EBV essential antigen EBNA3C attenuates H2AX expression
|
|
|
- Joshua Booker
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 15 January 2014 J. Virol. doi: /jvi Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 EBV essential antigen EBNA3C attenuates H2AX expression Hem C Jha ±, Mahadesh Prasad AJ ±, Abhik Saha, Shuvomoy Banerjee, Jie Lu, and Erle S Robertson* Department of Microbiology and Tumor Virology Program, Abramson Cancer Center, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, PA-19104, USA ± Equal Contribution * Corresponding Author: E. mail: erle@upenn.edu Phone: ; Fax: Running Title: EBNA3C deregulates H2AX Keywords: EBNA3C; H2AX; Ubiquitination; Ser139 H2AX mutation; EBV infection; Cell proliferation 1
2 ABSTRACT EBV latent antigen EBNA3C is implicated in B-cell immortalization and linked to several B-cell malignancies. Deregulation of H2AX can induce genomic instability with increased chromosomal aberrations which ultimately leads to tumorigenesis. Here we demonstrated that EBNA3C can attenuate H2AX expression at the transcript and protein levels. A reduction of total H2AX levels were clearly observed on infection of primary B-cells with wild type EBV, but not with EBNA3C knockout EBV recombinant virus. H2AX also interacted with EBNA3C through its N-terminal domain (residues 1-100). Furthermore, H2AX mutated at Ser139 failed to interact with EBNA3C. Luciferase based reporter assays also revealed that the binding domain of EBNA3C is sufficient for transcriptional inhibition of the H2AX promoter. EBNA3C also facilitated H2AX degradation through recruitment of components of the ubiquitin proteasome pathway. We further demonstrated that knockdown of H2AX in LCLs led to up-regulation of the Bub1 oncoprotein, and down-regulated expression of the p53. Overall, our study provides additional insights into EBV-associated B-cell lymphomas which is linked to regulation of the DNA damage response system in the infected cells. Importance: I. EBNA3C down-regulates H2AX expression at the protein and transcript levels in epithelial cells, B-cells and EBV transformed LCLs. II. EBNA3C binds with wild type H2AX but not with the ser139 mutant of H2AX. III. The N-terminal (residue amino acid) of EBNA3C is critical for binding to H2AX. 2
3 IV. Localization of H2AX is predominantly nuclear in the presence of EBNA3C. V. H2AX knocked down in LCLs led to enhanced expression of Bub1 and downregulation of the tumor suppressor p53 both important for driving the oncogenic process
4 INTRODUCTION Epstein Barr Virus (EBV) is a human gammaherpesvirus associated with infectious mononucleosis, and it is estimated that more than 95% of adults are carriers of EBV throughout their lifetime (1, 2). The contributory role of EBV in driving the oncogenic process is continually being explored. EBV transforms latently infected primary B-cells into constantly proliferating lymphoblastoid cell lines (LCLs) (3). EBV is also commonly involved in numerous malignancies, including Burkitt s Lymphoma (BL), Post transplant lympho-proliferative disorders (PTLDs), Nasopharyngeal carcinoma (NPC), HIV associated lymphomas, some types of T cell lymphomas, and Gastric Cancer (4, 5). Transformation of human B-cells into LCLs by EBV establishes a latent type of infection typically known as type III latency (6). Three major viral latency programs have been described with deferential expression profiles of specific viral gene transcription (7). EBV latency patterns are characterized by the expression of different EBV nuclear antigens (EBNAs) including, EBNA 1, 2, 3A, 3B, 3C, LP/5, and the latent membrane proteins (LMP) 1, 2A and 2B (8). Importantly, these latent proteins are significantly expressed during the latency III program (9) (10). Previous studies showed that EBNA2, EBNA3A, EBNA3C and LMP1 play critical roles in B-cell transformation (11, 12). Earlier studies showed that one of the essential EBV latent antigens, EBNA3C is important for modulating B-cell activation. For example, the B-cell activation marker CD21 was up-regulated in the presence of EBNA3C in Burkitt s lymphoma cell lines (13, 14). EBNA3C binds to RBP-Jk, an important regulator of the Notch signaling pathway, through an amino terminal motif, and the acidic domains are responsible for nuclear 4
5 translocation due to the presence of the nuclear localization signals (15). Recently, we reported that the p53 tumor suppressor is negatively regulated by EBNA3C both at the transcriptional and post-transcriptional level (16). Critically, EBNA3C has also been shown to regulate the major cell cycle checkpoints (17). Recently it was suggested that EBV has a potential role in inducing genomic instability and that viral proteins associated with the latency III program can regulate the DNA damage response (DDR) (18). In addition, earlier studies from our lab demonstrated that EBNA3C binds to Chk2, a major effecter of the DDR, which also deregulates the cell cycle of EBV infected cells at the G2/M phase (19, 20). EBV infection of primary B-cells was shown to activate the DDR by inducing phosphorylation of H2AX at Ser139 (-H2AX) (20). H2AX is a histone variant, which has a key regulatory function during induction of the DDR. Induction of -H2AX is a hallmark of the DDR and recruits various DNA damage proteins, repair proteins as well as cell cycle checkpoints (21). Recently, we found that H2AX phosphorylation is important for KSHV-induced oncogenesis, which is mediated through one of its major latent protein LANA (22). However, upon EBV infection, the mechanism by which cells trigger the DDR and proceed towards oncogenesis is still not clearly understood. Further, it is still not determined how the DDR progresses without repairing the damaged DNA to bypass cell cycle arrest or apoptosis (16, 23, 24). In this study, we now demonstrate that the EBV latent antigen EBNA3C deregulates total H2AX levels transcriptionally and post-translationally through involvement of the ubiquitin-mediated proteasome degradation pathway. Additionally, our study also showed dramatic changes in expression patterns of the tumor suppressor p53 and oncoprotein Bub1 in H2AX 5
6 knockdown LCLs. These results provide further clues as to the biological relevance of H2AX deregulation in EBV-induced oncogenesis. Overall, our study suggests that EBNA3C can play an important role in EBV-mediated oncogenesis through downmodulation of H2AX
7 MATERIALS AND METHODS Ethics Statement - The University of Pennsylvania School of Medicine CFAR (centre for AIDS research) Immunology core provided us human peripheral blood mononuclear cells (PBMC) from unidentified donors, with written consent, which is approved by IRB based on Helsinki recommendations. Constructs and Transfection - Full length EBNA3C plasmid or its various truncated domains were previously mentioned (25, 26). Plasmid expressing wild type H2AX was previously described (27). Constructs expressing Flag-tagged wild type H2AX and its mutated version H2AX Ser139 (H2AX S-A) were kindly provided by Dr Alvaro N.A. Monteiro (H. Lee Moffitt cancer center, Tampa, FL) (28). pgl3-h2ax wild type reporter construct was generously provided by Dr. Toru Ouchi (Roswell Park cancer Institute, Buffalo, NY) (29). All construct sequences were further confirmed by DNA sequencing at the core facility in University of Pennsylvania. All transfection procedures were as described earlier (30). Cell cultures and Antibodies EBV positive LCL1 and LCL2, BJAB stably expressing EBNA3C clones (BJAB#7 and BJAB#10) and EBV negative cell lines BJAB, DG75, and Ramos were cultured in RPMI 1640 medium and as described earlier (22). Rabbit anti-h2ax antibody was obtained from Bethyl laboratories Inc. (Montgomery, TX). GAPDH (anti-mouse) was purchased from US biological corp. (Swampscott, MA). Mouse antibodies reactive to the Myc epitope (9E10), Flag epitope (M2) and EBNA3C (A10) were described earlier (27). Induction and Infection of recombinant EBV - BAC GFP-EBV was generated as previously described (13). EBNA3C null mutant (ΔEBNA3C BAC GFP-EBV) was 7
8 generated by deleting EBNA3C region between bp from WT BAC GFP EBV Plasmid (JL, HCJ and ESR, Personal communications). Southern Blot analysis, PCR, and sequencing confirmed that the mutation was correct. Cells harboring BAC GFP-EBV and ΔEBNA3C GFP- EBV were induced with TPA (12-Otetradecanoylphorbol) and butyric acid (BA, 3mM: Sigma-Aldrich Corp., St. Louis, MO) for 4-5 days in DMEM. Cell suspension was centrifuged at 1,800 rpm for 15 min and filter through 0.45μ cellulose acetone filters. The viral particles were concentrated by ultracentrifugation at 23,500 rpm at 4 o C and viral particles were stored at -80 o C. For infection studies, we followed similar procedures as described earlier (31). Ten million PBMCs were mixed with either wild type or ΔEBNA3C virus supplemented with RPMI 1640 containing 10% FBS and incubated overnight at 37 o C. Cells were centrifuged at 2,000 rpm for 10 min. Pelleted cells were resuspended in 2 ml of cell culture medium supplemented with 10% FBS. GFP expression was visualized to monitor infection and RT-PCR of EBNA1 confirmed infection. RNA interference - Short-hairpin oligonucleotides directed against EBNA3C were designed (Dharmacon Research, Chicago, IL). All procedure and sequence for sh- EBNA3C- clone were followed as described earlier (28). To clone sh-h2ax, we used similar strategy and sequences described earlier (32). Cell cultures were selected with Puromycin. Immunoprecipitation and Western blotting - Transiently transfected cells were harvested and washed with ice-cold PBS followed by lysing cells using RIPA buffer (10mM Tris, ph 7.5, 150mM Sodium chloride, 2mM EDTA, 1% NP40) and the protease inhibitor cocktail was added before lysing the cells. The cell debris was removed by 8
