyopathy with Short-Term Doxorubicin Treatment
 Original Article C57BL/6 Mice are More Appropriate Acta Cardiol Sin 2012;28:236 240 Heart Failure & Cardiomyopathy C57BL/6 Mice are More Appropriate than BALB/C Mice in Inducing Dilated Cardiomyopathy
Original Article C57BL/6 Mice are More Appropriate Acta Cardiol Sin 2012;28:236 240 Heart Failure & Cardiomyopathy C57BL/6 Mice are More Appropriate than BALB/C Mice in Inducing Dilated Cardiomyopathy
The role of 14,15-dihydroxyeicosatrienoic acid levels in inflammation and its relationship to lipoproteins
 Yang et al. Lipids in Health and Disease 2013, 12:151 RESEARCH Open Access The role of 14,15-dihydroxyeicosatrienoic acid levels in inflammation and its relationship to lipoproteins Tian Yang, Ran Peng,
Yang et al. Lipids in Health and Disease 2013, 12:151 RESEARCH Open Access The role of 14,15-dihydroxyeicosatrienoic acid levels in inflammation and its relationship to lipoproteins Tian Yang, Ran Peng,
An epithelial-to-mesenchymal transition-inducing potential of. granulocyte macrophage colony-stimulating factor in colon. cancer
 An epithelial-to-mesenchymal transition-inducing potential of granulocyte macrophage colony-stimulating factor in colon cancer Yaqiong Chen, Zhi Zhao, Yu Chen, Zhonglin Lv, Xin Ding, Renxi Wang, He Xiao,
An epithelial-to-mesenchymal transition-inducing potential of granulocyte macrophage colony-stimulating factor in colon cancer Yaqiong Chen, Zhi Zhao, Yu Chen, Zhonglin Lv, Xin Ding, Renxi Wang, He Xiao,
Curriculum Vitae. Research Officer in Baker IDI Heart & Diabetes Institute from Aug
 Curriculum Vitae Yi Fu ( 付毅 ) (PhD, MBBS) Baker IDI Heart and Diabetes Institute 75 Commercial Road, Prahran, Melbourne, VIC 3004 Australia Phone: +61 3 8532 1288 Email: yi.fu@bakeridi.edu.au PROFILE Research
Curriculum Vitae Yi Fu ( 付毅 ) (PhD, MBBS) Baker IDI Heart and Diabetes Institute 75 Commercial Road, Prahran, Melbourne, VIC 3004 Australia Phone: +61 3 8532 1288 Email: yi.fu@bakeridi.edu.au PROFILE Research
INTRODUCTION. Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells
 Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells") Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
To determine the effect of over-expression and/or ligand activation of. PPAR / on cell cycle, cell lines were cultured as described above until ~80%
 Supplementary Materials and Methods Cell cycle analysis To determine the effect of over-expression and/or ligand activation of PPAR / on cell cycle, cell lines were cultured as described above until ~80%
Supplementary Materials and Methods Cell cycle analysis To determine the effect of over-expression and/or ligand activation of PPAR / on cell cycle, cell lines were cultured as described above until ~80%
Role of high mobility group box 1 in myocardial ischemia/reperfusion injury and the effect of ethyl pyruvate
 EXPERIMENTAL AND THERAPEUTIC MEDICINE 9: 1537-1541, 2015 Role of high mobility group box 1 in myocardial ischemia/reperfusion injury and the effect of ethyl pyruvate YUNLING LIN, LIANGLONG CHEN, WEIWEI
EXPERIMENTAL AND THERAPEUTIC MEDICINE 9: 1537-1541, 2015 Role of high mobility group box 1 in myocardial ischemia/reperfusion injury and the effect of ethyl pyruvate YUNLING LIN, LIANGLONG CHEN, WEIWEI
Dynamic changes of found in inflammatory zone 1 protein and mrna expression in the lung with experimental pulmonary fibrosis of the rat
 Acta Physiologica Sinica, August 25, 2005, 57 (4): 493-497 http://www.actaps.com.cn 493 FIZZ1 mrna 1 2, * 1 1 1 430022 2 430030 FIZZ1 (found in inflammatory zone 1) (5 mg/kg ) HE Masson FIZZ1 mrna (1)
Acta Physiologica Sinica, August 25, 2005, 57 (4): 493-497 http://www.actaps.com.cn 493 FIZZ1 mrna 1 2, * 1 1 1 430022 2 430030 FIZZ1 (found in inflammatory zone 1) (5 mg/kg ) HE Masson FIZZ1 mrna (1)
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Journal Club Semmler Lorenz
 Beer et al. 2015 - Analysis of the Secretome of Apoptotic Peripheral Blood Mononuclear Cells: Impact of Released Proteins and Exosomes for Tissue Regeneration Journal Club 13.11.2017 1 Introduction to
Beer et al. 2015 - Analysis of the Secretome of Apoptotic Peripheral Blood Mononuclear Cells: Impact of Released Proteins and Exosomes for Tissue Regeneration Journal Club 13.11.2017 1 Introduction to
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Prostaglandin E 2 reduces radiation-induced epithelial apoptosis through a mechanism involving AKT activation and bax translocation
 Research article Prostaglandin E 2 reduces radiation-induced epithelial apoptosis through a mechanism involving AKT activation and bax translocation Teresa G. Tessner, Filipe Muhale, Terrence E. Riehl,
Research article Prostaglandin E 2 reduces radiation-induced epithelial apoptosis through a mechanism involving AKT activation and bax translocation Teresa G. Tessner, Filipe Muhale, Terrence E. Riehl,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
Biol403 MAP kinase signalling
 Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Eicosapentaenoic Acid and Docosahexaenoic Acid: Are They Different?
