Inflammasome activation causes dual recruitment of NLRC4 and NLRP3 to the same macro-molecular complex
|
|
|
- Kerry Cornelia Daniels
- 5 years ago
- Views:
Transcription
1 Inflammasome activation causes dual recruitment of NLRC4 and NLRP3 to the same macro-molecular complex Si Ming Man, 1 Lee J. Hopkins, 1 Eileen Nugent, 2 Susan Cox, 3 Ivo M. Glück, 1 Panagiotis Tourlomousis, 1 John A. Wright, 1 Pietro Cicuta, 2 Tom P. Monie, 4 Clare E. Bryant 1* 1 Department of Veterinary Medicine, University of Cambridge, Madingley Road, CB3 0ES, United Kingdom. 2 Sector of Biological and Soft Systems, The Cavendish Laboratory, University of Cambridge, Madingley Road, Cambridge 3 Randall Division of Cell & Molecular Biophysics, Kings College, London, WC2R 2LS 4 Department of Biochemistry, University of Cambridge, 80 Tennis Court Road, Cambridge, CB2 1GA, United Kingdom. * Correspondence to: Clare E. Bryant Professor of Innate Immunity Department of Veterinary Medicine, University of Cambridge, Madingley Road, CB3 0ES, United Kingdom. ceb27@cam.ac.uk Phone: +44 (0) Fax: +44 (0) Running Title: NLRC4-NLRP3 inflammasome Keywords: caspase-1, caspase-8, bacteria, speck, innate immunity 1
2 Author contributions: S.M.M., L.J.H., E.N., S.C., I.G., P.T., and J.A.W. performed research; S.M.M., L.J.H., E.N., S.C., P.T., J.A.W., P.C., T.P.M., and C.E.B. analyzed data; and S.M.M., T.P.M., and C.E.B. wrote the paper. The authors declare no conflict of interest. 2
3 Abstract Pathogen recognition by nucleotide-binding oligomerization domain-like receptor (NLR) results in the formation of a macro-molecular protein complex (inflammasome) that drives protective inflammatory responses in the host. It is thought that the number of inflammasome complexes forms in a cell is determined by the number of NLRs being activated, with each NLR initiating its own inflammasome assembly independently of one another; however, here we show that the important food poisoning bacterium Salmonella enterica serovar Typhimurium (S. Typhimurium) simultaneously activates at least two NLRs whilst only a single inflammasome complex is formed in a macrophage. Both NLRC4 and NLRP3 are simultaneously present in the same inflammasome where both NLRs are required to drive IL-1β processing within the Salmonellainfected cell and regulate the bacterial burden in mice. Super resolution imaging of Salmonellainfected macrophages revealed a macro-molecular complex with an outer ring of ASC, an inner ring of NLRs, with active caspase effectors containing the pro-il-1β substrate localized internal to the ring structure. Our data reveal the spatial localization of different components of the inflammasome and how different members of the NLR family co-operate to drive robust IL-1 processing during Salmonella infection. 3
4 Significance Statement The nucleotide oligomerization domain-like receptor (NLR) family members, NLRC4 and NLRP3, activate the inflammasome to provide host defenses against infection. The precise molecular constituents of an inflammasome are unknown, however, it is believed that receptorspecific complexes containing ASC and caspase-1 are formed. Here we used confocal and superresolution microscopy to show that in macrophages infected with Salmonella Typhimurium, a pathogen that activates two distinct NLRs, ASC forms an outer ring-like structure that comprises NLRC4, NLRP3, caspase-1, caspase-8 and pro-il-1 within the same macro-molecular complex. These results suggest that the inflammasome is a highly dynamic macro-molecular protein complex capable of recruiting different NLRs and effectors to co-ordinate inflammasome responses to infection. 4
5 \body INTRODUCTION Inflammasomes are cytosolic multimeric protein complexes formed in the host cell in response to the detection of pathogen associated molecular patterns (PAMPs) or danger associated molecular patterns (DAMPs). Formation of the inflammasome in response to PAMPs is critical for host defense because it facilitates processing of the proinflammatory cytokines pro-il-1β and pro-il-18 into their mature forms (1). The inflammasome also initiates host cell death in the form of pyroptosis, releasing macrophage resident microbes to be killed by other immune mechanisms (2). The current paradigm is that there are individual, receptor specific inflammasomes consisting of one NLR (nucleotide-binding-and-oligomerization domain and leucine-rich-repeatcontaining) or PYHIN (pyrin domain and HIN domain-containing) receptor, the adaptor protein apoptosis-associated speck-like protein containing a CARD (ASC), and caspase-1 (3). How the protein constituents of the inflammasome are spatially orientated is unclear. NLRC4 and NLRP3 are the best characterized inflammasomes, especially with respect to their responses to pathogenic bacteria. The NLRC4 inflammasome is activated primarily by bacteria, including Aeromonas veronii (4), Escherichia coli (5), Listeria monocytogenes (6, 7), Pseudomonas aeruginosa (5), Salmonella enterica serovar Typhimurium (5, 8-10) and Yersinia species (11). In mouse macrophages, the NLRC4 inflammasome responds to flagellin and Type III secretion system (T3SS) associated needle or rod proteins (5, 8, 9) after their detection by NAIP5 or NAIP6 and NAIP1 or NAIP2, respectively (12-15). Phosphorylation of NLRC4 at a single, evolutionarily conserved residue, Ser 533, by PRKCD (PKCδ) kinase is required for NLRC4 inflammasome assembly (16). The NLRP3 inflammasome is activated by a large repertoire of DAMPs, including ATP, nigericin, maitotoxin, uric acid crystals, silica, aluminium hydroxide and muramyl dipeptide (17-20). NLRP3 is also activated by bacterial PAMPs from many species, including Aeromonas species (4, 21), L. monocytogenes (6, 7, 22), Neisseria gonorrhoe (23), S. Typhimurium (10), Streptococcus pneumoniae (24) and Yersinia species (11). The mechanisms by which NLRC4 and NLRP3 inflammasomes contribute to host defense against bacterial pathogens are emerging, however, little is known about the dynamics governing inflammasome assembly in infections caused by bacteria that activate multiple NLRs, such as S. Typhimurium (10), A. veronii (4) and Yersinia (11). 5
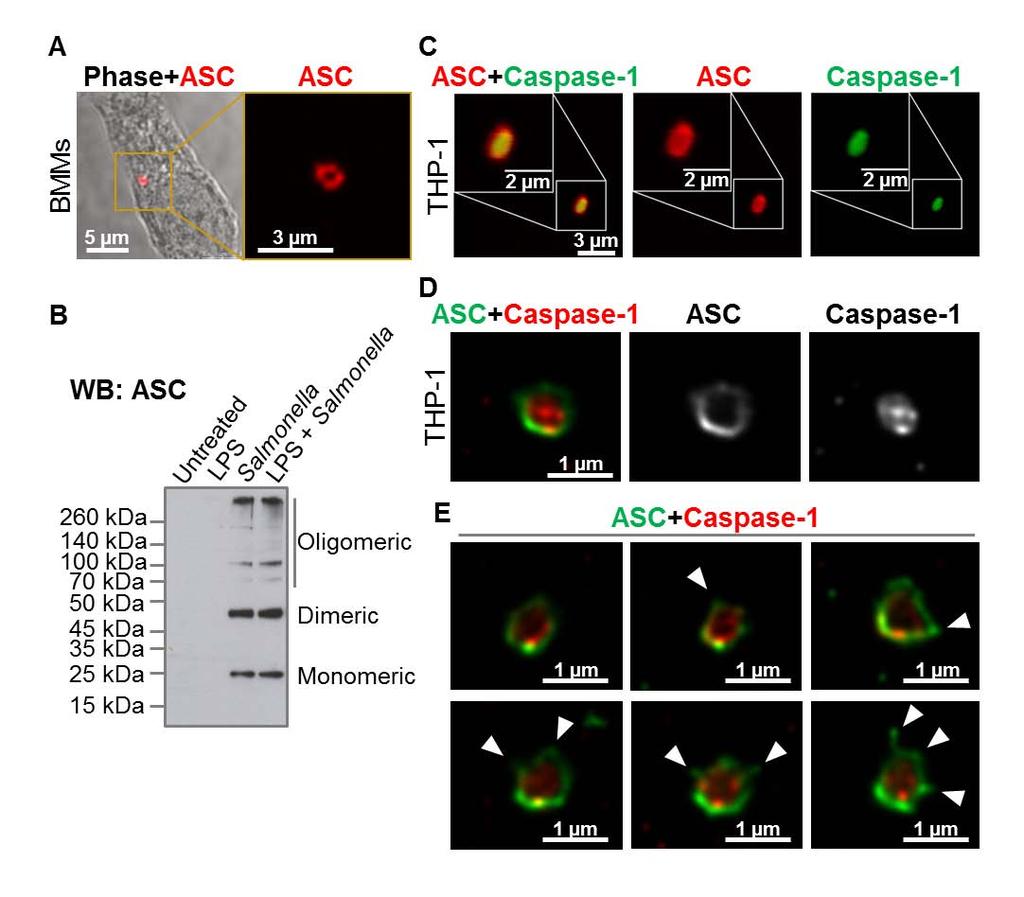
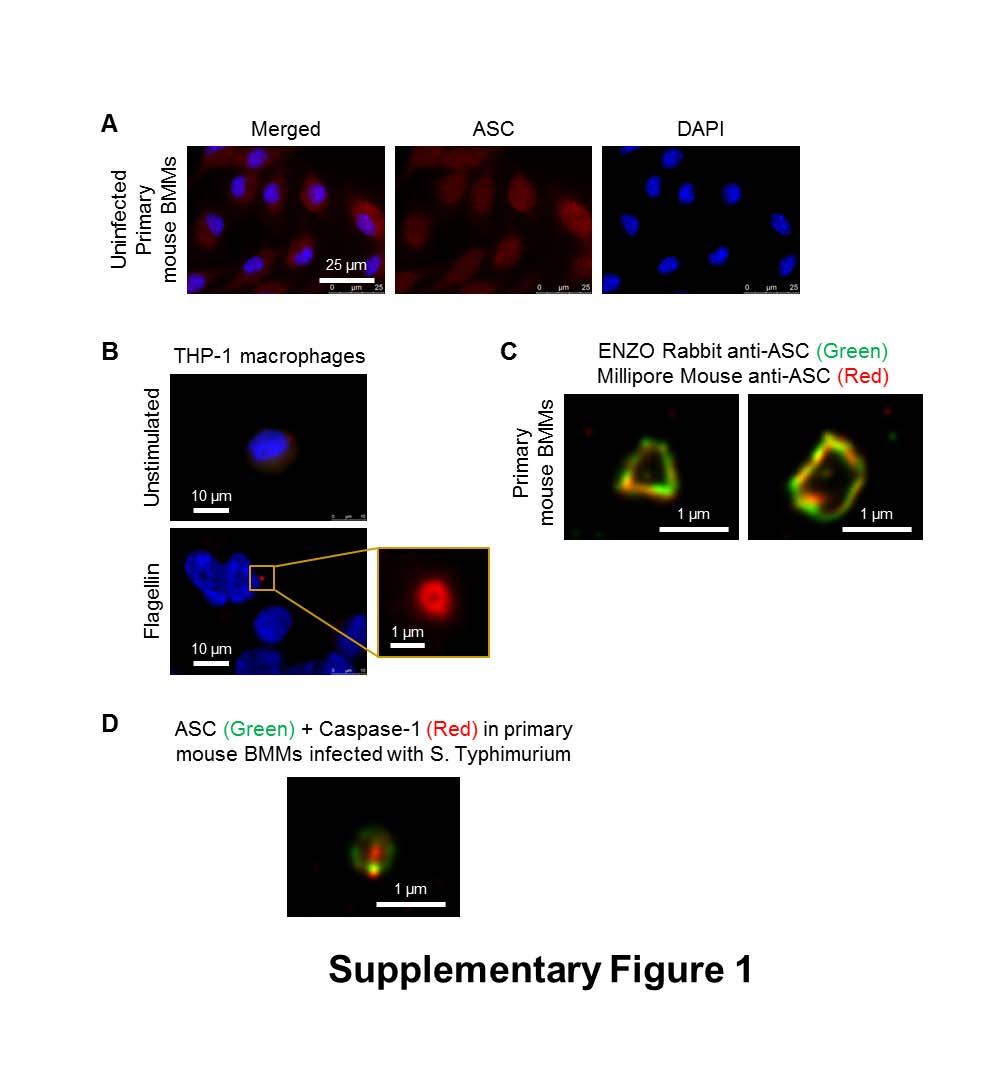
6 NLRP3 does not have a CARD (caspase activation and recruitment domain) and requires ASC to interact with the CARD domain of procaspase-1. This interaction requires a charged interface around Asp27 of the procaspase-1 CARD (25). Whether ASC is also required for the assembly of the NLRC4 inflammasome is less clear. NLRC4 contains a CARD which can interact directly with the CARD domain of procaspase-1 (26), however ASC is required for some of the responses driven by NLRC4 (27). Macrophages infected with S. Typhimurium or other pathogens exhibit formation of a distinct cytoplasmic ASC focus or speck, which can be visualized under the microscope and is indicative of inflammasome activation (10, 28, 29). Our lab and others have shown that only one ASC speck is formed per cell irrespective of the stimulus used (29-32). Yet many bacteria activate two or more NLRs and it is unclear whether a singular inflammasome is formed at a time or multiple inflammasomes are formed independently of each other, with each inflammasome containing one member of the NLR family. In this study we describe the endogenous molecular constituents of the Salmonella-induced inflammasome and their spatial orientation. In cross section, ASC forms a large external ring with the NLRs and caspases located internally. Critically NLRC4, NLRP3, caspase-1 and caspase-8 co-exist in the same ASC speck to coordinate pro-il-1β processing. All ASC specks observed contained both NLRC4 and NLRP3. These results suggest that Salmonella infection induces a single inflammasome protein complex containing different NLRs and recruiting multiple caspases to co-ordinate a multifaceted inflammatory response to infection. RESULTS NLR and caspase-1 are both spatially organized within the Salmonella-induced ASC speck The inflammasome adaptor protein ASC aggregates and forms a large speck in response to different inflammasome stimuli. Using confocal imaging we immunolocalized ASC after infection of primary bone marrow derived macrophages (BMMs) with S. Typhimurium (MOI 10) for 1 h. ASC redistributed to form a focus or speck in BMMs infected with S. Typhimurium (Fig. 1A), whereas endogenous ASC predominately distributed throughout the cytosol of unstimulated BMMs (Fig. S1A). Each infected cell contained one ASC speck of µm diameter after infection with S. Typhimurium (Fig. 1A). Protein oligomerization assays confirmed that ASC 6
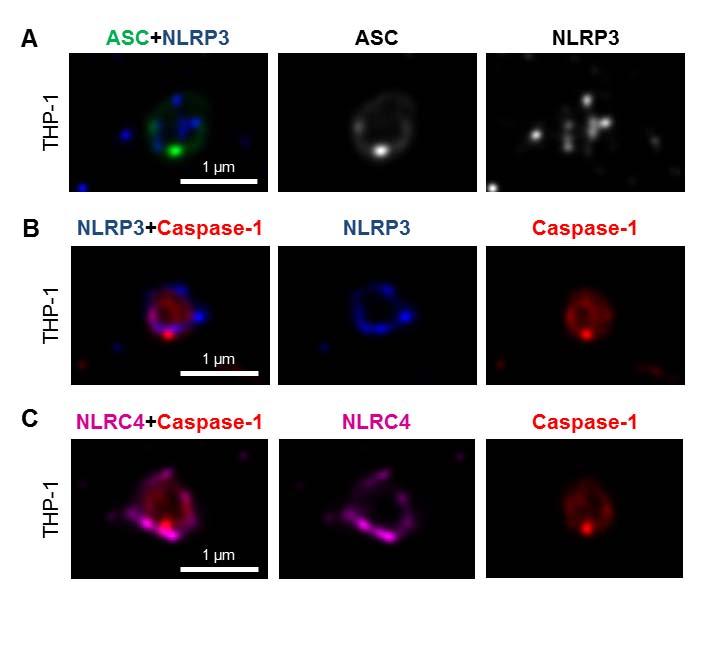
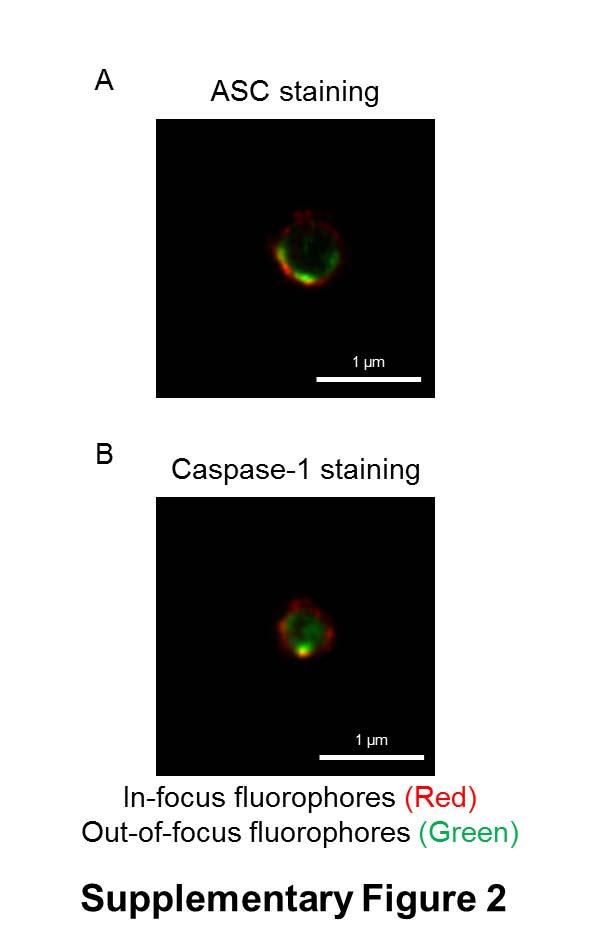
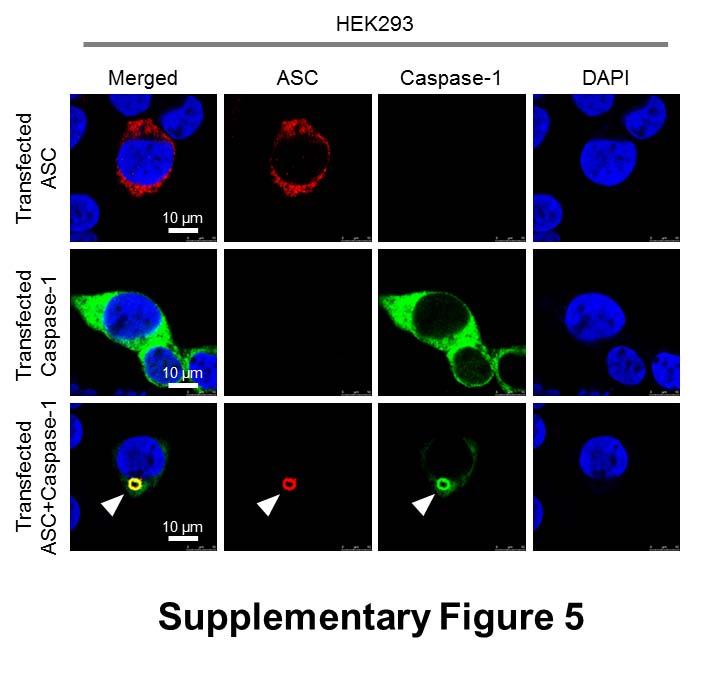
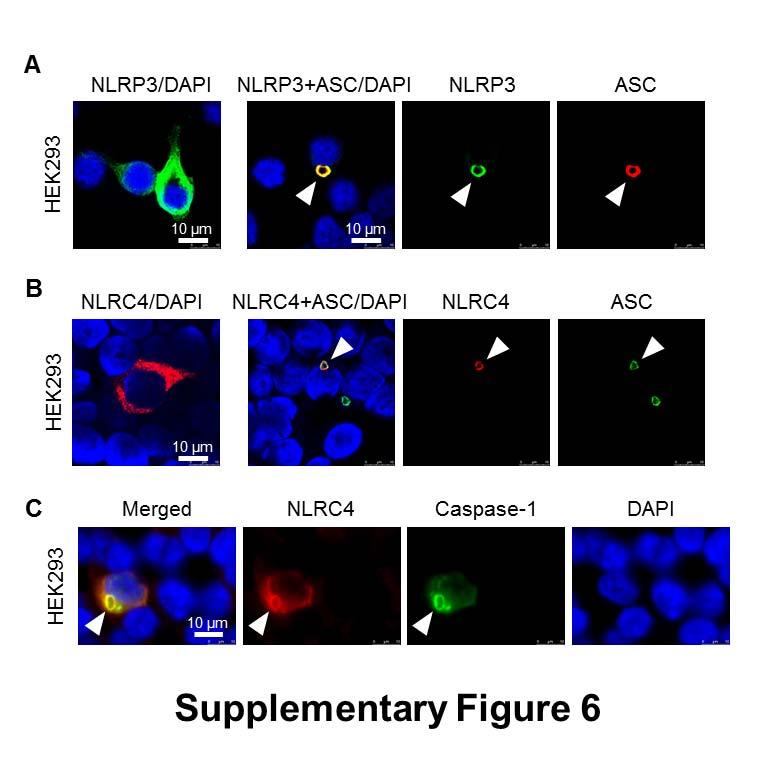
