Supporting Online Material for
|
|
|
- Moris Pierce
- 5 years ago
- Views:
Transcription
P 3 and PI(4,5)P 2 Lipids Target Proteins with Polybasic Clusters to the Plasma Membrane Won Do Heo, Takanari Inoue, Wei Sun")
1 Supporting Online Material for PI(3,4,5)P 3 and PI(4,5)P 2 Lipids Target Proteins with Polybasic Clusters to the Plasma Membrane Won Do Heo, Takanari Inoue, Wei Sun Park, Man Lyang Kim, Byung Ouk Park, Thomas J. Wandless, Tobias Meyer *To whom correspondence should be addressed. tobias1@stanford.edu This PDF file includes Materials and Methods Figs. S1 to S13 References Published 9 November 2006 on Science Express DOI: /science
2 Online Material Materials and Methods Cell culture and transfection. NIH3T3 and HeLa cells were purchased from ATCC. Cells were cultured in DMEM containing 10% FBS in 10% CO 2 and were split every three days by a 1/5 dilution at about 90% confluency. For confocal microscopy, 2.0 x10 4 cells were plated in each well of Lab-Tek 4-well chambered coverglasses (Nunc; #155383) coated with poly-d-lysine (0.1mg/ml) and cultured for 24 hrs prior to transfection. Cells were transfected with 0.25 μg/well of cdna using Lipofectamine 2000 (Invitrogen). Cloning of human small GTPases and generation of mutant constructs. A previously described library of human small GTPases (wildtype and constitutively active forms) (S1) was supplemented with twenty five additional human small GTPases using RT-PCR cloning with gene-specific primers containing attb sequences to create entry clones for the Gateway cloning system (Invitrogen). Wildtype and constitutively active forms were made for the 25 human small GTPases. Expression constructs were made by LR clonase (Invitrogen) to transfer the coding regions into CFP expression vectors. ARF subfamily members were N-terminally tagged and other subfamily members were C-terminally tagged with CFP. Tail mutants were designed using complementary oligonucleotides with restriction enzyme sites at both ends. p85 DN construct was kindly obtained from Dr. Daniel Storm (University of Washington). KRas tail was kindly provided by Dr. Marc Fivaz (Stanford).
3 Microscopy. All images were taken in live cells using Nipkow confocal and Zeiss epifluorescence microscopes (40x or 63x magnification). Images were analyzed using MetaMorph software (Molecular Devices). The detailed experimental protocol for the activation of the Inp54p PI(4,5)P 2 phosphatase is described in an accompanying paper (S2). In vitro lipid binding assay. YFP-PLCδ-PH and YFP-conjugated to the respective polybasic peptides were transfected into 293F cells in 293 Freestyle media. As a negative control, YFP-HRas tail was expressed. 48hrs after transfection, the cells were collected and total proteins were extracted from the cells with a lysis buffer (50mM Tris, ph 8.0; 10mM EDTA, ph8.0; 100mM NaCl; 0.5% Triton X-100; protease inhibitor cocktail). Each PIP Strip was preincubated for 1hr in 3% fatty-acid free BSA in TBS-T. Pre-incubated PIP strips were incubated with cell lysate in 3% BSA in TBS-T with gentle shaking overnight. After washing three times with 1% BSA in TBS-T, the strips were incubated with anti-gfp antibody (1:1000) in 1% BSA in TBS-T for an hour. After washing three times with 1% BSA in TBS-T, the strips were incubated with anti-rabbit IgG conjugated with alkaline phosphatase (Invitrogen; G-21079). The signals were detected with a NBT/BCIP detection kit (Biorad; ) after washing twice in 3% BSA in TBS-T, once in TBS-T, and once in 0.1M Tris, ph 9.5 for 5 min.
4 Supplementary Figure Legends Fig. S1. Subcellular localization of 125 human CFP-conjugated CA small GTPases in NIH3T3 and HeLa cells. Fluorescent images were acquired using a Nipkow confocal microscope (40x or 63x objective) and processed using MetaMorph software (Molecular Devices). The localization of each small GTPase was visually compared in at least five independent images. One representative image was chosen from each cell line, NIH3T3 (left) and HeLa (right). Prenylation consensus sequences are indicated by a box, palmitoylation consensus sequences are underlined, basic amino acid residues are marked in red, hydrophobic amino acid residues are marked in blue, and myristolylation consensus sequences are marked in green. Scale bars, 10 μm. Fig. S2. Differential localization of GDP and GTP bound ARF6. Arf6 was an outlier without a recognizable putative PM targeting domain. We tested whether PM targeting of Arf6 is regulated by a process that requires ARF6 GTP loading. CFP-tagged dominant negative ARF6 (T27N; GDP bound) was cotransfected in NIH3T3 cells with YFP-tagged constitutively active ARF6 (Q67L; GTP-bound) and the subcellular distribution of the two mutants was compared in the same cell using confocal imaging. Only the GTP bound Arf6 (Q67L) localized to the PM. The insets show a magnified part of the images and the arrow points to the PM. Scale bars, 10 μm. Fig. S3. LY (LY30), a kinase inactive analog of LY29, does not trigger the removal of Rin tail from the PM. (A) Structures of LY (LY29) and LY (LY30). LY30 contains a single atom substitution in the morpholine ring. LY30 has been reported to have similar chemical properties as LY29 but without a measurable inhibitory activity for PI3K (S3). (B) Effect of LY29 (50μM) and LY30 (50μM) on PDGF-induced
5 YFP-Akt-PH domain translocation. NIH3T3 cells were transfected with YFP-Akt-PH domains and cells were stimulated with 5nM PDGF for 3 min, followed by addition of LY29 or LY30. LY30 did not inhibit Akt-PH domain translocation or membrane ruffling induced by PDGF. (C) Effects of LY30, LY29, and irap on Rin tail localization in the same cell. LY30 was first added to the cells, followed by LY29 and irap. LY30 (50μM) did not trigger the removal of Rin tail from the PM, while LY29 (50μM) partially removed the Rin tail. Activation of CF-Inp by irap removed most of the remaining Rin tail from the PM. Scale bars, 10 μm. Fig. S4. Depletion of both PI(4,5)P 2 and PI(3,4,5)P 3 are required for the PM dissociation of MARCKS ED and Rin tail in HeLa cells. (A) Depletion of PI(4,5)P 2 by addition of irap triggered complete dissociation of the PI(4,5)P 2 biosensor PLCδ-PH and only a small dissociation of MARCKS ED and Rin tail from the PM of HeLa cells. Joint depletion of PI(4,5)P 2 and PI(3,4,5)P 3 by addition of irap and LY29 (irap + LY29) triggered a near complete dissociation of MARCKS ED and Rin tail from the PM. (B) Quantitative analysis of the PM dissociation of PLCδ-PH domain, MARCKS ED, and Rin tail. PLCδ-PH domain, MARCKS ED, or Rin tail were cotransfected with CF-Inp and Lyn- FRB in HeLa cells. The dissociation index was analyzed using MetaMorph software as described in Fig. 2B. Scale bars, 10 μm. Fig. S5. Kinetics of PM dissociation of different proteins with polybasic clusters. (A) Kinetics of the PM dissociation of Rit and Rin tails following the depletion of PI(4,5)P 2 and PI(3,4,5)P 3. (B) Kinetics of the PM dissociation of HRas tail (as a control), Arl7, Rab35, and DN RhoE (dominant negative version to minimize targeting by Rho-GTP binding proteins) following the depletion of PI(4,5)P 2 and PI(3,4,5)P 3.
6 Fig. S6. Control experiments monitoring Rin tail dissociation for different protocols of PI(4,5)P 2 and PI(3,4,5) depletion. (A) An alternative PI3K inhibitor, wortmannin (100nM), was added to the cells for 3min, followed by the addition of irap for 3 min and the change in localization of MARCKS ED and Rin tail were tracked over time. This protocol induced a similar PM dissociation as shown in Fig. 2 for the combination of irap and LY29. (B) Image series showing that LY29 alone had only a small effect on Rin tail PM dissociation while the subsequent addition of irap triggered a near complete PM dissociation of Rin tail. Scale bars, 10 μm. Fig. S7. Control experiments using expression of a peptide based inhibitor of PI3K, p85 DN. (A) Control experiments showing that PDGF-induced translocation of Akt-PH domain, a biosensor for PI(3,4,5)P 3, was nearly completely suppressed in cells expressing the PI3K inhibitor construct, p85 DN. (B) Cells expressing p85 DN showed a near complete removal of Rin tail from the PM when PI(4,5)P 2 concentration was depleted using CF-Inp activation. Together with fig. S6, this further supports the model that PI3K inhibition has to be combined with PI(4,5)P 2 depletion for polybasic clusters to dissociate from the PM. Scale bars, 10 μm. Fig. S8. PI(3,4,5)P 3 synthesis at the PM enhances PM localization of Rin tail. (A) Overexpression of a constitutively active PI3K (p110-caax) enhanced PI(3,4,5)P 3 synthesis at the PM and also enhanced the PM localization of Rin tail. CFP-Akt-PH domain and YFP-Rin tail were cotransfected in NIH3T3 cells together with p110-caax. In serum starved cells, p110-caax expression strongly increased Akt-PH domain localization at the PM. YFP-Rin tail also showed increased PM localization as evidenced by the recruitment of most nuclear Rin to the PM. There was also an
7 apparent co-localization of YFP-Rin with CFP-Akt-PH domain in lamellipodia. The presence of multiple lamellipodia correlated with p110-caax expression and Akt-PH localization to the PM. (B) Reversibility of the PI3K-dependent PM translocation of MARCKS ED and Rin tail. YFP-MARCKS ED or YFP-Rin tail was cotransfected with CFP-Akt-PH domain, together with p110-caax. To test for reversibility of PM targeting, cells were incubated with LY29 for 3min, followed by the removal of LY29 (in the presence of p110-caax). YFP-MARCKS ED and YFP-Rin tail partially dissociated from the PM by inhibition of p110-caax with LY29 (3min). After washout of LY29, PM targeting of YFP-MARCKS ED and YFP-Rin tail recovered within 1 min. The changes in the PM dissociation and PM binding of YFP-MARCKS ED or YFP-Rin tail coincided with the dissociation and PM binding of YFP-Akt-PH domains. Scale bars, 10 μm. Fig. S9. Effects of CF-Inp-activation and PDGF-stimulation on PI(3,4,5)P 3 production. (A) Activation of CF-Inp lowered not only PI(4,5)P 2 but also mediated a partial reduction in PI(3,4,5)P 3 as was evident from a partial dissociation of YFP-Akt-PH. Pre-stimulation with PDGF was used for better visualization of the effect of CF-Inp activation on PI(3,4,5)P 3. (B) PDGF still increases PI(3,4,5)P 3 concentration (as evidenced by YFP-Akt-PH translocation) in cells in which CF-Inp has been activated before to lower PI(4,5)P 2 concentration. Together, these control experiments suggest that there is still sufficient PI(4,5)P 2 substrate in cells with activated CF-Inp for significant PI3K-mediated PI(3,4,5)P 3 production. Fig. S10. Polybasic clusters with only 4 positively charged amino acids still dissociate from the PM following PI(4,5)P 2 and PI(3,4,5)P 3 depletion. (A) Rho E, which has only 4 positively charged amino acids in the 20 amino acid tail region, dissociated from the PM after depletion of PI(4,5)P 2 and PI(3,4,5)P 3. (B) Rho D, which
8 has also 4 positively charged amino acids, also dissociated from the PM after depletion of PI(4,5)P 2 and PI(3,4,5)P 3. Scale bars, 10 μm. Fig. S11. Pharmacological inhibition of calmodulin and protein kinase C does not alter Rin-tail PM localization and dissociation. (A) Rin-tail PM localization and dissociation by PI(4,5)P 2 and PI(3,4,5)P 3 is not affected by the calmodulin inhibitor W-7 (10μΜ). This supports the argument that increases in Ca 2+ concentration and Ca 2+ /CaM binding to the polybasic clusters are not responsible for the PM dissociation following PI(4,5)P 2 and PI(3,4,5)P 3 depletion. (B) Rin-tail PM localization and dissociation by PI(4,5)P 2 and PI(3,4,5)P 3 is not affected by the protein kinase C (PKC) inhibitor Ro This argues against the possibility that hydrolysis of PI(4,5)P 2 and inhibition of PI(3,4,5)P 3 may indirectly activate PKC and thereby phosphorylate polybasic clusters and cause their PM dissociation. A lack of such a role of protein phosphorylation is also supported by the finding that Rac1 (Fig. 4D) still dissociates from the PM even though it lacks a consensus phosphorylation site. Fig. S12. Polybasic peptides strongly bind phosphoinositides in vitro. (A) Schematic view of the lipid positions of 15 different lipids on a phosphoinositide lipid strip (Echelon; P-6001). These PIP strips are 2 x 6 cm hydrophobic membranes that have been spotted with 15 different biologically active lipids. (B) Proteins with polybasic clusters bind phosphoinositides and show selectivity for PI(3,4,5)P 3 over PI(3,4,5)P 3. The images show binding of PLCδ-PH domain and polybasic peptides to different phosphoinositides on the PIP Strip. YFP-PLCδ-PH domain and YFP-conjugated polybasic peptides were expressed in 293F cells and total proteins were extracted. Total proteins were incubated with the PIP Strips and detected with anti-gfp antibody as described in Materials and Methods.
9 Fig. S13. Hydrophobic amino acids are necessary for the PM targeting of the small GTPases GEM and RAD. Identification of essential hydrophobic amino acids for PM targeting in the polybasic regions of the small GTPases GEM and RAD. (A) Two hydrophobic amino acid residues, Phe and Trp, were replaced in GEM with the less hydrophobic amino acid residue Ala. A marked loss in PM targeting was observed for this GEM mutant. (B) The hydrophobic amino acid residue Phe in the RAD tail was replaced with Ala which also led to a loss in PM targeting of this RAD mutant. Scale bars, 10 μm.
10 Supplementary References S1. W. D. Heo, T. Meyer, Cell 113, 315 (2003). S2. B. C. Suh, T. Inoue, T. Meyer, B. Hille, Science (2006), this issue. S3. T. W. Poh, S. Pervaiz, Cancer Res 65, 6264 (2005).















11 Heo et al., Figure S1 Ras subfamily NIH3T3 HeLa NIH3T3 HeLa HRAS (NP_ ) KLNPPDESGPGCMSCKCVLS NRAS (NP_ ) KLNSSDDGTQGCMGLPCVVM KRAS4A (NP_ ) KISKEEKTPGCVKIKKCIIM KRAS4B (NP_ ) KMSKDGKKKKKKSKTKCVIM RRAS (NP_006261) PPSPPSAPRKKGGGCPCVLL RRAS2 (NP_036382) PSPEPTRKEKDKKGCHCVIF RRAS3 [MRAS] (NP_036382) KTKWRGDRATGTHKLQCVIL RIT [RIT1] (NP_008843) KNSVWKRLKSPFRKKKDSVT

















12 NIH3T3 HeLa NIH3T3 HeLa RIN [RIT2] (NP_002921) KDSLWKKLKGSLKKKRENMT RHEB (NP_005605) LEAEKMDGAASQGKSSCSVM RALA (NP_005393) NGKKKRKSLAKRIRERCCIL RALB (NP_002872) KNGKKSSKNKKSFKERCCLL RAP1A (NP_002875) INRKTPVEKKKPKKKSCLLL RAP1B (NP_ ) INRKTPVPGKARKKSSCQLL RAP2A (NP_066361) MNYAAQPDKDDPCCSACNIQ RAP2B (NP_116235) MNYAAQSNGDEGCCSACVIL RRP22 (NP_006468) PAHPALRLQGALHPARCSLM NKIRAS1 (NP_065078) SKSSFPLPGRKNKGNSNSEN














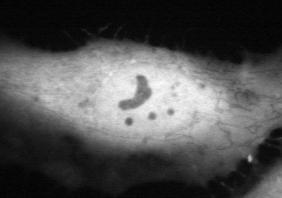




13 NIH3T3 HeLa NIH3T3 HeLa NKIRAS2 (NP_060065) QSKSAFPLSRKNKGSGSLDG RERG (NP_116307) RSSTTHVKQAINKMLTKISS RRAGA (NP_006561) RKHFEKLERVDGPKHSLLMR RRAGB (NP_006055) RKHFEKLERVDGPKQCLLMR RRAGC (NP_071440) QTSASSLKALTHNGTPRNAI RRAGD (NP_067067) QNRLQKKKRATPNGTPRVLL GEM [GEM1] (NP_005252) RKARRFWGKIVAKNNKNMAFKLKSKSCHDLSVL RAD [RRAD] (NP_004156) KKAKRFLGRIVARNSRKMAFRAKSKSCHDLSVL REM (NP_054731) QRARRFLARLTARSARRRALKARSKSCHNLAVL RASL12 (NP_057647) RAQSKRKAPTLTLLKGFKIF









14 NIH3T3 HeLa NIH3T3 HeLa RASL (NP_079006) GSKSMAKLINNVFGKRRKSV DIRAS3 [ARHI] (NP_004666) EKKSQMPNTTEKLLDKCIIM RASD1 [AGS1] (NP_057168) IREKASAGSQAKDKERCVIS RASD2 (NP_055125) IKAKVLREGQARERDKCTIQ DIRAS2 (NP_060064) GKKSKQQKRKEKLKGKCVIM RASL11B (NP_076429) DLKRRFKQALSAKVRTVTSV















15 Rho subfamily NIH3T3 HeLa NIH3T3 HeLa RHOA (NP_001655) ATRAALQARRGKKKSGCLVL RHOB (NP_004031) AALQKRYGSQNGCINCCKVL RHOC (NP_786886) ATRAGLQVRKNKRRRGCPIL RHO7 [RND2] (NP_005431) PDRGNEGEIHKDRAKSCNLM RHO6 [RND1] (NP_055285) RSELISSTFKKEKAKSCSIM RHOE [RND3] (NP_005159) ELSAVATDLRKDKAKSCTVM RHOH (NP_004301) NQARRRNRRRLFSINECKIF RIF [RHOF] (NP_061907) ALSALKKAQRQKKRRLCLLL
















16 NIH3T3 HeLa NIH3T3 HeLa TCL [RHOJ] (NP_065714) FHPKKKKKRCSEGHSCCSII TC10 [RHOQ] (NP_036381) KKHTVKKRIGSRCINCCLIT CDC42; isoform1 (NP_001782) AILAALEPPEPKKSRRCVLL RHOD (NP_055393) LSSRGRNFWRRITQGFCVVT CDC42H; isoform2 (NP_426359) AILAALEPPETQPKRKCCIF RAC1 (NP_008839) IRAVLCPPPVKKRKRKCLLL RAC2 (NP_002863) IRAVLCPQPTRQQKRACSLL RAC3 (NP_005043) IRAVLCPPPVKKPGKKCTVF RHOG (NP_001656) AVRAVLNPTPIKRGRSCILL














17 Rab subfamily NIH3T3 HeLa NIH3T3 HeLa RAB1A (NP_004152) EKSNVKIQSTPVKQSGGGCC RAB1B (NP_112243) ERPNLKIDSTPVKPAGGGCC RAB2 (NM_002865) ATNATHAGNQGGQQAGGGCC RAB3A (NP_002857) AKQGPQLSDQQVPPHQDCAC RAB3B (NP_002858) SSKNTRLSDTPPLLQQNCSC RAB3D (NP_004274) NGKGPAVGDAPAPQPSSCSC RAB4A (NP_004569) LRQLRSPRRTQAPNAQECGC RAB4B (NP_057238) LRQLRQPRSAQAVAPQPCGC







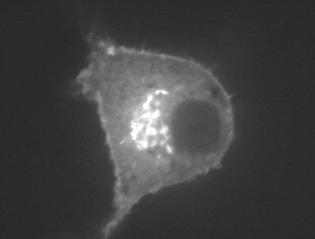











18 NIH3T3 HeLa NIH3T3 HeLa RAB5A (NP_004153) GGGVDLTEPTQPTRNQCCSN RAB5B (NP_002859) SRGVDLHEQSQQNKSQCCSN RAB5C (NP_004574) NRGVDLQENNPASRSQCCSN RAB6A (NP_002860) IDIKLEKPQEQPVSEGGCSC RAB6B (NP_057661) IDIKLDKPQEPPASEGGCSC RAB6C (NP_115520) IDIKLEKPQEQPVSEGGCSC RAB7 (NP_004628) EPIKLDKNDRAKASAESCSC RAB7L1 (NP_003920) LSTQGDYINLQTKSSSWSCC RAB8 [RAB8A] (NP_005361) GVKITPDQQKRSSFFRCVLL RAB8B (NP_057614) PVKITENRSKKTSFFRCSLL



















19 NIH3T3 HeLa NIH3T3 HeLa RAB9A (NP_004242) LIQTDTVNLHRKPKPSSSCC RAB9B (NP_057454) CMLGHTIDLNSGSKAGSSCC RAB10 (NP_057215) SENVDISSGGGVTGWKSKCC RAB11A (NP_004654) PIHVPPTTENKPKVQCCQNI RAB11B (NP_004209) SVPPTTDGQKPNKLQCCQNL RAB13 (NP_002861) PPSTDLKTCDKKNTNKCSLG RAB14 (NP_057406) APQGGRLTSEPQPQREGCGC RAB17 (NP_071894) GDAAVALNKGPARQAKCCAH RAB18 (NP_067075) LSHREEGQGGGACGGYCSVL RAB20 (NP_060287) HTVDISSHKPPKRTRSGCCA



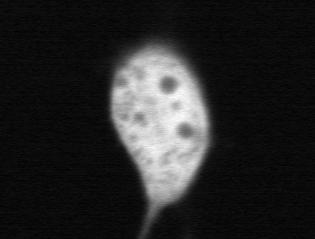















20 NIH3T3 HeLa NIH3T3 HeLa RAB21 (NP_055814) QIIDDEPQAQTSGGGCCSSG RAB22A (NP_065724) SGGKGFKLRRQPSEPKRSCC RAB22B [RAB31] (NP_006859) GNNGTIKVEKPTMQASRRCC RAB23 (NP_057361) RPNKQRTKKNRNPFSSCSIP RAB24 (NP_570137) DKGVDLGQKPNPYFYSCCHH RAB25 (P57735) SAQAGQEPGPGEKRACCISL RAB26 (NP_055168) RFRLHDYVKREGRGASCCRP RAB27A (NP_004571) NGHASTDQLSEEKEKGACGC RAB27B (NP_004154) NGGNSGNLDGEKPPEKKCIC RAB28 (NP_004240) YPEEENQHTTSTQSRICSVQ








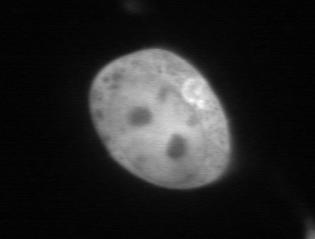










21 NIH3T3 HeLa NIH3T3 HeLa RAB30 (NP_055303) SSPLPGEGKSISYLTCCNFN RAB32 (NP_006825) VDKIKLDQETLRAENKSQCC RAB33A (NP_004785) GKVQKLEFPQEANSKTSCPC RAB33B (NP_112586) PPDNGIILKPEPKPAMTCWC RAB34 (NP_112586) NSDDSNLYLTASKKKPTCCP RAB35 (NP_006852) QQQNDVVKLTKNSKRKKRCC RAB38 (NP_071732) VVKPHLTSTKVASCSGCAKS RAB39 (NP_059986) NSDDSNLYLTASKKKPTCCP RAB39B (NP_741995) VPNVVHSSEEVVKSERRCLC RAB40A (NP_543155) IVCPPQSPPKNCTRNSCKIS











22 NIH3T3 HeLa NIH3T3 HeLa RAB40B (NP_006813) LVRPPQSPPKNCTRNSCKIS RAB40C (NP_066991) SIRPPQSPPQNCSRSNCKIS RABL2A (NP_009013) DQEQSSSIETPSEEVASPHS RABL2B (NP_009012) DQEQSSSIETPSEEAASPHS RABL4 (NP_006851) LAKQFHQLYREKVEVFRALA RBJ (NP_057628) SEDAFKAVVNARTALLKNIK













23 ARF subfamily NIH3T3 HeLa NIH3T3 HeLa ARF1 (NP_001649) MGNIFANLFK---SGDGLYEGLDWLSNQLRNQK ARF3 (NP_001650) MGNIFGNLLK---SGDGLYEGLDWLANQLKNKK ARF4 (NP_001651) MGLTISSLFS---TQGTGLYEGLDWLSNELSKR ARF5 (NP_001653) MGLTVSALFS---TQGTGLYDGLDWLSHELSKR ARF6 (NP_001654) MGKVLSKIFG---ATSGDGLYEGLTWLTSNYKS ARL1 (NP_001168) MGGFFSSIFS---KGTGLDEAMEWLVETLKSRQ ARL2 (NP_001658) MGLLTILKKM---LLPGIDWLLDDISSRIFTAD ARL3 (NP_004302) MGLLSILRKL---GEGVQDGMNWVCKNVNAKKK














24 NIH3T3 HeLa NIH3T3 HeLa ARL4 [ARL4A] (NP_005729) MGNGLSDQTS---KLHDMIIKRRKMLRQQKKKR ARL6 [ARL4D] (NP_001652) MGNHLTEMAP---RLYEMILKRKKAARGGKKRR ARL7 [ARL4C] (NP_005728) MGNISSNISA---DKLYEMILKRRKSLKQKKKR ARL5A (NP_036229) MGILFTRIWR---LTGEGLCQGLEWMMSRLKIR ARL6 (NP_115522) MGLLDRLSVL---GEGLQEGVDWLQDQIQTVKT ARL8B [ARL10C] (NP_060654) MAAIKSVVVG---KDNIDITLQWLIQHSKSRRS ARL14 (NP_079323) MGSLGSKNPQ---GFVKSHMKSRGDTLAFFKQN ARL15 (NP_061960) MSDLRITEAF---DSFSQLINLLEEKDHEAVRM ARFRP1 (NP_003215) MYTLLSGLYK---WMVKCVVRNVHRPPRQRDIT