 Eicosapentaenoic Acid and Docosahexaenoic Acid: Are They Different? Trevor A Mori, Ph.D., Professor, School of Medicine and Pharmacology, Royal Perth Hospital Unit, University of Western Australia, Perth,
Eicosapentaenoic Acid and Docosahexaenoic Acid: Are They Different? Trevor A Mori, Ph.D., Professor, School of Medicine and Pharmacology, Royal Perth Hospital Unit, University of Western Australia, Perth,
Role of Inflammation in Pulmonary Hypertension
 Role of Inflammation in Pulmonary Hypertension K. R. Stenmark University of Colorado Denver, USA Prominent Fibroproliferative Changes are Observed in the Lung Vasculature of Infants With Pulmonary Arterial
Role of Inflammation in Pulmonary Hypertension K. R. Stenmark University of Colorado Denver, USA Prominent Fibroproliferative Changes are Observed in the Lung Vasculature of Infants With Pulmonary Arterial
Supporting Information
 Supporting Information Muraski et al. 10.1073/pnas.0709135105 SI Text Generation of Transgenic Animals. Pim-WT and Pim-DN cdnas were subcloned NheI/SmaI from pegfp-c1 Pim-1 and pegfp-c1 Pim-DN plasmids
Supporting Information Muraski et al. 10.1073/pnas.0709135105 SI Text Generation of Transgenic Animals. Pim-WT and Pim-DN cdnas were subcloned NheI/SmaI from pegfp-c1 Pim-1 and pegfp-c1 Pim-DN plasmids
Supplemental Figure 1. Western blot analysis indicated that MIF was detected in the fractions of
 Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
European Society of Cardiology Congress DONOR AGE NEGATIVELY INFLUENCES THE CYTOPROTECTIVE PARACRINE EFFECTS EXERTED BY HUMAN MESENCHYMAL STEM CELLS
 European Society of Cardiology Congress 28 Aug - 01 Sep 2009, Stockholm - Sweden DONOR AGE NEGATIVELY INFLUENCES THE CYTOPROTECTIVE PARACRINE EFFECTS EXERTED BY HUMAN MESENCHYMAL STEM CELLS Massimiliano
European Society of Cardiology Congress 28 Aug - 01 Sep 2009, Stockholm - Sweden DONOR AGE NEGATIVELY INFLUENCES THE CYTOPROTECTIVE PARACRINE EFFECTS EXERTED BY HUMAN MESENCHYMAL STEM CELLS Massimiliano
CD14 + S100A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer
 CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
Paracrine Mechanisms in Adult Stem Cell Signaling and Therapy
 Paracrine Mechanisms in Adult Stem Cell Signaling and Therapy Massimiliano Gnecchi, Zhiping Zhang, Aiguo Ni, Victor J. Dzau Circulation Research 2008 Nov 21;103(11):1204-19 Introduction(1) After AMI all
Paracrine Mechanisms in Adult Stem Cell Signaling and Therapy Massimiliano Gnecchi, Zhiping Zhang, Aiguo Ni, Victor J. Dzau Circulation Research 2008 Nov 21;103(11):1204-19 Introduction(1) After AMI all