Fates-shifted is an F box protein that targets Bicoid for degradation and regulates developmental fate determination in Drosophila embryos
|
|
|
- Kristopher Williamson
- 5 years ago
- Views:
Transcription
1 ARTICLES Ftes-shifted is n F ox protein tht trgets Bioid for degrdtion nd regultes developmentl fte determintion in Drosophil emryos Juno Liu 1 nd Jun M 1,2,3 Bioid (Bd) is morphogeneti protein tht instruts ptterning long the nterior posterior (A P) xis in Drosophil melnogster emryos. Despite extensive studies, wht ontrols the formtion of norml onentrtion grdient of Bd remins n unresolved nd ontroversil question. Here, we show tht Bd protein degrdtion is medited y the uiquitin-protesome pthwy. We hve identified n F ox protein, enoded y ftes-shifted (fsd), tht hs n importnt role in Bd protein degrdtion y trgeting it for uiquityltion. Emryos from femles lking fsd hve n ltered Bd grdient profile, resulting in shift of the ftemp long the A P xis. Our study is n experimentl demonstrtion tht, ontrry to n lterntive hypothesis, Bd protein degrdtion is required for norml grdient formtion nd developmentl fte determintion. Morphogens re moleules tht form onentrtion grdients in developing emryo or tissue to provide positionl informtion to ells nd instrut their developmentl ftes 1 5. The Drosophil morphogeneti protein Bioid (Bd) forms onentrtion grdient long the nterior posterior (A P) xis of the emryo nd instruts ptterning y tivting its downstrem trget genes in onentrtion-dependent mnner 6 9. An importnt question out morphogens reltes to the moleulr mehnisms tht ontrol the formtion of norml grdient. Diffusion nd degrdtion of morphogen moleules re two physil properties ritil to oth the proess of grdient formtion nd the finl outome of the grdient 1,5,1,11. Although the diffusion onstnt for Bd moleules in emryos remins highly ontroversil 12 14, wht ontrols Bd degrdtion remins virtully unknown t this time. In this report, we investigte the moleulr mehnisms of Bd protein degrdtion. We present evidene tht Bd degrdtion is required for the formtion of norml onentrtion grdient in emryos nd for instruting ells to dopt their norml developmentl ftes. Results Bd degrdtion in Drosophil ells is sensitive to protesome inhiitors We generted Drosophil S2 ells tht stly express hemgglutinin (HA) tgged wild-type Bd protein from the tin promoter. Western lotting nlysis onfirmed the expression of the full-length Bd protein in these ells (Supplementry Informtion, Fig. S1). This HA Bd protein n tivte trnsription of Bd-dependent reporter gene in S2 ells 15, demonstrting tht it is funtionlly tive. To investigte mehnisms nd pthwys of Bd protein degrdtion, we treted the ells stly expressing HA Bd with vrious inhiitors. The protesome inhiitor MG132 inresed the totl mount of Bd (Fig. 1, lne 4 nd Supplementry Informtion, Fig. S1). Neither Z LL H, whih is n MG132 nlogue nd lpin inhiitor, nor hloroquine diphosphte, whih is lysosome inhiitor, hd suh n effet (Fig. 1, lnes 2 nd 3). Two dditionl protesome inhiitors, ltystin nd epoxomiin, whih inhiit protesome tivities through distint mehnisms 16,17, lso inresed the totl mount of Bd (Fig. 1, lnes 5 nd 6). These results suggest tht the protesome-dependent pthwy is importnt for Bd protein degrdtion. Bd degrdtion in Drosophil emryoni extrts revels role for uiquitin To further investigte Bd degrdtion mehnisms, we generted Drosophil emryoni extrts from emryos t 3 h. An HA Bd fusion protein ws effiiently degrded in these extrts (Fig. 1, lnes 1 6), ut suh degrdtion ws signifintly inhiited y MG132 (Fig. 1, lnes 7 12) or epoxomiin (Supplementry Informtion, Fig. S1, d). Bd degrdtion profiles in the presene or sene of MG132 oth fitted well with the first order dey funtion 18 (Fig. 1), with respetive Adjusted R 2 vlue of.999 nd.981. MG132 extends the estimted hlf-life of Bd y 13.1-fold (± 1.8; n = 3 from experiments performed s in Fig. 1, ). These results further support our onlusion tht Bd is degrded in protesome-dependent mnner. 1 Division of Biomedil Informtis, Cininnti Children s Reserh Foundtion, 3333 Burnet Avenue, Cininnti, OH 45229, USA. 2 Division of Developmentl Biology, Cininnti Children s Reserh Foundtion, 3333 Burnet Avenue, Cininnti, OH 45229, USA. 3 Correspondene should e ddressed to J.M. (e mil: jun.m@hm.org) Reeived 3 April 1; epted 17 Novemer 1; pulished online 19 Deemer 1; DOI: 1.138/n nture ell iology VOLUME 13 NUMBER 1 JANUARY 11
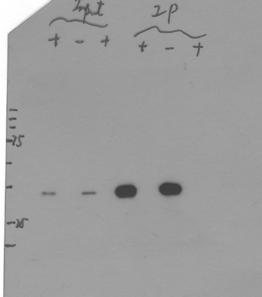
2 ARTICLES Proteins destined for protesome-medited degrdtion re often polyuiquitylted Unlike protein degrdtion in the lysosome, uiquityltion is proess tht requires energy from ATP hydrolysis 22. We therefore investigted whether Bd degrdtion in emryoni extrts is dependent on ATP. Addition of ATP into the emryoni extrts enhned Bd degrdtion, with 61% redution in the estimted hlf-life of Bd (Fig. 2, ). In ddition, depletion of ATP from the extrts through the use of hexokinse nd gluose led to the ttenution of Bd degrdtion (Fig. 2, d), extending the estimted hlf-life of Bd y 2.3-fold. To diretly investigte the role of uiquitin in Bd degrdtion, we used mutnt uiquitin, UM NK, whih hs ll of its seven lysine residues mutted to rginines. This mutnt uiquitin n prevent polyuiquityltion nd inhiit uiquitin-dependent protein degrdtion 23. If Bd degrdtion is indeed dependent on polyuiquityltion, we would expet this mutnt uiquitin to inhiit Bd degrdtion. Our results show tht when UM NK ws dded to the emryoni extrts Bd degrdtion ws ttenuted (Fig. 2e, f), extending its estimted hlf-life y 1.93-fold (s.d. ±.35, n = 3 experiments performed s in Fig. 2e, f). Together, these results suggest tht Bd is degrded in protesome- nd uiquitin-dependent mnner. Bd is uiquitylted To determine whether Bd is uiquitylted, we performed n in vivo uiquityltion ssy in humn emryoni kidney (HEK) 293T ells, whih hve n effiient uiquityltion system 24. To inrese the mount of uiquitinmodified produts, uiquitin-expressing plsmid ws o-trnsfeted into these ells, whih were then treted with MG132 efore hrvest. As shown in Figure 2g, uiquitin-modified Bd protein speies (lne 4) were deteted in Bd- nd MG132-dependent mnner (see other lnes for ontrols). These results support the hypothesis tht Bd is uiquitylted. Identifying n F ox protein involved in Bd degrdtion Uiquityltion is tlysed y three stepwise enzymti retions, with the uiquitin ligse E3 providing sustrte speifiity One group of E3 ligses, the SCF (Skp1 Cul1/Cd53 F-ox protein)-type E3 ligses, hve role in the turnover of mny trnsription ftors 3. The sustrte speifiity in these multi-protein omplexes is onferred y F ox proteins,31,32. We performed doule-strnded RNA interferene (dsrnai) sreen for 38 potentil Drosophil F ox proteins in the S2 ells stly expressing HA Bd. The ddition of dsrna trgeting the ftes-shifted (fsd, CG12765) gene used onsistent inrese in the totl mount of Bd (Supplementry Informtion, Fig. S2, lne 3; see lnes 1 nd 2 for speifiity). Next, we used yloheximide (CHX) to lok trnsltion in the HA Bd-expressing stle ells (Fig. 3) nd found tht fsd dsrna tretment inresed the estimted hlf-life of Bd y 1.5-fold (Fig. 3). In ddition, overexpressing Fsd in S2 ells redued the totl mount of Bd (42%; Fig. 3, lne 2; see lnes 1 nd 3 for ontrols). These results indite tht Fsd hs n importnt role in regulting Bd protein stility in Drosophil ells. Fsd is n F ox protein tht diretly interts with oth Skp1 nd Bd Bsed on the lignment of the 234 sequenes used to rete the F ox profile in the Pfm dtse, F-ox motifs re highly degenerte, lking strit primry onsensus sequene 31. Our lignment of ll potentil F ox domins in Drosophil derived from the Interpro dtse further onfirms the degenerte nture of the primry sequenes of the F ox Perentge Bd remining Time (min): HA Bd Anti-HA Anti- -tin DMSO DMSO Chloroquine Ltystin Epoxomiin HA Bd -tin motif (Supplementry Informtion, Fig. S3). A hllmrk of genuine F ox protein is its ility to intert with Skp1, nother omponent of the SCF omplex,31,32. To determine if Fsd hs this property, we performed o-immunopreipittion experiments using HEK 293T ells. HA Skp1 ws speifilly o-preipitted with Flg Fsd (Fig. 4, lne 6; see other lnes for ontrols), demonstrting tht Fsd nd Skp1 n physilly intert with eh other in ells. To further evlute the role of Skp1 in Bd degrdtion, we mnipulted Skp1 protein levels in the HA Bd-expressing stle ells. The totl mount of Bd in these ells ws deresed when dditionl Skp1 (Flg Skp1) ws expressed (Supplementry Informtion, Fig. S2, lne 3; see lne 1 for ontrol). As expeted, MG132, whih inhiits the downstrem step of protein degrdtion, inresed the totl mount of Bd to similr level regrdless of the sttus of dditionl Skp1 expression (Supplementry Informtion, Fig. S2, lnes 1 4). In ddition, when ells were treted with skpa dsrna, the estimted hlf-life of Bd ws extended y 2.5-fold (Supplementry Z-LL-H MG132 MG132 DMSO MG132 Figure 1 Bd is degrded through the protesome-dependent pthwy. () Effets of protese inhiitors on the totl mount of Bd in Drosophil S2 ells. Cells stly expressing HA Bd were treted with the indited inhiitors nd western lotting ws performed using the HA ntiody to determine the totl mount of HA Bd (top). β tin (ottom) represents the loding ontrol. () Time ourse of Bd degrdtion in emryoni extrts. HA Bd, synthesized in vitro, ws nlysed in emryoni extrts in the presene of MG132 or DMSO (s ontrol). Aliquots were tken from retion tues t the indited times, nd HA Bd ws deteted y western lotting. () Bd degrdtion kinetis. Intensity of the HA Bd nds in were quntified nd lulted s perentge of the intensity of the nd t time zero for eh tretment. The lines represent exponentil fitting to the experimentl dt. Throughout this study, ll kineti experiments shown were performed through side-y-side omprison of western lots (s shown in Fig. 1, ) nd only suh side-y-side results within single pnel n e ompred. Unropped imges of lots re shown in Supplementry Informtion, Fig. S9. nture ell iology VOLUME 13 NUMBER 1 JANUARY 11 23
 ATP + ATP 3 4 UM-NK + UM-NK 3 4 d g Perentge Bd remining 8 6 4 1 Time (min) HA Bd: + + FLAG U: + + + + MG132: + + M r (K) 2 1 1 2 3 4 hexokinse + hexokinse 3 4")
3 ARTICLES Time (min): HA Bd ATP + ATP hexokinse + hexokinse Time (min): HA Bd e Time (min): HA Bd f Perentge Bd remining Perentge Bd remining Time (min) UM-NK + UM-NK Time (min) ATP + ATP 3 4 UM-NK + UM-NK 3 4 d g Perentge Bd remining Time (min) HA Bd: + + FLAG U: MG132: + + M r (K) hexokinse + hexokinse 3 4 U-modified Bd Figure 2 Bd is uiquitylted. ( f) Time ourse of Bd degrdtion in emryoni extrts with or without ATP ddition (, ), ATP depletion (, d) or UM NK ddition (e, f). Hexokinse (together with gluose) ws used to deplete ATP. For eh experiment, liquots were tken from the retion tues t the indited times. (,, e) HA Bd ws deteted y western lotting. (, d, f) Frtion of remining Bd protein (quntified from intensity of the nds in the western lots) plotted ginst retion time. In eh se the lines represent exponentil fitting to the experimentl dt. (g) Bd uiquityltion deteted in Informtion, Fig. S2, d). These results suggest tht oth Fsd nd Skp1 prtiipte in the Bd degrdtion proess, presumly working together in novel Fsd-ontining SCF (SCF Fsd ) E3 ligse. F-ox proteins trget their sustrtes for degrdtion y reruiting them, through diret protein protein intertions, to the SCF E3 ligse omplexes for uiquityltion,31,32. To determine whether Bd is sustrte of the proposed SCF Fsd E3 ligse, we performed nother o-immunopreipittion experiment. Our results indite tht HA Bd ws speifilly o-preipitted y Flg Fsd (Fig. 4, lne 6; see other lnes for ontrols), demonstrting tht Fsd n physilly intert with Bd. Therefore Fsd seems to e on fide F ox protein tht n diretly intert with oth omponent (Skp1) within the proposed SCF Fsd omplex nd its sustrte Bd. Emryos lking Fsd exhiit posterior shifts in morphologil nd moleulr mrkers Sequene nlysis of n fsd DNA lone (NM_17.3) predited protein of 312 mino ids, with two potentil F ox domins (Fig. 5). Fsd does not seem to ontin ny other onserved domins. fsd trnsripts re uniformly distriuted in oth prelstoderm nd gstrulted emryos (Supplementry Informtion, Fig. S4, ), suggesting oth mternl nd zygoti expression. We otined pulilly ville Drosophil line with ells. HEK 293T ells were trnsfeted with plsmids expressing HA Bd nd were treted with MG132 s indited. HEK 293T ells were o-trnsfeted with plsmids enoding Flg-Uiquitin to inrese uiquitylted produts. Whole-ell extrts were immunopreipitted using n nti-ha ntiody nd nlysed y western lotting using the nti-uiquitin ntiody. Uiquitinmodified Bd produts re indited, nd moleulr weight stndrds re shown on the left. Unropped imges of lots re shown in Supplementry Informtion, Fig. S9. P element insertion tht disrupts the fsd gene, referred to s fsd KG2393. We onfirmed tht the insertion is 15 p downstrem of ATG, whih effetively disrupts the entire open reding frme (Fig. 5). The fsd KG2393 flies re homozygous vile, ut exhiit temperture-dependent hthing defet. Although the hthing rtes of emryos from wild-type nd fsd KG2393 flies were omprle t C (91.3% versus 81.7%, Supplementry Informtion, Fig. S4), emryos from mutnt flies hd signifintly redued hthing rte t 18 C (48.6%, ompred with 92% for wild-type). At 29 C, emryos from fsd KG2393 femles (referred to s fsd emryos from now on) hd hthing rte of %. Cutile exmintion deteted mutnt emryos with vrile A P ptterning defets when inuted t 29 C, inluding missing/fused dentile nds (Supplementry Informtion, Fig. S5 d). Whole-mount in situ hyridiztion nlysis lso deteted mutnt emryos with norml even-skipped (eve) expression ptterns with missing or fused stripes (Supplementry Informtion, Fig. S5f, g). These results suggest tht the hthing defet oserved t 29 C is onsequene of A P ptterning defets used y the fsd muttion. The hthing defet t 29 C is stritly ssoited with the mternl mutnt genotype (dt not shown) nd, furthermore, mutnt flies t non-optiml tempertures (18 C) do not exhiit signifint redution in survivl rtes from lrve to pupe or from pupe to dults (Supplementry Informtion, Fig. S4). 24 nture ell iology VOLUME 13 NUMBER 1 JANUARY 11
