Supplemental Data SUPPLEMENTAL EXPERIMENTAL PROCEDURES
|
|
|
- James Malone
- 6 years ago
- Views:
Transcription
1 Temporal transcription factors and their targets schedule the end of neural proliferation in Drosophila. Cédric Maurange, Louise Cheng and Alex P. Gould Supplemental Data SUPPLEMENTAL EXPERIMENTAL PROCEDURES Drosophila genetics Most genetic elements are described in Flybase (Drysdale and Crosby, 2005) or in the appropriate references provided below. The genotype of grh 370 mutants is b pr cn grh 370 bw/df(2r)pcl 7B. Genetic elements used in this study are elav-gal4 c155 ; tubp-gal80 ts ; en- GAL4; wor-gal4; UAS-CD8::GFP; UAS-GFP; UAS-YFP-pros (Choksi et al., 2006); His2AvDGFP (Clarkson and Saint, 1999); FRT19A npr 1 ; FRT40A chinmo 1 ; FRT2A Df(3L)H99; FRT82B pros 17 ; FRTG13 grh B37 ; cas 24 (Cui and Doe, 1992); UAS-cas; UAShb; UAS-Kr; UAS-pdm1; UAS-otd; UAS-kni; UAS-msh; UAS-D; svp e22 (Hiromi et al., 1993) and tubp-gal80 ts 7. nab-gal4 (NP1316 from GETDB) is a GAL4 trap inserted into nab (CG33545) that is active in late embryonic and larval neuroblast lineages (Clements et al., 2003). The clonemaker stocks used for mosaic analysis with a repressible cell marker (MARCM, Lee and Luo, 1999) were: for chromosome X: w hsflp 1, FRT19A, tubp-gal80 LL1 ; UAS-nlsLacZ 20b, UAS-mCD8::GFP LL5 ; tub-gal4 LL7. For chromosome II: w, elav-gal4 C155, hsflp 1 ; FRTG13, tubp-gal80 LL2 ; UASnlsLacZ J312, UAS-mCD8::GFP LL6 and w, elav-gal4 C155, hsflp 1 ; tubp-gal80 LL10 ; FRT40A, UAS-nlsLacZ J312, UAS-mCD8::GFP LL6. For chromosome III: w, tub-gal4, hsflp 122, UAS-GFP::6xMyc::NLS; tub-gal80 LL9, FRT2A and w, tubp-gal4, hsflp 122, UAS-GFP::6xMyc::NLS; FRT82B, tubp-gal80 LL3 and w, elav-gal4 C155, hsflp 1 ; UASnlsLacZ 20b, UAS-mCD8::GFP LL5 ; FRT82B, tubp-gal80 LL3 and w, elav-gal4 C155, hsflp 1 ; UAS-nlsLacZ 20b, UAS-mCD8::GFP LL5 ; tubp-gal80 LL9, FRT2A. Control (wildtype) genotypes were generated by crossing GAL4 drivers to yw 1118 or MARCM clonemakers to FRT2A, + or FRT82B, +. Five recombinant chromosomes were constructed for this study: UAS-cas, FRT2A; UAS-cas, UAS-abdA; FRT82B, svp e22 and FRT82B, cas 24. Due to the close proximity of FRT82B and cas, we scored recombinants between cas 24 and a
2 P{neoFRT}82B P{w +mc =lacw}l(3)l2100 L2100 chromosome (Chen et al., 2005). All 5 white - recombinants obtained from 3700 progeny raised in the presence of 625μg/ml of Geneticin were verified by failure to complement cas j1c2. Embryonic MARCM clones were induced using hsflp 122 at 37 C for 45 min, following either an 8 hr (for D analysis) or a 16 hr (for Chinmo and Br-C analyses) egg collection. Postembryonic MARCM clones were induced at 0-24 hr after larval hatching for 80 min at 37 C unless otherwise stated. Neural expression of UAS transgenes was temporally restricted to postembryonic stages by combining elav-gal4 C155 or nab-gal4 with tubp-gal80 ts (McGuire et al., 2003) and shifting from 18 C to 30 C from ~4 hr for induction during all larval stages. elav-gal4 C155 /+; UAS-cas/tub-GAL80 ts larvae do not detectably express UAS-cas when raised at 18 C but the switch to 30 C induces robust expression. For transient induction of Pros, UAS-YFP-pros/tub-GAL80 ts ; nab-gal4/+ larvae were raised at 18ºC and shifted to 30ºC during the or hr time windows. Antibodies The primary antibodies used for immunohistochemistry were: 1:2000 rabbit anti-cas (gift of W. Odenwald), 1:1000 rabbit anti-chinmo (gift of T. Lee), 1:100 rabbit anti-grh (gift of B. Bello), 1:2000 rabbit anti-corebr-c (gift of J-A. Lepesant), 1:1000 mouse anti-svp (gift of Y. Hiromi), 1:400 guinea-pig anti-eve (gift of D. Kosman and J. Reinitz), 1:10 mouse anti-pdm1/nubbin (gift of S. Cohen), 1:20 rat anti-brdu (Oxford Biotechnology), 1:100 mouse anti-core Br-C (Developmental Studies Hybridoma Bank, University of Iowa), 1:20 rat anti-pros (gift of F. Matsuzaki), 1:1000 rabbit anti-cnn (gift of T. Kaufman), 1:1000 rabbit anti-d (gift of J. Nambu), 1:400 rabbit anti-ph3 (Upstate Biotechnology), 1:300 rat anti-ph3 (Abcam), 1:50 mouse anti-mira (gift of F. Matsuzaki), 1:7000 rabbit anti- -galactosidase (Cappel), 1:100 mouse anti- galactosidase (Promega), 1:1000 rabbit anti-gfp (Molecular Probes), 1:200 mouse anti- GFP (Molecular Probes) and 1:100 rat anti-cd8 (Caltag). Secondary goat antibodies were conjugated to Alexa Fluor 488 or 568 (Molecular Probes) or to Cy5 (Jackson ImmunoResearch).
3 SUPPLEMENTAL REFERENCES Choksi, S. P., Southall, T. D., Bossing, T., Edoff, K., de Wit, E., Fischer, B. E., van Steensel, B., Micklem, G., and Brand, A. H. (2006). Prospero acts as a binary switch between self-renewal and differentiation in Drosophila neural stem cells. Dev Cell 11, Clarkson, M., and Saint, R. (1999). A His2AvDGFP fusion gene complements a lethal His2AvD mutant allele and provides an in vivo marker for Drosophila chromosome behavior. DNA Cell Biol 18, Clements, M., Duncan, D., and Milbrandt, J. (2003). Drosophila NAB (dnab) is an orphan transcriptional co-repressor required for correct CNS and eye development. Dev Dyn 226, Cui, X., and Doe, C. Q. (1992). ming is expressed in neuroblast sublineages and regulates gene expression in the Drosophila central nervous system. Development 116, Drysdale, R. A., and Crosby, M. A. (2005). FlyBase: genes and gene models. Nucleic Acids Res 33, D Grosskortenhaus, R., Robinson, K. J., and Doe, C. Q. (2006). Pdm and Castor specify late-born motor neuron identity in the NB7-1 lineage. Genes Dev 20, Hiromi, Y., Mlodzik, M., West, S. R., Rubin, G. M., and Goodman, C. S. (1993). Ectopic expression of seven-up causes cell fate changes during ommatidial assembly. Development 118, Isshiki, T., Pearson, B., Holbrook, S., and Doe, C. Q. (2001). Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny. Cell 106, Kambadur, R., Koizumi, K., Stivers, C., Nagle, J., Poole, S. J., and Odenwald, W. F. (1998). Regulation of POU genes by castor and hunchback establishes layered compartments in the Drosophila CNS. Genes Dev 12, Lee, T., and Luo, L. (1999). Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron 22, Ma, Y., Niemitz, E. L., Nambu, P. A., Shan, X., Sackerson, C., Fujioka, M., Goto, T., and Nambu, J. R. (1998). Gene regulatory functions of Drosophila fish-hook, a high mobility group domain Sox protein. Mech Dev 73, McGuire, S. E., Le, P. T., Osborn, A. J., Matsumoto, K., and Davis, R. L. (2003). Spatiotemporal rescue of memory dysfunction in Drosophila. Science 302, Zhu, S., Lin, S., Kao, C. F., Awasaki, T., Chiang, A. S., and Lee, T. (2006). Gradients of the Drosophila Chinmo BTB-zinc finger protein govern neuronal temporal identity. Cell 127,

4 Supplemental Figure 1
5 Figure S1. Early- and late-born neurons express Chinmo and Br-C respectively Panels in A-D show single confocal sections. (A) Confocal sections of the embryonic CNS (anterior to the left) showing most, if not all, neurons are Chinmo + at stage 17. Neuroblast 7-1 expresses Hb, Kr, Pdm1 and Cas in sequence to generate the U1-U5 Eve + motor neurons (Isshiki et al., 2001) and all five of these are Chinmo +. Dashed box is enlarged in triptych showing U2, U3 and U4 (dotted circles). (B) Confocal sections of the ventral thoracic CNS showing postembryonic neuroblasts (large cells) and their last-born progeny (small cells) marked with nab>mcd8::gfp (mgfp). At 48 hr, thoracic neuroblasts, postembryonic-born (mgfp + ) and embryonicborn (mgfp - ) cells express Chinmo and Br-C is absent from all postembryonic-born neural cells. This contrasts with mushroom-body neuroblasts which are reported not to express Chinmo (Zhu et al., 2006). At 55 hr, most but not all (dotted circle) neuroblasts weakly express Chinmo and some also begin to express Br-C (dotted circle). By 65 hr, most neuroblasts express Br-C but not Chinmo and begin to generate Br-C + neurons (data not shown). At 96 hr, Br-C (but not Chinmo) is expressed in neuroblasts and late-born neurons. Note that early-born Chinmo + progeny are located below the plane of this section (see Figure 1C) and that immature neurons do not express Br-C immediately after birth. (C-D) npr 1 (C) and chinmo 1 (D) MARCM clones, induced at 0-24 hr and marked with elav>mgfp, express the normal layer-specific patterns of Chinmo and Br-C respectively at 96 hr.
 and their progeny (small cells)")
6 Supplemental Figure 2 Figure S2. Thoracic postembryonic neuroblasts express Cas and Svp sequentially Single confocal sections of the ventral thoracic CNS with postembryonic neuroblasts (large cells) and their progeny (small cells) marked with nab>mcd8::gfp. At 30 hr, some larval neuroblasts leaving quiescence express Cas in a similar pattern in each segment (T1, T2 and T3) but no neuroblasts express Svp. At 40 hr, some neuroblasts (arrowheads and enlargement of dotted box) express Svp in a pattern resembling Cas at 30 hr. At 60 hr and 96 hr, neuroblasts no longer express Cas or Svp but new-born progeny continue to express Cas strongly, even though they must have been generated from Cas - neuroblasts. This contrasts with the temporally-linked Cas expression pattern observed in embryonic neuroblasts and their progeny (Isshiki et al., 2001).