7 formed oligomers following Salmonella infection (Fig. 1B). Specks that appeared in a cross section orientation had an apparent hole in the middle (Fig. 1A). A similar ASC ring structure was observed in human THP-1 macrophages (Fig. 1C and Fig. S1B). We used Bayesian localization super-resolution microscopy to obtain higher magnification images of the specks in cross section (33). After infection of THP-1 cells or BMMs with S. Typhimurium (MOI 10) for 30 min, immuno-labeled ASC specks from different cells were visualized and the images compared to each other. Analysis of multiple cross sections of an ASC speck revealed that it forms a ring-like structure (Fig. 1D). Immuno-labeling of ASC using two different antibodies revealed the same ring-like structure (Fig. S1C), suggesting that this is not due to the effect of a particular antibody. Further analysis of the ASC specks showed the presence of side filaments frequently decorating the external region of the ASC ring structure (Fig. 1E). How other endogenous proteins of the inflammasome associate with and spatially distribute relative to ASC is unclear. We used super-resolution microscopy to resolve the spatial orientation of ASC, caspase-1, and either NLRP3 or NLRC4 in macrophages infected with S. Typhimurium. Confocal imaging of caspase-1 using FLICA labeling in THP-1 cells or mouse BMMs identified caspase-1 specks of a smaller diameter (0.426 µm +/ µm; n=8) to the ASC specks (Fig. 1C E and Fig. S1D). Super-resolution imaging of ASC and caspase-1 colocalization showed that the ASC ring surrounded caspase-1, with the caspase-1 filling the central hole of the ASC ring (Fig. 1C E and Fig. S1D). We attempted to generate a three dimensional reconstruction of the super resolution ASC structure, however, the point spread function of the microscope is symmetric, which means that it is not possible to discriminate between point spread functions which are out-of-focus above the plane of focus, and those that are an equal distance low. However, we were able to discriminate between in-focus and out-of-focus point spread functions in the super resolution data. These show the in-focus fluorophores tending to be localized to the edge of the ring, with the out-of-focus fluorophores being more spread out (Fig. S2). This suggests that the outer ring structure is reasonably well defined in one plane, with some degree of narrowing occurring above or below. We next wondered how individual NLRs would spatially associate with ASC. LPS primed THP- 1 cells were infected with S. Typhimurium for 30 min at MOI 10 and immuno-labeled for endogenous NLRP3 and ASC. Confocal imaging of NLRP3 showed, like those seen for caspase-1, 7
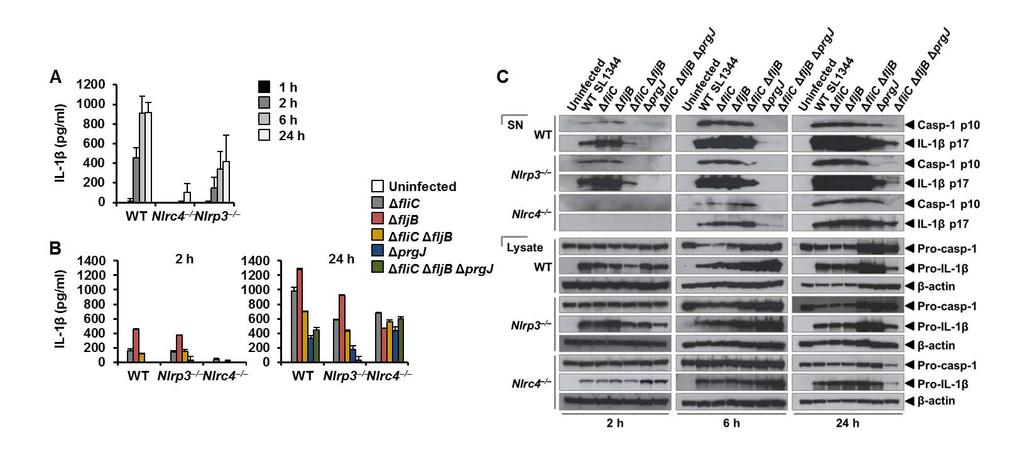
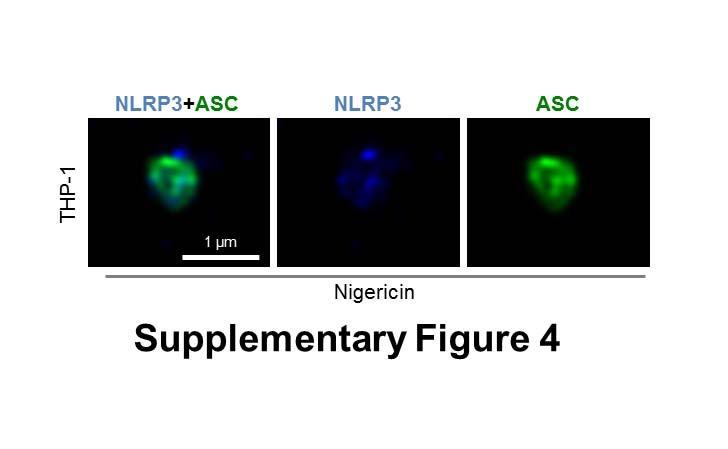
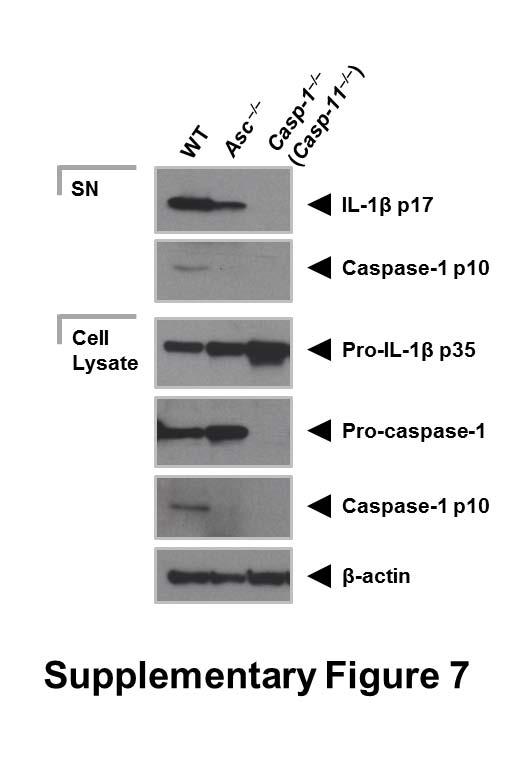
8 specks that were smaller (0.4 µm) than those seen with ASC immuno-labeling (Fig. 2A). Superresolution imaging of the co-labeled ASC and NLRP3 showed that NLRP3 formed a smaller ring-like structure within the ASC ring (Fig. 2A). This technique also showed that diffused caspase-1 predominantly localized within the NLRP3 ring-like speck in infected BMMs (Fig. 2B). Super-resolution microscopy of immuno-labeled NLRC4 also revealed smaller specks (0.578 µm +/ µm; n=8) than those formed by ASC (Fig. 2C and Fig. S3). Caspase-1 predominantly localized in a NLRC4 ring-like speck (Fig. 2C). Immunolocalization of NLRP3 and ASC in the nigericin-stimulated canonical NLRP3 inflammasome produced a similar pattern to that observed in the Salmonella-induced inflammasome structure (Fig S4). We confirmed our colocalization findings in HEK293 cells transfected with DNA constructs encoding different inflammasome proteins (Fig. S5 and S6). Taken together we provide evidence to show that following Salmonella infection of macrophages endogenous NLRC4 or NLRP3 reside in the ASC speck, within which caspase-1 is located to form an inflammasome complex. Co-ordinated activation of NLRs sustains infection-induced IL-1 production NLRC4 and NLRP3 can both be activated by Salmonella (10), but the dynamics of how NLRC4 and NLRP3 activation is coordinated with respect to their localization within the ASC speck during infection is unclear. Infection of BMMs from wildtype and Asc / mice with log phase grown wildtype S. Typhimurium for 2 h showed that Salmonella induction of caspase-1 proteolysis, pro-il-1β processing and IL-1β release required ASC (Fig. S7). Both NLRC4 and NLRP3 were required to drive robust IL-1β production (Fig. 3A). To further investigate the dynamics of NLRC4 and NLRP3-mediated IL-1β production, we infected primary BMMs from wildtype, Nlrc4 / or Nlrp3 / mice with S. Typhimurium lacking flagellin (ΔfliC ΔfljB), PrgJ (ΔprgJ) or all known activators of the NLRC4 inflammasome (ΔfliC ΔfljB ΔprgJ) (14, 15). We found that NLRC4 predominately drove caspase-1 proteolysis, pro-il-1β processing and IL-1β production in response to all Salmonella mutants after 2 h of infection, whereas NLRP3 responded to all mutants following 24 h of infection (Fig. 3B, 3C). Our results suggest that NLRC4 and NLRP3 cooperate to induce robust inflammasome activation following infection with S. Typhimurium. Our in vitro data suggest a role for both NLRC4 and NLRP3 in the host response to infection with S. Typhimurium. In lethal salmonellosis NLRP3 and NLRC4 are required to control the infection on day 5 post-infection (10), but the relative contributions of these NLRs in regulating 8
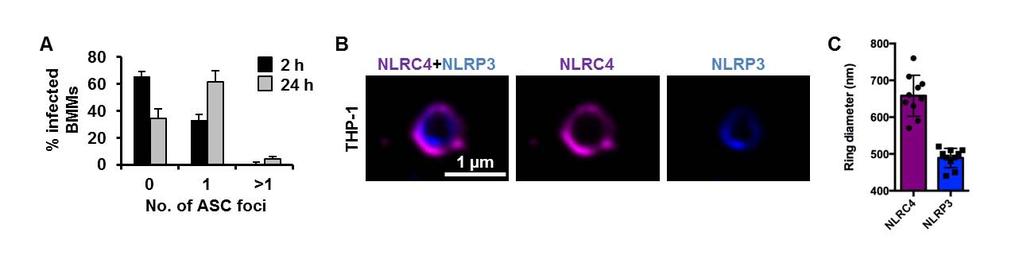
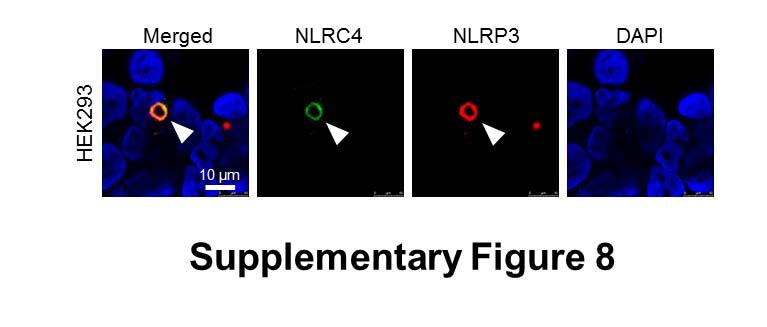
9 bacterial burden over time is unknown. We therefore used a sublethal model of murine salmonellosis where the mice control bacterial growth and eventually clear the infection over time, which allows us to monitor the impact of NLRs on Salmonella growth over a longer time course. Mice were infected with S. Typhimurium and killed at days 1, 7 and 13 post-infection. A similar bacterial burden was found in all mice at day 1 (Fig. 4). At day 7, both Nlrc4 / or Nlrp3 / mice had higher bacterial counts than wildtype mice (Fig. 4). Casp-1 / (casp-11 / ) mice had the highest bacterial burden compared to other mice on day 7, suggesting that both NLRs contribute to the host defense against Salmonella infection 7 days post-infection. On day 13, we observed elevated bacterial burden in Nlrc4 / or Nlrp3 / mice compared to wildtype mice (Fig. 4). Unlike day 7, a substantially higher bacterial numbers were found in Nlrc4 / mice compared to Nlrp3 / mice on day 13. Casp-1 / (casp-11 / ) mice still contained the highest bacterial burden compared to both Nlrc4 / or Nlrp3 / mice. These results suggest that the relative contribution between NLRC4 and NLRP3 in the host control of sub-lethal salmonellosis is comparable on day 7, whereas on day 13 NLRC4 exerts a greater antibacterial effect than NLRP3. The reason why we see distinct roles for NLRC4 and NLRP3, in comparison to Broz and colleagues (10), is simply because the differences in bacterial growth only become apparent as the infection progresses over time. This can only be seen if the animals can ultimately control bacterial growth. These data support the concept that NLRC4 and NLRP3 coordinate suppression of bacterial growth in vivo during Salmonella infection. Multiple NLRs and caspases can be recruited to the same Salmonella-induced inflammasome Activation of multiple NLRs in response to Salmonella infection occurs in vitro and in vivo, yet how is this achieved if cells only form a single inflammasome per cell? Infection of BMMs with S. Typhimurium for 24 h to induce the activation of both NLRC4 and NLRP3 still resulted in the formation of a single ASC speck per macrophage (Fig. 5A). A possible explanation for this is that triggering the formation of a single ASC speck by multiple NLRs could result in more than one NLR being present in the same inflammasome complex. Rather than having separate NLRP3 or NLRC4 inflammasomes occurring, we hypothesized that the inflammasome could be a single dynamic protein complex that is formed in response to different NLR inputs. NAIPs and NLRC4 are recruited to the same inflammasome so it is feasible that NLRP3 and NLRC4 may also be 9
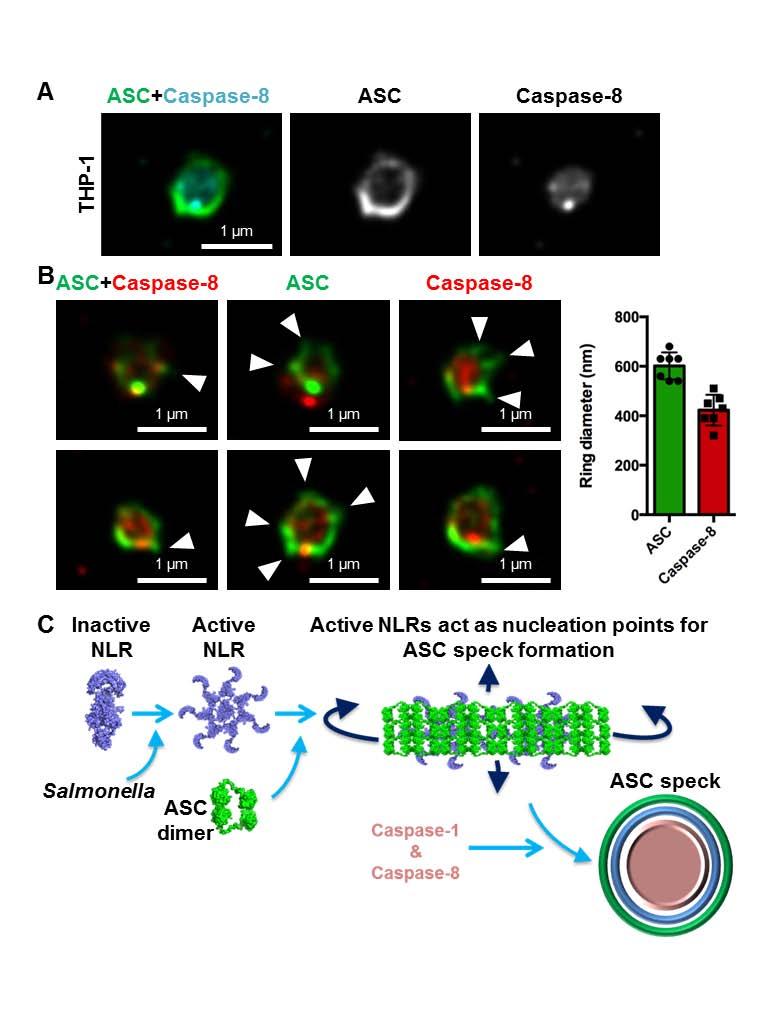