4 ARTICLES Time (h): Control fsd dsrna Perentge Bd remining Anti-HA Anti- -tin Control fsd dsrna Time (h) Flg Fsd: HA Skp1: Anti-HA Anti-Flg Flg Fsd: HA Bd: Anti-HA Anti-Flg Input + + Input Anti-Flg IP Anti-Flg IP HA Skp1 Flg Fsd HA Bd Flg Fsd Plsmid: Anti-HA Reltive Bd mount: Control vetor pa-fsd pa-cg1242 HA Bd Figure 4 Fsd interts with oth Skp1 nd Bd. () A o-immunopreipittion experiment deteting the intertion etween Fsd nd Skp1. HEK 293T ells were o-trnsfeted with the indited omintions of plsmids expressing Flg Fsd nd HA Skp1. Anti-Flg ntiody ws used to immunopreipitte (IP) Flg Fsd from whole-ell extrts, followed y western lotting nlyses using nti-ha (top) nd nti-flg (ottom) ntiodies to detet HA Skp1 nd Flg Fsd, respetively. () Experiments were performed s in, exept with the use of plsmid expressing HA Bd (in ple of HA Skp1) to detet the intertion etween Bd nd Fsd. Unropped imges of lots re shown in Supplementry Informtion, Fig. S9. Figure 3 Fsd hs role in Bd protein degrdtion. () CHX ssy using ells stly expressing HA Bd. Cells were treted with CHX, nd hrvested t indited times to detet the totl mount of Bd in ells y western lotting using nti-ha (left). Western lotting of β tin (right) ws used s loding ontrol. Top, ontrol ells without dsrna tretment; ottom ells inuted with fsd dsrna efore CHX. () Frtion of remining Bd protein in ontrol ells nd ells inuted with fsd dsrna (quntified from intensity of nds in nd normlized to β tin intensities) plotted ginst time fter CHX ddition. The lines represent exponentil fitting to the experimentl dt. () Drosophil S2 ells were trnsiently trnsfeted with plsmid enoding HA Bd. Cells were o-trnsfeted with ontrol vetor (pa5.1; vetor used for protein expression), plsmid enoding Fsd (pa-fsd), or ontrol F-ox protein (pa-cg1242). For eh lne, the loding mount ws djusted ording to the tivity of gltosidse expressed from lz ontrol plsmid trnsfeted t the sme time s the indited plsmids (to mesure trnsfetion effiieny). Reltive intensity of the nds is indited t the ottom. Unropped imges of lots re shown in Supplementry Informtion, Fig. S9. These results suggest tht the primry funtion of fsd is onferred mternlly to emryos. Ating s morphogen instruting A P ptterning, the mount of Bd diretly ontrols the ftemp of the entire emryo long the A P xis If Fsd hs role in promoting Bd protein degrdtion in emryos s oserved in our in vitro ssys, fsd emryos my exhiit posterior shift of the ftemp refletive of n ltered mount of Bd. To diretly evlute this possiility, we mesured the position of the ephli furrow, morphologil mrker long the A P xis nd strong inditor of Bd protein levels in emryos 13,34,36. For experiments desried from now on, ll emryos were olleted t C (unless stted otherwise) to void ny temperture influene on Bd grdient formtion nd emryoni ptterning. Figure 5, indites tht the ephli furrow position (in frtionl emryo length, ) ws shifted from.343 ±.5 (n = 1) in wild-type emryos to.2 ±.12 (n = 9) in fsd emryos (P = , Student s t-test). To evlute the effet of the fsd gene muttion on emryoni ptterning t the moleulr level, we mesured the expression pttern of eve using quntittive fluoresene in situ hyridiztion (FISH). To ensure diret omprison, experiments nd dt proessing for wildtype nd mutnt emryos were performed side-y-side. Figure 5d demonstrtes the men expression profiles of eve in wild-type nd mutnt emryos, reveling posterior shift for ll eve stripes in fsd emryos (see Supplementry Informtion Fig. S6, for rw dt). For exmple, the first eve stripe is shifted from.331 ±.14 (n = 7) in wild-type emryos to.353 ±.9 (n = 8) in fsd emryos (P = , Fig. 5d). To tre the origin of the oserved ftemp shift in mutnt emryos, we mesured expression ptterns of the gp genes hunhk (h) nd knirps (kni), oth of whih re diret trgets of Bd 8, Agin, we onduted side-y-side experiments in wild-type nd mutnt emryos for eh gene in our quntittive FISH experiments. As shown in Figure 5e, f, the expression oundries of oth gp genes exhiited signifint posterior shift in fsd emryos (see Supplementry Informtion, Fig. 6 f for rw dt). Speifilly, the h expression oundry hd posterior shift of pproximtely 3% of the emryo length, from of.4 ±.8 (n = 8) in wild-type emryos to.479 ±.18 (n = 7) in fsd emryos (P = , Fig. 5e). The kni expression oundry ws shifted towrd the posterior y pproximtely 2% of the emryo length, from.671 ±.1 (n = 9) in wild-type emryos to.689 ±.17 (n = 8) in fsd emryos (P = , Fig. 5f). These results demonstrte tht mternl loss of fsd funtion results in posterior shift of oth morphologil nd moleulr mrkers long the A P xis. fsd nd d intert genetilly The oserved ftemp shift in fsd emryos is supportive of diret role of Fsd in trgeting Bd for degrdtion in emryos. To further test this hypothesis, we onduted geneti intertion nlysis in emryos from d E1 /+ femles tht re either wild-type or mutnt for fsd. Our nture ell iology VOLUME 13 NUMBER 1 JANUARY 11
 Shemti representtion of the fsd gene (not to sle).")

5 ARTICLES CG4663 KG2393 F-ox F-ox fsd (CG12765) CG467 e Reltive h intensity WT fsd WT (n = 8) fsd (n = 7) d Reltive kni intensity Reltive eve intensity f WT (n = 9) fsd (n = 8) WT (n = 7) fsd (n = 8) Figure 5 Posterior shift of ftemp long the A P xis in fsd emryos. () Shemti representtion of the fsd gene (not to sle). The rrows under the genes show the diretion of trnsription nd the oxes represent nnotted exons. The lue setions of the oxes represent the nnotted fsd oding sequenes, wheres the unfilled setions represent untrnslted regions. The position of the KG2393 insertion nd the two F ox domins re mrked. Prts of the two nnotted neighouring genes re lso shown. (, ) Midsgittl imges of living emryos from w 1118 () nd fsd KG2393 () flies results show tht the nterior ftemp shift used y mternl d dosge redution is resued prtilly y eliminting fsd mternlly (Supplementry Informtion, Fig. S7 ). These results demonstrte n intertion, genetilly, etween fsd nd d in regulting developmentl fte speifition long the A P xis. fsd emryos hve n ltered Bd grdient profile To diretly determine whether Fsd hs role in Bd grdient formtion, we onduted quntittive immunostining nlysis using nti- Bd ntiodies on whole-mount wild-type nd mutnt emryos 45. For this nlysis we used rw, unproessed Bd fluoresene intensity dt within liner rnge, with kground intensities diretly mesured under identil experimentl onditions. Figure 6, show Bd intensity (B) dt extrted from individul wild-type nd fsd emryos tht were stined, imged nd nlysed side-y-side. Also shown in these figures re the mesured kground intensities. To evlute the shpe of the Bd grdient profiles, we lulted nd ompred the length onstnt (λ) vlues of either the verge Bd profiles from wild-type nd mutnt emryos or vlues from individul emryos. In simple diffusion model 1,5,46, λ of n exponentil profile is funtion of the effetive diffusion oeffiient (D) nd the effetive degrdtion rte of the morphogen moleules (ω): λ 2 = D/ω. As Bd protein itself hs not een ltered in fsd emryos, D is expeted to remin unffeted y fsd muttion, whih llows us to estimte the reltive ω vlues in wild-type nd mutnt emryos sed on the mesured λ vlues. Figure 6 shows ln (B/B mx ) plot s funtion of A P position. Both B nd B mx re from verge Bd profiles of wild-type nd fsd t C, imged under hloron oil. Arrowheds represent the ephli furrow positions. Sle r, μm. (d f) Averge normlized fluoresene intensities from FISH on whole-mount emryos, deteting the trnsripts of eve (d), h (e) nd kni (f) in wild-type (lue) nd fsd (red) emryos. See Supplementry Informtion, Fig. S6 for rw dt, extrted from individul emryos. The eve expression stripes 1 to 7 hve the following posterior shifts in their respetive posterior oundry positions in fsd emryos: 2.2%, 1.9%, 2.9%, 2.8%, 3.4%, 1.5% nd 1.1% of the emryo length. emryos nd re kground sutrted (s neessry) without ny further djustments. The ln trnsformtion onverts n exponentil funtion to liner funtion 42. Here, we perform liner fit within the rnge of.1 to.5, where the Bd intensity dt re lest sensitive to experimentl nd kground mesurement errors 45,46. We lso exlude dt from the most nterior prt of the emryo where Bd profiles re known to devite from n exponentil funtion 1,42,45. The liner fitting (Fig. 6) hd smller slope in fsd emryos thn in wildtype emryos, suggesting lrger λ vlue in fsd emryos. To further ompre Bd profiles, we lulted λ vlues for individul wild-type nd mutnt emryos (Fig. 6d). The lulted λ vlues inresed from ± μm (n = ) in wild-type emryos to ± 14.3 μm (n = 28) in fsd emryos (P = , Fig. 6d). These results (nd Fig. 5 nd Supplementry Informtion, Fig. S6) provide in vivo evidene tht Fsd hs role in regulting Bd protein stility nd determining developmentl ftes long the A P xis. Disussion Advning the morphogen onept requires n understnding of not only how ells respond to the positionl informtion enoded y the morphogen grdients, ut lso how suh grdients re estlished. Aording to widely epted view 1, the formtion of onentrtion grdient requires lolized soure of morphogen prodution, oupled with diffusion nd degrdtion of the morphogen moleules. However, reent studies question whether the formtion of norml onentrtion grdient of Bd in emryos requires either diffusion or dey of Bd moleules 47,48. Before our urrent work, it ws not known how Bd moleules re degrded, 26 nture ell iology VOLUME 13 NUMBER 1 JANUARY 11
6 ARTICLES WT (n = ) fsd (n = 28) 6 B 4 3 B In (B/B mx ) WT (n = ) fsd (n = 28).8 1. d λ (µm) P = WT fsd e B Figure 6 Bd grdient profiles in wild-type nd fsd emryos. (, ) Wholemount wild-type () nd fsd () emryos were immunostined with nti-bd ntiodies, imged y immunofluoresene mirosopy, nd the fluoresene intensity of Bd ws quntified long the A P xis. Eh olour represents dt from n individul emryo. The line t the ottom represents kground intensities. Dt re mens ± s.d. B, Bd intensity. () ln (B/B mx ) plotted ginst for verge Bd profiles from wild-type () nd fsd () emryos. Both B nd B mx re kground-sutrted s neessry, without ny further djustments. The solid lines represent liner fits for Bd profiles from wildtype emryos (y = 7.2x +.61, Adjusted R 2 =.997) nd fsd emryos mking it diffiult to evlute the role of Bd protein degrdtion in grdient formtion in vivo. We report for the first time tht Bd is degrded through the uiquitin-protesome pthwy. The identifition of novel F ox gene, fsd, mkes it possile to experimentlly pertur Bd degrdtion in vivo nd investigte its role on Bd grdient formtion nd developmentl fte determintion. The Bd grdient profile in fsd emryos hs lrger λ vlue thn in wildtype emryos, demonstrting tht, ontrry to reent proposl 47, Bd degrdtion is required for norml grdient formtion. Our oserved differene in λ vlues orresponds to, in simple diffusion model, n pproximtely 48% inrese in Bd stility in fsd emryos, differene suffiient to use signifint shifts in morphologil nd moleulr mrkers (Fig. 5). We note tht, unlike λ, B mx in wild-type nd fsd emryos did not exhiit signifint differene (44.32 ± 9.1 nd ± 9.19, respetively, P =.63, Student s t-test). Like λ, the stedy-stte mount of morphogen moleules t the soure is lso funtion of D nd ω in simple diffusion model, ut this mount is dditionlly dependent on the morphogen prodution rte J 1,5,46. As the Bd prodution site (tht is, d mrna lotion) is not restrited to single point s ssumed in the idelized simple diffusion model 1, λ should represent more relile prmeter (thn, for exmple, B mx ) in evluting the reltive effetive degrdtion rtes of Bd. (y = 6.31x +.45, Adjusted R 2 =.998). (d) λ vlues lulted from Bd intensity profiles of individul wild-type nd fsd emryos from nd. Individul dt points (unfilled irles) nd mens ± s.d. (oxes with error rs) re indited, with P vlue from Student s t-test indited t the top. (e) The verge intensity of Bd long the A P xis ws quntified for wildtype nd fsd emryos shown in nd. The positions t whih the Bd intensities (without djustments) from the wild-type nd fsd emryos ross onentrtion threshold re shown. The mesured verge h oundry positions in these emryos re mrked with solid rrowheds. Dt from wildtype emryos re shown in lue, nd fsd emryos in red. The h expression oundry in fsd emryos exhiits shift towrd the posterior y pproximtely 3% of the emryo length. This is onsistent with the shift of the positionl informtion enoded y the Bd grdient in fsd emryos (Fig. 6e). These results, together with those presented elsewhere 45,46,49, demonstrte tht the h expression oundry is primrily determined y the positionl informtion provided y the Bd grdient. Although gene regultory networks represent importnt mehnisms for mintining or refining gene expression ptterns 11,46,,51, our results suggest tht erly deisions in A P ptterning, suh s h expression, re ontrolled primrily y the positionl informtion enoded y the Bd grdient. The h oundry shift of pproximtely 3% of the emryo length in fsd emryos mthes the oserved ephli furrow shift (pproximtely 3% of the emryo length), inditing tht erly deisions in gene expression re fithfully pssed down to the morphologil level. We note tht the h oundry shift in fsd emryos, though seemingly modest, is signifint when ompred with the oserved shift (pproximtely 7% of the emryo length) used y the douling of the mternl d gene dose to four opies 42. A shift of pproximtely 3% of the emryo length, when expressed in solute length, is omprle to the differene etween our mesured λ vlues in wild-type nd mutnt emryos. We lso note tht the ftemp shift in nture ell iology VOLUME 13 NUMBER 1 JANUARY 11 27
7 ARTICLES fsd emryos ws deteted t C. The hthing defet of fsd emryos t 29 C represents mnifesttion of more severe A P ptterning defets (Supplementry Informtion, Fig. S5), proly refletive of the omplex effets of temperture on oth Bd grdient formtion (tht is, degrdtion nd diffusion) nd other relevnt proesses. Finlly, we note tht the fsd KG2393 llele used throughout our work is proly null llele euse it is indistinguishle from n llele, fsd 5 7, tht hs the entire fsd oding sequene deleted (Supplementry Informtion, Fig. S8). SCF E3 ligses regulte mny iologil proesses, suh s DNA replition, trnsription, signl trnsdution, ell prolifertion nd deth, nd tissue growth nd ptterning,26,31,32. Our study identifies n F ox protein (enoded y fsd) tht hs role in Bd grdient formtion nd developmentl fte speifition. Our study thus expnds the list of iologil proesses tht SCF E3 ligses regulte. Fsd diretly interts with oth its sustrte Bd nd n SCF omponent Skp1, suggesting tht Bd is trgeted for degrdtion y n Fsd-ontining SCF omplex, SCF Fsd, tht funtions s n E3 uiquitin ligse. We urrently do not know whether Fsd my hve other sustrtes. Identifition of dditionl sustrtes of Fsd, if ny, will provide more omprehensive understnding of the iologil funtions of this F ox protein. How norml Bd onentrtion grdient is formed in erly emryos remins n open question urrently under intense dete 1,13,14,47 49,52,53. One prtiulr spet of the dete fouses on the roles of the nuleus. Although some studies suggest tht nulei hve importnt roles in Bd degrdtion 7 nd proper grdient formtion 13,54, others suggest tht the shping of norml Bd grdient is independent of nulei 55. Fsd is rodly distriuted in ells with primry loliztion to the ytoplsm (Supplementry Informtion, Figs. S4d f), onsistent with the notion tht ytoplsm hs role in Bd degrdtion. However, our reent studies hve shown tht dcbp, Bd-interting trnsription oftor 56,57, ffets Bd grdient profile in emryos 46, suggesting tht nulei lso ontriute to Bd degrdtion nd grdient formtion. Although effiient Bd degrdtion requires Fsd oth in vitro nd in vivo, we onsider it unlikely tht SCF Fsd is the only E3 ligse for Bd. Identifying dditionl E3 ligses, prtiulrly those with primry loliztion to the nuleus, should help settle the ongoing ontroversy regrding the role of nulei in Bd grdient formtion. METHODS Methods nd ny ssoited referenes re ville in the online version of the pper t Note: Supplementry Informtion is ville on the Nture Cell Biology wesite Aknowledgements We thnk memers of our groups t CCHMC, in prtiulr F. He, D. Cheung, W. Dui, nd J. Deng, for disussion nd ssistne, nd we thnk Xinhu Lin s l for some of the primers used in our dsrnai sreening. This work ws supported in prt y grnts from NIH nd NSF (to J.M.). Author Contriutions J.L. nd J.M. oneived nd designed the study. J.L. performed ll experiments nd nlysis. J.L. nd J.M. interpreted the dt, J.L. generted ll figures nd J.L. nd J.M. wrote the pper. COMPETING FINANCIAL INTERESTS The uthors delre tht they hve no ompeting finnil interests. Pulished online t Reprints nd permissions informtion is ville online t reprintsndpermissions/ 1. Wolpert, L. Positionl informtion nd the sptil pttern of ellulr differentition. J. Theor. Biol, 1 47 (1969). 2. Kerszerg, M. & Wolpert, L. Speifying positionl informtion in the emryo: looking eyond morphogens. Cell 13, 5 9 (7). 3. Lnder, A. D. Morpheus unound: reimgining the morphogen grdient. Cell 128, (7). 4. Mrtinez Aris, A. & Hywrd, P. Filtering trnsriptionl noise during development: onepts nd mehnisms. Nt. Rev. Genet. 7, (6). 5. Wrtlik, O., Kihev, A. & Gonzlez-Gitn, M. Morphogen grdient formtion. Cold Spring Hr. Perspet. Biol. 1, 15 (9). 6. Ephrussi, A. & St. Johnston, D. Seeing is elieving. The ioid morphogen grdient mtures. Cell 116, (4). 7. Driever, W. & Nüsslein-Volhrd, C. A grdient of ioid protein in Drosophil emryos. Cell 54, (1988). 8. Struhl, G., Struhl, K. & Mdonld, P. The grdient morphogen ioid is onentrtion-dependent trnsriptionl tivtor. Cell 57, (1989). 9. Driever, W., Thom, G. & Nüsslein-Volhrd, C. Determintion of sptil domins of zygoti gene expression in the Drosophil emryo y the ffinity of inding site for the ioid morphogen. Nture 34, (1989). 1. Deng, J., Wng, W., Lu, L. J. & M, J. A two-dimensionl simultion model of the Bioid grdient in Drosophil. PLoS ONE 5, e12 (1). 11. Bergmnn, S. et l. Pre stedy stte deoding of the Bioid morphogen grdient. PLoS iology 5, e46 (7). 12. Gregor, T., Bilek, W., vn Stevenink, R. R., Tnk, D. W. & Wieshus, E. F. Diffusion nd sling during erly emryoni pttern formtion. Pro. Ntl Ad. Si. USA 12, (5). 13. Gregor, T., Wieshus, E. F., MGregor, A. P., Bilek, W. & Tnk, D. W. Stility nd nuler dynmis of the ioid morphogen grdient. Cell 13, (7). 14. Porher, A. et l. The time to mesure positionl informtion: mternl hunhk is required for the synhrony of the Bioid trnsriptionl response t the onset of zygoti trnsription. Development 1, (1). 15. Zho, C. et l. The tivity of the Drosophil morphogeneti protein Bioid is inhiited y domin loted outside its homeodomin. Development 129, (2). 16. Fenteny, G. et l. Inhiition of protesome tivities nd suunit-speifi minoterminl threonine modifition y ltystin. Siene 268, (1995). 17. Meng, L. et l. Epoxomiin, potent nd seletive protesome inhiitor, exhiits in vivo ntiinflmmtory tivity. Pro. Ntl Ad. Si. USA 96, (1999). 18. Belle, A., Tny, A., Bitink, L., Shmir, R. & O She, E. K. Quntifition of protein hlf-lives in the udding yest proteome. Pro. Ntl Ad. Si. USA 13, (6). 19. Pikrt, C. M. Mehnisms underlying uiquitintion. Annu. Rev. Biohem. 7, (1).. Pikrt, C. M. & Eddins, M. J. Uiquitin: strutures, funtions, mehnisms. Biohim. Biophys. At 1695, (4). 21. Herrmnn, J., Lermn, L. O. & Lermn, A. Uiquitin nd uiquitin-like proteins in protein regultion. Cir. Res., (7). 22. Ciehnover, A. Proteolysis: from the lysosome to uiquitin nd the protesome. Nt. Rev. Mol. Cell Biol. 6, (5). 23. Hershko, A., Gnoth, D., Pehrson, J., Plzzo, R. E. & Cohen, L. H. Methylted uiquitin inhiits ylin degrdtion in lm emryo extrts. J. Biol. Chem. 266, (1991). 24. Hsu, T., MRkn, D., Vinent, T. S. & Gert de Couet, H. Drosophil Pin1 prolyl isomerse Dodo is MAP kinse signl responder during oogenesis. Nt. Cell Biol. 3, (1).. Deshies, R. J. SCF nd Cullin/Ring H2-sed uiquitin ligses. Annu. Rev. Cell Dev. Biol. 15, (1999). 26. Conwy, R. C., Brower, C. S. & Conwy, J. W. Emerging roles of uiquitin in trnsription regultion. Siene 296, (2). 27. Murtni, M., Kung, C., Shokt, K. M. & Tnsey, W. P. The F ox protein Dsg1/Mdm3 is trnsriptionl otivtor tht stimultes Gl4 turnover nd otrnsriptionl mrna proessing. Cell 1, (5). 28. von der Lehr, N. et l. The F ox protein Skp2 prtiiptes in My proteosoml degrdtion nd ts s oftor for My regulted trnsription. Mol. Cell 11, (3). 29. Kornitzer, D., Roy, B., Kulk, R. G. & Fink, G. R. Regulted degrdtion of the trnsription ftor Gn4. EMBO J. 13, (1994). 3. Jing, J. & Struhl, G. Regultion of the Hedgehog nd Wingless signlling pthwys y the F ox/wd4-repet protein Slim. Nture 391, (1998). 31. Kipreos, E. T. & Pgno, M. The F ox protein fmily. Genome Biol 1, reviews32 reviews32.7 (). 32. Ho, M. S., Tsi, P. I. & Chien, C. T. F ox proteins: the key to protein degrdtion. J. Biomed. Si. 13, (6). 33. Frohnhöfer, H. G. & Nüsslein-Volhrd, C. Orgniztion of nterior pttern in the Drosophil emryo y the mternl gene ioid. Nture 324, 1 1 (1986). 34. Berleth, T. et l. The role of loliztion of ioid RNA in orgnizing the nterior pttern of the Drosophil emryo. EMBO J. 7, (1988). 35. Driever, W., Siegel, V. & Nüsslein-Volhrd, C. Autonomous determintion of nterior strutures in the erly Drosophil emryo y the ioid morphogen. Development 19, (199). 36. Driever, W. & Nüsslein-Volhrd, C. The ioid protein determines position in the Drosophil emryo in onentrtion dependent mnner. Cell 54, (1988). 28 nture ell iology VOLUME 13 NUMBER 1 JANUARY 11
8 ARTICLES. Smll, S., Krut, R., Hoey, T., Wrrior, R. & Levine, M. Trnsriptionl regultion of pir-rule stripe in Drosophil. Genes & Dev. 5, (1991). 38. RiverA Pomr, R. & Jkle, H. From grdients to stripes in Drosophil emryogenesis: filling in the gps. Trends Genet. 12, (1996). 39. Driever, W. & Nüsslein-Volhrd, C. Bioid protein is positive regultor of hunhk trnsription in the erly Drosophil emryo. Nture 3, (1989). 4. Perkins, T. J., Jeger, J., Reinitz, J. & Glss, L. Reverse engineering the gp gene network of Drosophil melnogster. PLoS Comput. Biol. 2, e51 (6). 41. Sheffer, V., Jnody, F., Loss, C., Despln, C. & Wimmer, E. A. Bioid funtions without its TATA-inding protein-ssoited ftor intertion domins. Pro. Ntl Ad. Si. USA 96, (1999). 42. Houhmndzdeh, B., Wieshus, E. & Leiler, S. Estlishment of developmentl preision nd proportions in the erly Drosophil emryo. Nture 415, (2). 43. Cruk, O. & Dosttni, N. Bioid determines shrp nd preise trget gene expression in the Drosophil emryo. Curr. Biol. 15, (5). 44. River-Pomr, R., Lu, X., Tuert, H., Perrimon, N. & Jkle, H. Ativtion of posterior gp gene expression in the Drosophil lstoderm. Nture 6, 3 6 (1995). 45. He, F. et l. Proing intrinsi properties of roust morphogen grdient in Drosophil. Dev. Cell 15, (8). 46. He, F. et l. Shping morphogen grdient for positionl preision. Biophys. J. 99, (1). 47. Coppey, M., Berezhkovskii, A. M., Kim, Y., Boettiger, A. N. & Shvrtsmn, S. Y. Modeling the ioid grdient: diffusion nd reversile nuler trpping of stle protein. Dev. Biol. 312, (7). 48. Spirov, A. et l. Formtion of the ioid morphogen grdient: n mrna grdient dittes the protein grdient. Development 136, (9). 49. He, F. et l. Distne mesurements vi the morphogen grdient of Bioid in Drosophil emryos. BMC Dev. Biol. 1, 8 (1).. Jeger, J. et l. Dynmi ontrol of positionl informtion in the erly Drosophil emryo. Nture 43, (4). 51. Mnu et l. Cnliztion of gene expression in the Drosophil lstoderm y gp gene ross regultion. PLoS iology 7, e49 (9). 52. Luhett, E. M., Vinent, M. E. & Ismgilov, R. F. A preise Bioid grdient is nonessentil during yles for preise ptterning in the Drosophil lstoderm. PLoS One 3, e3651 (8). 53. Heht, I., Rppel, W. J. & Levine, H. Determining the sle of the Bioid morphogen grdient. Pro. Ntl Ad. Si. USA 16, (9). 54. Gregor, T., MGregor, A. P. & Wieshus, E. F. Shpe nd funtion of the Bioid morphogen grdient in diptern speies with different sized emryos. Dev. Biol. 316, (8). 55. Grimm, O. & Wieshus, E. The Bioid grdient is shped independently of nulei. Development 1, (1). 56. Fu, D. & M, J. Interply etween positive nd negtive tivities tht influene the role of Bioid in trnsription. Nulei ids reserh 33, (5). 57. Fu, D., Wen, Y. & M, J. The o-tivtor CREB-inding protein prtiiptes in enhner-dependent tivities of Bioid. J. Biol. Chem. 279, (4). nture ell iology VOLUME 13 NUMBER 1 JANUARY 11 29