9 centrifugation at 21,000g for 12 min (4 o C), supernatant was transferred to autoclaved micro-centrifuge tubes. The lysate were then pre-cleared by spinning with normal mouse serum and 1:1 mixture of Protein A and Protein G conjugated Sepharose beads for 1.5 hours at 4 o C. The beads were spun at low speed and the supernatant transferred to a fresh micro-centrifuge tube. The specific protein of interest was incubated with 1 μg of suitable interacting antibody for overnight at 4 o C in a rotating chamber. Immune complexes were collected by binding with Protein A and G beads, pelleted and washed with ice cold RIPA buffer 3 times. WB was performed as described previously (33). RNA isolation and Quantitative real time PCR - Trizol reagent (Invitrogen Inc, Carlsbad, CA) was used to isolate total RNA as described by manufacturer's instruction. RNA to cdna was prepared by superscript II reverse transcription kit (Invitrogen Inc, Carlsbad, CA). The primers used for RT-PCR were as follows: H2AX 5 - GTTCCCAGTGGGCCGTGTA-3 and 5 -CGGTGAGGTACTCCAGCACT-3 for EBNA3C 5 -AGAAGGGGAGCGTGTTTGT-3 and 5 -GGCTCGTTTTGACGTCGGC- 3 for EBNA1 5 -CATTGAGTCGTCTCCCCTTTGGAAT-3 and 5 - TCATAACAAGGTCCTTAATCGCATC-3 and for GAPDH 5 - TGCACCACCAACTGCTTAG-3 and 5 -CATGCAGGGATGATGTTC-3. Quantitative RT-PCR was carried out using Step One Plus Real-time PCR system (Applied Bio Systems, Foster city, CA), and all experiments were performed in triplicates. 9
10 Reporter assay - 10 million HEK-293 cells were co-transfected with 10μg of pgl3h2ax reporter plasmids and increased concentration of EBNA3C, respectively. All procedures were followed as described earlier (22). Ubiquitination assay - HEK293 cells were transfected with vector, Flag- EBNA3C, HA-Ub and Myc-H2AX. Cells were treated with 20 μm MG132 drug (Enzo Life Sciences Inc, Plymouth meeting, PA). All procedures were essentially performed as described earlier (53). Briefly, after 36 hours post-transfection cell lysates were taken and immunoprecipitated with specific antibodies. Immunoprecipitated samples were resolved by SDS-PAGE electrophoresis and transferred onto nitrocellulose membranes. Levels of ubiquitination were evaluated by HA-specific antibody (12CA5). H2AX antibody was used for immunoprecipitation, and Western blots were performed using the anti-ubiquitin specific antibody. Immunofluorescence (IF) analysis - IF was carried out essentially as described previously (34). Briefly, U2OS cells layered on coverslips were transiently transfected with the indicated plasmids by using Lipofectamine 2000 (Invitrogen, Carlsbad, CA). B- cells BJAB, BJAB#7, BJAB#10, LCL1 and LCL2 were air dried and fixed. All procedures were followed as described earlier (22). GST pull down assay - Escherichia Coli BL21-DE3 was transformed (by heat shock method) with Glutathione-S-Transferase (GST) and GST H2AX plasmid constructs. All procedures were followed as described earlier (22). Colony formation assay - 10 million HEK-293 cells were transfected with EBNA3C and H2AX mutant and wild type plasmids by electroporation. 36 hours posttransfection cells were selected with G418 in DMEM supplemented with 5μg/ml 10
11 Puromycin. The selection media was changed on alternate days. After two weeks cells were fixed on the plates using 3% paraformaldehyde (PFA) for 30 min followed by staining with 0.1% crystal violet. The total intensity of colonies was calculated by scanning the dish using the Li-Cor Odyssey (LiCor Biosciences, Salt Lake city, Utah) system and quantified. 11
12 RESULTS H2AX expression is down-regulated in the presence of EBNA3C during primary infection and latency- It was recently demonstrated that the EBV essential antigen EBNA3C contributes to attenuation of the DNA damage response (DDR) during latency, and is particularly active during early infection of naive B-cells in vitro by EBV (31). However, the mechanism of attenuation has not been elucidated. H2AX is ubiquitously expressed in mammalian cells and the phosphorylation of H2AX at Ser139 is a well known marker for the DDR (35). This is supported by earlier studies which demonstrated that reduction in H2AX levels is associated with higher genomic instability and tumor predisposition (18, 23, 26). To investigate the effects of EBNA3C on H2AX regulation, we evaluated H2AX levels in the presence of EBNA3C expression as well as in EBV infected B-cells. We first compared H2AX expression in EBV-transformed lymphoblastoid cell lines (LCL1 and LCL2) with EBV-negative Burkitt s lymphoma (BL) cells, Ramos and DG75 (Fig. 1A). The results showed that cellular H2AX levels were down-regulated by approximately 3-4 fold in LCL1 and LCL2, compared to the EBV negative cell lines DG75 and Ramos by Western blot analysis (Fig. 1A). To determine if EBNA3C alone can affect H2AX expression, we next investigated the levels of H2AX in BJAB cells stably expressing EBNA3C cell lines (BJAB#7 and BJAB#10) compared to a BJAB vector control cell line (Fig. 1A). Our results from Western blot analysis showed changes in H2AX levels with a drop of at least 2-fold in BJAB#7, and BJAB#10 compared to BJAB cells (Fig. 1A). In addition, when EBNA3C expression was stably knocked down using specific lentiviral constructs in LCLs, a dramatic enhancement of approximately 5-fold in H2AX levels was observed (Fig. 1A). These 12
13 results strongly suggest that EBNA3C contributes directly to down-regulation of H2AX in EBNA3C-expressing cells. EBNA3C is a strong transcription factor, capable of regulating transcription of both viral and cellular genes (16, 30, 32, 34). To determine if H2AX was directly modulated by EBNA3C at the transcript level, we evaluated H2AX transcripts in EBV transformed, and EBNA3C stable cell lines compared to EBV negative B-cells. The cells include two lymphoblastoid cell lines (LCL1 and LCL2), two EBNA3C stably expressing BL cell lines (BJAB#7 and BJAB#10), and LCL1 stably knocked down for EBNA3C using a specific lentiviral construct along with its isogenic negative cell line (Fig. 1B). The results from real-time PCR analysis indicated that the H2AX levels were significantly reduced in both EBV positive as well as EBNA3C positive cell lines; whereas, this was mostly reversed when EBNA3C was knocked down in an LCL background (Fig. 1B). These results indicated that EBNA3C can block H2AX expression under normal physiological conditions. To provide another link to the physiological relevance of H2AX expression during primary infection, we performed an in vitro infection study using peripheral blood mononuclear cells (PBMCs) with recombinant viruses, including both wild type as well as an EBNA3C deleted EBV (ΔEBNA3C) (Fig. 1C). 10 million resting PBMCs were infected and grew over a period of seven days post EBV infection (Fig. 1C). H2AX expression level was attenuated within day 2 post-infection, and this decrease was recovered at day 7 to just over 50% based on quantitation using a LiCor Odyssey in the linear range (Fig. 1C). However, PBMC s infected with the ΔEBNA3C virus showed enhanced H2AX levels in post-infected cells to almost 2-fold which was similar up to 7 13
14 days (Fig. 1C). The Western blot results were also supported by subsequent real-time PCR results, where we observed an attenuation of H2AX at the transcript level in EBV infected PBMCs. However, an increase in H2AX transcript levels was observed using the EBNA3C deleted EBV recombinant virus (Fig. 1D). These results suggest that EBNA3C contributes to the reduction of total H2AX levels and may be important during the early stages of EBV-mediated B-cell infection which leads to transformation. Expression of EBNA3C leads to down-regulation of H2AX in Burkitt s lymphoma and epithelial cells To determine the involvement of other viral oncoproteins as the effects of EBNA3C on H2AX expression, we performed expression analysis in both Burkitt s lymphoma (BL) or epithelial cells. First, an EBNA3C expressing plasmid was co-transfected along with GFP-control vector into three viral negative BL cell lines namely BJAB, DG75 and Ramos (Fig. 2A). A GFP-expression vector was used to determine the overall transfection efficiency. Western blot results demonstrated that there was an approximately 2-4 fold reduction in H2AX protein levels in the presence of EBNA3C (Fig 2A). In agreement with these Western blots, H2AX transcripts analyzed using real-time PCR also showed a significant decrease of about 2-3 fold in the presence of EBNA3C compared to two EBV negative BL cell lines (Fig. 2B). To determine whether the effect of EBNA3C on H2AX is cell type specific, we performed similar experiments as described above in epithelial cells using HEK-293 cells in a dose dependent manner. As expected, H2AX levels were significantly attenuated with increasing expression of EBNA3C at the transcript and protein levels (Fig. 2C and 2D, respectively). 14