10 present in a single inflammasome complex (14, 15). To investigate this, we used super-resolution microscopy to visualize immuno-labeled endogenous NLRC4 and NLRP3 in LPS-primed THP-1 macrophages infected with S. Typhimurium. All of the ASC specks observed contained both NLRC4 and NLRP3 and image analysis of these showed co-localization of both NLRs in the same speck (Fig. 5B). NLRC4 localized in a ring-like structure (ring diameter µm +/ µm; n=10) whilst NLRP3 formed a smaller and more centralized ring-like structure (ring diameter µm +/ µm; n=10) (Fig. 5C). We confirmed these findings by coexpressing DNA constructs encoding human NLRC4 and NLRP3 in HEK293 cells and found that both receptors redistributed and colocalized into a single complex (Fig. S8). Our data suggest that NLRC4 and NLRP3 can be located to the same inflammasome. Given that the inflammasome platform can contain more than one NLR we wondered whether this also occurs for inflammasome-associated caspases. We have previously shown that the ASC speck is required to recruit the inflammasome effectors, caspase-1 and caspase-8, into a single complex during Salmonella infection (Fig. S9A) (32). In THP-1 cells super-resolution analysis of the ASC-caspase-8 speck showed a similar distribution to that seen for caspase-1 distribution within the ASC ring, with the diameter of the caspase-8 core measured at µm +/- 62 µm (n=7) (Fig. 6A,B). We performed additional cross-sectional analysis of the ASC-Caspase-8 specks (Fig. 6B) and confirmed these findings in primary BMMs (Fig. S9B). Taken together, we hypothesized that a single inflammasome platform provides a major proteolytic cleavage site within the cell where pro-il-1β must be recruited for processing. We stained pro-il-1β, ASC and caspase-1 and found that pro-il-1β formed a smaller speck-like structure (0.4 µm) that colocalized within the ASC-caspase-1 speck (Fig. S10), suggesting that pro-il-1β may be required to enter the inflammasome pore for proteolytic processing. It is not unreasonable to speculate that the ASC speck is a substrate-specific entity whereby only specific substrates such as pro-il-1β and pro-il-18 are able to enter for caspase-1 and -8 dependent processing while providing a controlled environment to prevent unregulated caspase-1 and -8 activity within the cell. The concentric organization of the speck suggests that ligand-mediated NLR activation provides a nucleation point for the rapid recruitment of ASC dimers and the subsequent assembly of the ASC speck, concomitant with the recruitment of caspase-1 and -8 (Fig. 6C). This results in the formation of an inflammasome platform containing more than one NLR family member which 10
11 drives the recruitment of multiple effectors to the same protein complex to process pro-il-1β and pro-il
12 DISCUSSION Bacterial infection activates different NLRs to drive inflammasome production of IL-1β and IL- 18 in the host tissue to help control the spread of the organism. Here we provide an answer to a central question in inflammasome biology. That is, in response to pathogens, like Salmonella, which activate multiple NLRs why does only a single ASC speck form? Our combined confocal and super-resolution imaging approach along with functional analysis shows, for the first time, that endogenous NLRC4 and NLRP3 colocalize in the same speck. Different NLRs can therefore be present in the same ASC speck to coordinate inflammasome functions after Salmonella infection. A single ASC speck can also contain multiple caspase effectors (caspase-1 and -8) at the same time, suggesting that the inflammasome platform is a dynamic protein complex that contains multiple NLRs and recruits more than one effector caspase to control the production of IL- 1β. Our work provides a clear picture of the gross organization of the endogenous ASC speck. The concentric organization of an external ASC layer surrounding the NLR proteins, which in turn surround the effector caspases, provides important insight into the function of the inflammasome. This arrangement permits the dynamic recruitment and movement of speck constituents following activation, thereby ensuring the inflammasome remains functionally active. For example, NLRP3 is critical for sustained pro-il-1β processing and is recruited to the ASC speck at a later time point than NLRC4, which is instead crucial during early infection. Hence, NLRP3 is positioned inside NLRC4 (Fig. 5B). The formation of an outer shell of ASC, rather than a solid or closed speck, is also consistent with continual dynamic recruitment and with the presence of a functional core consisting of active caspases and pro-il-1β (Fig. S10). A core that would be inaccessible if the ASC formed a closed shell. The outer ASC shell may also serve to stabilize and protect the internal structure. It is interesting to note that side filaments are often found decorating the ASC shell although the functional significance of this is currently unclear. The nature of the protein-protein interactions within the speck remains elusive at this resolution and it is intriguing that direct co-localization of components that reportedly interact is not always observed. This is likely to be a result of both the experimental reagents available and the functional organization of the speck. We predominantly co-stain ASC and active caspase-1 or -8 in our study, resulting in detection of active caspase-1/8 in the functional centre of the speck, pre- 12
13 sumably following its release from ASC (Fig. 6). Whereas where we stained ASC and caspase-8 using an antibody that detects both cleaved and pro-forms, more of the caspase is localized with the ASC ring (compare Fig. S9B and Fig. 6). Our results show that the spatial orientation of NLRP3 and ASC in the nigericin-stimulated canonical inflammasome resembles a similar pattern to that observed in the more complex Salmonella-induced inflammasome structure (Fig S4). We speculate that the composition of the AIM2 inflammasome would have a similar structure to the multi-nlr inflammasome. Indeed, analysis of the crystal structure of the HIN200 domain of AIM2 and dsdna revealed that the positively charged HIN domain embraces the dsdna while the AIM2 PYD domain located periphery to the HIN-dsDNA structure is thought to facilitate the recruitment of ASC (34). This proposed structure composed of dsdna-aim2 internal to ASC is similar to our observed structure comprised of NLRC4 and NLRP3 within an ASC shell. In contrast to our concentric ring structure where caspase-1 lies within the ASC ring, a recent study using Cryo-EM analysis suggests that caspase-1 forms filamentous structures external to ASC following nucleation by either NLRP3 or AIM2 (35). This difference is most likely due to the altered protein levels between an exogenous over-expression system and our own visualization of the endogenous inflammasome. Earlier EM-staining of endogenous ASC also reports a hollow, ring-like organization (36). The cryo-em analysis of the AIM2 ternary complex and the NLRP3 inflammasome represents a snapshot of inflammasome formation and may correlate with the starting point of full speck formation. This would be followed by additional NLR and ASC recruitment leading ultimately to the ring-like structure we observe. Salmonella activates both NLRC4 and NLRP3 which results in ASC focus formation and recruitment of caspases to the inflammasome (10, 32). Our data show that in vitro NLRP3 contributes to the macrophage response against Salmonella at a later stage than NLRC4 in unprimed conditions. This is unsurprising as NLRP3 expression in BMMs requires induction by LPS or cytokines such as TNF-α (37). In experiments using unprimed BMMs infected with Salmonella (Fig 2A and B), there will be a delay before NLRP3 is expressed, whereas NLRC4, which is constitutively expressed, will be activated immediately upon cellular infection. In vivo as infection progresses the effect of NLRC4 is slightly more important at day 13 in comparison to days 1 and 7. There are many factors that could explain the differences, including time-dependent effects of NLRC4 and NLRP3 in vitro and in vivo. These include the changing profile of cytokines, 13
14 the effects of NLRC4, but not NLRP3, in driving Salmonella-induced pyroptosis, and the switch in cell type dominance, over time during infection. Macrophages, for example, play a key role at day 7, the time point at which NLRC4 and NLRP3 have a similar effect in vivo, whereas by day 13 other cells such as NK cells and CD4 + T cells are starting to play a major role in controlling the infection. If NLRC4 and NLRP3 both contribute to the host defense against Salmonella infection, then mice lacking either of these receptors should show a greater susceptibility to Salmonella infection compared to wildtype mice. Our work supports this hypothesis because mice lacking NLRC4 or NLRP3 have higher bacterial loads than the wildtype mice at days 7 and 13 postinfection. Broz et al (10) previously reported that mice lacking NLRC4 or NLRP3 harbor similar bacterial numbers in the spleen, liver and mesenteric lymph nodes 5 days after Salmonella infection, however, consistent with our observations, they showed that mice lacking both NLRP3 and NLRC4 harbor a higher bacterial load compared to wildtype controls. We observed similar bacterial numbers in spleen and liver between wildtype mice and any of the knock-out strains at an early time point (day 1), but as the infection progresses (days 7 and 13) we see a clear increase in the bacterial burden in mice lacking NLRC4 or NLRP3. This observation highlights an important role for each of these receptors over the course of Salmonella infection, which is consistent with our in vitro data. Our data lead us to propose that the inflammasome is a dynamic multi-protein complex, where NLRC4, NLRP3, caspase-1, caspase-8 and pro-il-1β co-localize to the same ASC inflammasome in macrophages infected with Salmonella. Whether caspase-11 might associate with ASC in the non-canonical inflammasome is unclear and the spatial distribution of these proteins will be an interesting area for future research. In conclusion, our data reveal cooperative interactions between distinct members of the NLR and caspase families that form a dynamic inflammasome during Salmonella infection. MATERIALS AND METHODS Detailed information is presented in SI Materials and Methods. We used Bayesian localization microscopy to obtain super-resolution images of the ASC inflammasome structure (33). Super- 14