9 METHODS DOI: 1.138/n2141 Methods Fly lines, plsmids nd ells. The fsd KG2393 flies were otined from the Bloomington Drosophil Stok Center (stok numer 12983). The following vetors were used in this study. pgem3 (Promeg) ws used s templte for in vitro trnsription/trnsltion. The pa5.1/v5-his C vetor (Invitrogen) ontining the Drosophil tin 5C promoter ws used to onstitutively express proteins in Drosophil S2 ells. The pdna3 vetor (Invitrogen) ontining the humn Cytomeglovirus immedite-erly (CMV) promoter ws used for over-expressing proteins in HEK 293T ells. All plsmids used in this study were generted y stndrd loning methods. Drosophil S2 nd HEK 293T ells were ultured in Shneider s Drosophil medium (Invitrogen) nd Duleo s modifition of Egle s medium (Cellgro), respetively, oth ontining 1% (v/v) fetl lf serum (Invitrogen) nd 1 ntiioti-ntimyoti (Invitrogen). Trnsfetion in oth ells ws performed using the FuGENE HD trnsfetion regent (Rohe) ording to the mnufturer s instrutions. Stle Drosophil S2 ells nd protein stility ssy. Drosophil S2 ells tht stly express HA Bd were generted s desried previously 58. The following is list of inhiitors (soures; finl onentrtions) for treting ells: hloroquine diphosphte (Sigm; μm), Z LL H (Peptides Interntionl; μm), MG132 (Boston Biohem; μm), ltystin (Cliohem; 1 μm), epoxomiin (Cliohem; 3 nm), nd CHX (Sigm; μg ml 1 ). For trnsient ssys, lz reporter ws used to normlize trnsfetion effiieny 15. Protein degrdtion ssy in Drosophil emryoni extrts. Drosophil emryoni extrts (extrted from w1118 emryos t 3 h) were prepred s desried previously 59, diluted to finl onentrtion of 1 mg ml 1 nd stored t 8 C efore use. HA Bd protein used in protein degrdtion ssys ws trnslted in vitro using the T N T quik oupled trnsription/trnsltion system (Promeg). Degrdtion retions were performed t 3 C, with or without the following (t given finl onentrtions): MG132 ( μm), epoxomiin ( μm), Mg ATP (Boston Biohem; 5 mm), UM NK (Boston Biohem,.67 μg μl 1 ), gluose (Sigm; 1 mm) nd hexokinse (Sigm;.3 U μl 1 ). We hve notied tht the protein degrdtion tivities of the stored emryoni extrts vried somewht. Thus, ll kineti experiments with speifi tretments inluded side-y-side (t ll steps) no-tretment ontrol. Only omprisons etween side-y-side ssys re meningful nd, thus, no omprisons were mde (or should e mde) etween experiments. Despite this vriility, the effets of different tretments on Bd stility re highly reproduile etween independent experiments. Western lot nd protein quntifitions. For deteting totl protein mounts in ells, ells were diretly oiled in 1 SDS PAGE (sodium dodeyl sulfte polyrylmide gel eletrophoresis) loding uffer ( mm Tris-HCl t ph 6.8, mm dithiothreitol, 2% SDS, 1% glyerol nd.1% romophenol lue) for 8 min, with rief vortexing every 2 min. Proteins were seprted y SDS PAGE nd trnsferred to Immun-Blot PVDF (polyvinylidene fluoride) memrne (Bio-Rd) for western lotting using pproprite primry ntiodies nd HRP (horse rdish peroxidse)- onjugted seondry ntiodies. Western lotting signls were visulized y ECL (enhned hemiluminsesene) plus Western lotting detetion regents (GE Helthre). HA tgged nd Flg-tgged proteins were deteted y nti-ha (Covne) nd nti-flg (Sigm) primry ntiodies, respetively; nti β-tin ntiody (Am) ws used to detet β tin s loding ontrol whenever neessry. nti-ha (1:1,), nti-flg (1:1,) nd nti-β-tin (1:2,). The protein nds deteted in western lotting were quntified s follows. X ry films were snned with the HP Snjet 47 Photosmrt snner, followed y nlysis using the Sion imge softwre. A retngulr ox of the sme size ws hosen to over the individul nds of interest, with men intensities determined y the softwre. Bkground intensity ws otined y pling the sme-sized retngulr ox in lnk lne t the sme position s in experimentl lnes. To estimte the hlf-life of Bd in the emryoni extrts, we plotted the Bd remining mount (frtion of Bd intensity t time zero) ginst the retion time. The dt were then fitted to the exponentil dey eqution (y = e x ) using the Mtl softwre. The resulting ln2/ vlues represent the estimted hlf-life of Bd. In vivo uiquityltion nd o-immunopreipittion ssys. For uiquityltion ssy, the HA Bd-expressing plsmid ws trnsiently trnsfeted into HEK 293T ells, long with seond plsmid expressing Flg-tgged uiquitin. Cell lystes were prepred nd inuted with nti-ha monolonl ntiody (Rohe) nd protein G Sephrose 4 fst flow eds (GE Helthre). Immunopreipittes were resolved y SDS PAGE nd western lotting ws performed using nti-uiquitin ntiody (Zymed 1:). For o-immunopreipittion ssys, HEK 293T ells were o-trnsfeted with plsmids expressing Flg Fsd nd HA Skp1 (or HA Bd). Cell lystes were inuted with nti-flg ntiody (Sigm) nd protein G Sephrose 4 fst flow eds, followed y western lotting using indited ntiodies. dsrnai genertion nd sreening in the HA Bd-expressing stle S2 ells. For dsrnai sreening 6, 38 potentil Drosophil F ox proteins were determined from the Interpro dtse from the Europen Bioinformtis Institute. Primers used for PCR mplifition of eh gene frgment were hosen from the mplion dtse of the Drosophil RNAi Sreening Center t Hrvrd Medil Shool. To trnsrie dsrnas in vitro from the PCR frgments, eh primer ontined T7 RNA polymerse inding site (5ʹ-TAATACGACTCACTATAGGG-3ʹ) t 5ʹ. The dsrnas were generted from the MEGASCRIPT T7 trnsription kit (Amion) nd purified y the RNesy mini kit (Qigen). To minimize the effet of plte-to-plte vritions nd inrese the reliility of ssys during sreening, fold hnge in Bd levels for eh smple ws lulted y dividing the intensity of smple y the men of ll of the 12 smples on given plte, with ll experimentl nd quntifition steps performed stritly on side-y-side sis for ll smples on single plte. Emryo stining, imging nd intensity mesurement. Emryos were olleted t 4 h t the stted tempertures, fixed nd stined for mrna using digoxigenin (Rohe)-lelled ntisense RNA proe s previously desried 61,62 with the following modifitions: emryos were post-fixed with 1% (v/v) formldehyde (Fisher Sientifi) nd hyridiztion ws performed in uffer ontining.3% SDS (4 h inution) without protese K pre-tretment. Cy3-AffiniPure Donkey nti-mouse immunogloulin G (IgG; Jkson ImmunoReserh) ws used s the seondry ntiody. Imging nd quntifition were desried previously 45, with ll imges ptured within liner rnge under identil settings in single imge yle (for oth wild-type nd mutnt emryos tht were stined for gene of interest) to minimize mesurement errors. To mesure the FISH intensities in the ytoplsm, irulr window 61 pixels in size ws used to slide long the emryo edge immeditely outside of the nuler lyer (slly for h nd kni; pilly for eve) during the utomted imge proessing. It hs een shown tht gp gene nd segmenttion gene expression ptterns evolve during nuler yle 14 (refs, 63). To minimize suh effets nd llow urte omprison etween wild-type nd mutnt emryos, we seleted emryos within short developmentl windows with pek expression level for eh gene nlysed. Speifilly, we seleted emryos t erly nuler yle 14 with nuler height:width rtio of 1.3:1 to 1.7:1 for h nd kni expression ptterns, s desried previously 45. For h expression, emryos were further seleted to hve posterior h expression t normlized level etween. nd.. For kni expression, emryos were further seleted to ensure no detetle expression stripe t the position of pproximtely.. The segmenttion gene eve rehes its pek expression t lter time thn the gp genes, nd we seleted emryos t stge (efore the onset of gstrultion) shown to hve highly preise eve pttern 63. To further inrese the preision of developmentl time for eve nlysis, we seleted emryos tht hd the seventh stripe level > % of the first stripe level. The mesured oundry positions for ll three genes were highly preise within eh group of emryos, with stndrd devition tht is less thn tht of the oserved posterior shifts etween wild-type nd fsd emryos in eh se. Emryo stining, imging nd intensity mesurements with nti-bd ntiodies (Snt Cruz Biotehnology) were s desried previously 45. To ompre λ vl- ues of Bd profiles from wild-type nd fsd emryos, we first nlysed their men Bd intensity profiles s funtion of. Bd intensities used in this nlysis were kground-sutrted without further djustments. The liner fit of ln (B/B mx ) ws onduted using the Mtl softwre. The λ vlues in individul emryos were lulted s desried 45. In n lterntive fitting, we used the eqution of B = B mx e x/λ + C nd otined onsistent results; the men λ vlues lulted from Bd profiles of individul wild-type nd fsd emryos re.8 ± 9.9 μm nd 94. ± μm (P = ), respetively. Immunoytohemistry. Drosophil S2 ells were ultured on L-Tek II hmer slide nd trnsfeted with the HA Fsd-expressing plsmid. Cells were fixed, permeilized, inuted with nti-ha ntiody (1: dilution) nd Alex nture ell iology
10 ARTICLES Fluor 488 got nti-mouse IgG (1:4 dilution), nd imged using onfol mirosopy. TO PRO 3 dye (1 μm) ws used to stin nulei. Sttistil nlysis. All relevnt experimentl vlues represent men vlues nd stndrd devitions, with n representing the numer of independent smples. P vlues were lulted y the Student s t-test funtion (two-tiled) using the Mtl softwre (MthWorks). Exponentil nd liner fitting ws done through the urve fitting funtion of the softwre nd the Adjusted R 2 vlues 64 were lulted to determine how well the dt fit to the pplied model ( perfet fit hs vlue of 1). 58. Forler, D. et l. An effiient protein omplex purifition method for funtionl proteomis in higher eukryotes. Nt. Biotehnol. 21, (3). 59. Crevel, G. & Cotterill, S. DNA replition in ell-free extrts from Drosophil melnogster. EMBO J. 1, (1991). 6. Clemens, J. C. et l. Use of doule-strnded RNA interferene in Drosophil ell lines to disset signl trnsdution pthwys. Pro. Ntl Ad. Si. USA 97, (). 61. Tutz, D. & Pfeifle, C. A non-rdiotive in situ hyridiztion method for the loliztion of speifi RNAs in Drosophil emryos revels trnsltionl ontrol of the segmenttion gene hunhk. Chromosom 98, (1989). 62. Kosmn, D. et l. Multiplex detetion of RNA expression in Drosophil emryos. Siene 35, 846 (4). 63. Surkov, S. et l. Chrteriztion of the Drosophil segment determintion morphome. Developmentl iology 313, (8). 64. Lttin, J., Crroll, J. D. & Green, P. E. Anlyzing multivrite dt. (Thompson Books/ Cole, 3). nture ell iology VOLUME 13 NUMBER 1 JANUARY 11 31
 Time (min): HA-Bd DMSO epoxomiin 2 5 1 4 2 5 1 4 1 2 3 4 5 6 7 8 9 1 11 12 1 2 d HA-Bd WB: nti-β-tin β-tin 1 2 Figure S1 Bd degrdtion is")

11 supplementry informtion DOI: 1.138/n2141 HA-Bd 2 1 M r (K) Time (min): HA-Bd DMSO epoxomiin d HA-Bd WB: nti-β-tin β-tin 1 2 Figure S1 Bd degrdtion is sensitive to protesome inhiitors. () Fulllength HA-Bd protein deteted in the HA-Bd-expressing stle ells. Moleulr weight stndrds re shown. Lne 1 represents dt from ells with vetor lone. () Totl mount of HA-Bd in Drosophil S2 ells trnsiently trnsfeted with the HA-Bd expressing plsmid, with or without MG132 tretment. ( & d) HA-Bd degrdtion ws inhiited y protesome inhiitor epoxomiin in emryoni extrts. See Figs. 1 nd legends for further detils. 1

: Control skpa dsrna.5 1. 2. 4. 7.")
 The totl mount of Bd in HA-Bd-expressing stle ells is inresed y dsrna trgeting fsd (lne 3), ut not y dsrnas trgeting two F-ox ontrol genes (lnes 1 nd 2).")
12 supplementry informtion dsrna: FLAG-Skp1: WB: nti- β-tin Reltive Bd mount: HA-Bd β-tin WB: nti-β-tin WB: nti-flag HA-Bd β-tin FLAG-Skp1 d WB: nti-ha nti-β-tin Time (hr): Control skpa dsrna Control skpa dsrna skpa Figure S2 Skp1 prtiiptes in Bd degrdtion in Drosophil ells. () The totl mount of Bd in HA-Bd-expressing stle ells is inresed y dsrna trgeting fsd (lne 3), ut not y dsrnas trgeting two F-ox ontrol genes (lnes 1 nd 2). () Over-expression of Skp1 promotes Bd degrdtion. In this experiment, HA-Bd-expressing stle ells were trnsfeted with either the plsmid expressing FLAG-Skp1 or the vetor lone, nd the totl mount of HA-Bd ws nlyzed in the presene or sene of MG132. () CHX ssy in the HA-Bd-expressing stle ells in the presene or sene of skpa dsrna. See legend to Fig. 3 for further detils. (d) Stter plot of the experimentl dt from pnel ; see legend to Fig. 3 for further detils. 2
13 supplementry informtion Consensus: Figure S3 Alignment of F-ox domins in Drosophil. F-ox domins were derived from the Interpro dtse nd ligned using the Clustl W method. The two puttive F-ox domins of Fsd re inluded in this lignment. A loose onsensus is shown t the top of lignment, whih does not ontin ny invrile residues mong the sequenes nlyzed; the positions with the highest rte of identil residues re: 77% leuine t positions 3 nd 15, nd 72% isoleuine t position 11. Figur 3
 7/34 (.5%) 4/7 (57.1%) /45 (%) N/A pup dult 21/21 (%) 16/17 (94.1%) 15/15 (%) 39/42 (92.9%) 3/4 (.%) N/A d e f Figure S4 Chrteriztions of fsd gene.")
 Comprison of survivl rtes etween w 1118 nd fsd KG2393 flies t different tempertures nd stges. (d-f) Fsd is primrily lolized to the ytoplsm.")