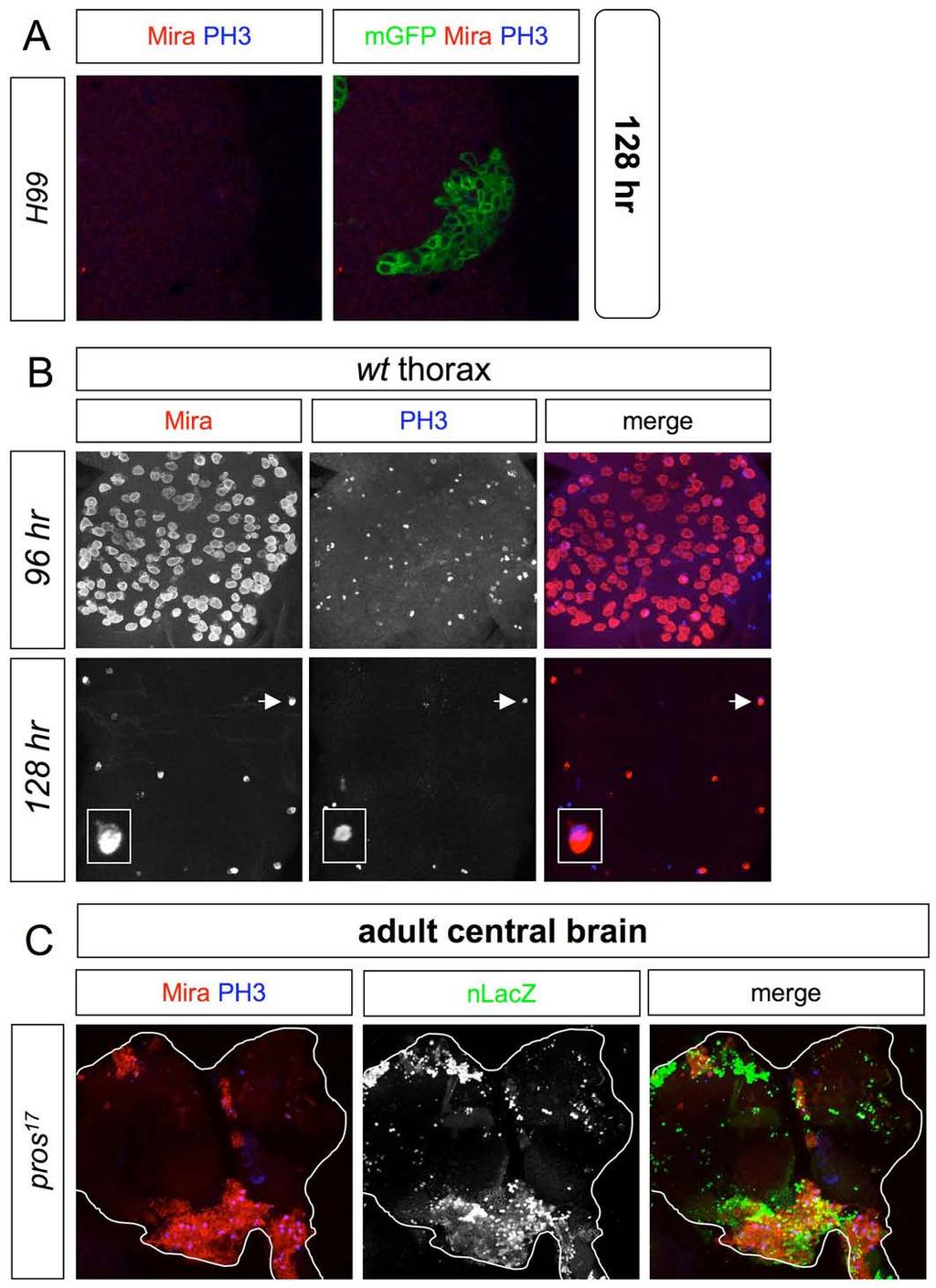
7 Supplemental Figure 3
8 Figure S3. Loss of Pros but not RHG activity prolongs thoracic and central brain neuroblast divisions Panels in A-C show ventral views of the CNS. (A) A thoracic MARCM clone lacking the activity of RHG proapoptotic genes (Df(3L)H99), induced at 0-24 hr and marked with elav>mgfp, does not contain PH3 + or Mira + progenitors at 128 hr. (B) Thoracic neuromeres contain numerous Mira + neuroblasts (some of which are PH3 + ) at 96 hr but lose most of these by 128 hr. Inset shows an enlargement of one of the last remaining Mira + neuroblasts to undergo cell-cycle exit (arrow). (C) Ventral view of 7-day adult central brain with several pros 17 MARCM clones, induced at 0-24 hr and marked with tub>nlacz, each containing numerous Mira + adult progenitors, many of which are PH3 +. Such progenitors are absent from control tissue surrounding the clones.
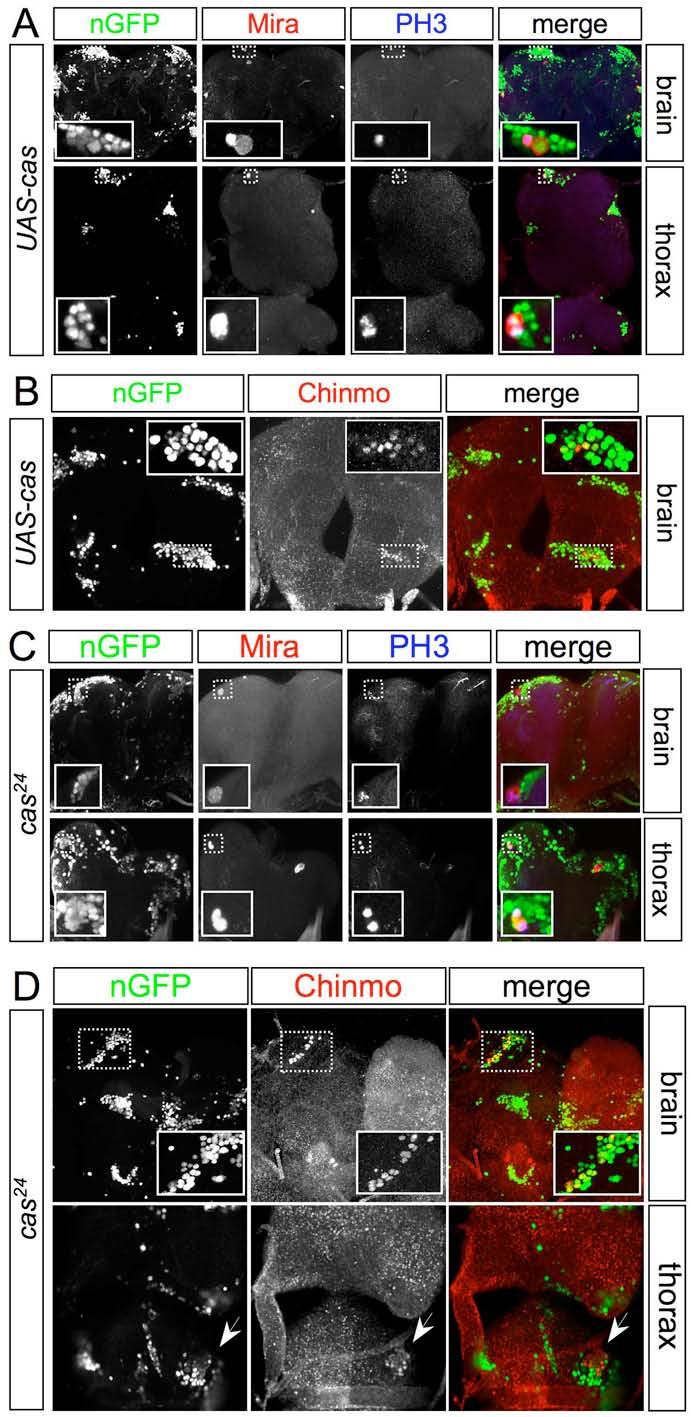
9 Supplemental Figure 4
10 Figure S4. Loss or mis-expression of Cas leads to neuroblast divisions and Chinmo + neurons in the adult Ventral views of the central brain and thoracic neuromeres of 3-day old adults. MARCM UAS-cas clones, induced at 0-24 hr (A-B) and cas 24 clones, induced during embryogenesis (C-D), were marked with tub>ngfp. Both types of clone often retain a single Mira + neuroblast that is mitotically active (some are PH3 + ) in the adult and continues to generate superficial (late-born) Chinmo + cells. Deep (early-born) neurons in surrounding control tissue (GFP - ) have down regulated Chinmo before adult eclosion.