15 resolution experiments were performed on a Nikon TI eclipse inverted microscope using a Nikon intensilight light source for fluorescence. ACKNOWLEDGEMENTS We would like to thank K.A. Fitzgerald (University of Massachusetts Medical School, USA) for critical review of our manuscript and for supplying the knockout mouse strains, G. Núñez (University of Michigan Medical School, USA) for the anti-nlrc4 antibody, and E. Creagh (Trinity College Dublin, Ireland) for providing technical advice and the ASC construct, F. Morgan, S. Achouri and I. Kazanis (University of Cambridge, UK) for providing assistance in microscopy. S.M.M was supported by a Cambridge International Scholarship. T.P.M was supported by a Wellcome Trust Research Career Development Fellowship (WT085090MA). This study was supported by Biotechnology and Biological Sciences Research Council (BBSRC) grants (BB/H003916/1 and BB/K006436/1) and a BBSRC Research Development Fellowship (BB/H021930/1) awarded to C.E.B. REFERENCES 1. Franchi L, Muñoz-Planillo R, & Núñez G (2012) Sensing and reacting to microbes through the inflammasomes. Nat Immunol 13(4): Miao EA, et al. (2010) Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat Immunol 11(12): Rathinam VAK, Vanaja SK, & Fitzgerald KA (2012) Regulation of inflammasome signaling. Nat Immunol 14(4): McCoy AJ, Koizumi Y, Higa N, & Suzuki T (2010) Differential regulation of caspase-1 activation via NLRP3/NLRC4 inflammasomes mediated by aerolysin and type III secretion system during Aeromonas veronii infection. J Immunol 185(11): Miao EA, et al. (2010) Innate immune detection of the type III secretion apparatus through the NLRC4 inflammasome. Proc Natl Acad Sci U S A 107: Wu J, Fernandes-Alnemri T, & Alnemri ES (2010) Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by Listeria monocytogenes. J Clin Immunol 30(5): Meixenberger K, et al. (2010) Listeria monocytogenes-infected human peripheral blood mononuclear cells produce IL-1beta, depending on listeriolysin O and NLRP3. J Immunol 184(2): Miao EA, et al. (2006) Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1beta via Ipaf. Nat Immunol 7(6):
16 9. Franchi L, et al. (2006) Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1beta in salmonella-infected macrophages. Nat Immunol 7(6): Broz P, et al. (2010) Redundant roles for inflammasome receptors NLRP3 and NLRC4 in host defense against Salmonella. J Exp Med 207(8): Brodsky IE, et al. (2010) A Yersinia effector protein promotes virulence by preventing inflammasome recognition of the type III secretion system. Cell Host Microbe 7(5): Yang J, Zhao Y, Shi J, & Shao F (2013) Human NAIP and mouse NAIP1 recognize bacterial type III secretion needle protein for inflammasome activation. Proc Natl Acad Sci U S A 110(35): Rayamajhi M, Zak DE, Chavarria-Smith J, Vance RE, & Miao EA (2013) Cutting Edge: Mouse NAIP1 Detects the Type III Secretion System Needle Protein. J Immunol 191(8): Kofoed EM & Vance RE (2011) Innate immune recognition of bacterial ligands by NAIPs determines inflammasome specificity. Nature 477(7366): Zhao Y, et al. (2011) The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature 477(7366): Qu Y, et al. (2012) Phosphorylation of NLRC4 is critical for inflammasome activation. Nature 490(7421): Li H, Willingham SB, Ting JP, & Re F (2008) Cutting edge: inflammasome activation by alum and alum's adjuvant effect are mediated by NLRP3. J Immunol 181(1): Mariathasan S, et al. (2006) Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440(7081): Dostert C, et al. (2008) Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 320(5876): Hornung V, et al. (2008) Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat Immunol 9(8): McCoy AJ, et al. (2010) Cytotoxins of the human pathogen Aeromonas hydrophila trigger, via the NLRP3 inflammasome, caspase-1 activation in macrophages. Eur J Immunol 40(10): Kim S, et al. (2010) Listeria monocytogenes is sensed by the NLRP3 and AIM2 inflammasome. Eur J Immunol 40(6): Duncan JA, et al. (2009) Neisseria gonorrhoeae activates the proteinase cathepsin B to mediate the signaling activities of the NLRP3 and ASC-containing inflammasome. J Immunol 182(10): McNeela EA, et al. (2010) Pneumolysin activates the NLRP3 inflammasome and promotes proinflammatory cytokines independently of TLR4. PLoS Pathog 6(11):e Kersse K, Lamkanfi M, Bertrand MJ, Vanden Berghe T, & Vandenabeele P (2011) Interaction patches of procaspase-1 caspase recruitment domains (CARDs) are differently involved in procaspase-1 activation and receptor-interacting protein 2 (RIP2)-dependent nuclear factor kappab signaling. J Biol Chem 286(41): Poyet JL, et al. (2001) Identification of Ipaf, a human caspase-1-activating protein related to Apaf-1. J Biol Chem 276(30): Proell M, Gerlic M, Mace PD, Reed JC, & Riedl SJ (2013) The CARD plays a critical role in ASC foci formation and inflammasome signalling. Biochem J 449(3):
17 28. Jones JW, et al. (2010) Absent in melanoma 2 is required for innate immune recognition of Francisella tularensis. Proc Natl Acad Sci U S A 107(21): Huang MT, et al. (2009) Critical role of apoptotic speck protein containing a caspase recruitment domain (ASC) and NLRP3 in causing necrosis and ASC speck formation induced by Porphyromonas gingivalis in human cells. J Immunol 182(4): Fernandes-Alnemri T, et al. (2007) The pyroptosome: a supramolecular assembly of ASC dimers mediating inflammatory cell death via caspase-1 activation. Cell Death Differ 14(9): Broz P, von Moltke J, Jones JW, Vance RE, & Monack DM (2010) Differential requirement for Caspase-1 autoproteolysis in pathogen-induced cell death and cytokine processing. Cell Host Microbe 8(6): Man SM, et al. (2013) Salmonella infection induces recruitment of Caspase-8 to the inflammasome to modulate IL-1beta production. J Immunol 191(10): Cox S, et al. (2011) Bayesian localization microscopy reveals nanoscale podosome dynamics. Nat Methods 9(2): Jin T, et al. (2012) Structures of the HIN domain:dna complexes reveal ligand binding and activation mechanisms of the AIM2 inflammasome and IFI16 receptor. Immunity 36(4): Lu A, et al. (2014) Unified Polymerization Mechanism for the Assembly of ASC- Dependent Inflammasomes. Cell 156(6): Masumoto J, et al. (1999) ASC, a novel 22-kDa protein, aggregates during apoptosis of human promyelocytic leukemia HL-60 cells. J Biol Chem 274(48): Bauernfeind FG, et al. (2009) Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol 183(2):
18 FIGURE LEGENDS Fig. 1 Spatial organization of the ASC speck. (A) Immuno-labeling of endogenous ASC in primary bone marrow-derived macrophages (BMMs) infected with S. Typhimurium (STm). (B) Endogenous ASC oligomerized in unprimed or LPS-primed BMMs following STm infection. (C) Confocal microscopy imaging of endogenous immuno-labeled ASC and FLICA staining of caspase-1 in human THP-1 macrophages infected with STm. (D) Bayesian localization super-resolution microscopy of endogenous ASC and caspase-1 in C. (E) Multiple cross-sections of the ASC Caspase-1 speck. Arrows indicate side filaments of ASC coming off the external region of the ring-like structure. Fig. 2 NLRs reside in the ASC speck following S. Typhimurium infection of macrophages. Bayesian localization super-resolution microscopy of (A) endogenous immuno-labeled ASC and NLRP3, (B) endogenous immuno-labeled NLRP3 and FLICA-stained caspase-1 and (C) endogenous immuno-labeled NLRC4 and FLICA-stained caspase-1 in human THP-1 macrophages infected with S. Typhimurium. Fig. 3 Co-ordinated activation of NLRs is required for IL-1β production induced by S. Typhimurium infection. WT, Nlrc4 / and Nlrp3 / primary bone marrow-derived macrophages (BMMs) were infected with (A) S. Typhimurium SL1344 or (B) its isogenic mutants ΔfliC, ΔfljB, ΔfliCΔfljB, ΔprgJ and ΔfliCΔfljBΔprgJ (multiplicity of infection [MOI] 1) for the indicated times. (A,B) Supernatant (SN) was collected and the levels of IL-1β were measured using ELISA. (C) Pro-caspase-1, cleaved caspase-1 p10, pro-il-1β and cleaved IL-1β p17 were detected in the supernatant or cell lysate using western blotting. β-actin was used as a loading control. Data shown are representatives of two (C) and three (A,B) experiments. Fig. 4 NLRC4 and NLRP3 have non-redundant roles in the host defense against sub-lethal murine S. Typhimurium (STm) infection. WT, Nlrc4 /, Nlrp3 / and casp-1 / (casp-11 / ) mice were infected intravenously (i.v) with CFUs STm strain M525P (a strain that establishes sub-lethal murine salmonellosis) and the mean bacterial load was determined in the spleen and liver after 1, 7 and 13 days of infection. Three to four animals of each genotype were 18