14 supplementry informtion 18 C C 29 C w 1118 fsd KG2393 w 1118 fsd KG2393 w 1118 fsd KG2393 egg lrv lrv pup 23/ (92%) 21/21 (%) 17/35 (48.6%) 17/17 (%) 21/23 (91.3%) 15/15 (%) 49/6 (81.7%) 42/45 (93.3%) 7/34 (.5%) 4/7 (57.1%) /45 (%) N/A pup dult 21/21 (%) 16/17 (94.1%) 15/15 (%) 39/42 (92.9%) 3/4 (.%) N/A d e f Figure S4 Chrteriztions of fsd gene. ( & ) fsd trnsripts in wt emryos deteted y whole mount in situ hyridiztion using n fsd-speifi proe t pre-lstoderm () stge-7 () emryos. Sle r: μm. () Comprison of survivl rtes etween w 1118 nd fsd KG2393 flies t different tempertures nd stges. (d-f) Fsd is primrily lolized to the ytoplsm. S2 ells trnsfeted with the HA-Fsd expressing plsmid were fixed y formldehyde nd stined with To-pro-3 (d) nd nti-ha ntiody (e). Pnel f is merged imge of d nd e. Sle r: 1 μm. 4



 Ptterns of eve expression in wt")

15 supplementry informtion d e f g Figure S5 Ptterning defets in fsd emryos t 29 o C. (-d) Cutile ptterns of wt emryo () nd fsd emryos tht exhiit vrile A-P ptterning defets (-d). Cutiles re oriented with the nterior towrds the left nd dorsl upwrds. (e-g) Ptterns of eve expression in wt emryo (e) nd fsd emryos tht exhiit fused/missing eve expression stripes (f nd g). Sle r: μm. 5
16 s u p p l e m e n t r y i n f o r m t i o n wt (n = 7) eve eve fsd (n = 8) wt (n = 8) h d fsd (n = 7) h e wt (n = 9) kni f fsd (n = 8) kni Figure S6 Figure S6 Normlized FISH intensity dt extrted from individul emryos. Dt for eve ( nd ), h ( nd d) nd kni (e nd f) re shown seprtely for wt nd fsd emryos. Eh olor represents dt from n individul emryo. The men intensity nd stndrd devition re lso shown. 6
17 supplementry informtion d E1 /+ (n = 9) fsd KG2393 ;d E1 /+ (n = 9) h h d E1 /+ fsd KG2393 ;d E1 /+ h Figure S7 Geneti intertion etween d nd fsd. ( nd ) Normlized FISH h intensity dt extrted from individul emryos from d E1 /+ () nd fsd KG2393 ; d E1 /+ () femles. () Averge h intensity profiles from pnels nd. The nteriorly shifted h oundry position in d E1 /+ emryos (.368 ±.13, n = 9) is resued, prtilly, y mternl depletion of fsd (.39 ±.12 n = 9; p = ). The nteriorly shifted CF position in d E1 /+ emryos (.277 ±.15, n = 7) is lso prtilly resued y mternl depletion of fsd (.3 ±.12, n = 8; p = ). 7
18 supplementry informtion CG4663 KG2393 F-ox F-ox fsd (CG12765) CG467 (133p deletion) fsd 5-7 : wt (n = 9) fsd 5-7 (n = 1) wt fsd 5-7 d h h h Figure S8 Anlysis of null llele of fsd. () A shemti representtion of the fsd 5-7 llele generted y P-element medited exision. Green oxes represent the nnotted oding sequenes. Like fsd KG2393 flies, fsd 5-7 flies re homozygous vile t C ut emryos fil to hth t 29 C. The mesured CF positions for wt emryos nd emryos from fsd 5-7 femles (referred to fsd 5-7 emryos; ll results shown in this figure were otined t C) re.347 ±.9 (n = 9) nd.6 ±.13 (n = 13), respetively (p = ). ( nd ) Normlized FISH intensity dt for h extrted from individul wt () nd fsd 5-7 () emryos. (d) Averge h intensity profiles from nd. The men h oundry positions in wt nd fsd 5-7 emryos re.447 ±.15 (n = 9) nd.4 ±.11 (n = 1), respetively (p = ). 8







19 supplementry informtion Fig.1 Fig.1 WB: nti-β-tin Fig.2 Fig Fig.2e Fig.2g WB: nti-uiquitin Figure S9 Unropped Western lot dt. 9








20 supplementry informtion Fig.3 WB: nti-β-tin WB: nti-β-tin Fig.3 Fig.4 WB: nti-flag Fig.4 WB: nti-flag Figure S9 ontinued 1
SUPPLEMENTARY INFORMATION
 doi: 1.138/nture862 humn hr. 21q MRPL39 murine Chr.16 Mrpl39 Dyrk1A Runx1 murine Chr. 17 ZNF295 Ets2 Znf295 murine Chr. 1 COL18A1 -/- lot: nti-dscr1 IgG hevy hin DSCR1 DSCR1 expression reltive to hevy
doi: 1.138/nture862 humn hr. 21q MRPL39 murine Chr.16 Mrpl39 Dyrk1A Runx1 murine Chr. 17 ZNF295 Ets2 Znf295 murine Chr. 1 COL18A1 -/- lot: nti-dscr1 IgG hevy hin DSCR1 DSCR1 expression reltive to hevy
Title of Experiment: Author, Institute and address:
 Title of Experiment: Trsfetion of murine mrophge RAW264.7 ells with METAFECTENE PRO. Author, Institute n ress: Ptrizi Pellegtti n Frneso Di Virgilio. Deprtment of Experimentl n Dignosti Meiine, Setion
Title of Experiment: Trsfetion of murine mrophge RAW264.7 ells with METAFECTENE PRO. Author, Institute n ress: Ptrizi Pellegtti n Frneso Di Virgilio. Deprtment of Experimentl n Dignosti Meiine, Setion
Cos7 (3TP) (K): TGFβ1(h): (K)
 (K): TGFβ1(h): (K)") IP#2: IP#1: Totl Lystes luiferse tivity (K): 6-4 - (K): luiferse tivity luiferse tivity (K): 2 1 RL-: - + + + + + Sm4-3F: + - + + + + MYC-Sm3: - - - - + + TβRI-HA(T204D): - - - + - + α-ha Luiferse Ativity
IP#2: IP#1: Totl Lystes luiferse tivity (K): 6-4 - (K): luiferse tivity luiferse tivity (K): 2 1 RL-: - + + + + + Sm4-3F: + - + + + + MYC-Sm3: - - - - + + TβRI-HA(T204D): - - - + - + α-ha Luiferse Ativity
Alimonti_Supplementary Figure 1. Pten +/- Pten + Pten. Pten hy. β-actin. Pten - wt hy/+ +/- wt hy/+ +/- Pten. Pten. Relative Protein level (% )
") Alimonti_Supplementry Figure 1 hy 3 4 5 3 Neo 4 5 5 Proe 5 Proe hy/ hy/ /- - 3 6 Neo β-tin d Reltive Protein level (% ) 15 1 5 hy/ /- Reltive Gene Expr. (% ) 15 1 5 hy/ /- Supplementry Figure 1 Chrteriztion
Alimonti_Supplementry Figure 1 hy 3 4 5 3 Neo 4 5 5 Proe 5 Proe hy/ hy/ /- - 3 6 Neo β-tin d Reltive Protein level (% ) 15 1 5 hy/ /- Reltive Gene Expr. (% ) 15 1 5 hy/ /- Supplementry Figure 1 Chrteriztion
EFFECT OF DIETARY ENZYME ON PERFORMANCE OF WEANLING PIGS
 EFFECT OF DIETARY ENZYME ON PERFORMANCE OF WEANLING PIGS Finl report sumitted to Dniso Animl Nutrition E. vn Heugten nd B. Frederik North Crolin Stte University, Deprtment of Animl Siene Summry The urrent
EFFECT OF DIETARY ENZYME ON PERFORMANCE OF WEANLING PIGS Finl report sumitted to Dniso Animl Nutrition E. vn Heugten nd B. Frederik North Crolin Stte University, Deprtment of Animl Siene Summry The urrent
SUPPLEMENTARY INFORMATION
 % ells with ili (mrke y A-Tu) Reltive Luiferse % ells with ili (mrke y Arl13) % ells with ili DOI: 1.138/n2259 A-Tuulin Hoehst % Cilite Non-ilite -Serum 9% 8% 7% 1 6% % 4% +Serum 1 3% 2% 1% % Serum: -
% ells with ili (mrke y A-Tu) Reltive Luiferse % ells with ili (mrke y Arl13) % ells with ili DOI: 1.138/n2259 A-Tuulin Hoehst % Cilite Non-ilite -Serum 9% 8% 7% 1 6% % 4% +Serum 1 3% 2% 1% % Serum: -
SUPPLEMENTARY INFORMATION
 DOI: 1.138/n358 TLR2 nd MyD88 expression in murine mmmry epithelil supopultions. CD24 min plus MRU Myo-epithelil Luminl progenitor (CD61 pos ) Mture luminl (CD61 neg ) CD49f CD61 Reltive expression Krt5
DOI: 1.138/n358 TLR2 nd MyD88 expression in murine mmmry epithelil supopultions. CD24 min plus MRU Myo-epithelil Luminl progenitor (CD61 pos ) Mture luminl (CD61 neg ) CD49f CD61 Reltive expression Krt5
SUPPLEMENTARY INFORMATION
 doi:.8/nture98 : hr NEMO :5 hr IKK IKK NF-κB p65 p5 p65/-rel NF-κB p65 p5 p65/-rel Cytoplsm Cytoplsm p65/p5 Nuleus Nuleus NEMO IKK IKK d : hr > : hr p65/-rel NF- p65 p5 Cytoplsm Cytoplsm p65/p5 p65/-rel
doi:.8/nture98 : hr NEMO :5 hr IKK IKK NF-κB p65 p5 p65/-rel NF-κB p65 p5 p65/-rel Cytoplsm Cytoplsm p65/p5 Nuleus Nuleus NEMO IKK IKK d : hr > : hr p65/-rel NF- p65 p5 Cytoplsm Cytoplsm p65/p5 p65/-rel
SUPPLEMENTARY INFORMATION
 DOI: 1.13/n7 Reltive Pprg mrna 3 1 1 Time (weeks) Interspulr Inguinl Epididyml Reltive undne..1.5. - 5 5-51 51-1 1-7 7 - - 1 1-1 Lipid droplet size ( m ) 1-3 3 - - - 1 1-1 1-1 1-175 175-3 3-31 31-5 >5
DOI: 1.13/n7 Reltive Pprg mrna 3 1 1 Time (weeks) Interspulr Inguinl Epididyml Reltive undne..1.5. - 5 5-51 51-1 1-7 7 - - 1 1-1 Lipid droplet size ( m ) 1-3 3 - - - 1 1-1 1-1 1-175 175-3 3-31 31-5 >5
LHb VTA. VTA-projecting RMTg-projecting overlay. Supplemental Figure 2. Retrograde labeling of LHb neurons. a. VTA-projecting LHb
 SUPPLEMENTARY INFORMATION Supplementl Figure 1 doi:10.1038/nture09742 Lterl 1.0 mm from midline mpfc BNST mpfc BNST Lterl 2.1 mm from midline LHA LHA Lterl 2.7 mm from midline SUPPLEMENTAL INFORMATION
SUPPLEMENTARY INFORMATION Supplementl Figure 1 doi:10.1038/nture09742 Lterl 1.0 mm from midline mpfc BNST mpfc BNST Lterl 2.1 mm from midline LHA LHA Lterl 2.7 mm from midline SUPPLEMENTAL INFORMATION
SUPPLEMENTARY INFORMATION
 { OI: 1.138/n31 Srifie n nlyze APs on week 1 s of iet 1 4 6 High-ft iet BrU High-ft iet BrU 4 High-ft iet BrU 6 High-ft iet BrU Lin - Lin - : C34 + : C9 + 1 1 3 1 4 1 5 C45 1 C34 1 1 1 1 3 1 4 1 5 S-1
{ OI: 1.138/n31 Srifie n nlyze APs on week 1 s of iet 1 4 6 High-ft iet BrU High-ft iet BrU 4 High-ft iet BrU 6 High-ft iet BrU Lin - Lin - : C34 + : C9 + 1 1 3 1 4 1 5 C45 1 C34 1 1 1 1 3 1 4 1 5 S-1
Supplementary Figure S1
 Supplementry Figure S1 - UTR m - 3HA - 2-1 hgh - 1 Uiquitin *! *! lk distl promoter m K3R/ K121R-3HA UTR hgh founder lines - HA - - founder lines TG- E1 L A2 B1 F9 G6 H4 H6 B C D2 G1 H3 J2 L - 7 IP: lk
Supplementry Figure S1 - UTR m - 3HA - 2-1 hgh - 1 Uiquitin *! *! lk distl promoter m K3R/ K121R-3HA UTR hgh founder lines - HA - - founder lines TG- E1 L A2 B1 F9 G6 H4 H6 B C D2 G1 H3 J2 L - 7 IP: lk
P AND K IN POTATOES. Donald A Horneck Oregon State University Extension Service
 P AND K IN POTATOES Donld A Hornek Oregon Stte University Extension Servie INTRODUCTION Phosphorous nd potssium re importnt to grow high yielding nd qulity pottoes. Muh of the northwest hs hd trditionlly
P AND K IN POTATOES Donld A Hornek Oregon Stte University Extension Servie INTRODUCTION Phosphorous nd potssium re importnt to grow high yielding nd qulity pottoes. Muh of the northwest hs hd trditionlly
WesternBright Quantum
 WesternBright Quntum Quntify hemiluminesent Western lots over wie ynmi rnge WesternBright Quntum is new hemiluminesent regent speilly formulte for CCD imging. This novel Horserish peroxise (HRP) sustrte
WesternBright Quntum Quntify hemiluminesent Western lots over wie ynmi rnge WesternBright Quntum is new hemiluminesent regent speilly formulte for CCD imging. This novel Horserish peroxise (HRP) sustrte
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION CD169 + MACROPHAGES PRESENT LIPID ANTIGENS TO MEDIATE EARLY ACTIVATION OF INVARIANT NKT CELLS IN LYMPH NODES Ptrii Brrl, Polo Polzell, Andres Brukuer, Nio vn Rooijen, Gurdyl S.
SUPPLEMENTARY INFORMATION CD169 + MACROPHAGES PRESENT LIPID ANTIGENS TO MEDIATE EARLY ACTIVATION OF INVARIANT NKT CELLS IN LYMPH NODES Ptrii Brrl, Polo Polzell, Andres Brukuer, Nio vn Rooijen, Gurdyl S.
Supplementary Figure 1. Scheme of unilateral pyramidotomy used for detecting compensatory sprouting of intact CST axons.
 () BDA 2 weeks fter Py () AAVs Cre or GFP t P1 BDA 2 weeks fter Py CSMN CST () Py t P7 or 2 months () Py t 2 months Supplementry Figure 1. Sheme of unilterl pyrmidotomy used for deteting ompenstory sprouting
() BDA 2 weeks fter Py () AAVs Cre or GFP t P1 BDA 2 weeks fter Py CSMN CST () Py t P7 or 2 months () Py t 2 months Supplementry Figure 1. Sheme of unilterl pyrmidotomy used for deteting ompenstory sprouting
SUPPLEMENTARY INFORMATION
 doi:1.138/nture1794 BR EPFs BRI1? ERECTA TMM BSKs YDA PP2A BSU1 BIN2 pbzr1/2 BZR1/2 MKK4/5/7/9 MPK3/6 SPCH Cell growth Stomtl production Supplementry Figure 1. The model of BR nd stomtl signling pthwys.
doi:1.138/nture1794 BR EPFs BRI1? ERECTA TMM BSKs YDA PP2A BSU1 BIN2 pbzr1/2 BZR1/2 MKK4/5/7/9 MPK3/6 SPCH Cell growth Stomtl production Supplementry Figure 1. The model of BR nd stomtl signling pthwys.
FAK integrates growth-factor and integrin signals to promote cell migration
 integrtes growth-ftor nd integrin signls to promote ell migrtion rtiles Dvid J. Sieg*, Christof R. Huk*, Dusko Ili, Cndie K. Klingeil*, Erik Shefer, Croline H. Dmsky nd Dvid D. Shlepfer* *Deprtment of
integrtes growth-ftor nd integrin signls to promote ell migrtion rtiles Dvid J. Sieg*, Christof R. Huk*, Dusko Ili, Cndie K. Klingeil*, Erik Shefer, Croline H. Dmsky nd Dvid D. Shlepfer* *Deprtment of
Interplay of LRRK2 with chaperone-mediated autophagy
 Interply of with hperone-medited utophgy Smnth J Orenstein,, Sheng-Hn Kuo,, Inmuld Tsset,,, Espernz Aris,, Hiroshi Kog,, Irene Fernndez-Crs, Etty Cortes,5, Lwrene S Honig,5, Willim Duer 6, Antonell Consiglio,7,
Interply of with hperone-medited utophgy Smnth J Orenstein,, Sheng-Hn Kuo,, Inmuld Tsset,,, Espernz Aris,, Hiroshi Kog,, Irene Fernndez-Crs, Etty Cortes,5, Lwrene S Honig,5, Willim Duer 6, Antonell Consiglio,7,
SUPPLEMENTARY INFORMATION
 DOI: 1.138/n2977 Numer of ells per field 6 4 2 P =.1 Orthotopi eum Normlized ventrl photon flux 1E7 1E6 1E5 1E4 1E3 1E2 n=8 n=9 1 2 3 4 5 6 Dys Dy54 1.5E5 2.4E7 d Mie with lymph node metstsis (%) 1 8 6
DOI: 1.138/n2977 Numer of ells per field 6 4 2 P =.1 Orthotopi eum Normlized ventrl photon flux 1E7 1E6 1E5 1E4 1E3 1E2 n=8 n=9 1 2 3 4 5 6 Dys Dy54 1.5E5 2.4E7 d Mie with lymph node metstsis (%) 1 8 6
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:.8/nture89 4 4 Ilr -/- Ilr -/- Ilr -/- Cspse- -/- As -/- Nlrp -/- Il8 -/- Ilr -/- Supplementl figure. Inresed severity of NASH in inflmmsome-defiient mie, ut not in Ilr-defiient