")
")


25 Other subfamily members NIH3T3 HeLa NIH3T3 HeLa SAR1A (NP_064535) SAR1B (NP_ ) SVLKRQGYGEGFRWLSQYID SVLKRQGYGEGFRWMAQYID RAN (NP_006316) YEHDLEVAQTTALPDEDDDL

26 Heo et al., Figure S2 ARF6 T27N ARF6 Q67L







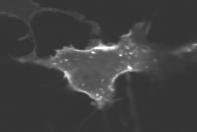
27 Heo et al., Figure S3 A B Akt-PH - LY29 - LY30 Akt-PH + LY29 + LY30 + PDGF + PDGF C Before LY30 LY29 Inp54 Rin tail



28 Heo et al., Figure S4 A PLCδ-PH MARCKS ED Rin tail Before After irap irap irap + LY29 irap irap + LY29 B PM dissociation index PLCδ-PH H-Ras tail MARCKS ED Rin tail irap LY
29 Heo et al., Figure S5 A PM dissociation index 2.5 Rin tail Rit tail irap+ly29 5 min B PM dissociation index RhoET37N Rab35 Arl7 HRastail 1 5 min irap+ly29 +












30 Heo et al., Figure S6 A Before Wortmannin Wortmannin + irap CF-Inp MARCKS ED CF-Inp Rin tail B Before LY29 LY29 + irap CF-Inp Rin tail






31 Heo et al., Figure S7 A Before PDGF Akt-PH Akt-PH + p85 DN B Before +irap CF-Inp Rin tail + p85 DN




32 Heo et al., Figure S8 A CFP-Akt-PH YFP-Rin tail Control p110-caax B Before LY29 for 3min LY29 washed out CFP-Akt-PH YFP-MARCKS ED + p110-caax CFP-Akt-PH YFP-Rin tail + p110-caax




")
")

33 Heo et al., Figure S9 A Before irap 3 min CF-Inp Akt-PH PDGF (3min) PDGF (6min) B Before PDGF 3min CF-Inp Akt-PH irap (3min) irap (6min)
34 Heo et al., Figure S10 A -1min irap + LY29 3min RhoE T37N B RhoD T31N -1min irap + LY29 3min








35 Heo et al., Figure S11 A CF-Inp irap + LY29 B CF-Inp irap + LY29-1min 3min -1min 3min YFP-Rin tail YFP-Rin tail 10μM W7 (Calmodulin inhibitor) 100nM Ro (PKC inhibitor)

 PI(3)P PI(4)P PI(5)P")
P 2 PI(3,5)P")

36 Heo et al., Figure S12 A B Lysophosphatidic acid Lysophosphatidylcholine Phosphatidylinositol (PI) PI(3)P PI(4)P PI(5)P Phosphatidylethanolamine Phosphatidylcholine Sphingosine-1-phosphate PI(3,4)P 2 PI(3,5)P 2 PI(4,5)P 2 PI(3,4,5)P 3 Phosphatidic acid Phosphatidylserine Blank PLC-δ-PH HRAS tail KRAS tail MARCKS ED Rin tail Rit tail PI(4,5)P 2 PI(3,4,5)P 3




37 Heo et al., Figure S13 A GEM GEM F269A W270A B RAD RAD F281A GEM RAD REM RKARRFWGKIVAKNNKNMAFKLKSKSCHDLSVL KKAKRFLGRIVARNSRKMAFRAKSKSCHDLSVL QRARRFLARLTARSARRRALKARSKSCHNLAVL
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Nature Biotechnology: doi: /nbt.3828
 Supplementary Figure 1 Development of a FRET-based MCS. (a) Linker and MA2 modification are indicated by single letter amino acid code. indicates deletion of amino acids and N or C indicate the terminus
Supplementary Figure 1 Development of a FRET-based MCS. (a) Linker and MA2 modification are indicated by single letter amino acid code. indicates deletion of amino acids and N or C indicate the terminus
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/7/334/rs4/dc1 Supplementary Materials for Rapidly rendering cells phagocytic through a cell surface display technique and concurrent Rac activation Hiroki Onuma,
www.sciencesignaling.org/cgi/content/full/7/334/rs4/dc1 Supplementary Materials for Rapidly rendering cells phagocytic through a cell surface display technique and concurrent Rac activation Hiroki Onuma,
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway
 Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Use of a camp BRET Sensor to Characterize a Novel Regulation of camp by the Sphingosine-1-phosphate/G 13 Pathway SUPPLEMENTAL DATA Characterization of the CAMYEL sensor and calculation of intracellular
Signal Transduction Cascades
 Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
Signal Transduction Cascades Contents of this page: Kinases & phosphatases Protein Kinase A (camp-dependent protein kinase) G-protein signal cascade Structure of G-proteins Small GTP-binding proteins,
nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering
 nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering Toru Komatsu, Igor Kukelyansky, J Michael McCaffery, Tasuku Ueno, Lidenys C Varela & Takanari Inoue Supplementary
nature methods Organelle-specific, rapid induction of molecular activities and membrane tethering Toru Komatsu, Igor Kukelyansky, J Michael McCaffery, Tasuku Ueno, Lidenys C Varela & Takanari Inoue Supplementary
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Validation & Assay Performance Summary
 Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Computational Systems Biology: Biology X
 Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#10:(November-22-2010) Cancer and Signals 1 1 Micro-Environment Story How does the rest of our body influences the cancer cell?
Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#10:(November-22-2010) Cancer and Signals 1 1 Micro-Environment Story How does the rest of our body influences the cancer cell?
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system
 Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Cell Biology Lecture 9 Notes Basic Principles of cell signaling and GPCR system Basic Elements of cell signaling: Signal or signaling molecule (ligand, first messenger) o Small molecules (epinephrine,
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human
 and HeLa (bottom) cells expressing for 24 h human") Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
Fig. S1. Subcellular localization of overexpressed LPP3wt-GFP in COS-7 and HeLa cells. Cos7 (top) and HeLa (bottom) cells expressing for 24 h human LPP3wt-GFP, fixed and stained for GM130 (A) or Golgi97
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
ab SREBP-2 Translocation Assay Kit (Cell-Based)
") ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Phosphoinositides Regulate Ciliary Protein Trafficking to Modulate Hedgehog Signaling
 Developmental Cell Supplemental Information Phosphoinositides Regulate Ciliary Protein Trafficking to Modulate Hedgehog Signaling Francesc R. Garcia-Gonzalo, Siew Cheng Phua, Elle C. Roberson, Galo Garcia
Developmental Cell Supplemental Information Phosphoinositides Regulate Ciliary Protein Trafficking to Modulate Hedgehog Signaling Francesc R. Garcia-Gonzalo, Siew Cheng Phua, Elle C. Roberson, Galo Garcia
Supplementary Figure S1 (a) (b)
 (b)") Supplementary Figure S1: IC87114 does not affect basal Ca 2+ level nor nicotineinduced Ca 2+ influx. (a) Bovine chromaffin cells were loaded with Fluo-4AM (1 μm) in buffer A containing 0.02% of pluronic
Supplementary Figure S1: IC87114 does not affect basal Ca 2+ level nor nicotineinduced Ca 2+ influx. (a) Bovine chromaffin cells were loaded with Fluo-4AM (1 μm) in buffer A containing 0.02% of pluronic
Supplemental information contains 7 movies and 4 supplemental Figures
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplemental information contains 7 movies and 4 supplemental Figures Movies: Movie 1. Single virus tracking of A4-mCherry-WR MV
Validation & Assay Performance Summary
 Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Beck et al., http://www.jcb.org/cgi/content/full/jcb.201011027/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Membrane binding of His-tagged proteins to Ni-liposomes.
Supplemental material Beck et al., http://www.jcb.org/cgi/content/full/jcb.201011027/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Membrane binding of His-tagged proteins to Ni-liposomes.
Nature Immunology doi: /ni.3268
 Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Nature Immunology: doi: /ni.3866
 Nature Immunology: doi:10.1038/ni.3866 Supplementary Figure 1 The effect of TIPE2 on chemotaxis. a, The expression of TIPE2 in dhl-60c, dhl-60t, TIPE2-expressing and 15/16Q-expressing dhl-60t neutrophils
Nature Immunology: doi:10.1038/ni.3866 Supplementary Figure 1 The effect of TIPE2 on chemotaxis. a, The expression of TIPE2 in dhl-60c, dhl-60t, TIPE2-expressing and 15/16Q-expressing dhl-60t neutrophils
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
File name: Supplementary Information Description: Supplementary figures and supplementary tables. File name: Peer review file Description:
 File name: Supplementary Information Description: Supplementary figures and supplementary tables. File name: Peer review file Description: Supplementary Figure 1. Schematic of Ras biochemical coupled assay.
File name: Supplementary Information Description: Supplementary figures and supplementary tables. File name: Peer review file Description: Supplementary Figure 1. Schematic of Ras biochemical coupled assay.
Supplemental Information. Autophagy in Oncogenic K-Ras. Promotes Basal Extrusion. of Epithelial Cells by Degrading S1P. Current Biology, Volume 24
 Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric*
 SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Name: Multiple choice questions. Pick the BEST answer (2 pts ea)
") Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
Exam 1 202 Oct. 5, 1999 Multiple choice questions. Pick the BEST answer (2 pts ea) 1. The lipids of a red blood cell membrane are all a. phospholipids b. amphipathic c. glycolipids d. unsaturated 2. The
Supplemental information
 Supplemental information PI(3)K p11δ controls the sucellular compartmentalization of TLR4 signaling and protects from endotoxic shock Ezra Aksoy, Salma Taoui, David Torres, Sandrine Delauve, Aderrahman
Supplemental information PI(3)K p11δ controls the sucellular compartmentalization of TLR4 signaling and protects from endotoxic shock Ezra Aksoy, Salma Taoui, David Torres, Sandrine Delauve, Aderrahman
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
www.sciencesignaling.org/cgi/content/full/6/283/ra57/dc1 Supplementary Materials for JNK3 Couples the Neuronal Stress Response to Inhibition of Secretory Trafficking Guang Yang,* Xun Zhou, Jingyan Zhu,
Supplementary Material and Methods
 Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
Online Supplement Kockx et al, Secretion of Apolipoprotein E from Macrophages 1 Supplementary Material and Methods Cloning of ApoE-GFP Full-length human apoe3 cdna (pcdna3.1/zeo + -apoe) was kindly provided
(A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a
 RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a") Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Structural basis of recognition of farnesylated and methylated KRAS4b by PDEδ
 Structural basis of recognition of farnesylated and methylated KRAS4b by PDEδ Srisathiyanarayanan Dharmaiah a, Lakshman Bindu a, Timothy H. Tran a, William K. Gillette a, Peter H. Frank a, Rodolfo Ghirlando
Structural basis of recognition of farnesylated and methylated KRAS4b by PDEδ Srisathiyanarayanan Dharmaiah a, Lakshman Bindu a, Timothy H. Tran a, William K. Gillette a, Peter H. Frank a, Rodolfo Ghirlando
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms. Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh
 A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh Abstract Phosphoinositide 3-kinases (PI 3-kinase) consist of a family
A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh Abstract Phosphoinositide 3-kinases (PI 3-kinase) consist of a family
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY FIGURE LEGENDS
 SUPPLEMENTARY FIGURE LEGENDS Supplemental FIG. 1. Localization of myosin Vb in cultured neurons varies with maturation stage. A and B, localization of myosin Vb in cultured hippocampal neurons. A, in DIV
SUPPLEMENTARY FIGURE LEGENDS Supplemental FIG. 1. Localization of myosin Vb in cultured neurons varies with maturation stage. A and B, localization of myosin Vb in cultured hippocampal neurons. A, in DIV
7.06 Cell Biology EXAM #3 April 24, 2003
 7.06 Spring 2003 Exam 3 Name 1 of 8 7.06 Cell Biology EXAM #3 April 24, 2003 This is an open book exam, and you are allowed access to books and notes. Please write your answers to the questions in the
7.06 Spring 2003 Exam 3 Name 1 of 8 7.06 Cell Biology EXAM #3 April 24, 2003 This is an open book exam, and you are allowed access to books and notes. Please write your answers to the questions in the
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
TECHNICAL BULLETIN. Phospho-Akt (pser 473 ) ELISA Kit for detection of human, mouse, or rat phospho-akt (pser 473 ) in cell and tissue lysates
 ELISA Kit for detection of human, mouse, or rat phospho-akt (pser 473 ) in cell and tissue lysates") Phospho-Akt (pser 473 ) ELISA Kit for detection of human, mouse, or rat phospho-akt (pser 473 ) in cell and tissue lysates Catalog Number RAB0011 Storage Temperature 20 C TECHNICAL BULLETIN Product Description
Phospho-Akt (pser 473 ) ELISA Kit for detection of human, mouse, or rat phospho-akt (pser 473 ) in cell and tissue lysates Catalog Number RAB0011 Storage Temperature 20 C TECHNICAL BULLETIN Product Description
Phosphoinositide Signaling
 Phosphoinositide Signaling Pietro De Camilli Department of Cell Biology Howard Hughes Medical Institute Program in Cellular Neuroscience, Neurodegeneration and Repair Yale University School of Medicine
Phosphoinositide Signaling Pietro De Camilli Department of Cell Biology Howard Hughes Medical Institute Program in Cellular Neuroscience, Neurodegeneration and Repair Yale University School of Medicine
Supplements. Figure S1. B Phalloidin Alexa488
 Supplements A, DMSO, PP2, PP3 Crk-myc Figure S1. (A) Src kinase activity is necessary for recruitment of Crk to Nephrin cytoplasmic domain. Human podocytes expressing /7-NephrinCD () were treated with
Supplements A, DMSO, PP2, PP3 Crk-myc Figure S1. (A) Src kinase activity is necessary for recruitment of Crk to Nephrin cytoplasmic domain. Human podocytes expressing /7-NephrinCD () were treated with
2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein
![2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein](/thumbs/86/94743397.jpg "2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein") Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
regulation of polarized membrane transport by rab GTPases Peter van der Sluijs;
 regulation of polarized membrane transport by rab GTPases Peter van der Sluijs; p.vandersluijs@umcutrecht.nl cytotoxic T cell recognizes complex of viral peptide with MHC class I and kills infected cell
regulation of polarized membrane transport by rab GTPases Peter van der Sluijs; p.vandersluijs@umcutrecht.nl cytotoxic T cell recognizes complex of viral peptide with MHC class I and kills infected cell
Supplementary material. Supplementary Figure legends
 Supplementary material Supplementary Figure legends Supplementary Figure 1: Senescence-associated proliferation stop in response to oncogenic N-RAS expression Proliferation of NHEM cells without (ctrl.)
Supplementary material Supplementary Figure legends Supplementary Figure 1: Senescence-associated proliferation stop in response to oncogenic N-RAS expression Proliferation of NHEM cells without (ctrl.)
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
DYNAMICS SIMULATION, MEMBRANE BINDING, AND CELL TRANSLOCATION STUDIES OF THE PKC
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 38, pp. 26047 26058, September 19, 2008 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Differential Roles
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 38, pp. 26047 26058, September 19, 2008 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Differential Roles
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Supplementary material Legends to Supplementary Figures Figure S1. Figure S2. Figure S3.
 Supplementary material Legends to Supplementary Figures. Figure S1. Expression of BICD-N-MTS fusion does not affect the distribution of the Golgi and endosomes. HeLa cells were transfected with GFP-BICD-N-MTS
Supplementary material Legends to Supplementary Figures. Figure S1. Expression of BICD-N-MTS fusion does not affect the distribution of the Golgi and endosomes. HeLa cells were transfected with GFP-BICD-N-MTS
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
1 ml e.a. USD μl 30 e.a. USD ml 10 e.a. USD ml 100 e.a. USD 1700
 Data Sheet Phosphatase Inhibitor Cocktail (Full spectrum) A blend of pan-phosphatase inhibitors for protection of phosphorylation state. Catalog No. Size Price K5000 1 ml e.a. USD 37 100 μl 30 e.a. USD
Data Sheet Phosphatase Inhibitor Cocktail (Full spectrum) A blend of pan-phosphatase inhibitors for protection of phosphorylation state. Catalog No. Size Price K5000 1 ml e.a. USD 37 100 μl 30 e.a. USD
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Ras and Cell Signaling Exercise
 Ras and Cell Signaling Exercise Learning Objectives In this exercise, you will use, a protein 3D- viewer, to explore: the structure of the Ras protein the active and inactive state of Ras and the amino
Ras and Cell Signaling Exercise Learning Objectives In this exercise, you will use, a protein 3D- viewer, to explore: the structure of the Ras protein the active and inactive state of Ras and the amino
RalA ACTIVATION ASSAY BIOCHEM KIT
 RalA ACTIVATION ASSAY BIOCHEM KIT Cat. # BK040 ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 Technical assistance: (303) - 322-2254 World Wide Web: Write to
RalA ACTIVATION ASSAY BIOCHEM KIT Cat. # BK040 ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 Technical assistance: (303) - 322-2254 World Wide Web: Write to
G-Protein Signaling. Introduction to intracellular signaling. Dr. SARRAY Sameh, Ph.D
 G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
Supplementary Figures
 Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643
 Data Sheet FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643 Background The PI3K/AKT signaling pathway is essential for cell growth and survival. Disruption of this pathway or its regulation has been linked
Data Sheet FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643 Background The PI3K/AKT signaling pathway is essential for cell growth and survival. Disruption of this pathway or its regulation has been linked
/07/$15.00/0 Molecular Endocrinology 21(12): Printed in U.S.A.
: Printed in U.S.A.") 0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
0888-8809/07/$15.00/0 Molecular Endocrinology 21(12):3087 3099 Printed in U.S.A. Copyright 2007 by The Endocrine Society doi: 10.1210/me.2006-0476 The Glucose Transporter 4 FQQI Motif Is Necessary for
Appendix. Table of Contents
 Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Principles of cell signaling Lecture 4
 Principles of cell signaling Lecture 4 Johan Lennartsson Molecular Cell Biology (1BG320), 2014 Johan.Lennartsson@licr.uu.se 1 Receptor tyrosine kinase-induced signal transduction Erk MAP kinase pathway
Principles of cell signaling Lecture 4 Johan Lennartsson Molecular Cell Biology (1BG320), 2014 Johan.Lennartsson@licr.uu.se 1 Receptor tyrosine kinase-induced signal transduction Erk MAP kinase pathway
TECHNICAL BULLETIN. Catalog Number RAB0447 Storage Temperature 20 C
 Phospho-Stat3 (ptyr 705 ) and pan-stat3 ELISA Kit for detection of human, mouse, or rat phospho-stat3 (ptyr 705 ) and pan-stat3 in cell and tissue lysates Catalog Number RAB0447 Storage Temperature 20
Phospho-Stat3 (ptyr 705 ) and pan-stat3 ELISA Kit for detection of human, mouse, or rat phospho-stat3 (ptyr 705 ) and pan-stat3 in cell and tissue lysates Catalog Number RAB0447 Storage Temperature 20
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
3.1 Activation of human peripheral blood T lymphocytes induces reversible association of cofilin with the actin cytoskeleton
 3. RESULTS 3.1 Activation of human peripheral blood T lymphocytes induces reversible association of cofilin with the actin cytoskeleton In unstimulated resting human peripheral blood T lymphocytes (PBTs)
3. RESULTS 3.1 Activation of human peripheral blood T lymphocytes induces reversible association of cofilin with the actin cytoskeleton In unstimulated resting human peripheral blood T lymphocytes (PBTs)
Enzyme-coupled Receptors. Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors
 Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Supplementary Figure 1. Validation of astrocytes. Primary astrocytes were
 Supplementary Figure 1. Validation of astrocytes. Primary astrocytes were separated from the glial cultures using a mild trypsinization protocol. Anti-glial fibrillary acidic protein (GFAP) immunofluorescent
Supplementary Figure 1. Validation of astrocytes. Primary astrocytes were separated from the glial cultures using a mild trypsinization protocol. Anti-glial fibrillary acidic protein (GFAP) immunofluorescent
El Azzouzi et al., http ://www.jcb.org /cgi /content /full /jcb /DC1
 Supplemental material JCB El Azzouzi et al., http ://www.jcb.org /cgi /content /full /jcb.201510043 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Acquisition of -phluorin correlates negatively with podosome
Supplemental material JCB El Azzouzi et al., http ://www.jcb.org /cgi /content /full /jcb.201510043 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Acquisition of -phluorin correlates negatively with podosome
Signaling Through Immune System Receptors (Ch. 7)
") Signaling Through Immune System Receptors (Ch. 7) 1. General principles of signal transduction and propagation. 2. Antigen receptor signaling and lymphocyte activation. 3. Other receptors and signaling
Signaling Through Immune System Receptors (Ch. 7) 1. General principles of signal transduction and propagation. 2. Antigen receptor signaling and lymphocyte activation. 3. Other receptors and signaling
The pi-value distribution of single-pass membrane proteins at the plasma membrane in immune cells and in total cells.
 Supplementary Figure 1 The pi-value distribution of single-pass membrane proteins at the plasma membrane in immune cells and in total cells. The PI values were measured for the first 10 amino acids in
Supplementary Figure 1 The pi-value distribution of single-pass membrane proteins at the plasma membrane in immune cells and in total cells. The PI values were measured for the first 10 amino acids in
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest.
 Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Life Science 1A Final Exam. January 19, 2006
 ame: TF: Section Time Life Science 1A Final Exam January 19, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use non-erasable
ame: TF: Section Time Life Science 1A Final Exam January 19, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use non-erasable
Supplementary Figure 1. Rab27a-KD inhibits speed and persistence of HEp3 cells migrating in the chick CAM. (a) Western blot analysis of Rab27a
 Western blot analysis of Rab27a") Supplementary Figure 1. Rab27a-KD inhibits speed and persistence of HEp3 cells migrating in the chick CAM. (a) Western blot analysis of Rab27a expression in GFP-expressing HEp3 cells. (b) Representative
Supplementary Figure 1. Rab27a-KD inhibits speed and persistence of HEp3 cells migrating in the chick CAM. (a) Western blot analysis of Rab27a expression in GFP-expressing HEp3 cells. (b) Representative
SelectScreen Biochemical Kinase Profiling Service
 Page 1 of 11 ASSAY THEORY 2 ADAPTA ASSAY CONDITIONS 3 ADAPTA ASSAY CONTROLS 4 ADAPTA DATA ANALYSIS 5 KINASE-SPECIFIC ASSAY CONDITIONS 6 CAMK1 (CaMK1) 6 CDK7/cyclin H/MNAT1 6 CDK9/cyclin T1 6 CHUK (IKK
Page 1 of 11 ASSAY THEORY 2 ADAPTA ASSAY CONDITIONS 3 ADAPTA ASSAY CONTROLS 4 ADAPTA DATA ANALYSIS 5 KINASE-SPECIFIC ASSAY CONDITIONS 6 CAMK1 (CaMK1) 6 CDK7/cyclin H/MNAT1 6 CDK9/cyclin T1 6 CHUK (IKK
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras
-based studies of receptor dynamics in living cells with Berthold s Mithras") Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Insulin stimulated glucose uptake in adipocytes requires the co-localization of
 STALL JR., RICHARD, M.S. Investigation of the Interaction Between Myosin IIA and Filamentous Actin During Insulin Stimulated Glucose Uptake in 3T3-L1 Adipocytes. (2012) Directed by Dr. Yashomati M. Patel
STALL JR., RICHARD, M.S. Investigation of the Interaction Between Myosin IIA and Filamentous Actin During Insulin Stimulated Glucose Uptake in 3T3-L1 Adipocytes. (2012) Directed by Dr. Yashomati M. Patel
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Membrane transport D. Endocytosis and Exocytosis
B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small cell lung cancer
, NCI-H460 (human non-small cell lung cancer") Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Cellular Biochemistry
 Cellular Biochemistry Fall Semester 2013 Sept. 23 Benoit Kornmann Institute of Biochemistry Introduction to biological membranes General functions and properties Membrane lipids Physical properties Distribution/asymmetry
Cellular Biochemistry Fall Semester 2013 Sept. 23 Benoit Kornmann Institute of Biochemistry Introduction to biological membranes General functions and properties Membrane lipids Physical properties Distribution/asymmetry
The role of the scaffolding protein Tks5 in EGF signaling
 The role of the scaffolding protein Tks5 in EGF signaling PhD Thesis Anna Fekete Doctoral School in Biology Head of the School: Dr. Anna Erdei Structural Biochemistry Doctoral Program Head of the Program:
The role of the scaffolding protein Tks5 in EGF signaling PhD Thesis Anna Fekete Doctoral School in Biology Head of the School: Dr. Anna Erdei Structural Biochemistry Doctoral Program Head of the Program:
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
ASSAY THEORY 2 ADAPTA ASSAY CONDITIONS 3 ADAPTA ASSAY CONTROLS 4 ADAPTA DATA ANALYSIS 5
 Page 1 of 11 ASSAY THEORY 2 ADAPTA ASSAY CONDITIONS 3 ADAPTA ASSAY CONTROLS 4 ADAPTA DATA ANALYSIS 5 KINASE-SPECIFIC ASSAY CONDITIONS 6 CAMK1 (CaMK1) 6 CDK4/cyclin D1 6 CDK4/cyclin D3 6 CDK6/cyclin D1
Page 1 of 11 ASSAY THEORY 2 ADAPTA ASSAY CONDITIONS 3 ADAPTA ASSAY CONTROLS 4 ADAPTA DATA ANALYSIS 5 KINASE-SPECIFIC ASSAY CONDITIONS 6 CAMK1 (CaMK1) 6 CDK4/cyclin D1 6 CDK4/cyclin D3 6 CDK6/cyclin D1