19 used per group per time point. Data are from one set of experiment representative of two. ns, no statistical significance, ** P < 0.01 and *** P < Fig. 5 NLRC4 and NLRP3 are recruited to the same Salmonella-induced ASC speck. (A) Unprimed WT primary bone marrow-derived macrophages (BMMs) were infected with S. Typhimurium (STm) expressing green fluorescent protein (GFP) for 2 or 24 h, and stained for ASC and DNA. The number of ASC foci per infected BMM was counted. At least 180 infected BMMs (indicated by the presence of GFP-expressing STm) were counted in each of the four independent experiments. (B) Bayesian localization super-resolution microscopy of endogenous NLRC4 and NLRP3 in THP-1 macrophages infected with STm. (C) Image analysis of the ring diameter of the NLRC4 and NLRP3 structures in B (n=10). Fig. 6 Caspase-8 is recruited to the same Salmonella-induced ASC speck. (A,B) Bayesian localization super-resolution microscopy of endogenous immuno-labeled ASC and FLICAstained caspase-8 in THP-1 macrophages infected with S. Typhimurium. (B) Multiple crosssections of the ASC Caspase-8 speck. Arrows indicate side filaments of ASC. Image analysis of the ring diameter of the ASC and caspase-8 structures (n=7). (C) A schematic of ASC speck assembly in response to Salmonella infection in a macrophage. 19
20
21
22
23
24
25
26 Supporting Information Materials and Methods Mice and infection Wildtype C57BL/6 mice (Harlan, Loughborough, UK), Nlrc4 / mice, Nlrp3 / mice, Asc / mice and Caspase-1(-11) / mice on the C57BL/6 background (all from Millennium Pharmaceuticals) were housed in a specific pathogen free facility according to the Animals Scientific Procedures outlined by the UK Home Office regulations. Six- to eight-week-old mice were infected with S. Typhimurium strain M525P grown overnight at 37 C in static culture in LB medium. Bacteria resuspended in PBS were injected into the tail veins of mice, and after 1, 7 and 13 days, killed by cervical dislocation. Spleens and livers were homogenized in a Colworth stomacher and the number of viable bacteria was determined by plating on LB agar. Bacterial infection, ligand stimulation and inhibitor study on primary bone marrow derived macrophages Primary bone marrow derived macrophages (BMMs) were cultivated in DMEM with 5 mm L-glutamine (Sigma), 10% FCS (Hyclone), 20% L929 conditioned media and 10 µg/ml gentamicin (Sigma) for 5 6 days before use. BMM were seeded in antibiotic free media at a concentration of cells onto 96-well plates and incubated overnight. THP-1 monocytes were grown in RPMI 1640 supplemented with 10% FCS, 5 mm L-glutamine 1% penicillin and streptomycin (10,000 units/ml) and 20 µm 2-mercaptoethanol. THP-1 cells were differentiated in Phorbol 12-myristate 13-acetate supplemented (250 nm, Sigma) media for 3 days. S. Typhimurium SL1344 was inoculated into LB broth and incubated for 17.5 h and subcultured (1:10) into fresh LB broth for 2 h. BMMs were infected with S. Typhimurium using the indicated Multiplicities of Infection (MOI). Bacteria were plated onto LB agar plates to confirm the number of inoculated bacteria. For 2 h infection, supernatant were removed after 1 h and replaced with media containing 50 µg/ml gentamicin (Sigma) for 1 h to kill extracellular bacteria. For 6 and 24 h infection, media containing 10 µg/ml gentamicin was used after 1 h incubation in media containing 50 µg/ml gentamicin. In experiments which required LPS priming, macrophages were stimulated using 200 ng/ml of ultrapure LPS
27 from E. coli (Invivogen) for 3 h. To activate the canonical NLRP3 inflammasome, 5 µm nigericin (Sigma) was used to stimulate cells for 30 min. Cytokine measurement Levels of mouse IL-1β secreted into the supernatant were measured using the BD OptEIA Mouse IL-1β Set according to the manufacturer s instructions. Transfection HEK293 cells were grown in DMEM (Sigma) supplemented with 10% FCS (Hyclone), 5 mm L-glutamine (Sigma) and penicillin and streptomycin (Sigma). Cells were seeded on cell-culture treated glass dishes (VWR) at a density of cells. HEK293 cells were transfected the next day with constructs encoding human NLRC4-HA, NLRP3-FLAG, ASC- FLAG, Caspase-1 or with the empty vector pcdna3. The ASC-FLAG construct was a gift from Emma Creagh (Trinity College Dublin, Ireland). Transfection was performed using 3 µl of Genejuice transfection reagent (Merck) per µg of DNA. Cells were incubated for 48 h. Immunofluorescence staining Macrophages or HEK293 cells were washed twice with PBS, fixed in -30 C methanol for 5 min or 4% (w/v) paraformaldehyde for 15 min. Blocking was performed using 10% normal goat serum or rabbit serum (Dako) in 0.1% saponin (Sigma) for 1 h. Cells were stained with primary and secondary antibodies for 50 min or overnight and 40 min, respectively. The primary antibodies used were rabbit anti-asc antibody (AL177; ENZO), mouse anti-mouse ASC antibody (clone 2EI-7; Millipore), rabbit anti-mouse cleaved caspase-8 (#5892; Cell Signaling Technology), goat anti-mouse IL1-β (AF-401-NA; R&D Systems), rabbit antihuman caspase-1 (sc-515; Santa Cruz Biotechnology) mouse anti-nlrp3 antibody (ALX C100; ENZO), rabbit anti-nlrc4 (gift from Gabriel Núñez), goat anti-nlrp3 (ab4207; Abcam), mouse or rat anti-flag antibody (Sigma) and mouse or rabbit anti-ha antibody (Sigma). The secondary antibodies used were Alexa Fluor 488, 568 or 647, antirabbit, anti-mouse, anti-goat or anti-rat IgG (Invitrogen). Cells were counterstained in DAPI mounting medium (Vecta Labs). To stain for active caspases, the supernatant from macrophages was removed following stimulation and replaced with media which contain
28 0.5 reconstituted FLICA active caspase-1 (FAM-YVAD-FMK), caspase-8 (FAM-IETD- FMK) or polycaspases (FAM-VAD-FMK) reagents (immunochemistry) and incubated for an additional 1 h. Cells were then fixed in 4% (w/v) paraformaldehyde and stained as described above. Cells and inflammasomes were visualized, counted and imaged using the Leica DM6000 B fluorescence microscope or the Leica TCS SP5 confocal microscope. Super-resolution microscopy We used Bayesian localization microscopy (34) to obtain super-resolution images of the ASC inflammasome structure in fixed cells. Macrophages were washed twice with PBS and fixed in 4% (w/v) paraformaldehyde for 15 min. Cells were permeabilised with 0.2% TritonX-100 in PBS for 20 min. Blocking was performed using 1% BSA and 0.1% Triton X-100 in PBS for 30 min. Cells were stained with primary and secondary antibodies as outlined above in 1% BSA and 0.1% Triton X-100 in PBS for overnight and 1 h, respectively. Super-resolution experiments were performed on a Nikon TI eclipse inverted microscope using a Nikon intensilight light source for fluorescence. Cells were imaged through an oil immersion objective ( 60, Numerical aperture 1.4) and a 2.5 Nikon relay lens. Images were collected on a low-noise Andor ixon EMCCD camera (pixel pitch 16 µm) at a frame rate of 43 frames per second. We applied the Bayesian localization analysis to sequences of 300 frames, which corresponds to an acquisition time of 7s. Western blotting Proteins from cell culture supernatants were precipitated using the methanol chloroform method as described.(20) Cell lysates were prepared by adding cell lysis buffer (150 mm NaCl, 50 mm Tris-HCl ph8, 1% Triton X-100, 1 mm PMSF, 10 µg/ml leupeptin, 1 µg/ml aprotinin) to cells and incubated for 10 min on ice before collecting the supernatant by centrifugation. Samples were separated by 4 20% gradient SDS PAGE and transferred onto PVDF membranes. The primary antibodies used were rabbit anti-mouse caspase-1 p10 (sc- 514; Santa Cruz Biotechnology), goat anti-mouse IL1-β (AF-401-NA; R&D Systems) and mouse anti-β-actin monoclonal antibody (ab3280; Abcam). The secondary antibodies used were goat anti-rabbit IgG-HRP, rabbit anti-goat IgG-HRP (sc-2004 and sc-2922, respectively; Santa Cruz Biotechnology) or polyclonal goat anti-mouse IgG-HRP (P0447; DAKO).