SUPPLEMENTARY INFORMATION doi:.8/nture89 4 4 Ilr -/- Ilr -/- Ilr -/- Cspse- -/- As -/- Nlrp -/- Il8 -/- Ilr -/- Supplementl figure. Inresed severity of NASH in inflmmsome-defiient mie, ut not in Ilr-defiient
(% of adherent cells) *** PBL firm adhesion. Frequency (% ) 4 1 L 2 CXCR3 DP-2
 *** PBL firm adhesion. Frequency (% ) 4 1 L 2 CXCR3 DP-2") Chemotxis (% of dded ells) PBL totl dhesion (N ells/mm 2 /1.1 6 PBL) Frequeny (% ) PBL firm dhesion Supplementry Figure 1 4 4 3 3 2 2 1.1-4 1-3 1.1.2. 1 1 8 6 4 2 Adiponetin ( g/ml) - + Adiponetin ( g/ml)
Chemotxis (% of dded ells) PBL totl dhesion (N ells/mm 2 /1.1 6 PBL) Frequeny (% ) PBL firm dhesion Supplementry Figure 1 4 4 3 3 2 2 1.1-4 1-3 1.1.2. 1 1 8 6 4 2 Adiponetin ( g/ml) - + Adiponetin ( g/ml)
YAP transcriptionally regulates COX-2 expression and GCCSysm-4 (G-4), a dual YAP/COX-2 inhibitor, overcomes drug resistance in colorectal cancer
, a dual YAP/COX-2 inhibitor, overcomes drug resistance in colorectal cancer") Li et l. Journl of Experimentl & Clinil Cner Reserh (7) 36:44 DOI.86/s346-7-6-3 RESEARCH Open Aess trnsriptionlly regultes expression nd GCCSysm-4 (G-4), dul / inhiitor, overomes drug resistne in oloretl
Li et l. Journl of Experimentl & Clinil Cner Reserh (7) 36:44 DOI.86/s346-7-6-3 RESEARCH Open Aess trnsriptionlly regultes expression nd GCCSysm-4 (G-4), dul / inhiitor, overomes drug resistne in oloretl
A p75 NTR and Nogo receptor complex mediates repulsive signaling by myelin-associated glycoprotein
 A p75 NTR nd Nogo reeptor omplex medites repulsive signling y myelin-ssoited glyoprotein Sott T. Wong 1, John R. Henley 1, Kevin C. Knning 2, Kuo-hu Hung 1, Mrk Bothwell 2 nd Mu-ming Poo 1 1 Division of
A p75 NTR nd Nogo reeptor omplex medites repulsive signling y myelin-ssoited glyoprotein Sott T. Wong 1, John R. Henley 1, Kevin C. Knning 2, Kuo-hu Hung 1, Mrk Bothwell 2 nd Mu-ming Poo 1 1 Division of
Nucleosome positioning as a determinant of exon recognition
 Nuleosome positioning s determinnt of exon reognition Hgen Tilgner 1,3, Christoforos Nikolou 1,3, Sonj Althmmer 1, Mihel Smmeth 1, Miguel Beto 1, Jun Vlárel 1,2 & Roderi Guigó 1 200 Nture Ameri, In. All
Nuleosome positioning s determinnt of exon reognition Hgen Tilgner 1,3, Christoforos Nikolou 1,3, Sonj Althmmer 1, Mihel Smmeth 1, Miguel Beto 1, Jun Vlárel 1,2 & Roderi Guigó 1 200 Nture Ameri, In. All
Chloride Nutrition Regulates Water Balance in Plants
 XII Portuguese-Spnish Symposium on Plnt Wter Reltions Chloride Nutrition Regultes Wter Blne in Plnts Frno-Nvrro JD 1, Brumós J, Rosles MA 1, Vázquez-Rodríguez A 1, Sñudo BJ 1, Díz- Rued P 1, Rivero C 1,
XII Portuguese-Spnish Symposium on Plnt Wter Reltions Chloride Nutrition Regultes Wter Blne in Plnts Frno-Nvrro JD 1, Brumós J, Rosles MA 1, Vázquez-Rodríguez A 1, Sñudo BJ 1, Díz- Rued P 1, Rivero C 1,
The GCN5-CITED2-PKA signalling module controls hepatic glucose metabolism through a camp-induced substrate switch
 Reeived 6 Apr 216 Aepted 8 Sep 216 Pulished 22 Nov 216 DOI: 1.138/nomms13147 OPEN The GCN5-CITED2-PKA signlling module ontrols hepti gluose metolism through AMP-indued sustrte swith Mshito Ski 1, Tomoko
Reeived 6 Apr 216 Aepted 8 Sep 216 Pulished 22 Nov 216 DOI: 1.138/nomms13147 OPEN The GCN5-CITED2-PKA signlling module ontrols hepti gluose metolism through AMP-indued sustrte swith Mshito Ski 1, Tomoko
Roles of the PI-3K and MEK pathways in Ras-mediated chemoresistance in breast cancer cells
 ritish Journl of Cner (23) 89, 18 191 & 23 Cner Reserh UK All rights reserved 7 92/3 $2. www.jner.om Roles of the PI-3K nd MEK pthwys in Rs-medited hemoresistne in rest ner ells W Jin 1,LWu 1, K Ling 1,
ritish Journl of Cner (23) 89, 18 191 & 23 Cner Reserh UK All rights reserved 7 92/3 $2. www.jner.om Roles of the PI-3K nd MEK pthwys in Rs-medited hemoresistne in rest ner ells W Jin 1,LWu 1, K Ling 1,
Whangarei District Council Class 4 Gambling Venue Policy
 Whngrei Distrit Counil Clss 4 Gmling Venue Poliy April 2013 Whngrei Distrit Counil Clss 4 Gmling Venue Poliy Tle of ontents Introdution... 3 1 Ojetives of the poliy in so fr s promoted y the Gmling At
Whngrei Distrit Counil Clss 4 Gmling Venue Poliy April 2013 Whngrei Distrit Counil Clss 4 Gmling Venue Poliy Tle of ontents Introdution... 3 1 Ojetives of the poliy in so fr s promoted y the Gmling At
SUPPLEMENTARY INFORMATION
 Prentl doi:.8/nture57 Figure S HPMECs LM Cells Cell lines VEGF (ng/ml) Prentl 7. +/-. LM 7. +/-.99 LM 7. +/-.99 Fold COX induction 5 VEGF: - + + + Bevcizum: - - 5 (µg/ml) Reltive MMP LM mock COX MMP LM+
Prentl doi:.8/nture57 Figure S HPMECs LM Cells Cell lines VEGF (ng/ml) Prentl 7. +/-. LM 7. +/-.99 LM 7. +/-.99 Fold COX induction 5 VEGF: - + + + Bevcizum: - - 5 (µg/ml) Reltive MMP LM mock COX MMP LM+
AUTHOR COPY ONLY. Glycogen synthase kinase 3b mediates high glucose-induced ubiquitination and proteasome degradation of insulin receptor substrate 1
 Glyogen synthse kinse 3 medites high gluose-indued uiquitintion nd protesome degrdtion of insulin reeptor sustrte 1 171 Snhu Leng, Wenshuo Zhng, Ynin Zheng, Ziv Liermn 1, Christopher J Rhodes, Hgit Eldr-Finkelmn
Glyogen synthse kinse 3 medites high gluose-indued uiquitintion nd protesome degrdtion of insulin reeptor sustrte 1 171 Snhu Leng, Wenshuo Zhng, Ynin Zheng, Ziv Liermn 1, Christopher J Rhodes, Hgit Eldr-Finkelmn
TNF-a Downregulates Filaggrin and Loricrin through c-jun N-terminal Kinase: Role for TNF-a Antagonists to Improve Skin Barrier
 ORIGINAL ARTICLE TNF- Downregultes Filggrin nd Loririn through -Jun N-terminl Kinse: Role for TNF- Antgonists to Improve Skin Brrier Byung Eui Kim, Mihel D. Howell,, Emm Guttmn,, Ptrii M. Gilleudeu, Irm
ORIGINAL ARTICLE TNF- Downregultes Filggrin nd Loririn through -Jun N-terminl Kinse: Role for TNF- Antgonists to Improve Skin Brrier Byung Eui Kim, Mihel D. Howell,, Emm Guttmn,, Ptrii M. Gilleudeu, Irm
Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling
 Vol 461 1 Otoer 29 doi:1.138/nture8356 Tnkyrse inhiition stilizes xin nd ntgonizes Wnt signlling Shih-Min A. Hung 1, Yuji M. Mishin 1, Shnming Liu 1, Atwood Cheung 1, rnk Stegmeier 1, Gregory A. Mihud
Vol 461 1 Otoer 29 doi:1.138/nture8356 Tnkyrse inhiition stilizes xin nd ntgonizes Wnt signlling Shih-Min A. Hung 1, Yuji M. Mishin 1, Shnming Liu 1, Atwood Cheung 1, rnk Stegmeier 1, Gregory A. Mihud
supplementary information
 DOI:.38/n83 k Mouse Ch8 lous 8 9 Stop CHD8L 75 CHD8L Chromoomins Helise/ATPse omin DNA ining omin 5 kd NIH 3T3 MEF 93T HeL HCT UOS SOS.. CHD8L IB: CHD8 8 5 L S Reltive mrna mount 3... Reltive mrna mount.8.
DOI:.38/n83 k Mouse Ch8 lous 8 9 Stop CHD8L 75 CHD8L Chromoomins Helise/ATPse omin DNA ining omin 5 kd NIH 3T3 MEF 93T HeL HCT UOS SOS.. CHD8L IB: CHD8 8 5 L S Reltive mrna mount 3... Reltive mrna mount.8.
Poultry No The replacement value of betaine for DL-methionine and Choline in broiler diets
 Poultry No. 1573 The replement vlue of etine for DL-methionine nd Choline in roiler diets Key Informtion In roiler diets defiient in sulfur mino ids ut dequtely supplemented with methyl groups vi dded
Poultry No. 1573 The replement vlue of etine for DL-methionine nd Choline in roiler diets Key Informtion In roiler diets defiient in sulfur mino ids ut dequtely supplemented with methyl groups vi dded
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nture07679 Emryonic Stem (ES) cell Hemngiolst Flk1 + Blst Colony 3 to 3.5 Dys 3-4 Dys ES differentition Sort of Flk1 + cells Supplementry Figure 1. Chrcteristion of lst colony development.
doi: 10.1038/nture07679 Emryonic Stem (ES) cell Hemngiolst Flk1 + Blst Colony 3 to 3.5 Dys 3-4 Dys ES differentition Sort of Flk1 + cells Supplementry Figure 1. Chrcteristion of lst colony development.
Tbp. Per Relative mrna levels Circadian Time. Liver weight/ body weight (%) n.s. Pernull
 n.s. Pernull") Liver weight/ ody weight (%) Dy Body weight (g) Reltive mrna levels Reltive mrna levels Reltive mrna levels Reltive mrna levels Dy Per1 Per2 Per3 Tp 8 2 8 2. 6 2 8 12162 Cirdin Time 3 2 1 2 1 1 8 12162
Liver weight/ ody weight (%) Dy Body weight (g) Reltive mrna levels Reltive mrna levels Reltive mrna levels Reltive mrna levels Dy Per1 Per2 Per3 Tp 8 2 8 2. 6 2 8 12162 Cirdin Time 3 2 1 2 1 1 8 12162
SUPPLEMENTARY INFORMATION
 2 weeks high holesterol diet 2 weeks high holesterol diet 2 weeks high holesterol diet 2 μm Mrophges Crystls Hoehst μm Mrophges Crystls Hoehst Hoehst Crystls Mrophges 2 μm 2 μm Supplementry Fig. 1: Erly
2 weeks high holesterol diet 2 weeks high holesterol diet 2 weeks high holesterol diet 2 μm Mrophges Crystls Hoehst μm Mrophges Crystls Hoehst Hoehst Crystls Mrophges 2 μm 2 μm Supplementry Fig. 1: Erly
Genome-wide nucleosome positioning during embryonic stem cell development
 Genome-wide nuleosome positioning during emryoni stem ell development Vldimir B Teif 1,2, Yevhen Vinshtein 2,3, Mïwen Cudron-Herger 1,2, Jn-Philipp Mllm 1,2, Croline Mrth 1,2, Thoms Höfer 2,3 & Krsten
Genome-wide nuleosome positioning during emryoni stem ell development Vldimir B Teif 1,2, Yevhen Vinshtein 2,3, Mïwen Cudron-Herger 1,2, Jn-Philipp Mllm 1,2, Croline Mrth 1,2, Thoms Höfer 2,3 & Krsten
The Hippo/YAP pathway interacts with EGFR signaling and HPV oncoproteins to regulate cervical cancer progression
 Reserh Artile The Hippo/ pthwy interts with EGFR signling nd HPV onoproteins to regulte ervil ner progression Chuno He 1,, Dgn Mo 1,3, Guohu Hu 1,, Xingmin Lv 1, Xingheng Chen, Peter C Angeletti 5, Jixin
Reserh Artile The Hippo/ pthwy interts with EGFR signling nd HPV onoproteins to regulte ervil ner progression Chuno He 1,, Dgn Mo 1,3, Guohu Hu 1,, Xingmin Lv 1, Xingheng Chen, Peter C Angeletti 5, Jixin
Ulk λ PPase. 32 P-Ulk1 32 P-GST-TSC2. Ulk1 GST (TSC2) : Ha-Ulk1 : AMPK. WB: Ha (Ulk1) : Glu. h CON - Glu - A.A WB: LC3 AMPK-WT AMPK-DKO
 : Ha-Ulk1 : AMPK. WB: Ha (Ulk1) : Glu. h CON - Glu - A.A WB: LC3 AMPK-WT AMPK-DKO") DOI: 10.1038/ncb2152 C.C + - + - : Glu b Ulk1 - - + λ PPse c AMPK + - + + : ATP P-GST-TSC2 WB: Flg (Ulk1) WB Ulk1 WB: H (Ulk1) GST (TSC2) C.C d e WT K46R - + - + : H-Ulk1 : AMPK - + - + + + AMPK H-Ulk1
DOI: 10.1038/ncb2152 C.C + - + - : Glu b Ulk1 - - + λ PPse c AMPK + - + + : ATP P-GST-TSC2 WB: Flg (Ulk1) WB Ulk1 WB: H (Ulk1) GST (TSC2) C.C d e WT K46R - + - + : H-Ulk1 : AMPK - + - + + + AMPK H-Ulk1
supplementary information
 DOI: 10.1038/nc2089 H3K4me1 H3K4me1 H3K4me1 H3K4me1 H3K4me1 H3K4me1 5 PN N1-2 PN H3K4me1 H3K4me1 H3K4me1 2-cell stge 2-c st cell ge Figure S1 Pttern of loclistion of H3K4me1 () nd () during zygotic development
DOI: 10.1038/nc2089 H3K4me1 H3K4me1 H3K4me1 H3K4me1 H3K4me1 H3K4me1 5 PN N1-2 PN H3K4me1 H3K4me1 H3K4me1 2-cell stge 2-c st cell ge Figure S1 Pttern of loclistion of H3K4me1 () nd () during zygotic development
SUPPLEMENTARY INFORMATION
 doi:.38/nture277 d 25 25 2 Time from sound onset (ms) 25 25 2 Time from sound onset (ms) Firing rte (spikes/s) Firing rte (spikes/s).8.6..2 e f g h.8.6..2 Frtion of neurons Frtion of neurons N = 53 2 2
doi:.38/nture277 d 25 25 2 Time from sound onset (ms) 25 25 2 Time from sound onset (ms) Firing rte (spikes/s) Firing rte (spikes/s).8.6..2 e f g h.8.6..2 Frtion of neurons Frtion of neurons N = 53 2 2
Introduction to Study Designs II
 Introdution to Study Designs II Commonly used study designs in publi helth & epidemiologi reserh Benjmin Rihrd H. Muthmbi, DrPH, MPH Stte HIV Epidemiologist HIV Epidemiology Investigtion Setion PA Deprtment
Introdution to Study Designs II Commonly used study designs in publi helth & epidemiologi reserh Benjmin Rihrd H. Muthmbi, DrPH, MPH Stte HIV Epidemiologist HIV Epidemiology Investigtion Setion PA Deprtment
GSK-3 is a master regulator of neural progenitor homeostasis
 GSK-3 is mster regultor of neurl progenitor homeostsis Woo-Yng Kim 1, Xinshuo Wng 1, Yohong Wu 1, Brdley W Dole 2, Stish Ptel 3, Jmes R Woodgett 3 & Willim D Snider 1 The development of the rin requires
GSK-3 is mster regultor of neurl progenitor homeostsis Woo-Yng Kim 1, Xinshuo Wng 1, Yohong Wu 1, Brdley W Dole 2, Stish Ptel 3, Jmes R Woodgett 3 & Willim D Snider 1 The development of the rin requires
Supplementary Information
 Supplementry Informtion A new lss of plnt lipid is essentil for protetion ginst phosphorus depletion Yozo Okzki 1, Hitomi Otsuki 1, Tomoko Nrisw 1, Mkoto Koyshi 1, Storu Swi 2, Yukiko Kmide 1, Miyko Kusno
Supplementry Informtion A new lss of plnt lipid is essentil for protetion ginst phosphorus depletion Yozo Okzki 1, Hitomi Otsuki 1, Tomoko Nrisw 1, Mkoto Koyshi 1, Storu Swi 2, Yukiko Kmide 1, Miyko Kusno
Osteoblasts secrete Cxcl9 to regulate angiogenesis in bone
 Reeived 2 De 215 Aepted 9 Nov 216 Pulished 14 De 216 DOI: 1.138/nomms13885 OPEN Osteolsts serete to regulte ngiogenesis in one Bin Hung 1,, Wenho Wng 1,, Qinghu Li 1,, Zhenyu Wng 1,BoYn 1, Zhongmin Zhng
Reeived 2 De 215 Aepted 9 Nov 216 Pulished 14 De 216 DOI: 1.138/nomms13885 OPEN Osteolsts serete to regulte ngiogenesis in one Bin Hung 1,, Wenho Wng 1,, Qinghu Li 1,, Zhenyu Wng 1,BoYn 1, Zhongmin Zhng
