Constitutively active Notch4 receptor elicits brain arteriovenous malformations through enlargement of capillary-like vessels.
|
|
|
- Dana Harrison
- 5 years ago
- Views:
Transcription
1 Constitutively active Notch4 receptor elicits brain arteriovenous malformations through enlargement of capillary-like vessels Patrick A. Murphy a,1,2, Tyson N. Kim a,1, Lawrence Huang a,1, Corinne M. Nielsen a, Michael T. Lawton b, Ralf H. Adams c, Chris B. Schaffer d, and Rong A. Wang a,3 a Laboratory for Accelerated Vascular Research, Department of Surgery, Division of Vascular Surgery, University of California, San Francisco, CA 94143; b Department of Neurosurgery, University of California, San Francisco, CA 94143; c Max Planck Institute for Molecular Biomedicine, Faculty of Medicine, Department of Tissue Morphogenesis, University of Münster, D Münster, Germany; and d Department of Biomedical Engineering, Cornell University, Ithaca, NY Edited by Michael A. Gimbrone, Brigham and Women s Hospital, Boston, MA, and approved November 11, 2014 (received for review August 13, 2014) Arteriovenous (AV) malformation (AVM) is a devastating condition characterized by focal lesions of enlarged, tangled vessels that shunt blood from arteries directly to veins. AVMs can form anywhere in the body and can cause debilitating ischemia and life-threatening hemorrhagic stroke. The mechanisms that underlie AVM formation remain poorly understood. Here, we examined the cellular and hemodynamic changes at the earliest stages of brain AVM formation by time-lapse two-photon imaging through cranial windows of mice expressing constitutively active Notch4 (Notch4*). AVMs arose from enlargement of preexisting microvessels with capillary diameter and blood flow and no smooth muscle cell coverage. AV shunting began promptly after Notch4* expression in endothelial cells (ECs), accompanied by increased individual EC areas, rather than increased EC number or proliferation. Alterations in Notch signaling in ECs of all vessels, but not arteries alone, affected AVM formation, suggesting that Notch functions in the microvasculature and/or veins to induce AVM. Increased Notch signaling interfered with the normal biological control of hemodynamics, permitting a positive feedback loop of increasing blood flow and vessel diameter and driving focal AVM growth from AV connections with higher blood velocity at the expense of adjacent AV connections with lower velocity. Endothelial expression of constitutively active Notch1 also led to brain AVMs in mice. Our data shed light on cellular and hemodynamic mechanisms underlying AVM pathogenesis elicited by increased Notch signaling in the endothelium. vascular anomaly angiogenesis stroke hereditary hemorrhagic telangiectasia Notch Proper vascular perfusion requires a capillary interface between arteries and veins. Establishing and preserving the arteriovenous (AV) interface are critical for tissue function. AV malformation (AVM) is a pathological perturbation of the AV interface and is characterized by enlarged, tangled vessels that shunt blood directly from arteries to veins lacking intervening capillaries (1). Brain AVMs can cause ischemia, intracerebral hemorrhage, disability, stroke, and death. About half are first diagnosed between 20 and 40 y of age (1), accounting for 50% of childhood stroke (2). The developmental mechanisms of AVM pathogenesis remain poorly understood, hindering the development of therapeutic treatment strategies. Clinical evidence has led to two theories about the origin of brainavm.first,brainavmsarecongenital and caused by failed regression of embryonic AV connections that persist into postnatal life (3). This theory is based on morphological interpretation of AVMs that resemble primitive vascular structures. However, the lack of detection by prenatal ultrasonography suggests that AVMs are either too small to detect in fetal stages or they develop after birth. The second theory suggests that AVMs result from improper remodeling of venules (4). Examination of biopsies from skin telangiectases in humans suggests that enlargement of postcapillary venules precedes AVM formation and can occur in postnatal life (5). However, the critical precipitating event remains unknown. Loss-of-function mutations identified in the familial disease Hereditary Hemorrhagic Telangectasia (HHT), including the TGF-β family receptor Alk1 (HHT2) (6) and its coreceptor endoglin (HHT1) (7), have provided valuable models to study the molecular basis of AVMs. Genetic deletion of each of these genes has revealed that they are required for normal vascular development and prevention of AVM formation (6 8). Mutations in SMAD4, RASA1, andpten have also been linked to AVMs in humans (9, 10), and mutations in Rasa1 lead to AV shunting in zebrafish (11). The Notch receptors are transmembrane proteins that promote arterial endothelial cell (EC) specification by enhancing expression of arterial molecular markers and suppressing the expression of venous markers (12 19). Abnormal signaling induces enlarged AV connections and shunting in mouse and zebrafish embryos (12 14). Notch activity is aberrantly increased in the endothelium of human brain AVMs (18, 20), suggesting that it may participate in the growth or maintenance of human AVMs. These findings offer exciting opportunities to understand the molecular mechanisms of AVM formation. We have previously reported a mouse model, Tie2-tTA;TRE-Notch4* or Tie2-Notch4* mice, wherein postnatal expression of constitutively active Notch4 (Notch4*) in Significance Brain arteriovenous malformations are focal lesions of enlarged, tangled vessels that shunt blood from arteries directly to veins. They can cause ischemia, hemorrhage, disability, and death, particularly in young people, accounting for 50% of childhood stroke. The molecular etiology of the disease remains poorly understood, hindering the development of therapeutic treatments. Here, we report that, in an animal model, the lesion arises from the enlargement of capillary-like vessels. Notch signaling in the endothelium of microvasculature and veins is critical for the disease initiation by increasing cell areas but not proliferation. Blood flow mediates disease progression by a positive feedback of increasing flow and vessel diameter. Our data shed light on the mechanism underlying the pathogenesis of this devastating disease. Author contributions: P.A.M., T.N.K., L.H., C.M.N., C.B.S., and R.A.W. designed research; P.A.M., T.N.K., L.H., and C.M.N. performed research; R.A.W. supervised the project; T.N.K., R.H.A., and C.B.S. contributed new reagents/analytic tools; P.A.M., T.N.K., L.H., C.M.N., M.T.L., C.B.S., and R.A.W. analyzed data; M.T.L. provided medical relevance; and P.A.M., T.N.K., L.H., and R.A.W. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 P.A.M., T.N.K., and L.H. contributed equally to this work. 2 Present address: Koch Institute for Integrative Cancer Research, Massachusetts Institute of Technology, Cambridge, MA To whom correspondence should be addressed. rong.wang@ucsfmedctr.org. This article contains supporting information online at /pnas /-/DCSupplemental. MEDICAL SCIENCES PNAS December 16, 2014 vol. 111 no

2 ECs results in spontaneous AVMs in mice (15 17, 21). Although this work has facilitated new research directions regarding the Notch pathway in AVMs, how aberrant Notch signaling leads to AVM remains unknown. Elucidating the initiating structural events that lead to AV shunt formation would provide insight into AVM pathogenesis. A major obstacle to understanding AVM pathogenesis has been the inability to observe AVM formation and blood flow over time with high-resolution in vivo imaging. Here, we examined AVM formation in Tie2-Notch4* mice from the initial genetic event, illuminating disease progression using 5D two-photon imaging, which allows high-resolution live imaging of vascular architecture (3D) and blood velocity (the fourth dimension) over time (the fifth dimension) (16, 22, 23). Our findings provide insights into the mechanism of brain AVM formation. Results Notch4*-Mediated AV Shunts Arise from the Enlargement of Capillary-Like Vessels. To determine the developmental origin of AVMs, we performed longitudinal live imaging in the brains of Tie2-Notch4* mice, in which Notch4* expression was repressed until birth by tetracycline treatment. We implanted cranial windows over the right parietal cortex of mice at postnatal day 7 (P7) and used in vivo two-photon microscopy to record vessel diameter and blood velocity over time. We measured lumen diameters of vessels connecting arteries and veins, or AV connections, at their narrowest point. We defined measurements 12.5 μm as AV shunts because AV connections of this diameter were not observed in control mice after P12 (SI Appendix, Fig. S1). We found that AV shunts developed from capillary-like vessels in Tie2- Notch4* mice beginning at P12 (Fig. 1 and SI Appendix,Figs.S2 and S3). Of 109 of the AV connections that we tracked, 41 (38%) grew to 12.5 μm between P14 and P25 in 12 mutants, and 4 of 109 (4%) grew to 30 μm (Fig. 1A and Movies S1 and S2). However, the initial diameter did not differ from that of controls (5.4 ± 1.6 vs. 5.3 ± 1.6 μm, Fig. 1B). We saw no correlation between the final and initial diameters of AV shunts (SI Appendix, Fig. S4). We conclude that AV shunts in Tie2-Notch4* mice enlarge from capillary-diameter AV connections, but that only a few continue to grow into large AVMs. We examined the hemodynamics of AV shunts at their onset and as they enlarged. Using 5D two-photon imaging (16, 23), we simultaneously analyzed red blood cell velocity and AV connection diameter (Fig. 1 C E and SI Appendix, Fig.S2). AV Fig. 1. Tie2-tTA;TRE-Notch4* mice developed AV shunts through enlargement of capillary-like vessels. (A and A ) Two-photon time-lapse imaging of FITCdextran labeled AV connections in the cerebral cortex through a cranial window. An AV shunt (red arrowheads and green arrows) developing from a capillary-diameter AV connection between P14 and P19. (B) Distribution of initial AV connection diameter (smallest lumen diameter) in AV connections in mutants (109 AV connections in 12 mice) and controls (36 AV connections in 5 mice, ages P8 P11). (C and D) Blood velocities (dotted lines) and lumen diameters (solid lines) measured over time in the capillary-like AV connections of (C1 C3) Notch4* mice and (D) controls. (E) Time of initial velocity increase vs. time of initial diameter increase for individual AV connections. Dotted red line represents simultaneous increase of blood velocity and vessel diameter. Points on the dotted vertical black line represent capillary-like vessels that enlarged but did not exhibit a significant increase in velocity. Increased velocity and diameter were defined by changes >2 the SD in control capillaries: 0.82 mm/s for velocity and 1.33 μm for diameter. (F and G) Whole-mount immunostaining of P12 surface cerebral cortex vasculature for α-smooth muscle actin (green) and VE-cadherin (red) in mutant (F) and control (G). Arrowheads indicate AV shunt (18 AV shunts in seven mice); corresponding lectin perfused vessels are shown (F and G ). (Scale bars, 50 μm.) Murphy et al.
, whereas others exhibited an increase in neither diameter nor velocity (Fig.")
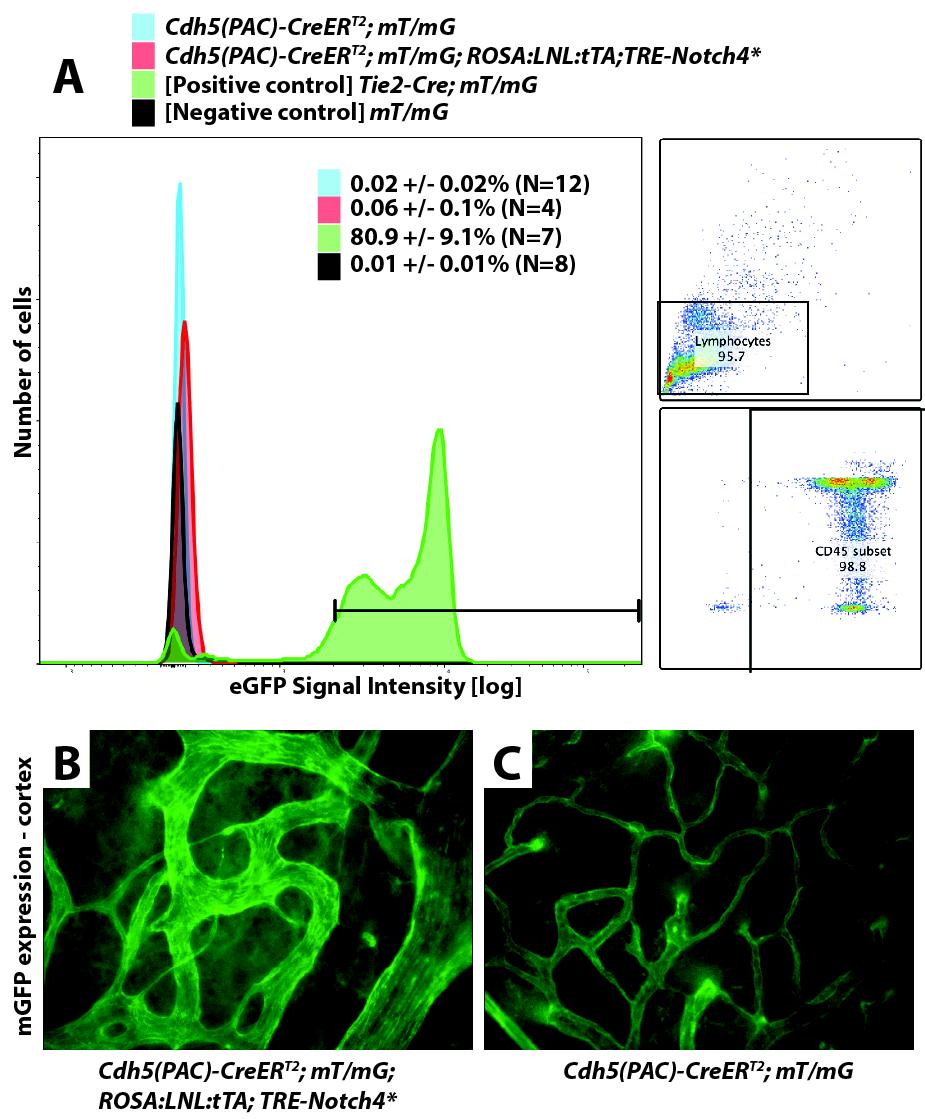
3 connection enlargement either coincided with or preceded the initial increase in velocity (Fig. 1C1, 8/18 vessels in four mice). Some capillary-like vessels enlarged without a significant increase in velocity (Fig. 1C2, 4/18 vessels in four mice), whereas others exhibited an increase in neither diameter nor velocity (Fig. 1C3, 6/18 vessels in four mice). Neither velocity nor diameter increased in controls (Fig. 1D, 18 vessels in four mice). Data are summarized in Fig. 1E. Thus, our data suggest that blood velocity in AV connections that grew into AV shunts is initially capillary-like and becomes abnormally elevated only when connections enlarge. As capillaries lack smooth muscle cell (SMC) coverage, we performed staining for α-smooth muscle actin (SMA) at P12, when AV shunts were first apparent. We identified 10 AV shunts from five mice that lacked SMA staining (Fig. 1F). In contrast, adjacent arteries were positive for SMA staining, similar to controls (Fig. 1G and SI Appendix, Fig. S5). We observed, as we have previously published (15 17), that advanced AVMs exhibited increased SMA staining. Thus, the absence of SMCs in the initial AV shunts is consistent with a capillary origin of AVMs in the Tie2-Notch4* mice. We next examined the expression of Notch4* during the initiation of AV shunting. We previously demonstrated that Tie2-tTA effectively and specifically drives gene expression throughout the brain endothelium, through activation of the tetracycline responsive element (TRE) by the tetracycline transactivator (tta) (17). Here we performed whole-mount staining of brain vasculature for the Notch4 intracellular domain (ICD), which also detects Notch4*, without the extracellular domain. We detected elevated Notch4-ICD throughout vasculature (reflecting Notch4* expression) in the mutants over the controls (SI Appendix, Fig. S6). To examine the kinetics of Notch4* expression in real time during AV shunt formation, we analyzed TRE-H2b-eGFP; Tie2- tta; TRE-Notch4* mice. TRE-H2b-eGFP reports tta activity and thus TRE-Notch4* expression. As expected, there was a positive correlation between H2b-eGFP intensity and Notch4* expression in fixed samples, validating the reporter assay (SI Appendix, Fig. S7). We found that tta activity preceded, by about 2 d, the enlargement in each segment (branch point to branch point) of the AV connections measured (Fig. 2 A C). Approximately 80% (159/ 198) of segments were positive for the TRE-H2b-eGFP reporter (Fig. 2D and SI Appendix, Fig.S8). About 50% (83/159) of the positive segments enlarged, and 40% (59/159) of these became part of AV shunts 12.5 μm in diameter. Only 15% (24/159) regressed (Fig. 2D and SI Appendix, Fig. S8). In contrast, only 10% of the H2b-eGFP negative segments enlarged (4/39); 80% (31/39) regressed. These data suggest that Notch4* expression promptly leads to AV shunting. EC Area, but Not Number or Proliferation, Increased in AV Shunt Formation. To examine the cellular changes underlying the initiation of AV shunts, we used the R26R-confetti reporter (24) to track the position and number of labeled cells. We activated the reporter from P1 to P5 by tamoxifen (TAM) induction of Cdh5 (PAC)-CreERT2 in Tie2-tTA;TRE-Notch4*; Cdh5(PAC)-CreERT2; R26R-Confetti mice and littermate controls (with either TRE- Notch4* or Tie2-tTA, but not both), allowing the tracking of ECs during AV shunt formation (Fig. 3A). Surprisingly, we saw no detectable increase in cell number in the initial 48 h of AV shunt formation (Fig. 3 A and B and SI Appendix, Fig. S9), despite a two- to threefold increase in AV shunt diameter (Fig. 3B). Because our analysis was powered to detect as little as a twofold difference in cell number, these data suggest that AV shunt growth does not occur through a simple increase in EC number. To investigate whether cell proliferation is involved in the initiation of AV shunts, we performed BrdU analysis of tomato-lectin labeled ECs. This assay showed no significant difference in the number of BrdU+ ECs (labeled for 48 h leading up to the analysis, Fig. 3 C and D and SI Appendix, Fig. S10). Furthermore, we found no BrdU+ EC cells in 10/15 AV shunts with diameters 12.5 μm, again suggesting that the initiation of the AV shunt does not require increased EC proliferation. Fig. 2. Notch4 expression increased in brain capillaries before enlargement of AV shunts. (A and A ) Two-photon imaging of an AV connection from P11 to P16; Texas-Red dextran labels plasma (red) and TRE-H2b-eGFP indicates tta activity (green). (B1 B4) Diameter vs. number of GFP+ cells in vessel segments: increased (B1 and B2), stable (B3), and decreased (B4) diameter. (C) Correlation between time of initial detection of GFP+ cells and initial diameter increase in capillary-diameter vessel segments of Notch4* mice. Gray regions indicate that GFP+ cells were not detected or diameter increase was not detected or both. Dotted line represents simultaneous detection of GFP+ cells and diameter increase. The indicates the means. GFP was detected before (125/198 vessel segments, five mice), or simultaneous with enlargement (19/198) in most cases. (D) Percentage of vessel segments (TREeGFP+ or negative) that demonstrated an increase, decrease, or no change in diameter or that completely regressed. Increased or decreased diameter is defined by >2 the SD of change in control vessels (1.1 μm) in the same period; segments are defined by branch points. (Scale bars: A, 100μm; C, 50μm.) We hypothesized that a change in the area covered by individual ECs might support the capillary enlargement in Tie2-Notch4* mice. To test this, we stained whole-mount cortical sections of mutants and controls for vascular endothelial cadherin (VE-cadherin, Cdh5) during the initial stages of AV shunt formation (P12). We found that the area covered by individual ECs increased more than threefold during the early stages of AV shunt formation (Fig. 3 E and F, mean cell area of 157 ± 24 μm 2 in three Tie2-tTA mice and 540 ± 46 μm 2 in five Tie2-tTA;TRE-Notch4* mice with AV shunts 12.5 μm; P = by two-tailed Student t test). Thus, an increase in EC area correlates with the enlargement of capillaries in AV shunt formation. Notch4* Expression in ECs of Arteries Was Not Sufficient to Induce AV Shunt Formation. We asked where in the vascular tree Notch4* acts to elicit AV shunt formation. Our in vivo imaging suggested that a defect in the capillaries or microcirculation led to AV shunts. Because no capillary EC-specific tta driver has been reported, we used Cdh5(PAC)-CreERT2 to induce Notch4* in all ECs and a recently developed arterial-specific line, BMX(PAC)-CreERT2, to examine the effects of Notch4* specifically in the arterial ECs. We first confirmed the expression of the two inducible Cre lines in our system. Cdh5(PAC)-CreERT2 efficiently activated a mt/mg Cre-reporter throughout the vascular endothelium, but not in circulating blood cells (SI Appendix, Figs. S11 and S12). The ROSA:LNL:tTA system effectively induced Notch4* expression, resulting in even higher protein levels than Tie2-tTA driven expression (SI Appendix, Fig. S13 and Movie S3) (25). As demonstrated in other tissues (26), BMX(PAC)-CreERT2 efficiently activated the same reporter throughout the arterial trunk MEDICAL SCIENCES Murphy et al. PNAS December 16, 2014 vol. 111 no
. Therefore, expression of Notch4* in the endothelium of BMX-CreERT2 positive arteries is not sufficient to induce AV shunts in the brain. Fig. 3.")
 Two-photon time-lapse imaging showing Cdh5 (PAC)-CreER; Confetti-marked nuclear GFP+ and cytoplasmic YFP+ ECs in cerebral cortex of mutants and controls. Texas-Red dextran labels plasma.")
4 connections were enlarged. By contrast, the BMX-CreERT2;ROSA: LNL:tTA;TRE-Notch4* mice resembled the negative littermate controls with only 2.1 ± 3.6% of total AV connections enlarged (Fig. 4 B and C and SI Appendix, Table S1). Therefore, expression of Notch4* in the endothelium of BMX-CreERT2 positive arteries is not sufficient to induce AV shunts in the brain. Fig. 3. Endothelial growth in the AV shunt accompanied by the expansion of endothelial cell surface area. (A) Two-photon time-lapse imaging showing Cdh5 (PAC)-CreER; Confetti-marked nuclear GFP+ and cytoplasmic YFP+ ECs in cerebral cortex of mutants and controls. Texas-Red dextran labels plasma. (B) Quantification of total cell number and AV connection diameter over the same interval (22 AV connections in five mutants, 11 AV connections in three controls). (C) Whole-mount BrdU staining/lectin perfusion. P12 AV connection from mutant cerebral cortex is shown. Blue dots indicate Hoechst+/lectin+ cells. White dots indicate BrdU+/lectin+ cells. (D) Quantification of BrdU+ ECs, total ECs, and AV connection diameter (34 connections in controls, 20 AV connections <12.5 μm in five mutants, and 15 AV connections 12.5 μm in four mutants). Gray indicates all Hoechst+/lectin+ cells, and the colored portion indicates the number of BrdU+/lectin+ cells. (E) Whole-mount staining of P12 cerebral cortex for VE-cadherin, counterstained by lectin perfusion. (F) Correlation of size of VE-cadherin traced cells in P12 control (blue) and Notch4* (red) connections with AV connection diameter (24 AV connections in 11 mutants, 6 AV connections in three controls). (Scale bars: A, 100μm; C and E, 25μm.) Rbpj in the ECs of BMX-CreERT2 Negative, but Not BMX-CreERT2 Positive Arteries, Was Required for Notch4*-Induced AV Shunt Formation. To investigate the molecular mediator of Notch4*- induced AV shunt formation, we examined Rbpj, a transcription factor required for canonical Notch signaling. We deleted Rbpj in Tie2-tTA;TRE-Notch4*;Cdh5(PAC)-CreERT2;Rbpj f/f mice before the onset of AV shunts by injecting TAM at P7 P8. Excision of Rbpj using Cdh5(PAC)-CreERT2 effectively suppressed AV shunt formation, and only 1.2 ± 1.7% of total AV connections were enlarged by P18 (Fig. 4D and SI Appendix, Table S2). In contrast, in controls with one remaining Rbpj allele, 22.3 ± 17.2% of total AV connections were enlarged (Fig. 4E and SI Appendix, Table S2). Thus, Notch4*-induced AV shunts require canonical Notch signaling via Rbpj. Because AV shunts were Rbpj-dependent, we tested whether blocking arterial Notch signaling suppresses AV shunt formation. We deleted Rbpj by injecting TAM at P7 P8 in arterial ECs of Tie2-tTA;TRE-Notch4*; BMX(PAC)-CreERT2;Rbpj f/f mice. Excision of both Rbpj alleles using BMX(PAC)-CreERT2 did not suppress the AV shunt phenotype, because we found that 62.7 ± 27.5% of total AV connections were enlarged by P18 (Fig. 4F and SI Appendix, Table S2), which is no different from mice with excision of only one Rbpj allele [60.1 ± 36.4% of total AV connections enlarged (SI Appendix, Table S2)]. Taken together, these data suggest that the onset of AV shunts is mediated through Rbpj in the ECs of microvasculature and veins but not BMX(PAC)-CreERT2 positive arteries. AV Shunts with Higher Velocity Increased in Velocity and Diameter, Whereas Adjacent Vessels with Lower Velocity Decreased in Velocity and Diameter. Despite Notch4* expression throughout the vascular endothelium, not all AV connections developed into AV shunts, and not all AV shunts continued to enlarge. We in the brain vasculature (SI Appendix, Fig. S11). Examination of mice with combined expression of the arterial marker ephrin-b2- H2b-eGFP and BMX(PAC)-CreERT2;mT/mG revealed that BMX (PAC)-CreERT2 excision overlapped with ephrin-b2-h2b-egfp in arteries but not in microvessels (SI Appendix, Fig. S14 and Movie S3). Therefore, BMX(PAC)-CreERT2 was uniformly active in arteries but not small arterioles. As such, the Cdh5(PAC)- CreERT2; ROSA:LNL:tTA driver induces Notch4* expression throughout the vascular endothelium, similar to the Tie2-tTA system, whereas the BMX(PAC)-CreERT2; ROSA:LNL:tTA driver induces Notch4* specifically in the endothelium of arteries. To evaluate the formation of AV shunts, we examined tomatolectin perfused whole-mount preparations of Cdh5(PAC)- CreERT2; ROSA:LNL:tTA;TRE-Notch4* and BMX-CreERT2; ROSA:LNL:tTA;TRE-Notch4* at P18, when most Tie2-Notch4* mice have developed AV shunts (17). As expected, we readily detected enlarged AV connections in Cdh5(PAC)-CreERT2; ROSA: LNL:tTA ;TRE-Notch4* mice by P18 after TAM injection at P7 P8 to induce Cre-mediated tta expression (Fig. 4A and SI Appendix, TableS1). In these mice, 82.8 ± 21.8% of total AV Fig. 4. Changes in Notch signaling in the artery alone did not affect AV shunt formation. (A F) Lectin-perfused vasculature on P18 surface cerebral cortex. (A and B) ROSA:LNL:tTA;Notch4* mice in which Notch4* was expressed at P7 with temporally inducible Cre-recombinase. (D F) Notch4* mutant or control mice with Cre-mediated RBPJ deletion at P7. (A) n = 83 AV connections in three mice. (B) n = 43 AV connections in three mice. (C) n = 103 AV connections in three mice. (D) n = 191 AV connections in five mice. (E) n = 186 AV connections in six mice. (F) n = 171 AV connections in seven mice. Blue arrowheads indicate AV connections. (Scale bars, 100 μm.) Murphy et al.
.")
 overtimeintie2-notch4*miceand compared the changes in velocity and diameter to control arteries (V1 and V2 in Fig. 5 C and D).")
5 investigated whether differences in blood flow velocity determine the growth of AV shunts. We identified arterial branch points in mutant mice in which one branch clearly led into an AV shunt (V1 in Fig. 5 A and B). We measured velocity and diameter in the higher-velocity AV shunt and the lower-velocity adjacent artery (V2 in Fig. 5 A and B) overtimeintie2-notch4*miceand compared the changes in velocity and diameter to control arteries (V1 and V2 in Fig. 5 C and D). The velocities in control arteries were more stable, varying by an average of 16% between time points vs. 45% in the mutants. In five of seven cases where the velocity in the AV shunt increased, the velocity in the adjacent artery with lower velocity decreased [compare Fig. 5 A to B; see summary in Fig. 5E when AV shunt velocity increases (see example in Movie S4)]. In three cases, the velocity was reducedinboththeavshuntanditsadjacentartery(fig.5e, when AV shunt velocity decreases ). The velocity in control arteries did not change (Fig. 5E, control ). Thus, increased velocity in AV shunts is often accompanied by decreased velocity in the adjacent arteries of Tie2-Notch4* mice. We then investigated whether changes in velocity correlated with changes in diameter in Tie2-Notch4* mutants. We focused on the AV shunts in which velocity increased and adjacent arteries in which velocity decreased (represented by red dots in Fig. 5E). We found that the adjacent downstream arteries indeed regressed, compared with either the AV shunt or arteries measured in control mice (Fig. 5F, P < 0.02 by two-tailed Student t test, V2 in Fig. 5 A and B, and SI Appendix, Fig. S15). Therefore, changes in velocity correlated with changes in diameter, so that the highest velocity connections, which also tended to be the most proximal AV connections in the Tie2-Notch4* mice, exhibited increasing velocity and diameter, whereas the lower velocity and typically distal arterial branches exhibited reduced velocity and diameter. For a broader perspective on AV shunt progression, we examined the whole brain by casting the vessels with radio-opaque microfil and imaging with microct. Despite widespread AV Fig. 5. AV shunts with higher velocity enlarged whereas adjacent arteries with lower velocity reduced in caliber. (A D) Two-photon time-lapse imaging of FITC-dextran labeled blood vessels on cerebral cortex through a cranial window. Average blood velocity, as determined by line-scan analysis of single RBCs is indicated for the AV shunt (V1) and the adjacent artery (V2) in a mutant (A and B) and control (C and D). (E) Change in velocity over h in distal arteries from Notch4* AV shunts when shunt velocity increased (n = 7) or decreased (n = 3), and in the smaller artery of similar branch points in controls (n = 5). (F) Change in diameter of AV shunts and adjacent arteries in the subgroup of Notch4* mice in which AV shunt velocity increased and distal velocity decreased (red points in E). (G and H) MicroCT of microfillabeled whole-brain vasculature in (G) Notch4* and (H) control mice. Arrow indicates focal aggregation of enlarged arteries and veins. *P < (Scale bars, 100 μm.) Fig. 6. Endothelial expression of Notch1-ICD induced AV shunts. Lectinperfused vasculature of P14 (A) Tie2-tTA; TRE-Notch1* mutant and (B) littermate control. (Scale bar, 100 μm.) shunting in mutant mice, AVM development appeared focal (Fig. 5G). Thus, our data suggest that the unchecked growth of higher-velocity AV connections results in focal AVM development. Endothelial Expression of Activated Notch1 Induced AV Shunt Formation. To determine whether expression of constitutively active Notch1 in the endothelium also causes AV shunt formation, we combined a constitutively active, tta-responsive Notch1* allele (16) with Tie2-tTA and activated Notch1* at birth. Five of five Tie2- tta;tre-notch1* mice exhibited signs of ataxia or lethargy by P14, as well as large brain AVMs, whereas none of six controls were affected (Fig. 6). Thus, endothelial expression of constitutively active Notch1 induces brain AV shunts during neonatal development. Discussion Here, we provide novel mechanistic insights into AVM formation. Our data suggest that AV shunts arise from the enlargement of capillary-like vessels and that cell-autonomous Notch signaling in the endothelium of microvasculature and veins is sufficient and required for AVM formation. AV shunt initiation is accompanied by the enlargement of EC area but not increased proliferation. Blood flow mediates AV shunt progression by positive feedback of increasing flow and increasing diameter, leading to the selective growth of focal AVMs. Our data shed light on the origin of AVMs. The vascular structure of brain AVMs has led to speculations that they arise from failed regression of primitive AV connections during development (3, 4). Extrapolation of histological analysis of skin AVMs led others to propose that brain AVMs arise from the dilation of postcapillary venules (5). In contrast to the existing models, our results demonstrate that Notch4*-induced AV shunts grow from preexisting capillary-like connections with no detectable differences from normal capillaries in either their diameter or their velocity. This suggests a new, potentially capillary-originated model of AVM initiation. Endothelial but not hematopoietic Notch signaling is critical in the formation of AV shunts. Previous work has demonstrated a critical function for blood cells in separating the blood and lymphatic vascular systems (27). Our data, confirmed by two independent induction systems, demonstrate that increased Notch signaling specifically within the endothelium is sufficient to cause AV shunt formation without obvious involvement of the blood cells. Conversely, expression of Notch4* specifically in Scl-tTA positive blood lineages did not lead to AV shunt formation (SI Appendix, Fig.S16). Furthermore, no increase in CD45+ cells was observed near AV shunts (SI Appendix, Fig. S17). Together, our data suggest that expression of Notch4* in the blood lineages is insufficient to induce AV shunts and that endothelial Notch4* elicits AVMs in a cell-autonomous fashion. Initial enlargement of AV shunts correlates with area expansion of individual ECs, rather than increased proliferation. Consistent with this finding, we found that the converse is true: EC area is reduced during AV shunt regression upon turning off the MEDICAL SCIENCES Murphy et al. PNAS December 16, 2014 vol. 111 no
6 Notch4* transgene (16). Further investigation is required to determine whether an altered cell area is a direct or indirect effect of Notch4*. Whether further enlargement of AV shunts beyond the initial stage depends on cell proliferation is currently unknown. We speculate that the growth of the AV shunts eventually requires cell proliferation because AV shunts in symptomatic mice can be >10-fold larger than normal capillaries. Notch signaling in the microvasculature and veins is both sufficient and required for AVM formation. Arterial BMX(PAC)- CreERT2 mediated up- or down-regulation of Notch signaling could not induce or suppress AV shunt formation, respectively. In the same genetic systems, global endothelial Cdh5(PAC)- CreERT2 mediated up- or down-regulation could. These findings are consistent with recent work showing that Rbpj-mediated arterial Notch signaling is dispensable for postnatal artery formation in the retina (26). Together, these data show that changes in Notch signaling in the BMX(PAC)-CreERT2 active cells are not critical in AV shunt formation, implying that it induces AVM through effects on the microvasculature and veins. Blood flow is a key regulator of vessel caliber (28). Based on clinical studies, it has been proposed that increased flow through low-resistance AV shunts encourages their growth while stealing blood flow from adjacent higher-resistance vessels (29). However, previous studies have typically focused on a single time point with lower resolution than we show here. We provide experimental evidence that endothelial Notch4* permits a steal and perpetuates a positive feedback loop, leading to selective growth of higher-velocity connections at the expense of lowervelocity connections. Our data also demonstrate that, in controls, blood velocity in adjacent two-branch arteries is maintained at a comparable rate over time. In mutants, the blood velocity in adjacent two-branch arteries varies greatly and with increasing disparity. Our earlier work shows that turning off Notch4* expression normalizes highvelocity AV shunts by rapidly reducing the flow and diameter of the AV shunt while increasing flow and diameter to the distal artery (16). Together, these findings suggest that sustained endothelial expression of Notch4* compromises biological checks that prevent AV connections from enlarging and stealing flow from adjacent vessels. We believe that normal AV specification is critical in maintaining proper hemodynamics in the vasculature. Because Notch signaling is a critical determinant of AV specification, it is conceivable that either gain or loss of Notch activity could disrupt AV specification, compromising normal vascular control of hemodynamic forces and leading to AVMs. Indeed, both gain- and loss-of-function Notch mutants develop abnormal shunting in mouse embryos (13, 30). Our model is consistent with recent work hinting that reduced Notch signaling in HHT type 2 could contribute to AVMs (31). We propose that increased Notch signaling in the endothelium of nonarterial vessels disturbs biological control of blood flow by allowing the persistent growth of high-flow AV connections at the expense of lower flow AV connections (SI Appendix, Fig. S18). Materials and Methods See SI Appendix for detailed information on materials and methods. Cranial windows, in vivo imaging, lectin-perfusion, Notch4-ICD, SMA, and VE-cadherin immunostaining was as we described (16). BrdU immunostaining was as described (32). For peripheral blood isolation, blood was removed from right ventricles, and erythrocytes were lysed. Flow cytometry was performed following CD45 immunostaining and DAPI labeling. For MicroCT, images of Microfil-labeled vasculature were acquired using a μct 40 system (Scanco Medical AG). All mouse lines used have been published. This study was carried out in strict accordance with National Institute of Health regulations and the Institutional Animal Care and Use Committee at the University of California, San Francisco. ACKNOWLEDGMENTS. This work was supported by National Institutes of Health (NIH) Grants R01 NS and R56NS06742, Vascular Cures (formerly the Pacific Vascular Research Foundation), the Frank A. Campini Foundation, the Mildred V. Strouss Trust, American Heart Association (AHA) Grant-in-Aid 10GRNT and GRNT , NIH Grant R01 HL (to R.A.W.), AHA Grant Y and Tobacco-Related Disease Research Program (TRDRP) 18DT-0009 Predoctoral Fellowships (to P.A.M.), TRDRP 19DT-007 and NIH F30 1F30HL A1 Predoctoral Fellowships (to T.N.K.), and TRDRP 20FT-0069 and NIH F32 1F32HL A1 Postdoctoral Fellowships (to C.M.N.). 1. Friedlander RM (2007) Clinical practice. Arteriovenous malformations of the brain. N Engl J Med 356(26): Meyer-Heim AD, Boltshauser E (2003) Spontaneous intracranial haemorrhage in children: Aetiology, presentation and outcome. Brain Dev 25(6): Mullan S, Mojtahedi S, Johnson DL, Macdonald RL (1996) Embryological basis of some aspects of cerebral vascular fistulas and malformations. J Neurosurg 85(1): Lasjaunias P (1997) A revised concept of the congenital nature of cerebral arteriovenous malformations. Interventional Neuroradiol 3(4): Braverman IM, Keh A, Jacobson BS (1990) Ultrastructure and three-dimensional organization of the telangiectases of hereditary hemorrhagic telangiectasia. J Invest Dermatol 95(4): Park SO, et al. (2009) Real-time imaging of de novo arteriovenous malformation in a mouse model of hereditary hemorrhagic telangiectasia. J Clin Invest 119(11): Mahmoud M, et al. (2010) Pathogenesis of arteriovenous malformations in the absence of endoglin. Circ Res 106(8): Chen W, et al. (2014) De novo cerebrovascular malformation in the adult mouse after endothelial Alk1 deletion and angiogenic stimulation. Stroke 45(3): Uebelhoer M, Boon LM, Vikkula M (2012) Vascular anomalies: From genetics toward models for therapeutic trials. Cold Spring Harb Perspect Med 2(8):a Whitehead KJ, Smith MC, Li DY (2013) Arteriovenous malformations and other vascular malformation syndromes. Cold Spring Harb Perspect Med 3(2):a Kawasaki J, et al. (2014) RASA1 functions in EPHB4 signaling pathway to suppress endothelial mtorc1 activity. J Clin Invest 124(6): Lawson ND, et al. (2001) Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 128(19): Kim YH, et al. (2008) Artery and vein size is balanced by Notch and ephrin B2/EphB4 during angiogenesis. Development 135(22): Lindskog H, et al. (2014) Molecular identification of venous progenitors in the dorsal aorta reveals an aortic origin for the cardinal vein in mammals. Development 141(5): Carlson TR, et al. (2005) Endothelial expression of constitutively active Notch4 elicits reversible arteriovenous malformations in adult mice. Proc Natl Acad Sci USA 102(28): Murphy PA, et al. (2012) Notch4 normalization reduces blood vessel size in arteriovenous malformations. Sci Transl Med 4(117):117ra Murphy PA, et al. (2008) Endothelial Notch4 signaling induces hallmarks of brain arteriovenous malformations in mice. Proc Natl Acad Sci USA 105(31): Murphy PA, Lu G, Shiah S, Bollen AW, Wang RA (2009) Endothelial Notch signaling is upregulated in human brain arteriovenous malformations and a mouse model of the disease. Lab Invest 89(9): Uyttendaele H, Ho J, Rossant J, Kitajewski J (2001) Vascular patterning defects associated with expression of activated Notch4 in embryonic endothelium. Proc Natl Acad Sci USA 98(10): ZhuGe Q, et al. (2009) Notch-1 signalling is activated in brain arteriovenous malformations in humans. Brain 132(Pt 12): Miniati D, et al. (2010) Constitutively active endothelial Notch4 causes lung arteriovenous shunts in mice. Am J Physiol Lung Cell Mol Physiol 298(2):L169 L Kitajewski J (2012) Arteriovenous malformations in five dimensions. Sci Transl Med 4(117):117fs Kim TN, et al. (2012) Line-scanning particle image velocimetry: An optical approach for quantifying a wide range of blood flow speeds in live animals. PLoS ONE 7(6): e Snippert HJ, et al. (2010) Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143(1): Wang L, et al. (2008) Restricted expression of mutant SOD1 in spinal motor neurons and interneurons induces motor neuron pathology. Neurobiol Dis 29(3): Ehling M, Adams S, Benedito R, Adams RH (2013) Notch controls retinal blood vessel maturation and quiescence. Development 140(14): Bertozzi CC, Hess PR, Kahn ML (2010) Platelets: Covert regulators of lymphatic development. Arterioscler Thromb Vasc Biol 30(12): Thoma R (1893) Untersuchungen über die Histogenese und Histomechanik des Gefässsytems (Ferdinand Enke, Stuttgart, Germany). German. 29. Fleischer LH, et al. (1993) Relationship of transcranial Doppler flow velocities and arteriovenous malformation feeding artery pressures. Stroke 24(12): Krebs LT, Starling C, Chervonsky AV, Gridley T (2010) Notch1 activation in mice causes arteriovenous malformations phenocopied by ephrinb2 and EphB4 mutants. Genesis 48(3): Larrivée B, et al. (2012) ALK1 signaling inhibits angiogenesis by cooperating with the Notch pathway. Dev Cell 22(3): Pitulescu ME, Schmidt I, Benedito R, Adams RH (2010) Inducible gene targeting in the neonatal vasculature and analysis of retinal angiogenesis in mice. Nat Protoc 5(9): Murphy et al.
7 Extended Materials and Methods: In vivo Imaging. Cranial windows for long-term imaging were as we described (1). Images and line-scan data were obtained using a locally constructed two-photon laser scanning microscope through a long working distance 1.0 NA water-immersion objective (Zeiss) as we described (2). Low-energy femtosecond laser pulses were centered at 870nm to simultaneously excite Texas Red-dextran (2000kDa) and GFP, or at 800nm to excite fluorescein-dextran (2000kDa, Sigma). Mice. This study was carried out in strict accordance with NIH regulations and the Institutional Animal Care and Use Committee at the University of California San Francisco. Tie2-tTA, TRE- Notch4* mice were generated as we described (3, 4). Tetracylcine (Tet) sucrose solution (0.5- mg/ml Tet, 50-mg/ml sucrose, Sigma) was administered to pregnant mothers from plugging, and withdrawn from pups at birth. Notch4* expression was regulated in TRE-Notch4* mutants as we described (5). Rbpj flox/flox (6), ROSA:LNL:tTA (7); TRE-H2b-eGFP (8), R26R-mT/mG (9), and R26R-confetti (10) mice have been published. Cdh5-CreERT 2 (11) and BMX-CreERT 2 (12) mice were kindly provided by the Adams lab. Tamoxifen (20mg/mL in corn oil) was given intragastrically (0.5mg at P7-P8) for induction of TRE-Notch4* by Cdh5(PAC)-CreERT 2 ; ROSA:LNL:tTA or BMX(PAC)-CreERT 2 ; ROSA:LNL:tTA or deletion of Rbpj by Cdh5(PAC)-CreERT 2 or BMX(PAC)- CreERT 2. For lineage tracing, Tamoxifen was given intragastrically (3x 0.1mg P1-P5) to Cdh5(PAC)-CreERT 2 ; R26R-confetti mice. Whole mount staining and fluorescence quantification. Immunostaining was performed as we described (1). Fluorescence quantification was performed using ImageJ and MATLAB (MathWorks). Notch4-ICD fluorescence staining intensity in mutants was normalized to the average staining intensity in littermate controls, and nuclear TRE-H2b-GFP intensities were normalized to background levels. Areas of VE-cadherin outlined ECs were quantified by tracing the outline of clearly visible individual ECs. For quantification of AV shunts in fixed samples, P18 mouse brains were sliced and flat-mounted to image surface vessels of the same region of the cortical surface. Images of tomato lectin-perfused vasculature were acquired (3-7 random fields of view per sample). AV connections within individual fields of view were identified and scored as enlarged if the minimum diameter was 12.5µm. The ratio of enlarged/total AV connections was computed for each animal, and the mean ratio (n = # of mice) was compared between groups.
8 BrdU incorporation and immunostaining. BrdU (5-bromo-2 -deoxyuridine)(100 mg/kg body weight in 0.9% saline)(fisher) was injected IP into pups on P10, P11, P12. Four hours postinjection on P12, 50ug biotinylated-lycopersicon esculentum (tomato) lectin (Vector Labs)/45 ug Alexa647-streptavidin (Jackson ImmunoResearch)/PBS was perfused via IVC. Whole mount immunostaining was followed according to Pitulescu et al. (2010), using anti-brdu (1:50)(BD Biosciences) and Cy3-conjugated secondary (Jackson ImmunoResearch). Tissue was incubated in 2ug/mL Hoechst (Molecular Probes) 30 min prior to imaging. Z stacks (1um steps) were captured with Yokogawa Spinning Disk confocal microscopy and ImageJ software at the UCSF Biological Imaging Development Center. Isolation of peripheral blood and flow cytometry. Blood ( uL) was removed from right ventricle of anesthetized mouse using 20G needle/syringe, dispensed into 5mM EDTA, and erythrocytes were lysed (155mM NH 4 Cl, 12mM NaHCO 3, 0.1mM EDTA, ph 7.5). Cells were pelleted, washed in PBS, fixed with 2% PFA, immunostained against CD45-Alexa647 (1:200)(BioLegend), and incubated in 2ug/mL DAPI (Sigma). The UCSF Liver Center Core performed flow cytometry using BD Biosciences LSRII cytometer. MicroCT. Mice were exsanguinated by cannulating the left ventricle and perfusing saline from the heart through the vasculature using an IV drip bag and butterfly needle. An incision was made in the right atrium allowing an outflow track for blood. The descending aorta was ligated with 6.0 suture after exsanguination to maximize brain perfusion. Without removing the butterfly needle from the left ventricle, a syringe containing a mixture of yellow Microfil MV122-yellow (1:10 catalyst to MV compound, Flow Tech Inc) was used to gently perfuse the vasculature until internal organs had a yellow appearance. Brains were harvested after 90 minutes allowing the Microfil to cure. Specimens were stored in 10% formalin until imaging. Brain specimens were scanned with a micro-computed tomography system (μct 40, Scanco Medical AG, Basserdorf, Switzerland) with a 12-μm isotropic nominal resolution. Images were acquired by scanning in the transaxial plane over a ~1.5cm section extending from the rostral to caudal edges of each brain. Data visualization was conducted at the microct workstation. Segmentation was performed by applying a predetermined threshold radio-attentuation value on all scans to eliminate signal voxels generated outside of the brain.
9 Statistics. Comparisons were made using a two-tailed Student s T-Test. Comparison of multiple groups was made using ANOVA with post-hoc Tukey s HSD analysis. References: 1. Murphy PA, et al. (2012) Notch4 normalization reduces blood vessel size in arteriovenous malformations. Science translational medicine 4(117):117ra Kim TN, et al. (2012) Line- scanning particle image velocimetry: an optical approach for quantifying a wide range of blood flow speeds in live animals. PLoS One 7(6):e Carlson TR, et al. (2005) Endothelial expression of constitutively active Notch4 elicits reversible arteriovenous malformations in adult mice. Proceedings of the National Academy of Sciences of the United States of America 102(28): Kim YH, et al. (2008) Artery and vein size is balanced by Notch and ephrin B2/EphB4 during angiogenesis. Development 135(22): Murphy PA, et al. (2008) Endothelial Notch4 signaling induces hallmarks of brain arteriovenous malformations in mice. Proceedings of the National Academy of Sciences of the United States of America 105(31): Tanigaki K, et al. (2002) Notch- RBP- J signaling is involved in cell fate determination of marginal zone B cells. Nat Immunol 3(5): Wang L, et al. (2008) Restricted expression of mutant SOD1 in spinal motor neurons and interneurons induces motor neuron pathology. Neurobiology of disease 29(3): Tumbar T, et al. (2004) Defining the epithelial stem cell niche in skin. Science 303(5656): Muzumdar MD, Tasic B, Miyamichi K, Li L, & Luo L (2007) A global double- fluorescent Cre reporter mouse. Genesis 45(9): Snippert HJ, et al. (2010) Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143(1): Pitulescu ME, Schmidt I, Benedito R, & Adams RH (2010) Inducible gene targeting in the neonatal vasculature and analysis of retinal angiogenesis in mice. Nat Protoc 5(9): Ehling M, Adams S, Benedito R, & Adams RH (2013) Notch controls retinal blood vessel maturation and quiescence. Development 140(14):
10 Supplemental Figure Legends Supplemental Figure 1. AV diameter increases over time in the cerebral vasculature of Notch4* expressing mutants. A) Mean AV diameter measured at the narrowest point of each of the indicated timepoints in Tie2-tTA (control) and Tie2-tTA; TRE-Notch4* (mutant) mice. Red dots indicate the diameter of all AV connections 12.5µm. B) The percentage of AV connections 12.5µm diameter at each timepoint. Supplemental Figure 2. Relationship between lumen diameter and blood velocity in capillary-like vessels studied over time. A) Plots of capillary-like vessels with paired increase of both lumen diameter (solid line) and blood velocity (dotted line) during the imaging period. B) Plots of capillary-like vessels with increase in diameter but minimal or no increase in blood velocity over the imaging period. C) Plots of capillary-like vessels with minimal or no increase in either diameter or blood velocity over the imaging period. D) Plots of capillaries in control mice demonstrating variation of lumen diameter and blood velocity over imaging period. Velocity and diameter analysis in all vessels that remained through the imaging interval (N=18 AV connection in 4 Notch4* mutant and N=18 AV connections in 4 control mice). Additional examples from these mice are shown in Figure 1. Supplemental Figure 3. Control mice did not develop AV shunts. Two-photon time-lapse imaging of FITC-dextran labeled AV connections on cerebral cortex through a cranial window. An AV connection (red arrowheads) in a control mouse remains capillary diameter between P14 to P19. Scale bar = 50µm. Supplemental Figure 4. No correlation between the initial diameter of the AV connection (smallest lumen diameter) and the final AV shunt diameter (largest lumen diameter). Supplemental Figure 5. Vascular smooth muscle cell coverage is grossly normal in Notch4* expressing mice at P12. Whole mount immunofluorescent staining of cortex from Notch4* mutant and control mice. Anti-α-smooth muscle actin in red and lectin perfusion in green. Scale bar = 200µm. Supplemental Figure 6. Strong expression of Notch4* transgene throughout endothelium of the microvasculature. Fluorescence immunostaining for Notch4 intracellular domain in cerebral cortex whole mount shows increased levels Notch4-ICD in the Notch4* expressing mice than in their littermate controls. Note that expression is elevated not only in the AV shunt (arrowhead), but also in the artery, vein and capillaries not obviously enlarged (arrows, magnified view in lower panels). Scale bars = 100um in upper panels and 25um in lower panels. Supplemental Figure 7. TRE-eGFP is a reporter of Notch4* expression. A) Notch4-ICD staining in sagittal brain sections of Tie2-tTA; TRE-Notch4*; TRE-H2B-GFP mice at P10 (a1), P12 (a2), and P18 (a3). (a4) Notch4-ICD staining in littermate control. (a5) Negative control of Notch4 staining using non-specific IgG on adjacent section in mutant specimen. B) Notch4-ICD staining intensity in lectin-perfused vessel segments of Notch4* mice, normalized to age-matched genetic controls. C) Correlation between Notch4 staining intensity and GFP intensity in lectin-perfused vessel segments of Notch4* mice over time. Supplemental Figure 8. In vivo assessment of the correlation between TRE-eGFP expression and diameter.
11 Time history of number of GFP+ cells and vessel segment diameter of Notch4* mice. Change in vessel segment diameter was scored as increase, constant, decrease, or regress. Each line trajectory corresponds to an individual vessel segment. Color transitions of line trajectory indicate time progression. Circle indicates vessel segment was part of an AV shunt. Cross indicates vessel segment regressed. Number of GFP+ cells was normalized to the initial vessel segment diameter and expressed as per unit length. Bold line trajectory corresponds to mean number of GFP+ cells and mean vessel segment diameter. Supplemental Figure 9. No significant difference in the number of Confetti labeled ECs in AV connections as they grow into AV shunts. Correlation plot showing the relationship between the change in diameter and the change in cell number over the 48hr interval. Triangle indicates mean change in diameter and cell number. Marker size represents number of observations. Supplemental Figure 10. No significant difference in BrdU labeled endothelium in Notch4* mutant mice. A) Quantification of BrdU labeled cells as a percentage of all DAPI+ lectin+ ECs in the indicated vessels (- Shunt indicates that the artery or vein is not directly connected to an AV shunt) and B) in the AV connections or AV shunts of the indicated mice. C) Quantification of minimal AV diameter in the measured connections. Error bars reflect standard deviation among individual animals (N=4 for AV shunt group and N=5 for all other groups). Supplemental Figure 11. Cdh5(PAC)-CreERT2 and BMX(PAC)-CreERT2 mediate endothelial- and arterial endothelial-specific gene excision, respectively. (A) Cdh5(PAC)-CreERT2 and (B) BMX(PAC)- CreERT2 activity was visualized in vivo using a membrane-targeted egfp Cre-reporter. Plasma in perfused vessels is labeled by Texas-red dextran. Higher magnification image shows differential labeling of the artery, capillary and vein (A & B ). Arrow indicates capillaries. Scale bars = (B) 150μm and (B ) 50μm. Supplemental Figure 12. Cdh5(PAC)-CreERT2 is active in vascular endothelium and AV shunts but not blood cells. (A) FACs showing the level of Cre-activated mgfp in Cdh5(PAC)-CreERT2 mice (with and without ROSA:LNL:tTA; TRE-Notch4*) and positive and negative controls. (B&C) Whole mount imaging showing strong mgfp expression throughout the vasculature of Notch4* expressing mice (B) and the control (C). Supplemental Figure 13. Cdh5(PAC)-CreERT 2 ; ROSA:LNL:tTA is a strong driver of TRE-Notch4* expression. Quantification of Notch4 immunofluorescence staining intensity in 10 fields per brain, normalized against littermate controls. (N=4 Cdh5(PAC)-CreERT 2 ; ROSA:LNL:tTA; TRE-Notch4* mice and N=6 Tie2-tTA; TRE-Notch4* mice.) Supplemental Figure 14. Variable BMX(PAC)-CreERT2-mediated excision in the distal ends of the arterioles. Two-photon imaging of the cerebellum of BMX(PAC)-CreERT2; mt/mg; ephrinb2-h2b-egfp mice with counter stain by tomato lectin perfusion. Note extension of the nuclear ephrinb2-h2b-egfp signal beyond the end of BMX(PAC)-CreERT2; mt/mg+ (cytoplasmic) arteries. Scale bar = 50μm. Supplemental Figure 15. Growth of an AV shunt and regression of a lower resistance distal vessel. 3D rendering of in vivo time-lapse data of an AV shunt and distal artery (arrowhead). Supplemental Figure 16. Scl-tTA; TRE-Notch4* does not result in AVM.
12 Vasculature of (A) Scl-tTA; TRE-Notch4* and (B) littermate Tie2-tTA; TRE-Notch4* mutants, revealed by fluorescent tomato lectin perfusion at P28 after tetracycline removal at birth. (A, B ) Magnified view of boxed regions in (A, B). Scale bar = 100μm (A, B) and 50μm (A, B ). Supplemental Figure 17. No increase in CD45+ hematopoetic cells in the AV shunts of Notch4* expressing mice. Fluorescence immunostaining for CD45 in cerebral cortex whole mount shows no detectable increase in hematopoetic cells in the AV shunts of Notch4* expressing mouse, and very few hematopoetic cells in general. Scale bars = 100µm. Supplemental Figure 18. Model for the flow-mediated selection of AV shunts Schematic depicting a normal cortical brain vascular network (A) and the remodeling of the vasculature induced by Notch4* expression (B-D). In the earliest stage, AV shunts throughout the brain are enlarged (arrowheads in B). Following initial enlargement, a few AV shunts in which flow is highest continue to enlarge (arrowheads in C). After sequential flow-mediated selections steps, AVM-like lesions develop, composed of dilated and tortuous arterial and venous vessels which shunt large amounts of blood from distal capillaries (arrowhead in D). Supplemental Table 1. Quantification of enlarged AV connections in mice with endothelial- or arterial endothelial-specific Notch4* expression. Cdh5(PAC)-CreERT2 and BMX(PAC)-CreERT2 were used to turn on Notch4* in all vessels and in arteries, respectively. Lectin-perfused AV connections on the surface of the cerebral cortex of Notch4* mice (see Fig. 4) were scored as enlarged if the minimum diameter was 12.5μm. The ratio of enlarged/total AV connections was computed for each animal (see Supplemental Table 1 raw data), and the mean ratio (n = # of mice) was compared between groups. See Supplemental Methods for detailed methodology. (Data are mean±sd) Supplemental Table 2. Quantification of Notch4*-mediated enlarged AV connections after endothelialor arterial endothelial-specific deletion of Rbpj. Cdh5(PAC)-CreERT2 and BMX(PAC)-CreERT2 were used to delete floxed-rbpj in all vessels and in arteries, respectively. Either both or a single Rbpj allele was excised. Lectin-perfused AV connections on the surface of the cerebral cortex of Notch4* and control mice (see Fig. 4) were scored as enlarged if the minimum diameter was 12.5μm. The ratio of enlarged/total AV connections was computed for each animal (see Supplemental Table 2 raw data), and the mean ratio (n = # of mice) was compared between groups. See Supplemental Methods for detailed methodology. (Data are mean±sd).
13 Supplemental Figure 1.
14 Supplemental Figure 2A-C.
15 Supplemental Figure 2D.

16 Supplemental Figure 3.
17 Supplemental Figure 4.

18 Supplemental Figure 5.

19 Supplemental Figure 6.

20 Supplemental Figure 7.
21 Supplemental Figure 8.
22 Supplemental Figure 9
23 Supplemental Figure 10

24 Supplemental Figure 11.
25 Supplemental Figure 12
26 Supplemental Figure 13

27 Supplemental Figure 14
28 Supplemental Figure 15.
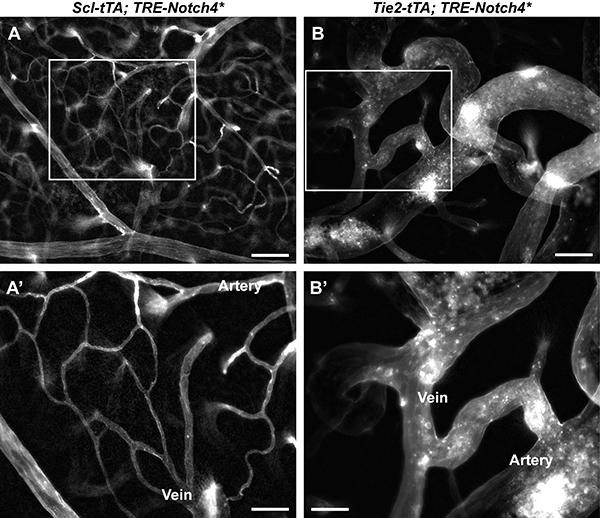
29 Supplemental Figure 16

30 Supplemental Figure 17

31 Supplemental Figure 18
32 Supplemental Table 1. Genotype n (Mice) Enlarged/Total (%) Mean SD Cdh5(PAC)-CreERT2; ROSA:LNL:tTA; TRE-Notch4* * ROSA:LNL:tTA; TRE-Notch4* (*, difference between groups, p = 0.02, t- test) Genotype n (Mice) Enlarged/Total (%) Mean SD BMX(PAC)-CreERT2; ROSA:LNL:tTA; TRE-Notch4* ROSA:LNL:tTA; TRE-Notch4* (Not significant, p = 0.42, t- test)
33 Supplemental Table 1 raw data Cdh5(PAC)-CreERT2; ROSA:LNL:tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) ROSA:LNL:tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) BMX(PAC)-CreERT2; ROSA:LNL:tTA; TRE-Notch4 Mouse Sum Total connections Enlarged connections Enlarged/Total (%) ROSA:LNL:tTA; TRE-Notch4 Mouse 1 2 Sum Total connections Enlarged connections Enlarged/Total (%) 0 0
34 Supplemental Table 2. Genotype n (Mice) Enlarged/Total (%) Mean SD Cdh5(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA; TRE-Notch4* Cdh5(PAC)-CreERT2; RBPJ +/fl ; Tie2-tTA; TRE-Notch4* * Cdh5(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA (*, difference vs. other groups, p = 0.004, ANOVA and post hoc multiple comparison test) Genotype n (Mice) Enlarged/Total (%) Mean SD BMX(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA; TRE-Notch4* BMX(PAC)-CreERT2; RBPJ +/fl ; Tie2-tTA; TRE-Notch4* BMX(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA * (*, difference vs. other groups, p = 0.001, ANOVA and post hoc multiple comparison test)
35 Supplemental Table 2 raw data Cdh5(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) Cdh5(PAC)-CreERT2; RBPJ +/fl ; Tie2-tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) Cdh5(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA Mouse Sum Total connections Enlarged connections Enlarged/Total (%) BMX(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) BMX(PAC)-CreERT2; RBPJ +/fl ; Tie2-tTA; TRE-Notch4* Mouse Sum Total connections Enlarged connections Enlarged/Total (%) BMX(PAC)-CreERT2; RBPJ fl/fl ; Tie2-tTA Mouse Sum Total connections Enlarged connections Enlarged/Total (%)

36 Supporting Information Murphy et al /pnas Movie S1. Vascular network in Notch4* mutant before AV shunt formation. Three-dimensional rendering of a two-photon imaging stack from the P14 Notch4* mutant shown in Fig. 1A. Movie S1 Murphy et al. 1of3

-CreERT 2 ; mtmg; ephrin-b2-h2b-egfp mutant mouse following perfusion with Alexa-647 labeled tomato lectin.")
37 Movie S2. Vascular network in Notch4* mutant after AV shunt formation. Three-dimensional rendering of an imaging stack from P19 Notch4* mutant showninfig.1aand Movie S1, 5 d later. An AV shunt can be observed in the center of the field, where only capillary connections had been seen previously. Movie S2 Movie S3. Overlapping expression of ephrinb2-h2b-egfp and BMX(PAC)-CreERT2 induced mt/mg. Optical z-stack from two-photon imaging of the cortical surface of the brain of a BMX(PAC)-CreERT 2 ; mtmg; ephrin-b2-h2b-egfp mutant mouse following perfusion with Alexa-647 labeled tomato lectin. Green channel: ephrin-b2-h2b-egfp and BMX(PAC)-CreERT2 activated mt/mg reporter; red channel: lectin perfusion. Note that the BMX(PAC)-CreERT2 reporter ends in the large arterioles before ephrin-b2-h2b-egfp capillaries. Movie S3 Murphy et al. 2of3

38 Movie S4. Dramatic reduction in distal blood flow with proximal steal effect. Single-plane movie from a two-photon imaging stack taken at a late time point in AV shunt progression in a Tie2-tTA; TRE-Notch4* mutant mouse. Movement of dark-red blood cells can be observed by contrast Texas-Red dextran labeled plasma. Slowly moving red blood cells can be observed as distinct spheres, and quickly moving blood cells appear as streaks. Blood flow and vessel diameter was reduced in this distal branch over time as the proximal connection grew. Movie S4 Other Supporting Information Files SI Appendix (PDF) Murphy et al. 3of3
Supplementary Materials for
 www.sciencetranslationalmedicine.org/cgi/content/full/4/117/117ra8/dc1 Supplementary Materials for Notch4 Normalization Reduces Blood Vessel Size in Arteriovenous Malformations Patrick A. Murphy, Tyson
www.sciencetranslationalmedicine.org/cgi/content/full/4/117/117ra8/dc1 Supplementary Materials for Notch4 Normalization Reduces Blood Vessel Size in Arteriovenous Malformations Patrick A. Murphy, Tyson
On blood vessels. Identification and Validation of Therapeutic Targets for Vascular Disease
 Identification and Valition of Therapeutic Targets for Vascular Disease UCSF LAVR Rong Wang, Ph. D. Director Laboratory for Accelerated Vascular Research Associate Professor Department of Surgery University
Identification and Valition of Therapeutic Targets for Vascular Disease UCSF LAVR Rong Wang, Ph. D. Director Laboratory for Accelerated Vascular Research Associate Professor Department of Surgery University
Supplementary Figure 1
 Supplementary Figure 1 The average sigmoid parametric curves of capillary dilation time courses and average time to 50% peak capillary diameter dilation computed from individual capillary responses averaged
Supplementary Figure 1 The average sigmoid parametric curves of capillary dilation time courses and average time to 50% peak capillary diameter dilation computed from individual capillary responses averaged
Supplementary Figure 1. EC-specific Deletion of Snail1 Does Not Affect EC Apoptosis. (a,b) Cryo-sections of WT (a) and Snail1 LOF (b) embryos at
 Cryo-sections of WT (a) and Snail1 LOF (b) embryos at") Supplementary Figure 1. EC-specific Deletion of Snail1 Does Not Affect EC Apoptosis. (a,b) Cryo-sections of WT (a) and Snail1 LOF (b) embryos at E10.5 were double-stained for TUNEL (red) and PECAM-1 (green).
Supplementary Figure 1. EC-specific Deletion of Snail1 Does Not Affect EC Apoptosis. (a,b) Cryo-sections of WT (a) and Snail1 LOF (b) embryos at E10.5 were double-stained for TUNEL (red) and PECAM-1 (green).
(a-r) Whole mount X-gal staining on a developmental time-course of hearts from
 Whole mount X-gal staining on a developmental time-course of hearts from") 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Supplementary Figure 1 (a-r) Whole mount X-gal staining on a developmental time-course of hearts from Sema3d +/- ;Ephb4 LacZ/+ and Sema3d -/- ;Ephb4 LacZ/+ embryos.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Supplementary Figure 1 (a-r) Whole mount X-gal staining on a developmental time-course of hearts from Sema3d +/- ;Ephb4 LacZ/+ and Sema3d -/- ;Ephb4 LacZ/+ embryos.
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Relative expression of K IR2.1 transcript to enos was reduced 29-fold in capillaries from knockout animals. Relative expression of K IR2.1 transcript to enos was reduced 29-fold
Supplementary Figure 1 Relative expression of K IR2.1 transcript to enos was reduced 29-fold in capillaries from knockout animals. Relative expression of K IR2.1 transcript to enos was reduced 29-fold
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the
 Schematic of Crb3 isoforms, ERLI and CLPI, indicating the") Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
Supplementary Figure 1 Expression of Crb3 in mouse sciatic nerve: biochemical analysis (a) Schematic of Crb3 isoforms, ERLI and CLPI, indicating the location of the transmembrane (TM), FRM binding (FB)
SUPPLEMENTARY INFORMATION
 b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
b 350 300 250 200 150 100 50 0 E0 E10 E50 E0 E10 E50 E0 E10 E50 E0 E10 E50 Number of organoids per well 350 300 250 200 150 100 50 0 R0 R50 R100 R500 1st 2nd 3rd Noggin 100 ng/ml Noggin 10 ng/ml Noggin
Supplementary Figure 1. Nature Neuroscience: doi: /nn.4547
 Supplementary Figure 1 Characterization of the Microfetti mouse model. (a) Gating strategy for 8-color flow analysis of peripheral Ly-6C + monocytes from Microfetti mice 5-7 days after TAM treatment. Living
Supplementary Figure 1 Characterization of the Microfetti mouse model. (a) Gating strategy for 8-color flow analysis of peripheral Ly-6C + monocytes from Microfetti mice 5-7 days after TAM treatment. Living
Santulli G. et al. A microrna-based strategy to suppress restenosis while preserving endothelial function
 ONLINE DATA SUPPLEMENTS Santulli G. et al. A microrna-based strategy to suppress restenosis while preserving endothelial function Supplementary Figures Figure S1 Effect of Ad-p27-126TS on the expression
ONLINE DATA SUPPLEMENTS Santulli G. et al. A microrna-based strategy to suppress restenosis while preserving endothelial function Supplementary Figures Figure S1 Effect of Ad-p27-126TS on the expression
Signaling Vascular Morphogenesis and Maintenance
 Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan Science 277: 48-50, in Perspectives (1997) Blood vessels are constructed by two processes: vasculogenesis, whereby a primitive vascular
TITLE: RBPJ and EphrinB2 as Molecular Targets to Treat Brain Arteriovenous Malformation in Notch4-Induced Mouse Model
 AWARD NUMBER: W81XWH-16-1-0665 TITLE: RBPJ and EphrinB2 as Molecular Targets to Treat Brain Arteriovenous Malformation in Notch4-Induced Mouse Model PRINCIPAL INVESTIGATOR: Rong Wang CONTRACTING ORGANIZATION:
AWARD NUMBER: W81XWH-16-1-0665 TITLE: RBPJ and EphrinB2 as Molecular Targets to Treat Brain Arteriovenous Malformation in Notch4-Induced Mouse Model PRINCIPAL INVESTIGATOR: Rong Wang CONTRACTING ORGANIZATION:
Supplemental Information. Tissue Myeloid Progenitors Differentiate. into Pericytes through TGF-b Signaling. in Developing Skin Vasculature
 Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
Bone Marrow Pop. (% Total) Mature Pool (Absolute %) Immature Pool (Absolute %) A10 EC Control A10 EC Control A10 EC Control
 Mature Pool (Absolute %) Immature Pool (Absolute %) A10 EC Control A10 EC Control A10 EC Control") Bone Marrow Pop. (% Total) Mature Pool (Asolute %) Immature Pool (Asolute %) A10 EC A10 EC A10 EC Myeloid 50.7 57.5 37.5 46.2 13.2 11.3 Erythroid 38.3 23.2 33.3 16.8 9.3 6.3 Lymphocytes 13.8 19.0 - - -
Bone Marrow Pop. (% Total) Mature Pool (Asolute %) Immature Pool (Asolute %) A10 EC A10 EC A10 EC Myeloid 50.7 57.5 37.5 46.2 13.2 11.3 Erythroid 38.3 23.2 33.3 16.8 9.3 6.3 Lymphocytes 13.8 19.0 - - -
Evaluation of directed and random motility in microslides Assessment of leukocyte adhesion in flow chambers
 Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Evaluation of directed and random motility in microslides Motility experiments in IBIDI microslides, image acquisition and processing were performed as described. PMN, which ended up in an angle < 180
Supplemental Information. Otic Mesenchyme Cells Regulate. Spiral Ganglion Axon Fasciculation. through a Pou3f4/EphA4 Signaling Pathway
 Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
mm Distance (mm)
") b a Magnet Illumination Coverslips MPs Objective 2575 µm 1875 µm 1575 µm 1075 µm 875 µm 545 µm 20µm 2 3 0.5 0.3mm 1 1000 100 10 1 0.1 1000 100 10 1 0.1 Field Induction (Gauss) 1.5 0 5 10 15 20 Distance
b a Magnet Illumination Coverslips MPs Objective 2575 µm 1875 µm 1575 µm 1075 µm 875 µm 545 µm 20µm 2 3 0.5 0.3mm 1 1000 100 10 1 0.1 1000 100 10 1 0.1 Field Induction (Gauss) 1.5 0 5 10 15 20 Distance
Supplemental Figure 1. Intracranial transduction of a modified ptomo lentiviral vector in the mouse
 Supplemental figure legends Supplemental Figure 1. Intracranial transduction of a modified ptomo lentiviral vector in the mouse hippocampus targets GFAP-positive but not NeuN-positive cells. (A) Stereotaxic
Supplemental figure legends Supplemental Figure 1. Intracranial transduction of a modified ptomo lentiviral vector in the mouse hippocampus targets GFAP-positive but not NeuN-positive cells. (A) Stereotaxic
John Nguyen, Nozomi Nishimura, Robert Fetcho, Costantino Iadecola, Chris B. Schaffer
 Supplemental figures and text for Occlusion of cortical ascending venules causes blood flow decreases, reversals in flow direction, and vessel dilation in upstream capillaries John Nguyen, Nozomi Nishimura,
Supplemental figures and text for Occlusion of cortical ascending venules causes blood flow decreases, reversals in flow direction, and vessel dilation in upstream capillaries John Nguyen, Nozomi Nishimura,
Angiogenesis in Human Development. Vascular Development
 Angiogenesis in Human Development Jan Kitajewski ICRC 217B, ph 851-4688, email: jkk9 BACKGROUND READING: Vascular Development Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan. Science 277:
Angiogenesis in Human Development Jan Kitajewski ICRC 217B, ph 851-4688, email: jkk9 BACKGROUND READING: Vascular Development Signaling Vascular Morphogenesis and Maintenance Douglas Hanahan. Science 277:
Supplementary information - Table (1), Figures (12), and Videos (5)
, Figures (12), and Videos (5)") Supplementary information - Table (1), Figures (12), and Videos (5) A soft, transparent, freely accessible cranial window for chronic imaging and electrophysiology Chaejeong Heo 1, Hyejin Park 1, 2, Yong-Tae
Supplementary information - Table (1), Figures (12), and Videos (5) A soft, transparent, freely accessible cranial window for chronic imaging and electrophysiology Chaejeong Heo 1, Hyejin Park 1, 2, Yong-Tae
Recruitment of pre-existing vessels. versus. Angiogenesis
 Pulmonary Arteriovenous Malformations After the Bidirectional Glenn and the Role of VEGF Background PAVMs first recognised during follow-up after classical Glenn shunt, in ipsilateral lung N.Sreeram. Heart
Pulmonary Arteriovenous Malformations After the Bidirectional Glenn and the Role of VEGF Background PAVMs first recognised during follow-up after classical Glenn shunt, in ipsilateral lung N.Sreeram. Heart
Supplementary Information
 Supplementary Information Title Degeneration and impaired regeneration of gray matter oligodendrocytes in amyotrophic lateral sclerosis Authors Shin H. Kang, Ying Li, Masahiro Fukaya, Ileana Lorenzini,
Supplementary Information Title Degeneration and impaired regeneration of gray matter oligodendrocytes in amyotrophic lateral sclerosis Authors Shin H. Kang, Ying Li, Masahiro Fukaya, Ileana Lorenzini,
Supporting Information
 Supporting Information Rock et al. 10.1073/pnas.1117988108 Fig. S1. Heterogeneity of stromal cells in normal and fibrotic mouse lungs. Sections of normal mouse lungs (A and D) and fibrotic lungs collected
Supporting Information Rock et al. 10.1073/pnas.1117988108 Fig. S1. Heterogeneity of stromal cells in normal and fibrotic mouse lungs. Sections of normal mouse lungs (A and D) and fibrotic lungs collected
Real-time imaging reveals the single steps of brain metastasis fo mation r
 Real-time imaging reveals the single steps of brain metastasis fo mation r Yvonne Kienast, Louisa von Baumgarten, Martin Fuhrmann, Wolfgang E.F. Klinkert, Roland Goldbrunner, Jochen Herms and Frank Winkler
Real-time imaging reveals the single steps of brain metastasis fo mation r Yvonne Kienast, Louisa von Baumgarten, Martin Fuhrmann, Wolfgang E.F. Klinkert, Roland Goldbrunner, Jochen Herms and Frank Winkler
Zhu et al, page 1. Supplementary Figures
 Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation
 SUPPLEMENTARY INFORMATION Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation Samantha Arokiasamy 1,2, Christian Zakian 1, Jessica Dilliway
SUPPLEMENTARY INFORMATION Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation Samantha Arokiasamy 1,2, Christian Zakian 1, Jessica Dilliway
Postn MCM Smad2 fl/fl Postn MCM Smad3 fl/fl Postn MCM Smad2/3 fl/fl. Postn MCM. Tgfbr1/2 fl/fl TAC
 A Smad2 fl/fl Smad3 fl/fl Smad2/3 fl/fl Tgfbr1/2 fl/fl 1. mm B Tcf21 MCM Tcf21 MCM Smad3 fl/fl Tcf21 MCM Smad2/3 fl/fl Tcf21 MCM Tgfbr1/2 fl/fl αmhc MCM C 1. mm 1. mm D Smad2 fl/fl Smad3 fl/fl Smad2/3
A Smad2 fl/fl Smad3 fl/fl Smad2/3 fl/fl Tgfbr1/2 fl/fl 1. mm B Tcf21 MCM Tcf21 MCM Smad3 fl/fl Tcf21 MCM Smad2/3 fl/fl Tcf21 MCM Tgfbr1/2 fl/fl αmhc MCM C 1. mm 1. mm D Smad2 fl/fl Smad3 fl/fl Smad2/3
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.
 Light microscopic pictures show mouse embryos at developmental stage E12.") Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Vascular Malformations of the Brain. William A. Cox, M.D. Forensic Pathologist/Neuropathologist. September 8, 2014
 Vascular Malformations of the Brain William A. Cox, M.D. Forensic Pathologist/Neuropathologist September 8, 2014 Vascular malformations of the brain are classified into four principal groups: arteriovenous
Vascular Malformations of the Brain William A. Cox, M.D. Forensic Pathologist/Neuropathologist September 8, 2014 Vascular malformations of the brain are classified into four principal groups: arteriovenous
Ahtiainen et al., http :// /cgi /content /full /jcb /DC1
 Supplemental material JCB Ahtiainen et al., http ://www.jcb.org /cgi /content /full /jcb.201512074 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Distinct distribution of different cell cycle phases in the
Supplemental material JCB Ahtiainen et al., http ://www.jcb.org /cgi /content /full /jcb.201512074 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Distinct distribution of different cell cycle phases in the
Macrophages form functional vascular mimicry channels in vivo. SI Figures and Legend
 Macrophages form functional vascular mimicry channels in vivo Authors: *Faith H. Barnett, *Mauricio Rosenfeld, Malcolm Wood, William Kiosses, Yoshihiko Usui, Valentina Marchetti, Edith Aguilar, and Martin
Macrophages form functional vascular mimicry channels in vivo Authors: *Faith H. Barnett, *Mauricio Rosenfeld, Malcolm Wood, William Kiosses, Yoshihiko Usui, Valentina Marchetti, Edith Aguilar, and Martin
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Formation of the AA5x. a, Camera lucida drawing of embryo at 48 hours post fertilization (hpf, modified from Kimmel et al. Dev Dyn. 1995 203:253-310). b, Confocal microangiogram
Supplementary Figure 1. Formation of the AA5x. a, Camera lucida drawing of embryo at 48 hours post fertilization (hpf, modified from Kimmel et al. Dev Dyn. 1995 203:253-310). b, Confocal microangiogram
Title: Smooth muscle cell-specific Tgfbr1 deficiency promotes aortic aneurysm formation by stimulating multiple signaling events
 Title: Smooth muscle cell-specific Tgfbr1 deficiency promotes aortic aneurysm formation by stimulating multiple signaling events Pu Yang 1, 3, radley M. Schmit 1, Chunhua Fu 1, Kenneth DeSart 1, S. Paul
Title: Smooth muscle cell-specific Tgfbr1 deficiency promotes aortic aneurysm formation by stimulating multiple signaling events Pu Yang 1, 3, radley M. Schmit 1, Chunhua Fu 1, Kenneth DeSart 1, S. Paul
a 0,8 Figure S1 8 h 12 h y = 0,036x + 0,2115 y = 0,0366x + 0,206 Labeling index Labeling index ctrl shrna Time (h) Time (h) ctrl shrna S G2 M G1
 Time (h) ctrl shrna S G2 M G1") (GFP+ BrdU+)/GFP+ Labeling index Labeling index Figure S a, b, y =,x +, y =,x +,,,,,,,, Time (h) - - Time (h) c d S G M G h M G S G M G S G h Time of BrdU injection after electroporation (h) M G S G M
(GFP+ BrdU+)/GFP+ Labeling index Labeling index Figure S a, b, y =,x +, y =,x +,,,,,,,, Time (h) - - Time (h) c d S G M G h M G S G M G S G h Time of BrdU injection after electroporation (h) M G S G M
Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex
 Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10188 Supplementary Figure 1. Embryonic epicardial genes are down-regulated from midgestation stages and barely detectable post-natally. Real time qrt-pcr revealed a significant down-regulation
doi:10.1038/nature10188 Supplementary Figure 1. Embryonic epicardial genes are down-regulated from midgestation stages and barely detectable post-natally. Real time qrt-pcr revealed a significant down-regulation
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTARY INFORMATION
 1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
What effects will proximal or distal disease have on a waveform?
 Spectral Doppler Interpretation Director of Ultrasound Education & Quality Assurance Baylor College of Medicine Division of Maternal-Fetal Medicine Maternal Fetal Center Imaging Manager Texas Children
Spectral Doppler Interpretation Director of Ultrasound Education & Quality Assurance Baylor College of Medicine Division of Maternal-Fetal Medicine Maternal Fetal Center Imaging Manager Texas Children
Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a,
 Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a, Transverse sections of E17.5 ovary and mesonephros from Gli1-LacZ reporter embryos (n=3) after LacZ staining (blue). The
Supplementary Figure 1: Expression of Gli1-lacZ in E17.5 ovary and mesonephros. a, Transverse sections of E17.5 ovary and mesonephros from Gli1-LacZ reporter embryos (n=3) after LacZ staining (blue). The
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3443 In the format provided by the authors and unedited. Supplementary Figure 1 TC and SC behaviour during ISV sprouting. (a) Predicted outcome of TC division and competitive Dll4-Notch-mediated
DOI: 10.1038/ncb3443 In the format provided by the authors and unedited. Supplementary Figure 1 TC and SC behaviour during ISV sprouting. (a) Predicted outcome of TC division and competitive Dll4-Notch-mediated
International Graduate Research Programme in Cardiovascular Science
 1 International Graduate Research Programme in Cardiovascular Science This work has been supported by the European Community s Sixth Framework Programme under grant agreement n LSHM-CT-2005-01883 EUGeneHeart.
1 International Graduate Research Programme in Cardiovascular Science This work has been supported by the European Community s Sixth Framework Programme under grant agreement n LSHM-CT-2005-01883 EUGeneHeart.
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Award Number: W81XWH TITLE: PRINCIPAL INVESTIGATOR: Michael Dellinger
 AD Award Number: W81XWH-10-1-0052 TITLE: PRINCIPAL INVESTIGATOR: Michael Dellinger CONTRACTING ORGANIZATION: UT Southwestern Medical Center Dallas, TX 75390 REPORT DATE: February 2013 TYPE OF REPORT: Annual
AD Award Number: W81XWH-10-1-0052 TITLE: PRINCIPAL INVESTIGATOR: Michael Dellinger CONTRACTING ORGANIZATION: UT Southwestern Medical Center Dallas, TX 75390 REPORT DATE: February 2013 TYPE OF REPORT: Annual
(A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a
 RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a") Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3-
 1 2 The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3- Vitelline vein from yolk sac 3 However!!!!! The left
1 2 The sinus venosus represent the venous end of the heart It receives 3 veins: 1- Common cardinal vein body wall 2- Umbilical vein from placenta 3- Vitelline vein from yolk sac 3 However!!!!! The left
Occlusive hyperemia: a theory for the hemodynamic complications following resection of intracerebral arteriovenous malformations
 J Neurosurg 78: 167-175, 1993 Occlusive hyperemia: a theory for the hemodynamic complications following resection of intracerebral arteriovenous malformations NAYEF R. F. AL-RODHAN, M.D., PH.D., THORALF
J Neurosurg 78: 167-175, 1993 Occlusive hyperemia: a theory for the hemodynamic complications following resection of intracerebral arteriovenous malformations NAYEF R. F. AL-RODHAN, M.D., PH.D., THORALF
Journal Club. 03/04/2012 Lama Nazzal
 Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
Journal Club 03/04/2012 Lama Nazzal NOTCH and the kidneys Is an evolutionarily conserved cell cell communication mechanism. Is a regulator of cell specification, differentiation, and tissue patterning.
Classification des Malformations vasculaires
 Classification des Malformations vasculaires Gilles Soulez, MD, MSc, FSIR Professeur Titulaire et Chairman Dpt Radiologie, Radio-Oncologie et Medecine Nucléaire Université de Montréal Basic principles
Classification des Malformations vasculaires Gilles Soulez, MD, MSc, FSIR Professeur Titulaire et Chairman Dpt Radiologie, Radio-Oncologie et Medecine Nucléaire Université de Montréal Basic principles
Title: Epigenetic mechanisms underlying maternal diabetes-associated risk of congenital heart disease
 1 Supplemental Materials 2 3 Title: Epigenetic mechanisms underlying maternal diabetes-associated risk of congenital heart disease 4 5 6 Authors: Madhumita Basu, 1 Jun-Yi Zhu, 2 Stephanie LaHaye 1,3, Uddalak
1 Supplemental Materials 2 3 Title: Epigenetic mechanisms underlying maternal diabetes-associated risk of congenital heart disease 4 5 6 Authors: Madhumita Basu, 1 Jun-Yi Zhu, 2 Stephanie LaHaye 1,3, Uddalak
Supplementary Figure S1 Enlarged coronary artery branches in Edn1-knockout mice. a-d, Coronary angiography by ink injection in wild-type (a, b) and
 and") Supplementary Figure S1 Enlarged coronary artery branches in Edn1-knockout mice. a-d, Coronary angiography by ink injection in wild-type (a, b) and Edn1-knockout (Edn1-KO) (c, d) hearts. The boxed areas
Supplementary Figure S1 Enlarged coronary artery branches in Edn1-knockout mice. a-d, Coronary angiography by ink injection in wild-type (a, b) and Edn1-knockout (Edn1-KO) (c, d) hearts. The boxed areas
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2610 Figure S1 FSMCs derived from MSLN CLN transgenic mice express smooth muscle-specific proteins. Beta-galactosidase is ubiquitously expressed within cultured FSMCs derived from MSLN
DOI: 10.1038/ncb2610 Figure S1 FSMCs derived from MSLN CLN transgenic mice express smooth muscle-specific proteins. Beta-galactosidase is ubiquitously expressed within cultured FSMCs derived from MSLN
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
F-actin VWF Vinculin. F-actin. Vinculin VWF
 a F-actin VWF Vinculin b F-actin VWF Vinculin Supplementary Fig. 1. WPBs in HUVECs are located along stress fibers and at focal adhesions. (a) Immunofluorescence images of f-actin (cyan), VWF (yellow),
a F-actin VWF Vinculin b F-actin VWF Vinculin Supplementary Fig. 1. WPBs in HUVECs are located along stress fibers and at focal adhesions. (a) Immunofluorescence images of f-actin (cyan), VWF (yellow),
Chapter 6. Villous Growth
 Core Curriculum in Perinatal Pathology Chapter 6 Villous Growth Overview of vasculogenesis and angiogenesis Vasculogenesis Extraembryonic Vasculogenesis Angiogenesis Branching angiogenesis Sprouting angiogenesis
Core Curriculum in Perinatal Pathology Chapter 6 Villous Growth Overview of vasculogenesis and angiogenesis Vasculogenesis Extraembryonic Vasculogenesis Angiogenesis Branching angiogenesis Sprouting angiogenesis
SUPPLEMENTAL DATA. Lumen area ( m 2 )
") Elastin Lumen area ( m 2 ) Media to lumen ratio (x1) H.E. Medium thickness ( m) Medium area ( m 2 ) SUPPLEMENTAL DATA A (Bmal1 flox/flox ) (SM-Bmal1 -/- ) B 1 8 8 6 6 4 4 2 2 1µm 5 8 4 6 3 2 4 1 2 Supplemental
Elastin Lumen area ( m 2 ) Media to lumen ratio (x1) H.E. Medium thickness ( m) Medium area ( m 2 ) SUPPLEMENTAL DATA A (Bmal1 flox/flox ) (SM-Bmal1 -/- ) B 1 8 8 6 6 4 4 2 2 1µm 5 8 4 6 3 2 4 1 2 Supplemental
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
SUPPLEMENTARY INFORMATION Supplementary Figure 1. Long-term protection studies. 45 minutes of ischemia was induced in wild type (S1pr2 +/+ ) and S1pr2 -/- by MCAO. A) 5 days later brains were harvested
Short title: Endothelial mtorc2 controls aberrant angiogenesis
 Endothelial Rictor is crucial for midgestational development and sustained and extensive FGF2 induced neovascularization in the adult Fabio Aimi 1+, Stavroula Georgiopoulou 1+, Ina Kalus 1, Fabienne Lehner
Endothelial Rictor is crucial for midgestational development and sustained and extensive FGF2 induced neovascularization in the adult Fabio Aimi 1+, Stavroula Georgiopoulou 1+, Ina Kalus 1, Fabienne Lehner
What effects will proximal or distal disease have on an waveform?
 Spectral Doppler Interpretation Director Director of of Ultrasound Ultrasound Education Education & & Quality Quality Assurance Assurance Baylor Baylor College College of of Medicine Medicine Division
Spectral Doppler Interpretation Director Director of of Ultrasound Ultrasound Education Education & & Quality Quality Assurance Assurance Baylor Baylor College College of of Medicine Medicine Division
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/10/487/eaag2476/dc1 Supplementary Materials for Gene expression profiles of brain endothelial cells during embryonic development at bulk and single-cell levels
www.sciencesignaling.org/cgi/content/full/10/487/eaag2476/dc1 Supplementary Materials for Gene expression profiles of brain endothelial cells during embryonic development at bulk and single-cell levels
Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice
 Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice Shinichi Hosokawa 1,3,Kenichiro Furuyama 1,3, Masashi Horiguchi 1,3,Yoshiki
Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice Shinichi Hosokawa 1,3,Kenichiro Furuyama 1,3, Masashi Horiguchi 1,3,Yoshiki
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Nature Neuroscience: doi: /nn Supplementary Figure 1. Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons
 Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
Department of Cell and Developmental Biology, Vanderbilt University, Nashville, TN
 UNC-6/Netrin mediates dendritic self-avoidance Cody J. Smith 1, Joseph D. Watson 1,4,5, Miri K. VanHoven 2, Daniel A. Colón-Ramos 3 and David M. Miller III 1,4,6 1 Department of Cell and Developmental
UNC-6/Netrin mediates dendritic self-avoidance Cody J. Smith 1, Joseph D. Watson 1,4,5, Miri K. VanHoven 2, Daniel A. Colón-Ramos 3 and David M. Miller III 1,4,6 1 Department of Cell and Developmental
Nature Neuroscience: doi: /nn Supplementary Figure 1. Splenic atrophy and leucopenia caused by T3 SCI.
 Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
Supplementary Information. Preferential associations between co-regulated genes reveal a. transcriptional interactome in erythroid cells
 Supplementary Information Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells Stefan Schoenfelder, * Tom Sexton, * Lyubomira Chakalova, * Nathan
Supplementary Information Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells Stefan Schoenfelder, * Tom Sexton, * Lyubomira Chakalova, * Nathan
Supplemental Figure 1. Quantification of proliferation in thyroid of WT, Ctns -/- and grafted
 Supplemental Figure 1. Quantification of proliferation in thyroid of WT, Ctns -/- and grafted Ctns -/- mice. Cells immunolabeled for the proliferation marker (Ki-67) were counted in sections (n=3 WT, n=4
Supplemental Figure 1. Quantification of proliferation in thyroid of WT, Ctns -/- and grafted Ctns -/- mice. Cells immunolabeled for the proliferation marker (Ki-67) were counted in sections (n=3 WT, n=4
The subcortical maternal complex controls symmetric division of mouse zygotes by
 The subcortical maternal complex controls symmetric division of mouse zygotes by regulating F-actin dynamics Xing-Jiang Yu 1,2, Zhaohong Yi 1, Zheng Gao 1,2, Dan-dan Qin 1,2, Yanhua Zhai 1, Xue Chen 1,
The subcortical maternal complex controls symmetric division of mouse zygotes by regulating F-actin dynamics Xing-Jiang Yu 1,2, Zhaohong Yi 1, Zheng Gao 1,2, Dan-dan Qin 1,2, Yanhua Zhai 1, Xue Chen 1,
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
Supplemental material Chen et al., http://www.jcb.org/cgi/content/full/jcb.201210119/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Lack of fast reversibility of UVR8 dissociation. (A) HEK293T
DEGREE (if applicable)
") OMB No. 0925-0001/0002 (Rev. 08/12 Approved Through 8/31/2015) NAME: Wang, Rong BIOGRAPHICAL SKETCH Provide the following information for the Senior/key personnel and other significant contributors. Follow
OMB No. 0925-0001/0002 (Rev. 08/12 Approved Through 8/31/2015) NAME: Wang, Rong BIOGRAPHICAL SKETCH Provide the following information for the Senior/key personnel and other significant contributors. Follow
Supplemental Information. Ciliary Beating Compartmentalizes. Cerebrospinal Fluid Flow in the Brain. and Regulates Ventricular Development
 Current Biology, Volume Supplemental Information Ciliary Beating Compartmentalizes Cerebrospinal Fluid Flow in the Brain and Regulates Ventricular Development Emilie W. Olstad, Christa Ringers, Jan N.
Current Biology, Volume Supplemental Information Ciliary Beating Compartmentalizes Cerebrospinal Fluid Flow in the Brain and Regulates Ventricular Development Emilie W. Olstad, Christa Ringers, Jan N.
Primary Cilia are Expressed on a Small Fraction of Cells in Trabecular Bone and Marrow
 Primary Cilia are Expressed on a Small Fraction of Cells in Trabecular Bone and Marrow Thomas R. Coughlin 1, Muriel Voisin, Ph.D. 2, Glen L. Niebur, PhD 1, Laoise M. McNamara 2. 1 University of Notre Dame,
Primary Cilia are Expressed on a Small Fraction of Cells in Trabecular Bone and Marrow Thomas R. Coughlin 1, Muriel Voisin, Ph.D. 2, Glen L. Niebur, PhD 1, Laoise M. McNamara 2. 1 University of Notre Dame,
Nature Neuroscience: doi: /nn Supplementary Figure 1. MADM labeling of thalamic clones.
 Supplementary Figure 1 MADM labeling of thalamic clones. (a) Confocal images of an E12 Nestin-CreERT2;Ai9-tdTomato brain treated with TM at E10 and stained for BLBP (green), a radial glial progenitor-specific
Supplementary Figure 1 MADM labeling of thalamic clones. (a) Confocal images of an E12 Nestin-CreERT2;Ai9-tdTomato brain treated with TM at E10 and stained for BLBP (green), a radial glial progenitor-specific
1. Microfluidic device characteristics and cell compartmentalization
 Supplementary figures: 1. Microfluidic device characteristics and cell compartmentalization Microfluidic channels were molded in PDMS (Fig. S1 A and B) over silicon masters, which were structured with
Supplementary figures: 1. Microfluidic device characteristics and cell compartmentalization Microfluidic channels were molded in PDMS (Fig. S1 A and B) over silicon masters, which were structured with
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/3/8/e1700521/dc1 Supplementary Materials for Functional vascularized lung grafts for lung bioengineering N. Valerio Dorrello, Brandon A. Guenthart, John D. O Neill,
advances.sciencemag.org/cgi/content/full/3/8/e1700521/dc1 Supplementary Materials for Functional vascularized lung grafts for lung bioengineering N. Valerio Dorrello, Brandon A. Guenthart, John D. O Neill,
Supplemental Figure S1. RANK expression on human lung cancer cells.
 Supplemental Figure S1. RANK expression on human lung cancer cells. (A) Incidence and H-Scores of RANK expression determined from IHC in the indicated primary lung cancer subgroups. The overall expression
Supplemental Figure S1. RANK expression on human lung cancer cells. (A) Incidence and H-Scores of RANK expression determined from IHC in the indicated primary lung cancer subgroups. The overall expression
CD31 5'-AGA GAC GGT CTT GTC GCA GT-3' 5 ' -TAC TGG GCT TCG AGA GCA GT-3'
 Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Supplementary. limb. bars
 Figure 1. CD163 -/- mice exhibit a similar phenotype ass WT mice in the absence of ischemic injury. a, Laser Doppler analysiss with perfusion quantitation at baseline (n= =10 per group). b, Immunostaining
Figure 1. CD163 -/- mice exhibit a similar phenotype ass WT mice in the absence of ischemic injury. a, Laser Doppler analysiss with perfusion quantitation at baseline (n= =10 per group). b, Immunostaining
Supplemental Experimental Procedures
 Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Supplementary Information
 Nature Immunology doi:1.138/ni.2477 Supplementary Information Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and instruct them with pattern recognition and motility
Nature Immunology doi:1.138/ni.2477 Supplementary Information Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and instruct them with pattern recognition and motility
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Quantification of myelin fragments in the aging brain (a) Electron microscopy on corpus callosum is shown for a 18-month-old wild type mice. Myelin fragments (arrows) were detected
Supplementary Figure 1 Quantification of myelin fragments in the aging brain (a) Electron microscopy on corpus callosum is shown for a 18-month-old wild type mice. Myelin fragments (arrows) were detected
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2697 Figure S1 Cytokeratin 5 is a specific marker for basal and intermediate cells in all mouse prostate lobes. (a) Immunofluorescence staining showing co-localization of YFP with p63 in
DOI: 10.1038/ncb2697 Figure S1 Cytokeratin 5 is a specific marker for basal and intermediate cells in all mouse prostate lobes. (a) Immunofluorescence staining showing co-localization of YFP with p63 in
Specimen. Humeral Head. Femoral Head. Objective. Femoral Condyle (medial) Supplementary Figure 1
 Supplementary Figure 1") A B Specimen Humeral Head 2 1 µm 76 µm Femoral Head Objective Femoral Condyle (medial) Supplementary Figure 1 A Femoral Head Global Cell Density Superficial Cell Density Cell Number at 1 µm Nuclei /.1
A B Specimen Humeral Head 2 1 µm 76 µm Femoral Head Objective Femoral Condyle (medial) Supplementary Figure 1 A Femoral Head Global Cell Density Superficial Cell Density Cell Number at 1 µm Nuclei /.1
Vessel malformation vascular malformation vascular malformation vascular malformation Vascular malformation vascular malformations
 A vascular malformation is another type of birthmark, or congenital (present at birth) growth, made up of arteries, veins, capillaries, or lymphatic vessels. There are several different types of malformations
A vascular malformation is another type of birthmark, or congenital (present at birth) growth, made up of arteries, veins, capillaries, or lymphatic vessels. There are several different types of malformations
Enhancement of Cranial US: Utility of Supplementary Acoustic Windows and Doppler Harriet J. Paltiel, MD
 Enhancement of Cranial US: Utility of Supplementary Acoustic Windows and Doppler Harriet J. Paltiel, MD Boston Children s Hospital Harvard Medical School None Disclosures Conventional US Anterior fontanelle
Enhancement of Cranial US: Utility of Supplementary Acoustic Windows and Doppler Harriet J. Paltiel, MD Boston Children s Hospital Harvard Medical School None Disclosures Conventional US Anterior fontanelle
Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were
 Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were electroporated with β- Catenin S33Y in PiggyBac expression
Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were electroporated with β- Catenin S33Y in PiggyBac expression
SUPPLEMENTAL INFORMATION FOR. PAX7 expression defines germline stem cells in the adult testis
 SUPPLEMENTAL INFORMATION FOR PAX7 expression defines germline stem cells in the adult testis Gina M. Aloisio, Yuji Nakada, Hatice D. Saatcioglu, Christopher G. Peña, Michael D. Baker, Edward D. Tarnawa,
SUPPLEMENTAL INFORMATION FOR PAX7 expression defines germline stem cells in the adult testis Gina M. Aloisio, Yuji Nakada, Hatice D. Saatcioglu, Christopher G. Peña, Michael D. Baker, Edward D. Tarnawa,
Pulmonary arterio-venous fistulas. Pulmonary arterio-venous fistulas. Pulmonary angiogram. Typical patient presentation
 Pulmonary arterio-venous fistulas Pulmonary arterio-venous fistulas No conflicts of interest to declare Julien I.E.Hoffman Department of Pediatrics UCSF Typical patient presentation KB was cyanotic at
Pulmonary arterio-venous fistulas Pulmonary arterio-venous fistulas No conflicts of interest to declare Julien I.E.Hoffman Department of Pediatrics UCSF Typical patient presentation KB was cyanotic at
SUPPLEMENTARY INFORMATION
 DOI:.38/ncb3399 a b c d FSP DAPI 5mm mm 5mm 5mm e Correspond to melanoma in-situ Figure a DCT FSP- f MITF mm mm MlanaA melanoma in-situ DCT 5mm FSP- mm mm mm mm mm g melanoma in-situ MITF MlanaA mm mm
DOI:.38/ncb3399 a b c d FSP DAPI 5mm mm 5mm 5mm e Correspond to melanoma in-situ Figure a DCT FSP- f MITF mm mm MlanaA melanoma in-situ DCT 5mm FSP- mm mm mm mm mm g melanoma in-situ MITF MlanaA mm mm
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. nrg1 bns101/bns101 embryos develop a functional heart and survive to adulthood (a-b) Cartoon of Talen-induced nrg1 mutation with a 14-base-pair deletion in
Supplementary Figures Supplementary Figure 1. nrg1 bns101/bns101 embryos develop a functional heart and survive to adulthood (a-b) Cartoon of Talen-induced nrg1 mutation with a 14-base-pair deletion in
Endothelial PGC 1 - α 1 mediates vascular dysfunction in diabetes
 Endothelial PGC-1α mediates vascular dysfunction in diabetes Reporter: Yaqi Zhou Date: 04/14/2014 Outline I. Introduction II. Research route & Results III. Summary Diabetes the Epidemic of the 21st Century
Endothelial PGC-1α mediates vascular dysfunction in diabetes Reporter: Yaqi Zhou Date: 04/14/2014 Outline I. Introduction II. Research route & Results III. Summary Diabetes the Epidemic of the 21st Century
sfigure 1: Detection of L-fucose in normal mouse renal cortex using the plant lectin LTL
 sfigure 1: Detection of L-fucose in normal mouse renal cortex using the plant lectin LTL LTL staining Negative control Fluorescence microscopy of normal (CL-11 +/+ ) mouse renal tissue after staining with
sfigure 1: Detection of L-fucose in normal mouse renal cortex using the plant lectin LTL LTL staining Negative control Fluorescence microscopy of normal (CL-11 +/+ ) mouse renal tissue after staining with
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References
 File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
SDF-1/CXCR4 Axis on Endothelial Progenitor Cells Regulates Bone Fracture Healing
 SDF-1/CXCR4 Axis on Endothelial Progenitor Cells Regulates Bone Fracture Healing Yohei Kawakami, M.D., Ph.D. 1,2, Masaaki Ii 3, Tomoyuki Matsumoto, M.D., Ph.D. 1, Astuhiko Kawamoto, M.D., Ph.D. 2, Yutaka
SDF-1/CXCR4 Axis on Endothelial Progenitor Cells Regulates Bone Fracture Healing Yohei Kawakami, M.D., Ph.D. 1,2, Masaaki Ii 3, Tomoyuki Matsumoto, M.D., Ph.D. 1, Astuhiko Kawamoto, M.D., Ph.D. 2, Yutaka
CONGENITAL HEART DISEASE (CHD)
") CONGENITAL HEART DISEASE (CHD) DEFINITION It is the result of a structural or functional abnormality of the cardiovascular system at birth GENERAL FEATURES OF CHD Structural defects due to specific disturbance
CONGENITAL HEART DISEASE (CHD) DEFINITION It is the result of a structural or functional abnormality of the cardiovascular system at birth GENERAL FEATURES OF CHD Structural defects due to specific disturbance
Supplementary Figure 1. Satellite cell contribution to myofibers in whole. gastrocnemius/plantaris/soleus, diaphragm, and EOM of 12 or 20 month
 Keefe et al. p. 1 Supplementary Figure 1. Satellite cell contribution to myofibers in whole muscles. (a-l) Representative cross-sections through whole TA/EDL, gastrocnemius/plantaris/soleus, diaphragm,
Keefe et al. p. 1 Supplementary Figure 1. Satellite cell contribution to myofibers in whole muscles. (a-l) Representative cross-sections through whole TA/EDL, gastrocnemius/plantaris/soleus, diaphragm,