29 Statistical analysis Statistical significance between values from two groups was determined using unpaired Student s t-test, and values between three or more groups was determined using One-way Analysis of Variance (ANOVA) with all values corrected using the Tukey-Kramer Multiple Comparisons Test (GraphPad Software). P<0.05 is considered significant. SUPPLEMENTARY FIGURE LEGENDS Fig. S1. Endogenous ASC forms a speck-like structure following Salmonella infection (related to Fig. 1). (A) Unprimed wildtype primary bone marrow derived macrophages (BMMs) were stained for ASC (red) and DNA (blue). (B) Endogenous ASC redistributed to form a distinct 1-µm pore shaped structure in THP-1 cells stimulated with intracellularlydelivered Salmonella flagellin. (C) Bayesian localization super-resolution microscopy of endogenous ASC immuno-labeled using two different antibodies from two different commercial suppliers in BMMs infected with STm. (D) The morphology of endogenous ASC caspase-1 specks visualized by Bayesian localization super-resolution microscopy in primary mouse BMDMs infected with S. Typhimurium. Fig. S2. Analysis of the in-focus and out-of-focus point spread functions of an ASCcaspase-1 speck using Bayesian localization microscopy. This analysis shows (A) the infocus fluorophores tending to be localized to the edge of the ring, with (B) the out-of-focus fluorophores being more spread out. Fig. S3. Caspase-1 is localized within a larger NLRC4 ring-like structure (related to Fig. 2). Image analysis of the ring diameter of the NLRC4 and caspase-1 structures observed by Bayesian localization super-resolution microscopy (n=8). Fig. S4. NLRP3 resides in the ASC speck following stimulation of the canonical NLRP3 inflammasome in macrophages. Bayesian localization super-resolution microscopy of
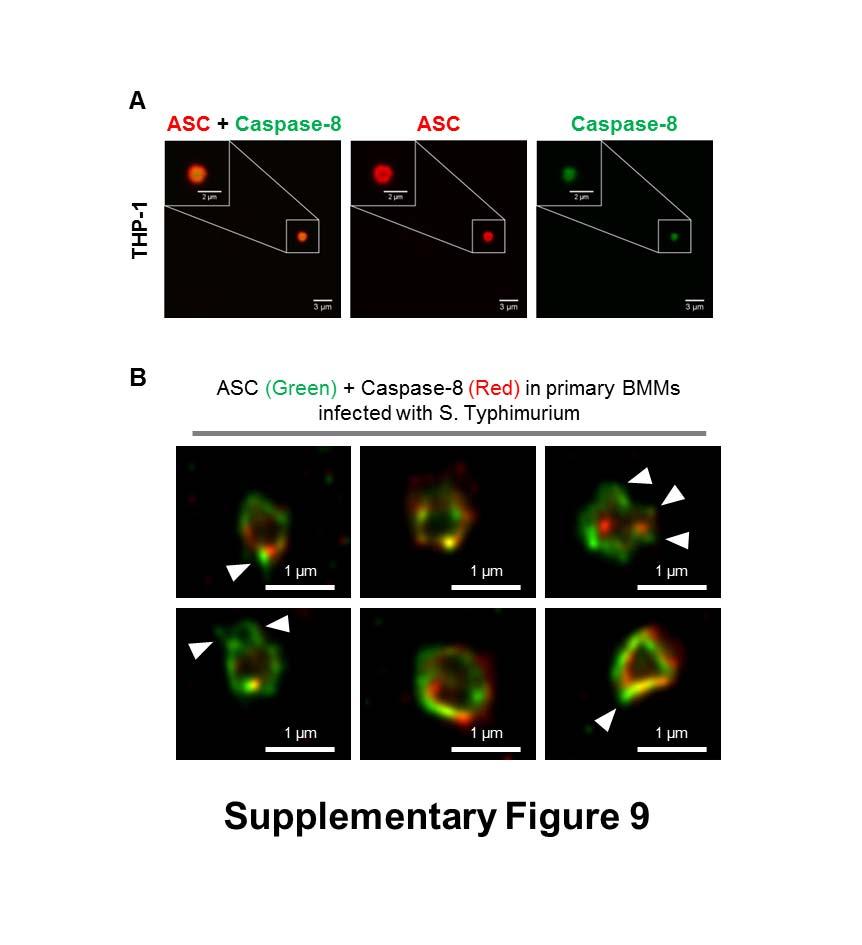
30 endogenous immuno-labeled NLRP3 and ASC in human THP-1 macrophages stimulated with 5 µm of nigericin for 30 min. Fig. S5. ASC and caspase-1 colocalize in a transfection system (related to Fig. 1). HEK293 cells transfected with hasc and hcaspase-1 DNA constructs. Cells were immunostained for the cytosolic distribution of these proteins. Fig. S6. NLRs and ASC colocalize in a transfection system (related to Fig. 2.). (A) HEK293 cells transfected with hnlrp3 alone or with hnlrp3 and hasc DNA constructs. (B) HEK293 cells transfected with hnlrc4 alone or with hnlrc4 and hasc DNA constructs. (C) HEK293 cells transfected with hnlrc4 and hcaspase-1 DNA constructs. Cells were immuno-stained for the cytosolic distribution of these proteins. Fig. S7. ASC is required for caspase-1 proteolysis in Salmonella infection (related to Fig. 3). Wildtype, Asc /, caspase-1 / (caspase-11 / ) LPS-primed primary bone marrow derived macrophages with S. Typhimurium (MOI 10) for 1 h. Cleaved IL-1β p17 subunit (processed IL-1β), cleaved caspase-1 p10 subunit (active caspase-1), pro-il-1β, pro-caspase- 1 and β-actin in the supernatant or cell lysate were detected by western blot analysis. Fig. S8. NLRC4 and NLRP3 colocalize in a transfection system (related to Fig. 5). HEK293 cells transfected with hnlrc4 and hnlrp3 DNA constructs. Cells were immunostained for the cytosolic distribution of these proteins. Fig. S9. Endogenous Caspase-8 is recruited to the same Salmonella-induced ASC speck (related to Fig. 6). (A) Confocal microscopy imaging of endogenous immuno-labeled ASC and FLICA staining of caspase-8 in human THP-1 macrophages infected with STm. (B) Bayesian localization super-resolution microscopy of endogenous immuno-labeled ASC and caspase-8 in primary mouse bone marrow-derived macrophages infected with STm. Multiple cross-sections of the ASC Caspase-8 speck. Arrows indicate side filaments of ASC coming off the external region of the ring-like structure.
31 Fig. S10. The endogenous ASC speck recruits caspase-1 and pro-il-1β. LPS-primed wildtype primary bone marrow derived macrophages (BMMs) were infected with STm for 30 min in the presence of caspase-1 FLICA stain (Green) and immuno-stained for ASC (magenta), pro-il-1β/il-1β (red) and DNA (blue).
32
33
34
35
36
37
38
39
40

41
A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System
 Cell Host & Microbe, Volume 7 Supplemental Information A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System Igor E. Brodsky, Noah
Cell Host & Microbe, Volume 7 Supplemental Information A Yersinia-Secreted Effector Protein Promotes Virulence by Preventing Inflammasome Recognition of the Type III Secretion System Igor E. Brodsky, Noah
Understanding the Role of the Inflammasome in Inflammation
 Understanding the Role of the Inflammasome in Inflammation Martha O Brien, Ph.D. Assay Design Group 215 Innate Immune Response in Inflammation LPS flagellin TLR-1/2/6 TLR-4 TLR-5 Adaptor proteins PAMPs,
Understanding the Role of the Inflammasome in Inflammation Martha O Brien, Ph.D. Assay Design Group 215 Innate Immune Response in Inflammation LPS flagellin TLR-1/2/6 TLR-4 TLR-5 Adaptor proteins PAMPs,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11429 S1a 6 7 8 9 Nlrc4 allele S1b Nlrc4 +/+ Nlrc4 +/F Nlrc4 F/F 9 Targeting construct 422 bp 273 bp FRT-neo-gb-PGK-FRT 3x.STOP S1c Nlrc4 +/+ Nlrc4 F/F casp1
SUPPLEMENTARY INFORMATION doi:10.1038/nature11429 S1a 6 7 8 9 Nlrc4 allele S1b Nlrc4 +/+ Nlrc4 +/F Nlrc4 F/F 9 Targeting construct 422 bp 273 bp FRT-neo-gb-PGK-FRT 3x.STOP S1c Nlrc4 +/+ Nlrc4 F/F casp1
Type of file: PDF Title of file for HTML: Supplementary Information Description: Supplementary Figures
 Type of file: PDF Title of file for HTML: Supplementary Information Description: Supplementary Figures Type of file: MOV Title of file for HTML: Supplementary Movie 1 Description: NLRP3 is moving along
Type of file: PDF Title of file for HTML: Supplementary Information Description: Supplementary Figures Type of file: MOV Title of file for HTML: Supplementary Movie 1 Description: NLRP3 is moving along
Innate Immunity & Inflammation
 Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
Journal club. Lama Nazzal
 Journal club Lama Nazzal Background Kidney stone disease affects about 12% of men and 5% of women during their lifetimes in the United States Intrarenal nephrocalcinosis is often asymptomatic, but can
Journal club Lama Nazzal Background Kidney stone disease affects about 12% of men and 5% of women during their lifetimes in the United States Intrarenal nephrocalcinosis is often asymptomatic, but can
Gout and Nucleic Acid Metabolism Vol.33 No
 Gout and Nucleic Acid Metabolism Vol.33 No.1 2009 1 1 2 3 in vitro 14 IgM 1 IgM IgM 1 PAMPs Pattern recognition receptors PRRs PRRs PRRs PAMPs Toll Toll-like receptor TLR PAMPs Nod Nod-like receptor NLR
Gout and Nucleic Acid Metabolism Vol.33 No.1 2009 1 1 2 3 in vitro 14 IgM 1 IgM IgM 1 PAMPs Pattern recognition receptors PRRs PRRs PRRs PAMPs Toll Toll-like receptor TLR PAMPs Nod Nod-like receptor NLR
Mice. Mice genetically deficient for IRAK-1, IRAK-2, IRAK-1/2, MyD88, TRIF, NLRP3,
 Supporting Information SI Text Mice. Mice genetically deficient for IRAK-1, IRAK-2, IRAK-1/2, MyD88, TRIF, NLRP3, MyD88/TRIF, TLR2/TLR4, TLR7, caspase-1, IL-1R1 and IL-18R, control C57BL/6 mice as well
Supporting Information SI Text Mice. Mice genetically deficient for IRAK-1, IRAK-2, IRAK-1/2, MyD88, TRIF, NLRP3, MyD88/TRIF, TLR2/TLR4, TLR7, caspase-1, IL-1R1 and IL-18R, control C57BL/6 mice as well
Supplementary information
 Supplementary information Supplementary Figure S1: Ex[Ca 2+ ]-induced IL-1ß production of monocytes primed with different TLR ligands IL-1ß release of CD14+ monocytes in response to stimulation for 16
Supplementary information Supplementary Figure S1: Ex[Ca 2+ ]-induced IL-1ß production of monocytes primed with different TLR ligands IL-1ß release of CD14+ monocytes in response to stimulation for 16
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration
 Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
TLR2 Signaling Contributes to Rapid Inflammasome Activation during F. novicida Infection
 TLR2 Signaling Contributes to Rapid Inflammasome Activation during F. novicida Infection Crystal L. Jones, Emory University David S Weiss, Emory University Journal Title: PLoS ONE Volume: Volume 6, Number
TLR2 Signaling Contributes to Rapid Inflammasome Activation during F. novicida Infection Crystal L. Jones, Emory University David S Weiss, Emory University Journal Title: PLoS ONE Volume: Volume 6, Number
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
LPS LPS P6 - + Supplementary Fig. 1.