AJ PUTT. Hematology. Chemistry. Species: Canine Gender: Female Year of Birth: 2013 Client: PUTT
 Speies: Cnine Gender: Femle Yer of Birth: 2013 Client: PUTT Requisition #: 9034-12 Aession #: W2152816 Aount Code: 72364 Veterinrin: CARTER Pnel/Profile: Tik Pnel Add-on Senior Profile with L 4Dx Plus
Speies: Cnine Gender: Femle Yer of Birth: 2013 Client: PUTT Requisition #: 9034-12 Aession #: W2152816 Aount Code: 72364 Veterinrin: CARTER Pnel/Profile: Tik Pnel Add-on Senior Profile with L 4Dx Plus
Inhibitory effect of p38 mitogen-activated protein kinase inhibitors on cytokine release from human macrophages
 British Journl of Phrmology (26) 149, 393 44 & 26 Nture Pulishing Group All rights reserved 7 1188/6 $3. www.rjphrmol.org RESEARCH PAPER Inhiitory effet of p38 mitogen-tivted protein kinse inhiitors on
British Journl of Phrmology (26) 149, 393 44 & 26 Nture Pulishing Group All rights reserved 7 1188/6 $3. www.rjphrmol.org RESEARCH PAPER Inhiitory effet of p38 mitogen-tivted protein kinse inhiitors on
Chow KD CR HFD. Fed Fast Refed
 Supplementry Figure 1 Control d/d Chow KD CR Fed Fst Refed Supplementry Figure 1: Liver expression in diet nd disese models. () expression in the livers of ontrol nd d/d mie. () expression in the livers
Supplementry Figure 1 Control d/d Chow KD CR Fed Fst Refed Supplementry Figure 1: Liver expression in diet nd disese models. () expression in the livers of ontrol nd d/d mie. () expression in the livers
Using Paclobutrazol to Suppress Inflorescence Height of Potted Phalaenopsis Orchids
 Using Pcloutrzol to Suppress Inflorescence Height of Potted Phlenopsis Orchids A REPORT SUBMITTED TO FINE AMERICAS Linsey Newton nd Erik Runkle Deprtment of Horticulture Spring 28 Using Pcloutrzol to Suppress
Using Pcloutrzol to Suppress Inflorescence Height of Potted Phlenopsis Orchids A REPORT SUBMITTED TO FINE AMERICAS Linsey Newton nd Erik Runkle Deprtment of Horticulture Spring 28 Using Pcloutrzol to Suppress
Acute and gradual increases in BDNF concentration elicit distinct signaling and functions in neurons
 nd grdul increses in BDNF concentrtion elicit distinct signling nd functions in neurons Yunyun Ji,, Yun Lu, Feng Yng, Wnhu Shen, Tin Tze-Tsng Tng,, Linyin Feng, Shumin Dun, nd Bi Lu,.. - Grdul (normlized
nd grdul increses in BDNF concentrtion elicit distinct signling nd functions in neurons Yunyun Ji,, Yun Lu, Feng Yng, Wnhu Shen, Tin Tze-Tsng Tng,, Linyin Feng, Shumin Dun, nd Bi Lu,.. - Grdul (normlized
SUPPLEMENTARY INFORMATION
 Supplementry Figure 1. Genertion of N- nd C-tgged cyclin D1 knock-in mice., N-tgged cyclin D1 gene trgeting construct, cyclin D1 genomic locus, cyclin D1 locus following homologous recomintion (trgeted
Supplementry Figure 1. Genertion of N- nd C-tgged cyclin D1 knock-in mice., N-tgged cyclin D1 gene trgeting construct, cyclin D1 genomic locus, cyclin D1 locus following homologous recomintion (trgeted
regulates stem cells through Wnt/β-catenin signalling
 LETTERS O regultes stem ells through Wnt/β-tenin signlling Jolly Mzumdr,,7, W. Timothy O Brien, Rndll S. Johnson, Joseph C. LMnn, Jun C. Chvez 5, Peter S. Klein nd M. Celeste Simon,, Stem ells reside in
LETTERS O regultes stem ells through Wnt/β-tenin signlling Jolly Mzumdr,,7, W. Timothy O Brien, Rndll S. Johnson, Joseph C. LMnn, Jun C. Chvez 5, Peter S. Klein nd M. Celeste Simon,, Stem ells reside in
Plant Physiology Preview. Published on February 21, 2017, as DOI: /pp
 Plnt Physiology Preview. Pulished on Ferury 21, 217, s DOI:1.114/pp.16.1928 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16 17 18 Running title: Spliing regultor STA1 in het stress dpttion Corresponding Author:
Plnt Physiology Preview. Pulished on Ferury 21, 217, s DOI:1.114/pp.16.1928 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16 17 18 Running title: Spliing regultor STA1 in het stress dpttion Corresponding Author:
larvi 2013 Epigenetic regulation of muscle development and growth in Senegalese sole larvae Catarina Campos
 lrvi 213 6th fish & shellfish lrviulture symposium Epigeneti regultion of musle development nd growth in Seneglese sole lrve Ctrin Cmpos ghent university, elgium, 2-5 septemer 213 EPIGENETIC REGULATION
lrvi 213 6th fish & shellfish lrviulture symposium Epigeneti regultion of musle development nd growth in Seneglese sole lrve Ctrin Cmpos ghent university, elgium, 2-5 septemer 213 EPIGENETIC REGULATION
Copy Number ID2 MYCN ID2 MYCN. Copy Number MYCN DDX1 ID2 KIDINS220 MBOAT2 ID2
 Copy Numer Copy Numer Copy Numer Copy Numer DIPG38 DIPG49 ID2 MYCN ID2 MYCN c DIPG01 d DIPG29 ID2 MYCN ID2 MYCN e STNG2 f MYCN DIPG01 Chr. 2 DIPG29 Chr. 1 MYCN DDX1 Chr. 2 ID2 KIDINS220 MBOAT2 ID2 Supplementry
Copy Numer Copy Numer Copy Numer Copy Numer DIPG38 DIPG49 ID2 MYCN ID2 MYCN c DIPG01 d DIPG29 ID2 MYCN ID2 MYCN e STNG2 f MYCN DIPG01 Chr. 2 DIPG29 Chr. 1 MYCN DDX1 Chr. 2 ID2 KIDINS220 MBOAT2 ID2 Supplementry
Lesions of prefrontal cortex reduce attentional modulation of neuronal responses. and synchrony in V4
 Lesions of prefrontl ortex reue ttentionl moultion of neuronl responses n synhrony in V4 Georgi G. Gregoriou,, Anrew F. Rossi, 3 Leslie G Ungerleier, 4 Roert Desimone 5 Deprtment of Bsi Sienes, Fulty of
Lesions of prefrontl ortex reue ttentionl moultion of neuronl responses n synhrony in V4 Georgi G. Gregoriou,, Anrew F. Rossi, 3 Leslie G Ungerleier, 4 Roert Desimone 5 Deprtment of Bsi Sienes, Fulty of
FRAMEstar. 2-Component PCR Plates
 FRAMEstr -Component Pltes FrmeStr two-omponent tehnology redues evportion from pltes, improving results nd llowing for volume redutions to sve on expensive regents. FrmeStr pltes mximise therml stility
FRAMEstr -Component Pltes FrmeStr two-omponent tehnology redues evportion from pltes, improving results nd llowing for volume redutions to sve on expensive regents. FrmeStr pltes mximise therml stility
Multisite phosphorylation of a CDK inhibitor sets a threshold for the onset of DNA replication
 Multisite phosphoryltion of CDK inhiitor sets threshold for the onset of DNA replition iers Nsh*², iojing Tng*², Stephen Orliky*, Qinghu Chen³, Frnk B. Gertler, Mihel D. Mendenhll³, Frnk Siheri*k, Tony
Multisite phosphoryltion of CDK inhiitor sets threshold for the onset of DNA replition iers Nsh*², iojing Tng*², Stephen Orliky*, Qinghu Chen³, Frnk B. Gertler, Mihel D. Mendenhll³, Frnk Siheri*k, Tony
LETTERS. Chfr is required for tumor suppression and Aurora A regulation
 25 Nture Pulishing Group http://www.nture.om/nturegenetis Chfr is required for tumor suppression nd Auror A regultion Xiohun Yu 1, Ktherine Minter-Dykhouse 1, Liviu Mlurenu 2, Wei-Meng Zho 3, Dongwei Zhng
25 Nture Pulishing Group http://www.nture.om/nturegenetis Chfr is required for tumor suppression nd Auror A regultion Xiohun Yu 1, Ktherine Minter-Dykhouse 1, Liviu Mlurenu 2, Wei-Meng Zho 3, Dongwei Zhng
Maintenance of protein synthesis reading frame by EF-P and m 1 G37-tRNA
 Reeived 23 Nov 214 Aepted 2 Apr 215 Pulished 26 My 215 DOI: 1.138/nomms8226 Mintenne of protein synthesis reding frme y EF-P nd m 1 G37-tRNA Howrd B. Gmper 1, *, Iso Msud 1, *, Miln Frenkel-Morgenstern
Reeived 23 Nov 214 Aepted 2 Apr 215 Pulished 26 My 215 DOI: 1.138/nomms8226 Mintenne of protein synthesis reding frme y EF-P nd m 1 G37-tRNA Howrd B. Gmper 1, *, Iso Msud 1, *, Miln Frenkel-Morgenstern
Differential expression of cyclin G2, cyclin dependent kinase inhibitor 2C and peripheral myelin protein 22 genes during adipogenesis..
 The Ohio Stte University From the SeletedWorks of Jiin Zhng Summer My 5, 214 Differentil expression of ylin G2, ylin dependent kinse inhiitor 2C nd peripherl myelin protein 22 genes during dipogenesis..pdf
The Ohio Stte University From the SeletedWorks of Jiin Zhng Summer My 5, 214 Differentil expression of ylin G2, ylin dependent kinse inhiitor 2C nd peripherl myelin protein 22 genes during dipogenesis..pdf
a3 Chains of type V collagen regulate breast tumour growth via glypican-1
 Reeive 5 Aug 16 Aepte De 16 Pulishe 19 Jn 17 3 Chins of type V ollgen regulte rest tumour growth vi glypin-1 Guorui Hung 1, Goxing Ge 1,w, Vlerio Izzi & Dniel S. Greenspn 1 DOI: 1.138/nomms1351 OPEN Periellulr
Reeive 5 Aug 16 Aepte De 16 Pulishe 19 Jn 17 3 Chins of type V ollgen regulte rest tumour growth vi glypin-1 Guorui Hung 1, Goxing Ge 1,w, Vlerio Izzi & Dniel S. Greenspn 1 DOI: 1.138/nomms1351 OPEN Periellulr
BTLA is a lymphocyte inhibitory receptor with similarities to CTLA-4 and PD-1
 BTLA is lymphoyte inhiitory reeptor with similrities to CTLA-4 nd PD-1 Norihiko Wtne 1,5, My Gvrieli 1, John R Sedy 1, Jinfei Yng 1,5, Frnes Fllrino 2, Susn K Loftin 1, Mihelle A Hurhl 1, Ntlie Zimmermn
BTLA is lymphoyte inhiitory reeptor with similrities to CTLA-4 nd PD-1 Norihiko Wtne 1,5, My Gvrieli 1, John R Sedy 1, Jinfei Yng 1,5, Frnes Fllrino 2, Susn K Loftin 1, Mihelle A Hurhl 1, Ntlie Zimmermn
EFFECTS OF AN ACUTE ENTERIC DISEASE CHALLENGE ON IGF-1 AND IGFBP-3 GENE EXPRESSION IN PORCINE SKELETAL MUSCLE
 Swine Dy 22 Contents EFFECTS OF AN ACUTE ENTERIC DISEASE CHALLENGE ON IGF-1 AND IGFBP-3 GENE EXPRESSION IN PORCINE SKELETAL MUSCLE B. J. Johnson, J. P. Kyser, J. D. Dunn, A. T. Wyln, S. S. Dritz 1, J.
Swine Dy 22 Contents EFFECTS OF AN ACUTE ENTERIC DISEASE CHALLENGE ON IGF-1 AND IGFBP-3 GENE EXPRESSION IN PORCINE SKELETAL MUSCLE B. J. Johnson, J. P. Kyser, J. D. Dunn, A. T. Wyln, S. S. Dritz 1, J.
Neural population coding of sound level adapts to stimulus statistics
 COMPUTATION AND SYSTEMS 25 Nture Pulishing Group http://www.nture.om/ntureneurosiene Neurl popultion oding of sound level dpts to stimulus sttistis Isel Den 1, Niol S Hrper 1,2 & Dvid MAlpine 1 Mmmls n
COMPUTATION AND SYSTEMS 25 Nture Pulishing Group http://www.nture.om/ntureneurosiene Neurl popultion oding of sound level dpts to stimulus sttistis Isel Den 1, Niol S Hrper 1,2 & Dvid MAlpine 1 Mmmls n
Other Uses for Cluster Sampling
 Other Uses for Cluster Smpling Mesure hnges in the level of n ttriute Hypothesis testing versus intervl estimtion Type I n 2 errors Power of the test Mesuring ttriute t sme time in ifferent sites Exmple:
Other Uses for Cluster Smpling Mesure hnges in the level of n ttriute Hypothesis testing versus intervl estimtion Type I n 2 errors Power of the test Mesuring ttriute t sme time in ifferent sites Exmple:
N6-methyladenosine (m6a) is the most prevalent messenger
 is the most prevalent messenger") https://oi.org/8/s556-8-7- m 6 A mrna methyltion regultes tivity to promote the prolifertion n tumorigeniity of enometril ner Jun Liu,,, Mrk A. Ekert,, Bryn T. Hr,,, Song-Mei Liu,, Zhike Lu,, Kngkng Yu,,5,
https://oi.org/8/s556-8-7- m 6 A mrna methyltion regultes tivity to promote the prolifertion n tumorigeniity of enometril ner Jun Liu,,, Mrk A. Ekert,, Bryn T. Hr,,, Song-Mei Liu,, Zhike Lu,, Kngkng Yu,,5,
The microrna mir-31 inhibits CD8 + T cell function in chronic viral infection
 A rt i l e s The mirorna mir-3 inhiits CD8 + T ell funtion in hroni virl infetion Howell F Moffett, Adm N R Crtwright, Hye-Jung Kim, Jernej Gode, Json Pyrdol, Trmo Äijö 3, Gustvo J Mrtinez,6, Anjn Ro,
A rt i l e s The mirorna mir-3 inhiits CD8 + T ell funtion in hroni virl infetion Howell F Moffett, Adm N R Crtwright, Hye-Jung Kim, Jernej Gode, Json Pyrdol, Trmo Äijö 3, Gustvo J Mrtinez,6, Anjn Ro,
Trans-dominant inhibition of RNA viral replication can slow growth of drug-resistant viruses
 25 Nture Pulishing Group http://www.nture.om/nturegenetis Trns-dominnt inhiition of RNA virl replition n slow growth of drug-resistnt viruses Sott Crowder & Krl Kirkegrd The high error rtes of virl RNA-dependent
25 Nture Pulishing Group http://www.nture.om/nturegenetis Trns-dominnt inhiition of RNA virl replition n slow growth of drug-resistnt viruses Sott Crowder & Krl Kirkegrd The high error rtes of virl RNA-dependent
Supplementary figure 1
 Supplementry figure 1 Dy 8 post LCMV infection Vsculr Assoc. Prenchym Dy 3 post LCMV infection 1 5 6.7.29 1 4 1 3 1 2 88.9 4.16 1 2 1 3 1 4 1 5 1 5 1.59 5.97 1 4 1 3 1 2 21.4 71 1 2 1 3 1 4 1 5 1 5.59.22
Supplementry figure 1 Dy 8 post LCMV infection Vsculr Assoc. Prenchym Dy 3 post LCMV infection 1 5 6.7.29 1 4 1 3 1 2 88.9 4.16 1 2 1 3 1 4 1 5 1 5 1.59 5.97 1 4 1 3 1 2 21.4 71 1 2 1 3 1 4 1 5 1 5.59.22
ARTICLE. I. Chopra & H. F. Li & H. Wang & K. A. Webster
 Dietologi (212) 55:783 794 DOI 1.17/s125-11-247-y ARTICLE Phosphoryltion of the insulin reeptor y AMP-tivted protein kinse (AMPK) promotes lignd-independent tivtion of the insulin signlling pthwy in rodent
Dietologi (212) 55:783 794 DOI 1.17/s125-11-247-y ARTICLE Phosphoryltion of the insulin reeptor y AMP-tivted protein kinse (AMPK) promotes lignd-independent tivtion of the insulin signlling pthwy in rodent
SUPPLEMENTARY INFORMATION
 DOI: 1.138/nc286 Figure S1 e f Medium DMSO AktVIII PP242 Rp S6K1-I Gr1 + + + + + + Strvtion + + + + + IB: Akt-pT38 IB: Akt K-pT389 K IB: Rptor Gr1 shs6k1-a shs6k1-b shs6k1-c shrictor shrptor Gr1 c IB:
DOI: 1.138/nc286 Figure S1 e f Medium DMSO AktVIII PP242 Rp S6K1-I Gr1 + + + + + + Strvtion + + + + + IB: Akt-pT38 IB: Akt K-pT389 K IB: Rptor Gr1 shs6k1-a shs6k1-b shs6k1-c shrictor shrptor Gr1 c IB:
TNF-α (pg/ml) IL-6 (ng/ml)
 IL-6 (ng/ml)") Xio, et l., Supplementry Figure 1 IL-6 (ng/ml) TNF-α (pg/ml) 16 12 8 4 1,4 1,2 1, 8 6 4 2 med Cl / Pm3CSK4 zymosn curdln Poly (I:C) LPS flgelin MALP-2 imiquimod R848 CpG TNF-α (pg/ml) IL-6 (ng/ml) 2 1.6
Xio, et l., Supplementry Figure 1 IL-6 (ng/ml) TNF-α (pg/ml) 16 12 8 4 1,4 1,2 1, 8 6 4 2 med Cl / Pm3CSK4 zymosn curdln Poly (I:C) LPS flgelin MALP-2 imiquimod R848 CpG TNF-α (pg/ml) IL-6 (ng/ml) 2 1.6
PDGF-BB secreted by preosteoclasts induces angiogenesis during coupling with osteogenesis
 Supplementry Informtion PDGF-BB secreted y preosteoclsts induces ngiogenesis during coupling with osteogenesis Hui Xie, Zhung Cui, Long Wng, Zhuying Xi, Yin Hu, Lingling Xin, Chngjun Li, Ling Xie, Jnet
Supplementry Informtion PDGF-BB secreted y preosteoclsts induces ngiogenesis during coupling with osteogenesis Hui Xie, Zhung Cui, Long Wng, Zhuying Xi, Yin Hu, Lingling Xin, Chngjun Li, Ling Xie, Jnet
 nestin ironetin p75 s1 CNS SKPs Dermo-1 +ve SKPs CNS H2O SCGs Skin Di. SKPs TH SHOX2 GAPDH NCAM D H Figure S1, Immunoytohemil nlysis o SKP spheres ulture rom neontl mouse (nestin, ironetin, S-1) or rt
nestin ironetin p75 s1 CNS SKPs Dermo-1 +ve SKPs CNS H2O SCGs Skin Di. SKPs TH SHOX2 GAPDH NCAM D H Figure S1, Immunoytohemil nlysis o SKP spheres ulture rom neontl mouse (nestin, ironetin, S-1) or rt
Protein tyrosine phosphatase 1B deficiency or inhibition delays ErbB2-induced mammary tumorigenesis and protects from lung metastasis
 Protein tyrosine phosphtse 1B defiieny or inhiition delys ErB2-indued mmmry tumorigenesis nd protets from lung metstsis Sofi G Julien 1,5, Ndi Dué 1,6, Mihelle Red 1, Jnie Penney 1, Mrilene Pquet 2, Yongxin
Protein tyrosine phosphtse 1B defiieny or inhiition delys ErB2-indued mmmry tumorigenesis nd protets from lung metstsis Sofi G Julien 1,5, Ndi Dué 1,6, Mihelle Red 1, Jnie Penney 1, Mrilene Pquet 2, Yongxin
Minimum effective dose of chenic acid for gallstone patients: reduction with bedtime administration and
 Gut, 1982, 23, 28-284 Minimum effetive dose of heni id for gllstone ptients: redution with bedtime dministrtion nd low holesterol diet D P MUDGL, R M KUPFER, ND T C NORTHFIELD* From the Normn Tnner Gstroenterology
Gut, 1982, 23, 28-284 Minimum effetive dose of heni id for gllstone ptients: redution with bedtime dministrtion nd low holesterol diet D P MUDGL, R M KUPFER, ND T C NORTHFIELD* From the Normn Tnner Gstroenterology
LETTERS. Disulphide-isomerase-enabled shedding of tumour-associated NKG2D ligands
 Vol 447 24 My 2007 doi:10.1038/nture05768 LETTERS Disulphide-isomerse-enled shedding of tumour-ssoited NKG2D lignds Brett K. Kiser 1 *, Desong Yim 1 *, I-Ting Chow 1 *, Segundo Gonzlez 1 {, Zhenpeng Di
Vol 447 24 My 2007 doi:10.1038/nture05768 LETTERS Disulphide-isomerse-enled shedding of tumour-ssoited NKG2D lignds Brett K. Kiser 1 *, Desong Yim 1 *, I-Ting Chow 1 *, Segundo Gonzlez 1 {, Zhenpeng Di
 DOI: 10.1038/nc2331 PCre;Ros26R 12 h induction 48 h induction Vegfr3 i EC c d ib4 24 h induction VEGFR3 e Fold chnge 1.0 0.5 P < 0.05 Vegfr3 i EC Vegfr3 Figure S1 Cre ctivtion leds to genetic deletion
DOI: 10.1038/nc2331 PCre;Ros26R 12 h induction 48 h induction Vegfr3 i EC c d ib4 24 h induction VEGFR3 e Fold chnge 1.0 0.5 P < 0.05 Vegfr3 i EC Vegfr3 Figure S1 Cre ctivtion leds to genetic deletion
Oocytes determine cumulus cell lineage in mouse ovarian follicles
 133 Reserh tile Ooytes determine umulus ell linege in mouse ovrin folliles Frniso J. Diz, Kren Wigglesworth nd John J. Eppig* The Jkson Lortory, 6 Min Street, Br Hror, ME 469, USA *Author for orrespondene
133 Reserh tile Ooytes determine umulus ell linege in mouse ovrin folliles Frniso J. Diz, Kren Wigglesworth nd John J. Eppig* The Jkson Lortory, 6 Min Street, Br Hror, ME 469, USA *Author for orrespondene
The soy isoflavone genistein promotes apoptosis in mammary epithelial cells by inducing the tumor suppressor PTEN
 Crinogenesis vol. no.1 pp.1793 183, 5 doi:1.193/rin/gi131 dvne ess pulition My 19, 5 The soy isoflvone genistein promotes poptosis in mmmry epithelil ells y induing the tumor suppressor huvnesh Dve 1,,
Crinogenesis vol. no.1 pp.1793 183, 5 doi:1.193/rin/gi131 dvne ess pulition My 19, 5 The soy isoflvone genistein promotes poptosis in mmmry epithelil ells y induing the tumor suppressor huvnesh Dve 1,,
SUPPLEMENTARY INFORMATION
 TM TM tip link horizontl top connectors 1 leucine-rich (21 %) otoncorin-like 1809 ntigenic peptides B D signl peptide hydrophoic segment proline/threonine-rich (79 %) Supplementry Figure 1. () The outer
TM TM tip link horizontl top connectors 1 leucine-rich (21 %) otoncorin-like 1809 ntigenic peptides B D signl peptide hydrophoic segment proline/threonine-rich (79 %) Supplementry Figure 1. () The outer
Learning to see: experience and attention in primary visual cortex
 2 Nture Pulishing Group http://neurosi.nture.om rtiles Lerning to see: experiene nd ttention in primry visul ortex 2 Nture Pulishing Group http://neurosi.nture.om Roy E. Crist, Wu Li nd Chrles D. Gilert
2 Nture Pulishing Group http://neurosi.nture.om rtiles Lerning to see: experiene nd ttention in primry visul ortex 2 Nture Pulishing Group http://neurosi.nture.om Roy E. Crist, Wu Li nd Chrles D. Gilert
NappHS. rrna. transcript abundance. NappHS relative con W+W 0.8. nicotine [µg mg -1 FM]
![NappHS. rrna. transcript abundance. NappHS relative con W+W 0.8. nicotine [µg mg -1 FM]](/thumbs/92/110177253.jpg "NappHS. rrna. transcript abundance. NappHS relative con W+W 0.8. nicotine [µg mg -1 FM]") (A) W+OS 3 min 6 min con L S L S RNA loding control NppHS rrna (B) (C) 8 1 k NppHS reltive trnscript undnce 6 4.5 *** *** *** *** 3 k. + + + line 1 line (D) nicotine [µg mg -1 FM] 1..8.4. con W+W Supplementl
(A) W+OS 3 min 6 min con L S L S RNA loding control NppHS rrna (B) (C) 8 1 k NppHS reltive trnscript undnce 6 4.5 *** *** *** *** 3 k. + + + line 1 line (D) nicotine [µg mg -1 FM] 1..8.4. con W+W Supplementl
Open Access RESEARCH ARTICLE. Genetics Selection Evolution
 DOI 10.1186/s12711-016-0222-0 Genetis Seletion Evolution RESEARCH ARTICLE Open Aess Comprison of host geneti ftors influening pig response to infetion with two North Amerin isoltes of porine reprodutive
DOI 10.1186/s12711-016-0222-0 Genetis Seletion Evolution RESEARCH ARTICLE Open Aess Comprison of host geneti ftors influening pig response to infetion with two North Amerin isoltes of porine reprodutive
Role of HCP5-miR-139-RUNX1 Feedback Loop in Regulating Malignant Behavior of Glioma Cells
 originl rtile The Amerin Soiety of Gene & Cell Therpy Role of HCP-miR-19-RUNX1 Feedk Loop in Regulting Mlignnt Behvior of Gliom Cells Ho Teng 1,2, Ping Wng, Yixue Xue, Xioi Liu 1,2, Jun M, Heng Ci 1,2,
originl rtile The Amerin Soiety of Gene & Cell Therpy Role of HCP-miR-19-RUNX1 Feedk Loop in Regulting Mlignnt Behvior of Gliom Cells Ho Teng 1,2, Ping Wng, Yixue Xue, Xioi Liu 1,2, Jun M, Heng Ci 1,2,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.18/nture129 ontrol-dna -DNA CD49 Blood Lung e.98 +/-.9.71 +/-.2.29+/-.1 2.9 +/-.6 Bsophils (x1 )/ml 4 Bsophils ( x1 ) d f 45. 22.5 15 75 ontrol-dna ontrol-dna -DNA -DNA
SUPPLEMENTARY INFORMATION doi:1.18/nture129 ontrol-dna -DNA CD49 Blood Lung e.98 +/-.9.71 +/-.2.29+/-.1 2.9 +/-.6 Bsophils (x1 )/ml 4 Bsophils ( x1 ) d f 45. 22.5 15 75 ontrol-dna ontrol-dna -DNA -DNA
SUPPLEMENTARY INFORMATION
 doi:0.08/nture078 RNse VifHA VifHA βctin 6 Cell lyste IP: ntiha MG VifHA VifHA β ctin 6 7 Cell lyste IP: ntiha Supplementry Figure. Effect of RNse nd MG tretment on the Vif interction., RNse tretment does
doi:0.08/nture078 RNse VifHA VifHA βctin 6 Cell lyste IP: ntiha MG VifHA VifHA β ctin 6 7 Cell lyste IP: ntiha Supplementry Figure. Effect of RNse nd MG tretment on the Vif interction., RNse tretment does
Insulin regulation of heart function in aging fruit flies
 4 Nture Pulishing Group http://www.nture.om/nturegenetis Insulin regultion of hert funtion in ging fruit flies Roert J Wessells, Erin Fitzgerld, Jmes R Cypser, Mr Ttr & Rolf Bodmer Insulin-IGF reeptor
4 Nture Pulishing Group http://www.nture.om/nturegenetis Insulin regultion of hert funtion in ging fruit flies Roert J Wessells, Erin Fitzgerld, Jmes R Cypser, Mr Ttr & Rolf Bodmer Insulin-IGF reeptor
Supplementary Figure 1
 Roles of endoplsmic reticulum stress-medited poptosis in -polrized mcrophges during mycocteril infections Supplementry informtion Yun-Ji Lim, Min-Hee Yi, Ji-Ae Choi, Jung-hwn Lee, Ji-Ye Hn, Sung-Hee Jo,
Roles of endoplsmic reticulum stress-medited poptosis in -polrized mcrophges during mycocteril infections Supplementry informtion Yun-Ji Lim, Min-Hee Yi, Ji-Ae Choi, Jung-hwn Lee, Ji-Ye Hn, Sung-Hee Jo,
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/nc2824 Hcn4 Tx5 Mlc2 c Hcn4- ISH d Tx5- ISH e Mlc2-ISH Hcn4-ISH f e Tx5-ISH f -ISH Figure S1 Section in situ hyridistion nlysis of crescent stge mouse emryos (E7.5). () More nterior section
DOI: 10.1038/nc2824 Hcn4 Tx5 Mlc2 c Hcn4- ISH d Tx5- ISH e Mlc2-ISH Hcn4-ISH f e Tx5-ISH f -ISH Figure S1 Section in situ hyridistion nlysis of crescent stge mouse emryos (E7.5). () More nterior section
Effects of exercise training on hepatic steatosis in high fat diet-induced obese mice
 Effets of exerise trining on hepti stetosis in high ft diet-indued oese mie Hyunsik Kng, PhD Sungkyunkwn University Non-Aloholi Ftty Liver Disese (NAFLD) A reversile ondition tht is hrterized y hepti lipid
Effets of exerise trining on hepti stetosis in high ft diet-indued oese mie Hyunsik Kng, PhD Sungkyunkwn University Non-Aloholi Ftty Liver Disese (NAFLD) A reversile ondition tht is hrterized y hepti lipid
DHRS3, a retinal reductase, is differentially regulated by retinoic acid and lipopolysaccharide-induced inflammation in THP-1 cells and rat liver
 Am J Physiol Gstrointest Liver Physiol 33: G78 G88, 212. First pulished July 12, 212; doi:1.112/jpgi.23.212. DHRS3, retinl redutse, is differentilly regulted y retinoi id nd lipopolyshride-indued inflmmtion
Am J Physiol Gstrointest Liver Physiol 33: G78 G88, 212. First pulished July 12, 212; doi:1.112/jpgi.23.212. DHRS3, retinl redutse, is differentilly regulted y retinoi id nd lipopolyshride-indued inflmmtion
TNFa Signaling Exposes Latent Estrogen Receptor Binding Sites to Alter the Breast Cancer Cell Transcriptome
 Artile TNF Signling Exposes Ltent Estrogen Reeptor Binding Sites to Alter the Brest Cner Cell Trnsriptome Grphil Astrt Authors Hetor L. Frno, Anush Ngri, W. Lee Krus Correspondene lee.krus@utsouthwestern.edu
Artile TNF Signling Exposes Ltent Estrogen Reeptor Binding Sites to Alter the Brest Cner Cell Trnsriptome Grphil Astrt Authors Hetor L. Frno, Anush Ngri, W. Lee Krus Correspondene lee.krus@utsouthwestern.edu
SUPPLEMENTARY INFORMATION
 doi:10.1038/nture10754 Supplementry note 1 To ompre our dt with previous studies, we mesured the width of spikes from identified dopminergi neurons nd unidentified neurons from DATCre mie. Previous studies
doi:10.1038/nture10754 Supplementry note 1 To ompre our dt with previous studies, we mesured the width of spikes from identified dopminergi neurons nd unidentified neurons from DATCre mie. Previous studies
Activation of Akt as a Mechanism for Tumor Immune Evasion
 The Amerin Soiety of Gene Therpy originl rtile Ativtion of Akt s Mehnism for Tumor Immune Evsion Kyung Hee Noh 1, Te Heung Kng 1, Jin Hee Kim 1, Sr I Pi 2, Ken Y Lin 3, Chien-Fu Hung 4, T-C Wu 4 7 nd Te
The Amerin Soiety of Gene Therpy originl rtile Ativtion of Akt s Mehnism for Tumor Immune Evsion Kyung Hee Noh 1, Te Heung Kng 1, Jin Hee Kim 1, Sr I Pi 2, Ken Y Lin 3, Chien-Fu Hung 4, T-C Wu 4 7 nd Te
SUPPLEMENTARY INFORMATION
 ~ 5 % ltion > 99 % ltion 25 2 15 5 Gly yemi (mmol/l) 3 7 15 3 Time fter -ell ltion (dys) Survivl (%) & ~ 5 % ltion 75 > 99% ltion 5 25 25 5 (dys) 7.4 d 1.25 lood ph 73 7.3 7.2 7.1 7. 7d ody weight h nge
~ 5 % ltion > 99 % ltion 25 2 15 5 Gly yemi (mmol/l) 3 7 15 3 Time fter -ell ltion (dys) Survivl (%) & ~ 5 % ltion 75 > 99% ltion 5 25 25 5 (dys) 7.4 d 1.25 lood ph 73 7.3 7.2 7.1 7. 7d ody weight h nge
Technical Report GIT-CERCS The Sleepy Keeper Approach: Methodology, Layout and Power Results for a 4-bit Adder
 Tehnil Report GIT-CERCS-06-03 The Sleepy Keeper Approh: Methodology, Lyout nd Power Results for 4-it Adder Se Hun Kim, Vinent J. Mooney III nd Jun Cheol Prk Center for Reserh on Emedded Systems nd Tehnology
Tehnil Report GIT-CERCS-06-03 The Sleepy Keeper Approh: Methodology, Lyout nd Power Results for 4-it Adder Se Hun Kim, Vinent J. Mooney III nd Jun Cheol Prk Center for Reserh on Emedded Systems nd Tehnology