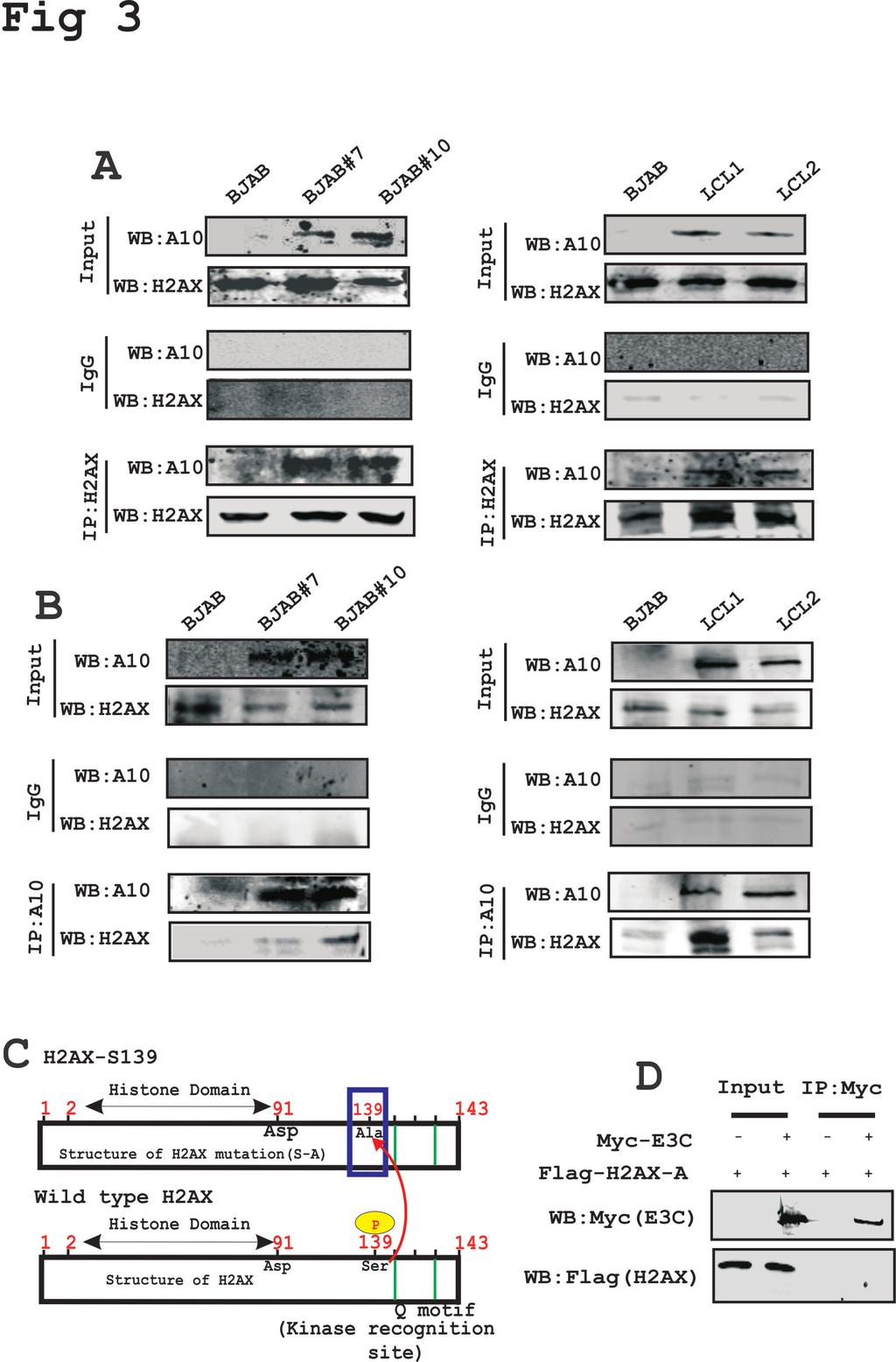
15 Importantly, co-expression of H2AX tagged with the Myc epitope and EBNA3C tagged with the Flag epitope from heterologous promoters also showed attenuation of H2AX at the protein level as using the Myc specific mouse monoclonal antibody (Fig. 2E). These results indicated that EBNA3C can also deregulate H2AX at the transcriptional level. EBNA3C forms a complex with H2AX - In an attempt to understand whether this attenuation of H2AX levels in the presence of EBNA3C expression is regulated through an interaction between these two molecules, we further investigated their binding activities in B-cells. To support this, we performed immunoprecipitation assays with anti- H2AX antibody in BJAB, BJAB#7, BJAB#10 and LCL1, LCL2 cells (Fig. 3A). The results demonstrated that H2AX was stably associated with EBNA3C in both EBNA3C stably expressing cells (BJAB#7 and BJAB#10) as well as in LCL1 and LCL2 (Fig. 3A). Further to support our interaction results, we performed a reverse immunoprecipitation assay, where we used the A10 mouse monoclonal antibody specific for EBNA3C using similar cell lines as described above (Fig. 3B). These association studies provided strong evidence that EBNA3C interacted with H2AX in both EBV-transformed cells as well as in EBNA3C positive Burkitt's lymphoma cells (Fig. 3A). H2AX autophosphorylation plays a critical role in interaction between H2AX with EBNA3C - H2AX phosphorylation at Ser139 has previously been shown to be critical for maintenance of latency during multiple gammaherpesviruses infection including mouse gammaherpesvirus 68 (MHV-68) and KSHV (22, 36). Interaction between 15
16 EBNA3C and H2AX prompted us to further evaluate whether mutation at the Ser139 residue of H2AX was critical for complex formation with EBNA3C (Fig. 3C). To address this question, we performed an immunoprecipitation assay using HEK-293 cells where the Flag-tagged Ser139 H2AX (where the Serine residue was replaced by an Alanine residue) was co-expressed with Myc-tagged EBNA3C (Fig. 3D). Subsequently, immunoprecipitation was performed by using anti-myc antibody to target the Myctagged EBNA3C (Fig. 3D). The immunoprecipitated bands were visualized with anti- Flag antibody. Interestingly, no signal was detected for EBNA3C when H2AX was mutated at the Ser139 residue (Fig. 3D). This indicated that the Ser139 residue of H2AX was important for complex formation of H2AX with EBNA3C. Moreover, we found reduced expression levels of gamma-h2ax in EBV positive LCL1 and LCL2 compared to EBV negative DG75 and Ramos cells (data not shown). The first 100 residues of EBNA3C are responsible for the interaction of EBNA3C with H2AX - To identify the binding residues of EBNA3C important for interaction with H2AX, we further performed similar immunoprecipitation experiments in a heterologous expression system using HEK-293 cells. Cells were transfected with plasmids expressing Flag tagged full-length EBNA3C (residues 1-992), N-terminal region (residues 1-365), middle region (residues ), and the C-terminal region (residues ) along with a Myc-tagged H2AX expression vector (Fig. 4A). Immunoprecipitation assays were performed using a Flag specific mouse monoclonal antibody (M2). The results revealed that H2AX forms a complex with residues within the N- terminal region of EBNA3C, but not with the middle or carboxyl-terminal domains (Fig. 4A). 16
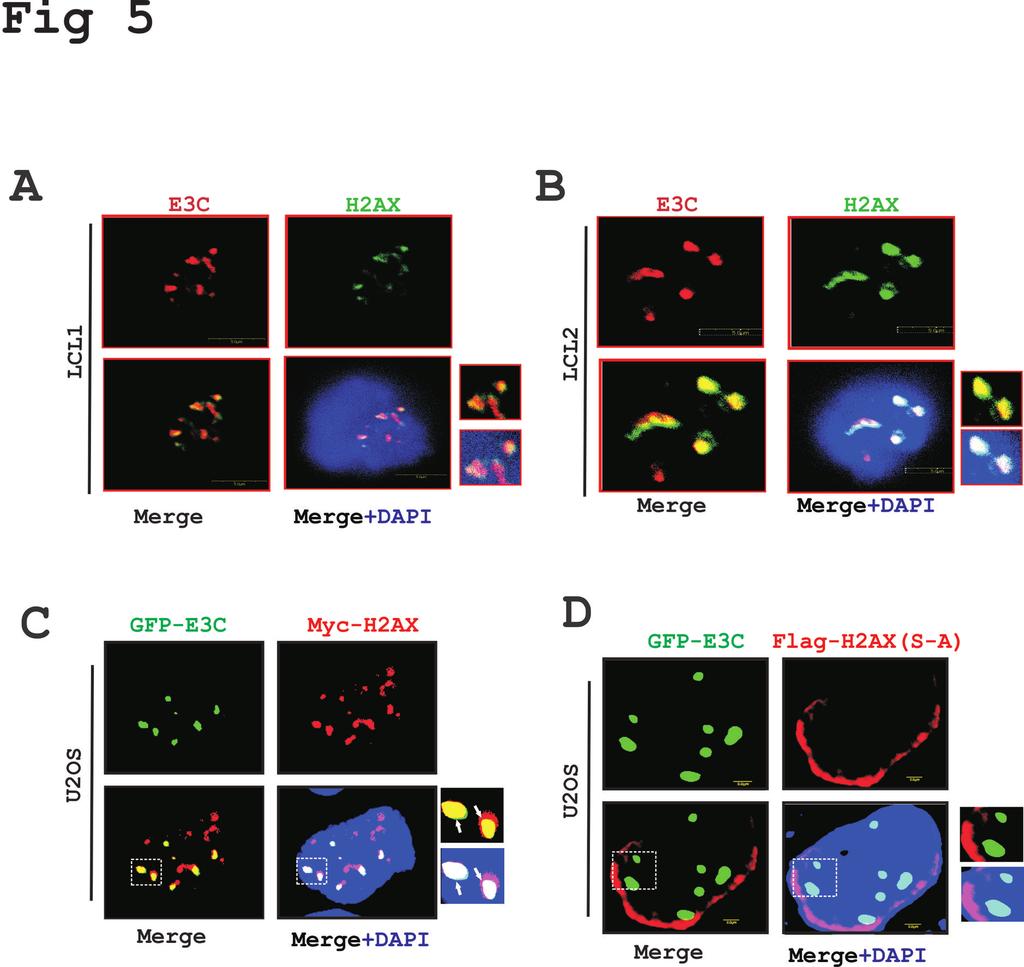
17 To further confirm the above association studies, we performed GST pull-down experiments using bacterially purified GST-fused H2AX with the HEK-293 cell extracts transiently transfected with plasmids expressing full length EBNA3C and the three major truncated domains of EBNA3C (N-terminal, M-domain and C-terminal) (Fig. 4B). The results from GST pull-down assays also confirmed a strong interaction between fulllength EBNA3C and the N- terminal (1-365aa) residues with H2AX (Fig. 4B, top panels). The amount of control GST and GST-H2AX fusion protein used in the GST pull down experiment was similar as shown by corresponding Coomassie brilliant blue stained SDS-PAGE (Fig. 4B, bottom panel). Additionally, to narrow down the binding residues with the N-terminal domain of EBNA3C, we further performed GST pull-down assays using various truncated regions of the EBNA3C N-terminal region (Fig. 4C). A strong association was clearly detected between GST-fused H2AX with EBNA3C residues 1-300, and 1-100, while none were found beyond the first 100 amino acid residues of EBNA3C (Fig. 4C). These results demonstrate that the EBNA3C residues are responsible for formation of a stable complex with H2AX in cells. The schematic represents the mapping of the H2AX interaction domain within EBNA3C and further outlines the studies to identify the interacting residues (Fig. 4D). EBNA3C colocalizes with H2AX in EBV-transformed LCLs - Our results from both immunoprecipitation as well as GST pull-down assays strongly supported the formation of a complex between EBNA3C and H2AX under physiological conditions using EBVtransformed B cells. This prompted us to investigate whether EBNA3C and H2AX can colocalize within similar cellular compartments. Immunofluorescence (IF) studies were 17
18 performed using EBNA3C stably expressing B-cells as well as EBV transformed LCLs and EBV-negative controls (Fig. 5). First, IF was carried out in BJAB control cells along with the isogenic EBNA3C expressing cells (BJAB#7 and BJAB#10) (data not shown). The assay was carried out using specific antibodies against EBNA3C (A10, mouse monoclonal), and H2AX (rabbit polyclonal) along with Texas-Red mouse and FITC-anti rabbit secondary antibodies for visualization of the complexes. The results showed a strong colocalization between H2AX and EBNA3C in similar nuclear compartments (data not shown). Furthermore, an analysis of the fluorescence intensity for H2AX signals in EBNA3C expressing BJAB cells demonstrated a significant reduction compared to control BJAB cells (data not shown). This further corroborated our biochemical studies as discussed above, which showed reduced H2AX levels in the presence of either EBV or EBNA3C. Importantly, IF analysis using LCL1 and LCL2 showed a similar co-localization pattern of H2AX and EBNA3C (Fig. 5A-B). To further validate this observation, we performed similar IF experiments using GFP-tagged EBNA3C and Myc-tagged H2AX in U2OS epithelial cells at 36 hours post-transfection (Fig. 5C). Monitoring the GFP signal of EBNA3C and a specific mouse monoclonal antibody against H2AX tagged with the Myc epitope clearly showed a punctuate pattern of co-localization between H2AX molecules and EBNA3C in similar nuclear compartments (Fig. 5C). Our binding results previously demonstrated that phosphorylation of H2AX at Ser139 is crucial for binding with EBNA3C. To further determine whether this residue was also important for colocalization, we next performed an IF study with GFP-tagged EBNA3C and a Flag tagged mutated version of H2AX (S-A) in U2OS cells (Fig. 5D). At 18
19 hours post transfection, IF was performed using the specific mouse monoclonal Flagantibody (M2) along with GFP fluorescence at 488 nm excitation (Fig. 5D). Interestingly, the IF-signals for H2AX (S-A) were observed mostly at the periphery of the nucleus instead of punctuate dots seen for the wild type version (compare Fig. 5C and 5D). Consequently, little or no association of EBNA3C was observed in nuclear compartments with H2AX (S-A) (Fig. 5D). In agreement with our binding studies, the IF results once again showed that Ser139 of H2AX may play a critical role in formation of a complex as well as colocalization with EBNA3C in similar nuclear compartments. EBNA3C inhibits the transcription activity of the H2AX promoter - The above results showed that H2AX transcripts were down-regulated in the presence of EBNA3C. To more specifically investigate this phenomena we performed Luciferase based reporter assays using the wild type H2AX promoter linked to the luciferase reporter gene in the presence of EBNA3C (Fig. 6). First, the H2AX reporter plasmid H2AX-pGL3 was transiently transfected into HEK-293 cells in the presence of vector or full length Myctagged EBNA3C (Fig. 6A). Cells were also transfected with GFP control plasmid to monitor the transfection efficiency in this experiment. The results demonstrated that EBNA3C dramatically attenuated the transcriptional activity from the H2AX promoter in a dose dependent fashion in HEK-293 cells (Fig. 6A). The levels of EBNA3C, and GAPDH were analyzed by WB (Fig. 6A). Further, to evaluate whether the binding domain of EBNA3C is necessary to control the H2AX transcription, we performed similar reporter assays using various truncation domains of EBNA3C (Fig. 6B-F). The results showed that the N-terminal binding region (residues 1-365), but not the non- 19