 P6 LPS - - - + + + - LPS + + - - P6 + Supplementary Fig. 1. Pharmacological inhibition of the JAK/STAT blocks LPS-induced HMGB1 nuclear translocation. RAW 267.4 cells were stimulated with LPS in the absence
P6 LPS - - - + + + - LPS + + - - P6 + Supplementary Fig. 1. Pharmacological inhibition of the JAK/STAT blocks LPS-induced HMGB1 nuclear translocation. RAW 267.4 cells were stimulated with LPS in the absence
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Caspase-11 Activation in Response to Bacterial Secretion Systems That Access the Host Cytosol
 University of Pennsylvania ScholarlyCommons Department of Microbiology Perelman School of Medicine 6-6-2013 Caspase-11 Activation in Response to Bacterial Secretion Systems That Access the Host Cytosol
University of Pennsylvania ScholarlyCommons Department of Microbiology Perelman School of Medicine 6-6-2013 Caspase-11 Activation in Response to Bacterial Secretion Systems That Access the Host Cytosol
Structure-function analysis of Nlrp1b reveals a link. between metabolism and inflammation: energy stress. triggers inflammasome activation
 Structure-function analysis of Nlrp1b reveals a link between metabolism and inflammation: energy stress triggers inflammasome activation by Kuo-Chieh Liao A thesis submitted in conformity with the requirements
Structure-function analysis of Nlrp1b reveals a link between metabolism and inflammation: energy stress triggers inflammasome activation by Kuo-Chieh Liao A thesis submitted in conformity with the requirements
Innate immunity. Abul K. Abbas University of California San Francisco. FOCiS
 1 Innate immunity Abul K. Abbas University of California San Francisco FOCiS 2 Lecture outline Components of innate immunity Recognition of microbes and dead cells Toll Like Receptors NOD Like Receptors/Inflammasome
1 Innate immunity Abul K. Abbas University of California San Francisco FOCiS 2 Lecture outline Components of innate immunity Recognition of microbes and dead cells Toll Like Receptors NOD Like Receptors/Inflammasome
The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1b secretion but dispensable for adjuvant activity
 Eur. J. Immunol. 2008. 38: 2085 2089 DOI 10.1002/eji.200838549 Highlights 2085 The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1b secretion but dispensable for adjuvant activity
Eur. J. Immunol. 2008. 38: 2085 2089 DOI 10.1002/eji.200838549 Highlights 2085 The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1b secretion but dispensable for adjuvant activity
Human SH-SY5Y neuroblastoma cells (A.T.C.C., Manassas, VA) were cultured in DMEM, F-12
 were cultured in DMEM, F-12") SUPPLEMENTARY METHODS Cell cultures Human SH-SY5Y neuroblastoma cells (A.T.C.C., Manassas, VA) were cultured in DMEM, F-12 Ham with 25 mm HEPES and NaHCO 3 (1:1) and supplemented with 10% (v/v) FBS, 1.0
SUPPLEMENTARY METHODS Cell cultures Human SH-SY5Y neuroblastoma cells (A.T.C.C., Manassas, VA) were cultured in DMEM, F-12 Ham with 25 mm HEPES and NaHCO 3 (1:1) and supplemented with 10% (v/v) FBS, 1.0
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
2. Innate immunity 2013
 1 Innate Immune Responses 3 Innate immunity Abul K. Abbas University of California San Francisco The initial responses to: 1. Microbes: essential early mechanisms to prevent, control, or eliminate infection;
1 Innate Immune Responses 3 Innate immunity Abul K. Abbas University of California San Francisco The initial responses to: 1. Microbes: essential early mechanisms to prevent, control, or eliminate infection;
TLR2/MyD88/NF-kB Pathway, Reactive Oxygen Species, Potassium Efflux Activates NLRP3/ASC Inflammasome during Respiratory Syncytial Virus Infection
 TLR2/MyD88/NF-kB Pathway, Reactive Oxygen Species, Potassium Efflux Activates NLRP3/ASC Inflammasome during Respiratory Syncytial Virus Infection Jesus Segovia 1., Ahmed Sabbah 1., Victoria Mgbemena 1,
TLR2/MyD88/NF-kB Pathway, Reactive Oxygen Species, Potassium Efflux Activates NLRP3/ASC Inflammasome during Respiratory Syncytial Virus Infection Jesus Segovia 1., Ahmed Sabbah 1., Victoria Mgbemena 1,
Sensing and reacting to microbes through the inflammasomes
 The inflammasome Sensing and reacting to microbes through the inflammasomes Luigi Franchi, Raul Muñoz-Planillo & Gabriel Núñez Inflammasomes are multiprotein complexes that activate caspase-1, which leads
The inflammasome Sensing and reacting to microbes through the inflammasomes Luigi Franchi, Raul Muñoz-Planillo & Gabriel Núñez Inflammasomes are multiprotein complexes that activate caspase-1, which leads
Supplementary Figure 1.
 Supplementary Figure 1. Increased β cell mass and islet diameter in βtsc2 -/- mice up to 35 weeks A: Reconstruction of multiple anti-insulin immunofluorescence images showing differences in β cell mass
Supplementary Figure 1. Increased β cell mass and islet diameter in βtsc2 -/- mice up to 35 weeks A: Reconstruction of multiple anti-insulin immunofluorescence images showing differences in β cell mass
Nature Immunology doi: /ni.3268
 Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
THE ROLE OF AIM2 IN INFLAMMATORY GENE EXPRESSION
 MQP-JXR-1218 THE ROLE OF AIM2 IN INFLAMMATORY GENE EXPRESSION A Major Qualifying Project Report submitted to the Faculty of WORCESTER POLYTECHNIC INSTITUTE in partial fulfillment of the requirements for
MQP-JXR-1218 THE ROLE OF AIM2 IN INFLAMMATORY GENE EXPRESSION A Major Qualifying Project Report submitted to the Faculty of WORCESTER POLYTECHNIC INSTITUTE in partial fulfillment of the requirements for
Comparison of Two Mice Strains, A/J and C57BL/6, in Caspase-1 Activity and IL-1
 Mediators of Inflammation Volume, Article ID 7873, 5 pages doi:.55//7873 Research Article Comparison of Two Mice Strains, A/J and C57BL/6, in Caspase- Activity and IL-β SecretionofMacrophage to Mycobacterium
Mediators of Inflammation Volume, Article ID 7873, 5 pages doi:.55//7873 Research Article Comparison of Two Mice Strains, A/J and C57BL/6, in Caspase- Activity and IL-β SecretionofMacrophage to Mycobacterium
Intrinsic cellular defenses against virus infection
 Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Newly Recognized Components of the Innate Immune System
 Newly Recognized Components of the Innate Immune System NOD Proteins: Intracellular Peptidoglycan Sensors NOD-1 NOD-2 Nod Protein LRR; Ligand Recognition CARD RICK I-κB p50 p65 NF-κB Polymorphisms in Nod-2
Newly Recognized Components of the Innate Immune System NOD Proteins: Intracellular Peptidoglycan Sensors NOD-1 NOD-2 Nod Protein LRR; Ligand Recognition CARD RICK I-κB p50 p65 NF-κB Polymorphisms in Nod-2
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Material
 Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Supplementary Material accompanying the manuscript Interleukin 37 is a fundamental inhibitor of innate immunity Marcel F Nold, Claudia A Nold-Petry, Jarod A Zepp, Brent E Palmer, Philip Bufler & Charles
Supplementary Methods
 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16 17 18 19 2 21 22 23 24 25 26 27 28 29 3 31 32 Supplementary Methods Gentamicin protection assay THP-1 monocytes were seeded onto a 12-well plate and differentiated
1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 16 17 18 19 2 21 22 23 24 25 26 27 28 29 3 31 32 Supplementary Methods Gentamicin protection assay THP-1 monocytes were seeded onto a 12-well plate and differentiated
Asc and Ipaf Inflammasomes Direct Distinct Pathways for Caspase-1 Activation in Response to Legionella pneumophila
 INFECTION AND IMMUNITY, May 2009, p. 1981 1991 Vol. 77, No. 5 0019-9567/09/$08.00 0 doi:10.1128/iai.01382-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Asc and Ipaf Inflammasomes
INFECTION AND IMMUNITY, May 2009, p. 1981 1991 Vol. 77, No. 5 0019-9567/09/$08.00 0 doi:10.1128/iai.01382-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Asc and Ipaf Inflammasomes
Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection
 Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection Victoria L. Smith, Yong Cheng, Barry R. Bryant and Jeffrey S. Schorey Supplementary Figure 1: Unprocessed
Exosomes function in antigen presentation during an in vivo Mycobacterium tuberculosis infection Victoria L. Smith, Yong Cheng, Barry R. Bryant and Jeffrey S. Schorey Supplementary Figure 1: Unprocessed
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Role of Innate Immunity in Control of Adaptive Immunity
 Role of Innate Immunity in Control of Adaptive Immunity Innate Immunity The burden of pathogen sensing is placed on the innate immune system Danger hypothesis Missing Self Based on the detection of molecular
Role of Innate Immunity in Control of Adaptive Immunity Innate Immunity The burden of pathogen sensing is placed on the innate immune system Danger hypothesis Missing Self Based on the detection of molecular
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH
 FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Inflammation - Molecular events
 Academic lectures for students of medical schools 3rd Year updated 2004-2015 GENERAL PATHOPHYSIOLOGY Inflammation - Molecular events R. A. Benacka, MD, PhD Department of Pathophysiology Medical faculty
Academic lectures for students of medical schools 3rd Year updated 2004-2015 GENERAL PATHOPHYSIOLOGY Inflammation - Molecular events R. A. Benacka, MD, PhD Department of Pathophysiology Medical faculty
Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC
 Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC Supplementary Table 2. Drug content and loading efficiency estimated with F-NMR and UV- Vis Supplementary Table 3. Complete
Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC Supplementary Table 2. Drug content and loading efficiency estimated with F-NMR and UV- Vis Supplementary Table 3. Complete
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
F-actin VWF Vinculin. F-actin. Vinculin VWF
 a F-actin VWF Vinculin b F-actin VWF Vinculin Supplementary Fig. 1. WPBs in HUVECs are located along stress fibers and at focal adhesions. (a) Immunofluorescence images of f-actin (cyan), VWF (yellow),
a F-actin VWF Vinculin b F-actin VWF Vinculin Supplementary Fig. 1. WPBs in HUVECs are located along stress fibers and at focal adhesions. (a) Immunofluorescence images of f-actin (cyan), VWF (yellow),
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Masmudur M. Rahman and Grant McFadden* Department of Molecular Genetics and Microbiology, University of Florida, Gainesville, Florida 32610
 JOURNAL OF VIROLOGY, Dec. 2011, p. 12505 12517 Vol. 85, No. 23 0022-538X/11/$12.00 doi:10.1128/jvi.00410-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Myxoma Virus Lacking
JOURNAL OF VIROLOGY, Dec. 2011, p. 12505 12517 Vol. 85, No. 23 0022-538X/11/$12.00 doi:10.1128/jvi.00410-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Myxoma Virus Lacking
Molecular Mechanisms in Inflammasome Signalling: From NLRP1 to Pyroptosis. Sebastian Hiller
 Molecular Mechanisms in Inflammasome Signalling: From NLRP1 to Pyroptosis Sebastian Hiller How do host cells detect pathogens? The inflammasome: a caspase-1 activation platform Inflammasome receptors:
Molecular Mechanisms in Inflammasome Signalling: From NLRP1 to Pyroptosis Sebastian Hiller How do host cells detect pathogens? The inflammasome: a caspase-1 activation platform Inflammasome receptors:
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Within the gastrointestinal (GI) tract, innate immune REVIEWS IN BASIC AND CLINICAL GASTROENTEROLOGY AND HEPATOLOGY
 tract, innate immune REVIEWS IN BASIC AND CLINICAL GASTROENTEROLOGY AND HEPATOLOGY") GASTROENTEROLOGY 2011;141:1986 1999 IN BASIC CLINICAL GASTROENTEROLOGY HEPATOLOGY Robert F. Schwabe and John W. Wiley, Section Editors Inflammasomes in Intestinal Inflammation and Cancer GRACE Y. CHEN*
GASTROENTEROLOGY 2011;141:1986 1999 IN BASIC CLINICAL GASTROENTEROLOGY HEPATOLOGY Robert F. Schwabe and John W. Wiley, Section Editors Inflammasomes in Intestinal Inflammation and Cancer GRACE Y. CHEN*
NOD-like Receptors (NLRs) and Inflammasomes
 and Inflammasomes") International Edition NOD-like Receptors (NLRs) and Inflammasomes In mammals, germ-line encoded pattern recognition receptors (PRRs) detect the presence of pathogens through recognition of pathogen-associated
International Edition NOD-like Receptors (NLRs) and Inflammasomes In mammals, germ-line encoded pattern recognition receptors (PRRs) detect the presence of pathogens through recognition of pathogen-associated
Page 39 of 44. 8h LTA & AT h PepG & AT h LTA
 Page 39 of 44 Fig. S1 A: B: C: D: 8h LTA 8h LTA & AT7519 E: F: 8h PepG G: 8h PepG & AT7519 Fig. S1. AT7519 overrides the survival effects of lipoteichoic acid (LTA) and peptidoglycan (PepG). (A) Human
Page 39 of 44 Fig. S1 A: B: C: D: 8h LTA 8h LTA & AT7519 E: F: 8h PepG G: 8h PepG & AT7519 Fig. S1. AT7519 overrides the survival effects of lipoteichoic acid (LTA) and peptidoglycan (PepG). (A) Human
Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival
 Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Identification of ethyl pyruvate as a NLRP3 inflammasome inhibitor that preserves mitochondrial integrity
 Li et al. Molecular Medicine (2018) 24:8 https://doi.org/10.1186/s10020-018-0006-9 Molecular Medicine RESEARCH ARTICLE Open Access Identification of ethyl pyruvate as a NLRP3 inflammasome inhibitor that
Li et al. Molecular Medicine (2018) 24:8 https://doi.org/10.1186/s10020-018-0006-9 Molecular Medicine RESEARCH ARTICLE Open Access Identification of ethyl pyruvate as a NLRP3 inflammasome inhibitor that
Supporting Information. FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce. apoptosis
 1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
a b G75 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server.