11 Supplemental Figure 5 Figure S5. Svp is required for nuclear Pros in neuroblasts at 120 hr A svp e22 MARCM clone, induced at 0-24 hr, marked with tub>ngfp and analyzed at 120 hr, lacks Br-C + neurons, indicating that the postembryonic temporal series is stalled. Although Pros is absent from the nucleus of the interphase Mira + neuroblast (arrow), it remains normally expressed in the nucleus of the GMC (arrowhead).

12 Supplemental Figure 6
13 Figure S6. Dichaete is down regulated in most wild-type neuroblasts during development (A) Single confocal sections of the embryonic CNS at stages At stage 9, D is expressed in most medial (m) and intermediate (i) but not lateral (l) Mira + neuroblasts. From stage 10 onwards, many lateral neuroblasts express D. As previously reported, D is expressed in both the nucleus and cytoplasm (Ma et al., 1998). (B) At 55 hr, a small subset of thoracic Mira + neuroblasts (and other cells) expresses D. (C) At 65 hr, D is not expressed in wild-type Mira + abdominal neuroblasts. Three dl lineages (marked with nab>mgfp) in segments A3-A5 are shown. (D) Histogram showing that the percentage of thoracic Mira + neuroblasts expressing D progressively decreases. It is 47% in stage 12 embryos (n=12 hemisegments, sd=11.3), 50.7% in stage 14 embryos (n=10, sd=11.2), 41.5% in stage 15 embryos (n=8,sd=10.5), 23.6% in 55 hr larvae (n=18, sd=3.5) and only 14.6% in 96 hr larvae (n=18, sd=3). Asterisk, P < for stage 14 embryos versus 96 hr larvae.
 Ventral views of T2 and T3 neuromeres at embryonic stage 14 showing that early continuous expression of Hb within the Engrailed")
14 Supplemental Figure 7 Figure S7. Embryonic Cas activates Grh and down regulates D in neuroblasts (A) Ventral views of T2 and T3 neuromeres at embryonic stage 14 showing that early continuous expression of Hb within the Engrailed stripes (brackets) of en>hb embryos or loss-of-function of Cas in cas 24 embryos prevents Grh expression in Mira + neuroblasts. (B) Ventral views of T2 and T3 neuromeres at embryonic stage 12. In most Mira + neuroblasts, Grh has yet to be expressed in wild-type (wt) embryos but within the Engrailed stripes (brackets) of en>cas embryos it is prematurely activated. Consistent with previous reports, we also observe that Cas represses Pdm (Grosskortenhaus et al., 2006; Kambadur et al., 1998). (C-D) Ventral views of T2-A1 neuromeres at embryonic stages 14 and 15. (C) At embryonic stage 15, D down regulation in Mira + neuroblasts, particularly in lateral
15 progenitors (arrows) is prevented by early and continuous expression of Hb within the Engrailed stripes (brackets) of en>hb embryos, or by loss-of-function of Cas in cas 24 embryos. More neuroblasts with M-phase Mira crescents are found in en>hb and cas 24 than in wild-type embryos at this stage, suggesting that entry into quiescence is delayed. (D) At embryonic stage 14, early and persistent expression of Cas in wor>cas embryos leads to precocious down regulation of D in lateral Mira + neuroblasts (arrows).


16 Supplemental Figure 8 Figure S8. Persistent adult neuroblasts in svp e22 clones retain Grh expression Postembryonic svp e22 MARCM clones were induced at 0-24 hr, marked with tub>ngfp, and analyzed in the central brain of 3-day old adults. Inset shows enlargement of a clone (arrow) containing a Mira + Grh + neuroblast. Supplemental Figure 9 Figure S9. The Chinmo->Br-C switch is not blocked in grh B37 clones Single confocal section of a grh B37 MARCM lineage in a thoracic neuromere, induced at 0-24hr and marked with elav>mgfp, showing that although these postembryonic lineages are smaller-than-normal they nevertheless contain both Chinmo + and Br-C + cells at 96 hr (n=34/34 clones).
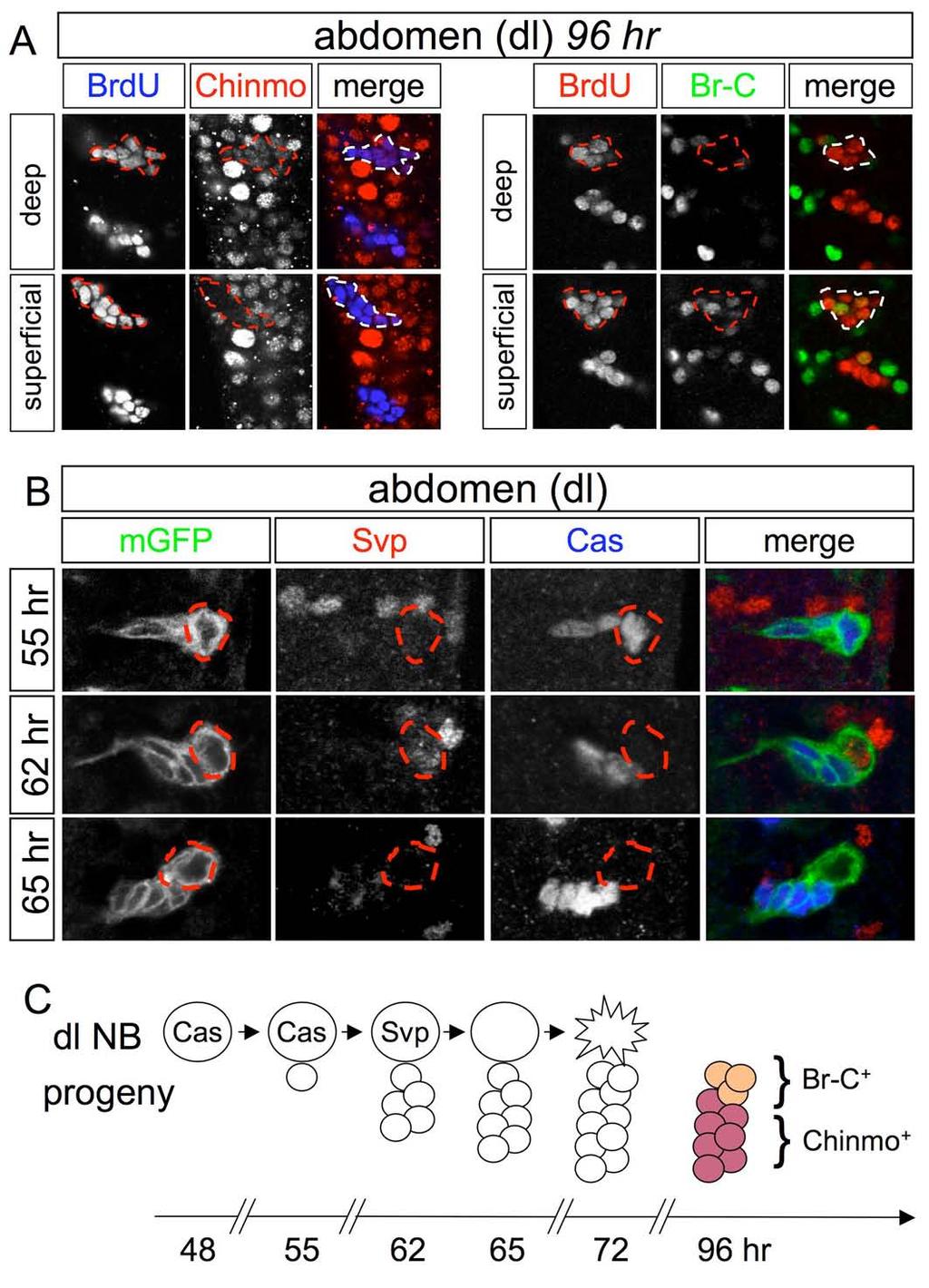
17 Supplemental Figure 10
18 Figure S10. dl neuroblasts express Cas and Svp sequentially and generate Chinmo + and Br-C + progeny (A) Single confocal sections of two wild-type dl lineages in the central abdomen at 96 hr. The postembryonic component of these lineages is labeled with BrdU (the most anterior dl lineage is dotted) and contains Br-C + cells and somewhat deeper Chinmo + cells. (B) Single confocal sections of a wild-type dl lineage, labeled with nab>mgfp. Both the dl neuroblast and its progeny express Cas at 55 hr but only the progeny expresses Cas at 62 hr and 65 hr. Svp is expressed in dl neuroblasts at 62 hr but not at 55 hr or 65 hr. (C) Schematic representation of Cas and Svp expression in the dl neuroblast (large circle) and its progeny (small circles). For clarity, Cas expression is omitted from postmitotic cells and Chinmo and Br-C are only shown at 96 hr.

19 Supplemental Figure 11
20 Figure S11. Cas and Svp are required for timely apoptosis of dl neuroblasts (A) Abdominal neuromeres containing several svp e22 clones, induced at 0-24 hr and marked with elav>mgfp or elav>nlslacz. At 96 hr, dl clones retain a single Mira + neuroblast (arrow), PH3 + progenitors (inset) and supernumerary mgfp + cells. (B) Abdominal neuromeres containing a single cas 24 clone, induced during embryogenesis and marked with tub>ngfp. At 96 hr, this dl clone retains a single Mira + PH3 + neuroblast, a PH3 + GMC (inset) and supernumerary mgfp + cells. (C) Abdominal neuromeres continuously expressing neural Cas during postembryonic stages (UAS-Cas, referring to elav-gal4 C155 ; UAS-cas/tub-GAL80 ts ). At 96 hr, dl and other abdominal lineages contain more postembryonic cells (BrdU + ) than normal and retain Mira + neuroblasts (some of which are PH3 + ). (D) Quantification of supernumerary (GFP + ) cells generated in dl MARCM clones marked with tub>ngfp that are wild-type (0 or 1 cell only) or svp e22 (m=22.5 cells, sd=5.5, n=12). Asterisk indicates P < (E) Numbers of postembryonic-born (BrdU + ) cells in dl lineages for wt, UAS-cas (elav- GAL4 C155 ; UAS-cas/tub-GAL80 ts ) and UAS-D (elav-gal4 C155 ; UAS-D; tub-gal80 ts ) larvae at 96 hr. Asterisks indicate P < for wt (m=9.5, sd=2, n=17) versus UAS-cas (m=16.6, sd=2.4 and n=18) or UAS-D (m=23.8, sd=5.6 and n=10). (F) Single confocal sections of Df(3L)H99, svp e22 and UAS-cas clones, all induced at 0-24hr, and cas 24 clones, induced during embryogenesis. All four types of MARCM clone generate larger-than-normal dl lineages by 96 hr but the supernumerary (tub>ngfp labeled) cells in Df(3L)H99 clones are Br-C + but do not express Chinmo, whereas those in svp e22, UAS-cas and cas 24 clones are Chinmo + but do not express Br-C. This indicates that the Chinmo->Br-C transition is blocked in svp e22, UAS-cas and cas 24 clones but not in Df(3L)H99 clones.
 Abdominal neuromeres at 96 hr from elav C155 -GAL4; tub-gal80 ts larvae")
21 Supplemental Figure 12 Figure S12. Postembryonic mis-expression of Hb, Kr or Pdm1 blocks neuroblast apoptosis and the Chinmo->Br-C switch in dl lineages (A-E) Abdominal neuromeres at 96 hr from elav C155 -GAL4; tub-gal80 ts larvae carrying either UAS-otd (A), UAS-hb (B,C), UAS-kr (D) or UAS-pdm1 (E) transgenes. Continuous postembryonic neural expression of Hb, Kr, Pdm1 but not Otd produces supernumerary cells, some of which are PH3 +, in the vm, vl and dl lineages (BrdU + ). Control elav C155 - GAL4; tub-gal80 ts ; UAS-hb larvae raised at the non-induced temperature (18 C) display no such over proliferation (B). UAS-knirps and UAS-msh gave similar negative results to UAS-otd (data not shown). (F) A superficial confocal section of a dl lineage (BrdU + ) from an elav C155 -GAL4; tub- GAL80 ts ; UAS-hb larva at 96hr. Many of the supernumerary late-born neurons generated after continuous postembryonic neural expression of Hb are Chinmo + but none are Br-C +.
 Postembryonic mis-expression of D (UAS-D refers to elav-gal4 C155 ; UAS-D; tub-")
22 Supplemental Figure 13 Figure S13. Postembryonic mis-expression of D blocks dl neuroblast apoptosis but not Grh expression nor the Chinmo->Br-C switch (A) Postembryonic mis-expression of D (UAS-D refers to elav-gal4 C155 ; UAS-D; tub- GAL80 ts larvae) leads to persistent dl neuroblasts that are Grh + (and PH3 + in some cases) at 96 hr. Postembryonic cells are labeled with BrdU. (B) Postembryonic mis-expression of D (UAS-D refers to UAS-D/tub-GAL80 ts ; nab- GAL4, UAS-mGFP larvae) leads to persistent dl neuroblasts that generate supernumerary Chinmo - Br-C + neurons at 96 hr.

23 Supplemental Figure 14 Figure S14. Postembryonic blockade of the temporal series does not prevent dl neuroblasts from expressing AbdA At 96 hr, AbdA expression is retained by the persistent dl neuroblasts in UAS-cas (elav C155 -GAL4; tub-gal80ts/uas-cas) larvae (A) and also in svp e22 MARCM clones (B), induced at 0-24 hr and marked with elav>mgfp.
24 Supplemental Movie 1. Self-renewing asymmetric division of a neuroblast at 96 hr Time-lapse movie of a 96 hr thoracic neuroblast (large mitotic cell, ~10μm diameter) and smaller surrounding cells expressing the chromatin label His2AvDGFP. The interval between frames (single confocal sections) is 30 seconds and anaphase and telophase of this type-i neuroblast take ~3 min to complete. Supplemental Movie 2. Atypical division of a neuroblast at 120 hr Time-lapse movie of a 120 hr thoracic neuroblast (intermediate-size mitotic cell, ~7μm diameter) and smaller surrounding cells expressing the chromatin label His2AvDGFP. The interval between frames (single confocal sections) is 30 seconds and anaphase and telophase of this type-i neuroblast take ~16 min to complete.
glial cells missing and gcm2 Cell-autonomously Regulate Both Glial and Neuronal
 glial cells missing and gcm2 Cell-autonomously Regulate Both Glial and Neuronal Development in the Visual System of Drosophila Carole Chotard, Wendy Leung and Iris Salecker Supplemental Data Supplemental
glial cells missing and gcm2 Cell-autonomously Regulate Both Glial and Neuronal Development in the Visual System of Drosophila Carole Chotard, Wendy Leung and Iris Salecker Supplemental Data Supplemental
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature07173 SUPPLEMENTARY INFORMATION Supplementary Figure Legends: Supplementary Figure 1: Model of SSC and CPC divisions a, Somatic stem cells (SSC) reside adjacent to the hub (red), self-renew
doi: 10.1038/nature07173 SUPPLEMENTARY INFORMATION Supplementary Figure Legends: Supplementary Figure 1: Model of SSC and CPC divisions a, Somatic stem cells (SSC) reside adjacent to the hub (red), self-renew
The Tumor Suppressors Brat and Numb Regulate Transit-Amplifying Neuroblast Lineages in Drosophila
 Article The Tumor Suppressors Brat and Numb Regulate Transit-Amplifying Neuroblast Lineages in Drosophila Sarah K. Bowman, 1 Vivien Rolland, 1 Joerg Betschinger, 1,3 Kaolin A. Kinsey, 2 Gregory Emery,
Article The Tumor Suppressors Brat and Numb Regulate Transit-Amplifying Neuroblast Lineages in Drosophila Sarah K. Bowman, 1 Vivien Rolland, 1 Joerg Betschinger, 1,3 Kaolin A. Kinsey, 2 Gregory Emery,
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Expression of escargot (esg) and genetic approach for achieving IPC-specific knockdown. (a) esg MH766 -Gal4 UAS-cd8GFP (green) and esg-lacz B7-2-22 (red) show similar expression
Supplementary Figure 1 Expression of escargot (esg) and genetic approach for achieving IPC-specific knockdown. (a) esg MH766 -Gal4 UAS-cd8GFP (green) and esg-lacz B7-2-22 (red) show similar expression
Supplementary Figure S1: TIPF reporter validation in the wing disc.
 Supplementary Figure S1: TIPF reporter validation in the wing disc. a,b, Test of put RNAi. a, In wildtype discs the Dpp target gene Sal (red) is expressed in a broad stripe in the centre of the ventral
Supplementary Figure S1: TIPF reporter validation in the wing disc. a,b, Test of put RNAi. a, In wildtype discs the Dpp target gene Sal (red) is expressed in a broad stripe in the centre of the ventral
The Hunchback temporal transcription factor establishes, but is not required to maintain, early-born neuronal identity
 Hirono et al. Neural Development (2017) 12:1 DOI 10.1186/s13064-017-0078-1 RESEARCH ARTICLE Open Access The Hunchback temporal transcription factor establishes, but is not required to maintain, early-born
Hirono et al. Neural Development (2017) 12:1 DOI 10.1186/s13064-017-0078-1 RESEARCH ARTICLE Open Access The Hunchback temporal transcription factor establishes, but is not required to maintain, early-born
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2419 Figure S1 NiGFP localization in Dl mutant dividing SOPs. a-c) time-lapse analysis of NiGFP (green) in Dl mutant SOPs (H2B-RFP, red; clones were identified by the loss of nlsgfp) showing
DOI: 10.1038/ncb2419 Figure S1 NiGFP localization in Dl mutant dividing SOPs. a-c) time-lapse analysis of NiGFP (green) in Dl mutant SOPs (H2B-RFP, red; clones were identified by the loss of nlsgfp) showing
Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster
 RESEARCH ARTICLE 53 Development 137, 53-61 (2010) doi:10.1242/dev.041749 Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster James W. Truman*,, Wanda
RESEARCH ARTICLE 53 Development 137, 53-61 (2010) doi:10.1242/dev.041749 Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster James W. Truman*,, Wanda
Brat Is a Miranda Cargo Protein that Promotes Neuronal Differentiation and Inhibits Neuroblast Self-Renewal
 Developmental Cell 10, 441 449, April, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.devcel.2006.01.017 Brat Is a Miranda Cargo Protein that Promotes Neuronal Differentiation and Inhibits Neuroblast Self-Renewal
Developmental Cell 10, 441 449, April, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.devcel.2006.01.017 Brat Is a Miranda Cargo Protein that Promotes Neuronal Differentiation and Inhibits Neuroblast Self-Renewal
Department of Cell and Developmental Biology, Vanderbilt University, Nashville, TN
 UNC-6/Netrin mediates dendritic self-avoidance Cody J. Smith 1, Joseph D. Watson 1,4,5, Miri K. VanHoven 2, Daniel A. Colón-Ramos 3 and David M. Miller III 1,4,6 1 Department of Cell and Developmental
UNC-6/Netrin mediates dendritic self-avoidance Cody J. Smith 1, Joseph D. Watson 1,4,5, Miri K. VanHoven 2, Daniel A. Colón-Ramos 3 and David M. Miller III 1,4,6 1 Department of Cell and Developmental
Control of neuronal cell fate and number by integration of distinct daughter cell proliferation modes with temporal progression
 Control of neuronal cell fate and number by integration of distinct daughter cell proliferation modes with temporal progression Carina Ulvklo, Ryan MacDonald, Caroline Bivik, Magnus Baumgardt, Daniel Karlsson
Control of neuronal cell fate and number by integration of distinct daughter cell proliferation modes with temporal progression Carina Ulvklo, Ryan MacDonald, Caroline Bivik, Magnus Baumgardt, Daniel Karlsson
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
let-7-complex MicroRNAs Regulate the Temporal Identity of Drosophila Mushroom Body Neurons via chinmo
 Short Article let-7-complex MicroRNAs Regulate the Temporal Identity of Drosophila Mushroom Body Neurons via chinmo Yen-Chi Wu, 1 Ching-Huan Chen, 1 Adam Mercer, 1 and Nicholas S. Sokol 1, * 1 Department
Short Article let-7-complex MicroRNAs Regulate the Temporal Identity of Drosophila Mushroom Body Neurons via chinmo Yen-Chi Wu, 1 Ching-Huan Chen, 1 Adam Mercer, 1 and Nicholas S. Sokol 1, * 1 Department
marker. DAPI labels nuclei. Flies were 20 days old. Scale bar is 5 µm. Ctrl is
 Supplementary Figure 1. (a) Nos is detected in glial cells in both control and GFAP R79H transgenic flies (arrows), but not in deletion mutant Nos Δ15 animals. Repo is a glial cell marker. DAPI labels
Supplementary Figure 1. (a) Nos is detected in glial cells in both control and GFAP R79H transgenic flies (arrows), but not in deletion mutant Nos Δ15 animals. Repo is a glial cell marker. DAPI labels
Supplementary Figure 1 Madm is not required in GSCs and hub cells. (a,b) Act-Gal4-UAS-GFP (a), Act-Gal4-UAS- GFP.nls (b,c) is ubiquitously expressed
 Act-Gal4-UAS-GFP (a), Act-Gal4-UAS- GFP.nls (b,c) is ubiquitously expressed") Supplementary Figure 1 Madm is not required in GSCs and hub cells. (a,b) Act-Gal4-UAS-GFP (a), Act-Gal4-UAS- GFP.nls (b,c) is ubiquitously expressed in the testes. The testes were immunostained with GFP
Supplementary Figure 1 Madm is not required in GSCs and hub cells. (a,b) Act-Gal4-UAS-GFP (a), Act-Gal4-UAS- GFP.nls (b,c) is ubiquitously expressed in the testes. The testes were immunostained with GFP
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. nrg1 bns101/bns101 embryos develop a functional heart and survive to adulthood (a-b) Cartoon of Talen-induced nrg1 mutation with a 14-base-pair deletion in
Supplementary Figures Supplementary Figure 1. nrg1 bns101/bns101 embryos develop a functional heart and survive to adulthood (a-b) Cartoon of Talen-induced nrg1 mutation with a 14-base-pair deletion in
Cell Birth and Death. Chapter Three
 Cell Birth and Death Chapter Three Neurogenesis All neurons and glial cells begin in the neural tube Differentiated into neurons rather than ectoderm based on factors we have already discussed If these
Cell Birth and Death Chapter Three Neurogenesis All neurons and glial cells begin in the neural tube Differentiated into neurons rather than ectoderm based on factors we have already discussed If these
Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice
 Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice Shinichi Hosokawa 1,3,Kenichiro Furuyama 1,3, Masashi Horiguchi 1,3,Yoshiki
Impact of Sox9 Dosage and Hes1-mediated Notch Signaling in Controlling the Plasticity of Adult Pancreatic Duct Cells in Mice Shinichi Hosokawa 1,3,Kenichiro Furuyama 1,3, Masashi Horiguchi 1,3,Yoshiki
100 mm Sucrose. +Berberine +Quinine
 8 mm Sucrose Probability (%) 7 6 5 4 3 Wild-type Gr32a / 2 +Caffeine +Berberine +Quinine +Denatonium Supplementary Figure 1: Detection of sucrose and bitter compounds is not affected in Gr32a / flies.
8 mm Sucrose Probability (%) 7 6 5 4 3 Wild-type Gr32a / 2 +Caffeine +Berberine +Quinine +Denatonium Supplementary Figure 1: Detection of sucrose and bitter compounds is not affected in Gr32a / flies.
Supplementary Figures
 J. Cell Sci. 128: doi:10.1242/jcs.173807: Supplementary Material Supplementary Figures Fig. S1 Fig. S1. Description and/or validation of reagents used. All panels show Drosophila tissues oriented with
J. Cell Sci. 128: doi:10.1242/jcs.173807: Supplementary Material Supplementary Figures Fig. S1 Fig. S1. Description and/or validation of reagents used. All panels show Drosophila tissues oriented with
Viktorin, G. and Riebli, N. and Popkova, A. and Giangrande, A. and Reichert, H.
 Institutional Repository of the University of Basel University Library Schoenbeinstrasse 18-20 CH-4056 Basel, Switzerland http://edoc.unibas.ch/ Year: 2011 Multipotent neural stem cells generate glial
Institutional Repository of the University of Basel University Library Schoenbeinstrasse 18-20 CH-4056 Basel, Switzerland http://edoc.unibas.ch/ Year: 2011 Multipotent neural stem cells generate glial
BIOL2005 WORKSHEET 2008
 BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
Supplementary Figure 1. Nature Neuroscience: doi: /nn.4547
 Supplementary Figure 1 Characterization of the Microfetti mouse model. (a) Gating strategy for 8-color flow analysis of peripheral Ly-6C + monocytes from Microfetti mice 5-7 days after TAM treatment. Living
Supplementary Figure 1 Characterization of the Microfetti mouse model. (a) Gating strategy for 8-color flow analysis of peripheral Ly-6C + monocytes from Microfetti mice 5-7 days after TAM treatment. Living
Nature Neuroscience: doi: /nn Supplementary Figure 1. Neuron class-specific arrangements of Khc::nod::lacZ label in dendrites.
 Supplementary Figure 1 Neuron class-specific arrangements of Khc::nod::lacZ label in dendrites. Staining with fluorescence antibodies to detect GFP (Green), β-galactosidase (magenta/white). (a, b) Class
Supplementary Figure 1 Neuron class-specific arrangements of Khc::nod::lacZ label in dendrites. Staining with fluorescence antibodies to detect GFP (Green), β-galactosidase (magenta/white). (a, b) Class
mir-7a regulation of Pax6 in neural stem cells controls the spatial origin of forebrain dopaminergic neurons
 Supplemental Material mir-7a regulation of Pax6 in neural stem cells controls the spatial origin of forebrain dopaminergic neurons Antoine de Chevigny, Nathalie Coré, Philipp Follert, Marion Gaudin, Pascal
Supplemental Material mir-7a regulation of Pax6 in neural stem cells controls the spatial origin of forebrain dopaminergic neurons Antoine de Chevigny, Nathalie Coré, Philipp Follert, Marion Gaudin, Pascal
Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells.
 Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells. a. Schematic of the V-ATPase proton pump macro-complex structure. The V1 complex is composed of
Supplementary Fig. 1 V-ATPase depletion induces unique and robust phenotype in Drosophila fat body cells. a. Schematic of the V-ATPase proton pump macro-complex structure. The V1 complex is composed of
A Genetic Program for Embryonic Development
 Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Supplementary Figure 1
 Supplementary Figure 1 Kif1a RNAi effect on basal progenitor differentiation Related to Figure 2. Representative confocal images of the VZ and SVZ of rat cortices transfected at E16 with scrambled or Kif1a
Supplementary Figure 1 Kif1a RNAi effect on basal progenitor differentiation Related to Figure 2. Representative confocal images of the VZ and SVZ of rat cortices transfected at E16 with scrambled or Kif1a
The subcortical maternal complex controls symmetric division of mouse zygotes by
 The subcortical maternal complex controls symmetric division of mouse zygotes by regulating F-actin dynamics Xing-Jiang Yu 1,2, Zhaohong Yi 1, Zheng Gao 1,2, Dan-dan Qin 1,2, Yanhua Zhai 1, Xue Chen 1,
The subcortical maternal complex controls symmetric division of mouse zygotes by regulating F-actin dynamics Xing-Jiang Yu 1,2, Zhaohong Yi 1, Zheng Gao 1,2, Dan-dan Qin 1,2, Yanhua Zhai 1, Xue Chen 1,
Early cell death (FGF) B No RunX transcription factor produced Yes No differentiation
 B No RunX transcription factor produced Yes No differentiation") Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Solution Key - Practice Questions Question 1 a) A recent publication has shown that the fat stem cells (FSC) can act as bone stem cells to repair cavities in the skull, when transplanted into immuno-compromised
Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were
 Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were electroporated with β- Catenin S33Y in PiggyBac expression
Supplementary Figure 1. Electroporation of a stable form of β-catenin causes masses protruding into the IV ventricle. HH12 chicken embryos were electroporated with β- Catenin S33Y in PiggyBac expression
Control of neuronal cell fate and number by integration of distinct daughter cell proliferation modes with temporal progression
 678 AND STEM CELLS Development 139, 678-689 (2012) doi:10.1242/dev.074500 2012. Published by The Company of Biologists Ltd Control of neuronal cell fate and number by integration of distinct daughter cell
678 AND STEM CELLS Development 139, 678-689 (2012) doi:10.1242/dev.074500 2012. Published by The Company of Biologists Ltd Control of neuronal cell fate and number by integration of distinct daughter cell
SUPPLEMENTARY INFORMATION
 DOI: 1.138/ncb222 / b. WB anti- WB anti- ulin Mitotic index (%) 14 1 6 2 T (h) 32 48-1 1 2 3 4 6-1 4 16 22 28 3 33 e. 6 4 2 Time (min) 1-6- 11-1 > 1 % cells Figure S1 depletion leads to mitotic defects
DOI: 1.138/ncb222 / b. WB anti- WB anti- ulin Mitotic index (%) 14 1 6 2 T (h) 32 48-1 1 2 3 4 6-1 4 16 22 28 3 33 e. 6 4 2 Time (min) 1-6- 11-1 > 1 % cells Figure S1 depletion leads to mitotic defects
Zhu et al, page 1. Supplementary Figures
 Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
Zhu et al, page 1 Supplementary Figures Supplementary Figure 1: Visual behavior and avoidance behavioral response in EPM trials. (a) Measures of visual behavior that performed the light avoidance behavior
SUPPLEMENTARY DATA. Supplementary Table 2. Antibodies used for Immunofluoresence. Supplementary Table 3. Real-time PCR primer sequences.
 Supplementary Table 2. Antibodies used for Immunofluoresence. Antibody Dilution Source Goat anti-pdx1 1:100 R&D Systems Rabbit anti-hnf6 1:100 Santa Cruz Biotechnology Mouse anti-nkx6.1 1:200 Developmental
Supplementary Table 2. Antibodies used for Immunofluoresence. Antibody Dilution Source Goat anti-pdx1 1:100 R&D Systems Rabbit anti-hnf6 1:100 Santa Cruz Biotechnology Mouse anti-nkx6.1 1:200 Developmental
Supplemental Information. Otic Mesenchyme Cells Regulate. Spiral Ganglion Axon Fasciculation. through a Pou3f4/EphA4 Signaling Pathway
 Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Supplemental Information. Tissue Myeloid Progenitors Differentiate. into Pericytes through TGF-b Signaling. in Developing Skin Vasculature
 Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Wang and Page-McCaw, http://www.jcb.org/cgi/content/full/jcb.201403084/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Extracellular anti-wg staining is specific. Note
Supplemental material Wang and Page-McCaw, http://www.jcb.org/cgi/content/full/jcb.201403084/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Extracellular anti-wg staining is specific. Note
Programmed Cell Death (apoptosis)
") Programmed Cell Death (apoptosis) Stereotypic death process includes: membrane blebbing nuclear fragmentation chromatin condensation and DNA framentation loss of mitochondrial integrity and release of
Programmed Cell Death (apoptosis) Stereotypic death process includes: membrane blebbing nuclear fragmentation chromatin condensation and DNA framentation loss of mitochondrial integrity and release of
Postembryonic Development of Amplifying Neuroblast Lineages in the Drosophila Brain: Proliferation, Differentiation and Projection Patterns.
 Postembryonic Development of Amplifying Neuroblast Lineages in the Drosophila Brain: Proliferation, Differentiation and Projection Patterns. Inauguraldissertation zur Erlangung der Würde eines Doktors
Postembryonic Development of Amplifying Neuroblast Lineages in the Drosophila Brain: Proliferation, Differentiation and Projection Patterns. Inauguraldissertation zur Erlangung der Würde eines Doktors
A Cxcl12-Cxcr4 Chemokine Signaling Pathway Defines
 Supplemental Data A Cxcl12-Cxcr4 Chemokine Signaling Pathway Defines the Initial Trajectory of Mammalian Motor Axons Ivo Lieberam, Dritan Agalliu, Takashi Nagasawa, Johan Ericson, and Thomas M. Jessell
Supplemental Data A Cxcl12-Cxcr4 Chemokine Signaling Pathway Defines the Initial Trajectory of Mammalian Motor Axons Ivo Lieberam, Dritan Agalliu, Takashi Nagasawa, Johan Ericson, and Thomas M. Jessell
Supporting Information
 Supporting Information Fig. S1. Overexpression of Rpr causes progenitor cell death. (A) TUNEL assay of control intestines. No progenitor cell death could be observed, except that some ECs are undergoing
Supporting Information Fig. S1. Overexpression of Rpr causes progenitor cell death. (A) TUNEL assay of control intestines. No progenitor cell death could be observed, except that some ECs are undergoing
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.
 Light microscopic pictures show mouse embryos at developmental stage E12.") Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Supplementary Figure 1: Hsp60 / IEC mice are embryonically lethal (A) Light microscopic pictures show mouse embryos at developmental stage E12.5 and E13.5 prepared from uteri of dams and subsequently genotyped.
Supplemental Information. Myocardial Polyploidization Creates a Barrier. to Heart Regeneration in Zebrafish
 Developmental Cell, Volume 44 Supplemental Information Myocardial Polyploidization Creates a Barrier to Heart Regeneration in Zebrafish Juan Manuel González-Rosa, Michka Sharpe, Dorothy Field, Mark H.
Developmental Cell, Volume 44 Supplemental Information Myocardial Polyploidization Creates a Barrier to Heart Regeneration in Zebrafish Juan Manuel González-Rosa, Michka Sharpe, Dorothy Field, Mark H.
50mM D-Glucose. Percentage PI. L-Glucose
 Monica Dus, Minrong Ai, Greg S.B Suh. Taste-independent nutrient selection is mediated by a brain-specific Na+/solute cotransporter in Drosophila. Control + Phlorizin mm D-Glucose 1 2mM 1 L-Glucose Gr5a;Gr64a
Monica Dus, Minrong Ai, Greg S.B Suh. Taste-independent nutrient selection is mediated by a brain-specific Na+/solute cotransporter in Drosophila. Control + Phlorizin mm D-Glucose 1 2mM 1 L-Glucose Gr5a;Gr64a
Ahtiainen et al., http :// /cgi /content /full /jcb /DC1
 Supplemental material JCB Ahtiainen et al., http ://www.jcb.org /cgi /content /full /jcb.201512074 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Distinct distribution of different cell cycle phases in the
Supplemental material JCB Ahtiainen et al., http ://www.jcb.org /cgi /content /full /jcb.201512074 /DC1 THE JOURNAL OF CELL BIOLOGY Figure S1. Distinct distribution of different cell cycle phases in the
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
Control of proliferation activation in quiescent neuroblasts of the Drosophila central nervous system
 Development 121, 1173-1182 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1173 Control of proliferation activation in quiescent neuroblasts of the Drosophila central nervous system
Development 121, 1173-1182 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1173 Control of proliferation activation in quiescent neuroblasts of the Drosophila central nervous system
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
Cell migration in Drosophila optic lobe neurons is controlled by eyeless/pax6
 687 Development 138, 687-693 (2011) doi:10.1242/dev.056069 2011. Published by The Company of Biologists Ltd Cell migration in Drosophila optic lobe neurons is controlled by eyeless/pax6 Javier Morante
687 Development 138, 687-693 (2011) doi:10.1242/dev.056069 2011. Published by The Company of Biologists Ltd Cell migration in Drosophila optic lobe neurons is controlled by eyeless/pax6 Javier Morante
Developmental Biology
 Developmental Biology 327 (2009) 288 300 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology The HLH protein Extramacrochaetae is required
Developmental Biology 327 (2009) 288 300 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology The HLH protein Extramacrochaetae is required
effects on organ development. a-f, Eye and wing discs with clones of ε j2b10 show no
 Supplementary Figure 1. Loss of function clones of 14-3-3 or 14-3-3 show no significant effects on organ development. a-f, Eye and wing discs with clones of 14-3-3ε j2b10 show no obvious defects in Elav
Supplementary Figure 1. Loss of function clones of 14-3-3 or 14-3-3 show no significant effects on organ development. a-f, Eye and wing discs with clones of 14-3-3ε j2b10 show no obvious defects in Elav
Biology Developmental Biology Spring Quarter Midterm 1 Version A
 Biology 411 - Developmental Biology Spring Quarter 2013 Midterm 1 Version A 75 Total Points Open Book Choose 15 out the 20 questions to answer (5 pts each). Only the first 15 questions that are answered
Biology 411 - Developmental Biology Spring Quarter 2013 Midterm 1 Version A 75 Total Points Open Book Choose 15 out the 20 questions to answer (5 pts each). Only the first 15 questions that are answered
Neurogenesis and neuronal circuit formation in the Drosophila visual center
 The Japanese Society of Developmental Biologists Develop. Growth Differ. (2014) 56, 491 498 doi: 10.1111/dgd.12151 Review Article Neurogenesis and neuronal circuit formation in the Drosophila visual center
The Japanese Society of Developmental Biologists Develop. Growth Differ. (2014) 56, 491 498 doi: 10.1111/dgd.12151 Review Article Neurogenesis and neuronal circuit formation in the Drosophila visual center
Early Embryonic Development
 Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Nature Neuroscience: doi: /nn Supplementary Figure 1. MADM labeling of thalamic clones.
 Supplementary Figure 1 MADM labeling of thalamic clones. (a) Confocal images of an E12 Nestin-CreERT2;Ai9-tdTomato brain treated with TM at E10 and stained for BLBP (green), a radial glial progenitor-specific
Supplementary Figure 1 MADM labeling of thalamic clones. (a) Confocal images of an E12 Nestin-CreERT2;Ai9-tdTomato brain treated with TM at E10 and stained for BLBP (green), a radial glial progenitor-specific
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Ras V12 expression in the entire eye-antennal disc does not cause invasive tumours. a, Eye-antennal discs expressing Ras V12 in all cells (marked with GFP, green) overgrow moderately
Supplementary Figure 1. Ras V12 expression in the entire eye-antennal disc does not cause invasive tumours. a, Eye-antennal discs expressing Ras V12 in all cells (marked with GFP, green) overgrow moderately
Midterm 1. Number of students Score. Mean: 73 Median: 75 Top Score: 98
 Midterm 1 14 12 Number of students 10 8 6 4 2 0 35-40 41-45 Mean: 73 Median: 75 Top Score: 98 46-50 51-55 56-60 61-65 66-70 71-75 Score 76-80 81-85 86-90 91-95 96-100 Write your name and student ID# on
Midterm 1 14 12 Number of students 10 8 6 4 2 0 35-40 41-45 Mean: 73 Median: 75 Top Score: 98 46-50 51-55 56-60 61-65 66-70 71-75 Score 76-80 81-85 86-90 91-95 96-100 Write your name and student ID# on
Bidirectional Notch signaling regulates Drosophila intestinal stem cell multipotency
 RESEARCH RESEARCH ARTICLE SUMMARY STEM CELL REGULATION Bidirectional Notch signaling regulates Drosophila intestinal stem cell multipotency Zheng Guo and Benjamin Ohlstein* INTRODUCTION: In the Drosophila
RESEARCH RESEARCH ARTICLE SUMMARY STEM CELL REGULATION Bidirectional Notch signaling regulates Drosophila intestinal stem cell multipotency Zheng Guo and Benjamin Ohlstein* INTRODUCTION: In the Drosophila
Lineage diversity in the Drosophila nervous system Yohanns Bellaïche* and François Schweisguth
 418 Lineage diversity in the Drosophila nervous system Yohanns Bellaïche* and François Schweisguth The detailed descriptions of cellular lineages in the Drosophila nervous system have provided the foundations
418 Lineage diversity in the Drosophila nervous system Yohanns Bellaïche* and François Schweisguth The detailed descriptions of cellular lineages in the Drosophila nervous system have provided the foundations
Control of neural stem cell self-renewal and differentiation in Drosophila
 Institutional Repository of the University of Basel University Library Schoenbeinstrasse 18-20 CH-4056 Basel, Switzerland http://edoc.unibas.ch/ Year: 2014 Control of neural stem cell self-renewal and
Institutional Repository of the University of Basel University Library Schoenbeinstrasse 18-20 CH-4056 Basel, Switzerland http://edoc.unibas.ch/ Year: 2014 Control of neural stem cell self-renewal and
CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions
 CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions Yi Feng, Peng Cui, Xiaowei Lu, Brian Hsueh, Fredrik Möller Billig, Livia Zarnescu Yanez, Raju Tomer, Derek
CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions Yi Feng, Peng Cui, Xiaowei Lu, Brian Hsueh, Fredrik Möller Billig, Livia Zarnescu Yanez, Raju Tomer, Derek
Capu and Spire Assemble a Cytoplasmic Actin Mesh
 Developmental Cell 13 Supplemental Data Capu and Spire Assemble a Cytoplasmic Actin Mesh that Maintains Microtubule Organization in the Drosophila Oocyte Katja Dahlgaard, Alexandre A.S.F. Raposo, Teresa
Developmental Cell 13 Supplemental Data Capu and Spire Assemble a Cytoplasmic Actin Mesh that Maintains Microtubule Organization in the Drosophila Oocyte Katja Dahlgaard, Alexandre A.S.F. Raposo, Teresa
Article. The Color-Vision Circuit in the Medulla of Drosophila. Javier Morante 1 and Claude Desplan 1, * 1
 Current Biology 18, 1 13, April 22, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.02.075 The Color-Vision Circuit in the Medulla of Drosophila Article Javier Morante 1 and Claude Desplan
Current Biology 18, 1 13, April 22, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.02.075 The Color-Vision Circuit in the Medulla of Drosophila Article Javier Morante 1 and Claude Desplan
Compartmental organization of the Drosophila genital imaginal discs
 Development 124, 205-218 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV8366 205 Compartmental organization of the Drosophila genital imaginal discs Elizabeth H. Chen 1 and Bruce
Development 124, 205-218 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV8366 205 Compartmental organization of the Drosophila genital imaginal discs Elizabeth H. Chen 1 and Bruce
Linkage Mapping in Drosophila Melanogaster
 Linkage Mapping in Drosophila Melanogaster Genetics: Fall 2012 Joshua Hanau Introduction: An experiment was performed in order to determine the presence and degree of gene linkage in Drosophila Melanogaster.
Linkage Mapping in Drosophila Melanogaster Genetics: Fall 2012 Joshua Hanau Introduction: An experiment was performed in order to determine the presence and degree of gene linkage in Drosophila Melanogaster.
Karen L.P. McNally, Amy S. Fabritius, Marina L. Ellefson, Jonathan R. Flynn, Jennifer A. Milan, and Francis J. McNally
 Developmental Cell, Volume 22 Supplemental Information Kinesin-1 Prevents Capture of the Oocyte Meiotic Spindle by the Sperm Aster Karen L.P. McNally, Amy S. Fabritius, Marina L. Ellefson, Jonathan R.
Developmental Cell, Volume 22 Supplemental Information Kinesin-1 Prevents Capture of the Oocyte Meiotic Spindle by the Sperm Aster Karen L.P. McNally, Amy S. Fabritius, Marina L. Ellefson, Jonathan R.
Segmentation. Serial Homology. Hox genes
 Segmentation Hox genes Serial Homology William Bateson 1861-1926 Homeosis: a variation in which something has been changed into the likeness of something else Calvin Bridges at Columbia bithorax 1923 1
Segmentation Hox genes Serial Homology William Bateson 1861-1926 Homeosis: a variation in which something has been changed into the likeness of something else Calvin Bridges at Columbia bithorax 1923 1
Developmental Biology
 Developmental iology 356 (2011) 553 565 ontents lists available at ScienceDirect Developmental iology journal homepage: www.elsevier.com/developmentalbiology Multipotent neural stem cells generate glial
Developmental iology 356 (2011) 553 565 ontents lists available at ScienceDirect Developmental iology journal homepage: www.elsevier.com/developmentalbiology Multipotent neural stem cells generate glial
Supplementary Fig. 1 Blocking shh function at the protein level confirms its role as a guidance cue for postcommissural axons.
 Supplementary Fig. 1 Blocking shh function at the protein level confirms its role as a guidance cue for postcommissural axons. As an alternative method to demonstrate the role of shh as a guidance cue
Supplementary Fig. 1 Blocking shh function at the protein level confirms its role as a guidance cue for postcommissural axons. As an alternative method to demonstrate the role of shh as a guidance cue
Polarity and Segmentation. Chapter Two
 Polarity and Segmentation Chapter Two Polarization Entire body plan is polarized One end is different than the other Head vs. Tail Anterior vs. Posterior Front vs. Back Ventral vs. Dorsal Majority of neural
Polarity and Segmentation Chapter Two Polarization Entire body plan is polarized One end is different than the other Head vs. Tail Anterior vs. Posterior Front vs. Back Ventral vs. Dorsal Majority of neural
Neuroepithelial Cells and Neural Differentiation
 Neuroepithelial Cells and Neural Differentiation Neurulation The cells of the neural tube are NEUROEPITHELIAL CELLS Neural crest cells migrate out of neural tube Neuroepithelial cells are embryonic stem
Neuroepithelial Cells and Neural Differentiation Neurulation The cells of the neural tube are NEUROEPITHELIAL CELLS Neural crest cells migrate out of neural tube Neuroepithelial cells are embryonic stem
Tramtrack controls glial number and identity in the Drosophila embryonic CNS
 Development 128, 4093-4101 (2001) Printed in Great Britain The Company of Biologists Limited 2001 DEV7891 4093 Tramtrack controls glial number and identity in the Drosophila embryonic CNS Paul Badenhorst
Development 128, 4093-4101 (2001) Printed in Great Britain The Company of Biologists Limited 2001 DEV7891 4093 Tramtrack controls glial number and identity in the Drosophila embryonic CNS Paul Badenhorst
PEER REVIEW FILE. Reviewers' Comments: Reviewer #1 (Remarks to the Author):
:") PEER REVIEW FILE Reviewers' Comments: Reviewer #1 (Remarks to the Author): In this manuscript entitled "Neural circuits for long-term water-reward memory processing in thirsty Drosophila", the authors
PEER REVIEW FILE Reviewers' Comments: Reviewer #1 (Remarks to the Author): In this manuscript entitled "Neural circuits for long-term water-reward memory processing in thirsty Drosophila", the authors
HHS Public Access Author manuscript Curr Opin Neurobiol. Author manuscript; available in PMC 2018 February 01.
 Timing temporal transitions during brain development Anthony M. Rossi 1,*, Vilaiwan M. Fernandes 1,*, and Claude Desplan 1,2, 1 Department of Biology, New York University, New York, New York 10003, USA
Timing temporal transitions during brain development Anthony M. Rossi 1,*, Vilaiwan M. Fernandes 1,*, and Claude Desplan 1,2, 1 Department of Biology, New York University, New York, New York 10003, USA
Isoform-Specific Control of Male Neuronal Differentiation and Behavior in Drosophila by the fruitless Gene
 Current Biology 16, 1063 1076, June 6, 2006 ª2006 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2006.04.039 Isoform-Specific Control of Male Neuronal Differentiation and Behavior in Drosophila by
Current Biology 16, 1063 1076, June 6, 2006 ª2006 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2006.04.039 Isoform-Specific Control of Male Neuronal Differentiation and Behavior in Drosophila by
Problem Set 5 KEY
 2006 7.012 Problem Set 5 KEY ** Due before 5 PM on THURSDAY, November 9, 2006. ** Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. You are studying the development
2006 7.012 Problem Set 5 KEY ** Due before 5 PM on THURSDAY, November 9, 2006. ** Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. You are studying the development
Regionalization of the nervous system. Paul Garrity 7.68J/9.013J February 25, 2004
 Regionalization of the nervous system Paul Garrity 7.68J/9.013J February 25, 2004 Patterning along: Rostral/Caudal (AP) axis Dorsal/Ventral (DV) axis Start with DV axial patterning in Spinal Cord Dorsal/Ventral
Regionalization of the nervous system Paul Garrity 7.68J/9.013J February 25, 2004 Patterning along: Rostral/Caudal (AP) axis Dorsal/Ventral (DV) axis Start with DV axial patterning in Spinal Cord Dorsal/Ventral
Temporal Patterning of Neuroblasts Controls Notch-Mediated Cell Survival through Regulation of Hid or Reaper
 Temporal Patterning of Neuroblasts Controls Notch-Mediated Cell Survival through Regulation of Hid or Reaper Claire Bertet, 1 Xin Li, 1 Ted Erclik, 1 Matthieu Cavey, 1 Brent Wells, 1 and Claude Desplan
Temporal Patterning of Neuroblasts Controls Notch-Mediated Cell Survival through Regulation of Hid or Reaper Claire Bertet, 1 Xin Li, 1 Ted Erclik, 1 Matthieu Cavey, 1 Brent Wells, 1 and Claude Desplan
Nature Neuroscience: doi: /nn Supplementary Figure 1. Splenic atrophy and leucopenia caused by T3 SCI.
 Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
C. elegans Embryonic Development
 Autonomous Specification in Tunicate development Autonomous & Conditional Specification in C. elegans Embryonic Development Figure 8.36 Bilateral Symmetry in the Egg of the Tunicate Styela partita Fig.
Autonomous Specification in Tunicate development Autonomous & Conditional Specification in C. elegans Embryonic Development Figure 8.36 Bilateral Symmetry in the Egg of the Tunicate Styela partita Fig.
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3443 In the format provided by the authors and unedited. Supplementary Figure 1 TC and SC behaviour during ISV sprouting. (a) Predicted outcome of TC division and competitive Dll4-Notch-mediated
DOI: 10.1038/ncb3443 In the format provided by the authors and unedited. Supplementary Figure 1 TC and SC behaviour during ISV sprouting. (a) Predicted outcome of TC division and competitive Dll4-Notch-mediated
Gene Regulation. Bacteria. Chapter 18: Regulation of Gene Expression
 Chapter 18: Regulation of Gene Expression A Biology 2013 1 Gene Regulation rokaryotes and eukaryotes alter their gene expression in response to their changing environment In multicellular eukaryotes, gene
Chapter 18: Regulation of Gene Expression A Biology 2013 1 Gene Regulation rokaryotes and eukaryotes alter their gene expression in response to their changing environment In multicellular eukaryotes, gene
SUPPLEMENTARY INFORMATION
 DOI: 0.038/ncb33 a b c 0 min 6 min 7 min (fixed) DIC -GFP, CenpF 3 µm Nocodazole Single optical plane -GFP, CenpF Max. intensity projection d µm -GFP, CenpF, -GFP CenpF 3-D rendering e f 0 min 4 min 0
DOI: 0.038/ncb33 a b c 0 min 6 min 7 min (fixed) DIC -GFP, CenpF 3 µm Nocodazole Single optical plane -GFP, CenpF Max. intensity projection d µm -GFP, CenpF, -GFP CenpF 3-D rendering e f 0 min 4 min 0
Meigo governs dendrite targeting specificity by modulating Ephrin level and N-glycosylation
 Meigo governs dendrite targeting specificity by modulating Ephrin level and N-glycosylation Sayaka U. Sekine, Shuka Haraguchi, Kinhong Chao, Tomoko Kato, Liqun Luo, Masayuki Miura, and Takahiro Chihara
Meigo governs dendrite targeting specificity by modulating Ephrin level and N-glycosylation Sayaka U. Sekine, Shuka Haraguchi, Kinhong Chao, Tomoko Kato, Liqun Luo, Masayuki Miura, and Takahiro Chihara
Supplementary Figure 1
 Supplementary Figure 1 Global TeNT expression effectively impairs synaptic transmission. Injection of 100 pg tent mrna leads to a reduction of vesicle mediated synaptic transmission in the spinal cord
Supplementary Figure 1 Global TeNT expression effectively impairs synaptic transmission. Injection of 100 pg tent mrna leads to a reduction of vesicle mediated synaptic transmission in the spinal cord
Cellular Reproduction
 9 Cellular Reproduction section 1 Cellular Growth Before You Read Think about the life cycle of a human. On the lines below, write some of the stages that occur in the life cycle of a human. In this section,
9 Cellular Reproduction section 1 Cellular Growth Before You Read Think about the life cycle of a human. On the lines below, write some of the stages that occur in the life cycle of a human. In this section,
Erzsebet Kokovay, Susan Goderie, Yue Wang, Steve Lotz, Gang Lin, Yu Sun, Badrinath Roysam, Qin Shen,
 Cell Stem Cell, Volume 7 Supplemental Information Adult SVZ Lineage Cells Home to and Leave the Vascular Niche via Differential Responses to SDF1/CXCR4 Signaling Erzsebet Kokovay, Susan Goderie, Yue Wang,
Cell Stem Cell, Volume 7 Supplemental Information Adult SVZ Lineage Cells Home to and Leave the Vascular Niche via Differential Responses to SDF1/CXCR4 Signaling Erzsebet Kokovay, Susan Goderie, Yue Wang,
Negative regulation of atonal in proneural cluster formation of Drosophila R8 photoreceptors
 Proc. Natl. Acad. Sci. USA Vol. 96, pp. 5055 5060, April 1999 Developmental Biology Negative regulation of atonal in proneural cluster formation of Drosophila R8 photoreceptors CHIEN-KUO CHEN AND CHENG-TING
Proc. Natl. Acad. Sci. USA Vol. 96, pp. 5055 5060, April 1999 Developmental Biology Negative regulation of atonal in proneural cluster formation of Drosophila R8 photoreceptors CHIEN-KUO CHEN AND CHENG-TING
Genesis of cerebellar interneurons and the prevention of neural DNA damage require XRCC1.
 Genesis of cerebellar interneurons and the prevention of neural DNA damage require XRCC1. Youngsoo Lee, Sachin Katyal, Yang Li, Sherif F. El-Khamisy, Helen R. Russell, Keith W. Caldecott and Peter J. McKinnon.
Genesis of cerebellar interneurons and the prevention of neural DNA damage require XRCC1. Youngsoo Lee, Sachin Katyal, Yang Li, Sherif F. El-Khamisy, Helen R. Russell, Keith W. Caldecott and Peter J. McKinnon.
Development Supplementary information
 Supplemental Materials and Methods Mosaic clonal analysis GSC and SP clones were induced with the FLP/FRT-mediated mitotic recombination technique (Xu and Rubin, 1993) in files with following genotypes:
Supplemental Materials and Methods Mosaic clonal analysis GSC and SP clones were induced with the FLP/FRT-mediated mitotic recombination technique (Xu and Rubin, 1993) in files with following genotypes:
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Courtheoux et al., http://www.jcb.org/cgi/content/full/jcb.200902093/dc1 S1 Figure S2. Visualization of multiple merotelic attachments.
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Courtheoux et al., http://www.jcb.org/cgi/content/full/jcb.200902093/dc1 S1 Figure S2. Visualization of multiple merotelic attachments.
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
Drosophila Hey is a target of Notch in asymmetric divisions during embryonic and larval neurogenesis
 AND STEM CELLS 191 Development 137, 191-201 (2010) doi:10.1242/dev.043604 Drosophila Hey is a target of Notch in asymmetric divisions during embryonic and larval neurogenesis Maria Monastirioti 1, Nikolaos
AND STEM CELLS 191 Development 137, 191-201 (2010) doi:10.1242/dev.043604 Drosophila Hey is a target of Notch in asymmetric divisions during embryonic and larval neurogenesis Maria Monastirioti 1, Nikolaos
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES 1 Supplementary Figure 1, Adult hippocampal QNPs and TAPs uniformly express REST a-b) Confocal images of adult hippocampal mouse sections showing GFAP (green), Sox2 (red), and REST
SUPPLEMENTARY FIGURES 1 Supplementary Figure 1, Adult hippocampal QNPs and TAPs uniformly express REST a-b) Confocal images of adult hippocampal mouse sections showing GFAP (green), Sox2 (red), and REST
Supplementary information
 Supplementary information 1 Supplementary Figure 1. CALM regulates autophagy. (a). Quantification of LC3 levels in the experiment described in Figure 1A. Data are mean +/- SD (n > 3 experiments for each
Supplementary information 1 Supplementary Figure 1. CALM regulates autophagy. (a). Quantification of LC3 levels in the experiment described in Figure 1A. Data are mean +/- SD (n > 3 experiments for each
Haematopoietic stem cells
 Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Haematopoietic stem cells Neil P. Rodrigues, DPhil NIH Centre for Biomedical Research Excellence in Stem Cell Biology Boston University School of Medicine neil.rodrigues@imm.ox.ac.uk Haematopoiesis: An
Nature Medicine doi: /nm.2860
 Supplemental Figure Legends Supplemental Figure 1: Hypomorphic expression of IFT88 results in olfactory signaling proteins no longer localizing to the ciliary layer. (a) ACIII localizes to the cilia and
Supplemental Figure Legends Supplemental Figure 1: Hypomorphic expression of IFT88 results in olfactory signaling proteins no longer localizing to the ciliary layer. (a) ACIII localizes to the cilia and
Fertilization. mitosis. N sperm pronucleus. fertilization. N pronucleus (after sperm enters egg) 2N zygote nucleus
 2N zygote nucleus") Fertilization 2N pronucleus (suspended in meiotic metaphase) N polar body reduction division N pronucleus (after sperm enters egg) mitosis 2N zygote nucleus N polar bodies (2): disintegrate membrane formation
Fertilization 2N pronucleus (suspended in meiotic metaphase) N polar body reduction division N pronucleus (after sperm enters egg) mitosis 2N zygote nucleus N polar bodies (2): disintegrate membrane formation
Nature Genetics: doi: /ng Supplementary Figure 1. Immunofluorescence (IF) confirms absence of H3K9me in met-2 set-25 worms.
 confirms absence of H3K9me in met-2 set-25 worms.") Supplementary Figure 1 Immunofluorescence (IF) confirms absence of H3K9me in met-2 set-25 worms. IF images of wild-type (wt) and met-2 set-25 worms showing the loss of H3K9me2/me3 at the indicated developmental
Supplementary Figure 1 Immunofluorescence (IF) confirms absence of H3K9me in met-2 set-25 worms. IF images of wild-type (wt) and met-2 set-25 worms showing the loss of H3K9me2/me3 at the indicated developmental