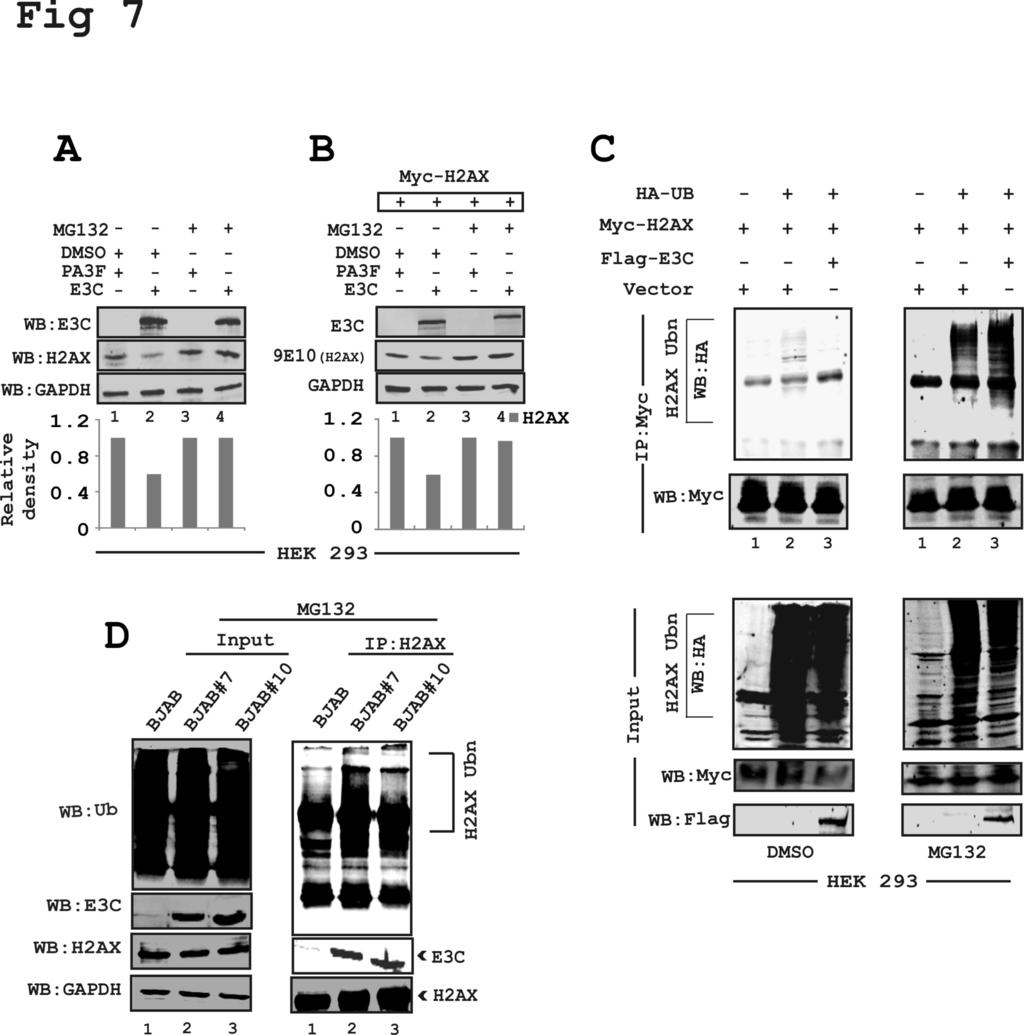
20 binding domains (residues ) of EBNA3C substantially attenuated transcription from the H2AX promoter (Fig. 6B-D). In agreement, the smaller N-terminal binding residues amino acid binding region of EBNA3C also showed a similar inhibitory trend of transcriptional regulation at the H2AX promoter (Fig. 6E). This indicated that the amino-terminal binding residues of EBNA3C is required, as well as sufficient for transcriptional inhibition of H2AX promoter. To further validate our results above we transfected HEK-293 cells with the N-terminal deleted region of EBNA3C (ΔN EBNA3C) along with the reporter and GFP expression plasmids (Fig. 6F). This result supported our above data that the N-terminal binding residues of EBNA3C are important for regulating H2AX transcription. All reporter experiments were carefully examined by WB analysis to monitor the expression levels of EBNA3C and GAPDH as an internal loading control (Fig. 6). EBNA3C can induce degradation of H2AX by recruiting the ubiquitin-proteasome system - To establish latency EBV has evolved strategies for targeted inactivation of cellular factors to escape from the DDR induced during early infection (37-40). However, the underlying mechanism is still not clear. Overall, our results described above, strongly suggested that in addition to transcriptional regulation, EBNA3C can also attenuate H2AX at the protein level. Moreover, it has also been reported that the H2AX protein level can be regulated through poly-ubiquitination (23). Interestingly, EBNA3C has been previously shown to utilize a ubiquitin-mediated proteosome degradation strategy for targeting and degrading multiple tumor suppressor proteins to establish successful latent infection in infected B-cells (31, 32). Along these lines, we wanted to determine whether down-regulation of H2AX protein levels were associated with EBNA3C-mediated 20
21 degradation through the ubiquitin-proteasome pathway. To explore this possibility, HEK- 293 cells were transfected with EBNA3C or vector alone and subsequently treated with either DMSO control or MG132 over a period of 12 hours at 48 hr post-transfection (Fig. 7A). MG132 blocks the degradation of ubiquitin-conjugated proteins in mammalian cells by the 26S proteasome complex (41). The results demonstrated that the reduced levels of endogenous H2AX in the presence of EBNA3C was rescued by MG132 when compared to control cells treated with DMSO (Fig. 7A, compare lanes 2 and 4). This observation was further validated using transient expression of Myc-tagged H2AX expressed from a heterologous promoter in HEK-293 cells (Fig. 7B). These results also demonstrated a rescue of H2AX protein levels from EBNA3C-mediated reduction in the presence of MG132 (Fig 7B). These results indicate that EBNA3C can contribute to H2AX degradation through recruitment of the ubiquitin-proteasome machinery, the function of which was blocked by MG132 (7A-B). To support these results, we next performed in vivo ubiquitination experiments. HEK-293 cells were co-transfected with plasmids expressing HA-tagged ubiquitin moiety (HA-Ub), Flag-tagged EBNA3C and Myc-tagged H2AX (Fig. 7C). The levels of H2AX-ubiquitination was monitored by performing specific immunoprecipitation of H2AX molecules using anti-myc mouse monoclonal antibody (9E10) followed by WB analysis (Fig. 7C). The results demonstrated a significant difference in polyubiquitination levels of H2AX in the MG132 treated compared to DMSO control group (Fig. 7C, compare left and right panels). This is due to impairment in proteasomal function which resulted in an increase in levels of ubiquitinated proteins. A dramatic enhancement in H2AX poly-ubiquitination was reproducibly observed in the presence of 21
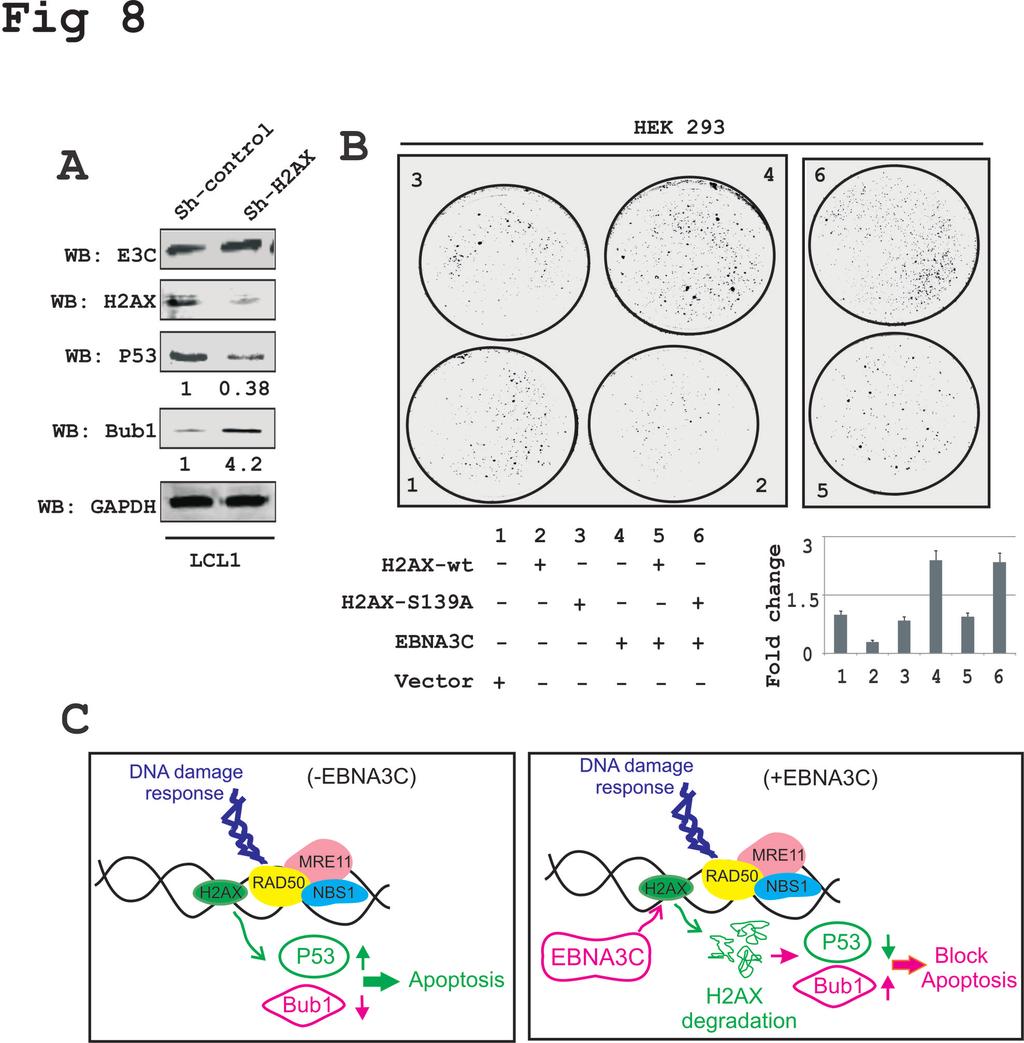
22 EBNA3C with MG132 treatment, but not with DMSO control (Fig. 7C, compare left and right panels, lane 3). To support the above findings, ubiquitination experiments were also carried out in BJAB, BJAB#7 and BJAB#10 cells by performing H2AX immunoprecipitation using an anti-h2ax specific antibody (Fig. 7D). The results further demonstrated a relatively higher level of H2AX poly-ubiquitination in EBNA3C expressing BJAB#7 and BJAB#10 cells compared to the BJAB control cells (Fig. 7D, compare lane 1, with lanes 2 and 3). This provided additional evidence to suggest that EBNA3C can facilitate H2AX destabilization through ubiquitin-mediated proteasomal degradation. EBNA3C inhibits the growth suppressive activity of H2AX The spindle assembly checkpoint (SAC) and DDR are two critical mechanisms by which mammalian cells can maintain genomic stability (42). Moreover, H2AX was shown to induce cell cycle arrest via the p53/p21 pathway (43). In addition, knockdown of H2AX was shown to strongly suppress apoptosis in lung cancer cells (44). To elucidate the biological significance of H2AX degradation through EBNA3C in LCLs as well as to explore the underlying molecular mechanism of this DNA damage protein in EBV-induced B-cell lymphomagenesis, we used H2AX knockdown LCL1 cells compared to control vector LCL1. Interestingly, H2AX knockdown resulted in a reduction in levels of the p53 tumor suppressor protein of about 2-3 fold, while the level of Bub1, a critical kinetochore protein essential for SAC was upregulated about 3-5 fold (Fig. 8A). These results now provide a potential molecular strategy for H2AX downregulation in LCLs, and highlights a critical role for EBNA3C in regulating the DDR during B-cell lymphomagenesis. 22
23 To specifically explore the inhibitory effects of EBNA3C on H2AX-mediated growth suppressive properties, we carried out colony formation assays (CFA) in HEK- 293 cells (Fig 8B). Cells were transfected with either vector control or vectors expressing wild type H2AX, mutated H2AX (S139A), or EBNA3C alone or co-transfected with combinations of EBNA3C and wild type H2AX, or EBNA3C with mutated H2AX (S139A) (Fig. 8B). 24 hours post-transfection, cells were further selected with G418 for 2 weeks. Both media and antibiotics were changed every 2-days. The total colony formation was scanned and quantitated using the Odyssey Image analysis software (Fig. 8B). The results showed that, expression of wild type H2AX caused an approximately 70% reduction in colony formation of HEK-293 cells when compared to vector control. However, minimal effects were observed in the presence of the mutated version of H2AX (Fig. 8B, compare panels 1, 2 and 3). This again signifies the importance of the Ser139 residue to the growth suppressive properties of H2AX. Importantly, co-expression of EBNA3C with wild type H2AX demonstrated an almost complete reversion of H2AXmediated growth suppressive activity which was similar to vector control (Fig. 8B, compare panels 1, 3 and 5). Interestingly, EBNA3C expression alone showed a dramatic enhancement of colony formation (Fig. 8B, panels 4 and 6), which further strengthens a role for EBNA3C as a dominant viral oncoprotein. Overall, these results describe a model where EBNA3C can play a critical role in regulating H2AX stability through ubiquitinmediated degradation and can therefore contribute to modulating its activity during the DDR induced by viral infection. This ultimately drives establishment of the EBV latency program in infected primary B-cells. 23
24 DISCUSSION Viral infection and the cellular DNA damage response (DDR) have been widely debated and explored in recent years (24, 45). Viruses have evolved complex regulatory mechanisms to persist in infected cells by manipulating several DNA repair mechanisms (46). H2AX plays a key role in the DDR (47). H2AX is well known for its positive regulation of DNA repair and activation of the DNA damage checkpoints (48). H2AX also varies from H2A with a unique C-terminal region (23), which contains a highly conserved SQE motif (49). However, it has been suggested that H2AX is not utterly important for recognition of DNA damage but facilitates the damage response through interaction with proteins that recognizes the phosphorylated form of H2AX (γ-h2ax) (48). However, previous studies by Bassing et al reported that cells from H2AX deficient mice are more prone to chromosomal instability and less efficient in DNA double stranded break repair (37). Earlier it was reported that the EBV latent proteins EBNAs and LMPs can contribute towards chromosomal instability and cellular DNA damage by inhibiting as well as facilitating degradation of DNA repair elements, or through mis-localization (50, 51). Reports from our lab and others also revealed that EBNA3C can contribute to the oncogenic process through degradation of p53 or interaction with Chk2 (16, 20). However, limited information is available for demonstrating a role for EBNA3C in modulating H2AX. Here, we identified H2AX as an interacting partner with EBNA3C during EBV infection. We showed that in both EBV as well as EBNA3C positive B cell lines, cellular H2AX levels were significantly down-regulated. This initial observation was further 24
25 corroborated using RNA interference strategy by knocking down EBNA3C expression in EBV transformed LCLs. Using an in vitro model system for resting B cell infection with an EBNA3C deleted recombinant virus in comparison to wild type EBV infection, we confirmed that EBNA3C was directly involved in regulating H2AX levels. The attenuation of H2AX levels was clearly more significant in the wild type virus infection, whereas, the EBNA3C deleted EBV did not show any significant change in H2AX expression. These results supported our hypothesis that through modulation of the total H2AX levels, EBNA3C can contribute to EBV latency and cell proliferation. These initial findings were further corroborated in HEK-293 cells, further strengthening our initial hypothesis that EBNA3C is directly involved in deregulating H2AX expression. The reduction of H2AX levels were found to be inversely proportional to the increase in EBNA3C levels at the protein and transcript levels. While unable to bind DNA directly, EBNA3C was previously shown to be a strong transcriptional factor that can regulate transcription of multiple cellular and viral genes. EBNA3C formed a stable complex with H2AX through interaction with its amino-terminal first 100 amino acid residues. Interestingly, we know that the N-terminal region of EBNA3C interacts with various cell cycle regulatory molecules such as Cyclin D1, Cyclin A, p53, Mdm2, E2F1, IRF4 and Aurora Kinase B (16, 30, 32, 34, 52, 53). Phosphorylation of Ser139 of H2AX (referred to as gamma-h2ax) is indispensible for recruitment of DNA repair elements (54). Furthermore, H2AX is phosphorylated by ATM in response to double stranded breaks (54). More specifically, mutation at Ser139 completely abolished the interaction between H2AX and EBNA3C. This suggests, that H2AX phosphorylation may play an important role in EBNA3C 25
26 regulated cell proliferation. This also suggests the involvement of EBNA3C in regulating the DDR and consequently also opens a new avenue for further exploration. Our reporter analysis of the H2AX promoter demonstrated that the N-terminal binding residues were sufficient for blocking H2AX transcription. Furthermore, H2AX localization is strictly confined to the nucleosomes (55). However, some reports demonstrated that H2AX also functions outside the nucleosomes (56, 57). Our results demonstrated that EBNA3C can localize with H2AX within similar nuclear compartments in EBV transformed LCLs as well as EBNA3C expressing stable cell lines. The staining intensity of H2AX in EBNA3C negative and EBNA3C positive cell lines showed a reduction in the levels of H2AX in the presence of EBNA3C further supporting our hypothesis that EBNA3C can attenuate H2AX expression. In addition to transcriptional regulation, our results also indicated that EBNA3C can regulate H2AX protein levels. Previous reports showed that H2AX is degraded by poly-ubiquitination through the proteosome degradation pathway, and that inhibition of the proteosome can stabilize H2AX (40, 58, 59). We hypothesized that EBNA3C mediated H2AX deregulation may be through poly-ubiquitination and degradation since our previous studies showed that EBNA3C recruits the ubiquitin-proteasome machinery for targeted proteolysis of multiple tumor suppressor proteins through its N-terminal residues (31, 60). Recently Feng et al hypothesized that Ubc13 and RNF8 are involved in DDR-induced H2AX ubiquitination (61). How these proteins are involved in the functional regulation of H2AX levels in EBV positive cells remains undetermined. In addition to this, mono-ubiquitination of H2AX was shown to be an essential epigenetic marker during the DNA damage response (62, 63). This would be interesting to 26
27 determine whether EBNA3C could also initiate H2AX mono-ubiquitination to regulate nucleosomal activity and thereby gene transcription. For now, our results demonstrated that EBNA3C efficiently induces H2AX degradation most likely through polyubiquitination. To determine the biological implications of H2AX deregulation in EBV-driven oncogenesis, we generated LCLs knocked down for H2AX and evaluated the expression levels of the p53 tumor suppressor and the oncoprotein Bub1. Earlier Fragkos et al suggested that H2AX was required for cell cycle arrest through p53 (43). There is a growing body of evidence for crosstalk between the DDR elements and SAC components (42). Moreover, Bub1 was shown to be an essential mitotic protein in gammaherpesvirus induced oncogenesis (64). Depletion of H2AX expression in LCLs led to a significant down-regulation of p53 and up-regulation of Bub1. This supports an anti-proliferative role for H2AX in the context of EBV-induced transformation of B- cells. Subsequent colony formation assays further supported the anti-proliferative properties of H2AX. Our results also clearly demonstrated that EBNA3C can effectively neutralize the growth suppressive effects of H2AX. Therefore this essential latent antigen EBNA3C may have evolved to modulate the DDR and so contribute to viral latency in addition to all its other functions. Overall this study provides strong evidence to support a role for EBNA3Cmediated attenuation of total H2AX levels upon EBV infection (Fig. 8C) and may be important for targeted therapeutic strategies against EBV associated B-cell lymphomas 27
28 FIGURE LEGENDS Figure 1: EBNA3C down-regulates total H2AX levels in B cells - (A) Endogenous expression of H2AX in EBV positive cell lines (LCL1 and LCL2) and EBV negative Burkitt s lymphoma cell lines (Ramos and DG75) were analyzed at the protein level (Fig 1A-left panel). Endogenous expression of H2AX in BJAB (EBV negative) BJAB#7, BJAB#10 (EBNA3C stably expressing) cells were analyzed by WB (Fig. 1A, mid panel). LCL1 cells stably transduced for sh-control and sh-ebna3c were harvested to determine the protein expression levels of H2AX. The relative density of H2AX was analyzed by normalizing with GAPDH. (B) All groups of cells were extracted using Trizol reagent and RNA concentration was estimated by using Nanodrop (Eppendorf Inc., Hamburg, Germany). Transcript levels for the H2AX gene is represented by fold change and the experiments were performed in triplicates. Results of LCL1 and LCL2 compared to Ramos and DG75 are shown in Fig. 1B left panel. Results of BJAB compared to BJAB#7 and BJAB#10 are shown in Fig. 1B mid panel and LCL1 stably knocked down for EBNA3C and control vector results were shown in Fig. 1B right panel. (C, D) PBMC s were infected with wild type (WT) EBV virus and ΔEBNA3C virus up to 7 days. Cells were harvested and expression of H2AX was monitored in a time-dependent manner for protein and transcript levels by performing WB analysis and real-time PCR. All panels show a representative picture of the repeated experiments. Figure 2: EBNA3C attenuates H2AX levels in Burkitt s lymphoma and epithelial cells - (A) Approximately 30 million BJAB, DG75 and Ramos cells were harvested for WB analysis of H2AX, EBNA3C, GAPDH and GFP levels after transient transfection of EBNA3C and GFP constructs (B) Endogenous expression of H2AX levels 28
29 in Burkitt s lymphoma cell lines transfected with EBNA3C was analyzed for transcript levels by real-time PCR. Approximately 15 million cells of BJAB, DG75 and Ramos were co-transfected with the GFP expressing vector either with EBNA3C or control vector. Transcript levels of the H2AX gene are represented as fold change. The experiments were performed in triplicate. (C) H2AX levels in HEK-293 cells were transfected with EBNA3C in a dose-dependent manner and analyzed for transcript levels by RT-PCR. Transcript levels of the H2AX gene was represented as fold change performed in triplicate. (D) H2AX expression levels in HEK-293 cells transfected with EBNA3C in a dose-dependent manner was analyzed for protein level. (E) H2AX and EBNA3C constructs were transfected in HEK-293 cells and analyzed for the detection of H2AX protein levels. HEK-293 cells were transfected with GFP, EBNA3C Flag or vector with Myc-H2AX. All panels are representative pictures from repeat experiments. Figure 3: H2AX associates with EBNA3C in cells - (A) Complex formation between EBNA3C and H2AX was evaluated using anti-h2ax antibody in Burkitt's lymphoma (Fig. 3A, left panel) and LCLs (Fig. 3A, right panel). (B) Association between EBNA3C and H2AX was evaluated using the anti-ebna3c (A10) antibody in Burkitt's lymphoma (Fig. 3B, left panel) and LCLs (Fig. 3B, right panel). (C) A schematic showing H2AX wild type and specific serine139 mutation with alanine in full length protein is shown. (D) Mutant Flag-H2AX Ser139 (H2AX-S-A) was co-transfected either with Myc-EBNA3C or control vector. WB was performed with anti-myc and anti-flag antibodies. Figure 4: The first 100 amino acid residues of EBNA3C protein are critical for complex formation with H2AX - (A) Complex formation between EBNA3C and 29
30 H2AX was investigated in HEK-293 cells by immunoprecipitation. Approximately 10 million HEK-293 cells were transfected either with control vector, full length EBNA3C or different truncations of EBNA3C (Amino, Middle and carboxy-terminal) region tagged with Flag vector. (B) GST pull down assay was performed by using EBNA3C truncations (Full-length EBNA3C aa, N-terminal aa, Middle domain aa and C- terminal aa) fused to GST. Smaller truncations of the N-terminal region of EBNA3C (1-300, 1-200, , 1-100, 1-50 and aa) were cloned in pa3m vector and monitored for binding activity with H2AX in the right panel. Coomassie stained gel is shown for GST-control and GST-H2AX fusion protein. (C) A schematic shows different EBNA3C domains associated with other reported proteins as well as its binding domain for H2AX. Figure 5: EBNA3C colocalizes with H2AX in nuclear compartments - (A) In vitro EBV transformed B-cell lines LCL1 and LCL2 were plated on slides and subsequently air-dried at room temperature. Cells were fixed using 3% PFA and blocked with 5% skimmed milk. Protein expression were detected by using anti-mouse EBNA3C A10 monoclonal antibody (1:150), and anti-rabbit H2AX antibody (1:500) followed by anti-rabbit Alexa flour 594 (red) for H2AX and anti-mouse Alexa flour 488 (green) for EBNA3C. DAPI was used for nuclear staining. (B) 0.3 million epithelial U2OS cells were transfected with GFP-EBNA3C and Myc-H2AX plasmids using Lipofectamine. H2AX was detected using Anti-Myc (9E10) antibody (1:200) followed by Anti-rabbit Alexa flour 594 (red) secondary antibody. Co-localization of H2AX with GFP-EBNA3C was monitored using an Olympus confocal microscope. (C) 3x10 5 U2OS cells were seeded on coverslip and transfected with GFP-EBNA3C and mutant H2AX S-A plasmid 30
31 using Lipofectamine. (D) HEK-293 cells were co-transfected with plasmids expressing Flag-EBNA3C and Myc-H2AX. Figure 6: The N-terminal binding residues of EBNA3C are sufficient for suppressing H2AX transcription - HEK-293 cells were transfected with 10 μg of pgl3-h2ax and 1, 2 and 4 μg of EBNA3C truncations (EBNA3C aa, aa, aa and aa). The N-terminal deleted region of EBNA3C (ΔEBNA3C-N) was also included. Data from the reporter assays are represented as relative Luciferase unit (RLU) with mean and standard deviation represented by error bars. The fold activation was calculated by comparison of the promoter activity in the presence of promoter pgl3-h2ax with the value of the PA3M vector alone. Figure 7: EBNA3C facilitates H2AX degradation through the ubiquitinmediated proteasomal pathway - (A) HEK-293 cells were transfected with plasmids and treated with either MG132 (20μg/ml) or DMSO as control for 30 hours posttransfection to inhibit the proteasome activity. (B) HEK-293 cells were transfected with plasmids expressing EBNA3C-Flag or Myc-H2AX. (C) HEK-293 cells were transfected with various combinations of expression vectors as showed. (D) In vivo ubiquitination assays were carried out using 50 million BJAB (EBNA3C negative) BJAB#7 and BJAB#10 (EBNA3C positive stable cell lines). H2AX antibody (2μg) was used for pull down experiments and subsequently detected using ubiquitin and H2AX specific antibodies. The input was also analyzed for loading control using the anti-gapdh antibody. Figure 8: EBNA3C blocks H2AX-mediated growth suppressive activity - (A) Knockdown for H2AX cells were evaluated to determine H2AX, EBNA3C, Bub1 and 31
32 p53 expression levels using their respective antibodies. Experiments were independently performed three times with similar results. (B) Colony formation assays was performed in HEK-293 cells. Cells were transfected with control vector, wild type H2AX, mutant H2AX (S-A), EBNA3C and EBNA3C combined with wild type H2AX or H2AX (S-A). 24 hours post-transfection the cells were selected with G418 for 2 weeks. On alternative days the media and antibiotics were replaced. Cells were fixed by 3% PFA and stained with 0.1% crystal violet. The total number of colonies was measured using the Odyssey Image analysis software. (C) A schematic showing the role of EBNA3C in degradation of H2AX which results in modulation of p53 and Bub1 levels. These changes can contribute to EBV-induced oncogenesis. 32
33 ACKNOWLEDGMENTS We are grateful to Dr Alvaro N.A. Monteiro (H. Lee Moffitt cancer center, Tampa, FL), Dr. Toru Ouchi (Roswell Park cancer Institute, Buffalo, NY), and Elliott Kieff (Harvard Medical School, Boston, MA) for kindly providing reagents. The work was supported by NCI grant CA , NIDDK grant DK ESR is a scholar of the Leukemia and Lymphoma Society of America. 33
34 REFERENCES 1. Epstein MA, Achong BG, Barr YM Virus Particles in Cultured Lymphoblasts from Burkitt's Lymphoma. Lancet 1: Young LS, Murray PG Epstein-Barr virus and oncogenesis: from latent genes to tumours. Oncogene 22: Pegtel DM, Middeldorp J, Thorley-Lawson DA Epstein-Barr virus infection in ex vivo tonsil epithelial cell cultures of asymptomatic carriers. J Virol 78: Chan AS, To KF, Lo KW, Ding M, Li X, Johnson P, Huang DP Frequent chromosome 9p losses in histologically normal nasopharyngeal epithelia from southern Chinese. Int J Cancer 102: Cho WC Nasopharyngeal carcinoma: molecular biomarker discovery and progress. Mol Cancer 6:1. 6. Thompson MP, Kurzrock R Epstein-Barr virus and cancer. Clin Cancer Res 10: Cohen JI Epstein-Barr virus infection. N Engl J Med 343: Amon W, Farrell PJ Reactivation of Epstein-Barr virus from latency. Rev Med Virol 15: Rezk SA, Weiss LM Epstein-Barr virus-associated lymphoproliferative disorders. Hum Pathol 38: Gandhi MK, Tellam JT, Khanna R Epstein-Barr virus-associated Hodgkin's lymphoma. Br J Haematol 125: Marchini A, Kieff E, Longnecker R Marker rescue of a transformation-negative Epstein-Barr virus recombinant from an infected Burkitt lymphoma cell line: a method useful for analysis of genes essential for transformation. J Virol 67: Maruo S, Wu Y, Ishikawa S, Kanda T, Iwakiri D, Takada K Epstein- Barr virus nuclear protein EBNA3C is required for cell cycle progression and growth maintenance of lymphoblastoid cells. Proc Natl Acad Sci U S A 103: Halder S, Murakami M, Verma SC, Kumar P, Yi F, Robertson ES Early events associated with infection of Epstein-Barr virus infection of primary B-cells. PLoS One 4:e Wang F, Tsang SF, Kurilla MG, Cohen JI, Kieff E Epstein-Barr virus nuclear antigen 2 transactivates latent membrane protein LMP1. J Virol 64: Maruo S, Wu Y, Ito T, Kanda T, Kieff ED, Takada K Epstein-Barr virus nuclear protein EBNA3C residues critical for maintaining lymphoblastoid cell growth. Proc Natl Acad Sci U S A 106: Saha A, Bamidele A, Murakami M, Robertson ES EBNA3C attenuates the function of p53 through interaction with inhibitor of growth family proteins 4 and 5. J Virol 85: McClellan MJ, Khasnis S, Wood CD, Palermo RD, Schlick SN, Kanhere AS, Jenner RG, West MJ Downregulation of integrin receptor-signaling 34
35 genes by Epstein-Barr virus EBNA 3C via promoter-proximal and -distal binding elements. J Virol 86: Nikitin PA, Luftig MA At a crossroads: human DNA tumor viruses and the host DNA damage response. Future Virol 6: Choudhuri T, Verma SC, Lan K, Murakami M, Robertson ES The ATM/ATR signaling effector Chk2 is targeted by Epstein-Barr virus nuclear antigen 3C to release the G2/M cell cycle block. J Virol 81: Nikitin PA, Yan CM, Forte E, Bocedi A, Tourigny JP, White RE, Allday MJ, Patel A, Dave SS, Kim W, Hu K, Guo J, Tainter D, Rusyn E, Luftig MA An ATM/Chk2-mediated DNA damage-responsive signaling pathway suppresses Epstein-Barr virus transformation of primary human B cells. Cell Host Microbe 8: Summers KC, Shen F, Sierra Potchanant EA, Phipps EA, Hickey RJ, Malkas LH Phosphorylation: the molecular switch of double-strand break repair. Int J Proteomics 2011: Jha HC, Upadhyay SK, M AJP, Lu J, Cai Q, Saha A, Robertson ES H2AX phosphorylation is important for LANA-mediated Kaposi's sarcomaassociated herpesvirus episome persistence. J Virol 87: Ikura T, Tashiro S, Kakino A, Shima H, Jacob N, Amunugama R, Yoder K, Izumi S, Kuraoka I, Tanaka K, Kimura H, Ikura M, Nishikubo S, Ito T, Muto A, Miyagawa K, Takeda S, Fishel R, Igarashi K, Kamiya K DNA damage-dependent acetylation and ubiquitination of H2AX enhances chromatin dynamics. Mol Cell Biol 27: Turnell AS, Grand RJ DNA viruses and the cellular DNA-damage response. J Gen Virol 93: Bajaj BG, Murakami M, Cai Q, Verma SC, Lan K, Robertson ES Epstein-Barr virus nuclear antigen 3C interacts with and enhances the stability of the c-myc oncoprotein. J Virol 82: Knight JS, Sharma N, Robertson ES SCFSkp2 complex targeted by Epstein-Barr virus essential nuclear antigen. Mol Cell Biol 25: Lu J, Verma SC, Murakami M, Cai Q, Kumar P, Xiao B, Robertson ES Latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus (KSHV) upregulates survivin expression in KSHV-Associated B- lymphoma cells and contributes to their proliferation. J Virol 83: Rios-Doria J, Fay A, Velkova A, Monteiro AN DNA damage response: determining the fate of phosphorylated histone H2AX. Cancer Biol Ther 5: Kang MA, So EY, Ouchi T Deregulation of DNA damage response pathway by intercellular contact. J Biol Chem 287: Jha HC, Lu J, Saha A, Cai Q, Banerjee S, Prasad MA, Robertson ES EBNA3C-mediated regulation of aurora kinase B contributes to Epstein-Barr virus-induced B-cell proliferation through modulation of the activities of the retinoblastoma protein and apoptotic caspases. J Virol 87: Saha A, Lu J, Morizur L, Upadhyay SK, Aj MP, Robertson ES E2F1 mediated apoptosis induced by the DNA damage response is blocked by EBV nuclear antigen 3C in lymphoblastoid cells. PLoS Pathog 8:e
36 Yi F, Saha A, Murakami M, Kumar P, Knight JS, Cai Q, Choudhuri T, Robertson ES Epstein-Barr virus nuclear antigen 3C targets p53 and modulates its transcriptional and apoptotic activities. Virology 388: Gao J, Cai Q, Lu J, Jha HC, Robertson ES Upregulation of cellular Bcl-2 by the KSHV encoded RTA promotes virion production. PLoS One 6:e Saha A, Halder S, Upadhyay SK, Lu J, Kumar P, Murakami M, Cai Q, Robertson ES Epstein-Barr virus nuclear antigen 3C facilitates G1-S transition by stabilizing and enhancing the function of cyclin D1. PLoS Pathog 7:e Paull TT, Rogakou EP, Yamazaki V, Kirchgessner CU, Gellert M, Bonner WM A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Curr Biol 10: Tarakanova VL, Stanitsa E, Leonardo SM, Bigley TM, Gauld SB Conserved gammaherpesvirus kinase and histone variant H2AX facilitate gammaherpesvirus latency in vivo. Virology 405: Bassing CH, Suh H, Ferguson DO, Chua KF, Manis J, Eckersdorff M, Gleason M, Bronson R, Lee C, Alt FW Histone H2AX: a dosagedependent suppressor of oncogenic translocations and tumors. Cell 114: Celeste A, Fernandez-Capetillo O, Kruhlak MJ, Pilch DR, Staudt DW, Lee A, Bonner RF, Bonner WM, Nussenzweig A Histone H2AX phosphorylation is dispensable for the initial recognition of DNA breaks. Nat Cell Biol 5: Fernandez-Capetillo O, Lee A, Nussenzweig M, Nussenzweig A H2AX: the histone guardian of the genome. DNA Repair (Amst) 3: Ikura T, Ogryzko VV, Grigoriev M, Groisman R, Wang J, Horikoshi M, Scully R, Qin J, Nakatani Y Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis. Cell 102: Nawaz Z, Lonard DM, Dennis AP, Smith CL, O'Malley BW Proteasome-dependent degradation of the human estrogen receptor. Proc Natl Acad Sci U S A 96: Yang C, Wang H, Xu Y, Brinkman KL, Ishiyama H, Wong ST, Xu B The kinetochore protein Bub1 participates in the DNA damage response. DNA Repair (Amst) 11: Fragkos M, Jurvansuu J, Beard P H2AX is required for cell cycle arrest via the p53/p21 pathway. Mol Cell Biol 29: Lu C, Xiong M, Luo Y, Li J, Zhang Y, Dong Y, Zhu Y, Niu T, Wang Z, Duan L Genome-wide transcriptional analysis of apoptosis-related genes and pathways regulated by H2AX in lung cancer A549 cells. Apoptosis 18: Nikitin PA, Luftig MA The DNA damage response in viral-induced cellular transformation. Br J Cancer 106: Chaurushiya MS, Weitzman MD Viral manipulation of DNA repair and cell cycle checkpoints. DNA Repair (Amst) 8:
37 Chadwick BP, Willard HF Histone H2A variants and the inactive X chromosome: identification of a second macroh2a variant. Hum Mol Genet 10: Kinner A, Wu W, Staudt C, Iliakis G Gamma-H2AX in recognition and signaling of DNA double-strand breaks in the context of chromatin. Nucleic Acids Res 36: Li A, Eirin-Lopez JM, Ausio J H2AX: tailoring histone H2A for chromatin-dependent genomic integrity. Biochem Cell Biol 83: Kamranvar SA, Masucci MG The Epstein-Barr virus nuclear antigen- 1 promotes telomere dysfunction via induction of oxidative stress. Leukemia 25: Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K, Linn S Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem 73: Jha HC, Lu J, Saha A, Cai Q, Banerjee S, Prasad MA, Robertson ES EBNA3C-mediated regulation of Aurora Kinase B contributes to EBV-induced B-cell proliferation through modulation of the activities of Rb and apoptotic Caspases. J Virol 87: Banerjee S, Lu J, Cai Q, Saha A, Jha HC, Dzeng RK, Robertson ES The EBV Latent Antigen 3C Inhibits Apoptosis through Targeted Regulation of Interferon Regulatory Factors 4 and 8. PLoS Pathog 9:e Burma S, Chen BP, Murphy M, Kurimasa A, Chen DJ ATM phosphorylates histone H2AX in response to DNA double-strand breaks. J Biol Chem 276: Downs JA, Nussenzweig MC, Nussenzweig A Chromatin dynamics and the preservation of genetic information. Nature 447: Bewersdorf J, Bennett BT, Knight KL H2AX chromatin structures and their response to DNA damage revealed by 4Pi microscopy. Proc Natl Acad Sci U S A 103: Liu Y, Tseng M, Perdreau SA, Rossi F, Antonescu C, Besmer P, Fletcher JA, Duensing S, Duensing A Histone H2AX is a mediator of gastrointestinal stromal tumor cell apoptosis following treatment with imatinib mesylate. Cancer Res 67: Bassing CH, Alt FW H2AX may function as an anchor to hold broken chromosomal DNA ends in close proximity. Cell Cycle 3: Bassing CH, Chua KF, Sekiguchi J, Suh H, Whitlow SR, Fleming JC, Monroe BC, Ciccone DN, Yan C, Vlasakova K, Livingston DM, Ferguson DO, Scully R, Alt FW Increased ionizing radiation sensitivity and genomic instability in the absence of histone H2AX. Proc Natl Acad Sci U S A 99: Saha A, Murakami M, Kumar P, Bajaj B, Sims K, Robertson ES Epstein-Barr virus nuclear antigen 3C augments Mdm2-mediated p53 ubiquitination and degradation by deubiquitinating Mdm2. J Virol 83: Feng L, Chen J The E3 ligase RNF8 regulates KU80 removal and NHEJ repair. Nat Struct Mol Biol 19:
38 Bergink S, Jentsch S Principles of ubiquitin and SUMO modifications in DNA repair. Nature 458: Lukas J, Bartek J DNA repair: New tales of an old tail. Nature 458: Xiao B, Verma SC, Cai Q, Kaul R, Lu J, Saha A, Robertson ES Bub1 and CENP-F can contribute to Kaposi's sarcoma-associated herpesvirus genome persistence by targeting LANA to kinetochores. J Virol 84: Downloaded from on December 15, 2018 by guest 38
39
40
41
42
43
44
45
46
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
EBV infection B cells and lymphomagenesis. Sridhar Chaganti
 EBV infection B cells and lymphomagenesis Sridhar Chaganti How EBV infects B-cells How viral genes influence the infected B cell Differences and similarities between in vitro and in vivo infection How
EBV infection B cells and lymphomagenesis Sridhar Chaganti How EBV infects B-cells How viral genes influence the infected B cell Differences and similarities between in vitro and in vivo infection How
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
SUPPLEMENTAL FIGURE LEGENDS
 SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh-
 or GSK3b-specific shrna (sh-") 1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Supplementary Figure 1
 Supplementary Figure 1 a γ-h2ax MDC1 RNF8 FK2 BRCA1 U2OS Cells sgrna-1 ** 60 sgrna 40 20 0 % positive Cells (>5 foci per cell) b ** 80 sgrna sgrna γ-h2ax MDC1 γ-h2ax RNF8 FK2 MDC1 BRCA1 RNF8 FK2 BRCA1
Supplementary Figure 1 a γ-h2ax MDC1 RNF8 FK2 BRCA1 U2OS Cells sgrna-1 ** 60 sgrna 40 20 0 % positive Cells (>5 foci per cell) b ** 80 sgrna sgrna γ-h2ax MDC1 γ-h2ax RNF8 FK2 MDC1 BRCA1 RNF8 FK2 BRCA1
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Supplementary Information
 Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
Supplementary Information Supplementary Figure 1. EBV-gB 23-431 mainly exists as trimer in HEK 293FT cells. (a) Western blotting analysis for DSS crosslinked FLAG-gB 23-431. HEK 293FT cells transfected
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v)
") SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
Supplementary material. Supplementary Figure legends
 Supplementary material Supplementary Figure legends Supplementary Figure 1: Senescence-associated proliferation stop in response to oncogenic N-RAS expression Proliferation of NHEM cells without (ctrl.)
Supplementary material Supplementary Figure legends Supplementary Figure 1: Senescence-associated proliferation stop in response to oncogenic N-RAS expression Proliferation of NHEM cells without (ctrl.)
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis
 Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis") Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer
 AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
SUPPLEMENTARY INFORMATION
 Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Supplementary Figure 1
 Supplementary Figure 1 YAP negatively regulates IFN- signaling. (a) Immunoblot analysis of Yap knockdown efficiency with sh-yap (#1 to #4 independent constructs) in Raw264.7 cells. (b) IFN- -Luc and PRDs
Supplementary Figure 1 YAP negatively regulates IFN- signaling. (a) Immunoblot analysis of Yap knockdown efficiency with sh-yap (#1 to #4 independent constructs) in Raw264.7 cells. (b) IFN- -Luc and PRDs
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
Supplementary Information. Supplementary Figure 1
 Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Supplementary Figure 1. MAT IIα is Acetylated at Lysine 81.
 IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
(A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939 (5 µm)
 SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939 (5 µm)") Supplementary Figure Legends Figure S1. Tankyrase inhibition suppresses cell proliferation in an axin/β-catenin independent manner. (A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939
Supplementary Figure Legends Figure S1. Tankyrase inhibition suppresses cell proliferation in an axin/β-catenin independent manner. (A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators
 /, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
/, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
S1a S1b S1c. S1d. S1f S1g S1h SUPPLEMENTARY FIGURE 1. - si sc Il17rd Il17ra bp. rig/s IL-17RD (ng) -100 IL-17RD
 -100 IL-17RD") SUPPLEMENTARY FIGURE 1 0 20 50 80 100 IL-17RD (ng) S1a S1b S1c IL-17RD β-actin kda S1d - si sc Il17rd Il17ra rig/s15-574 - 458-361 bp S1f S1g S1h S1i S1j Supplementary Figure 1. Knockdown of IL-17RD enhances
SUPPLEMENTARY FIGURE 1 0 20 50 80 100 IL-17RD (ng) S1a S1b S1c IL-17RD β-actin kda S1d - si sc Il17rd Il17ra rig/s15-574 - 458-361 bp S1f S1g S1h S1i S1j Supplementary Figure 1. Knockdown of IL-17RD enhances
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
supplementary information
 DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Electron micrograph of phosphotungstanic acid-stained exosomes derived from murine
 1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplemental Materials. STK16 regulates actin dynamics to control Golgi organization and cell cycle
 Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
Supplemental Materials STK16 regulates actin dynamics to control Golgi organization and cell cycle Juanjuan Liu 1,2,3, Xingxing Yang 1,3, Binhua Li 1, Junjun Wang 1,2, Wenchao Wang 1, Jing Liu 1, Qingsong
Mitochondrial DNA Isolation Kit
 Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Activation of cellular proto-oncogenes to oncogenes. How was active Ras identified?
 Dominant Acting Oncogenes Eugene E. Marcantonio, M.D. Ph.D. Oncogenes are altered forms of normal cellular genes called proto-oncogenes that are involved in pathways regulating cell growth, differentiation,
Dominant Acting Oncogenes Eugene E. Marcantonio, M.D. Ph.D. Oncogenes are altered forms of normal cellular genes called proto-oncogenes that are involved in pathways regulating cell growth, differentiation,
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
Supplemental material Chairoungdua et al., http://www.jcb.org/cgi/content/full/jcb.201002049/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Expression of CD9 and CD82 inhibits Wnt/ -catenin
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Functional characterisation of hepatitis B viral X protein/microrna-21 interaction in HBVassociated hepatocellular carcinoma
 RESEARCH FUND FOR THE CONTROL OF INFECTIOUS DISEASES Functional characterisation of hepatitis B viral X protein/microrna-21 interaction in HBVassociated hepatocellular carcinoma CH Li, SC Chow, DL Yin,
RESEARCH FUND FOR THE CONTROL OF INFECTIOUS DISEASES Functional characterisation of hepatitis B viral X protein/microrna-21 interaction in HBVassociated hepatocellular carcinoma CH Li, SC Chow, DL Yin,
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Mammalian Tissue Protein Extraction Reagent
 Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
33VASTVNGATSANNHGEPPS51PADARPR58
 Pro-rich region Trans-membrane region 214 246 359 381 UL50 1 397 211SSRTAS216PPPPPR222 NLS CR1 CR2 CR3 CR4 UL53 1 376 11RERRS15ALRS19LLRKRRR25 33VASTVNGATSANNHGEPPS51PADARPR58 FIG S1. UL97 phosphorylation
Pro-rich region Trans-membrane region 214 246 359 381 UL50 1 397 211SSRTAS216PPPPPR222 NLS CR1 CR2 CR3 CR4 UL53 1 376 11RERRS15ALRS19LLRKRRR25 33VASTVNGATSANNHGEPPS51PADARPR58 FIG S1. UL97 phosphorylation
MOLECULAR BASIS OF ONCOGENESIS
 MOLECULAR BASIS OF ONCOGENESIS MUDr. Jiří Vachtenheim, CSc. 1 Cell processes which result also in cell cycle effects. Differentiation. Differentiated cells are usually in the G0 phase of the cell cycle.
MOLECULAR BASIS OF ONCOGENESIS MUDr. Jiří Vachtenheim, CSc. 1 Cell processes which result also in cell cycle effects. Differentiation. Differentiated cells are usually in the G0 phase of the cell cycle.
BIO360 Quiz #1. September 14, Name five of the six Hallmarks of Cancer (not emerging hallmarks or enabling characteristics ): (5 points)
: (5 points)") Name: BIO360 Quiz #1 September 14, 2012 1. Name five of the six Hallmarks of Cancer (not emerging hallmarks or enabling characteristics ): (5 points) 2. The controversial hypothesis that only a small subset
Name: BIO360 Quiz #1 September 14, 2012 1. Name five of the six Hallmarks of Cancer (not emerging hallmarks or enabling characteristics ): (5 points) 2. The controversial hypothesis that only a small subset
Supplemental information
 Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental
Carcinoemryonic antigen-related cell adhesion molecule 6 (CEACAM6) promotes EGF receptor signaling of oral squamous cell carcinoma metastasis via the complex N-glycosylation y Chiang et al. Supplemental
293T cells were transfected with indicated expression vectors and the whole-cell extracts were subjected
 SUPPLEMENTARY INFORMATION Supplementary Figure 1. Formation of a complex between Slo1 and CRL4A CRBN E3 ligase. (a) HEK 293T cells were transfected with indicated expression vectors and the whole-cell
SUPPLEMENTARY INFORMATION Supplementary Figure 1. Formation of a complex between Slo1 and CRL4A CRBN E3 ligase. (a) HEK 293T cells were transfected with indicated expression vectors and the whole-cell
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3076 Supplementary Figure 1 btrcp targets Cep68 for degradation during mitosis. a) Cep68 immunofluorescence in interphase and metaphase. U-2OS cells were transfected with control sirna
DOI: 10.1038/ncb3076 Supplementary Figure 1 btrcp targets Cep68 for degradation during mitosis. a) Cep68 immunofluorescence in interphase and metaphase. U-2OS cells were transfected with control sirna
Supplementary Figures
 Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
Supplementary Figures Supplementary Figure 1 Characterization of stable expression of GlucB and sshbira in the CT26 cell line (a) Live cell imaging of stable CT26 cells expressing green fluorescent protein
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Cell Lysis Buffer. Catalog number: AR0103
 Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
supplementary information
 DOI: 10.1038/ncb2153 Figure S1 Ectopic expression of HAUSP up-regulates REST protein. (a) Immunoblotting showed that ectopic expression of HAUSP increased REST protein levels in ENStemA NPCs. (b) Immunofluorescent
DOI: 10.1038/ncb2153 Figure S1 Ectopic expression of HAUSP up-regulates REST protein. (a) Immunoblotting showed that ectopic expression of HAUSP increased REST protein levels in ENStemA NPCs. (b) Immunofluorescent
Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535
 Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535 Product Description Recombinant Jurkat T cell expressing firefly luciferase gene under the control of NFAT response elements with constitutive
Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535 Product Description Recombinant Jurkat T cell expressing firefly luciferase gene under the control of NFAT response elements with constitutive
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts
 Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts (TIG-3 cells) were rendered senescent by either serial passage
Supplementary Figure 1 Induction of cellular senescence and isolation of exosome. a to c, Pre-senescent primary normal human diploid fibroblasts (TIG-3 cells) were rendered senescent by either serial passage
Supplemental Information
 Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Nature Immunology doi: /ni.3268
 Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
Supplementary Figure 1 Loss of Mst1 and Mst2 increases susceptibility to bacterial sepsis. (a) H&E staining of colon and kidney sections from wild type and Mst1 -/- Mst2 fl/fl Vav-Cre mice. Scale bar,
BIT 120. Copy of Cancer/HIV Lecture
 BIT 120 Copy of Cancer/HIV Lecture Cancer DEFINITION Any abnormal growth of cells that has malignant potential i.e.. Leukemia Uncontrolled mitosis in WBC Genetic disease caused by an accumulation of mutations
BIT 120 Copy of Cancer/HIV Lecture Cancer DEFINITION Any abnormal growth of cells that has malignant potential i.e.. Leukemia Uncontrolled mitosis in WBC Genetic disease caused by an accumulation of mutations
Appendix. Table of Contents
 Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway
 Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Expanded View Figures
 MO reports PR3 dephosphorylates TZ Xian-o Lv et al xpanded View igures igure V1. PR3 dephosphorylates and inactivates YP/TZ., Overexpression of tight junction proteins Pals1 () or LIN7 () has no effect
MO reports PR3 dephosphorylates TZ Xian-o Lv et al xpanded View igures igure V1. PR3 dephosphorylates and inactivates YP/TZ., Overexpression of tight junction proteins Pals1 () or LIN7 () has no effect
Protocol for purification of recombinant protein from 300 ml yeast culture
 Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Pre-made Reporter Lentivirus for NF-κB Signal Pathway
 Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
Pre-made Reporter for NF-κB Signal Pathway Cat# Product Name Amounts LVP965-P or: LVP965-P-PBS NFKB-GFP (Puro) LVP966-P or: LVP966-P-PBS NFKB-RFP (Puro) LVP967-P or: LVP967-P-PBS NFKB-Luc (Puro) LVP968-P
condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1%
 FIGURE LEGENDS Supplementary Fig 1 (A) sumoylation pattern detected under denaturing condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1% SDS in the presence and absence
FIGURE LEGENDS Supplementary Fig 1 (A) sumoylation pattern detected under denaturing condition. Left panel, the HCT-116 cells were lysed with RIPA buffer containing 0.1% SDS in the presence and absence
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3
 Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
Purification and Fluorescent Labeling of Exosomes Asuka Nanbo 1*, Eri Kawanishi 2, Ryuji Yoshida 2 and Hironori Yoshiyama 3 1 Graduate School of Medicine, Hokkaido University, Sapporo, Japan; 2 Graduate
Genome of Hepatitis B Virus. VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department
 Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters.
 The Nuclear Chaperone Nucleophosmin Escorts an Epstein-Barr Virus Nuclear Antigen to Establish Transcriptional Cascades for Latent Infection in Human B Cells The Harvard community has made this article
The Nuclear Chaperone Nucleophosmin Escorts an Epstein-Barr Virus Nuclear Antigen to Establish Transcriptional Cascades for Latent Infection in Human B Cells The Harvard community has made this article
p.r623c p.p976l p.d2847fs p.t2671 p.d2847fs p.r2922w p.r2370h p.c1201y p.a868v p.s952* RING_C BP PHD Cbp HAT_KAT11
 ARID2 p.r623c KMT2D p.v650fs p.p976l p.r2922w p.l1212r p.d1400h DNA binding RFX DNA binding Zinc finger KMT2C p.a51s p.d372v p.c1103* p.d2847fs p.t2671 p.d2847fs p.r4586h PHD/ RING DHHC/ PHD PHD FYR N
ARID2 p.r623c KMT2D p.v650fs p.p976l p.r2922w p.l1212r p.d1400h DNA binding RFX DNA binding Zinc finger KMT2C p.a51s p.d372v p.c1103* p.d2847fs p.t2671 p.d2847fs p.r4586h PHD/ RING DHHC/ PHD PHD FYR N
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document
 Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
(A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a
 RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a") Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the
 Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the") Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the genome-wide methylation microarray data. Mean ± s.d.; Student
Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the genome-wide methylation microarray data. Mean ± s.d.; Student