 a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
- 1 - Cell types Monocytes THP-1 cells Macrophages. LPS Treatment time (Hour) IL-6 level (pg/ml)
 IL-6 level (pg/ml)") Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the
 Schematic of Crb3 isoforms, ERLI and CLPI, indicating the") Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Doctoral Degree Program in Marine Biotechnology, College of Marine Sciences, Doctoral Degree Program in Marine Biotechnology, Academia Sinica, Taipei,
 Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
Supplementary Figure 1: Co-localization of reconstituted L-PTC and dendritic cells
 a CD11c Na + K + ATPase Na + K + ATPase CD11c x-y CD11c Na + K + ATPase Na + K + ATPase CD11c x-z c b x-y view BoNT NAPs CD11c BoNT CD11c NAPs BoNT NAPs CD11c 90 x-z view Apical Basolateral Supplementary
a CD11c Na + K + ATPase Na + K + ATPase CD11c x-y CD11c Na + K + ATPase Na + K + ATPase CD11c x-z c b x-y view BoNT NAPs CD11c BoNT CD11c NAPs BoNT NAPs CD11c 90 x-z view Apical Basolateral Supplementary
TCR, MHC and coreceptors
 Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
The oxidized phospholipid oxpapc protects from septic shock by targeting the non-canonical inflammasome in macrophages
 ARTICLE DOI: 1.138/s41467-18-39-3 OPEN The oxidized phospholipid protects from septic shock by targeting the non-canonical inflammasome in macrophages Lan H. Chu 1,2, Mohanalaxmi Indramohan 1, Rojo A.
ARTICLE DOI: 1.138/s41467-18-39-3 OPEN The oxidized phospholipid protects from septic shock by targeting the non-canonical inflammasome in macrophages Lan H. Chu 1,2, Mohanalaxmi Indramohan 1, Rojo A.
Toll-like Receptor Signaling
 Toll-like Receptor Signaling 1 Professor of Medicine University of Massachusetts Medical School, Worcester, MA, USA Why do we need innate immunity? Pathogens multiply very fast We literally swim in viruses
Toll-like Receptor Signaling 1 Professor of Medicine University of Massachusetts Medical School, Worcester, MA, USA Why do we need innate immunity? Pathogens multiply very fast We literally swim in viruses
Identification of Microbes
 Identification of Microbes Recognition by PRR (pattern recognition receptors) Recognize conserved molecular patterns on microbes called pathogen associated molecular patterns (PAMPs) which are not present
Identification of Microbes Recognition by PRR (pattern recognition receptors) Recognize conserved molecular patterns on microbes called pathogen associated molecular patterns (PAMPs) which are not present
Fang et al. NMuMG. PyVmT unstained Anti-CCR2-PE MDA-MB MCF MCF10A
 A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small cell lung cancer
, NCI-H460 (human non-small cell lung cancer") Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Evaluation of directed and random motility in microslides Assessment of leukocyte adhesion in flow chambers
 Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Supporting Information
 Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Supporting Information Pang et al. 10.1073/pnas.1322009111 SI Materials and Methods ELISAs. These assays were performed as previously described (1). ELISA plates (MaxiSorp Nunc; Thermo Fisher Scientific)
Prolonged mitotic arrest induces a caspase-dependent DNA damage
 SUPPLEMENTARY INFORMATION Prolonged mitotic arrest induces a caspase-dependent DNA damage response at telomeres that determines cell survival Karolina O. Hain, Didier J. Colin, Shubhra Rastogi, Lindsey
SUPPLEMENTARY INFORMATION Prolonged mitotic arrest induces a caspase-dependent DNA damage response at telomeres that determines cell survival Karolina O. Hain, Didier J. Colin, Shubhra Rastogi, Lindsey
Innate Immunity II. Integration. Lindsay Nicholson Advanced Immunology L2
 Innate Immunity II Integration Lindsay Nicholson Advanced Immunology L2 l.nicholson@bristol.ac.uk Lecture 1 Defining Innate Immunity Recognition and effector mechanisms (I) Lecture 2 Recognition and effector
Innate Immunity II Integration Lindsay Nicholson Advanced Immunology L2 l.nicholson@bristol.ac.uk Lecture 1 Defining Innate Immunity Recognition and effector mechanisms (I) Lecture 2 Recognition and effector
Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines
 CORRECTION NOTICE Nat. Med. doi:10.1038/nm.3547; corrected online 25 August 2014 Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines Christine Schauer, Christina
CORRECTION NOTICE Nat. Med. doi:10.1038/nm.3547; corrected online 25 August 2014 Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines Christine Schauer, Christina
Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death
 www.impactjournals.com/oncotarget/ Oncotarget, Supplementary Materials 2016 Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death Supplementary
www.impactjournals.com/oncotarget/ Oncotarget, Supplementary Materials 2016 Tumor suppressor Spred2 interaction with LC3 promotes autophagosome maturation and induces autophagy-dependent cell death Supplementary
Immunology - Lecture 2 Adaptive Immune System 1
 Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Supplementary Figure 1
 Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
Primer sequences Target Sequence F Sequence R TNF-α (Tnfa) TCAGCCGATTTGCTATCTCAT A
 TCAGCCGATTTGCTATCTCAT A") Supplementary Table 1. Q- and RT-PR primers used in this study. Primer sequences Target Sequence F Sequence R TNF-α (Tnfa) TGGTTTGTTTT GTTTGGGGTTG T hemokine (- motif) ligand 5 (cl5) GTGTTTGTTT TGGTGGTG
Supplementary Table 1. Q- and RT-PR primers used in this study. Primer sequences Target Sequence F Sequence R TNF-α (Tnfa) TGGTTTGTTTT GTTTGGGGTTG T hemokine (- motif) ligand 5 (cl5) GTGTTTGTTT TGGTGGTG
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
INFLAMMASOME IN INTESTINAL INFLAMMATION AND CANCER. Laura Stronati ENEA - Roma
 INFLAMMASOME IN INTESTINAL INFLAMMATION AND CANCER Laura Stronati ENEA - Roma Inflammasome: definition, components and activation TLRs NODs RLRs CLRs PRRs LPS, Flagellin, DNA, RNA, etc) PAMPs DAMPs ATP,
INFLAMMASOME IN INTESTINAL INFLAMMATION AND CANCER Laura Stronati ENEA - Roma Inflammasome: definition, components and activation TLRs NODs RLRs CLRs PRRs LPS, Flagellin, DNA, RNA, etc) PAMPs DAMPs ATP,
Cryopyrin and pyrin activate caspase-1, but not NF-jB, via ASC oligomerization
 (2006) 13, 236 249 & 2006 Nature Publishing Group All rights reserved 1350-9047/06 $30.00 www.nature.com/cdd Cryopyrin and pyrin activate caspase-1, but not NF-jB, via ASC oligomerization J-W Yu 1,3,JWu
(2006) 13, 236 249 & 2006 Nature Publishing Group All rights reserved 1350-9047/06 $30.00 www.nature.com/cdd Cryopyrin and pyrin activate caspase-1, but not NF-jB, via ASC oligomerization J-W Yu 1,3,JWu
COURSE: Medical Microbiology, MBIM 650/720 - Fall TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12
 COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS
: DEVELOPMENT OF ANTIVESICANT DRUGS") STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS Xiannu Jin 1, Radharaman Ray 2, Guang Xu 1 and Prabhati Ray 1 1 Department
STUDIES ON MUSTARD-STIMULATED PROTEASES AND INHIBITORS IN HUMAN EPIDERMAL KERATINOCYTES (HEK): DEVELOPMENT OF ANTIVESICANT DRUGS Xiannu Jin 1, Radharaman Ray 2, Guang Xu 1 and Prabhati Ray 1 1 Department
Immune receptors Y Y. Multiple immune responses CLR TLR NLR ITAM RLR. C-type lectin. ITAM: Immunoreceptor Tyrosine-based Activation Motif
 C Immune receptors Innate immunity Acquired immunity TLR CLR FcR BCR TCR C-type lectin Y Y Ig Ig NLR ITAM ITAM ITAM RLR Multiple immune responses ITAM: Immunoreceptor Tyrosine-based Activation Motif Immune
C Immune receptors Innate immunity Acquired immunity TLR CLR FcR BCR TCR C-type lectin Y Y Ig Ig NLR ITAM ITAM ITAM RLR Multiple immune responses ITAM: Immunoreceptor Tyrosine-based Activation Motif Immune
Supplemental Information. Autophagy in Oncogenic K-Ras. Promotes Basal Extrusion. of Epithelial Cells by Degrading S1P. Current Biology, Volume 24
 Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
Current Biology, Volume 24 Supplemental Information Autophagy in Oncogenic K-Ras Promotes Basal Extrusion of Epithelial Cells by Degrading S1P Gloria Slattum, Yapeng Gu, Roger Sabbadini, and Jody Rosenblatt
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes
 Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes Siew Leong Chan Protein Analytics, AbbVie Bioresearch Center Worcester MA Acknowledgement
Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes Siew Leong Chan Protein Analytics, AbbVie Bioresearch Center Worcester MA Acknowledgement
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth.
 Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
PKR is required for macrophage apoptosis after activation of TLR-4. By Christina, Ania and Xiaofu
 PKR is required for macrophage apoptosis after activation of TLR-4 By Christina, Ania and Xiaofu Introduction Macrophage PAMP Bacteria LPS Gram -ve LTA Gram +ve activation Clearance Macrophage PAMP Bacteria
PKR is required for macrophage apoptosis after activation of TLR-4 By Christina, Ania and Xiaofu Introduction Macrophage PAMP Bacteria LPS Gram -ve LTA Gram +ve activation Clearance Macrophage PAMP Bacteria
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Integrated Innate Immunity Combining Activation and Effector Functions
 Section 4 Integrated Innate Immunity Combining Activation and Effector Functions 4.1 THE DETECTION OF BACTERIAL LIPOPOLYSACCHARIDE LPS is one of the most potent immune stimuli known. It is found in the
Section 4 Integrated Innate Immunity Combining Activation and Effector Functions 4.1 THE DETECTION OF BACTERIAL LIPOPOLYSACCHARIDE LPS is one of the most potent immune stimuli known. It is found in the
Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).
 Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).") Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary